
Comparative Studies on the Distribution and Population of ...
7
FULL PAPER Anatomy Comparative Studies on the Distribution and Population of Immunocompetent Cells in Bovine Hemal Node, Lymph Node and Spleen Weidong ZHANG 1) , Tetsuo NASU 1) , Yoshinao Z. HOSAKA 2) and Masahiro YASUDA 1) * 1) Department of Veterinary Anatomy, Faculty of Agriculture, University of Miyazaki, 1–1 Gakuen Kibanadai-Nishi, Miyazaki 889–2192, Japan 2) Department of Veterinary Anatomy, Faculty of Agriculture, Tottori University, 4–101 Koyama-Minami, Tottori 680–8550, Japan (Received 31 August 2011/Accepted 30 October 2011/Published online in J-STAGE 11 November 2011) ABSTRACT. The distribution and population of immunocompetent cells in bovine hemal node, mesenteric lymph node and spleen were analyzed comparatively by immunohistochemistry and flow cytometry. Many CD8 + cells, CD172a + cells and γδ T cells were found in the lymphatic cord along the sinus of the hemal node and the splenic red pulp. A few CD8 + cells and γδ T cells were distributed dif- fusely in the paracortex and medullary cord of the mesenteric lymph node. Many germinal centers were recognized in the lymphatic regions such as the cortex and white pulp of these lymphoid organs. The populations of CD8 + cells and γδ T cells in the hemal node and the spleen were higher than those of the mesenteric lymph node. In addition, the populations of CD21 + cells and MHC class II + cells in the hemal node and the mesenteric lymph node were higher than those of the spleen. The results suggest that the hemal node has an important role in both cellular and humoral immunity as well as the lymph node and the spleen in cattle. KEY WORDS: comparative study, flow cytometry, hemal node, immunocompetent cells, immunohistochemistry. doi: 10.1292/jvms.11-0405; J. Vet. Med. Sci. 74(4): 405–411, 2012 The hemal node (HN) is a hematopoietic and lymphoid organ that is found in some mammals such as humans, rats and ruminants. In ruminants, HNs are located in the sub- cutaneous region of the head, the mesenteric region and along the large blood vessels such as the aorta in the tho- rax and abdomen; however, the distribution and number of HNs in these regions seem to be diverse [1, 8, 28, 36]. The histological structure of HNs resembles that of lymph nodes and consists of a lymphoid region, in which the cor- tex and paracortex are recognized, and a sinusoid region, in which the lymphatic cord and sinus are formed [8, 9, 33]. The HN sinus is filled with numerous erythrocytes, lacks lymphatic vessels and only connects small blood vessels [1, 26, 32, 42]. In addition, the HN structure also resembles that of the spleen, which consists of white pulp, in which the germinal center is formed, and red pulp, in which the splenic sinus is filled with numerous erythrocytes [26, 42]. The antibody-producing ability and immunological competence of rat HN after antigen challenge have been described [26, 34]. The possibilities of erythropoiesis and erythrophagocytosis in bovine HNs were described by Cerutti and Guerrero [10]. However, the immunological role of the bovine HN has not yet been determined. In the present paper, we analyzed the immunological function of bovine HNs by comparing the distribution and population of immunocompetent cells in the HNs of calves with those of the mesenteric lymph nodes (MLNs) and spleen. MATERIALS AND METHODS Animals and collection of specimens: Japanese black calves (5–8 months old, n=10) were obtained from local farms in Miyazaki Prefecture. The animals were killed by electric shock following the combination of xylazine (0.2 mg/kg) and pentobarbital (15 mg/kg) intravenous injection. All animal procedures were approved by the Institutional Animal Care and Use Committee of the University of Mi- yazaki. The HNs, MLNs and spleens were collected for immunohistochemistry (IHC) and flow cytometry (FACS). The specimens for IHC were mounted in OCT embedding compound (Sakura Finetek Japan Co., Ltd., Tokyo, Japan) on Cryomold (Sakura Finetek Japan Co., Ltd.), frozen on dry ice, and then stored at −80°C until used. The speci- mens for FACS were transported on ice and then treated as described below. For histological analysis, HNs were fixed with 10% buffered formalin. The samples were passed through etha- nol and toluene and embedded in paraffin. The samples were cut into 4-µm thick sections and stained with hema- toxylin-eosin (HE) stain. Monoclonal antibodies (mAbs): mAbs specific for calf CD4 (IL-A11), CD8 (CACT80c), CD21 (GB25A), CD172a (DH59B), TCR1-N24: δ chain specific (GB21A) and MHC class II (TH14B) were purchased from VMRD Inc. (Pull- man, WA, U.S.A.). A 1:500 dilution of each mAb was used for IHC and FACS. IHC: Cryostat sections were stained with mAbs by us- ing the indirect immunoperoxidase technique described by Yasuda et al. [40]. In brief, the sections (7–10 µm thick) were air-dried on slides and fixed with ice-cold acetone for 10 min. To block any nonspecific binding, the sections *CORRESPONDENCE TO: YASUDA, M., Department of Veterinary Anatomy, Faculty of Agriculture, University of Miyazaki, 1–1 Gakuen Kibanadai-Nishi, Miyazaki 889–2192, Japan. e-mail: [email protected] ©2012 The Japanese Society of Veterinary Science
Transcript of Comparative Studies on the Distribution and Population of ...

FULL PAPER Anatomy
Comparative Studies on the Distribution and Population of Immunocompetent Cells in Bovine Hemal Node, Lymph Node and Spleen
Weidong ZHANG1), Tetsuo NASU1), Yoshinao Z. HOSAKA2) and Masahiro YASUDA1)*
1)Department of Veterinary Anatomy, Faculty of Agriculture, University of Miyazaki, 1–1 Gakuen Kibanadai-Nishi, Miyazaki 889–2192, Japan
2)Department of Veterinary Anatomy, Faculty of Agriculture, Tottori University, 4–101 Koyama-Minami, Tottori 680–8550, Japan
(Received 31 August 2011/Accepted 30 October 2011/Published online in J-STAGE 11 November 2011)
abstract. The distribution and population of immunocompetent cells in bovine hemal node, mesenteric lymph node and spleen were analyzed comparatively by immunohistochemistry and flow cytometry. Many CD8+ cells, CD172a+ cells and γδ T cells were found in the lymphatic cord along the sinus of the hemal node and the splenic red pulp. A few CD8+ cells and γδ T cells were distributed dif-fusely in the paracortex and medullary cord of the mesenteric lymph node. Many germinal centers were recognized in the lymphatic regions such as the cortex and white pulp of these lymphoid organs. The populations of CD8+ cells and γδ T cells in the hemal node and the spleen were higher than those of the mesenteric lymph node. In addition, the populations of CD21+ cells and MHC class II+ cells in the hemal node and the mesenteric lymph node were higher than those of the spleen. The results suggest that the hemal node has an important role in both cellular and humoral immunity as well as the lymph node and the spleen in cattle.KEY WORDS: comparative study, flow cytometry, hemal node, immunocompetent cells, immunohistochemistry.
doi: 10.1292/jvms.11-0405; J. Vet. Med. Sci. 74(4): 405–411, 2012
The hemal node (HN) is a hematopoietic and lymphoid organ that is found in some mammals such as humans, rats and ruminants. In ruminants, HNs are located in the sub-cutaneous region of the head, the mesenteric region and along the large blood vessels such as the aorta in the tho-rax and abdomen; however, the distribution and number of HNs in these regions seem to be diverse [1, 8, 28, 36]. The histological structure of HNs resembles that of lymph nodes and consists of a lymphoid region, in which the cor-tex and paracortex are recognized, and a sinusoid region, in which the lymphatic cord and sinus are formed [8, 9, 33]. The HN sinus is filled with numerous erythrocytes, lacks lymphatic vessels and only connects small blood vessels [1, 26, 32, 42]. In addition, the HN structure also resembles that of the spleen, which consists of white pulp, in which the germinal center is formed, and red pulp, in which the splenic sinus is filled with numerous erythrocytes [26, 42]. The antibody-producing ability and immunological competence of rat HN after antigen challenge have been described [26, 34]. The possibilities of erythropoiesis and erythrophagocytosis in bovine HNs were described by Cerutti and Guerrero [10]. However, the immunological role of the bovine HN has not yet been determined. In the present paper, we analyzed the immunological function of bovine HNs by comparing the distribution and population of immunocompetent cells in the HNs of calves with those of the mesenteric lymph nodes (MLNs) and spleen.
MATERIALS AND METHODS
Animals and collection of specimens: Japanese black calves (5–8 months old, n=10) were obtained from local farms in Miyazaki Prefecture. The animals were killed by electric shock following the combination of xylazine (0.2 mg/kg) and pentobarbital (15 mg/kg) intravenous injection. All animal procedures were approved by the Institutional Animal Care and Use Committee of the University of Mi-yazaki. The HNs, MLNs and spleens were collected for immunohistochemistry (IHC) and flow cytometry (FACS). The specimens for IHC were mounted in OCT embedding compound (Sakura Finetek Japan Co., Ltd., Tokyo, Japan) on Cryomold (Sakura Finetek Japan Co., Ltd.), frozen on dry ice, and then stored at −80°C until used. The speci-mens for FACS were transported on ice and then treated as described below.
For histological analysis, HNs were fixed with 10% buffered formalin. The samples were passed through etha-nol and toluene and embedded in paraffin. The samples were cut into 4-µm thick sections and stained with hema-toxylin-eosin (HE) stain.
Monoclonal antibodies (mAbs): mAbs specific for calf CD4 (IL-A11), CD8 (CACT80c), CD21 (GB25A), CD172a (DH59B), TCR1-N24: δ chain specific (GB21A) and MHC class II (TH14B) were purchased from VMRD Inc. (Pull-man, WA, U.S.A.). A 1:500 dilution of each mAb was used for IHC and FACS.
IHC: Cryostat sections were stained with mAbs by us-ing the indirect immunoperoxidase technique described by Yasuda et al. [40]. In brief, the sections (7–10 µm thick) were air-dried on slides and fixed with ice-cold acetone for 10 min. To block any nonspecific binding, the sections
*correspondence to: Yasuda, M., Department of Veterinary Anatomy, Faculty of Agriculture, University of Miyazaki, 1–1 Gakuen Kibanadai-Nishi, Miyazaki 889–2192, Japan.
e-mail: [email protected]©2012 The Japanese Society of Veterinary Science

W. ZHANG, T. NASU, Y. Z. HOSAKA AND M. YASUDA406
were rehydrated in PBS and incubated with 10% normal horse serum in PBS for 30 min. The sections were stained with mAbs for 60 min and washed three times with PBS. They were then incubated with a secondary antibody ab-sorbed with acetone powder from calves HN, MLN and spleen. After incubation with the secondary antibody, endogenous peroxidase was quenched with 0.3% H2O2 in methanol for 30 min followed by incubation with ABC complex (Vector Lab., Burlingame, CA, U.S.A.) for 15 min. After the sections were rinsed three times in PBS, the reactions were made visible with metal-enhanced diami-nobenzidine (DAB, Pierce, Rockford, IL, U.S.A.). All im-munohistochemical staining was performed at room tem-perature in a moist chamber. Control staining, in which the primary antibody was replaced with PBS, was performed simultaneously. No positive staining was found in the con-trol slides (data not shown).
FACS: Cell suspensions were obtained from HNs, MLNs and spleens by teasing the minced organs through nylon mesh for removing the debris. The cell suspensions from each lymphoid organ and peripheral blood mononuclear cells (PBMC) were further purified by density gradient centrifugation with Ficoll-Paque (GE Healthcare UK Ltd., Little Chalfont, Buckinghamshire, England). For immuno-fluorescence, the cells were suspended in PBS supplement-ed with 0.5% bovine serum albumin and 0.05% sodium azide. Viable cells were incubated with mAbs at 4°C for 30 min. After washing, a second incubation was conducted with fluorescein isothiocyanate (FITC)-conjugated sheep anti-mouse IgG (H+L) F (ab)2 (Silenus Lab., Boronia, Vic-toria, Australia) at 4°C for 30 min. Relative immunofluo-rescence intensities were determined by flow cytometry with a FACSCantoTMII system (Becton Dickinson, Frank-lin Lakes, NJ, U.S.A.).
Statistical analysis: The statistical analysis consisted of an analysis of variance (ANOVA) and multiple comparison tests performed with Origin Pro 8.0 software (OriginLab Co., Northampton, MA, U.S.A.). P-values less than 0.05 were regarded as statistically significant.
RESULTS
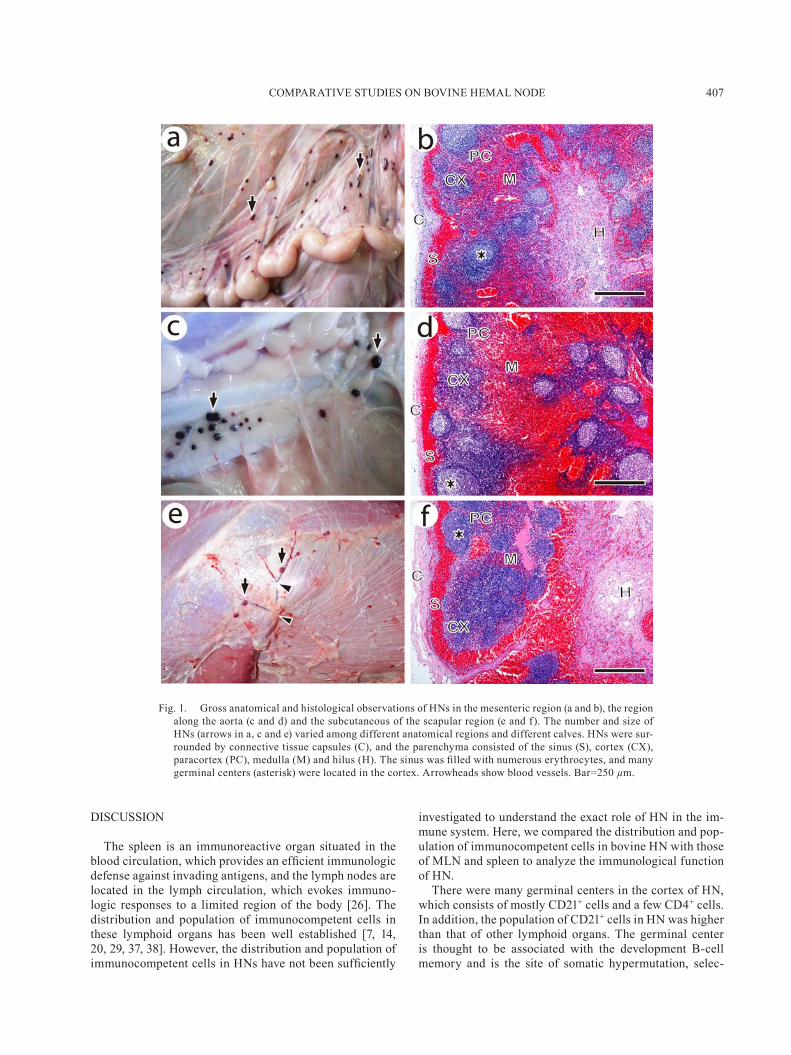
Distribution of HNs and general histological structure: HNs in Japanese black cattle were located mainly in the mesenteric region and along the aorta of the thoracic, ab-dominal and pelvic cavities (Fig. 1a and 1c). In addition, HNs also could be found in the axillary region and the subcutaneous of the scapular region and abdominal region, which located along with the blood vessels (Fig. 1e). The HNs were encapsulated by connective tissues, in which many small blood vessels were found, and the sinus was filled with numerous erythrocytes. There were many varia-tions of lymphoid regions in HNs. The most developed lymphoid regions in HNs consisted of three main regions: the follicular region in which many germinal centers were located; the paracortex which consisted of the lymphatic region and located between cortex and medulla; and the si-nusoid region which consisted of the lymphatic cord and
sinus (Fig. 1b, 1d and 1f).Distribution of immunocompetent cells in HNs, MLNs
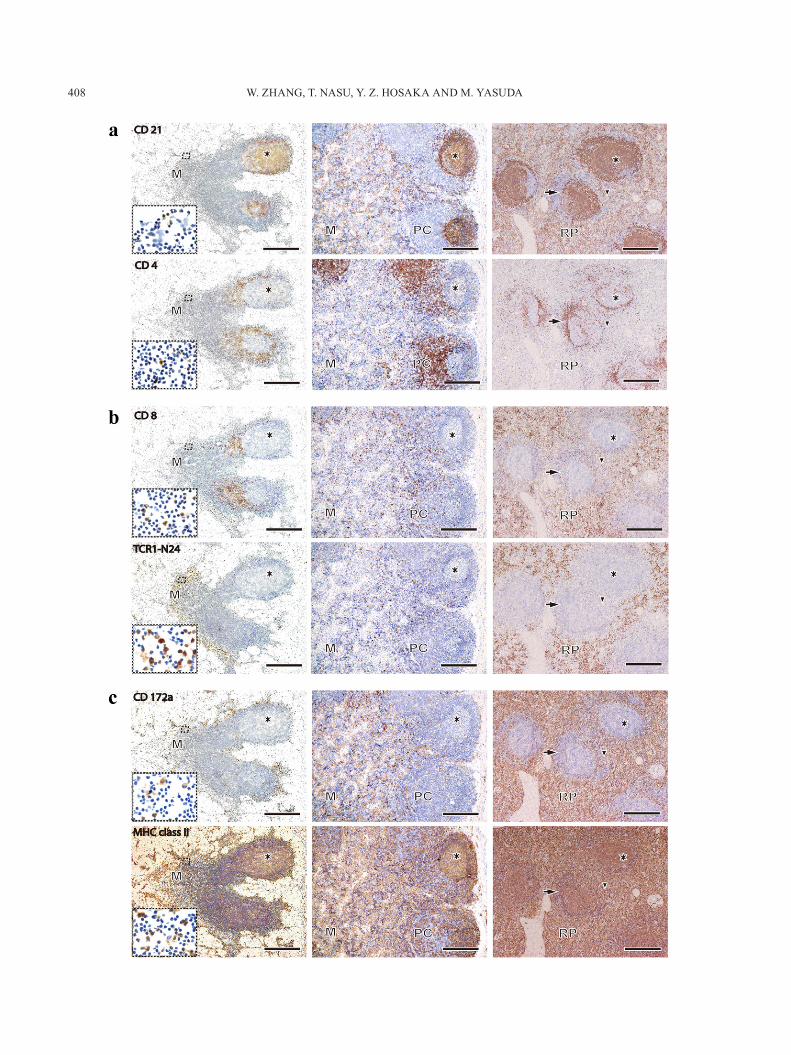
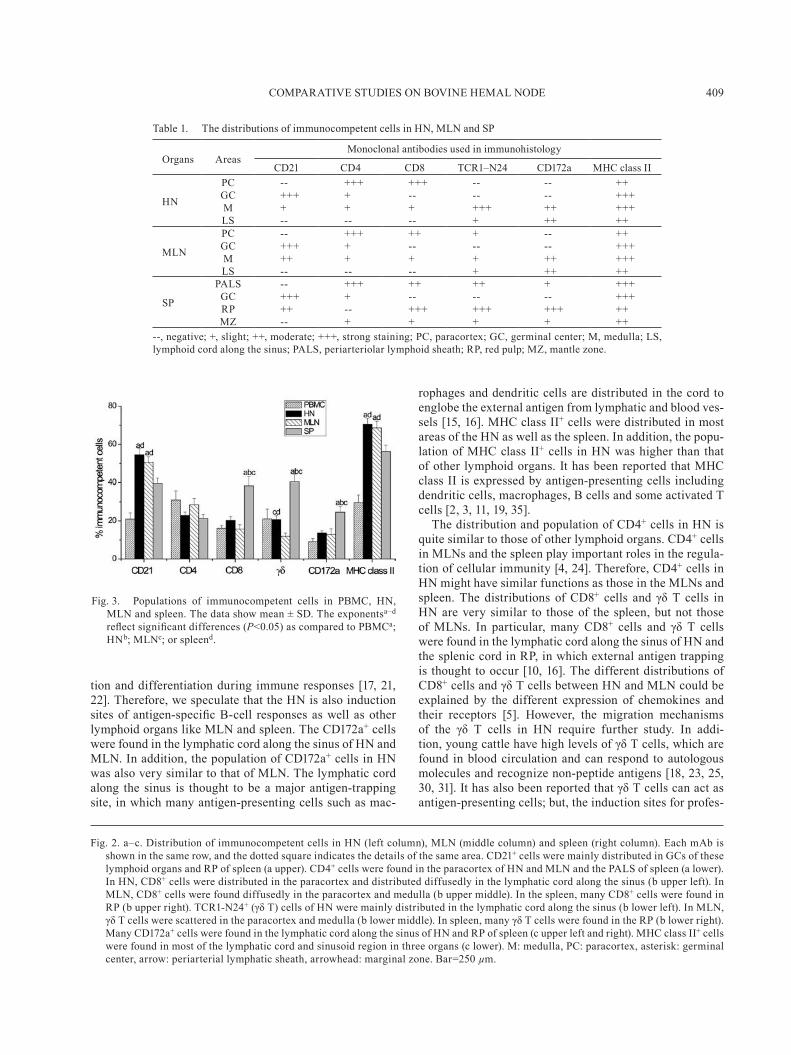
and spleen: The distributions of immunocompetent cells in these lymphoid organs were summarized in Table 1. In HNs, CD21+ cells were distributed in the lymphatic fol-licles, especially in the germinal center (GC), and a few positive cells were also located in the medulla (M) (upper left figure in Fig. 2a). The distribution of CD21+ cells in MLNs was similar to those of HNs (upper middle figure in Fig. 2a). But, many CD21+ cells were also found in the medulla of MLNs. In the spleen, many CD21+ cells were located in the GC and red pulp (RP) (upper right figure in Fig. 2a). CD4+ cells in HN and MLN were mainly present in the paracortex, and a few positive cells were found in the GC and M (lower left and middle figures in Fig. 2a). In the spleen, many CD4+ cells were distributed in the periarteri-olar lymphoid sheath (PALS), and a few positive cells were also present in the GC, the marginal zone (MZ) and the RP (lower right figure in Fig. 2a). In HNs, many CD8+ cells were found in the paracortex, and a few positive cells were found in M (upper left figure in Fig. 2b). In MLN, CD8+ cells were infiltrated diffusely in the paracortex and M (up-per middle figure in Fig. 2b). In the spleen, many CD8+ cells were located in RP, and a few positive cells were found in PALS and MZ (upper right figure in Fig. 2b). Many TCR1-N24+ (γδ T) cells in HNs were distributed in the lymphatic cord along the sinus of M (lower left figure in Fig. 2b). A few γδ T cells were scattered in the paracortex and M of MLN (lower middle figure in Fig. 2b). In the spleen, many γδ T cells were found in RP, and very few positive cells were found in MZ (lower right figure in Fig. 2b). CD172a+ cells, which are expressed on the macrophages and granu-locytes, were distributed in the lymphatic cord along the sinus and M in HN (upper left figure in Fig. 2c). In MLN, the distribution of CD172a+ cells was very similar to those of HN (upper middle figure in Fig. 2c). In the spleen, many positive cells were also distributed in RP, and a few posi-tive cells were recognized in PALS and MZ. MHC class II+ cells were mainly distributed in the lymphatic follicles, the paracortex and the sinusoid regions of HN (lower left figure in Fig. 2c). In MLN, the distribution of positive cells was very similar to those of HN (lower middle figure in Fig. 2c). In spleen, many positive cells were distributed in RP and the white pulp (lower right figure in Fig. 2c).
Population of immunocompetent cells in PBMC, HN, MLN and spleen: As shown in Fig. 3, the populations of CD21+ cells and MHC class II+ cells in HN and MLN were significantly higher than those of the spleen and PBMC (P<0.05); but these vales for HN was similar to those of MLN. In addition, the populations of CD8+ cells, γδ T cells and CD172a+ cells in the spleen were significantly higher than those in HN, MLN and PBMC (P<0.05); but, these values of CD8+ cells and γδ T cells for HN were higher than those for MLN. In particular, the population of γδ T cells in HN was significantly higher than that of MLN (P<0.05). The population of CD4+ cells was not significant different among these lymphoid tissues.
COMPARATIVE STUDIES ON BOVINE HEMAL NODE 407
DISCUSSION
The spleen is an immunoreactive organ situated in the blood circulation, which provides an efficient immunologic defense against invading antigens, and the lymph nodes are located in the lymph circulation, which evokes immuno-logic responses to a limited region of the body [26]. The distribution and population of immunocompetent cells in these lymphoid organs has been well established [7, 14, 20, 29, 37, 38]. However, the distribution and population of immunocompetent cells in HNs have not been sufficiently
investigated to understand the exact role of HN in the im-mune system. Here, we compared the distribution and pop-ulation of immunocompetent cells in bovine HN with those of MLN and spleen to analyze the immunological function of HN.
There were many germinal centers in the cortex of HN, which consists of mostly CD21+ cells and a few CD4+ cells. In addition, the population of CD21+ cells in HN was higher than that of other lymphoid organs. The germinal center is thought to be associated with the development B-cell memory and is the site of somatic hypermutation, selec-
Fig. 1. Gross anatomical and histological observations of HNs in the mesenteric region (a and b), the region along the aorta (c and d) and the subcutaneous of the scapular region (e and f). The number and size of HNs (arrows in a, c and e) varied among different anatomical regions and different calves. HNs were sur-rounded by connective tissue capsules (C), and the parenchyma consisted of the sinus (S), cortex (CX), paracortex (PC), medulla (M) and hilus (H). The sinus was filled with numerous erythrocytes, and many germinal centers (asterisk) were located in the cortex. Arrowheads show blood vessels. Bar=250 µm.
W. ZHANG, T. NASU, Y. Z. HOSAKA AND M. YASUDA408
COMPARATIVE STUDIES ON BOVINE HEMAL NODE 409
tion and differentiation during immune responses [17, 21, 22]. Therefore, we speculate that the HN is also induction sites of antigen-specific B-cell responses as well as other lymphoid organs like MLN and spleen. The CD172a+ cells were found in the lymphatic cord along the sinus of HN and MLN. In addition, the population of CD172a+ cells in HN was also very similar to that of MLN. The lymphatic cord along the sinus is thought to be a major antigen-trapping site, in which many antigen-presenting cells such as mac-
rophages and dendritic cells are distributed in the cord to englobe the external antigen from lymphatic and blood ves-sels [15, 16]. MHC class II+ cells were distributed in most areas of the HN as well as the spleen. In addition, the popu-lation of MHC class II+ cells in HN was higher than that of other lymphoid organs. It has been reported that MHC class II is expressed by antigen-presenting cells including dendritic cells, macrophages, B cells and some activated T cells [2, 3, 11, 19, 35].
The distribution and population of CD4+ cells in HN is quite similar to those of other lymphoid organs. CD4+ cells in MLNs and the spleen play important roles in the regula-tion of cellular immunity [4, 24]. Therefore, CD4+ cells in HN might have similar functions as those in the MLNs and spleen. The distributions of CD8+ cells and γδ T cells in HN are very similar to those of the spleen, but not those of MLNs. In particular, many CD8+ cells and γδ T cells were found in the lymphatic cord along the sinus of HN and the splenic cord in RP, in which external antigen trapping is thought to occur [10, 16]. The different distributions of CD8+ cells and γδ T cells between HN and MLN could be explained by the different expression of chemokines and their receptors [5]. However, the migration mechanisms of the γδ T cells in HN require further study. In addi-tion, young cattle have high levels of γδ T cells, which are found in blood circulation and can respond to autologous molecules and recognize non-peptide antigens [18, 23, 25, 30, 31]. It has also been reported that γδ T cells can act as antigen-presenting cells; but, the induction sites for profes-
Table 1. The distributions of immunocompetent cells in HN, MLN and SP
Organs AreasMonoclonal antibodies used in immunohistology
CD21 CD4 CD8 TCR1–N24 CD172a MHC class II
HN
PC -- +++ +++ -- -- ++GC +++ + -- -- -- +++M + + + +++ ++ +++LS -- -- -- + ++ ++
MLN
PC -- +++ ++ + -- ++GC +++ + -- -- -- +++M ++ + + + ++ +++LS -- -- -- + ++ ++
SP
PALS -- +++ ++ ++ + +++GC +++ + -- -- -- +++RP ++ -- +++ +++ +++ ++MZ -- + + + + ++
--, negative; +, slight; ++, moderate; +++, strong staining; PC, paracortex; GC, germinal center; M, medulla; LS, lymphoid cord along the sinus; PALS, periarteriolar lymphoid sheath; RP, red pulp; MZ, mantle zone.
Fig. 3. Populations of immunocompetent cells in PBMC, HN, MLN and spleen. The data show mean ± SD. The exponentsa–d reflect significant differences (P<0.05) as compared to PBMCa; HNb; MLNc; or spleend.
Fig. 2. a–c. Distribution of immunocompetent cells in HN (left column), MLN (middle column) and spleen (right column). Each mAb is shown in the same row, and the dotted square indicates the details of the same area. CD21+ cells were mainly distributed in GCs of these lymphoid organs and RP of spleen (a upper). CD4+ cells were found in the paracortex of HN and MLN and the PALS of spleen (a lower). In HN, CD8+ cells were distributed in the paracortex and distributed diffusedly in the lymphatic cord along the sinus (b upper left). In MLN, CD8+ cells were found diffusedly in the paracortex and medulla (b upper middle). In the spleen, many CD8+ cells were found in RP (b upper right). TCR1-N24+ (γδ T) cells of HN were mainly distributed in the lymphatic cord along the sinus (b lower left). In MLN, γδ T cells were scattered in the paracortex and medulla (b lower middle). In spleen, many γδ T cells were found in the RP (b lower right). Many CD172a+ cells were found in the lymphatic cord along the sinus of HN and RP of spleen (c upper left and right). MHC class II+ cells were found in most of the lymphatic cord and sinusoid region in three organs (c lower). M: medulla, PC: paracortex, asterisk: germinal center, arrow: periarterial lymphatic sheath, arrowhead: marginal zone. Bar=250 µm.

W. ZHANG, T. NASU, Y. Z. HOSAKA AND M. YASUDA410
sional antigen-presenting γδ T cells have not been fully de-termined in vivo [6, 12, 31, 39]. We speculate that γδ T cells, which are distributed in the lymphatic cord along the sinus of HN, have a potential for antigen presenting cell (manu-script in preparation). In addition, there are two types of nodes form: hemal nodes containing blood-filled sinusoids, which are peculiar to ruminants, and hemolymph nodes containing both blood and lymph sinusoids, which occur in other animal [10, 13, 27]. However, it has been described that the ruminant hemal nodes also have both blood ves-sel and lymphatics [41, 42]. Therefore, the functional and morphological differences between two types of ruminant hemal nodes should be addressed in near future.
Taken together, these results suggest that the HN has the potential to be an antigen-capturing site and the induc-tion site of antigen-specific humoral and cellular immunity. However, further studies are needed to determine if HNs provide local immunity like MLNs, systemic immunity like the spleen, or both local and systemic immunity.
ACKNOWLEDGMENTS. This work was supported by Grants-in-Aid for Scientific Research (Nos. 20580323 and 23405042) from the Ministry of Education, Science, Sport and Culture, Japan.
REFERENCES
1. Bacha, W. J. and Bacha, L. M. 2000. Color Atlas of Veterinary Histology, 2nd ed., Lippincott Williams and Wilkins, London.
2. Baldwin, C. L., Taele, A., Naessens, J., Goddeeris, B., Ma-chugh, N. and Morrison, W. 1986. Characterization of a subset of bovine T lymphocytes that express BoT4 by monoclonal antibodies and function: similarity to lymphocytes defined by human T4 and murine L3T4. J. Immunol. 136: 4385–4391. [Medline]
3. Banchereau, J., Briere, F., Caux, C., Davous, J., Lebecque, S., Liu, Y. J., Pulendran, B. and Palucka, K. 2000. Immuno-biology of dendritic cells. Annu. Rev. Immunol. 18: 767–811. [Medline] [CrossRef]
4. Bennett, S. R., Carbone, F. R., Karamalis, F., Miller, J. F. and Heath, W. R. 1997. Induction of a CD8+ cytotoxic T lympho-cyte response by cross-priming requires cognate CD4+ T cell help. J. Exp. Med. 186: 65–70. [Medline] [CrossRef]
5. Blumerman, S. L., Wang, F., Herzig, C. T. A. and Bladwin, C. L. 2007. Molecular cloning of bovine chemokine receptors and expression by WC1+ γδ T cells. Dev. Comp. Immunol. 31: 87–102. [Medline] [CrossRef]
6. Brandes, M., Williams, K. and Moser, B. 2005. Professional antigen-presentation function by human gammadelta T cells. Science 309: 264–268. [Medline] [CrossRef]
7. Caro, M. R., Gallego, M. C., Buendia, A. J. and Navarro, E. 1998. Postnatal evolution of lymphocyte subpopulations in pe-ripheral blood and lymphoid organs in the goat. Res. Vet. Sci. 65: 145–148. [Medline] [CrossRef]
8. Casteleyn, C. R., Breugelmans, S., Simoens, P. and Van den Broeck, W. 2008. Morphological and immunological char-acteristics of the bovine temporal lymph node and hemal node. Vet. Immunol. Immunopathol. 126: 339–350. [Medline] [CrossRef]
9. Cerutti, P. 1998. A scanning and immunohistochemical study in bovine haemal node. Anat. Histol. Embryol. 27: 387–392.
[Medline] [CrossRef] 10. Cerutti, P. and Guerrero, F. 2008. Erythropoiesis and eryth-
rophagocytosis in bovine haemal nodes. Int. J. Morphol. 26: 557–562. [CrossRef]
11. Cheng, L., Cui, Y., Shao, H., Han, G. C., Zhu, L., Huang, Y. F., O’Brien, R. L., Born, W. K., Kaplan, H. J. and Sun, D. 2008. Mouse γδ T cells are capable of expressing MHC class II mol-ecules, and of functioning as antigen-presenting cells. J. Neu-roimmunol. 203: 3–11. [Medline] [CrossRef]
12. Collins, R. A., Werling, D., Duggan, S. E., Bland, A. P., Par-sons, K. R. and Howard, C. J. 1998. γδ T cells present antigen to CD4+ αβ T cells. J. Leukoc. Biol. 63: 707–714. [Medline]
13. Constantinescu, G. M., Brown, E. M. and McClure, R. C. 1988. Accessory parotid lymph nodes and hemal nodes in the temporal fossa in three oxen. Cornell Vet. 78: 147–154. [Med-line]
14. Faldyna, M., Sinkora, J., Knotigova, P., Leva, L. and Toman, M. 2005. Lymphatic organ development in dogs: major lym-phocyte subsets and activity. Vet. Immunol. Immunopathol. 104: 239–247. [Medline] [CrossRef]
15. Galeotti, M., Sarli, G., Eleni, C. and Marcato, P. S. 1993. Iden-tification of cell types present in bovine haemolymph nodes and lymph nodes by immunostaining. Vet. Immunol. Immuno-pathol. 36: 319–331. [Medline] [CrossRef]
16. Goldsby, R. A., Kindt, T. J., Osborne, B. A. and Kuby, J. 2002. Immunology, 5th ed., W. H. Freeman, New York.
17. Grandien, A., Modigliani, Y., Freitas, A., Anderson, J. and Coutinho, A. 1994. Positive and negative selection of antibody repertoires during B-cell differentiation. Immunol. Rev. 137: 53–89. [Medline] [CrossRef]
18. Hein, W. R. and Mackay, C. R. 1991. Prominence of γδ T cells in the ruminant immune system. Immunol. Today 12: 30–34. [Medline] [CrossRef]
19. Janeway, C. A. Jr., Travers, P., Walport, M. and Shlomchik, M. J. 2001. Immunobiology, 5th ed., Garland Science, New York.
20. Kampen, A. H., Olsen, I., Tollersrud, T., Storset, A. K. and Lund, A. 2006. Lymphocyte subpopulations and neutrophil function in calves during the first 6 months of life. Vet. Im-munol. Immunopathol. 113: 53–63. [Medline] [CrossRef]
21. Leanderson, T., Kalleberg, E. and Gray, D. 1992. Expansion, selection and mutation of antigen-specific B cells in germinal centers. Immunol. Rev. 126: 47–61. [Medline] [CrossRef]
22. Liu, Y. J., Joshua, D. E., Williams, G. T., Amith, C. A., Gor-don, J. and MacLennan, I. C. M. 1989. Mechanism of antigen-derived selection in germinal centers. Nature 342: 929–931. [Medline] [CrossRef]
23. Martino, A., Casetti, R., D’Alessandri, A., Sacchi, A. and Poccia, F. 2005. Complementary function of gamma delta T-lymphocytes and dendritic cells in the response to isopente-nyl-pyrophosphate and lipopolysaccharide antigens. J. Clin. Immunol. 25: 230–237. [Medline] [CrossRef]
24. Ossendorp, F., Toes, R. E. M., Offringa, R., Burg, S. H. and Melief, C. J. M. 2000. Importance of CD4+ T helper cell re-sponses in tumor immunity. Immunol. Lett. 74: 75–79. [Med-line] [CrossRef]
25. Pollock, J. M. and Welsh, M. D. 2002. The WC1+ γδ T-cell population in cattle: a possible role in resistance to intracel-lular infection. Vet. Immunol. Immunopathol. 89: 105–114. [Medline] [CrossRef]
26. Sakita, K., Fujino, M., Koshikawa, T., Ohmiya, N., Ohbayashi, M. and Asai, J. 1997. Structure and function of the hemolymph node in rats. Nagoya J. Med. Sci. 60: 129–137. [Medline]
27. Singh, A. 1959. On the microscopic structure of the haemal

COMPARATIVE STUDIES ON BOVINE HEMAL NODE 411
nodes of buffalo calves. Br. Vet. J. 115: 271–275. 28. Snider, T. G. III., Coats, K. S., Storts, R. W., Graves, K. F.,
Cooper, C. R., Hoyt, P. G., Luther, D. G. and Jenny, B. F. 2003. Natural bovine lentivirus type 1 infection in Holstein dairy cattle. II. Lymphoid tissue lesions. Comp. Immunol. Micro-biol. Infect. Dis. 26: 1–15. [Medline] [CrossRef]
29. Sopp, P. and Howard, C. J. 2001. IFN-γ and IL-4 production by CD4, CD8 and WC1 γδ T cells from cattle lymph nodes and blood. Vet. Immunol. Immunopathol. 81: 85–96. [Medline] [CrossRef]
30. Tanaka, Y., Morita, C. T., Tanaka, Y., Nieves, E., Brenner, M. B. and Bloom, B. R. 1995. Natural and synthetic non-peptide antigens recognized by human γδ T cells. Nature 375: 155–158. [Medline] [CrossRef]
31. Toka, F. N., Kenney, M. A. and Golde, W. T. 2011. Rapid and transient activation of γδ T cells to IFN-γ production, NK cell-like killing, and antigen processing during acute virus infec-tion. J. Immunol. 186: 4853–4861. [Medline] [CrossRef]
32. Turner, D. R. 1969. The vascular tree of the haemal node in the rat. J. Anat. 104: 481–493. [Medline]
33. Turner, D. R. 1971. The reticulo-endothelial components of the haemal node: a light and electron microscopic study. J. Anat. 108: 13–22. [Medline]
34. Turner, D. R. 1971. Immunological competence of the haemal node. J. Anat. 110: 17–24. [Medline]
35. Van Kampen, C. and Mallard, B. A. 1997. Effects of peri-partum stress and health on circulating bovine lymphocyte subsets. Vet. Immunol. Immunopathol. 59: 79–91. [Medline]
[CrossRef] 36. Vincent, S. and Harrison, S. 1897. On the haemolymph glands
of some vertebrates. J. Anat. Physiol. 31: 176–198. [Medline] 37. Wilson, R. A., Zolnai, A., Rudas, P. and Frenyo, L. V. 1996.
T-cell subsets in blood and lymphoid tissues obtained from fetal calves, maturing calves, and adult bovine. Vet. Immunol. Immunopathol. 53: 49–60. [Medline] [CrossRef]
38. Wu, D., Takahashi, K., Liu, N., Koguchi, A., Makara, M., Sasaki, J., Goryo, M. and Okada, K. 1999. Distribution of T-lymphocyte subpopulation in blood and spleen of normal cat-tle and cattle with enzootic bovine leukosis. J. Comp. Pathol. 120: 117–127. [Medline] [CrossRef]
39. Wu, Y., Wu, W., Wong, W. M., Ward, E., Thrasher, A. J., Gold-blatt, D., Osman, M., Digard, P., Canaday, D. H. and Gustafs-son, K. 2009. Human γδ T cells: a lymphoid lineage cell capa-ble of professional phagocytosis. J. Immunol. 183: 5622–5629. [Medline] [CrossRef]
40. Yasuda, M., Tanaka, S., Arakawa, H., Taura, Y., Yokomizo, Y. and Ekino, S. 2002. A comparative study of gut-associated tissue in calf and chicken. Anat. Rec. 266: 207–217. [Medline] [CrossRef]
41. Zidan, M. and Pabst, R. 2004. Histological, histochemical and immunohistochemical study of the haemal nodes of the drom-edary camel. Anat. Histol. Embryol. 33: 284–289. [Medline] [CrossRef]
42. Zidan, M. and Pabst, R. 2010. Histology of hemal nodes of the water buffalo (Bos bubalus). Cell Tissue Res. 340: 491–496. [Medline] [CrossRef]


















