
Chapitre II. Les mutations et les systèmes enzymatiques … : les "mutations adaptatives" de J....
24
Chapitre II. Les mutations et les systèmes enzymatiques de réparation et de recombinaison de l’ADN 1
Transcript of Chapitre II. Les mutations et les systèmes enzymatiques … : les "mutations adaptatives" de J....

Chapitre II. Les mutations et les systèmes enzymatiques de réparation et de recombinaison de
l’ADN
1

2
1. Les mutations2. Les systèmes de réparation de l’ADN3. La recombinaison homologue
Sommaire
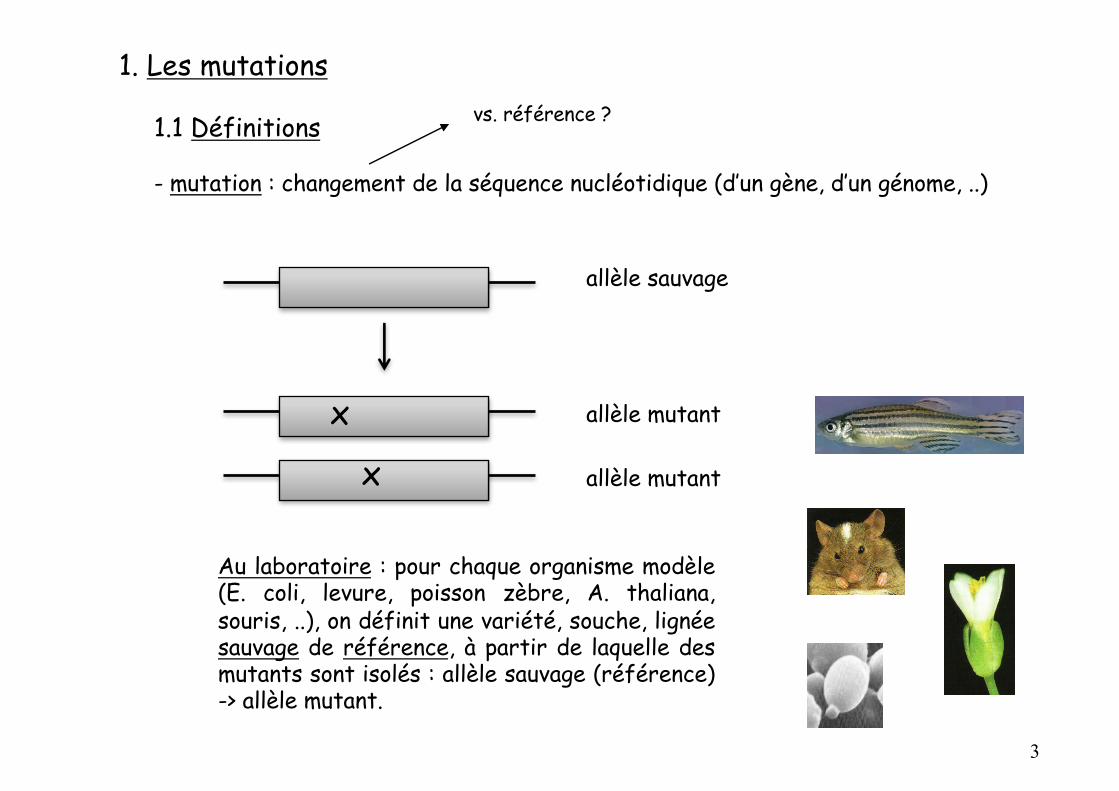
1. Les mutations
1.1 Définitions
- mutation : changement de la séquence nucléotidique (d’un gène, d’un génome, ..)
Au laboratoire : pour chaque organisme modèle(E. coli, levure, poisson zèbre, A. thaliana,souris, ..), on définit une variété, souche, lignéesauvage de référence, à partir de laquelle desmutants sont isolés : allèle sauvage (référence)-> allèle mutant.
3
vs. référence ?
x
x
allèle sauvage
allèle mutant
allèle mutant

Dans la nature : chaque gène d'une espèce est typiquement représenté par plusieurs allèles (chaque allèle est présent à une certaine fréquence au sein de la population), c'est le polymorphisme génétique naturel. Un allèle particulier est parfois associé à une pathologie.
Référence ? L’allèle le plus fréquent d'un gène pourrait en principe êtreconsidéré comme l’allèle de référence, mais dans la pratique, l’allèle deréférence est en général celui qui a été séquencé en premier, ou séquencélors d’un programme de séquençage du génome complet. Ex. génomehumain, version GRC38h = référence
- génotype : définition des allèles - ex. individu diploïde A B cA b c
- phénotype : propriétés ou caractères visibles
déterminé par le génotype (génétique) mais aussi par le niveau d’expression des gènes (épigénétique)
4
Projet 1000 genomes : recensement de ≈ 88 millions de variationsgénétiques. Typiquement, le génome d’un individu présente entre 4 et 5millions de différences par rapport à la séquence de référence (GRC38h).
® cad : sans rapport avec l’éventuel effet qu’aura la mutation sur l’organisme
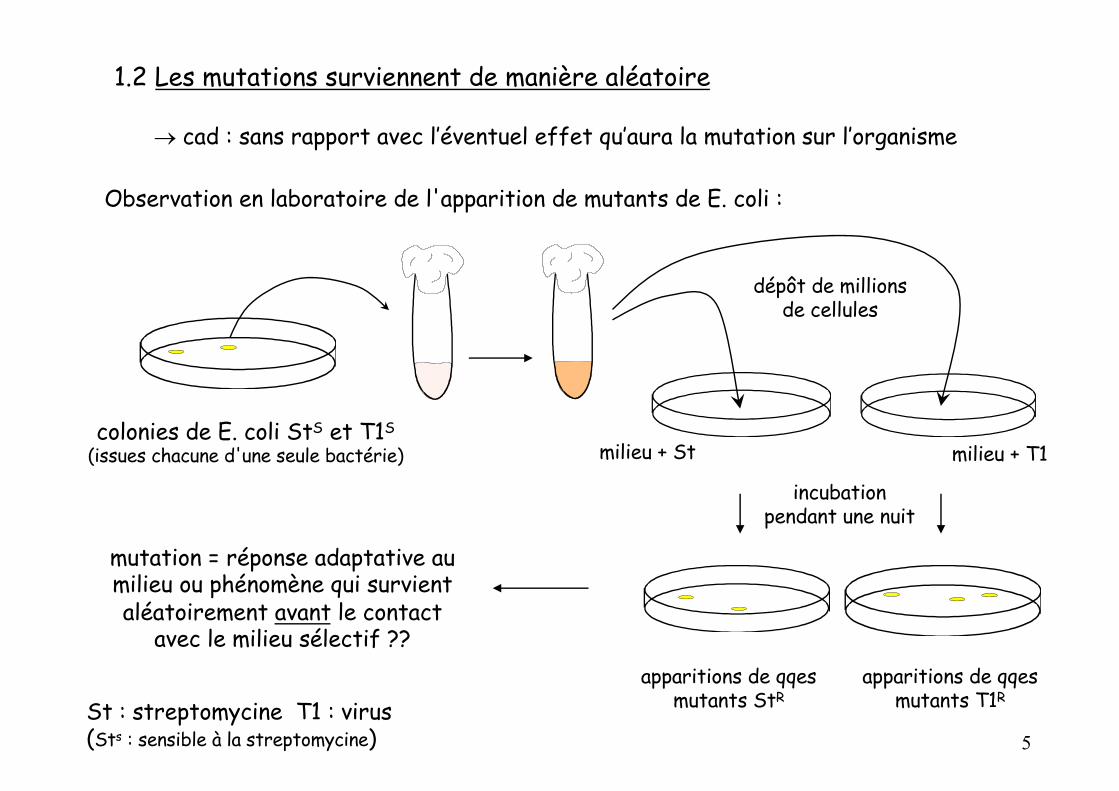
1.2 Les mutations surviennent de manière aléatoire
milieu + St
apparitions de qqesmutants StR
apparitions de qqesmutants T1R
dépôt de millions de cellules
milieu + T1
mutation = réponse adaptative au milieu ou phénomène qui survient aléatoirement avant le contact
avec le milieu sélectif ??
St : streptomycine T1 : virus(Sts : sensible à la streptomycine)
incubation pendant une nuit
colonies de E. coli StS et T1S
(issues chacune d'une seule bactérie)
Observation en laboratoire de l'apparition de mutants de E. coli :
5
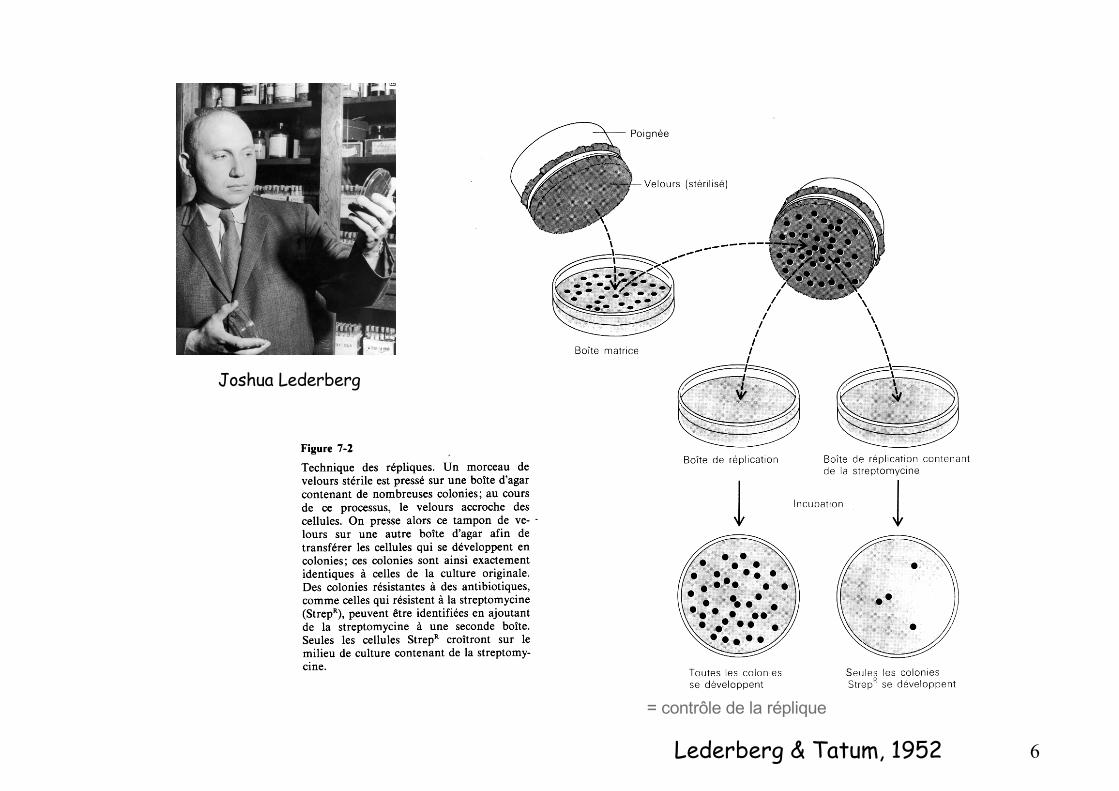
Lederberg & Tatum, 1952
Joshua Lederberg
6
= contrôle de la réplique
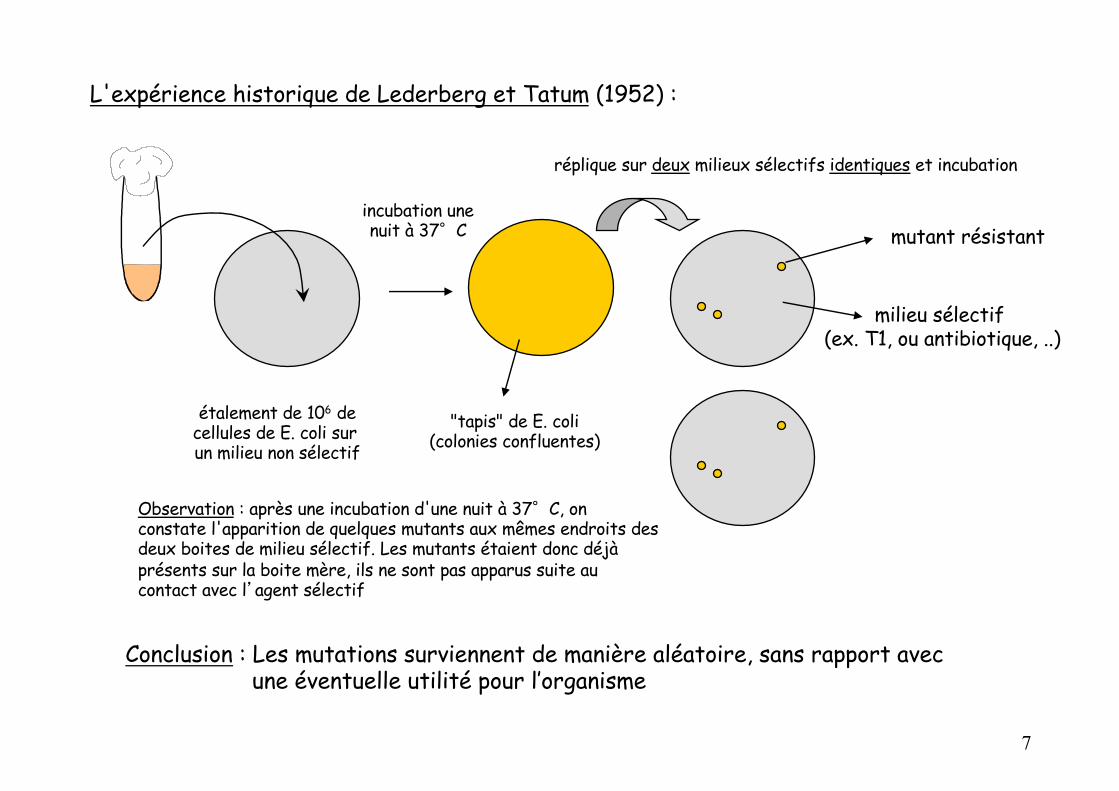
Conclusion : Les mutations surviennent de manière aléatoire, sans rapport avec une éventuelle utilité pour l’organisme
milieu sélectif (ex. T1, ou antibiotique, ..)
réplique sur deux milieux sélectifs identiques et incubation
étalement de 106 decellules de E. coli sur un milieu non sélectif
"tapis" de E. coli(colonies confluentes)
incubation unenuit à 37°C
L'expérience historique de Lederberg et Tatum (1952) :
mutant résistant
Observation : après une incubation d'une nuit à 37°C, on constate l'apparition de quelques mutants aux mêmes endroits des deux boites de milieu sélectif. Les mutants étaient donc déjà présents sur la boite mère, ils ne sont pas apparus suite au contact avec l’agent sélectif
7

rem : dans sa théorie, C Darwin avait déjà proposé que les variations héréditaires surviennent au sein des espèces de manière aléatoire … c'est l'un des principaux fondements de sa théorie et de tous les développements scientifiques qui s'en sont suivis
Les divers mouvements créationnistes, au contraire, rejettent ce principe :
Bruce ChapmanDiscovery Institute, USA
Théorie de l' "Intelligent design" :
Les mutations surviennent de manière "orientée", "intentionnelle" pour générer à terme une fonction, un mécanisme, un organe, ... selon un plan prédéfini ("design")
Harun YahyaScientific ResearchFoundation, Turquie
"Toutes les espèces (et leurs génomes) ont été créées, l'évolution des espèces est un mythe"
"Le seul effet des mutations est d'altérer, de détruire les fonctions biologiques … "
http://www.discovery.org/http://www.harunyahya.fr/
8
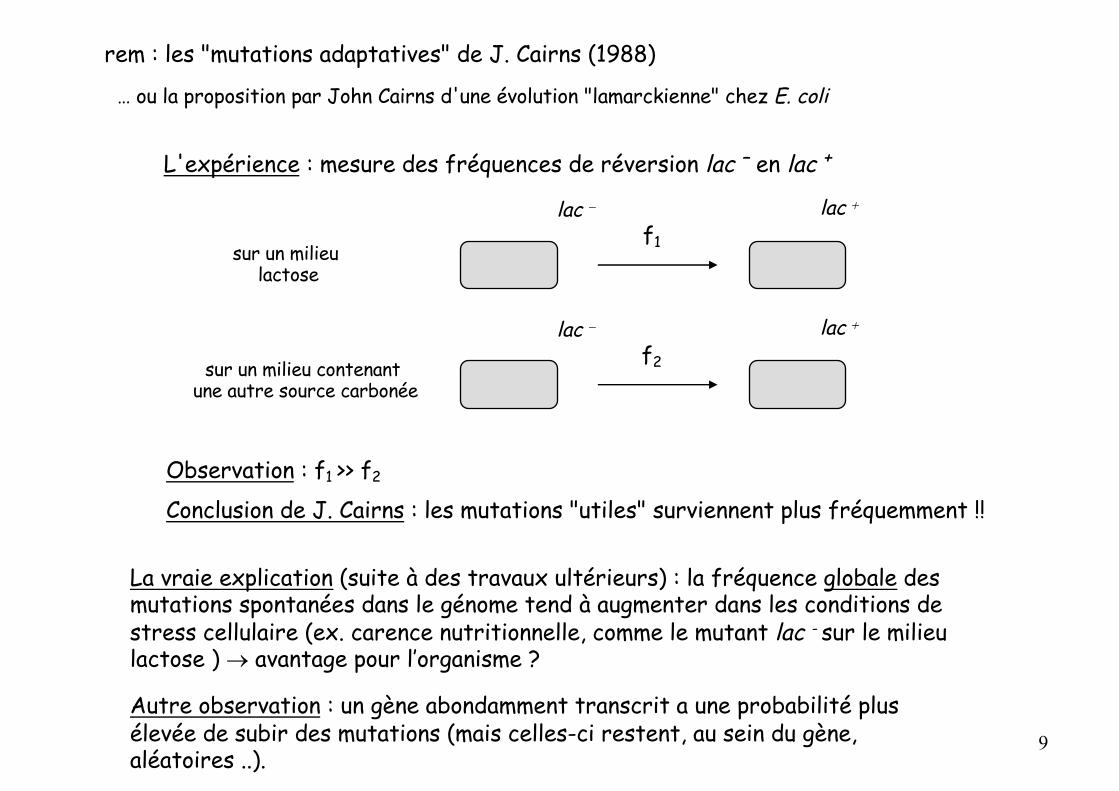
rem : les "mutations adaptatives" de J. Cairns (1988)
… ou la proposition par John Cairns d'une évolution "lamarckienne" chez E. coli
La vraie explication (suite à des travaux ultérieurs) : la fréquence globale des mutations spontanées dans le génome tend à augmenter dans les conditions de stress cellulaire (ex. carence nutritionnelle, comme le mutant lac - sur le milieu lactose ) ® avantage pour l’organisme ?
Autre observation : un gène abondamment transcrit a une probabilité plus élevée de subir des mutations (mais celles-ci restent, au sein du gène, aléatoires ..).
lac - lac +
sur un milieu lactose
f1
lac - lac +
sur un milieu contenant une autre source carbonée
f2
L'expérience : mesure des fréquences de réversion lac – en lac +
Observation : f1 >> f2
Conclusion de J. Cairns : les mutations "utiles" surviennent plus fréquemment !!
9
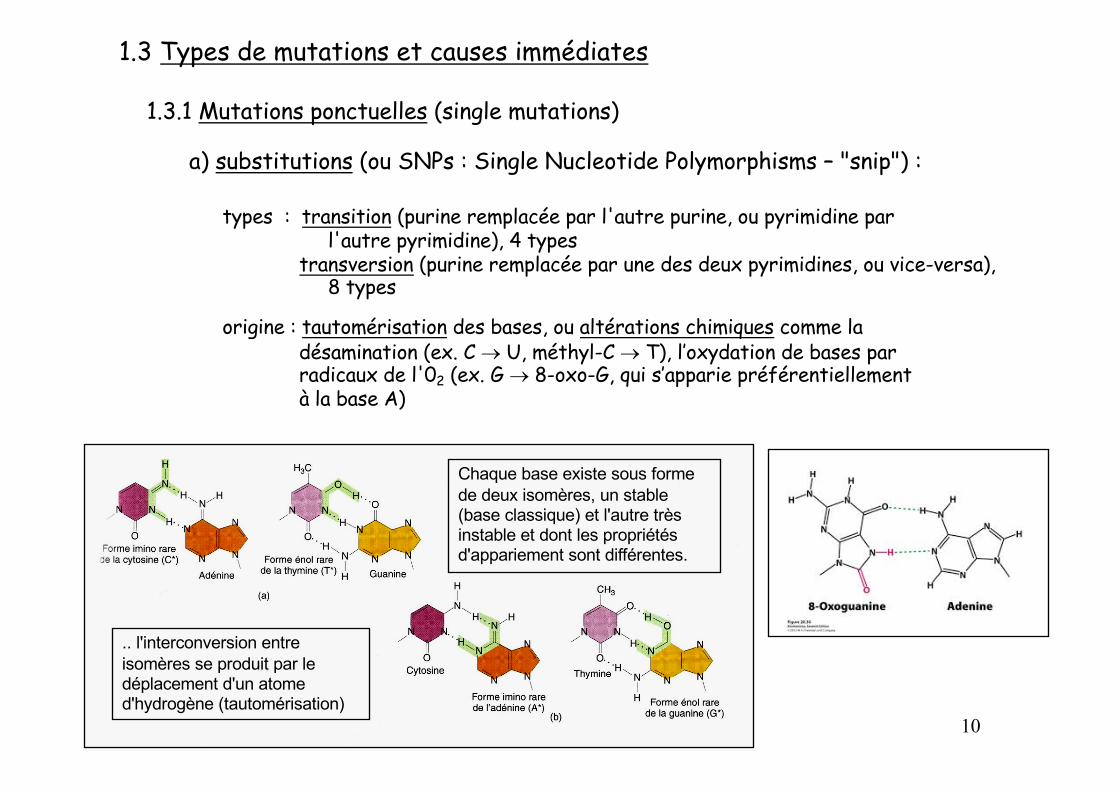
1.3 Types de mutations et causes immédiates
1.3.1 Mutations ponctuelles (single mutations)
a) substitutions (ou SNPs : Single Nucleotide Polymorphisms – "snip") :
types : transition (purine remplacée par l'autre purine, ou pyrimidine par l'autre pyrimidine), 4 types
transversion (purine remplacée par une des deux pyrimidines, ou vice-versa), 8 types
origine : tautomérisation des bases, ou altérations chimiques comme la désamination (ex. C ® U, méthyl-C ® T), l’oxydation de bases parradicaux de l'02 (ex. G ® 8-oxo-G, qui s’apparie préférentiellementà la base A)
10
Chaque base existe sous forme de deux isomères, un stable (base classique) et l'autre très instable et dont les propriétés d'appariement sont différentes.
.. l'interconversion entre isomères se produit par le déplacement d'un atome d'hydrogène (tautomérisation)
C
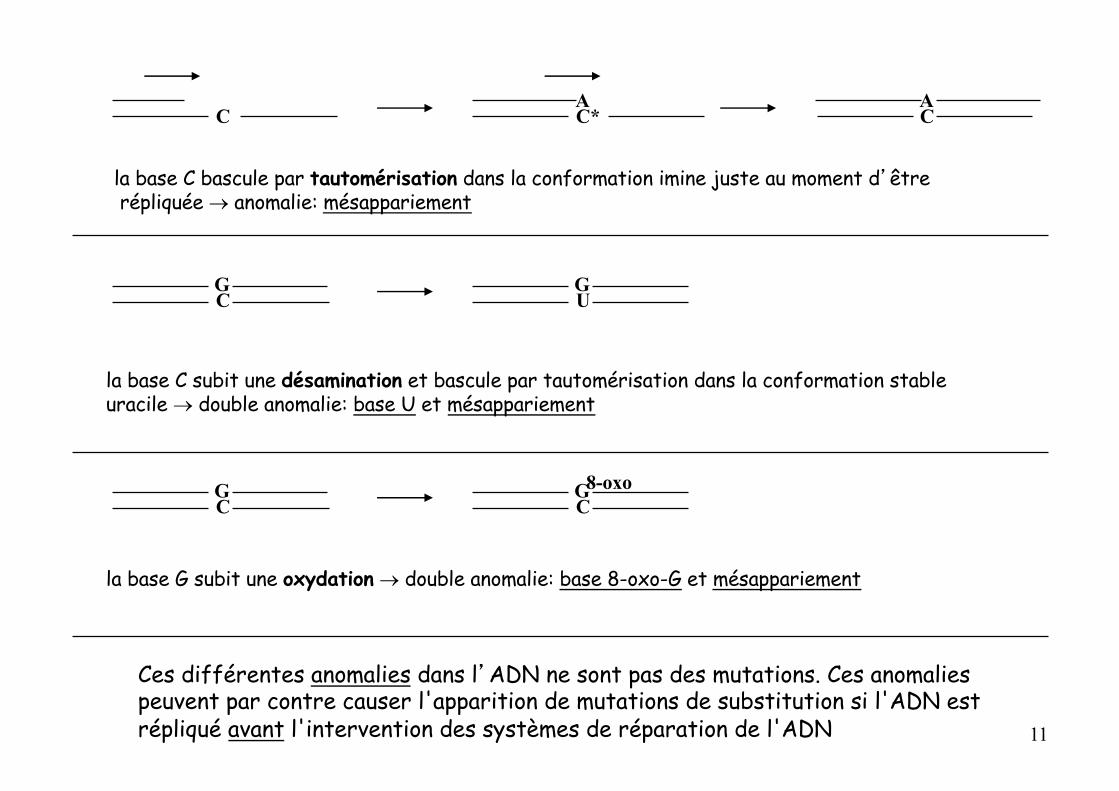
la base C bascule par tautomérisation dans la conformation imine juste au moment d’être répliquée ® anomalie: mésappariement
UG
C*A
CA
CG8-oxo
CG
la base C subit une désamination et bascule par tautomérisation dans la conformation stable uracile ® double anomalie: base U et mésappariement
CG
la base G subit une oxydation ® double anomalie: base 8-oxo-G et mésappariement
Ces différentes anomalies dans l’ADN ne sont pas des mutations. Ces anomalies peuvent par contre causer l'apparition de mutations de substitution si l'ADN est répliqué avant l'intervention des systèmes de réparation de l'ADN 11
CA
TA
CG
mutationanomalie
UG
CG
UA
anomalie
anomalie
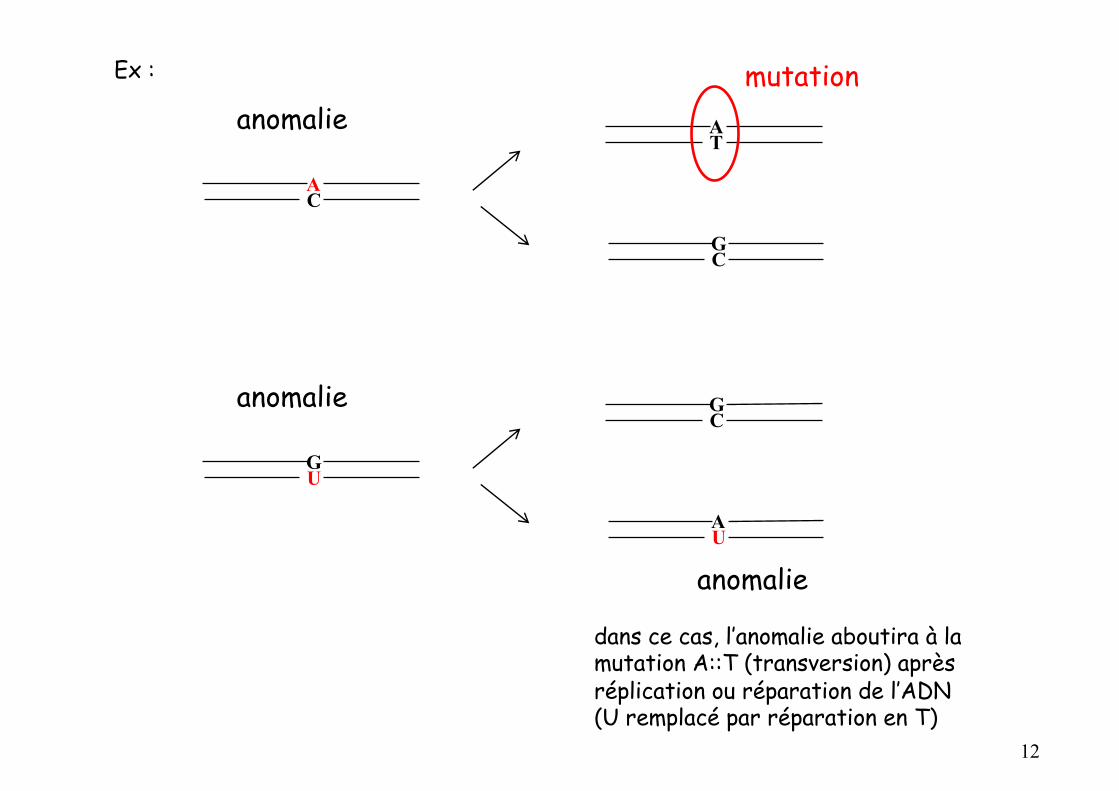
dans ce cas, l’anomalie aboutira à la mutation A::T (transversion) après réplication ou réparation de l’ADN(U remplacé par réparation en T)
12
Ex :
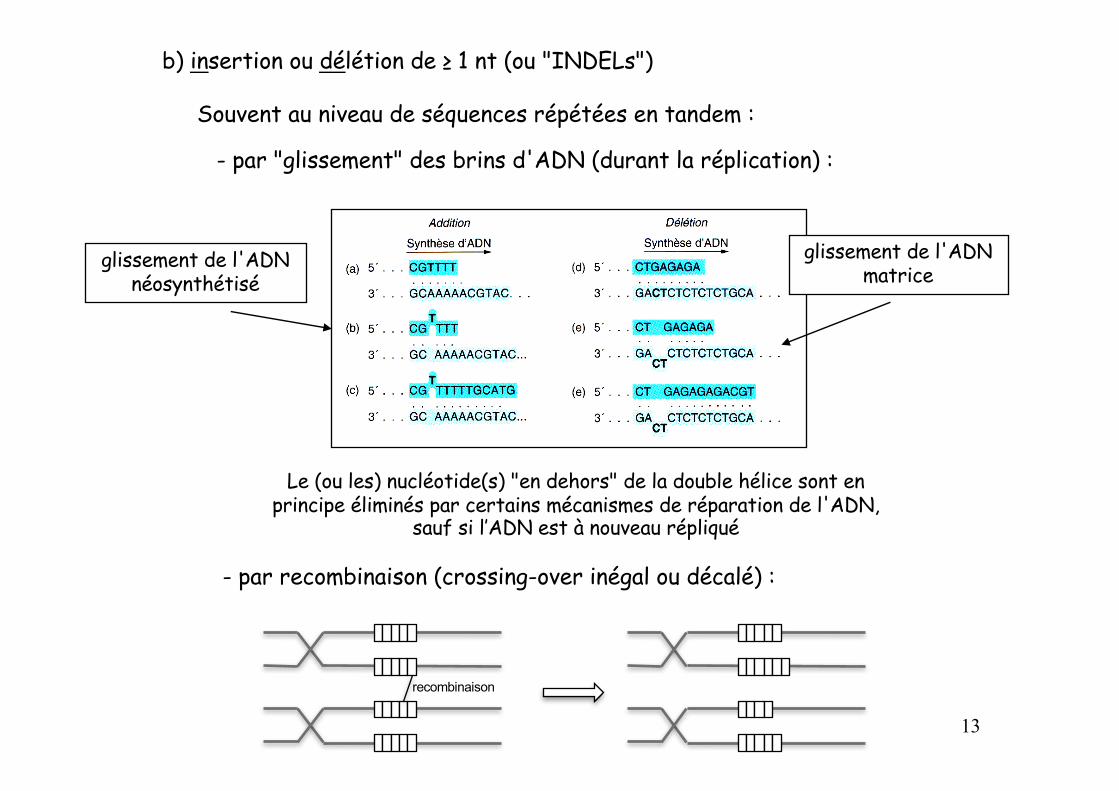
b) insertion ou délétion de ≥ 1 nt (ou "INDELs")
- par "glissement" des brins d'ADN (durant la réplication) :
glissement de l'ADN néosynthétisé
glissement de l'ADN matrice
- par recombinaison (crossing-over inégal ou décalé) :
Souvent au niveau de séquences répétées en tandem :
Le (ou les) nucléotide(s) "en dehors" de la double hélice sont en principe éliminés par certains mécanismes de réparation de l'ADN,
sauf si l’ADN est à nouveau répliqué
13
recombinaison
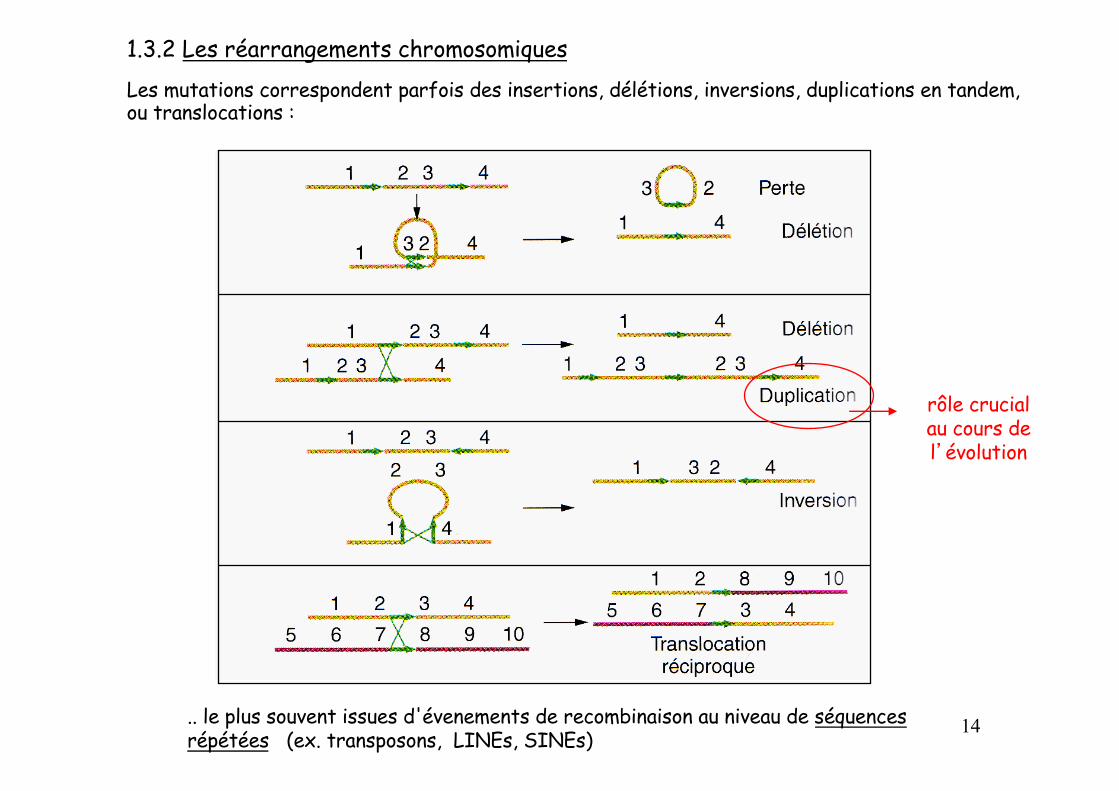
1.3.2 Les réarrangements chromosomiquesLes mutations correspondent parfois des insertions, délétions, inversions, duplications en tandem, ou translocations :
.. le plus souvent issues d'évenements de recombinaison au niveau de séquences répétées (ex. transposons, LINEs, SINEs)
rôle crucial au cours de l’évolution
14

1.4 Conséquences des mutations
- m. silencieuse : aa inchangé (ex. codon AGG > Arg ® codon CGG > Arg)- m. faux-sens ("missense") : aa remplacé par un autre (ex. AGG > AGC > Ser)- m. non-sens : aa remplacé par "stop" (si codon remplacé par UAG = mutation amber, par UAA = m. ochre, par UGA = m. umber ou opal)
- Mutations dans l'ORF d'un gène :
Substitutions ponctuelles (SNPs) :
Délétions/insertions d'un ou qqes nt (INDELs) :
- provoque le plus souvent un décalage du cadre de lecture (frameshift) :
NH2
NH2
COOH
COOH
- Les mutations peuvent aussi altérer les séquences régulatrices, le promoteur, les sites d'épissage d'un gène, …
NH2
NH2
COOH
COOH protéine tronquée du côté C-terminal
15
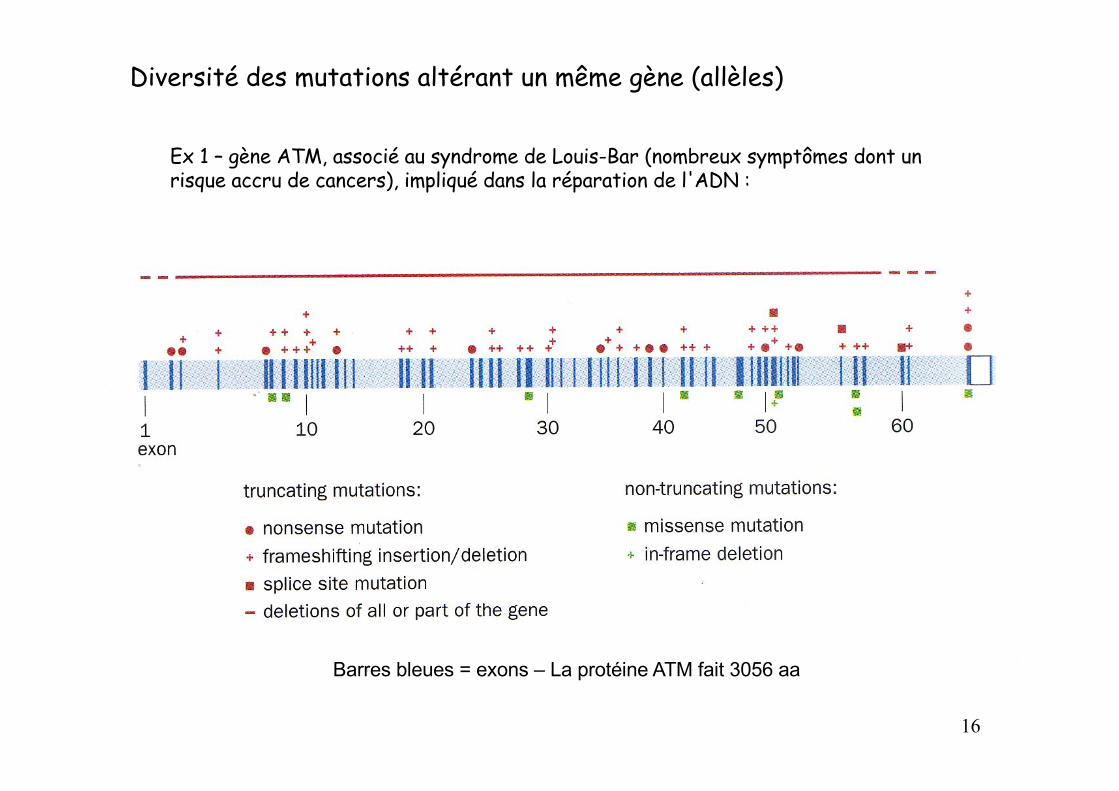
Diversité des mutations altérant un même gène (allèles)
Ex 1 – gène ATM, associé au syndrome de Louis-Bar (nombreux symptômes dont un risque accru de cancers), impliqué dans la réparation de l'ADN :
Barres bleues = exons – La protéine ATM fait 3056 aa
16
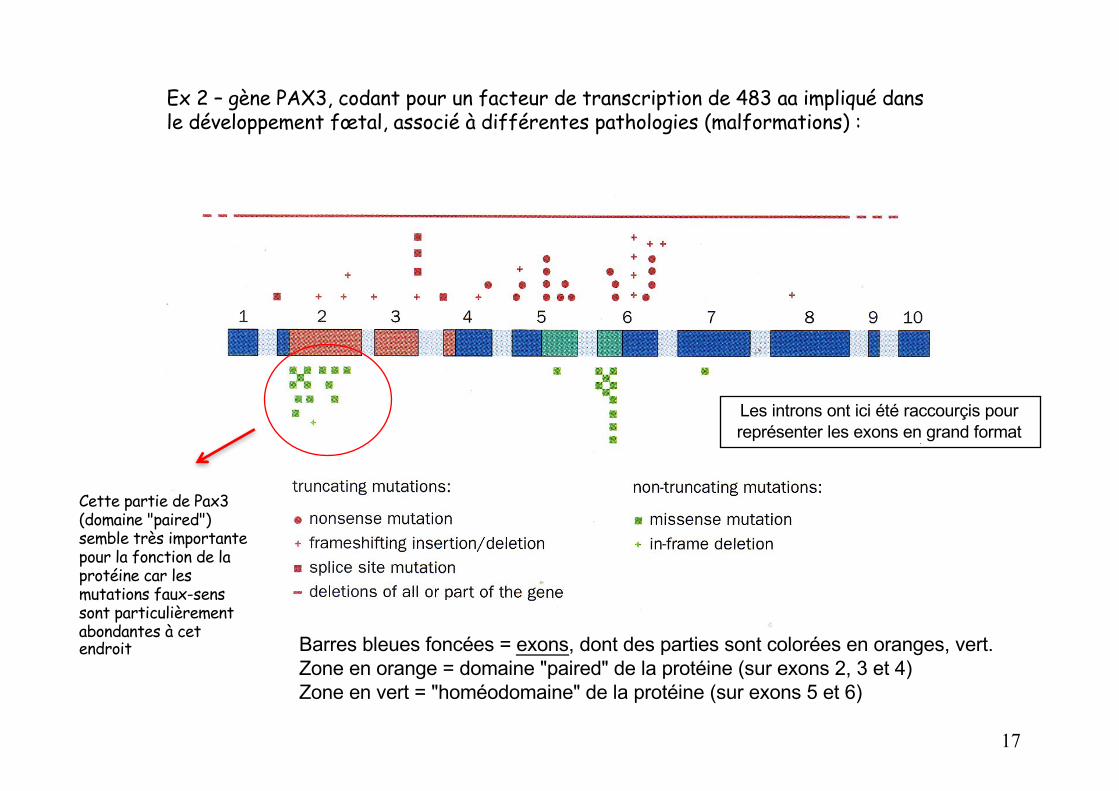
Ex 2 – gène PAX3, codant pour un facteur de transcription de 483 aa impliqué dans le développement fœtal, associé à différentes pathologies (malformations) :
Barres bleues foncées = exons, dont des parties sont colorées en oranges, vert.Zone en orange = domaine "paired" de la protéine (sur exons 2, 3 et 4)Zone en vert = "homéodomaine" de la protéine (sur exons 5 et 6)
Cette partie de Pax3 (domaine "paired") semble très importante pour la fonction de la protéine car les mutations faux-sens sont particulièrement abondantes à cet endroit
17
Les introns ont ici été raccourçis pour représenter les exons en grand format

18
rem : diversité des mutations recensées dans le génome humain (projet 1000 K) :
- recensement d’environ 88,4 millions de mutations / variations génétiques
~ 95% = SNPs~ 4% = INDELs~ 0,05% = réarrangements chromosomiques
- la plupart de ces variations sont rares :
64 millions f < 0,5%12 millions 0,5 < f < 5%~ 8 millions f > 5%
- un génome séquencé contient typiquement :
4-5 millions de variations (vs. séquence de référence)la plupart sont communes, seulement 40.000 à 200.000 présent une f < 0,5%> 99 % des variations = SNPs et INDELs Le reste : ~ 1000 délétions, ~ 160 variations du nombre de copies, ~900 insertions Alu,~130 insertions L1, ~10 inversions, ...
Parmi ces variations, environ 500.000 altèrent la séquence d’acides aminés de protéines, dont 150 à 200 qui sont de type ” protein truncating ”
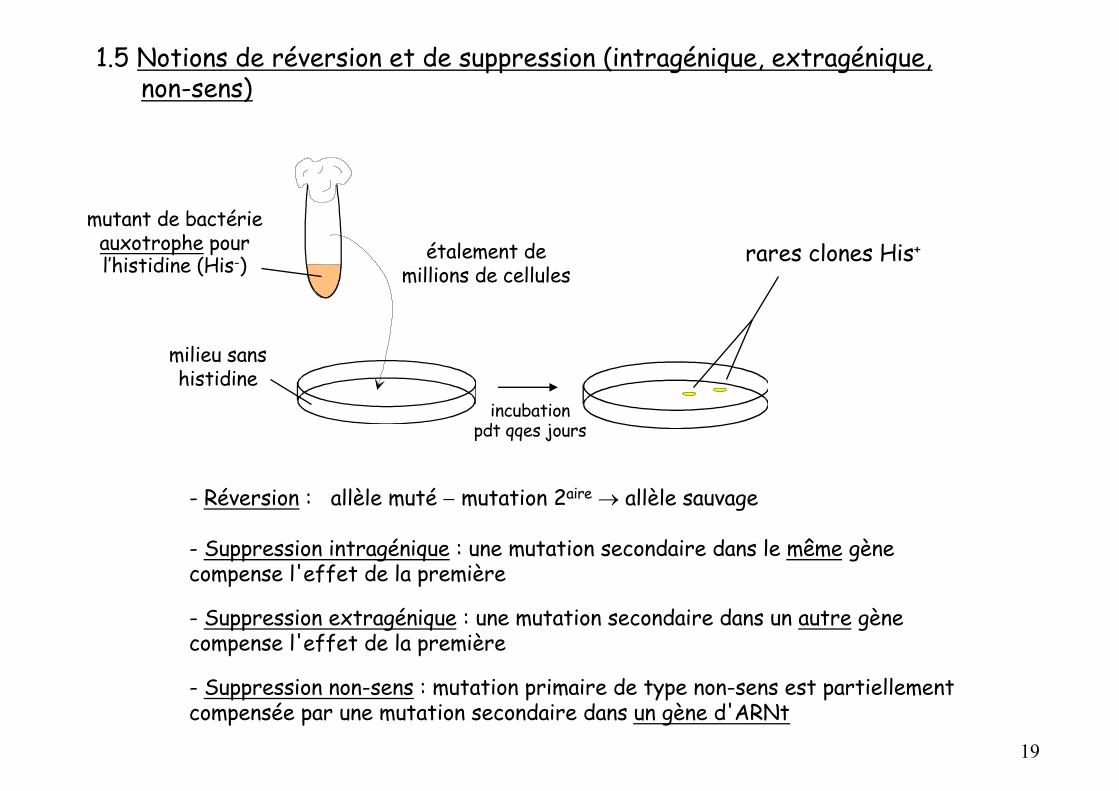
1.5 Notions de réversion et de suppression (intragénique, extragénique,non-sens)
rares clones His+étalement de millions de cellules
mutant de bactérie auxotrophe pour l’histidine (His-)
milieu sans histidine
- Réversion : allèle muté - mutation 2aire ® allèle sauvage
- Suppression intragénique : une mutation secondaire dans le même gène compense l'effet de la première
- Suppression extragénique : une mutation secondaire dans un autre gène compense l'effet de la première
- Suppression non-sens : mutation primaire de type non-sens est partiellement compensée par une mutation secondaire dans un gène d'ARNt
incubationpdt qqes jours
19
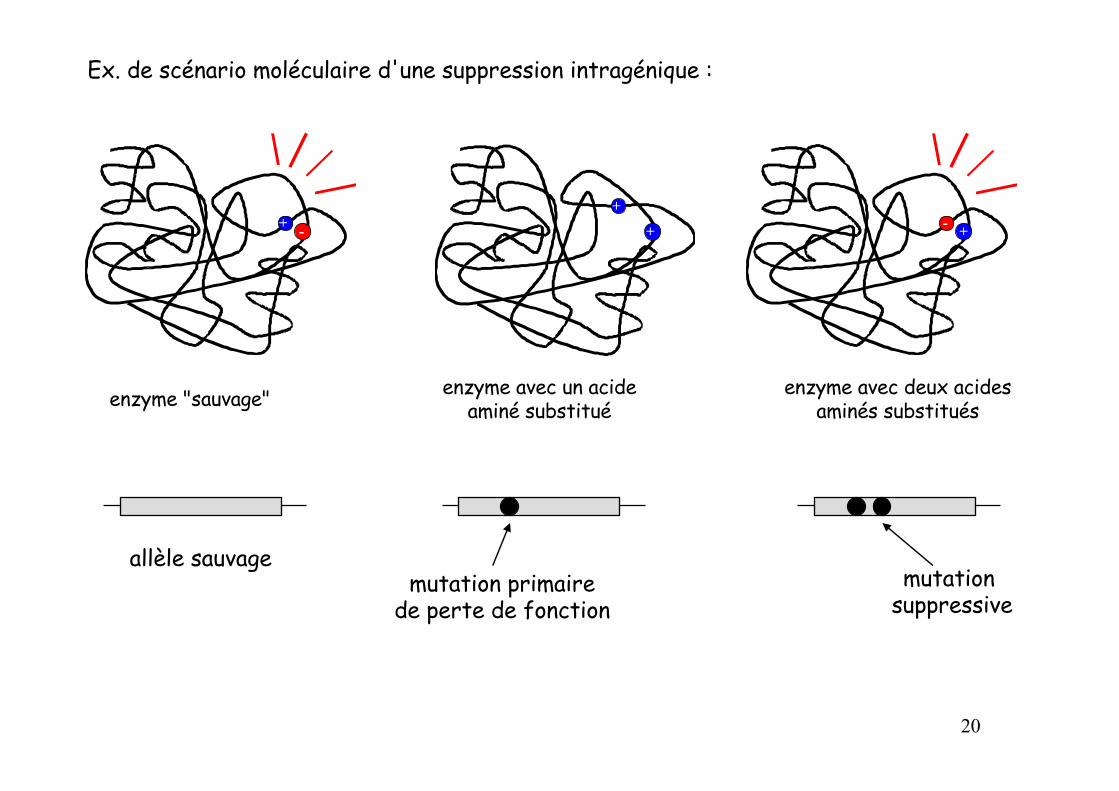
enzyme "sauvage" enzyme avec un acide aminé substitué
enzyme avec deux acides aminés substitués
allèle sauvagemutation primaire
de perte de fonction
+ -
+
+ +-
mutation suppressive
Ex. de scénario moléculaire d'une suppression intragénique :
20
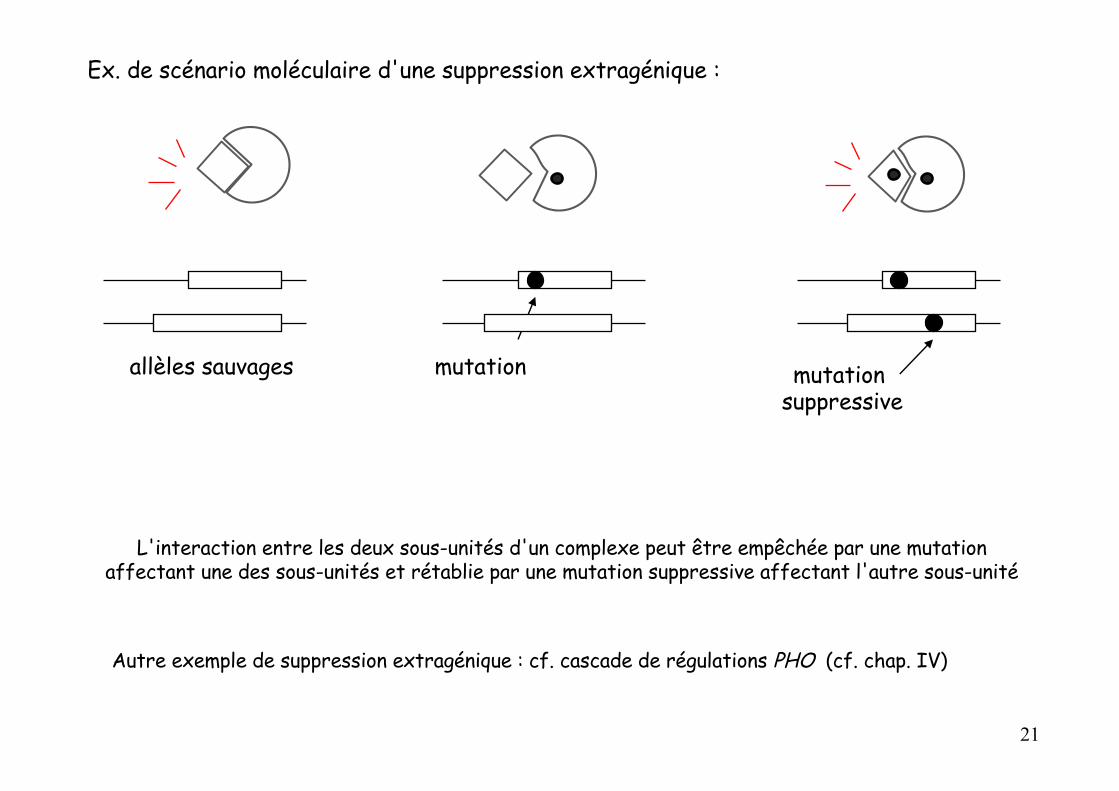
Ex. de scénario moléculaire d'une suppression extragénique :
allèles sauvages mutation mutation suppressive
L'interaction entre les deux sous-unités d'un complexe peut être empêchée par une mutation affectant une des sous-unités et rétablie par une mutation suppressive affectant l'autre sous-unité
Autre exemple de suppression extragénique : cf. cascade de régulations PHO (cf. chap. IV)
21
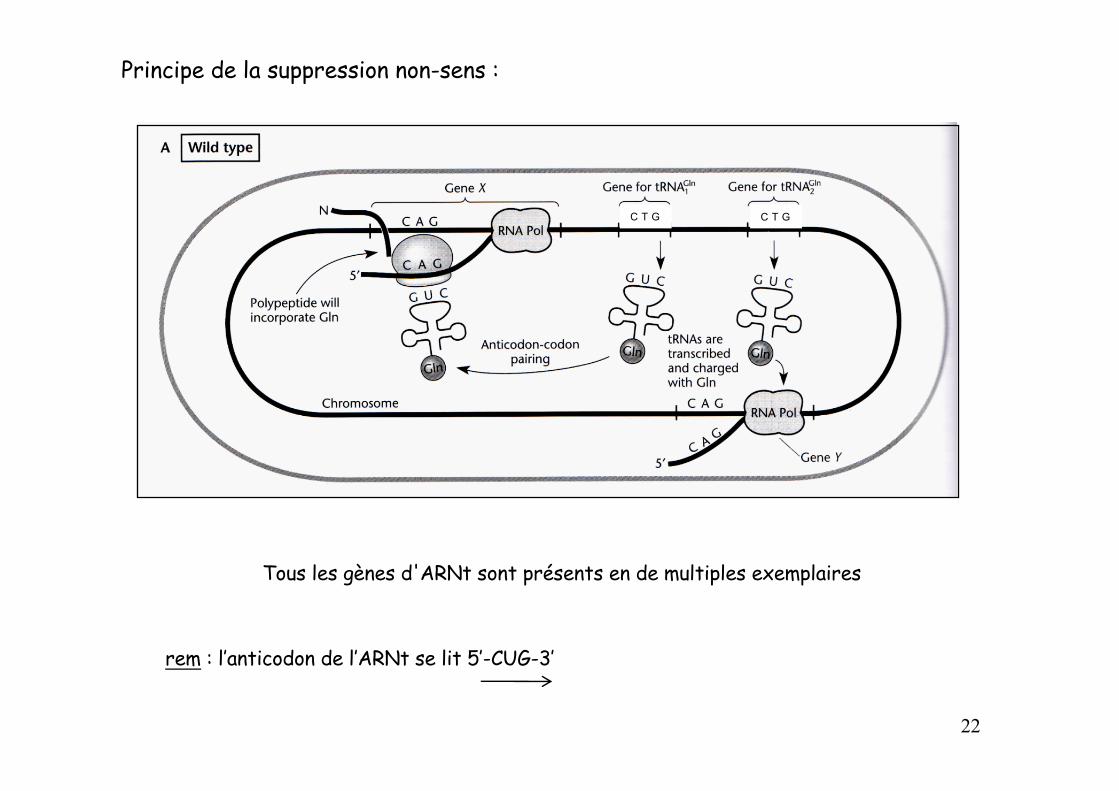
Principe de la suppression non-sens :
Tous les gènes d'ARNt sont présents en de multiples exemplaires
rem : l’anticodon de l’ARNt se lit 5’-CUG-3’
C T G C T G
22
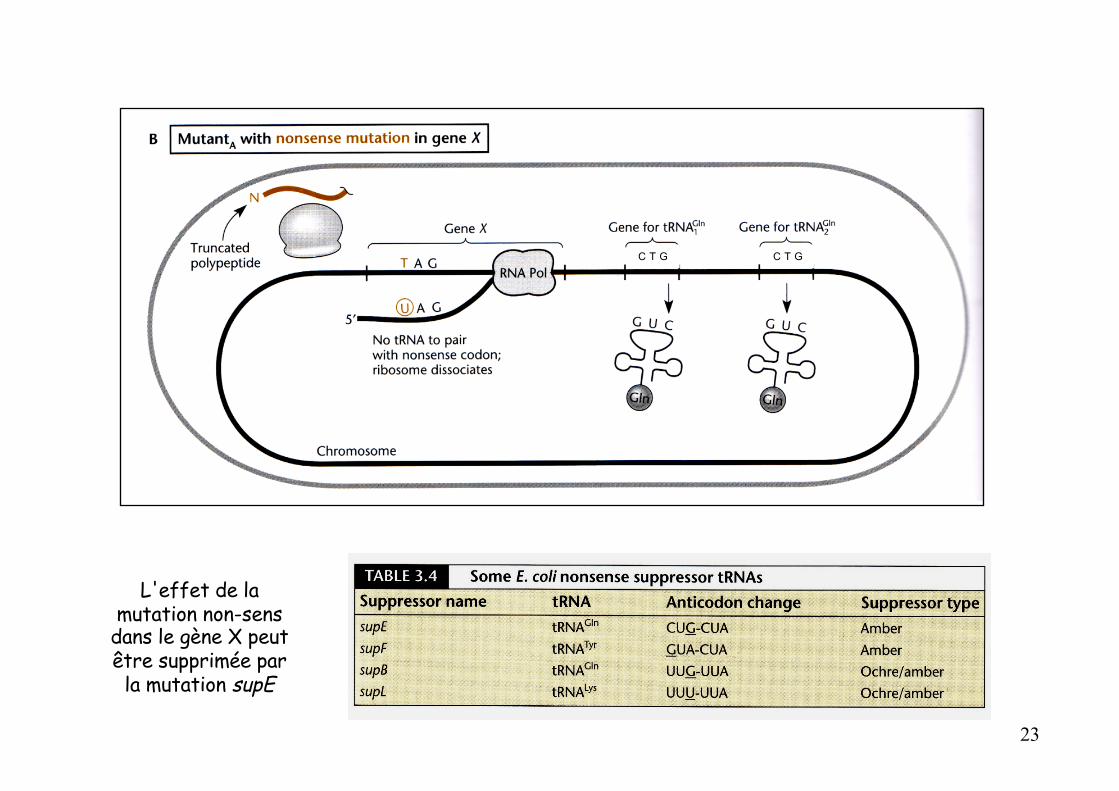
L'effet de la mutation non-sens
dans le gène X peut être supprimée par
la mutation supE
C T G C T G
23
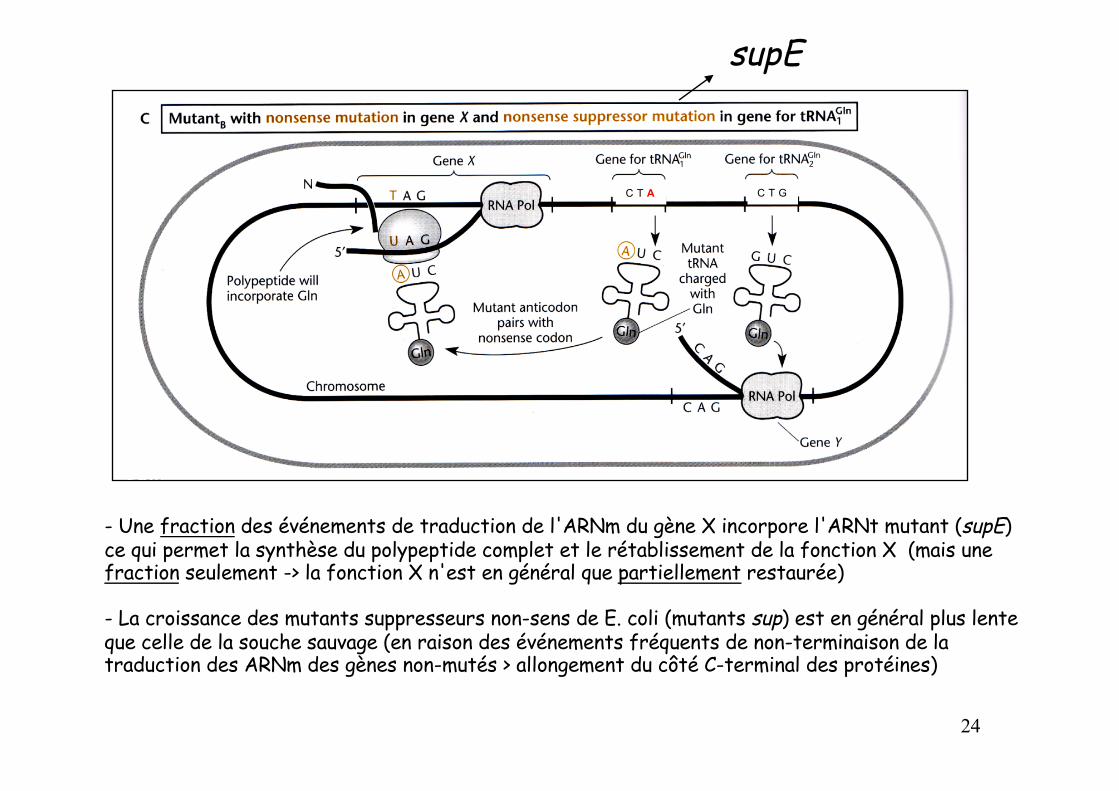
- Une fraction des événements de traduction de l'ARNm du gène X incorpore l'ARNt mutant (supE) ce qui permet la synthèse du polypeptide complet et le rétablissement de la fonction X (mais une fraction seulement -> la fonction X n'est en général que partiellement restaurée)
- La croissance des mutants suppresseurs non-sens de E. coli (mutants sup) est en général plus lente que celle de la souche sauvage (en raison des événements fréquents de non-terminaison de la traduction des ARNm des gènes non-mutés > allongement du côté C-terminal des protéines)
supE
C T A C T G
24


















