
C. Renard UMR SQPOV, Avignon - posalim.ufpr.br pectines_4.pdf · Jus de pommes à cidre: utilisé...
35
Les enzymes pectolytiques C. Renard UMR SQPOV, Avignon Fév 2010
Transcript of C. Renard UMR SQPOV, Avignon - posalim.ufpr.br pectines_4.pdf · Jus de pommes à cidre: utilisé...

Les enzymes pectolytiques
C. RenardUMR SQPOV, Avignon
Fév 2010

Plan
• Les pectines: définition et structure• Méthodes d'étude des pectines• Les pectines en tant qu'additifs• Les enzymes pectolytiques
– Enzymes dégradant la chaaîne homogalacturonique– Enzymes dégradant les zones rhamnogalacturoniques– Enzymes pectolytiques et jus de fruits
• Les pectines in situ: biosynthèse, rôles dans les parois et dans la transformation des fruits et légumes
• Quelques exemples de nos travaux

Les principales pectinases
Type
Hydrolase
HydrolaseHydrolase
Lyase; endo
LyaseLyase
Nom
Pectineméthylestérase
Polygalacturonaseendoexo
Pectine-lyase
Pectate-lyaseendoexo
N°
EC 3.1.1.11
EC 3.2.1.15EC 3.2.1.67
EC 4.2.2.10
EC 4.2.2.2EC 4.2.2.9
Produits
Méthanol + GalA
Chaînes plus courtes, oligosMono OU dimère
Chaînes plus courtes
Dimères
Autres
Fongique (pH 4.5) ou plantes (pH 7)
Viscosité; pH 4-5Extrémité non-réductrice
pH opt: 6
pH opt: 8-9
Substrat
Haut DM
PectatePectate
Haut DM
DM bas
Type
Hydrolase
HydrolaseHydrolase
Lyase; endo
LyaseLyase
Nom
Pectineméthylestérase
Polygalacturonaseendoexo
Pectine-lyase
Pectate-lyaseendoexo
N°
EC 3.1.1.11
EC 3.2.1.15EC 3.2.1.67
EC 4.2.2.10
EC 4.2.2.2EC 4.2.2.9
Produits
Méthanol + GalA
Chaînes plus courtes, oligosMono OU dimère
Chaînes plus courtes
Dimères
Autres
Fongique (pH 4.5) ou plantes (pH 7)
Viscosité; pH 4-5Extrémité non-réductrice
pH opt: 6
pH opt: 8-9
Substrat
Haut DM
PectatePectate
Haut DM
DM bas
Nom
Pectineméthylestérase
Polygalacturonaseendoexo
Pectine-lyase
Pectate-lyaseendoexo
N°
EC 3.1.1.11
EC 3.2.1.15EC 3.2.1.67
EC 4.2.2.10
EC 4.2.2.2EC 4.2.2.9
Produits
Méthanol + GalA
Chaînes plus courtes, oligosMono OU dimère
Chaînes plus courtes
Dimères
Autres
Fongique (pH 4.5) ou plantes (pH 7)
Viscosité; pH 4-5Extrémité non-réductrice
pH opt: 6
pH opt: 8-9
Substrat
Haut DM
PectatePectate
Haut DM
DM bas
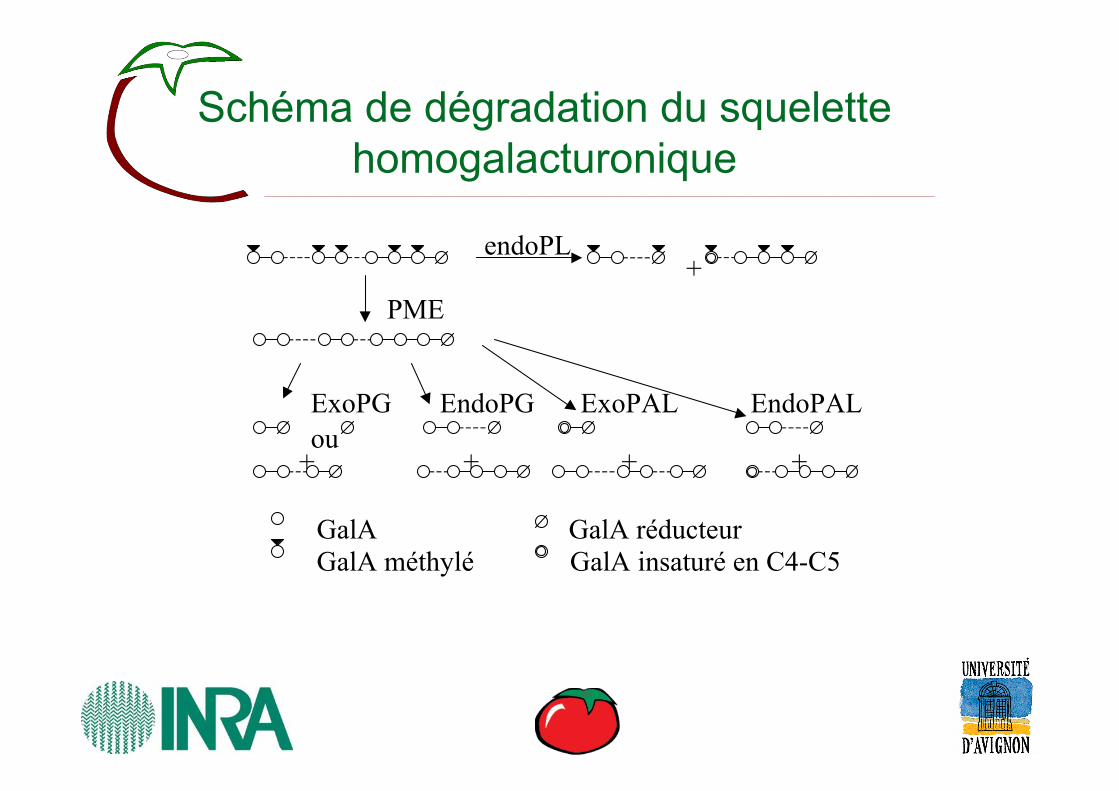
Schéma de dégradation du squelette homogalacturonique
GalAGalA méthylé
GalA réducteur GalA insaturé en C4-C5
endoPL+
PME
ExoPGou
+
EndoPG
+
EndoPALExoPAL
++
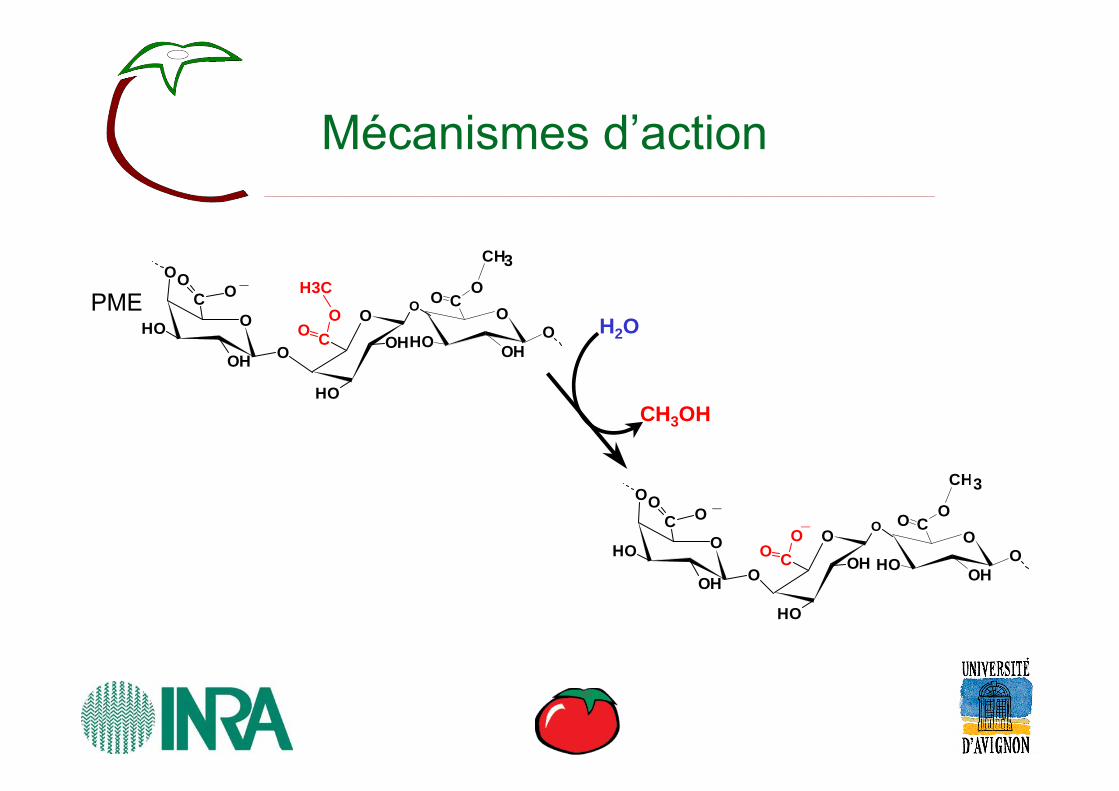
Mécanismes d’action
O
OH
HO
O
O
C OO _
_O
OH
HO
CO
OO
O
O
CH3
OC
HO OH
O
O
OH
HO
O
O
C OO _
O
O
CH3
OC
HO OH
OOOH
HO
CO
OO
H3C
H2O
CH3OH
3
3
PME
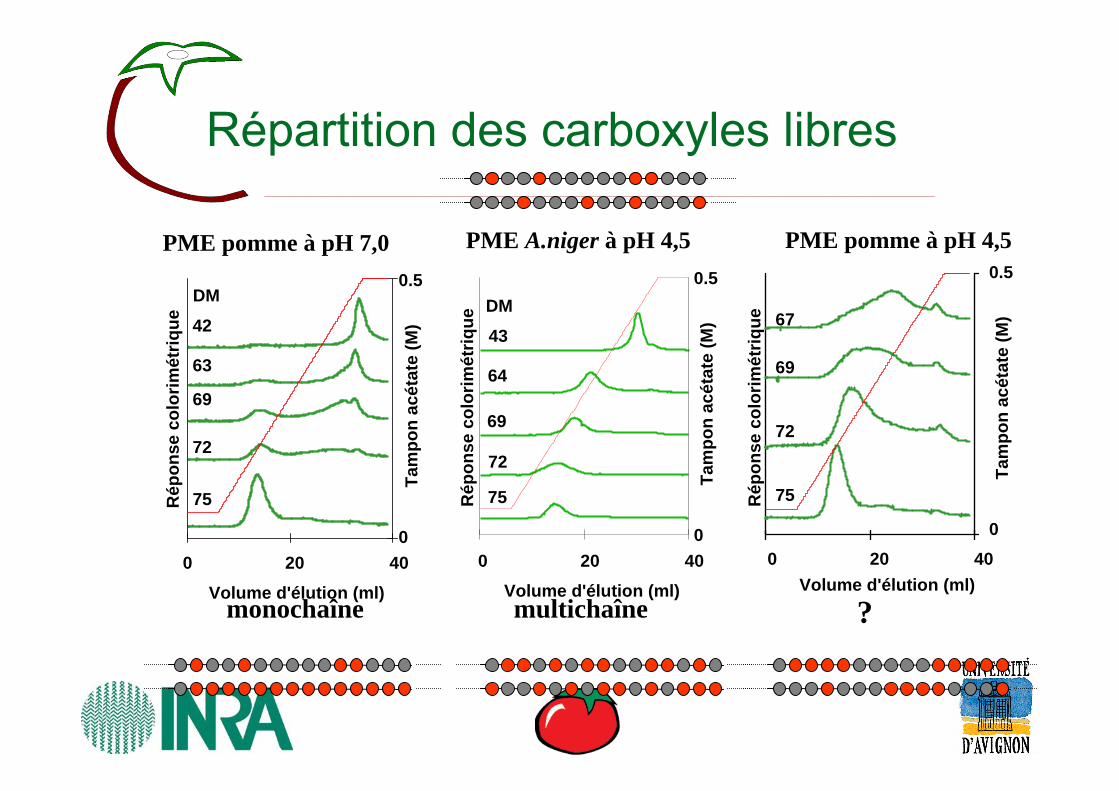
Répartition des carboxyles libres
multichaînemonochaîne ?
0 20 40Volume d'élution (ml)
Rép
onse
col
orim
étriq
ue
0
0.5
Tam
pon
acét
ate
(M)
75
72
69
64
43
DM
PME A.niger à pH 4,5 PME pomme à pH 4,5
0 20 40Volume d'élution (ml)
Rép
onse
col
orim
étriq
ue
0
0.5
Tam
pon
acét
ate
(M)
75
72
69
63
42
DM
PME pomme à pH 7,0
0 20 40Volume d'élution (ml)
0
0.5
75
72
69
Rép
onse
col
orim
étriq
ue
Tam
pon
acét
ate
(M)67

Effets des PMEs endogènes
Déstabilisation des troubles (en présence de calcium)PMEs endogènes, très thermo (et baro)-résistantes
Ca++
Ca++
Ca++
Ca++
Ca++
Ca++
Ca++
Ca++
PE
Apparition de zones à haute densité de charge, fixant le calcium: gélification des pectines suivie de rétraction du gel avec les particules
Jus d'agrumes, ananas... : déphasageJus de pommes à cidre: utilisé en clarification préfermentaire.

Déstabilisation des jus troubles
• Action de la PME : – Diminution du DM des pectines solubles
• Gelification des pectines déméthoxylées– Présence d’ions Calcium dans le jus
• Rétraction du gel de pectate de calcium

Déstabilisation des jus troubles
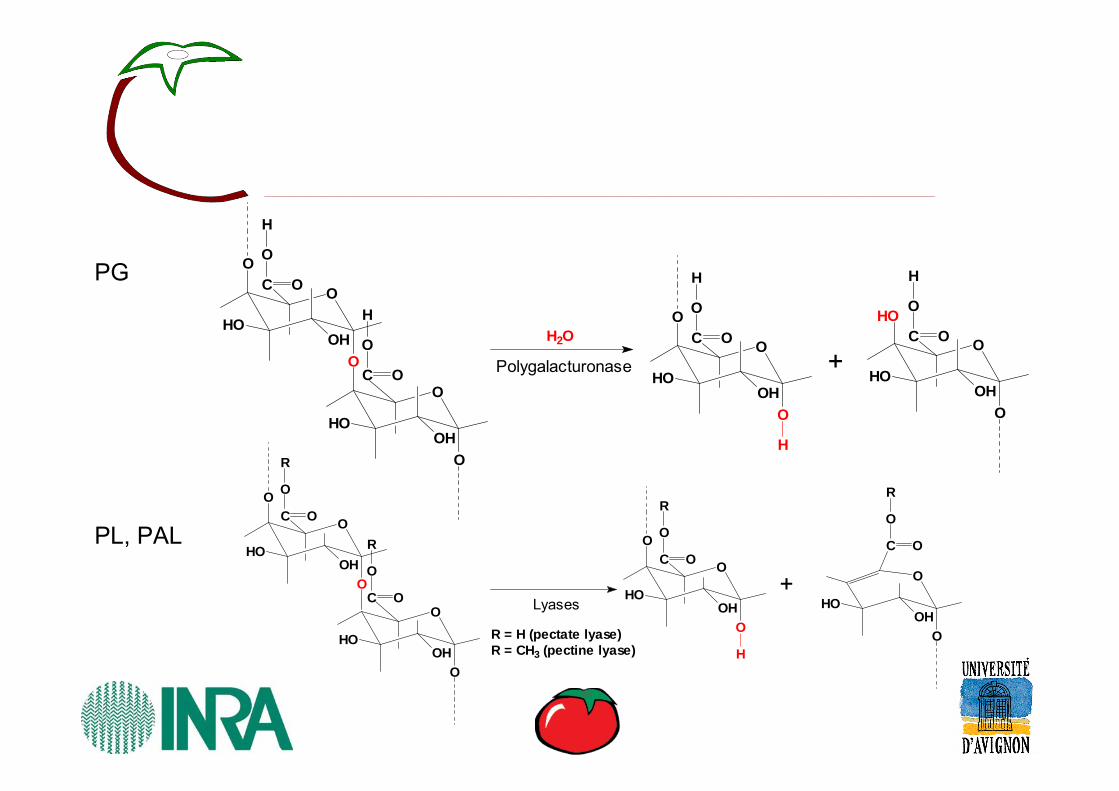
Polygalacturonase
O
HO
O
O
OH
H
C O
O
H
C O
O
H
O
HO
O
O
OH
C O
O
H
O
HO
O
OH
H2O
+
O
HO
O
HO
OH
C O
O
H
+
O
HO
O
OH
O
HO
O
O
OH
C O
O
R
C O
O
R
O
HO
O
O
OH
H
C O
O
R
Lyases
R = H (pectate lyase)R = CH3 (pectine lyase)
O
HO
O
OH
C O
O
R
PG
PL, PAL

Enzymes mineures
• Pectineacétylestérase
• Endo xylogalacturonan hydrolase (XGH; EC 3.2.1.-)– Action endo, sur GalA substitué– Isolée de A. tubingensis– pH opt 3.0–3.5, à 50 °C– Certaines exo-PG agissent sur le XG

Inactivation des enzymes endogènes
• Surtout pectineméthylestérases– Industrie des jus de fruits– Déstabilisation des particules– Enzymes souvent résistantes: thermo et baro-résistantes
• Pour production de jus en pasteurisation athermique– Hautes pressions– Champs électriques pulsés
• Polygalacturonases– Nettement plus labiles– PG1 (100 KDa), PG2 (45 KDa) et sous-unité β
(glycoprotéine, 38 KDa)
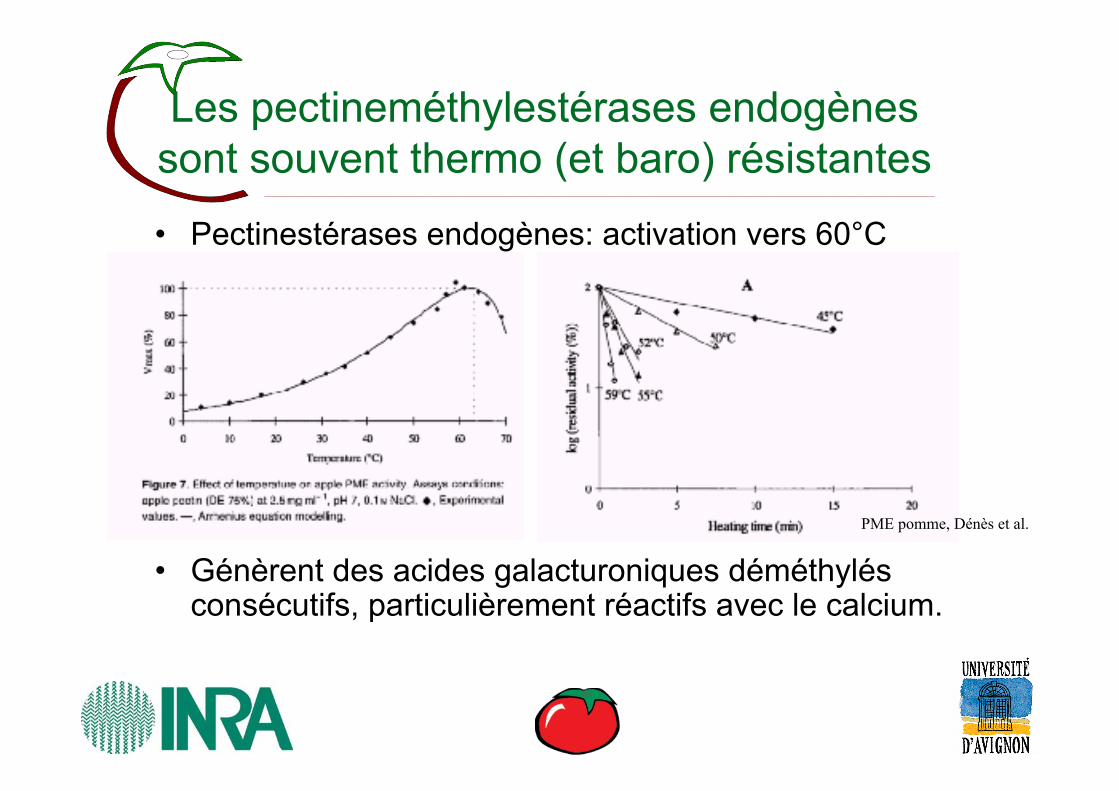
Les pectineméthylestérases endogènes sont souvent thermo (et baro) résistantes
• Pectinestérases endogènes: activation vers 60°C
• Génèrent des acides galacturoniques déméthylésconsécutifs, particulièrement réactifs avec le calcium.
PME pomme, Dénès et al.

Inactivation thermique de PMEs d'orange
0
1
2
0 5 10 15 20 25 30 35 40Heating time (min)
Log
(res
idua
l act
ivity
(%))
50°C
60°C58°C56°C 54°C
52°C

Inactivation thermique des PMEs de plantes
Source System Kinetic model T-range (°C) studied
z/Ea-value Reference
Apple citrate-phosphate pH 4.0 first-order 45 to 55 z = 9.2 °C Denès and others 2000 Banana Tris buffer pH 7.0 first-order 65 to 72.5 z = 5.89 °C Ly Nguyen and others 2002a Carrot carrot tissue first-order 50 to 95 nd Tijskens and others 1997 Carrot Tris buffer pH 7.0 fractional conversion 48 to 60 nd Ly Nguyen and others 2002b Carrot citrate buffer pH 4.5, 6.0 first-order 50 to 60 z pH 6.0= 5.37 °C
z pH 4.5= 6.54 °C Balogh and others 2004
Carrot juice, pH 6.0 first-order 50 to 60 z = 5.73 °C Balogh and others 2004 Carrot pieces first-order 66 to 74 z = 4.13 °C Balogh and others 2004 Grapefruit Tris buffer pH 7.0 fractional conversion 56 to 62 nd Guiavarc'h and others 2005 Guava concentrate pH 8.0 not given 50 to 98 nd Leite and others 2006 Hawthorn pH 7.0 not given 50 to 80 Ea = 146.2 kJ/mol Vivar-Vera and others 2007 Orange water fractional conversion 60 to 67 nd Van den Broeck and others
1999 Orange pulp biphasic 60 to 90 z L= 10.8 °C
z S= 6.5 °C Wicker and Temmelli 1988
Orange juice–milk mixture
citric buffer pH 3.2 to 4.2 biphasic 65 to 80 EaS = 605955 kJ/mol ; EaL = 523603 kJ/mol
Sampedro and others 2008
Papaya citric acid-phosphate buffer, pH 7.5
first-order 60 to 75 z = 7.8 °C Fayyaz and others 1995
Pepper citrate buffer pH 5.6 biphasic 56 to 64 EaL = 371.6 kJ/mol Castro and others 2006 Tomato puree biphasic 60 to 75 z L= 1.7 °C
z S= 7.5 °C Crelier and others 1995
Tomato water biphasic 60 to 75 z L= 9.8 °C ; z S= 6.2 °C
Crelier and others 1995
Tomato water first-order 57 to 65 z = 6.5 °C Crelier and others 2001

Inactivation par les hautes pressionsSource System Kinetic model p–T range Reference Banana 20 mM Tris buffer, pH 7.0 fractional conversion 600 to 700 MPa ; 10 °C Ly Nguyen and others
2002a Carrot 20 mM Tris buffer, pH 7.0 fractional conversion 600 to 700 MPa ; 10 °C Ly Nguyen and others
2002b Carrot pH 6.0 citrate buffer first-order 650 to 800 MPa ; 10 to 25
°C Balogh and others 2004
Carrot juice first-order 700 to 800 MPa ; 10 °C Balogh and others 2004 Carrot pieces first-order 700 to 800 MPa ; 40 °C Balogh and others 2004 Orange juice first-order 100 to 400 MPa ; 10 to 40
°C Basak and Ramaswamy 1996
Orange water fractional conversion 600 to 900 MPa ; 20 to 30 °C
Van den Broeck and others 1999
Orange juice first-order 400 to 600 MPa ; 25 to 50 °C
Nienaber and Shellhammer 2001
Orange juice nd 600 MPa, 20 °C Bull and others 2004 Pepper citrate buffer pH 5.6 fractional
conversion/biphasic 100 to 800 MPa ; 10-64°C Castro and others 2006
Plum Tris buffer pH 7.5 first-order 650 to 800 MPa Nunes and others 2006 Tomato puree first-order 400 to 800 MPa ; 30 to 75
°C Crelier and others 1995
Tomato water first-order 100 to 900 MPa ; 40 to 60 °C
Van den Broeck and others 2000b
Tomato citrate buffer pH 6.0 first-order 100 to 800 MPa ; 20°C, 40 °C
Plaza and others 2007
White grapefruit
20 mM Tris buffer, pH 7.0 fractional conversion 100 to 800 MPa ; 10 to 62 °C
Guiavarc'h and others 2005
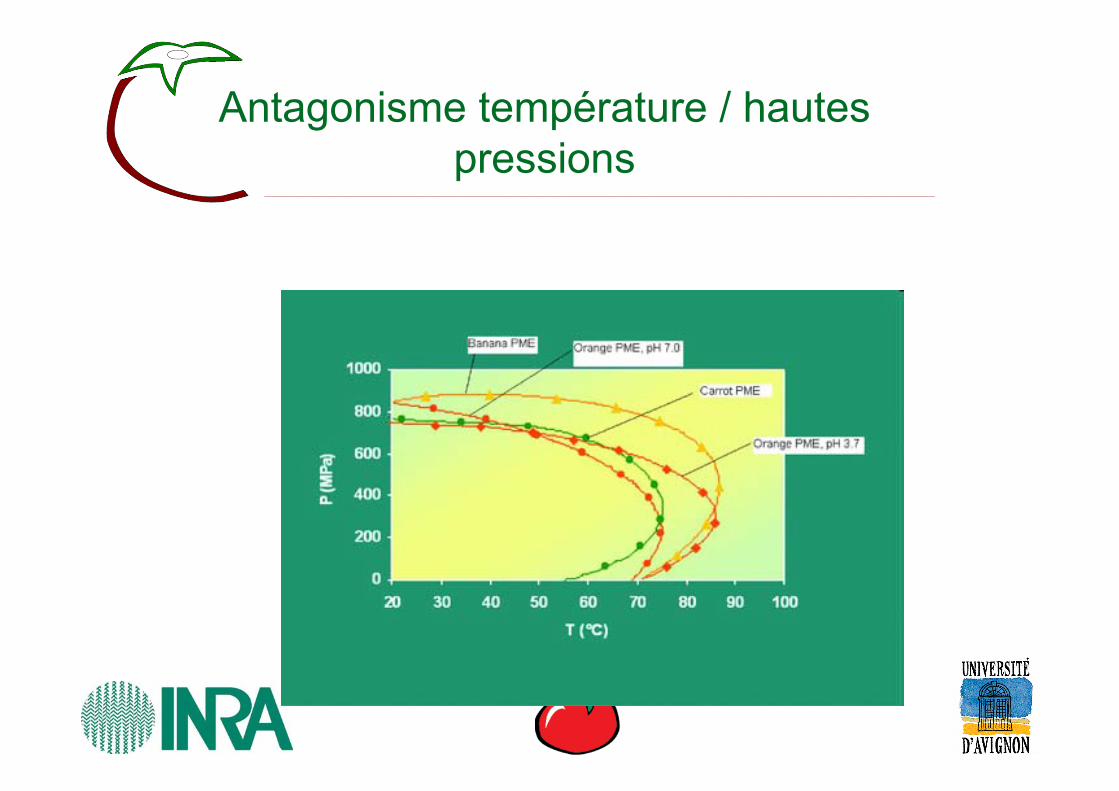
Antagonisme température / hautes pressions

Champs électriques pulsésEnzyme Medium Treatment conditions Inactivation (%) Reference Tomato PME Distilled water 30 kV/cm, 400 μs ≤10 Van Loey and others 2002 Tomato PME Tomato juice 35 kV/cm, 1500 μs, <35 °C 82 Aguilo-Aguayo and others 2008 Tomato PME Tris-HCl buffer (pH 7.0) 16.8 kV/cm, 1600 μs, ≤35.1 °C 87 Espachs-Barroso and others 2006 Tomato PME Tomato extract 24 kV/cm, 8 ms, ≤15 °C 93.8 Giner and others 2000 Orange PME Freshly squeezed, frozen,
and thawed Valencia orange juice
35 kV/cm, 59 μs, ≤60 °C 90 Yeom and others 2000
Orange PME Orange juice 25 kV/cm, 250 ms, ≤64 °C 90 Yeom and others 2002 Orange PME Distilled water and
McIlvaine buffer (pH 3.7) 30 kV/cm, 400 μs ≤10 Van Loey and others 2002
Orange PME Orange juice 35 kV/cm, 1000 μs ≤10 Van Loey and others 2002 Orange PME Tris-HCl buffer (pH 7.0) 17.5 kV/cm, 400 μs, ≤67.2 °C 87 Espachs-Barroso and others 2006 Carrot PME Tris-HCl buffer (pH 7.0) 15 kV/cm, 600 μs, ≤62.1 °C 83 Espachs-Barroso and others 2006 Banana PME Tris-HCl buffer (pH 7.0) 16.8 kV/cm, 1600 μs, ≤35.1 °C 45 Espachs-Barroso and others 2006 Orange PME Orange juice 35 kV/cm, 1000 μs, ≤40 °C 81.6 Elez-Martinez and others 2006 Orange PME Orange juice 35 kV/cm, 1500 μs, ≤37.5 °C 80 Elez-Martinez and others 2007b PME Citrus juices: Clementine
mandarines, Ortanique fruits, Valencia oranges
25 kV/cm, 330 μs, ≤72 °C 88.3, 85.2, 90.6 Sentandreu and others 2006
PME Orange–carrot juice (80% to 20%) (batch A)
25 kV/cm, 280 μs, ≤68 °C 75.6 Rivas and others 2006
PME Orange-carrot juice (80% to 20%) (batch B)
25 kV/cm, 330 μs, ≤70 °C 81 Rivas and others 2006
PME Commercial preparation 38 kV/cm, 340 μs, <40 °C 86.8 Giner and others 2005 PME Orange–carrot juice (80%
to 20%) 25 kV/cm, 340 μs, 63 °C 81.4 Rodrigo and others 2003
Enzyme Medium Treatment conditionsa Inactivation (%) Reference Tomato PME Distilled water 30 kV/cm, 400 μs ≤10 Van Loey and others 2002 Tomato PME Tomato juice 35 kV/cm, 1500 μs, <35 °C 82 Aguilo-Aguayo and others 2008 Tomato PME Tris-HCl buffer (pH 7.0) 16.8 kV/cm, 1600 μs, ≤35.1 °C 87 Espachs-Barroso and others 2006
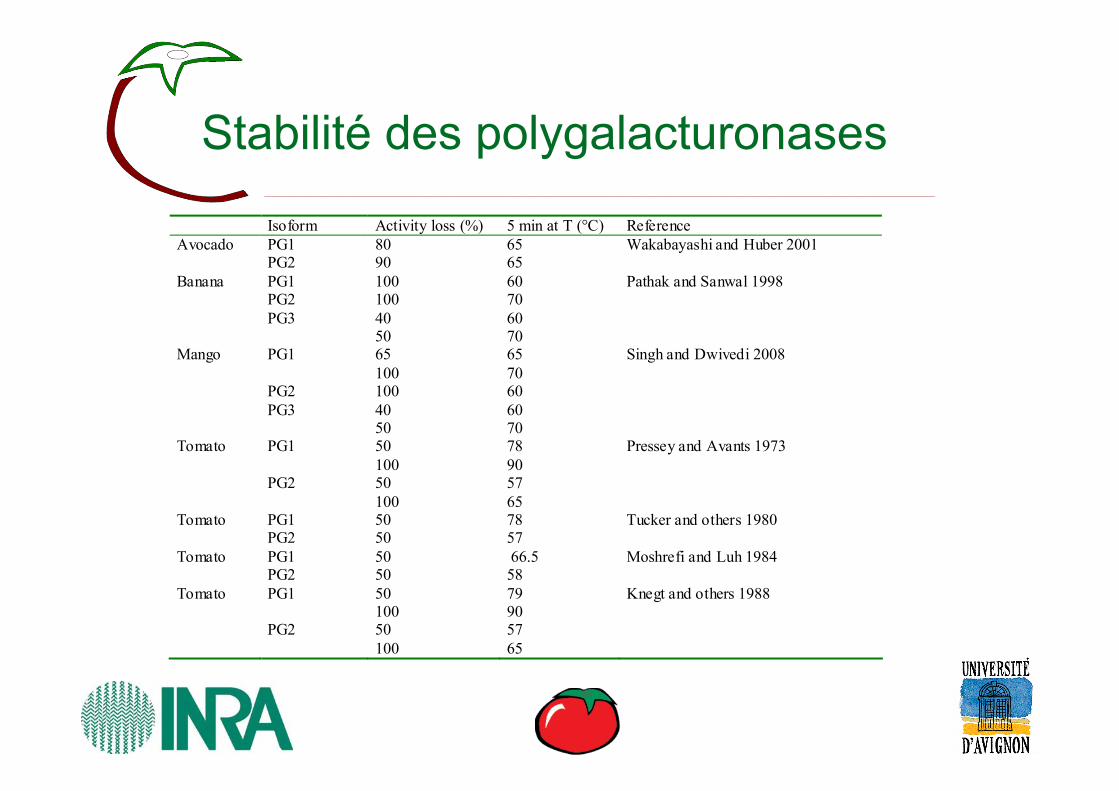
Stabilité des polygalacturonases
Isoform Activity loss (%) 5 min at T (°C) Reference Avocado PG1 80 65 Wakabayashi and Huber 2001 PG2 90 65 Banana PG1 100 60 Pathak and Sanwal 1998 PG2 100 70 PG3 40 60 50 70 Mango PG1 65 65 Singh and Dwivedi 2008 100 70 PG2 100 60 PG3 40 60 50 70 Tomato PG1 50 78 Pressey and Avants 1973 100 90 PG2 50 57 100 65 Tomato PG1 50 78 Tucker and others 1980 PG2 50 57 Tomato PG1 50 66.5 Moshrefi and Luh 1984 PG2 50 58 Tomato PG1 50 79 Knegt and others 1988 100 90 PG2 50 57 100 65
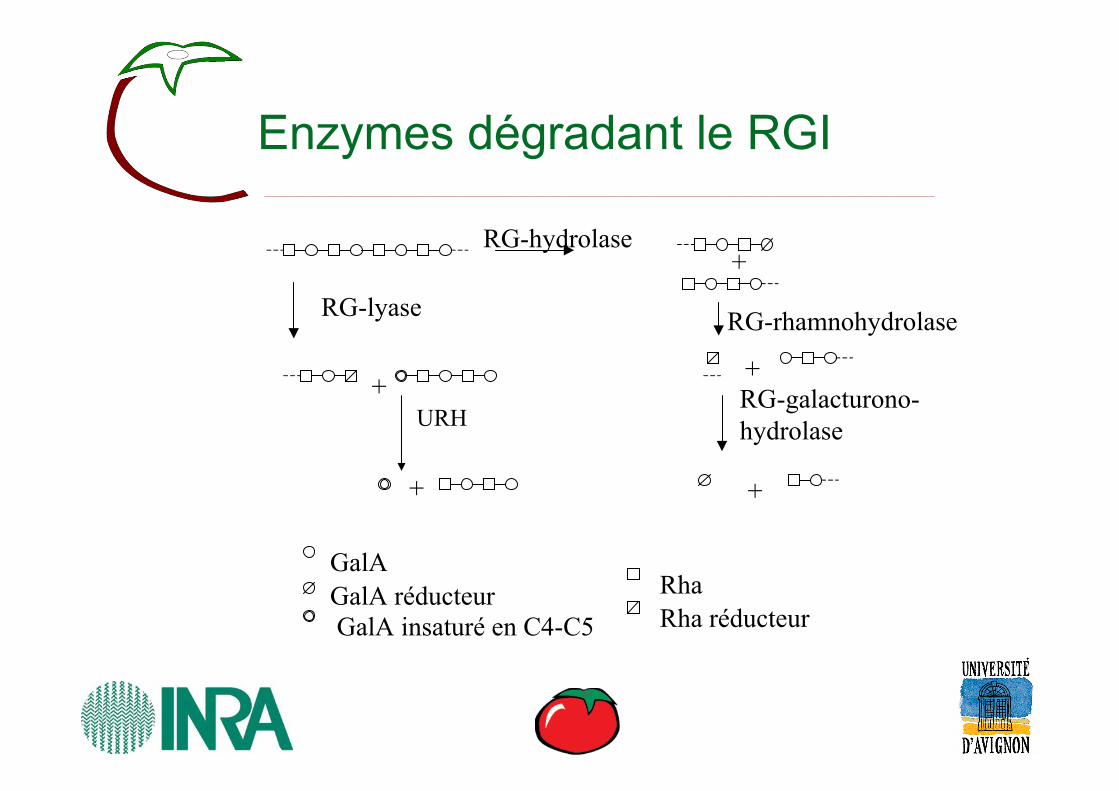
Enzymes dégradant le RGI
RG-lyase
+
RG-hydrolase+
RG-rhamnohydrolase
+RG-galacturono-hydrolase
+
GalAGalA réducteurGalA insaturé en C4-C5
RhaRha réducteur
URH
+

• Deux endo– (1) Rhamnogalacturonan α-d-galactopyranosyuronide-(1→2)-α-l-
rhamnopyranosyl hydrolase (rhamnogalacturonan hydrolase; endo-rhamnogalacturonan hydrolase; RG-hydrolase)
– (2) Rhamnogalacturonan α-l-rhamnopyranosyl-(1→4)-α-d-galactopyranosyluronide lyase (Rhamnogalacturonan lyase; endo-rhamnogalacturonan lyase; RG-lyase).
• Trois exo– (1) Unsaturated rhamnogalacturonyl hydrolase (URH) – (2) Rhamnogalacturonan α-l-rhamnopyranohydrolase (RG-
rhamnohydrolase) – (3) Rhamnogalacturonan α-d-galactopyranosyluronohydrolase
(RG-galacturonohydrolase)• Une estérase
– Rhamnogalacturonan acetylestérase
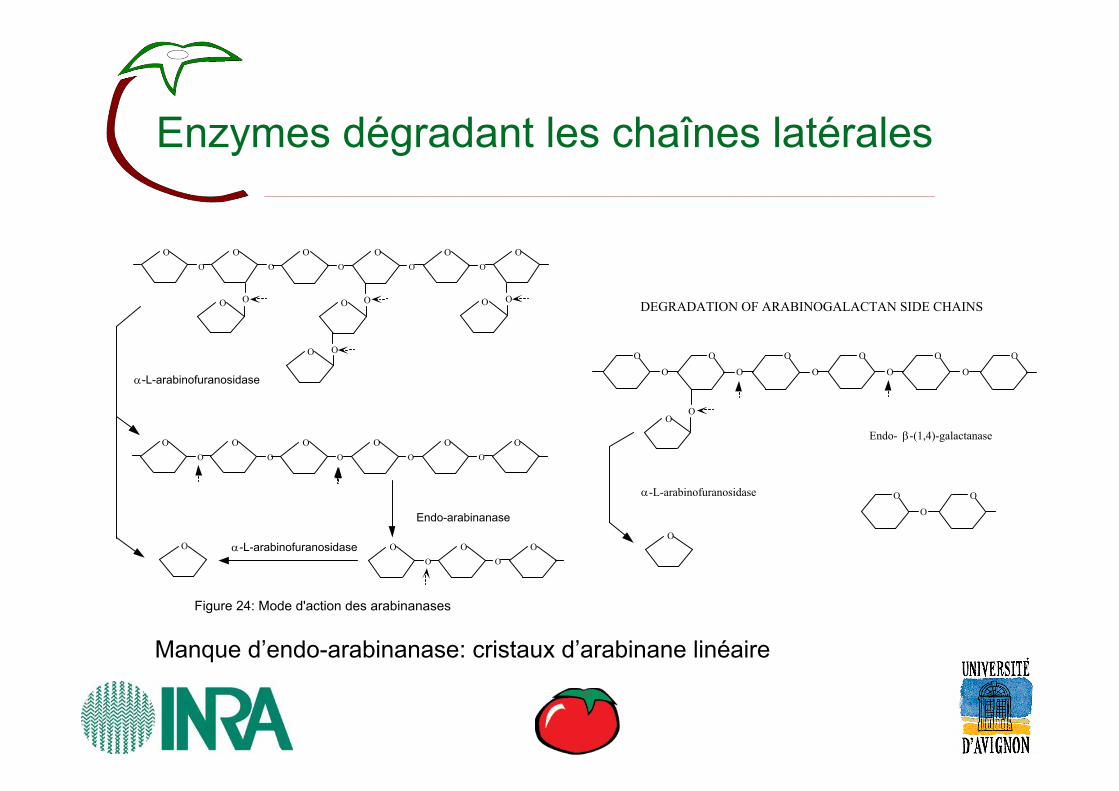
Enzymes dégradant les chaînes latérales
O
OO
OO
OO
OOO
OO
OO
OO
OO
OOO
OO
OO
O
O OO
O
O
OO
OO
O
α-L-arabinofuranosidase
Endo-arabinanase
α-L-arabinofuranosidase
Figure 24: Mode d'action des arabinanases
OO
O
O
O
O
O
O
OO
O
O
O
O
O
O
O
α-L-arabinofuranosidase
Endo- β-(1,4)-galactanase
DEGRADATION OF ARABINOGALACTAN SIDE CHAINS
Manque d’endo-arabinanase: cristaux d’arabinane linéaire

Utilisation d’enzymes en technologie des jus de fruits
• Clarification– Pectinases– PME (cidre)
• Aide au pressage– Pectinases
• Macération– Pectinases
• Liquéfaction– Pectinases et cellulases

Déstabilisation des troubles par dépolymérisation des pectines
Avantage supplémentaire: dégradation des pectines solubles, ce qui évite: viscosité et gélification lors de la concentrationformation d'une couche de diffusion lors de la filtration
Les pectines dégradées dévoilent les charges + des particules. En l'absence de macromolécules ces dernières s'agrègent par attraction électrostatique et précipitent.
PG + PE, PL
O
O
OO
OO O
OO
O
O O
O
O
OO
O
OpH 3,5
pH 5,0
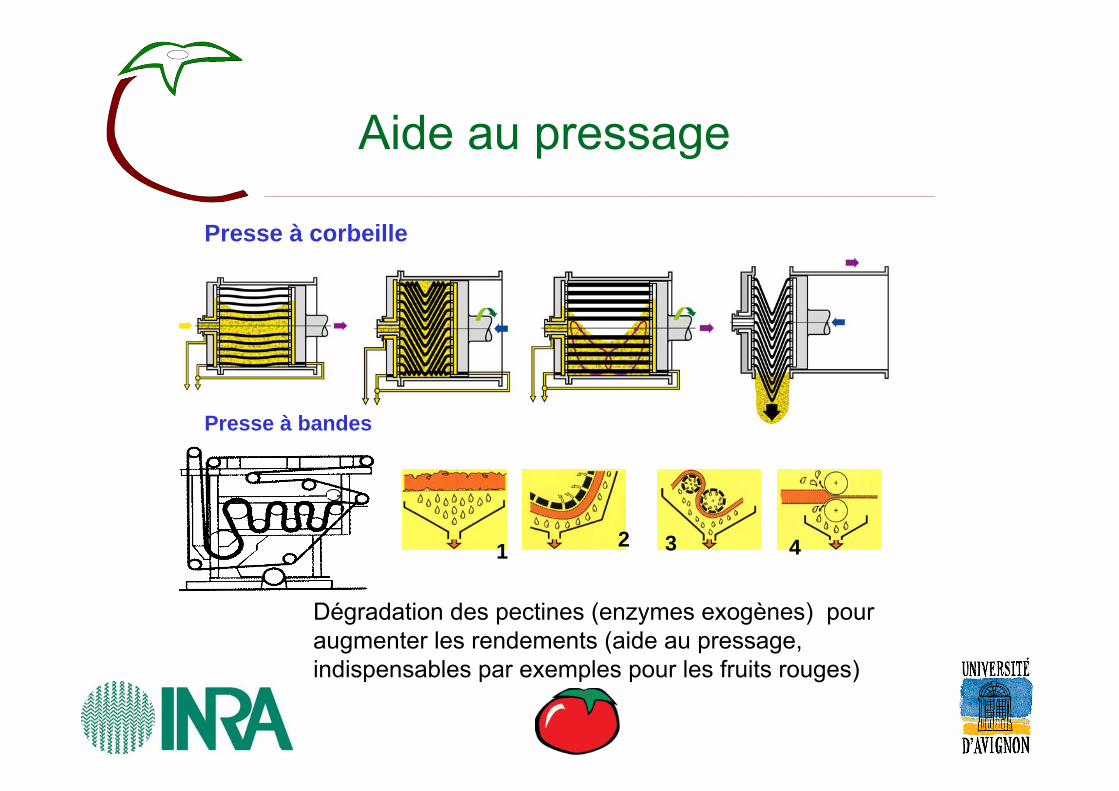
Aide au pressage
Dégradation des pectines (enzymes exogènes) pour augmenter les rendements (aide au pressage, indispensables par exemples pour les fruits rouges)
Presse à corbeille
1 2 3 4
Presse à bandes
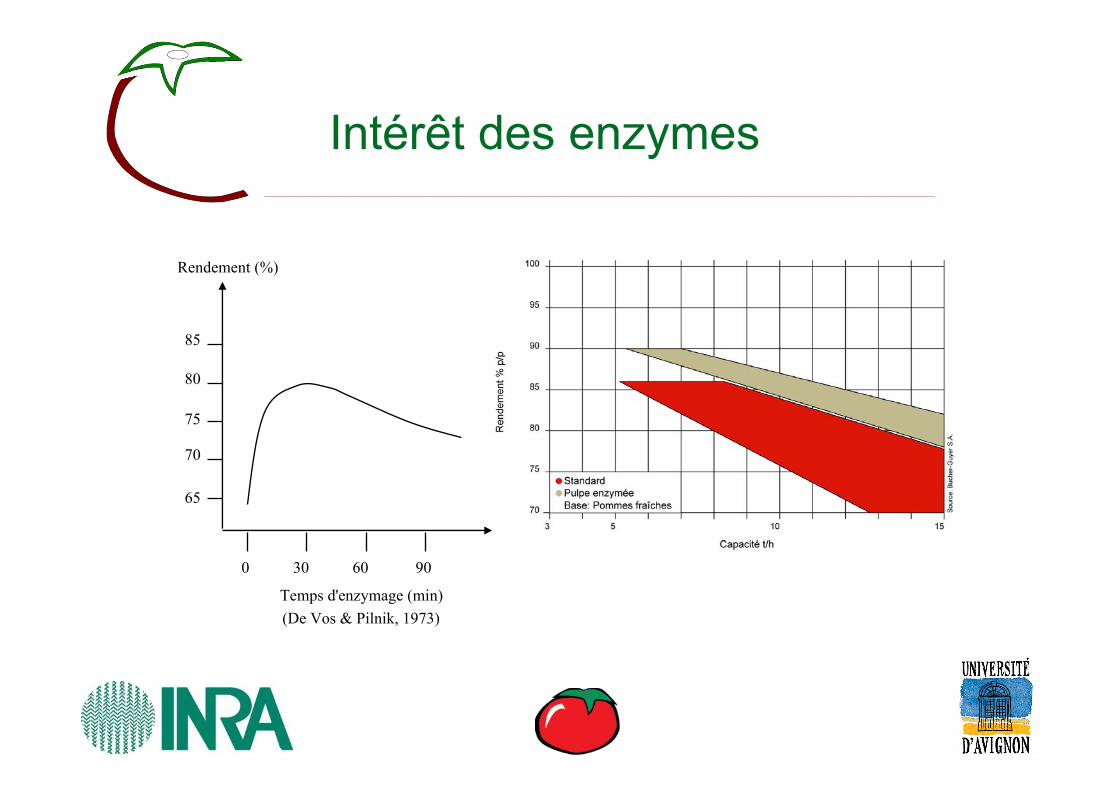
Intérêt des enzymes
(De Vos & Pilnik, 1973)
Rendement (%)
Temps d'enzymage (min)
0 30 60 90
65
70
75
80
85

Procédés nectars
BroyageLavage
2 - 3 heures35 °C
PG
Cuve sous agitation
Tamis vibrant
Cellules
Jus visqueux
Embouteillage
Pasteurisation
Procédé enzymatique: macérationProcédé classique
Washing bowl
Cores remove
Helicoidal crusher
Heat treatment
Finisher
Concentrated sediment
Nectar
Clear juice
Dilution
Concentrate
Aroma
Ascorbic acid(300 mg / kg)
Apples
Dégradation mesurée des parois pour aboutir à la libération de cellules intactes.Enzymes pectolytiques, qui se retrouvent très vite essentiellement dans la phase liquide.Produits stables, à seuil d'écoulement
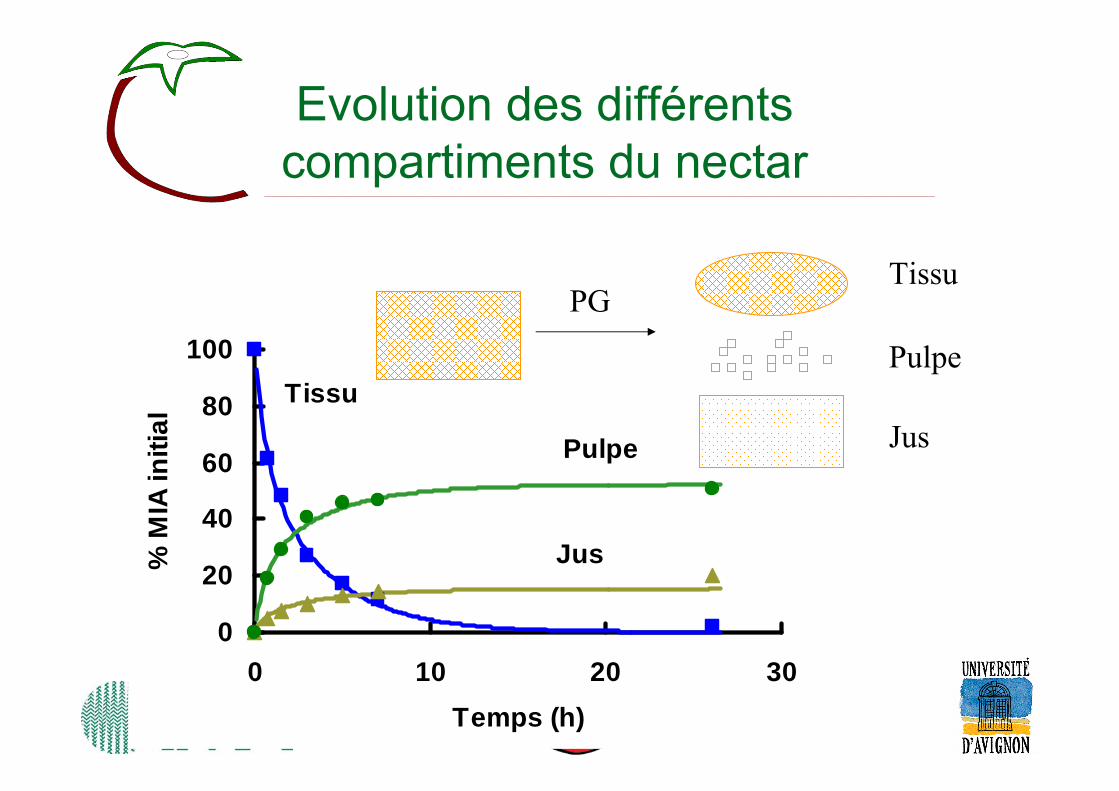
Evolution des différentscompartiments du nectar
0
20
40
60
80
100
0 10 20 30Temps (h)
% M
IA in
itial
Pulpe
Jus
Tissu
PGTissu
Pulpe
Jus
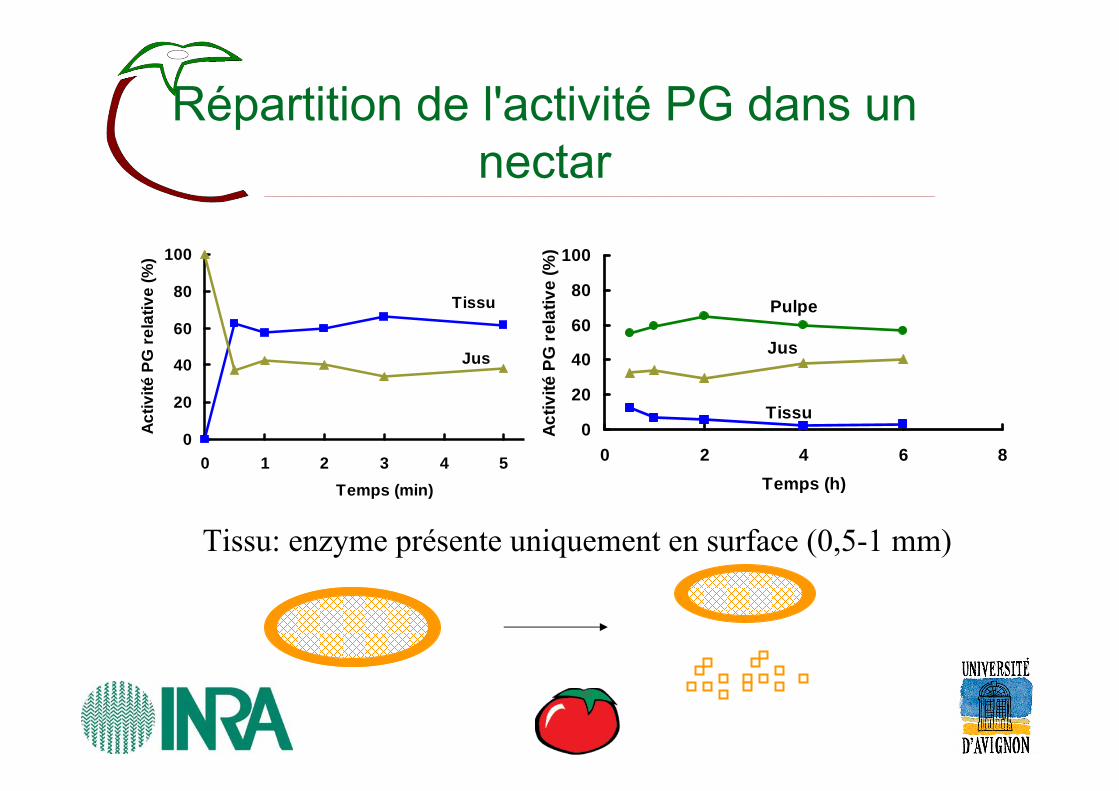
Répartition de l'activité PG dans un nectar
0
20
40
60
80
100
0 1 2 3 4 5 6Temps (min)
Activ
ité P
G re
lativ
e (%
)
Tissu
Jus
0
20
40
60
80
100
0 2 4 6 8Temps (h)
Activ
ité P
G re
lativ
e (%
)
Pulpe
Jus
Tissu
Tissu: enzyme présente uniquement en surface (0,5-1 mm)

Cellulases
Cellulose Gn G2 GEndoglucanaseEndoglucanase CellobiohydrolaseCellobiohydrolase ββ--GlucosidaseGlucosidase
GlucohydrolaseGlucohydrolase
Nom
endoglucanase (= endo-1,4-β-glucanase)
cellobiohydrolase(=exoglucanase)
β-glucosidase
N°
E.C. 3.2.1.4
E.C. 3.2.1.91
E.C. 3.2.1.21
Mw (T. viride)
20-50 Kda
50-60 Kda
76 KDa
pH opt (T. viride)
3.5-7.7
3.5-5.3
3.9
Eads (mg/g)(T. viride)
126-2.8
63-6.6
ND
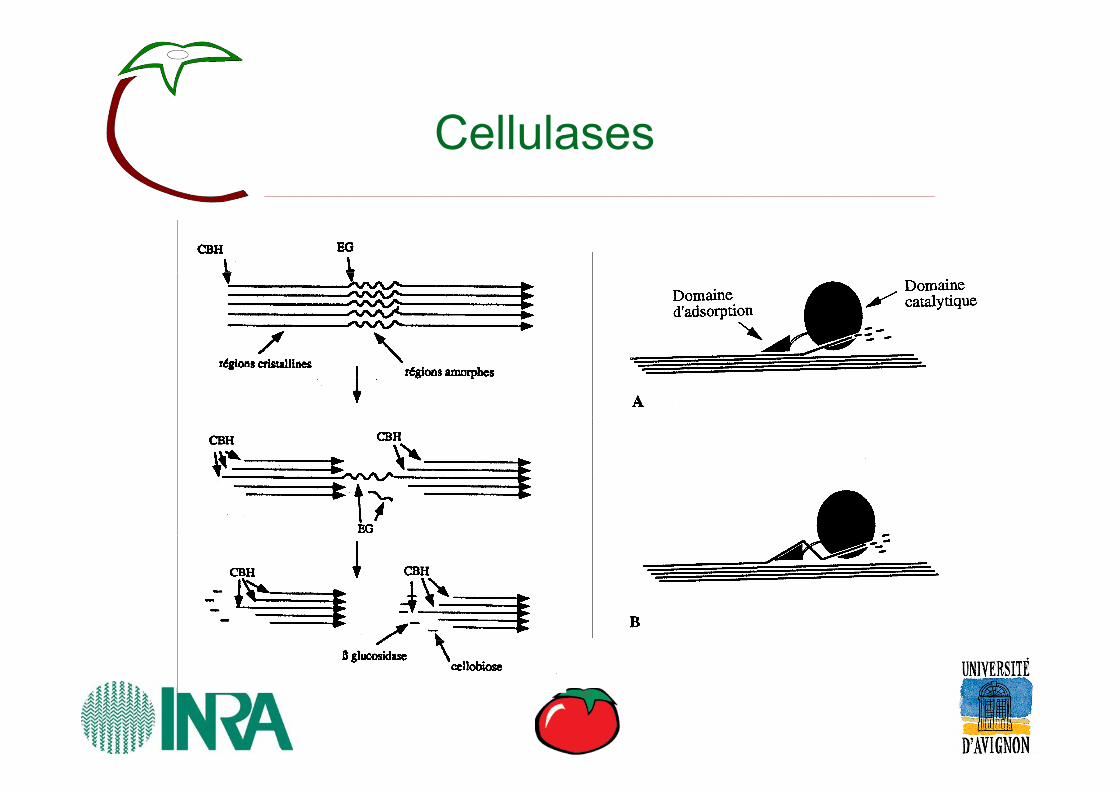
Cellulases

Liquéfaction
Dégradation des parois pour fabrication de jus sans pressage.
Enzymes pectolytiques + cellulases
Dans le jus: acide galacturonique (RG II), cellobiose.
Broyage
PME + PG, PLCellulases
Lavage Traitement thermique
Cuve sous agitation
50 °C
90 °C3 min
2 - 3heures
DécanteurMicro filtration
Concentratration Jus concentré
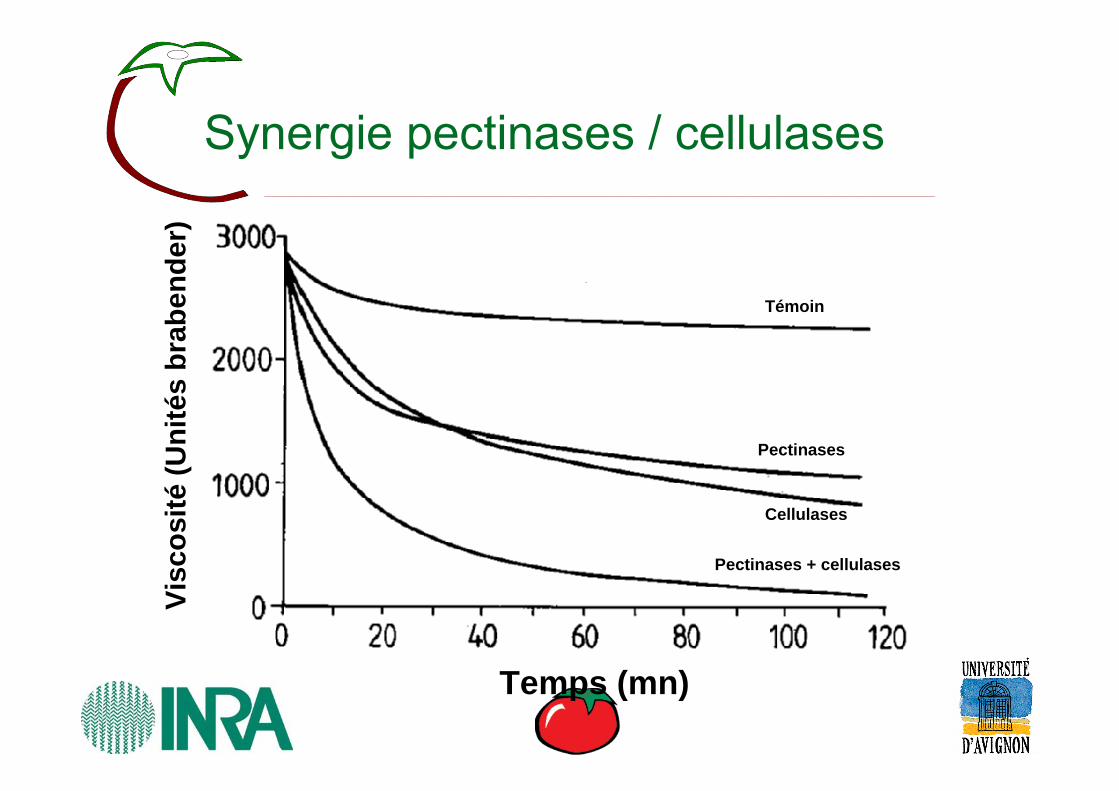
Synergie pectinases / cellulases
Témoin
Pectinases
Cellulases
Pectinases + cellulases
Temps (mn)
Visc
osité
(Uni
tés
brab
ende
r)

Aspect des produitsParenchyme de pomme Nectar commercial de pomme
Nectar de pomme macéréeParenchyme liquéfié de pomme

De très nombreuses enzymes commerciales
• Source: – Aspergillus niger
• Variation:– Ratio PME /PG– Intensité relative des activités secondaires
• Quelques marques– Boehringer Ingelheim, Allemagne Panzym– Ciba-Geigy, A.G. Basel, Suisse Ultrazyme– Grinsted, Danemark Pectolase– Kikkoman Shoyu, Co.Tokyo, JapanSclase– Schweizerische Ferment, A.G. Basel, Suisse Pectinex– DSM (Rapidase) , Pays-Bas Rapidase, Clairzyme– AB Enzymes (Rohm), Darmstadt, Allemagne Pectinol, Rohament– Biocon Pvt LtdBangalore, IndiaPectinase


















