C D Dissémination et domestication du cocotier à la ...
13
233 C OCOTIER Juillet - Août 1998 Plantations, recherche, développement Dissémination et domestication du cocotier à la lumière des marqueurs RFLP Lebrun P., Grivet L., Baudouin L. CIRAD, BP 5035, 34032 Montpellier Cedex 1, France L ’extension géographique et la diversité du cocotier pour le port de l’arbre, la forme et la couleur du fruit (planche photos) ont conduit à identifier un grand nombre de variétés. Dans une liste interna- tionale, établie pour le compte de l’Ipgri 1 , nous avons recensé plus de 300 noms de cul- tivars correspondant à des populations tra- ditionnelles cultivées ou améliorées par sélection massale. Si certains d’entre eux représentent, sans doute, des populations mal définies ou même éteintes, il n’en reste pas moins que près de 60 ont fait l’objet d’échanges internationaux entre stations de recherche dans les 50 dernières années. Ceci traduit l’intérêt des sélectionneurs pour la collecte et l’utilisation des ressources géné- tiques du cocotier. Dix-sept pays du réseau Cogent 2 ont contribué à la base de donnée Cgrd 3 , ils maintiennent plus de 900 acces- sions, et se sont engagés à fournir une liste prioritaire de descripteurs. Dans les programmes d’amélioration, les ressources génétiques jouent un rôle clé, dans la mesure où elles constituent une source de variation utilisable par sélection. Dans certains cas, la réalisation d’hybrides entre populations a mis en évidence une forte hétérosis (Nucé de Lamothe et Bénard, 1985). Enfin, elles constituent un réservoir de variabilité pour la recherche de carac- tères d’adaptation tant aux conditions édaphoclimatiques qu’aux maladies et rava- geurs (Whitehead, 1968). Si la richesse des collections de cocotiers est un atout considérable pour l’amélio- ration, son importance même rend indis- pensable l’établissement d’une stratégie raisonnée d’exploitation. Le nombre de combinaisons possibles entre cultivars dé- passe largement les possibilités du dispositif d’amélioration mondial. La conservation de l’ensemble des cultivars occupe un espace très important et sa description représente un travail considérable. L’identification d’un sous-ensemble de populations représentatif de la diversité de la plante s’avère néces- saire. L’utilisation de caractères morpholo- 1 Ipgri : International Plant Genetic Resources Institute. 2 Cogent : Coconut Genetic Resources Network. 3 Cgrd : Coconut Genetic Resources Database. D eux grands groupes de cocotiers se distinguent clairement par leur profil RFLP. L’un correspond aux populations d’Asie du Sud-Est et du Pacifique et a donné naissance aux Nains. L’autre s’est individualisé dans le sous- continent indien et a été disséminé par l’Homme vers l’Afrique de l’Ouest et la région Caraïbe. Le contenu de cet article a fait l’objet d’un exposé présenté à l’International symposium on coconut biotechnology, à Mérida, Yucatán (Mexique), du 1 er au 5 décembre 1997. Retour au menu
Transcript of C D Dissémination et domestication du cocotier à la ...

233CO C O T I E R
Juil let - Août 1998 Plantations, recherche, développement
Dissémination et domestication
du cocotier à la lumière
des marqueursRFLP
Lebrun P., Grivet L., Baudouin L.CIRAD, BP 5035, 34032 Montpellier Cedex 1, France
L ’extension géographique et la diversitédu cocotier pour le port de l’arbre, laforme et la couleur du fruit (planche
photos) ont conduit à identifier un grandnombre de variétés. Dans une liste interna-tionale, établie pour le compte de l’Ipgri1,nous avons recensé plus de 300 noms de cul-tivars correspondant à des populations tra-ditionnelles cultivées ou améliorées parsélection massale. Si certains d’entre euxreprésentent, sans doute, des populationsmal définies ou même éteintes, il n’en restepas moins que près de 60 ont fait l’objetd’échanges internationaux entre stations derecherche dans les 50 dernières années. Cecitraduit l’intérêt des sélectionneurs pour lacollecte et l’utilisation des ressources géné-tiques du cocotier. Dix-sept pays du réseauCogent2 ont contribué à la base de donnéeCgrd3, ils maintiennent plus de 900 acces-sions, et se sont engagés à fournir une listeprioritaire de descripteurs.
Dans les programmes d’amélioration, lesressources génétiques jouent un rôle clé,dans la mesure où elles constituent unesource de variation utilisable par sélection.Dans certains cas, la réalisation d’hybridesentre populations a mis en évidence uneforte hétérosis (Nucé de Lamothe et Bénard,1985). Enfin, elles constituent un réservoirde variabilité pour la recherche de carac-tères d’adaptation tant aux conditionsédaphoclimatiques qu’aux maladies et rava-geurs (Whitehead, 1968).
Si la richesse des collections de cocotiersest un atout considérable pour l’amélio-ration, son importance même rend indis-pensable l’établissement d’une stratégie raisonnée d’exploitation. Le nombre de combinaisons possibles entre cultivars dé-passe largement les possibilités du dispositifd’amélioration mondial. La conservation del’ensemble des cultivars occupe un espacetrès important et sa description représenteun travail considérable. L’identification d’unsous-ensemble de populations représentatifde la diversité de la plante s’avère néces-saire. L’utilisation de caractères morpholo-
1 Ipgri : International Plant Genetic Resources Institute.2 Cogent : Coconut Genetic Resources Network.3 Cgrd : Coconut Genetic Resources Database.
Deux grandsgroupes
de cocotiers se distinguentclairement parleur profil RFLP.L’un correspondaux populationsd’Asie du Sud-Estet du Pacifiqueet a donnénaissance aux Nains.L’autre s’estindividualisédans le sous-continent indienet a étédisséminé par l’Hommevers l’Afrique de l’Ouest et la régionCaraïbe.
Le contenu de cet article a fait l’objet d’un exposéprésenté à l’International symposium on coconutbiotechnology, à Mérida, Yucatán (Mexique), du 1er au 5 décembre 1997.
Retour au menu

2. Grand d’Asie du Sud-Est (Grand Tagnanan) Tall from South East Asia (Tagnanan Tall)
1. Grand du groupe Indo-atlantique(Grand Sri Lanka) Tall from Indo-Atlantic group (Sri Lanka Tall)
4. Nain (Nain Jaune Malais)Dwarf (Malayan Yellow Dwarf)
Plantations, recherche, développement Juil let - Août 1998
234 CO C O T I E R
Aspect général (A) et forme du fruit (B) de quatre cultivars de cocotier General appearance (A) and fruit shape (B) of four coconut cultivars
A
B
A
B
B
A
B
R. B
ou
rdei
x
3. Grand du Pacifique Sud(Grand Karkar) Tall from South Pacific (Karkar Tall)
A
Retour au menu
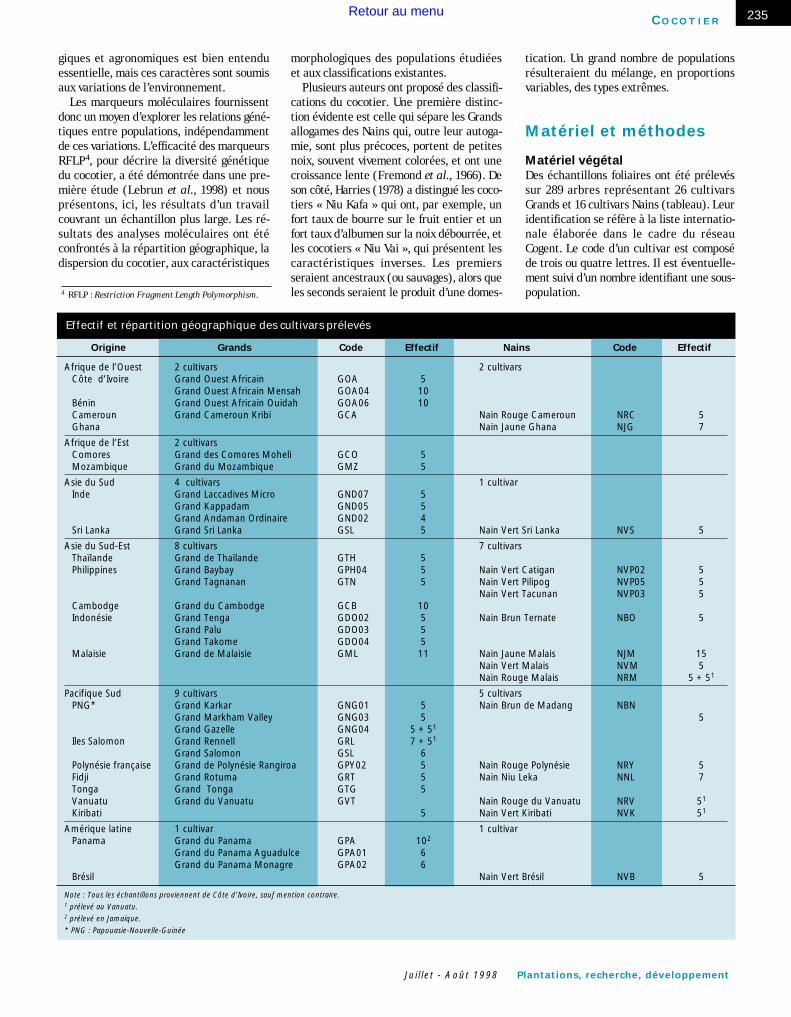
Effectif et répartition géographique des cultivars prélevés
Origine Grands Code Effectif Nains Code Effectif
Afrique de l’Ouest 2 cultivars 2 cultivarsCôte d’Ivoire Grand Ouest Africain GOA 5
Grand Ouest Africain Mensah GOA04 10 Bénin Grand Ouest Africain Ouidah GOA06 10 Cameroun Grand Cameroun Kribi GCA Nain Rouge Cameroun NRC 5Ghana Nain Jaune Ghana NJG 7
Afrique de l’Est 2 cultivarsComores Grand des Comores Moheli GCO 5 Mozambique Grand du Mozambique GMZ 5
Asie du Sud 4 cultivars 1 cultivar Inde Grand Laccadives Micro GND07 5
Grand Kappadam GND05 5 Grand Andaman Ordinaire GND02 4
Sri Lanka Grand Sri Lanka GSL 5 Nain Vert Sri Lanka NVS 5
Asie du Sud-Est 8 cultivars 7 cultivarsThaïlande Grand de Thaïlande GTH 5 Philippines Grand Baybay GPH04 5 Nain Vert Catigan NVP02 5
Grand Tagnanan GTN 5 Nain Vert Pilipog NVP05 5Nain Vert Tacunan NVP03 5
Cambodge Grand du Cambodge GCB 10 Indonésie Grand Tenga GDO02 5 Nain Brun Ternate NBO 5
Grand Palu GDO03 5 Grand Takome GDO04 5
Malaisie Grand de Malaisie GML 11 Nain Jaune Malais NJM 15Nain Vert Malais NVM 5Nain Rouge Malais NRM 5 + 51
Pacifique Sud 9 cultivars 5 cultivarsPNG* Grand Karkar GNG01 5 Nain Brun de Madang NBN
Grand Markham Valley GNG03 5 5 Grand Gazelle GNG04 5 + 51
Iles Salomon Grand Rennell GRL 7 + 51
Grand Salomon GSL 6 Polynésie française Grand de Polynésie Rangiroa GPY02 5 Nain Rouge Polynésie NRY 5Fidji Grand Rotuma GRT 5 Nain Niu Leka NNL 7 Tonga Grand Tonga GTG 5 Vanuatu Grand du Vanuatu GVT Nain Rouge du Vanuatu NRV 51
Kiribati 5 Nain Vert Kiribati NVK 51
Amérique latine 1 cultivar 1 cultivarPanama Grand du Panama GPA 102
Grand du Panama Aguadulce GPA01 6 Grand du Panama Monagre GPA02 6
Brésil Nain Vert Brésil NVB 5
Note : Tous les échantillons proviennent de Côte d’Ivoire, sauf mention contraire.1 prélevé au Vanuatu.2 prélevé en Jamaïque.
* PNG : Papouasie-Nouvelle-Guinée
giques et agronomiques est bien entenduessentielle, mais ces caractères sont soumisaux variations de l’environnement.
Les marqueurs moléculaires fournissentdonc un moyen d’explorer les relations géné-tiques entre populations, indépendammentde ces variations. L’efficacité des marqueursRFLP4, pour décrire la diversité génétiquedu cocotier, a été démontrée dans une pre-mière étude (Lebrun et al., 1998) et nousprésentons, ici, les résultats d’un travail couvrant un échantillon plus large. Les ré-sultats des analyses moléculaires ont étéconfrontés à la répartition géographique, ladispersion du cocotier, aux caractéristiques
morphologiques des populations étudiées et aux classifications existantes.
Plusieurs auteurs ont proposé des classifi-cations du cocotier. Une première distinc-tion évidente est celle qui sépare les Grandsallogames des Nains qui, outre leur autoga-mie, sont plus précoces, portent de petitesnoix, souvent vivement colorées, et ont unecroissance lente (Fremond et al., 1966). Deson côté, Harries (1978) a distingué les coco-tiers « Niu Kafa » qui ont, par exemple, unfort taux de bourre sur le fruit entier et unfort taux d’albumen sur la noix débourrée, etles cocotiers « Niu Vai », qui présentent lescaractéristiques inverses. Les premiersseraient ancestraux (ou sauvages), alors queles seconds seraient le produit d’une domes-
235CO C O T I E R
Juil let - Août 1998 Plantations, recherche, développement
tication. Un grand nombre de populationsrésulteraient du mélange, en proportionsvariables, des types extrêmes.
Matériel et méthodes
Matériel végétalDes échantillons foliaires ont été prélevéssur 289 arbres représentant 26 cultivarsGrands et 16 cultivars Nains (tableau). Leuridentification se réfère à la liste internatio-nale élaborée dans le cadre du réseauCogent. Le code d’un cultivar est composéde trois ou quatre lettres. Il est éventuelle-ment suivi d’un nombre identifiant une sous-population.4 RFLP : Restriction Fragment Length Polymorphism.
Retour au menu

Analyses RFLPPour chaque arbre, l’ADN 5 a été extrait defolioles lyophilisées, prélevées sur la feuillenuméro 1, par la méthode dite « Ctab »adaptée par Hoisington (1992) sur le maïs.Pour chaque échantillon, 10 µg d’ADN ontété digérés par les quatre enzymes de res-triction suivantes : Eco RI, Eco RV, Pst I etBgl II. Chaque digestion a, ensuite, été dépo-sée sur un gel d’agarose à 0,8 % dans du tam-pon TAE. Après migration, l’ADN est trans-féré sur une membrane de Nylon. Les sondessont marquées au P32 grâce à l’utilisation dukit Amersham « Megaprime ». La préhybri-dation ainsi que l’hybridation ont été réali-sées selon le protocole de Hoisington (1992).
Sondes utiliséesCette étude a été réalisée à l’aide de 20 sondes ADNc présentant un profil simpled’hybridation sur le cocotier, parmi lesquelson ne trouve qu’une sonde de cocotier, le reste étant des sondes hétérologues de riz, de palmier à huile et de maïs. Trois sondes de riz proviennent du labora-toire japonais du Rice Genome Program,Niar/Staff, Japan (Kurata et al., 1994), uneautre de l’université de Cornell, Etats-Unis(Causse et al., 1994). La sonde de maïs vientde l’université du Missouri à Colombia,Etats-Unis (Gardiner et al., 1993). J. Traegeret A. Rival, Cirad/Orstom6, Montpellier,France nous ont procuré les 15 sondes depalmiers à huile. Enfin, nous avons aussiutilisé la sonde cytoplasmique Cox 1 de blé(Lejeune et al., 1988).
Analyse des donnéesChaque bande révélée par une combinaisonenzyme–sonde particulière a été codéecomme un marqueur dominant : 10 pour saprésence et 01 pour son absence, ceci afin dedonner le même poids à tous les individus.Nous avons, ainsi, obtenu une matricebinaire bande x individu, point de départ desAfc (analyse factorielle des correspon-dances) (Benzecri, 1973) qui nous ont per-mis de décrire la structuration génétique dugermoplasme. Les analyses multivariablesont été réalisées grâce au logiciel Addad(Addad, 1983).
L’examen des analyses factorielles descorrespondances et les informations géogra-phiques permettent de constituer des
groupes de cultivars relativement homo-gènes. Ainsi, il a été possible d’identifier desmarqueurs spécifiques de chaque groupe.
RésultatsSur les 289 arbres analysés, 60 bandes poly-morphes ont été révélées par les 25 couplesenzyme–sonde et 2 profils cytoplasmiquespar la sonde Cox 1.
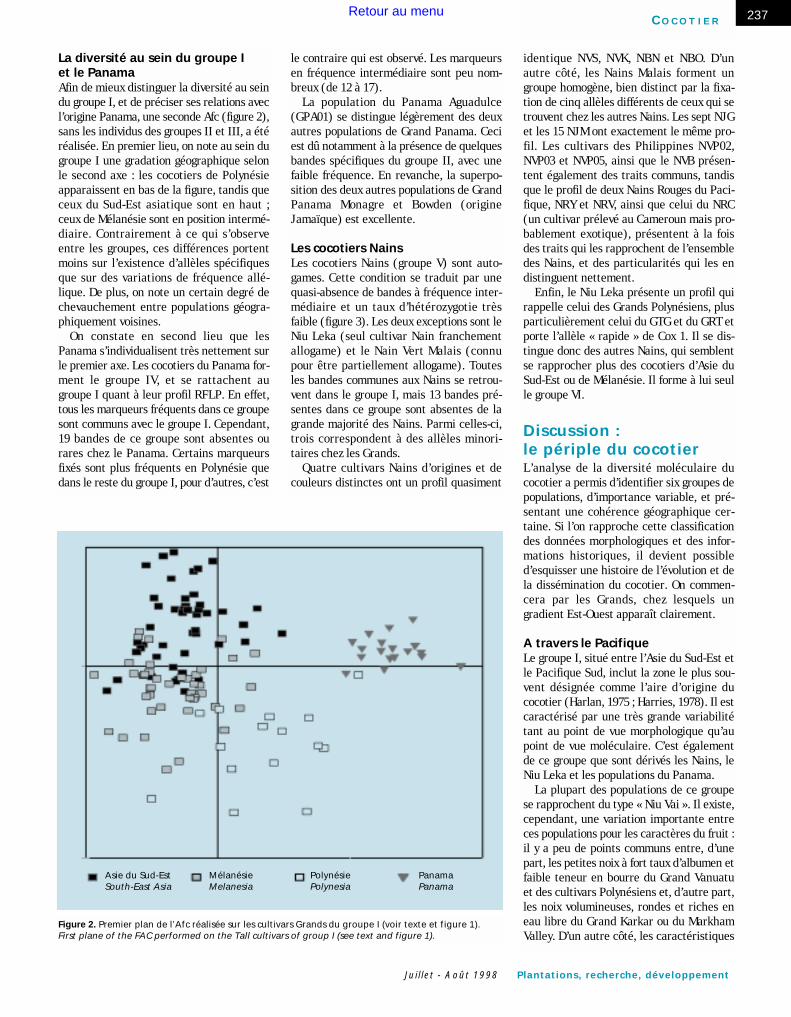
Les deux zones majeures de diversité des cocotiers GrandsOn distingue (figure 1) sur la droite du gra-phique un premier groupe, correspondant àl’ensemble des Grands de l’Asie du Sud-Estau Pacifique Sud (groupe I), auquel se rat-tachent tous les cultivars Nains, et les coco-tiers du Panama. Un autre ensemble, situé àgauche, correspond aux cocotiers d’Inde, duSri Lanka et d’Afrique de l’Ouest (groupeII). Ils sont séparés par un ensemble de troiscultivars, riverains de l’Océan Indien(groupe III).
En termes de marqueurs, le groupe II sedistingue par la présence prédominante dequatre marqueurs qui sont pratiquementabsents du groupe I, et de trois autres, qui
sont partagés avec les seules populationsd’Asie du Sud-Est. Vingt-et-un marqueurssont absents ou à faible fréquence dans legroupe II, alors qu’ils sont bien représentésdans la plupart des cultivars du groupe I.Enfin, le groupe II possède un allèle spéci-fique pour le marqueur cytoplasmique Cox 1.Tous ses membres présentent en effetl’allèle « rapide » à quelques exceptionsprès. Trois individus sur 10 du GOA06, deuxsur cinq du GSL et tous les individus duGND05 portent en effet l’allèle « lent »,caractéristique du groupe I.
Ces deux groupes se distinguent égale-ment par leur degré de polymorphisme : legroupe I présente un plus grand nombre demarqueurs communs aux différents cultivars(42 contre 33) et un plus grand nombre demarqueurs à fréquence intermédiaire (20 à38 dont la fréquence est comprise entre 10 %et 90 % selon le cultivar, contre 11 à 20 dansle groupe II).
Le groupe III présente presque tous lesmarqueurs nucléaires des groupes I et II,c’est donc celui qui regroupe la plus largediversité moléculaire. L’allèle cytoplasmiqueest celui du groupe I (sauf pour deux indivi-dus GMZ).
Plantations, recherche, développement Juil let - Août 1998
236 CO C O T I E R
5 ADN : acide désoxyribonucléique.6 Cirad/Orstom : Centre de coopération internationale enrecherche agronomique pour le développement / Institutfrançais de recherche scientifique pour le développementen coopération.
Afrique de l’OuestWest Africa
Asie du Sud-EstSouth-East Asia
Afrique de l’EstEast Africa
Pacifique SudSouth Pacific
Asie du SudSouth Asia
AmériqueAmerica
Figure 1. Premier plan de l’analyse factorielle des correspondances (Afc) réaliséesur les cultivars Grands. / First plane of the Factorial Analysis of Correspondances (FAC) performed on the Tall cultivars.
0,5
– 0,5
– 1,5 – 1 – 0,5 0 0,5 1
0
1
groupe II
groupe III
groupe I
Retour au menu
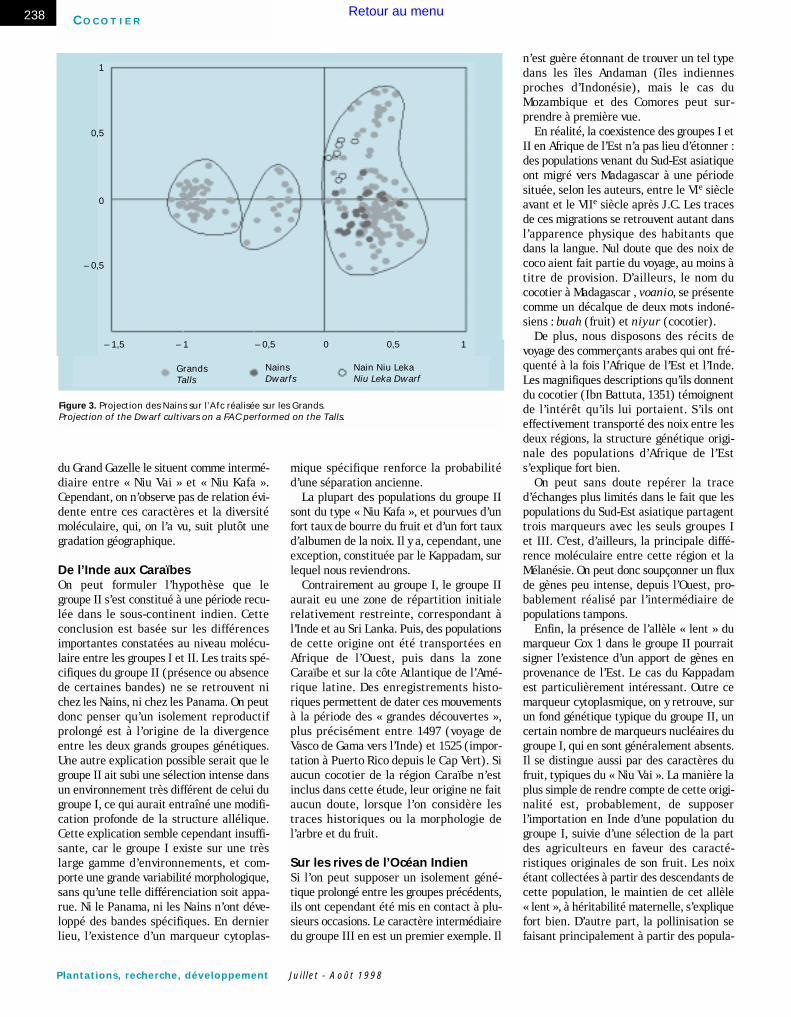
La diversité au sein du groupe I et le PanamaAfin de mieux distinguer la diversité au seindu groupe I, et de préciser ses relations avecl’origine Panama, une seconde Afc (figure 2),sans les individus des groupes II et III, a étéréalisée. En premier lieu, on note au sein dugroupe I une gradation géographique selonle second axe : les cocotiers de Polynésieapparaissent en bas de la figure, tandis queceux du Sud-Est asiatique sont en haut ;ceux de Mélanésie sont en position intermé-diaire. Contrairement à ce qui s’observeentre les groupes, ces différences portentmoins sur l’existence d’allèles spécifiquesque sur des variations de fréquence allé-lique. De plus, on note un certain degré dechevauchement entre populations géogra-phiquement voisines.
On constate en second lieu que lesPanama s’individualisent très nettement surle premier axe. Les cocotiers du Panama for-ment le groupe IV, et se rattachent augroupe I quant à leur profil RFLP. En effet,tous les marqueurs fréquents dans ce groupesont communs avec le groupe I. Cependant,19 bandes de ce groupe sont absentes ourares chez le Panama. Certains marqueursfixés sont plus fréquents en Polynésie quedans le reste du groupe I, pour d’autres, c’est
le contraire qui est observé. Les marqueursen fréquence intermédiaire sont peu nom-breux (de 12 à 17).
La population du Panama Aguadulce(GPA01) se distingue légèrement des deuxautres populations de Grand Panama. Ceciest dû notamment à la présence de quelquesbandes spécifiques du groupe II, avec unefaible fréquence. En revanche, la superpo-sition des deux autres populations de GrandPanama Monagre et Bowden (origineJamaïque) est excellente.
Les cocotiers NainsLes cocotiers Nains (groupe V) sont auto-games. Cette condition se traduit par unequasi-absence de bandes à fréquence inter-médiaire et un taux d’hétérozygotie trèsfaible (figure 3). Les deux exceptions sont leNiu Leka (seul cultivar Nain franchementallogame) et le Nain Vert Malais (connupour être partiellement allogame). Toutesles bandes communes aux Nains se retrou-vent dans le groupe I, mais 13 bandes pré-sentes dans ce groupe sont absentes de lagrande majorité des Nains. Parmi celles-ci,trois correspondent à des allèles minori-taires chez les Grands.
Quatre cultivars Nains d’origines et decouleurs distinctes ont un profil quasiment
identique NVS, NVK, NBN et NBO. D’unautre côté, les Nains Malais forment ungroupe homogène, bien distinct par la fixa-tion de cinq allèles différents de ceux qui setrouvent chez les autres Nains. Les sept NJGet les 15 NJM ont exactement le même pro-fil. Les cultivars des Philippines NVP02,NVP03 et NVP05, ainsi que le NVB présen-tent également des traits communs, tandisque le profil de deux Nains Rouges du Paci-fique, NRY et NRV, ainsi que celui du NRC(un cultivar prélevé au Cameroun mais pro-bablement exotique), présentent à la foisdes traits qui les rapprochent de l’ensemble des Nains, et des particularités qui les endistinguent nettement.
Enfin, le Niu Leka présente un profil quirappelle celui des Grands Polynésiens, plusparticulièrement celui du GTG et du GRT etporte l’allèle « rapide » de Cox 1. Il se dis-tingue donc des autres Nains, qui semblentse rapprocher plus des cocotiers d’Asie duSud-Est ou de Mélanésie. Il forme à lui seulle groupe VI.
Discussion : le périple du cocotier L’analyse de la diversité moléculaire ducocotier a permis d’identifier six groupes depopulations, d’importance variable, et pré-sentant une cohérence géographique cer-taine. Si l’on rapproche cette classificationdes données morphologiques et des infor-mations historiques, il devient possibled’esquisser une histoire de l’évolution et dela dissémination du cocotier. On commen-cera par les Grands, chez lesquels un gradient Est-Ouest apparaît clairement.
A travers le PacifiqueLe groupe I, situé entre l’Asie du Sud-Est etle Pacifique Sud, inclut la zone le plus sou-vent désignée comme l’aire d’origine ducocotier (Harlan, 1975 ; Harries, 1978). Il estcaractérisé par une très grande variabilitétant au point de vue morphologique qu’aupoint de vue moléculaire. C’est égalementde ce groupe que sont dérivés les Nains, leNiu Leka et les populations du Panama.
La plupart des populations de ce groupese rapprochent du type « Niu Vai ». Il existe,cependant, une variation importante entreces populations pour les caractères du fruit :il y a peu de points communs entre, d’unepart, les petites noix à fort taux d’albumen etfaible teneur en bourre du Grand Vanuatuet des cultivars Polynésiens et, d’autre part,les noix volumineuses, rondes et riches eneau libre du Grand Karkar ou du MarkhamValley. D’un autre côté, les caractéristiques
237CO C O T I E R
Juil let - Août 1998 Plantations, recherche, développement
Asie du Sud-EstSouth-East Asia
MélanésieMelanesia
PolynésiePolynesia
PanamaPanama
Figure 2. Premier plan de l’Afc réalisée sur les cultivars Grands du groupe I (voir texte et figure 1). First plane of the FAC performed on the Tall cultivars of group I (see text and figure 1).
Retour au menu
du Grand Gazelle le situent comme intermé-diaire entre « Niu Vai » et « Niu Kafa ».Cependant, on n’observe pas de relation évi-dente entre ces caractères et la diversitémoléculaire, qui, on l’a vu, suit plutôt unegradation géographique.
De l’Inde aux CaraïbesOn peut formuler l’hypothèse que legroupe II s’est constitué à une période recu-lée dans le sous-continent indien. Cetteconclusion est basée sur les différencesimportantes constatées au niveau molécu-laire entre les groupes I et II. Les traits spé-cifiques du groupe II (présence ou absencede certaines bandes) ne se retrouvent nichez les Nains, ni chez les Panama. On peutdonc penser qu’un isolement reproductifprolongé est à l’origine de la divergenceentre les deux grands groupes génétiques.Une autre explication possible serait que legroupe II ait subi une sélection intense dansun environnement très différent de celui dugroupe I, ce qui aurait entraîné une modifi-cation profonde de la structure allélique.Cette explication semble cependant insuffi-sante, car le groupe I existe sur une trèslarge gamme d’environnements, et com-porte une grande variabilité morphologique,sans qu’une telle différenciation soit appa-rue. Ni le Panama, ni les Nains n’ont déve-loppé des bandes spécifiques. En dernierlieu, l’existence d’un marqueur cytoplas-
mique spécifique renforce la probabilitéd’une séparation ancienne.
La plupart des populations du groupe IIsont du type « Niu Kafa », et pourvues d’unfort taux de bourre du fruit et d’un fort tauxd’albumen de la noix. Il y a, cependant, uneexception, constituée par le Kappadam, surlequel nous reviendrons.
Contrairement au groupe I, le groupe IIaurait eu une zone de répartition initialerelativement restreinte, correspondant àl’Inde et au Sri Lanka. Puis, des populationsde cette origine ont été transportées enAfrique de l’Ouest, puis dans la zoneCaraïbe et sur la côte Atlantique de l’Amé-rique latine. Des enregistrements histo-riques permettent de dater ces mouvementsà la période des « grandes découvertes »,plus précisément entre 1497 (voyage deVasco de Gama vers l’Inde) et 1525 (impor-tation à Puerto Rico depuis le Cap Vert). Siaucun cocotier de la région Caraïbe n’estinclus dans cette étude, leur origine ne faitaucun doute, lorsque l’on considère lestraces historiques ou la morphologie del’arbre et du fruit.
Sur les rives de l’Océan IndienSi l’on peut supposer un isolement géné-tique prolongé entre les groupes précédents,ils ont cependant été mis en contact à plu-sieurs occasions. Le caractère intermédiairedu groupe III en est un premier exemple. Il
n’est guère étonnant de trouver un tel typedans les îles Andaman (îles indiennesproches d’Indonésie), mais le cas duMozambique et des Comores peut sur-prendre à première vue.
En réalité, la coexistence des groupes I etII en Afrique de l’Est n’a pas lieu d’étonner :des populations venant du Sud-Est asiatiqueont migré vers Madagascar à une périodesituée, selon les auteurs, entre le VIe siècleavant et le VIIe siècle après J.C. Les tracesde ces migrations se retrouvent autant dansl’apparence physique des habitants quedans la langue. Nul doute que des noix decoco aient fait partie du voyage, au moins àtitre de provision. D’ailleurs, le nom ducocotier à Madagascar , voanio, se présentecomme un décalque de deux mots indoné-siens : buah (fruit) et niyur (cocotier).
De plus, nous disposons des récits devoyage des commerçants arabes qui ont fré-quenté à la fois l’Afrique de l’Est et l’Inde.Les magnifiques descriptions qu’ils donnentdu cocotier (Ibn Battuta, 1351) témoignentde l’intérêt qu’ils lui portaient. S’ils onteffectivement transporté des noix entre lesdeux régions, la structure génétique origi-nale des populations d’Afrique de l’Ests’explique fort bien.
On peut sans doute repérer la traced’échanges plus limités dans le fait que lespopulations du Sud-Est asiatique partagenttrois marqueurs avec les seuls groupes I et III. C’est, d’ailleurs, la principale diffé-rence moléculaire entre cette région et la Mélanésie. On peut donc soupçonner un fluxde gènes peu intense, depuis l’Ouest, pro-bablement réalisé par l’intermédiaire depopulations tampons.
Enfin, la présence de l’allèle « lent » dumarqueur Cox 1 dans le groupe II pourraitsigner l’existence d’un apport de gènes enprovenance de l’Est. Le cas du Kappadamest particulièrement intéressant. Outre cemarqueur cytoplasmique, on y retrouve, surun fond génétique typique du groupe II, uncertain nombre de marqueurs nucléaires dugroupe I, qui en sont généralement absents.Il se distingue aussi par des caractères dufruit, typiques du « Niu Vai ». La manière laplus simple de rendre compte de cette origi-nalité est, probablement, de supposerl’importation en Inde d’une population dugroupe I, suivie d’une sélection de la partdes agriculteurs en faveur des caracté-ristiques originales de son fruit. Les noixétant collectées à partir des descendants decette population, le maintien de cet allèle « lent », à héritabilité maternelle, s’expliquefort bien. D’autre part, la pollinisation sefaisant principalement à partir des popula-
Plantations, recherche, développement Juil let - Août 1998
238 CO C O T I E R
GrandsTalls
NainsDwarfs
Nain Niu LekaNiu Leka Dwarf
Figure 3. Projection des Nains sur l’Afc réalisée sur les Grands. Projection of the Dwarf cultivars on a FAC performed on the Talls.
0,5
– 0,5
– 1,5 – 1 – 0,5 0 0,5 1
0
1
Retour au menu

tions locales, les gènes du groupe II ontpresque totalement supplanté ceux dugroupe I. En résumé, si cette interprétationest correcte, le Kappadam résulte de l’intro-duction dans le groupe I du phénotype « NiuVai » par rétrocroisements successifs.
L’isthme de PanamaL’origine de la population Panama se trouvedans le groupe I, Pacifique-Asie du Sud-Est.Il est difficile de dire exactement à partir dequel matériel de base elle est issue, car elleprésente des traits qui la rapprochent tantôtdes populations polynésiennes, tantôt decelles d’Asie du Sud-Est et de Mélanésie. Lafaible variabilité de cette population est unargument fort à l’encontre de l’origine amé-ricaine du cocotier : cette population a dûtraverser un goulot d’étranglement qui aconsidérablement réduit la taille efficacede la population.
On peut alors formuler deux hypothèsesentre lesquelles il est difficile de trancher.Dans la première, le cultivar Panama seraitassez représentatif des populations préco-lombiennes reconnues par Oviedo en 1530.Dans ce cas, les importations ultérieures duPacifique, en particulier Salomon et Philip-pines (Zizumbo-Villareal, 1996), auraientpeu modifié leur structure génétique. Dansla seconde, ces populations post-colom-biennes joueraient un rôle prépondérant,expliquant l’aspect composite du profilRFLP des populations du Panama. Pourrendre compte de la pauvreté alléliqueobservée, il faudrait alors faire intervenirun goulet d’étranglement, qui pourrait êtredû à une pression de sélection intense desmaladies, des ravageurs, des cyclones ou àun effet fondateur.
C’est en Amérique que se referme lepériple du cocotier autour de la ceinturetropicale. Au bout d’un long voyage, lesreprésentants des deux branches majeuresde son évolution sont parvenus sur chacunede ses rives, et c’est là qu’elles se réunis-sent à nouveau. La population de GrandPanama Aguadulce (GPA01) en est uneillustration : son profil RFLP est très prochede celui qu’on peut attendre de l’introgres-sion d’environ 10 % de gènes du groupe II(représenté dans la région par les popula-tions dites « Atlantico » ou « Tres Picos »)dans des populations « Redondo » similairesà celles de Monagre ou de Bowden.
La domestication du cocotier NainToute hypothèse sur l’origine et la diversifi-cation des cocotiers Nains doit tenir comptedes ressemblances frappantes qui existent
entre leurs profils RFLP. Il est très pro-bable, en effet, qu’ils soient apparus en uneseule fois, ou du moins à l’intérieur d’uneseule population. La présence de 13 mar-queurs communs à presque tous les Nains,alors même que trois d’entre eux sont mino-ritaires chez les Grands semble plaider pourune apparition unique.
Les cocotiers Nains se trouvent rarementen larges peuplements homogènes (si l’onexcepte la réalisation, au cours de ce siècle,de quelques plantations industrielles), maison en retrouve souvent à proximité immé-diate des habitations. Il est donc permis dedécrire le nanisme comme l’étape ultime dela domestication du cocotier, dont l’élémentdéterminant aurait été l’apparition del’autogamie et, par suite, la possibilité dereproduction conforme d’un type attrayant.
Les caractères recherchés compren-draient la croissance lente, la précocité ou,chez certains d’entre eux, l’excellente qua-lité gustative de l’eau des noix immatures.En outre, la couleur souvent vive des fruits,sa forme caractéristique et le port de l’arbreconstituent des moyens efficaces d’identifi-cation variétale.
La première conséquence de l’autogamiea été la fixation aléatoire d’un allèle àchaque locus (généralement — mais pastoujours — le plus fréquent), et la conser-vation, par la suite, de l’essentiel de sastructure génétique. On peut penser quel’histoire ultérieure des Nains est constituéede longues périodes de fixité relative, entre-coupées d’échanges de gènes sporadiquesavec les populations de Grands locaux oud’autres Nains, qui ont contribué à différen-cier les cultivars. En réalité, le croisementd’un Nain et d’un Grand donne un hybrideF1 dont les caractéristiques diffèrent net-tement du Nain. La plupart des descendantsde tels croisements ont toutes les chancesde se fondre dans la masse des populationsGrand environnantes. Seul le croisementd’un Nain par un hybride Nain x Grand oupar un autre Nain a des chances de fournirun cocotier de type Nain, et de contribuer,ainsi, à la diversification des populations deNains. Ceci implique, d’une part que de telsévénements sont relativement rares, d’autrepart, que chacun d’entre eux se traduit parun nombre limité de substitutions allé-liques.
Dans ces conditions, il semble possiblede proposer un schéma d’évolution desNains à partir de la population originellehypothétique :• la branche correspondant aux Nains Phi-
lippins et Malais se serait individualiséeassez tôt, avec deux allèles communs. Les
Malais forment un groupe homogène,ayant trois autres allèles en commun etne se sont sans doute individualisés quetardivement ;
• une structure proche de celle de la popu-lation initiale se serait maintenue, dansun ensemble de cultivars prélevés dansdes régions très diverses : NVS, NVK,NBO, NBN (tableau) ;
• au contraire, une divergence plus mar-quée s’observe chez les Nains Rouges du Pacifique : NRY, NRV, auxquelss’ajoute le NRC qui, bien que prélevé au Cameroun, pourrait bien avoir une origine Pacifique.Bien que les cas malais et philippins
témoignent d’une structure géographiquede la variation des Nains, celle-ci est beau-coup moins évidente que chez les Grands.Contrairement à ce qui se passe chez lesGrands, un cultivar Nain importé dans unenouvelle région tend à rester génétiquementisolé des populations locales. Ceci expliqueque des génotypes semblables se retrouventdans des zones éloignées, ainsi le NVB pré-sente un profil voisin de celui des cultivarsphilippins, et les cultivars apparentés auNVS sont dispersés dans des régions éloi-gnées. En ce sens, les Nains témoignent de l’intensité des échanges qui ont eu lieu à l’intérieur de la zone de culture du cocotier.
Il reste à situer la région d’origine descocotiers Nains. Il n’est pas possible de lefaire très précisément, car la fixation aléa-toire des allèles due à l’homozygotie a puentraîner dès le début une modification pro-fonde de la structure génétique de la popu-lation initiale. Les cultivars Grands qui s’enrapprochent le plus se trouvent dans unerégion qui va du Sud-Est asiatique à laPapouasie-Nouvelle-Guinée.
Un type original : le Niu LekaBien qu’il ait une croissance lente commeles Nains, le Niu Leka constitue une variététrès particulière, par son allogamie, sonaspect et le fait qu’un hybride Niu Leka x Grand ressemble à un Niu Leka. Son profilRFLP confirme sa singularité. Clairementdistinct de celui des Nains, il est, aucontraire, extrêmement similaire à celui des Grands du Pacifique, le plus procheétant le GTG, suivi du GRT, ce qui corres-pond précisément à la région d’origine duNiu Leka. En revanche, aucun élément dis-ponible ne permet de rendre compte de laprésence de l’allèle « rapide » de Cox 1, quine se retrouve chez aucun autre cultivar dela région.
239CO C O T I E R
Juil let - Août 1998 Plantations, recherche, développement
Retour au menu

Conclusion-perspectives
L’exploitation des données RFLP n’est pasachevée, et il conviendrait de compléterl’échantillonnage par un petit nombre decultivars, pour combler certains vides, oupour vérifier quelques hypothèses. Ces culti-vars, au nombre d’une douzaine, seraient desGrands à collecter en Amérique latine, enMicronésie, en Polynésie (Niu Kafa), enAfrique de l’Est, ainsi que quelques cultivarsNains ou semi-Nains tels que King Coconut.Pareillement, la recherche d’un certainnombre de marqueurs supplémentairespourrait être utile, notamment pour distin-guer certains Nains. Cependant, les résul-tats disponibles à ce jour fournissent uneimage relativement précise de la diversitédes cocotiers, des voies suivies lors de leurdissémination et de leur évolution.
Dans une certaine mesure, le schéma pro-posé s’accorde avec celui d’Harries (1978),notamment en ce qui concerne la situationobservée en Amérique latine et dans lesCaraïbes. En revanche, le rôle joué par ladomestication et par les isolements géogra-phiques y est nettement différent. Ainsi,malgré un certain degré de ressemblance,une origine unique n’est pas démontrée pour
les « Niu Kafa » du groupe I et le groupe II.Les données suggèrent fortement une ori-gine unique des Nains, et ils apparaissentcomme l’étape ultime de la domesticationdu cocotier.
Dans certaines situations, le polymor-phisme RFLP permet d’effectuer un dia-gnostic extrêmement précis d’identificationvariétale. Il a même été possible de détecter,dans certaines populations, la présence degènes exogènes, et d’en expliquer la causeprobable. Compte tenu des effectifs très res-treints utilisés, cette technique s’est révé-lée très puissante.
Elle doit apporter une contribution impor-tante dans la gestion des ressources géné-tiques, notamment pour identifier les redon-dances, ou le degré de représentativité decertains échantillons. Lors du rajeunisse-ment des collections, on peut, par exemple,se demander s’il vaut mieux reproduire lapopulation originale ou un sous-échantillonissu d’une sélection massale. L’étude del’évolution des fréquences alléliques per-mettra d’y répondre. En sélection, on dis-pose maintenant d’un moyen de limiter lenombre de combinaisons à tester, en se fon-dant à la fois sur les similarités relevées aux niveaux phénotypique et moléculaire.
Une étude de l’hétérosis observée chez leshybrides interpopulations en fonction de ladistance génétique reste à faire.
L’étude des populations de la côte Paci-fique d’Amérique latine, en relation avec latolérance au jaunissement mortel, noussemble une des applications importantespossibles. Les trois populations de GrandPanama étudiées paraissent issues d’unepopulation initiale de petite taille. Ainsi,l’étude, sur de plus larges échantillonnages,de la diversité et de la dynamique des popu-lations chez le Grand Pacifique permettraitde caractériser plus précisément la naturedes apports initiaux, et d’identifier avec plusde certitude les populations susceptibles detransmettre une tolérance à la maladie. ■
Plantations, recherche, développement Juil let - Août 1998
240 CO C O T I E R
ADDAD, 1983. Manuel de référence. Paris, France.Association pour le développement et la diffu-sion de l’analyse des données, 250 p.
BENZECRI J.P., 1973. L’analyse des données. TomeII. Paris, France, Dunod, 619 p.
CAUSSE M.A., FULTON T.M., CHO Y.G., AHN S.N., CHUN-WONGSE J., WU K., XIAO J., YU Z., RONALD P.C.,HARRINGTON S.E., SECOND G., MCCOUCH S.R.,TANKSLEY S., 1994. Saturated molecular map ofthe rice genome based on an interspecific back-cross population. Genetics 138 : 1251-1274.
FREMOND Y., ZILLER R., NUCÉ DE LAMOTHE M. de,1966. Le cocotier. Paris, France, Maisonneuveet Larose, coll. Techniques agricoles et pro-ductions tropicales, 267 p.
GARDINER J.M., COE E.H., MELIA-HANCOCK S., HOISINGTON D.A., CHAO S., 1993. Development ofa core RFLP map in maize using an immortali-zed F2 population. Genetics 134 : 917-930.
HARLAN J.R., 1975. Crops and Man. AmericanSociety of Agronomy, Madison, Wisconsin.
Bibliographie / References
HARRIES H.C., 1978. The evolution, disseminationand classification of Cocos nucifera L. Bot. Rev.44 : 265-320.
HOISINGTON D., 1992. Laboratory protocols. CIM-MYT applied molecular genetics laboratory.Mexico, Mexique, CIMMYT, 44 p.
IBN BATTUTA, 1351. Voyages et périples. In : Lesvoyageurs arabes. Collection “La Pléiade”, Gal-limard, France. 1408 p.
KURATA N., NAGAMURA Y., YAMAMOTO K. , HARU-SHIMA Y. , SUE N. , WU J., ANTONIO B.A., SHOMURA
A., SHIMIZU T., LIN S.Y., INOUE T., FUKUDA A., SHIMANO T., KUBOKI Y., TOYAMA T., MIYAMOTO Y.,KIRIHARA T., HAYASAKA K., MIYAO A., MONNA L.,ZHONG H.S., TAMURA Y., WANG Z.X., MOMMA T.,UMEHARA Y., YANO M., SASAKI T., MINOBE Y., 1994.A 300 kilobase interval genetic map of riceincluding 883 expressed sequences. Nat. Genet.8 : 365-372.
LEBRUN P., N’CHO Y.P., SEGUIN M., GRIVET L., BAUDOUIN L., 1998. Genetic diversity in coconut
(Cocos nucifera L.) Revealed by restrictionfragment length polymorphism (RFLP) markers. Euphytica 101 : 103-108.
LEJEUNE B., QUETIER F., JUBIER M.F., FALCONET D.,RODE A., HARTMANN C., 1988. Le génome mitochondrial des plantes supérieures : orga-nisation moléculaire et expression. Bull. Soc.Bot. Fr. 135 : 49-55.
NUCÉ DE LAMOTHE M. de, BÉNARD G., 1985. L’hybridede cocotier PB-121 (ou Mawa) (NJM x GOA).Oléagineux 40 (5) : 261-266.
WHITEHEAD R.A., 1968. Selecting and breedingcoconuts palms (Cocos nucifera L.) resistant tolethal yellowing disease. A review of recentwork in Jamaica. Euphytica 17 : 81-101.
ZIZUMBO-VILLAREAL D., 1996. History of coconut(Cocos nucifera L.) in Mexico: 1539-1810.Genet. Resour. Crop Evol. 43: 505-515.
Remerciements
Cette étude n’aurait pas été possible sans lacollaboration des stations de recherche quicollectent et entretiennent le matérielgénétique étudié ici. Nous remercions les directeurs ainsi que les équipes de l’Idefor-Dpo (Côte d’Ivoire), du Carfv(Vanuatu) et du Cib (Jamaïque) qui nousont approvisionné en matériel végétal.
Retour au menu

The geographical spread and diversity of thecoconut palm in terms of growth habit and fruitshape and colour (photos) have led to theidentification of a large number of varieties. Forinstance, the international list drawn up forIPGRI1 contains over 300 names of cultivarscorresponding to traditionally grown populationsor those improved by mass selection. Althoughsome of them undoubtedly represent poorlydefined or even extinct populations, almost 60 have been exchanged internationally byresearch stations over the past 50 years. Thisreflects breeders’ interest in collecting and usingcoconut germplasm. As a result, 17 countries inthe COGENT2 network that have contributed to the CGRD3 database now have over 900 acces-sions, and have undertaken to provide a prioritylist of descriptors.
Germplasm in fact plays a key role in breedingprogrammes, since it is a source of variation thatcan be used by selection. Moreover, creatingbetween-population hybrids has in some casesrevealed substantial heterosis (Nucé de Lamotheand Bénard, 1985). Lastly, germplasm is a sourceof variability enabling a search for characters ofadaptation to soil and climatic conditions as wellas pests and diseases (Whitehead, 1968).
Although the wealth of these coconut collec-tions is a considerable asset for breeding, theirvery size makes it essential to draw up a soundstrategy for their use. The number of possiblecombinations between cultivars far exceeds thepossibilities of the worldwide breeding structure.Conserving and describing all the cultivars ishighly labour and land-intensive, and it isnecessary to identify a sub-set of populationsrepresentative of the diversity of the plant. It isobviously essential to use morphological andagronomic characters, but these characters aresubject to environmental variations.
The spread and domestication of the coconut palm in the light of RFLP markers
Lebrun P., Grivet L., Baudouin L.CIRAD, BP 5035, 34032 Montpellier Cedex 1, France
Two main groups of coconut palms can be distinguished clearly from their RFLP profile. One corresponds topopulations from Southeast Asia and the Pacific and gave rise to Dwarfs. The other developed on the Indiansubcontinent and was disseminated by man to West Africa and the Caribbean.
Molecular markers are a way of exploring thegenetic relations between populations, irrespec-tive of these disturbances. The efficacy of RFLP4
was demonstrated in an initial study (Lebrun et al., 1998), and we shall discuss here the results of a study covering a wider range of coconutdiversity. We compared the results of molecularanalyses with what is known of the geograph-ical distribution and spread of coconut, themorphological characteristics of the populationsstudied and existing classifications.
Several authors have attempted to proposecoconut classifications. An initial clear distinctioncan be drawn between the allogamous Talls andDwarfs, which besides being allogamous are alsomore precocious, bear small, often brightlycoloured nuts and grow slowly (Fremond et al.,1966). For his part, Harries (1978) proposeddistinguishing between “Niu Kafa” palms, whichamongst other things have a high husk: whole nutratio and meat:husked nut ratio, and “Niu Vai”palms, which have the opposite characteristics.The former are apparently ancestral (or wild),whilst the latter were apparently produced bydomestication. Many populations are in fact amixture, in varying proportions, of extreme types.
Material and methods
Planting materialLeaf samples were taken from 289 palms repre-senting 26 Tall and 16 Dwarf cultivars or subpopulations. These cultivars and their geograph-ical origins are listed in table. Their identificationrefers to the international list drawn up withinthe COGENT network. The cultivar code com-prises three or four letters, sometimes followedby a figure identifying the sub-population.
RFLP analysisFor each palm, the DNA5 was extracted from freshleaflets of frond number one following the CTABmethod developed by Hoisington (1992) on maize.Blots were prepared using 10 µg of DNA for
each plant after digestion with a restrictionenzyme. The restriction fragments were separatedby electrophoresis on 0.8% agarose gels in TAEbuffer and transferred onto Nylon membranes.Probes were labelled with P32 using theAmersham Megaprime commercial kit. Pre-hybridization and hybridization were performedaccording to Hoisington’s protocol (1992). Theblots were washed four times in 0.1 SSC, 0.1 SDSat 65˚C.
Sources of RFLP probesTwenty nuclear cDNA probes selected for theirsimple hybridization pattern on coconut wereused in this study. One was an anonymous cDNAfrom coconut and the others were heterologousprobes from rice, maize and oil palm. Three riceprobes were provided by the Rice GenomeResearch Program, NIAR/STAFF, Japan (Kurataet al, 1994) and one by Cornell University, USA (Causse et al., 1994). One maize probe wasprovided by the University of Missouri, Colombia,USA (Gardiner et al., 1993) and the 15 oil palmprobes were provided by I. Traeger and A. Rival,CIRAD/ORSTOM6, Montpellier, France.
One cytoplasmic DNA clone from wheat, Cox 1,was also used as a probe (Lejeune et al., 1988).
The polymorphism of each probe was revealedon one to four restriction enzymes among EcoRI,EcoRV, BglII and SstI.
Data analysisEach polymorphic band revealed by a givenprobe/enzyme combination was scored as 10 forpresence and 01 for absence among individualsto give each individual the same importance. Thisproduced a band x individual binary matrix.Factorial analyses of correspondence (Benzecri,1973) were performed on several samples ofindividuals to describe the genetic structuring ofthe germplasm. Computation was performedusing the Addad software (Addad, 1983).
241CO C O N U T
Juil let - Août 1998 Plantations, recherche, développement
The content of this article was covered in a paper presentedat the International symposium on coconut biotechnology,held in Mérida, Yucatán, Mexico, from 1st to 5th December1997.1 IPGRI: International Plant Genetic Resources Institute.2 COGENT: Coconut Genetic Resources Network.3 CGRD: Coconut Genetic Resources Database.
4 RFLP: Restriction Fragment Length Polymorphism.5 DNA: Desoxyribonucleic acid.
6 CIRAD/ORSTOM: Centre de coopération internationale enrecherche agronomique pour le développement. / Institutfrançais de recherche scientifique pour le développementen coopération.
Retour au menu
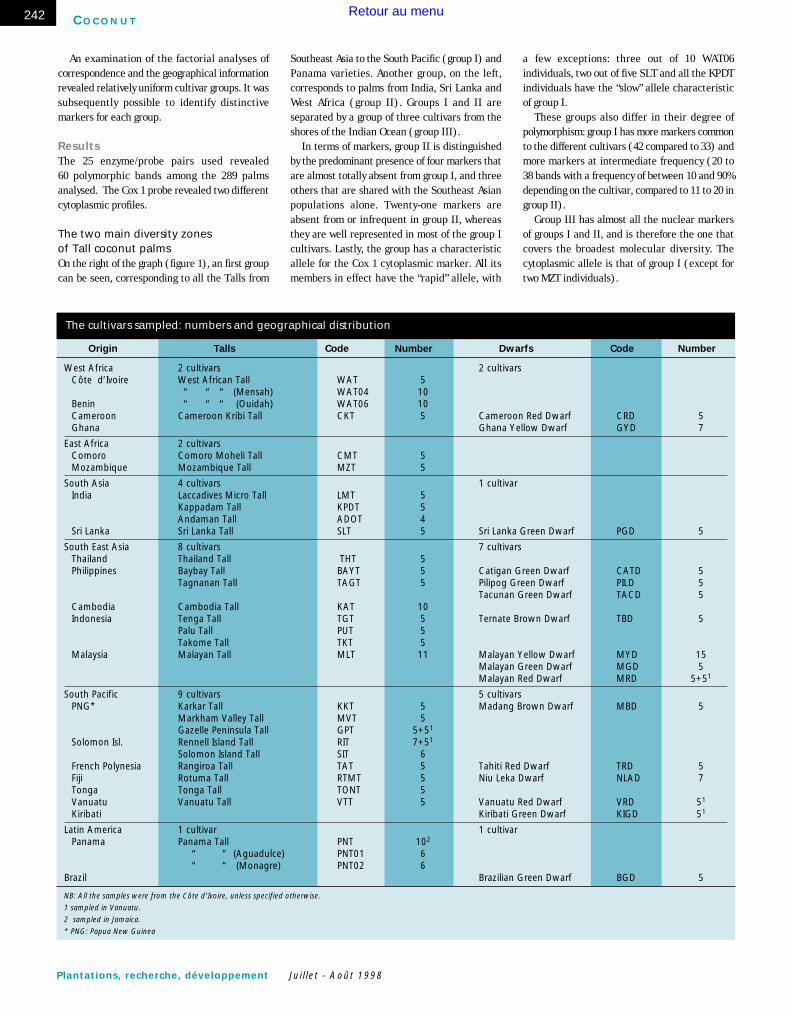
Origin Talls Code Number Dwarfs Code Number
West Africa 2 cultivars 2 cultivarsCôte d’Ivoire West African Tall WAT 5
“ “ “ (Mensah) WAT04 10Benin “ “ “ (Ouidah) WAT06 10Cameroon Cameroon Kribi Tall CKT 5 Cameroon Red Dwarf CRD 5Ghana Ghana Yellow Dwarf GYD 7
East Africa 2 cultivarsComoro Comoro Moheli Tall CMT 5Mozambique Mozambique Tall MZT 5
South Asia 4 cultivars 1 cultivar India Laccadives Micro Tall LMT 5
Kappadam Tall KPDT 5 Andaman Tall ADOT 4
Sri Lanka Sri Lanka Tall SLT 5 Sri Lanka Green Dwarf PGD 5
South East Asia 8 cultivars 7 cultivarsThailand Thailand Tall THT 5Philippines Baybay Tall BAYT 5 Catigan Green Dwarf CATD 5
Tagnanan Tall TAGT 5 Pilipog Green Dwarf PILD 5Tacunan Green Dwarf TACD 5
Cambodia Cambodia Tall KAT 10Indonesia Tenga Tall TGT 5 Ternate Brown Dwarf TBD 5
Palu Tall PUT 5 Takome Tall TKT 5
Malaysia Malayan Tall MLT 11 Malayan Yellow Dwarf MYD 15Malayan Green Dwarf MGD 5Malayan Red Dwarf MRD 5+51
South Pacific 9 cultivars 5 cultivarsPNG* Karkar Tall KKT 5 Madang Brown Dwarf MBD 5
Markham Valley Tall MVT 5Gazelle Peninsula Tall GPT 5+51
Solomon Isl. Rennell Island Tall RIT 7+51
Solomon Island Tall SIT 6French Polynesia Rangiroa Tall TAT 5 Tahiti Red Dwarf TRD 5Fiji Rotuma Tall RTMT 5 Niu Leka Dwarf NLAD 7Tonga Tonga Tall TONT 5Vanuatu Vanuatu Tall VTT 5 Vanuatu Red Dwarf VRD 51
Kiribati Kiribati Green Dwarf KIGD 51
Latin America 1 cultivar 1 cultivarPanama Panama Tall PNT 102
“ “ (Aguadulce) PNT01 6“ “ (Monagre) PNT02 6
Brazil Brazilian Green Dwarf BGD 5
NB: All the samples were from the Côte d’Ivoire, unless specified otherwise.
1 sampled in Vanuatu.
2 sampled in Jamaica.
* PNG: Papua New Guinea
Southeast Asia to the South Pacific (group I) andPanama varieties. Another group, on the left,corresponds to palms from India, Sri Lanka andWest Africa (group II). Groups I and II areseparated by a group of three cultivars from theshores of the Indian Ocean (group III).
In terms of markers, group II is distinguishedby the predominant presence of four markers thatare almost totally absent from group I, and threeothers that are shared with the Southeast Asianpopulations alone. Twenty-one markers areabsent from or infrequent in group II, whereasthey are well represented in most of the group Icultivars. Lastly, the group has a characteristicallele for the Cox 1 cytoplasmic marker. All itsmembers in effect have the “rapid” allele, with
a few exceptions: three out of 10 WAT06individuals, two out of five SLT and all the KPDTindividuals have the “slow” allele characteristicof group I.
These groups also differ in their degree ofpolymorphism: group I has more markers commonto the different cultivars (42 compared to 33) andmore markers at intermediate frequency (20 to38 bands with a frequency of between 10 and 90%depending on the cultivar, compared to 11 to 20 ingroup II).
Group III has almost all the nuclear markersof groups I and II, and is therefore the one thatcovers the broadest molecular diversity. Thecytoplasmic allele is that of group I (except fortwo MZT individuals).
Plantations, recherche, développement Juil let - Août 1998
242
An examination of the factorial analyses ofcorrespondence and the geographical informationrevealed relatively uniform cultivar groups. It wassubsequently possible to identify distinctivemarkers for each group.
ResultsThe 25 enzyme/probe pairs used revealed 60 polymorphic bands among the 289 palmsanalysed. The Cox 1 probe revealed two differentcytoplasmic profiles.
The two main diversity zones of Tall coconut palmsOn the right of the graph (figure 1), an first groupcan be seen, corresponding to all the Talls from
CO C O N U T
The cultivars sampled: numbers and geographical distribution
Retour au menu

Diversity within group I and PanamaIn order to gain a clearer picture of the diversitywithin group I and its relations with the Panamaorigin, a second FAC (figure 2) was carried out.Firstly, there is a geographical gradation alongthe y axis: Polynesian cultivars are at the bottomof the figure and Southeast Asian cultivars at thetop. Unlike the differences seen between groups,these differences relate less to the existence ofspecific alleles than to variations in frequency.Moreover, there is a degree of overlap betweengeographically neighbouring populations.
Secondly, the Panama cultivars are clearlymarked out along the x axis. The Panama popu-lations make up group IV, linked to group I bytheir RFLP profile. In fact, all the frequentmarkers in this group are shared with group I.However, 19 group I bands are not found in Panama cultivars. Some fixed markers are morefrequent in Polynesia than in the rest of group I,but for others, the opposite is the case. There are few intermediate frequency markers (from 12 to 17).
Population PNT01 (Aguadulce) differs slightlyfrom the other two. In particular, this is due to the existence of some group II-specific bands,with a low frequency. On the other hand, thesuperimposition of the Monagre and Bowdenpopulations (Jamaica) is excellent.
Dwarf cultivarsDwarfs cultivars (group V) are clearly related togroup I (figure 3). Their autogamy results in thealmost total absence of intermediate frequencybands and a very low heterozygoty rate. The twoexceptions are the Niu Leka (the only trulyallogamous Dwarf cultivar) and the MalayanGreen Dwarf (known to be partly allogamous).All the common Dwarf bands are found in group I,but 13 of the bands in this group are not foundin the vast majority of Dwarfs. Of these, threecorrespond to minority alleles in Talls.
Four Dwarf cultivars of different origins andcolours have an almost identical banding pattern:PGD, KID, MBD and TBD. On the other hand, theMalayan Dwarfs form a uniform group, clearlydistinguished by the fixation of five alleles diffe-rent from those found in other Dwarfs. The sevenGYD and 15 MYD have exactly the same bandingpattern. The Philippine cultivars CATD, TACDand PILD, and the BGD also have shared traits,whilst the banding patterns of two Pacific redDwarfs, TRD and VRD, and that of CRD (acultivar collected in Cameroon but probablyexotic), have both traits making them similar toDwarfs as a whole and specificities that markthem out clearly.
Lastly, the NLAD has a banding pattern similarto that of the Polynesian Talls, particularly TONTand RTMT, and bears the “rapid” Cox 1 allele. It
therefore differs from the other Dwarfs, whichseem to be more similar to Southeast Asian orMelanesian populations. On its own, it makes upgroup VI.
Discussion: the route taken by the coconut palmAn analysis of the genetic diversity of the coco-nut palm reveals six population groups of varyingsizes, with a degree of geographical coherence.If this classification is compared with morpho-logical data and historical information, it ispossible to trace the history of the evolution and spread of the coconut palm. We shall startwith Talls, for which there is a clear East-Westgradient.
Throughout the PacificGroup I, between Southeast Asia and the SouthPacific, includes the zone most often reputed tobe the origin of the coconut palm (Harlan, 1975;Harries, 1978). It is characterized by substantialvariability, both morphological and molecular. It is also from this group that the Dwarfs, NiuLeka and Panama populations are derived.
Most of the populations in this group aresimilar to the “Niu Vai” type. However, there issubstantial variation between these popula-tionswith regard to fruit characters: the small nutswith a high meat percentage and low huskpercentage produced by the VTT and Polynesiancultivars represented in this study have little in common with the large, round nuts with a high free water content produced by the KKT and MVT. The characteristics of the GPT place it somewhere between “Niu Vai” and “Niu Kafa”.However, there is no clear link between thesecharacters and molecular diversity which, asalready seen, follows a geographical gradation.
From India to the CaribbeanIt can be suggested that group II developed in the distant past in the Indian sub-continent. This conclusion is based on the extent of themolecular differences observed between groups Iand II. The group II-specific traits (existence or absence of certain bands) are not found ineither Dwarfs or Panama populations. We cantherefore assume that extended reproductiveisolation is behind the divergence between thetwo groups. Another possible reason is that groupII may have undergone intense selection in anenvironment very different from that of group I,which caused a significant modification of itsallelic structure. However, this explanation seemsto be insufficient, as group I exists in a wide rangeof environments and includes substantial morph-ological variability, with no such differentiation.Neither the Panama nor the Dwarfs havedeveloped specific bands. Lastly, the existence of a specific cytoplasmic marker backs up the
probability of separation some considerable timeago.
Most of the group II populations are “Niu Kafa”type, with a high husk to fruit and meat to nutratio. However, there is one exception, theKappadam, which we shall come back to.
Unlike group I, group II apparently had arelatively limited initial distribution, corres-ponding to India and Sri Lanka. Populations ofthis origin were subsequently transported to WestAfrica and then the Caribbean and the Atlanticcoast of Latin America. Historical records putthese movements at the time of the “greatdiscoveries”, more precisely between 1497 (Vascode Gama’s voyage to India) and 1525 (importsinto Puerto Rico from Cape Verde). Although thisstudy did not include any Caribbean cultivars,their origin is beyond doubt, considering thehistory of the species or the morphology of thepalm and its fruit.
On the shores of the Indian OceanAlthough we can assume that there was extendedgenetic isolation of the previous groups, they did come into contact on several occasions. The intermediate character group III is the first example of this. It is hardly surprising to find such a type in the Andaman Islands (Indianislands near Indonesia), but the case of theMozambique and Comoro is initially moreunexpected.
In reality, the co-existence of groups I and II inEast Africa is not surprising: South-East Asianpopulations migrated to Madagascar some timebetween the sixth century BC and seventhcentury AD, depending on which author onereads. The traces of these migrations are foundas much in the physical appearance of theinhabitants as in their language. There is nodoubt that they took coconuts with them, at leastas food on the trip. In any event, it is a fact thatthe name of the coconut palm in Madagascar —voanio— is a combination of two Indonesianwords: buah (fruit) and niyur (coconut palm).Moreover, there are accounts written by Arabtraders working in both East Africa and India.Their magnificent descriptions of the coconutpalm (Ibn Battuta, 1351) reveal their interest init. If they did transport nuts between the tworegions, the original genetic structure of the EastAfrican populations is easily explained.
Signs of more limited exchanges could be seenin the fact that the Southeast Asian populationsshare three markers with groups I and III alone.Moreover, this is the main molecular differencebetween this region and Melanesia. We cantherefore suspect a very slight influx of genes,from the West, probably via buffer populations.
Lastly, the existence of the “slow” allele of the Cox 1 marker in group II could be the sign
243CO C O N U T
Juil let - Août 1998 Plantations, recherche, développement
Retour au menu

of an influx from the East. The case of theKappadam is particularly interesting. Besides thiscytoplasmic marker, on a genetic base typical ofgroup II, there are also some nuclear markers,which are not generally found in group II. It isalso distinguished by fruit characters typical of“Niu Vai”. The easiest way of explaining thisoriginality is probably to assume that a group Ipopulation was imported to India, followed by breeding on the part of growers in favour ofthe original characteristics of its fruit. As the nutswere collected from progenies of this population,the maintenance of the “slow” allele is easilyexplained. Moreover, as pollination is primarilyfrom local populations, the genes of group II havealmost entirely supplanted those of group I. Inshort, if this interpretation is correct, theKappadam results from the introduction intogroup I of the “Niu Vai” phenotype by successivebackcrossing.
The Isthmus of PanamaThe origin of the Panama population lies in groupI. It is difficult to say exactly from which basicmaterial, as the population has traits making itsimilar to both Polynesian and Southeast Asianor Melanesian populations. The low variability ofthis population is a strong argument in favour ofthe American origin of the coconut palm: thispopulation must have been through a period ofisolation, with a limited number of effectiveindividuals.
There are two possible explanations, and it is hard to choose between them: in the first, the Panama cultivar would seem to be quiterepresentative of the pre-Colombian populationsrecognized by Oviedo in 1530. In this case,subsequent imports from the Pacific (particularlythe Solomon Islands and the Philippines,(Zizumbo-Villareal, 1996) would only slightlyhave modified their genetic structure. In thesecond, these post-Colombian populations couldhave played a predominant role, which wouldexplain the composite appearance of the RFLPbanding pattern of Panama populations. Toexplain the allelic poverty observed, there wouldhave to have been a bottleneck, which could beput down to intense breeding for disease, pest orcyclone resistance.
It is in the Americas that the coconut palm’sspread around the Tropics came to an end. Aftera long voyage, the representatives of the twomajor branches of its evolution arrived on eachof the continent’s shores and met up again. TheAguadulce population (PNT01) is proof of this:its RFLP banding pattern is very similar to thatwhich could be expected from introgressingaround 10% of the genes of group II (representedin the region by so-called “Atlantico” or “TresPicos” populations) into “Redondo” populationssimilar to Monagre or Bowden.
Domestication of the Dwarf coconut palmAny hypotheses as to the origin and spread ofDwarfs has to take account of the strikingresemblances between the RFLP bandingpatterns. In fact, it is highly likely that they allappeared at the same time, or at least within asingle population. The existence of 13 markersshared by almost all Dwarfs, whereas three ofthem are in the minority in Talls, seems to favouran appearance at the same time.
Dwarf coconut palms are rarely found in large,uniform stands (except for some commercialplantations set up this century), but they areoften found in the immediate vicinity of houses.Dwarfism can therefore be seen as the last stageof coconut palm domestication, in which thedetermining factor was the appearance ofautogamy, and the subsequent possibility ofreproducing true-to-type worthwhile palms. Thecharacters sought would have included slowgrowth, precocity or, in some varieties, theexcellent taste of the water in unripe nuts.Moreover, the often bright colour of the fruits,their characteristic shape and the palm growthhabit are effective varietal identification factors.
The first consequence of autogamy was therandom fixation of one allele to each locus(generally —but not always— the mostfrequent), and the subsequent conservation of themajor part of its genetic structure. We couldassume that the subsequent history of Dwarfscomprised long periods of relative stability,interspersed with sporadic gene exchanges withlocal Tall populations or other Dwarfs, whichhelped to differentiate between cultivars. Inreality, crossing a Dwarf and a Tall produces anF1 hybrid whose characteristics differ markedlyfrom those of the Dwarf. Most of the progenies ofsuch crosses have every chance of blending intothe mass of surrounding Tall populations. Onlycrossing a Dwarf with a Dwarf x Tall hybrid orwith another Dwarf has any chance of producing aDwarf type coconut palm, hence helping todiversify the Dwarf populations. This implies onthe one hand that such events are relatively rare,and on the other hand that each of them resultsin a limited number of allelic substitutions.
Under these conditions, it seems possible topropose a hypothetical diagram of Dwarf evolutionfrom the hypothetical original population:• the branch corresponding to Philippine and
Malayan Dwarfs apparently individualized quiteearly, with two shared alleles. The Malayancultivars form a uniform group with three otheralleles in common, and no doubt did notindividualize until later;
• a structure similar to that of the initialpopulation apparently subsisted, in a set ofcultivars collected from very diverse regions:PGD, KIGD, TBD, MBD;
• on the other hand, there is more markeddiversity amongst Pacific Red Dwarfs: TRD,VRD, plus the CRD, which could have a Pacificorigin although it was sampled in Cameroon.Although the Malayan and Philippine cases
are proof of a geographical structure to Dwarfvariability, it is much less clear-cut than for Talls:contrary to Talls, Dwarf cultivars imported intonew regions tend to remain genetically isolatedfrom local populations. This explains why similargenotypes are found in distant zones; for instance,the BGD has a banding pattern similar to that of the Philippine cultivars, and the cultivarssimilar to the PGD are spread throughout widelyscattered regions. In this respect, Dwarfs areproof of the intense exchanges that have takenplace within the zone in which coconut is grown.
What remains is to determine the region oforigin of Dwarf coconut varieties. This cannotnecessarily be done very accurately, since therandom fixation of alleles due to homozygositymay have profoundly modified the geneticstructure of the initial population right from thestart. The Tall cultivars that are most similar to itare found in a region stretching from SoutheastAsia to Papua New Guinea.
An original type: the Niu LekaAlthough it grows slowly like Dwarfs, the Niu Lekais a very specific variety by virtue of its allogamy,appearance and the fact that a Niu Leka x Tallhybrid resembles a Niu Leka. Its RFLP bandingpattern confirms its originality: it is clearlydifferent from that of Dwarfs, but extremelysimilar to Pacific Talls, particularly the TONT,followed by the RTMT, which correspondsprecisely to the zone of origin of the Niu Leka.The presence of the “rapid” Cox 1 allele in the NiuLeka and in one TONT individual is also in favourof a Tongan origin of the Niu Leka. However,there is no explanation for the existence in thiscountry of this allele, which is not found in anyother cultivars from the region.
Conclusion-prospectsRFLP data analysis is not finished, and it wouldbe wise to complete sampling with a smallnumber of cultivars to fill in a few gaps or checkcertain hypotheses. Around twelve cultivarsshould be used: Talls to be collected in LatinAmerica, Micronesia, Polynesia (Niu Kafa) andEast Africa, plus a few Dwarf or semi-Dwarfcultivars such as the King Coconut. It may alsobe worth seeking additional markers, notably todistinguish between certain Dwarfs. However, theresults available to date give a relatively precisepicture of the diversity of the species and of theways in which it has spread and evolved.
To some extent, the proposed scheme tallieswith that of Harries (1978), particularly asregards the situation observed in Latin America
Plantations, recherche, développement Juil let - Août 1998
244 CO C O N U TRetour au menu

RésuméLa diversité génétique du cocotier a été étu-diée à l’aide de marqueurs Rflp. Deux-cent-quatre-vingt-neuf cocotiers répartis entre 26 écotypes Grands et 16 écotypes Nains ontainsi été analysés. Ces cultivars proviennentdes principales zones de culture du cocotier.Vingt sondes ADNc de palmier à huile, deriz, de maïs et de cocotier ainsi qu’une sondecytoplasmique de blé ont été hybridées surl’ADN digéré à l’aide de quatre enzymes derestriction. A partir du polymorphisme molé-culaire, une Afc (analyse factorielle des cor-respondances) nous a permis de donner unereprésentation de la structure génétique despopulations. Ces résultats, ainsi que desconsidérations géographiques et historiques,nous ont conduit à individualiser deuxgroupes majeurs pour les cocotiers Grands,le premier contenant les écotypes originairesdu Sud-Est Asiatique et de l’Océan Pacifiquetandis que le second regroupe les écotypesdu sous-continent Indien et de l’Afrique del’Ouest. Les cultivars de l’Afrique de l’Estainsi que celui des îles Andamans possèdentdes marqueurs des deux groupes alors que le Grand Panama dérive vraisemblablementdu premier groupe. Tous les Nains (exceptéle Niu Leka) forment un ensemble trèshomogène relié au premier groupe desGrands. Ces résultats, qui nous permettentd’esquisser une histoire de la disséminationet de la domestication du cocotier, illustrentl’intérêt des marqueurs moléculaires pourla gestion et l’utilisation des ressourcesgénétiques.
AbstractA study of the genetic diversity in coconutwas performed using RFLP markers. A totalof 289 palms, representative of 26 Tall and16 Dwarf ecotypes, were analysed. The culti-vars originated from the major coconut cul-tivation areas. Twenty cDNA probes from oilpalm, rice, maize and coconut, and one cyto-plasmic probe from wheat were hybridizedon digested DNA using four restrictionenzymes. Based on the molecular polymor-phism, FCA (Factorial Correspondence Ana-lysis) was used to represent the structure ofthe populations. These results, along withgeographical and historical considerations,led us to define two main groups of Tall coconut palms, originating from South-EastAsia and the Pacific Ocean, and from theIndian sub-continent and West Africa. Thecultivars from East Africa and from theAndamans shared markers from both groups,whereas the Panama Tall appeared to bederived from the first one. All the Dwarfs(except Niu Leka) formed a highly homoge-neous group related to the first group ofTalls. These results give some idea of thehistory of the dissemination and domestica-tion of the coconut palm and illustrate themerits of molecular markers for the mana-gement and utilization of genetic resources.
ResumenSe estudió la diversidad genética del coco-tero mediante marcadores RFLP. De estemodo, se analizaron 289 cocoteros distribui-dos entre 26 ecotipos Grandes y 16 ecotiposEnanos. Estos cultivares proceden de lasprincipales zonas de cultivo del cocotero. Sehibridaron 20 sondas cDNA de palma deaceite, de arroz, de maíz y de cocotero asícomo una sonda citoplasmica de trigo sobreel ADN digerido mediante cuatro enzimasde restricción. A partir del polimorfismomolecular, una AFC (Análisis Factorial deCorrespondencias) nos ha permitido dar unarepresentación de la estructura de la diver-sidad de las poblaciones. Estos resultados,así como consideraciones geográficas e histó-ricas, nos llevaron a individualizar dos gru-pos mayores para los cocoteros Grandes, elprimero que contiene los ecotipos proce-dentes del Sureste Asiático y del OcéanoPacífico mientras que el segundo reúne losecotipos del subcontinente Indio y de Áfricadel Oeste. Los cultivares de África del Esteasí como aquel de las islas Andamans poseenmarcadores de ambos grupos mientras queel Grande Panamá deriva probablemente delprimer grupo. Todos los Enanos (salvo el NiuLeka) forman un conjunto muy homogéneorelacionado con el primer grupo de losGrandes. Estos resultados, que nos permi-ten esbozar una reseña histórica de la dise-minación y de la domesticación del cocotero,ilustran el interés de los marcadores mole-culares para la gestión y el uso de los recur-sos genéticos.
245CO C O N U T
Juil let - Août 1998 Plantations, recherche, développement
Acknowledgements
This study would not have been possiblewithout the collaboration of the research stationwhich collected and maintened the geneticmaterial studied here. We are very grateful tothe Directors and the staff members of IDEFOR-DPO (Côte d’Ivoire), CARFV (Vanuatu) and CIB(Jamaica) who provided us with the necessaryexperimental material.
and the Caribbean. However, the role played by domestication and by geographical isolation is markedly different. Despite a degree of resem-blance, the “Niu Kafa” of group I, and group II donot necessarily have the same origin. The dataavailable clearly suggest a single origin for Dwarfs,which would seem to be the ultimate stage indomesticating the coconut palm.
In certain situations, RFLP can provideextremely accurate varietal identification. It haseven been possible to detect exogenous genes incertain populations, and to explain the likelyreason. Given the very limited numbers ofindividuals used, this technique has provedextremely powerful.
It should make a significant contribution to germplasm management, particularly inidentifying redundancies or the degree of
representativeness of certain samples. Whenrejuvenating, for instance, it is not always clearwhether it is better to reproduce the originalpopulation or a sub-sample produced by massselection. A study of allelic frequency trendsshould provide an answer. In breeding, there isnow a way of limiting the number of combinationsto be tested, by working on both phenotypic and molecular similarities. A study of the hete-rosis observed in between-population hybridsdepending on genetic distance has yet to beconducted.
One possible application which we feel to beimportant is a study of populations from thePacific coast of Latin America, in relation toLethal yellowing tolerance. The three populationsstudied seem to stem from a small initialpopulation. A study of Pacific Tall population
diversity and dynamics, using larger samples,would provide a clearer picture of the type of initial inputs and ensure a more accurateidentification of the populations likely to transmitdisease tolerance. ■
Retour au menu