Bactériophage lambda - 2009.igem.org2009.igem.org/wiki/images/6/6a/Phage_lambda.pdf · pour les...
17
I. Généralité sur le phage lambd 1) Description du bactério Le bactériophage lambda est un bactériophage dit tempéré, ca conditions. Pendant le cycle lyt bactérie des progénies viraux so cycle lysogène, le génome du ba et sera transmit à la descendanc hôte. Le bactériophage lambda possèd particule virale, l’ADN est liné nucléotides simple brin complé injecter son génome au sein de l former un génome circulaire et a 2) Infection du bactériop Le bactériophage lambda se lie à l’extrémité de la queue. Cette de membrane d’E.Coli. Ensuite, le Bactériophage lambda da ophage lambda : n virus procaryote qui infecte la bactérie Esc ar il peut alterner phase lytique et lysog tique, l’ADN circulaire est répliqué en gra ont formés, conduisant à la mort de la cellu actériophage infectant, est intégré dans le ch ce. La réplication se fait alors au rythme des de un ADN double brin de 48 502 paire de éaire, mais ce dernier possède des extré émentaire. Au cours de l’infection, le bact la bactérie. Les extrémités cohésives peuvent ainsi permettre sa réplication. phage lambda : à sa cellule cible, Escherichia Coli, par la prot ernière se lie à une porine, faisant parti de génome linéaire du phage est injecté par cherichia Coli. C’est un gène selon certaines ande quantité dans la ule hôte. Au cours du hromosome bactérien, s divisions de la cellule e bases. Au sein de la émités cohésives : 12 tériophage lambda va t alors s’apparier pour téine J qui se trouve à l’opéron maltose, à la r diffusion passive (le
Transcript of Bactériophage lambda - 2009.igem.org2009.igem.org/wiki/images/6/6a/Phage_lambda.pdf · pour les...

I. Généralité sur le phage lambda
1) Description du bactériophage lambda
Le bactériophage lambda est un virus procaryote qui infecte la bactérie bactériophage dit tempéré, car il peut alterner phase lytique et lysogène selon certaines conditions. Pendant le cycle lytique, l’ADN circulaire est répliqué en grande quantité dans la bactérie des progénies viraux sont formés, conduisant à la mocycle lysogène, le génome du bactériophage infectant, est intégré dans le chromosome bactérien, et sera transmit à la descendance. La réplication se fait alors au rythme des divisions de la cellule hôte. Le bactériophage lambda possède unparticule virale, l’ADN est linéairenucléotides simple brin complémentaireinjecter son génome au sein de la bactérie. Les extrémités cohésives peuvent alors s’apparier pour former un génome circulaire et ainsi
2) Infection du bactériophage lambda
Le bactériophage lambda se lie à sa cellule cible, l’extrémité de la queue. Cette dernière se lie à une porine, faisant parti de l’opéron maltose, à la membrane d’E.Coli. Ensuite, le génome linéaire du phage est injecté par diffusion passive (le
Bactériophage lambda
Généralité sur le phage lambda
Description du bactériophage lambda :
Le bactériophage lambda est un virus procaryote qui infecte la bactérie Escherichia Coli.
bactériophage dit tempéré, car il peut alterner phase lytique et lysogène selon certaines cycle lytique, l’ADN circulaire est répliqué en grande quantité dans la
bactérie des progénies viraux sont formés, conduisant à la mort de la cellule hôte. Ale génome du bactériophage infectant, est intégré dans le chromosome bactérien,
sera transmit à la descendance. La réplication se fait alors au rythme des divisions de la cellule
possède un ADN double brin de 48 502 paire de particule virale, l’ADN est linéaire, mais ce dernier possède des extrémités
ides simple brin complémentaire. Au cours de l’infection, le bactériophage lambda vinjecter son génome au sein de la bactérie. Les extrémités cohésives peuvent alors s’apparier pour
circulaire et ainsi permettre sa réplication.
Infection du bactériophage lambda :
Le bactériophage lambda se lie à sa cellule cible, Escherichia Coli, par la protéine J qui se trouve à l’extrémité de la queue. Cette dernière se lie à une porine, faisant parti de l’opéron maltose, à la
Ensuite, le génome linéaire du phage est injecté par diffusion passive (le
Escherichia Coli. C’est un bactériophage dit tempéré, car il peut alterner phase lytique et lysogène selon certaines
cycle lytique, l’ADN circulaire est répliqué en grande quantité dans la rt de la cellule hôte. Au cours du
le génome du bactériophage infectant, est intégré dans le chromosome bactérien, sera transmit à la descendance. La réplication se fait alors au rythme des divisions de la cellule
ADN double brin de 48 502 paire de bases. Au sein de la ier possède des extrémités cohésives : 12
. Au cours de l’infection, le bactériophage lambda va injecter son génome au sein de la bactérie. Les extrémités cohésives peuvent alors s’apparier pour
, par la protéine J qui se trouve à l’extrémité de la queue. Cette dernière se lie à une porine, faisant parti de l’opéron maltose, à la
Ensuite, le génome linéaire du phage est injecté par diffusion passive (le
Bactériophage lambda
bactériophage lambda ne possédant pas de queue contractile) dans la cellule à travers la membrane. L’ADN se circularise immédiatement grâce aux sites cos par l’intermédiaire d’une ligase bactérienne. Par la suite, une gyrase bactérienne va permettre l’introduction de surenroulement de l’ADN, favorable aux transcriptions et réplications par les ARN et ADN polymérases de la cellule hôte.
3) Structure du génome :
Nom de la protéine
Fonction de protéines
Cro Contrôle de l’opérateur de cI, inhibiteur de la transcription qui se lie à OR1, OR2, OR3 (mais préférentiellement à OR3 = bloquage de cI).
cI Inhibiteur de la trancription, se lie à tous les opérateurs, même à OR3 (autorégulation de sa propre synthèse).
cII Activateur de la trancription (promoteurs pAQ, pRE, pI), sensible aux protéases bactérienne (HflB).
cIII Se lie aux HflB et protège ainsi cII d’être dégradé par les protéases.
N Antiterminateur, se lie à l’ARN polymérase, permet à cette dernière d’ignorer les sites de terminaison.
Q Se lie à l’ARN et aux cofacteurs de l’ARN polymérase, les sites de terminaison sont ignorés, seul la sequence de terminaison de Q est
efficace.
Xis Entraîne l’excision du génome du phage dans le chromosome bactérien.
Int Entraîne l’insertion du génome du phage dans le chromosome bactérien.
D Protéines majeures de la capside du phage, rôle dans la stabilité de cette dernière, souvent associés à des protéines de ciblage.
A, B, C, E, F, Protéines participant à la formation de la tête du phage.
J Protéines majeures de la queue du phage, souvent associé à d’éventuelles protéines de ciblage.
Z, U, V, G, T, H, M, L, K, I
Protéines participant à la formation de la queue du phage.
S, R Autrement appeler holine et endolysine, ils permettent à d’importante concentration, de provoquer la lyse de la cellule hôte.
O, P Protéines impliquées dans la réplication du génome du phage.

Nom de la protéine
attP Point d’action de int et xis, correspond à attB
4) Le cycle lytique:
a) Transcription primaire de l’ADN phagique:
Très tôt au cours de l’infection, l’ARN polymérase se fixe sur l’ADN phagique et initie la transcription par 2 promoteurs, p«précoce» issue de cette transcription, se termine à la fin des gènes tL1 et tR1, respectivement. Le produit du gène N est un facteur d’antiterminaison, qui en se fixantbactérienne, permet la transcription de gènes se trouvant conséquence, la transcription des gènes suivant, très important pour le cycle lytique
- O : codant pour une protéine phage,
- P : codant pour une protéine qui va interagir avec O, pour permettre sa fixation sur ORI, et inhiber certaines protéines bactériennes.
- Q : autrement appeler protéine de lyse, comme N, Q est unmodifie l’ARN polymérase et initie la transcription au niveau du promoteur fort pour les protéines de lyse et structure du phage.
D’autre part, à la droite de cI, l’accumulation du produit du gène OR3, permet de bloquer sa trancription, et d’ainsi d’inhiber complètement l’entrée en phase lysogène.
Bactériophage lambda
Fonction de protéines
Point d’action de int et xis, correspond à attB qui est le point d’insertion du génome du phage.
Transcription primaire de l’ADN phagique:
Très tôt au cours de l’infection, l’ARN polymérase se fixe sur l’ADN phagique et initie la pL et pR situé à la gauche et la droite du gène
«précoce» issue de cette transcription, se termine à la fin des gènes N et cro,
Le produit du gène N est un facteur d’antiterminaison, qui en se fixant sur l’ARN polymérase la transcription de gènes se trouvant au-delà de tL1 et
conséquence, la transcription des gènes suivant, très important pour le cycle lytiquecodant pour une protéine qui va se fixer sur l’origine de réplication du génome du
: codant pour une protéine qui va interagir avec O, pour permettre sa fixation sur ORI, et certaines protéines bactériennes.
: autrement appeler protéine de lyse, comme N, Q est un facteur d’modifie l’ARN polymérase et initie la transcription au niveau du promoteur fort
les protéines de lyse et structure du phage.
I, l’accumulation du produit du gène cro, sur l’opérateur du gène 3, permet de bloquer sa trancription, et d’ainsi d’inhiber complètement l’entrée en phase
qui est le point d’insertion du génome du phage.
Très tôt au cours de l’infection, l’ARN polymérase se fixe sur l’ADN phagique et initie la du gène cI (répresseur). L’ARN
cro, au niveau des sites
sur l’ARN polymérase et tR1. Cela a donc pour
conséquence, la transcription des gènes suivant, très important pour le cycle lytique : l’origine de réplication du génome du
: codant pour une protéine qui va interagir avec O, pour permettre sa fixation sur ORI, et
facteur d’antiterminaison. Il modifie l’ARN polymérase et initie la transcription au niveau du promoteur fort pR’, codant
sur l’opérateur du gène cI, 3, permet de bloquer sa trancription, et d’ainsi d’inhiber complètement l’entrée en phase

Bactériophage lambda
b) Transcription secondaire de l’ADN phagique:
Une accumulation de la protéine Q, permet la transcription des gènes tardifs (protéines de structures de la tête, de la queue, etc…) de commencer. La protéine N, en neutralisant l’activité du facteur de terminaison d’E-Coli ρ, permet l’assemblage des complexes de transcription pour le cycle lytique. Le complexe est formé au niveau des ARNs sur les sites nutL et nutR et est arrangé en ARN polymérase et autres protéines nommées Nus. La protéine N est donc un régulateur important dont l’activité est nécessaire pour la phase lytique du bactériophage lambda.
c) Réplication de l’ADN phagique:
Durant la phase précoce de l’infection, l’ADN circulaire du phage lambda est répliquée de manière bidirectionnelle à partir d’une origine de réplication unique (ori), dont l’activité requière les protéines virales 0 et P. Plus tard, la réplication en «cercle roulant» commence, plusieurs génomes entiers sont ainsi générés, formant un concatémère. Durant l’encapsidation, le concatémère est clivé au niveau des sites cos, par le produit du gène phagique A (Terminase phagique), afin de former un monomère d’ADN avec des extrémités cohésives, présent dans les particules de bactériophages matures.

Bactériophage lambda
Synthèse du concatémère : Polymère du génome du phage lambda
d) Transcription de l’ADN phagique:
L’un des produits de la transcription précoce de pR code pour une protéine appelée cro. Comme cro s’accumule durant l’infection, ce dernier va se fixer sur OL et OR, inhibant l’attachement de l’ARN polymérase et ainsi, bloque la transcription de pL et pR. A ce moment, un taux suffisant d’une protéine contrôle, appelé Q, est synthétisé pour permettre la transcription des gènes tardifs de commencer. Q est un régulateur positive de la synthèse d’ARN à partir de p’R, le promoteur utilisé pour la transcription de la partie tardive du génome entier, contenant les gènes impliqués dans la tête et la queue.
e) Assemblage du bactériophage
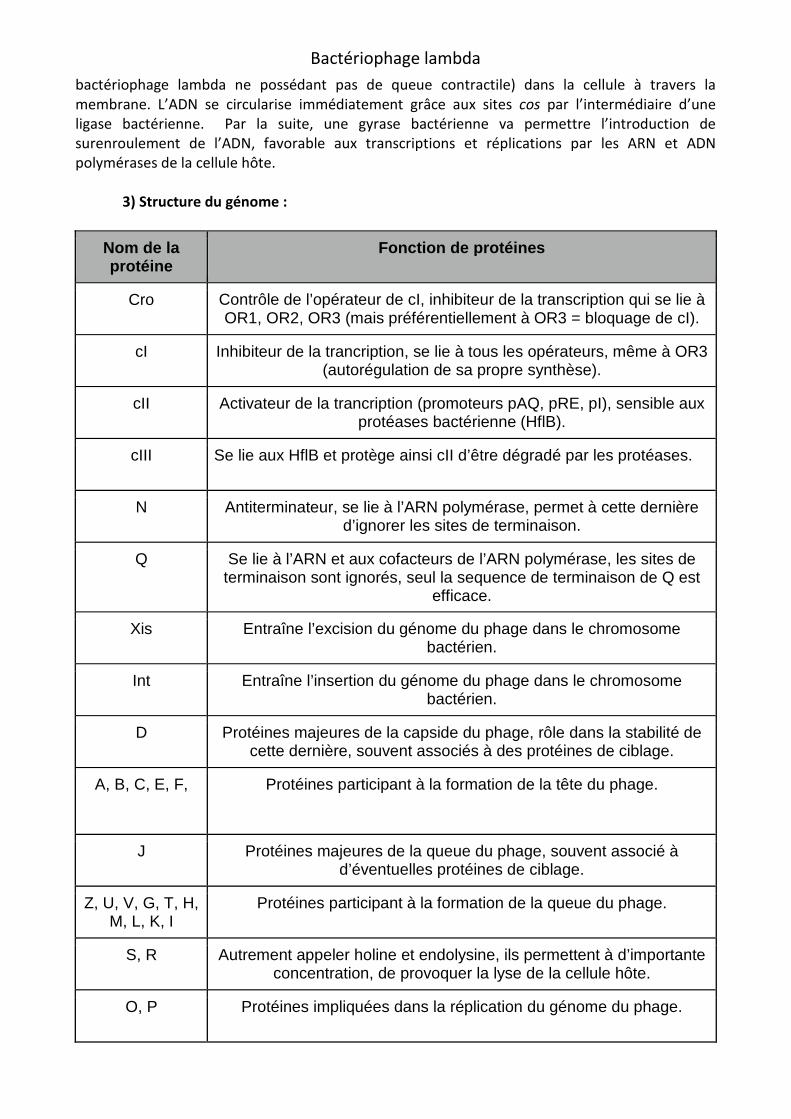
Les deux parties majeures de la particule mature, la tête et la queue, sont assemblées séparément. Tout d’abord débute la formation de la capside. Le bactériophage va coder pour les protéines B, C et Nu3, vont former un premier complexe sans forme définie (1). Ensuite, les protéines de l’hôte GroEL et GroES, permettent la maturation de la celle-ci, et la formation d’une procapside (2). Puis l’ajout de la protéine E, permet de former la tête du phage immature (3), puis
la fusion entre les protéines C et phage est ainsi mature (4). Le clivage de la protéine B permet la formation d’uncapside (5). L’encapsidation du génome phagique requièrepartir du concatémère d’ADN. Ceci est réalisé grâce à la présence des séquences cos à la jonction de deux génomes du concatémèrecos sont également les sites de terminaison de l’encapsidation.
Formation de la capside
L’enzyme terminase du phage lambda, reconnaît l’ADN viral, et va préparer une extrémité du génome pour le packaging. La terminasel’activité endonucléase, et est donc responsable de l’initiation/terminaison de l’ADN à encapsider. Ceci est réalisé par l’introduction d’une coupure dans les séquences également appelée «hélicase», puisqu’il va permettre la séparation des brins d’ADN au niveau des coupures, formant un génome mature ayant des extrémités cohésives. Cette même souscontient également une activité translocase, responsable de l’encapsidatiATPase, permettant la translocation. La sousLa sous-unité gpNu1, et donc essentiel à son activité, permettant la fixation sur les séquences cos, l’assemblage et la stabilité de la machinerie d’encapsidation ( En résumé, la terminase se fixe sur une séquence ce qui permet la libération de l’extrémité gauche du génome à encapsider. La liaison du complexe enzyme-DNA à une procapside vide permet l’initiation de l’encapsidation.
(1)
(5)
Bactériophage lambda
et E, permet la formation des protéines X1
Le clivage de la protéine B permet la formation d’unL’encapsidation du génome phagique requière qu’un monomère d’ADN soit généré à
d’ADN. Ceci est réalisé grâce à la présence des séquences cos à la jonction concatémère, et servent de site d’initiation de l’encapsidation. Les séquences
ites de terminaison de l’encapsidation.
L’enzyme terminase du phage lambda, reconnaît l’ADN viral, et va préparer une extrémité du génome pour le packaging. La terminase est composé des sous-unités gpNu1
l’activité endonucléase, et est donc responsable de l’initiation/terminaison de l’ADN à encapsider. Ceci est réalisé par l’introduction d’une coupure dans les séquences cos. La sous
«hélicase», puisqu’il va permettre la séparation des brins d’ADN au niveau des coupures, formant un génome mature ayant des extrémités cohésives. Cette même souscontient également une activité translocase, responsable de l’encapsidation de l’ADN, et activité ATPase, permettant la translocation. La sous-unité gpA seule a une activité catalytique très faible.
, et donc essentiel à son activité, permettant la fixation sur les séquences cos, de la machinerie d’encapsidation (gpA+gpNu1).
En résumé, la terminase se fixe sur une séquence cos du concatémère et provoque une coupure, ce qui permet la libération de l’extrémité gauche du génome à encapsider. La liaison du complexe
ocapside vide permet l’initiation de l’encapsidation.
(2) (3)
(4)
X1 et X2, la capside du Le clivage de la protéine B permet la formation d’une entrée dans la
qu’un monomère d’ADN soit généré à d’ADN. Ceci est réalisé grâce à la présence des séquences cos à la jonction
, et servent de site d’initiation de l’encapsidation. Les séquences
L’enzyme terminase du phage lambda, reconnaît l’ADN viral, et va préparer une extrémité du gpNu1 et gpA. La gpA porte
l’activité endonucléase, et est donc responsable de l’initiation/terminaison de l’ADN à encapsider. . La sous-unité gpA, est
«hélicase», puisqu’il va permettre la séparation des brins d’ADN au niveau des coupures, formant un génome mature ayant des extrémités cohésives. Cette même sous-unité
on de l’ADN, et activité seule a une activité catalytique très faible.
, et donc essentiel à son activité, permettant la fixation sur les séquences cos,
et provoque une coupure, ce qui permet la libération de l’extrémité gauche du génome à encapsider. La liaison du complexe
Bactériophage lambda
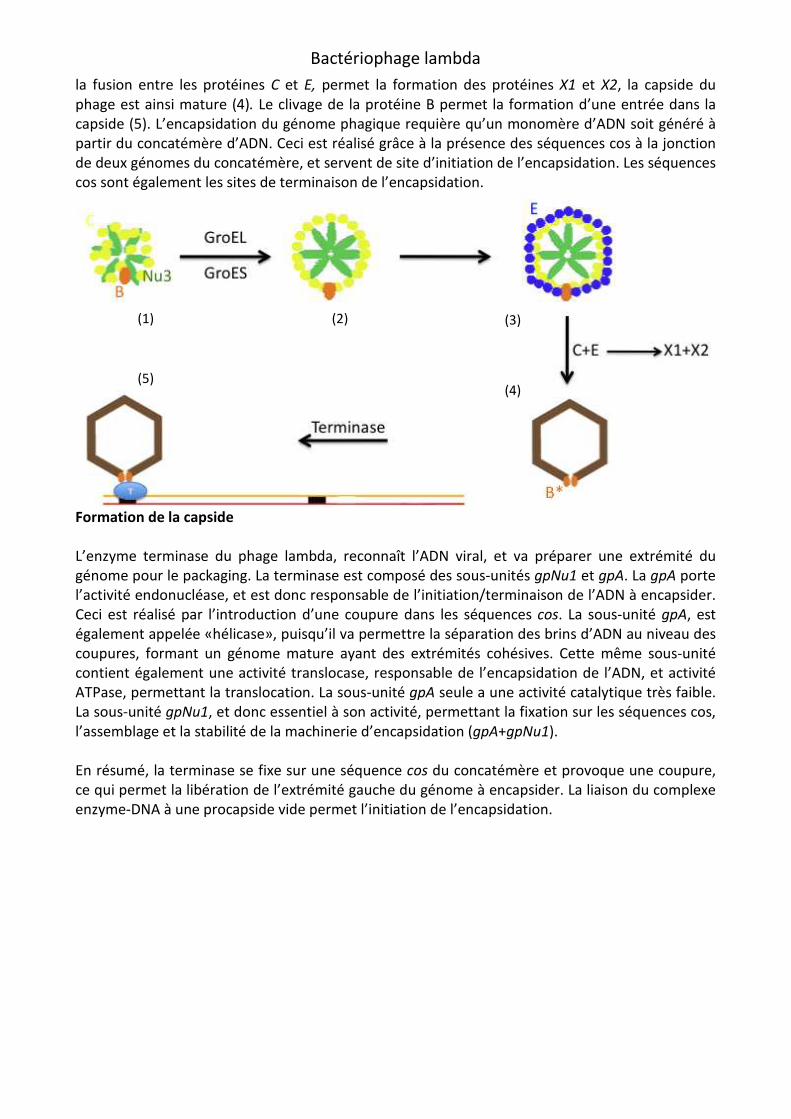
Fixation du complexe gpA/gpNu1 sur l’ADN du bactériophage lambda
=>> Molecular Cloning: A Laboratory Manual
(Third Edition) =>> A New Look at Bacteriophage lambda Genetic Networks
f) Les séquences cos du bactériophage:
Le site cos est une séquence d’environ 200 pb. Environ 40 pb sont localisés à la fin du génome linéaire du phage lambda (48462-48502) et le reste de la séquence se trouve au début du génome linéaire (1-132). Le site où la terminase introduit des coupures génératives des extrémités cohésives est appelé cosN. Une initiation efficace nécessite la présence de la sous-unité cosB, situé après cosN. Une terminaison efficace nécessite la présence de la sous-unité cosQ, sous-unité présente avant cosN. -CosN: La terminase va introduire des coupures dans le complexe cos du concatémère au niveau du cosN et vont générer des extrémités cohésives de l’ADN mature, et initier ainsi l’encapsidation. CosN est fonctionnellement asymétrique. CosNL (coté gauche) a une mutation G2C, CosNR (coté droit) a une mutation C11G. Les positions des coupures sont disposées

Bactériophage lambda
symétriquement dans la séquence. Une première dans CosNL appelé N2. Une dans CosNR appelé N1. -CosB: Site de fixation de la terminase sur le complexe cos, cosB est localisé à la droite de cosN. cosB possède 3 sites distinctes de liaison à gpNu1: R1, R2, R3 (position 53 à 166). Cette liaison aux sites R est médiée par un site spécifique de gpNu1. Entre R2 et R3 se trouve I1, un site de fixation pour le facteur d’intégration de l’E-coli (IHF). -CosQ: Rôle important dans la terminaison de l’ADN empaqueté. cosQ est constitué de 7 pb et est localisé 17 pb après cosN. Une mutation sévère de cosQ peut affecter les coupures dans cosN, Et de l’ADN présent après le complexe cos peut alors être encapsider en plus du génome. cosQ est donc nécessaire en tant que séquence d'arrêt de l’encapsidation, mais cosQ seule est incapable d’arrêter cette encapsidation. Il agit donc avec cosN et I2. I2 est une séquence située entre cosN et cosB, et semble jouer un rôle important dans l’initiation et la terminaison du clivage de cosN (des délétions dans la séquence de I2 ont provoqué des baisses de la fréquence des coupures). Il semblerait que I2 est un rôle également déterminant dans l’espace qui sépare cosN et cosB (le remplacement de la séquence s’est révélé létale). Au final, I2 a une rôle spécifique dans le bon déroulement de la terminaison de l’encapsidation (séquence cosQ-cosN-I2 doit être présent). =>> Bactériophage Lambda Terminase and the Mechanism of Viral DNA Packaging. =>> The Functional Asymmetry of cosNcosNcosNcosN, the Nicking Site for Bacteriophage λ DNA Packaging, Is
Dependent on the Terminase Binding Site, cosBcosBcosBcosB).
=>> Bacteriophage lambda DNA : The begginning of the End.
La terminase reste lié au concatémère une fois que l’ADN («un monomère» est été encapsidé). L’ensemble enzyme-DNA, se fixe à une procapside vide afin d’initier un second tour d’encapsidation.
g) Mécanisme d’encapsidation:
Il a été démontré que l’IHF (Integration Host Factor) agit en symbiose avec la gpNu1, lors de l’assemblage du mécanisme d’encapsidation. Mais ce phénomène n’est pas encore très bien décrit. Il existe cependant un modèle décrivant assez bien le phénomène d’initiation de l’encapsidation. L’IHF se fixe au niveau de I1, se qui provoque une courbure de l’ADN, et permet À gpNu1 de se fixer sur les sites R(1,2,3) de cosB. On ne sait pas lequel de IHF ou gpNu1 se fixe en premier. Un autre modèle décrit le phénomène de terminaison. =>> Bacteriophage Lambda gpNu1 and Escherichia coli IHF Proteins Cooperatively Bind and Bend
Viral DNA: Implications for the Assembly of a Genome-Packaging Motor.
h) Lyse de la bactérie:
Deux protéines du phages, R (endolysine) et S (holine), sont requises pour lyser la cellules hôte et libérer les progénies du phages. La membrane cytoplasmique est rompue par la holine, qui réalise des liaisons non-spécifique assez grande pour permettre le passage de l’endolysine, qui va digérer la couche peptidoglycane.

Bactériophage lambda
== Schéma possible== =>> Holin of bacteriphage lambda: structural insights into a membrane lesion.
2) Cycle lysogène:
Dans une petite proportion de cellules infectées, le cycle lytique est abandonné et un pathway alternatif est activé, menant à l’intégration de l’ADN du phage lambda dans le génome de la bactérie hôte. Deux protéines d’origine virale, Int et cI, sont requises pour établir une lysogénie stable. Int permet l’intégration du génome du phage dans le chromosome bactérien, et cI represse l’initiation des promoteurs pR et pL, bloquant ainsi toute expression de gènes impliqués dans le cycle lytique. a) cII et cIII, l’établissement de la répression:
Les protéines cII et cIII, sont synthétisés suite à la transcription primaire. Ils sont uniquement requis pour activer initialement la synthèse de cI. Par la suite, cI seul assure le maintien de la lysogénie, et assure sa propre synthèse. La protéine cII lance la transcription à partir de 3 promoteurs pI (qui transcrit le gène int), pRE (qui transcrit le gène cI), et pAQ (qui transcrit un ARN antisens dérivant du gène Q). b) cI, le répresseur de la transcription phagique:
Le produit du gène cI est un represseur de la transcription primaire et va donc bloquer l’expression des gènes tardifs (gènes de structure : capside et queue). La protéine int reconnaît les séquences au niveau des sites att dans les génomes bactériens et viraux, et catalyse une cassure puis une liaison, menant à l’insertion de l’ADN viral dans le chromosome de l’hôte. cI contient deux domaines distinctes qui sont connecté par une région sensible aux protéases. La région N terminal, est un domaine de liaison à l’ADN, et possède également une zone de contact avec l’ARN polymérase, bloquant l’activation de la transcription. La région C terminal permet la liaison entre plusieurs cI, et ainsi la liaison à plusieurs sites opérateurs. A l’état prophage (état intégré au génome), toutes transcriptions est réprimées, uniquement le gène cI est exprimé, et permet donc le maintient du cycle lysogène. L’expression de cI permet la formation d’un oligomère, qui va se fixer sur les différents sites opérateurs du génome du phage lambda. Ce dernier possède 6 sites opérateurs : 3 dans chaque région contrôle : OR (OR1, OR2, et OR3) et OL (OL1, OL2, et OL3). La fixation de cI sur les 3 opérateurs de la région de contrôle OR, qui sont bordés par les promoteurs pR et pRM, bloque la transcription des gènes de lyse précoce et permet donc le maintien de la lysogénie. La fixation de cI sur les 3 opérateurs de la région de contrôle OL, bloque également les gènes de lyse précoce promus par pL cI bloque donc son propre promoteur pRM, permettant ainsi une autorégulation de sa transcription.

Etat prophage : Fixation de cI sur les sites opérateur
Le plus souvent le cycle lytique est réactivé par un état de stress de la bactérie causé par la dégradation de son ADN ce qui induit une réponse SOS et le retour à un cycle bactérienne RecA (dommage de l’ADN) se lie site sensible aux protéasex reliant les deux domaines de et C terminal. Ce clivage va libérer les sites opérateurs et permettre la transcription des promoteurs pR et pL. Le premier gène transcrit, sous le contrôle du promoteur qui se lie aux mêmes opérateurrepressant ainsi la trancription synthèse de cI. =>> The bactériophage lambda cI protein finds an asymmetric solution
Bactériophage lambda
I sur les sites opérateurs
Le plus souvent le cycle lytique est réactivé par un état de stress de la bactérie causé par la dégradation de son ADN ce qui induit une réponse SOS et le retour à un cycle bactérienne RecA (dommage de l’ADN) se lie alors à cI, provoquant un autosite sensible aux protéasex reliant les deux domaines de cI, séparant ainsi les domaines N terminal
va libérer les sites opérateurs et permettre la transcription des
Le premier gène transcrit, sous le contrôle du promoteur pR code pour un autre répresseur (opérateurs que cI. Mais Cro se lie de manière plus
repressant ainsi la trancription de cI partant du promoteur pRM, réprimant ainsi une nouvelle
The bactériophage lambda cI protein finds an asymmetric solution.
Le plus souvent le cycle lytique est réactivé par un état de stress de la bactérie causé par la dégradation de son ADN ce qui induit une réponse SOS et le retour à un cycle lytique. La protéine
I, provoquant un auto-clivage au niveau du I, séparant ainsi les domaines N terminal
va libérer les sites opérateurs et permettre la transcription des
R code pour un autre répresseur (Cro), se lie de manière plus spécifique à OR3,
, réprimant ainsi une nouvelle
Bactériophage lambda
II. Construction du vecteur phage lambda:
Il y a de nombreux avantage à utiliser comme vecteur un bactériophage lambda recombinant pour la thérapie génique. En effet, d’un point de vue connaissances, le bactériophage lambda a été très étudié: entièrement séquencé, très utilisé en tant que vecteur de clonage, le génome du bactériophage lambda est même utilisé, afin de réaliser des banques d’ADN génomique (insertion d’ADN exogène entre les séquences cos: cosmide). De plus, au niveau éthique, le phage lambda n’étant naturellement pas infectieux pour les cellules eucaryotes, il ne peut pas se répliquer. D’un point de vue industrielle, le bactériophage lambda est très peu coûtant à produire, facile à réaliser et en quantité importante, grâce à sa réplication naturelle dans les bactéries Escherichia
Coli. D’un point de vue stratégie thérapeutique, le phage lambda est un vecteur intéressant puisque n’importe quel gène thérapeutique peut-être insérer entre les séquences cos, et sera ainsi encapsider dans la tête du phage. En travaillant donc sur la manière dont on va rendre le bactériophage lambda spécifique des cellules eucaryotes, on peut donc espérer obtenir un vecteur en grande quantité, peu coûteux, non-réplicatif, spécifique à un type cellulaire, donc quasi idéal pour la thérapie génique. Il existe quelques données expérimentales sur l’utilisation de phages lambda recombinants en thérapie génique avec des résultats plutôt prometteurs.
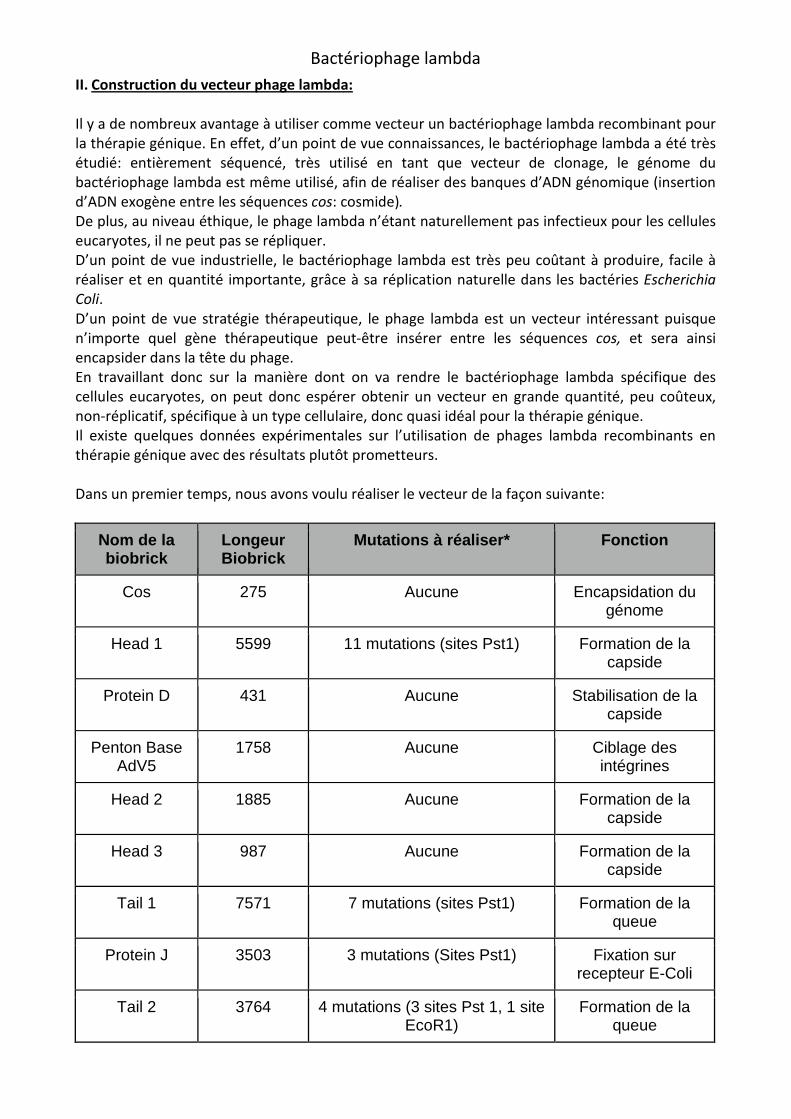
Dans un premier temps, nous avons voulu réaliser le vecteur de la façon suivante:
Nom de la biobrick
Longeur Biobrick
Mutations à réaliser* Fonction
Cos 275 Aucune Encapsidation du génome
Head 1 5599 11 mutations (sites Pst1) Formation de la capside
Protein D 431 Aucune Stabilisation de la capside
Penton Base AdV5
1758 Aucune Ciblage des intégrines
Head 2 1885 Aucune Formation de la capside
Head 3 987 Aucune Formation de la capside
Tail 1 7571 7 mutations (sites Pst1) Formation de la queue
Protein J 3503 3 mutations (Sites Pst1) Fixation sur recepteur E-Coli
Tail 2 3764 4 mutations (3 sites Pst 1, 1 site EcoR1)
Formation de la queue
Bactériophage lambda
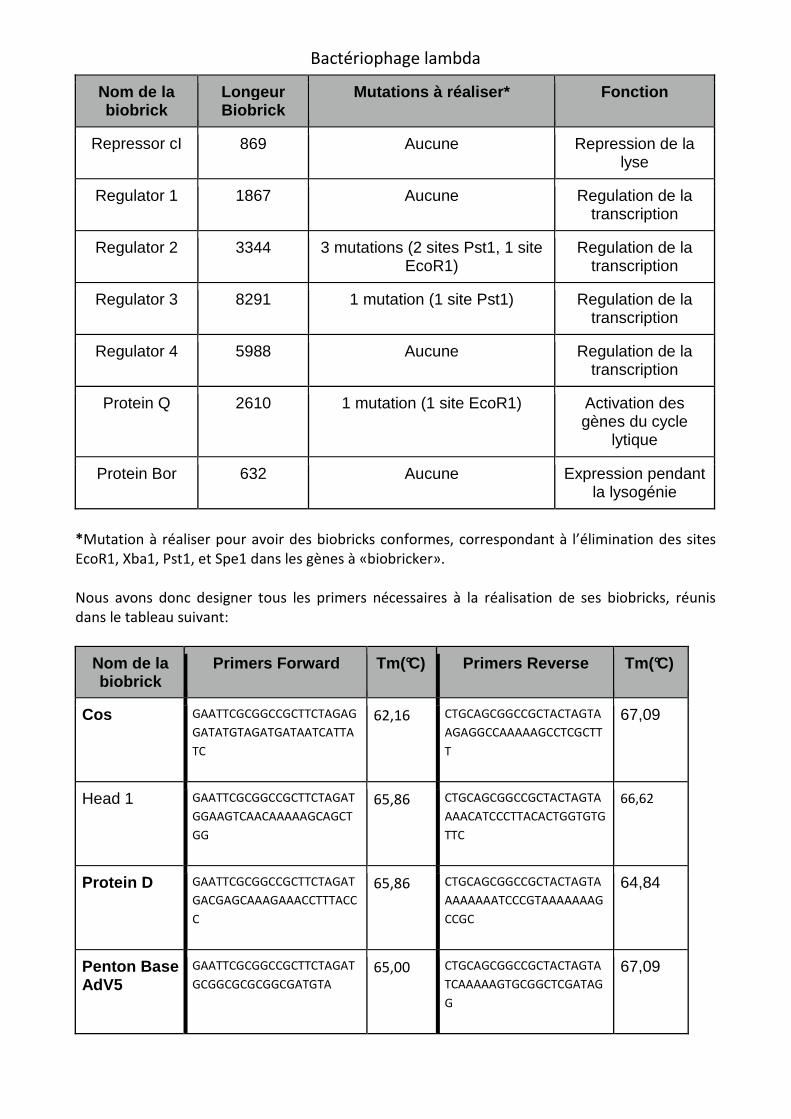
Nom de la biobrick
Longeur Biobrick
Mutations à réaliser* Fonction
Repressor cI 869 Aucune Repression de la lyse
Regulator 1 1867 Aucune Regulation de la transcription
Regulator 2 3344 3 mutations (2 sites Pst1, 1 site EcoR1)
Regulation de la transcription
Regulator 3 8291 1 mutation (1 site Pst1) Regulation de la transcription
Regulator 4 5988 Aucune Regulation de la transcription
Protein Q 2610 1 mutation (1 site EcoR1) Activation des gènes du cycle
lytique
Protein Bor 632 Aucune Expression pendant la lysogénie
*Mutation à réaliser pour avoir des biobricks conformes, correspondant à l’élimination des sites EcoR1, Xba1, Pst1, et Spe1 dans les gènes à «biobricker».
Nous avons donc designer tous les primers nécessaires à la réalisation de ses biobricks, réunis dans le tableau suivant:
Nom de la biobrick
Primers Forward Tm(°C) Primers Reverse Tm(°C)
Cos GAATTCGCGGCCGCTTCTAGAG
GATATGTAGATGATAATCATTA
TC
62,16 CTGCAGCGGCCGCTACTAGTA
AGAGGCCAAAAAGCCTCGCTT
T
67,09
Head 1 GAATTCGCGGCCGCTTCTAGAT
GGAAGTCAACAAAAAGCAGCT
GG
65,86 CTGCAGCGGCCGCTACTAGTA
AAACATCCCTTACACTGGTGTG
TTC
66,62
Protein D GAATTCGCGGCCGCTTCTAGAT
GACGAGCAAAGAAACCTTTACC
C
65,86 CTGCAGCGGCCGCTACTAGTA
AAAAAAATCCCGTAAAAAAAG
CCGC
64,84
Penton Base AdV5
GAATTCGCGGCCGCTTCTAGAT
GCGGCGCGCGGCGATGTA 65,00 CTGCAGCGGCCGCTACTAGTA
TCAAAAAGTGCGGCTCGATAG
G
67,09

Bactériophage lambda
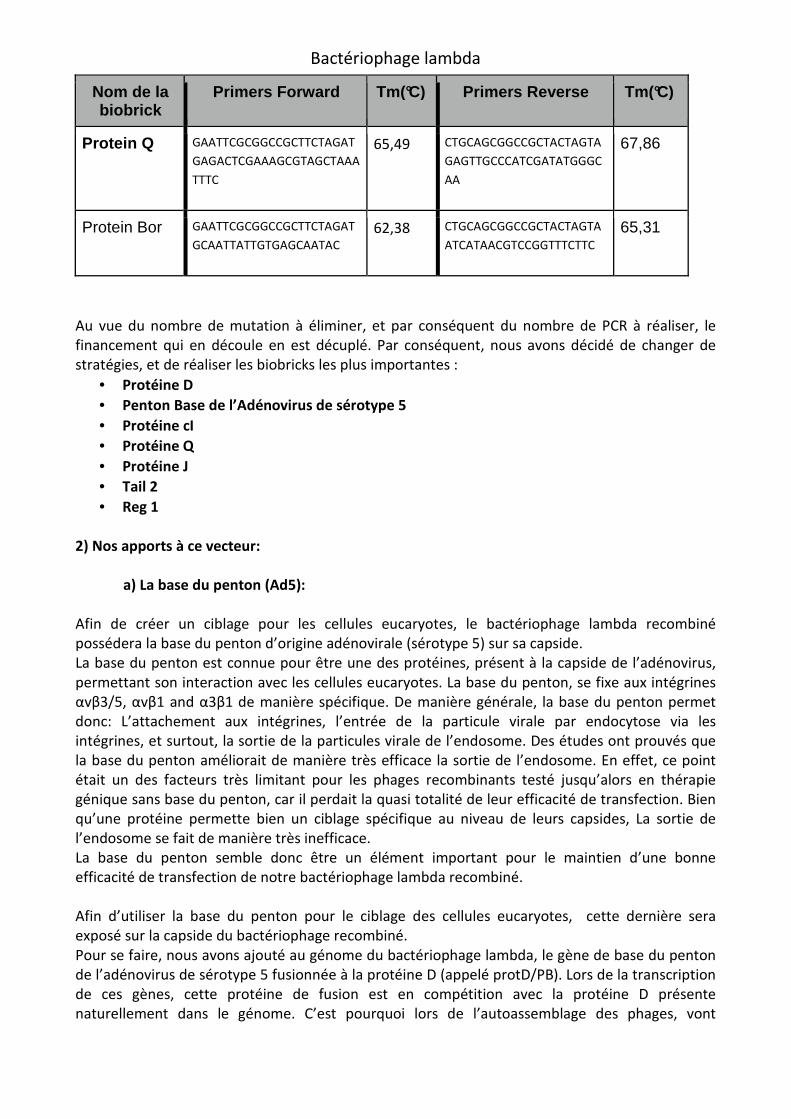
Nom de la biobrick
Primers Forward Tm(°C) Primers Reverse Tm(°C)
Head 2 GAATTCGCGGCCGCTTCTAGAT
GTCGATGTACACAACCGCCCAA
C
67,68 CTGCAGCGGCCGCTACTAGTA
ACATCCCCCTTTCAGCGGCGA
69,21
Head 3 GAATTCGCGGCCGCTTCTAGAG
TTAGCGTGGAAAGATTTGTGTA 64,13 CTGCAGCGGCCGCTACTAGTA
ATGATTAGCAGTTATGGTGAT
C
64,23
Tail 1 GAATTCGCGGCCGCTTCTAGAT
GGCCATAAAAGGTCTTGAGCAG
G
66,77 CTGCAGCGGCCGCTACTAGTA
TCCTCACGCTCCATAAATGACA
AAA
65,73
Protein J GAATTCGCGGCCGCTTCTAGAT
GGGTAAAGGAAGCAGTAAGGG 66,14 CTGCAGCGGCCGCTACTAGTA
CGTTCACCTCTCACTGTTTTATA
AT
64,86
Tail 2 GAATTCGCGGCCGCTTCTAGAT
GCGTAATGTGTGTATTGCCGTT 65,06 CTGCAGCGGCCGCTACTAGTA
GTCATAGAACTTCCATTATTCT
C
64,13
Repressor cI
GAATTCGCGGCCGCTTCTAGAG
AATTCAATCCATTTACTATGTTA
TG
62,12 CTGCAGCGGCCGCTACTAGTA
ATGAGCACAAAAAAGAAACCA
TTAA
63,06
Regulator 1 GAATTCGCGGCCGCTTCTAGAG
TCAATTGTTCATAGTGTTTACAT 62,21 CTGCAGCGGCCGCTACTAGTA
AGAATATGATGAAAGATAGAA
AA
61,33
Regulator 2 GAATTCGCGGCCGCTTCTAGAG
AGATCCCTCTGAAAAAATCTTC 64,13 CTGCAGCGGCCGCTACTAGTA
TGATTTTATTTTGACTGATAGT
G
62,27
Regulator 3 GAATTCGCGGCCGCTTCTAGAG
CGCAAAGTCTCCGCAATTACAC
GCA
68,22 CTGCAGCGGCCGCTACTAGTA
TCGGCAAGGTGTTCTGGTCGG
C
69,95
Regulator 4 GAATTCGCGGCCGCTTCTAGAG
ACGTTAAATCTATCACCGCAAG
GGA
66,48 CTGCAGCGGCCGCTACTAGTA
GCGCCTTCTCCCTGTACCTGAA
69,00
Bactériophage lambda
Nom de la biobrick
Primers Forward Tm(°C) Primers Reverse Tm(°C)
Protein Q GAATTCGCGGCCGCTTCTAGAT
GAGACTCGAAAGCGTAGCTAAA
TTTC
65,49 CTGCAGCGGCCGCTACTAGTA
GAGTTGCCCATCGATATGGGC
AA
67,86
Protein Bor GAATTCGCGGCCGCTTCTAGAT
GCAATTATTGTGAGCAATAC 62,38 CTGCAGCGGCCGCTACTAGTA
ATCATAACGTCCGGTTTCTTC
65,31
Au vue du nombre de mutation à éliminer, et par conséquent du nombre de PCR à réaliser, le financement qui en découle en est décuplé. Par conséquent, nous avons décidé de changer de stratégies, et de réaliser les biobricks les plus importantes :
• Protéine D
• Penton Base de l’Adénovirus de sérotype 5
• Protéine cI
• Protéine Q
• Protéine J
• Tail 2
• Reg 1
2) Nos apports à ce vecteur:
a) La base du penton (Ad5):
Afin de créer un ciblage pour les cellules eucaryotes, le bactériophage lambda recombiné possédera la base du penton d’origine adénovirale (sérotype 5) sur sa capside. La base du penton est connue pour être une des protéines, présent à la capside de l’adénovirus, permettant son interaction avec les cellules eucaryotes. La base du penton, se fixe aux intégrines αvβ3/5, αvβ1 and α3β1 de manière spécifique. De manière générale, la base du penton permet donc: L’attachement aux intégrines, l’entrée de la particule virale par endocytose via les intégrines, et surtout, la sortie de la particules virale de l’endosome. Des études ont prouvés que la base du penton améliorait de manière très efficace la sortie de l’endosome. En effet, ce point était un des facteurs très limitant pour les phages recombinants testé jusqu’alors en thérapie génique sans base du penton, car il perdait la quasi totalité de leur efficacité de transfection. Bien qu’une protéine permette bien un ciblage spécifique au niveau de leurs capsides, La sortie de l’endosome se fait de manière très inefficace. La base du penton semble donc être un élément important pour le maintien d’une bonne efficacité de transfection de notre bactériophage lambda recombiné. Afin d’utiliser la base du penton pour le ciblage des cellules eucaryotes, cette dernière sera exposé sur la capside du bactériophage recombiné. Pour se faire, nous avons ajouté au génome du bactériophage lambda, le gène de base du penton de l’adénovirus de sérotype 5 fusionnée à la protéine D (appelé protD/PB). Lors de la transcription de ces gènes, cette protéine de fusion est en compétition avec la protéine D présente naturellement dans le génome. C’est pourquoi lors de l’autoassemblage des phages, vont

coexister ceux qui auront la protéine naturelle à la capside, et des phages recombinés avec la protéine de fusion ProtD/PB.
=>> Identification of a conserved receptor
adenovirus. =>> Binding Properties, Cell Delivery, and Gene Transfer of Adenoviral Penton Base Displaying
Bacteriophage.
Génome du phage modifié
b) Le contrôle du répresseur
Pour notre projet, nous avons souhaité modifier notre bactériophage lambda pour entrée en phase lytique. Le phage lambda est naturellement tempéré, alternant cycle lytique et lysogène. Afin de contrôler l’entrée en phase lytiquecI, le répresseur du cycle lytique. Ce dernier va ainsi bloquerpermettre l’inhibition de l’expression des gènes lytiquesNotre but est donc d’activer l’expression devoulu. Pour cela, nous utilisons le système régulant l’opéron Le répresseur tetR se fixe en continu surdevant le gène dont la trancription est à contrôlerl’inhibition est levé, ce qui permet la trancription Dans notre système, le gène dont nous voulons contrôler l’expression est le répresseur l’opéron Lac. Ce dernier, une fois transcrit, ira inhiber l’opérateur de l’opéron Lac, qui sera associé au répresseur cI (séparé de son opérateur et promoteur d’origine OR1 et pRM)En l’absence du répresseur LacI, le gène seront ainsi réprimés, maintenant l’état «Par contre, en présence de LacI, l’opérateur de l’opéron transcription de cI. Les sites opérateurslytique, et la production des phages.
Plasmide contrôle de lyse et génome du phage modifié
Bactériophage lambda
la protéine naturelle à la capside, et des phages recombinés avec la
Identification of a conserved receptor-binding site on the fiber proteins of CAR
Binding Properties, Cell Delivery, and Gene Transfer of Adenoviral Penton Base Displaying
du répresseur cI, système de l’opéron tet:
Pour notre projet, nous avons souhaité modifier notre bactériophage lambda pour Le phage lambda est naturellement tempéré, alternant cycle lytique et
Afin de contrôler l’entrée en phase lytique, nous avons souhaité contrôler la trancriptionle répresseur du cycle lytique. Ce dernier va ainsi bloquer les promoteurs pL et pR,
pression des gènes lytiques. l’expression de cI en continue, puis d’inhiber son expression en temps
nous utilisons le système régulant l’opéron tet d’Escherichia C
se fixe en continu sur l’opérateur tetO, afin de l’inhiber, ce dernier se trouvant trancription est à contrôler. Lors de l’ajout de tétracycline dans le système,
l’inhibition est levé, ce qui permet la trancription du gène collé à l’opérateur Dans notre système, le gène dont nous voulons contrôler l’expression est le répresseur
. Ce dernier, une fois transcrit, ira inhiber l’opérateur de l’opéron Lac, qui sera associé ré de son opérateur et promoteur d’origine OR1 et pRM)
I, le gène cI sera exprimé, les sites opérateurs du génome du phage seront ainsi réprimés, maintenant l’état « non-lytique extrachromosomal ».
I, l’opérateur de l’opéron Lac sera alors réprimé, bloquant ainsi la I. Les sites opérateurs du génome du phage, une fois libérés, initieront le cycle
lytique, et la production des phages.
Plasmide contrôle de lyse et génome du phage modifié
la protéine naturelle à la capside, et des phages recombinés avec la
binding site on the fiber proteins of CAR-recognizing
Binding Properties, Cell Delivery, and Gene Transfer of Adenoviral Penton Base Displaying
Pour notre projet, nous avons souhaité modifier notre bactériophage lambda pour contrôler son Le phage lambda est naturellement tempéré, alternant cycle lytique et
ouhaité contrôler la trancription du gène les promoteurs pL et pR, et donc
e, puis d’inhiber son expression en temps Escherichia Coli :
ce dernier se trouvant Lors de l’ajout de tétracycline dans le système,
du gène collé à l’opérateur tetO. Dans notre système, le gène dont nous voulons contrôler l’expression est le répresseur LacI de
. Ce dernier, une fois transcrit, ira inhiber l’opérateur de l’opéron Lac, qui sera associé ré de son opérateur et promoteur d’origine OR1 et pRM).
opérateurs du génome du phage
sera alors réprimé, bloquant ainsi la , une fois libérés, initieront le cycle
Plasmide contrôle de lyse et génome du phage modifié :

Système sans tétracycline
Système avec tétracycline
Bactériophage lambda
Système sans tétracycline :
Système avec tétracycline :

=>> Switches in bacteriophage lambda development.
Plasmide contrôle de Lyse
c) Séquence Cos extérieur au plasmide phage
Nous avons vu précédemment que les séquences séquence à encapsider. En partant de ce fait, il est possible de réaliser un cosmide. Ce dernier est un plasmide contenant, outre une origine de réplication et un gène sélectionnable, le site cos du bactériophage lambda. 40 à 50 kb d’ADN étranger peuvent être intégrer. On peut dtype de séquence dans la tête de notre bactériophage recombinant. Cependant, aucune donnée n’existe sur la taille minimum de la séquence à introduire
Bactériophage lambda
in bacteriophage lambda development.
c) Séquence Cos extérieur au plasmide phage :
Nous avons vu précédemment que les séquences cos étaient présentes en début et en fin de la
En partant de ce fait, il est possible de réaliser un cosmide. Ce dernier est un plasmide contenant, outre une origine de réplication et un gène sélectionnable, le site cos du bactériophage lambda. 40 à 50 kb d’ADN étranger peuvent être intégrer. On peut donc espérer encapsider n’importe quel type de séquence dans la tête de notre bactériophage recombinant. Cependant, aucune donnée n’existe sur la taille minimum de la séquence à introduire en compagnie des séquences cos.
en début et en fin de la
En partant de ce fait, il est possible de réaliser un cosmide. Ce dernier est un plasmide contenant, outre une origine de réplication et un gène sélectionnable, le site cos du bactériophage lambda.
onc espérer encapsider n’importe quel type de séquence dans la tête de notre bactériophage recombinant. Cependant, aucune donnée
en compagnie des séquences cos.