
Autophagie et maladies respiratoires
5
© 2011 SPLF. Publié par Elsevier Masson SAS. Tous droits réservés. Revue des Maladies Respiratoires Actualités (2011) 3, 461-465 Actualités Maladies Respiratoires Revue des Organe Officiel de la Société de Pneumologie de Langue Française Congrès annuel de l’American Thoracic Society Denver, États-Unis - 13-18 mai 2011 Mission ATS 2011 2011 5 Correspondance. Adresse e-mail : [email protected] (A.-T. Dinh-Xuan). A.-T. Dinh-Xuan Service de physiologie – explorations fonctionnelles, hôpital Cochin et Faculté de Médecine, Université Paris Descartes, Paris, France Autophagie et maladies respiratoires Autophagy and respiratory illnesses L’ autophagie, que l’on peut littéralement traduire par manger (űŞŠŢnIũ, phagein) soi-même (ŞŸůŷ, auto), reÁète la capacité singulière des cellules euca- ryotes à éliminer par digestion enzymatique certains de ses propres constituants. Ce phénomène, encore assez souvent assimilé à de l’autocannibalisme [1], est connu depuis plus de 50 ans. En effet, ce terme a été introduit au milieu du siècle dernier par le médecin biologiste belge, Christian de Duve, avec sa description princeps des mécanismes de dégradation de certains organites intracellulaires par les lysosomes [2], dont la découverte lui a permis de parta- ger par la suite le prix Nobel avec Albert Claude et George- Émile Palade en 1974 (http://nobelprize.org/nobel_prizes/ medicine/laureates/1974/). L’autophagie et ses trois formes biologiques Il existe en réalité non pas une mais trois formes d’autopha- gies [3] : 1) la macro-autophagie ; 2) la micro-autophagie ; et 3) l’autophagie dépendante des molécules chaperon (CMA) (Fig. 1). La macro-autophagie, encore communément appelée autophagie, est la forme la plus anciennement connue (déjà décrite par Christian de Duve dans les années soixante [2]) et la mieux caractérisée. La première étape de ce processus passe par la formation d’une vacuole délimitée par une double membrane, appelée autophagosome, qui séquestre de façon non sélective des macromolécules (agrégats protéiques) et des organites (mitochondries, peroxysomes) (Figs. 1 et 2). L’autophagosome fusionne ensuite avec le lysosome pour former le phago-lysosome au sein duquel les constituants macromoléculaires vont être dégradés par hydrolyse enzymatique (Fig. 2). La macro-autophagie est directement impliquée dans la protection contre les tumeurs et la défense contre les invasions pathogènes (virales et bac- tériennes), et joue un rôle essentiel dans le développement embryonnaire et la différenciation tissulaire. La micro-autophagie a lieu directement au niveau du lyso- some, avec séquestration de petites portions du cytoplasme suite à l’invagination de la membrane du lysosome (Fig. 1). La micro-autophagie est mise en jeu lors de la dégradation des protéines à longue durée de vie. L’autophagie dépendante des molécules chaperon (ADC) est caractérisée par une translocation de la protéine à dégrader qui passe directement du cytosol à l’intérieur du lysosome, à travers la membrane de celui-ci, par l’inter- médiaire de complexes protéiques membranaires capables de reconnaître des motifs spéciÀques de la protéine à dégrader. Du fait de ce passage transmembranaire, seules les protéines solubles sont importées dans le lysosome pour y être dégradées. Les organites intracellulaires (mitochon- dries, peroxysomes) sont ainsi exclus de ce mécanisme de
Transcript of Autophagie et maladies respiratoires

© 2011 SPLF. Publié par Elsevier Masson SAS. Tous droits réservés.
Revue des Maladies Respiratoires Actualités (2011) 3, 461-465ISSN 1877-1203
www.splf.org
Actualités
Maladies
RespiratoiresRevue
des
Organe Officiel de la Société de Pneumologie de Langue Française
Congrès annuel de l’American
Thoracic Society
Denver, États-Unis - 13-18 mai 2011
Numéro réalisé avec le soutien institutionnel du laboratoire
Coordination de la mission ATS de la SPLF : J.-C. Meurice, A.-T. Dinh-Xuan, D. ValeyreCoordination du numéro pour le Comité de Rédaction : A. Cuvelier
Mission ATS 2011
6865
7
OctobreVol 3 2011 N° 5
Correspondance. Adresse e-mail : [email protected] (A.-T. Dinh-Xuan).
A.-T. Dinh-Xuan
Service de physiologie – explorations fonctionnelles, hôpital Cochin et Facultéde Médecine, Université Paris Descartes, Paris, France
Autophagie et maladies respiratoiresAutophagy and respiratory illnesses
L’autophagie, que l’on peut littéralement traduire par manger ( nI , phagein) soi-même ( , auto),re ète la capacité singulière des cellules euca-
ryotes à éliminer par digestion enzymatique certains de ses propres constituants. Ce phénomène, encore assez souvent assimilé à de l’autocannibalisme [1], est connu depuis plus de 50 ans. En effet, ce terme a été introduit au milieu du siècle dernier par le médecin biologiste belge, Christian de Duve, avec sa description princeps des mécanismes de dégradation de certains organites intracellulaires par les lysosomes [2], dont la découverte lui a permis de parta-ger par la suite le prix Nobel avec Albert Claude et George-Émile Palade en 1974 (http://nobelprize.org/nobel_prizes/medicine/laureates/1974/).
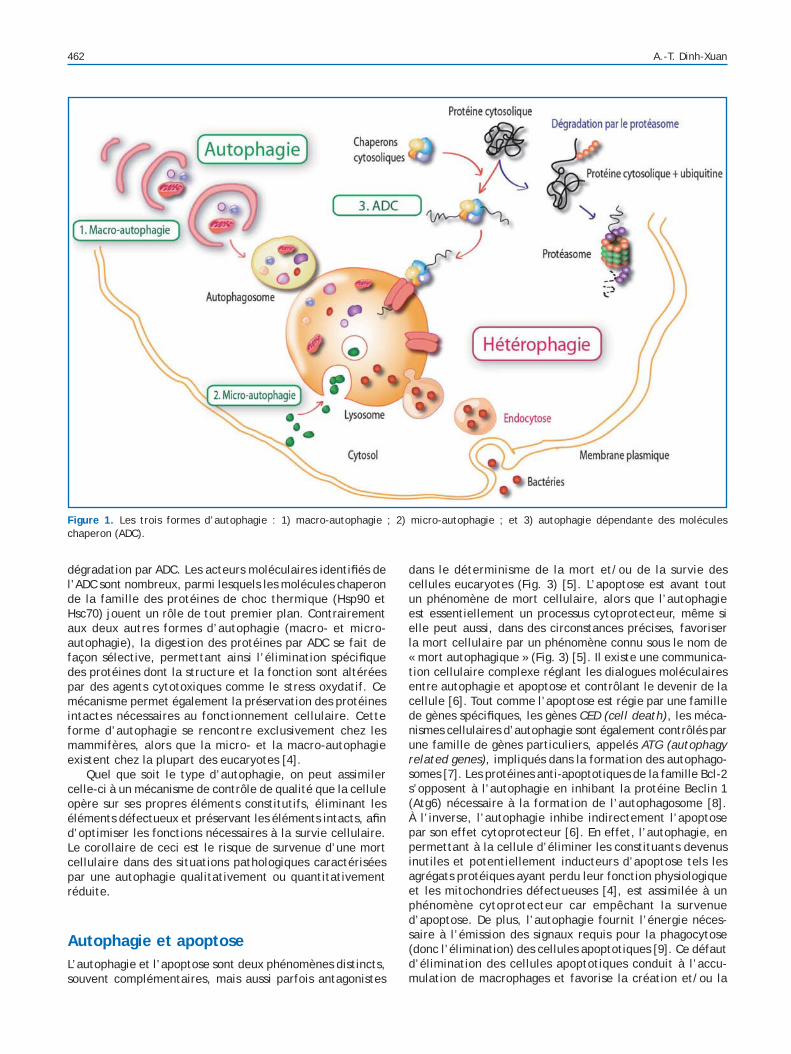
L’autophagie et ses trois formes biologiquesIl existe en réalité non pas une mais trois formes d’autopha-gies [3] : 1) la macro-autophagie ; 2) la micro-autophagie ; et 3) l’autophagie dépendante des molécules chaperon (CMA) (Fig. 1).
La macro-autophagie, encore communément appelée autophagie, est la forme la plus anciennement connue (déjà décrite par Christian de Duve dans les années soixante [2]) et la mieux caractérisée. La première étape de ce processus passe par la formation d’une vacuole délimitée par une
double membrane, appelée autophagosome, qui séquestre de façon non sélective des macromolécules (agrégats protéiques) et des organites (mitochondries, peroxysomes) (Figs. 1 et 2). L’autophagosome fusionne ensuite avec le lysosome pour former le phago-lysosome au sein duquel les constituants macromoléculaires vont être dégradés par hydrolyse enzymatique (Fig. 2). La macro-autophagie est directement impliquée dans la protection contre les tumeurs et la défense contre les invasions pathogènes (virales et bac-tériennes), et joue un rôle essentiel dans le développement embryonnaire et la différenciation tissulaire.
La micro-autophagie a lieu directement au niveau du lyso-some, avec séquestration de petites portions du cytoplasme suite à l’invagination de la membrane du lysosome (Fig. 1). La micro-autophagie est mise en jeu lors de la dégradation des protéines à longue durée de vie.
L’autophagie dépendante des molécules chaperon (ADC) est caractérisée par une translocation de la protéine à dégrader qui passe directement du cytosol à l’intérieur du lysosome, à travers la membrane de celui-ci, par l’inter-médiaire de complexes protéiques membranaires capables de reconnaître des motifs spéci ques de la protéine à dégrader. Du fait de ce passage transmembranaire, seules les protéines solubles sont importées dans le lysosome pour y être dégradées. Les organites intracellulaires (mitochon-dries, peroxysomes) sont ainsi exclus de ce mécanisme de
462 A.-T. Dinh-Xuan
dégradation par ADC. Les acteurs moléculaires identi és de l’ADC sont nombreux, parmi lesquels les molécules chaperon de la famille des protéines de choc thermique (Hsp90 et Hsc70) jouent un rôle de tout premier plan. Contrairement aux deux autres formes d’autophagie (macro- et micro-autophagie), la digestion des protéines par ADC se fait de façon sélective, permettant ainsi l’élimination spéci que des protéines dont la structure et la fonction sont altérées par des agents cytotoxiques comme le stress oxydatif. Ce mécanisme permet également la préservation des protéines intactes nécessaires au fonctionnement cellulaire. Cette forme d’autophagie se rencontre exclusivement chez les mammifères, alors que la micro- et la macro-autophagie existent chez la plupart des eucaryotes [4].
Quel que soit le type d’autophagie, on peut assimiler celle-ci à un mécanisme de contrôle de qualité que la cellule opère sur ses propres éléments constitutifs, éliminant les éléments défectueux et préservant les éléments intacts, a nd’optimiser les fonctions nécessaires à la survie cellulaire. Le corollaire de ceci est le risque de survenue d’une mort cellulaire dans des situations pathologiques caractérisées par une autophagie qualitativement ou quantitativement réduite.
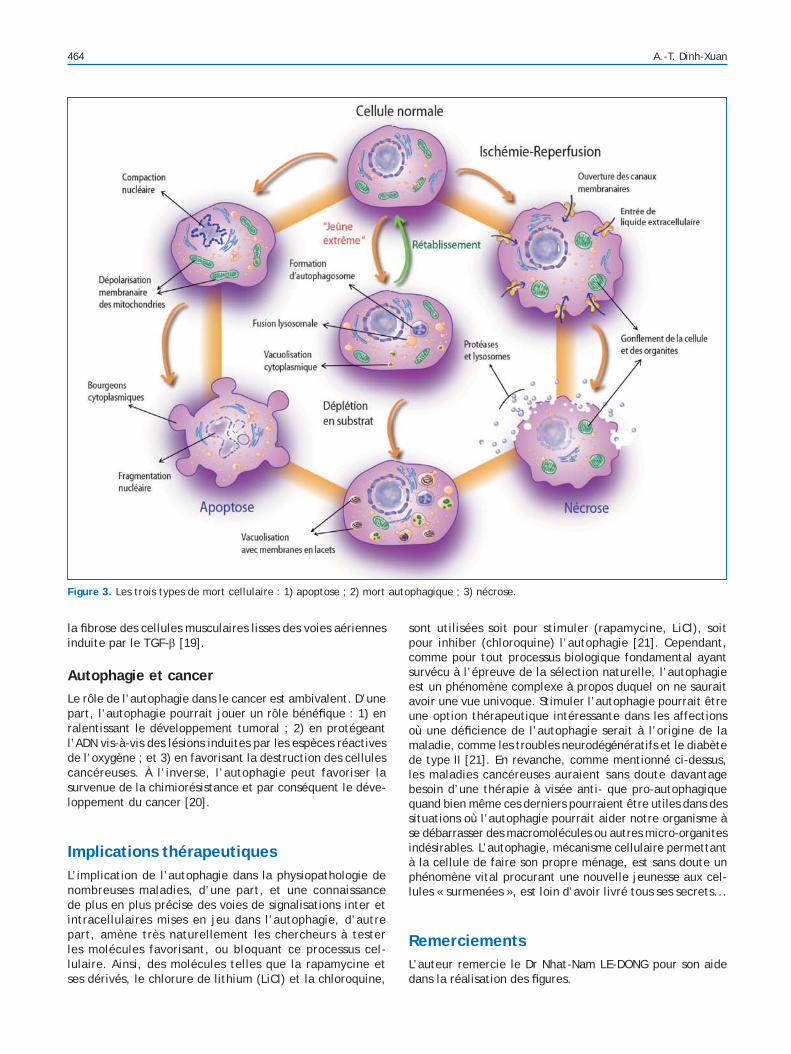
Autophagie et apoptoseL’autophagie et l’apoptose sont deux phénomènes distincts, souvent complémentaires, mais aussi parfois antagonistes
dans le déterminisme de la mort et/ou de la survie des cellules eucaryotes (Fig. 3) [5]. L’apoptose est avant tout un phénomène de mort cellulaire, alors que l’autophagie est essentiellement un processus cytoprotecteur, même si elle peut aussi, dans des circonstances précises, favoriser la mort cellulaire par un phénomène connu sous le nom de « mort autophagique » (Fig. 3) [5]. Il existe une communica-tion cellulaire complexe réglant les dialogues moléculaires entre autophagie et apoptose et contrôlant le devenir de la cellule [6]. Tout comme l’apoptose est régie par une famille de gènes spéci ques, les gènes CED (cell death), les méca-nismes cellulaires d’autophagie sont également contrôlés par une famille de gènes particuliers, appelés ATG (autophagyrelated genes), impliqués dans la formation des autophago-somes [7]. Les protéines anti-apoptotiques de la famille Bcl-2 s’opposent à l’autophagie en inhibant la protéine Beclin 1 (Atg6) nécessaire à la formation de l’autophagosome [8]. À l’inverse, l’autophagie inhibe indirectement l’apoptose par son effet cytoprotecteur [6]. En effet, l’autophagie, en permettant à la cellule d’éliminer les constituants devenus inutiles et potentiellement inducteurs d’apoptose tels les agrégats protéiques ayant perdu leur fonction physiologique et les mitochondries défectueuses [4], est assimilée à un phénomène cytoprotecteur car empêchant la survenue d’apoptose. De plus, l’autophagie fournit l’énergie néces-saire à l’émission des signaux requis pour la phagocytose (donc l’élimination) des cellules apoptotiques [9]. Ce défaut d’élimination des cellules apoptotiques conduit à l’accu-mulation de macrophages et favorise la création et/ou la
Figure 1. Les trois formes d’autophagie : 1) macro-autophagie ; 2) micro-autophagie ; et 3) autophagie dépendante des molécules chaperon (ADC).

Autophagie et maladies respiratoires 463
persistance de foyers in ammatoires tissulaires secondaires à la libération du contenu des corps apoptotiques dans l’espace extracellulaire (nécrose secondaire).
Rôle immunitaire de l’autophagieLe rôle de l’autophagie dans l’immunité, désormais admis [10], est basé sur un ensemble d’arguments expérimentaux. L’autophagie participe à la défense innée de l’organisme en éliminant certains micro-organismes pathogènes, un processus cellulaire encore connu sous le nom de « xénopha-gie » [11,12]. La famille des récepteurs Toll-like, protéines membranaires activées par des antigènes microbiens, stimule l’autophagie. En n, l’autophagie participe à l’immunité adaptative en contrôlant le tra c intracellulaire des peptides provenant de la dégradation d’antigènes cytosoliques et leur présentation par les molécules du complexe majeure d’histocompatibilité de classe II aux lymphocytes T CD4+ [13].
Autophagie et maladies respiratoiresPhénomène biologique connu depuis près d’un demi-siècle [2], et bien qu’étant déjà l’objet d’une brève mise au point dans la Revue des Maladies Respiratoires en 2008 [1], le rôle de l’autophagie dans la physiopathologie des maladies respira-toires ne commence à être mieux connu de la communauté pneumologique que depuis près d’un an [14,15].
Autophagie et BPCOL’autophagie, au même titre que l’apoptose, joue un rôle essentiel dans la physiopathologie de la BPCO [14-16]. La fumée de cigarette provoque la mort autophagique des cellules épithéliales [17] et favorise le développement d’un emphysème en activant une protéine microtubulaire (microtubule-associated protein 1 light chain B, LC3B) codé par le gène Atg6 (de la famille des gènes autophagiques ATG, voir plus haut) [18]. De plus, l’autophagie est impliquée dans
Figure 2. Rôle du lysosome dans la phagocytose et l’autophagie.
464 A.-T. Dinh-Xuan
Figure 3. Les trois types de mort cellulaire : 1) apoptose ; 2) mort autophagique ; 3) nécrose.
la brose des cellules musculaires lisses des voies aériennes induite par le TGF- [19].
Autophagie et cancerLe rôle de l’autophagie dans le cancer est ambivalent. D’une part, l’autophagie pourrait jouer un rôle béné que : 1) en ralentissant le développement tumoral ; 2) en protégeant l’ADN vis-à-vis des lésions induites par les espèces réactives de l’oxygène ; et 3) en favorisant la destruction des cellules cancéreuses. À l’inverse, l’autophagie peut favoriser la survenue de la chimiorésistance et par conséquent le déve-loppement du cancer [20].
Implications thérapeutiquesL’implication de l’autophagie dans la physiopathologie de nombreuses maladies, d’une part, et une connaissance de plus en plus précise des voies de signalisations inter et intracellulaires mises en jeu dans l’autophagie, d’autre part, amène très naturellement les chercheurs à tester les molécules favorisant, ou bloquant ce processus cel-lulaire. Ainsi, des molécules telles que la rapamycine et ses dérivés, le chlorure de lithium (LiCl) et la chloroquine,
sont utilisées soit pour stimuler (rapamycine, LiCl), soit pour inhiber (chloroquine) l’autophagie [21]. Cependant, comme pour tout processus biologique fondamental ayant survécu à l’épreuve de la sélection naturelle, l’autophagie est un phénomène complexe à propos duquel on ne saurait avoir une vue univoque. Stimuler l’autophagie pourrait être une option thérapeutique intéressante dans les affections o une dé cience de l’autophagie serait à l’origine de la maladie, comme les troubles neurodégénératifs et le diabète de type II [21]. En revanche, comme mentionné ci-dessus, les maladies cancéreuses auraient sans doute davantage besoin d’une thérapie à visée anti- que pro-autophagique quand bien même ces derniers pourraient être utiles dans des situations où l’autophagie pourrait aider notre organisme à se débarrasser des macromolécules ou autres micro-organites indésirables. L’autophagie, mécanisme cellulaire permettant à la cellule de faire son propre ménage, est sans doute un phénomène vital procurant une nouvelle jeunesse aux cel-lules « surmenées », est loin d’avoir livré tous ses secrets...
RemerciementsL’auteur remercie le Dr Nhat-Nam LE-DONG pour son aide dans la réalisation des gures.

Autophagie et maladies respiratoires 465
A.-T. Dinh-Xuan : l’auteur déclare n’avoir aucun con it d’intérêt potentiel en rapport avec le thème de l’article.
Références[1] Bonniaud P. Autophagie : autocanibalisme ou autodéfense ?
Rev Mal Respir 2008;25:8-10.[2] De Duve C, Wattiaux R. Functions of lysosomes. Annu Rev
Physiol 1966;28:435-92.[3] Klionsky DJ. Autophagy: from phenomenology to molecular
understanding in less than a decade. Nat Rev Mol Cell Biol 2007;8:931-7.
[4] Mizushima N, Levine B, Cuervo AM, Klionsky DJ. Autophagy ghts disease through cellular self-digestion. Nature 2008;
451:1069-75.[5] Hotchkiss RS, Strasser A, McDunn JE, Swanson PE. Cell death.
N Engl J Med 2009;361:1570-83.[6] Maiuri MC, Zalckvar E, Kimchi A, Kroemer G. Self-eating and
self-killing: crosstalk between autophagy and apoptosis. Nat Rev Mol Cell Biol 2007;8:741-52.
[7] Codogno P. Les gènes ATG et la macro-autophagie. Med Sci (Paris) 2004;20:734-6.
[8] Pattingre S, Tassa A, Qu X, Garuti R, Liang XH, Mizushima N, Packer M, Schneider MD, Levine B. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 2005;122:927-39.
[9] Qu X, Zou Z, Sun Q, Luby-Phelps K, Cheng P, Hogan RN, Gilpin C, Levine B. Autophagy gene-dependent clearance of apopto-tic cells during embryonic development. Cell 2007;128:931-46.
[10] Levine B, Deretic V. Unveiling the roles of autophagy in innate and adaptive immunity. Nat Rev Immunol 2007;7:767-77.
[11] Nakahira K, Haspel JA, Rathinam VA, Lee SJ, Lam HC, Rabi-novitch M, Cernadas M, Kim HP, Fitzgerald K, Ryter SW, Choi
AM. Autophagy proteins regulate innate immune response by inhibiting NALP3 in ammasome-mediated mitochondrial DAN release. Am J Respir Crit Care Med 2011;183:A1077.
[12] Glasser AL, Lapaquette P, Darfeuille-Michaud A. Altération de l’autophagie chez les patients atteints de maladie de Crohn : Une porte ouverte pour des bactéries invasives ? Med Sci (Paris) 2009;25:349-51.
[13] Xu Y, Eissa NT. Autophagy in innate and adaptive immunity. Proc Am Thorac Soc 2010;7:22-8.
[14] Ryter SW, Choi AM. Autophagy in the lung. Proc Am Thorac Soc 2010;7:13-21.
[15] Pandit L, Bonilla DL, Eissa NT. Autophagy: a new frontier in research in lung diseases. Am J Respir Crit Care Med 2011;183:566-8.
[16] Ryter SW, Lam HC, Chen ZH, Choi AM. Deadly triplex: smoke, autophagy and apoptosis. Autophagy 2011;7:436-7.
[17] Chen ZH, Kim HP, Sciurba FC, Lee SJ, Feghali-Bostwick C, Stolz DB, Dhir R, Landreneau RJ, Schuchert MJ, Yousem SA, Nakahira K, Pilewski JM, Lee JS, Zhang Y, Ryter SW, Choi AM. EGR-1 regu-lates autophagy in cigarette smoke-induced chronic obstructive pulmonary disease. PLoS One 2008;3:e3316.
[18] Chen ZH, Lam HC, Jin Y, Kim HP, Cao J, Lee SJ, Ifedigbo E, Parameswaran H, Ryter SW, Choi AM. Autophagy protein micro-tubule-associated protein 1 light chain-3B (LC3B) activates extrinsic apoptosis during cigarette smoke-induced emphy-sema. Proc Natl Acad Sci USA 2010;107:18880-5.
[19] Ghavami S, Yeganeh B, Serebrin A, Mutawe MM, Sharma P, McNeill KD, Stelmack G, Kashani H, Dixon IM, Klonisch T, Nachtigal MW, Halayko AJ. Autophagy regulates TGF-Beta1 induced brosis in human airway smooth muscle cells. Am J Respir Crit Care Med 2011;183:A2110.
[20] Maycotte P, Thorburn A. Autophagy and cancer therapy. Cancer Biol Ther 2011;11:127-37.
[21] Rubinsztein DC, Gestwicki JE, Murphy LO, Klionsky DJ. Poten-tial therapeutic applications of autophagy. Nat Rev Drug Discov 2007;6:304-12.


















