1 Le cytosquelette I - Les principes communs aux trois types de filaments, assemblage,...
95
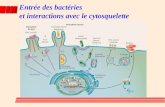
1 Le cytosquelette • I - Les principes communs aux trois types de filaments, assemblage, désassemblage • II - Les protéines associées et leur rôle • III - Les moteurs moléculaires • IV - Fonctions du cytosquelette dans la cellule
-
Upload
lydie-remond -
Category
Documents
-
view
104 -
download
0
Transcript of 1 Le cytosquelette I - Les principes communs aux trois types de filaments, assemblage,...

1
Le cytosquelette
• I - Les principes communs aux trois types de filaments, assemblage, désassemblage
• II - Les protéines associées et leur rôle
• III - Les moteurs moléculaires
• IV - Fonctions du cytosquelette dans la cellule

IV - Fonctions du cytosquelette dans la cellule

3
Coordination des éléments du cytosquelette
• Forme de la cellule
• Déplacement de la cellule
• Division pendant la mitose (cf. infra)

4
Plan
I. La levure comme modèle d'étudeII. Locomotion des cellules (filipodes,
lamellipodes et pseudopodes)III. Action du cytosquelette sur
l'environnementIV. Action de l'environnement sur le
cytosqueletteV. Cas particulier : le neurone

5
I - La levure comme modèle d'étude
• Levure Saccharomyces cerevisiae

6
Asymétrie de Saccharomyces cerevisiae
• Grosse cellule mère + petite cellule fille après division
• Due à l'orientation des faisceaux d'actine• Très peu de microtubules polarité
supportée par l'actine• Deux types d'assemblage de filaments
d'actine– câbles– Patchs associés au cortex (lieux d'endo- et exo-
cytose)

7
Polarité de Saccharomyces cerevisiae
• De la cellule mère naît un bourgeon
• À l'extrémité du bourgeon nombreux câbles d'actine

8
Fig 16-81
• Polarité des patchs et câbles d'actine au cours du cycle cellulaire– Actine-phalloïdine* : cables + patches
Cellule mère avant la formation du bourgeon
Aymétrie des patches et des cables
Naissance du bourgeon
Les patches sont principalement
dans le bourgeon
Les patches sont principalement
dans le bourgeon
Apparition de nouveaux patches
Apparition de nouveaux patches

10
• Fig. 1. Rotating images of 3-D reconstructions of the actin cytoskeleton. (A) A small-budded cell. (B) A budding zygote. Lines point to features of interest. 1, patches that overlap at certain angles. 2, an elongated patch that appears round at a certain angle of view. 3, a cable of uneven thickness along its length. 4, round patches at the ends of thin cables. 5, an elongated patch at the end of a thick cable. 6, a cable in the bud ending on a patch. 7, a cable end that is free but appears to overlap with a patch at certain angles of view. 8, a cable, both ends of which are not associated with a patch. These frames are sequential 2-D projections of rotating images of a vegetative cell and of a zygote with a step size of 24°. Angles of view are marked. The full data set in each case contains 180 frames with sequential projections at 2° angles. The movies are available at the authors' Web site, . Cells were stained with rhodamine-phalloidin. Deconvolution was with the EM method. Bar, 10 µm.
J. Cell Biol., Volume 142, Number 6, September 21, 1998 1501-1517 (Tatiana S. Karpova, James G. McNally, Samuel L. Moltz, and John A. Cooper)

11
• Fig. 2. Filamentous actin disassembly in cells treated with latrunculin. Time points as labeled on the figure. Cells were stained with rhodamine-phalloidin. Bar, 5 µm.
J. Cell Biol., Volume 142, Number 6, September 21,
1998 1501-1517 (Tatiana S. Karpova, James G. McNally, Samuel L. Moltz, and John A.
Cooper)

12
• Fig. 3. Stereopairs of the actin cytoskeleton in sequential stages of the cell cycle (A-D). Arrows indicate cells with thin cables in a random meshwork. Cells were stained with rhodamine-phalloidin. Deconvolution was with the LLS method (A and D) or the EM method (B and C). Bar, 10 µm.
J. Cell Biol., Volume 142, Number 6, September 21,
1998 1501-1517 (Tatiana S. Karpova, James G. McNally, Samuel L. Moltz, and John A.
Cooper)

13
• Fig. 4. Optical sectioning of a large-budded cell with a meshwork of thin cables. The distance (µm) of sequential planes along the z-axis is indicated. This cell is also shown as a stereopair in Fig. 3 D. The cell was stained with rhodamine-phalloidin. Deconvolution was with the PWLS method. Bar, 10 µm.
J. Cell Biol., Volume 142, Number 6, September 21, 1998 1501-1517 (Tatiana S. Karpova, James G. McNally, Samuel L. Moltz, and John A. Cooper)

14
• Fig. 5. Cables after heat shock. (A) Control cells, no heat shock applied. (B) Cells after 5 min of heat shock. (C) Cells after 30 min of heat shock. Single-focal plane images of cells stained with rhodamine-phalloidin are shown. Bar, 10 µm.
J. Cell Biol., Volume 142, Number 6,
September 21, 1998 1501-1517 (Tatiana S. Karpova, James G. McNally, Samuel L. Moltz, and John A.
Cooper)

15
• Fig. 6. The actin cytoskeleton in cap2 (A) and tpm1 (B and C) mutants. Single-focal plane images of cells at different stages of the cell cycle are presented in A and B. Asterisks mark the cells with partially delocalized patches, in which cables are thinner and less well oriented than the cables in wild-type cells. C shows a stereopair of a tpm1 cell. A rotating movie of the data set for C, with 180 frames at sequential projections of 2° angles, is available at the authors' Web site, . Cells were stained with rhodamine-phalloidin. Deconvolution was with the EM method (C). Bar, 10 µm
J. Cell Biol., Volume 142, Number 6, September 21,
1998 1501-1517 (Tatiana S. Karpova, James G. McNally, Samuel L. Moltz, and John A.
Cooper)
(A) The actin cytoskeleton in cap2 mutant : single-focal plane images of cells at different stages
(B) The actin cytoskeleton in tpm1 mutant : single-focal plane images of cells at different stages
(C) The actin cytoskeleton in tpm1 mutant : stereopair of a tpm1 cell
*cells with partially
delocalized patches, in which
cables are thinner and less well
oriented than the cables in wild-type
cells
Cells were stained with rhodamine-phalloidin.

16
• Fig. 7. The actin cytoskeleton of the bee1/las17 mutant. (A) Single-focal plane images of four cells at different stages of the cell cycle with a completely random cable meshwork. (B) A stereopair of another such cell. Movie 3 shows rotating images of this cell, with 180 frames at sequential projections of 2° angles. Movies are available at . (C) Single- focal plane images of two cells with partial orientation of cables. Cells were stained with rhodamine-phalloidin. Deconvolution was with the EM method. Bar, 10 µm.
J. Cell Biol., Volume 142, Number 6, September 21, 1998 1501-1517
(Tatiana S. Karpova, James G. McNally, Samuel L.
Moltz, and John A. Cooper)
(A) Single-focal plane images of four cells at different stages of the cell cycle with a completely random cable meshwork.
(B) A stereopair of another such cell. Movie 3 shows rotating images of this cell, with 180 frames at sequential projections of 2° angles.
C) Single- focal plane images of two cells with partial orientation of cables.
The actin cytoskeleton of the bee1/las17 mutant.
Cells were stained with rhodamine-phalloidin. Bar, 10 µm.

17
• Fig. 8. Diagram summarizing results testing for correlations between patch and cable rearrangement
J. Cell Biol., Volume 142, Number 6, September 21,
1998 1501-1517 (Tatiana S. Karpova, James G. McNally, Samuel L. Moltz, and John A.
Cooper)
Diagram summarizing results testing for correlations between patch and cable rearrangement

18
• Fig. 9. The random meshwork of thin cables in large-budded cells. A-C show three examples. D shows three small-budded polarized cells for comparison. Single-focal plane images of cells stained with rhodamine-phalloidin are shown. Bar, 5 µm.
J. Cell Biol., Volume 142, Number 6, September 21,
1998 1501-1517
(Tatiana S. Karpova, James G. McNally,
Samuel L. Moltz, and John A.
Cooper)
The random meshwork of thin cables in large-budded cells. Single-focal plane images of cells stained with rhodamine-phalloidin are shown. Bar, 5 µm.
A-C show three examples
D shows three small-budded polarized cells for comparison

19
• Fig. 10. Cables in zygote formation and development. (A) Cells with mating projections after 1 h of -factor treatment. (B) Cells with mating projections after 3 h of -factor treatment. (C- E) Sequential stages of zygote formation and development. Single-focal plane images of cells stained with rhodamine-phalloidin are shown. Bar, 10 µm.
J. Cell Biol., Volume 142, Number 6,
September 21, 1998 1501-1517 (Tatiana S. Karpova, James G. McNally, Samuel L. Moltz, and John A.
Cooper)
(A) Cells with mating projections after 1 h of -factor treatment
(B) Cells with mating projections after 3 h of -factor treatment
(C- E) Sequential stages of zygote formation and development
Cables in zygote formation and development. Single-focal plane images of cells stained with rhodamine-phalloidin are shown. Bar, 10 µm.

20
Levure Saccharomyces cerevisiae
• Forme haploïde– Prolifère– Si conditions difficiles doivent s'accoupler pour
devenir diploïde et sporuler – Facteurs d'accouplement a et (récepteurs
correspondants)
• Forme diploïde– Seule peut sporuler si conditions difficiles

21
Facteurs d'accouplement (a ou )
• Se lient à la surface de la cellule (haploïde)
• Entraîne la polarisation de la cellule • Modification de la forme de la cellule en
"shmoo" • Préparation à l'accouplement

22
Fig 16-82
• Saccharomyces cerevisiae devenant polarisé après traitement par le facteur d'accouplement a ou

23
Fig 16-83
• Voie de signalisation de la réponse de la levure au facteur d'accouplement
GTPase

24
Transmission au réseau de microtubules
• Polymérisation de l'actine • COMT (inclus dans la membrane nucléaire
chez S. cerevisiae) se place sur le côté du noyau près de l'extrémité du shmoo
• Alignement du noyau lui permettant de fusionner avec son partenaire

25
Fig16-50
• Effets de Rac, Rho et Cdc42 sur l'organisation de l'actine dans le fibroblaste

26
Même mécanisme pour la croissance du bourgeon qui deviendra une cellule fille
• Construction d'un nouveau bourgeon juste à côté du bourgeon précédent
• Intervention de Cdc 42 pour la transmission du message au cytosquelette

27
Localisation des molécules dans la cellule
• ARN
• Protéines

28
Exemple de la protéine de régulation de gène Ash1
• Le messager de Ash1 et sa protéine sont localisés exclusivement dans le bourgeon en croissance et ne se retrouveront donc que dans la cellule fille
• Nécessité d'une des deux myosines V pour cette distribution asymétrique de l'ARNm de Ash1
• Nécessité d'au moins 6 autres produits de gène pour cette polarité (tropomyosine, profiline, actine, protéines intermédiaires entre ARN et myosine V,[She2 & She3])

29
Fig 16-84
• Localisation d'ARNm à l'extrémité du bourgeonnement de la levure (répartition asymétrique de l'ARNm de la protéine ash1)
ARNm de ash1

30
Autres applications du rôle du cytosquelette dans la
polarité de la cellule• Embryon de drosophile : axe antéro-
postérieur–ARN localisés à l'extrémité
postérieure uniquement grâce au…
–… transport des ARN le long de microtubules

31
II – Locomotion des cellules (filipodes, lamellipodes et
pseudopodes)

32
Presque toutes les cellules sont capables de migrer sur un support• Amibe
• Exception : le spermatozoïde
• Migrations chez l'embryon
• Cellules de la crête neurale cellules pigmentaires de la peau, cellules ganglionnaires,
• Macrophages, neutrophiles, ostéoclastes, fibroblastes, entérocytes de la villosité intestinale, cellules cancéreuses, …

33
Mécanismes de la locomotion de la cellule
• A – Expansion = protrusion de structures riches en actine
• B - Attachement
• C - Traction

34
Fig 16-85
• Forces déployées au cours du déplacement de la cellule– Parfois
impression de mouvement continu
– Parfois impression de mouvement irrégulier

35
Forces déployées au cours du déplacement de la cellule
• Protrusion du bord avant par polymérisation de l'actine
• Fixation du lamellipode au support force d'étirement sur le cortex d'actine contraction à l'arrière pour soulager la tension• Création de nouveaux contacts focaux à l'avant• Désassemblage de contacts focaux à l'arrière • Déplacement de la cellule sur le support

36
A - Rôle de l'actine dans la création d'expansions membranaires
• Filopodes (= microspikes = spicules)
• Lamellipodes (les plus étudiés)
• Pseudopodes

37
Les différentes formes de "bord avant"
• Filipodes, lamellipodes, pseudopodes
• La structure est fonction de la cellule
• Rempli de faisceau d'actine pas d'organites
• 1, 2 ou 3 –D (cf supra)

38
Filopodes
• Cônes de croissance des neurones
• Fibroblastes
• Ressemblent à des microvillosités mais plus longs, plus fins et plus dynamiques
• 1-D

39
Lamellipodes
• Cellules épithéliales de l'épiderme de poisson ou grenouille = keratocyte
• Fibroblastes
• Certains neurones
• Damier orthogonal de filaments d'actine parallèle au support
• Les plus faciles à étudier
• 2-D

40
Pseudopodes
• Amibes et neutrophiles
• Gel de filaments d'actine
• 3-D

41
Les "kératocytes"
• Cellules de l'épiderme des poissons et de la grenouille
• Riches en filament de kératine
• Peuvent se déplacer très rapidement pour réparer une blessure (jusqu'à 30 µ/minute)
• Aspect particulier en culture– Grand lamellipode– Petit corps cellulaire contenant le noyau et non
adhérent au support

42
Fig 16-86
• Kératocytes en migration (épiderme de poisson)
Culture toutes les 15 sec (15 m/sec)
MEBActineMicrotubulesFilaments intermédiaires
Petit corps cellulaire contenant le noyau et non adhérent au support

43
Fig 16-87
Comportement de fragment de lamellipode séparé par microchirurgie
A Continue à migrer avec la même organisationActine réseau dans le bord avantMyosine II bande à l'arrière
B S'arrête de migrer et devient rondActine répartie en périphérieMyosine II spots dans tout le cytoplasme

44
Verkovski,AB1999p11
Verkovski,AB1999p11 : Cytoskeletal organization of (a–d) a keratocyte fragment undergoing locomotion and (e–g) a stationary keratocyte fragment. The distribution of myosin II (red), as revealed by antibody staining, and that of actin (cyan), as revealed by staining with tetramethylrhodamine–phalloidin, is shown (a,e). (b–d,f,g) Electron microscopy of platinum replicas. (b) An overview and (c,d) regions of the moving fragment reveal a crisscross actin network at the leading edge (c) and a bundle at the trailing edge (d). A cluster of myosin II minifilaments revealed after the extraction of actin from the cytoskeleton of a similar fragment is shown in the inset in (d) at an approximate position that it would occupy in the intact cytoskeleton. (f) An overview and (g) a region of a stationary fragment reveals a narrow actin brush (indicated with brackets) at the edge of the fragment. (h) Triple staining for actin (cyan), myosin (red) and tubulin (yellow) in several fragments reveals the presence of few or no microtubules in the fragments. Bars represent 2 mm in (a,b,h) and 200 nm in (c).

45
Fig 16-88
• Nucléation des filaments d'actine et formation du réseau par le complexe ARP dans un lamellipode de kératocyte
• Actine (rouge) + ARP (vert) jaune sur le bord avant
• tapis roulant s'assemblant en avant et se désassemblant en arrière
• (C) FG de (B) embranchement à 70° du complexe ARP
ActineARP Actine + ARP

46
Comportement du réseau d'actine
• Étude dans le lamellipode du kératocyte
• Les filaments d'actine restent fixes par rapport au support au fur et à mesure que le kératocyte se déplace
• Extrémité– Extrémité + : distale vers l'avant– Extrémité – : fixée au complexe ARP
• Impression de tapis roulant de l'ensemble du bord avant

47
Rôle de la cofiline(Actine Depolymeristion Factor)
• Le nucléation a lieu au bord avant et pousse la membrane plasmique
• La dépolymérisation a lieu en arrière
• Cofiline se lie préférentiellement à la forme D (ADP) de l'actine le bord avant (riche en T-actine) résiste à la dépolymérisation par cofiline
• Le filament vieillit hydrolyse de l'ATP désassemblage du filament par la cofiline

48
Fig 16-89
• Cofiline (=actine depolymerizing factor) dans les lamellipodes de kératocyte
Actine
Cofiline
Actine + cofiline = Actine-Cofiline(en jaune)

49
Fig 16-90
• Protrusion du réseau d'actine au bord avant
Nucléation médiée par les complexes ARP du bord avant

50
• Fig. 1. Multiple branching of actin filaments in lamellipodia. EM of lamellipodia of Xenopus keratocytes (a-g) and fibroblasts (h-o) showing overviews of the leading edge (a and h) and enlargements of the boxed regions (b-g and i-o). Many examples of filaments with tightly spaced multiple branches (cyan) can be visualized in lamellipodia despite the high overall density of the actin network. Bar, 0.5 µm.
Svitkina,TM1999p1009
Kératocyte Fibroblaste

51
• Fig. 2. Improved visualization of actin filament branching in lamellipodia. EM of keratocyte or fibroblast lamellipodial actin network after CD treatment (a and b, 0.2 µM for 30 min or 0.5 µM for 10 min), 1 min recovery from serum starvation of a mouse fibroblast (c), LA treatment (0.2 µM for 10 min), or unprotected extraction (d). All examples demonstrate frequent branching of actin filaments. Bars, 0.1 µm; b and d are shown at the same magnification as c
Svitkina,TM1999p1009Embranchements de filaments d'actine

52
• Fig. 3. Localization of Arp2/3 complex in lamellipodia. (a-c and e-g) Fluorescence microscopy of Xenopus keratocyte (a-c) or fibroblast (e-g). Staining with p21 antibody (green) and TRITC-phalloidin (red) shows ARP2/3 complex highly enriched in lamellipodia. Boxed region in g is enlarged in insets; it shows several filopodia lacking and only one filopodium containing Arp2/3 complex. (d and h) Immuno-EM of lamellipodia of Xenopus keratocyte (d) or fibroblast (h) stained with p21 primary antibody and 10-nm gold-conjugated secondary antibody after glutaraldehyde fixation and SDS treatment of detergent-extracted cells. Gold particles are highlighted in yellow. Bars: (a and e) 10 µm; (d and h) 0.1 µm
Svitkina,TM1999p1009
Kera
tocy
te X
en
opu
s fib
robla
ste
Lamellipode riche en ARP
Lamellipode riche en ARP

53
• Fig. 4. Localization of Arp2/3 complex at actin filament branching points. Xenopus keratocytes and fibroblasts were treated with CD (0.2 µM for 30 min or 0.5 µM for 10 min), extracted in the presence of phalloidin, fixed with glutaraldehyde, treated with 33% methanol, and immunostained with p21 antibody followed by 10-nm gold-conjugated secondary antibody. Gold particles are highlighted in yellow. Bar, 50 nm
Svitkina,TM1999p1009

54
• Fig. 5. Localization of cross-linking proteins in fibroblast cytoskeleton. (a-c) Fluorescence microscopy and corresponding intensity profiles (a'-c') of Xenopus (a and c) or human 356 (b) fibroblast lamellipodia double stained with TRITC-phalloidin (red) and either p21 (a and a'), ABP-280 (b and b'), or -actinin (c and c') antibodies (green). The protein/actin ratio at the leading edge of the lamellipodium is high for Arp2/3 complex (a and a'), medium for ABP-280 (b and b'), and low for -actinin (c and c') compared with internal actin structures. (d-i) Immuno-EM of the cell edge (d-f) or interior (g-i) of CD-treated Xenopus (d, f, g, and i) or human 356 (e and h) fibroblasts stained with p21 (d and g), ABP-280 (e and h), or -actinin (f and i) primary antibody and 10-nm (d, e, g, and h) or 18-nm (f and i) gold-conjugated secondary antibody. Gold particles (yellow) reveal Arp2/3 complex at Y-junctions at cell edge and ABP-280 and -actinin at filament crossovers in the cell interior. Bars: (a-c) 1 µm; (d-i) 0.1 µm
Svitkina,TM1999p1009

55
• Fig. 6. Structural differentiation of actin network in lamellipodium. EM of Xenopus keratocytes (a-c) or Xenopus fibroblasts (d-f) after regular extraction in the presence of PEG and phalloidin (a and d) or after unprotected extraction without PEG and phalloidin (b, c, e, and f). Boxed areas from b and e are enlarged in c and f, respectively. Actin network at lamellipodial rear disassembled in the course of unprotected extraction, whereas front zone remained as dense as in control cells. Bar, 1 µm
Svitkina,TM1999p1009

56
• Fig. 7. Differential response of lamellipodial actin network to LA. (A) Phase-contrast sequence of a locomoting Xenopus keratocyte. After addition of 0.1 µm LA at 9 min time point, the cell continued to translocate, retaining the crescent-like shape. (B) Plot showing rate of front edge protrusion versus time of the cell shown in A. LA addition (arrow) decreased rate of protrusion from ~4 µm/min before LA application to 1 µm/min at the end of the sequence. (C) Fluorescence microscopy of the boxed region of the cell shown in A, which was lysed at 34 min time point, fixed, and stained with TRITC-phalloidin. Actin-staining reveals narrow bright lamellipodium at the leading edge, separated by the wide actin-depleted zone from the internal actin structures. (D and E) EM of a Xenopus keratocyte (D) or Xenopus fibroblast (E) lamellipodium treated with LA (D, 0.1 µM for 30 min; E, 0.25 µM for 10 min) reveals actin depletion from the lamellipodial rear. Bars: (A) 10 µm; (D and E) 1 µm
Svitkina,TM1999p1009

57
Svitkina,TM1999p1009
• Fig. 8. Localization of XAC in Xenopus keratocytes. (a-e) Fluorescence microscopy of a whole cell (a-c) and the intensity profile (d) of the enlarged lamellipodium (e) from the boxed region in c, double stained with XAC antibody (green) and TRITC-phalloidin (red). XAC in lamellipodium is excluded from the narrow zone at the extreme leading edge. (f) Phase-contrast sequence of a locomoting cell; time is shown in min. (g-i) Immuno-EM with XAC antibody of the cell shown in f, which was lysed and processed at 8 min time point. (g) Cell overview. (h) Intermediate magnification of the boxed region from g showing distribution of gold particles (yellow) in lamellipodia. (i) High magnification of the boxed region from d showing leading edge. XAC is excluded from the extreme front of the lamellipodium (h and i). Bars: (a and f) 10 µm; (h) 10 µm

58
• Fig. 9. Localization of XAC to posterior regions of depolymerization-resistant actin brush. Electron (a and c) and fluorescence (b) microscopy of lamellipodia of Xenopus keratocytes after unprotected extraction (a) or LA treatment (b, 0.15 µM for 30 min; c, 0.25 µM for 10 min) and subsequent staining with XAC antibody (a and c) or double staining with TRITC-phalloidin and XAC antibody (b).
Svitkina,TM1999p1009

59
• Fig. 10. Localization of XAC in Xenopus fibroblasts. (a-e) Fluorescence microscopy of a cell fragment (a-c) and the intensity profile (d) of the enlarged lamellipodium (e) from the boxed region in c, double stained with XAC antibody (green) and TRITC-phalloidin (red). XAC is distributed throughout the entire lamellipodium. (f and g) Phase-contrast sequence showing protruding lamellipodium of a motile cell (f) and the overview of the same cell (g) at 80 s time point. (h and i) Immuno-EM with XAC antibody of the cell shown in g. (h) Cell overview. Nucleus and surrounding regions look bright because of high content of cellular proteins, which creates high electron density (brightness in inverted contrast). Immunogold-labeling in these central regions is very low. (i) High magnification of the protruding region pointed to by arrow in h. Gold particles (yellow) revealing XAC localization are found at the extreme leading edge, as well as throughout the lamellipodia
Svitkina,TM1999p1009

60
• Fig. 11. Two treadmilling models for actin turnover in lamellipodia. Left, treadmilling of individual filaments suggests that each actin filament in the actin network simultaneously assembles subunits at its barbed end and releases subunits from the pointed end, thus continuously reproducing itself by treadmilling mechanism. Treadmilling of individual filaments collectively results in the treadmilling of the lamellipodial network. Right, treadmilling of the dendritic array suggests frequent formation of new filaments by de novo nucleation, which occurs within the narrow zone at the leading edge, the actin brush. Newborn filaments become immediately incorporated into the actin array as branches of pre-existing filaments. Within the actin brush, filaments are protected from depolymerization at pointed ends. Nucleation, cross-linking, and pointed end capping are proposed to be mediated by the Arp2/3 complex. Many barbed ends are predicted to be capped to prevent exponential increase in filament mass. Release of the Arp2/3 complex from Y-junctions behind the actin brush, followed by ADF/cofilin-mediated dissociation of actin subunits from pointed ends, may be a major pathway for actin array disassembly. By this model, an individual filament in the dendritic array does not treadmill, but rather first grows at the barbed end and later shrinks at the pointed end. However, the actin filament array as a whole treadmills, reproducing itself at the cell front and dismantling itself at the lamellipodial rear. Growing barbed ends are shaded in gray
Svitkina,TM1999p1009

61
Modification de l'orientation des filaments d'actine
• Intervention de filaments bipolaires de myosine II
• Les filaments d'actine deviennent parallèles au bord avant
• Évite la formation d'une protrusion

62
Les cellules se déplacent par B - adhésion et C - traction
• Rôle très important de l’environnement sur la locomotion donc de l’adhésion
• Adhésion est inversement proportionnel à migration– kératocyte : faiblement adhérent lamellipode
facile à créer– neurone de Aplysia : très adhérent gros
lamellipode ne permettant pas la migration– mais même cycle pour l'actine l’actine se
retrouve en arrière de la cellule

63
Fig 16-91
• Mouvement rétrograde du réseau d'actine dans un lamellipode (neurones de la limace Aplysia) – (A) Culture sur un support
collant• Organites et MT localisé à
l’arrière• Réseau de filaments d’actine
dans le lamellipode
– (B) Traitement par la cytochalasine (capping des +) le réseau de filaments d’actine dans le lamellipode se retire
– (C) id (B)actine – phalloïdine

64
Répartition de l’actine
• Si support très collant actine concentrée en arrière de la cellule (eg neurone de Aplysia)
• Si support peu adhérent actine dans les lamellipodes (eg kératocyte)
• En général entre ces deux extrêmes la répartition de l’actine dépend de
l’environnement

65
Sites d’attachement
• Se forment en avant du lamellipode• Identiques aux contacts focaux• Si absence de fixation du
lamellipode, bascule sur le bord supérieur de la cellule et forme des ondulations

66
Fig 16-92
Lamellipodes et ondulations au bord avant d’un fibroblaste
Mouvement de « vagues » vers l’arrière en cas
d’échec d’attachement du lamellipode

67
Rôle des contacts focaux
• Sites d’ancrage sur le support pour tirer le corps cellulaire en avant
• Rôle de la myosine II comme moteur
• Moteurs … fonctionnement mal connu ...

68
Fig 16-93
• Localisation de la myosine I et II dans l'amibe Dictyostelium– Myosine I localisée
dans le bord avant du pseudopode
– Myosine II localisée dans le cortex postérieur riche en actine
Myosine I
Myosine II
5m

69
Rôle des myosines
• Myosine I pour projeter le bord avant• Myosine II pour faire avancer le corps
cellulaire• Amibe Dictyostelium déficiente en
myosine II– Peut pousser des pseudopodes– Mais ramène lentement le reste du corps
cellulaire… comment ?? affaiblissement des contacts focaux à la suite de l'étirement ?

70
III - Action du cytosquelette sur l'environnement

71
Fig 16-94
• Forces de traction exercées par les cellules sur le support : plissement du support de silicone

72
Fig 16-95
• Explants de deux cœurs de poulet sur un gel de collagène au bout de 4 jours
– modification de la matrice

73
IV - Action de l'environnement sur le cytosquelette
• Polarisation = préalable indispensable à la locomotion de la cellule
• Polarisation organisation pluri cellulaire• Réponse à des modifications de
l'environnement– physique
– ou chimique chimiotactisme

74
Chimiotactisme
• "Mouvement d'une cellule dans une direction donnée contrôlée par un gradient d'une substance chimique diffusible"
• eg : mouvement du neutrophile vers une source bactérienne – Des récepteurs à la surface des neutrophiles
peuvent détecter (différence de 1% de chaque côté de la cellule) de très faibles concentrations de peptides N- formylés (N-formyl-méthionine)

75
Fig 16-96
Polarisation et chimiotactismedu neutrophile
Libération par la pipette de formyl-Met-Leu-Phe apparition d'un lamellipode vers la source du chémoattractant
Agrandissement du lamellipode et polarisation du cytosquelette. Localisation de la myosine II à l’arrière de la cellule
Déplacement de la cellule vers le peptide

76
Interprétation moléculaire
• Fixation du ligand à un récepteur– Diffusible (chimiotactisme)– Non diffusible (molécules fixées sur la matrice)
• Activation d’une GTPase de la famille Rho
• Polymérisation d’actine
• Création d’une expansion
• Migration de la cellule dans cette direction

77
Rôle des microtubules• La polymérisation d’actine dans le bord avant • Activation de moteurs qui tirent sur le centrosome via
les microtubules • Position du centrosome• Qui nuclée de nouveaux microtubules dont les
extrémités + vont dans le bord avant • Modification de l’adhésion locale • Activation de Rac GTPase • Augmentation de la polymérisation d’actine dans le bord
avant • Allongement de ce bord avant • Feed back positif • Renforcement de la polarité de la cellule • Actine agit sur les MT qui agissent en retour sur l’actine

78
Autres exemples
• Mort d’une cellule cible par les lymphocytes T cytotoxiques
• Reconnaissance de l’antigène par des récepteurs de la surface des lymphocytes T
• Activation des GTPases de la famille Rho • Polarisation de l’actine corticale sous la zone de contact • Réorientation du centrosome vers la zone de contact • Repositionnement du Golgi sous la zone de contact

79
Fig 16-97(A)
• Modifications du cytosquelette d'un lymphocyte T cytotoxique à la suite d'un contact avec une cible (modification du cytosquelette des deux cellules)

80
Fig 16-97(B)
Cellule T
Cellule cible
• Cellule T et cible colorées par un AC anti-microtubule– Dans la cellule T
irradiation des microtubules dirigés vers le point de contact entre les deux cellules
– Dans la cellule cible le réseau de microtubules n’est pas polarisé
10 m

81
V – Cas particulier : le neurone
• Les grosses différences entre pro et eucaryote sont des conséquences du cytosquelette– Pro : transport par diffusion– Eu : transport par moteurs
cellules plus grosses et plus complexes
• Le plus gros (long) et le plus complexe : le neurone

82
Le neurone
• Cellule embryonnaire
• Migration +++
• Apparition de longs prolongements (= neurites)– Axones– Dendrites– Remplis de microtubules

83
Axone
• Tous les microtubules sont orientés selon la même direction– Extrémité - vers le corps cellulaire– Extrémité + vers l’extrémité de l’axone
• Un microtubule quelques microns :un microtubule ne traverse pas tout l’axone d’un bout à l’autre : il y a chevauchement
• Autoroute pour le transport des protéines et des vésicules ( vésicules synaptiques)

84
Dendrites
• Beaucoup plus courts que l'axone
• Reçoivent les signaux
• Les microtubules sont parallèles mais de polarité mixte
• Se forment comme les axones

85
Vésicules synaptiques
– Utilité à l’extrémité des axones– Construites dans le corps cellulaire et les
dendrites où sont les ribosomes– Très grande longueur (jusqu’à 1 mètre)

86
Transport axonal antérograde
• Concerne les mitochondries, les protéines, les vésicules synaptiques
• Se fait le long des microtubules
• Par des moteurs moléculaires de la famille des kinésines– Jusqu'à un mètre par jour– Sinon 8 ans pour une mitochondrie !
• Nombreux types de moteur spécificité du transport

87
Transport axonal rétrograde
• Concerne le retour des vieux organites
• Se fait le long des mêmes microtubules
• Par des moteurs moléculaires de la famille des dynéines cytoplasmiques (moteur qui se déplace dans le sens moins)

88
Fig 16-98
• Organisation des microtubules dans les fibroblastes et les neurones
Tous les MT ont la même polarité
Les MT ont une polarité mixte

89
L'organisation des neurones dépend du cytosquelette
• Actine, filaments intermédiaires, microtubules
• Actine– Localisée sous le cortex de l'axone– Moteurs de myosine V
• Filaments intermédiaires– = neurofilaments– Rôle de structure– Attachés aux microtubules et aux filaments
d'actine par des protéines

90
Développement du neurone(= neurogenèse) (1/4)
• Repose sur la mobilité liée à l'actine
• Apparition d'un cône de croissance : structure mobile riche en actine

91
Fig 16-99
• Croissance du cône de croissance neuronal
Deux cônes à l'extrémité d'un neurite
Neurite
Nombreux filopodes et
un gros lamellipode
Aspect raide du neurite dû àla traction de l'extrémité
Cône de croissance d'un neurone sensoriel à la surface interne de l'épiderme

92
Développement du neurone(= neurogenèse) (2/4)
• Les cônes de croissance produisent des filopodes voire des lamellipodes
• Ces filopodes sont stabilisés exclusivement par l'environnement– Nétrine protéine soluble qui attire ou repousse
les cônes de croissance– Marqueurs de surface sur la matrice extra
cellulaire ou les cellules

93
Développement du neurone(= neurogenèse) (3/4)
• Formation rapide de contacts focaux • Collapsus du réseau d'actine des cônes non
stabilisés
• Combinaison complexe de signaux positifs et négatifs, solubles et insolubles

94
Développement du neurone(= neurogenèse) (4/4)
• Les microtubules suivent avec leur instabilité dynamique
• Stabilisé par les signaux d'adhésion (contacts focaux)

96
Fig 16-100
• Complexité de l'architecture d'un neurone (rétine de singe)

97
Conclusion
Le cytosquelette est à la base de la construction du neurone, donc du système nerveux