







Langages
Pages
Légal


Transfert d’électrons et oxydations phosphorylantes en un clin d’œil
-
Dans chaque cellule se déroulent des centaines de réactions chimiques, qui
constituent le métabolisme. Les composés chimiques impliqués sont appelés
métabolites.
-
Pratiquement toutes ces transformations ne se dérouleraient que très lentement en
dehors de la cellule et de plus dans toutes les directions. Des successions
ordonnées de réactions chimiques avec une vitesse élevée (= voies métaboliques),
ne sont possibles que grâce à
l’existence d’enzymes spécifiques.
A.
Vue d’ensemble sur le métabolisme intermédiaire-
La plupart des cellules et des organismes ont en commun un ensemble de voies
métaboliques centrales.
-
Ces voies qui servent à
la synthèse, la dégradation
et la transformation de
métabolites importants
ainsi qu’à
la conservation de l’énergie, forment le
métabolisme intermédiaire, dont nous allons examiner ici un schéma simplifié.

-
Pour vivre, les cellules ont besoin en permanence de matières organiques et
inorganiques ainsi que d’énergie chimique, qu’elles tirent essentiellement de l’ATP.
-
Selon la façon dont ces besoins seront couverts, on distingue les organismes
autotrophes
ou hétérotrophes:
Les espèces autotrophes
(végétaux et nombreux micro-organismes), peuvent
synthétiser des molécules organiques à
partir de
précurseurs inorganiques
(CO2
).
Les espèces hétérotrophes
(animaux et champignons), dépendent de l’apport de
substances organiques par la nourriture.
-
La plupart des substances nutritives (protéines, sucres, acides nucléiques et
lipides) ne peuvent être utilisées directement →
doivent d’abord être dégradées
en éléments plus petits
par des voies cataboliques
(flèches roses, Figure 1).
-
Les métabolites ainsi formés (= compartiment métabolique) vont:1) soit aboutir à
la formation d’énergie
après de nouvelles transformation
cataboliques
2) soit être de nouveau intégrés dans des molécules complexes
par des voies
anaboliques
(flèches bleues, Figure 1).
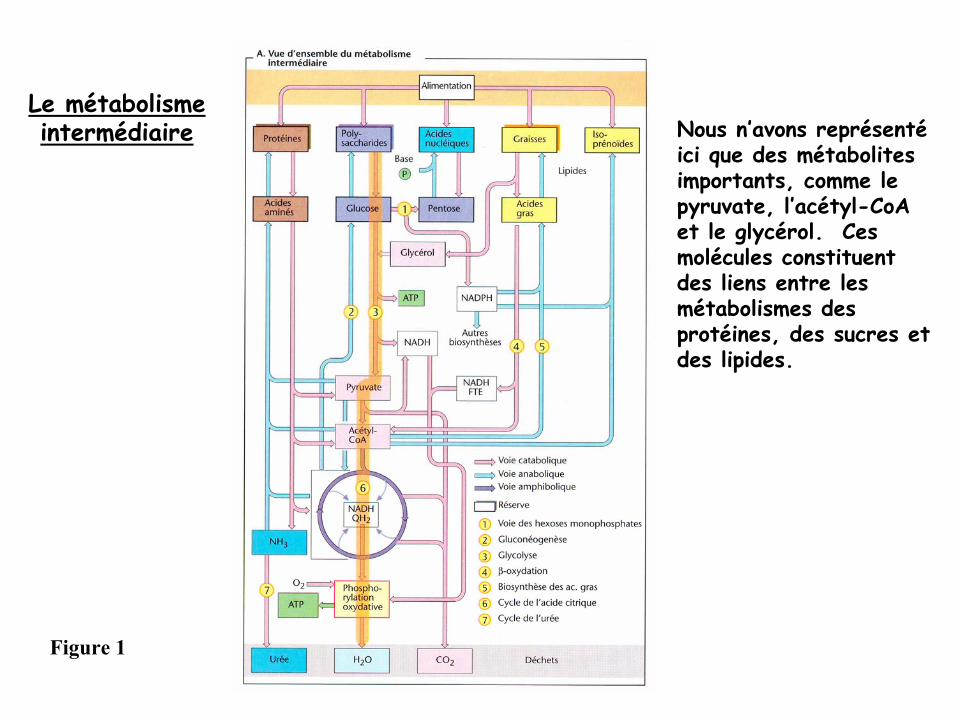
Le métabolismeintermédiaire Nous n’avons représenté
ici que des métabolites importants, comme le pyruvate, l’acétyl-CoAet le glycérol. Ces molécules constituent des liens entre les métabolismes des protéines, des sucres et des lipides.
Figure 1
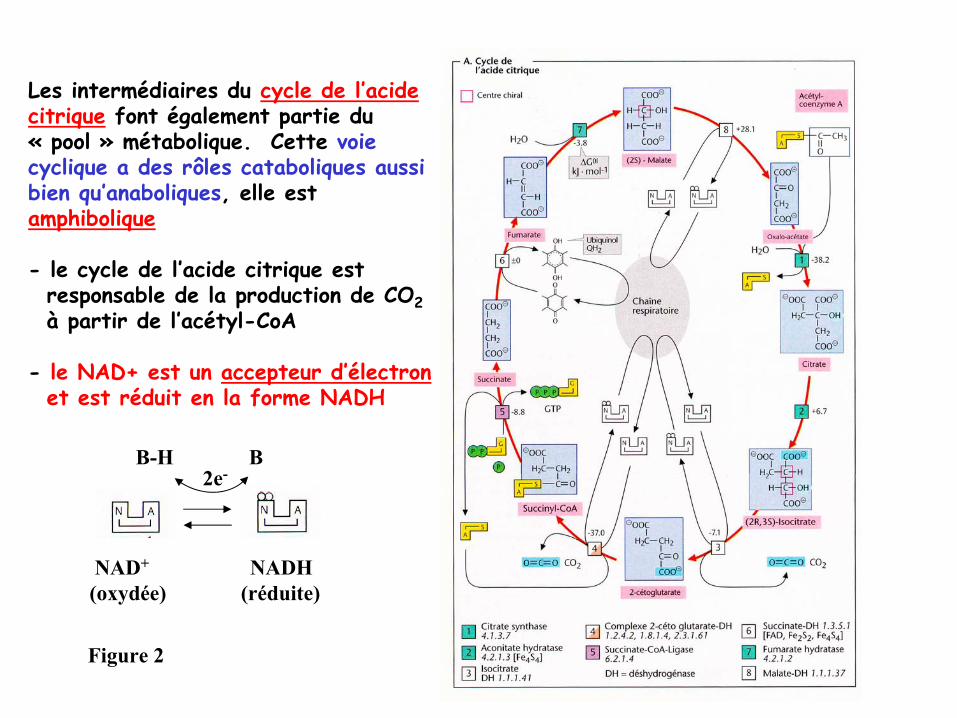
Les intermédiaires du cycle de l’acide citrique
font également partie du «
pool
»
métabolique. Cette voie cyclique a des rôles cataboliques aussibien qu’anaboliques, elle est amphibolique
-
le cycle de l’acide citrique est responsable de la production de CO2à
partir de l’acétyl-CoA
-
le NAD+ est un accepteur d’électronet est réduit en la forme NADH
NAD+
NADH(oxydée) (réduite)
Figure 2
B-H B2e-

-
La forme de réserve de l’énergie chimique dans toutes les cellules est l’adénosine triphosphate
(ATP). - La formation d’ATP nécessite l’utilisation d’énergie, le processus est donc
endergonique.
- de l’énergie sera libérée par la coupure de l’ATP en ADP et en phosphate.
L’hydrolyse de l’ATP est exergonique.
• cette hydrolyse permet, via un couplage énergétique, le déroulement de
phénomènes nécessitant de l’énergie
comme la plupart des voies métaboliques
anaboliques ainsi que les phénomènes de transport et de mouvement.
-
La voie la plus importante de synthèse d’ATP est la phosphorylation oxydative
(Figure 1).
-
les voies cataboliques du métabolisme génèrent des cofacteurs réduits
(NADH, QH2). Les électrons seront ensuite transportés jusqu’à
l’oxygène.
-
ce processus fortement exergonique
est catalysé
par la chaîne respiratoire
et
utilisé
indirectement pour la synthèse d’ATP.
-
plusieurs organismes peuvent, dans des conditions anaérobiques (en absence
d’oxygène) se rabattre sur l’ATP provenant de la glycolyse. Ce mode de
formation de l’ATP, plus rapide mais moins efficace, est la fermentation

Les réactions d’oxydo-réduction
-
les réactions d’oxydo-réduction, qui se traduisent par des transferts d’électrons, fournissent la plus grande partie de l’énergie libre aux organismes
Réduction
-> gain d’électrons
NAD+
+ H+
+ 2e-
NADH
le NAD+
est un accepteur d’électrons ou agent oxydant
ou forme oxydéele NADH est un donneur d’électrons ou agent réducteur
ou forme réduite
Oxydation
-> perte d’électrons
H2O ½O2
+ 2H+
+ 2e-
(photosynthèse)
-
l’efficacité
thermodynamique du transfert d’électron peut être déterminé
par les valeurs de potentiel d’oxydo-réduction
standard
(E°)
-
l’affinité
d’un agent oxydant pour les électrons augmente avec la valeur de son potentiel d’oxydo-réduction
standard
(E°)
NAD+
+ H+
+ 2e-
NADH E° = -0.315 VH2
O ½O2
+ 2H+
+ 2e-
E° = 0.815 V
•
plus le potentiel d’oxydo-réduction
standard est positif, plus la forme oxydéea tendance à
accepter les électrons et à
devenir une forme réduite

-
de ces deux réactions, c’est la réaction O2
/H2
O qui a la valeur de E° la plus élevéeet donc la plus grande affinité
pour les électrons
-
si ces deux réactions sont maintenant couplées en une réaction d’oxydo-réduction, c’est le NADH qui sera le donneur d’électrons
et O2
l’accepteur
½O2
+ NADH + H+
H2
O + NAD+
la différence de potentiel d’oxydo-réduction
(ΔE°) de cette réaction est:
ΔE°
= E° (l’accepteur d’électron) -
E° (donneur d’électron)ΔE°
= 0.815 –
(-0.315) = 1.130 V
On peut calculer la variation d’énergie libre de cette réaction par l’équation:
ΔG°
= -nFΔE° où
n = nombre d’électrons transférés par mole de substratF = constante de Faraday (96.494 C/mol)
-
pour la réaction ci-haut, le ΔG°
sera de -218 kJ/mol
=> réaction très exergonique
- pour produire 1mol d’ATP à
partir d’ADP et de Pi
, la quantité
d’énergie libre requiseest de 30,5 kJ/mol
=> la variation d’énergie libre de l’oxydation du NADH par O2est amplement suffisante pour produire plusieurs moles d’ATP

Respiration et fermentation
A.
Oxydation aérobique
et anaérobique du glucose
-
En présence d’oxygène (conditions aérobies) la plupart des cellules animales sont
capables de «
respirer
», c’est-à-dire de dégrader complètement par des processus
oxydatifs divers types d’aliments (lipides, acides aminés et sucres).
-
Si l’oxygène manque (conditions anaérobies), seul le glucose peut encore servir à
la
formation d’ATP. Bien que la dégradation du glucose dans ces conditions se termine
précocement au niveau du lactate, et ne fournisse que de faibles quantités d’ATP, elle
a une importance considérable pour la survie des cellules en l’absence d’oxygène.
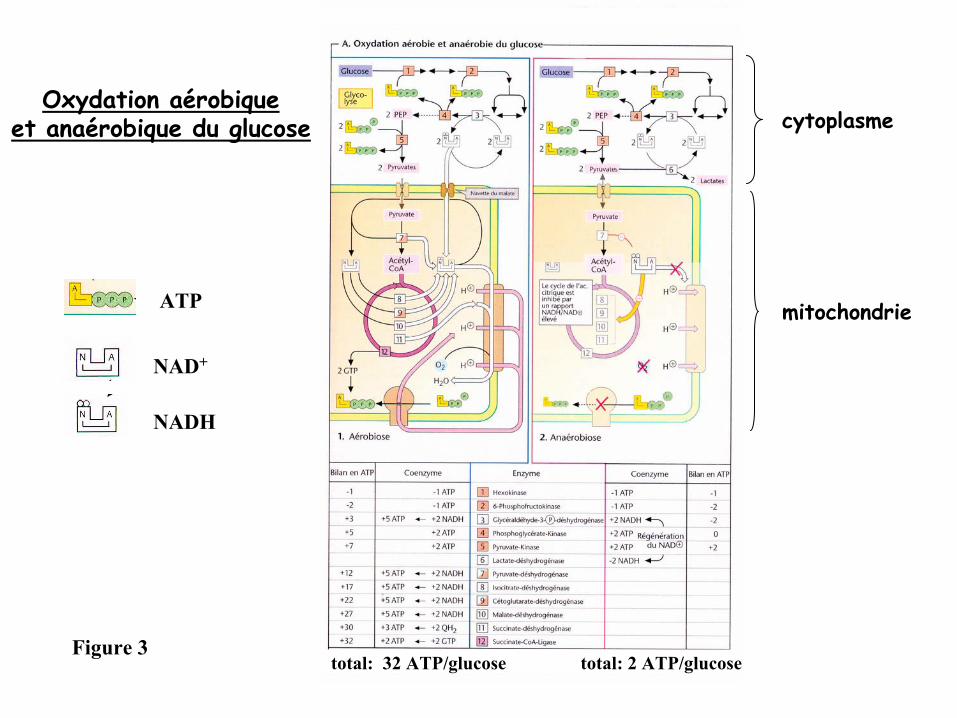
total: 32 ATP/glucose total: 2 ATP/glucose
Oxydation aérobiqueet anaérobique du glucose cytoplasme
mitochondrieATP
NAD+
NADH
Figure 3

Conditions aérobiques
(Figure 3 à
gauche)
-
l’ATP est obtenu presque exclusivement par phosphorylation oxydative.
- les acides gras parviennent dans les mitochondries avec l’aide de la carnitine
et y seront dégradés par β-oxydation
en groupements acétyls
liés au coenzyme A.
- Le glucose sera transformé
en pyruvate dans le cytoplasme par la glycolyse.
-le pyruvate sera transporté
dans la matrice mitochondriale pour y
être décarboxylé
de façon oxydative en acétyl-CoA
par le complexe
pyruvate-déshydrogénase.
-
Les équivalents réducteurs (2 NADH par glucose) apparus durant la glycolyse
atteignent la matrice mitochondriale grâce à
la navette du malate.
-
Les groupements acétyls
formés seront oxydés en CO2
dans le cycle de l’acide
citrique.
-
La dégradation des acides aminés génère également des groupements acétyls
ou des
produits qui peuvent s’insérer directement dans le cycle de l’acide citrique.
- Les équivalents réducteurs obtenus seront en cas de besoin transférés à
l’oxygène
via la chaîne respiratoire, ce qui libère de l’énergie libre utilisée pour former de
l’ATP par l’intermédiaire d’un gradient de protons.

Conditions anaérobiques
(Figure 3 à
droite)
-
comme l’accepteur d’électrons de la chaîne respiratoire (l’oxygène) manque, le NADH et le QH ne peuvent plus être réoxydés.
-
Ce n’est donc pas seulement la synthèse mitochondriale d’ATP qui se bloque mais presque tout le métabolisme de la matrice mitochondriale.
-
Une des causes majeures de ce phénomène est l’élévation de la concentration de NADH, qui inhibe entre autres le cycle de l’acide citrique et la pyruvate-déshydrogénase.
-
La β-oxydation et la navette du malate, qui ont besoin de NAD+, sont également arrêtées.
-
Comme la dégradation des acides aminés ne peut plus conduire à
la formation d’énergie, la cellule est complètement dépendante de la formation d’ATP via la dégradation du glucose par la glycolyse.
-
Si ce processus doit se poursuivre continuellement, le NADH formé
doit être réoxydéen permanence. Comme ceci ne peut plus s’effectuer dans les mitochondries, les cellules animales réduisent le pyruvate en lactate dans des conditions anaérobies et libèrent ce lactate dans le sang.
-
Ce type de processus est appelé
fermentation
et leur rendement en ATP est faible
: c’est ainsi que lors de la formation du lactate on n’obtient que 2 ATP par glucose.

-
Si l’on veut évaluer le nombre de molécules d’ATP formées dans des conditions
aérobiques, il est nécessaire de connaître le quotient P/O,
-
le rapport molaire entre l’ATP formé
(«
P
») et l’eau formée («
O
»).
-
Au cours du transport de deux électrons du NADH à
l’O2
, on estime aujourd’hui que
10
protons seront accumulés dans l’espace intermembranaire, tandis qu’à
partir de
l’ubiquinone
(QH2
) il n’y en aura que 6.
-
L’ATP-synthase
a besoin de trois H+
pour la synthèse d’un ATP, de sorte que les
rapports P/O maximaux possibles sont environ de 3 ou 2.
-
Cependant, le transport des métabolites dans la matrice mitochondriale et l’échange
de ATP4-
contre ADP3-
dans l’espace intermembranaire
utilisent aussi des protons.
- donc, les rapports P/O pour l’oxydation de NADH et du QH2
sont plutôt de l’ordre de
2,5 ou 1,5 selon le cas.
-
Si l’on évalue le bilan énergétique de la glycolyse aérobie sur la base de ces données,
on aboutit à
un rendement de 32 ATP par glucose
(certains livres rapportent des rapports P/O de 3 pour le NADH et 2 pour le QH2
,
ce qui donne un rendement de 38 ATP par glucose)

Mitochondries : organisation et rôles (Chapitre 20, Voet et Voet)
1.
Organisation des mitochondries
(Figure 4)
-
Les mitochondries sont des organites de la taille d’une bactérie (taille d’environ
1 x 2 μm), présents en grand nombre dans pratiquement toutes les cellules
eucaryotes.
-
il en existe à
peu près 2,000 par cellule et occupent jusqu’à
25% du volume
d’une cellule.
-
les mitochondries sont bordées par deux membranes:
- une membrane externe
lisse
- une membrane interne
plissée qui présente donc une surface importante et
traverse l’espace matriciel. Les replis de la membrane interne s’appellent
des
crêtes.
-
l’espace intermédiaire entre la membrane externe et la membrane interne
est désigné
le plus souvent sous le terme d’espace intermembranaire.
-
les tissus ayant un métabolisme oxydatif intense comme par exemple le muscle
cardiaque possèdent des mitochondries contenant de nombreuses crêtes.
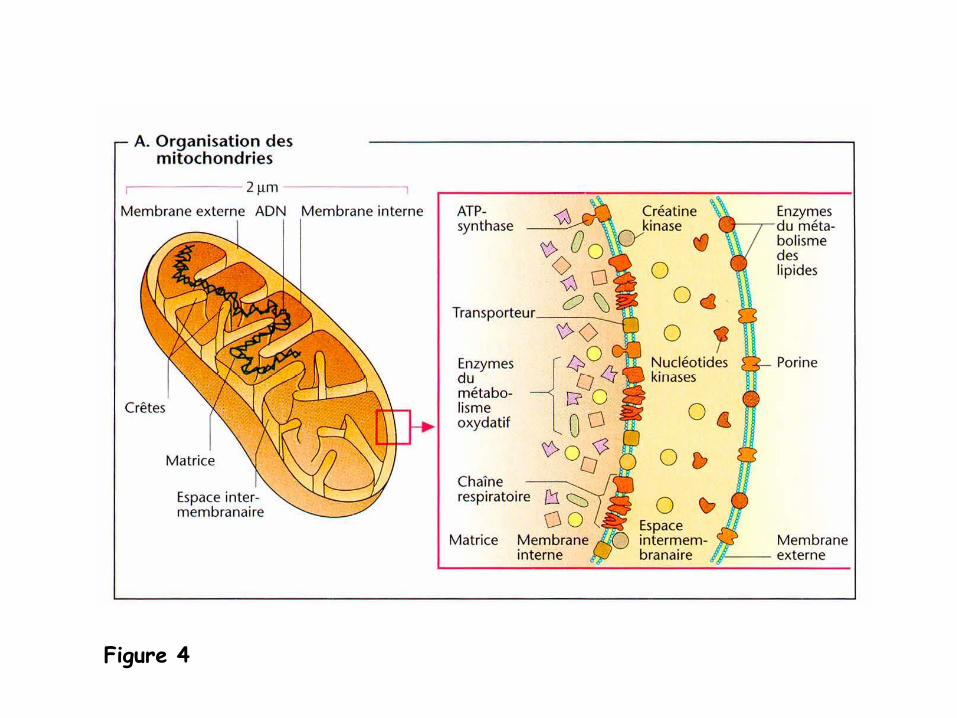
Figure 4

Les membranes mitochondriales contiennent des protéines intégrales.
- Dans la membrane externe, sont présentes:
- des porines, formant des pores qui rendent la membrane perméable aux
molécules de taille inférieure à
10 kDa
ainsi permettant l’échange de ces
molécules entre le cytoplasme et l’espace intermembranaire.
-
La membrane interne des mitochondries est au contraire imperméable à
la plupart
des molécules
(à
l’exceptions de l’O2
, CO2
, et H2
O).
-
elle contient des cardiolipines, un type de phospholipide particulier. -
parmi les protéines de la membrane interne, on va trouver des
transporteurs spécifiques
qui permettent un transport contrôlé
de
substances, des enzymes
et d’autres composants de la chaîne
respiratoire
ainsi que l’ATP-synthase.
-
La matrice est également riche en protéines, et en particulier en enzymes du
cycle de l’acide citrique.

2.
Rôles métaboliques
(Figure 5)
-
Les mitochondries sont appelées les «
usines biochimiques »
de la cellule car elles
génèrent au cours de la dégradation oxydative des substances alimentaires via la
phosphorylation oxydative la majeure partie de l’ATP.
-
dans la matrice sont localisé
la pyruvate déshydrogénase (transformation du
pyruvate en acétyl-CoA), les enzymes du cycle de l’acide citrique, de la
dégradation des acides gras par β-oxydation, et d’une partie du cycle de l’urée.
-
la chaîne respiratoire couplée à
la synthèse
d’ATP
(dont l’ensemble est désigné
sous
le nom de phosphorylation oxydative) ainsi que les enzymes de la biosynthèse des
hèmes sont associées à
la membrane interne.
-
les mitochondries fournissent aux cellules les produits intermédiaires de leur
métabolisme et constituent une réserve de calcium qui maintient la concentration
cytoplasmique à
une valeur constante faible (~ 1 μm).
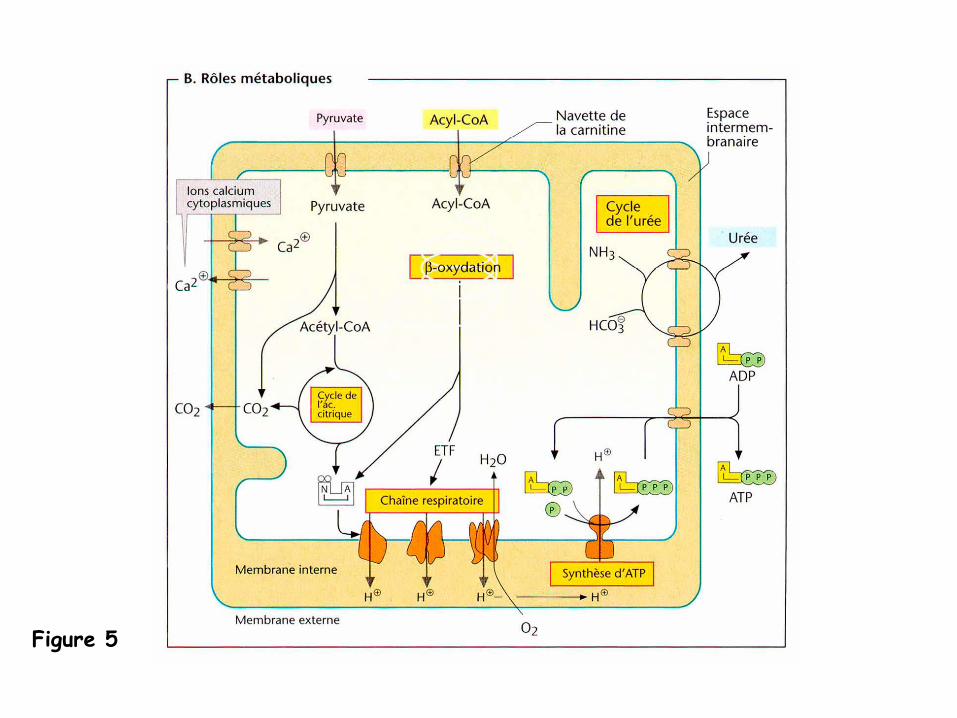
Figure 5
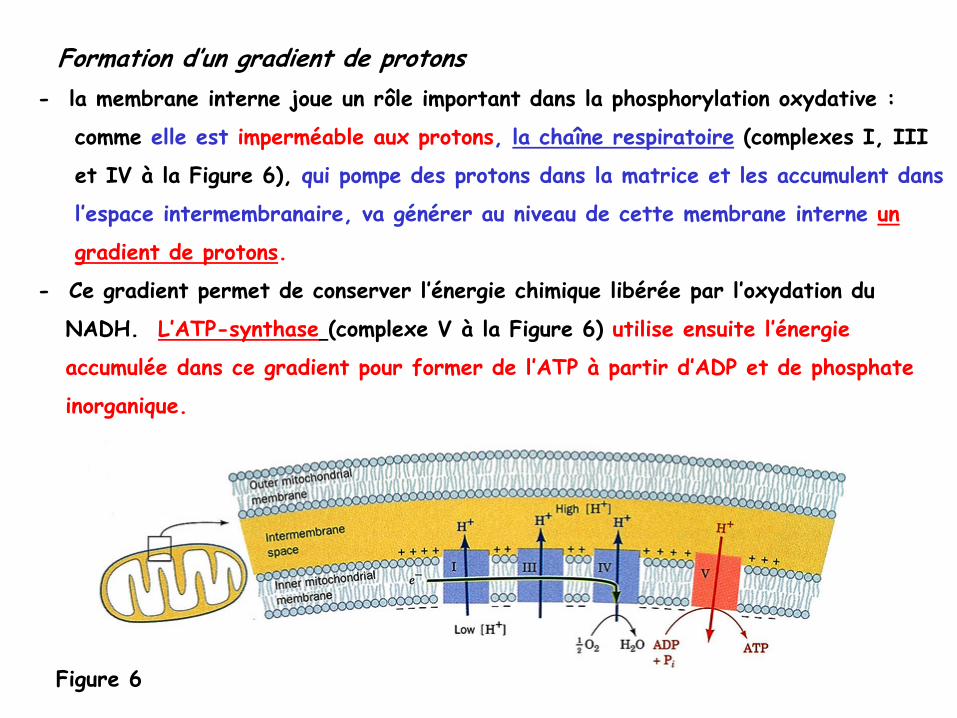
Formation d’un gradient de protons-
la membrane interne joue un rôle important dans la phosphorylation oxydative
:
comme elle est imperméable aux protons, la chaîne respiratoire
(complexes I, III
et IV à
la Figure 6), qui pompe des protons dans la matrice et les accumulent dans
l’espace intermembranaire, va générer au niveau de cette membrane interne
un
gradient de protons.
-
Ce gradient permet de conserver l’énergie chimique libérée par l’oxydation du
NADH. L’ATP-synthase
(complexe V à
la Figure 6) utilise ensuite l’énergie
accumulée dans ce gradient pour former de l’ATP à
partir d’ADP et de phosphate
inorganique.
Figure 6
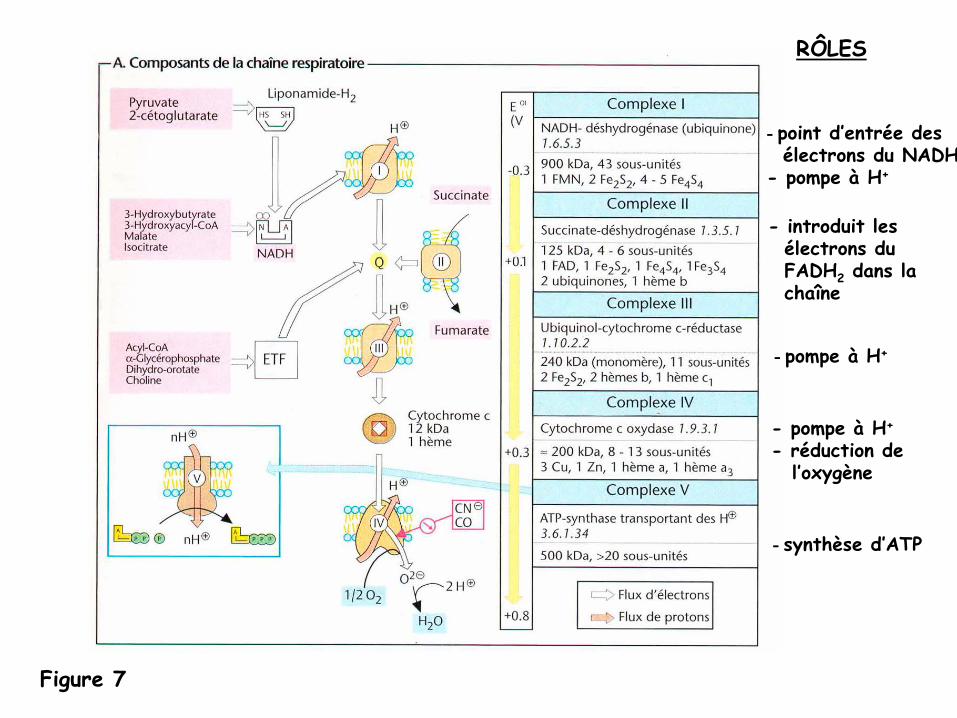
Figure 7
-
point d’entrée desélectrons du NADH
- pompe à H+
-
pompe à
H+
- pompe à H+
- réduction del’oxygène
-
synthèse d’ATP
RÔLES
- introduit lesélectrons duFADH2
dans lachaîne
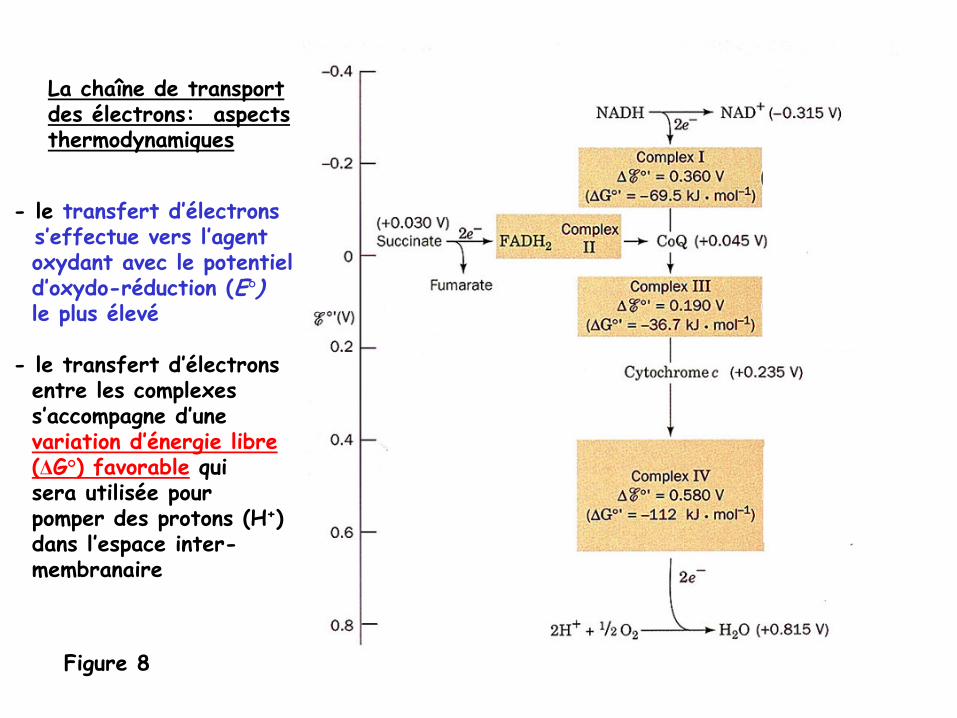
Figure 8
La chaîne de transportdes électrons: aspectsthermodynamiques
- le transfert d’électronss’effectue vers l’agentoxydant avec le potentield’oxydo-réduction
(E°)le plus élevé
-
le transfert d’électronsentre les complexess’accompagne d’unevariation d’énergie libre(ΔG°) favorable
qui sera utilisée pour pomper des protons (H+)dans l’espace inter-membranaire
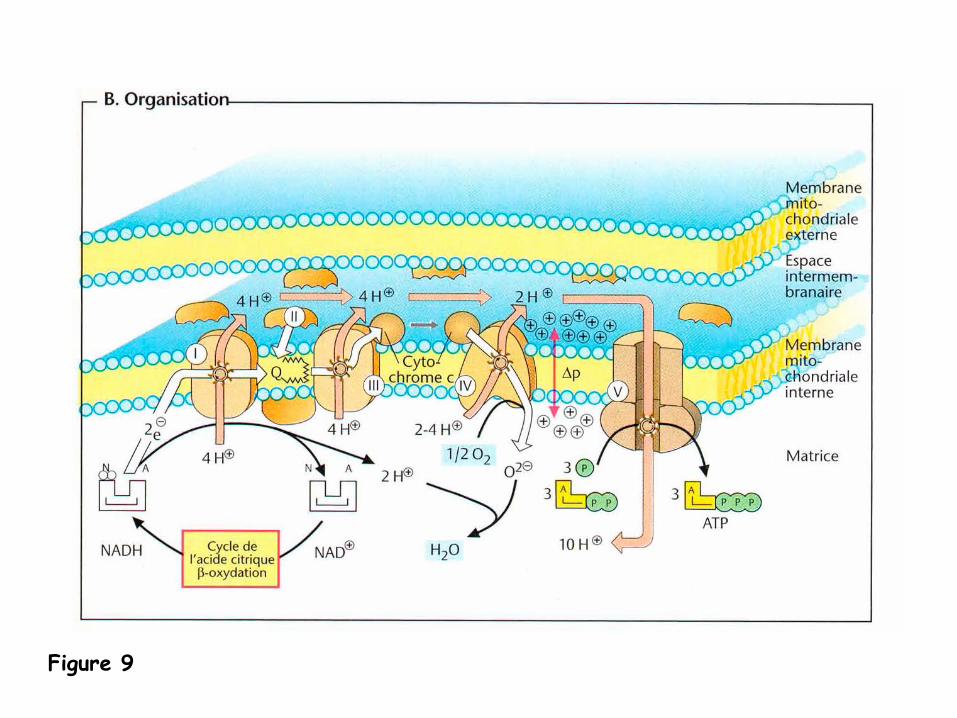
Figure 9

Récapitulation : Rôles métaboliques des mitochondries
Les mitochondries sont appelées les « usines biochimiques »de la cellule car elles produisent la majeure partie de l’ATP, au cours de la dégradation oxydative des substances alimentaires.
Les processus suivants se déroulent dans les mitochondries : transformation du pyruvate en acétyl-CoA, cycle de l’acide citrique, chaîne respiratoire couplée à la synthèse d’ATP (dont l’ensemble est désigné sous le nom de phosphorylation oxydative), dégradation des acides gras par β-oxydation et une partie du cycle de l’urée.
La tâche fondamentale des mitochondries est de préleverdans le cytoplasme les substrats du métabolisme énergétique (acides gras, pyruvate, squelette carboné des acides aminés) et de réaliser une dégradation oxydative de ces substances en CO2et H2O en échange d’ATP.
Les réactions du cycle de l’acide citrique qui se déroulent principalement dans la matrice fournissent du carbone complètement oxydé (CO2) et des équivalents réducteurs liés àdes coenzymes (NADH et QH2).
Récapitulation : Rôles métaboliques des mitochondries
Les mitochondries sont appelées les « usines biochimiques »de la cellule car elles produisent la majeure partie de l’ATP, au cours de la dégradation oxydative des substances alimentaires.
Les processus suivants se déroulent dans les mitochondries : transformation du pyruvate en acétyl-CoA, cycle de l’acide citrique, chaîne respiratoire couplée à la synthèse d’ATP (dont l’ensemble est désigné sous le nom de phosphorylation oxydative), dégradation des acides gras par β-oxydation et une partie du cycle de l’urée.
La tâche fondamentale des mitochondries est de préleverdans le cytoplasme les substrats du métabolisme énergétique (acides gras, pyruvate, squelette carboné des acides aminés) et de réaliser une dégradation oxydative de ces substances en CO2et H2O en échange d’ATP.
Les réactions du cycle de l’acide citrique qui se déroulent principalement dans la matrice fournissent du carbone complètement oxydé (CO2) et des équivalents réducteurs liés àdes coenzymes (NADH et QH2).

Récapitulation : La chaîne respiratoire
La chaîne respiratoire qui réoxyde les coenzymes est localisée dans la membrane interne des mitochondries.
Elle nécessite comme agents réducteurs (fournissant des électrons) du NADH et du FADH2 liés à un enzyme pour réduire l’oxygène et former l’H2O.
Cette réaction fortement exergonique est divisée en plusieurs étapes et couplée avec le transport de protons (H+), depuis la matrice jusqu’à l’espace intramembranaire à travers la membrane interne des mitochondries.
Il se forme ainsi de part et d’autre de la membrane interne un gradient électrochimique.
Les mitochondries utilisent ces gradients pour la synthèse d’ATP à partir d’ADP et de Pi, catalysée par l’ATP-synthase; par ailleurs, certains systèmes de transport spécifique seront stimulés.
Récapitulation : La chaîne respiratoire
La chaîne respiratoire qui réoxyde les coenzymes est localisée dans la membrane interne des mitochondries.
Elle nécessite comme agents réducteurs (fournissant des électrons) du NADH et du FADH2 liés à un enzyme pour réduire l’oxygène et former l’H2O.
Cette réaction fortement exergonique est divisée en plusieurs étapes et couplée avec le transport de protons (H+), depuis la matrice jusqu’à l’espace intramembranaire à travers la membrane interne des mitochondries.
Il se forme ainsi de part et d’autre de la membrane interne un gradient électrochimique.
Les mitochondries utilisent ces gradients pour la synthèse d’ATP à partir d’ADP et de Pi, catalysée par l’ATP-synthase; par ailleurs, certains systèmes de transport spécifique seront stimulés.

Études expérimentales de la chaîne de transport des électrons
-
l’élucidation de la séquence des transporteurs de la chaîne de transport d’électrons est due, pour une large part, à
l’utilisation d’inhibiteurs spécifiquesde composantes de cette chaîne:
-
la roténone
(une toxine végétale)-
l’amytal
(un barbiturique)-
l’antimycine
A
(un antibiotique)-
le cyanure
- ces inhibiteurs bloquent le transport des électrons le long de la chaîne
(Figure 11)
-
l’activité
de la chaîne de transfert d’électrons peut être mesurée grâce à
savitesse de consommation d’oxygène
-
utilise une électrode à
oxygène pour mesurerla concentration en O2
=>
Figure 10
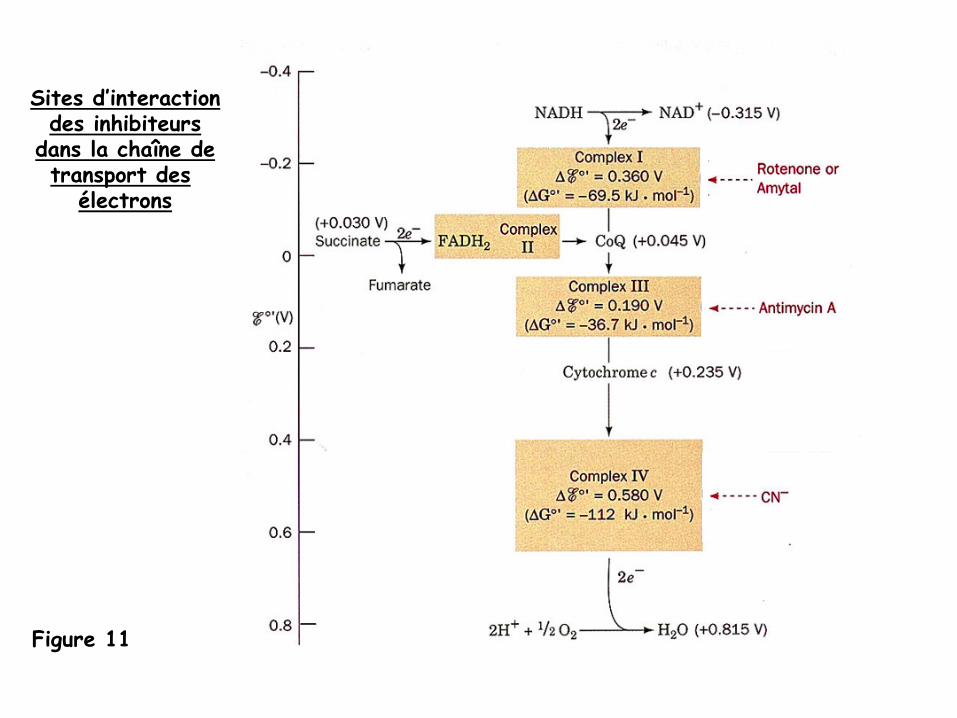
Figure 11
Sites d’interactiondes inhibiteurs
dans la chaîne detransport des
électrons
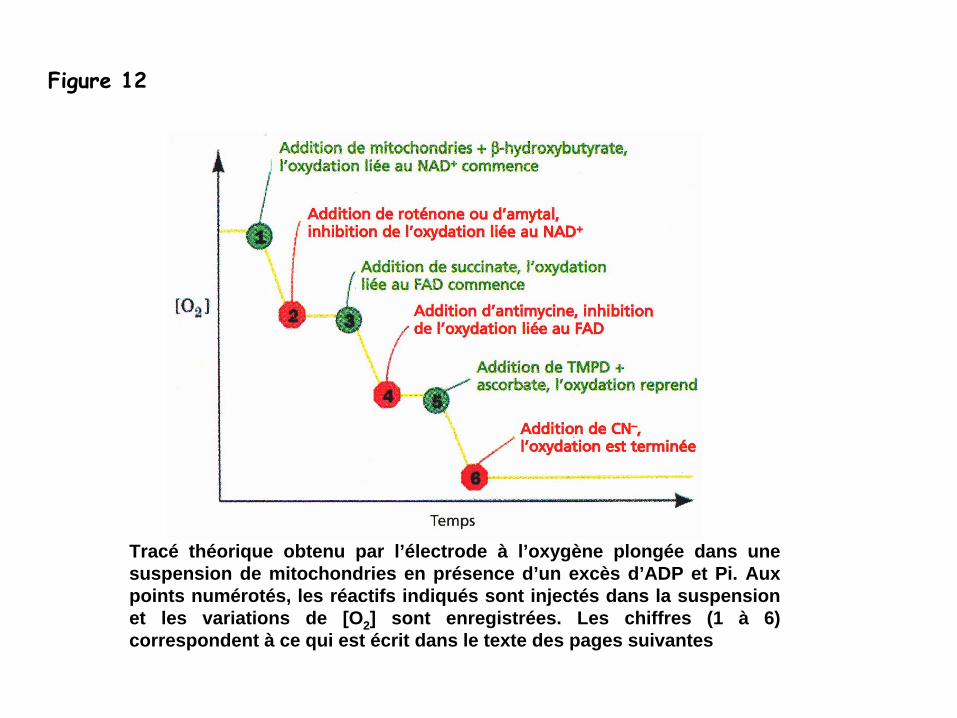
Tracé théorique obtenu par l’électrode à l’oxygène plongée dans une suspension de mitochondries en présence d’un excès d’ADP et Pi. Aux points numérotés, les réactifs indiqués sont injectés dans la suspension et les variations de [O2 ] sont enregistrées. Les chiffres (1 à 6) correspondent à ce qui est écrit dans le texte des pages suivantes
Figure 12
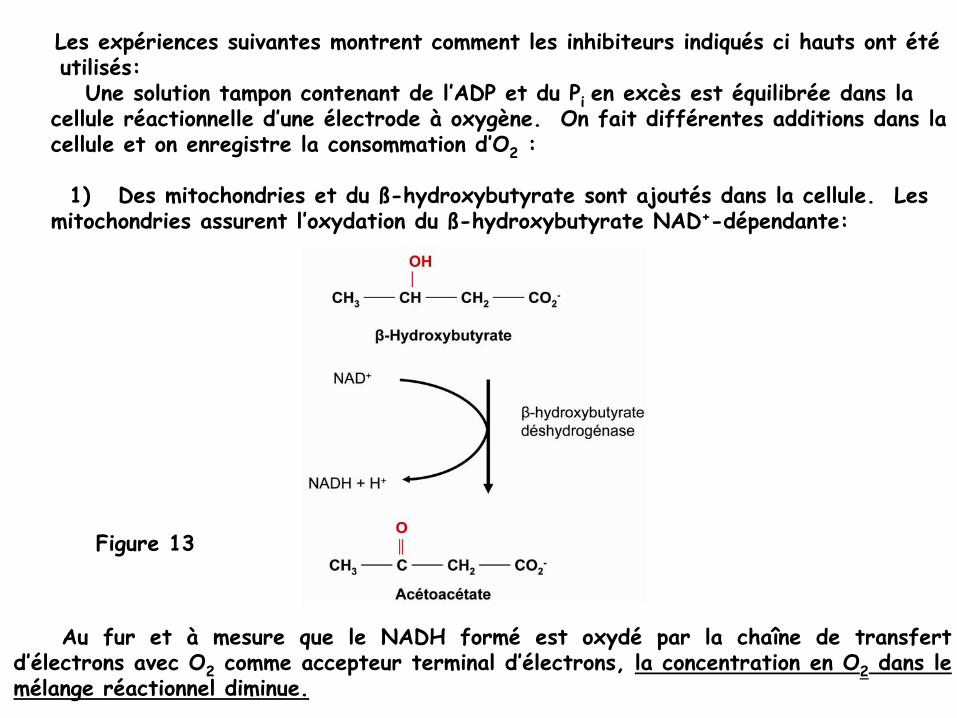
Les expériences suivantes montrent comment les inhibiteurs indiqués ci hauts ont étéutilisés:
Une solution tampon contenant de l’ADP et du Pi en excès est équilibrée dans la cellule réactionnelle d’une électrode à
oxygène. On fait différentes additions dans la cellule et on enregistre la consommation d’O2
:
1)
Des mitochondries et du ß-hydroxybutyrate
sont ajoutés dans la cellule. Les mitochondries assurent l’oxydation du ß-hydroxybutyrate
NAD+-dépendante:
Au fur et à
mesure que le NADH formé
est oxydé
par la chaîne de transfert d’électrons avec O2
comme accepteur terminal d’électrons, la concentration en O2
dans le mélange réactionnel diminue.
Figure 13

2) L’addition de roténone ou d’amytal
arrête complètement l’oxydation du ß-
hydroxybutyrate.
3) L’addition de succinate, qui s’oxyde par l’intermédiaire du FAD, rétablit la
consommation d’O2
. Les électrons du FADH2
peuvent donc réduire l’oxygène en
présence de roténone ; cela signifie que les électrons du FADH2
entrent dans la chaîne de transfert d’électrons après le site d’inhibition de la roténone.
4) L’addition d’antimycine
A inhibe le transfert d’électrons provenant du FADH2
.
5)
Bien que le NADH et le FADH2
soient les donneurs d’électrons physiologiques de
la chaîne respiratoire, on peut utiliser aussi des donneurs d’électrons artificiels
pour explorer le cheminement des électrons.
• Le tétraméthyl-p-phénylène-diamine (TMPD)
est un transporteur d’électrons
réduit par l’ascorbate
et qui transfère ses électrons directement au
cytochrome c.
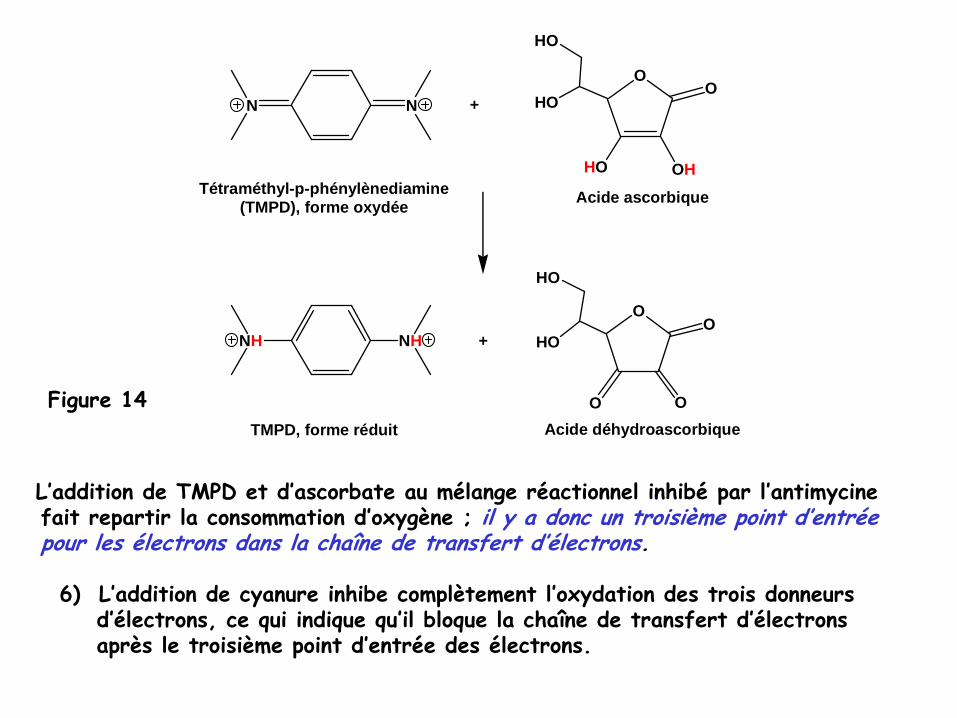
L’addition de TMPD et d’ascorbate
au mélange réactionnel inhibé
par l’antimycinefait repartir la consommation d’oxygène ; il y a donc un troisième point d’entréepour les électrons dans la chaîne de transfert d’électrons.
6)
L’addition de cyanure inhibe complètement l’oxydation des trois donneurs d’électrons, ce qui indique qu’il bloque la chaîne de transfert d’électrons après le troisième point d’entrée des électrons.
O
O
O
OHHO
NN
NHNHO
OO
HO
HO
HO
HO
+
+
Tétraméthyl-p-phénylènediamine(TMPD), forme oxydée Acide ascorbique
Acide déhydroascorbiqueTMPD, forme réduit
Figure 14
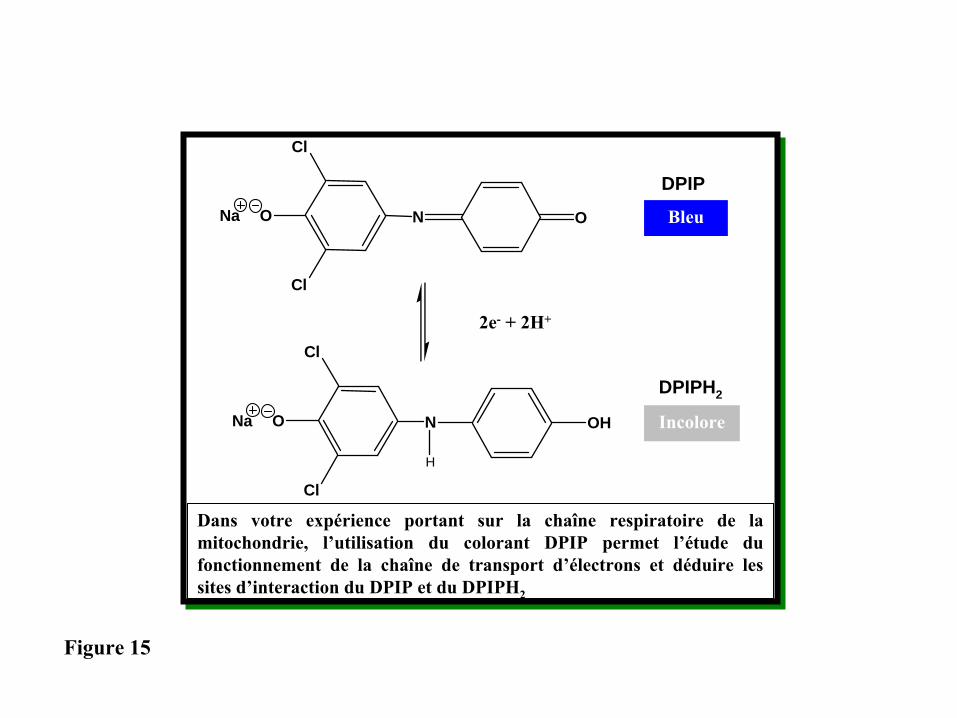
Dans votre expérience portant sur la chaîne respiratoire de la mitochondrie, l’utilisation du colorant DPIP permet l’étude du fonctionnement de la chaîne de transport d’électrons et déduire les sites d’interaction du DPIP et du DPIPH2
DPIP
DPIPH2
N O
Cl
O
Cl
Na
N OH
Cl
O
Cl
Na
H
2e-
+ 2H+
Bleu
Incolore
Figure 15
Top Related