






Langages
Pages
Légal



Définitions
• Addiction: consommation compulsive en dépit de toutes conséquences négatives
•Dépendance: survenu d’un syndrome de sevrage à l’arrêt brusque de la drogue
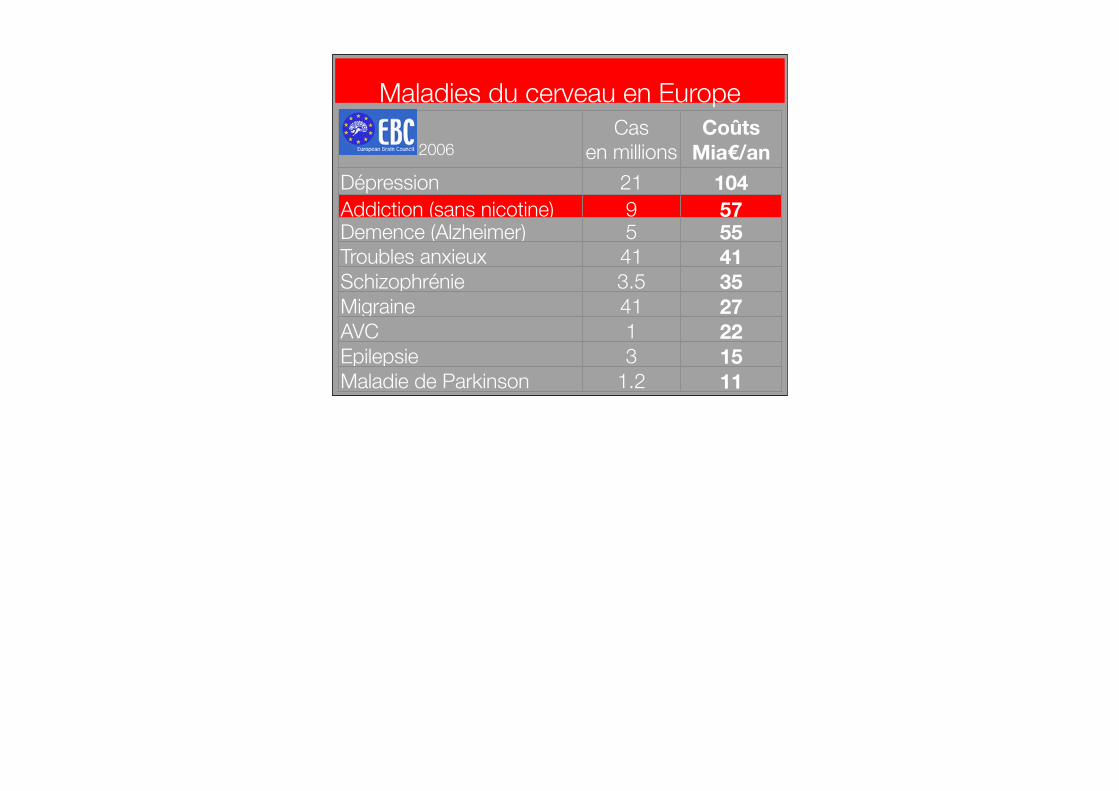
Casen millions
CoûtsMia€/an
Dépression 21 104Addiction (sans nicotine) 9 57Demence (Alzheimer) 5 55Troubles anxieux 41 41Schizophrénie 3.5 35Migraine 41 27AVC 1 22Epilepsie 3 15Maladie de Parkinson 1.2 11
2006
Maladies du cerveau en Europe

• Hallucinogènes (LSD) 1
• Cannabis (THC) 2
• Benzodiazépines (BDZ) 2
• Alcool 3
• Nicotine 3
•Opiacés (Morphine, Héroïne) 4
• Amphétamines 5
• Cocaïne 5
Risque relatif d’addictionGoldstein & Kalant, Science 1990
20
15
10
5
0
Cum
ulat
ive
proa
bilit
y (%
)403020100
Years since first use
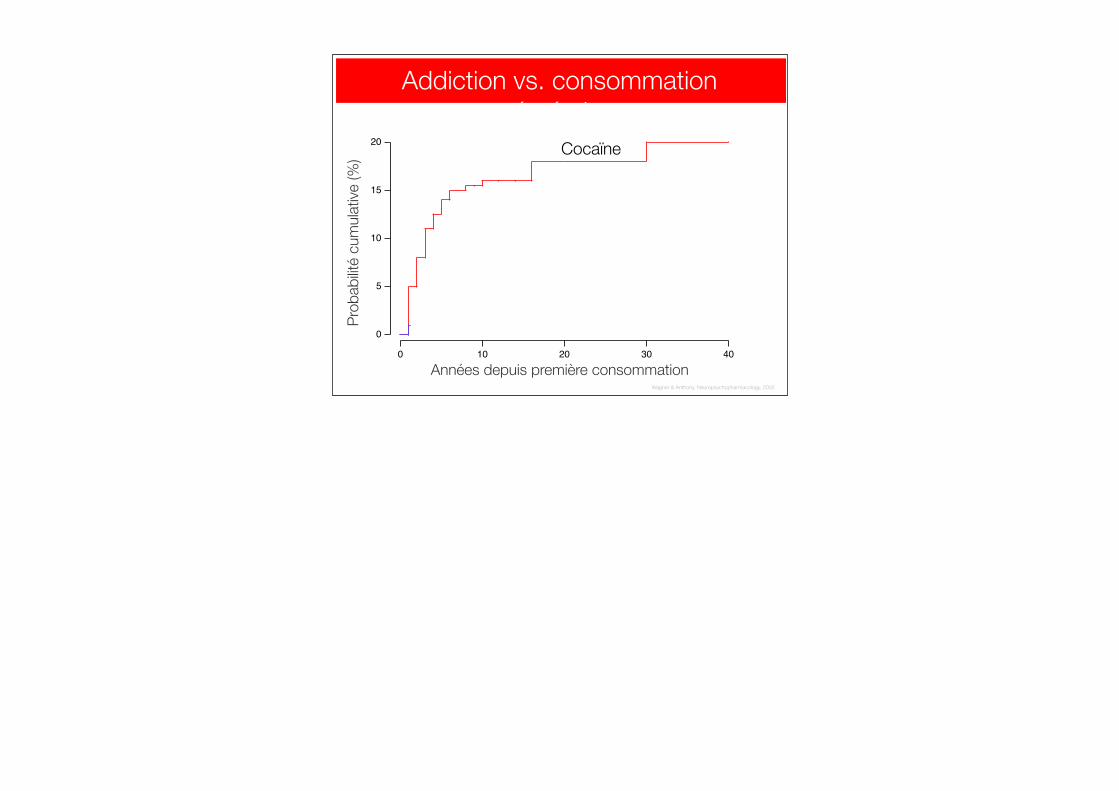
Addiction vs. consommation récréative
Cocaïne
Cannabis
Wagner & Anthony, Neuropsychopharmacology, 2002
Cum
mul
ativ
e pr
obab
ility
(%)
Années depuis première consommationPr
obab
ilité
cum
ulativ
e (%
)
Cocaïne
100
80
60
40
20
0
Cum
ulat
ive
proa
bilit
y (%
)403020100
Years since first useC
umul
ativ
e pr
obab
ility
(%)
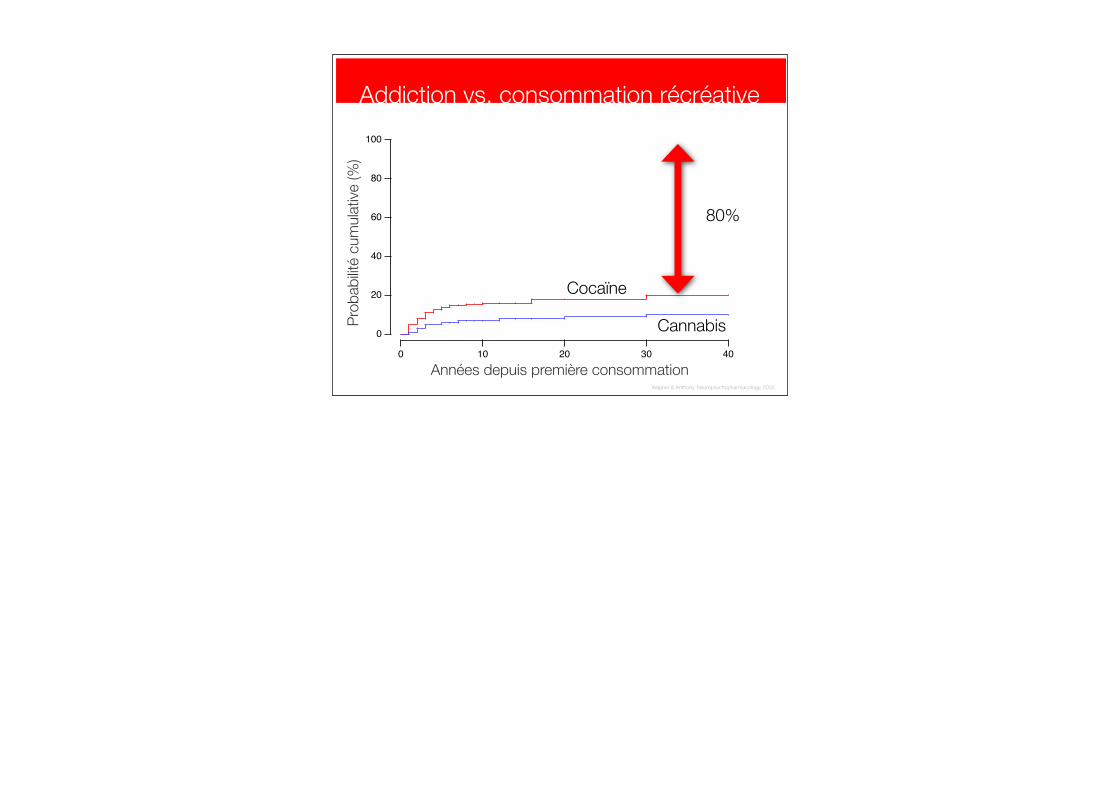
80%
Années depuis première consommationPr
obab
ilité
cum
ulativ
e (%
)
Wagner & Anthony, Neuropsychopharmacology, 2002
Addiction vs. consommation récréative
Cannabis

Dangers de la cocaïne
• Pression sanguine ↑↑
• Infarctus du myocarde
• Arythmie cardiaque
• Hémorragie cérébrale
• Addiction
Vulnérabilité individuelle ?

Nicotine Gambling ChocolateCocaine Morphine
AddictionAddiction

-----------
Drugs Gambling Food
Leyton et al., 2002 Boileau et al., 2003 Small et al., 2003
e.g.Poker e.g.Hamburgere.g.Amphetamines
Activation of the ventral striatum

-----------
Drugs Gambling Food
Leyton et al., 2002 Boileau et al., 2003 Small et al., 2003
Activation of the ventral striatum
e.g.Poker e.g.Hamburgere.g.Amphetamines
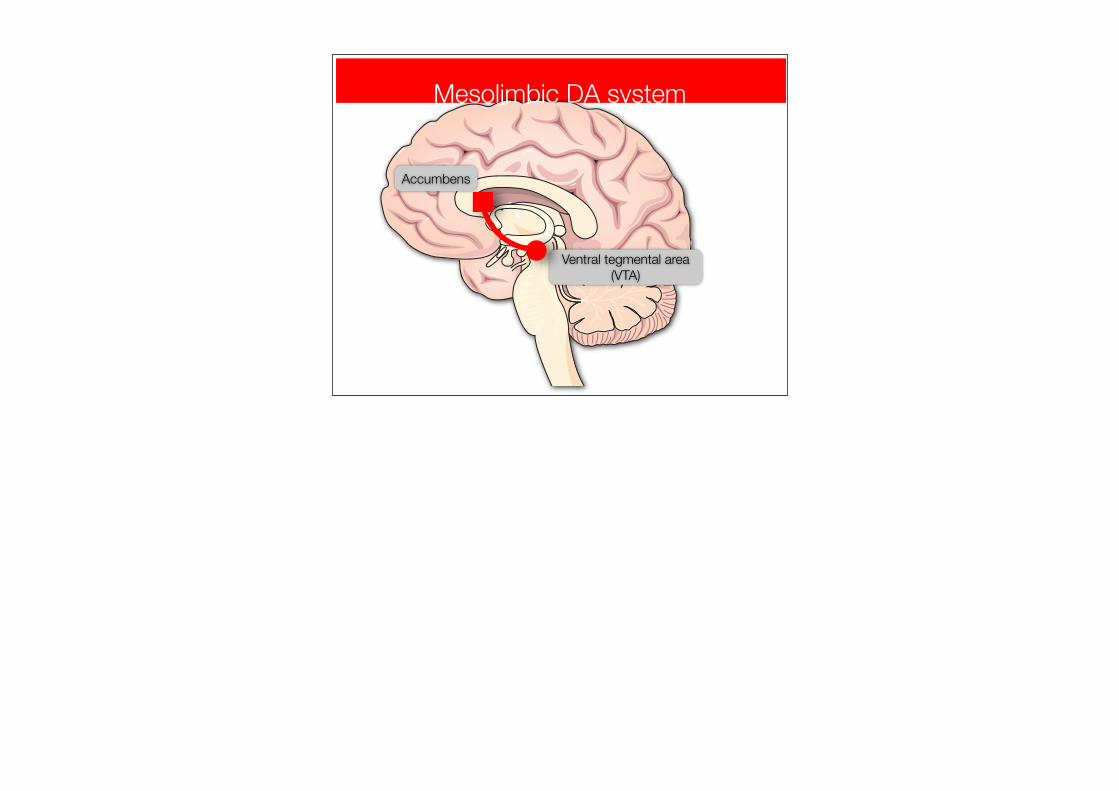
Mesolimbic DA system
Ventral tegmental area (VTA)
Accumbens

DA
GABA
NAcVTA
i
DA
GABA
NAcVTA
i
Increased dopamine(all addictive drugs)
Class I (morphine, THC, GHB)
G
G
Ventral tegmental area
Dopamine transporterG-protein coupled receptor Ionotropic receptorLüscher & Ungless, PLoS, 2006

DA
GABA
NAcVTA
i
DA
GABA
NAcVTA
i
Increased dopamine(all addictive drugs)
Class I (morphine, THC, GHB)
G
G
DA
GABA
Class II (benzodiazepines, nicotine, ethanol)
NAcVTA
i
i
Increased dopamine(all addictive drugs)
Class I (morphine, THC, GHB)
G
G
Ventral tegmental area
Dopamine transporterG-protein coupled receptor Ionotropic receptorLüscher & Ungless, PLoS, 2006

DA
GABA
NAcVTA
i
DA
GABA
Class III(cocaine, amphetamine, ecstacy)
Class II (benzodiazepines, nicotine, ethanol)
NAcVTA
i
i
T
Increased dopamine
Class I (morphine, THC, GHB)
G
G
DA
GABA
NAcVTA
i
Increased dopamine(all addictive drugs)
Class I (morphine, THC, GHB)
G
G
DA
GABA
Class II (benzodiazepines, nicotine, ethanol)
NAcVTA
i
i
Increased dopamine(all addictive drugs)
Class I (morphine, THC, GHB)
G
G
Ventral tegmental area
Dopamine transporterG-protein coupled receptor Ionotropic receptorLüscher & Ungless, PLoS, 2006
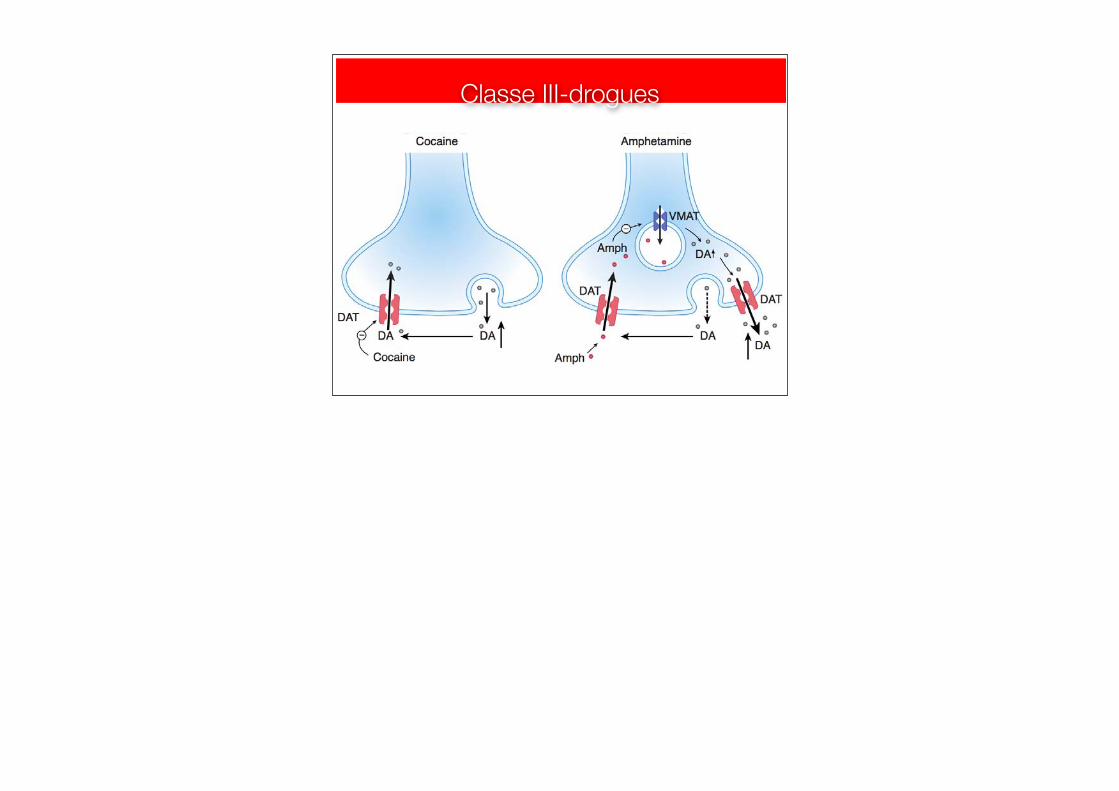
Classe III-drogues


Nicotine Gambling ChocolateCocaine Morphine
Addiction
Mesolimbic dopamine ↑
Addiction

0 1 2s
Schultz et al., Science 1997
Prediction error
-1 0 1 2s
0 1 2 s
-1 0 1 2s
R

0 1 2s
Schultz et al., Science 1997
0 1 2 s
-1 0 1 2s
no R
0 1 2
Image
-1 0 1 2s
Image R
0 1 2
R
Prediction error
1857
In a resting animal, midbrain dopamineneurons show a slow, steady (“tonic”) rateof firing; certain meaningful stimuli pro-voke brief, abrupt (“phasic”) changes indischarge. In previous work, Schultz andco-workers (2, 3) recorded the activity ofsingle dopamine neurons in awake mon-keys with microelectrodes. When themonkey received an unexpected reward,such as a drop of juice, most dopaminecells responded with a burst of firing.However, if the monkey learned that astimulus, such as a particular pattern on acomputer monitor, always preceded deliv-ery of the reward, the dopamine neuronsno longer responded to the reward butfired instead in response to the predictive(“conditioned”) stimulus. Omission of apredicted reward caused slowing or cessa-tion of dopamine firing around the timethat the reward was expected.
The phasic responses of midbraindopamine neurons resemble a key signalin computer models based on animallearning (4–7). Through adjustment ofconnection weights in a neural network(“reinforcement”), these models are ableto predict the achievement of a goal state(“reward”) and result in optimization ofactions. The incremental improvement inpredictions is driven by a “prediction er-ror”: the difference between expected andexperienced rewards. The new findings ofFiorillo et al. both strengthen and chal-lenge the reinforcement-learning notion of
midbrain dopamine neuronal activity whileraising many fascinating new questions.
The novelty of the Fiorillo et al. studylies in their systematic variation of the pro-portion of conditioned stimuli that werefollowed by a reward. The conditionedstimulus associated with each reward prob-ability (0.0, 0.25, 0.50, 0.75, 1.0) consistedof a unique visual pattern displayed on acomputer monitor for 2 seconds (see thefigure). Delivery of the reward, a drop ofsyrup, coincided with the offset of the con-ditioned stimulus. The dopamine neuronsresponded to the conditioned stimuli withphasic increases in firing that correlatedpositively with reward probability. In con-trast, responses to reward delivery showeda strong negative correlation with rewardprobability. On trials when an expected re-ward was omitted, the firing of dopamineneurons tended to decrease at the time ofpotential reward delivery, and the magni-tude of this dip tended to increase with theprobability of a reward. The systematicvariation in the strength of the phasic re-sponses to conditioned stimuli and to re-ward delivery or omission supports and ex-tends previous findings obtained at the twoextreme probabilities (0.0 and 1.0): Thehigher the likelihood of reward, the
stronger the firing to the conditioned stim-ulus, the larger the decrease in firing to re-ward omission, and the weaker the firing toreward delivery.
Their most provocative results concernthe activity of the dopamine cells beforethe time of potential reward delivery. Inprevious work carried out at the two ex-treme probabilities, the firing rate was sta-ble. By exploring intermediate probabili-ties, Fiorillo et al. reveal a striking new pat-tern: The activity of dopamine neurons in-creased before the potential delivery of anuncertain reward. In contrast to the briefupswings in firing triggered by reward-predicting stimuli and unexpected re-wards, the population firing rate rosesteadily throughout most of the 2-secondpresentation of the conditioned stimuluswhen the probability of reward was 0.5, at-taining a higher rate than when the rewardprobability was 0.25 or 0.75.
The authors propose that the sustainedfiring preceding the time of potential re-ward delivery tracked the uncertainty ofthe reward. The onset of the conditionedstimuli signaling probabilities of either 0.0or 1.0 provided the monkey with definitiveinformation as to whether the rewardwould be delivered; if the meaning of thestimuli had been fully learned, then uncer-tainty about reward occurrence followingstimulus onset would be zero. In contrast, areward was equally likely to be delivered oromitted when the probability was 0.5, andthe monkey should have been maximallyuncertain about reward occurrence. Whenthe reward probability was 0.25 or 0.75,uncertainty was intermediate; if the mon-key had bet on the occurrence of the rewardfollowing stimulus onset, it could havewon, on average, three trials out of everyfour.
Regarding reinforcement learning mod-els, the phasic response of the dopamineneurons encodes a prediction error that isused as a “teaching signal” to improve fu-ture predictions. The incremental adjust-ment of the weights causes the teachingsignal to move backward in time toward theonset of the earliest stimulus that reliablypredicts the occurrence of a reward (and/orits potential time of delivery). How the sus-tained response preceding potential rewarddelivery could be incorporated into suchmodels is hard to see. If the dopamine sig-nal serves as the teacher, and the sustainedcomponent is not filtered out, how couldthe sustained component remain stationaryin time and amplitude over many trials?This problem is a knotty one because thesustained and phasic signals do not appearto be carried by independent populations ofneurons. Thus, postsynaptic elements arelikely to register the combined impact of
CS
Reward
P U
1.0
0.5
0.0
0.0
1.0
0.0
Anticipating a reward. Electrophysiological re-sponses of single midbrain dopamine neurons(yellow) were recorded while monkeys viewed acomputer monitor. Unique visual stimuli wereassociated with different probabilities of a re-ward (a drop of syrup). Rewards were presentedwith different probabilities (P) when the condi-tioned stimulus was switched off (CS offset).Theuncertainty (U) of the reward varied as an in-verted U-shaped function of probability. When P= 0 or P = 1, the monkey is certain that rewarddelivery will or will not accompany CS offset. Incontrast, when P = 0.5, the onset of the CS pro-vides no information about whether reward willor will not occur, but it does predict the potentialtime of reward delivery. The interval betweensuccessive CSs varied unpredictably (not shown),and thus the onset of the CS is the earliest reli-able predictor of the occurrence and/or the po-tential time of reward delivery. Once the mean-ing of the stimuli has been learned, the popula-tion of dopamine neurons responds to CS onsetwith a brief increase in activity when P = 1.0, butreceipt of the expected reward does not provokea strong change in firing. When P = 0, CS onsetproduced little response; if, however, the investi-gator violated expectation by delivering a re-ward, the dopamine cells responded with a brief,vigorous increase in firing.When P = 0.5 (and un-certainty about reward occurrence is maximal), aslow, steady increase in firing is seen prior to thetime of potential reward delivery.C
RED
IT:K
ATH
ARI
NE
SUTL
IFF/
SCIE
NC
E
www.sciencemag.org SCIENCE VOL 299 21 MARCH 2003
P E R S P E C T I V E S
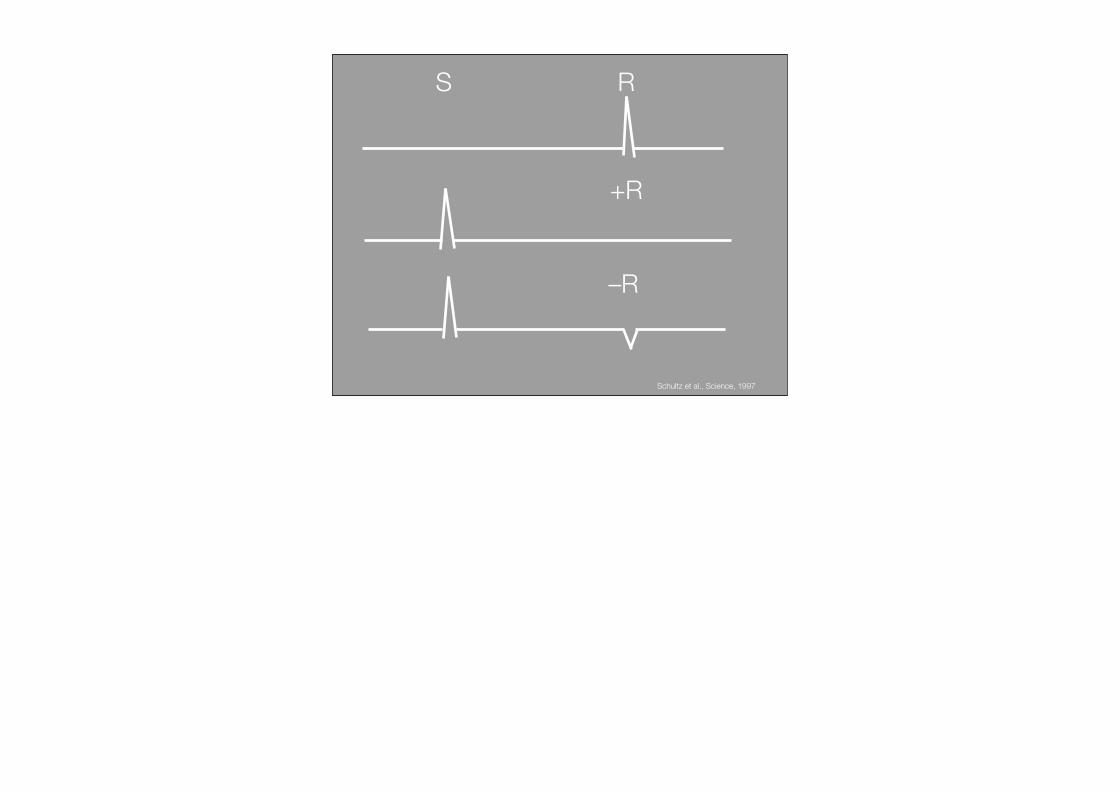
–R
S R
+R
Schultz et al., Science, 1997
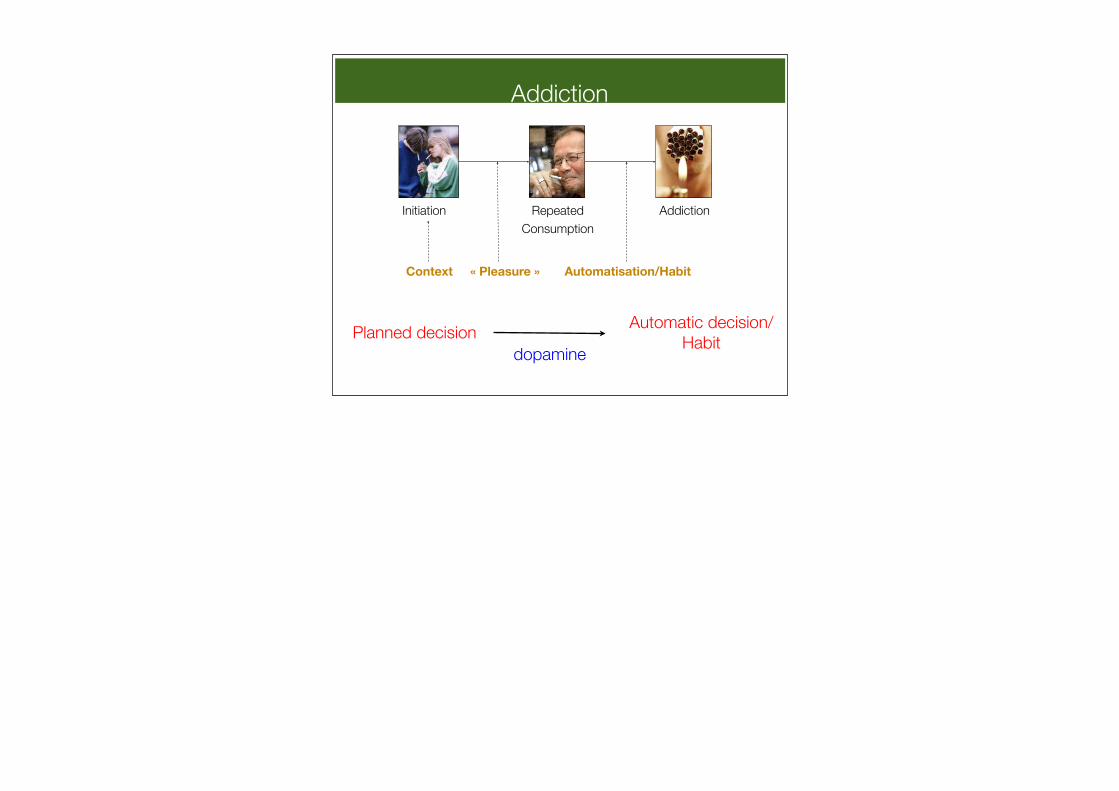
Addiction
Planned decision Automatic decision/Habit
Context
Initiation AddictionRepeated Consumption
« Pleasure » Automatisation/Habit
dopamine

Nicotine Gambling ChocolateCocaine Morphine
Addiction
Compulsion/habit
Mesolimbic dopamine ↑
Addiction

Animal models to study addiction

Memory trace in rodents600
400
200
0
seconds
PRE TEST
**A B A B
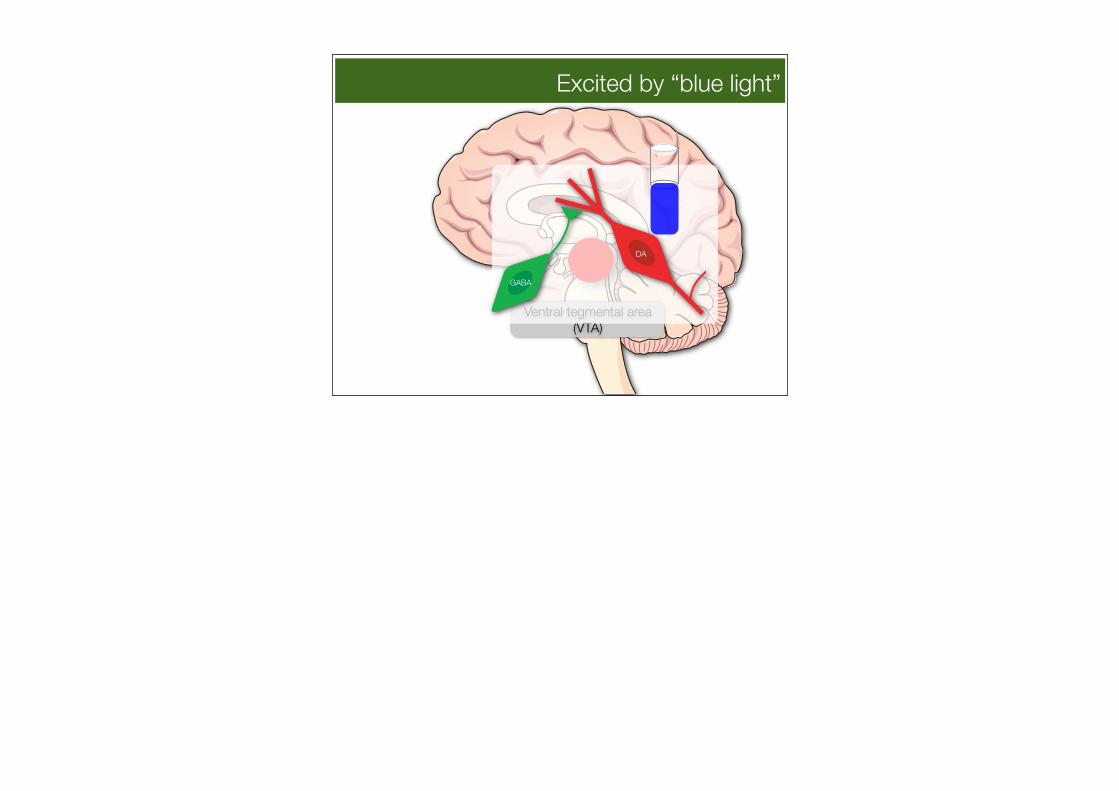
Excited by “blue light”
Ventral tegmental area (VTA)
GABA
DA

Nicotine Gambling ChocolateCocaine Morphine
Addiction
Compulsion/habit
Hijacking of learning and memory
Mesolimbic dopamine ↑
Addiction
Synaptic plasticity
And here is what we have done over the last few years to experimentally test this hypothesis.
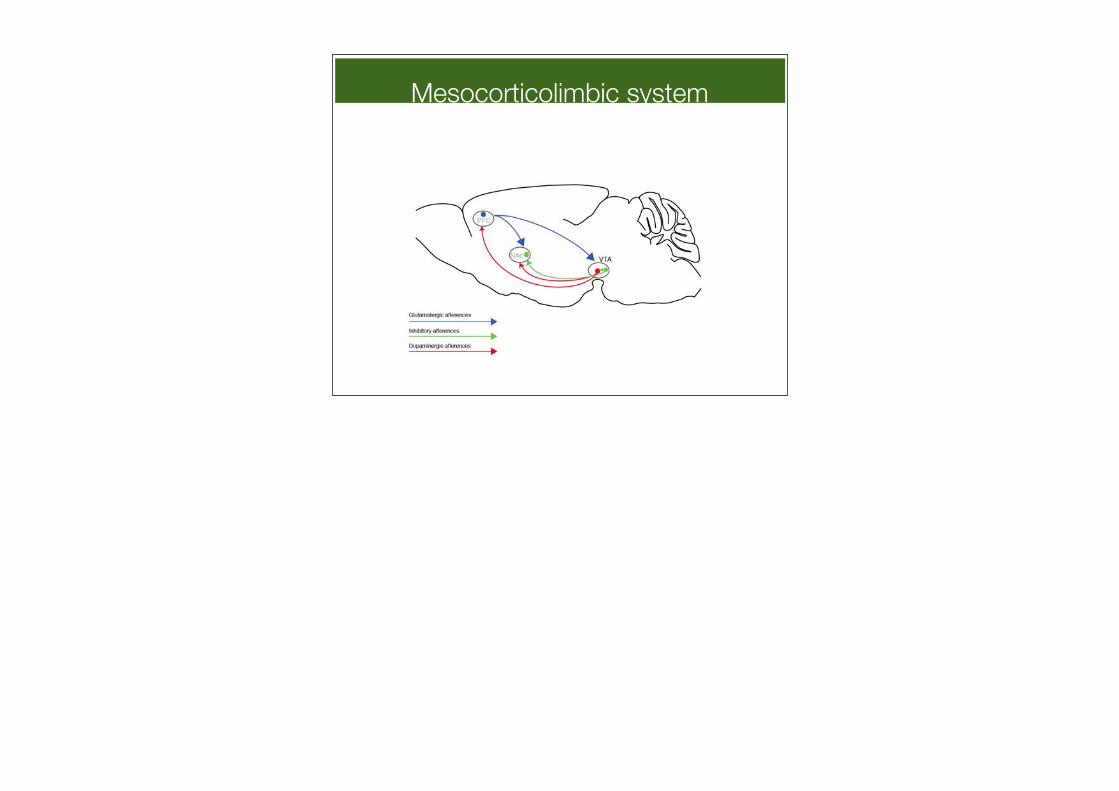
Mesocorticolimbic system
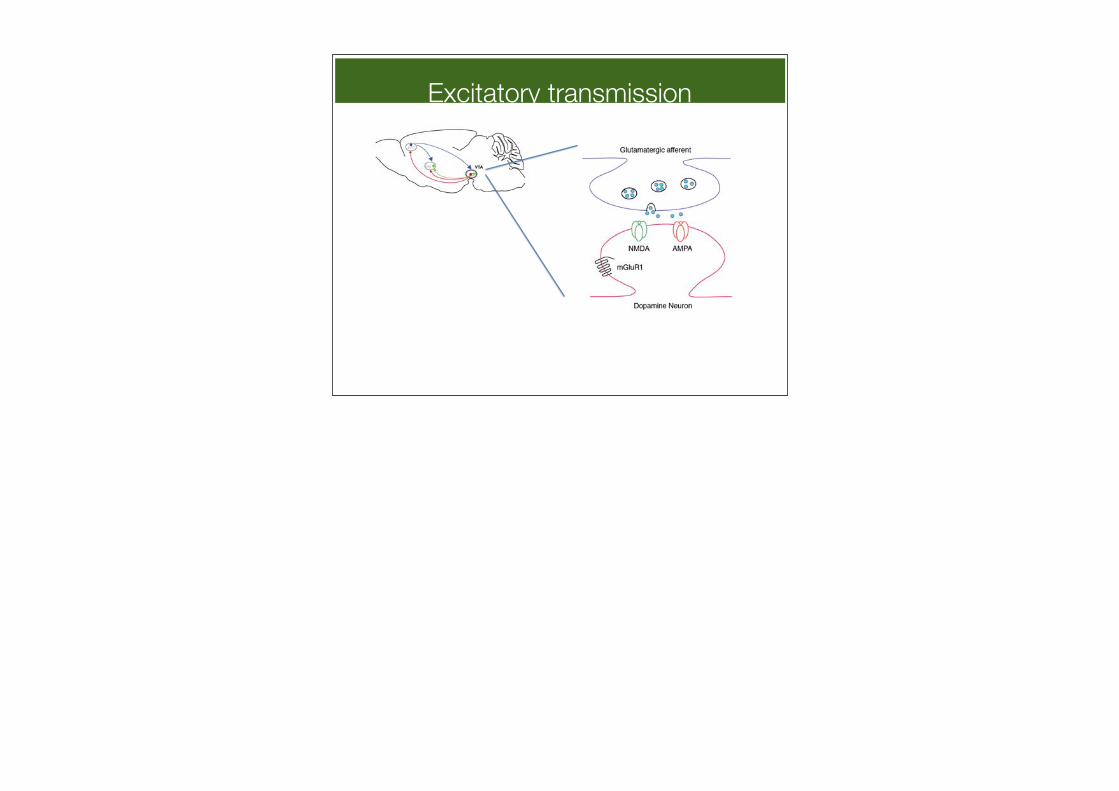
Excitatory transmission
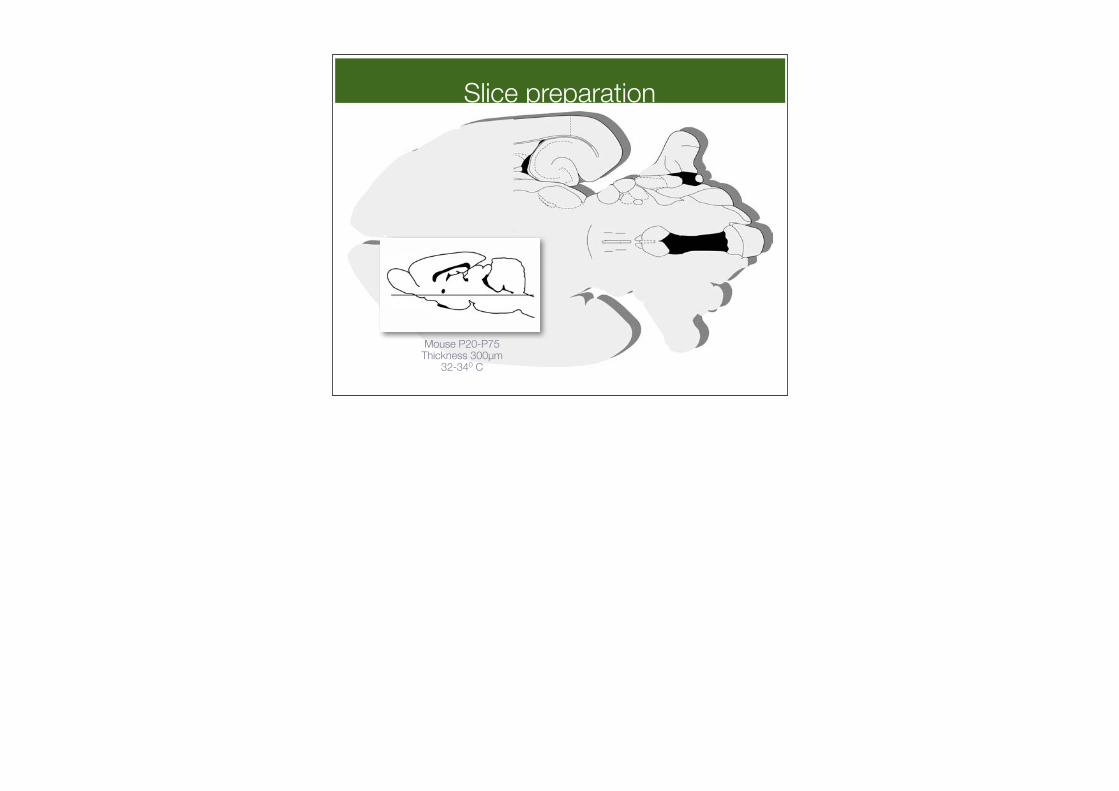
Rat age 10-18 daysThickness 300µm
Mouse P20-P75Thickness 300µm
32-340 C
Slice preparation

Slice preparation
• pitx3-GFP+/–
• Meng Li, London
VTA

The synapse
Boutonsynaptique
Dendrite
Récepteur Glutamate
0 mV
10 ms
5 mV

Increase in neurotransmitter
Glutamate
0 mV
10 ms
5 mV
Récepteur

Increase in number of Receptors
Glutamate
0 mV
10 ms
5 mV
Récepteur
Synaptic plasticity

Cocaine:increase in AMPA/NMDA
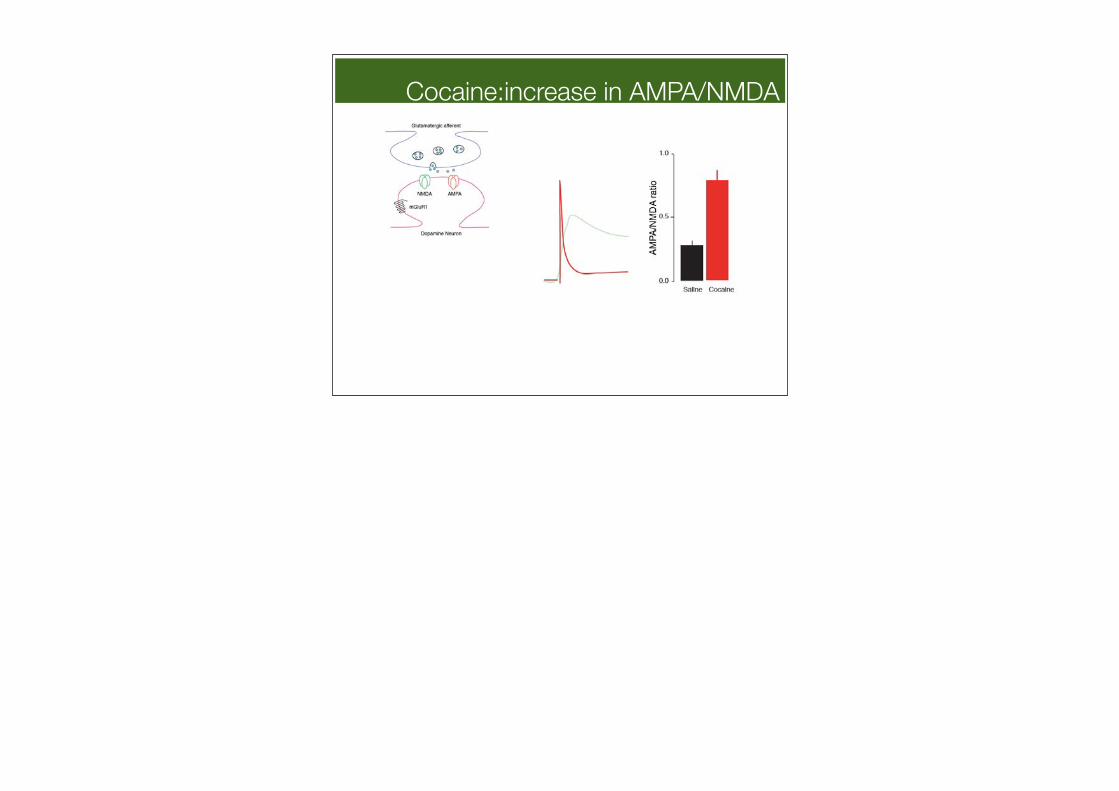
Cocaine:increase in AMPA/NMDA
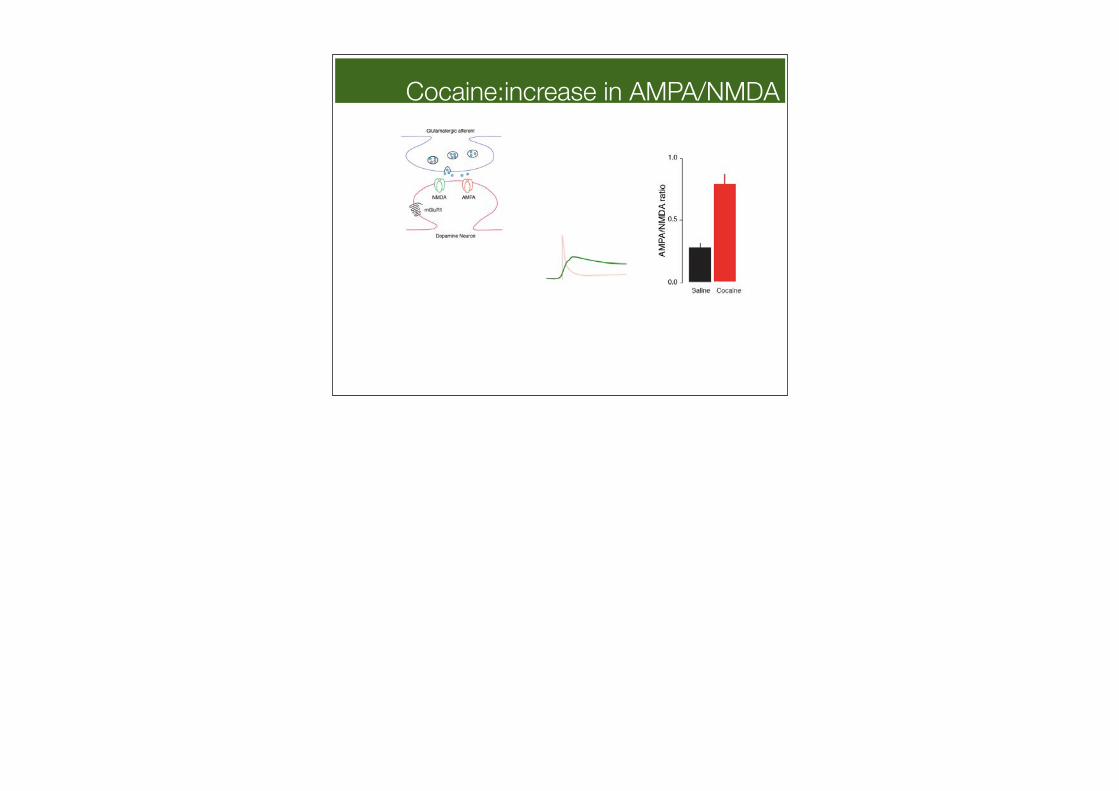
Cocaine:increase in AMPA/NMDA
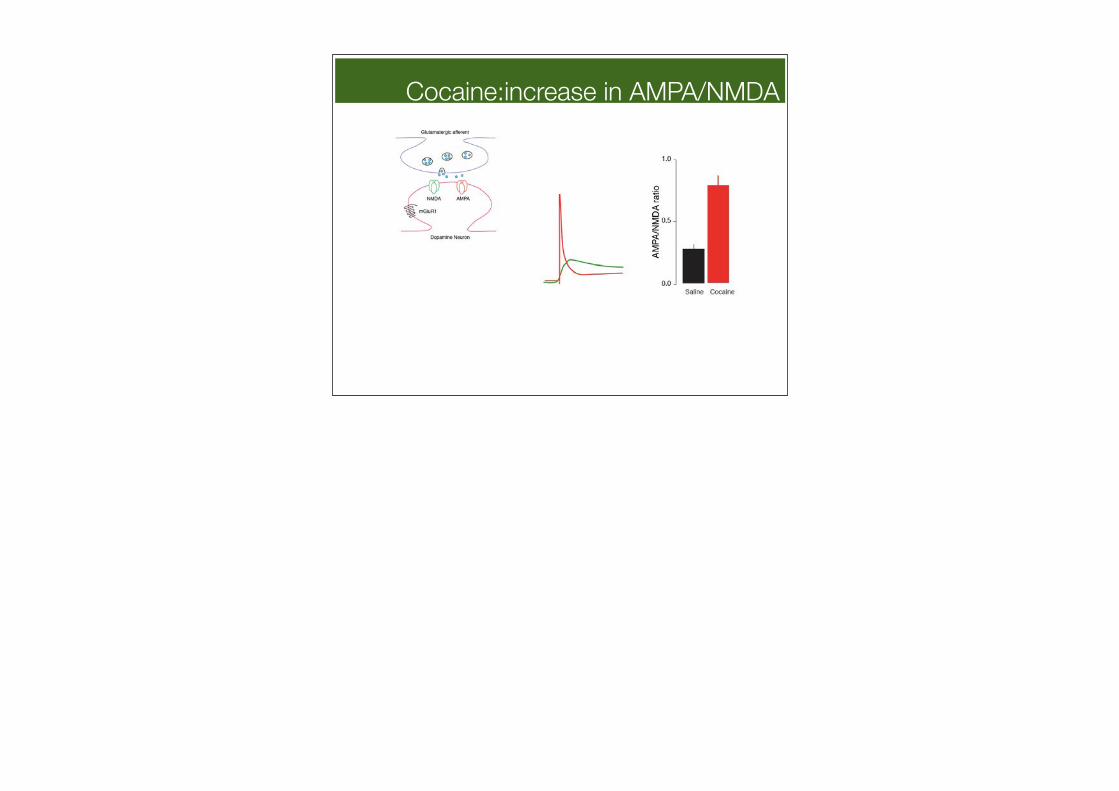
Cocaine:increase in AMPA/NMDA
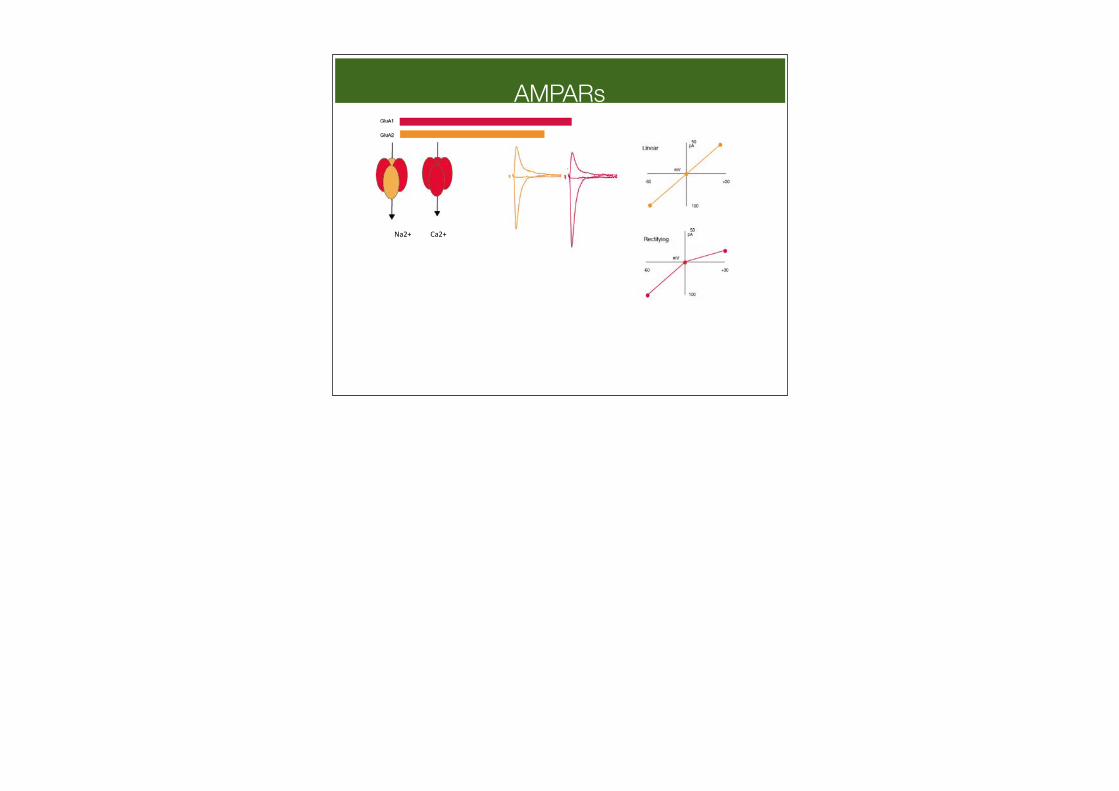
AMPARs
Na2+% Ca2+%

Common synaptic adaptation
stresssaline cocainemorphine nicotine
-100
-50
50
100
-80 -40 40
n = 8 -100
-50
50
100
-80 -40 40
n = 8 -100
-50
50
100
-80 -40 40
n = 9 -100
-50
50
100
-80 -40 40
n = 10 -100
-50
50
100
-80 -40 40
n = 9
pA
mV
in d-APV

Common synaptic adaptation
stresssaline cocainemorphine nicotine
-100
-50
50
100
-80 -40 40
n = 8 -100
-50
50
100
-80 -40 40
n = 8 -100
-50
50
100
-80 -40 40
n = 9 -100
-50
50
100
-80 -40 40
n = 10 -100
-50
50
100
-80 -40 40
n = 9
pA
mV
Class IIIClass I Class II
Bellone et al., 2006; Brown et al., 2010

AMPAR subunit composition
Top Related