Transcranial Low-Level Laser Therapy Improves Brain
11
Transcranial low-level laser therapy improves brain mitochondrial function and cognitive impairment in D-galactoseeinduced aging mice Farzad Salehpour a, b , Nahid Ahmadian a , Seyed Hossein Rasta b, c, d , Mehdi Farhoudi a , Pouran Karimi a , Saeed Sadigh-Eteghad a, * a Neurosciences Research Center (NSRC), Tabriz University of Medical Sciences, Tabriz, Iran b Department of Medical Physics, Tabriz University of Medical Sciences, Tabriz, Iran c Department of Medical Bioengineering, Tabriz University of Medical Sciences, Tabriz, Iran d School of Medical Sciences, University of Aberdeen, Aberdeen, UK article info Article history: Received 5 December 2016 Received in revised form 11 May 2017 Accepted 29 June 2017 Available online 6 July 2017 Keywords: Transcranial low-level laser therapy D-galactose Aging Mitochondrial function Episodic-like memory Spatial memory Apoptosis abstract Mitochondrial function plays a key role in the aging-related cognitive impairment, and photo- neuromodulation of mitochondria by transcranial low-level laser therapy (LLLT) may contribute to its improvement. This study focused on the transcranial LLLT effects on the D-galactose (DG)einduced mitochondrial dysfunction, apoptosis, and cognitive impairment in mice. For this purpose, red and near- infrared (NIR) laser wavelengths (660 and 810 nm) at 2 different fluencies (4 and 8 J/cm 2 ) at 10-Hz pulsed wave mode were administrated transcranially 3 d/wk in DG-received (500 mg/kg/subcutane- ous) mice model of aging for 6 weeks. Spatial and episodic-like memories were assessed by the Barnes maze and What-Where-Which (WWWhich) tasks. Brain tissues were analyzed for mitochondrial func- tion including active mitochondria, adenosine triphosphate, and reactive oxygen species levels, as well as membrane potential and cytochrome c oxidase activity. Apoptosis-related biomarkers, namely, Bax, Bcl- 2, and caspase-3 were evaluated by Western blotting method. Laser treatments at wavelengths of 660 and 810 nm at 8 J/cm 2 attenuated DG-impaired spatial and episodic-like memories. Also, results showed an obvious improvement in the mitochondrial function aspects and modulatory effects on apoptotic markers in aged mice. However, same wavelengths at the fluency of 4 J/cm 2 had poor effect on the behavioral and molecular indexes in aging model. This data indicates that transcranial LLLT at both of red and NIR wavelengths at the fluency of 8 J/cm 2 has a potential to ameliorate aging-induced mitochondrial dysfunction, apoptosis, and cognitive impairment. Ó 2017 Elsevier Inc. All rights reserved. 1. Introduction Aging is a complicated and multifactorial biological process that is characterized by a gradual decline in many organ functions. Brain aging is one of the important aspects of the aging progression which is associated with behavioral deficits, memory loss, and cognitive impairment (Grady, 2012). Mitochondria are a main apparatus in the cell respiratory chain, the dysfunction of which during aging is associated with cognitive impairment (Bishop et al., 2010). The evidence also suggests that perturbation in mitochondrial markers such as adenosine triphosphate (ATP) and membrane potential reduction or reactive oxygen species (ROS) generation, play the active role in the brain aging and age-related cognitive impairments (Jang and Van Remmen, 2009; Navarro et al., 2002). Furthermore, an imbalance between ROS synthesis and antioxidant activities during aging, triggers mitochondrial apoptotic pathways via Bax-, Bcl-2e, and caspase-3edependent mechanisms in the brain which finally leads to neuronal death (Niu et al., 2010; Pollack et al., 2002). D-galactose (DG) is a reducing sugar in the body that can be metabolized at the normal physiological concentrations. Never- theless, at high concentration, it is able to convert into hydrogen peroxide and aldose which results in the generation of ROS and causes oxidative stress (Ho et al., 2003). Chronic administration of DG has been used to induce brain oxidative stress and cognitive dysfunction to mimic natural aging in rodent (Lei et al., 2008; Pourmemar et al., 2017; Tsai and Yin, 2012; Zhang et al., 2008). Moreover, long-term administration of DG triggers mitochondrial * Corresponding author at: Neurosciences Research Center (NSRC), Tabriz Uni- versity of Medical Sciences, 5166614756 Tabriz, Iran. Tel.: þ984133340730; fax: 984133340730. E-mail address: [email protected] (S. Sadigh-Eteghad). Contents lists available at ScienceDirect Neurobiology of Aging journal homepage: www.elsevier.com/locate/neuaging 0197-4580/$ e see front matter Ó 2017 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.neurobiolaging.2017.06.025 Neurobiology of Aging 58 (2017) 140e150
Transcript of Transcranial Low-Level Laser Therapy Improves Brain

Transcranial low-level laser therapy improves brain mitochondrialfunction and cognitive impairment in D-galactoseeinducedaging mice
Farzad Salehpour a,b, Nahid Ahmadian a, Seyed Hossein Rasta b,c,d, Mehdi Farhoudi a,Pouran Karimi a, Saeed Sadigh-Eteghad a,*
aNeurosciences Research Center (NSRC), Tabriz University of Medical Sciences, Tabriz, IranbDepartment of Medical Physics, Tabriz University of Medical Sciences, Tabriz, IrancDepartment of Medical Bioengineering, Tabriz University of Medical Sciences, Tabriz, Irand School of Medical Sciences, University of Aberdeen, Aberdeen, UK
a r t i c l e i n f o
Article history:Received 5 December 2016Received in revised form 11 May 2017Accepted 29 June 2017Available online 6 July 2017
Keywords:Transcranial low-level laser therapyD-galactoseAgingMitochondrial functionEpisodic-like memorySpatial memoryApoptosis
a b s t r a c t
Mitochondrial function plays a key role in the aging-related cognitive impairment, and photo-neuromodulation of mitochondria by transcranial low-level laser therapy (LLLT) may contribute to itsimprovement. This study focused on the transcranial LLLT effects on the D-galactose (DG)einducedmitochondrial dysfunction, apoptosis, and cognitive impairment in mice. For this purpose, red and near-infrared (NIR) laser wavelengths (660 and 810 nm) at 2 different fluencies (4 and 8 J/cm2) at 10-Hzpulsed wave mode were administrated transcranially 3 d/wk in DG-received (500 mg/kg/subcutane-ous) mice model of aging for 6 weeks. Spatial and episodic-like memories were assessed by the Barnesmaze and What-Where-Which (WWWhich) tasks. Brain tissues were analyzed for mitochondrial func-tion including active mitochondria, adenosine triphosphate, and reactive oxygen species levels, as well asmembrane potential and cytochrome c oxidase activity. Apoptosis-related biomarkers, namely, Bax, Bcl-2, and caspase-3 were evaluated by Western blotting method. Laser treatments at wavelengths of 660and 810 nm at 8 J/cm2 attenuated DG-impaired spatial and episodic-like memories. Also, results showedan obvious improvement in the mitochondrial function aspects and modulatory effects on apoptoticmarkers in aged mice. However, same wavelengths at the fluency of 4 J/cm2 had poor effect on thebehavioral and molecular indexes in aging model. This data indicates that transcranial LLLT at both of redand NIR wavelengths at the fluency of 8 J/cm2 has a potential to ameliorate aging-induced mitochondrialdysfunction, apoptosis, and cognitive impairment.
! 2017 Elsevier Inc. All rights reserved.
1. Introduction
Aging is a complicated and multifactorial biological process thatis characterized by a gradual decline in many organ functions. Brainaging is one of the important aspects of the aging progressionwhichis associated with behavioral deficits, memory loss, and cognitiveimpairment (Grady, 2012). Mitochondria are a main apparatus inthe cell respiratory chain, the dysfunction of which during aging isassociated with cognitive impairment (Bishop et al., 2010). Theevidence also suggests that perturbation in mitochondrial markerssuch as adenosine triphosphate (ATP) and membrane potential
reduction or reactive oxygen species (ROS) generation, play theactive role in the brain aging and age-related cognitive impairments(Jang andVan Remmen, 2009; Navarro et al., 2002). Furthermore, animbalance between ROS synthesis and antioxidant activities duringaging, triggers mitochondrial apoptotic pathways via Bax-, Bcl-2e,and caspase-3edependent mechanisms in the brain which finallyleads to neuronal death (Niu et al., 2010; Pollack et al., 2002).
D-galactose (DG) is a reducing sugar in the body that can bemetabolized at the normal physiological concentrations. Never-theless, at high concentration, it is able to convert into hydrogenperoxide and aldose which results in the generation of ROS andcauses oxidative stress (Ho et al., 2003). Chronic administration ofDG has been used to induce brain oxidative stress and cognitivedysfunction to mimic natural aging in rodent (Lei et al., 2008;Pourmemar et al., 2017; Tsai and Yin, 2012; Zhang et al., 2008).Moreover, long-term administration of DG triggers mitochondrial
* Corresponding author at: Neurosciences Research Center (NSRC), Tabriz Uni-versity of Medical Sciences, 5166614756 Tabriz, Iran. Tel.: þ984133340730; fax:984133340730.
E-mail address: [email protected] (S. Sadigh-Eteghad).
Contents lists available at ScienceDirect
Neurobiology of Aging
journal homepage: www.elsevier .com/locate/neuaging
0197-4580/$ e see front matter ! 2017 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.neurobiolaging.2017.06.025
Neurobiology of Aging 58 (2017) 140e150

dysfunction and cytochrome C release which eventually causeneuronal apoptosis (Cui et al., 2006; Lan et al., 2012). Many studiesshowed that mitochondria-protective components are able toimprove the functioning of the nervous system and delay brainaging (Liu and Ames, 2005; Liu et al., 2002). Furthermore,nonpharmaceutical therapies for treatment or prevention ofage-related disorders have also aroused significant interest amongresearchers in the recent years (Gonzalez-Lima et al., 2014; Salgadoet al., 2015).
Transcranial low-level laser therapy (LLLT) with far-red to near-infrared (NIR) light is a novel noninvasive modality used tomodulate neuronal activity in animal and human studies (Barrettand Gonzalez-Lima, 2013; Chung et al., 2012; Salehpour andRasta, 2017; Xuan et al., 2014). Over the past decade, transcranialLLLT has been suggested to treat different neurodegenerative(Lapchak, 2012; Purushothuman et al., 2014) and psychiatric dis-orders (Salehpour et al., 2016; Schiffer et al., 2009), given its goodsafety profile and efficiency. Light at specific wavelengths between600 and 1100 nm has maximum penetration depth in skull(Bashkatov et al., 2006; Firbank et al., 1993). In addition, thisspectral region corresponds to light absorption by mitochondrialchromophores such as cytochrome c oxidase (CCO), which is theterminal enzyme of the respiratory chain (Karu, 2014). This pho-toabsorption process causes more electron transfer through themitochondrial respiratory chain and leading to increasing of ATPproduction (Mochizuki-Oda et al., 2002; Oron et al., 2007). Conse-quently, further secondary cellular effects such as modulation ofROS and increase in the mitochondrial membrane potential (MMP)could occur (de Freitas and Hamblin, 2016). There is some evidencethat shows photoneuromodulation can exert neuroprotective ef-fects via reduction of neuronal cells apoptosis in the braincognition-related compartments (De Taboada et al., 2011; Grilloet al., 2013; Lu et al., 2016; Uozumi et al., 2010). Also, a clinicalstudy has showed procognitive effects of photoneuromodulation byred/NIR transcranial LLLT in traumatic brain injury (TBI) patients(Naeser et al., 2011). An improvement in the neurologic condition ofpatients in a persistent vegetative state following NIR light treat-ment was also reported (Nawashiro et al., 2012). Furthermore, NIRtranscranial LLLT enhanced some aspects of cognition in healthysubjects (Barrett and Gonzalez-Lima, 2013).
The aim of this study is to evaluate the effects of transcranialLLLT using red and NIR lights on the DG-induced learning andmemory impairment and possible impact of mitochondrialdysfunction and apoptosis.
2. Materials and methods
2.1. Animals
Seventy-two adult male BALB/c mice from the animal center ofTabriz University of Medical Sciences (TUOMS), weighing 25e30 gwere used for this study. Before and during the study, mice weresocially housed in standard cages (5 in each cage) and kept on a 12/12-h light/dark cycle at a temperature of 23 "C # 2 "C, with freeaccess to tap water and standard pellet food ad libitum. All animalswere weighed weekly during the experiment. All of the procedureswere conducted in conformity with the Guide for the Care and Useof Laboratory Animals of the National Institutes of Health (NIH;Publication No. 85-23, revised 1985) and approved by the regionalethics committee of TUOMS (No: IR.TBZMED.REC.1395.445).
2.2. Experimental design
After a week of adaptation, mice were randomized into thecontrol (n ¼ 12) and aging-induced groups (n ¼ 60). The control
group received normal saline (0.9% NaCl, 0.2 mL/mice). Aging wasinduced by injection of 500 mg/kg of DG (Sigma, St. Louis, MO, USA)through the subcutaneous (s.c.) route once daily for 6 weeks.Separate groups of DG-injected mice received sham, red, or NIRlaser irradiation transcranially at 2 fluencies of 4 or 8 J/cm2, 3 d/wk,which was named as following: (1) DGþsham (DG injection withsham laser irradiation); (2) DGþred4 (DG injection with 4 J/cm2 ofred laser irradiation); (3) DGþNIR4 (DG injection with 4 J/cm2 ofNIR laser irradiation); (4) DGþred8 (DG injectionwith 8 J/cm2 of redlaser irradiation); and (5) DGþNIR8 (DG injection with 8 J/cm2 ofNIR laser irradiation).
All tests and analysis were performed by an experimenter whowas unaware of the entity of experimentations. Fig. 1 shows thestudy design, including procedures and time periods.
2.3. Laser treatment
A commercially available therapeutic Diode laser (Thor Photo-medicine, Chesham, UK) consisting of a red probe (AlGaInP) with660 nm wavelength and an NIR probe (GaAlAs) with 810 nmwavelength were used for irradiation sessions. The laser wasoperated at the maximum output power of 200 mW, a powerdensity of 4.75W/cm2, and 10-Hz pulsed wavemodewith 88% dutycycle for both probes. For laser irradiation, each mouse was heldfirmly by the dorsal skin of the neck, and its head was carefullylocated. The laser probe was positioned over the head in contactwith the scalp that covered the entire mouse brain. To determinethe delivered dose on the cortex, transmission values of red and NIRlight through the fresh postmortem scalp and skull were measuredby a 7-mm diameter power meter sensor of the laser device.Average energy densities (fluencies) of 4 and 8 J/cm2 per eachsessionwere delivered to the animal’s cortical surfacewith both redand NIR lasers. Transcranial LLLT was administered 3 times a week,with a minimum of 48 hours between the sessions, over a period of6 weeks. Laser treatment was done at least 5 hours after DG in-jections. Sham-treated mice (DGþsham laser group) were under-gone identical to laser treatment procedure except that the laserdevice was not turned on.
2.4. Barnes maze test
2.4.1. ApparatusThe spatial learning andmemory test was performed in a Barnes
maze (Sunyer et al., 2007). The maze was made of wood and con-sisted of a circular platform 100 cm in diameter raised 50 cm fromthe ground. Twenty circular holes, 5 cm in diameter, were locatedthrough the platform 3 cm from the perimeter. A movable plasticescape box (20 % 15 % 5 cm) was placed under the escape hole. Themaze was placed in a roomwhere distinct spatial cues were locatedon the walls. An 80-dB white noise was used as negative stimuliduring the test.
Fig. 1. Timescale of D-galactose or normal saline administration, laser treatments,behavioral tests, and sampling. Figure was created in the Mind the Graph platform,www.mindthegraph.com. Abbreviation: LLLT, low-level laser therapy.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150 141
2.4.2. Task sessionsThe task consisted of 3 sessions including adaptation, training,
and probe sessions which lasted for 5 days. The first day consistedof an adaptation session and 4 trials and 3 subsequent daysconsisting of 4 trials per day, each separated by a 3-minute in-terval. In the adaptation session, a mouse was located in thecenter of the maze in a start chamber. After 10 seconds, simul-taneous with white noise turning on, the start chamber wasremoved and the mouse was set free to explore the maze for3 minutes to enter into the escape box. When the mouse enteredthe escape box, the white noise was turned off and the animal wasallowed to remain in the box for 1 minute. On the fifth day, aprobe trial was carried out without the escape box. Between alltest sessions, the maze and escape box were cleaned with 70%alcohol to remove odor cues. A digital video camera was securedabove the maze. The latency time (the period it takes the animalto find the scape box), and time spent in error holes (the sum ofperiods that animal spent around all of the holes except targethole) during the training session, and time spent in the targetquadrant (where the escape box was located during trainingsession), and relative error time (a ratio of time spending in targethole to time spending in error holes) during the probe session
were extracted using a video tracking program Etho Vision (Nol-dus, The Netherlands).
2.5. What-Where-Which (WWWhich) task
2.5.1. ApparatusThe episodic-like memory was evaluated by WWWhich task
(Davis et al., 2013). The experimental apparatus consisted of 2open-field arena made of Plexiglas (30 % 30 % 25 cm). The arenaswere altered into two different contexts. In context 1, the floor wasattached to a LEGO base plate and the walls were painted mattblack. While context 2 consisted of a smooth matte black floor andwalls were painted with black and white vertical stripes. All objectsfor testing were assembled from LEGO.
2.5.2. Task sessionsAfter 24 hours of the last treatments, the WWWhich task was
performed. The task consisted of 3 sessions including habituation,exposure, and test sessions (Davis et al., 2013). In the habituationsession, mice were habituated singly to each context for 5 minutesfor 1 day, and locomotor activity was recorded through the sessionin one of the contexts (same context for all animals). During the
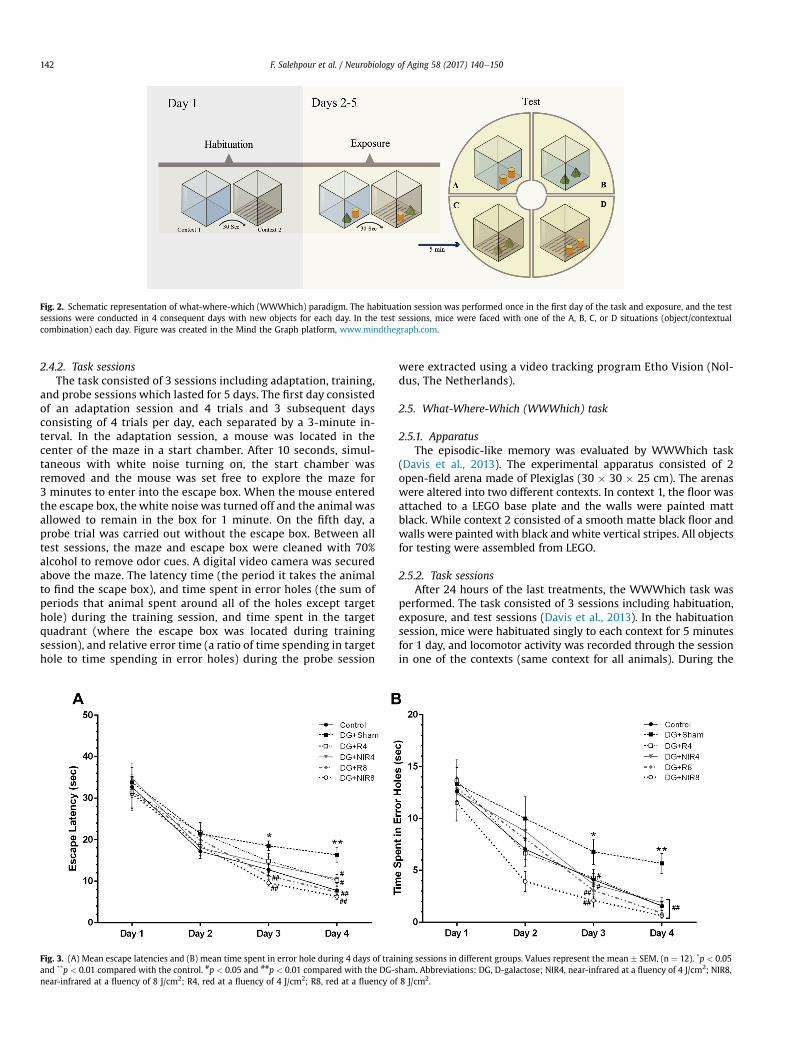
Fig. 2. Schematic representation of what-where-which (WWWhich) paradigm. The habituation session was performed once in the first day of the task and exposure, and the testsessions were conducted in 4 consequent days with new objects for each day. In the test sessions, mice were faced with one of the A, B, C, or D situations (object/contextualcombination) each day. Figure was created in the Mind the Graph platform, www.mindthegraph.com.
Fig. 3. (A) Mean escape latencies and (B) mean time spent in error hole during 4 days of training sessions in different groups. Values represent the mean # SEM, (n ¼ 12). *p < 0.05and **p < 0.01 compared with the control. #p < 0.05 and ##p < 0.01 compared with the DG-sham. Abbreviations: DG, D-galactose; NIR4, near-infrared at a fluency of 4 J/cm2; NIR8,near-infrared at a fluency of 8 J/cm2; R4, red at a fluency of 4 J/cm2; R8, red at a fluency of 8 J/cm2.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150142

exposure sessions, 2 different objects (A and B) were first placed inthe context 1 and mice were given 3 minutes to explore it. Next,mouse was placed in the holding cage for 30 seconds and the ob-jects reversely (B and A) placed in context 2. Next, mice wereallowed to explore it for 3 minutes and then mice were returned tothe holding cage. The test session was performed after 5 minutesinterval. Mice were encountered with 2 copies of one of the objects(A or B) were placed in one of the 2 contexts (1 or 2) for 3 minutes.Exposure and test sessions were performed on one trail for 4consequence days with new objects for each day (Fig. 2). Explora-tion was defined as time spent with the nose oriented toward andwithin 1 cm of the object. Circling or sitting on the object were notconsidered as exploratory behavior. The episodic-like memory(object/contextual combination) was evaluated by displacementindex (DI). The DI was calculated for each group as follow: DI¼ (N&F)/(Nþ F), where N is the exploration time of the novel object and Fis the exploration time of the familiar object. After each trial, thecontexts and subjects were cleaned thoroughly using 70% ethanolto eliminate the presence of any olfactory cues. All experimentalsessions were videotaped from above and behavioral data wereextracted from the videos using a video-tracking program EthoVision (Noldus, The Netherlands).
2.6. Brain mitochondrial isolation
Mice from all experimental groups were decapitated after deepanesthesia with a mixture of ketamine (100 mg/kg) and xylazine(10 mg/kg), at 24 hours after last behavioral test. The whole brain(excluding cerebellum) were quickly taken out from the skull andplaced in ice-cold isolation buffer containing 200-mM mannitol,70-mM sucrose, 10-mM HEPES, and 2-mM EDTA, pH 7.5. Then,entire brain tissues were homogenized in ice-cold extraction buffercontaining 2 mg/mL albumin (10% w/v). The samples were centri-fuged at 600 % g in 4 "C for 5 minutes. The supernatant liquid wastransferred into a new tube and centrifuged at 12,000% g in 4 "C for15 minutes. The pellet was resuspended in storage buffer contain-ing 10-mM HEPES, pH 7.4, 250-mM sucrose, 1-mM ATP, 0.08-mMADP, 5-mM sodium succinate, 2-mM K2HPO4, and 1-mM DTT.Protein in tissue or mitochondrial suspensions was determined byBradford method (Bradford, 1976).
2.7. Active mitochondria levels measurement
Mitochondria labeling were done using MitoTracker probe (Cellsignaling, USA) in the brain cells. The probe passively diffusedacross the plasma membrane and accumulates in active mito-chondria. A 100 mg of entire brain tissue was cut into 2e4 mmpieces and an appropriate amount of trypsin (0.1%) was added.Tissue fragments were washed 3 times with PBS containing 25%bovine serum albumin. A single cell suspension was obtained bytrituration of fragments in DMEM (Life technologies, Germany)containing 10% fetal calf serum (Integro, The Netherlands), 25,000IU/L penicillin, and 25 mg/L streptomycin (Sigma, Germany). TheMitoTracker Green was directly added into growth media at aconcentration 100e400 nM and incubated for 15e30 minutes at 37"C in 5% CO2. Next, mitochondria were isolated according to “brainmitochondrial isolation” session. The fluorescence intensity wasdetermined (lex ¼ 490 nm, lem ¼ 516 nm) in a fluorescencemicroplate reader. The active mitochondria level was representedas fluorescence intensity and normalized to samples protein.
2.8. Mitochondrial membrane potential assay
Change of MMP (Djm) in brain cells was evaluated using JC-1dye (Mitochondria Staining Kit [Sigma-Aldrich, St. Louis, MO,
USA]). In normal condition, JC-1 concentrates in the mitochondrialmatrix, where it forms red fluorescent aggregates. Any event thatdissipates the MMP leads to shifting from red to green fluorescence(JC-1 monomers). Stain and buffers were prepared based on the kitinstruction. Mitochondria were suspended in JC-1 stain in the finalconcentration of 0.6 mM. Fluorescence intensity in samples wasmeasured by fluorimetry method. The Djm calculated by the ratioof red (lex ¼ 490 nm, lem ¼ 590 nm) to green (lex ¼ 488 nm, lem ¼530 nm) fluorescence intensity in themitochondria suspension andnormalized to samples protein.
2.9. Mitochondrial cytochrome c oxidase activity
The CCO activity was determined using the commercial kit(CYTOCOX1, Sigma, USA). The assay in this kit is based on Lembergmethod (Lemberg, 1969) in which spectrophotometrically followedthe decrease in absorbance at 550 nm of ferrocytochrome c causedby its oxidation to ferricytochrome c by CCO. Cytochrome c wasmixed with a 0.1-M DTT solution to prepare its reduced form. Thereduction was confirmed by the change of color from dark orange-red to pale purple-red. Change in absorbance per min wasmeasured and results were expressed as nmol per min per mgprotein.
2.10. ATP colorimetric assay
The colorimetric assay kit (MAK190, Sigma, USA) was used formeasuring of ATP. Briefly, 10-mg tissue was homogenized in 100 mLof ATP assay buffer (provided in the kit). After adding ATP probe inthe presence of the developer (provided in the kit), the absorbanceat 570 nm were measured. The standard curve was used to obtainthe ATP concentration and it was presented in nmol per mg protein.
2.11. Brain mitochondrial ROS production
The fluorescent dye dichlorohydrofluorescein diacetate (DCFDA)was used to determine the ROS production level in brain mito-chondria (Novalija et al., 2003). The mitochondria were incubatedwith 2-mM DCFDA at 37 "C for 20 minutes. The fluorescence in-tensity was determined (lex ¼ 485 nm, lem ¼ 530 nm) in a fluo-rescence microplate reader. The ROS levels were represented asfluorescence intensity and normalized to samples protein.
2.12. Western blot
Expression of proteins in the brain tissue was analyzed byWestern blot method as described previously (Sadigh-Eteghadet al., 2015). Briefly, brain tissue was homogenized in radio-immunoprecipitation assay buffer containing a protease inhibitorcocktail. Proteins were separated using 12.5% polyacrylamide geland transferred onto a polyvinylidenedifluoride membrane (Roche,UK). Then, the membranes were incubated with anti-Bax (1:500,sc-493), Bcl-2 (1:500, sc-492), and caspase-3 (1:500, sc-7148) an-tibodies. Next, the membrane was incubated with the horseradishperoxidaseeconjugated secondary antibody (1:5000, sc-2004).Finally, membranes were placed in ECL prime Western blottingdetection reagent (Amersham, UK) and the signals visualized byexposure to autoradiography film (Kodak, USA). For internal con-trol, antieb-actin (1:500, sc-130656) antibody was used. All anti-bodies were purchased from Santa Cruz (USA). The signal intensityof each band was quantitated using ImageJ 1.62 software (NationalInstitutes of Health, USA).
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150 143
2.13. Statistical analysis
Descriptive data were expressed as mean # SEM. Comparisonof different groups was carried out by a one-way ANOVA fol-lowed by the post hoc Tukey test. All analyses were performedusing IBM SPSS statistics software (version 22 forWindows; SPSSInc, USA). p-values <0.05 were considered significant in allcomparisons.
3. Results
3.1. Barnes test
3.1.1. Training sessionThe one-way ANOVA followed by Tukey test showed that DG-
treated sham mice were delayed in finding the escape boxcompared with control group on the third and fourth days (p <
0.05, and p< 0.01, respectively) of the training session. Both the redand NIR laser treatments (at a fluency of 8 J/cm2) significantlyreduced latency time on the third day of training compared withDG-shammice (p < 0.01). On the fourth day of the training session,significant differences were also observed in 4 J/cm2 (p < 0.05) and8 J/cm2 (p < 0.01) of red and NIR laser groups compared with DG-sham group (Fig. 3A).
In addition, as shown in Fig. 3B, chronic administration of DG insham group significantly increased time spending in error holesover the last 2 days of the training session (p < 0.05, and p < 0.01,respectively). Time spent in error holes significantly diminished on
days 3 (p< 0.05 for 4 J/cm2; and p< 0.01 for 8 J/cm2) and 4 (p< 0.01for both doses) of the training session in red and NIR laser groupscompared with DG-sham group.
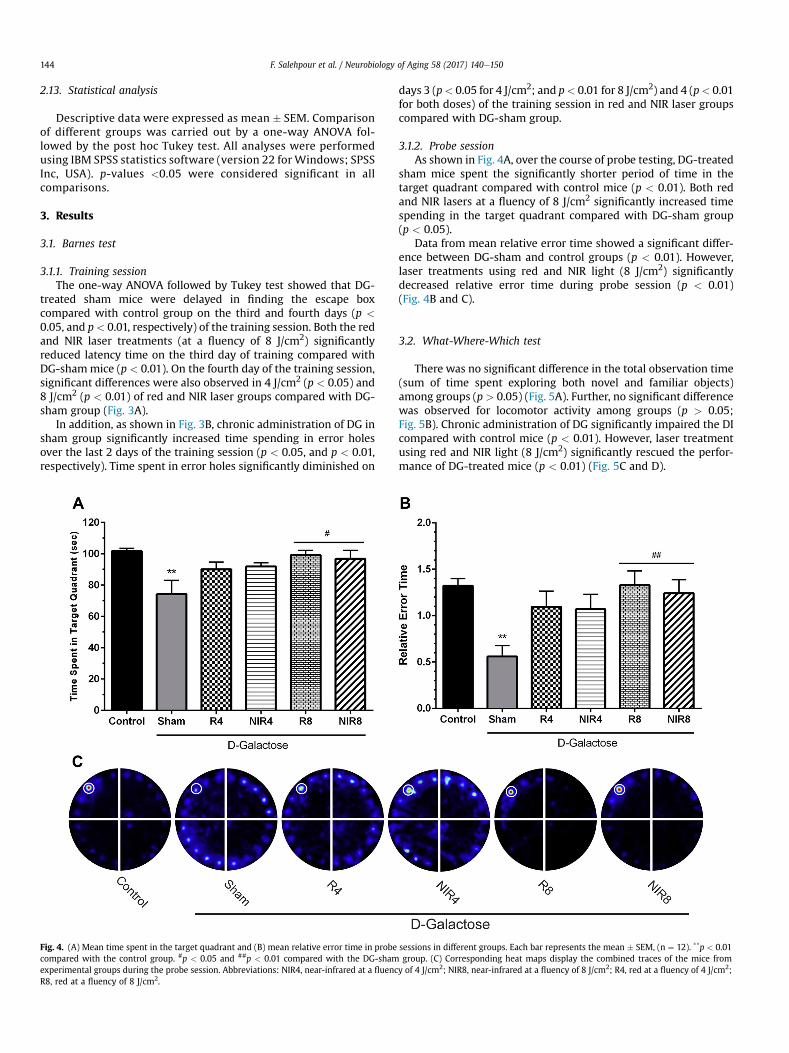
3.1.2. Probe sessionAs shown in Fig. 4A, over the course of probe testing, DG-treated
sham mice spent the significantly shorter period of time in thetarget quadrant compared with control mice (p < 0.01). Both redand NIR lasers at a fluency of 8 J/cm2 significantly increased timespending in the target quadrant compared with DG-sham group(p < 0.05).
Data from mean relative error time showed a significant differ-ence between DG-sham and control groups (p < 0.01). However,laser treatments using red and NIR light (8 J/cm2) significantlydecreased relative error time during probe session (p < 0.01)(Fig. 4B and C).
3.2. What-Where-Which test
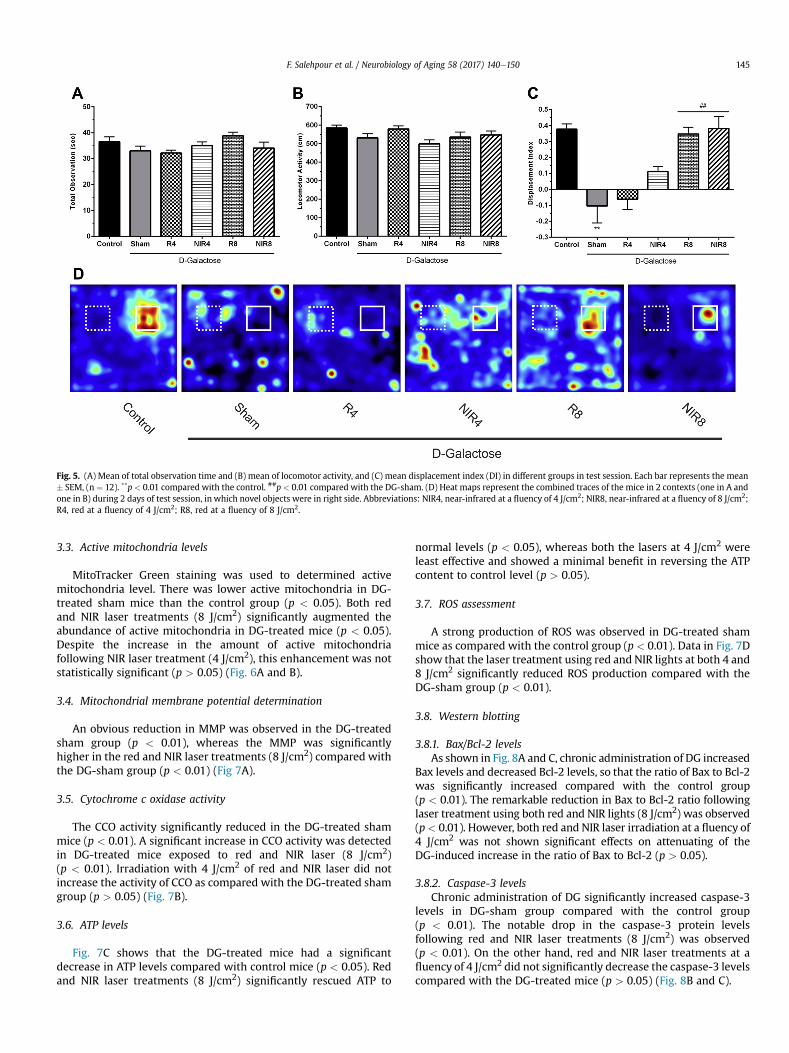
There was no significant difference in the total observation time(sum of time spent exploring both novel and familiar objects)among groups (p > 0.05) (Fig. 5A). Further, no significant differencewas observed for locomotor activity among groups (p > 0.05;Fig. 5B). Chronic administration of DG significantly impaired the DIcompared with control mice (p < 0.01). However, laser treatmentusing red and NIR light (8 J/cm2) significantly rescued the perfor-mance of DG-treated mice (p < 0.01) (Fig. 5C and D).
Fig. 4. (A) Mean time spent in the target quadrant and (B) mean relative error time in probe sessions in different groups. Each bar represents the mean # SEM, (n ¼ 12). **p < 0.01compared with the control group. #p < 0.05 and ##p < 0.01 compared with the DG-sham group. (C) Corresponding heat maps display the combined traces of the mice fromexperimental groups during the probe session. Abbreviations: NIR4, near-infrared at a fluency of 4 J/cm2; NIR8, near-infrared at a fluency of 8 J/cm2; R4, red at a fluency of 4 J/cm2;R8, red at a fluency of 8 J/cm2.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150144
3.3. Active mitochondria levels
MitoTracker Green staining was used to determined activemitochondria level. There was lower active mitochondria in DG-treated sham mice than the control group (p < 0.05). Both redand NIR laser treatments (8 J/cm2) significantly augmented theabundance of active mitochondria in DG-treated mice (p < 0.05).Despite the increase in the amount of active mitochondriafollowing NIR laser treatment (4 J/cm2), this enhancement was notstatistically significant (p > 0.05) (Fig. 6A and B).
3.4. Mitochondrial membrane potential determination
An obvious reduction in MMP was observed in the DG-treatedsham group (p < 0.01), whereas the MMP was significantlyhigher in the red and NIR laser treatments (8 J/cm2) compared withthe DG-sham group (p < 0.01) (Fig 7A).
3.5. Cytochrome c oxidase activity
The CCO activity significantly reduced in the DG-treated shammice (p < 0.01). A significant increase in CCO activity was detectedin DG-treated mice exposed to red and NIR laser (8 J/cm2)(p < 0.01). Irradiation with 4 J/cm2 of red and NIR laser did notincrease the activity of CCO as compared with the DG-treated shamgroup (p > 0.05) (Fig. 7B).
3.6. ATP levels
Fig. 7C shows that the DG-treated mice had a significantdecrease in ATP levels compared with control mice (p < 0.05). Redand NIR laser treatments (8 J/cm2) significantly rescued ATP to
normal levels (p < 0.05), whereas both the lasers at 4 J/cm2 wereleast effective and showed a minimal benefit in reversing the ATPcontent to control level (p > 0.05).
3.7. ROS assessment
A strong production of ROS was observed in DG-treated shammice as compared with the control group (p < 0.01). Data in Fig. 7Dshow that the laser treatment using red and NIR lights at both 4 and8 J/cm2 significantly reduced ROS production compared with theDG-sham group (p < 0.01).
3.8. Western blotting
3.8.1. Bax/Bcl-2 levelsAs shown in Fig. 8A and C, chronic administration of DG increased
Bax levels and decreased Bcl-2 levels, so that the ratio of Bax to Bcl-2was significantly increased compared with the control group(p < 0.01). The remarkable reduction in Bax to Bcl-2 ratio followinglaser treatment using both red and NIR lights (8 J/cm2) was observed(p< 0.01). However, both red and NIR laser irradiation at a fluency of4 J/cm2 was not shown significant effects on attenuating of theDG-induced increase in the ratio of Bax to Bcl-2 (p > 0.05).
3.8.2. Caspase-3 levelsChronic administration of DG significantly increased caspase-3
levels in DG-sham group compared with the control group(p < 0.01). The notable drop in the caspase-3 protein levelsfollowing red and NIR laser treatments (8 J/cm2) was observed(p < 0.01). On the other hand, red and NIR laser treatments at afluency of 4 J/cm2 did not significantly decrease the caspase-3 levelscompared with the DG-treated mice (p > 0.05) (Fig. 8B and C).
Fig. 5. (A) Mean of total observation time and (B) mean of locomotor activity, and (C) mean displacement index (DI) in different groups in test session. Each bar represents the mean# SEM, (n ¼ 12). **p< 0.01 compared with the control. ##p < 0.01 compared with the DG-sham. (D) Heat maps represent the combined traces of the mice in 2 contexts (one in A andone in B) during 2 days of test session, in which novel objects were in right side. Abbreviations: NIR4, near-infrared at a fluency of 4 J/cm2; NIR8, near-infrared at a fluency of 8 J/cm2;R4, red at a fluency of 4 J/cm2; R8, red at a fluency of 8 J/cm2.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150 145

4. Discussion
Since the role of mitochondria as a main target of bioenergeticsand oxidative damage in the neuronal aging process and conse-quent cognitive impairment (Sastre et al., 2002), the present studyfocused on the photoneuromodulation response of mitochondrialfunction, apoptosis, and different learning and memory aspects inDG-induced mice aging model.
Brain bioenergetics impairment, active mitochondria reduction,CCO dysregulation, and ATP decline have been suggested asmechanisms underlying memory and cognitive impairment duringaging (Navarro et al., 2002; Reddy, 2009). Recently, regulation ofneuronal function following LLLT have been shown in cell cultures(Huang et al., 2013), animal models (Lu et al., 2016), and clinicalstudies (Barrett and Gonzalez-Lima, 2013; Blanco et al., 2017). Toinvestigate the neuroenhancement effects of laser in terms ofwavelengths, 2 different wavelengths of 660 nm (red) and 810 nm(NIR) were applied, which matches to the peak absorption spec-trum of CCO (Karu and Kolyakov, 2005).
Results showed chronic administration of DG significantlyreduced the active mitochondria marker. Previous studies in Alz-heimer’s disease (AD) had also shown the loss of brain mitochon-drial number (Hirai et al., 2001; Xie et al., 2013). Respiration andATP production in brain is correlated with the numbers of activemitochondria (Moyes and Battersby, 1998; Perkins and Ellisman,2011) and its diminution could lead to a decline in energy meta-bolism in aged conditions (Stoll et al., 2011). Red and NIR lights (8 J/cm2) increased the abundance of active mitochondria in aged mice.
Higher concentration of active mitochondria in ischemic fibroblastcells irradiated with 660 nm light (5 J/cm2) has been also reported(Houreld et al., 2012).
Mitochondria are the primary site of photon absorption incellular level, and CCO is the key molecule which is responsible forthis photoreaction (Karu and Afanaseva, 1995). Photo-neuromodulation is based on light absorption by the metal centersof CCO, which results in acceleration of mitochondrial electrontransfer and, consequently, ATP production (Karu, 2010).
In this study, administration of DG resulted in deficits in CCOactivity, which was in agreement with previous reports (Prakashand Kumar, 2013; Zhang et al., 2010). On the other hand, red andNIR lasers (8 J/cm2) caused a significant increase in CCO activity.Also, augmentation of CCO activity in neuronal mitochondriafollowing light irradiation has been shown (Lu et al., 2016;Purushothuman et al., 2014; Rojas et al., 2012).
The reduced CCO activity and subsequent low ATP production inthe hippocampus of the normal aged brain were demonstrated(Bertoni-Freddari et al., 2004). Our data showed that red and NIRlights (8 J/cm2) could reverse the ATP levels to normal andcompensate energy deficiency. This is consistent with the previousstudies showing that LLLT increase ATP production in AD (DeTaboada et al., 2011; Lu et al., 2016; Sommer et al., 2012) and TBI(Dong et al., 2015) models. It should be stated that the effectivenessof both the 660 and 810 nm lights in the improvement of CCO ac-tivity and ATP content in this study is in good agreement withWong-Riley et al. study which demonstrated a direct relationshipbetween light irradiation at 670 and 830 nm and CCO absorption
Fig. 6. (A) Mean of MitoTracker Green (MTG) intensity as active mitochondria level index in brain tissue of different groups animals. Each bar represents the mean # SEM, (n ¼ 8).*p< 0.05 compared with the control. #p < 0.05 compared with the DG-sham. (B) MTG staining in cell and correspond isolated mitochondria in different groups. Abbreviations: NIR4,near-infrared at a fluency of 4 J/cm2; NIR8, near-infrared at a fluency of 8 J/cm2; R4, red at a fluency of 4 J/cm2; R8, red at a fluency of 8 J/cm2.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150146

spectrum in primary neurons (Wong-Riley et al., 2005). For normalmemory and cognitive functions, synaptic assembly and activity,ATP supply has a crucial role (Reddy, 2009). Due to the high con-centration of mitochondria in the neuronal tissue (Schwarz, 2013),it seems that mitochondrial CCO could be a potential mediator inphotoneuromodulation of brain bioenergetics and cognitiveimprovement.
Administration of DG decreased brain MMP levels which wereconsistent with the previous in vitro (Shen et al., 2014) and in vivo(Du et al., 2015; Zhu et al., 2014) reports. The age-related oxidativestress is believed to be the main cause of MMP collapse (Hagenet al., 1997) and it causes cell respiratory deficiency and decline ofATP synthesis (Beal et al., 1993). In this study, red and NIR lights(8 J/cm2) increased theMMP levels. This is in linewith other studies
Fig. 7. (A) Mean of JC-1 red-to-green ratio intensity as mitochondrial membrane potential (Djm) index, (B) mean of cytochrome c oxidase activity, (C) mean of ATP content, and (D)mean of DCFDA intensity as reactive oxygen species (ROS) level index in brain tissue of different groups animals. Each bar represents the mean # SEM, (n ¼ 8). *p < 0.05 and **p <
0.01 compared with the control. #p < 0.05 and ##p < 0.01 compared with the DG-sham. Abbreviations: NIR4, near-infrared at a fluency of 4 J/cm2; NIR8, near-infrared at a fluency of8 J/cm2; R4, red at a fluency of 4 J/cm2; R8, red at a fluency of 8 J/cm2.
Fig. 8. (A) Mean ratio of Bax to Bcl-2 and (B) caspase-3 protein levels in the brain tissue of different groups of animals. Each bar represents the mean # SEM, (n ¼ 8). **p < 0.01compared with the control. ##p < 0.01 compared with the DG-sham. (C) Representative images of Bax, Bcl-2, and caspase-3 protein level detected by Western blot. b-actin wasused as an internal control. Abbreviations: NIR4, near-infrared at a fluency of 4 J/cm2; NIR8, near-infrared at a fluency of 8 J/cm2; R4, red at a fluency of 4 J/cm2; R8, red at afluency of 8 J/cm2.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150 147

showing that NIR (w810 nm) rescue MMP decline that is inducedby beta-amyloid (Ab) (Lu et al., 2016), oxidative stresses (Huanget al., 2013), and oxygen-glucose deprivation (Yu et al., 2015a).Absorption of light by mitochondrial inner membrane enzymesthereby enhancing the MMP would be a possible explanation forthis beneficial effect (Gavish et al., 2004).
Mitochondria are pacemakers of cellular aging due to theongoing generation of free radicals such as ROS (Leutner et al.,2001). Increased ROS production affects neurons in part bydamaging mitochondrial components, which ultimately results insenescence (Loeb et al., 2005). Augmentation of mitochondrial ROSproduction following administration of DG has been shown (Haoet al., 2014; Yu et al., 2015a; Zhang et al., 2010). Our data fromROS levels in the brain of DG-treated mice also showed a remark-able increase. In contrast, NIR and red lights suppressed mito-chondrial ROS production. Our findings are in line with previousstudies reporting that NIR laser (810 nm, 3 J/cm2) (Huang et al.,2013) and red LEDs (635 nm, 18 J/cm2) (Lim et al., 2011) reducedbrain ROS levels. Although the biphasic dose response for LLLT-induced regulation of ROS levels has been reported (Huang et al.,2009; Sharma et al., 2011), exact correspond cellular mechanismis unknown. While, reduction in ROS levels may occur viaenhancement of total antioxidant capacity in neuronal cells (Luet al., 2016), studies showed that apoptosis initiate by releasingcytochrome c frommitochondria into the cytoplasmvia a caspase-3activity following a reduction in ATP levels and MMP (Green andKroemer, 2004; Hengartner, 2000). The increased Bax/Bcl-2 ratioalso promotes the activation of caspases and results in cellapoptosis (Qian et al., 2008). In this study, administration of DGincreased Bax/Bcl-2 ratio and caspase-3 level, which is in consistentwith previous findings (Gao et al., 2015; Lu et al., 2010; Ullah et al.,2015; Zhu et al., 2014).
The red and NIR lights at 8 J/cm2, but not 4 J/cm2 notablyregulated Bax, Bcl-2, and caspase-3 activity and rescue neuronsfrom apoptosis. Thus, it suggests that red and NIR LLLT at the op-timum dose of 8 J/cm2 has a potential anti-apoptotic effect. Thepossible reason for this anti-apoptotic effect may be associatedwiththe increasing effect of LLLT on the MMP, which avoid the openingof the mitochondrial permeability, cytochrome c release, andfinally, caspase-3 activation (Xuan et al., 2014). Further, the abilityof LLLT to reduce the ROS production and consequent decrease inapoptotic signaling may be key to realizing the neuroprotectivepotential of photoneuromodulation. The anti-apoptotic effects oflight were also reported in Ab (Duan et al., 2003; Zhang et al., 2012)and other neurotoxins in vitro (Liang et al., 2006; Yu et al., 2015b) ortransient cerebral ischemia (Yip et al., 2011) and TBI (Xuan et al.,2014) in vivo models.
Impaired spatial learning and memory have been reported inanimal DG brain aging model (Kumar et al., 2010; Lu et al., 2010;Wei et al., 2005). In the present study, administration of DGimpaired spatial and episodic-like memories. Michalikova et al.showed improved working memory of normal middle-aged mice ina 3D maze following NIR LED irradiation (Michalikova et al., 2008).The present study showed that red and NIR lights (8 J/cm2) signif-icantly reversed spatial learning andmemory impairment in Barnestest. These are in line with recent report indicating that NIR laser(808 nm, 15 J/cm2) improves performance of AD mice model inBarnes test (Lu et al., 2016). In another study, mitigated spatialmemory deficit in AD mice following NIR laser treatment (808 nm,6 J/cm2) has been shown (De Taboada et al., 2011).We also observedthat red and NIR lights (8 J/cm2) notably improved the episodic-likememory in the DG-treatedmice. This data is also in agreement withthe work of Lu et al., which showed improvement of recognitionmemory of Ab-received mice after laser treatment in novel objectrecognition test (Lu et al., 2016).
Data from mice showed that 2.5% and 3% of light with corre-sponding wavelengths of 670 and 810 nm, respectively, could reacha depth in the brain of 5 mm (Reinhart et al., 2017). Based on this, itcould be assumed that in our studyw0.25 J/cm2 of red or NIR lights(at cortical fluence of 8 J/cm2) reach brainstem which is somewhatin the biostimulatory range for LLLT (Moro et al., 2014; Sharmaet al., 2011). In the human cases, using high-power laser device,2.9% of 810-nm light reach 30-mmdepth of brain tissue (Hendersonand Morries, 2015). This is a clear limitation in the human use oftranscranial LLLT in brain conditions that are associated to deeperparts dysfunction. Given this, to deliver light to the deeper regionsof the brain which are involved in AD and PD pathologies, mostrecently there have been attempts to develop some new ap-proaches such as the combination of transcranial and intranasalLLLT (Saltmarche et al., 2017) as well as intracranial LLLT method(Darlot et al., 2016).
It could conclude that transcranial LLLT using red and NIR lights(8 J/cm2) rescued mice brain fromDG-induced spatial and episodic-likememory impairment probably via modulation of mitochondrialfunction and bioenergetics as well as ROS production and neuronalapoptosis. In addition, we applied fluency of 4 J/cm2, but this maynot be the sufficient therapeutic regimen, in terms of fluency.Moreover, the NIR laser showed slightly, but not significantly betterresults than red and effects did not appear to be dependent on laserwavelength. This piece of result suggests that photo-neuromodulation of brain could be a promising candidate forneuroenhancement of age-related cognitive impairments. Never-theless, the optimum treatment parameters of transcranial LLLTwith the best efficacy and safety should be determined in futurestudies.
Disclosure statement
The authors have no conflicts of interest to disclose.
Acknowledgements
This research was supported in part by Neurosciences ResearchCenter (NSRC), TUOMS (grant number: 5/88/1587).
References
Barrett, D., Gonzalez-Lima, F., 2013. Transcranial infrared laser stimulation producesbeneficial cognitive and emotional effects in humans. Neuroscience 230, 13e23.
Bashkatov, A.N., Genina, E.A., Kochubey, V.I., Tuchin, V.V., 2006. Optical properties ofhuman cranial bone in the spectral range from 800 to 2000 nm. Opt. Technol.Biophys. Med. VII. Int. Soc. Opt. Photon. 1e11.
Beal, M.F., Hyman, B.T., Koroshetz, W., 1993. Do defecs in mitochondrial energymetabolism underlie the pathology of neurodegenerative diseases? TrendsNeurosci. 16, 125e131.
Bertoni-Freddari, C., Patrizia, F., Giorgetti, B., Solazzi, M., Marta, B., Casoli, T.,Stefano, G., 2004. Cytochrome oxidase activity in hippocampal synaptic mito-chondria during aging: a quantitative cytochemical investigation. Ann. N. Y.Acad. Sci. 1019, 33e36.
Bishop, N.A., Lu, T., Yankner, B.A., 2010. Neural mechanisms of ageing and cognitivedecline. Nature 464, 529e535.
Blanco, N.J., Maddox, W.T., Gonzalez-Lima, F., 2017. Improving executive functionusing transcranial infrared laser stimulation. J. Neuropsychol. 11, 14e25.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem. 72, 248e254.
Chung, H., Dai, T., Sharma, S.K., Huang, Y.-Y., Carroll, J.D., Hamblin, M.R., 2012. Thenuts and bolts of low-level laser (light) therapy. Ann. Biomed. Eng. 40, 516e533.
Cui, X., Zuo, P., Zhang, Q., Li, X., Hu, Y., Long, J., Packer, L., Liu, J., 2006. Chronicsystemic D-galactose exposure induces memory loss, neurodegeneration, andoxidative damage in mice: protective effects of R-a-lipoic acid. J. Neurosci. Res.83, 1584e1590.
Darlot, F., Moro, C., Massri, N., Chabrol, C., Johnstone, D.M., Reinhart, F., Agay, D.,Torres, N., Bekha, D., Auboiroux, V., Costecalde, T., 2016. Near-infrared light isneuroprotective in a monkey model of Parkinson disease. Ann. Neuro 79,59e75.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150148

Davis, K.E., Easton, A., Eacott, M.J., Gigg, J., 2013. Episodic-like memory for what-where-which occasion is selectively impaired in the 3xTgAD mouse model ofAlzheimer’s disease. J. Alzheimers. Dis. 33, 681e698.
de Freitas, L.F., Hamblin, M.R., 2016. Proposed mechanisms of photobiomodulationor low-level light therapy. IEEE. J. Sel. Top. Quan. Electron 22, 1e17.
De Taboada, L., Yu, J., El-Amouri, S., Gattoni-Celli, S., Richieri, S., McCarthy, T.,Streeter, J., Kindy, M.S., 2011. Transcranial laser therapy attenuates amyloid-bpeptide neuropathology in amyloid-b protein precursor transgenic mice.J. Alzheimers. Dis. 23, 521e535.
Dong, T., Zhang, Q., Hamblin, M.R., Wu, M.X., 2015. Low-level light in combinationwith metabolic modulators for effective therapy of injured brain. J. Cereb. BloodFlow. Metab. 35, 1435e1444.
Du, Z., Yang, Q., Liu, L., Li, S., Zhao, J., Hu, J., Liu, C., Qian, D., Gao, C., 2015. NADPHoxidase 2-dependent oxidative stress, mitochondrial damage and apoptosis inthe ventral cochlear nucleus of d-galactose-induced aging rats. Neuroscience286, 281e292.
Duan, R., Zhu, L., Liu, T.C.Y., Li, Y., Liu, J., Jiao, J., Xu, X., Yao, L., Liu, S., 2003. Lightemitting diode irradiation protect against the amyloid beta 25e35 inducedapoptosis of PC12 cell in vitro. Lasers. Surg. Med. 33, 199e203.
Firbank, M., Hiraoka, M., Essenpreis, M., Delpy, D., 1993. Measurement of the opticalproperties of the skull in the wavelength range 650-950 nm. Phys. Med. Biol. 38,503e510.
Gao, J., He, H., Jiang, W., Chang, X., Zhu, L., Luo, F., Zhou, R., Ma, C., Yan, T., 2015.Salidroside ameliorates cognitive impairment in a d-galactose-induced ratmodel of Alzheimer’s disease. Behav. Brain Res. 293, 27e33.
Gavish, L., Asher, Y., Becker, Y., Kleinman, Y., 2004. Low level laser irradiationstimulates mitochondrial membrane potential and disperses subnuclear pro-myelocytic leukemia protein. Lasers. Surg. Med. 35, 369e376.
Gonzalez-Lima, F., Barksdale, B.R., Rojas, J.C., 2014. Mitochondrial respiration as atarget for neuroprotection and cognitive enhancement. Biochem. Pharmacol.88, 584e593.
Grady, C., 2012. The cognitive neuroscience of ageing. Nat. Rev. Neurosci. 13,491e505.
Green, D.R., Kroemer, G., 2004. The pathophysiology of mitochondrial cell death.Science 305, 626e629.
Grillo, S., Duggett, N., Ennaceur, A., Chazot, P., 2013. Non-invasive infra-red therapy(1072nm) reduces b-amyloid protein levels in the brain of an Alzheimer’sdisease mouse model, TASTPM. J. Photochem. Photobiol. B 123, 13e22.
Hagen, T.M., Yowe, D.L., Bartholomew, J.C., Wehr, C.M., Do, K.L., Park, J.-Y., Ames, B.N.,1997. Mitochondrial decay in hepatocytes from old rats: membrane potentialdeclines, heterogeneity and oxidants increase. Proc. Natl. Acad. Sci. 94,3064e3069.
Hao, L., Huang, H., Gao, J., Marshall, C., Chen, Y., Xiao, M., 2014. The influence ofgender, age and treatment time on brain oxidative stress and memoryimpairment induced by d-galactose in mice. Neurosci. Lett. 571, 45e49.
Henderson, T.A., Morries, L.D., 2015. Near-infrared photonic energy penetration: caninfrared phototherapy effectively reach the human brain. Neuropsychiatr. Dis.Treat 11, 2191e2208.
Hengartner, M.O., 2000. The biochemistry of apoptosis. Nature 407, 770e776.Hirai, K., Aliev, G., Nunomura, A., Fujioka, H., Russell, R.L., Atwood, C.S., Johnson, A.B.,
Kress, Y., Vinters, H.V., Tabaton, M., 2001. Mitochondrial abnormalities in Alz-heimer’s disease. J. Neurosci. Nurs. 21, 3017e3023.
Ho, S.-C., Liu, J.-H., Wu, R.-Y., 2003. Establishment of the mimetic aging effect inmice caused by D-galactose. Biogerontology 4, 15e18.
Houreld, N.N., Masha, R.T., Abrahamse, H., 2012. Low-intensity laser irradiation at660 nm stimulates cytochrome c oxidase in stressed fibroblast cells. Lasers.Surg. Med. 44, 429e434.
Huang, Y.-Y., Chen, A.C.-H., Carroll, J.D., Hamblin, M.R., 2009. Biphasic dose responsein low level light therapy. Dose Response 7, 358e383.
Huang, Y.Y., Nagata, K., Tedford, C.E., McCarthy, T., Hamblin, M.R., 2013. Low-levellaser therapy (LLLT) reduces oxidative stress in primary cortical neurons in vitro.J. Biophotonics 6, 829e838.
Jang, Y.C., Van Remmen, H., 2009. The mitochondrial theory of aging: insight fromtransgenic and knockout mouse models. Exp. Gerontol. 44, 256e260.
Karu, T., 2010. Mitochondrial mechanisms of photobiomodulation in context of newdata about multiple roles of ATP. Photomed. Laser Surg. Photomed. Laser Surg. 2,159e160.
Karu, T., Afanaseva, N., 1995. Cytochrome c oxidase acts as a primary photoacceptorin cell cultures subjected to visible and near IR laser irradiation. Dokl. Biochem.84e86.
Karu, T., Kolyakov, S., 2005. Exact action spectra for cellular responses relevant tophototherapy. Photomed. Laser Surg. 23, 355e361.
Karu, T.I., 2014. Cellular and molecular mechanisms of photobiomodulation (low-power laser therapy). IEEE. J. Sel. Top. Quan. Electron 20, 143e148.
Kumar, A., Prakash, A., Dogra, S., 2010. Naringin alleviates cognitive impairment,mitochondrial dysfunction and oxidative stress induced by D-galactose in mice.Food Chem. Toxicol. 48, 626e632.
Lan, Z., Liu, J., Chen, L., Fu, Q., Luo, J., Qu, R., Kong, L., Ma, S., 2012. Danggui-Shaoyao-San ameliorates cognition deficits and attenuates oxidative stress-relatedneuronal apoptosis in d-galactose-induced senescent mice. J. Ethnopharmacol141, 386e395.
Lapchak, P.A., 2012. Transcranial near-infrared laser therapy applied to promoteclinical recovery in acute and chronic neurodegenerative diseases. Expert Rev.Med. Devices 9, 71e83.
Lei, M., Su, Y., Hua, X., Ding, J., Han, Q., Hu, G., Xiao, M., 2008. Chronic systemicinjection of D-galactose impairs the septohippocampal cholinergic system inrats. Neuroreport 19, 1611e1615.
Lemberg, M., 1969. Cytochrome oxidase. Physiol. Rev. 49, 48e121.Leutner, S., Eckert, A., Müller, W., 2001. ROS generation, lipid peroxidation and
antioxidant enzyme activities in the aging brain. J. Neural Transm. 108,955e967.
Liang, H., Whelan, H., Eells, J., Meng, H., Buchmann, E., Lerch-Gaggl, A., Wong-Riley, M., 2006. Photobiomodulation partially rescues visual cortical neuronsfrom cyanide-induced apoptosis. Neuroscience 139, 639e649.
Lim, W.B., Kim, J.S., Ko, Y.J., Kwon, H., Kim, S.W., Min, H.K., Kim, O., Choi, H.R.,Kim, O.J., 2011. Effects of 635nm light-emitting diode irradiation on angiogen-esis in CoCl2-exposed HUVECs. Lasers. Surg. Med. 43, 344e352.
Liu, J., Ames, B.N., 2005. Reducing mitochondrial decay with mitochondrial nutri-ents to delay and treat cognitive dysfunction, Alzheimer’s disease, and Parkin-son’s disease. Nutr. Neurosci. 8, 67e89.
Liu, J., Atamna, H., Kuratsune, H., Ames, B.N., 2002. Delaying brain mitochondrialdecay and aging with mitochondrial antioxidants and metabolites. Ann. N. Y.Acad. Sci. 959, 133e166.
Loeb, L.A., Wallace, D.C., Martin, G.M., 2005. The mitochondrial theory of aging andits relationship to reactive oxygen species damage and somatic mtDNA muta-tions. Proc. Natl. Acad. Sci. U. S. A. 102, 18769e18770.
Lu, J., Wu, D.M., Zheng, Y.L., Hu, B., Zhang, Z.F., 2010. Purple Sweet Potato color al-leviates d-galactose-induced brain aging in old mice by promoting survival ofneurons via PI3K pathway and inhibiting cytochrome C-mediated apoptosis.Brain Pathol. 20, 598e612.
Lu, Y., Wang, R., Dong, Y., Tucker, D., Zhao, N., Ahmed, M.E., Zhu, L., Liu, T.C.-Y.,Cohen, R.M., Zhang, Q., 2016. Low-level laser therapy for beta-amyloid toxicityin rat hippocampus. Neurobiol. Aging 49, 165e182.
Michalikova, S., Ennaceur, A., van Rensburg, R., Chazot, P., 2008. Emotional re-sponses and memory performance of middle-aged CD1 mice in a 3D maze:effects of low infrared light. Neurobiol. Learn. Mem. 89, 480e488.
Mochizuki-Oda, N., Kataoka, Y., Cui, Y., Yamada, H., Heya, M., Awazu, K., 2002. Effectsof near-infra-red laser irradiation on adenosine triphosphate and adenosinediphosphate contents of rat brain tissue. Neurosci. Lett. 323, 207e210.
Moro, C., Massri, N.E., Torres, N., Ratel, D., De Jaeger, X., Chabrol, C., Perraut, F.,Bourgerette, A., Berger, M., Purushothuman, S., Johnstone, D., 2014. Photo-biomodulation inside the brain: a novel method of applying near-infrared lightintracranially and its impact on dopaminergic cell survival in MPTP-treatedmice: Laboratory investigation. J. Neurosurg. 120, 670e683.
Moyes, C., Battersby, B., 1998. Regulation of muscle mitochondrial design. J. Exp.Biol. 201, 299e307.
Naeser, M.A., Saltmarche, A., Krengel, M.H., Hamblin, M.R., Knight, J.A., 2011.Improved cognitive function after transcranial, light-emitting diode treatmentsin chronic, traumatic brain injury: two case reports. Photomed. Laser Surg. 29,351e358.
Navarro, A., Del Pino, M.J.S., Gómez, C., Peralta, J.L., Boveris, A., 2002. Behavioraldysfunction, brain oxidative stress, and impaired mitochondrial electrontransfer in aging mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 282,R985eR992.
Nawashiro, H., Wada, K., Nakai, K., Sato, S., 2012. Focal increase in cerebral bloodflow after treatment with near-infrared light to the forehead in a patient in apersistent vegetative state. Photomed. Laser Surg. 30, 231e233.
Niu, Y.-L., Zhang, W.-J., Wu, P., Liu, B., Sun, G.-T., Yu, D.-M., Deng, J.-B., 2010.Expression of the apoptosis-related proteins caspase-3 and NF-kB in the hip-pocampus of Tg2576 mice. Neurosci. Bull 26, 37e46.
Novalija, E., Kevin, L.G., Eells, J.T., Henry, M.M., Stowe, D.F., 2003. Anesthetic pre-conditioning improves adenosine triphosphate synthesis and reduces reactiveoxygen species formation in mitochondria after ischemia by a redox dependentmechanism. J. Am. Soc. Anesthesiol 98, 1155e1163.
Oron, U., Ilic, S., De Taboada, L., Streeter, J., 2007. Ga-As (808 nm) laser irradiationenhances ATP production in human neuronal cells in culture. Photomed. LaserSurg. 25, 180e182.
Perkins, G.A., Ellisman, M.H., 2011. Mitochondrial configurations in peripheral nervesuggest differential ATP production. J. Struct. Biol. 173, 117e127.
Pollack, M., Phaneuf, S., Dirks, A., Leeuwenburgh, C., 2002. The role of apoptosis inthe normal aging brain, skeletal muscle, and heart. Ann. N. Y. Acad. Sci. 959,93e107.
Pourmemar, E., Majdi, A., Haramshahi, M., Talebi, M., Karimi, P., Sadigh-Eteghad, S.,2017. Intranasal cerebrolysin attenuates learning and memory impairments ind-galactose-induced senescence in mice. Exp. Gerontol. 87, 16e22.
Prakash, A., Kumar, A., 2013. Pioglitazone alleviates the mitochondrial apoptoticpathway and mito-oxidative damage in the d-galactose-induced mouse model.Clin. Exp. Pharmacol. Physiol. 40, 644e651.
Purushothuman, S., Johnstone, D.M., Nandasena, C., Mitrofanis, J., Stone, J., 2014.Photobiomodulation with near infrared light mitigates Alzheimer’s disease-related pathology in cerebral cortexeevidence from two transgenic mousemodels. Alzheimers Res. Ther. 6, 2.
Qian, Y.-f., Wang, H., Yao, W.-b., Gao, X.-d., 2008. Aqueous extract of the Chinesemedicine, Danggui-Shaoyao-San, inhibits apoptosis in hydrogen peroxide-induced PC12 cells by preventing cytochrome c release and inactivating ofcaspase cascade. Cell. Biol. Int. 32, 304e311.
Reddy, P.H., 2009. Role of mitochondria in neurodegenerative diseases: mitochon-dria as a therapeutic target in Alzheimer’s disease. CNS. Spectr. 14, 8e13.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150 149

Reinhart, F., El Massri, N., Torres, N., Chabrol, C., Molet, J., Johnstone, D.M., Stone, J.,Benabid, A.L., Mitrofanis, J., Moro, C., 2017. The behavioural and neuroprotectiveoutcomes when 670nm and 810nm near infrared light are applied together inMPTP-treated mice. Neurosci. Res. 117, 42e47.
Rojas, J.C., Bruchey, A.K., Gonzalez-Lima, F., 2012. Low-level light therapy improvescortical metabolic capacity andmemory retention. J. Alzheimers. Dis. 32, 741e752.
Sadigh-Eteghad, S., Talebi, M., Mahmoudi, J., Babri, S., Shanehbandi, D., 2015. Se-lective activation of a 7 nicotinic acetylcholine receptor by PHA-543613 im-proves Ab 25e35-mediated cognitive deficits in mice. Neuroscience 298, 81e93.
Salehpour, F., Rasta, S.H., 2017. The potential of transcranial photobiomodulationtherapy for treatment of major depressive disorder. Rev. Neurosci. 28, 441e453.
Salehpour, F., Rasta, S.H., Mohaddes, G., Sadigh-Eteghad, S., Salarirad, S., 2016. Ther-apeutic effects of 10-HzPulsed wave lasers in rat depression model: a comparisonbetween near-infrared and red wavelengths. Lasers. Surg. Med. 48, 695e705.
Salgado, A.S., Zângaro, R.A., Parreira, R.B., Kerppers, I.I., 2015. The effects of trans-cranial LED therapy (TCLT) on cerebral blood flow in the elderly women. Lasers.Med. Sci. 30, 339e346.
Saltmarche, A.E., Naeser, M.A., Ho, K.F., Hamblin, M.R., Lim, L., 2017. Significantimprovement in cognition in mild to moderately severe dementia cases treatedwith transcranial plus intranasal photobiomodulation: case series report.Photomed. Laser Surg. http://dx.doi.org/10.1089/pho.2016.4227 [Epub aheadof print].
Sastre, J., Borrás, C., GARCÍA-SALA, D., Lloret, A., Pallardó, F.V., Viña, J., 2002. Mito-chondrial damage in aging and apoptosis. Ann. N. Y. Acad. Sci. 959, 448e451.
Schiffer, F., Johnston, A.L., Ravichandran, C., Polcari, A., Teicher, M.H., Webb, R.H.,Hamblin, M.R., 2009. Psychological benefits 2 and 4 weeks after a singletreatment with near infrared light to the forehead: a pilot study of 10 patientswith major depression and anxiety. Behav. Brain Funct. 5, 46.
Schwarz, T.L., 2013. Mitochondrial trafficking in neurons. Cold. Spring Harb. Per-spect. Biol. 5, a011304.
Sharma, S.K., Kharkwal, G.B., Sajo, M., Huang, Y.Y., De Taboada, L., McCarthy, T.,Hamblin, M.R., 2011. Dose response effects of 810 nm laser light on mouseprimary cortical neurons. Lasers. Surg. Med. 43, 851e859.
Shen, Y., Gao, H., Shi, X., Wang, N., Ai, D., Li, J., Ouyang, L., Yang, J., Tian, Y., Lu, J., 2014.Glutamine synthetase plays a role in d-galactose-induced astrocyte agingin vitro and in vivo. Exp. Gerontol. 58, 166e173.
Sommer, A.P., Bieschke, J., Friedrich, R.P., Zhu, D., Wanker, E.E., Fecht, H.J.,Mereles, D., Hunstein, W., 2012. 670 nm laser light and EGCG complementarilyreduce amyloid-b aggregates in human neuroblastoma cells: basis for treatmentof alzheimer’s disease? Photomed. Laser Surg. 30, 54e60.
Stoll, E.A., Cheung, W., Mikheev, A.M., Sweet, I.R., Bielas, J.H., Zhang, J.,Rostomily, R.C., Horner, P.J., 2011. Aging neural progenitor cells have decreasedmitochondrial content and lower oxidative metabolism. J. Biol. Chem. 286,38592e38601.
Sunyer, B., Patil, S., Höger, H., Lubec, G., 2007. Barnes maze, a useful task to assessspatial reference memory in the mice. Nat. Protoc. 390, 10e38.
Tsai, S.-J., Yin, M.-C., 2012. Anti-oxidative, anti-glycative and anti-apoptotic effects ofoleanolic acid in brain of mice treated by D-galactose. Eur. J. Pharmacol. 689,81e88.
Ullah, F., Ali, T., Ullah, N., Kim, M.O., 2015. Caffeine prevents d-galactose-inducedcognitive deficits, oxidative stress, neuroinflammation and neurodegenerationin the adult rat brain. Neurochem. Int. 90, 114e124.
Uozumi, Y., Nawashiro, H., Sato, S., Kawauchi, S., Shima, K., Kikuchi, M., 2010. Tar-geted increase in cerebral blood flow by transcranial near-infrared laser irra-diation. Lasers. Surg. Med. 42, 566e576.
Wei, H., Li, L., Song, Q., Ai, H., Chu, J., Li, W., 2005. Behavioural study of theD-galactose induced aging model in C57BL/6J mice. Behav. Brain Res. 157,245e251.
Wong-Riley, M.T., Liang, H.L., Eells, J.T., Chance, B., Henry, M.M., Buchmann, E.,Kane, M., Whelan, H.T., 2005. Photobiomodulation directly benefits primaryneurons functionally inactivated by toxins role of cytochrome c oxidase. J. Biol.Chem. 280, 4761e4771.
Xie, H., Guan, J., Borrelli, L.A., Xu, J., Serrano-Pozo, A., Bacskai, B.J., 2013. Mito-chondrial alterations near amyloid plaques in an Alzheimer’s disease mousemodel. J. Neurosci. Nurs. 33, 17042e17051.
Xuan, W., Vatansever, F., Huang, L., Hamblin, M.R., 2014. Transcranial low-level lasertherapy enhances learning, memory, and neuroprogenitor cells after traumaticbrain injury in mice. J. Biomed. Opt. 19, 108003.
Yip, K., Lo, S., Leung, M., So, K., Tang, C., Poon, D., 2011. The effect of low-energy laserirradiation on apoptotic factors following experimentally induced transientcerebral ischemia. Neuroscience 190, 301e306.
Yu, Y., Bai, F., Wang, W., Liu, Y., Yuan, Q., Qu, S., Zhang, T., Tian, G., Li, S., Li, D.,2015a. Fibroblast growth factor 21 protects mouse brain against d-galactoseinduced aging via suppression of oxidative stress response and advancedglycation end products formation. Pharmacol. Biochem. Behav. 133,122e131.
Yu, Z., Li, Z., Liu, N., Jizhang, Y., McCarthy, T.J., Tedford, C.E., Lo, E.H., Wang, X., 2015b.Near infrared radiation protects against oxygen-glucose deprivation-inducedneurotoxicity by down-regulating neuronal nitric oxide synthase (nNOS)activity in vitro. Metab. Brain Dis. 30, 829e837.
Zhang, H., Wu, S., Xing, D., 2012. Inhibition of Ab 25e35-induced cell apoptosis bylow-power-laser-irradiation (LPLI) through promoting Akt-dependent YAPcytoplasmic translocation. Cell. Signal 24, 224e232.
Zhang, X., Liu, W., Niu, X., An, L., 2010. Systemic administration of catalpol preventsd-galactose induced mitochondrial dysfunction in mice. Neurosci. Lett. 473,224e228.
Zhang, X.-L., An, L.-J., Bao, Y.-M., Wang, J.-Y., Jiang, B., 2008. D-galactose adminis-tration induces memory loss and energy metabolism disturbance in mice:protective effects of catalpol. Food Chem. Toxicol. 46, 2888e2894.
Zhu, S.Y., Dong, Y., Tu, J., Zhou, Y., Zhou, X.H., Xu, B., 2014. Silybum marianum oilattenuates oxidative stress and ameliorates mitochondrial dysfunction in micetreated with D-galactose. Pharmacogn. Mag. 10 (Suppl 1), S92.
F. Salehpour et al. / Neurobiology of Aging 58 (2017) 140e150150