Tenorio et al., Human Mutation - imprinting disorders€¦ · Tenorio et al., Human Mutation 2...
19
Tenorio et al., Human Mutation 1 SUPP. MATERIALS AND METHODS Karyotypes, arrayCGH, SNP-arrays and custom designed MLPA Karyotypes were carried out following routine techniques. Array-CGH was performed using the KaryoArray™ v3.0 customized array (60,000 genome-wide oligonucleotide array (Vallespin, et al., 2013). The array has an average probe density of 43 kb (Vallespin, et al., 2013). Array experiments were performed as recommended by the manufacturer (Agilent Technologies, Santa Clara, CA, USA) and then scanned by the Agilent Microarray Scanner and analyzed with the Feature Extraction software (v9.1 Agilent Technologies) and Genomic Work Bench 5.0. For SNP-arrays we applied the Illumina Human610-Quad BeadChip according to the manufacturer's specifications (Illumina, San Diego, CA, USA) and analyzed the results using the Chromosome Viewer tool contained in GenomeStudio (Illumina). GenCall scores <0.15 at any locus were considered ‘no calls’. The log R ratio was employed, which is the log (base 2) ratio of the observed normalized R value for a SNP divided by the expected normalized R value. Following the observation of a RNF125 deletion, we custom designed a multiplex ligation probe amplification (MLPA) assay to screen the remaining patients for deletions or duplications within RNF125. Basically 4 probes localized in the gene and three control probes were designed. The protocol has been previously described for other genes. Subsequently the ratios of the probands’ peak areas vs. controls’ samples were determined using an Excel data sheet (Microsoft Corp., Richmond, CA). Normal peaks were classified as showing a ratio of 0.65–1.35, whereas deletions and duplications were classified as having a ratio less than 0.65 or greater than 1.35, respectively. High Resolution Melting Analysis and Sanger Sequencing After the finding of a microdeletion at 18q encompassing two RNF genes (RNF125 and RNF138) in patient 1, 270 single individuals with no known overgrowth were analyzed for the presence of mutations in these two genes and also in RNF135, a gene already linked to overgrowth and macrocephaly at chromosome 17 (Douglas, et al., 2007). After PCR amplification of all exons (primers in Supp. Table S4) of RNF125, RNF135 and RNF138 we applied high resolution melting (HRM) analysis to detect DNA variants. Sanger sequencing was performed for all abnormal fragments cosegregation analysis of the variants with the phenotype were undertaken where possible. PCR products were purified with Exostar (GE Healthcare, UK) and sequenced using BigDye terminator 3.1 (Life Technologies, USA). Sequences were analyzed using Sequencher V5.0® software (Gene Codes, USA). The frequency of each variant was determined in a total of 300 healthy Spanish controls (600 chromosomes).
Transcript of Tenorio et al., Human Mutation - imprinting disorders€¦ · Tenorio et al., Human Mutation 2...

Tenorio et al., Human Mutation 1
SUPP. MATERIALS AND METHODS
Karyotypes, arrayCGH, SNP-arrays and custom designed MLPA
Karyotypes were carried out following routine techniques. Array-CGH was performed using
the KaryoArray™ v3.0 customized array (60,000 genome-wide oligonucleotide array (Vallespin, et al.,
2013). The array has an average probe density of 43 kb (Vallespin, et al., 2013). Array experiments
were performed as recommended by the manufacturer (Agilent Technologies, Santa Clara, CA, USA)
and then scanned by the Agilent Microarray Scanner and analyzed with the Feature Extraction
software (v9.1 Agilent Technologies) and Genomic Work Bench 5.0. For SNP-arrays we applied the
Illumina Human610-Quad BeadChip according to the manufacturer's specifications (Illumina, San
Diego, CA, USA) and analyzed the results using the Chromosome Viewer tool contained in
GenomeStudio (Illumina). GenCall scores <0.15 at any locus were considered ‘no calls’. The log R
ratio was employed, which is the log (base 2) ratio of the observed normalized R value for a SNP
divided by the expected normalized R value.
Following the observation of a RNF125 deletion, we custom designed a multiplex ligation
probe amplification (MLPA) assay to screen the remaining patients for deletions or duplications
within RNF125. Basically 4 probes localized in the gene and three control probes were designed. The
protocol has been previously described for other genes. Subsequently the ratios of the probands’
peak areas vs. controls’ samples were determined using an Excel data sheet (Microsoft Corp.,
Richmond, CA). Normal peaks were classified as showing a ratio of 0.65–1.35, whereas deletions and
duplications were classified as having a ratio less than 0.65 or greater than 1.35, respectively.
High Resolution Melting Analysis and Sanger Sequencing
After the finding of a microdeletion at 18q encompassing two RNF genes (RNF125 and RNF138) in
patient 1, 270 single individuals with no known overgrowth were analyzed for the presence of
mutations in these two genes and also in RNF135, a gene already linked to overgrowth and
macrocephaly at chromosome 17 (Douglas, et al., 2007). After PCR amplification of all exons
(primers in Supp. Table S4) of RNF125, RNF135 and RNF138 we applied high resolution melting
(HRM) analysis to detect DNA variants. Sanger sequencing was performed for all abnormal
fragments cosegregation analysis of the variants with the phenotype were undertaken where
possible. PCR products were purified with Exostar (GE Healthcare, UK) and sequenced using BigDye
terminator 3.1 (Life Technologies, USA). Sequences were analyzed using Sequencher V5.0® software
(Gene Codes, USA). The frequency of each variant was determined in a total of 300 healthy Spanish
controls (600 chromosomes).

Tenorio et al., Human Mutation 2
Pyrosequencing confirmation
We applied pyrosequencing in selected samples to confirm dosage of each allele and to rule out any
degree of mosaicism. The pyrosequencing assays were designed to analyze the sequence just a few
nucleotides before the mutation sites of RNF125. Primers used for PCR amplification and
pyrosequencing are listed in Supp. Table S4. Each PCR reaction contained forward and reverse
primers (final concentration of 0.3 μM), 0.125 mmol/L of each dNTP, 1.5 mmol/L MgCl2, 1× PCR
buffer, 1.5 U of Taq polymerase (Roche, catalog no. 11435094001) and 30ng of template DNA in a
total volume of 30μl. PCR conditions consisted of initial denaturing at 95ºC for 5 minutes; 45 cycles
of 95ºC for 30 seconds, 60ºC for 30 seconds, and 72ºC for 30 seconds; and a final extension at 72ºC
for 10 minutes. The PCR products were separated by agarose gel electrophoresis in 1.5% of agarose
to confirm successful amplification. The PCR products (20 μl) were pyrosequenced on a PSQ96 HS
system (Biotage AB, Uppsala, Sweden) following the manufacturer's instructions (Supp. Table S4).
Negative and blank controls were included in each run. A sequential nucleotide dispensation
protocol was used reflecting the expected order of nucleotide incorporation and the potential base
changes. In addition, negative nucleotide dispensations were inserted to serve as internal controls
for nucleotide misincorporation. Peak heights are proportional to the number of nucleotides that are
incorporated with each dispensation.
Whole exome sequencing
To exclude the presence of other pathogenic mutations causes of the overgrowth observed in
patients 2 and 5, we performed whole exome sequencing (WES) in these individuals. Samples were
prepared and captured with the Agilent SureSelect Human All Exon 50 Mb kit. The adapter-ligated
sample was purified using Agencourt AMPure XP beads (Beckman Coulter, Brea, CA, USA) and
analyzed on a Bioanalyzer High Sensitivity DNA chip. Sequencing was performed in Illumina GA II
equipment. Data analysis was performed as previously described (Medina, et al., 2012).
Whole-genome methylation arrays
We generated methylation datasets using the Illumina Infinium 450K BeadChip arrays which
simultaneously quantifies ~2% of all CpG dinucleotides. Bisulphite conversion of 600 ng of DNA was
performed according to the manufacturer’s recommendations for the Illumina Infinium Assay (EZ
DNA methylation kit, ZYMO, Orange, CA). Subsequently, the bisulphite-converted DNA was used for
hybridization. The resulting data for the three genome-wide DNA from leukocytes of patients 1, 2
and 5 have been deposited in the GEO database with the accession number GSE54335.

Tenorio et al., Human Mutation 3
Whole genome expression arrays
Micro-array expression analysis was performed using RNA samples of patients 1, 2 and 5 and RNA of
controls (age and sex-matched). RNA was extracted from peripheral blood following the
manufacturer’s procedure using TRIReagent Solution (Applied Biosystems, USA). RNA quality
assessment was measured by both spectrophotometry (NanoDrop Spectrophotometer) and with
Agilent 2100 bioanalyzer and Nano6000. For expression analysis we used SurePrint G3 Human GE
8x60K one-color Microarray Kit (Agilent, USA). Data arrays outputs were analyzed using Gene Spring
software (Agilent, USA). Student T-test (with 0.05 significance) was performed as a statistic analysis
to discriminate genes with differentially mRNA levels. Data of the expression arrays are deposited in
GEO (Gene Expression Omnibus) with the accession number GSE53756.
Quantitative RT-PCR analysis
We conducted quantitative PCR confirmation for a selected number of genes obtained from the
expression array studies (included in the RNF125-RNF135-RIG-I pathway). To confirm the expression
array results, single RT-qPCR were performed for RNF125, RIG1, MDA5, LGP2, ISG15 and IPS1.
Primers for the qPCR were obtained from Toyoda et al. (38). As a control, we employed β-actin using
the following primers: forward 5´GAGCGGGAAATCGTGCGTGACATT3´ and reverse
5´GAAGGTAGTTTCGTGGATGCC3´. The High Capacity cDNA Reverse Transcription Kit (Applied
Biosystems, USA) were used to perform the reverse transcription from the RNA samples. The
reaction mixture was incubated at 25ºC for 10 minutes and 120 minutes at 37ºC. cDNA products
were treated with RNAase (Qiagen, USA) for 15 minutes to eliminate any RNA residues. Then, qPCR
was performed with SYBR Premix Ex Taq (perfect real time) (Takara, Japan) following the
manufacturer’s recommendations. Age and sex-matched cDNA control were also used as a calibrator
to calculate the 2-ΔΔCT ratio. Standard deviations were calculated using the mean of three measures
of each sample from three biological replicates.
In order to validate the whole genome microarray results of the PI3K-AKT pathway, a
commercial qPCR plate of the PI3K-AKT pathway real-time RT-PCR arrays containing 84 genes
(Qiagen, USA) was used to analyze transcript expression level differences between patients and
controls. Total RNA was isolated as described previously. cDNA synthesis and real-time PCR were
conducted using the manufacturer’s recommended reagents and protocols for the ABi 7300
thermocycler instrument. Results were normalized to the average Ct for five house-keeping genes
contained on the RT-PCR array (ACTB, B2M, GAPDH, HPRT1 and RPLP0) and ΔΔCt values were

Tenorio et al., Human Mutation 4
calculated to determine expression changes. Genes that reached the Ct maximum of 35 in either
trial were excluded from the analysis.
In silico analysis of RNF125 variants
We computed the predicted effects of RNF125 non-synonymous variants on the protein function
using several bioinformatics tools. We applied SIFT (Ng and Henikoff, 2001), Mutation
Taster(Schwarz, et al., 2014), MutPred(Li, et al., 2009) and Polyphen-2 (Adzhubei, et al., 2010) to
evaluate the putative pathogenicity of the identified variants. We also searched the Exome Variant
Server (EVS) and the 1000 genomes to check the presence of these variants in these databases.
Additionally, to assess the potential functional consequences of the substitutions, a 3D structural
analysis of RNF125 was modeled by using the PHYRE2 server (Kelley and Sternberg, 2009).
Cell cultures and Western blotting
Lymphoblast cells were extracted from peripheral blood from patient 1, 2, 5, and 6. Then, these cells
were immortalized by using Epstein-Barr virus transformation (at the CIBERER Biobank, Valencia,
Spain). Control lymphoblast cells were obtained from Coriell Institute for Medical Research (New
Jersey, USA) (Catalog ID: GM14907; GM17095) and from the CIBERER Biobank from sex and age-
matched healthy individuals. Cells were growing in RPMI1640 medium; 2mM L-glutamine; 15% fetal
bovine serum; 1% Fungizone; 1% Penicillin/Streptomycin. When indicated, Poly (I:C) (Sigma-Aldrich,
USA) at 100 µg/ml final concentration was used for 16 hours. Poly (I:C) is a synthetic analog of
double-stranded RNA that activates TLR3 receptor40. For protein synthesis inhibition Cycloheximide
(Sigma-Aldrich, USA) was used at 50 μg/ml for the indicated periods of time. Cells were lysed using
cell-lysis buffer (RIPA). Standard procedures for SDS-PAGE and protein transfer on to membranes
were used. Membranes were incubated with RIG-1 (1:1000; #3743 Cell Signaling, USA) or the β-Actin
control (1:5000; #A5316 Sigma-Aldrich, USA) antibodies and rabbit or mouse HRP conjugated
antibodies (1:5000; Sigma-Aldrich, USA) as secondary antibodies were used. The signals were
detected with SuperSignal West Pico Chemiluminescent Substrate (Thermo Scientific, USA) and were
quantified by densitometry using the Quantity-1 software (BioRad), normalizing values to actin
content.
HLA genotyping
HLA typing was performed using commercial panels (INNO-LiPA HLA-DRB1 Plus kit and INNO-LiPA
HLA-B Update Plus; INNOGENETICS, Ghent, Belgium) based on the reverse hybridization principle.
Briefly, after a PCR round with specific primers, amplified biotinylated DNA material was chemically

Tenorio et al., Human Mutation 5
denatured and the separated strands are hybridized against a total of 37 sequence-specific DNA
probes (INNO-LiPA HLA-DRB1 Plus kit) or 66 sequence-specific probes (INNO-LiPA HLA-B Update
Plus) and two control probes. The strips were stringently washed to remove any mismatched
amplified material and then streptavidin-conjugated with alkaline phosphatase was added to bind to
any biotinylated hybrids. Results are observed following Incubation with a substrate chromogen
solution resulting in a purple/brown precipitate.

Tenorio et al., Human Mutation 6
Supp. Figure S1. Conservation of amino acids at the three RNF125 mutations. The changes observed
in patients 2, 3 and 5 are green-shaded. Amino acids of the zinc fingers are grey-shaded (partial view
in the first and third fingers).

Tenorio et al., Human Mutation 7
Supp. Figure S2. Three-dimensional modelling of RNF125 protein and the three mutants using
PHYRE2 software (Kelley and Sternberg, 2009). Comparison of 3D structure reveals differences
between the wild type protein and the three different mutations. The p.M112I mutation cause the
formation of a B-lamin in residues 197-169 (Pro-Val-Phe) which is not present in the wild type
protein. The p.S163L mutant lacks the B-lamin residues in the amino acids 138-140 as in the wild
type, and also triggers the expansion of the alpha-helix from amino acids 150-161 to 150-164, now
including the mutated 163 residue. The p.R174C mutation causes the formation of a novel B-lamin
motif at positions 168-169 which is not present in the WT. Additionally the alpha-helix in which the
missense mutation is located, changes from ProLeuCysArg to LeuCysCys. Confidence key ranges from
red (high) to dark blue (low).
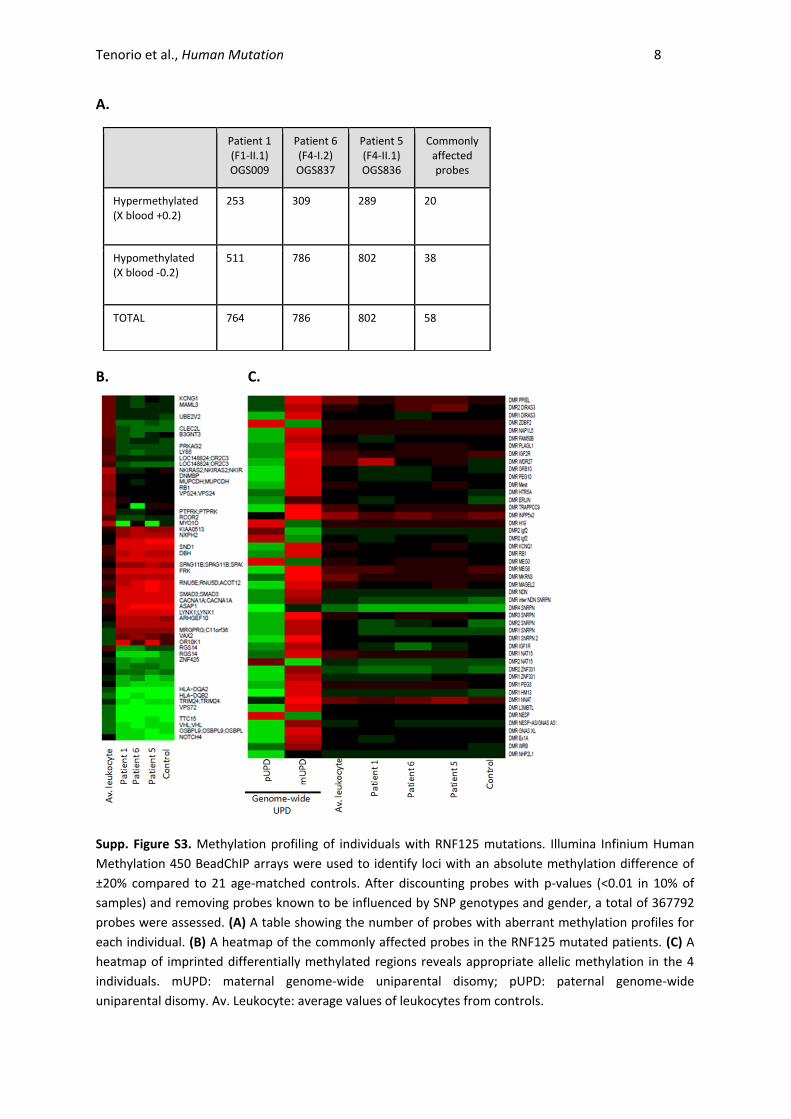
Tenorio et al., Human Mutation 8
A.
B. C.
Supp. Figure S3. Methylation profiling of individuals with RNF125 mutations. Illumina Infinium Human
Methylation 450 BeadChIP arrays were used to identify loci with an absolute methylation difference of
±20% compared to 21 age-matched controls. After discounting probes with p-values (<0.01 in 10% of
samples) and removing probes known to be influenced by SNP genotypes and gender, a total of 367792
probes were assessed. (A) A table showing the number of probes with aberrant methylation profiles for
each individual. (B) A heatmap of the commonly affected probes in the RNF125 mutated patients. (C) A
heatmap of imprinted differentially methylated regions reveals appropriate allelic methylation in the 4
individuals. mUPD: maternal genome-wide uniparental disomy; pUPD: paternal genome-wide
uniparental disomy. Av. Leukocyte: average values of leukocytes from controls.
Patient 1 (F1-II.1) OGS009
Patient 6 (F4-I.2) OGS837
Patient 5 (F4-II.1) OGS836
Commonly affected probes
Hypermethylated (X blood +0.2)
253 309 289 20
Hypomethylated (X blood -0.2)
511 786 802 38
TOTAL 764 786 802 58
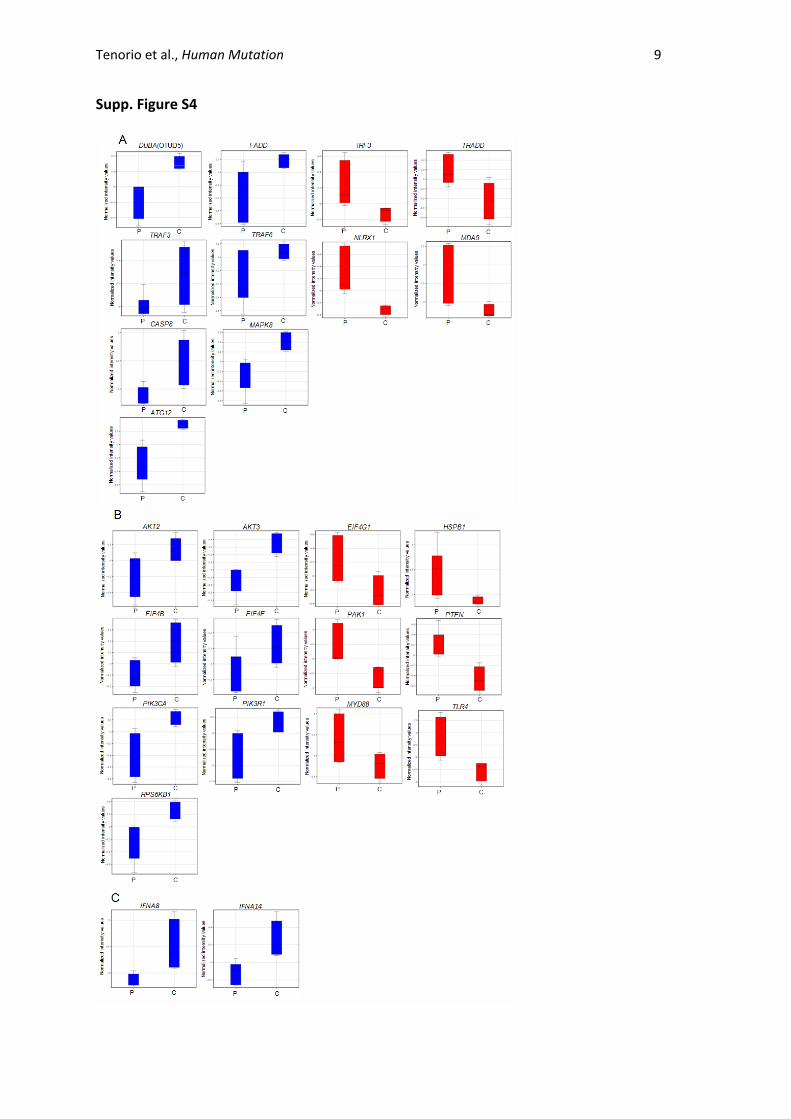
Tenorio et al., Human Mutation 9
Supp. Figure S4
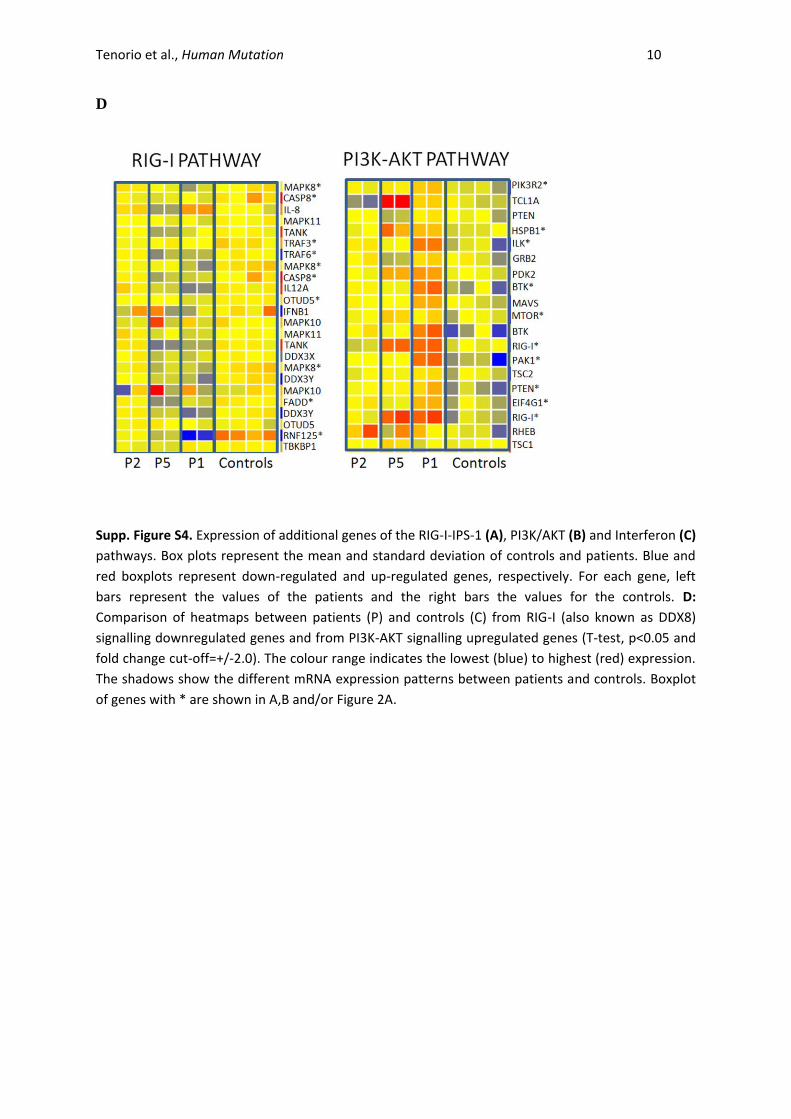
Tenorio et al., Human Mutation 10
D
Supp. Figure S4. Expression of additional genes of the RIG-I-IPS-1 (A), PI3K/AKT (B) and Interferon (C)
pathways. Box plots represent the mean and standard deviation of controls and patients. Blue and
red boxplots represent down-regulated and up-regulated genes, respectively. For each gene, left
bars represent the values of the patients and the right bars the values for the controls. D:
Comparison of heatmaps between patients (P) and controls (C) from RIG-I (also known as DDX8)
signalling downregulated genes and from PI3K-AKT signalling upregulated genes (T-test, p<0.05 and
fold change cut-off=+/-2.0). The colour range indicates the lowest (blue) to highest (red) expression.
The shadows show the different mRNA expression patterns between patients and controls. Boxplot
of genes with * are shown in A,B and/or Figure 2A.
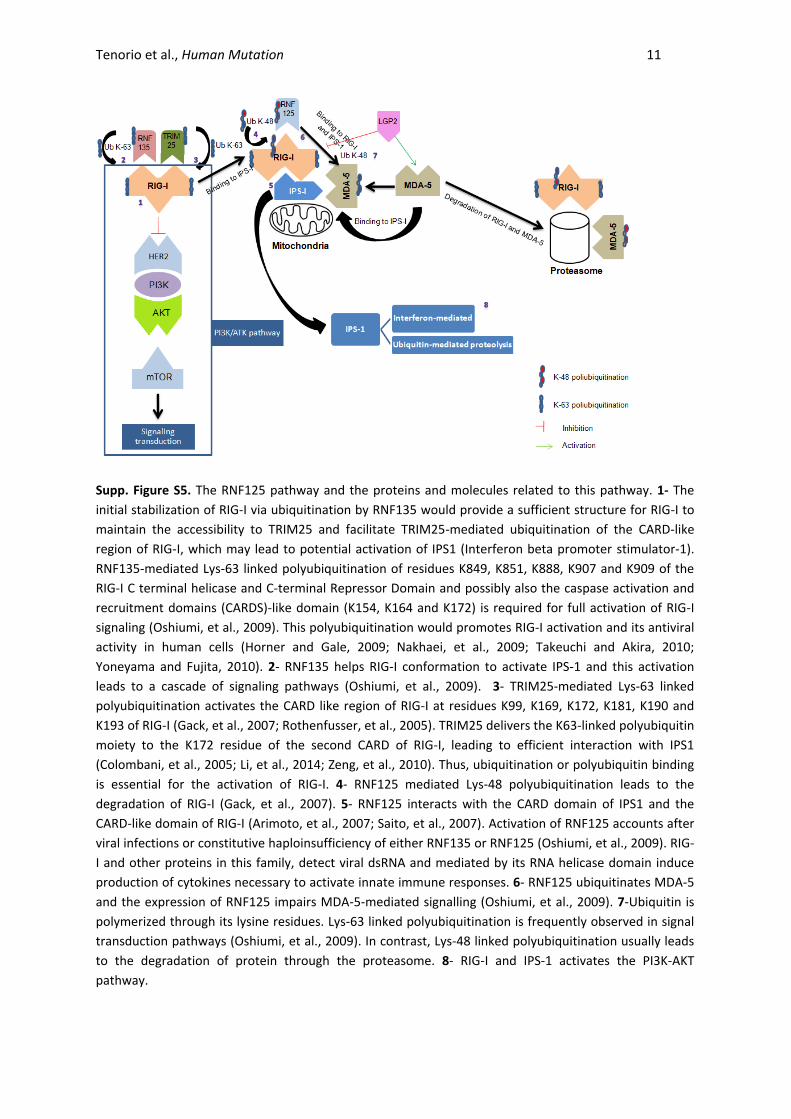
Tenorio et al., Human Mutation 11
Supp. Figure S5. The RNF125 pathway and the proteins and molecules related to this pathway. 1- The
initial stabilization of RIG-I via ubiquitination by RNF135 would provide a sufficient structure for RIG-I to
maintain the accessibility to TRIM25 and facilitate TRIM25-mediated ubiquitination of the CARD-like
region of RIG-I, which may lead to potential activation of IPS1 (Interferon beta promoter stimulator-1).
RNF135-mediated Lys-63 linked polyubiquitination of residues K849, K851, K888, K907 and K909 of the
RIG-I C terminal helicase and C-terminal Repressor Domain and possibly also the caspase activation and
recruitment domains (CARDS)-like domain (K154, K164 and K172) is required for full activation of RIG-I
signaling (Oshiumi, et al., 2009). This polyubiquitination would promotes RIG-I activation and its antiviral
activity in human cells (Horner and Gale, 2009; Nakhaei, et al., 2009; Takeuchi and Akira, 2010;
Yoneyama and Fujita, 2010). 2- RNF135 helps RIG-I conformation to activate IPS-1 and this activation
leads to a cascade of signaling pathways (Oshiumi, et al., 2009). 3- TRIM25-mediated Lys-63 linked
polyubiquitination activates the CARD like region of RIG-I at residues K99, K169, K172, K181, K190 and
K193 of RIG-I (Gack, et al., 2007; Rothenfusser, et al., 2005). TRIM25 delivers the K63-linked polyubiquitin
moiety to the K172 residue of the second CARD of RIG-I, leading to efficient interaction with IPS1
(Colombani, et al., 2005; Li, et al., 2014; Zeng, et al., 2010). Thus, ubiquitination or polyubiquitin binding
is essential for the activation of RIG-I. 4- RNF125 mediated Lys-48 polyubiquitination leads to the
degradation of RIG-I (Gack, et al., 2007). 5- RNF125 interacts with the CARD domain of IPS1 and the
CARD-like domain of RIG-I (Arimoto, et al., 2007; Saito, et al., 2007). Activation of RNF125 accounts after
viral infections or constitutive haploinsufficiency of either RNF135 or RNF125 (Oshiumi, et al., 2009). RIG-
I and other proteins in this family, detect viral dsRNA and mediated by its RNA helicase domain induce
production of cytokines necessary to activate innate immune responses. 6- RNF125 ubiquitinates MDA-5
and the expression of RNF125 impairs MDA-5-mediated signalling (Oshiumi, et al., 2009). 7-Ubiquitin is
polymerized through its lysine residues. Lys-63 linked polyubiquitination is frequently observed in signal
transduction pathways (Oshiumi, et al., 2009). In contrast, Lys-48 linked polyubiquitination usually leads
to the degradation of protein through the proteasome. 8- RIG-I and IPS-1 activates the PI3K-AKT
pathway.
Tenorio et al., Human Mutation 12
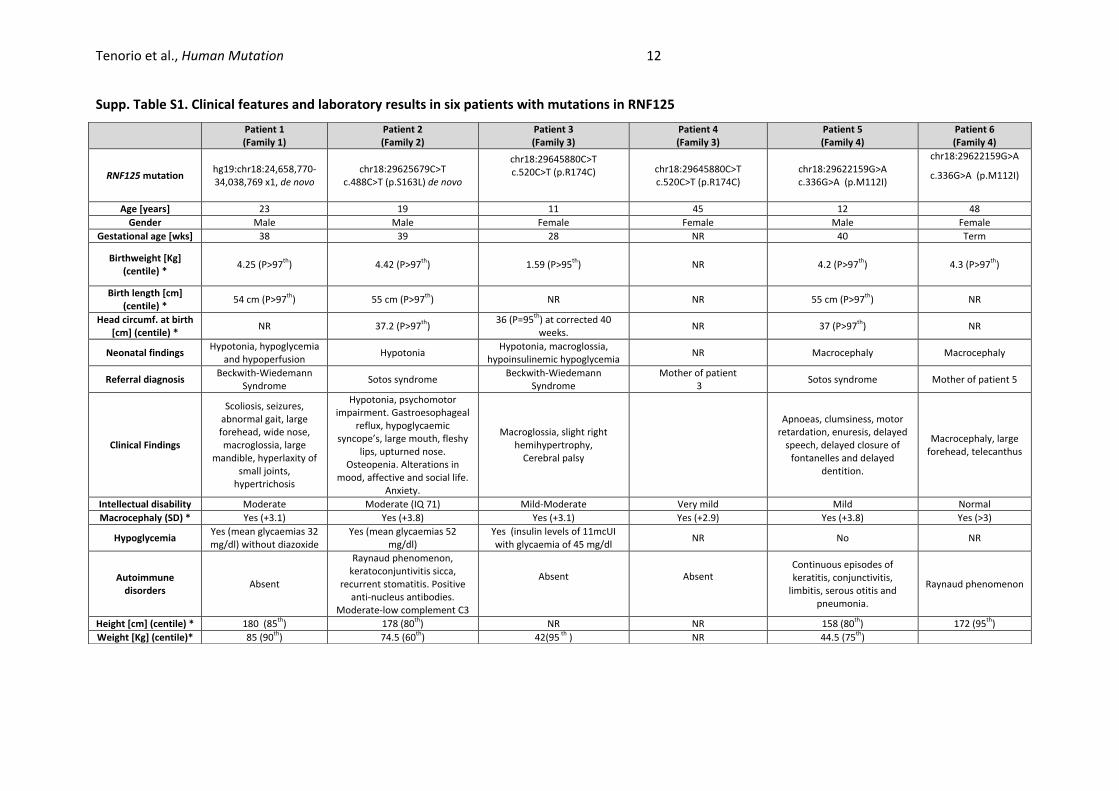
Supp. Table S1. Clinical features and laboratory results in six patients with mutations in RNF125
Patient 1 (Family 1)
Patient 2 (Family 2)
Patient 3 (Family 3)
Patient 4 (Family 3)
Patient 5 (Family 4)
Patient 6 (Family 4)
RNF125 mutation hg19:chr18:24,658,770-34,038,769 x1, de novo
chr18:29625679C>T c.488C>T (p.S163L) de novo
chr18:29645880C>T c.520C>T (p.R174C) chr18:29645880C>T
c.520C>T (p.R174C) chr18:29622159G>A c.336G>A (p.M112I)
chr18:29622159G>A
c.336G>A (p.M112I)
Age [years] 23 19 11 45 12 48
Gender Male Male Female Female Male Female
Gestational age [wks] 38 39 28 NR 40 Term
Birthweight [Kg] (centile) *
4.25 (P>97th
) 4.42 (P>97th
) 1.59 (P>95th
) NR 4.2 (P>97th
) 4.3 (P>97th
)
Birth length [cm] (centile) *
54 cm (P>97th
) 55 cm (P>97th
) NR NR 55 cm (P>97th
) NR
Head circumf. at birth [cm] (centile) *
NR 37.2 (P>97th
) 36 (P=95
th) at corrected 40 weeks.
NR 37 (P>97th
) NR
Neonatal findings Hypotonia, hypoglycemia
and hypoperfusion Hypotonia
Hypotonia, macroglossia, hypoinsulinemic hypoglycemia
NR Macrocephaly Macrocephaly
Referral diagnosis Beckwith-Wiedemann
Syndrome Sotos syndrome
Beckwith-Wiedemann Syndrome
Mother of patient 3
Sotos syndrome Mother of patient 5
Clinical Findings
Scoliosis, seizures, abnormal gait, large forehead, wide nose, macroglossia, large
mandible, hyperlaxity of small joints,
hypertrichosis
Hypotonia, psychomotor impairment. Gastroesophageal
reflux, hypoglycaemic syncope’s, large mouth, fleshy
lips, upturned nose. Osteopenia. Alterations in
mood, affective and social life. Anxiety.
Macroglossia, slight right hemihypertrophy,
Cerebral palsy
Apnoeas, clumsiness, motor retardation, enuresis, delayed
speech, delayed closure of fontanelles and delayed
dentition.
Macrocephaly, large forehead, telecanthus
Intellectual disability Moderate Moderate (IQ 71) Mild-Moderate Very mild Mild Normal
Macrocephaly (SD) * Yes (+3.1) Yes (+3.8) Yes (+3.1) Yes (+2.9) Yes (+3.8) Yes (>3)
Hypoglycemia Yes (mean glycaemias 32 mg/dl) without diazoxide
Yes (mean glycaemias 52 mg/dl)
Yes (insulin levels of 11mcUI with glycaemia of 45 mg/dl
NR No NR
Autoimmune disorders
Absent
Raynaud phenomenon, keratoconjuntivitis sicca,
recurrent stomatitis. Positive anti-nucleus antibodies.
Moderate-low complement C3
Absent Absent Continuous episodes of keratitis, conjunctivitis,
limbitis, serous otitis and pneumonia.
Raynaud phenomenon
Height [cm] (centile) * 180 (85th
) 178 (80th
) NR NR 158 (80th
) 172 (95th
)
Weight [Kg] (centile)* 85 (90th
) 74.5 (60th
) 42(95 th
) NR 44.5 (75th
)
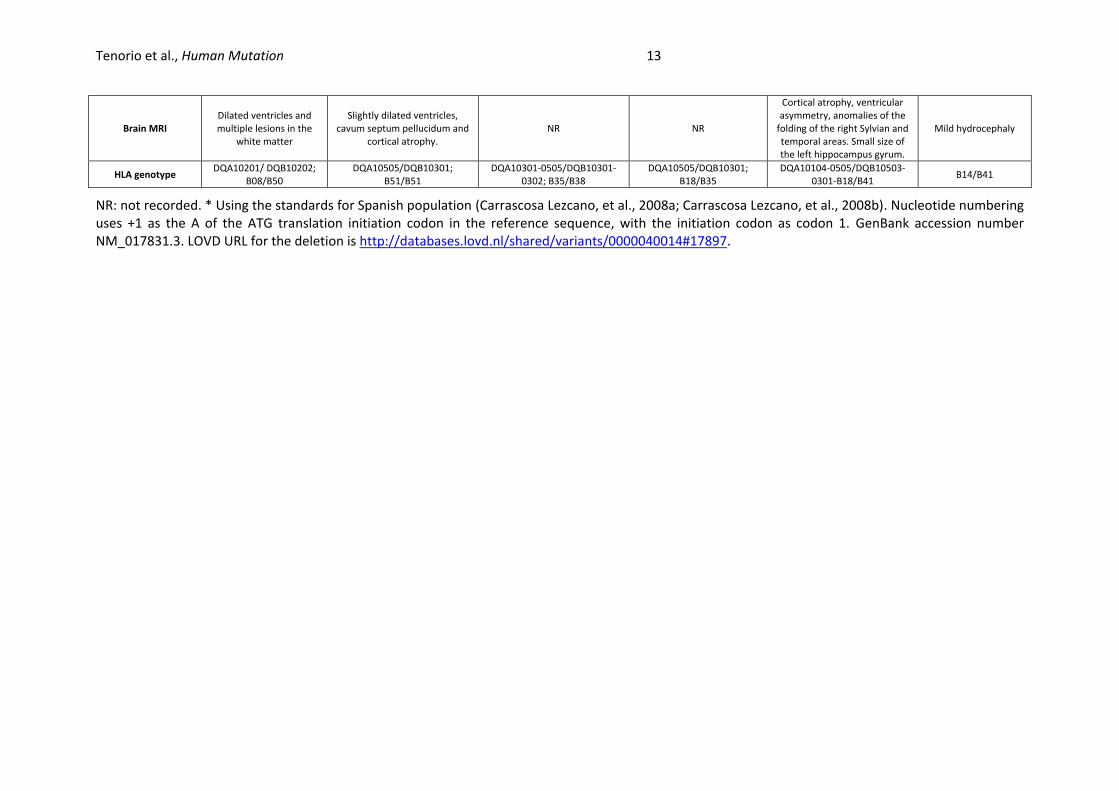
Tenorio et al., Human Mutation 13
NR: not recorded. * Using the standards for Spanish population (Carrascosa Lezcano, et al., 2008a; Carrascosa Lezcano, et al., 2008b). Nucleotide numbering uses +1 as the A of the ATG translation initiation codon in the reference sequence, with the initiation codon as codon 1. GenBank accession number NM_017831.3. LOVD URL for the deletion is http://databases.lovd.nl/shared/variants/0000040014#17897.
Brain MRI Dilated ventricles and multiple lesions in the
white matter
Slightly dilated ventricles, cavum septum pellucidum and
cortical atrophy. NR NR
Cortical atrophy, ventricular asymmetry, anomalies of the
folding of the right Sylvian and temporal areas. Small size of the left hippocampus gyrum.
Mild hydrocephaly
HLA genotype DQA10201/ DQB10202;
B08/B50 DQA10505/DQB10301;
B51/B51 DQA10301-0505/DQB10301-
0302; B35/B38 DQA10505/DQB10301;
B18/B35 DQA10104-0505/DQB10503-
0301-B18/B41 B14/B41
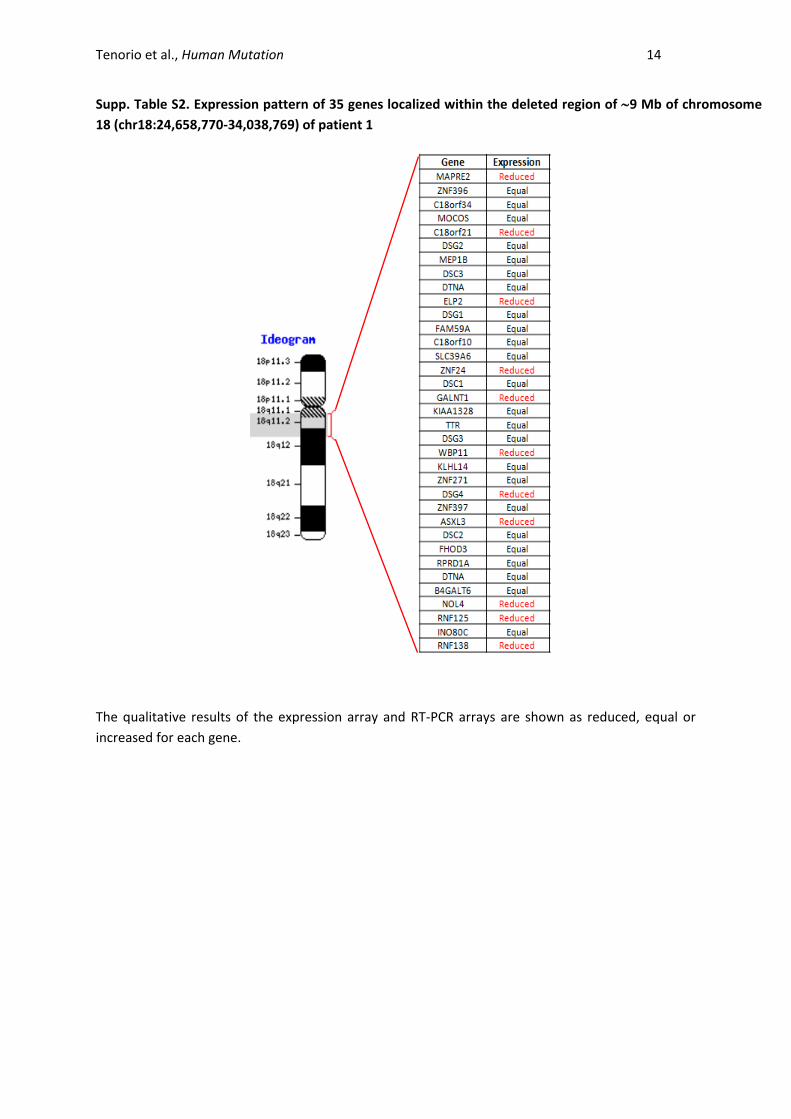
Tenorio et al., Human Mutation 14
Supp. Table S2. Expression pattern of 35 genes localized within the deleted region of 9 Mb of chromosome
18 (chr18:24,658,770-34,038,769) of patient 1
The qualitative results of the expression array and RT-PCR arrays are shown as reduced, equal or
increased for each gene.
Tenorio et al., Human Mutation 15
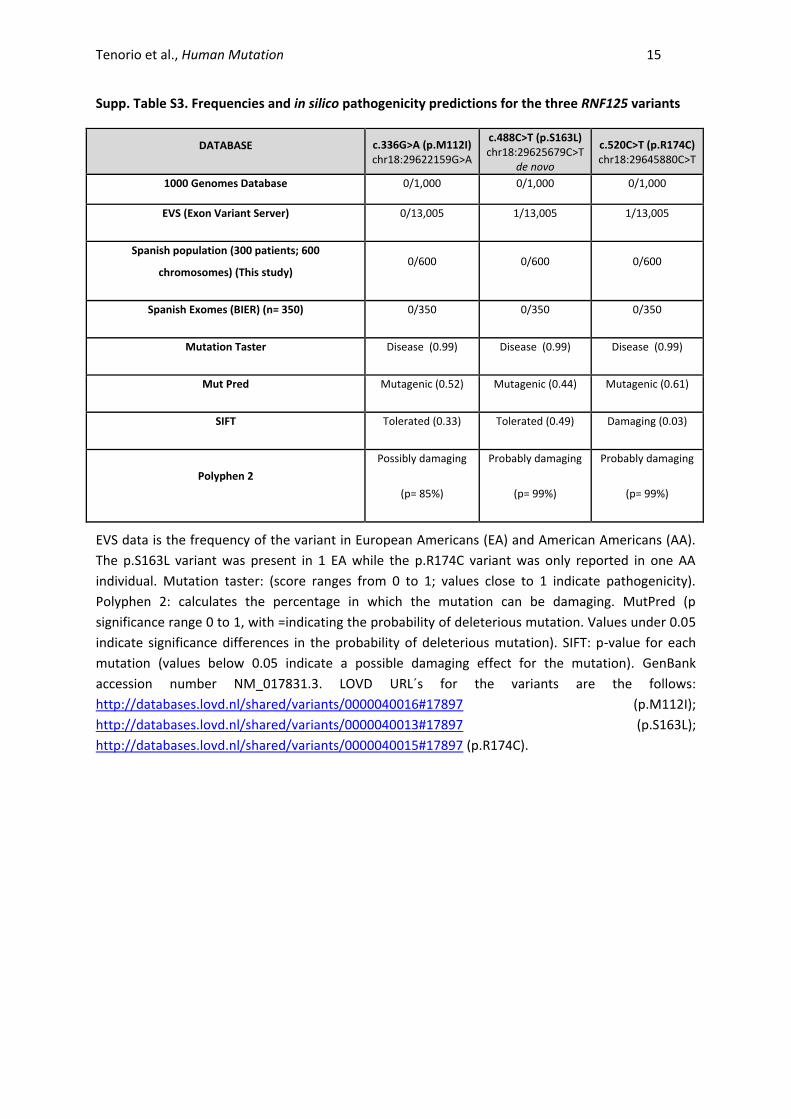
Supp. Table S3. Frequencies and in silico pathogenicity predictions for the three RNF125 variants
DATABASE c.336G>A (p.M112I) chr18:29622159G>A
c.488C>T (p.S163L) chr18:29625679C>T
de novo
c.520C>T (p.R174C) chr18:29645880C>T
1000 Genomes Database 0/1,000 0/1,000 0/1,000
EVS (Exon Variant Server) 0/13,005 1/13,005 1/13,005
Spanish population (300 patients; 600
chromosomes) (This study) 0/600 0/600 0/600
Spanish Exomes (BIER) (n= 350) 0/350 0/350 0/350
Mutation Taster Disease (0.99) Disease (0.99) Disease (0.99)
Mut Pred Mutagenic (0.52) Mutagenic (0.44) Mutagenic (0.61)
SIFT Tolerated (0.33) Tolerated (0.49) Damaging (0.03)
Polyphen 2
Possibly damaging
(p= 85%)
Probably damaging
(p= 99%)
Probably damaging
(p= 99%)
EVS data is the frequency of the variant in European Americans (EA) and American Americans (AA).
The p.S163L variant was present in 1 EA while the p.R174C variant was only reported in one AA
individual. Mutation taster: (score ranges from 0 to 1; values close to 1 indicate pathogenicity).
Polyphen 2: calculates the percentage in which the mutation can be damaging. MutPred (p
significance range 0 to 1, with =indicating the probability of deleterious mutation. Values under 0.05
indicate significance differences in the probability of deleterious mutation). SIFT: p-value for each
mutation (values below 0.05 indicate a possible damaging effect for the mutation). GenBank
accession number NM_017831.3. LOVD URL´s for the variants are the follows:
http://databases.lovd.nl/shared/variants/0000040016#17897 (p.M112I);
http://databases.lovd.nl/shared/variants/0000040013#17897 (p.S163L);
http://databases.lovd.nl/shared/variants/0000040015#17897 (p.R174C).
Tenorio et al., Human Mutation 16
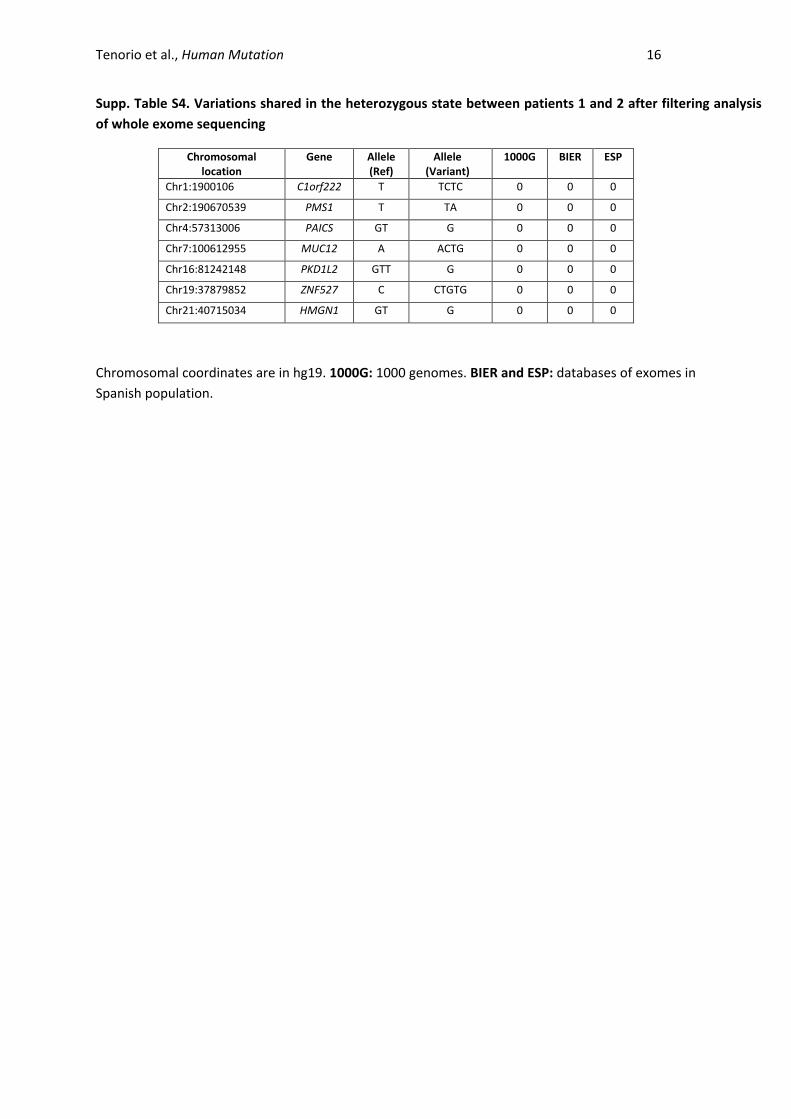
Supp. Table S4. Variations shared in the heterozygous state between patients 1 and 2 after filtering analysis
of whole exome sequencing
Chromosomal location
Gene Allele (Ref)
Allele (Variant)
1000G BIER ESP
Chr1:1900106 C1orf222 T TCTC 0 0 0
Chr2:190670539 PMS1 T TA 0 0 0
Chr4:57313006 PAICS GT G 0 0 0
Chr7:100612955 MUC12 A ACTG 0 0 0
Chr16:81242148 PKD1L2 GTT G 0 0 0
Chr19:37879852 ZNF527 C CTGTG 0 0 0
Chr21:40715034 HMGN1 GT G 0 0 0
Chromosomal coordinates are in hg19. 1000G: 1000 genomes. BIER and ESP: databases of exomes in
Spanish population.
Tenorio et al., Human Mutation 17
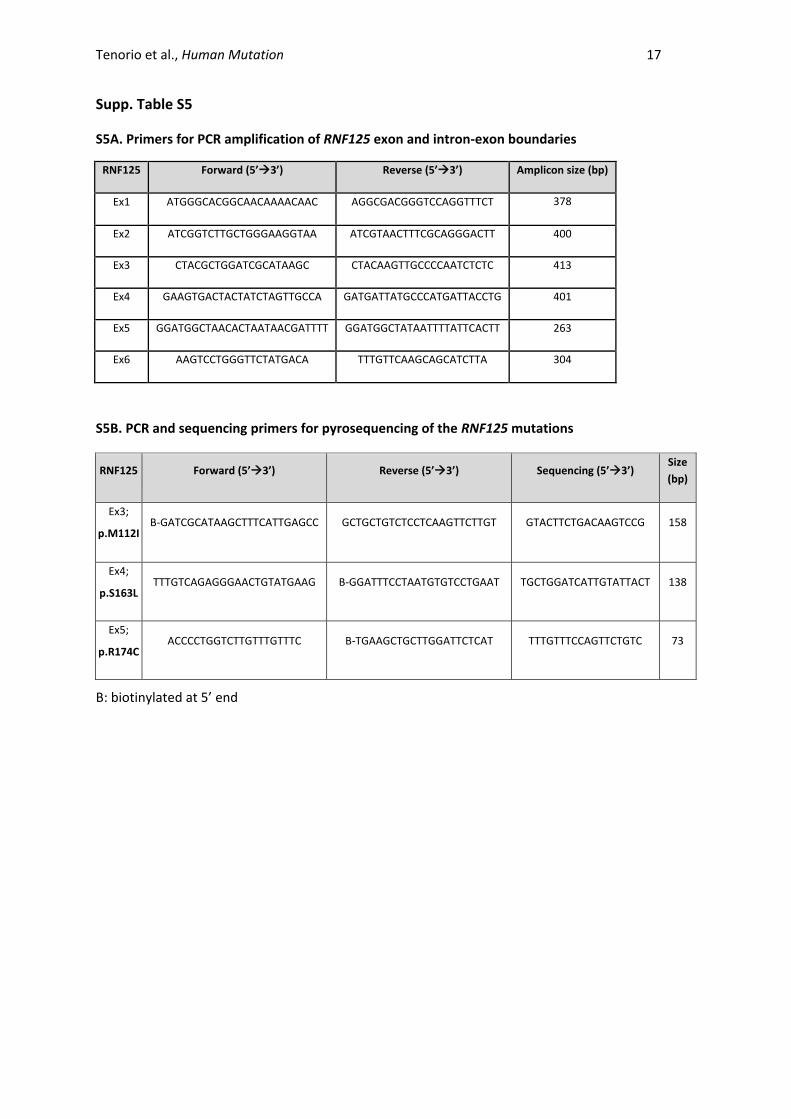
Supp. Table S5
S5A. Primers for PCR amplification of RNF125 exon and intron-exon boundaries
RNF125 Forward (5’3’) Reverse (5’3’) Amplicon size (bp)
Ex1 ATGGGCACGGCAACAAAACAAC AGGCGACGGGTCCAGGTTTCT 378
Ex2 ATCGGTCTTGCTGGGAAGGTAA ATCGTAACTTTCGCAGGGACTT 400
Ex3 CTACGCTGGATCGCATAAGC CTACAAGTTGCCCCAATCTCTC 413
Ex4 GAAGTGACTACTATCTAGTTGCCA GATGATTATGCCCATGATTACCTG 401
Ex5 GGATGGCTAACACTAATAACGATTTT GGATGGCTATAATTTTATTCACTT 263
Ex6 AAGTCCTGGGTTCTATGACA TTTGTTCAAGCAGCATCTTA 304
S5B. PCR and sequencing primers for pyrosequencing of the RNF125 mutations
RNF125 Forward (5’3’) Reverse (5’3’) Sequencing (5’3’) Size
(bp)
Ex3;
p.M112I B-GATCGCATAAGCTTTCATTGAGCC GCTGCTGTCTCCTCAAGTTCTTGT GTACTTCTGACAAGTCCG 158
Ex4;
p.S163L TTTGTCAGAGGGAACTGTATGAAG B-GGATTTCCTAATGTGTCCTGAAT TGCTGGATCATTGTATTACT 138
Ex5;
p.R174C ACCCCTGGTCTTGTTTGTTTC B-TGAAGCTGCTTGGATTCTCAT TTTGTTTCCAGTTCTGTC 73
B: biotinylated at 5’ end

Tenorio et al., Human Mutation 18
Supp. References
Adzhubei IA, Schmidt S, Peshkin L, Ramensky VE, Gerasimova A, Bork P, Kondrashov AS, Sunyaev SR.
2010. A method and server for predicting damaging missense mutations. Nat Methods
7(4):248-9.
Arimoto K, Takahashi H, Hishiki T, Konishi H, Fujita T, Shimotohno K. 2007. Negative regulation of the
RIG-I signaling by the ubiquitin ligase RNF125. Proc Natl Acad Sci U S A 104(18):7500-5.
Carrascosa Lezcano A, Fernandez Garcia JM, Fernandez Ramos C, Ferrandez Longas A, Lopez-Siguero
JP, Sanchez Gonzalez E, Sobradillo Ruiz B, Yeste Fernandez D, Grupo Colaborador E. 2008a.
[Spanish cross-sectional growth study 2008. Part II. Height, weight and body mass index
values from birth to adulthood]. An Pediatr (Barc) 68(6):552-69.
Carrascosa Lezcano A, Ferrandez Longas A, Yeste Fernandez D, Garcia-Dihinx Villanova J, Romo
Montejo A, Copil Copil A, Almar Mendoza J, Salcedo Abizanda S, Gussinye Canadell M,
Baguer Mor L. 2008b. [Spanish cross-sectional growth study 2008. Part I: weight and height
values in newborns of 26-42 weeks of gestational age]. An Pediatr (Barc) 68(6):544-51.
Colombani J, Bianchini L, Layalle S, Pondeville E, Dauphin-Villemant C, Antoniewski C, Carre C, Noselli
S, Leopold P. 2005. Antagonistic actions of ecdysone and insulins determine final size in
Drosophila. Science 310(5748):667-70.
Douglas J, Cilliers D, Coleman K, Tatton-Brown K, Barker K, Bernhard B, Burn J, Huson S, Josifova D,
Lacombe D and others. 2007. Mutations in RNF135, a gene within the NF1 microdeletion
region, cause phenotypic abnormalities including overgrowth. Nat Genet 39(8):963-5.
Gack MU, Shin YC, Joo CH, Urano T, Liang C, Sun L, Takeuchi O, Akira S, Chen Z, Inoue S and others.
2007. TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity.
Nature 446(7138):916-920.
Horner SM, Gale M, Jr. 2009. Intracellular innate immune cascades and interferon defenses that
control hepatitis C virus. J Interferon Cytokine Res 29(9):489-98.
Kelley LA, Sternberg MJ. 2009. Protein structure prediction on the Web: a case study using the Phyre
server. Nat Protoc 4(3):363-71.
Li B, Krishnan VG, Mort ME, Xin F, Kamati KK, Cooper DN, Mooney SD, Radivojac P. 2009. Automated
inference of molecular mechanisms of disease from amino acid substitutions. Bioinformatics
25(21):2744-50.
Li S, Wang C, Sandanaraj E, Aw SS, Koe CT, Wong JJ, Yu F, Ang BT, Tang C, Wang H. 2014. The
SCFSlimb E3 ligase complex regulates asymmetric division to inhibit neuroblast overgrowth.
EMBO Rep 15(2):165-74.

Tenorio et al., Human Mutation 19
Medina I, De Maria A, Bleda M, Salavert F, Alonso R, Gonzalez CY, Dopazo J. 2012. VARIANT:
Command Line, Web service and Web interface for fast and accurate functional
characterization of variants found by Next-Generation Sequencing. Nucleic Acids Res
40(Web Server issue):W54-8.
Nakhaei P, Genin P, Civas A, Hiscott J. 2009. RIG-I-like receptors: sensing and responding to RNA
virus infection. Semin Immunol 21(4):215-22.
Ng PC, Henikoff S. 2001. Predicting deleterious amino acid substitutions. Genome Res 11(5):863-74.
Oshiumi H, Matsumoto M, Hatakeyama S, Seya T. 2009. Riplet/RNF135, a RING finger protein,
ubiquitinates RIG-I to promote interferon-beta induction during the early phase of viral
infection. J Biol Chem 284(2):807-17.
Rothenfusser S, Goutagny N, DiPerna G, Gong M, Monks BG, Schoenemeyer A, Yamamoto M, Akira
S, Fitzgerald KA. 2005. The RNA helicase Lgp2 inhibits TLR-independent sensing of viral
replication by retinoic acid-inducible gene-I. J Immunol 175(8):5260-8.
Saito T, Hirai R, Loo YM, Owen D, Johnson CL, Sinha SC, Akira S, Fujita T, Gale M, Jr. 2007. Regulation
of innate antiviral defenses through a shared repressor domain in RIG-I and LGP2. Proc Natl
Acad Sci U S A 104(2):582-7.
Schwarz JM, Cooper DN, Schuelke M, Seelow D. 2014. MutationTaster2: mutation prediction for the
deep-sequencing age. Nat Methods 11(4):361-2.
Takeuchi O, Akira S. 2010. Pattern recognition receptors and inflammation. Cell 140(6):805-20.
Vallespin E, Palomares Bralo M, Mori MA, Martin R, Garcia-Minaur S, Fernandez L, de Torres ML,
Garcia-Santiago F, Mansilla E, Santos F and others. 2013. Customized high resolution CGH-
array for clinical diagnosis reveals additional genomic imbalances in previous well-defined
pathological samples. Am J Med Genet A 161A(8):1950-60.
Yoneyama M, Fujita T. 2010. Recognition of viral nucleic acids in innate immunity. Rev Med Virol
20(1):4-22.
Zeng W, Sun L, Jiang X, Chen X, Hou F, Adhikari A, Xu M, Chen ZJ. 2010. Reconstitution of the RIG-I
pathway reveals a signaling role of unanchored polyubiquitin chains in innate immunity. Cell
141(2):315-30.