Structural flexibility of the Gαs α-helical domain in the ... · Diane Calinski d, Gisselle...
6
Structural flexibility of the Gαs α-helical domain in the β 2 -adrenoceptor Gs complex Gerwin H. Westfield a,1 , Søren G. F. Rasmussen b,c,1 , Min Su a,1 , Somnath Dutta a,1 , Brian T. DeVree d , Ka Young Chung b , Diane Calinski d , Gisselle Velez-Ruiz d , Austin N. Oleskie a , Els Pardon e,f , Pil Seok Chae g , Tong Liu h , Sheng Li h , Virgil L. Woods, Jr. h , Jan Steyaert e,f , Brian K. Kobilka b,2 , Roger K. Sunahara d,2 , and Georgios Skiniotis a,2 a Life Sciences Institute and Department of Biological Chemistry, d Department of Pharmacology, University of Michigan Medical School, Ann Arbor, MI 48109; b Department of Molecular and Cellular Physiology, Stanford University School of Medicine, Stanford, CA 94305; c Department of Neuroscience and Pharmacology, The Panum Institute, University of Copenhagen, 2200 Copenhagen N, Denmark; e Structural Biology Brussels and f VIB Department of Structural Biology, Vrije Universiteit Brussels, 1050 Brussels, Belgium; g Department of Chemistry, University of Wisconsin, Madison, WI 53706; and h Department of Chemistry, University of California at San Diego, La Jolla, CA 92093 Contributed by Brian K. Kobilka, August 19, 2011 (sent for review August 9, 2011) The active-state complex between an agonist-bound receptor and a guanine nucleotide-free G protein represents the fundamental signaling assembly for the majority of hormone and neurotrans- mitter signaling. We applied single-particle electron microscopy (EM) analysis to examine the architecture of agonist-occupied β 2 - adrenoceptor (β 2 AR) in complex with the heterotrimeric G protein Gs (Gαsβγ). EM 2D averages and 3D reconstructions of the deter- gent-solubilized complex reveal an overall architecture that is in very good agreement with the crystal structure of the active-state ternary complex. Strikingly however, the α-helical domain of Gαs appears highly flexible in the absence of nucleotide. In contrast, the presence of the pyrophosphate mimic foscarnet (phosphono- formate), and also the presence of GDP, favor the stabilization of the α-helical domain on the Ras-like domain of Gαs. Molecular modeling of the α-helical domain in the 3D EM maps suggests that in its stabilized form it assumes a conformation reminiscent to the one observed in the crystal structure of Gαs-GTPγS. These data argue that the α-helical domain undergoes a nucleotide- dependent transition from a flexible to a conformationally stabilized state. G protein-coupled receptor | negative stain electron microscopy | random conical tilt T he majority of hormones and neurotransmitters communi- cate information to cells via G protein-coupled receptors (GPCRs), which instigate intracellular signaling by activating their cognate heterotrimeric G proteins on the cytoplasmic side. GPCRs constitute the largest family of membrane proteins and play essential roles in regulating every aspect of normal physi- ology, thereby representing major pharmacological targets. De- spite a wealth of biochemical and biophysical studies on inactive and active conformations of several heterotrimeric G proteins, the molecular underpinnings of G-protein activation remain elusive. The β 2 -adrenergic receptor (β 2 AR) and its complex with heterotrimeric stimulatory G-protein Gs (Gαsβγ) represent an ideal model system for the large family of GPCRs activated by diffusible ligands. Agonist binding to the β 2 AR promotes inter- actions with GDP-bound Gsαβγ heterotrimer, leading to the exchange of GDP for GTP, and the functional dissociation of Gs into Gα-GTP and Gβγ subunits. To examine the architecture of agonist occupied β 2 AR in complex with Gαsβγ under different conditions, we used electron microscopy (EM) and single-parti- cle analysis. Because of the limited size of the protein complex (∼148 kDa), we visualized specimens embedded in negative stain, which provides sufficient contrast from relatively small protein assemblies (1). This approach allowed us to obtain 2D projection averages and 3D reconstructions that provided new insights into dynamic features of the β 2 AR-Gs complex, and helped guide a successful approach to crystallize the complex enabling a high-resolution structure (2). Results and Discussion In a first step, we sought to examine the architecture of com- plexes in the nucleotide-free state of Gαs. Before coupling with an agonist-bound receptor, the nucleotide binding pocket of the α-subunit of the Gαsβγ heterotrimer is occupied by GDP. Upon forming a complex with the β 2 AR, GDP dissociates, and the resulting nucleotide-free β 2 AR-Gs complex is highly stable (2). EM visualization of the nucleotide-free complex showed a monodisperse particle population (Fig. 1A, and SI Appendix, Fig. S1). Reference-free alignment and classification of ∼17,000 particle projections revealed characteristic class averages with an overall density that is in very good agreement with the crystal structure of the complex (2). Because of its shape, the complex adsorbs on the carbon support with small variations (± 20°) of mainly two diametrically opposite preferred orientations that generate practically identical, mirror-related 2D projections (SI Appendix, Figs. S2 and S3). The distinct features of the class averages in these preferred orientations allowed us to assign the negative stain projection profiles from specific components of the complex (Fig. 1 B and C). A central oval density represents the β 2 AR in a detergent micelle, with a small protruding density corresponding to T4 lysozyme (T4L) that replaces the unstructured extracellular N terminus of the receptor and serves as an orienting landmark. This interpretation was confirmed by EM analysis of complexes lacking T4L (SI Appendix, Fig. S4). Some class averages of the T4L-β 2 AR-Gs complex do not reveal a density corresponding to the T4L. Besides the presence of a relatively flexible linker connecting T4L and the β 2 AR, this effect is mostly because T4L lies at an angle to the longitudinal axis of the complex, as shown in the X-ray structure (2). Because of this geometry, even a 10° variation in the way the particle adsorbs on the carbon support drastically reduces the visibility of the T4L projection profile, as demonstrated by projection simulation experiments (SI Appen- dix, Fig. S5). Thus, the visibility of the T4L projection profile is very sensitive to even limited out-of-plane particle tilting (e.g., because of particle “rock” and “roll” or because of variations in the flatness of the carbon support). Because we observe a single density corresponding to T4L, the detergent micelle contains only a single copy of the β 2 AR, in agreement with the crystal structure. Therefore, the significant additional density around Author contributions: V.L.W., J.S., B.K.K., R.K.S., and G.S. designed research; G.H.W., S.G.F.R., M.S., S.D., B.T.D., K.Y.C., D.C., G.V.-R., A.N.O., E.P., P.S.C., T.L., S.L., and G.S. performed research; and B.K.K., R.K.S., and G.S. wrote the paper. The authors declare no conflict of interest. 1 G.H.W., S.G.F.R., M.S., and S.D. contributed equally to this work. 2 To whom correspondence may be addressed. E-mail: [email protected], sunahara@ umich.edu, or [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1113645108/-/DCSupplemental. 16086–16091 | PNAS | September 20, 2011 | vol. 108 | no. 38 www.pnas.org/cgi/doi/10.1073/pnas.1113645108 Downloaded by guest on May 10, 2021
Transcript of Structural flexibility of the Gαs α-helical domain in the ... · Diane Calinski d, Gisselle...

Structural flexibility of the Gαs α-helical domainin the β2-adrenoceptor Gs complexGerwin H. Westfielda,1, Søren G. F. Rasmussenb,c,1, Min Sua,1, Somnath Duttaa,1, Brian T. DeVreed, Ka Young Chungb,Diane Calinskid, Gisselle Velez-Ruizd, Austin N. Oleskiea, Els Pardone,f, Pil Seok Chaeg, Tong Liuh, Sheng Lih,Virgil L. Woods, Jr.h, Jan Steyaerte,f, Brian K. Kobilkab,2, Roger K. Sunaharad,2, and Georgios Skiniotisa,2
aLife Sciences Institute and Department of Biological Chemistry, dDepartment of Pharmacology, University of Michigan Medical School, Ann Arbor, MI 48109;bDepartment of Molecular and Cellular Physiology, Stanford University School of Medicine, Stanford, CA 94305; cDepartment of Neuroscience andPharmacology, The Panum Institute, University of Copenhagen, 2200 Copenhagen N, Denmark; eStructural Biology Brussels and fVIB Department of StructuralBiology, Vrije Universiteit Brussels, 1050 Brussels, Belgium; gDepartment of Chemistry, University of Wisconsin, Madison, WI 53706; and hDepartment ofChemistry, University of California at San Diego, La Jolla, CA 92093
Contributed by Brian K. Kobilka, August 19, 2011 (sent for review August 9, 2011)
The active-state complex between an agonist-bound receptor anda guanine nucleotide-free G protein represents the fundamentalsignaling assembly for the majority of hormone and neurotrans-mitter signaling. We applied single-particle electron microscopy(EM) analysis to examine the architecture of agonist-occupied β2-adrenoceptor (β2AR) in complex with the heterotrimeric G proteinGs (Gαsβγ). EM 2D averages and 3D reconstructions of the deter-gent-solubilized complex reveal an overall architecture that is invery good agreement with the crystal structure of the active-stateternary complex. Strikingly however, the α-helical domain of Gαsappears highly flexible in the absence of nucleotide. In contrast,the presence of the pyrophosphate mimic foscarnet (phosphono-formate), and also the presence of GDP, favor the stabilization ofthe α-helical domain on the Ras-like domain of Gαs. Molecularmodeling of the α-helical domain in the 3D EM maps suggeststhat in its stabilized form it assumes a conformation reminiscentto the one observed in the crystal structure of Gαs-GTPγS. Thesedata argue that the α-helical domain undergoes a nucleotide-dependent transition from a flexible to a conformationallystabilized state.
G protein-coupled receptor | negative stain electron microscopy | randomconical tilt
The majority of hormones and neurotransmitters communi-cate information to cells via G protein-coupled receptors
(GPCRs), which instigate intracellular signaling by activatingtheir cognate heterotrimeric G proteins on the cytoplasmic side.GPCRs constitute the largest family of membrane proteins andplay essential roles in regulating every aspect of normal physi-ology, thereby representing major pharmacological targets. De-spite a wealth of biochemical and biophysical studies on inactiveand active conformations of several heterotrimeric G proteins,the molecular underpinnings of G-protein activation remainelusive. The β2-adrenergic receptor (β2AR) and its complex withheterotrimeric stimulatory G-protein Gs (Gαsβγ) represent anideal model system for the large family of GPCRs activated bydiffusible ligands. Agonist binding to the β2AR promotes inter-actions with GDP-bound Gsαβγ heterotrimer, leading to theexchange of GDP for GTP, and the functional dissociation of Gsinto Gα-GTP and Gβγ subunits. To examine the architecture ofagonist occupied β2AR in complex with Gαsβγ under differentconditions, we used electron microscopy (EM) and single-parti-cle analysis. Because of the limited size of the protein complex(∼148 kDa), we visualized specimens embedded in negativestain, which provides sufficient contrast from relatively smallprotein assemblies (1). This approach allowed us to obtain 2Dprojection averages and 3D reconstructions that provided newinsights into dynamic features of the β2AR-Gs complex, andhelped guide a successful approach to crystallize the complexenabling a high-resolution structure (2).
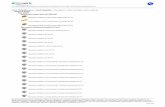
Results and DiscussionIn a first step, we sought to examine the architecture of com-plexes in the nucleotide-free state of Gαs. Before coupling withan agonist-bound receptor, the nucleotide binding pocket of theα-subunit of the Gαsβγ heterotrimer is occupied by GDP. Uponforming a complex with the β2AR, GDP dissociates, and theresulting nucleotide-free β2AR-Gs complex is highly stable (2).EM visualization of the nucleotide-free complex showed amonodisperse particle population (Fig. 1A, and SI Appendix, Fig.S1). Reference-free alignment and classification of ∼17,000particle projections revealed characteristic class averages with anoverall density that is in very good agreement with the crystalstructure of the complex (2). Because of its shape, the complexadsorbs on the carbon support with small variations (± 20°) ofmainly two diametrically opposite preferred orientations thatgenerate practically identical, mirror-related 2D projections (SIAppendix, Figs. S2 and S3).The distinct features of the class averages in these preferred
orientations allowed us to assign the negative stain projectionprofiles from specific components of the complex (Fig. 1 B andC). A central oval density represents the β2AR in a detergentmicelle, with a small protruding density corresponding to T4lysozyme (T4L) that replaces the unstructured extracellular Nterminus of the receptor and serves as an orienting landmark.This interpretation was confirmed by EM analysis of complexeslacking T4L (SI Appendix, Fig. S4). Some class averages of theT4L-β2AR-Gs complex do not reveal a density corresponding tothe T4L. Besides the presence of a relatively flexible linkerconnecting T4L and the β2AR, this effect is mostly because T4Llies at an angle to the longitudinal axis of the complex, as shownin the X-ray structure (2). Because of this geometry, even a 10°variation in the way the particle adsorbs on the carbon supportdrastically reduces the visibility of the T4L projection profile, asdemonstrated by projection simulation experiments (SI Appen-dix, Fig. S5). Thus, the visibility of the T4L projection profile isvery sensitive to even limited out-of-plane particle tilting (e.g.,because of particle “rock” and “roll” or because of variations inthe flatness of the carbon support). Because we observe a singledensity corresponding to T4L, the detergent micelle containsonly a single copy of the β2AR, in agreement with the crystalstructure. Therefore, the significant additional density around
Author contributions: V.L.W., J.S., B.K.K., R.K.S., and G.S. designed research; G.H.W., S.G.F.R.,M.S., S.D., B.T.D., K.Y.C., D.C., G.V.-R., A.N.O., E.P., P.S.C., T.L., S.L., and G.S. performedresearch; and B.K.K., R.K.S., and G.S. wrote the paper.
The authors declare no conflict of interest.1G.H.W., S.G.F.R., M.S., and S.D. contributed equally to this work.2To whom correspondence may be addressed. E-mail: [email protected], [email protected], or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1113645108/-/DCSupplemental.
16086–16091 | PNAS | September 20, 2011 | vol. 108 | no. 38 www.pnas.org/cgi/doi/10.1073/pnas.1113645108
Dow
nloa
ded
by g
uest
on
May
10,
202
1

the receptor stems from the large micelle formed by the de-tergent (3). Diametrically opposite to the T4L domain, two maininteracting densities representing the Gs trimer appear in closeproximity to the receptor on its intracellular surface. One of thetwo domains appears to extensively interact with the receptordensity, suggesting it corresponds to the Ras-like domain of Gαs,while its neighboring domain has a profile consistent with theside view of Gβγ. Interestingly however, several class averagesrevealed an additional small globular density bound on the Ras-like domain of Gαs (Fig. 1B, Right, and SI Appendix, Fig. S2). Inthis location, the additional density could only be attributed tothe α-helical (AH) domain of Gαs, occupying a position expectedfrom the crystal structure of Gαs-GTPγS alone (4) and thestructure of the Gi heterotrimer (5) (Fig. 1C, and SI Appendix,Fig. S6), but in entirely different location from that observed inthe crystal structure of the β2AR-Gs complex (2). To assess thefraction of particles displaying the AH domain in this location,we selected and classified only projections clearly displaying theprofiles of Ras-like, Gβγ, β2AR, and T4L domain densities in thesame position, thereby restricting the range of particle projectionorientations. The classification revealed that the AH domain wasordered on the Ras-like domain in ∼35% of the particles, but inmost other particle projections this density was absent (Fig. 1B,and SI Appendix, Fig. S3). It should also be noted that in contrast
to the T4L domain, the projection profile of the AH domain inthis position (SI Appendix, Fig. S3) is not sensitive to the rela-tively limited out-of-plane tilts (± 20°) of the preferred particleorientation on the carbon support (SI Appendix, Fig. S5). ThisEM analysis provided the initial evidence for a high degree ofmobility of the AH domain relative to the Ras domain in thenucleotide-free β2AR-Gs complex. Furthermore, the structuralheterogeneity observed provided insights to the challenges inobtaining 3D crystals of the complex.To promote complex stabilization for high-resolution struc-
tural studies, we generated and screened llama antibodies(nanobodies) to the purified complex (2). Nanobodies are small(∼15 kDa), clonable variable domains of a heavy chain-onlyantibody, obtained by immunizing a llama with purified de-tergent-solubilized β2AR-Gs complex stabilized with a shorthomobifunctional crosslinker (2). By screening samples withnegative-stain EM, we identified two nanobodies (Nb35 andNb37) that bound to the complex, but not the receptor alone.Class averages of particles incubated with Nb35 indicated a ho-mogeneous protein complex displaying increased density be-tween Gαs and Gβγ. Even though the nanobody projectionprofile was not clearly distinguished in the preferred particleorientations, its presence appeared to enhance the uniformity inthe disposition of Gαs-ras and Gβγ domains. The use of Nb35
C
D
A B
T4Lβ2AR
Gs-βγGαs
m m
Ras G
AH
Ras
T4Lβ2AR
Gs-βγαs
m m
37%63%
nucleotide-free T4L-β2AR-Gs
nucleotide-free T4L-β2AR-Gs + Nb37
Gαs
T4L
Gs-βγ
β2AR
Gs-βγ AH
T4L
Gαs
Gs-βγ Gαs
T4Lβ2AR
AH
m m m m
Gs-βγ Gαs
T4Lβ2AR
m m m mβ2AR
Fig. 1. Two-dimensional projection analysis of the T4L-β2AR-Gs complex in the nucleotide-free state. (A) Raw EM image of detergent-solubilized T4L-β2AR-Gscomplex embedded in negative stain. (Scale bar, 50 nm.) (B) Representative EM class averages of the nucleotide-free complex with the projection profile ofthe AH domain not visible (Left), or visible on the Ras domain (Right, AH indicated by arrow). The cartoon models represent the conformations reflected bythe EM averages, with the one on the left depicting the variable positioning of the AH domain, suggesting flexibility or multiple conformations (the positionof the detergent micelle is indicated by gray shaded arcs and labeled with “m”). (Scale bar, 10 nm.) (C) Reprojections (Upper) of the crystal structure (2)(Lower) in the same overall orientation as B reveal the identity of each EM density component. The crystal structure on the Right shows the AH domain in thesame position (relative to the Ras-like domain) as the one determined in the crystal structure of Gαs-GTPγS alone (4). (D) Representative class averages ofnucleotide-free complex with nanobody Nb37 bound on the AH domain (arrows) reveal its flexibility. (Scale bar, 10 nm.)
Westfield et al. PNAS | September 20, 2011 | vol. 108 | no. 38 | 16087
PHARM
ACO
LOGY
Dow
nloa
ded
by g
uest
on
May
10,
202
1

indeed allowed us to obtain the crystal structure of the T4L-β2AR-Gs complex, which showed that the nanobody binds at theinterface of the Gαs-Ras and Gβγ. In this location, Nb35 wouldnot be predicted to interact with or stabilize the AH domain (2).Accordingly, the classification of Nb35-bound complexes revealeda similar distribution of particles with an ordered AH domain onthe Ras-like domain as in the absence of Nb35 (SI Appendix,Figs. S7–S9).In contrast to Nb35, Nb37 appears bound directly to the AH
domain [as also determined by deuterium-exchange MS(DXMS)]) (SI Appendix, Fig. S10) and could be distinguished insingle-particle EM class averages of the β2AR-Gs complex as anextension of the AH domain. Using Nb37 as a domain markerallowed us to track the variable positioning of the small AH re-gion of Gαs (SI Appendix, Figs. S11 and 12). The 2D class av-erages from this preparation reveal an enhanced and elongateddensity adopting different orientations around the Ras-like do-main, ranging from close proximity to the Gβγ module to ex-tending much further out of the complex in the oppositedirection (Fig. 1D and SI Appendix, Fig. S12). Collectively, thesefindings suggest that in the absence of nucleotide, the AH do-main is flexible, thereby sampling different positions around theRas-like domain. Deuterium-exchange studies are consistentwith a dynamic interface between the Gαs Ras and AH domains(6). Therefore, the unexpected position of the “open” AH do-main in the crystal structure (2) represents just one of the pos-sible conformations.To obtain a more detailed view of the complex architecture,
we used the random conical tilt approach (7) to calculate initial3D reconstructions of complexes with and without ordered AHdomain on the Ras-like domain (SI Appendix, Fig. S13). Theseinitial 3D models were subsequently used for multireferencesupervised alignment (8, 9) to separate particle projections fromour entire dataset according to the AH positioning (see SI Ap-pendix). This approach allowed us to obtain quality 3D recon-structions from particle projections with and without densitycorresponding to AH domain on the Ras-like domain. Thereconstructions are in excellent agreement with the corre-sponding 2D averages (Fig. 2A and SI Appendix, Fig. S14). In 3Dreconstructions of particles where density for AH domain isobserved, its orientation relative to the Ras-like domain appearsto be similar to that found in the crystal structure of Gαs-GTPγS(4) (Fig. 2A and SI Appendix, Fig. S6). In addition, we obtained3D reconstructions of nucleotide-free complexes with boundNb37 marking the positioning of the AH domain. The 3D mapsclearly reveal that the Nb37-enhanced density of the AH domaincan adopt different conformations around the Ras-like domain,indicative of a relative flexibility in the interaction between thetwo domains (Fig. 2B and SI Appendix, Figs. S15 and S16). Thisvariability in the 3D conformation of the AH domain or the AH/Nb37 module around the Ras-like domain was further confirmedby cross-validating 3D reconstructions (SI Appendix, Fig. S17).As noted above, previous crystal structures of G proteins show
that bound nucleotides contribute to the stability of interactionsbetween the Ras and AH domains. We therefore investigatedthe positioning of the AH domain in the presence of guaninenucleotides and nucleotide fragments. Pyrophosphate (PPi),representing two phosphates in GTP or GDP, has been shownto bind Ras in a Mg2+-dependent manner, presumably at the β-and γ-phosphate positions (10). PPi and its chemically morestable analog, foscarnet, more known for its antiviral properties(11), also bind to heterotrimeric Gαs with an apparent affinity of∼0.5 and 1.6 mM, respectively, as determined by competitionbinding with a fluorescent GTPγS probe (Bodipy-GTPγS) (SIAppendix, Fig. S18). Binding of PPi and foscarnet most likelysubstitutes for the α- and β-phosphates of GDP rather than the β-and γ-phosphates of GTP. Binding of β- and γ-phosphates wouldresult in modification of the switch II domain with subsequent
Gβγ dissociation and dissolution of the complex (12). However,PPi (with or without Mg2+) does not disrupt the receptor-Gprotein complex. This finding is in contrast to dissociation of thecomplex observed with the GTP mimetic GTPγS (2).Although the presence of PPi does not appear to affect the AH
domain positioning, in the presence of Mg2+·foscarnet we observea significantly higher proportion (∼70%) of complexes with anordered AH region on the Ras-like domain (Fig. 3A and SI Ap-pendix, Figs. S19–S28). The observations in 2D class averages were
A
B
AH+Nb37AAAAAA
β2AR
Gs-βγAHGαsRas
4L
m m
T4L
β2AR
Gs-βγ Gαs
AH
AH
AH
Ras
Fig. 2. Three-dimensional reconstructions of the T4L-β2AR-Gs complex inthe nucleotide-free state. (A) Representative class averages and corre-sponding 3D reconstructions of particles in each category show the vari-ability in the positioning of the AH domain in the nucleotide-free complex.In the reconstruction to the left, the AH domain (orange ribbon) is shown inthe same position as found in the docked crystal structure (2). Absence ofsufficient density to accommodate this domain indicates that its position ishighly variable in this particle population. In the reconstruction to the right,the AH domain is modeled within the available EM density right below theRas-like domain of Gαs, as also suggested by the 2D averages. (B) Three-di-mensional reconstructions of distinct conformations of nucleotide-free T4L-β2AR-Gs complex with bound nanobody Nb37. The Nb37-enhanced densityof the AH domain (marked with an oval) shows variable positioning aroundthe Ras-like domain of Gαs. (Scale bars, 5 nm.)
16088 | www.pnas.org/cgi/doi/10.1073/pnas.1113645108 Westfield et al.
Dow
nloa
ded
by g
uest
on
May
10,
202
1

also reproduced by individual 3D reconstructions for the differentconformers in each state (SI Appendix, Figs. S24 and S28). Theseresults further confirm that the variability in the visibility of the AHdomain is indeed because of its variable positioning and not be-cause of negative stain artifacts, such as incomplete embedding.The ability of foscarnet to stabilize the AH domain on the Rasdomain suggests it is acting as a ligand fragment that binds to thenucleotide binding pocket. Given its low affinity, it is not sur-prising that the stabilization is incomplete.In contrast to PPi and foscarnet, addition of GDP or the
nonhydrolyzable GTP analog GTPγS leads to dissociation of theβ2AR-Gs complex (2). To examine the effect of GDP andGTPγS, we rapidly mixed the complex with nucleotide and im-
mediately fixed the sample by negative stain embedding. Addi-tion of either of these nucleotides at concentrations above 10 μMresulted in significant amounts of partially dissociated complexes(SI Appendix, Fig. S29). This result is expected because a largeexcess of either of these nucleotides would uncouple the Gprotein from the receptor. However, short incubation with lowerGDP concentrations (1 μM) and immediate sample fixation forEM allowed to us to examine intact complexes, revealing that theAH region was ordered in ∼60% of the intact particles (SI Ap-pendix, Figs. S30–S32). In contrast, even low concentrations ofGTPγS (1 μM) showed a significant amount of destabilizedcomplexes, and we were able to capture an array of intermediatedissociation states (Fig. 3B and SI Appendix, Figs. S30 and S33).Collectively, these data strongly suggest that the presence ofnucleotide, or nucleotide fragments such as foscarnet, results inAH domain stabilization against the Ras-like domain of Gαs. Inthe absence of nucleotide, the position of the AH domain ishighly variable (Figs. 3 and 4).Our results are in agreement with a recent study of the com-
plex formed by Gi and rhodopsin. Using double electron-elec-tron resonance spectroscopy, Hamm, Hubbell, and colleaguesdocumented large (up to 20 Å) changes in distance betweennitroxide probes positioned on the Ras and AH domains of Giupon formation of a complex with light-activated rhodopsin (13).The broad distance distributions observed for several labelingpairs are compatible with multiple conformations in dynamicequilibrium. Our findings are also consistent with results fromDXMS that show increased deuterium exchange at both the nu-cleotide binding pocket and at sites of interaction between the Rasand AH domains upon formation of the β2AR-Gs complex (6).Support for the open conformation in vivo may come from
studies on the action mechanism of the cholera toxin, the en-terotoxin secreted by the pathogen Vibrio cholerae. Cholera toxin,together with ADP ribosylation factor (ARF), ADP ribosylatesGαs at R201, rendering the residue catalytically ineffective andthe G protein GTPase-deficient (14). The ADP ribosylated andconstitutively active Gαs will continue to stimulate adenylyl cy-clase, cAMP production, and protein kinase A (PKA) activation.Activated PKA opens intestinal Cl− channels and leads to in-creased water secretion that results in diarrhea (15). Crystallo-graphic studies of G proteins in GDP and GTPγS-bound formsindicate that the catalytic arginine (R201 in Gs) is buried andlikely inaccessible to cholera toxin and ARF (4). However, for-mation of the nucleotide-free form of Gαs and opening of the Rasand AH domains would facilitate accessibility to R201.
GTP uncouplingNucleotide free dissociation
β2AR
Gs-βγ Gα-ras
Gα-H
4321
extracellular
Fig. 4. Model of conformational transitions in the β2AR-Gs complex. The nucleotide-free β2AR-Gs complex is characterized by a highly flexible AH domain(model 1). GTP binding promotes stabilization of the AH domain on the Ras-like domain of Gαs (model 2). In this model, the AH domain has the same positionas the one observed in the crystal structure of Gαs-GTPγS (4). GTP binding results in subsequent uncoupling of Gs and β2AR (model 3), with eventual dis-sociation of Gαs and Gβγ (model 4).
T4L
β2AR
complex in the presence of 1 μM GTPγS
A
B
Intact - 47% Receptor only - 15%Partially dissociated - 38%
noit al upopel citr ap
α−Helical domain stabilization on Ras
37%34% 33%
40%
69%62%
20%
0%
40%
60%
80%
10%
30%
50%
70%
Nuc. - free Nuc. - free+Nb35
1 mM PPi 10 mM PPi 10 mMFoscarnet
1 μMGDP
AHRas
T4L
β2AR
Gs-βγGαs
Fig. 3. Nucleotide-dependent positioning of the Gαs AH domain. (A) Dis-tribution of particles with a distinct projection profile of the AH domainstabilized on the Ras-like domain across different conditions (Inset Right,marked with a white dot). A class average of a particle with a nonvisible AHdomain is shown for comparison (Inset Left). The presence of foscarnet andGDP significantly increases the number of particles with stabilized AH do-main. (B) Representative class averages of the T4L-β2AR-Gs complex afterrapid mixing with GTPγS (1 μM) and immediate stain embedding reveal bothintact as well as partially or fully dissociated complexes. The fraction ofcorresponding subpopulations is indicated. (Scale bars, 10 nm.)
Westfield et al. PNAS | September 20, 2011 | vol. 108 | no. 38 | 16089
PHARM
ACO
LOGY
Dow
nloa
ded
by g
uest
on
May
10,
202
1

In conclusion, single-particle EM analysis of the β2AR-Gscomplex has provided novel structural insights into the dynamicnature of the assembly (Fig. 4). EM visualization of nanobodybound complexes allowed us to clearly reveal the variable posi-tioning of the AH domain under nucleotide-free conditions and,additionally, to identify conditions that were key for the suc-cessful characterization of the complex by X-ray crystallography.Furthermore, single-particle EM examination of the β2AR-Gscomplexes in varying concentrations of GDP and GTPγS en-abled us to capture transient intermediate dissociation statesbetween β2AR and Gs. This approach should prove useful forstudying other signaling complexes involving GPCRs and othermembrane proteins. The combination and integration of thesetechnologies will be crucial for studying structural aspects ofchallenging macromolecular complexes at large.
Experimental ProceduresSpecimen Preparation and EM Imaging of Negative-Stained Samples. The T4L-β2AR-Gs complex and nanobodies (Nb) were prepared as described in Ras-mussen et al. (2). Specimens were visualized by EM in the following con-ditions: (i) T4L-β2AR-Gs complex alone (nucleotide-free) (SI Appendix, Fig.S1); (ii) T4L-β2AR-Gs in presence of Nb35 (SI Appendix, Fig. S7); (ii) T4L-β2AR-Gs in presence of Nb37 (SI Appendix, Fig. S11); (iv) T4L-β2AR-Gs in presenceof 1 mM PPi (SI Appendix, Fig. S19 A and B); (v) T4L-β2AR-Gs in the presenceof 10 mM PPi (SI Appendix, Fig. S19 C and D); (vi) T4L-β2AR-Gs in the pres-ence of 10 mM foscarnet and 10 mM MgCl2 (SI Appendix, Fig. S25); (vii) T4L-β2AR-Gs in the presence of 1 μM GDP and 10 mM MgCl2 (SI Appendix, Fig.S30 A and B); (viii) T4L-β2AR-Gs in presence of 1 μM GTPγs and 10 mM MgCl2(SI Appendix, Fig. S30 C and D). All samples were prepared for EM using theconventional negative staining protocol (1). For nanobody labeling, the T4L-β2AR-Gs complex was incubated for 15 min at room temperature with ap-proximately equimolar concentrations of Nb35 or Nb37, and subsequentlyprepared by negative staining. For nucleotide (GDP or GTPγS) and nucleotidefragment (PPi or foscarnet) incubations, these components were rapidlymixed with the complex and the sample was immediately fixed by negative-stain embedding.
Specimens were imaged at room temperature with a Tecnai T12 electronmicroscope operated at 120 kV using low-dose procedures. Images wererecorded at a magnification of 71,138× and a defocus value of ∼1.5 μm ona Gatan US4000 CCD camera (SI Appendix, Figs. S1, S7, S11, S19, S25, S29,and S30). All images were binned (2 × 2 pixels) to obtain a pixel size of 4.16 Åon the specimen level. Tilt-pair particles from 60° and 0° images were se-lected using WEB (16). Particles for only 2D classification of 0° projectionswere excised using Boxer (part of the EMAN 1.9 software suite) (17). Thenumber of particles or tilt-pair particle projections per condition is providedin SI Appendix, Table S1.
Two-Dimensional Classifications and 3D Reconstructions of T4L-β2AR-Gs. The2D reference-free alignment and classification of particle projections wereperformed using SPIDER (16). For all conditions, the 0° particle projectionswere iteratively classified into multiple classes for 10 cycles (SI Appendix,Figs. S2, S8, S12, S20, S22, S26, S31, and S33). SI Appendix, Table S1 providesthe number of classes for each condition. For AH conformation assignments,we used the first classification to select only the particles from averagesclearly displaying the profiles of Ras-like, Gβγ, β2AR, and T4L domain den-sities in the same position, thereby restricting the range of particle pro-jection orientations. These projections were pulled together and subjectedto a second iterative classification (referred to as the “secondary” classifi-cation) (SI Appendix, Figs. S3, S9, S21, S23, S27, and S32). SI Appendix, TableS2 provides the number of classes and particle projections for each conditionin the secondary classification. For counting the numbers of particles withand without stabilized ΑΗ domain on the Ras-like domain, three differentoperators examined each secondary classification and assigned each classaverage according to the projection profile of the specific region (SI Ap-pendix, Figs. S3, S9, S21, S23, S27, S32, and S33). The assignment from thedifferent operators was in good agreement, and the particle numbers be-longing to individual classes were added to calculate percentages for eachconformation. Assignments for each full individual dataset were done inaddition to the secondary classification, and the results were in agreement.To test any bias, the particles from nucleotide-free, 1 mM PPi, 10 mM PPi,and foscarnet conditions were combined into a single dataset of 15,753particles and were classified into 200 classes. The individual class averageswere assigned as before according to the visibility of the AH domain, and
the percentage of projections from each condition was determinedaccording to the number of projections contributing to the assigned classaverages (SI Appendix, Fig. S34). The results of this “blind” test showed verygood agreement with our assignments from individual classifications.
For 3D reconstructions, in a first step we used the random conical tilttechnique (7) to determine initial 3D maps by back-projection of tiltedparticle images belonging to individual classes. After a first round of an-gular refinement, corresponding particles from the images of the untiltedspecimen were added, and the images were subjected to another cycle ofrefinement. We thus generated reliable initial models for complexes withvariability in the positioning of the ΑΗ domain of Gαs (SI Appendix, Fig.S13). After contrast transfer function (CTF) correction according to localdefocus values obtained by CTFTILT (18), the full dataset from each con-dition was subjected to multiple reference-supervised alignment (8, 9) withthe “multirefine” routine in EMAN (1.9) by using our initial models asreference maps. This approach allowed us to separate particles from theentire dataset (of each condition) according to the positioning of the ΑΗdomain of Gαs. The number of contributing particles in each condition andconformation is provided in SI Appendix, Table S3. For final maps, we usedthe separated datasets, as provided by the multiple reference-supervisedalignment, and used FREALIGN (19) for further refinement of the orien-tation parameters and reconstruction (SI Appendix, Fig. S14–S16, S24, andS28). The resolution for each map was determined at FSC = 0.5 and isprovided in SI Appendix, Table S3.
Molecular Modeling. The crystal structure of T4L-β2AR-Gs (2) was fit in the EMdensity as a rigid body. Because of the presence of the detergent micelle,which accounted for significant density surrounding β2AR, all dockingoperations were performed manually with visual inspection of the best fit.The docking of the T4L-β2AR-Gs complex revealed that the EM density cor-responding to Gβγwas shifted further away from the receptor in all 3D mapscompared with the crystal structure. The most likely cause for this variation isthe presence of the planar lipid bilayer provided by the cubic lipid phase inthe crystals, which most likely maintains a different interaction with Gβγcompared with the detergent micelle. However, it is also possible that thisdifference is attributed to the crystal packing, or a limited deformation ofthe complex because of the presence of the carbon support on the EM grid,or both. Accordingly, Gβγ was translated manually by 9 Å to best fit itsdensity yet retain all interactions with Gαs. This final model is shown un-modified for all fittings in 3D reconstructions (Fig. 2, and SI Appendix, Figs.S14–S16, S24, and S28). For maps showing the ΑΗ domain of Gαs on the Ras-like domain, we manually modeled the ΑΗ domain in its correspondingposition by taking into account steric constraints. In this conformation, theposition of the ΑΗ domain is very similar to the one observed in the crystalstructure of Gαs-GTPγS (4) (SI Appendix, Fig. S6).
Deuterium-ExchangeMS. For DXMS, 1.5mL of R:G complex or 1.5mL of R:G:NBwas mixed with 4.5 mL of D2O buffer (20 mMHepes, pH 7.5, 100 mMNaCl, 10mM BI-167107, 100 mM TCEP, 0.0015%MNG-3 in D2O) and incubated for 10,100, 1,000, and 10,000 s on ice. At the indicated times, the sample wasquenched by 15 mL of ice-cold Quench solution (0.1 M NaH2PO4, 20 mMTCEP, 16.6% glycerol, pH 2.4), immediately frozen on dry ice, and stored at –80 °C. Nondeuterated control was prepared in H2O buffer (20 mM Hepes, pH7.5, 100 mM NaCl, 10 μM BI-167107, 100 μM TCEP, 0.0015% MNG-3 in H2O),mixed with Quench solution, and snap-frozen on dry ice. Samples werethawed and immediately passed through an immobilized porcine pepsincolumn (16 mL bed volume) at a flow rate of 20 mL/min of 0.05% tri-fluoroacetic acid. Peptide fragments were collected contemporaneously ona C18 trap column for desalting and separated by a Magic C18AQ column(Michrom BioResources Inc.) using a linear gradient of acetonitrile from 6.4%to 38.4% over 30 min. MS analysis was performed using LCQ Classic massspectrometer from Thermo Finnigan, with capillary temperature of 20 °C.Deuterium quantification data were collected in MS1 profile mode, andpeptide identification data were collected in data-dependent MS/MS mode.Recovered peptide identification and analysis were carried out using DXMSExplorer (Sierra Analytics Inc.), a software specialized in processing DXMSdata (SI Appendix, Fig. S10).
Bodipy-GTPγS Binding. The effect of foscarnet and PPiwasmeasured using 100nM bodipy-GTPγS-FL (Invitrogen). Fluorescence intensity of bodipy-GTPγS-FL(lex ∼470 nm) increases upon G-protein binding, as demonstrated by McE-wan et al. (20). A wavelength scan of bodipy-GTPγS-FL (100 nM) in the ab-sence (dotted) or presence (solid) of a molar excess of purified Gαs (1 mM)was determined to assess optimal spectroscopy conditions. The capacity ofPPi and the chemically stable pyrophosphate analog of PPi, foscarnet, to
16090 | www.pnas.org/cgi/doi/10.1073/pnas.1113645108 Westfield et al.
Dow
nloa
ded
by g
uest
on
May
10,
202
1

inhibit bodipy-GTPγS-FL (lex∼470 nm, lem∼515 nm) was measured as de-scribed in SI Appendix, Fig. S18 B and C in both Gαs (B and C) and hetero-trimeric Gαsβγ (C). The fluorescence of 100 nM bodipy-GTPγS-FL wasmeasured in the presence of 1 mM G protein. PPi or foscarnet were addedtogether with bodipy-GTPγS-FL and initiated by the addition of G protein (1mM) in 20 mM Tris-HCl, pH 8.0, 3 mM MgCl2, 1 mM DTT in a final volume of200 mL bodipy-GTPγS-FL binding to heterotrimeric G protein included 0.1%dodecylmaltoside (final). Fluorescence was measured on a short time scale(600 s) to minimize the accumulation of hydrolysis product bodipy-phos-phate (21). Hydrolysis, which also appears as an increase in fluorescence, wasdetermined simply by chelating Mg2+ with 10 mM EDTA following the 600-sincubation. Inhibition of bodipy-GTPγS-FL by PPi and foscarnet can be re-versed with the subsequent addition of high Mg2+ (25 mM), which enhancesbodipy-GTPγS-FL binding, indicating that PPi and foscarnet are not irre-versibly binding or denaturing the G protein. Gαs was purified as describedin Sunahara et al. (4). Gαsβγwas purified as described by Rasmussen et al. (2).
Fluorescence was measured in a 96-well microtiter plate format on a M5fluorescence plate reader (Molecular Precision).
ACKNOWLEDGMENTS. We thank J. Tesmer for suggestions. This work wassupported by the Lundbeck Foundation Junior Group Leader Fellowship (toS.G.F.R.); the Fund for Scientific Research of Flanders (Fonds Wetenschappe-lijk Onderzoek-Vlaanderen) and the Institute for the encouragement ofScientific Research and Innovation of Brussels (to E.P. and J.S.); the NationalInstitute of General Medical Sciences (NIGMS) Molecular Biophysics TrainingGrant GM008270 (to G.H.W. and B.T.D.); the National Institute of NeuralDisorders and Stroke Grant R01-NS28471 (to B.K.K.); the Mather CharitableFoundation (to B.K.K.); NIGMS Grants R01-GM083118 (to B.K.K. and R.K.S.)and R01-GM068603 (to R.K.S.); National Institute of Diabetes and Digestiveand Kidney Diseases (NIDDK) Grant R01-DK090165 (to G.S.); NationalInstitutes of Health Grants AI076961, AI08192, AI2008031, CA118595,GM20501, GM066170, GM093325, and RR029388 (to V.L.W.); MichiganDiabetes Research and Training Center Grant, NIDDK, P60DK-20572 (toR.K.S.); and the University of Michigan Biological Sciences Scholars Program(R.K.S. and G.S). G.S. is a Pew Scholar of Biomedical Sciences.
1. Ohi M, Li Y, Cheng Y, Walz T (2004) Negative staining and image classification—Powerful tools in modern electron microscopy. Biol Proced Online 6(1):23–34.
2. Rasmussen SG, et al. (2011) Crystal structure of the β(2) adrenergic receptor-Gs proteincomplex. Nature, 10.1038/nature10361.
3. Rubinstein JL (2007) Structural analysis of membrane protein complexes by singleparticle electron microscopy. Methods 41:409–416.
4. Sunahara RK, Tesmer JJ, Gilman AG, Sprang SR (1997) Crystal structure of the adenylylcyclase activator Gsalpha. Science 278:1943–1947.
5. Wall MA, et al. (1995) The structure of the G protein heterotrimer Gi alpha 1 beta 1gamma 2. Cell 83:1047–1058.
6. Chung KY, et al. (2011) beta(2) adrenergic receptor-induced conformational changesin the heterotrimeric G protein Gs. Nature, 10.1038/nature10488.
7. Radermacher M, Wagenknecht T, Verschoor A, Frank J (1987) Three-dimensional re-construction from a single-exposure, random conical tilt series applied to the 50S ri-bosomal subunit of Escherichia coli. J Microsc 146(1):113–136.
8. Brink J, et al. (2004) Experimental verification of conformational variation of humanfatty acid synthase as predicted by normal mode analysis. Structure 12(2):185–191.
9. Ménétret JF, et al. (2005) Architecture of the ribosome-channel complex derived fromnative membranes. J Mol Biol 348:445–457.
10. Zhang B, Zhang Y, Shacter E, Zheng Y (2005) Mechanism of the guanine nucleotideexchange reaction of Ras GTPase—Evidence for a GTP/GDP displacement model.Biochemistry 44:2566–2576.
11. Sundquist B, Oberg B (1979) Phosphonoformate inhibits reverse transcriptase. J GenVirol 45:273–281.
12. Sprang SR (1997) G protein mechanisms: Insights from structural analysis. Annu RevBiochem 66:639–678.
13. Van Eps N, et al. (2011) Interaction of a G protein with an activated receptor opensthe interdomain interface in the alpha subunit. Proc Natl Acad Sci USA 108:9420–9424.
14. Freissmuth M, Gilman AG (1989) Mutations of GS alpha designed to alter the re-activity of the protein with bacterial toxins. Substitutions at ARG187 result in loss ofGTPase activity. J Biol Chem 264:21907–21914.
15. Gabriel SE, Brigman KN, Koller BH, Boucher RC, Stutts MJ (1994) Cystic fibrosis het-erozygote resistance to cholera toxin in the cystic fibrosis mouse model. Science 266(5182):107–109.
16. Frank J, et al. (1996) SPIDER and WEB: Processing and visualization of images in 3Delectron microscopy and related fields. J Struct Biol 116:190–199.
17. Ludtke SJ, Baldwin PR, Chiu W (1999) EMAN: Semiautomated software for high-res-olution single-particle reconstructions. J Struct Biol 128(1):82–97.
18. Mindell JA, Grigorieff N (2003) Accurate determination of local defocus and specimentilt in electron microscopy. J Struct Biol 142:334–347.
19. Grigorieff N (2007) FREALIGN: High-resolution refinement of single particle struc-tures. J Struct Biol 157(1):117–125.
20. McEwen DP, Gee KR, Kang HC, Neubig RR (2001) Fluorescent BODIPY-GTP analogs:Real-time measurement of nucleotide binding to G proteins. Anal Biochem 291(1):109–117.
21. Jameson EE, et al. (2005) Real-time detection of basal and stimulated G proteinGTPase activity using fluorescent GTP analogues. J Biol Chem 280:7712–7719.
Westfield et al. PNAS | September 20, 2011 | vol. 108 | no. 38 | 16091
PHARM
ACO
LOGY
Dow
nloa
ded
by g
uest
on
May
10,
202
1