Soutenance de thèse de Lorraine BOTTIN 27 février 2006
67
Déterminants de la variation moléculaire et phénotypique d’une espèce forestière en milieu insulaire Cas de Santalum austrocaledonicum en Nouvelle- Calédonie Soutenance de thèse de Lorraine BOTTIN 27 février 2006
description
Déterminants de la variation moléculaire et phénotypique d’une espèce forestière en milieu insulaire Cas de Santalum austrocaledonicum en Nouvelle-Calédonie. Soutenance de thèse de Lorraine BOTTIN 27 février 2006. - PowerPoint PPT Presentation
Transcript of Soutenance de thèse de Lorraine BOTTIN 27 février 2006

Déterminants de la variation moléculaire et phénotypique d’une espèce forestière en milieu insulaire
Cas de Santalum austrocaledonicum en Nouvelle-Calédonie
Soutenance de thèse de Lorraine BOTTIN
27 février 2006

1. Diversité et structuration génétiques sur la base de marqueurs microsatellites
2. Diversité des caractères phénotypiques
Introduction : les systèmes insulaires
Perspectives de recherche
Conclusion
Déterminants de la variation moléculaire et phénotypique d’une espèce forestière en milieu insulaire
Cas de Santalum austrocaledonicum en Nouvelle-Calédonie

Les systèmes insulaires: des « laboratoires évolutifs »
Introduction
Nouvelle Calédonie, île de Grande Terre
altitude
Écosystèmes uniques
Diversité des biotopes / taille île :

Introduction
Les systèmes insulaires: des « laboratoires évolutifs »
GRANDE TERRE
OUVEA
LIFOU
MARE
ILE DES PINS
ILES LOYAUTE Origine géologique :
Grande Terre + île des Pins: Gondwana
Îles Loyauté: anciens volcans
ennoyés, comblés par calcaire corallien
composition du sol
altitude
Écosystèmes uniques
Diversité des biotopes / taille île :

climat
Introduction
Les systèmes insulaires: des « laboratoires évolutifs »
composition du sol
altitude
Écosystèmes uniques
Diversité des biotopes / taille île :
2000 mm/an
900-1000 mm/an
1500 mm/an
Introduction
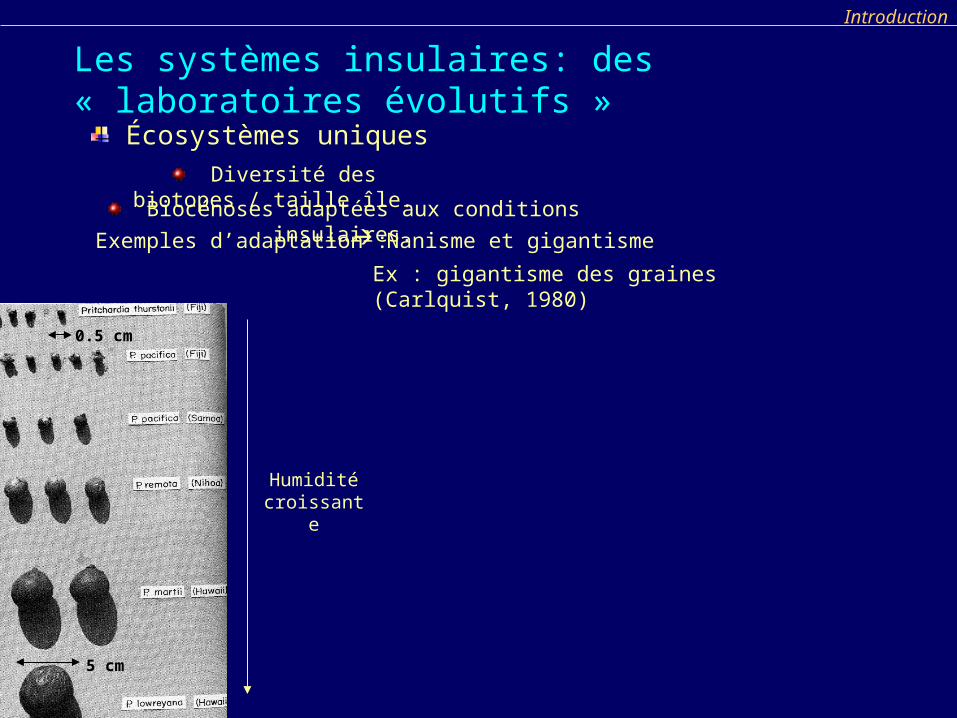
Ex : gigantisme des graines (Carlquist, 1980)
Les systèmes insulaires: des « laboratoires évolutifs »
Humidité croissante
Biocénoses adaptées aux conditions insulaires. Nanisme et gigantismeExemples d’adaptation :
Écosystèmes uniques
Diversité des biotopes / taille île.
5 cm
0.5 cm

Introduction
Les systèmes insulaires: des « laboratoires évolutifs »
Autogamie (plantes) : adaptation au manque de pollinisateurs
Ex : Cyclamen balearicum vs Cyclamen hederifolium
Cyclamen balearicum (insulaire) Cyclamen hederifolium
Biocénoses adaptées aux conditions insulaires. Nanisme et gigantismeExemples d’adaptation :
Écosystèmes uniques
Diversité des biotopes / taille île.
Ex : gigantisme des graines (Carlquist, 1980)

Introduction
Les systèmes insulaires: des « laboratoires évolutifs »
Perte des structures de dissémination et des facultés de vol
Biocénoses adaptées aux conditions insulaires. Nanisme et gigantisme Exemples d’adaptation :
Écosystèmes uniques
Diversité des biotopes / taille île.
Cagou (Nouvelle
Calédonie)
Bidens pilosa (Amérique continentale)
Bidens populifolia (Hawaï)
Ex : gigantisme des graines (Carlquist, 1980)
Autogamie (plantes) : adaptation au manque de pollinisateurs
Ex : Cyclamen balearicum vs Cyclamen hederifolium

Introduction
Les systèmes insulaires: des « laboratoires évolutifs »
Biocénoses adaptées aux conditions insulaires. Nanisme et gigantisme :Exemples d’adaptation :
Écosystèmes uniques
Diversité des biotopes / taille île.
Ensemble d’espèces adaptés au milieu, vulnérables face aux invasions
donc particulièrement soumis aux extinctions
Perte des structures de dissémination et des facultés de vol
Ex : gigantisme des graines (Carlquist, 1980)
Autogamie (plantes) : adaptation au manque de pollinisateurs
Ex : Cyclamen balearicum vs Cyclamen hederifolium

Statut isolé Migration réduite diminution / rupture du flux de gènes
Introduction
Les systèmes insulaires: des « laboratoires évolutifs »
1000 km100 km
Écosystèmes uniques
Forces évolutives particulières par rapport au continent Sélection naturelle pour s’adapter aux écosystèmes insulaires
150 km1500 km

Faible taille des populations Phénomènes stochastiques : dérive génétique
Introduction
Faible nombre d’individus fondateurs
Les systèmes insulaires: des « laboratoires évolutifs »
100 km
Statut isolé Migration réduite
Écosystèmes uniques
Forces évolutives particulières par rapport au continent Sélection naturelle pour s’adapter aux écosystèmes insulaires
diminution / rupture du flux de gènes
5 km

Espèces insulaires
plus différenciées que les espèces continentales
possèdent moins de diversité
Introduction
Les systèmes insulaires: des « laboratoires évolutifs »
Écosystèmes uniques
Faible taille des populations Phénomènes stochastiques : dérive génétique
Faible nombre d’individus fondateurs
Statut isolé Migration réduite
Forces évolutives particulières par rapport au continent Sélection naturelle pour s’adapter aux écosystèmes insulaires
diminution / rupture du flux de gènes

Plantes insulaires : modèles pour aborder de nombreux problèmes en biologie évolutive
Les bénéfices adaptatifs de l’autogamie et l’allogamie
Rôle des processus stochastiques, de la sélection et de la migration dans l’évolution des espèces
Processus de spéciation
Processus d’extinction
Introduction
Les systèmes insulaires: des « laboratoires évolutifs »
Écosystèmes uniques
Forces évolutives particulières par rapport au continent

Introduction
Choix d’une espèce végétale de milieu insulaire : Santalum austrocaledonicum, espèce à forte valeur patrimoniale en Nouvelle-Calédonie, menacée après surexploitation.
Les systèmes insulaires: des « laboratoires évolutifs »
Écosystèmes uniques
Plantes insulaires : modèles pour aborder de nombreux problèmes en biologie évolutive
Forces évolutives particulières par rapport au continent
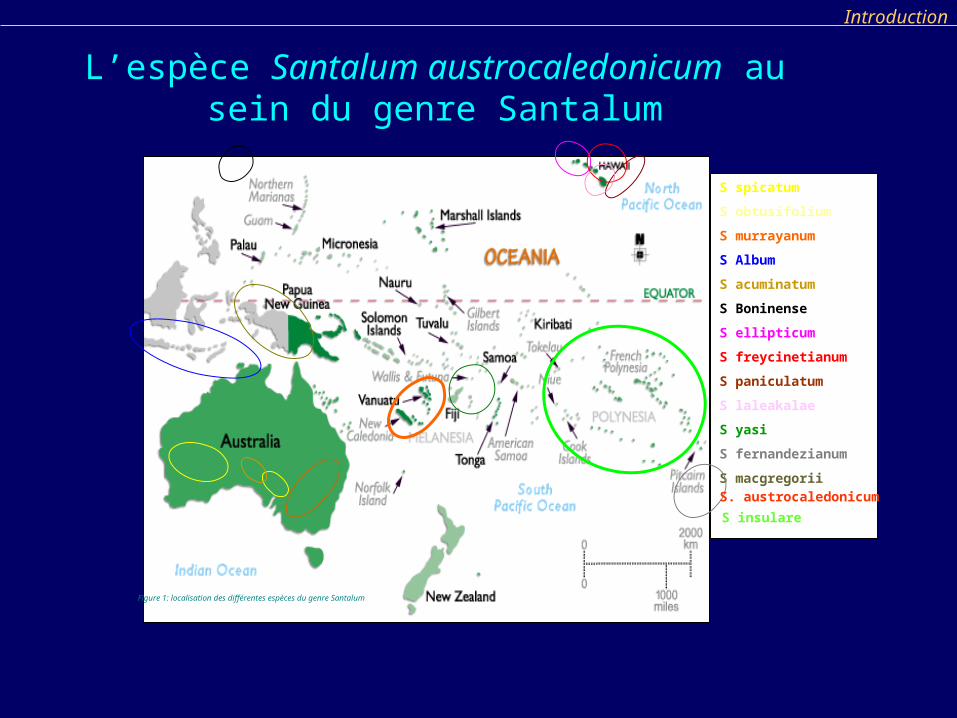
S spicatum
S obtusifolium
S murrayanum
S Album
S acuminatum
S Boninense
S ellipticum
S freycinetianum
S paniculatum
S laleakalae
S yasi
S fernandezianum
S macgregorii
Figure 1: localisation des différentes espèces du genre Santalum
S insulare
S. austrocaledonicum
L’espèce Santalum austrocaledonicum au sein du genre Santalum
Introduction

Osyris albaColpoon compressumS. acuminatum WA Australia S. acuminatum WA AustraliaS. acuminatum SA AustraliaS. acuminatum NSW AustraliaS. acuminatum NSWAustraliaS. acuminatum VIC AustraliaS. fernandezianum Juan FernandezS. spicatum SA AustraliaS. spicatum WA AustraliaS. spicatum WA AustraliaS. murrayanum VIC AustraliaS. murrayanum SA Australia S. lanceolatum NSW AustraliaS. lanceolatum QNS AustraliaS. lanceolatum SA Australia S. austrocaledonicum VanuatuS. lanceolatum NT AustraliaS. ellipticum Oahu S. ellipticum OahuS. ellipticum OahuS. ellipticum var. littorale OahuS. paniculatum Hawaii S. paniculatum HawaiiS. macgregorii PNGS. macgregorii PNGS. macgregorii PNG
S. insulare var. raivanse RaivaveS. insulare var. raiateense RaiateaS. insulare var. hendersonense HendersonS. insulare var. mitiaro CooksS. insulare HendersonS. insulare var. marchionense Marquesas S. insulare TahitiS. boninense BoninS. freycinetinaum var. pyrularium KauaiS. freycinetianum var. lanaiense MauiS. freycinetianum MolokaiS. freycinetianum OahuS. freycinetianum MolokaiS. haleakalae MauiS. obtusifolium NSW AustraliaS. austrocaledonicum New CaledoniaS. obtusifolium QNS AustraliaS. yasi FijiS. yasi TongaS. yasi TongaS. album NT AustraliaS. album India
100
100
66
98
63
97
949267
88
81
81
88
6366
91
92
98
88
95 67
74
8368
75
Australie
Hawaï
Juan FernandezVanuatu
PolynésieBoninNlle CalédonieFiji, Tonga Inde
L’espèce Santalum austrocaledonicum au sein du genre Santalum
Introduction
ADN chloroplastique (TrnK)
ADN ribosomal : séquences ITS et ETS de la zone 18S-26S
Phylogénie obtenue avec
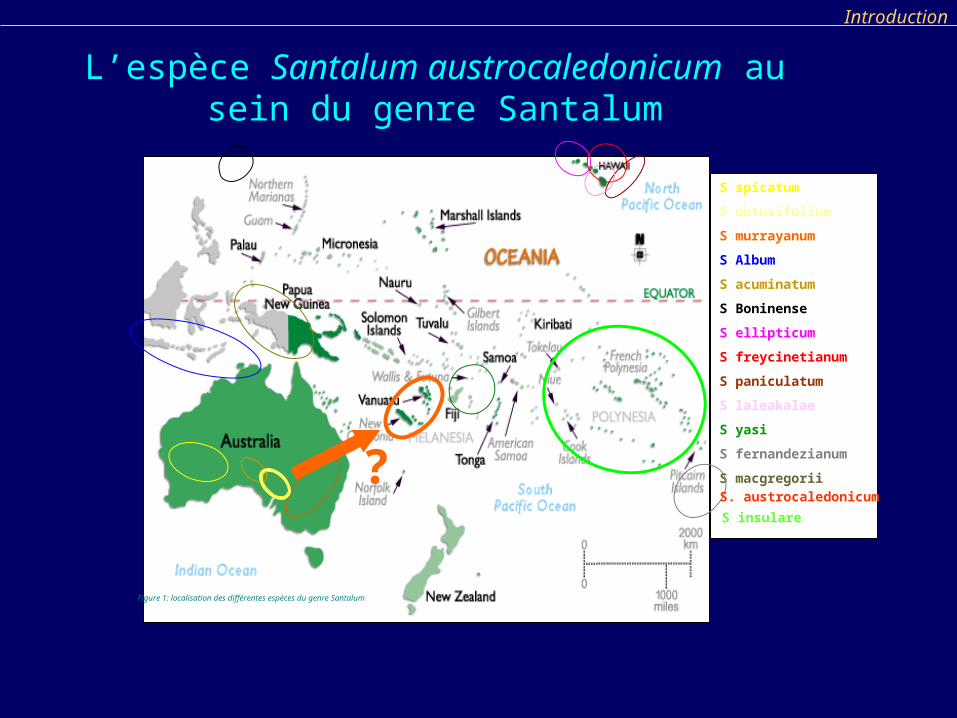
S spicatum
S obtusifolium
S murrayanum
S Album
S acuminatum
S Boninense
S ellipticum
S freycinetianum
S paniculatum
S laleakalae
S yasi
S fernandezianum
S macgregorii
Figure 1: localisation des différentes espèces du genre Santalum
S insulare
S. austrocaledonicum?
L’espèce Santalum austrocaledonicum au sein du genre Santalum
Introduction

Caractéristiques botaniques-Arbre, Angiospermes
-Taille moyenne : 6 à 8m, diamètre à 1,3m : 30 cm à maturité.
-Feuilles : individus juvéniles (50 x 6 mm) différents ind. adultes (50 x 30 mm).-Fleurs : blanches verdâtres, apétales. Plusieurs périodes de floraison.
-Fruit : drupe charnue (10 x 15 mm).
Feuilles d’individus juvéniles
L’espèce Santalum austrocaledonicum
Introduction

Caractéristiques botaniques
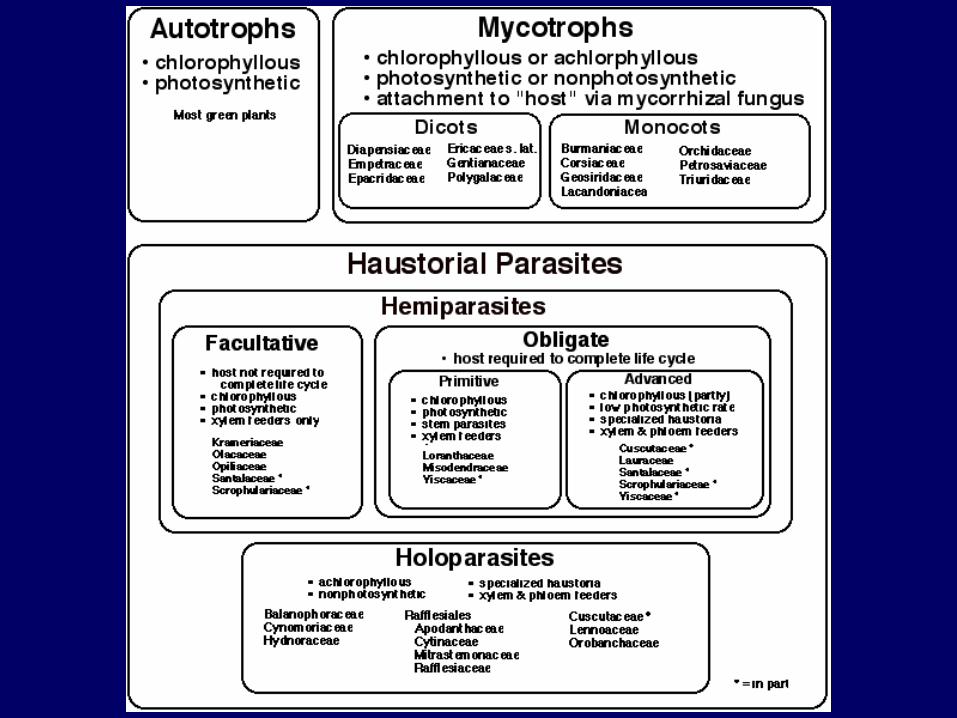
-Racines : munies d’haustoria (suçoirs) font du Santal un hémiparasite non spécifique
-Arbre, Angiospermes
-Taille moyenne : 6 à 8m, diamètre à 1,3m : 30 cm à maturité.
-Feuilles : individus juvéniles (50 x 6 mm) différents ind. adultes (50 x 30 mm).-Fleurs : blanches verdâtres, apétales. Plusieurs périodes de floraison.
-Fruit : drupe charnue (10 x 15 mm).
Introduction
L’espèce Santalum austrocaledonicum
Racine de Citrus reticulata parasitée par un Santal
Haustoria
Traces laissées par des
haustoria

Le « bois de Santal » = bois de cœur
Exploitable lorsque l’arbre a 30 ans
Utilisé en parfumerie et cosmétique (huile essentielle) et en artisanat (bois)
Santalum austrocaledonicum: surexploitée au 19ème siècle, toujours exploitée actuellement
1. La Nouvelle-Calédonie et l’espèce Santalum austrocaledonicum
L’espèce Santalum austrocaledonicum
Aubier
Bois de cœur

-Reproduction sexuée :
Entomophile, dispersion des graines par zoochorie (oiseaux)
A priori allogame et autogame: étude en cours
Reproduction
-Reproduction asexuée :
Rejet de souche après une coupe
Drageons
Introduction
L’espèce Santalum austrocaledonicum

Répartition dans l’archipel
Différence Grande Terre / îles Loyauté
Sur Grande Terre : petites populations reliques: une forme d’insularité
Îles Loyauté
Îles Loyauté
Île des Pins
Ouvéa
Lifou
Maré
Grande Terre<1 000
242 000
100 000
127 000
61 000
L’espèce Santalum austrocaledonicum
Introduction

Questions de recherche
Introduction
Comment s’établissent les flux de gènes au sein de l’archipel calédonien?
Existe t-il des variations phénotypiques de l’espèce au sein de l’archipel?
Si oui, quel est le déterminisme génétique de cette variation?
Cette variation est-elle adaptative?
Dans quelle mesure l’isolement des îles influence les phénomènes de dérive et migration?
Si oui, quelles sont les pressions de sélection spécifiques à l’écosystème calédonien?
Quels sont les patrons de diversité entre et à l’intérieur des populations?
Comment élaborer une stratégie de gestion de l’espèce dans un but de conservation?

caractères moléculaires neutres non soumis à la sélection
caractères a priori liés à l’adaptation
Introduction
Marqueurs microsatellites nucléaires et chloroplastiques
Taille des graines et des feuilles juvéniles, composition chimique du bois de cœur
Stratégie
Combiner l’analyse de différents caractères
Établir la relation entre cette variabilité et l’isolement des îles ou populations, leur petite taille impact de la dérive et de la
migration
les variables abiotiques (climat (pluviométrie, température), sol ) impact de la sélection

1. Diversité et structuration génétiques sur la base de marqueurs
microsatellites
Objectif: comprendre les phénomènes de dérive et de migration au sein de l’archipel

ÉchantillonnageSur l’ensemble de l’archipel, plusieurs campagnes de récolte.
Iles Loyauté : 240 Grande Terre : 306 Ile des Pins : 69 Vanuatu : 11
Total : 626 individus
Îles Loyauté
Déséquilibré Exhaustif sur Grande Terre (sauf Ouen Toro), non exhaustif sur Loyautés et île des Pins
Surfaces recouvertes différentes.
Nombre d’individus par population différent.
Grande Terre
Ouen Toro (22)
Païta (53)
Dumbéa (9) Mont Dore (2) Nouville (34) Vavouto (34)
Pindai (26)
Tiéai (10)
Oundjo (2)
Koumak (10)
Malhec (40)
Tangadiou (5)
Île des Pins (69)
Ouvéa (48)
Lifou (103)
Maré (89)
1. Diversité et structuration génétique sur la base de marqueurs microsatellites

300 m
10 km
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
ÉchantillonnageSur l’ensemble de l’archipel, plusieurs campagnes de récolte. Déséquilibré Exhaustif sur Grande Terre (sauf Ouen Toro), non exhaustif sur Loyautés et île des Pins
Surfaces recouvertes différentes.
Nombre d’individus par population différent.

Diversité et structuration ENTRE populations Diversité et structuration chloroplastique : 15 chlorotypes
Différence Grande Terre / îles Loyauté rupture très ancienne du flux de gènes.
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
Arbre des distances génétiques (chloroplastes)
Fst = 0.6
Malhec
Pindaï
Païta
Ouen Toro Ile des Pins
Maré
Lifou
Ouvéa Hienghène
Tiéa
Répartition des chlorotypes dans l’archipel
îles Loyauté
île des Pins
Grande Terre
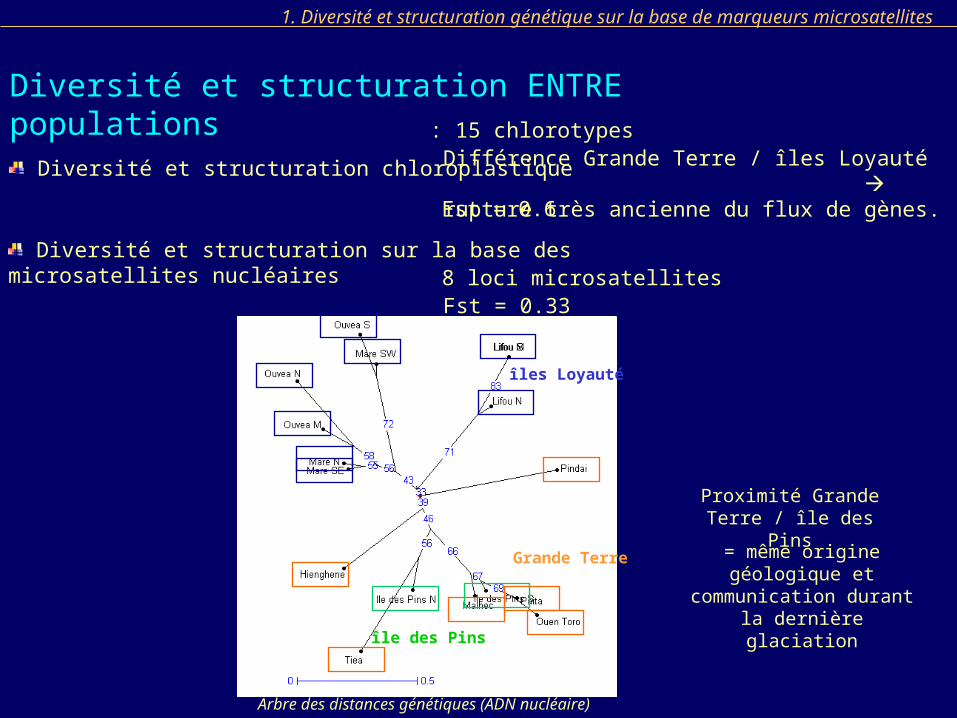
: 15 chlorotypesDifférence Grande Terre / îles Loyauté rupture très ancienne du flux de gènes.
Diversité et structuration sur la base des microsatellites nucléaires8 loci microsatellites
Fst = 0.6
Arbre des distances génétiques (ADN nucléaire)
Fst = 0.33
îles Loyauté
Grande Terre
île des Pins
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
Diversité et structuration ENTRE populations Diversité et structuration chloroplastique
Proximité Grande Terre / île des Pins
= même origine géologique et communication durant la
dernière glaciation

: 15 chlorotypesDifférence Grande Terre / îles Loyauté rupture très ancienne du flux de gènes.
Diversité et structuration sur la base des microsatellites nucléaires8 loci microsatellites
Fst = 0.6
Fst = 0.33
Fis = 0.17 ***
Fis = 0.0 ns
Fis = 0.25 ***
Fis = 0.62 ***
Fis = 0.14 *
Ile des Pins
Maré
Ouvéa
Lifou
Grande Terre Fis = 0.17 ***
Effet Wahlund
0.09***
0.03*** 0.00ns
-0.02ns
-0.05ns
-0.03 ns
Nombreux Fis positifs
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
Diversité et structuration ENTRE populations Diversité et structuration chloroplastique
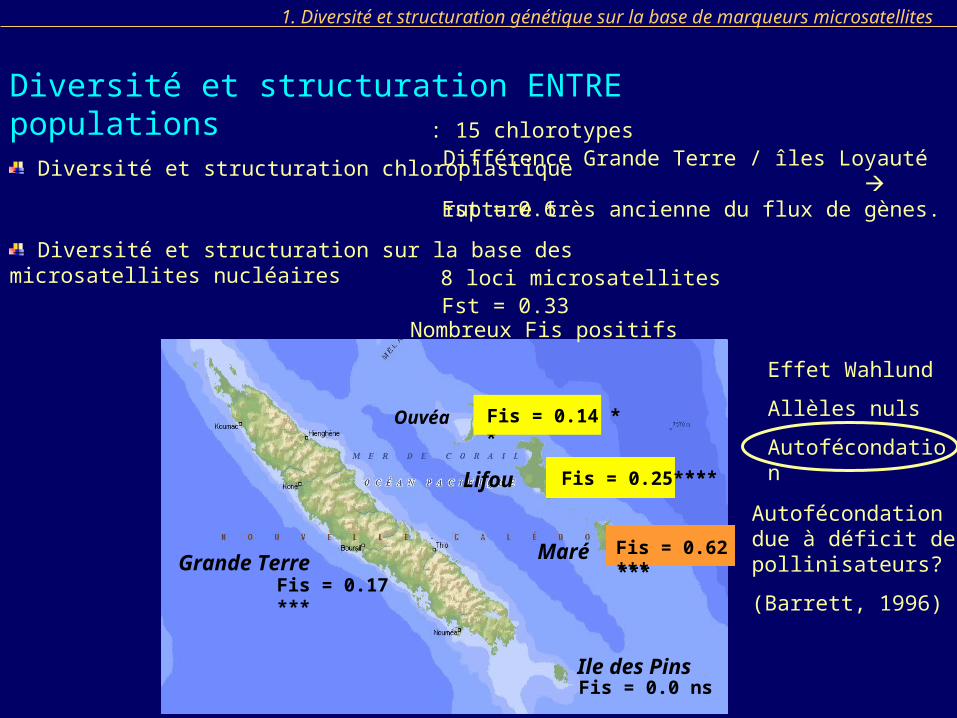
Fis = 0.17 ***
Fis = 0.0 ns
Fis = 0.25 ***
Fis = 0.62 ***
Fis = 0.14 *
Ile des Pins
Maré
Ouvéa
Lifou
Grande Terre
Fis = 0.14 *
Fis = 0.25***
Fis = 0.62 ***
Effet Wahlund
Allèles nuls
Autofécondation
Autofécondation due à déficit de pollinisateurs?
(Barrett, 1996)
: 15 chlorotypesDifférence Grande Terre / îles Loyauté rupture très ancienne du flux de gènes.
Diversité et structuration sur la base des microsatellites nucléaires8 loci microsatellites
Fst = 0.6
Fst = 0.33Nombreux Fis positifs
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
Diversité et structuration ENTRE populations Diversité et structuration chloroplastique

: 15 chlorotypesDifférence Grande Terre / îles Loyauté rupture très ancienne du flux de gènes.
Diversité et structuration sur la base des microsatellites nucléaires8 loci microsatellites
Fst = 0.6
Fst = 0.33Nombreux Fis positifs
Comparaison avec d’autres espèces
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
Diversité et structuration ENTRE populations Diversité et structuration chloroplastique
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
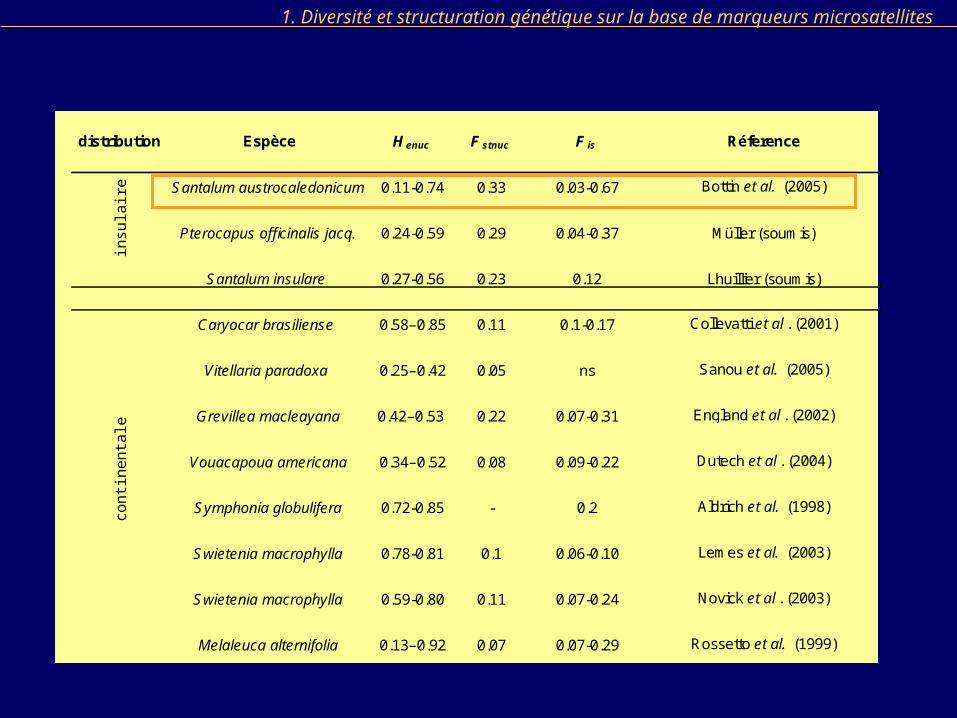
distribution Espèce H enuc F stnuc F is Réference
Santalum austrocaledonicum 0.11-0.74 0.33 0.03-0.67 Bottin et al. (2005)
Pterocapus officinalis jacq. 0.24-0.59 0.29 0.04-0.37 Müller (soumis)
Santalum insulare 0.27-0.56 0.23 0.12 Lhuillier (soumis)
Caryocar brasiliense 0.58–0.85 0.11 0.1-0.17 Collevatti et al . (2001)
Vitellaria paradoxa 0.25–0.42 0.05 ns Sanou et al. (2005)
Grevillea macleayana 0.42–0.53 0.22 0.07-0.31 England et al . (2002)
Vouacapoua americana 0.34–0.52 0.08 0.09-0.22 Dutech et al . (2004)
Symphonia globulifera 0.72-0.85 - 0.2 Aldrich et al. (1998)
Swietenia macrophylla 0.78-0.81 0.1 0.06-0.10 Lemes et al. (2003)
Swietenia macrophylla 0.59-0.80 0.11 0.07-0.24 Novick et al . (2003)
Melaleuca alternifolia 0.13–0.92 0.07 0.07-0.29 Rossetto et al. (1999)
con
tine
nta
lein
sula
ire
: 15 chlorotypesDifférence Grande Terre / îles Loyauté rupture très ancienne du flux de gènes.
Diversité et structuration sur la base des microsatellites nucléaires8 loci microsatellites
Fst = 0.6
Fst = 0.33Nombreux Fis positifs
Comparaison avec d’autres espèces
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
Diversité et structuration ENTRE populations Diversité et structuration chloroplastique
CONCLUSIONS : Un fort impact de la dérive, exacerbée par l’isolement des îles
Un régime de reproduction ayant évolué vers l’autogamie
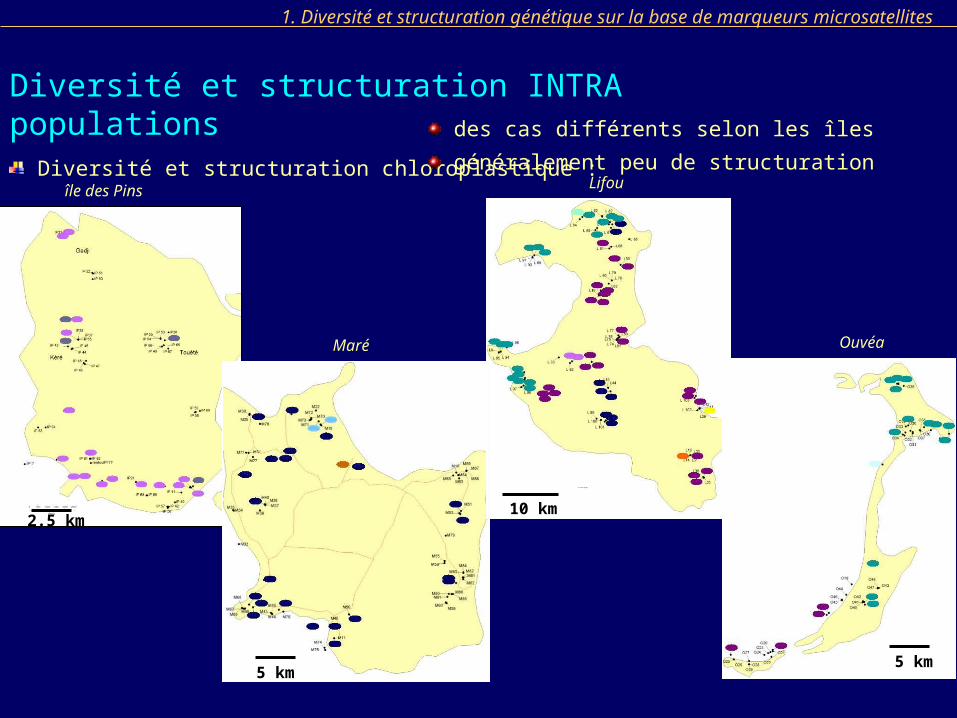
1. Diversité et structuration génétique sur la base de marqueurs microsatellites
Diversité et structuration INTRA populations Diversité et structuration chloroplastique : des cas différents selon les îles
généralement peu de structurationîle des Pins
Maré
Lifou
Ouvéa
2.5 km10 km
5 km5 km

1. Diversité et structuration génétique sur la base de marqueurs microsatellites
Diversité et structuration INTRA populations Diversité et structuration chloroplastique : des cas différents selon les îles
généralement peu de structuration
Structuration sur la base des microsatellites nucléaires :
CONCLUSION : Faible structuration intra population / intra îles Loyauté
Mantel absence d’isolement par la distance
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 500 1000 1500 2000 2500
distances géographiques
dis
tan
ces
gén
étiq
ues
Païta

2. Diversité des caractères phénotypiques
Objectif: identifier les déterminants (biotiques, abiotiques, génétiques) de la variation des graines, des feuilles juvéniles et de la composition chimique
du bois de cœur
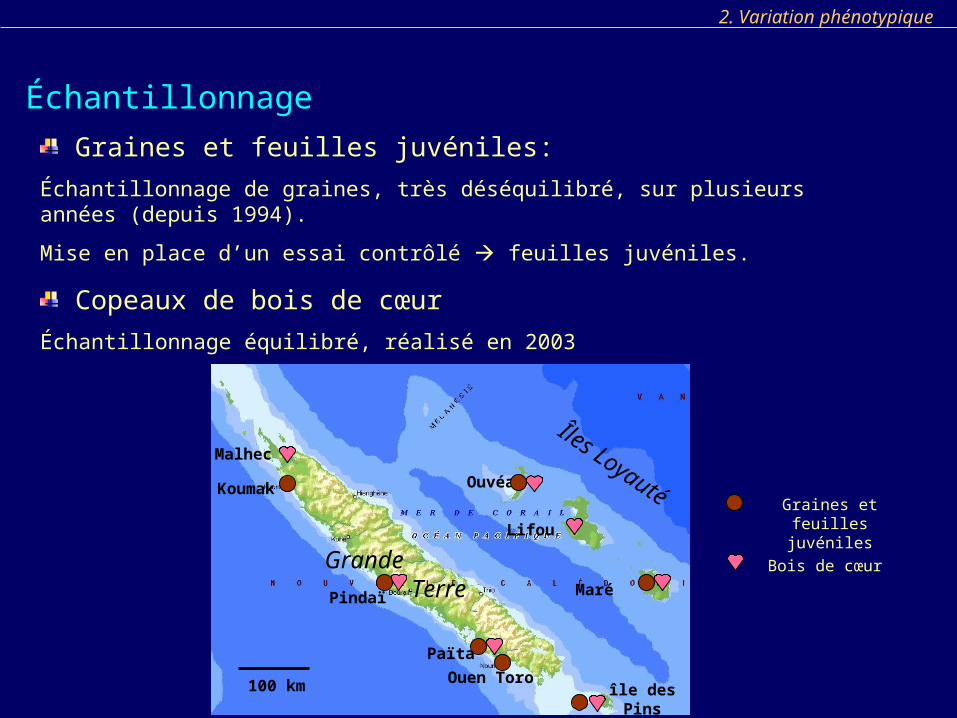
Échantillonnage
2. Variation phénotypique
Graines et feuilles juvéniles:
Échantillonnage de graines, très déséquilibré, sur plusieurs années (depuis 1994).
Mise en place d’un essai contrôlé feuilles juvéniles.
Copeaux de bois de cœur
Échantillonnage équilibré, réalisé en 2003
HienghèneGraines et feuilles
juvéniles
Îles Loyauté
Grande Terre
100 km
Lifou
Malhec
Bois de cœur
Ouen Toro
Ouvéa
île des Pins
Païta
Koumak
Pindaï Maré
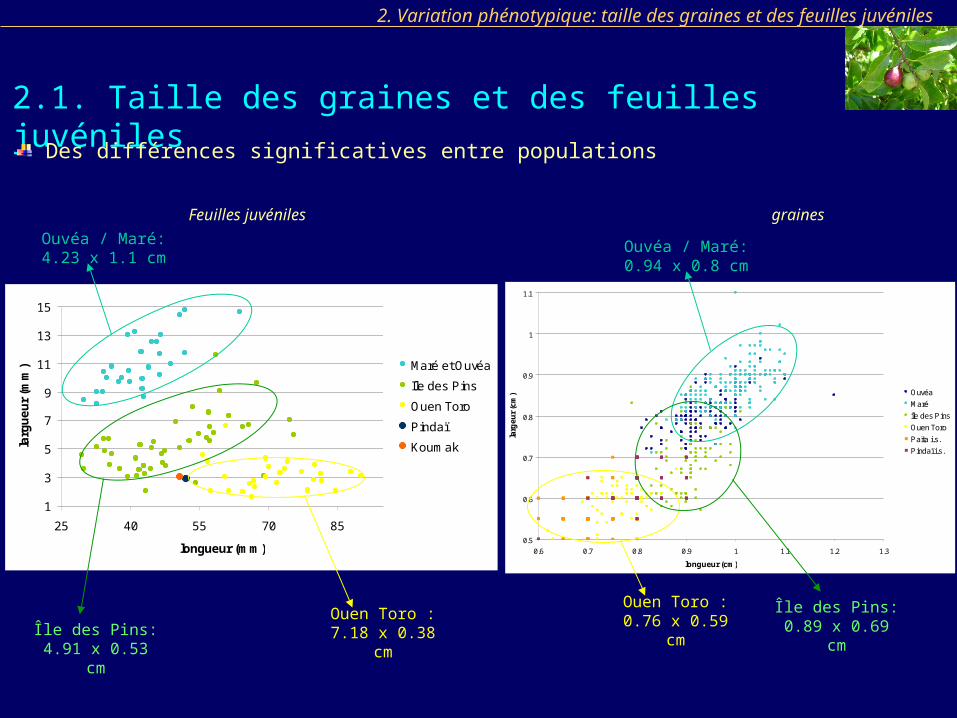
2.1. Taille des graines et des feuilles juvéniles
2. Variation phénotypique: taille des graines et des feuilles juvéniles
1
3
5
7
9
11
13
15
25 40 55 70 85
longueur (mm)
larg
ue
ur
(mm
) Maré et Ouvéa
Ile des Pins
Ouen Toro
Pindaï
Koumak
0.5
0.6
0.7
0.8
0.9
1
1.1
0.6 0.7 0.8 0.9 1 1.1 1.2 1.3
longueur (cm)
larg
eu
r (c
m) Ouvéa
Maré
île des Pins
Ouen Toro
Païta i.s.
Pindaï i.s.
Feuilles juvéniles graines
Des différences significatives entre populations
Ouen Toro : 7.18 x 0.38 cm
Ouvéa / Maré: 4.23 x 1.1 cm
Île des Pins: 4.91 x 0.53 cm
Ouvéa / Maré: 0.94 x 0.8 cm
Ouen Toro : 0.76 x 0.59 cm
Île des Pins: 0.89 x 0.69 cm

0
4
8
12
16
20
24
Dis
sim
ilar
ité
Dendrogramme
Ouv
éa
Mar
é
Paî
ta
Oue
ntor
o
Ile d
es P
ins
Pin
daï
Kou
mac
îles LoyautéGrande Terre
2.1. Taille des graines et des feuilles juvéniles
2. Variation phénotypique: taille des graines et des feuilles juvéniles
Des différences significatives entre populations
Ile des Pins
Maré
Ouvéa
Lifou Grande Terre

1500 mm/an2000 mm/an900 m
m/an
2. Variation phénotypique: taille des graines et des feuilles juvéniles
Déterminants de la variation de la taille des feuilles et des grainesUn déterminisme génétique = héritabilité de 42% à 85%
Effet de la dérive / faible migration? Structuration Grande Terre / Loyautés comme pour les microsatellites (marqueurs neutres) un effet possible
Effet de l’adaptation?
Pluviométrie? Grandes différences selon les localités. Corrélation significative et positive de la pluviométrie avec taille des graines et largeur des feuilles
Graines + grosses en zone humide car compétition
Feuilles + fines pour limiter la transpiration en zone sèche
Oui car Qst > Fst
Influence possible d’autres facteurs (non testés) :
Un déterminisme génétique = héritabilité de 42% à 85%
Déterminants de la variation de la taille des feuilles et des graines
Effet de la dérive / faible migration? Structuration Grande Terre / Loyautés comme pour les microsatellites (marqueurs neutres) un effet de dérive possible
Effet de l’adaptation? Oui car Qst > Fst
2. Variation phénotypique: taille des graines et des feuilles juvéniles
Pluviométrie? Grandes différences selon les localités. Corrélation significative et positive de la pluviométrie avec taille des graines et largeur des feuilles
Graines + grosses en zone humide car compétition
Feuilles + fines pour limiter la transpiration en zone sèche
altitude,
température,
sol (sols calcaires de mauvaise qualité sur Loyautés),
cortège d’oiseaux disperseurs,
plante hôte.
2.2. Composition chimique du bois de cœur
2. Variation phénotypique: composition chimique du bois de coeur
Le bois de cœur de Santal, zone productrice d’huiles essentielles
Essences très volatiles, non miscibles à l'eau et souvent parfumées.
Santal: Formation dans la zone de transition aubier / bois de cœur.
Huiles essentielles = composés secondaires des plantes: rôle défensif, et moteur évolutif.
Santalum austrocaledonicum: un rôle de résistance aux champignons et termites
Pression de sélection biotique?

Extraction au solvant (CHCl3) de la « concrète » : mêmes molécules que l’huile essentielle + autres molécules
Chromatographie en phase gazeuse
Analyse statistique des pics : l’aire d’un pic sur le chromatogramme est représentative de sa quantité dans la concrète pics exprimés en pourcentage d’aire de concrète
Etalon octanolEtalon octanol hydrocarbureshydrocarbures
Sesquiterpènes oxygénés = mono-alcoolsSesquiterpènes oxygénés = mono-alcools
diolsdiols
Huile essentielle
Concrète
2. Variation phénotypique: composition chimique du bois de coeur
Analyses chimiques
0
10
20
30
40
50
60
(+)-(Z)-a-santalol (30) (-)-(Z)-a-trans-bergamotol (34) (-)-(Z)-b-santalol (40) (E)-lanceol (50)
Hienghene
Malhec
Pindaï
Paita
Ile Des Pins
Lifou
Mare
Ouvea
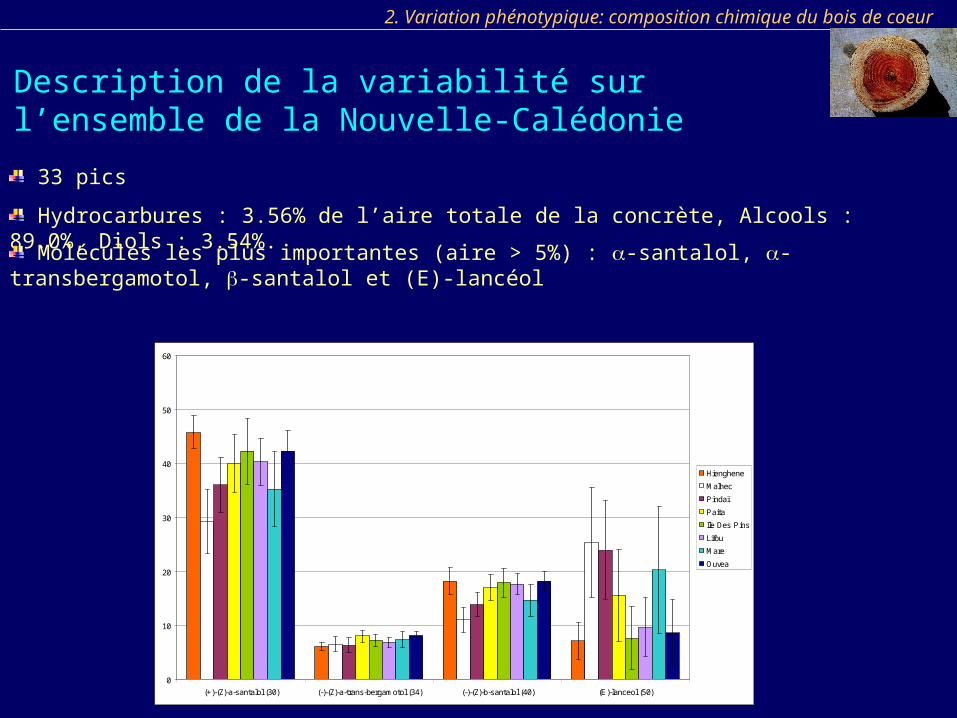
33 pics
Hydrocarbures : 3.56% de l’aire totale de la concrète, Alcools : 89.0%, Diols : 3.54%.
Molécules les plus importantes (aire > 5%) : -santalol, -transbergamotol, -santalol et (E)-lancéol
2. Variation phénotypique: composition chimique du bois de coeur
Description de la variabilité sur l’ensemble de la Nouvelle-Calédonie
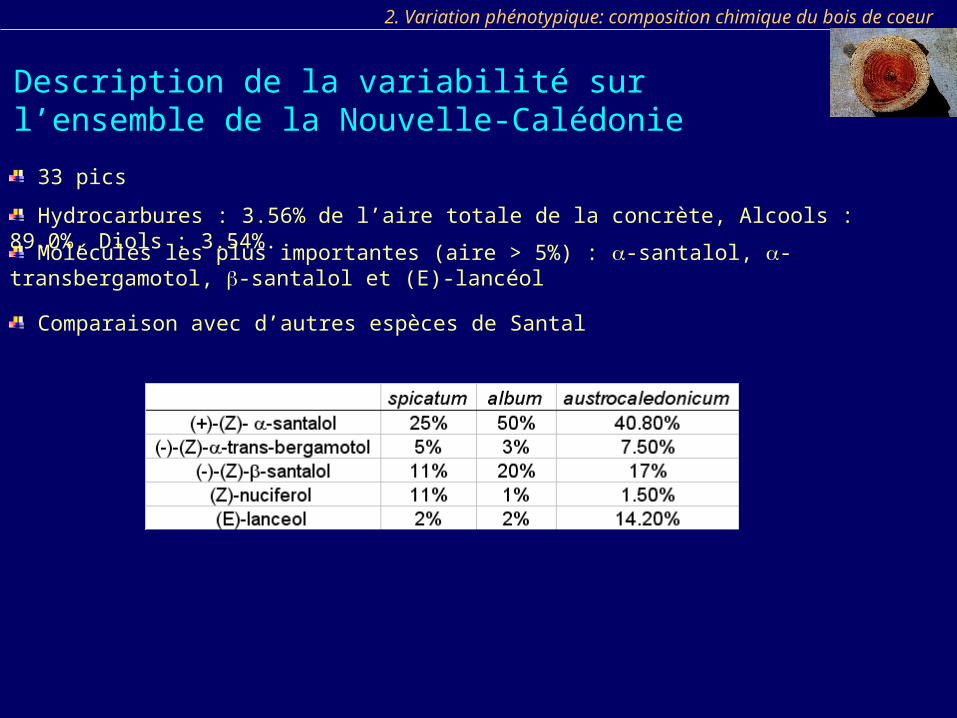
Comparaison avec d’autres espèces de Santal
Molécules les plus importantes (aire > 5%) : -santalol, -transbergamotol, -santalol et (E)-lancéol
2. Variation phénotypique: composition chimique du bois de coeur
Description de la variabilité sur l’ensemble de la Nouvelle-Calédonie
33 pics
Hydrocarbures : 3.56% de l’aire totale de la concrète, Alcools : 89.0%, Diols : 3.54%.

Individus (axes F1 et F2 : 59.30 %)
-5
0
5
10
-5 0 5 10
-- axe F1 (39.43 %) -->
-- a
xe F
2 (1
9.87
%)
-->
Hienghene
Ile Des Pins
Lifou
Malhec
Mare
Ouvea
Paita
Pindaï
Individus (axes F1 et F3 : 55.68 %)
-4
-2
0
2
4
6
-4 -2 0 2 4 6 8
-- axe F1 (39.43 %) -->
-- a
xe F
3 (1
6.25
%)
-->
Hienghene
Ile Des Pins
Lifou
Malhec
Mare
Ouvea
Paita
Pindaï
Variation entre populations par approche multivariée
Malhec
Pindaï
Malhec
Hienghène
Malhec: une faible teneur en-santalol et forte teneur en (E)-lancéol
Pindaï: forte teneur en molécule du pic 79
Hienghène: forte teneurs en dérivés de l’-santalène
2. Variation phénotypique: composition chimique du bois de coeur
Variation inter et intra-population par modèle à effets aléatoires
18.9% de la variance totale des pics est due aux différences entre populations.
-santalol : 19.68%, -trans-bergamotol : 9.06 %, -santalol : 22.52%, (E)-lanceol : 13.26%.

Variation entre groupes d’îles par approche multivariée
Individus (axes F1 et F2 : 100.00 %)
-3
-1
1
3
5
-5 -3 -1 1 3
-- axe F1 (77.72 %) -->
-- a
xe F
2 (2
2.28
%)
-->
Grande Terre
Ile Des Pins
LoyautésDifférences significatives
entre Grande Terre, île des Pins et Loyautés
2. Variation phénotypique: composition chimique du bois de coeur

Déterminants de la variation chimique
Pluviométrie: non corrélée à 31 pics sur 33.
Sol: Malhec (sol fersiallitique) : effet différent sur la composition chimique. Influence du sol ou autre facteur?
Effet de la dérive et de l’isolementSi même effet sur les huiles et les microsatellites structuration similaire des populations.arbre des distances génétiques (microsatellites nucléaires) arbre des distances chimiques
Arbres de distance différents: dérive et isolement ont moins d’effet que l’adaptation
2. Variation phénotypique: composition chimique du bois de coeur
Effet des facteurs abiotiques

Adaptation à des cortèges d’insectes particuliers selon les localités à tester
Effet des plantes parasitées: nature et quantité d’espèces parasitées à tester
2. Variation phénotypique: composition chimique du bois de coeur
Effet des facteurs biotiques

Conclusions principales

Une structuration génétique marquée (Fstnucl = 0.33, Fstchl = 0.66), particulièrement entre les îles Loyauté et Grande Terre / île des Pins.
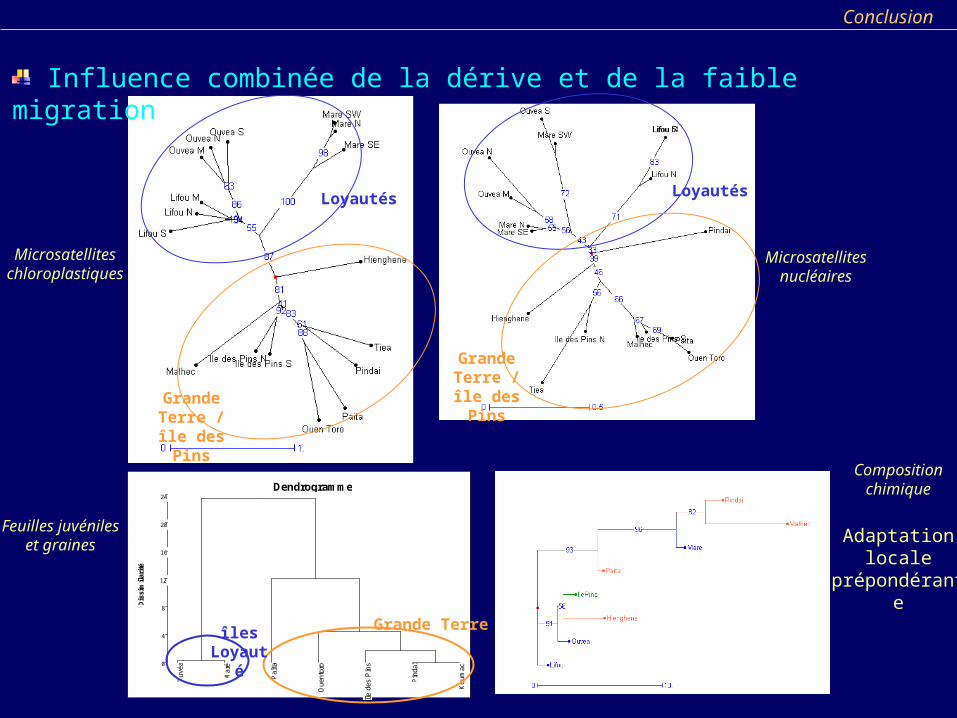
Influence combinée de la dérive et de la faible migration
Microsatellites chloroplastiques
Loyautés Loyautés
Grande Terre / île des Pins
Grande Terre / île des Pins
Microsatellites nucléaires
Conclusion
Causes :
Dérive = effets de fondation (îles Loyauté)
petites populations (Grande Terre)
Migration faible due à l’isolement des îles et des populations de Grande Terre

Loyautés Loyautés
Grande Terre / île des Pins
Grande Terre / île des Pins
Microsatellites chloroplastiques
Microsatellites nucléaires
0
4
8
12
16
20
24
Dis
sim
ilar
ité
Dendrogramme
Ouv
éa
Mar
é
Paî
ta
Oue
ntor
o
Ile d
es P
ins
Pin
daï
Kou
mac
îles Loyauté
Grande Terre
Feuilles juvéniles et graines Un effet de la dérive et l’isolement possible
Mais ces facteurs n’agissent pas seuls: Qst > Fst un effet de la sélection
Conclusion
Influence combinée de la dérive et de la faible migration
Loyautés Loyautés
Grande Terre / île des Pins
Grande Terre / île des Pins
0
4
8
12
16
20
24
Dis
sim
ilar
ité
Dendrogramme
Ouv
éa
Mar
é
Paî
ta
Oue
ntor
o
Ile d
es P
ins
Pin
daï
Kou
mac
îles Loyauté
Grande Terre
Microsatellites chloroplastiques
Microsatellites nucléaires
Feuilles juvéniles et graines
Composition chimique
Adaptation locale
prépondérante
Conclusion
Influence combinée de la dérive et de la faible migration
Taille des graines et des feuilles juvéniles : adaptation à la pluviométrie, au type de sol et au espèces d’oiseaux disperseurs présentes.
Influence de l’adaptation locale
Composition chimique : pas d’adaptation aux facteurs abiotiques, adaptation au cortège d’insectes, et peut-être plante hôte.
Mais échantillonnage lacunaire et peu précis : résultats à prendre avec précaution.
Conclusion

Conclusion
Une évolution caractéristique des espèces insulaires
Faible taille des populationsdérive génétique
Faible nombre d’individus fondateurs
Statut isolé Migration réduite
Sélection naturelle particulière (hypothèses probables)
diminution / rupture du flux de gènes
Forte structuration des populations
Diminution des disperseurs variation de taille des graines
Diminution des pollinisateurs autogamie
Particularité des sols insulaires taille des graines
Particularité des communautés végétales plantes hôtes huiles, graines, feuilles
Particularité des insectes et champignons ravageurs du bois huiles
Application à la conservation de l’espèce
Pourquoi conserver Santalum austrocaledonicum ?
Forte exploitation au 19ème siècle : diminution drastique des populations, particulièrement à Grande Terre
Mise en place de quotas (1988), basés uniquement sur des résultats démographiques
Nécessité de prise en compte de la génétique et de l’adaptation au milieu
Méthode de Crandall (2000) : combiner des données moléculaires et adaptatives à différents niveaux temporels pour établir des unités de conservation.
Recommandation de gestion: Grande Terre / Loyauté : ne pas permettre le flux de gènes
Résultats à appliquer sur le terrain mesures de gestion concrètes
Conclusion

Perspectives de recherche
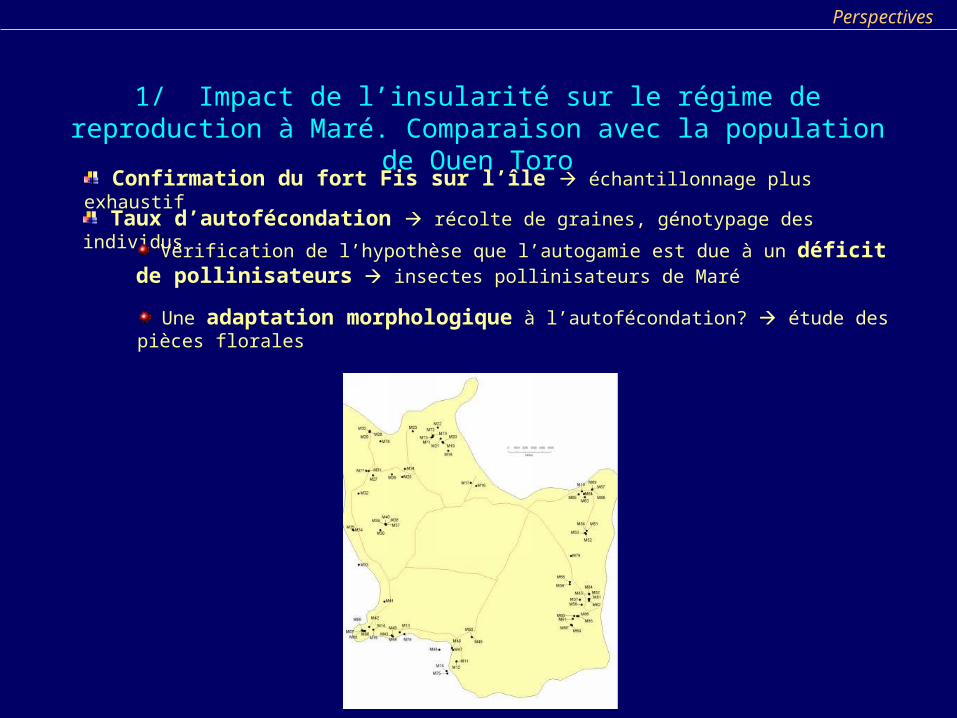
Confirmation du fort Fis sur l’île échantillonnage plus exhaustif
1/ Impact de l’insularité sur le régime de reproduction à Maré. Comparaison avec la population de Ouen Toro
Taux d’autofécondation récolte de graines, génotypage des individus
Une adaptation morphologique à l’autofécondation? étude des pièces florales
Vérification de l’hypothèse que l’autogamie est due à un déficit de pollinisateurs insectes pollinisateurs de Maré
Perspectives
Meilleure connaissance de la phénologie de Santalum austrocaledonicum
Suivi phénologique d’une 50aine de Santals
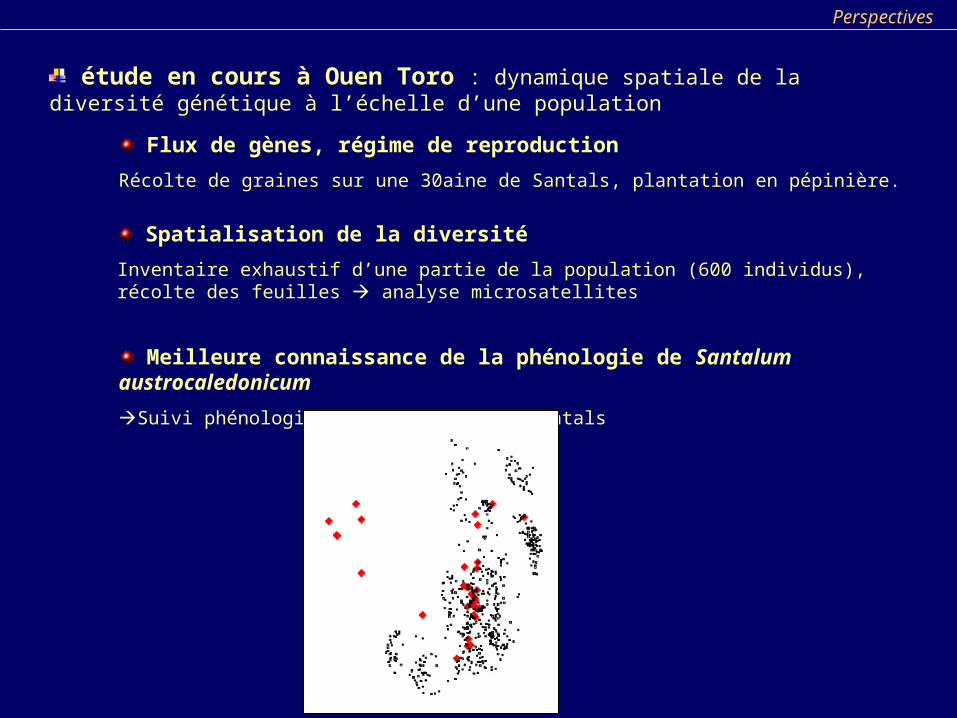
Spatialisation de la diversité
Inventaire exhaustif d’une partie de la population (600 individus), récolte des feuilles analyse microsatellites
Flux de gènes, régime de reproduction
Récolte de graines sur une 30aine de Santals, plantation en pépinière.
étude en cours à Ouen Toro : dynamique spatiale de la diversité génétique à l’échelle d’une population
Perspectives

Un échantillonnage plus exhaustif, avec une meilleure traçabilité des provenances.
Mesure d’autres traits adaptatifs et traits liés à l’histoire de vie
2/ Déterminants de la variabilité des caractères adaptatifs
Perspectives
Vitesse de croissance des semis
Production de graines
Mesure des traits le long d’un gradient (ex: pluviométrie)
Mise en place d’un essai plus rigoureux, éventuellement multi-sites.

3/ Étude spatiale de la dynamique intra-île
Hétérogénéité du milieu Dynamique des populations différentes selon les localités
Bonne régénération dans les zones de jachère (groupe 1)
Régénération faible dans les zones récemment fermées (groupe 2)
Régénération faible et nombreux arbres de gros diamètres fermeture du milieu très ancienne (groupe 3)
Objectif: évaluer la structure de la diversité génétique selon les caractéristiques démographiques des populations.
Microsatellites sur l’une des îles Loyauté
Perspectives
0-1010-2020-3030-4040-5050-60
De
nsi
té N
/ha
00
20
40
60
80
Groupes démographiques sur Maré
1 2 3


Et des « merci » particuliers…
Aux membres du jury
Hery Emeline Haby SophieMireille
RoselyneAlex
Et Jean-Marc BOUVET
Liva
À toute l’équipe du CIRAD-forêt
Au DEA BEE 2001… un
grand crû!
À Sophie, Fred, Thomas, Guillaume,Cédric et Matthieu
À Pascal ;)
À Manon et Sylvain
Et un trèèès grand merci à mes parents!
Daniel VERHAGEN
Frédéric MORTIER
Jean-Marc GION
Olivier MONTEUUIS
Robert NASI
Au personnel de Nouvelle-Calédonie
Jacques TASSIN
Alexandre LAGRANGE
Géraldine DERROIRE
Camille ISNARD
Colin GODEFFROY
Au laboratoire de chimie des substances naturelles
de Tahiti
Jean François BUTAUD
Phila RAHARIVELOMANANA
Jean-Pierre BIANCHINI
Jean-Christophe GLASZMANN
Isabelle OLIVIERI
Frédéric AUSTERLITZ
Frédéric HOSPITAL
Claire BILLOT
À toutes les personnes qui m’ont aidé au cours de ma thèse
Mais aussi…


Conclusion
Forces évolutives
Caractères étudiés
Marqueurs neutres Marqueurs potentiellement adaptatifs
Microsat. nucléaires
Microsat. chlorop.
Taille des graines
Taille des feuilles juv.
Composition chimique.
Dérive (effet fondation, petites tailles pop)
Migration
Sélection
Pluviométrie
Sol
Altitude
Température
Oiseaux disperseurs
Insectes phytophages
Plante hôte
??
??
??
??
? ? ?
??
? ?