
Situations de challenge m...
22
- Le jeûne - La naissance - L’exercice musculaire Situations de challenge métabolique Substrats énergétiques Glucose Utilisable par tous les tissus Certains tissus n’utilisent que le glucose (cerveau) : 120 g/jour Acides gras Utilisation par le foie, les muscles, le coeur Le cerveau n ’utilise pas d ’acides gras (barrière hémato-encéphalique) Corps cétoniques Composés hydrophiles formés lors de l ’oxydation des acides gras et pouvant être utilisés par le cerveau
Transcript of Situations de challenge m...

- Le jeûne
- La naissance
- L’exercice musculaire
Situations de challenge métabolique
Substrats énergétiques
Glucose
Utilisable par tous les tissus
Certains tissus n’utilisent que le glucose (cerveau) : 120 g/jour
Acides gras
Utilisation par le foie, les muscles, le coeur
Le cerveau n!’utilise pas d!’acides gras (barrière hémato-encéphalique)
Corps cétoniques
Composés hydrophiles formés lors de l!’oxydation des acides gras et pouvant être utilisés par le cerveau

PROBLEME CENTRAL
Maintenir un apport de glucose constant aux tissus
dépendants de ce substrat
Réserves énergétiques de l!’organisme
Carbohydrates
Lipides
Protéines
Quantité pour une personne
de 65 kg
Equivalent
énergétique
Réserves
Glucose libre
Glycogène
Triglycérides
12 g
450 g
15 kg
12,5 kg *
0,2 MJ
550 MJ
210 MJ
7,65 MJ
30 min
18 h
55 j
21 j

Les différentes phases du jeûne
1- Période post-absorptivecommence quelques heures après la consommation d’un repas lorsque le contenuintestinal a été absorbé
2- Jeûne courtSuccède à la période post-absorptiveJeûne de 1 à 3 jours chez l!’homme
3- Jeûne intermédiaireJeûne de 3 jours à 3 semaines chez l!’homme
4- Jeûne prolongéAu delà de 3 semaines
Les modèles expérimentaux
Modèles humains :
• Jeûne total : données expérimentales obtenues après quelques jours de jeûne
• Grévistes de la faim Irlandais : 60/70 jours de jeûne
• Quelques données chez l!’obèse : 200 à 300 jours de jeûne
Modèles animaux
• manchot royal
• Oiseaux migrateurs
• Rongeurs (rat)
Les différentes phases du jeûne
1- Période post-absorptivecommence quelques heures après la consommation d’un repas lorsque le contenuintestinal a été absorbé
2- Jeûne courtSuccède à la période post-absorptiveJeûne de 1 à 3 jours chez l!’homme
3- Jeûne intermédiaireJeûne de 3 jours à 3 semaines chez l!’homme
4- Jeûne prolongéAu delà de 3 semaines
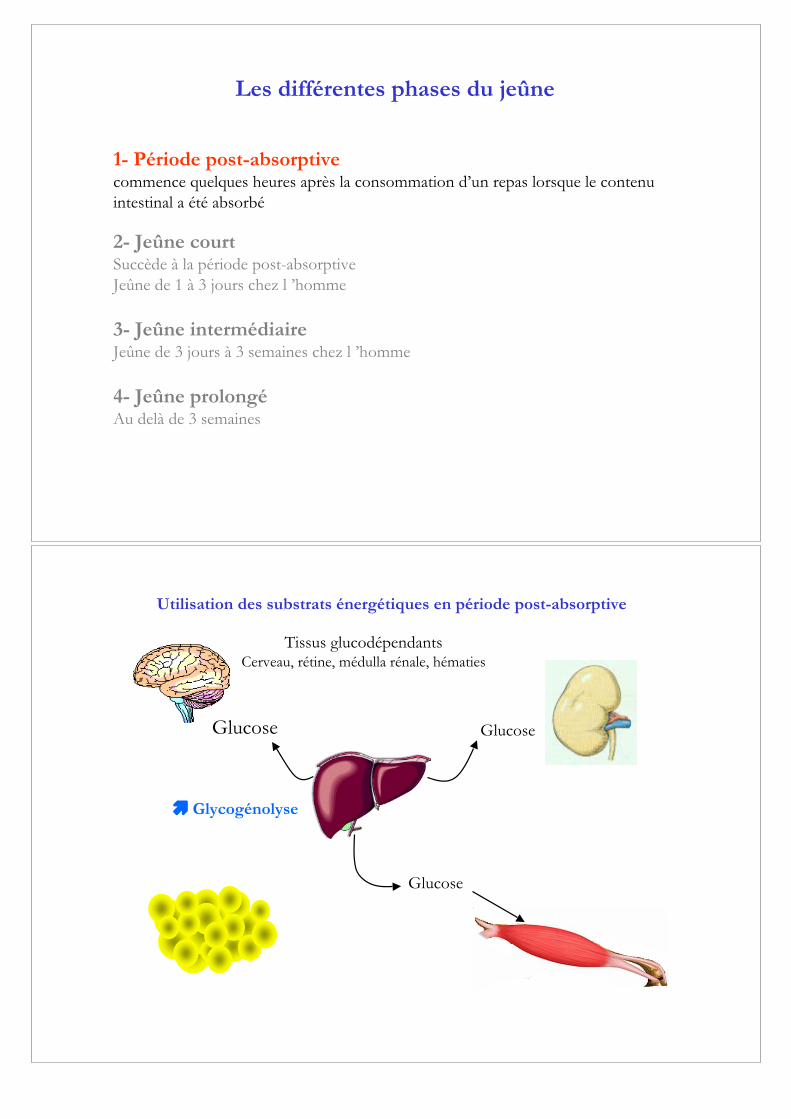
Utilisation des substrats énergétiques en période post-absorptive
Glucose
Tissus glucodépendantsCerveau, rétine, médulla rénale, hématies
Glucose
Glucose
! Glycogénolyse
Glucose
Utilisation des substrats énergétiques en période post-absorptive
Glucose
Tissus glucodépendantsCerveau, rétine, médulla rénale, hématies
Glucose
! Glycogénolyse
! Néoglucogenèse
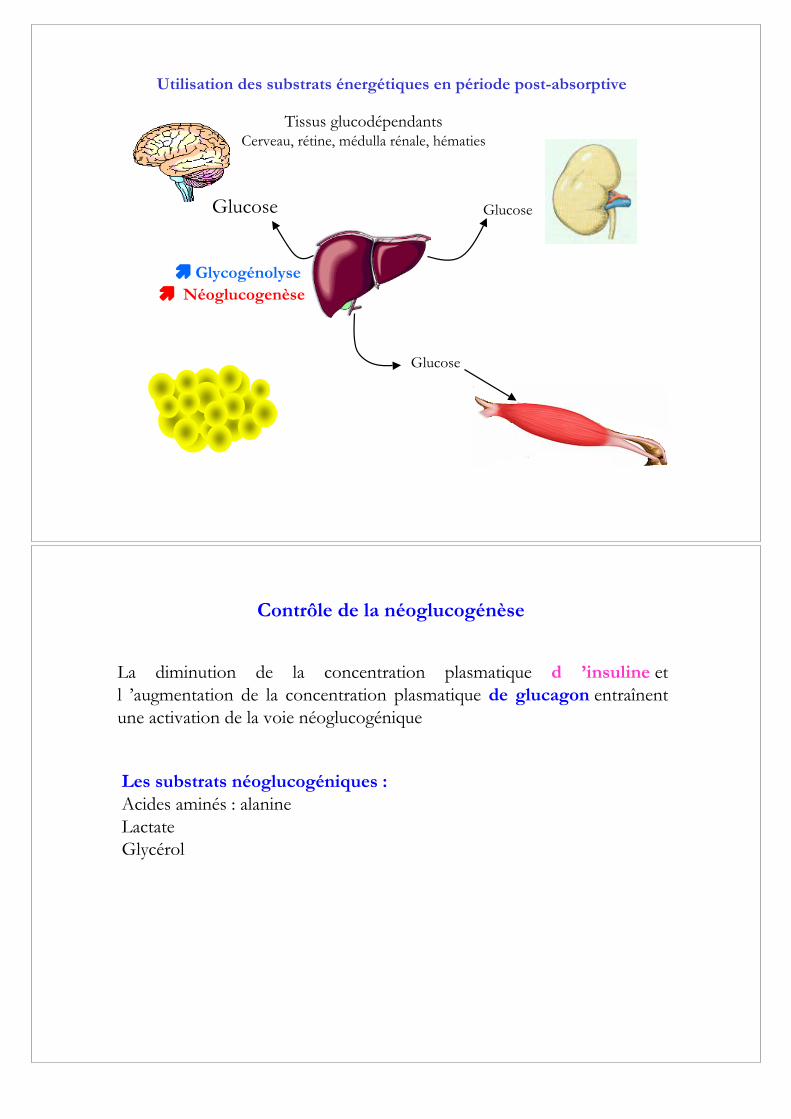
Contrôle de la néoglucogénèse
La diminution de la concentration plasmatique d! ’insuline etl! ’augmentation de la concentration plasmatique de glucagon entraînentune activation de la voie néoglucogénique
Les substrats néoglucogéniques :
Acides aminés : alanineLactateGlycérol

Les différentes phases du jeûne
1- Période post-absorptivecommence quelques heures après la consommation d’un repas lorsque le contenuintestinal a été absorbé
2- Jeûne courtSuccède à la période post-absorptiveJeûne de 1 à 3 jours chez l!’homme
3- Jeûne intermédiaireJeûne de 3 jours à 3 semaines chez l!’homme
4- Jeûne prolongéAu delà de 3 semaines
1- Intensification de la néoglucogenèse
A partir principalement d!’acides aminés dits glucoformateurs : alanine, glutamine
Jeûne court
2- Protéolyse musculaire intense
Due à la chute des concentrations plasmatiques d!’insuline
exemple : 1,75 g de protéines donnent 1 g de glucose. 210 g de protéines pour le fonctionnement du cerveau en une journée
besoin d!’adaptations pour économiser les protéines musculaires

1- Utilisation de substrats de remplacement : corps cétoniques
en particulier par le cerveau, acides gras. Conséquence : La production hépatique de glucose décroit
Adaptations métaboliques mises en place pour économiser
la protéolyse musculaire
3- Utilisation d!’autres précurseurs pour la néoglucogenèse : lactate
2- la lipolyse augmente : le glycérol devient un important substrat glucogénique
4- Les corps cétoniques pourraient s!’opposer à la protéolyse
musculaire
Utilisation des substrats énergétiques lors de l!’adaptation au jeûne
! NéoglucogenèseGlucose
Tissus gluco-dépendantsCerveau, rétine, médulla rénale, hématies
TG
Acides gras
! Lipolyse

Lors d! ’une période de jeûne, on observe une chute de l! ’insulinémie et une augmentation des concentrations
locales de catécholamines dans le tissu adipeux.
AGAGAG
TG
Gouttelette
de triglycérides
Récepteur aux catécholamines
Phosphodiestérase
AMP
Récepteur à l!’insuline
+
ATP AMPc
Adénylate cyclase
Protéine kinase A
Régulation de la lipolyse
LHS
Périlipine
P
P
PP
Utilisation des substrats énergétiques lors d’un jeûne court
NéoglucogenèseGlucose
Tissus gluco-dépendantsCerveau, rétine, médulla rénale, hématies
TG
Acides gras ! Oxydation
des AG
! Oxydation
des AG
Lipolyse
! Oxydation
des AG : cétogenèse
Utilisation des substrats énergétiques lors d’un jeûne court
Néoglucogenèse
Glucose
Tissus gluco-dépendantsCerveau, rétine, médulla rénale, hématies
TG
Fatty acids
Fatty acids " CO2
! Oxydation
des AG
Lipolyse
! Cétogénèse(Corps cétoniques)
Corps cétoniques
Corps cétoniques " CO2
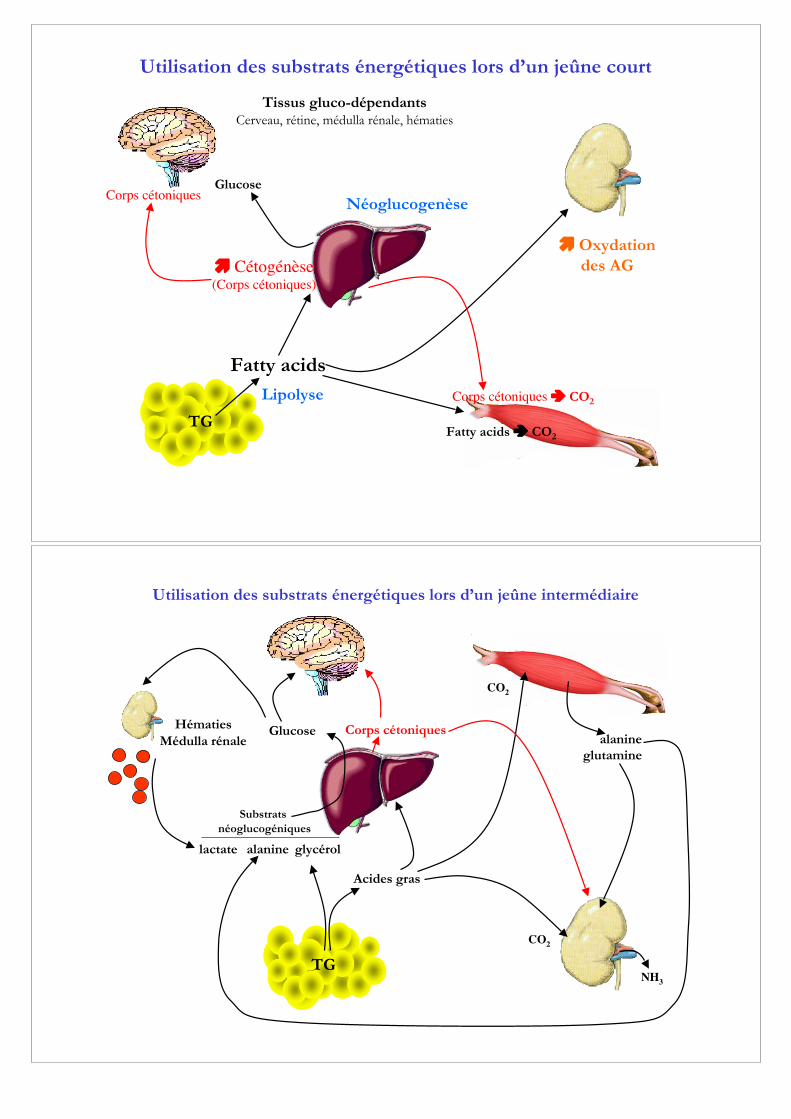
Utilisation des substrats énergétiques lors d’un jeûne intermédiaire
TG
Acides gras
CO2
CO2
alanineglutamine
NH3
glycérolalaninelactate
Hématies
Médulla rénaleCorps cétoniques
Substrats
néoglucogéniques
Glucose

Les différentes phases du jeûne
1- Période post-absorptivecommence quelques heures après la consommation d’un repas lorsque le
contenu intestinal a été absorbé
2- Jeûne courtSuccède à la période post-absorptive
Jeûne de 1 à 3 jours chez l!’homme
3- Jeûne intermédiaireJeûne de 3 jours à 3 semaines chez l!’homme
4- Jeûne prolongéAu delà de 3 semaines
• étudié chez l!’animal
• jeûne intermédiaire se caractérise par :Réduction de la glycémieÉlévation des concentrations plasmatiques d!’acides gras et de corps cétoniques circulantsBaisse du catabolisme protéique qui se maintient à un niveau faible
• jeûne prolongé se caractérise par :Augmentation de la concentration plasmatique de glucoseDiminution des concentrations d!’acides gras et corps cétoniques circulantsAugmentation du catabolisme protéique
• reprise de l!’activité motrice
Jeûne prolongé

Une situation exceptionnelle : la naissance
Nourri continuellement par lecordon ombilical :Utilise essentiellement du glucose(métabolisme oxydatif).
La fourniture de substrat cessebrutalement.Le nouveau-né est confronté àune période de jeûne glucidiqueplus ou moins longue suivie d ’unrégime plutôt riche en graisse.
BESOINS EN GLUCOSEELEVÉS :
(cerveau: 12% du poids ducorps versus 2% chez l’adulte)
Glucides LipidesProtéines
« Régime »foetal
Composition du lait
Femme Brebis Truie Ratte
% d
es c
alor
ies
tota
les
0
0,5
1G
lycé
mie
(g/l
)
Heures après la naissance
0 4 8 12 16
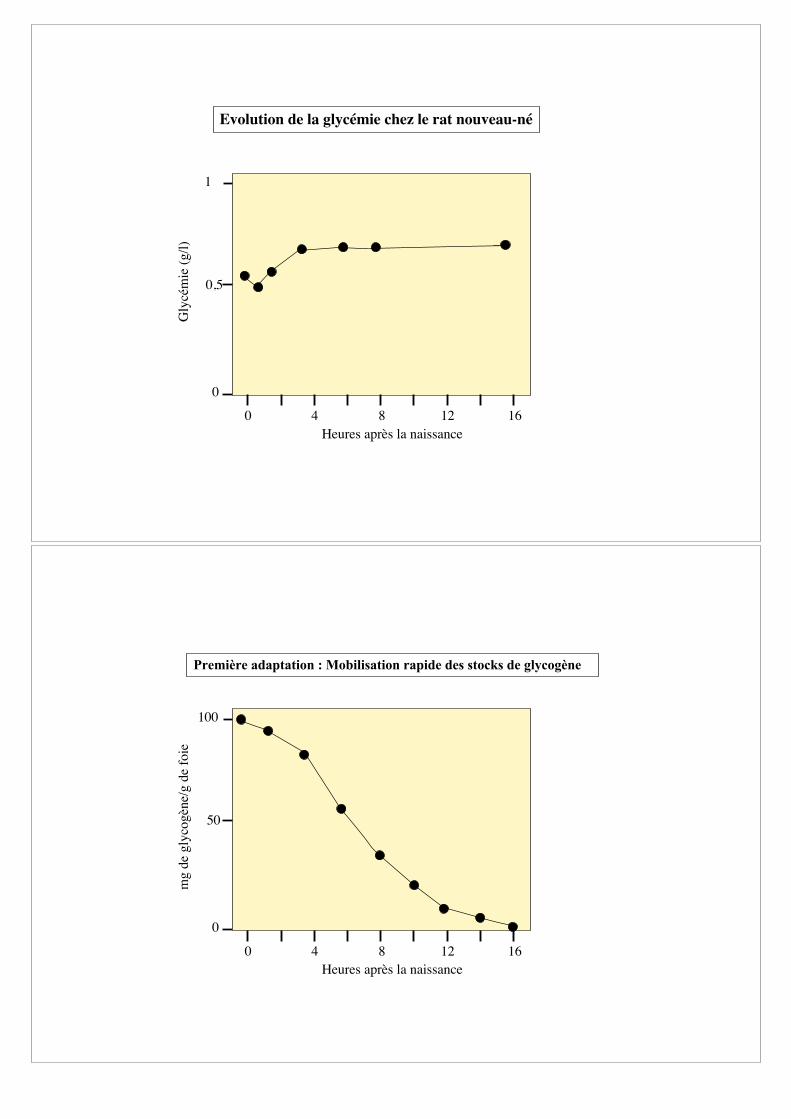
Evolution de la glycémie chez le rat nouveau-né
Première adaptation : Mobilisation rapide des stocks de glycogène
0
50
100
mg d
e gly
cogèn
e/g d
e fo
ie
Heures après la naissance
0 4 8 12 16

0
50
100
Heures après la naissance
0 4 8 12 16
Deuxième adaptation : Mise en route de la gluconéogénèse
Cap
acit
é g
luco
néo
gén
ique
du f
oie
(unit
és a
rbit
rair
es)
Act
ivit
é de
la P
EP
CK
(unit
és a
rbit
rair
es)
0
1,5
3
Heures après la naissance
0 4 8 12 16
Troisième adaptation : Mise en route de la beta-oxydationhépatique et de la production de corps cétoniques
Corp
s cé
toniq
ues
pla
smar
iques
(mm
ol/
l)
0
0,5
1
Aci
des
gra
s pla
smat
iques
(mol/
l)

0
1
2
0 2 4 6
Effets d’un blocage de l’oxydation hépatique des acides graschez des rats nouveau-nés allaités (I).
Corp
s cé
ton
iqu
es p
lasm
atiq
ues
(mm
ol/
l) Allaités
Allaités+ McN 3716
McN 3716 McN 3716 + TCM
Allaités+ McN 3716
+ TCM
0
2
4
0 2 4 6
Effets d’un blocage de l’oxydation hépatique des acides graschez des rats nouveau-nés allaités (II).
Gly
cém
ie (
mm
ol/
l)
Allaités
Allaités+ McN 3716
McN 3716 McN 3716 + TCM
Allaités+ McN 3716
+ TCM

0
2
4
0 2 4 6
Effets d’un blocage de l’oxydation hépatique des acides graschez des rats nouveau-nés allaités (III).
Glu
con
éogén
èse
(mm
ol/
l) Allaités
Allaités+ McN 3716
McN 3716 McN 3716 + TCM
Allaités+ McN 3716
+ TCM
Nouveau-né humain de 24 heures, 6h après la dernière tétée
Acides gras Corps cétoniques Glycémie
Vitesse de
renouvellement
(µmol/min/kg)
0.95 ± 0.1
0.34
1.25 ± 0.2
0.15
4 ± 0.06
4.2
15 ± 1
5
5.2 ± 0.5
1
27 ± 2
11
Concentration
(mmol/l)
Valeurs chez un adulte 48h à jeun

Quels sont les facteurs responsables de ces variations?
0
75
150
Heures après la naissance
0 4 8 12 16
Insu
line
pla
smat
ique
(µU
/ml)
0
0,5
1
Glu
cagon p
lasm
atiq
ue
(ng/m
ll)
Stress fœtal (hypoxie) ==> adrénaline + noradrénaline ==> inhibition de la sécrétiond ’insuline, activation de la sécrétion de glucagon :
Quelles sont les adaptations métaboliques mises en jeu?Le fœtus stocke de grande quantité de glycogène pendant le derniertrimestre de la gestation et des triglycérides dans son tissu adipeux.Le nouveau-né humain est un des nouveau-nés les plus gras à lanaissance
Stress fœtal (hypoxie) ==> adrénaline + noradrénaline ==> inhibition de la sécrétion d ’insuline,activation de la sécrétion de glucagon :
- Glycogénolyse- Activation de la gluconéogenèse==> production hépatique de glucose
- Lipolyse dans le tissu adipeux blanc ==> libération d’AGL et de glycérol dans la circulation- Activation de la ß-oxydation et de la cétogénèse hépatique ==> libération de corps cétoniques pourle cerveau==> épargne de glucose (les besoins en glucose sont élevés alors que la masse musculaire permettantde fournir des précurseurs gluconéogéniques est faible).
- Chez le nouveau-né, un intervalle entre deux tétées (environ 5-6h) correspond en termesd’adaptations énergétiques à un jeûne de 48h chez l ’adulte.
- Les nouveaux-nés prématurés, qui n ’ont pas stocké de glycogène hépatique et de lipides sont à risquede développer des hypoglycémies néonatales.

L’exercice musculaire
D’un point de vue énergétique, il existe différents types de fibres musculaires.
Fibres glycolytiques (blanches) II Fibres oxydatives (rouges) I
- Vitesse de contraction Rapide Lente
- Capacité glycolytique Elevée Faible
- Capacité oxydative (mitochondries) Faible Elevée
- Réseau capillaire Pauvre Dense
- Myoglobine Faible Elevée
- Réserve de glycogène Elevée Modérée
- Type d ’exercice Intense, rapide Longue durée(saut, sprint)

Exercice court et intense : Sprint
Puissance élevée : 100m : 6 kcal =>10s, 36Kcal/min (50000 Kcal/jour!!)
Mise en jeu des fibres rapides glycolytiques (conduction nerveuse rapide, mise en jeu dpresque toutes les fibres, cycles contraction/relaxation très courts).
Energie immédiatement disponible :- ATP régénéré par l’hydrolyse de la phosphocréatine:H+ +ADP + phosphocréatine => ATP + créatine.Ce système peut assurer 5-10 s de contraction.
• Le glucose est le seul substrat pouvantfournir l'énergie nécessaire à laformation de l'ATP en anaérobie (5% dela quantité totale d'énergie produite parson métabolisme aérobie).
• L’origine du glucose (glucose-6-P) est leglycogène, dégradé après quelquessecondes d’exercice.
• Mais production d’ions H+ et baisse dupH intracellulaire ==> arrêt de lacontraction.
Exercice en endurance
Marathon: 42,2 km : 2800 kcal =>2h 15 min, (20cal/min)Mise en jeu des fibres lentes oxydatives.- Après une phase initiale anaérobique, les adaptations circulatoires et ventilatoirespermettent d’augmenter les apports en oxygène de 0,3 à 3-4 L d’oxygène par minute.- Multiplication du débit cardiaque (fréquence + volume d’éjection) par 4-5 (de 5 L à25 L/min).-Augmentation de la fréquence respiratoire.-Redistribution du débit sanguin vers les muscles (de 1 L/min à 20 L/min)
ATP
Créatine
Glycolyse
anaérobie
Métabolisme
aérobie
2s 10s 90s
Durée de l’exercice
Anaérobie Aérobie

Les acides gras libres
-La phase aérobique permet l’oxydation du glucose mais également des acides graslibres.
- L’utilisation des acides gras libres commence vers 10 min et augmenteprogressivement avec la durée de l’exercice pour atteindre 60% des besoinsénergétiques après 3 heures.
- L’utilisation des acides gras libres est favorisé par la chute intracellulaire dumalonyl-CoA et par une lipolyse qui atteint son maximum après 2-3 heures d’effort.
- L’utilisation des lipides permet une épargne du glycogène, indispensable pour lapoursuite de l’effort (rôle anaplérotique).
Dépassement des capacités aérobies : la notion de VO2 max
VO2 max : Volume maximal d’oxygène prélevé au niveau des poumons et utilisé par les
muscles par unité de temps (35-50 ml/min/kg).
Lors d’un effort qui dépasse les 50-70 % de VO2 max, le seuil aérobique est dépassé et la
filière anaérobique lactique est sollicitée, conduisant après quelques minutes à l’arrêt de
l’effort. .
La VO2 max peut être considérablement augmentée par l’entraînement et le seuilanaérobique retardé jusqu’à 90% de la VO2 max chez des sportifs exceptionnels.

Les concentrations circulantes de lactate
Puissance (Watt)
Lact
ate
san
gu
in (
mm
ol/
L)
1
5
10
Le lactate produit pendant l’exercice devra être oxydé à l’arrêt de l’exercice,
entraînant une «!dette!» d’oxygène.
0 100 200 300
Et le glucose circulant ?
Glucose
La contraction musculaire s’accompagne d’une augmentation du captage du
glucose circulant. Le recrutement des transporteurs GLUT 4 à la membrane n’est
pas dans ces conditions dépendante de l’insuline et pourrait impliquer l’AMP-
activated protein kinase.

La production hépatique de glucose
Afin de maintenir l’homéostasie glucidique lors de l’exercice prolongé:
# La glycogénolyse est stimulée.
#La gluconéogénèse est également stimulée.
Elle utilise :
-Le lactate provenant du métabolisme anaérobie du glucose.
-Le glycérol provenant de la lipolyse.
-L’alanine dans les efforts très prolongé où une protéolyse se met en place.
Lactate
Alanine
GlucoseTG Glycérol
Quels sont les mécanismes régulateurs mis en jeu ?
L’exercice musculaire est un stress : augmentation des concentrations circulantes
de catécholamines :
# Adaptations circulatoires et respiratoires.
# Modifications des sécrétions pancréatiques : baisse del’insuline et augmentation
du glucagon favorisant la glycogénolyse et la gluconéogenèse hépatiques et la
lipolyse dans le tissu adipeux.

Quels sont les mécanismes régulateurs mis en jeu ?
Adaptations musculaires locales :
# Baisse de l’ATP, augmentation de l’AMP (quelques secondes). Activation du
transport de glucose par mobilisation de GLUT4. Activation de la glycogène
phosphorylase. Activation de la glycolyse.
#Augmentation des concentrations cytoplasmiques de calcium (quelques secondes).
Activation de la glycogène phosphorylase.
#Vasodilatation des capillaires permettant un apport accru d’oxygène et de
substrats.
#A plus long terme, diminution des concentrations de malonyl-CoA. Augmentation
de la ß-oxydation.


















