
Rôle de la mécanotransduction du poil racinaire … · Le poil absorbant joue un rôle crucial...
21
1 ÉCOLE DOCTORALE 145 "SCIENCES DU VÉGÉTAL : DU GENE A L’ECOSYSTEME " A envoyer à [email protected] pour le 12 février 2015 PROPOSITION DE SUJET AU CONCOURS 2015 A - TITRE DE LA THESE Rôle de la mécanotransduction du poil racinaire pendant les étapes précoces de la symbiose entre Medicago et son microsymbiote bactérien Role of mechanotransduction in root hair during early steps of symbiosis between Medicago and its microsymbiont B - DIRECTEUR DE THESE Jean-Marie FRACHISSE http://www.isv.cnrs-gif.fr/recherche/st/sttheme3-msl.html Tél: 01 69 82 36 87 e-mail: [email protected]gif.fr Habilitation à diriger des recherches C - UNITE DE RATTACHEMENT ET EQUIPE D’ACCUEIL Institut de Biologie Intégrative de la Cellule (I2BC), UMR 9198, Département de Biologie Cellulaire, équipe "Approches intégrées des transports d'ions" dirigée par Sébastien Thomine D – NOMBRE DE THESES ENCADREES PAR LE DIRECTEUR DE THESE Doctorants encadrés actuellement par le directeur de thèse, date de 1 ère inscription et type de financement de la thèse: (ajuster le nombre de lignes si nécessaire) Nom, Prénom Date 1ère inscription Financement Guichard Marjorie Octobre 2013 Bourse MESR Doctorants ayant soutenu leur thèse au cours des 5 dernières années : (ajuster le nombre de lignes si nécessaire) Nom, Prénom Date 1ère inscription Date de soutenance Situation actuelle (si possible)
Transcript of Rôle de la mécanotransduction du poil racinaire … · Le poil absorbant joue un rôle crucial...

1
ÉCOLE DOCTORALE 145 "SCIENCES DU VÉGÉTAL : DU GENE A L’ECOSYSTEME "
A e n v o y e r à e c o d o c . s d v@ u - p s u d . f r p o u r l e 1 2 f é v r i e r 2 0 1 5
P RO POS I T ION D E S U J E T A U CONCOURS 2 0 1 5 A - TITRE DE LA THESE Rôle de la mécanotransduction du poil racinaire pendant les étapes précoces de la symbiose entre Medicago et son microsymbiote bactérien Role of mechanotransduction in root hair during early steps of symbiosis between Medicago and its microsymbiont B - DIRECTEUR DE THESE Jean-Marie FRACHISSE http://www.isv.cnrs-gif.fr/recherche/st/sttheme3-msl.html Tél: 01 69 82 36 87 e-mail: [email protected] Habilitation à diriger des recherches C - UNITE DE RATTACHEMENT ET EQUIPE D’ACCUEIL Institut de Biologie Intégrative de la Cellule (I2BC), UMR 9198, Département de Biologie Cellulaire, équipe "Approches intégrées des transports d'ions" dirigée par Sébastien Thomine D – NOMBRE DE THESES ENCADREES PAR LE DIRECTEUR DE THESE Doctorants encadrés actuellement par le directeur de thèse, date de 1ère inscription et type de financement de la thèse: (ajuster le nombre de lignes si nécessaire) Nom, Prénom Date 1ère inscription Financement Guichard Marjorie Octobre 2013 Bourse MESR Doctorants ayant soutenu leur thèse au cours des 5 dernières années : (ajuster le nombre de lignes si nécessaire) Nom, Prénom Date 1ère
inscription Date de soutenance
Situation actuelle (si possible)

2
Liste complète des publications des doctorants ayant soutenu leur thèse au cours des 5 dernières années, références complètes à donner en soulignant les noms des doctorants encadrés (et * pour M2 encadrés) Le dernier doctorant ayant soutenu sa thèse que j’ai encadré a obtenu celle-ci il y a plus de 5 ans (Rémi Peyronnet, janvier 2009) je porte ci-dessous pour information les articles que nous avons co-publiés
Haswell, E., Peyronnet, R., Barbier-Brygoo, H., Meyerowitz, E. M. & Frachisse, J. M. (2008) Two MscS homologues provide mechanosensitive channel activities in the Arabidopsis root. Curr. Biol., 18: 730-734
Peyronnet R, Haswell ES, Barbier-Brygoo H, and Frachisse JM (2008) AtMSL9 and AtMSL10: Sensors of plasma membrane tension in Arabidopsis roots. Plant Signaling & Behavior 3, 726-729.
Colcombet J., Mathieu Y., Peyronnet R., Agier N., Lelièvre F., Barbier-Brygoo H., Frachisse J.M. (2009) R-type anion channel activation is an essential step for ROS-dependent innate immune response in Arabidopsis suspension cells Functional Plant Biology, 36:832-843
Diatloff E., Peyronnet R., Colcombet J., Thomine S., Barbier-Brygoo H., Frachisse JM. (2010) R type anion channel: a multifunctional channel seeking its molecular identity. Plant Signaling and Behavior, 5:22-27. Peyronnet R., Tran D., Tiffanie G., Frachisse JM. (2014) Mechanosensitive channels: feeling tension in a world under pressure. Front. Plant Sci. doi: 10.3389/fpls.2014.00558 Nombre total de thèses que vous avez encadrées : 3 + 1 en cours
F - COLLABORATIONS (Collaborations développées dans le cadre du projet) - H. Sentenac/ A-A Very, Biochimie et Physiologie Moléculaire des Plantes, INRA, Montpellier (électrophysiologie sur Medicago) - N. Pauly, Interactions Biotiques et Santé Végétale, INRA, Sophia Antipolis (outils génétique et BM chez Medicago) - J-M Alain, Laboratoire de Mécanique des Solides, Polytechnique (modélisation du poil racinaire) -A. Genre, Departement of life Sciences and System Biology, Université de Turin (signalisation calcique Medicago) G – FINANCEMENTS ASSOCIES AU PROJET ANR Blanc CAROLS (2012-2015) H –RESUMES DU PROJET (200 mots maximum, ces résumés seront mis en ligne sur le site de l’ED) Français : Le poil absorbant joue un rôle crucial dans l’absorption de l’eau et des ions minéraux, et ainsi dans le développement de la plante. Chez les Légumineuses, il est également le lieu où s’établit l’interaction symbiotique avec la bactérie fixatrice d’azote du genre Rhizobium. Cette symbiose qui permet une forte réduction des intrants azotés présente une importance considérable dans le cadre d’une agriculture durable. De plus, le poil absorbant constitue un modèle unicellulaire

3
prometteur pour une approche de type "biologie des systèmes" ce qui est un défi actuel pour les végétalistes. La croissance cellulaire polarisée ainsi que la réorientation de la croissance du poil absorbant suite à l’interaction physique de celui-ci avec la bactérie symbiotique rhizobium sont des processus complexes, dans lesquels les canaux ioniques, dont les canaux mécanosensibles, jouent des rôles clefs. Le but du projet de thèse est d’obtenir une vision d’ensemble de l’implication des canaux ioniques mécanosensibles de la membrane plasmique du poil et de leur régulation par la tension de membrane dans les premières étapes de la symbiose entre la Légumineuse modèle Medicago truncatula et son microsymbiote Sinorhizobium meliloti. Anglais : The root hair plays crucial roles in water and nutrient uptake, and thus in plant development. In Legumes, it also plays a central role in the establishment of the nitrogen-fixing symbiotic interaction with rhizobia, which has a crucial importance for sustainable agriculture as it largely contributes to limitation of nitrogen fertilizers. Furthermore, the root hair provides a model to investigate the biology of a single differentiated plant cell type, one of the present challenges for plant scientists, and is considered as a highly promising model for plant cell systems biology. This so-called tip growth and the re-orientation of the tip growth occurring during the initial physical interaction with the rhizobial microsymbiont are highly complex processes, in which ion channels including mechanosensitive channels are key actors. The aim of the PhD project is to obtain a holistic view of the involvement of root hair plasma membrane mechanosensitive ion channels and of their regulation by membrane stretching during the first steps of the symbiotic interaction between the model Legume Medicago truncatula and its microsymbiont Sinorhizobium meliloti. I – MOTS CLES (choisir 5 mots clés maximum représentant la thématique et la méthodologie) Medicago, poil absorbant, canaux mécanosensibles, symbiose, calcium J – INTRODUCTION ET CONTEXTE SCIENTIFIQUE (1 page maxi) Le poil absorbant joue un rôle prépondérant dans l’alimentation minérale et hydrique de la plante. Chez les légumineuses il constitue le point d’entrée des bactéries conduisant à l’établissement d’une symbiose entre la plante et les bactéries fixatrices d’azote. Ces deux aspects à eux seuls prouvent l’intérêt d’une bonne connaissance de la physiologie et des fonctions du poil absorbant. De plus, étant une structure unicellulaire facilement accessible, le poil absorbant, au même titre que les cellules de garde pour les parties aériennes, représente un système modèle très prometteur. Il permet d’étudier le développement et l’élongation cellulaire, l’absorption hydrique et minérale et les réponses à des signaux biotiques et abiotiques dans le cadre d’une approche de "biologie des systèmes" (Libault et al., 2010). Les canaux ioniques, aussi bien chez les animaux que chez les plantes, sont des éléments précoces de la perception des stimuli biotiques et abiotiques délivrés par l’environnement (Hetherington and Brownlee, 2004 ; Maathuis, 2009, Monshausenand Gilroy, 2009). Le rôle de ces canaux a particulièrement été bien analysé chez les cellules de garde où il a été montré que ces derniers interviennent dans la transpiration de la plante par le contrôle qu’ils exercent sur l’ouverture du stomate (Schröder et al., 2001 ; Fan et al., 2004). Concernant leur implication dans une réponse

4
mécanique, on peut avancer trois faisceaux d’arguments : (i) d’une part la présence d’activités canal mécanosensible a été observée sur la membrane plasmique sur tous types de protoplastes isolés à partir de différents organes provenant de différentes espèces végétales (Haswell 2007), d’autre part (ii) l’identification de deux canaux mécanosensibles MSL9 et MSL10 (Haswell et al., 2007) homologues du MscS de E. coli (Mechano-Sensitive Channel of Small Conductance) actifs sur la racine d’Arabidopsis et finalement (iii) la capacité du poil à "ressentir" une interaction physique (le toucher) ceci étant illustré par la réponse exacerbée que présente le poil du mutant dmi2 (Does not Make Infections) de Medicago qui est par ailleurs incapable d’entrer en symbiose avec la bactérie (Esseling 2004). Le programme de thèse s’inscrit dans la continuité du projet ANR CAROLS (partenaires : H. Sentenac/ A-A Very, Montpellier et N. Pauly, Sophia Antipolis) qui a pour but d’obtenir une vision d’ensemble de l’implication des canaux ioniques de la membrane plasmique du poil racinaire et de leur régulation dans les premières étapes de la symbiose entre la légumineuse modèle M. truncatula et son microsymbiote S. meliloti. Dans le cadre de ce projet de thèse, nous nous intéresserons au rôle des canaux mécanosensibles du poil absorbant. Pour cela nous développerons une caractérisation fonctionnelle des canaux mécanosensibles appartenant aux familles MSL, Piezo et MCA, spécifiquement exprimés dans le poil absorbant. Notamment nous préciserons d’une part (i) les conditions mécaniques d’activation et les propriétés fonctionnelles des canaux mécanosensibles (ii) des éléments de régulation de l’activité de ces canaux tels le cytosquelette et les EAO (espèces activées d’oxygène), (iii) l’action des canaux mécanosensibles sur la déformation et les propriétés mécaniques du poil. Ce projet fournira des informations indispensables sur les acteurs de la perception des signaux mécaniques lors de la déformation et de la pénétration du cordon bactérien symbiontes dans le poil racinaire (Jayaraman et al., 2014).
K – OBJECTIFS DU PROJET SCIENTIFIQUE (quelques lignes)
Les objectifs de ce projet sont dans la continuité de l’étude déjà entreprise par D. Tran (post doc dans la cadre du projet ANR CAROLS) et par M. Guichard (étudiante en 2éme année de thèse). Dans un premier temps la caractérisation fonctionnelle des canaux mécanosensibles appartenant aux familles MSL, MCA et Piezo et exprimés dans le poil racinaire sera conduite. Notamment le rôle du cytosquelette dans l’activation du canal sera étudié ainsi que la régulation par les messagers secondaires Ca2+ et EAO. Par une approche de génétique inverse le rôle de ces canaux dans la capacité du poil à se déformer (massue, crosse de berger) consécutivement à une microstimulation mécanique ou en réponse à la bactérie symbionte sera précisé. Ceci nous conduira à une vision d’ensemble de l’implication des canaux mécanosensibles dans les premières étapes de la symbiose entre la Légumineuse modèle M. truncatula et son microsymbiote S. meliloti.
L – APPROCHES PROPOSEES (détailler : stratégie, outils disponibles ou à construire, méthodologie, faisabilité, le cas échéant contribution des collaborations mentionnées en F) (2 pages maxi)
Le programme proposé s’appuie sur les outils et résultats déjà obtenus sur cette thématique, il se décline selon les trois taches explicitées ci-dessous. Les travaux de M. Guichard (2eme année de thèse) couplé à l’analyse d’expression des gènes dans le poil racinaire par RNAseq (ANR CAROLS) ont permis de préciser que les canaux mécanosensibles, MtMSLII-1, MtMSLII-4 et MtMCA1 et MtPiezo2 étaient présents dans le poil racinaire. De plus il a été montré pour certain des MSL appartenant à la classe II (fusion traductionnelle GFP::MSL) une localisation de ceux-ci sur la membrane plasmique.

5
Tâche 1 Clonage des canaux MtPiezo2 et MtMCA1 et sélection de lignée invalidées pour ces gènes
Tâche 1.1 : Le canal MSLII-4 a été cloné dans le cadre de la thèse de M. Guichard et différents outils moléculaires sont disponibles (MSL avec GFP en fusion N-ter et C-ter, gènes rapporteurs GUS et GFP sous promoteurs MSL, …) par contre les canaux Piezo2 et MCA1 seront clonés en utilisant la technologie Gateway. Cette technologie largement utilisée dans notre laboratoire permettra par la réaction de recombinaison LR de transférer le gène candidat dans un vecteur permettant soit de réaliser une fusion avec une protéine reportrice GFP pour faire de la localisation subcellulaire, soit de faire de l’expression transitoire du canal afin d’en réaliser la caractérisation électrophysiologique (tâche 2).
Tâche 1.2 : Des lignées invalidées pour les deux gènes MSL (insertion du transposon Tnt-1) ont été obtenues auprès de la "Noble Foundation" et la descendance de ces plantes est en cours de génoptypage. Il sera nécessaire dans le cadre de ce projet de se procurer les graines pour lesquelles les gènes Piezo2 et MCA1 ont étés invalidés afin de générer un outil équivalent à celui que nous allons obtenir pour les MSLII-1, MSLII-4. Ces plantes permettront de conduire l’analyse phénotypique décrite dans la tache 3.
Tâche 2 Caractérisation fonctionnelle des canaux candidats
Nous avons montré au laboratoire que des protoplastes provenant d’un mutant d’Arabidopsis délété pour cinq gènes codants pour différents MSL (quintuple mutant msl4; msl5; msl6; msl9; msl10) constituaient un système d’expression permettant de caractériser les canaux mécanosensibles (Haswell et al., 2008). Nous utiliserons ce système d’expression transitoire pour effectuer la caractérisation fonctionnelle des canaux Piezo-2 et MCA-1 exprimés dans le poil racinaire de Medicago.
Une telle caractérisation fonctionnelle qui à l’heure actuelle est en cours pour le canal MSLII-4, devrait permettre de répondre au moins à quatre questions majeures dans le cadre de ce projet :
(i) quelle est la mecanosensibilité des canaux Piezo2 et MCA1 du poil absorbant de Medicago? Des éléments de réponse seront amenés en étudiant la probabilité d’ouverture (Po) de ces canaux en fonction de la tension mécanique appliquée à la membrane plasmique. L’appareil d’imposition rapide de pression "High Speed Pressure Clamp" récemment acquis au laboratoire permettra de conduire cette étude.
(ii) quelle est la sélectivité des canaux mécanosensibles? Celle-ci devrait être en faveur des anions pour les MtMSL si ils se comportent comme leur homologues décrits chez Arabidopsis (Haswell et al., 2008). Concernant Piezo et MCA les informations disponibles dans la littérature indiquent que ces deux canaux seraient sélectifs des cations. Piezo chez la souris est perméable aux cations monovalents alors que MCA1 d’Arabidopsis serait perméable au calcium (Kurusu et al., 2013 ; Peyronnet et al., 2014). Une telle approche permettra de définir la sélectivité de ces canaux et de préciser si celle-ci est en faveur du calcium, comme l’on pourrait s’y attendre dans le cadre d’une réponse rapide à une stimulation mécanique.
(iii) l’activité du canal est-elle modulée par la présence d’effecteurs de type EAO (espèces activées d’oxygène), par des molécules symbiotiques (facteur Nod), et par l’intégrité du cytosquelette ? Une approche pharmacologique consistant à faire agir dans la bain de perfusion de l’H2O2 ou des effecteurs du cytosquelette (Zang et al., 2007) amènera des éléments de réponse à ces questions.
6
Tâche 3 Identification du rôle des canaux impliqués dans la réponse de mécanotransduction du poil.
Une analyse par imagerie de la forme du poil chez des plantes sauvages ou invalidées pour les gènes codants pour les canaux MSLII-1, MSLII-4, Piezo2 et MCA1 sera conduite dans un premier temps. Cette étude sera complétée par une analyse de la déformation du poil (chez les différentes lignées mentionnées ci-dessus) soumis à différentes stimulations tels que : (i) une microstimulation mécanique, (ii) le facteur Nod, (iii) la bactérie microsymbionte. L’analyse et l’interprétation des données sera conduite en collaboration avec J-M Allain (Laboratoire de Mécanique des Solides, Polytechnique) et devrait aboutir à une modélisation sur le rôle des canaux mecanosensibles du poil dans les premières étapes de la symbiose.
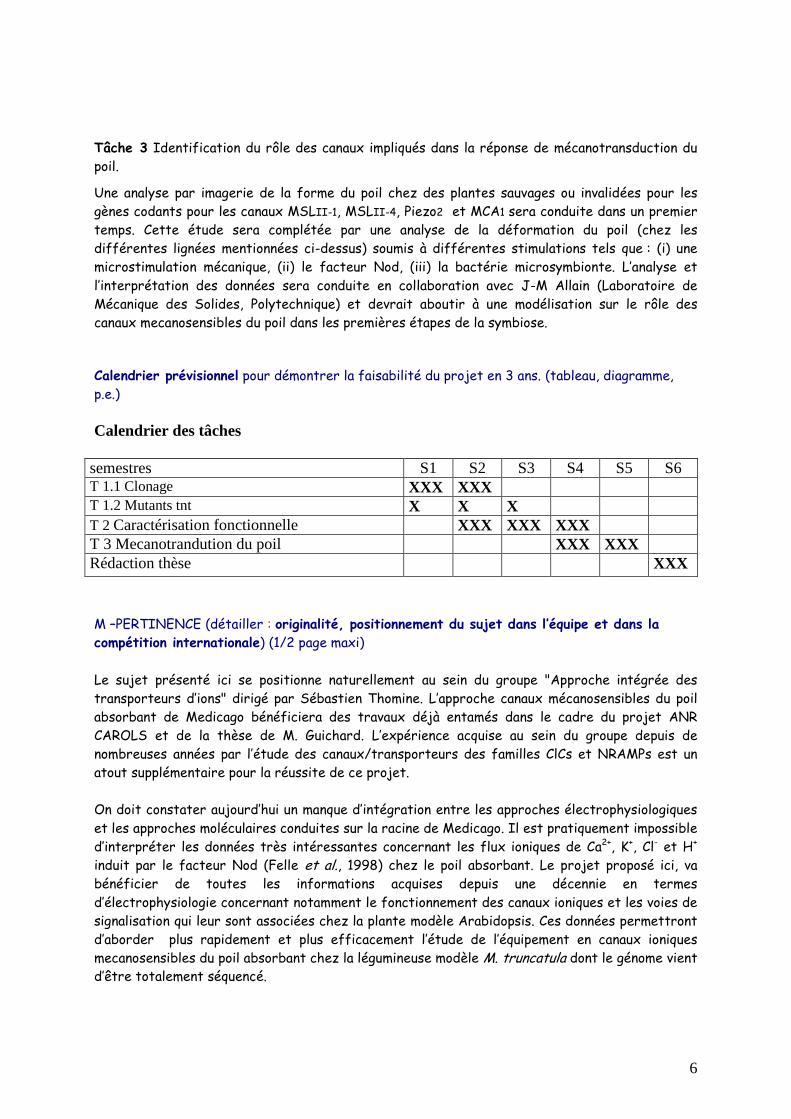
Calendrier prévisionnel pour démontrer la faisabilité du projet en 3 ans. (tableau, diagramme, p.e.)
Calendrier des tâches
semestres S1 S2 S3 S4 S5 S6 T 1.1 Clonage XXX XXX T 1.2 Mutants tnt X X X T 2 Caractérisation fonctionnelle XXX XXX XXX T 3 Mecanotrandution du poil XXX XXX Rédaction thèse XXX
M –PERTINENCE (détailler : originalité, positionnement du sujet dans l’équipe et dans la compétition internationale) (1/2 page maxi)
Le sujet présenté ici se positionne naturellement au sein du groupe "Approche intégrée des transporteurs d’ions" dirigé par Sébastien Thomine. L’approche canaux mécanosensibles du poil absorbant de Medicago bénéficiera des travaux déjà entamés dans le cadre du projet ANR CAROLS et de la thèse de M. Guichard. L’expérience acquise au sein du groupe depuis de nombreuses années par l’étude des canaux/transporteurs des familles ClCs et NRAMPs est un atout supplémentaire pour la réussite de ce projet.
On doit constater aujourd’hui un manque d’intégration entre les approches électrophysiologiques et les approches moléculaires conduites sur la racine de Medicago. Il est pratiquement impossible d’interpréter les données très intéressantes concernant les flux ioniques de Ca2+, K+, Cl- et H+ induit par le facteur Nod (Felle et al., 1998) chez le poil absorbant. Le projet proposé ici, va bénéficier de toutes les informations acquises depuis une décennie en termes d’électrophysiologie concernant notamment le fonctionnement des canaux ioniques et les voies de signalisation qui leur sont associées chez la plante modèle Arabidopsis. Ces données permettront d’aborder plus rapidement et plus efficacement l’étude de l’équipement en canaux ioniques mecanosensibles du poil absorbant chez la légumineuse modèle M. truncatula dont le génome vient d’être totalement séquencé.

7
Ce projet est conduit en étroite collaboration avec les deux autres partenaires (H. Sentenac, Montpellier et N. Pauly, Sophia Antipolis) du programme ANR CAROLS. Les données moléculaires et fonctionnelles obtenues à Gif-sur-Yvette sur les canaux mécanosensibles seront complétées par celles obtenues par le partenaire de Montpellier. Elles concernent notamment les canaux potassiques de la famille shaker (Very and Sentenac, 2003), les canaux anioniques de la famille SLAC (Geiger et al., 2009) et les canaux des familles CNGC et Glutamate-récepteur dont la sélectivité reste controversée. Le troisième partenaire quant à lui s’intéressera plus spécifiquement aux EAO et à leur effet sur les patrons d’expression de ces différentes familles de canaux. Cette approche conduite en parallèle par trois laboratoires devrait permettre d’aboutir à une vision la plus complète en termes d’équipement en canaux ioniques du poil absorbant et du rôle de ceux-ci dans l’interaction précoce poil/Rhizobium.
Par les rencontres et les échanges qu’il génère, ce projet aura un effet stimulant sur les différents doctorants impliqués dans chaque équipe. Cet aspect interactif fait partie intégrante de la formation du doctorant et l’initiera ainsi à la recherche dans un contexte collaboratif.
N – BIBLIO ANNEXE (10-12 références)
Esseling JJ, Emons AM. (2004) Anonsymbiotic root hair tip growth phenotype in NORKmutated legumes: implications for nodulation factor-induced signaling and formation of a multifaceted root hair pocket for bacteria. Plant Cell. 16:933-944
Fan LM, Zhao Z, Assmann SM (2004). Guard cells: a dynamic signaling model. CurrOpin Plant Biol. 7: 537-546.
Felle H. H., Kondorosi E., Kondorosi A., Schultze M. (1998). The role of ion fluxes in Nodfactor signalling in Medicago sativa. Plant J. 13: 455–463.
Geiger D., Scherzer S., Mumm P., Stange A., Marten I., Bauer H., Ache P., Matschi S., Liese A., Al-Rasheid K.A.S., Romeis T., Hedrich R. (2009). Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase-phosphatase pair. Proc. Natl. Acad. Sci. U. S. A 106: 21425–21430.
HaswellE S (2007). MscS-like Proteins in Plants. Current Topics in Membranes 58: 329-359.
Haswell ES., Peyronnet R., Barbier-Brygoo, H., Meyerowitz EM,.Frachisse JM. (2008) Two MscS homologues provide mechanosensitive channel activities in the Arabidopsis root.Curr. Biol., 18: 730-734
Hetherington AM and Brownlee C (2004). The generation of Ca2+ signals in plants. Annu.Rev. Plant Biol. 55: 401-427.
Kurusu,T.,Kuchitsu,K.,Nakano,M.,Nakayama,Y.,andIida,H.(2013). Plant mechanosensingandCa2+ transport. TrendsPlantSci. 18, 227–233.doi: 10.1016/j.tplants.2012.12.002
Libault M, Brechenmacher L, Cheng J, Xu D and Stacey G. (2010) Root hair systems biology. Trends in Plant Science 11:641-50.
Maathuis FJM (2009). Physiological functions of mineral macronutrients. Curr.Opin. PlantBiol. 12:250-258.
Monshausen GB, Gilroy S. (2009).Feeling green: mechanosensing in plants.Trends Cell Biol.;19(5):228-35
Peyronnet R., Tran, D., Girault T,.Frachisse JM. (2014) Mechanosensitivechannels:feelingtensioninaworldunderpressure
Peyronnet R.,Tran, D., Girault T, Frachisse JM. (2014) Mechanosensitive channels: feeling tension in a world under pressure. Front. In Plant Sciences, doi: 10.3389/fpls.2014.00558
Schroeder JI, Kwak JM, Allen GJ (2001). Guard cell abscisic acid signalling and engineeringdrought hardiness in plants.Nature 410: 327-330.

8
VéryA.-A., and Sentenac H. (2003). Molecular mechanisms and regulation of K+ transport inhigher plants. Annu. Rev. Plant Biol. 54: 575-603.
Zhang W, Fan LM, Wu WH. (2007) Osmo-sensitive and stretch-activated calcium-permeable channels in Viciafaba guard cells are regulated by actin dynamics. Plant physiol.143:1140-1151
SVP donner 1-2 revues récentes sur le sujet pour aider le conseil et le jury. Merci d’engendrer un seul fichier avec le projet et les articles de synthèse en pdf.
Jayaraman D., Gilroy S., Ané J-M. (2014) Staying in touch: mechanical signals in plant–microbe interactions. Current Opinion in Plant Biology. 01/2014; 20:104–109.
Kurusu,T., Kuchitsu,K., Nakano,M., Nakayama, Y.,and Iida,H.(2013). Plant mechanosensing and Ca2+ transport. Trends Plant Sci. 18, 227–233.doi: 10.1016/j.tplants.2012.12.002
O – APPRECIATION DU CONSEIL D’UNITE (OBLIGATOIRE)
Je soussigné Renaud Legouis, chef du département de Biologie Cellulaire de l’I2BC (UMR9198) et représentant du directeur Thierry Meinnel, donne un avis favorable à la proposition de sujet au concours de l’école doctorale 145 "SCIENCES DU VÉGÉTAL : DU GENE A L’ECOSYSTEME ". Le sujet proposé au concours s’intègre parfaitement avec les axes de recherches du Département et les conditions d’encadrement d’un étudiant en thèse dans l’équipe d’accueil sont tout à fait satisfaisantes.

Staying in touch: mechanical signals in plant–microbeinteractionsDhileepkumar Jayaraman1, Simon Gilroy2 and Jean-Michel Ane1
Mechanical stimulations play a significant role in the day to day
existence of plants. Plants exhibit varied responses depending
on the nature and intensity of these stimuli. In this review, we
present recent literature on the responses of plants to
mechanical stimuli, focusing primarily on those exerted during
plant–microbe interactions. We discuss how microbes are able
to apply mechanical stimuli on plants and how some plant
responses to pathogenic and symbiotic microbes present
striking similarities with responses to mechanical stimuli applied,
for instance, using micro-needles. We hypothesize that
appropriate responses of plants to pathogenic and symbiotic
microbes may require a tight integration of both chemical and
mechanical stimulations exerted by these microbes.
Addresses1 Department of Agronomy, University of Wisconsin, Madison, WI 53706,
United States2 Department of Botany, University of Wisconsin, Madison, WI 53706,
United States
Corresponding author: Ane, Jean-Michel ([email protected])
Current Opinion in Plant Biology 2014, 20:104–109
This review comes from a themed issue on Biotic interactions
Edited by Makoto Hayashi and Martin Parniske
For a complete overview see the Issue and the Editorial
Available online 27th May 2014
http://dx.doi.org/10.1016/j.pbi.2014.05.003
1369-5266/# 2014 Elsevier Ltd. All rights reserved.
BackgroundPlants are exposed to a myriad of stimuli during their life
cycle and are characterized by their inability to relocate to
avoid stressful environments [1]. Mechanical stimulation
(MS), which includes the effects of gravity, wind, turgor,
touch, pathogens or wounding is an inescapable com-
ponent of a plant’s surroundings that significantly affects
plant growth and development, morphogenesis, repro-
duction, and survival [2]. Plants perceive and respond to
these mechanical cues at the cellular and organismal
levels [3]. Plants such as the Venus flytrap (Dionaeamuscipula), Sundew (Drosera rotundifolia), and Sensitive
Plant (Mimosa pudica) respond within seconds to MS due
to the presence of specialized mechanosensory cells [1].
Most other plants, despite lacking these specialized cells,
also respond to MS by altering their growth rate, growth
direction, and morphology [1]. ‘Thigmomorphogenesis’ is
the term collectively used to denote plant growth and
developmental responses to MS [2,3]. Recent evidence
shows that MS signaling is closely linked with defense
signaling pathways. Mutants in jasmonic acid synthesis
and response have been found to display reduced MS
responses, whereas jasmonic acid overproduction mimics
MS [4�]. MS results in the accumulation of jasmonic acid
and promotes defense against the fungal pathogen Botry-tis cinerea [4�]. Jasmonic acid also acts as a commonly
induced signaling molecule in wounding and MS. The
transient elevation of endogenous jasmonic acid followed
by defense gene activation is a common response to this
wounding, and also in plants subjected to repeated MS
exposure [5]. In addition, wounding and MS also lead to a
swift increase in the cytosolic calcium concentrations, and
it is widely believed that these changes play a role in
activating many downstream defense responses [6]. MS is
known to trigger widespread changes in gene expression,
including upregulation of a classic set of ‘touch-inducible’
(TCH) genes that encode calmodulin and the calmodulin-
like proteins [7]. The calmodulins and calmodulin-like
proteins have also been linked to the maintenance of
innate immunity with, for example, mutants in TCH2showing impaired resistance to Pseudomonas syringae [8].
Interestingly, some mutants affected in plant–microbe
symbioses also present defects in their responses to MS.
For instance, in the model legume Medicago truncatula,
does not make infections 2 and 3 (dmi2 and dmi3) mutants are
affected in both nodulation and mycorrhization. The dmi2mutant is further affected in its ability to resume growth
after MS [9��], whereas the dmi3 mutant showed an
altered cytoplasmic response after MS [10��]. In addition,
dmi2 mutants show more symbiotic responses to rhizobial
Nod factors in the absence than in the presence of MS,
indicating that MS plays a critical role in regulating the
response to symbiotic signals [9��]. These observations
raise the question of relationships between systems
responsible for MS sensing and response, and those
mediating plant–microbe interactions; are the appearance
of similar response components in both systems function-
ally linked or just coincidence?
Pathogenic and symbiotic microbes can exertMS on plantsPlants interact with a variety of microbes (from beneficial
to pathogenic) in the environment. Microbes colonize
plants to satisfy their environmental, nutritional, or water
needs [11]. Arbuscular mycorrhizal fungi form hyphopo-
dia on the roots, and most pathogenic fungi and
Available online at www.sciencedirect.com
ScienceDirect
Current Opinion in Plant Biology 2014, 20:104–109 www.sciencedirect.com
oomycetes form appressoria on the leaves and hyphopodia
on the roots, at certain points of contact with the plant
epidermis [12,13]. The appressoria develop a penetration
peg at their base that aids in the penetration of the plant
cell, whereas in the case of hyphopodia, the hyphae that
develop aid in invading plant tissues [14]. The turgor
pressure of the hyphae provides the mechanical force
necessary for breaching the plant cell wall and entering
plant tissues [11,14,15]. It has been estimated that Magna-porthe grisea and Colletotrichum species produce a force of 8–25 mN to facilitate invasion of host tissues [14,16��]. The
appressoria or hyphopodia vary in size, which in turn may
affect the force generated for penetration. For instance, in
Medicago truncatula, three fungi have been reported to
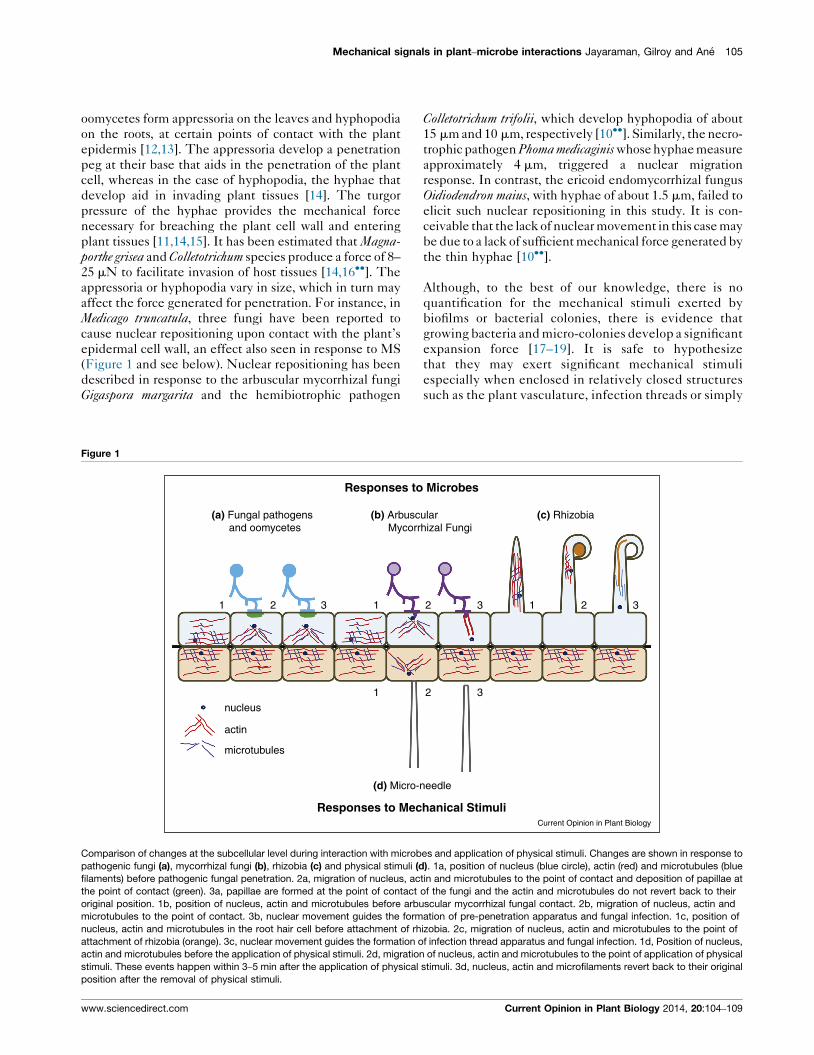
cause nuclear repositioning upon contact with the plant’s
epidermal cell wall, an effect also seen in response to MS
(Figure 1 and see below). Nuclear repositioning has been
described in response to the arbuscular mycorrhizal fungi
Gigaspora margarita and the hemibiotrophic pathogen
Colletotrichum trifolii, which develop hyphopodia of about
15 mm and 10 mm, respectively [10��]. Similarly, the necro-
trophic pathogen Phoma medicaginis whose hyphae measure
approximately 4 mm, triggered a nuclear migration
response. In contrast, the ericoid endomycorrhizal fungus
Oidiodendron maius, with hyphae of about 1.5 mm, failed to
elicit such nuclear repositioning in this study. It is con-
ceivable that the lack of nuclear movement in this case may
be due to a lack of sufficient mechanical force generated by
the thin hyphae [10��].
Although, to the best of our knowledge, there is no
quantification for the mechanical stimuli exerted by
biofilms or bacterial colonies, there is evidence that
growing bacteria and micro-colonies develop a significant
expansion force [17–19]. It is safe to hypothesize
that they may exert significant mechanical stimuli
especially when enclosed in relatively closed structures
such as the plant vasculature, infection threads or simply
Mechanical signals in plant–microbe interactions Jayaraman, Gilroy and Ane 105
Figure 1
nucleus
actin
microtubules
(d) Micro-needle
(a) Fungal pathogens and oomycetes
1 1
1
12 2
2
23 3
3
3
(b) Arbuscular Mycorrhizal Fungi
(c) Rhizobia
Responses to Mechanical Stimuli
Responses to Microbes
Current Opinion in Plant Biology
Comparison of changes at the subcellular level during interaction with microbes and application of physical stimuli. Changes are shown in response to
pathogenic fungi (a), mycorrhizal fungi (b), rhizobia (c) and physical stimuli (d). 1a, position of nucleus (blue circle), actin (red) and microtubules (blue
filaments) before pathogenic fungal penetration. 2a, migration of nucleus, actin and microtubules to the point of contact and deposition of papillae at
the point of contact (green). 3a, papillae are formed at the point of contact of the fungi and the actin and microtubules do not revert back to their
original position. 1b, position of nucleus, actin and microtubules before arbuscular mycorrhizal fungal contact. 2b, migration of nucleus, actin and
microtubules to the point of contact. 3b, nuclear movement guides the formation of pre-penetration apparatus and fungal infection. 1c, position of
nucleus, actin and microtubules in the root hair cell before attachment of rhizobia. 2c, migration of nucleus, actin and microtubules to the point of
attachment of rhizobia (orange). 3c, nuclear movement guides the formation of infection thread apparatus and fungal infection. 1d, Position of nucleus,
actin and microtubules before the application of physical stimuli. 2d, migration of nucleus, actin and microtubules to the point of application of physical
stimuli. These events happen within 3–5 min after the application of physical stimuli. 3d, nucleus, actin and microfilaments revert back to their original
position after the removal of physical stimuli.
www.sciencedirect.com Current Opinion in Plant Biology 2014, 20:104–109

the plant apoplast. These forces may be required for
penetration into the host tissue in pathogenic and sym-
biotic interactions.
Some responses to microbes are similar toresponses to MSAs mentioned previously, nuclear repositioning within
the plant cell towards the site of contact is a common
response to MS, fungi, and symbiotic bacteria. This
nuclear repositioning appears to be a relatively generic
response to MS but it is affected by the size of the
touching object. For example, in Medicago truncatula, this
response is observed only in cells that are in contact with
objects larger than 2 mm irrespective of the object — be it
a micropipette, fungal hyphae, or appressoria [10��].During plant–microbe interactions, such nuclear move-
ments occur before cell penetration. Indeed, in the case of
arbuscular mycorrhizal fungi or rhizobia, nuclear move-
ment initiates the formation of a pre-penetration appar-
atus and infection thread, respectively, that guides the
infection process [10��,20�]. At later stages of symbiotic
associations with arbuscular mycorrhizal fungi, nuclear
repositioning was also observed during arbuscule devel-
opment suggesting a possible role for MS in this process,
too [13,21].
Cytoplasmic streaming and aggregation in the plant cell
beneath the invading pathogen, including the reorganiza-
tion of cytoskeletal and endomembrane components, is
also a response common to several plant–microbe inter-
actions [10��,22–24], although, the precise dynamics of
the response of the plant’s microtubule response differs
with the invading microorganism [25]. MS exerted on
plant cells using glass or tungsten needles in leaf epider-
mal cells or cotyledons caused a similar cytoskeletal
reorganization, including the aggregation of actin micro-
filaments beneath the contact site, the formation of a
microtubule-depleted zone, and the accumulation of
endomembrane components, such as the endoplasmic
reticulum and peroxisomes, beneath the contact point.
Although the force generated by these needles is not
known, these responses were rapid, occurring within 3–5 min of stimulus application, and migrated to the region
where the stimulus (needle) was moved. In addition,
these responses were abolished once the stimulus was
removed but so far no cytoplasmic aggregation has been
reported in roots upon application of MS. Because plants
are able to perceive and respond to the MS applied by the
needles with changes that mimic those triggered in
response to pathogens, it is likely that, in plant–microbe
interactions, MS exerted by the pathogens may be trig-
gering these responses [23��].
A further similarity between microbial and MS-induced
responses in plants lies in their link to a common signaling
molecule, extracellular adenosine-50-triphosphate, eATP.
Although eATP elicits several cellular responses in
plants, only recently has it been recognized as a bona fidesignal [26]. eATP has been implicated in stress responses
(both biotic and abiotic), including thigmotropism [26]. In
Arabidopsis, eATP is released in response to touch, and
this release is modulated by the heteromeric G-protein
complex [27]. In legumes, lectin nucleotide phosphohy-
drolase (LNP) are Nod factor-binding proteins induced
following rhizobial infection. Antibodies directed towards
the LNPs decreased nodulation in Dolichos fluorus and
soybean [28,29]. Knockdown of these LNPs resulted in
reduced nodulation in soybean and Lotus japonicus [30,31]
but also reduced mycorrhization in L. japonicus [31]. LNPs
are cell wall-located ecto-apyrases that hydrolyze ATP
into ADP, suggesting that the knockdown in LNP should
cause elevated eATP levels. Addition of eADP partially
rescued the nodulation phenotype of soybean knock-
down lines suggesting that eADP could be a signal.
Reciprocally, overexpression of the LNP resulted in
increased rhizobial infection, consistent with reduced
ATP or increased ADP promoting the development of
symbiosis [30,32]. Rhizobial diffusible signals, Nod fac-
tors, may trigger eATP release as these Nod factors are
similar to fungal chitin elicitors, which have been impli-
cated in ATP release [26]. As discussed further, MS may
also reinforce this eATP release. The associated increase
in eATP would then be used to fuel eADP production by
the LNP [30]. Very recently a lectin receptor kinase-I.9
(LecRK-I.9) was identified as a receptor for eATP in
Arabidopsis and named P2K [33��]. The ecto-apyrases
may occur alongside the P2K receptor (as seen in animal
systems) and together may compete for the nucleotides
that are released from plant cells in response to signals
such as mechanical, environmental, or pathogenic stress.
The balance between these two systems, ecto-apyrase
and P2K could then determine the magnitude of the
eATP signal generated by these stimuli and so govern
whether the plant would mount an appropriate response
[34–36].
MS work in concert with chemical stimuli toregulate defense and symbiotic reactionsResponses to pathogens
Lipopolysaccharides are major components of the outer
membrane of Gram-negative bacteria and are essential for
processes such as establishing contact with the host
during the early stages of infection and bacterial viru-
lence. Lipopolysaccharide mutants are affected not only
in their ability to form biofilms, but also in their virulence
for phytopathogenic bacteria such as Pseudomonas syringaeand Xanthomonas citri [37]. Because lipopolysaccharide
mutants often are impaired in biofilm formation, they may
lack the mechanical force necessary to penetrate the host
tissue. Arabidopsis leaves subjected to mechanical stress
exhibit enhanced resistance to the fungal pathogen Botry-tis cinerea. Here, the perception of MS leads to changes in
the calcium concentration and production of reactive
106 Biotic interactions
Current Opinion in Plant Biology 2014, 20:104–109 www.sciencedirect.com

oxygen species, two signaling molecules well character-
ized as being generated by a wide range of MS in plants.
Indeed, reactive oxygen species have been directly impli-
cated in resistance to B. cinerea [38�]. In the case of
Phytophtora, in addition to mechanical stimuli, this species
secretes elicitins and effectors. Elicitins are 104 kDa
proteins which play a role in the uptake of sterols from
plant membranes and are recognized by lectin-like re-
ceptor kinases, triggering the hypersensitive response. In
Phytophtora, four effectors have been identified so far and
are recognized by nucleotide-binding site–leucine-rich
repeat (NBS–LRR) proteins, leading to hypersensitive
reactions. Taken together, a combination of chemical and
mechanical stimuli is produced by oomycetes and fungi,
and both of these stimuli are detected by plants, leading
to the onset of the basal defense mechanisms. A success-
ful invasion occurs if the fungi can overcome the basal
defenses and subsequent hypersensitive cell death that
are triggered by their elicitors and effectors [24].
Responses to symbionts
In many legumes, rhizobia enter the root through intra-
cellular infection of root hairs via an infection thread.
Entrapment of rhizobia in the root hair curl (shepherd’s
crook) and the formation of a micro-colony inside of this
curl are required for the initiation of the infection thread.
Nod factors produced by the rhizobia trigger root hair
curling and the purpose of the root hair curl may be to
provide MS [9��]. Possible origins for the MS include: (i)
the root hair curls on itself and pushes on its own shank; or
(ii) the curl is necessary to entrap bacteria and form a
micro-colony, which through its growth provides the MS.
From an evolutionary standpoint, it is possible that the
root hair curl may ‘mimic’ hyphopodia from arbuscular
mycorrhizal fungi. Interestingly, the size of the micro-
colony in the root hair curl is approximately 10–15 mm,
which is roughly the size of the arbuscular mycorrhizal
fungi hyphopodia and we hypothesize that it may there-
fore provide a similar force [10��]. In many tropical
legumes, instead of entering through the root hairs, the
rhizobia pass through the epidermis via cracks, in a
process known as crack entry. For instance, in Sesbaniarostrata bacterial colonies, named infection pockets, are
formed at the site of infection and are required for the
further formation of infection threads. These infection
pockets contain a dense bacterial population that can be
seen as analogous to a micro-colony in root hair curls and
may provide the necessary MS for further entry into host
cells [39].
A precise combination of chemical and mechanical
stimuli may be essential for successful symbiotic coloni-
zation. For example, during colonization by arbuscular
mycorrhizal fungi, plant cells show a characteristic repeti-
tive increase in nuclear and perinuclear calcium concen-
trations, named calcium spiking, as part of the signaling
system allowing symbiotic associations. Calcium spiking
can be induced by Myc factors, diffusible signals pro-
duced by arbuscular mycorrhizal fungi. Interestingly, the
frequency of calcium spiking is greater in root cells
containing arbuscular mycorrhizal fungi hyphae than in
the adjacent cells. It is conceivable that the arbuscular
mycorrhizal fungi hyphae not only produce Myc factors
but also exert MS in the infected cells, whereas the
adjacent cells perceive only the Myc factors [13,40�]and so may generate a response of lower intensity. Sim-
ilarly, in rhizobium–legume associations, diffusible Nod
factors trigger calcium spiking in their legume host.
Knocking-down LNP expression in L. japonicus reduced
nodulation, mycorrhization and calcium spiking in
response to Nod factors but not root hair deformations,
suggesting that MS from the root hair curl or the hyphae
may regulate the plant response to Nod and Myc factors
(including calcium spiking) via eATP release and the
LNP [31]. Addition of eATP also caused oscillations of
the cytoplasmic calcium concentration in Arabidopsis
roots. However, there is no evidence that these calcium
waves originate from the nuclear envelope like calcium
spiking [41]. In addition, Arabidopsis lacks the CASTOR
protein which is required for calcium spiking in L. japo-nicus. It is therefore unclear at this point that these
oscillations of cytoplasmic calcium concentration are
related.
In addition rhizobia produce exopolysaccharides that are
chief components of the biofilm matrix. For instance,
Sinorhizobium meliloti produces two distinct exopolysac-
charides: a succinoglycan and a galactoglucan. Both
succinoglycan and Nod factors are essential for the
formation of an active infection thread. Rhizobial exo-
polysaccharides may play a role in infection thread
formation via signaling or by changing the physicochem-
ical conditions within the infection thread and we
hypothesize that this change may affect the MS exerted
by the bacteria and/or exopolysaccharides itself inside
the thread exerting MS, on host plant cells. Rhizobial
exopolysaccharides may alter free calcium concen-
trations by binding to these ions, or the acidic exopo-
lysaccharide alginate may change the hydration of the
infection thread matrix by binding to water. Taken
together, exopolysaccharides may affect host–bacterial
interactions by chemical but also mechanical means
[37,42–44]. Within the infection thread, it is speculated
that physical movement followed by concomitant
division of rhizobial cells occurs and exopolysaccharides
may facilitate such movement. Furthermore, presence
of colonizing bacteria in close vicinity to the tip is
essential for progression of the infection thread
suggesting that coordinated development of the infec-
tion thread is governed by some form of bacterium–host
communication. Nod factors in conjunction with MS
generated by the growing infection thread may be key
signals controlling infection thread progression in rhi-
zobium–legume symbioses [20�].
Mechanical signals in plant–microbe interactions Jayaraman, Gilroy and Ane 107
www.sciencedirect.com Current Opinion in Plant Biology 2014, 20:104–109

ConclusionMechanical forces play a key role in plant growth and
development and although the influence of MS on plant
growth has long been recognized, our understanding of
the molecular mechanisms involved in MS perception
and transduction is still in the nascent stages. The realiz-
ation that MS may be a key signal monitored by plants
and their interacting microbes to establish symbioses or
initiate pathogenesis should help pinpoint potential roles
for orphan plant mechanosensors. This neglected field of
mechanosensing and how MS regulate plant responses to
pathogenic and symbiotic microbes and interact with
diverse signaling systems is a missing component in
our understanding of many plant response pathways.
The advent of new and refined techniques in the field
of cell imaging, biomechanics, next-generation sequenc-
ing, and bioinformatics, in addition to an interdisciplinary
research approach, should help to unravel the signal
transduction mechanism(s) of MS. This, in turn, will
aid in future efforts to engineer defenses against patho-
gens and associations with beneficial microbes.
AcknowledgementsThis work was supported by a grant from the NASA (NNX13AM50G) to SGand a grant from the National Science Foundation (NSF#0701846) to JMA.
References and recommended readingPapers of particular interest, published within the period of review,have been highlighted as:
� of special interest
�� of outstanding interest
1. Chehab E, Eich E, Braam J: Thigmomorphogenesis: a complexplant response to mechano-stimulation. J Exp Bot 2009, 60:43-56.
2. Scippa G, Trupiano D, Rocco M, Di Iorio A, Chiatante D:Unravelling the response of poplar (Populus nigra) roots tomechanical stress imposed by bending. Plant Biosyst 2008,142:401-413.
3. Braam J: In touch: plant responses to mechanical stimuli. NewPhytol 2005, 165:373-389.
4.�
Chehab E, Yao C, Henderson Z, Kim S, Braam J: Arabidopsistouch-induced morphogenesis is jasmonate mediated andprotects against pests. Curr Biol 2012, 22:701-706.
The results from this study shows that plants impaired in jasmonic acidbiosynthesis and response are affected in their response to mechanicalstimuli. In addition, this paper also revealed that mechanical stimuli leadto enhanced jasmonic acid accumulation and results in enhanced resis-tance to fungal pathogens.
5. Tretner C, Huth U, Hause B: Mechanostimulation of Medicagotruncatula leads to enhanced levels of jasmonic acid. J Exp Bot2008, 59:2847-2856.
6. Romeis T, Herde M: From local to global: CDPKs in systemicdefense signaling upon microbial and herbivore attack. CurrOpin Plant Biol 2014, 20C:1-10.
7. Braam J, Davis RW: Rain-, wind-, and touch-inducedexpression of calmodulin and calmodulin-related genes inArabidopsis. Cell 1990, 60:357-364.
8. Ma W, Smigel A, Tsai Y, Braam J, Berkowitz G: Innate immunitysignaling: cytosolic Ca2+ elevation is linked to downstreamnitric oxide generation through the action of calmodulin or acalmodulin-like protein. Plant Physiol 2008, 148:818-828.
9.��
Esseling JJ, Lhuissier FG, Emons AM: A nonsymbiotic root hairtip growth phenotype in NORK-mutated legumes: implicationsfor nodulation factor-induced signaling and formation of amultifaceted root hair pocket for bacteria. Plant Cell 2004,16:933-944.
This paper provides the first experimental evidence that mechanicalstimuli cause a nonsymbiotic root hair tip growth phenotype in the norkmutants of the model legume Medicago truncatula. It also suggests thatthese mutants are unable to entrap the bacteria due to the mechanicalstimuli that are exerted by the plant’s own root hair cells upon curling andtouching its shank. In addition, this paper also suggests a dual signalingrole for Nod factors: one involved in root hair curling and the other insymbiotic gene expression.
10.��
Genre A, Ortu G, Bertoldo C, Martino E, Bonfante P: Biotic andabiotic stimulation of root epidermal cells reveals commonand specific responses to arbuscular mycorrhizal fungi. PlantPhysiol 2009, 149:1424-1434.
This paper demonstrates the non-specific nuclear repositioning of epi-dermal cells when subjected to mechanical stimuli greater than a certainmagnitude and also demonstrates that nuclear repositioning can beuncoupled from cytoplasmic aggregation. This is supported by the lackof cytoplasmic aggregation in the epidermal cell when touched with amicromanipulator or a non-host fungi.
11. Nezhad A, Geitmann A: The cellular mechanics of an invasivelifestyle. J Exp Bot 2013, 64:4709-4728.
12. Bonfante P, Genre A: Mechanisms underlying beneficial plant–fungus interactions in mycorrhizal symbiosis. Nat Commun2010, 1:48.
13. Gutjahr C, Parniske M: Cell and developmental biology ofarbuscular mycorrhiza symbiosis. Annu Rev Cell Dev Biol 2013,29:593-617.
14. Bastmeyer M, Deising HB, Bechinger C: Force exertion in fungalinfection. Annu Rev Biophys Biomol Struct 2002, 31:321-341.
15. Money PN: Insights on the mechanics of hyphal growth. FungalBiol Rev 2008, 22:71-76.
16.��
Bechinger C, Giebel K, Schnell M, Leiderer P, Deising H,Bastmeyer M: Optical measurements of invasive forcesexerted by appressoria of a plant pathogenic fungus. Science1999, 285:1896-1899.
Using optical waves, this paper directly measured the force applied by asingle appresorium, indicating that the mechanical force exerted by somefungi is sufficient to infect the plant hosts.
17. Su PT, Liao CT, Roan JR, Wang SH, Chiou A, Syu WJ: Bacterialcolony from two-dimensional division to three-dimensionaldevelopment. PLoS ONE 2012, 7:e48098.
18. Farrell FD, Hallatschek O, Marenduzzo D, Waclaw B:Mechanically driven growth of quasi-two-dimensionalmicrobial colonies. Phys Rev Lett 2013, 111:168101.
19. Tuson HH, Auer GK, Renner LD, Hasebe M, Tropini C, Salick M,Crone WC, Gopinathan A, Huang KC, Weibel DB: Measuring thestiffness of bacterial cells from growth rates in hydrogels oftunable elasticity. Mol Microbiol 2012, 84:874-891.
20.�
Fournier J, Timmers A, Sieberer B, Jauneau A, Chabaud M,Barker D: Mechanism of infection thread elongation in roothairs of Medicago truncatula and dynamic interplay withassociated rhizobial colonization. Plant Physiol 2008, 148:1985-1995.
This paper demonstrates that infection thread development is a discon-tinuous process and suggests that a combination of a physical stimulusprovided by bacterial movement inside the thread and a chemical sti-mulus provided by the bacterially secreted Nod factors is essential for theprogression of the infection thread.
21. Genre A, Chabaud M, Faccio A, Barker D, Bonfante P:Prepenetration apparatus assembly precedes and predictsthe colonization patterns of arbuscular mycorrhizal fungiwithin the root cortex of both Medicago truncatula andDaucus carota. Plant Cell 2008, 20:1407-1420.
22. Genre A, Chabaud M, Timmers T, Bonfante P, Barker D:Arbuscular mycorrhizal fungi elicit a novel intracellularapparatus in Medicago truncatula root epidermal cells beforeinfection. Plant Cell 2005, 17:3489-3499.
108 Biotic interactions
Current Opinion in Plant Biology 2014, 20:104–109 www.sciencedirect.com

23.��
Hardham A, Takemoto D, White R: Rapid and dynamicsubcellular reorganization following mechanical stimulationof Arabidopsis epidermal cells mimics responses to fungaland oomycete attack. BMC Plant Biol 2008:8.
This paper provides evidence that the application of mechanical stimulion the epidermal cell surface elicits responses similar to those induced byfungal or oomycete pathogens. This paper suggests that the perceptionof the mechanical force exerted by the pathogen during invasion may leadto the mounting of the basal defense response by the plants.
24. Hardham A, Blackman L: Molecular cytology of Phytophthora–plant interactions. Austral Plant Pathol 2010, 39:29-35.
25. Hardham AR: Microtubules and biotic interactions. Plant J2013, 75:278-289.
26. Tanaka K, Gilroy S, Jones A, Stacey G: Extracellular ATPsignaling in plants. Trends Cell Biol 2010, 20:601-608.
27. Weerasinghe RR, Swanson SJ, Okada SF, Garrett MB, Kim SY,Stacey G, Boucher RC, Gilroy S, Jones AM: Touch induces ATPrelease in Arabidopsis roots that is modulated by theheterotrimeric G-protein complex. FEBS Lett 2009, 583:2521-2526.
28. Etzler M, Kalsi G, Ewing N, Roberts N, Day R, Murphy J: A nodfactor binding lectin with apyrase activity from legume roots.Proc Natl Acad Sci U S A 1999, 96:5856-5861.
29. Day R, McAlvin C, Loh J, Denny R, Wood T, Young N, Stacey G:Differential expression of two soybean apyrases, one of whichis an early nodulin. Mol Plant–Microbe Interact 2000, 13:1053-1070.
30. Govindarajulu M, Kim SY, Libault M, Berg RH, Tanaka K, Stacey G,Taylor CG: GS52 ecto-apyrase plays a critical role duringsoybean nodulation. Plant Physiol 2009, 149:994-1004.
31. Roberts NJ, Morieri G, Kalsi G, Rose A, Stiller J, Edwards A, Xie F,Gresshoff PM, Oldroyd GE, Downie JA et al.: Rhizobial andmycorrhizal symbioses in Lotus japonicus require lectinnucleotide phosphohydrolase, which acts upstream ofcalcium signaling. Plant Physiol 2013, 161:556-567.
32. McAlvin C, Stacey G: Transgenic expression of the soybeanapyrase in Lotus japonicus enhances nodulation. Plant Physiol2005, 137:1456-1462.
33.��
Choi J, Tanaka K, Cao Y, Qi Y, Qiu J, Liang Y, Lee SY, Stacey G:Identification of a plant receptor for extracellular ATP. Science2014, 343:290-294.
This paper used the ATP-induced calcium response to identify thereceptor for extracellular ATP in plants, suggesting that these receptorsin plants are different than those present in animals.
34. Song CJ, Steinebrunner I, Wang X, Stout SC, Roux SJ: ExtracellularATP induces the accumulation of superoxide via NADPHoxidases in Arabidopsis. Plant Physiol 2006, 140:1222-1232.
35. Joseph SM, Buchakjian MR, Dubyak GR: Colocalizationof ATP release sites and ecto-ATPase activity at theextracellular surface of human astrocytes. J Biol Chem 2003,278:23331-23342.
36. Alvarado-Castillo C, Harden TK, Boyer JL: Regulation of P2Y1receptor-mediated signaling by the ectonucleosidetriphosphate diphosphohydrolase isozymes NTPDase1 andNTPDase2. Mol Pharmacol 2005, 67:114-122.
37. Bogino PC, Oliva Mde L, Sorroche FG, Giordano W: The role ofbacterial biofilms and surface components in plant–bacterialassociations. Int J Mol Sci 2013, 14:15838-15859.
38.�
Benikhlef L, L’Haridon F, Abou-Mansour E, Serrano M, Binda M,Costa A, Lehmann S, Metraux J: Perception of soft mechanicalstress in Arabidopsis leaves activates disease resistance.BMC Plant Biol 2013:13.
This paper demonstrates that plants can perceive mechanical stimuli,which in turn leads to increased resistance to fungi. This paper alsosuggests that this resistance is mediated by the enhanced production ofROS, which is implicated in the activation of defenses to biotic and abioticstress.
39. D’Haeze W, De Rycke R, Mathis R, Goormachtig S, Pagnotta S,Verplancke C, Capoen W, Holsters M: Reactive oxygen speciesand ethylene play a positive role in lateral root base nodulationof a semiaquatic legume. Proc Natl Acad Sci U S A 2003,100:11789-11794.
40.�
Sieberer BJ, Chabaud M, Fournier J, Timmers AC, Barker DG: Aswitch in Ca2+ spiking signature is concomitant withendosymbiotic microbe entry into cortical root cells ofMedicago truncatula. Plant J 2012, 69:822-830.
This study determined that the calcium spiking pattern is different in thecortical cells during rhizobial infection thread progression and arbuscularmycorrhizal pre-penetration apparatus progression. This paper alsoshows that these two microbes elicit a similar frequency of calciumoscillations during their entry into the cortical cells.
41. Tanaka K, Swanson SJ, Gilroy S, Stacey G: Extracellularnucleotides elicit cytosolic free calcium oscillations inArabidopsis. Plant Physiol 2010, 154:705-719.
42. Jones KM, Kobayashi H, Davies BW, Taga ME, Walker GC: Howrhizobial symbionts invade plants: the Sinorhizobium–Medicago model. Nat Rev Microbiol 2007, 5:619-633.
43. Mendis HC, Queiroux C, Brewer TE, Davis OM, Washburn BK,Jones KM: The succinoglycan endoglycanase encoded byexoK is required for efficient symbiosis of Sinorhizobiummeliloti 1021 with the host plants Medicago truncatula andMedicago sativa (Alfalfa). Mol Plant–Microbe Interact 2013,26:1089-1105.
44. Rinaudi L, Giordano W: An integrated view of biofilm formationin rhizobia. FEMS Microbiol Lett 2010, 304:1-11.
Mechanical signals in plant–microbe interactions Jayaraman, Gilroy and Ane 109
www.sciencedirect.com Current Opinion in Plant Biology 2014, 20:104–109

Plant mechanosensing and Ca2+
transportTakamitsu Kurusu1,2,3, Kazuyuki Kuchitsu1,2, Masataka Nakano4,Yoshitaka Nakayama4, and Hidetoshi Iida4
1 Department of Applied Biological Science, Tokyo University of Science, 2641 Yamazaki, Noda, Chiba 278-8510, Japan2 Research Institute for Science and Technology, Tokyo University of Science, 2641 Yamazaki, Noda, Chiba 278-8510, Japan3 School of Bioscience and Biotechnology, Tokyo University of Technology, 1404-1 Katakura, Hachioji, Tokyo 192-0982, Japan4 Department of Biology, Tokyo Gakugei University, 4-1-1 Nukui kita-machi, Koganei, Tokyo 184-8501, Japan
Mechanical stimuli generate Ca2+ signals and influencegrowth and development in plants. Recently, candidatesfor Ca2+-permeable mechanosensitive (MS) channelshave been identified. These channels are thought tobe responsible for sensing osmotic shock, touch, andgravity. One candidate is the MscS-like (MSL) proteinfamily, a homolog of the typical bacterial MS channels.Some of the MSL proteins are localized to plastids tomaintain their shape and size. Another candidate is themid1-complementing activity (MCA) protein family,which is structurally unique to the plant kingdom.MCA proteins are localized in the plasma membraneand are suggested to be involved in mechanosensingand to be functionally related to reactive oxygen species(ROS) signaling. Here, we review their structural featuresand role in planta.
Plants sense mechanical stimuliCharles Darwin described how, when the petioles of someclimbing plants were touched with sticks, the petiolesclasped them and increased in thickness [1]. Rubbingthe internode for approximately 10 s once or twice a dayresulted in morphological changes and inhibition of growth[2]. Recently, this response, thigmomorphogenesis, wasshown to play a role in protection against pests [3]. Touch-ing twice a day was sufficient to retard elongation ofinflorescences and delay flowering in Arabidopsis thaliana[4]. These reports are examples of how mechanical stimuli,such as touching and shaking, can affect morphogenesis,growth, and development in plants.
The examples given above represent mechanical stimu-lus-induced gradual alterations, which are often hardlyvisible to the human eye. However, plants also show rapidresponses. The two lobes of the leaf of the Venus flytrapDionaea muscipula rapidly close to capture insects thatcontact the trigger-hairs at the center of the leaf [5]. Theleaflet of Mimosa pudica folds after mechanical stimula-tion. In both cases, the responses occur within 1 s.
Whether the responses are fast or slow, plants are ableto sense and respond to mechanical stimuli. There has beena long history of studies on these processes, but the molec-ular mechanisms by which mechanical sensing leads to
responses have only recently started to be elucidated(reviewed in [3,4,6–9]).
Mechanosensing and Ca2+ transientsHow do plants sense mechanical stimuli? Some progresstowards answering this question was made using Arabi-dopsis plants expressing a Ca2+-sensitive photoprotein,aequorin, from the jellyfish Aequorea victoria [10]. Therecombinant plants emitted light immediately after touch,osmotic stress [6], and gravistimulation [11,12], suggestingthat an immediate early event in the mechanoresponse isto increase cytosolic Ca2+ concentration ([Ca2+]cyt). Hypo-osmotic shock, as well as trinitrophenol, an activator of MSchannels in Escherichia coli [13], induced a rapid andtransient rise in [Ca2+]cyt, predominantly due to plasmamembrane Ca2+ influx in plant cells [14,15]. Althoughmechanical stimuli, such as touch, bending, and barriersto growth, all elicited rapid and transient increases in[Ca2+]cyt, their spatiotemporal patterns (Ca2+ signatures)were stimulus specific [16].
Consistent with these observations, cellular Ca2+-bind-ing regulatory proteins, such as calmodulin and calmodu-lin-like proteins, were induced by mechanical stimuli [17].These downstream events, as well as adaptive responsesincluding the induction of transcription factors, could besuppressed when Ca2+ influx was compromised by Ca2+
chelators, such as ethylene glycol tetraacetic acid (EGTA) or1,2-bis(o-aminophenoxy)ethane-N,N,N0,N0-tetraacetic acid(BAPTA), or by Ca2+-channel blockers [15,18,19]. Therefore,it is possible that plasma membrane Ca2+-permeable MSchannels are key molecules that sense mechanical stimuli.
In this review, we describe the properties and themolecular basis of Ca2+-permeable MS channels in variousplant species. We also highlight the important roles of theMS channels in mechanical signaling and stress adapta-tion in plant cells.
Plant MS channels and candidatesMSL protein family
Genome-wide screening to search for plant MS channelshomologous to eukaryotic and prokaryotic MS channelsrevealed the MSL protein family [20,21]. MscS, one of threetypes of prokaryotic MS channel, has MS channel activitywith small conductance, and functions as a safety valve toavoid cell rupture by releasing cytoplasmic solutes upon
Review
Corresponding authors: Iida, H. ([email protected]);Kuchitsu, K. ([email protected])
1360-1385/$ – see front matter � 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tplants.2012.12.002 Trends in Plant Science, April 2013, Vol. 18, No. 4 227

hypo-osmotic shock [22]. MSLs are found in land plants,algae, and fungi, but not in animals (Table 1). The overallamino acid sequences of MSLs are not homologous to thoseof prokaryotic MscS; only a pore-forming transmembranesegment and the secondary structure adjacent to thissegment have been conserved. Conserved motifs ofMSL2 have recently been characterized [23].
Based on the sequence of this conserved region, eukary-otic MSLs can be grouped into two main classes; Class Iand Class II [24]. Several Class I MSLs contain organelle-targeting sequences predicted to localize to mitochondriaor plastids. By contrast, Class II MSLs do not have atargeting sequence and are found in fungi and protists,which are organisms that lack plastids. MSLs in fissionyeast are localized at the endoplasmic reticulum (ER)membrane and regulate [Ca2+]cyt to support the survivalof cells upon hypo-osmotic shock [25]. Arabidopsis has tenMSL genes (MSL1–10) in its genome. MSL1–3 and MSL4–10 are assigned to Class I and Class II, respectively.
Several lines of evidence indicate that some of the MSLfamily proteinshaveMSchannelactivity.MSL9 and MSL10,two of seven Class II Arabidopsis MSLs localized in theplasma membrane in roots, provide MS channel activity,although their physiological function is not yet known [26].Phenotypically, the double (msl9/10) and quintuple (msl4/5/6/9/10) mutant plants grow normally. Electrophysiolog-ical studies suggest that MSL9 and MSL10 permeate Cl–
rather than Ca2+ [26,27]. Recently, electrophysiologicalproperties of MSL10 were clearly demonstrated [28].Chlamydomonas reinhardtii MSC1, a chloroplast-localizedClassIMSL,wasshownelectrophysiologicallytohaveanion-selective MS channel activity in E. coli giant spheroplasts[29]. Fission yeast Msy1, an ER-localized Class II MSL, hasalso been demonstrated electrophysiolologically to have MSchannel activity [25]. Genetically, MSL2 and MSL3 aresuggested to act as MS channels because both proteins cancomplement the lethality of an E. coli mutant lacking MscSunder hypo-osmotic conditions [30]. It is still unknownwhether these MSLs are involved in mechanical stimuli-induced generation of Ca2+ signals as Ca2+-permeable MSchannels.
The physiological evidence obtained so far suggests thatMSL2 and MSL3 are plastid MS channels that participatein maintaining the shape and size of the plastid [30]. Bothproteins are also required to protect plastids from hypo-osmotic stress during normal plant growth [31]. The Ara-bidopsis msl2 msl3 double mutant has large, round plas-tids that lack dynamic tubular structures, known asstromules, on the surface. This phenotype can be rescuedby increasing cytoplasmic osmolarity through exogenouslyprovided osmolytes, or by withholding water. The enlargedplastids in the msl2 msl3 double mutant contain multiple
filamentous temperature-sensitive Z (FtsZ) rings [32].MSL2 and MSL3 are suggested to function as componentsof the plastid division machinery in the pathway to regulatesize and division of plastids, such as the Min system, servingto restrict FtsZ ring formation to the middle of plastids.
The Piezo protein family
Piezo proteins in mouse and Drosophila are pore-formingsubunits of MS channels and respond to mechanical sti-muli. Drosophila larvae lacking the DmPiezo gene showeda reduced behavioral response to noxious mechanical sti-muli, compared with their wild type counterparts [33,34].Many eukaryotic species, including plant species, have asingle Piezo protein, but yeast and bacteria have no clearhomologs [35]. Studies on plant Piezo homologs have notyet been reported.
MCA protein family
A genetic screen of Arabidopsis cDNAs, which comple-ments the conditional lethality of a yeast mutant lackingthe functional MID1 gene, resulted in the identification ofMCA1 as a candidate MS channel [14]. The yeast Mid1protein shows MS channel activity when expressed inmammalian cells [36]. Ca2+ uptake activity has beenshown for MCA1 and its paralog MCA2, as well as rice(Oryza sativa) OsMCA1 and tobacco (Nicotiana tabacum)NtMCA1 and NtMCA2 [14,15,37–39]. MCA proteins arepresent in all land plants, including ferns and mosses, butnot in algae, animals, protists, and fungi, suggesting thatthe function of MCA proteins is fundamental to landplants. Phylogenetic analyses indicate that genetic diver-sity of MCA genes occurred relatively recently, and onlyone MCA gene is present in various Poaceae [38](Figure 1a). Interestingly, among the MCA proteins indicots, those in Brassicaceae, including Arabidopsis andBrassica, are diverged from those in other dicots, and existas two distinct paralogs (MCA1 and MCA2), suggestingtheir functional divergence. Functional diversity betweenArabidopsis MCA1 and MCA2 has indeed been suggested[37]. Localization, ion permeability, and physiological func-tions of MSLs and MCAs characterized so far are summa-rized in Table 2.
Structural features of MCA proteinsThe amino acid sequences of MCA proteins show relativelyhigh similarity throughout the entire sequence, exceptfor the serine-rich region in the middle, which showssubstantial diversity among these proteins (Figure 1b).The N-terminal half has a region similar to the putativeregulatory domain of protein kinase candidates found ingrass species, and coiled-coil and EF-hand-like motifs[14,38,40].
Despite their activity in mediating Ca2+ uptake, MCAproteins have no homology to other known ion channels ortransporters. Truncation analysis showed that the N-ter-minal half without the coiled-coil motif of MCA1 and MCA2(MCA11-173 and MCA21-173) is necessary and sufficient forCa2+ uptake activity [40]. This region has one putativetransmembrane segment that contains a highly conservedAsp21. Replacement of Asp21 with Asn resulted in completeand partial loss of Ca2+ uptake activity in MCA1 and
Table 1. Presence or absence of MS channels in variouskingdoms and a domain
Kingdom/domain MscL MscS MCA TRP Piezo
Plants – + + – +
Animals – – – + +
Fungi – + – + ?
Bacteria + + – – –
Review Trends in Plant Science April 2013, Vol. 18, No. 4
228
MCA2, respectively. Both MCA1 and MCA2 form a tetra-mer, suggesting that this region is the functional motif ofthe MCA protein, whereas the putative transmembranesegments of the MCA tetramer form a Ca2+-permeablepore (Figure 1b; see also Figure 1 of [40]).
The EF-hand-like and coiled-coil motifs are suggested tohave distinct effects on the Ca2+ influx activity of Arabi-dopsis MCA1 and MCA2 expressed in yeast cells, asassessed by truncation analysis [40]. A truncated form ofMCA1 lacking the EF-hand-like motif (MCA11-135) did notshow activity, whereas that of MCA2 (MCA21-135) did,suggesting that this motif positively regulates MCA1.Another truncated form of MCA1 with the coiled-coil motif(MCA11-237) did not have activity, whereas that of MCA2(MCA21-237) did. As mentioned above, the truncated formof MCA1 lacking the coiled-coil motif but having the EF-hand-like motif (MCA11-173) showed activity, suggestingthat the coiled-coil motif negatively regulates MCA1. Thus,it is possible that the two motifs regulate MCA1 and MCA2differently. Another explanation is that both proteins arenot properly regulated in yeast cells, which do not containintrinsic plant regulatory proteins.
The C-terminal half of MCA proteins has a cysteine-richdomain called the Plac8 or DUF614 motif. The function ofthis motif is unknown [41]. In general, the functions ofproteins containing this motif (the Plac8 super-family) arelargely unknown, but some are suggested to mediate cad-mium resistance [42,43] and are localized in plasma mem-brane microdomains [44,45].
All of the MCA proteins examined so far are localizedat the plasma membrane [14,15,37,38]. Some are local-ized at Hechtian strands or the plasma membrane–cellwall interface and at punctuated structures on the cellsurface [38], which may be related to plasma membranemicrodomains. Plant MCA proteins may form clusters orcomplexes with other signaling molecules via the Plac8motif.
MCA proteins mediate mechanical signal-inducedCa2+ transport and regulation of cell growth anddevelopmentSeveral lines of evidence suggest that the MCA proteinfunctions as a Ca2+-permeable MS channel component.Firstly, ectopic overexpression of MCA proteins increasedCa2+ uptake in Arabidopsis seedlings as well as in culturedrice cells, and enhanced the hypo-osmotic shock-inducedincrease in [Ca2+]cyt [14,15,38]. Secondly, hypo-osmoticshock-induced changes in [Ca2+]cyt and Ca2+ influx werepartially impaired in OsMCA1-suppressed cultured ricecells, whereas changes triggered by chitin fragments, amajor microbe-associated molecular pattern, were not af-fected [15]. Thirdly, a cell stretching-induced increase in[Ca2+]cyt was enhanced in Chinese hamster ovary cells het-erologously expressing MCA1 [14]. Fourthly, electrophysio-logical characterization of Xenopus oocytes expressingMCA1 suggested that MCA1 is a possible MS channel witha conductance of 34 pS [46]. MCA2 is also shown to generatemembrane stretch-activated currents in oocytes [46]. Lastly,whole-cell patch-clamp recordings on Arabidopsis mesophyllprotoplasts have shown that the anionic amphipath trini-trophenol, which penetrates the outer leaflet of the plasmamembrane and thus causes membrane distortion, inducesCa2+ currents more in MCA1-overexpressing protoplaststhan in their wild type counterparts [14].
In a knockout mutant of Arabidopsis MCA1 (but notMCA2), the roots were less able to penetrate a layer of hardagar, suggesting some defect in the mechanical response[14]. Rice OsMCA1-RNAi lines showed stunted growth andshortened rachises [15], which were reminiscent of mor-phological changes under drought stress [47]. OsMCA1may play a role in the drought stress response throughsensing changes in membrane distortion. Growth of theArabidopsis mca1 mca2 double mutant showed enhancedsensitivity to Mg2+ [37]. Growth of OsMCA1-suppressedcultured rice cells was significantly restricted under Ca2+
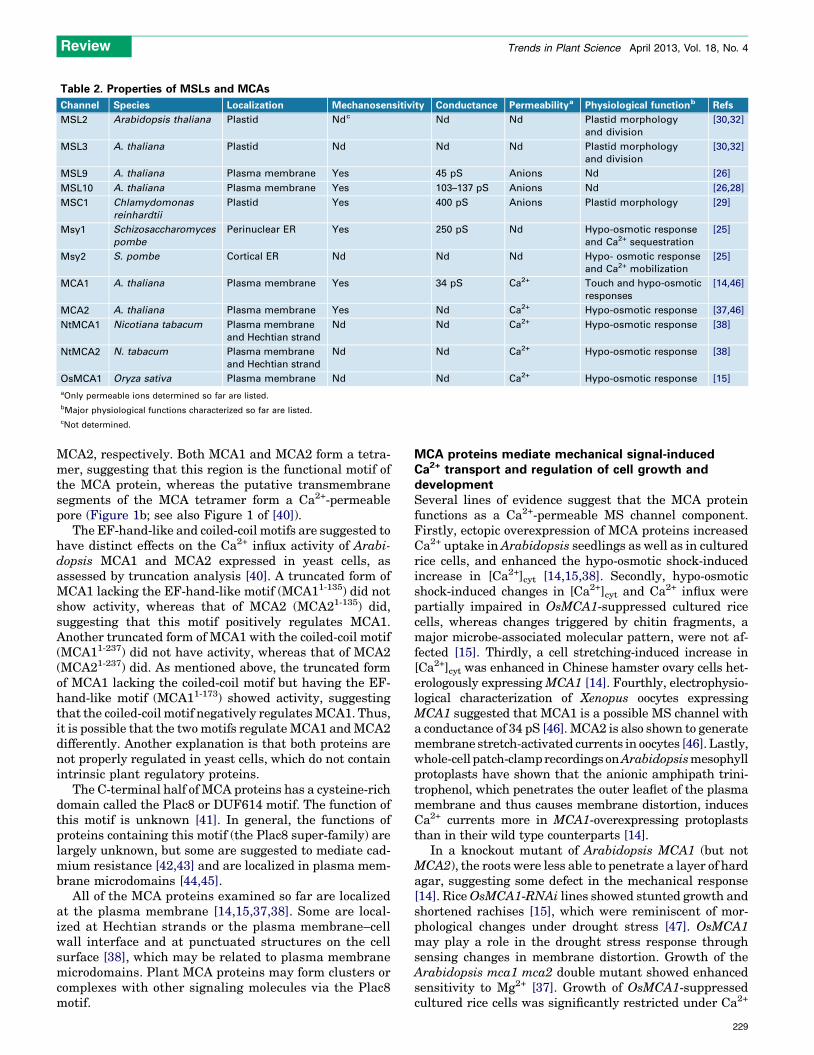
Table 2. Properties of MSLs and MCAs
Channel Species Localization Mechanosensitivity Conductance Permeabilitya Physiological functionb Refs
MSL2 Arabidopsis thaliana Plastid Ndc Nd Nd Plastid morphology
and division
[30,32]
MSL3 A. thaliana Plastid Nd Nd Nd Plastid morphology
and division
[30,32]
MSL9 A. thaliana Plasma membrane Yes 45 pS Anions Nd [26]
MSL10 A. thaliana Plasma membrane Yes 103–137 pS Anions Nd [26,28]
MSC1 Chlamydomonas
reinhardtii
Plastid Yes 400 pS Anions Plastid morphology [29]
Msy1 Schizosaccharomyces
pombe
Perinuclear ER Yes 250 pS Nd Hypo-osmotic response
and Ca2+ sequestration
[25]
Msy2 S. pombe Cortical ER Nd Nd Nd Hypo- osmotic response
and Ca2+ mobilization
[25]
MCA1 A. thaliana Plasma membrane Yes 34 pS Ca2+ Touch and hypo-osmotic
responses
[14,46]
MCA2 A. thaliana Plasma membrane Yes Nd Ca2+ Hypo-osmotic response [37,46]
NtMCA1 Nicotiana tabacum Plasma membrane
and Hechtian strand
Nd Nd Ca2+ Hypo-osmotic response [38]
NtMCA2 N. tabacum Plasma membrane
and Hechtian strand
Nd Nd Ca2+ Hypo-osmotic response [38]
OsMCA1 Oryza sativa Plasma membrane Nd Nd Ca2+ Hypo-osmotic response [15]
aOnly permeable ions determined so far are listed.
bMajor physiological functions characterized so far are listed.
cNot determined.
Review Trends in Plant Science April 2013, Vol. 18, No. 4
229

MCA 2M. esculenta
MCA 1
(OsMCA1: AB601973)
(MCA2: At2G17780)
0.05
S. bicolor
L. usita�ssimum
Z. mays
A. thaliana
A. lyrata MCA2C. rubella MCA2
A. lyrata MCA1C. rubella MCA1A. thaliana (MCA1: At4G35920)
B. rapa MCA1T. halophila MCA1B. napus
T. halophila MCA2
B. rapa MCA2
B. distachyonH. vulgare
T. aes�vum
S. italicaO. sa�va
P. sitchensis
P. patens MCA1P. patens MCA2
S. moellendorffii
N. tabacum(NtMCA1: AB6228 11)
S. lycopersicumM. esculenta
M. domes�caP. persica
C. si
nens
isC.
cle
men
�na
C. papaya
G. max MCA1
M. truncatula MCA1
M. truncatula MCA2P. vulgaris MCA2
G. max MCA3G. max MCA4
E. grandis
C. sa�vusA. coerulea
P. trichocarpa
G. max MCA2
N. tabacum(NtMCA2: AB622812)
Poaceae
Brassicaceae
Mosses and/or lycophytes
Most dicots
(a)
(b)
Plac8
Putaitve func�onal domain of MCA
TM
EF-hand-like Coiled-coil
MCA1 1MCA2 1
NtMCA1 1OsMCA1 1G. max 1
L. usitatissimum 1M. esculenta 1
P. trichocarpa 1E. grandis 1
P. sitchensis 1S. moellendorffii 1
P. patens 1
MCA1 121MCA2 121
NtMCA1 120OsMCA1 120G. max 120
L. usitatissimum 120M. esculenta 120
P. trichocarpa 120E. grandis 120
P. sitchensis 117S. moellendorffii 118
P. patens 119
MCA1 237MCA2 238
NtMCA1 236OsMCA1 235G. max 235
L. usitatissimum 236M. esculenta 236
P. trichocarpa 236E. grandis 238
P. sitchensis 232S. moellendorffii 235
P. patens 233
MCA1 346MCA2 338
NtMCA1 344OsMCA1 344G. max 341
L. usitatissimum 344M. esculenta 344
P. trichocarpa 344E. grandis 335
P. sitchensis 337S. moellendorffii 325
P. patens 339
MSHSWDGLGEIASVAQLTGLDAVKLIGLIVKAANTAWMHKKNCRQFAQHLKLIGNLLEQLKISEMKKYPETREPLEGLEDALRRSYLLVNSCRDRSYLYLLAMGWNIVYQFRKHQDEIDR 12 0MANSWDQLGEIASVAQLTGIDALKLIGMIVNAANTARMHKKNCRQFAHHLKLIRNLLEQIKNSEMNQRSEILEPLQGLDDALRRSYILVKSCQEKSYLYLLAMGWNIVNQFEKAQNEIDL 12 0MA-TWEHFGEVANFAQLAGLDAVRLIGMIVKAASTARMHKKNCRQFAQHLKLIGNLLEQLKITELKKYPETREPLEYLEDALRRSYMLVHSCHDRSYLYLLAMGWNIVYQFRKAQNEIDQ 11 9MA-SWENLGDVATVVQLTGLDAVRLISMIVKAASTARLHKRNCRRFAQHLKLIGGLLEQLRVSELKKYPETREPLEQLEDALRRAYLLVHSCQDRSYLYLLAMGWNIVYQFRKAQNEIDN 11 9MA-SWDQMGELANVAQLTGVDAVRLIGMIVRAASTARMHKKNCRQFAQHLKLIGNLLEQLKISELKKYPETREPLEQLEDALRRSYILVNSCQDRSYLYLLAMGWNIVYQFRKAQNEIDR 11 9MA-AWGNFGEIANVAQITGVDAVRLIGLIVKAASTARMHKKNCRQFAQHLKLIGGLLEQLKISELKKYPETREPLEQLEDSLRKSYLLVNSCQDRSYLYLLAMGWNIVYQFRKAQNEIDR 11 9MS-SWEHFGEIANVAQLTGVDAVRLIGMIVKAASTARMHKKNCRQFAQHLKLIGNLLEQLKISELKKYPETREPLEQLEDALRRSYILVNSCQDRSYLYLLAMGWNIVYQFRKAQNEIDR 11 9MA-TWEHLGEVANVVQLTGIDAVRLIAMIGKAATTARMHKKNCRQFAQHLKLIGNLLEQLKISELKRYPETREPLEQLEDALRRSYLLVNSCQDRSYLYLLAMGWNIVYQFRKAQNEIDR 11 9MA-TWEAFGEVANVAQLTGLDATRLIGMIVQAANTARMHKRNCRQFAQHLKLIGNLLEQLKISELKRYPETREPIEQLEDALRRSYILVNSCQDRSYLYLLAMGWNIVYQFRRAQNEIDR 11 9----WENVGDLANVTQLTGLNAVSLIALIVKAASNARMHKKNCRQFAQHLKLIGNLLEQLKATELKKYPETREPLEQLEDALRRSYVLVDSCQNRSYLYLLAMGWTIVNQFRQAHAEIDR 11 6MV-F--PVGDVATVAQIAGLDSLKLIAAVAAAAKNARMHKKNCRNFAQHLKLIGNLLEKLRLSELREHPETSEPLERLEEALRKAYILVNSCKNKSYLYLLAMGWNIVNQFKLRQAEIDR 11 7MP--WLALGDVASFGQLAGINAVQLIAMIVKAANNARMHKKNCRQFAQHLKLIANLLEQLNLTDLKERPECREPLEHLEQALRKAVVLVESCRDKSYLYLVAMGWMYVTKFRDYQDEIDK 11 8
FLKIIPLITLVDNARI--R-ERFEYIDRDQREYTLDEEDRHVQDVILKQESTRE-AASVLKKTLSCSYPNLRFCEALKTENEKLQIELQRSQEHYDVAQCEVIQRLIGVTQAAAAVEPDS 23 6FLKIVPLINMADNARI--R-ERLEAIERDQREYTLDEEDRKVQDVILKQESTREAATSVLKKTLSRSYPNMGFCEALKTEEEKLQLELQRSRARYDADQCEVIQRLIDVTQTAATVEPNL 23 7YLKIIPLITLVDNARV--R-ERFEIIEKDQCEYTLEAEDMKVQEVILKREPSKH-DTVVLKKTLSRSYPSMPINEAIQKENEKLQLELQRSQANLDVSHCEFIQHLLEVTEVVASESLSE 23 5YLRLVPLITLVDNARV--R-ERMEYIERDQCEYSFDDEDKEVQDALLNPDPSTN-PTVVLKKTLSCSYPNLPFNEALRKESEKLQVELQRSQSNMDMGQCEVIQHLLGVTKTVAS-SIPE 23 4YLRLVPLITLVDNARV--R-ERLEVIEMDQREYTLDDEDQKAQTVIFKPEPDKD-DTAVLKKTLSCSYPNCSFTEALKKENEKLKLELQRSQANLDMNQCEVIQRLLDVTEVAA-YSVPA 23 4YLRLIPLITLVDNTRI--R-DRLEYIERDQCEYTLDEEDRKVQDLILKSESVKD-QTVVLKKTLSCSYPNMCFNEALKKESEKLQIELQRSQSNMDVNPSEVIQHLLEVTEVAAASVVPE 23 5YLRLVPLITLVDNARV--R-ERLEDIQKDQHEYTLDEEDRKVQDVILKPEPLKD-QTIVLKKTLSSSYPNLGFNEALQKESQKLQLELQRSQANLDVKQCEVIQHLIDVTEAAAANSLPQ 23 5YLRLVPLITLVDNSRV--R-ERLEDIERDQREYTLDDEDRRVQDVILKPDCSGE-HTTMLKKTLSCSYPNMCFNEALRKENEKLQLELQRSQAHLDVNQCEVIQHLIEVTEVAAASSLPE 23 5YLKLVPLITLVDNARVRVR-ERLEVIERDQREYTLDDEDQKVQTVILTHEPSKN-DAMMLKKTLSCSYPNLCFNEALKKENEKLRMELQRSQTNMDMNQCEVIQRLLEVTETVASNSLPD 23 7YLKIVPLINLLDNHRV--K-DRLQAIEKDQREYTLDEEEEKVHDAILHPESSVL-NSSMLRKSLSRRYPYLGFEEALQKENEKLQVELHRSRANMDVDQCDVIRHLIEVTENVANI-PPE 23 1LLQIIPLISLVDNNRV--RWQQLREIQRDQKEYTLDEDELRLQETVLKPDLSVN-ESRELRRNLSRNYPGLALEEALRKENKKLKKELQKMQSLMEEDQCDVIRRLIDITETDVKGTYEK 23 4YLKLIPLISLVENS----R-ERIRAIVKDKRSYTMERSDVKVQETLLKPEHTRR-DSMRLSRQLSRRYPGMPLDIALREENAKLQKELEHMRACMELEKCGVIEHLIDFTEAAAADPVIL 23 2
EKELTKKASKKS-ERSSSMK-T---EYSYDEDSPK----KSSTRAASRSTSNVSSGHDLLSRR--ASQAQHHEEWHTDLLACCSEPSLCFKTFFFPCGTLAKIATAASNRHISSAEACNE 34 5EKVLTKK----E-ELTSSKK-R---DDLYDTDS-------SSIRADSRSTSYVSSGHELLSGR--SLQ--HRGNWHADLLDCCSEPCLCLKTLFFPCGTLAKISTVATSRQISSTEVCKN 33 7KSSPTKPTKKLE-HSHSDVN-SD--KEHNDRSYTK----SDEKQSTSRITSSVSSQRELLSSK--GSD--RYDEWHSDLLGCCSEPLLCIKTCFFPCGTFSKVASAAADRHISSADACNE 34 3KCATPKVSEKAD-SNHTKVS-EDSAKTYHDDSPKK----QKDACTAPRSSPPSSYGHDLVSSR--GSY--S-DEWHADLLGCCSEPSLCLRTFFFPCGTFSKIASIAKNRPMSSSEACND 34 3KCSPEKSHKKEE-YNYSDANSDK--DHSSDEKYHA----KIDKHSPSRYSV---AQKDLASTG--GSY--QQEDWHTDLLACCSEPSLCMKTFFYPCGTFSKIASVARNRPISSGEACND 34 0KRSSKTLSKKLE-PNYSDMN-VK--EHTYDDSYSR----KSDSRTISRDTSSVSSGQDLLSRK--GSS--HDEEWQTDLLGCCSEPLLCVKTLFFPCGTFSKIATVTSNRHMASAEACNE 34 3KTSPMKASKKLE-SNNSDVS-EK--NDSSDENYPK----KSDSRTTSRNTSSFSSGDNLLSHR--NSY--QHEEWHSDLLGCCSEPSLCLKTFFYPCGTFSKIATVATNKHMSPAEACNE 34 3KSSSTKSSKKLE-PAYSDAS-EN--KHSFDDSYST----KSDSHKTSRNTSSVSSRDDLLSSR--GSH--QQEEWHADLLGCCSEPYLCIKTLFYPCGTFAKIATVAKNRHISSAEACNE 34 3YGSPVKGSKKKE-KYTSDADSDR--EQSTGETYP-------------NNSS---LDNDLLPSK--GSY--GHYEWHTDLLGCCSEPLLCIKTFFCPCDTFSKIATVATNRHMSKGEACND 33 4RKHFQHSNCEEN-HPCADCCEEQ--PSPMDGSYLR----KQD----GRQDSSISSGRDLISQR--GSH--RYEEWKTDLFGCCMEPYLCFKTCIYPCGTFSNIAAVASNGKISPEQACND 33 6DPDYMKEAKKYEQSNYEQSN---------SDAYTY----Q-E--------S---HKSNETAS---KSW--QLQDWHHDLYGCCGSPLLCVGTFLCPCCTFATVAATATNGIMPKHAACTN 32 4AEAMAEIPEE--------DN-EE--EERHRKPHTRTEVVESSKRHMKHLKPSFST-LSLSSQKVVQCR--HEDNWRYDLFDCCVDPCLCIETFCYPCGTFTLVASSVTDGGTSEDSACSQ 33 8
LMAYSLILSCCCYTCCVRRKLRKTLNITGGFIDDFLSHVMCCCCALVQELREVEIRGAY--GTEK--TKISP-PSSQFMEH 42 1LIVYSLILSCCCYTCCIRKKLRKTLNITGGCIDDFLSHLMCCCCALVQELREVEIHRASYAGTEKSNKEMSP-PTPQFMEE 41 7LMAYSLILSCCCYTCCIRKKLRKKLNITGGIIDDFLSHLMCCCCALVQELREVEIRGAH--GTGK--TKTSP-PPSQFMES 41 9IMAYSLILSCCCYTCCVRRKLRQKLNIAGGCIDDFLSHLMCCCCALVQEWREVEIRGAY--G-ER--TKISP-PSFQYMEH 41 8LMAYSLILSCCCYTCCVRRKLRKMLNITGGFIDDFLSHLMCCCCALVQEWREVEIRGLS--G-SK--TKTSP-PPSQYMES 41 5LMAYSLILSCCCYTCCIRRKLRNMFNISGGFIDDFLSHLMCCCCALVQEWREVEIRGAY--G-QK--TKTSP-PVSQSME- 41 7LMAYSLILSCCCYTCCIRRKLRKTLNISGGFVDDFLSHLMCCCCALVQEWREVETRGVY--GPEK--TKTSP-PPSQFMES 41 9LMAYSMMLSCCCYTCCVRRELRKTLNITGGFIDDFLSHLMCCCCALVQEWREVEIRGVY--GPEK--TKTSP-PPSQFMES 41 9LLAYSLILSCCCYTCCVRRKLRKMLNI-GGWFDDFLSHFMCCCCALVQEWREVEIRGLS--G------------------K 39 4LMTYSLVFGCCCYTCCMRRKLRKLLNIAGGMCDDFLTHMTCCCCALVQEWREIECRGLD--DSHM--KKMSP-PPSQQME- 41 1CLIYTMVLSCCFYTCCFRRKLRKLYNIEGGSCDDCWAHFLCFCCALVQEAREIKARERD--GGNC--H------TLLLFIT 39 5LAFHSLYGGCCCYTSCIRRKVRRRFDIPGDCFSDYWAHVCCCCCAVLQELHELRFREKQ--GKYT--TQFFLYSLSGMQNV 41 5
TRENDS in Plant Science
L. japonicus P. vulgarisMCA1
Figure 1. Phylogenetic study and domain structure of the mid1-complementing activity (MCA) protein family. (a) A phylogenetic tree of the MCA protein family based on
amino acid sequences of the MCA functional domain constructed using the MEGA5.1 software by the Neighbor-Joining method. The alignment was performed by MTRAP
[67]. The scale bar (0.05) represents the relative branch length. (b) Alignment of the MCA protein family from Arabidopsis thaliana, tobacco (Nicotiana tabacum), rice (Oryza
sativa), soybean (Glycine max), flax (Linum usitatissimum), cassava (Manihot esculenta), poplar (Populus trichocarpa), eucalyptus (Eucalyptus grandis), spruce (Picea
sitchensis), lycophyte (Selaginella moellendorffii), and moss (Physcomitrella patens). The black or gray boxes indicate identical or conserved residues in the whole
Review Trends in Plant Science April 2013, Vol. 18, No. 4
230
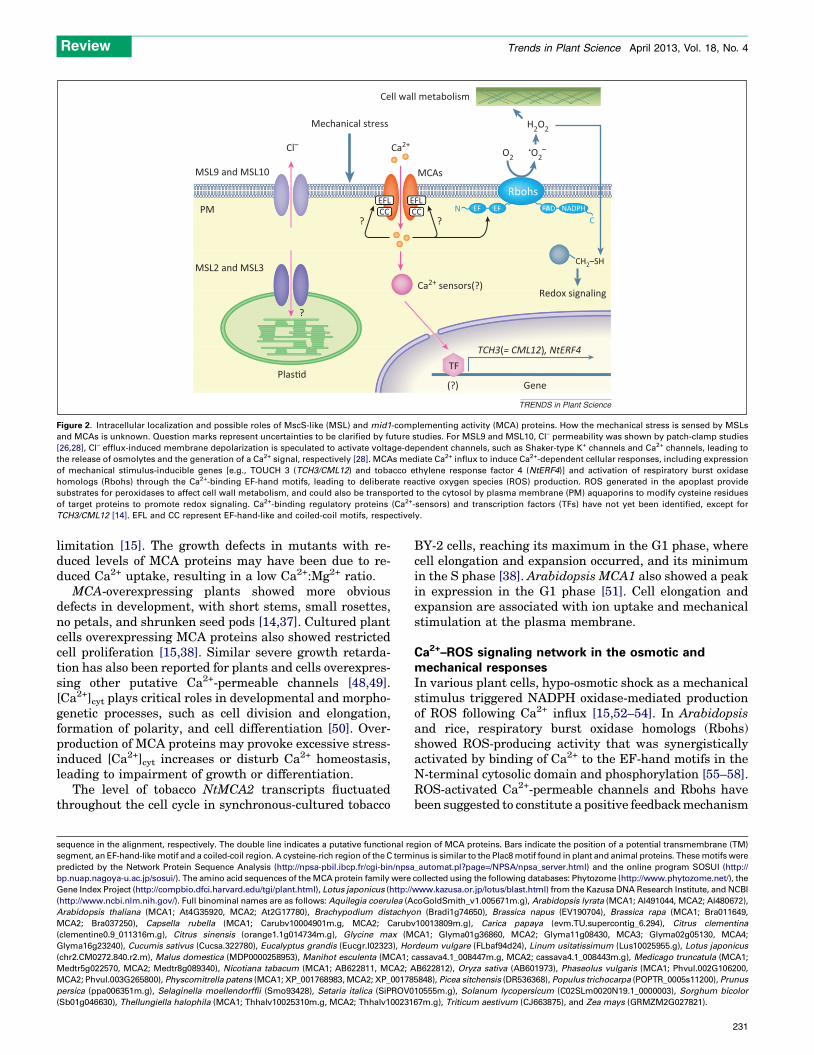
limitation [15]. The growth defects in mutants with re-duced levels of MCA proteins may have been due to re-duced Ca2+ uptake, resulting in a low Ca2+:Mg2+ ratio.
MCA-overexpressing plants showed more obviousdefects in development, with short stems, small rosettes,no petals, and shrunken seed pods [14,37]. Cultured plantcells overexpressing MCA proteins also showed restrictedcell proliferation [15,38]. Similar severe growth retarda-tion has also been reported for plants and cells overexpres-sing other putative Ca2+-permeable channels [48,49].[Ca2+]cyt plays critical roles in developmental and morpho-genetic processes, such as cell division and elongation,formation of polarity, and cell differentiation [50]. Over-production of MCA proteins may provoke excessive stress-induced [Ca2+]cyt increases or disturb Ca2+ homeostasis,leading to impairment of growth or differentiation.
The level of tobacco NtMCA2 transcripts fluctuatedthroughout the cell cycle in synchronous-cultured tobacco
BY-2 cells, reaching its maximum in the G1 phase, wherecell elongation and expansion occurred, and its minimumin the S phase [38]. Arabidopsis MCA1 also showed a peakin expression in the G1 phase [51]. Cell elongation andexpansion are associated with ion uptake and mechanicalstimulation at the plasma membrane.
Ca2+–ROS signaling network in the osmotic andmechanical responsesIn various plant cells, hypo-osmotic shock as a mechanicalstimulus triggered NADPH oxidase-mediated productionof ROS following Ca2+ influx [15,52–54]. In Arabidopsisand rice, respiratory burst oxidase homologs (Rbohs)showed ROS-producing activity that was synergisticallyactivated by binding of Ca2+ to the EF-hand motifs in theN-terminal cytosolic domain and phosphorylation [55–58].ROS-activated Ca2+-permeable channels and Rbohs havebeen suggested to constitute a positive feedback mechanism
sequence in the alignment, respectively. The double line indicates a putative functional region of MCA proteins. Bars indicate the position of a potential transmembrane (TM)
segment, an EF-hand-like motif and a coiled-coil region. A cysteine-rich region of the C terminus is similar to the Plac8 motif found in plant and animal proteins. These motifs were
predicted by the Network Protein Sequence Analysis (http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_server.html) and the online program SOSUI (http://
bp.nuap.nagoya-u.ac.jp/sosui/). The amino acid sequences of the MCA protein family were collected using the following databases: Phytozome (http://www.phytozome.net/), the
Gene Index Project (http://compbio.dfci.harvard.edu/tgi/plant.html), Lotus japonicus (http://www.kazusa.or.jp/lotus/blast.html) from the Kazusa DNA Research Institute, and NCBI
(http://www.ncbi.nlm.nih.gov/). Full binominal names are as follows: Aquilegia coerulea (AcoGoldSmith_v1.005671m.g), Arabidopsis lyrata (MCA1; Al491044, MCA2; Al480672),
Arabidopsis thaliana (MCA1; At4G35920, MCA2; At2G17780), Brachypodium distachyon (Bradi1g74650), Brassica napus (EV190704), Brassica rapa (MCA1; Bra011649,
MCA2; Bra037250), Capsella rubella (MCA1; Carubv10004901m.g, MCA2; Carubv10013809m.g), Carica papaya (evm.TU.supercontig_6.294), Citrus clementina
(clementine0.9_011316m.g), Citrus sinensis (orange1.1g014734m.g), Glycine max (MCA1; Glyma01g36860, MCA2; Glyma11g08430, MCA3; Glyma02g05130, MCA4;
Glyma16g23240), Cucumis sativus (Cucsa.322780), Eucalyptus grandis (Eucgr.I02323), Hordeum vulgare (FLbaf94d24), Linum usitatissimum (Lus10025955.g), Lotus japonicus
(chr2.CM0272.840.r2.m), Malus domestica (MDP0000258953), Manihot esculenta (MCA1; cassava4.1_008447m.g, MCA2; cassava4.1_008443m.g), Medicago truncatula (MCA1;
Medtr5g022570, MCA2; Medtr8g089340), Nicotiana tabacum (MCA1; AB622811, MCA2; AB622812), Oryza sativa (AB601973), Phaseolus vulgaris (MCA1; Phvul.002G106200,
MCA2; Phvul.003G265800), Physcomitrella patens (MCA1; XP_001768983, MCA2; XP_001785848), Picea sitchensis (DR536368), Populus trichocarpa (POPTR_0005s11200), Prunus
persica (ppa006351m.g), Selaginella moellendorffii (Smo93428), Setaria italica (SiPROV010555m.g), Solanum lycopersicum (C02SLm0020N19.1_0000003), Sorghum bicolor
(Sb01g046630), Thellungiella halophila (MCA1; Thhalv10025310m.g, MCA2; Thhalv10023167m.g), Triticum aestivum (CJ663875), and Zea mays (GRMZM2G027821).
Cell wall metabolism
Mechanical stress
Cl–
MSL9 and MSL10
MSL2 and MSL3
Plas�d(?)
TF
Gene
TCH3(= CML12), NtERF4
PM
?
? ?
MCAs
EFLCC CC
EFL
Ca2+O2
H2O2
•O2–
Ca2+ sensors(?)
CH2–SH
CN NADPHFADEFEF
Rbohs
Redox signaling
TRENDS in Plant Science
Figure 2. Intracellular localization and possible roles of MscS-like (MSL) and mid1-complementing activity (MCA) proteins. How the mechanical stress is sensed by MSLs
and MCAs is unknown. Question marks represent uncertainties to be clarified by future studies. For MSL9 and MSL10, Cl– permeability was shown by patch-clamp studies
[26,28], Cl– efflux-induced membrane depolarization is speculated to activate voltage-dependent channels, such as Shaker-type K+ channels and Ca2+ channels, leading to
the release of osmolytes and the generation of a Ca2+ signal, respectively [28]. MCAs mediate Ca2+ influx to induce Ca2+-dependent cellular responses, including expression
of mechanical stimulus-inducible genes [e.g., TOUCH 3 (TCH3/CML12) and tobacco ethylene response factor 4 (NtERF4)] and activation of respiratory burst oxidase
homologs (Rbohs) through the Ca2+-binding EF-hand motifs, leading to deliberate reactive oxygen species (ROS) production. ROS generated in the apoplast provide
substrates for peroxidases to affect cell wall metabolism, and could also be transported to the cytosol by plasma membrane (PM) aquaporins to modify cysteine residues
of target proteins to promote redox signaling. Ca2+-binding regulatory proteins (Ca2+-sensors) and transcription factors (TFs) have not yet been identified, except for
TCH3/CML12 [14]. EFL and CC represent EF-hand-like and coiled-coil motifs, respectively.
Review Trends in Plant Science April 2013, Vol. 18, No. 4
231

to enhance both Ca2+ and ROS signals [56,57]. A functionalNADPH oxidase, AtRbohC/RHD2, affected mechanicalstress-induced ROS generation, and regulated root hairelongation in a Ca2+-dependent manner [16,56].
It is plausible that MCAs play a role in the regulation ofmechanical responses via signal transduction pathwaysdependent on Ca2+ and ROS. Overexpression of riceOsMCA1 enhanced NADPH oxidase-mediated ROS pro-duction in cultured cells [15]. Both Arabidopsis MCA1 andROS generated by AtRbohD and/or AtRbohF were sug-gested to be involved in regulating osmosensitive metabol-ic changes [59]. Apoplastic ROS production plays criticalroles in the modification of cell wall metabolism, includingcrosslinking to regulate its rigidity [60,61]. MCA1 wasrequired for cell wall damage-induced deposition of lignin,which confers mechanical strength to the cell wall [62].Expression of the Arabidopsis touch-inducible geneTOUCH 3 (TCH3/CML12), which encodes a calmodulin-like protein, and tobacco ethylene response factor 4(NtERF4), which is responsive to mechanical stress, areupregulated in the overexpressors of MCAs in Arabidopsisplants and tobacco BY-2 cells [14,38].
These findings suggest the following initial plasmamembrane signaling in response to osmotic and mechani-cal stimulation: activation of the plasma membrane MSCa2+-permeable channels, such as the MCA protein family,induces the influx of Ca2+, leading to activation of Rboh(s)through the Ca2+-binding EF-hand motifs to generate ROS(Figure 2). The signaling network dependent on Ca2+ andROS may play a crucial role in regulating downstreamevents, such as regulation of the mechanical properties ofthe cell wall, metabolic changes, and gene expression. ROSgenerated in the apoplast provide substrates for peroxi-dases to affect cell wall metabolism, such as crosslinking,and are also transported to the cytosol by plasma mem-brane aquaporins [63] to modify cysteine residues of targetproteins to promote redox signaling [64].
Concluding remarks and future perspectivesMechanical stresses greatly affect plant growth and de-velopment. To better understand such general phenome-na of life, it is crucial to understand the molecularmechanisms of mechanosensing and mechanotransduc-tion through elucidating the structure and function of MSchannels and their interacting molecules [9,65,66]. Re-cent molecular and genetic studies represent a new erafor studies on MS channels and their candidates, includ-ing MSLs and MCAs. For example, molecular characteri-zation of MSLs and MCAs would facilitate determinationof the structural basis of mechanosensory machineriesthat might comprise molecules in the cell wall and/orextracellular matrix, the plasma membrane, intracellu-lar regulatory proteins, cytoskeletons, and MS channelitself. Importantly, such studies would help elucidate as-yet-unknown mechanisms of how plant MS channelssense a variety of mechanical stresses. MS channels havebeen suggested to be mechanosensors that transduce theshear forces in the cell wall–membrane–cytoskeletonsystem [66]. We anticipate that the study of MSLs andMCAs will help decisively to unravel the mechanismsinvolved.
Both extracellular and intracellular mechanical stressescan be sensed via various types of MS channel. Indeed, someMSLs are specific to organellar membranes, whereas othersare localized in the plasma membrane. So far, plant MSLshave been studied only in Arabidopsis. It will be of greatinterest to investigate the MSLs of other plant species,including monocots. By contrast, all the MCA proteins ex-amined to date are localized in the plasma membrane andare involved in the hypo-osmotic response and probably alsoin the touch response. Other physical and chemical stresses,including gravity, might also activate MS channels. There-fore, not only MSLs and MCAs, but also the plant Piezoproteins should provide further clues for understanding howplants perceive and respond to such a variety of stresses.
AcknowledgmentsWe would like to thank Kazuo Shinozaki (RIKEN Plant Science Center,Yokohama, Japan) for his critical reading of the manuscript. This workwas supported in part by Grants-in-Aid for Scientific Research on PriorityArea No. 21026009 (to H.I.), 21117516 (to K.K.), 23117718 (to K.K.), andNo. 23120509 (to H.I.) from the Ministry of Education, Culture, Sports,Science, and Technology of Japan, a Grant-in-Aid for Scientific ResearchB No. 19370023 (to K.K.), 21370017 (to H.I.) and 23380027 (to K.K.) fromthe Japan Society for the Promotion of Science (JSPS), and a Grant-in-Aidfor JSPS Fellows No. 10J02008 (to Y.N.).
References1 Darwin, C. (1865) On the movements and habits of climbing plants.
J. Linn. Soc. Lond. (Bot.) 9, 1–1182 Jaffe, M.J. (1973) Thigmomorphogenesis: the response of plant growth
and development to mechanical stimulation. Planta 114, 143–1573 Chehab, E.W. et al. (2009) Thigmomorphogenesis: a complex plant
response to mechano-stimulation. J. Exp. Bot. 60, 43–564 Braam, J. (2005) In touch: plant responses to mechanical stimuli. New
Phytol. 165, 373–3895 Escalante-Perez, M. et al. (2011) A special pair of phytohormones
controls excitability, slow closure, and external stomach formationin the Venus flytrap. Proc. Natl. Acad. Sci. U.S.A. 108, 15492–15497
6 Trewavas, A. and Knight, M. (1994) Mechanical signalling, calcium andplant form. Plant Mol. Biol. 26, 1329–1341
7 Jaffe, M.J. et al. (2002) Thigmo responses in plants and fungi. Am. J.Bot. 89, 375–382
8 Telewski, F.W. (2006) A unified hypothesis of mechanoperception inplants. Am. J. Bot. 93, 1466–1476
9 Monshausen, G.B. and Gilroy, S. (2009) Feeling green: mechanosensingin plants. Trends Cell Biol. 19, 228–235
10 Knight, M.R. et al. (1991) Transgenic plant aequorin reports the effectsof touch and cold-shock and elicitors on cytoplasmic calcium. Nature352, 524–526
11 Plieth, C. and Trewavas, A.J. (2002) Reorientation of seedlings in theearth’s gravitational field induces cytosolic calcium transients. PlantPhysiol. 129, 786–796
12 Toyota, M. et al. (2008) Cytoplasmic calcium increases in response tochanges in the gravity vector in hypocotyls and petioles of Arabidopsisseedlings. Plant Physiol. 146, 505–514
13 Martinac, B. et al. (1990) Mechanosensitive ion channels of E. coliactivated by amphipaths. Nature 348, 261–263
14 Nakagawa, Y. et al. (2007) Arabidopsis plasma membrane proteincrucial for Ca2+ influx and touch sensing in roots. Proc. Natl. Acad.Sci. U.S.A. 104, 3639–3644
15 Kurusu, T. et al. (2012) Plasma membrane protein OsMCA1 is involvedin regulation of hypo-osmotic shock-induced Ca2+ influx and modulatesgeneration of reactive oxygen species in cultured rice cells. BMC PlantBiol. http://dx.doi.org/10.1186/1471-2229-12-11
16 Monshausen, G.B. et al. (2009) Ca2+ regulates reactive oxygen speciesproduction and pH during mechanosensing in Arabidopsis roots. PlantCell 21, 2341–2356
17 Braam, J. and Davis, R.W. (1990) Rain-, wind-, and touch-inducedexpression of calmodulin and calmodulin-related genes in Arabidopsis.Cell 60, 357–364
Review Trends in Plant Science April 2013, Vol. 18, No. 4
232

18 Lee, J.S. et al. (1983) Reversible loss of gravitropic sensitivity in maizeroots after tip application of calcium chelators. Science 220, 1375–1376
19 Fujita, Y. et al. (2011) ABA-mediated transcriptional regulation inresponse to osmotic stress in plants. J. Plant Res. 124, 509–525
20 Pivetti, C.D. et al. (2003) Two families of mechanosensitive channelproteins. Microbiol. Mol. Biol. Rev. 67, 66–85
21 Haswell, E.S. et al. (2011) Mechanosensitive channels: what can theydo and how do they do it? Structure 19, 1356–1369
22 Hurst, A.C. et al. (2008) MscS, the bacterial mechanosensitive channelof small conductance. Int. J. Biochem. Cell Biol. 40, 581–585
23 Jensen, G.S. and Haswell, E.S. (2012) Functional analysis of conservedmotifs in the mechanosensitive channel homolog MscS-like2 fromArabidopsis thaliana. PLoS ONE 7, e40336
24 Haswell, E.S. (2007) MscS-like proteins in plants. Curr. Top. Membr.58, 329–359
25 Nakayama, Y. et al. (2012) Organellar mechanosensitive channels infission yeast regulate the hypo–osmotic shock response. Nat. Commun.http://dx.doi.org/10.1038/ncomms2014
26 Haswell, E.S. et al. (2008) Two MscS homologs providemechanosensitive channel activities in the Arabidopsis root. Curr.Biol. 18, 730–734
27 Peyronnet, R. et al. (2008) AtMSL9 and AtMSL10: sensors of plasmamembrane tension in Arabidopsis roots. Plant Signal. Behav. 3,726–729
28 Maksaev, G. and Haswell, E.S. (2012) MscS-Like10 is a stretch-activated ion channel from Arabidopsis thaliana with a preferencefor anions. Proc. Natl. Acad. Sci. U.S.A. 109, 19015–19020
29 Nakayama, Y. et al. (2007) Molecular and electrophysiologicalcharacterization of a mechanosensitive channel expressed in thechloroplasts of Chlamydomonas. Proc. Natl. Acad. Sci. U.S.A. 104,5883–5888
30 Haswell, E.S. and Meyerowitz, E.M. (2006) MscS-like proteinscontrol plastid size and shape in Arabidopsis thaliana. Curr. Biol.16, 1–11
31 Veley, K.M. et al. (2012) Mechanosensitive channels protect plastidsfrom hypoosmotic stress during normal plant growth. Curr. Biol. 22,408–413
32 Wilson, M.E. et al. (2011) Two mechanosensitive channel homologsinfluence division ring placement in Arabidopsis chloroplasts. PlantCell 23, 2939–2949
33 Coste, B. et al. (2012) Piezo proteins are pore-forming subunits ofmechanically activated channels. Nature 483, 176–181
34 Kim, S.E. et al. (2012) The role of Drosophila Piezo in mechanicalnociception. Nature 483, 209–212
35 Coste, B. et al. (2010) Piezo1 and Piezo2 are essential components ofdistinct mechanically activated cation channels. Science 330, 55–60
36 Kanzaki, M. et al. (1999) Molecular identification of a eukaryotic,stretch-activated nonselective cation channel. Science 285, 882–886[report clarification in Science (2000) 288, 1374]
37 Yamanaka, T. et al. (2010) MCA1 and MCA2 that mediate Ca2+ uptakehave distinct and overlapping roles in Arabidopsis. Plant Physiol. 152,1284–1296
38 Kurusu, T. et al. (2012) Involvement of the putative Ca2+-permeablemechanosensitive channels, NtMCA1 and NtMCA2, in Ca2+ uptake,Ca2+-dependent cell proliferation and mechanical stress-induced geneexpression in tobacco (Nicotiana tabacum) BY-2 cells. J. Plant Res. 125,555–568
39 Kurusu, T. et al. (2012) Roles of a putative mechanosensitive plasmamembrane Ca2+-permeable channel OsMCA1 in generation of reactiveoxygen species and hypo-osmotic signaling in rice. Plant Signal. Behav.7, 796–798
40 Nakano, M. et al. (2011) Determination of structural regions importantfor Ca2+ uptake activity in Arabidopsis MCA1 and MCA2 expressed inyeast. Plant Cell Physiol. 52, 1915–1930
41 Galaviz-Hernandez, C. et al. (2003) Plac8 and Plac9, novel placental-enriched genes identified through microarray analysis. Gene 309,81–89
42 Song, W.Y. et al. (2004) A novel family of cys-rich membrane proteinsmediates cadmium resistance in Arabidopsis. Plant Physiol. 135,1027–1039
43 Tanaka, S. et al. (2007) A novel gene with antisalt and anticadmiumstress activities from a halotolerant marine green alga Chlamydomonassp W80. FEMS Microbiol. Lett. 271, 48–52
44 Iomini, C. et al. (2006) Two flagellar genes, AGG2 and AGG3, mediateorientation to light in Chlamydomonas. Curr. Biol. 16, 1147–1153
45 Lefebvre, B. et al. (2007) Characterization of lipid rafts from Medicagotruncatula root plasma membranes: a proteomic study reveals thepresence of a raft-associated redox system. Plant Physiol. 144, 402–418
46 Furuichi, T. et al. (2012) Expression of Arabidopsis MCA1 enhancedmechanosensitive channel activity in the Xenopus laevis oocyte plasmamembrane. Plant Signal. Behav. 7, 1022–1026
47 Ji, X.M. et al. (2005) Tissue-specific expression and droughtresponsiveness of cell-wall invertase genes of rice at flowering. PlantMol. Biol. 59, 945–964
48 Kim, S.A. et al. (2001) Overexpression of the AtGluR2 gene encoding anArabidopsis homolog of mammalian glutamate receptors impairscalcium utilization and sensitivity to ionic stress in transgenicplants. Plant Cell Physiol. 42, 74–84
49 Kurusu, T. et al. (2004) Identification of a putative voltage-gated Ca2+-permeable channel (OsTPC1) involved in Ca2+ influx and regulation ofgrowth and development in rice. Plant Cell Physiol. 45, 693–702
50 White, P.J. and Broadley, M.R. (2003) Calcium in plants. Ann. Bot. 92,487–511
51 Menges, M. et al. (2002) Cell cycle-regulated gene expression inArabidopsis. J. Biol. Chem. 277, 41987–42002
52 Beffagna, N. et al. (2005) Modulation of reactive oxygen speciesproduction during osmotic stress in Arabidopsis thaliana culturedcells: involvement of the plasma membrane Ca2+-ATPase and H+-ATPase. Plant Cell Physiol. 46, 1326–1339
53 Hayashi, T. et al. (2006) Ca2+ transient induced by extracellular changesin osmotic pressure in Arabidopsis leaves: differential involvement of cellwall-plasma membrane adhesion. Plant Cell Environ. 29, 661–672
54 Cazale, A.C. et al. (1998) Oxidative burst and hypoosmotic stress intobacco cell suspensions. Plant Physiol. 116, 659–669
55 Ogasawara, Y. et al. (2008) Synergistic activation of the ArabidopsisNADPH oxidase AtrbohD by Ca2+ and phosphorylation. J. Biol. Chem.283, 8885–8892
56 Takeda, S. et al. (2008) Local positive feedback regulation determinescell shape in root hair cells. Science 319, 1241–1244
57 Kimura, S. et al. (2012) Protein phosphorylation is a prerequisite forthe Ca2+-dependent activation of Arabidopsis NADPH oxidases andmay function as a trigger for the positive feedback regulation of Ca2+
and reactive oxygen species. Biochim. Biophys. Acta 1823, 398–40558 Takahashi, S. et al. (2012) Reactive oxygen species production and
activation mechanism of the rice NADPH oxidase OsRbohB. J.Biochem. 152, 37–43
59 Wormit, A. et al. (2012) Osmosensitive changes of carbohydratemetabolism in response to cellulose biosynthesis inhibition. PlantPhysiol. 159, 105–117
60 Brady, J.D. and Fry, S.C. (1997) Formation of di-isodityrosine and lossof isodityrosine in the cell walls of tomato cell-suspension culturestreated with fungal elicitors or H2O2. Plant Physiol. 115, 87–92
61 O’Brien, J.A. et al. (2012) Reactive oxygen species and their role inplant defence and cell wall metabolism. Planta 236, 765–779
62 Denness, L. et al. (2011) Cell wall damage-induced lignin biosynthesisis regulated by a reactive oxygen species- and jasmonic acid-dependentprocess in Arabidopsis. Plant Physiol. 156, 1364–1374
63 Dynowski, M. et al. (2008) Plant plasma membrane water channelsconduct the signalling molecule H2O2. Biochem. J. 414, 53–61
64 Spoel, S.H. and Loake, G.J. (2011) Redox-based protein modifications:the missing link in plant immune signalling. Curr. Opin. Plant Biol. 14,358–364
65 Perbal, G. and Driss-Ecole, D. (2003) Mechanotransduction ingravisensing cells. Trends Plant Sci. 8, 498–504
66 Ding, J.P. and Pickard, B.G. (1993) Mechanosensory calcium-selectivecation channels in epidermal cells. Plant J. 3, 83–110
67 Hara, T. et al. (2010) MTRAP: pairwise sequence alignment algorithmby a new measure based on transition probability between twoconsecutive pairs of residues. BMC Bioinformatics http://dx.doi.org/10.1186/1471-2105-11-235
Review Trends in Plant Science April 2013, Vol. 18, No. 4
233




![Le Poil & la Plume [BaT-BR].indd 1 22/07/11 14:36 · Anny Duperey Le Poil et la Plume récit Éditions du Seuil 25, bd Romain-Rolland, Paris XIVe Le Poil & la Plume [BaT-BR].indd](https://static.fdocuments.fr/doc/165x107/60e594b7409d3f2ec665e41d/le-poil-la-plume-bat-brindd-1-220711-1436-anny-duperey-le-poil-et-la.jpg)












