![Costa rica [enregistrement automatique] [enregistrement automatique]](https://static.fdocuments.fr/doc/165x107/557d7c91d8b42a2c428b4ffd/costa-rica-enregistrement-automatique-enregistrement-automatique.jpg)
Costa rica [enregistrement automatique] [enregistrement automatique]
Upload
lehoux-hugoCategory
view
220download
0description

LEHOUX Hugo
9, rue Duboys des Sauzais
35150 Corps-Nuds FRANCE
« Limnologie et entomologie tropicales » Pré-projets pour le monitoring des rivières de la région d’Esquinas
Costa Rica
Tuteur entreprise Tuteur IUT Friedrich SCHIEMER David LANDRY Département Limnologie et hydro-botanique Maître de conférences Université de Vienne, Autriche Université/IUT, France
Entreprise Centre de recherche Biologique Tropicale
La Gamba, Parque nacional de las Piedras Blancas
Costa-Rica

Remerciements
Je tiens à remercier énormément toutes les personnes qui ont participé à la réalisation de ce
rapport, de près ou de loin, à commencer par M. Friedrich SCHIEMER mon tuteur de stage, pour m’avoir
offert la possibilité d’effectuer cette étude. Egalement toute l’équipe de la Tropenstation, qui m’a réservée
un accueil chaleureux en tout temps et tout particulièrement Victor, Eduardo, Roy, Mari, Eduvijes, Lizbeth,
Beatriz, Vanessa, Luiz, Daniel et Paul. Je remercie humblement les étudiants et chercheurs qui ont séjourné
à la station, et qui m’ont donné leur avis et conseillé sur le déroulement du stage et tout particulièrement
à Laura RIBA-HERNANDEZ (Université de São Paulo) et Dangelo SANDOVAL (CATIE), pour la partie
géologique. Un grand remerciement pour leur participation, à Frank SUHLING, Carlos ESQUIVEL et Rodolfo
NOVELLO-GUTTIEREZ, experts en Odonatologie tropicale, sans qui les déterminations seraient restées
parfois approximatives. Un grand merci au MINAE pour m’avoir autorisé à mener cette étude et m’avoir
délivré le précieux permis scientifique. Je remercie également l’Université, le conseil général d’Ille et
Vilaine (Jeunes à travers le monde), ainsi que la mairie de la commune de Corps-Nuds, pour leur soutien
matériel et financier, nécessaire à la bonne réalisation de ce stage.
Sans oublier toute l’équipe pédagogique de l’IUT, et plus particulièrement mon tuteur David LANDRY pour
le soutien important apporté durant le stage.
Un grand merci à ma famille et mes proches, qui m’ont permis de réaliser ce stage dans les
meilleures conditions possibles.

Sommaire
Introduction 1
1) Présentation de l’environnement écologique, économique et social de la station 2
1.1 ) La station de recherche tropicale La Gamba 2
1.2 ) Présentation du tuteur 3
1.3 ) Le Costa-Rica 4
1.3.1) Généralités 4
1.3.2) Situation du secteur agricole 4
1.3.3) Déforestation, mise en culture de terres et qualité de l’eau 5
2) Problématique et axes de recherche 6
2.1) Problématique de stage 6
2.2) Aires d’étude 7
2.2.1) Zones de relevés limnologiques 7
2.2.2) Zones de relevés entomologiques 8
2.3) Matériel et méthodes 9
2.3.1) Limnologie 9
2.3.2) Entomologie 11
2.3.4) Statistiques 12
3) Résultats 13
3.1) Limnologie 13
3.1.1) Généralités 13
3.1.2) Quebrada Negra 15
3.2) Entomologie 20
3.2.1) Alpha et gamma diversités 20
3.2.2) Beta diversité 23

4) Discussion 27
4.1) Limnologie 27
4.2) Entomologie 28
4.2.1) Richesse en espèces 28
4.2.2) Similarités 29
4.3) Limites de l’étude 30
4.4) Les Odonates comme bio-indicateurs 31
5) Bilan personnel 33
Bibliographie 34
Annexes 38

1
Introduction
La problématique universelle de la qualité de
l’eau, intéresse les politiques du Costa-Rica depuis
déjà une vingtaine d’année. Un rapport de 1990,
soulignait déjà les problèmes que posent la mise en
place d’un plan national de gestion de la qualité des
eaux (CALVO C., 1990). La création, cette même
année, du Ministère des Ressources Nationales et du
Développement Durable a permis d’établir une
stratégie nationale qui tend à répondre à une
demande croissante, qui a quadruplé ces quarante
dernières années.
La gestion de la quantité n’est pas un
problème en soi pour le pays (3ème pays d’Amérique
centrale, en terme d’offre hydrique par habitant),
cependant la qualité des eaux tend à se détériorer
avec le développement du tourisme, nouvelles
technologies agricoles et industries (Rapport exécutif
de 2004).
Le secteur agricole est le deuxième plus gros
utilisateur d’eau, après la production d’hydro-énergie.
Si on tient compte du fait que l’hydro-énergie reverse
sans modification, l’eau dans le système
hydrologique, l’agriculture est réellement le premier
consommateur d’eau, avec 28.3% du total de l’offre
d’eau superficielle (FAO 2009). Les déforestations
massives et l’intensification des cultures telles que
l’ananas (Ananas comosus), la palme à huile (Elays
guineensis) ou encore le riz (Oryza sp.), favorisent la
dégradation des masses d’eau, qui peuvent
rapidement avoir une influence sur la santé de la
population, de l’économie du pays et pourrait
engendrer d’importants problèmes sociaux. Les
travaux sur la reforestation menés par la station
biologique tropicale la Gamba, entrent en accord avec
la nécessité de préserver cet environnement
écologique, social et économique.
Dans le cadre d’une collaboration avec une
association brésilienne « P P bio Manaos », la station
s’est engagée à effectuer une étude complète sur les
neuf cours d’eau du Bassin versant d’Esquinas (La
« Cuenca Esquinas » contient La Quebrada Negra,
Q.Mari, Q.Chorro, Q.Gamba, Q.Bolsa, Q. Sardinal, Rio
Oro, R.Bonito, R. Esquinas). L’objectif d’une telle
étude est de connaître la structure fluviale et la bio-
structure de régions néo-tropicales, pour comprendre
leur fonctionnement, et pouvoir installer un
« monitoring » permanent d’indicateurs de bonne
santé écologique.
Qu’est-ce qu’un monitoring ?
La création d’un monitoring signifie la mise en
place d’un suivi des paramètres physico-chimiques
(concentrations en solutés…), des êtres vivants
(macro-invertébrés…) ainsi que des phénomènes
(décomposition de la matière organique…) qui se
rattachent aux cours d’eau du bassin versant. Il
permet d’évaluer la qualité écologique d’un système
hydrologique.

2
1) Présentation de l’environnement écologique, économique et social de la station.
1.1) La station de recherche tropicale La Gamba
La station de recherche biologique tropicale
de La Gamba est située dans la Parc National de Las
Piedras Blancas, dans le Sud-Ouest du Costa-Rica.
Celle-ci a une histoire un peu particulière qui a, en
partie, participé au choix du lieu de stage.
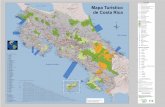
Fig. 1 Carte du Costa-rica , réalisée avec ArcGIS.
En 1991, Michael Schnitzer, professeur de
musique et d’arts à l’université de Vienne, fonda
l’association « Rainforest of the Austrians ». Durant
plusieurs années, il a collecté des fonds importants en
Autriche, pour les reverser au gouvernement du
Costa-Rica. Ce dernier racheta des terres, qu’il plaça
sous la tutelle du Parc National de « Las Piedras
Blancas », dans l’objectif de préserver la forêt
Esquinas, sur la côte Pacifique ouest du Costa-Rica.
Cet endroit est rapidement devenu une zone
privilégiée pour promouvoir des programmes de
recherche scientifique.
Afin que l’accès à cette zone soit facilité,
l’association acheta en 1993, une vieille ferme (une
« finca »). Avec l’aide du ministère de la science et de
la recherche, et de l’université de Vienne, le noyau a
commencé à se durcir près du parc national et de la
ville paisible de La Gamba. Aujourd’hui la station peut
accueillir jusqu’à 30 personnes, et dispose d’un jardin
botanique impressionnant. En 2008, fut installé le
laboratoire avec air conditionné, et tout le matériel
nécessaire pour effectuer quelques analyses basiques.
Depuis sa création, la station s’attache à découvrir et
à préserver les forêts néotropicales pluvieuses
(Neotropical rainforest), et à reforester les zones
saccagées.
Au-delà des aspects de recherches, la station
participe au développement de la Gamba, employant
plus d’une quinzaine de locaux à l’année (Annexe 1),
et offre tous les fruits du jardin à l’école de la ville.
L’association a également créé un hôtel à proximité
de la station, le «Esquinas Rainforest Lodge hotel »,
qui permet de développer l’éco-tourisme dans la
région (Certifié « Three-leaf Certificate of Sustainable
Tourism »).
Photographie 1 Vue de l'entrée de la Tropenstation La Gamba, Costa Rica

3
Le diagramme météorologique ci-contre, résume
les conditions météorologiques particulières de la région.
La partie en noir illustre la pluviométrie de l’année 2009,
qui atteint 5816mm, pour une moyenne de température
de 27,9°C. On peut nettement distinguer la saison de pluies
qui commence en Mai et termine en Septembre
(Graduation mensuelle de Janvier 2009 à Janvier 2010).
Cette combinaison de fortes températures et de pluies
abondantes, favorise la grande diversité des êtres vivants
Figure 2 Diagramme météorologique Janvier 2009 à Janvier 2010 Station HOBO
1.2) Présentation du tuteur et de ses recherches à la station
Friedrich SCHIEMER dispose d’une chaire à l’Université de Vienne en Autriche, en
temps que Limnologue. En 2005, il a pris la tête du département de limnologie et
d’hydro-botanique de l’université de Vienne. Depuis 1982, il a dirigé, coordonné et
participé à de nombreux programmes de recherches aquatiques sur tous les continents. Il
dirige actuellement un programme financé par l’Union Européenne, qui vise à stabiliser le
lit du Danube, de Vienne à Bratislava. Il a jusqu’aujourd’hui publié plusieurs livres, dont
un considéré comme une référence de la limnologie tropicale, « Perspectives in tropical
Limnology » en 1996.
Il a participé à plusieurs projets limnologiques et entomologiques sur les cours d’eau du bassin
versant d’Esquinas du Costa-Rica. Une première étude a été publiée en 2007 sur le réseau des rivières du
Parc national de las Piedras Blancas1, et une autre sur les invertébrés et sur la décomposition de la litière² en
2008. Une recherche a également été faite sur les poissons de la rivière Quebrada Negra³, à proximité de la
station de recherche. 2
1 The river network of the Piedras Blancas National Park, Costa-Rica », J.Tschelaut, C.Pichler and F.Schiemer, 2007
2 Macroinvertebrates and leaf litter decomposition in a neotropical stream Quebrada Negra, J. Tschelaut, and F.Schiemer, 2008
³ Ecology of fishes of Quebrada Negra, Costa Rica, a first order neotropical lowland stream , C.Pichler and F.Schiemer, 2008

4
1.3) Le Costa-Rica
1.3.1) Généralités
Le Costa Rica est une république démocratique (51 000 km² et 4,2
millions d'habitants) située entre le Panama et le Nicaragua en Amérique
centrale. Son revenu moyen par habitant de 3 646 dollars US, (en 2006) est
supérieur à la moyenne des pays d'Amérique latine.
Ce pays est souvent admiré pour sa longue tradition démocratique, et une bonne stabilité
économique. Il a aboli son armée en 1949. Depuis le 8 Mai 2010, Laura Chinchilla Mirinda est la première
femme du pays au pouvoir (Partido de Liberacíon Nacionál). La biodiversité très importante qui existe au Costa-
Rica, fait de ce pays une destination touristique privilégiée. Ce secteur important a largement augmenté cette
dernière décennie (CIA World Factbook, 2009).
1.3.2) Situation agricole
L’agriculture primaire du Costa-Rica est bien diversifiée: le café, les bananes, les cultures à cycle court,
l'élevage de boucherie, de laiterie, et les plantations de forêts représentent l'essentiel de l'utilisation des terres.
Pour ce qui est de la valeur de la production par hectare, d'autres cultures sont plus importantes, notamment
les fruits et légumes (Ananas), les fleurs et les plantes ornementales.
L'élevage destiné à la viande de boucherie est l'activité qui utilise les terres de la manière la plus extensive.
Son importance dans le secteur agricole a diminué tout comme sa productivité. Les exportations de viande
bovine, qui étaient considérables par le passé, sont maintenant très limitées du fait de la faible rentabilité. La
production laitière, par contre, est plus intensive; elle a augmenté et représente une part importante des
exportations. Les volailles et les œufs sont des sous-secteurs importants, mais dépendent du maïs et du soja,
importés majoritairement depuis les Etats-Unis.
Café29%
Riz18%
Bananes13%
Canne à sucre15%
Haricots8%
Palme à huile10%
Oranges7%
Soles nationales des grandes cultures
L'agriculture primaire représente
6.5% du PNB intérieur, et sa part continue de
reculer. Elle emploie 14% de la main
d’œuvre nationale. Ce chiffre atteint 32% si
on y ajoute les services para-agricoles
(Country profile FAO, 2008).
Fig. 3 Diagramme circulaire des soles nationales des
grandes cultures

5
Les ouvriers agricoles et les paysans dont les exploitations sont inférieures à 3 hectares (le plus souvent situées
en zone montagneuse et qui pratiquent l'agriculture pluviale) disposent souvent de revenus inférieurs au seuil de
pauvreté. La majorité ne dépend pas seulement de la vente des récoltes pour leurs revenus, c’est pourquoi les
emplois d'appoint non agricoles sont une pratique courante. Les exploitations moyennes, de 3 à 10 hectares sont
les plus courantes. Elles sont généralement consacrées à l'élevage des bœufs de boucherie, à l'huile de palme, au riz,
à l'ananas et aux bananes, et sont parfois propriétés d'entreprises ou de sociétés internationales (Chiquita, ex-United
Fruit company).
Du fait de l'orientation de l'agriculture costaricaine vers des marchés d'exportation, il a été nécessaire
d'accroître les quantités de semences importées, d'engrais et de produits agrochimiques, ce dont s'est chargé le
secteur privé. Dans certains cas, les coopératives agricoles participent à la commercialisation de ces produits.
1.3.3) Déforestation, mise en culture de terres et qualité de l’eau
Le Costa-Rica, comme beaucoup d’autres pays richement pourvus en surfaces forestières, a subit de plein fouet
les vagues de déforestation, qui submergent et dévastent encore certaines côtes et rives, de cours d’eau d’Amérique
centrale et latine. Cependant le pays voit ses dernières années, sa surface forestière augmenter légèrement.
Les conséquences écologiques sur l’eau et sa bio-structure peuvent être importantes, créant des chemins
préférentiels d’écoulement des eaux, mais surtout de transferts de polluants et fertilisants… Ces problèmes ont
longtemps été tamponnés par un renouvellement rapide des nappes d’eau du fait des précipitations importantes,
mais depuis quelques années surgissent des problèmes sanitaires importants. Un de ceux-ci nous concerne
directement, puisqu’une bactérie s’est développée dans le Rio Bonito, et a causé de graves lésions oculaires chez
plusieurs personnes (Annexe 2 Lettre au Ministre de la Santé et traduction linéaire).
Le développement d’algues vertes dans les canaux de drainages de certaines palmeraies, et cultures
d’ananas, laisse préjuger des atteintes écologiques d’une fertilisation mal contrôlée. On peut évaluer l’importance
du maintien des zones sylvicoles existantes, dans le schéma consigné en annexe 3. Il résume grossièrement les
conséquences écologiques, économiques et sociales d’une déforestation massive et d’une uniformisation des
cultures.
Le stage entre dans le cadre des préoccupations écologiques du Costa-Rica, par le biais d’une étude sur le
Bassin Versant d’Esquinas.

6
2) Problématique et axes de recherche
2.1) Problématique de stage
Les méthodes d’évaluation biologique,
utilisant les organismes macro-invertébrés, ont été
largement utilisées au sein de « monitoring » de la
qualité des eaux, dans de nombreux pays
(TRIHADININGRUM et DE PAUW, 1994). L’utilisation
d’indicateurs biologiques présente l’avantage, par
rapport aux méthodes physico-chimiques, de prendre
en compte de nombreux paramètres
environnementaux non-mesurables, et améliore
l’efficience du monitoring. Ils permettent également
d’observer une évolution de l’environnement
écologique, plus pertinente vis-à-vis de la protection
des organismes aquatiques (donc de la biodiversité) et
de la santé humaine (LOUGHEED et al. 2007).
Les insectes font souvent de bons indicateurs,
car ils sont présents, dans une certaine mesure, dans
tous les types d’habitats (WHITEHOUSE et al. 2008) et
vivent parfois dans des habitats spécialisés (LEWIS et
GRIPENBERG 2008). Bien que beaucoup de données
d’études aient exclu de nombreux taxons comme
« indicateurs potentiels » (SAHLEN & EKESTUBBE
2001), certaines études tendent à montrer que leur
utilisation est possible.
L’ordre des Odonates est étudié depuis
longtemps, car il joue un rôle important dans
l’équilibre des communautés d’invertébrés aquatiques
(macro-invertébrés benthiques), et son abondance et
sa diversité font de lui un candidat bio-indicateur
idéal. Etant très sensibles à leur habitat, les
populations d’Odonates peuvent être indicatrices
d’abondance d’autres invertébrés et de macrophytes
(CORBET 1999, BRIED et ERVIN 2005).
Des études ont déjà été effectuées sur
l’utilisation des adultes Odonates, en tant
qu’indicateurs de pollution d’un milieu par des
insecticides (SCHULIN F. et al., 1999), ou par les
métaux lourds (STRINIKI A. 2010), montrant une
sensibilité avérée de certaines espèces indicatrices à
ces produits.
Dans le cadre d’un monitoring, ces organismes
peuvent être plus facilement utilisés que d’autres
organismes aquatiques (micro-organismes) :
- Ils sont facilement récoltables, comparés aux
micro-organismes.
- Ils vivent et se reproduisent dans tout le spectre
d’habitats possibles.
- Ils sont très hétérogènes. La probabilité que
certains organismes réagissent à un changement
particulier de conditions environnementales est
importante.
- Ils sont plus ou moins sensibles et réactifs à
différents types de polluants (SCHULIN et al., 1999).
- Ils sont relativement faciles à déterminer en
comparaison aux micro-organismes (études
bactériologiques).
L’objectif de ce stage est de faire une première
approche de la mise en place d’un suivi de la qualité
des eaux du Bassin versant d’Esquinas, par l’étude de
l’influence anthropique sur les populations
d’Odonates. Elle se fait dans un premier temps par la
connaissance plus précise du réseau fluvial. Certains
paramètres abiotiques des cours d’eau du bassin
versant sont analysés, tels que la morphologie,
l’hydrochimie, la sédimentologie, leur proximité avec
les cultures, ce qui permet une meilleur appréhension
du lieu de vie des Odonates (1). Dans un second
temps, on donnera une idée de l’influence de la
déforestation et de la mise en culture de terres, sur
les populations d’Odonates, comme bio-indicateur(2).

7
2.2) Aires d’étude
2.2.1) Zone de relevés limnologiques
Le bassin versant d’Esquinas s’étend sur 424km² (BERGOEING 1998). La zone étudiée est
composée de neuf cours d’eau principaux, d’ordre 1 à 5. Les deux rivières importantes de la vallée
de La Gamba, les Rio Bonito et Rio Esquinas, traversent des zones agricoles importantes et des
forêts secondaires à différents stades de maturité. Le Rio Esquinas forme une bordure naturelle de
la Forêt Esquinas au Nord et du Parc Naturel protégé à l’Ouest, puis se jette dans l’Océan Pacifique,
au niveau du Golfo Dulce.
Du fait de la déforestation importante, on ne trouve presque plus de forêt primaire dans
cette zone. Les parties en amont de ces deux rivières présentent peu d’influence anthropique, et la
majorité des prises d’eaux (ordre 1) coule dans l’unique forêt primaire de la zone. Les coordonnées
GPS des points de relevés se trouvent en annexe 4.
Figure 4 Réseau fluvial du Bassin versant Esquinas, réalisé avec ArcGIS.

8
2.2.2) Zones de relevés entomologiques
Afin d’étudier les différences entre les habitats primaires et les habitats modifiés, les relevés de libellules
sont faits le long de cours d’eau en forêt primaire, en marge de forêt, et dans les zones cultivées.
Les sites de la forêt primaire se trouvent le long de la Quebrada negra (ordre 1), ainsi qu’à proximité d’un
petit affluent de celui-ci, que l’on rejoint par le Riverbed trail. Ce dernier correspond à la prise d’eau potable
de la station de recherche. Ces sites seront affublés du sigle F (pour Forêt) suivi d’un chiffre, qui permettra
de les identifier.
Les deux sites en marge de forêt, sont situés à proximité de la station de recherche. Le premier se trouve
dans le jardin botanique proche d’une plantation de cacao abandonnée (Theobroma cacao), et le second sur
le « Laguna trail ». Ils seront signalés par le sigle FM (pour « Forest margin » : en marge de forêt).
- Les sites en zone cultivée se trouvent le long de plusieurs rivières : Q.Negra, Q.Gamba, et Rio bonito, proche
du village de la Gamba, traversant rizières (Oryza sp.) et champs de palme à huile (Elaeis guineensis). Les
zones de relevés sont recensées sur la carte suivante par le sigle CA (pour « Cultivated areas »).
Fig. 5 Carte des zones de relevés entomologiques
Ceux-ci correspondent aux trois types d’habitats étudiés pour la répartition des Odonates, et l’évaluation
de l’influence de la déforestation et mise en culture de terres.
Les coordonnées GPS
des points de relevés
sont consignées en
Annexe 5.
1km

9
2.3) Matériel et méthodes
2.3.1) Limnologie
La connaissance du lieu de vie des macro-invertébrés aquatiques, est nécessaire à la compréhension de
leur développement, de leur interaction avec ce milieu (nutrition, reproduction…). Le stage consiste à
construire et mettre en place les protocoles suivants. En premier lieu est réalisé un profil morphologique
précis de portions représentatives de certains cours d’eau dans des milieux de vies aquatiques distincts. Des
zones de relevés (longues de 100m) sont mises en place, au sein de la forêt primaire, tandis que d’autres sont
installées en zones cultivées (riz et palme). Les paramètres physiques et chimiques des eaux du Bassin versant
sont analysés, afin d’établir une base de données pour le monitoring.
Morphologie du cours d’eau
- Les cartes sont réalisées grâce aux programmes ArcGIS et Photoshop, à partir de photographies aériennes et
de relevés GPS.
- Les zones de relevés sont définies à partir d’une reconnaissance de terrain, sur des tronçons représentatifs
du cours d’eau.
- Des points de relevés du niveau d’eau sont établies tous les 10 mètres, sur les tronçons de 100 mètres. Des
tubes en PVC (Ø50mm) gradués tous les 5cm, et d’une taille moyenne d’1,50m, y sont implantés, et sont
maintenus au sol par un piquet de fer, et arrimés aux rives par des ficelles hydrofuges.
- Le niveau 0 de référence est établi le samedi 24/04/2010 entre 14h15 et 14h45, sur la Quebrada Negra.
- A l’aide d’un tube mobile gradué tous les centimètres, des mesures de profondeur sont effectuées tous les
20cm sur les lignes d’eaux, perpendiculaires à l’écoulement de l’eau.
- Pour la mesure des parties émergées, une ficelle horizontale est tendue à 1,5 mètre au-dessus du niveau
d’eau de référence. La hauteur de la rive par rapport au niveau de l’eau est obtenue par différence. Ces
mesures permettent de donner un profil assez précis du tronçon.
Niveau d’eau
Réponse d’une rivière aux fortes pluies à court terme
La réponse du bassin versant se calculera en variation de hauteur des cours d’eau, sur la Quebrada
Negra. Le relevé du niveau d’eau est effectué toutes les 5 minutes. Grâce à la station météorologique HOBO
présente dans le jardin de la station, on pourra mettre en relation les précipitations, et la réponse du cours
d’eau.
Sur la durée du stage
Le relevé du niveau d’eau est journalier pour la Quebrada Negra (entre 8h00 et 10h00), et deux relevés sont
effectués sur les autres rivières.

10
Température de l’eau, pH, Oxygène dissout
A l’aide d’une sonde, ces valeurs sont relevées dans l’ensemble des cours d’eau. Ces mesures sont plus
régulières pour la Quebrada Negra, dans les deux tronçons étudiés. Chaque cours d’eau est prospecté deux fois.
- pH mètre « Hanna HI-8314 »
- Dissolved oxygen meter “Keithley 874C”
- GPS system “Garmin 60CSX”
- Station météorologique “Hobo” (humidité de l’air et température, température du sol et pluviomètre,
intensité de lumière)
Sédimentologie
Dans un cadre en bois de 40x40cm, une appréciation visuelle est donnée
de la proportion de sédiments (en termes de couverture du lit), selon trois tailles
distinctes, qui permettent de classer les zones selon le triangle suivant. Les valeurs
permettent de faire une carte des sédiments du fond des différents tronçons
étudiés, grâce au programme de cartographie ArcGIS.
Fig. 6 Triangle des sédiments
Des mesures de solides en suspension sont effectuées pour donner une première idée de la charge des cours
d’eau [L’expérience a failli, le filtre Buchner ne fonctionnant pas]
Hydro-chimie
Des échantillons d’eau sont prélevés dans les zones prospectées. Les analyses chimiques sont effectuées au
laboratoire du CATIE de Turrialba. Ont uniquement été dosés les nitrates et phosphates. Les échantillons sont
transportés dans un bac réfrigéré, et filtrés avant le dosage des nitrates. La conductimétrie est également mesurée.
Vitesse du cours d’eau
Une mesure de la vitesse du cours d’eau est effectuée, à l’aide d’un bâton et de deux cordes. Ces dernières
sont tendues perpendiculairement à l’écoulement du cours d’eau, et séparées de 10 mètres. Le bâton est jeté 10
mètres avant le premier fil, et on mesure le temps de passage sous chaque fil. Du fait de l’imprécision du protocole,
celui-ci est répété 10 fois sur chaque tronçon, pour diminuer au maximum le biais.
Agriculture locale
La connaissance des milieux situés en zone agricoles passe aussi par la connaissance des pratiques agricoles
locales. Le propriétaire des rizières dont les canaux de drainage débouchent dans la Quebrada negra a été audité.
Cette rencontre vise à connaître les façons agricoles (travail du sol, fertilisation, traitements, insecticides…) associés
à la culture de riz, et à discuter avec les exploitants des problématiques de l’eau.
La connaissance globale du milieu est nécessaire au suivi de populations d’êtres vivants.

11
2.3.2) Entomologie
D’après les études menées sur la Quebrada negra en 2004, les Odonates représentent en nombre d’individus,
3,7% des organismes macro-invertébrés aquatiques.
L’ordre des Odonates (libellules et demoiselles) se divise en deux sous-ordres facilement reconnaissables à
l’état adulte. Les Anisoptères, d’une taille moyenne à grande (5 à 12mm), sont les plus habiles et ont une tête ronde
avec les yeux fortement rapprochés. Lorsqu’ils se posent sur une branche, ils gardent les ailes ouvertes. L’autre sous-
ordre est celui des Zygoptères, à la tête aplatie, de taille plus petite (2 à 7mm) avec les yeux bien espacés. Lorsqu’ils
se posent, ils mettent leurs ailes dans l’alignement du corps.
Méthode de relevés
Les relevés sont effectués, entre le 23/04 et le 18/05, de 08h00 et 12h30 (GMT -06 :00). Chaque site défini a
été sondé deux fois, durant 50 minutes précisément, sur des zones longues de 100 mètres, sur les rivières. Les
Odonates perchées ou volantes ont toutes été prises en photo avec un appareil CANON zoom optique x26 avec
stabilisateur d’image, ce qui permet d’avoir une qualité d’images permettant une reconnaissance parfaitement
fiable. Un travail préalable sur les espèces probablement présentes a été réalisé, et quelques captures ont été
effectuées pour une détermination exacte. Les photos sont référencées dans un catalogue électronique (Compact
disque ci-joint). Certaines Odonates sont trop rapides pour pouvoir être prises en photographie, et sont donc
capturées à l’aide d’un filet rond, de diamètre 30cm, avec des mailles de 2,5mm, et muni d’un manche d’1,50m.
Les libellules ont été identifiées selon la clé de Förster (FORSTER 1999), et celle de Libélulas de Mesoamérica
y el Caribe (ESQUIVEL C. 2006), puis sont relâchées si elles avaient été capturées. L’identification s’est toujours faite
jusqu’au genre et jusqu’à l’espèce lorsque ceci a été possible. Le sexe est également précisé quand c’est possible. Il a
été nécessaire, de contacter des spécialistes pour déterminer Erythrodiplax kimminsii et Dythemis multiponctata.
Couverture lumineuse
Pour chaque station de prélèvement d’Odonates, la couverture lumineuse (Canopy cover : cc) est mesurée, à
l’aide d’un densiomètre (en pourcentage). On cherchera à mettre en valeur une relation entre les valeurs mesurées
et la présence d’Odonates. Dix relevés par station permettent d’établir une moyenne représentative du tronçon.
Autres paramètres relevés
Pour chaque prélèvement sont notés l’heure, la profondeur du cours d’eau, la hauteur à laquelle les
Odonates sont perchées, la largeur du cours d’eau, et des commentaires comportementaux peuvent être ajoutés,
pour une meilleure compréhension de la répartition des espèces.
Cette première approche va introduire d’autres recherches sur le sujet. La création d’une clé simplifiée, adaptée
aux libellules de la région a été créée lors de ce stage, pour faciliter les recherches (CD et annexe)

12
2.3.3) Statistiques
Richesse en espèces (Diversité Alpha)
Du fait de contraintes de temps, les nombres d’espèces observées représente une fraction non connue du
nombre réel d’espèces d’Odonates pour un habitat donné, dont le taux peut varier d’un habitat à l’autre. Bien
que la méthode soit standardisée, il peut y avoir des différences considérables dans les taux de captures entre les
différents habitats, du fait que les densités de populations diffèrent, ou que les relevés soient plus faciles dans
les habitats ouverts qu’en forêt, où certaines libellules volent dans la canopée. Afin de comparer le nombre
d’espèces, il est nécessaire de prendre en compte ces « effets de relevés ». Pour cette raison, les courbes
d’accumulation d’espèces et l’indice estimateur non-paramétrique d’abondance (ACE), ont été calculés à partir
du logiciel gratuit Estimates 7.0 package (COLWELL, 2006), utilisant les formules Chao classic et 100
randomisations. La richesse en espèce n’a pas été évaluée pour les marges de forêt (FM), du fait du faible nombre
de répétitions (n=2 sites).
Afin d’identifier les caractéristiques des différents habitats influant sur la richesse en espèces, ont été testés
plusieurs paramètres. La couverture lumineuse (cc : canopy cover), ainsi que la profondeur (sd: stream depth) et
la largeur moyenne (sw: stream width) des cours d’eau. Différents tests de corrélation et régression ont été
appliqués à l’aide du programme R (COLWELL, 2006), et de ses packages VEGAN (OKSANEN J. 2010) et MASS
(RIPLEY B. 2010). Les deux tests de corrélation de Spearman et Kendall sont appliqués sur les résultats. D’autres
valeurs ont été mesurées durant cette période, mais ont été écartées du modèle (pH, Température, Oxygène
dissout, conductivité) car elles varient très peu d’un point à l’autre. La vitesse du courant n’a pas pu être testée
du fait de l’imprécision du protocole.
Similarités (Diversité Beta).
Dans l’objectif de comparer la composition en espèces des différents habitats, une matrice standardisée des
similarités Bray-Curtis a été calculée à l’aide du programme R et Estimates 7.0 Package. Un graphe NMDS (Non-
metric multidimensional scaling) a été effectué grâce au programme de traitement statistique R, muni du
package VEGAN et cluster (MAECHLER M. 2010). Il a été démontré que le NMDS est une méthode d’ordination
pertinente de relevés incomplets pour la faune entomologique tropicale (RIPLEY B. 2010).
Pour tester la significativité des différences dans la composition d’espèces des milieux, un test ANOSIM
(Analysis of similarity) a été effectué. L’influence des paramètres sur la composition est évaluée par un test de
Mantel, réalisé grâce aux lignes de codes de Zt Software (BONNET E. et VAN de PEER K., 2002). Afin d’identifier
les espèces qui caractérisent les différents types d’habitats, la contribution des espèces aux similarités a été
calculée grâce au programme PRIMER (PRIMER-E, 2007), à l’aide de l’algorithme SIMPER (Similarity percentages).
L’analyse SIMPER n’est pas appliquée à l’habitat FM, du fait du faible nombre de relevés.

13
3) Résultats
3.1) Limnologie
3.1.1) Généralités
Les neuf cours d’eau étudiés sont tous des affluents du Rio Esquinas qui traverse le Parc National de Las Piedras
blancas, et dont l’embouchure se trouve dans le Golfo Dulce (Golfito). Chaque cours d’eau a été prospecté deux fois,
sauf la Quebrada Mari, Quebrada Negra et le Rio Esquinas. Les descriptions des cours d’eau et résultats détaillés
sont consignés en annexe 6.
Rivière Ordre Largeur
m
Profondeur
m
Température
°C
pH Oxygène
%/mg.L¯¹
Conductivité
μS/cm
PO4
μg.L¯¹
NO3
μg.L¯¹
Q Negra ca 1 2,9 0,2 24,8 7,83 96/7,8 200 48 95
Q Negra f 1 2,8 0,2 23,7 7,90 98/7 ,9 185 26 64
Q Mari 1 2,4 0,2 22,3 8,05 100/8,0 260 3 285
Q Chorro 2 4,5 0,2 26,3 7,97 102/8,16 128 5 9
Q Gamba 2 6,8 0,4 26,4 7,85 98/8,05 154 28 62
Q Bolsa 2 10 0,1 25,2 7,92 - 158 31 120
Q Sardinal 2 5,9 0,2 27,6 7,73 - 126 - -
R Oro 3 16,8 0,2 26,8 7,75 - 367 36 269
R Bonito 3-4 5,1 0,2 26,2 8,3 - 155 13 47
R Esquinas 5 30* 1,5 29,4 7,78 - 299 - -
*estimé Tableau 1 Récapitulatif des données morphologiques et physico-chimiques des cours d’eau
La température des cours d’eau se trouve généralement autour de 26°C, sauf pour la Quebrada Mari à 22,3°C,
représentative des cours d’eaux des zones néo-tropicales. Les valeurs de pH varient peu et se trouvent globalement
autour de 8, qui reflètent également le pH des sols de la région. Les valeurs d’oxygène dissout varient très peu
également, autour de 8mg/L. La conductivité de la Quebrada Mari, du Rio Oro et du Rio Esquinas est nettement
supérieure, sûrement due au fait que ces cours d’eau traversent une zone géologique différente (« Fila »).
Les analyses chimiques effectuées en laboratoire, ne montrent que les phosphates et nitrates, dont les fuites
sont souvent associées à l’exploitation de terres agricoles. On peut tout d’abord mettre en valeur les concentrations
en phosphates importantes dans la Quebrada Marí et dans le Rio Oro, que l’on associe à leur origine géologique, le
Fila Cruces. Pour le Fila Gamba, on peut voir que les valeurs de phosphore sont relativement élevées dans les
Quebrada Negra, Gamba et Bolsa, toutes situées dans des zones cultivées et fertilisées (voir carte des cultures en
page 19).
On peut également observer la différence qui existe entre les deux relevés d’eau sur la Quebrada Negra. L’eau
en zone cultivée dispose d’une concentration en phosphates deux fois supérieure à celle de la forêt, et le niveau de
nitrates passe de 64 à 95 μg.L¯¹. Il est nécessaire de préciser que les relevés d’eau ont été effectués peu après la
fertilisation et la plantation du riz de Monsieur Cheppo. On précise également que les canaux de drainage de 3ha
champs de palme à huile, débouchent dans la Quebrada Negra, 300 mètres en aval du lieu de relevé.

14
Les rivières sont classées de gauche à droite,
selon leur ordre. On peut voir que la largeur
augmente de manière importante, pour atteindre
plus de 40 mètres de large à l’embouchure. On
rappelle que ces données correspondent uniquement
à des relevés en saison des pluies. La largeur peut
être nettement inférieure en saison sèche pour
certains cours d’eau.
Fig. 7 Histogramme des largeurs des cours d’eau
Les valeurs de profondeur ne sont pas
corrélées avec l’ordre de la rivière. On peut
voir que la Quebrada Marí (ordre 1), égale
quasiment la profondeur du Rio Oro (ordre
3). La variation de profondeur entre les
deux saisons diffère assurément d’un cours
d’eau à un autre.
Fig. 8 Histogramme des largeurs des cours d’eau
La vitesse du courant des différents cours
d’eau est très variable. La Quebrada Marí est plus
rapide que le Rio Oro, ce qui montre qu’il n’y a pas
de corrélation entre l’ordre et la vitesse du
courant. La vitesse donnée dépend beaucoup des
tronçons sur lesquels ont été mesurés la vitesse. La
succession de pool-riffle influe sur la vitesse
moyenne.
Fig. 9 Histogramme des vitesses des cours d’eau
On rappelle que le protocole de mesure de la vitesse n’est pas extrêmement précis, mais qu’il est
suffisant pour comparer les rivières entre elles. On ne peut cependant pas se permettre d’évaluer le
débit précis du fait de l’erreur importante.

15
3.1.2) Quebrada negra
L’étude s’est concentrée sur la Quebrada negra, cours d’eau naturel directement accessible
depuis la station. Les cours d’eau néo-tropicaux sont des environnements très hétérogènes qui varient
sur de très courtes distances.
Dans un cours d’eau naturel, il existe plus ou moins une alternance entre des zones peu profondes,
avec un courant rapide et un substrat graveleux (« riffles »), et des zones plus profondes, avec un
courant plus lent et un substrat plus fin (« pools »). Sur les graphes suivants, on peut voir qu’une faible
profondeur entraîne une vitesse élevée et qu’une zone profonde, induit une faible vitesse en surface.
f
On rappelle que les valeurs des
vitesses données ne sont pas
extrêmement précises, mais qu’elles sont
suffisantes pour montrer l’enchaînement
des « pool» et « riffle ».
Cet enchaînement est présent dans tous
les cours d’eau. Les « pool » semblent
être plus profonds dans les cours d’eau
plus importants.
Fig. 10,11 et 12 Graphes de la vitesse et de la
profondeur de la Quebrada Negra (25/05/2010).

16
On peut observer les « pool » et « riffle » sur ces deux cartes. On peut voir que les zones les plus
profondes en bleu foncé, correspondent globalement aux zones sédimentologiques de substrat fin (<2mm)
en jaune. Les autres cartes et le détail des profils morphologiques sont consignée en annexe 7 et 8
Fig. 13 Carte morphologique d’un tronçon de 100 mètres de la Quebrada Negra, à proximité de la Tropenstation
Fig. 14 Carte sédimentologique d’un transect de 100 mètres de la Quebrada Negra, à proximité de la Tropenstation

17
Le graphe suivant permet de montrer la réponse de la Quebrada Negra aux précipitations, du 25/04/2010
au 10/06/2010. On rappelle que le niveau zéro, correspond à la référence du 24/04. Depuis cette date, le
niveau d’eau n’est pas descendu sous cette valeur. Les valeurs sont consignées en annexe 9.
On peut observer l’évolution des précipitations du début de saison des pluies, et l’augmentation
progressive du niveau d’eau moyen. On peut souligner la pluviométrie du 1er juin, journée durant laquelle,
il a plu 144mm, ce qui représente plus d’un dixième des précipitations annuelles de la Bretagne. Les
flèches signalent des augmentations du niveau d’eau, qui ne correspondent pas aux précipitations, du fait
de précipitations en amont du lieu de relevé. La baisse moyenne d’une journée sans pluie est de 0,62 cm,
cependant cette valeur est peu exploitable sur une si courte durée.
0
5
10
15
20
25
30
35
40
45
50
1 3 5 7 9 11 13 15 17 19 21
No
mb
re d
e jo
urn
ée
s
Niveau d'eau atteint [mm]
0
5
10
15
20
25
0
20
40
60
80
100
120
140
160
25
-avr
.
27
-avr
.
29
-avr
.
01
-mai
03
-mai
05
-mai
07
-mai
09
-mai
11
-mai
13
-mai
15
-mai
17
-mai
19
-mai
21
-mai
23
-mai
25
-mai
27
-mai
29
-mai
31
-mai
02
-ju
in
04
-ju
in
06
-ju
in
08
-ju
in
Niv
eau
d'e
au [
cm]
Pré
cip
itat
ion
s [m
m]
Ce graphique représente le nombre de
journées ayant atteint un certain niveau d’eau
(x), durant les 46 jours étudiés.
Plus de la moitié des journées se trouve au
dessus de 6mm. Le niveau le plus élevé observé
atteint 22mm. Si des relevés continus sont mis
en place sur tous les cours d’eau, ce type de
graphique permet de comparer leur réponse
aux intempéries.

18
L’évolution du niveau d’eau à court terme de la Quebrada negra est très typique et joue un rôle
plus important que la fluctuation à long terme. Les niveaux d’eau augmentent rapidement après le début
des précipitations, pour un retard de réponse (ρ = « rho ») calculé à 20 minutes. Le retard correspond à
l’écart entre le pic des précipitations et le pic du niveau d’eau.
Fig. 16 Graphe d’évolution du niveau d’eau, en fonction des précipitations du 13/05/2010
Beaucoup de sols de sont très argileux, ou sont rocheux imperméables, et très peu d’eau de pluie
rejoint les nappes souterraines. Un cours d’eau calme peut très rapidement se changer en torrent très
rapide et boueux. Les niveaux augmentent rapidement mais redescendent également très rapidement,
dus à la haute résilience de la Rainforest. Ceci constitue la règle générale des cours d’eaux de cette région
du Costa-Rica (SCHIEMER).
[La mesure des solides en suspension n’a pas fonctionné du fait du filtre Buchner non utilisable. Les premiers
résultats obtenus ne sont pas exploitables. Cette mesure pourrait être facile à mener, mais nécessite de racheter
un tube plastique solide. Le temps d’attente avant la réception du tube, n’a pas permis de réaliser cette
expérience].

19
Figure 18 Carte macro-échelonnée des relevés entomologiques, avec indications agricoles.

20
3.2) Entomologie
3.2.1) Alpha et Gamma diversités
Sur les 10 zones de relevés, 427 individus d’Odonates adultes ont été relevés, représentant 26 espèces et
6 familles dont 5 Zygoptères et 1 Anisoptère.
Tableau synthétique des relevés
Sites
Famille Espèce F1 F2 F3 F4 CA1 CA2 CA3 CA4 FM1 FM2
Calopterygidae Hetaerina fuscoguttata (Selys 1878) - - - - 4 3 - 13 - -
Hetaerina occisa (Hagen 1853) 6 7 6 1 21 15 13 - - -
Coenagrionidae Acanthagrion inexpectum (Selys 1876) 1 - 1 - 4 5 - - 2 17
Argia cupraurea (Calvert 1902) - - 2 - 26 6 2 - - -
Argia sp (Rambur 1842) - - 3 - 15 9 9 - - 13
Enallagma novaehispaniae (Calvert 1907) - - - - 6 2 - - - -
Ischnura capreola (Hagen 1861) - - - - - - - - - 10
Megapodagrionidae Heteragrion erythrogastrum (Selys 1886) - 8 9 1 - - - - - -
Philogenia zeteki (?) - - - 7 - - - - - -
Protoneuridae Neoneura esthera (Williamson 1917) - - - - - 2 1 - - -
Pseudostigmatidae Mecitogaster modesta (?) - 1 - - - - 1 - - -
Libellulidae Cannaphila insularis (Kirby 1889) - - - - - 1 - - - -
Cannaphila vibex (Hagen 1861) - - - - - - - 1 - -
Dythemis multiponctata (Kirby 1894) 1 - - - - - - - 6 6
Dythemis sterilis (Hagen 1861) - - - - 4 2 1 3 - -
Erythrodiplax funerea (Burmeister 1839) - - - - - - - 3 - -
Erythrodiplax fervida (Erichson 1848) - - - - - - - 1 1 -
Erythrodiplax fusca (Rambur 1842) - - - - 1 - 1 4 - 9
Erythrodiplax kimminsi (Borror 1942) - - - - - 2 - 1 - -
Eurythemis plebeja (?) - - - - - - - - - 1
Macrothemis imitans (Karsch 1890) - 1 - - 1 - - - - -
Micrathyria pseudeximia(?) - - - - - - - - 17 11
Orthemis ferruginea (Fabricius 1775) - - - - - - - 2 5 11
Perithemis mooma (Kirby 1889) - - - - - - - 3 2 1
Rhodopygia (Kirby 1889) sp. - - - - - - - - 12 23
Uracis imbutata (Burmeister 1839) - - - - - 1 1 11 13 -
Tramea binotata (?) - - - - - 1 1 - - 9 Tableau 2 Tableau synthétique des relevés d’Odonates
Sur le disque compact ci-joint, vous pourrez trouver le guide d’identification des Odonates des
cours d’eau de la région. Dans le cadre du monitoring, ce guide doit permettre de faciliter le suivi des
populations d’Odonates, en donnant les caractères de reconnaissance, ainsi que des informations sur le
comportement, les lieux de vie ainsi que leur répartition sur les continents américains. Un exemple de ces
fiches est donné en annexe 11.

21
Un nombre d’espèces plus important a été rencontré en
zones cultivées (ca = 15 espèces) qu’en forêt (f = 9 espèces),
(Welch N=4, t=7.3 ddl¹=5 p<0.001, Détail du test en annexe 12),
ce qui a été également prouvé par l’estimateur de richesse en
espèces.
Fig.19 Boîtes à moustache des espèces identifiées par habitat.
Fig. 20 Courbes d’accumulation d’espèces (Mao Tau) avec leur intervalle de confiance à 95%
Habitat Nb de relevés Espèces identifiées ACE² Inventaire effectué à Familles Genres
F 8 9 13 70% 5 7
FM 4 13 - - 2 12
CA 8 20 25 80% 5 14
Total 20 27 30 90% 6 20 Tableau 3 Récapitulatif de l’inventaire, et estimation de richesse en espèces
Le nombre total d’individus observés est plus important dans les zones cultivées (203), qu’en forêt
(55). Le nombre d’individus observés par site est également plus important en zone cultivée
(moyenne= 50.75 écart-type=22,2) qu’en forêt (moyenne = 13,75 écart-type=6,3). (Welch : N=4,
t=3,9, ddl¹=3.5, p=0.04).3
3 ddl : Degré de liberté obtenu par le test ²ACE : Abundance-based coverage estimator
Plus globalement les trois courbes
d’accumulation montrent qu’un effort plus
important aurait mené à un nombre plus
important d’espèces. L’inventaire
d’espèces est effectué à 80% pour CA et
75% pour F. L’estimation totale est de 30
espèces, qui représentent 11% des 286
espèces connues au Costa-Rica (ESQUIVEL,
2006).
0
5
10
15
20
25
F
FM
CA
f ca fm
F CA FM
Note : Les points montrent la médiane du nombre d’espèces, la boîte montre l’intervalle 25-75% des espèces et les moustaches montrent les maximums et minimums.

22
Les espèces considérées comme « autochtones », sont celles uniquement réparties en Amérique
centrale. On peut voir sur ces graphes, que la proportion d’espèces et d’individus « généralistes », parfois
appelés « opportunistes » augmente avec la dégradation des habitats.
Fig. 21 et 22 Histogramme des proportions relatives d’espèces et individus, autochtones et généralistes.
La distribution des espèces à travers les deux continents américains est reportée en Annexe 13, à
partir des travaux du bio-géographe Carlos-Esquivel.
5
2 3
4
11 17
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
F FM CA
Espèces autochtones Espèces généralistes
29
54
11
26
115
192
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
F FM CA
Individus autochtones Individus généralistes

23
3.2.2) Beta diversité
Dans l’objectif de comparer les paramètres influant sur les populations d’Odonates, différents tests de
corrélations ont été menés. Explication des tests en annexe 14.
Les tests de corrélation (Spearman et Kendall) donnent
une corrélation négative entre le nombre d’espèces et la
couverture lumineuse significative au risque de 5%. Attention
de ne pas y attribuer un lien de causalité. Une telle
corrélation peut venir d’un autre paramètre indépendant
non mesuré. Ce test affirme juste qu’on trouve plus de
libellules au soleil qu’à l’ombre, de manière significative.
Fig. 23 Graphique de la couverture lumineuse en fonction du nombre d’espèces.
Les tests de corrélation (Spearman et Kendall)
ne donnent pas de relation significative entre le
nombre d’espèces et la profondeur du cours d’eau, au
risque de 5%. (p>0,05).
Fig. 24 Graphique de la profondeur du cours d’eau, en fonction du
nombre d’espèces.
Les tests de corrélation (Spearman et Kendall) ne
donnent pas de corrélation significative entre le nombre
d’espèces et la largeur du cours d’eau, au risque de 5%.
Ces paramètres seront ensuite testés sur l’assemblage
des espèces de chaque milieu.
Fig. 25 Graphique de la largeur du cours d’eau, en fonction du nombre
d’espèces.
Seule la couverture lumineuse a été déterminée, comme facteur corrélé au nombre d’espèces.

24
On compare la composition en espèces des différents milieux, à l’aide des distances de Bray-Curtis
(Annexe 15)
Le graphe NMDS (Non metric dimensional scaling),
met dans l’espace les distances (dissimilarités) calculées
par la méthode de Bray-Curtis. Deux sites ayant une
composition très semblable seront proches (distance
proche de 0), tandis que deux sites avec une composition
différente, seront éloignés (distance de 1).
On peut voir que les sites de forêt et de zones cultivées
forment deux clusters bien distincts. Les sites en marge de
forêt forment un intermédiaire.
Fig. 26 Non metric dimensional scaling (NMDS), basé sur les indices de similarités de Bray-Curtis.
Fig. 27 Graphes des similarités de bray-Curtis, en fonction des différences de canopy-cover.
Le Mantel test a prouvé que la couverture
lumineuse a une influence significative sur la
composition en espèce (p<0.05), tandis que les
autres facteurs n’influent pas (p>>0.05)
Le test compare la matrice de similarités de
Bray-Curtis, avec une matrice de différence de
couverture lumineuse. Le test s’appuie ensuite
sur les rangs de Spearman, pour montrer la
corrélation (Annexe 17). Pour le graphique, les
valeurs sont arrondies à la dizaine pour une
meilleure lisibilité.
La composition en espèce diffère significativement entre les différents milieux (ANOSIM :
Rglobal=0,692 p=0,02). Ceci reste vrai pour la forêt et les zones cultivées au risque de 5%
(ANOSIM : test pour le couple F-CA, R=0.406 et p=0.029) Annexe 16. Ceci permet alors de réaliser
les tests suivants pour connaître l’influence des différents paramètres sur l’assemblage des
espèces.

25
Après avoir montré les facteurs influant ou non sur la composition en espèces des milieux, on
s’intéresse de plus près à cette dernière, et aux similarités entre milieux. L’objectif est de définir des
groupes (familles, genre, espèces) représentatifs des différents habitats, pour en faire un suivi plus
précis dans le cadre du monitoring. Les toiles d’araignée représentent la répartition des individus des
différents habitats, selon leur famille. On peut voir qu’il existe des différences significatives, dès ce
niveau taxonomique. Le diagramme central présente le nombre d’espèces présentes dans chaque
milieu, les nombre d’espèces partagées par deux milieux, et au centre, partagées par les trois milieux.
Fig. 28 Résumé de la composition en familles, espèces et espèces partagées entre habitats
6 9
3
F CA
FM
27
15
8
13

26
Analyse SIMPER
Elle permet d’observer les espèces qui caractérisent les différents habitats, par le calcul de
contribution des espèces à la similarité (Annexe 18).
Contribution des espèces à la similarité en Forêt (F)
Espèces Abondance moyenne
Similarité moyenne
Contribution [%]
Contribution cumulée [%]
Hetaerina occisa 5,00 25,88 69,39 69,39
Heteragrion erythrogastrum 4,50 10,14 27,17 96,56
Acanthagrion inexpectum 0,50 1,28 3,44 100
Similarité moyenne : 37,30 Tableau 4 Contributions aux similarités en fôret
Pour les zones cultivées (CA). Les faibles contributions sont cachées à partir de 90%
Espèces Abondance moyenne
Similarité moyenne
Contribution [%]
Contribution cumulée [%]
Hetaerina occisa 12,25 16,70 50,09 50,09
Dythemis sterilis 2,50 3,83 11,50 61,59
Argia cupraurea 8,50 3,72 11,16 72,75
Hetaerina fuscoguttata 5,00 3,40 10,21 82,96
Uracis imbuta 3,25 1,50 4,49 87,44
Acanthagrion inexpectum 2,25 1,25 3,74 91,18
Similarité moyenne : 33,34 Tableau 5 Contributions aux similarités en zone cultivée
Dissimilarités du couple F-CA
Espèces Abondance Moyenne F
Abondance Moyenne CA
Dissimilarité moyenne
Contribution [%]
Contribution cumulée
Hetaerina occisa 5,00 12,25 18,20 22,09 22,09
Argia cupraurea 0,50 8,50 12,02 14,59 36,68
Hetaerina fuscoguttata 0,00 5,00 8,63 10,47 47,15
Heteragrion erythrogastrum 4,50 0,00 8,19 9,94 57,09
Uracis imbuta 0,00 3,25 6,26 7,59 64,68
Dythemis sterilis 0,00 2,50 4,33 5,25 69,93
Acanthagrion inexpectum 0,50 2,25 3,84 4,66 74,59
Philogenia zeteki 1,75 0,00 3,78 4,59 79,18
Erythrodiplax fusca 0,00 1,50 2,89 3,51 82,69
Ennalagma novahispaniae 0,00 2,00 2,83 3,44 86,13
Neoneura esthera 0,00 0,75 1,70 2,06 88,19
Erhytrodiplax kimminsi 0,00 0,75 1,41 1,71 89,90
Erythrodiplax funerea 0,00 0,75 1,37 1,67 91,57
Dissimilarité moyenne : 82,40
Tableau 6 contribution aux dissimilarités entre forêt et zone cultivée
Cette analyse doit être complétée par l’analyse suivante, des dissimilarités de composition
entre les milieux, pour définir quelles sont les espèces qui les définissent le mieux.

27
4) Discussion
4.1) Limnologie
Les données récoltées représentent un premier pas dans la connaissance des eaux du Bassin
versant d’Esquinas. Les résultats des mesures hydrologiques et morphométriques, correspondent à ce que
l’on aurait pu imaginer. Les cours d’eau d’ordre faible ont une largeur moins importante que les cours
d’eau d’ordre plus élevé. La vitesse moyenne et la profondeur ne montrent pas ces tendances. Ceci est
sûrement dû au choix aléatoire des zones de relevés. Les tronçons étaient parfois situés dans une zone de
« pool », et parfois sur des tronçons de type « riffle ». Ces deux paramètres ne sont donc pas très
comparables au sein de l’étude.
Les résultats de la sédimentologie ont mis en valeur la différence de composition du lit de la rivière,
entre les prises d’eau du Fila Golfito et les prises du Fila Cruces. La taille des sédiments dépend de
nombreux facteurs très localisés. Ils dépendent de la vitesse du courant, de la profondeur et de
l’inclinaison (« slope »), de la présence d’obstacles au cours d’eau, entre autres.
Les rivières tropicales ont en général de faibles concentrations en constituants chimiques,
comparées aux rivières des zones tempérées. Ceci est dû à un temps de résidence faible, du fait des fortes
pluies journalières, mais la géologie locale joue également un rôle important. On a pu observer de nouveau
une différence entre les rivières du Fila Cruces (Quebrada Mari, Rio Oro) et les autres, du point de vue de la
composition chimique.
Les valeurs de nitrates et phosphates sont élevées dans les zones cultivées, et l’exemple de la
Quebrada Negra montre une influence certaine de l’utilisation des terres sur la qualité des eaux. Certains
phénomènes d’eutrophisation ont été observés en aval de la Quebrada Gamba, lorsqu’elle rejoint le Rio
Bonito. Ce phénomène semblerait dépendre d’un rapport modifié entre le phosphore et l’azote,
notamment par l’apport de fertilisants. Le concept de zones tampons, ou de bandes enherbées pourrait
assurément diminuer ces fuites de fertilisants vers les eaux.
Les cartes créées de la morphologie, et de la sédimentologie des cours d’eau, ont mis en évidence
la succession de pool et riffles de plusieurs portions de rivières du Bassin versant. Leur intérêt sera expliqué
dans la discussion sur la notion de bio-indicateur. Le gros défaut de cette partie est le manque de matériel
pour mesurer la vitesse du cours d’eau et pour pouvoir estimer le débit des cours d’eau. Cet élément sera
nécessaire pour le monitoring dans l’objectif de définir clairement les paramètres des différents milieux.

28
4.2) Entomologie
4.2.1) Richesse en espèces
Les résultats obtenus vont dans le sens d’études comparables sur le sujet. Il semblerait qu’il y ait
une diversité d’espèces plus importante en zone cultivée, qu’en forêt primaire. Estimates (Estimateur de
richesse en espèces) permet de voir que l’inventaire qui a été effectué dans les deux milieux est
relativement complet (F 70% et CA 80%), et laisse supposer qu’un effort de relevé plus important
n’affecterait pas les conclusions générales de cette étude. L’inventaire effectué instaure une base de
référence pour le monitoring des rivières du Bassin versant. Des relevés en saison sèche permettraient
assurément d’accroître le nombre d’espèces identifiées, dans l’aire d’étude. Certaines espèces endémiques
du Costa-Rica, comme Cora semiopaca, sont rapportées pour se révéler en saison sèche.
L’augmentation de richesse en espèces, des forêts primaires vers des zones
plus perturbées a été trouvée dans plusieurs études, et a été expliquée par
l’accroissement de la diversité d’habitats créés par la perturbation anthropique.
Durant l’étude, on a pu trouver en très grande quantité Uracis imbuta dans les
champs de riz. Les flaques créées, le peu de mouvement et la présence de végétation,
sont un habitat idéal pour leurs larves, qui profiteraient même des fertilisants
apportés (SUHLING 1999).
Photographie 2 Vue macroscopique d’une exuvie d’Uracis imbuta rencontrée dans un champ de riz (Oryza sp.)
La végétation des rives, adaptée à une atmosphère très lumineuse, a été citée plusieurs fois
comme facteur déterminant pour l’assemblage des espèces. Lors de cette étude, la couverture lumineuse
est significativement influente sur cet assemblage. Une étude plus précise des macrophytes des rives
seraient pertinentes dans le cadre du monitoring.

29
4.2.2) Similarités
Comme attendues, de fortes dissimilarités existent entre les assemblages des différents milieux.
Certaines espèces peuvent être attribuées à un habitat particulier (Heteragrion erythrogastrum en forêt),
tandis que d’autres s’adaptent aux conditions de plusieurs milieux (Hetaerina occisa).
Les compositions en espèce des sites F1, F2 et F3 sont très semblables, sûrement dû au fait que les
zones soient situées sur le même cours d’eau. Le site F4 est caractérisé par des conditions différentes. Il est
constitué d’un cours d’eau étroit au fond d’une vallée pentue, avec plusieurs cascades, dans lesquelles se
reproduit une espèce rare, qu’est Philogenia zeteki.
L’analyse SIMPER montre que Heteragrion erythrogastrum est responsable de la dissimilarité
calculée entre F et CA. Cette dernière est signalée comme vivant dans des zones sombres, dans lesquels
elle peut s’immobiliser pendant près de quatre heures (ESQUIVEL C., 2004). Cette espèce est donc
considérée comme représentative de la forêt. Les espèces Argia cupraurea et Hetaerina fuscoguttata
peuvent être considérées, comme représentatives des zones cultivées. On ne peut rattacher directement
Hetaerina occisa aux zones cultivées, du fait de son abondance relative importante en forêt, mais elle s’y
développe encore plus facilement.
Les zones cultivées disposent également de surfaces d’ombre importantes, notamment sur le site
CA3 à la couverture lumineuse (cc%) relativement importante, cependant on notera l’absence totale
d’Heteragrion erythrogastrum. On ne connaît pas dans quelle mesure la fragmentation et l’isolation de ces
zones d’ombres et de forêts, empêche sa colonisation, et celle d’autres espèces « forestières ». Le cas
particulier de Mecitogaster modesta peut être discuté. Cette espèce, rapportée pour être strictement
associée à un habitat de type Forêt mature, a été retrouvée en zone cultivée (CA3). Il existe cependant
une « galerie » d’arbres qui rejoint les sites de la forêt, au site CA3. Il serait intéressant d’approfondir cette
notion d’isolation de zones d’ombres forestières, afin de discuter la nécessité d’un continuum forestier le
long des cours d’eau.
La communauté de libellules des zones cultivées est caractérisée par un nombre important
d’espèces et d’individus généralistes (distribués à travers les deux continents Américains). On peut
observer que les proportions relatives d’individus autochtones et généralistes forment un gradient,
représentatif d’une perturbation anthropique croissante de la forêt vers les zones cultivées. Il existe des
différences d’assemblage qui résident à un niveau taxonomique supérieur. En effet, on peut attribuer la
présence de certaines familles, à certains milieux :
- Les Megapodagrionidae sont très fortement rattachés à la forêt.
- Les Libellulidae (Anisoptères), dominent amplement les marges de forêt.
- Les Protoneuridae sont rattachés aux zones cultivées.
La capacité d’autorégulation de température corporelle pourrait être un facteur déterminant de la
répartition des familles (Position de l’obélisque et ventilation importante des Anisoptères).

30
4.3) Limites de l’étude
L’absence du tuteur professionnel aura été une des difficultés majeures de cette étude, cependant
la collaboration d’Odonatologues et de M.Landry (tuteur IUT), aura permis de mener à bien cette
recherche.
Bien qu’on obtienne des résultats significatifs, certains paramètres non mesurés devraient avoir un impact
supplémentaire sur les assemblages en espèces des différents milieux :
1- L’étude a été menée sur une période très courte en saison des pluies. Une investigation en saison
sèche pourrait assurément améliorer l’efficience du monitoring.
2- PICHLER (2006) a reporté 23 espèces de poissons sur la Quebrada Negra, dont certains sont
insectivores. La prédation des poissons sur les larves de libellules devrait également influer sur la
répartition des adultes. Aucune étude n’a été menée sur les poissons des zones cultivées,
cependant une composition différente de poissons insectivores aura un effet différent sur les
communautés d’insectes aquatiques. L’étude de répartition des larves seraient particulièrement
intéressante, bien que peu pratique dans le cadre d’un suivi de population.
3- Les zones de relevés entomologiques étaient disposées sur des cours d’eau d’ordre faible et les
populations d’êtres vivants varient le long d’un cours d’eau (HAWKING 1999). Dans le cadre du
monitoring, la base d’Odonates du Bassin versant doit s’étendre à des zones de relevés sur tous les
cours d’eau.
4- Un certain nombre de paramètres n’ont pu être mesurés, tels que le débit de l’eau, qui influe
assurément sur la présence d’insectes dans l’eau (qui constitue l’alimentation des Odonates), ou
encore la turbidité (influant sur la présence poissons (PICHLER C., 2003). Les données chimiques des
cours d’eau sont en faible quantité du fait du coût important de ces analyses. Une étude globale de
la chimie des eaux du Bassin versant est nécessaire, avant la mise en place du monitoring.
5- Dans les zones où les libellules sont en très grand nombre, il est difficile d’affirmer que chaque
individu relevé était différent. La capture des animaux est une méthode bien plus précise,
cependant les dégâts écologiques provoqués sont incommensurables (Communication personnelle
C. ESQUIVEL).
L’étude présente le désavantage d’un faible effort de relevé, avec une restriction temporelle à la saison des
pluies, cependant ce travail représente une base pour la mise en place du monitoring.

31
4.4) Les Odonates comme bio-indicateur
La mise en place d’un bio-indicateur de la qualité de l’eau, semble être bien plus complexe que de
suivre les populations d’Odonates sur des zones de relevés d’un Bassin versant. Les Odonates entrent dans
un schéma plus global d’êtres vivants du réseau fluvial, et dépendent indirectement de la qualité propre de
l’eau. Aucune relation directe n’a été établie entre les composantes chimiques de l’eau et les populations
d’Odonates. Les facteurs influents mesurés, montrent que l’utilisation des terres a un effet significatif sur
la qualité de l’eau, ainsi que sur les populations d’Odonates, mais les paramètres des différents habitats
restent relativement méconnus à la fin de cette étude.
Cette étude permet de justifier une fois encore, le besoin de protéger les zones sylvestres, comme
réservoir de biodiversité important. L’augmentation de diversité de milieux, créée par les façons agricoles
n’autorise que la colonisation d’espèces généralistes (Uracis imbuta, Dythemis sterilis…), plus tolérantes.
Cependant la présence de Mecitogaster modesta en zone cultivée, permet de se poser quelques questions.
Dans quelle mesure l’isolation de forêts secondaires le long des cours d’eau empêche la colonisation
d’espèce locale ? N’y-a-t’il pas un intérêt à garder un continuum d’arbres le long des cours d’eau ? Ce
concept de continuum est basé sur le concept de corridors biologiques qui autorisent des échanges d’êtres
vivants, assez peu tolérants aux conditions des zones anthropisées (persistance de la dynamique de
métapopulations, RIEDL I., 2000). Ces questions intéressent la station, dans son objectif de reforester des
rives des cours d’eau, par des essences locales et protectrices des rives contre l’érosion importante [non-
encore évaluée, mais possible grâce aux mesures des solides en suspension]. Dans l’objectif de protéger la
qualité de l’eau et de préserver la diversité de sa bio-structure, il serait intéressant d’observer l’effet de la
mise en place de « zones tampons », au sein du continuum préalablement décrit.
Pour que le monitoring d’un Bassin Versant s’appuie sur des données biologiques, il est nécessaire
d’avoir une vue globale de la bio-structure qui lui est associée, c’est pourquoi d’autres macro-invertébrés
benthiques doivent être prospectés, avant de mettre en place un suivi régulier. Il existe des indices qui
permettent de caractériser l’état écologique des masses d’eau, comme l’IGBN (Indice biologique Global
normalisé) (Eaux et rivières Bretagne). Ce dernier utilise plusieurs organismes aquatiques, comme témoins
de la capacité de l’eau à maintenir et entretenir la vie. Le suivi régulier des populations de plusieurs bio-
indicateurs serait nécessaire au bon fonctionnement du monitoring.

32
La méthode de Rapid Bioassessment (RBP III), développée par l’Environmental Protection Agency
(EPA, Etats-Unis, URL : www.epa.gov/OWOW/monitoring/techmon.html), correspond parfaitement aux
besoins du monitoring. Ce protocole est une synthèse des méthodes existantes, utilisées par des agences
de l’eau des Etats-Unis et d’Europe. Il préconise une approche « mono-habitat », qui consiste à
n’échantillonner que les habitats à écoulement rapide, les « riffles ». Les méthodes de bio-surveillance
correspondent au suivi de tous les organismes macro-invertébrés benthiques, sur des tronçons de 100
mètres. Suite à cela, une note de qualité écologique peut être appliquée, en affectant un coefficient de
sensibilité à chaque famille, genre ou espèce présent (donnant l’IGBN). Une note de 1 à 20 est également
attribuée aux paramètres physiques et chimiques, par rapport à une référence que sont les têtes de Bassin
Versant (« sites de référence spécifique »). Il serait intéressant de mettre en place de tels protocoles, après
une étude sur les populations présentes. Pour plus d’informations : Guide de surveillance biologique basée
sur les macro-invertébrés benthiques d’eau douce du Québec, Plan Saint-Laurent.
Il serait également préconisé de mettre en place un suivi permanent des paramètres physico-
chimiques de l’eau, par l’installation d’un matériel enregistreur. La mesure du débit est un paramètre
important à ne pas négliger. Ce matériel permettra d’étudier l’influence d’évènement particulier sur les
populations. Le protocole d’évaluation doit rester relativement simple et rapide. Pour ce faire, un guide
des macro-invertébrés benthiques présents dans les cours d’eau pourrait être réalisé, dans le but de
permettre un suivi continu par les étudiants qui viennent à la station.
Conclusion
Le projet de monitoring n’en est qu’à ses premiers pas, et les limites et questionnements
rencontrés vont permettre d’en améliorer l’efficience. Ces pré-projets établissent une première base de
réflexion, pour la mise en place de protocoles d’évaluation de qualité écologique des eaux. Le suivi des
Odonates aura montré l’influence de l’occupation des terres sur leur répartition, el les relevés
limnologiques forment une première base de données, qu’il sera nécessaire d’alimenter par un suivi
régulier. Le seul suivi de la population des odonates ne peut suffire aux besoins du monitoring, c’est
pourquoi d’autres bio-indicateurs devront être prospectés, dans le cadre d’un protocole bien défini. Un
travail conséquent reste à effectuer avant la mise en place du monitoring, mais le projet semble
prometteur, et compte désormais parmi les objectifs importants de la station de recherche en biologie
tropicale de La Gamba.

33
5) Bilan personnel
Ces pré-travaux auront été particulièrement stimulants durant les onze semaines de stage.
L’absence d’aide du tuteur professionnel était d’abord effrayante, puis s’est rapidement transformée en un
défi d’autonomie. La démarche de recherche est particulièrement plaisante, avec l’établissement de la
problématique, les recherches bibliographiques, les contraintes matérielles et les relevés de terrain. La
nécessité de réajuster en permanence les objectifs est très plaisante, et les contraintes matérielles ont
toujours été essuyées grâce à l’aide des employés de la station.
La thématique étudiée entre parfaitement en accord avec mes attirances. Les problématiques de
l’Eau ont souvent composé mes questionnements. Il m’a paru nécessaire dans le cadre de mes études
d’agronomie, d’approcher les notions d’écologie et de s’en servir comme outil d’évaluation pertinent.
L’idée d’un monitoring permanent me semble être une clé nécessaire à l’étude de l’influence agricole sur
les cours d’eau. J’ai cependant quelques regrets quant aux objectifs de celui-ci. Une simple étude
écologique de la qualité de l’eau, sans autre objectif que la connaissance scientifique m’a paru être une
vision restrictive du monitoring. Au terme de cette étude, on constate encore une fois l’influence
importante de l’agriculture sur la qualité des eaux, cependant aucune initiative de protection ne sera
imaginée par la station. La ferme expérimentale de la station, pourrait jouer un rôle important dans la
protection du réseau fluvial, mais l’absence totale de contacts avec les exploitants locaux lui ferait défaut.
Cette étude aura était extrêmement enrichissante, avec de nombreuses rencontres, de nombreux
relevés, de par l’utilisation de nouveau logiciels et de méthodes statistiques très intéressantes. J’ajouterai
à tout cela, l’immense plaisir d’apprendre la langue espagnole aux couleurs costaricaines ! La participation
à d’autres études comme le projet OMV (Reforestation d’anciennes zones cultivées), ou encore l’étude des
sols de la région avec un groupe d’étudiants de l’Université nationale du Costa-Rica, m’ont permis de voir
la diversité des applications de l’agronomie dans un environnement tropical. La prise en compte de leur
environnement écologique, social, économique et politique, m’auront fait apprécier pleinement ces
projets.
Ce stage de fin d’IUT, m’a confirmé une fois encore, que l’Agronomie est une science très large. Elle
fait appel à de nombreux champs de compétences qui s’entremêlent, et qui constituent sa grande richesse,
avec de multiples applications à travers le monde.

34
Bibliographie
APPENDIX H (2009), Limnology, Water Quality parameters, and conditions and ecoregions, Appendix H,
dernier accès: 5 Avril 2010, http://www.usbr.gov/uc/envdocs/eis/navajo/pdfs/feis-vol2/Append_H.pdf
BARBOUR T. et GERRITSEN J. (1999), Rapid Bioassesment Protocols for use in streams ans wadeable
Rivers: Periphyton, Benthic Macroinvertebrates and fish, Office of Water in Washington D.C.
BERGOEING J.P (1998), Geomorfologia de Costa Rica, San Jose Costa Rica
BOISSEAUNAULT Y. (2007), Bio-indicateurs Eau PDF 2, dernier accès : 15 Avril 2010,
http://openpdf.com/ebook/bio-indicateurs-eau-pdf-2.html
BONNET E. et VAN DE PEER Y. (2002), Zt :a software tool for simple and partial Mantel tests, Journal of
Statistical Software, URL: http://www.psb.ugent.be/~erbon/mantel
BORROR D.J. et WHITE E. (1970), Insects, Peterson field guides
BRIED J.T., ERVIN G.N.(2005), Distribution of Adult Odonata among Localized Wetlands in East- central
Mississippi, Southeastern Naturalist
CALVO C. (1990), Rapporte ejecutivo, dernier accès 5 Avril 2010, http://openpdf.com/ebook/agriculture-
costa-rica-water-pdf-2.html
Centro agronomico technico y de ensenanza (CATIE), http://www.catie.uc.cr
CIA World Factbook, CIA - The World Factbook -- Costa Rica, 2009
CLARKE K.R. et GORLEY R.N (2001), PRIMER V5-User manual/tutorial, PRIMER E
CORBET P., SUHLING F. et SOENDGERATH (2005), Voltinism of Odonata: a review, University of
Edimburgh, transmis par l’auteur le 05/05/2010
CORBET P. (1999), Dragonflies: Behaviour and Ecology of Odonata, Co mstock Publishing ,Associates,
Ithaca, New York.
COLWELL, R. K. (2009), EstimateS: Statistical estimation of species richness and shared species from
samples, Version 8.2. User's Guide and application published at: http://purl.oclc.org/estimates.s
DELBEKE L. et VAN DEUN K. (2000), Multidimensional scaling, University of Leuven, dernier accès le 13
Mai 2010, http://cran.r project.org/web/packages/HSAUR/vignettes/Ch_multidimensional_scaling.pdf
Eaux et rivières Bretagne (Non daté), Les bio-indicateurs, sentinelles de la rivière, dour ha Stêroù Breizh

35
ESQUIVEL C. (2006), Libélulas de Mesoamérica y el Caribe, Editorial INBio
FAO country profile, dernier accès 3 Avril 2010, http://www.fao.org/fi/website
FÖRSTER S. (1999), The dragonflies of Central America, Gunnar rehfeldt braunschweig
HAWKING J.H. (1999), The distribution patterns of dragonflies (Insecta:Odonata) along the Kiewa river,
Australia, and their relevance in conservation assessment, Hydrobiologia
HOLLAND M. (2008), Non metric dimensional scaling, University of Georgia, http://cran.r-
project.org/doc/contrib/Faraway-PRA.pdf
LEWIS O.T., GRIPENBERG, S. (2008), Insect seed predators and environmental change, Journal of applied
ecology,20: pp 24-32
LOUGHEED V.L., PARKER C.A., STEVENSON, R.J. (2007), Using non-linear responses of multiple
taxonomic groups to establish criteria indicative of wetland biological condition, Wetlands
MAECHLER M. (2010), Package cluster for R, R-project CRAN
dernier accès: 13 Mai 2010, http://cran.r-project.org/web/packages/cluster/cluster.pdf
MOOG O. (2006), Key of Odonata (Dragonflies and Damselflies), BOKU/Univ. of Vienna
NOVELO-GUTTIEREZ R. (2008), A comparative study of Odonata (Insecta) assemblages, along an
altitudinal gradient in the Sierra de Coalcoman Mountains, Michoacan, Mexico, Springer Science, transmis
par l’auteur le 13/05/2010
OKSANEN J. (2010), Multivariate Analysis of Ecological Communities in R:Vegan tutorial, R-project CRAN
dernier accès: 13 Mai 2010 http://cc.oulu.fi/~jarioksa/opetus/metodi/vegantutor.pdf
RIEDL I. (2008), Gallery forests in the lowland countryside of Costa-Rica: Corridors for tropical birds?, Univ.
of Vienna
RIPLEY B. (2010), Package MASS for R, R-project CRAN
dernier accès: 13 Mai 2010, http://cran.r-project.org/web/packages/MASS/MASS.pdf
ROLDAN-PEREZ G. (1989), Guío para el estudio de los macroinvertebrates acuáticos, Fondo Fen Colombia
SAHLEN G., EKESTUBBE K. (2001), Identification of dragonflies (Odonata) as indicators of general species
richness in boreal forest lakes, Biodiversity and conservation, 10: pp 119-124
SCHIEMER F., BOLAND K.T. (1996), Perspectives in tropical Limnology, SPB Academic publishing
SCHNEEWEIS S. (2009), Dragonflies of the golfo dulce Region, Costa Rica, INBio
SFO (2008), Code déontologique de l’odonatologue, société française de l’Odonatologie, dernier accès le
07/05/2010, URL : www.libellules.org

36
SUHLING F. et al. (1999), Effects of insecticide applications on macroinvertebrate density and biomass in
rice-fields in the Rhône-delta, France, transmis par l’auteur le 05/05/2010
TRIHADININGRUM et DE PAUW (1994), Use of benthic macroinvertebrates for water quality assessment
of the Blawi river (East Java, Indonesia), Limnology, SPB Academic Publishing, pp 199-219
TSCHELAUT J., PICHLER C. (2005), the river network of the Piedras blancas National Park, BOKU/Univ. of
Vienna
TSCHELAUT J. (2005), Leaf litter decomposition macroinvertebrates in a neotropical lowland stream, Q
Negra Costa-Rica, Diplomarbeit BOKU
WHITEHOUSE et al. (2008), Odonata Exuviae as Indicators of Wetland Restoration Success In Waterloo Region, Ontario Science

Résumé
Les problématiques de la qualité de l’eau sont aujourd’hui omniprésentes à travers le monde entier.
L’Agriculture, les industries et les pollutions urbaines sont majoritairement responsables des perturbations des lacs
et rivières, qui constituent nos zones de captage d’eau potable. L’étude menée à La Gamba, Costa Rica a pour
objectif de montrer l’impact des activités anthropiques, sur l’intégrité écologique du Bassin Versant d’Esquinas
(ordre 5). Une première partie traite de la structure géographique, ainsi que de paramètres physico-chimiques
généraux des cours du réseau fluvial, et dans une seconde partie, sont observées les différences dans la structure
des populations de Libellules et Demoiselles (Odonates), dans différents habitats du Bassin Versant. L’étude a été
réalisée entre les mois d’Avril et Juin 2010, au début de la saison des pluies. Dans l’objectif d’analyser la structure
physique du réseau fluvial, ont été mesurées les profondeurs, largeurs et vitesses moyennes de neuf cours d’eau.
Des échantillons d’eau des différentes rivières ont été analysés selon différents paramètres tels que la température,
l’oxygène dissout, les solides en suspension, le pH, la conductivité, la concentration en nitrates (NO3) et phosphates
(PO4). En plus de ces mesures a été conduite une étude plus précise de la morphologie d’un des cours d’eau, la
Quebrada Negra(ordre 1), avec un suivi journalier du niveau d’eau, l’étude de la réponse à une forte pluie, la
structure sédimentologie, et la vitesse du courant le long d’un tronçon de 100mètres. Le suivi des Odonates a été
effectué en forêt primaire (4 sites), en marge de forêt (2 sites) et en zones cultivées (4 sites), dans l’objectif de
mettre en valeur un gradient de perturbation entre les habitats. Ont été relevés 427 individus de 26 espèces, lors des
vingt relevés. Les assemblages des espèces des différents habitats ont été comparés, et les différents paramètres
relevés ont été testés, afin d’expliquer les différences de répartition. Cette étude aboutie avec une discussion sur les
protocoles pertinents à suivre ou tester pour le monitoring, en questionnant la pertinence des Odonates comme bio-
indicateurs de la qualité des eaux.
Abstract
In today’s world, questions about water quality are a constant issue. Agriculture, industry and urban
pollution are majorly responsible for the disturbance of lakes and rivers, our freshwater basins. This model study in
La Gamba, Costa Rica, shall show the impact of human activities on the biological integrity in the Esquinas’ drainage
basin. While the first part of the study talks about the geographical structure of the river network as well as the
general chemical and physical features of its water, the second, main part of the study assesses the different
population structures of dragonflies and damselflies (odonata) in different habitats within the drainage basin. The
study took place between April and June 2010, therefore in the beginning of the rainy season. In order to examine
the physical structure of the river network, I measured current velocity, depth and width at different spots of nine
rivers within the drainage basin. Of the same nine rivers, I took samples to learn about the water features, checking
temperature, DO, suspended solids, pH, conductivity, NO3 and PO4. In addition to these measurements, I conducted
a more detailed study of the morphology of one of the nine rivers, the Quebrada Negra, analyzing the daily change
of water levels as well as level changes caused by rainfall, the sediment structure and the current velocity
throughout a transect of 100 meters. The study of the dragonflies and damselflies’ distribution took place in the
primary forest (4 sites), at forest margin (2 sites) and in cultivated areas (4 sites). 427 individuals of 26 species were
identified during the twenty sampling periods. The species’ assemblages were compared, and different sampled
parameters were tested, in order to explain the differences in the distribution. In the end, the study leads to a
discussion about the suitability of various protocols for the monitoring.