
RAPPORT DE STAGE Croissance radiale de trois …tree-ajporte.fr/etudiants/Bizet 2009 Licence...
22
Licence 3 : Biologie des Organismes Année universitaire 2008/2009 RAPPORT DE STAGE Croissance radiale de trois espèces arborées de la dune boisée girondine. François BIZET Projet de recherche : Structuration fonctionnelle des peuplements forestiers de la dune girondine et colonisation du Chêne vert Maîtres de stage : Didier BERT - Annabel PORTE (UMR BIOGECO) Quercus ilex Quercus robur Pinus pinaster
Transcript of RAPPORT DE STAGE Croissance radiale de trois …tree-ajporte.fr/etudiants/Bizet 2009 Licence...

Licence 3 : Biologie des Organismes
Année universitaire 2008/2009
RAPPORT DE STAGE
Croissance radiale de trois espèces arborées de la dune boisée girondine.
François BIZET
Projet de recherche :
Structuration fonctionnelle des peuplements forestiers de la dune girondine et colonisation du Chêne vert
Maîtres de stage : Didier BERT - Annabel PORTE (UMR BIOGECO)
Quercus ilex
Quercus robur
Pinus pinaster


Remerciements
La voici donc la joyeuse page de remerciements que tout le monde regarde avec curiosité, celle que je peux me permettre d'écrire avec un peu plus de fantaisie. Je vais tâcher d'être bref, mais sans oublier personne.
À tout seigneur, tout honneur, je me dois de commencer par le « maître des cernes ». Comment ça, tu ne t'es pas reconnu Didier ? Je ne suis pas un expert, mais je n'en connais pas d'autres capables de débusquer aussi bien que toi un cerne sauvage, surtout quand le fourbe se cache derrière une mauvaise année. Merci pour ton aide et pour avoir partagé avec moi une partie de tes connaissances via d'innombrables explications et schémas improvisés. D'ailleurs je ne doute plus que ta thèse fasse un jour partie des classiques de la dendrologie. Travailler avec toi aura été un réel plaisir.
Bien entendu je ne peux pas oublier mon deuxième maître de stage. Merci Annabel de m’avoir accompagné comme tu l’as fait lors de mon arrivée. J'ai beaucoup apprécié nos longues conversations jusque bien au-delà de tes horaires et je n'oublierai pas de faire appel à toi l'an prochain pour m'aider dans mon orientation.
Je continue avec Jean-Sébastien. Merci à toi aussi de m'avoir aidé à m'intégrer dans mes débuts, et bon courage pour la suite, en particulier musicalement. Tu as toute la motivation nécessaire pour réaliser tes ambitions.
Un petit coucou aussi à Francisco, qui m'a fait comprendre que mine de rien, l'anglais, ça se travaille.
Je remercie également toute l'équipe BIOGECO pour son accueil et sa sympathie.
Et pour terminer en beauté, je voudrais saluer respectueusement le cerne 1985. Merci à toi mon ami, sans toi je n'aurais peut être pas eu le temps d'aller jusqu’au bout ou en tout cas pas avec les mêmes résultats. D'ailleurs en y pensant j'aurais peut être craqué et allumé un bon feu avec toute les carottes de Chêne vert.

Sommaire
Structure d’accueil………………………………………………………………………. page 1
Introduction………………………………………………………………………………page 2
Matériels et méthodes…………………………………………………………………… page 3
Inventaire des carottes……………………………………………………………. page 3
Datation des carottes……………………………………………………………... page 3
Mise en forme des données...……………………………………………………... page 4
Estimation de l’âge de chaque individu…………………………………………... page 4
Calcul des surfaces de cernes…………………………………………………….. page 4
Courbes de croissance en fonction de l’âge………………………………………. page 5
Standardisation des courbes……………………………………………………… page 5
Comparaison aux données climatiques…………………………………………... page 5
Résultats…………………………………………………………………………………. page 6
Âge des populations étudiées……………………………………………………... page 6
Comparaison de la croissance de chaque espèce.................................................... page 6
Analyse de la croissance par faciès............................................…………………. page 10
Rôle du climat……………………………………………………………………. page 11
Discussion………………………………………………………………………………... page 13
Conclusion……………………………………………………………………………….. page 14
Bibliographie…………………………………………………………………………….. page 15
Annexes…………………………………………………………………………………... page 16

1
Structure d’accueil
L’intégralité du stage s’est déroulé dans l'Unité Mixte de Recherches (UMR) "Biodiversité, Gènes et Communautés" (BIOGECO). Je n’avais pas un, mais deux maîtres de stage appartenant chacun à une équipe différente : Annabel Porté, rattachée à l’équipe d’Ecologie des Communautés située à Talence sur le campus universitaire de Bordeaux 1 et Didier Bert, qui appartient à l’équipe de Génétique basée sur le site forêt-bois de Pierroton. Le stage a eu lieu pendant les mois de juin et juillet 2009 sur le site de Pierroton, dans le laboratoire de dendrochronologie/microscopie équipé de tout le matériel nécessaire pour faire de la dendroécologie.
L’UMR BIOGECO fait partie de l’Institut National de Recherche Agronomique (INRA). Elle est composée de quatre équipes. En plus des deux équipes dont j’ai fait parti, il existe une équipe d’Entomologie elle aussi basée sur Pierroton, et une équipe de Pathologie, sur le site de la Grande Ferrade à Villenave d’Ornon.
Le programme de recherche de l’UMR est « orienté vers l’analyse des mécanismes régissant l’évolution de la diversité à différents niveaux hiérarchiques (communautés, espèces, populations, gènes) dans une perspective de gestion durable des ressources et des milieux » (site de BIOGECO).
Pour répondre à cet objectif, les thèmes de recherches sont orientés autour de 5 grands axes : 1. Description et organisation de la diversité 2. Dynamique et évolution de la diversité 3. Impacts anthropiques sur la diversité 4. Rôle fonctionnel de la diversité 5. Valorisation
Ce stage s’inscrit dans le deuxième axe grâce à l’étude de la colonisation du Chêne vert et de son impact potentiel sur les populations de chênes pédonculés, ainsi que dans le troisième axe par l’observation de l’impact des pratiques sylvicoles sur les populations étudiées, ainsi que celui des changements climatiques.

2
Introduction
Ce rapport s’inscrit dans la continuité d’une étude visant à comprendre les mécanismes de la colonisation du Chêne vert dans la dune boisée du littoral Atlantique. À l’intérieur du site d’étude, au Nord du lac d’Hourtin, le Chêne vert (Quercus ilex) est actuellement en limite Nord de son aire de répartition. Dans le contexte actuel de réchauffement global, les modélisations climatiques pour 2100 issues du scénario B2 (IPCC, 2001) ont montré une large extension vers l’Ouest et le Nord de la France de cette aire de répartition (équipe CARBOFOR, Badeau et al, 2004).
Le site a été divisé en trois faciès censés présenter un gradient d’humidité croissant de A vers C avec le faciès A en haut de la pente Ouest de la dune (exposé aux vents dominants), le faciès B en haut de la pente Est et le faciès C dans la lède en bas de la pente Est (annexe 1a). Il s’est avéré au final que seul le faciès C est significativement plus humide que les deux autres qui sont identiques d’un point de vue microclimatique et édaphique (Lamy, 2008 ; Urli, 2008).
Les précédents rapports ont mis en évidence que la colonisation du milieu dunaire littoral par le Chêne vert a débuté à la fin du XIXème siècle. Des mesures de discrimination isotopique entre années sèches et humides ont montré que cette espèce présente une sensibilité moindre à la sécheresse que les deux autres espèces arborées du milieu : le Pin maritime (Pinus pinaster) et le Chêne pédonculé (Quercus robur). La distance à la nappe s’est également révélée être un bon critère pour modéliser les répartitions des deux espèces de chênes : le Chêne pédonculé est plus abondant dans le faciès le plus humide (C), alors que la tendance s’inverse dans les faciès plus secs (A et B), où le Chêne vert domine. Concernant la strate de régénération, le Chêne vert domine partout et le Chêne pédonculé n’est présent qu’en faciès B et C, en faible proportion. Cette colonisation pourrait ainsi mener, à plus ou moins long terme, à l’extirpation du Chêne pédonculé de ce milieu alors qu’il s’agit d’une espèce qui contribue à sa valeur patrimoniale.
L’objectif de ce stage est d’analyser et de comparer la croissance radiale du Chêne vert, du Chêne pédonculé et du Pin maritime le long du gradient de sécheresse induit par les faciès dunaires grâce à des analyses dendrochronologiques et statistiques, afin de mieux comprendre les dynamiques forestières de la dune boisée du littoral Atlantique.
Dans une première partie, nous comparerons la croissance des trois espèces entre elles en fonction de l’âge des individus. Puis nous nous intéresserons, au sein de chaque espèce, à l’effet du faciès et donc du stress hydrique sur la croissance. Enfin, nous étudierons la manière dont va réagir chaque espèce en fonction de divers facteurs climatiques (températures, précipitations) et paramètres associés (évapotranspiration, bilan hydrique).

3
Matériels et méthodes
• Inventaire des carottes
Les carottes ont été prélevées durant l’hiver 2008 sur des pins maritimes, chênes pédonculés et chênes verts le long de quatre transects Est-Ouest perpendiculaires à la troisième dune barkhanoïde au nord du lac d’Hourtin (Annexe 1b). Il y a au total 60 carottes par espèces : 15 par transects et dans chacun d’eux, 5 par faciès. Remarque : le Chêne pédonculé étant absent du faciès A, à une exception près, il n’y a que 41 carottes provenant de cette espèce.
• Datation des carottes
Les carottes ont été planées à l’aide d’un cutter pendant qu’elles étaient maintenues dans un étau. Elles ont ensuite été scannées à une résolution de 1200 dpi pour le Pin maritime et le Chêne pédonculé, 1600 dpi pour le Chêne vert (scanner Epson 1640XL). La datation a été faite avec le logiciel d’analyse Windendro (version 2009a, Regent Instrument, Canada) permettant de placer les limites de cernes sur les images scannées afin d’obtenir les largeurs de cernes. L’exactitude des mesures a été vérifiée par interdatation en créant pour chaque espèce une courbe de référence de plus en plus solide au fur et à mesure de l’avancement dans les mesures sur les carottes. Une courbe de référence reflète la croissance moyenne d’une espèce au cours du temps.
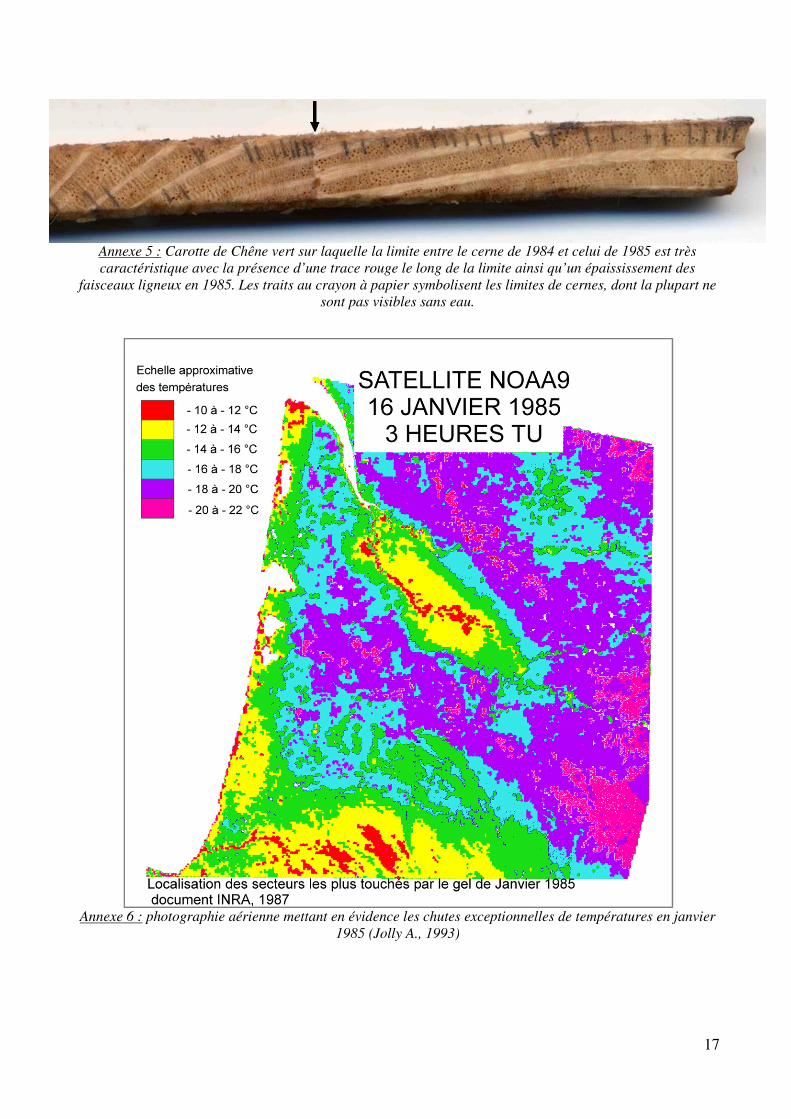
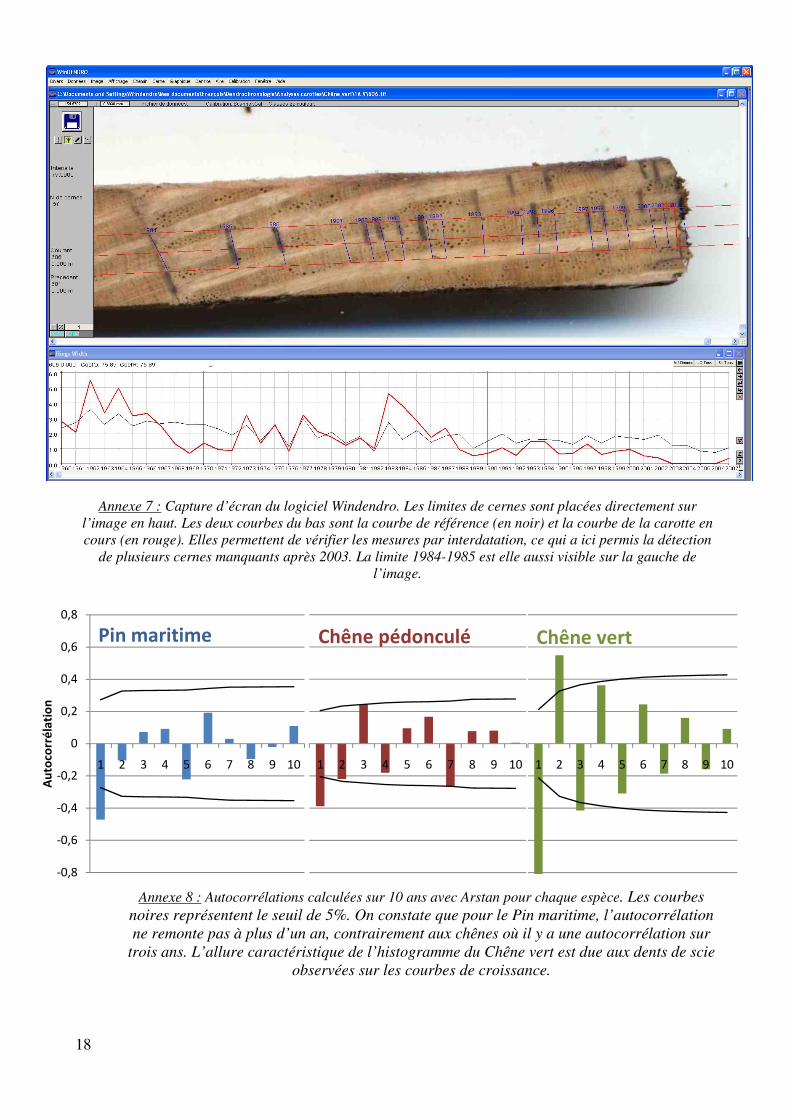
Le Pin maritime et le Chêne pédonculé n’ont pas posé de problèmes majeurs pour la datation. Il faut toutefois se méfier de la présence régulière de faux cernes chez le premier (annexe 2) et faire attention chez le second à ne pas oublier des cernes quand ils sont très serrés, ce qui n’est pas toujours évident (annexe 3). Il en va tout autrement du chêne vert qui a posé nombre de difficultés. En premier lieu, les cernes sont souvent difficiles à voir, ce qui a posé de sérieux problèmes dans les premières carottes pour obtenir une bonne courbe de référence. Les meilleures armes se sont révélées être la persévérance… et l’eau. En effet, plongées sous l’eau, les limites de cernes apparaissent beaucoup mieux (pas toujours hélas). Il faut alors constamment jongler entre la loupe binoculaire où l’on observe la carotte immergée et l’écran pour placer les limites sur l’image. Cela n’a pas empêché dans 90% des cas de devoir planer et observer les deux faces de la carotte pour vérifier certaines limites de cernes. Heureusement, une aide précieuse s’est rapidement révélée : le cerne 1985 (annexe 5). Dans une grande majorité de carottes, ce cerne se retrouve très facilement car il est caractérisé par une trace rouge à la limite 1984-1985 ainsi qu’un épaississement des rayons ligneux en 1985. C’est donc un repère très pratique pour vérifier la datation. Il s’est formé suite au mois de janvier 1985 extrêmement rigoureux avec des températures inférieures à -10°C sur le littoral (annexe 6). Enfin, il arrive de trouver dans certaines carottes des « cernes manquants », c'est-à-dire qu’il manque des cernes qui auraient dû être là en raison d’un stress important dans le développement de l’arbre. Ces cernes sont indétectables sans l’interdatation (annexe 7). Cette méthode utilise une courbe de référence que l’on compare avec la courbe de croissance issue de la carotte en cours d’analyse, il est ainsi possible de découvrir à quels endroits il faut enlever/ajouter des cernes.

4
• Mise en forme des données
Largeur de cerne, largeur du bois initial (ou bois de printemps), largeur de bois final (ou bois d’été), pourcentage de bois initial et de bois final sont sauvegardés depuis Windendro sous format texte (txt). Il n’a toutefois pas été possible de distinguer les limites bois initial/bois final sur le Chêne vert et seules les largeurs de cernes ont été sauvegardées sur cette espèce. Ces fichiers de données se présentent sous forme de lignes qui correspondent chacune à un type de données (comme par exemple les largeurs de cernes) sur une carotte. Pour la réalisation des analyses statistiques sous SAS (version 8.1, SAS Institute, Cary, NC, USA), il a fallu réorganiser ces données sous Excel afin de faire correspondre à chaque ligne un seul cerne. Par manque de temps, toutes les analyses ultérieures n’ont portées que sur les largeurs de cernes.
• Estimation de l’âge de chaque individu
Dans la plupart des cas, la carotte ne passe pas par le cœur de l’arbre. La distance au cœur (dc), est alors déterminée avec une mire. Elle permet une estimation du nombre de cernes de l’arbre qui n’ont pas été mesurés.
������ �� ���� ��� ����é ������ �� ����������� ������� �� 5 ������� ����
L’ensemble « cernes mesurés » + « cernes non mesurés » permet d’obtenir l’âge de l’arbre. Le calcul de ces données a permis de tracer les diagrammes de répartition des arbres en classe d’âge pour chaque espèce.
• Calcul des surfaces de cerne ou BAI (Basal Area Increment, en cm2)
Les surfaces de cernes ont été calculées avec SAS à partir des largeurs de cernes de la manière suivante :
- pour le cerne contenant le cœur de l’arbre, sa surface (S1, cm2) est égale à celle d’un cercle dont le rayon serait la largeur du cerne (L1, cm) : �� �. ���
- Pour le cerne suivant, sa surface (S2, cm2) est égale à celle d’un cercle dont le rayon serait la largeur du cerne (L2, cm) ajoutée à la largeur du cerne précédent (L1, cm), à laquelle on soustrait la surface du cerne précédent (S1, cm2) : �� ���� � ���� � ��
- Ainsi de suite jusqu’au cerne le plus récent :
� ! "# $
%&'
(� � )&

5
• Courbes de croissance en fonction de l’âge
Connaissant le nombre de cernes non mesurés, l’âge de l’arbre et sa dernière année de croissance, il est possible d’attribuer un âge à chaque cerne et ainsi de tracer les courbes moyennes de croissance en fonction de l’âge de l’arbre. La courbe a été tracée en surfaces de cerne pour chaque espèce à partir de la moyenne arithmétique des valeurs pour l’ensemble des carottes. Ces courbes ont ensuite été ajustées sous SAS à l’aide de différents modèles. L’ajustement pour les deux espèces de Chênes a été fait avec un modèle monomoléculaire. Pour le Pin maritime, c’est un polynôme de degré 5 qui s’est avéré le plus pertinent.
• Standardisation des courbes
La courbe de croissance de chaque arbre en fonction de l’âge a été standardisée en faisant le rapport entre la croissance en surface de cernes d’une année donnée et l’ajustement de la courbe de croissance de l’espèce en fonction de l’âge, ce qui donne un indice de croissance (Ic) exprimé en pourcentage.
* +,�-./-0-/1,â45+,�671895:5.9,â45 . 100
Cette méthode permet d’éliminer l’effet de l’âge, qui tend à diminuer la croissance quand l’arbre vieillit, tout en conservant l’effet d’autres facteurs à long terme tels que par exemple de grandes tendances climatiques. On obtient un indice de croissance, exprimé en pourcentage, qui sert à tracer les courbes standardisées de croissance moyenne par espèce et par faciès.
Les courbes de croissance par faciès ont été analysées avec SAS en utilisant un test de comparaison de moyennes multiples (test de Bonferroni) au seuil d’erreur α=0,05 afin de déterminer quelles sont les années où un faciès se démarque des deux autres.
• Comparaison aux données climatiques
Les courbes standardisées de croissance moyenne de chaque espèce ont été comparées sous SAS à de nombreuses variables climatiques (station météo de Bordeaux-Mérignac) afin de trouver les meilleurs coefficients de relation. Les données utilisées sont les moyennes mensuelles de température et pluviométrie depuis janvier 1949. De nombreux indices d’évapotranspiration et de bilan hydrique ont été calculés à partir de ces données, et principalement :
=>?>@AB C@DEFCG�mm/mois� &H I JK&LICM NOE I P
t = température moyenne mensuelle (°C)
I = indice thermique annuel, somme des 12 indices mensuels *7 K9QN�,Q�R
a = fonction de l’indice I = 6,75.10)U�*V� � 7,71.10)Q�*��1,792.10)��*� � 0,49239 F = terme correctif fonction de la durée théorique de l’insolation variant avec la latitude et la saison
=>?>[B\ I L, L&] I �^_ � `L� I � CC � &`�
n = période considérée (jour : n=1 ; décade : n=10 ; mois : n=28, 29, 30 ou 31) t = température moyenne de l’air de la période Rg = rayonnement global en cal/cm2/jour
Puis pour chaque formule, le bilan hydrique a été calculé : a@ ?Bé\FbFCECFA c � =>?
6
Résultats
• Âge des populations étudiées
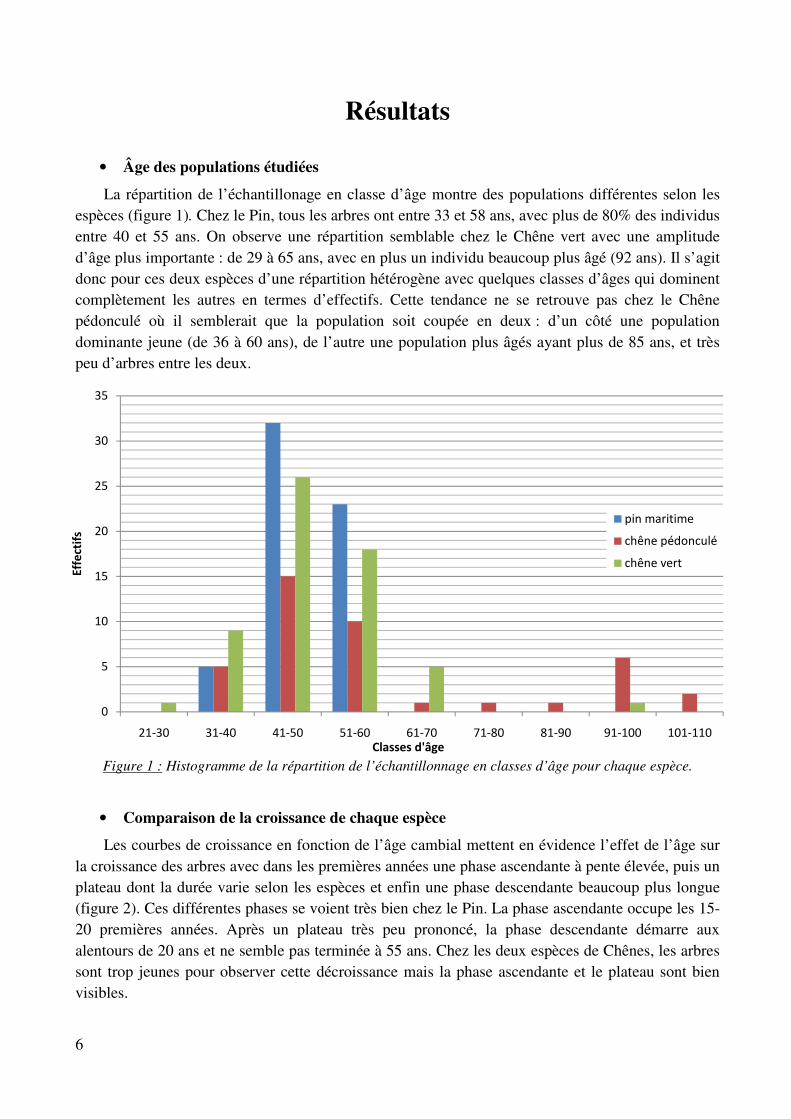
La répartition de l’échantillonage en classe d’âge montre des populations différentes selon les espèces (figure 1). Chez le Pin, tous les arbres ont entre 33 et 58 ans, avec plus de 80% des individus entre 40 et 55 ans. On observe une répartition semblable chez le Chêne vert avec une amplitude d’âge plus importante : de 29 à 65 ans, avec en plus un individu beaucoup plus âgé (92 ans). Il s’agit donc pour ces deux espèces d’une répartition hétérogène avec quelques classes d’âges qui dominent complètement les autres en termes d’effectifs. Cette tendance ne se retrouve pas chez le Chêne pédonculé où il semblerait que la population soit coupée en deux : d’un côté une population dominante jeune (de 36 à 60 ans), de l’autre une population plus âgés ayant plus de 85 ans, et très peu d’arbres entre les deux.
Figure 1 : Histogramme de la répartition de l’échantillonnage en classes d’âge pour chaque espèce.
• Comparaison de la croissance de chaque espèce
Les courbes de croissance en fonction de l’âge cambial mettent en évidence l’effet de l’âge sur la croissance des arbres avec dans les premières années une phase ascendante à pente élevée, puis un plateau dont la durée varie selon les espèces et enfin une phase descendante beaucoup plus longue (figure 2). Ces différentes phases se voient très bien chez le Pin. La phase ascendante occupe les 15-20 premières années. Après un plateau très peu prononcé, la phase descendante démarre aux alentours de 20 ans et ne semble pas terminée à 55 ans. Chez les deux espèces de Chênes, les arbres sont trop jeunes pour observer cette décroissance mais la phase ascendante et le plateau sont bien visibles.
0
5
10
15
20
25
30
35
21-30 31-40 41-50 51-60 61-70 71-80 81-90 91-100 101-110
Eff
ect
ifs
Classes d'âge
pin maritime
chêne pédonculé
chêne vert
7
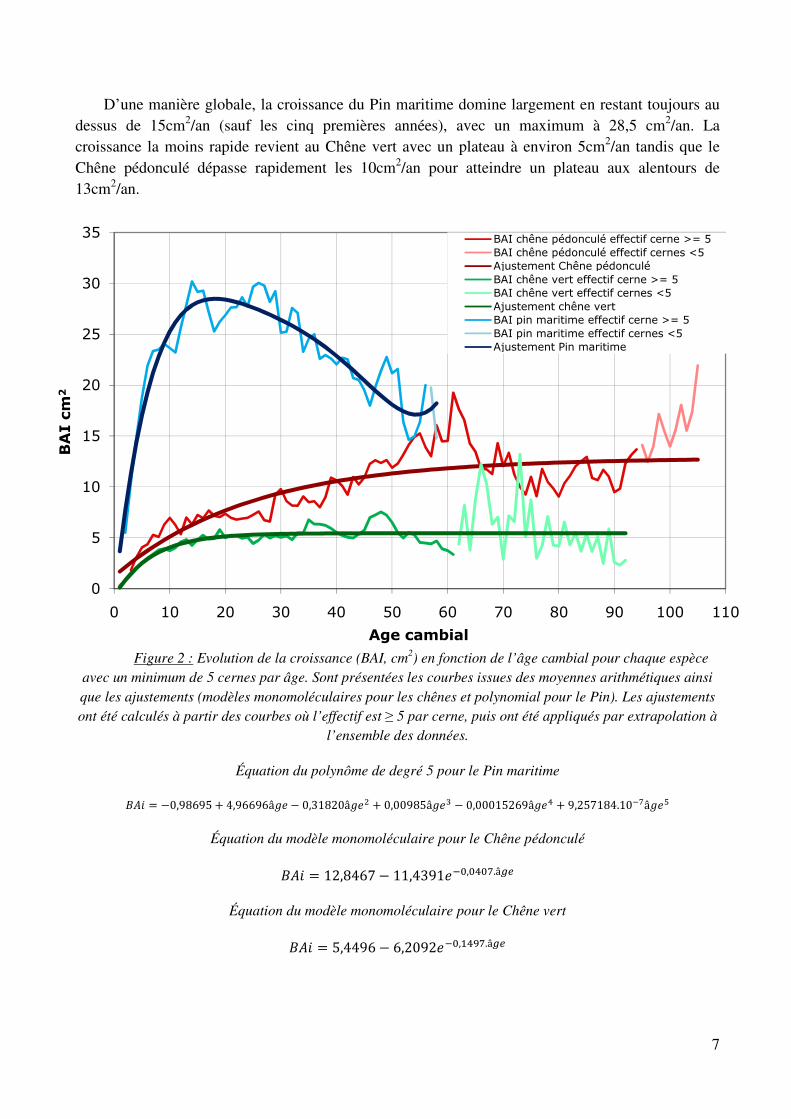
D’une manière globale, la croissance du Pin maritime domine largement en restant toujours au dessus de 15cm2/an (sauf les cinq premières années), avec un maximum à 28,5 cm2/an. La croissance la moins rapide revient au Chêne vert avec un plateau à environ 5cm2/an tandis que le Chêne pédonculé dépasse rapidement les 10cm2/an pour atteindre un plateau aux alentours de 13cm2/an.
Figure 2 : Evolution de la croissance (BAI, cm2) en fonction de l’âge cambial pour chaque espèce avec un minimum de 5 cernes par âge. Sont présentées les courbes issues des moyennes arithmétiques ainsi que les ajustements (modèles monomoléculaires pour les chênes et polynomial pour le Pin). Les ajustements ont été calculés à partir des courbes où l’effectif est ≥ 5 par cerne, puis ont été appliqués par extrapolation à
l’ensemble des données.
Équation du polynôme de degré 5 pour le Pin maritime
+,� �0,98695 � 4,96696� � 0,31820�� � 0,00985�V � 0,00015269�R � 9,257184.10)U�Q
Équation du modèle monomoléculaire pour le Chêne pédonculé
+,� 12,8467 � 11,4391�)e,eReU.â45
Équation du modèle monomoléculaire pour le Chêne vert
+,� 5,4496 � 6,2092�)e,�RfU.â45
0
5
10
15
20
25
30
35
0 10 20 30 40 50 60 70 80 90 100 110
BAI cm2
Age cambial
BAI chêne pédonculé effectif cerne >= 5BAI chêne pédonculé effectif cernes <5Ajustement Chêne pédonculéBAI chêne vert effectif cerne >= 5BAI chêne vert effectif cernes <5Ajustement chêne vertBAI pin maritime effectif cerne >= 5BAI pin maritime effectif cernes <5Ajustement Pin maritime

8
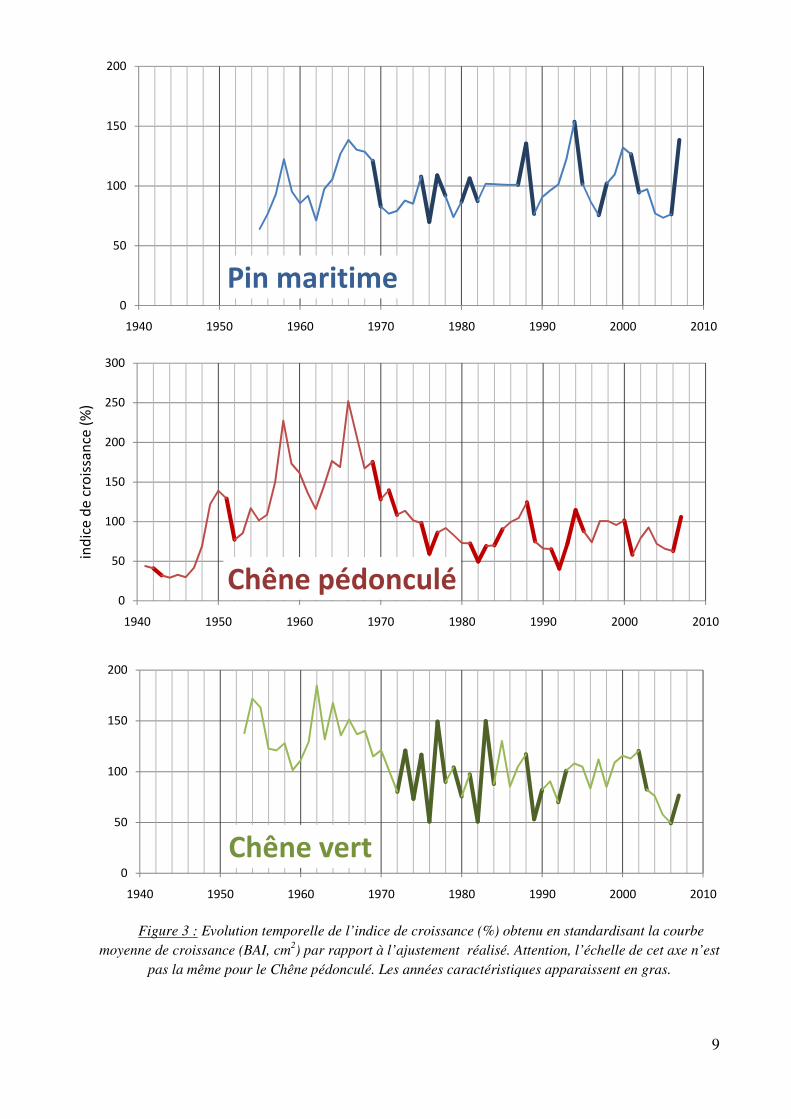
Après standardisation, nous ne pouvons plus comparer les courbes sur un même graphique car elles n’ont pas été standardisées à partir des mêmes courbes d’ajustement. Il est cependant possible d’identifier et de comparer les années caractéristiques pour chaque espèce (figure 3). Une année caractéristique est une année durant laquelle au moins 70% des arbres ont réagi de la même manière, avec une différence de croissance d’au moins 10% par rapport à l’année précédente. Les pourcentages d’années caractéristiques sont de :
- 24% pour le Pin maritime - 24,2% pour le Chêne pédonculé - 27,8% pour le Chêne vert
On retrouve cinq années caractéristiques communes aux trois espèces. 1977 et 2007, avec des étés humides, induisent une croissance forte. Ce n’est pas le cas de 1976 et 1989 aux étés chauds et secs, ni celui de 1982, caractérisé par un mois d’avril pratiquement sans eau qui a pu retarder le démarrage de la saison de croissance. La plupart des années ne sont caractéristiques que chez une, voir deux espèces comme par exemple 1983 et 1993 qui sont des années humides induisant une forte croissance positive chez les chênes
Certaines années se démarquent également car elles ont un effet qui reste observable pendant plusieurs années. C’est le cas par exemple de 1970 qui induit une forte chute de croissance chez le Pin maritime et le Chêne pédonculé : il faut attendre 1975 pour que le Pin retrouve une croissance normale, et 1977 pour que le Chêne pédonculé ait une forte croissance positive. On retrouve un effet similaire, bien que moins prononcé, de 1995 sur ces deux espèces.
En restant dans cette même échelle de temps, deux périodes majeures sont à retenir car elles ont fortement marqué sur plusieurs années la croissance des trois espèces :
- L’année 1989 a induit une très forte chute de croissance (-50% pour le Pin maritime et le Chêne pédonculé, -65% pour le Chêne vert) qui a handicapé les arbres sur une période allant de 3 à 5 ans.
- Le début des années 2000, succession d’années aux étés chauds et secs où seul le Chêne vert semblait résister, jusqu’en 2003 où la canicule de l’été l’a durement fait chuter. Il faut attendre 2007 pour observer un retour à une croissance normale, seulement chez deux des trois espèces (Pin maritime et Chêne pédonculé).
Enfin, il est possible de dégager les grandes tendances de chaque courbe. La croissance ne du Pin maritime est restée stable. Celle du Chêne pédonculé également, malgré une croissance globalement plus faible depuis 1976. Toutefois, chez le Chêne vert, il est possible de visualiser une baisse de la croissance avec une première chute au début des années 70 et une seconde en 2003. Il serait intéressant d’ici quelques années d’analyser de nouvelles carottes pour voir si l’espèce aura fini par se remettre de cette seconde chute.
9
Figure 3 : Evolution temporelle de l’indice de croissance (%) obtenu en standardisant la courbe moyenne de croissance (BAI, cm2) par rapport à l’ajustement réalisé. Attention, l’échelle de cet axe n’est
pas la même pour le Chêne pédonculé. Les années caractéristiques apparaissent en gras.
0
50
100
150
200
1940 1950 1960 1970 1980 1990 2000 2010
Chêne vert
0
50
100
150
200
1940 1950 1960 1970 1980 1990 2000 2010
Pin maritime
0
50
100
150
200
250
300
1940 1950 1960 1970 1980 1990 2000 2010
ind
ice
de
cro
issa
nce
(%
)
Chêne pédonculé
10
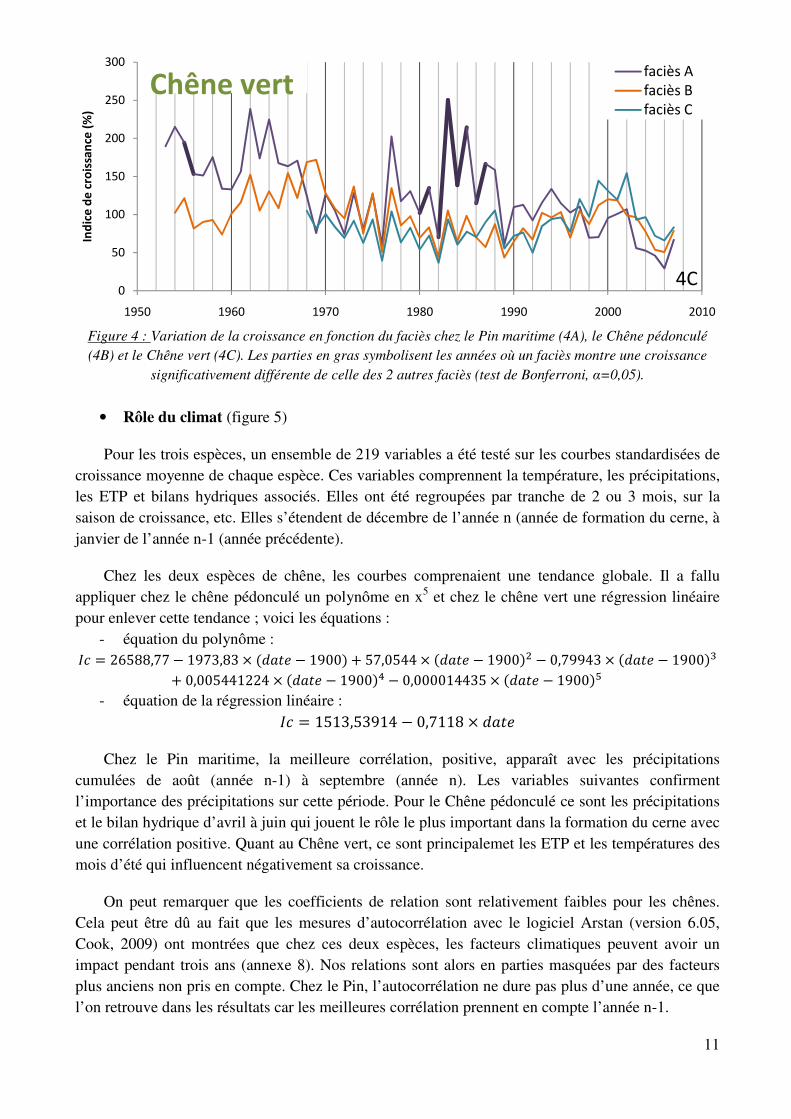
• Analyse de la croissance par faciès
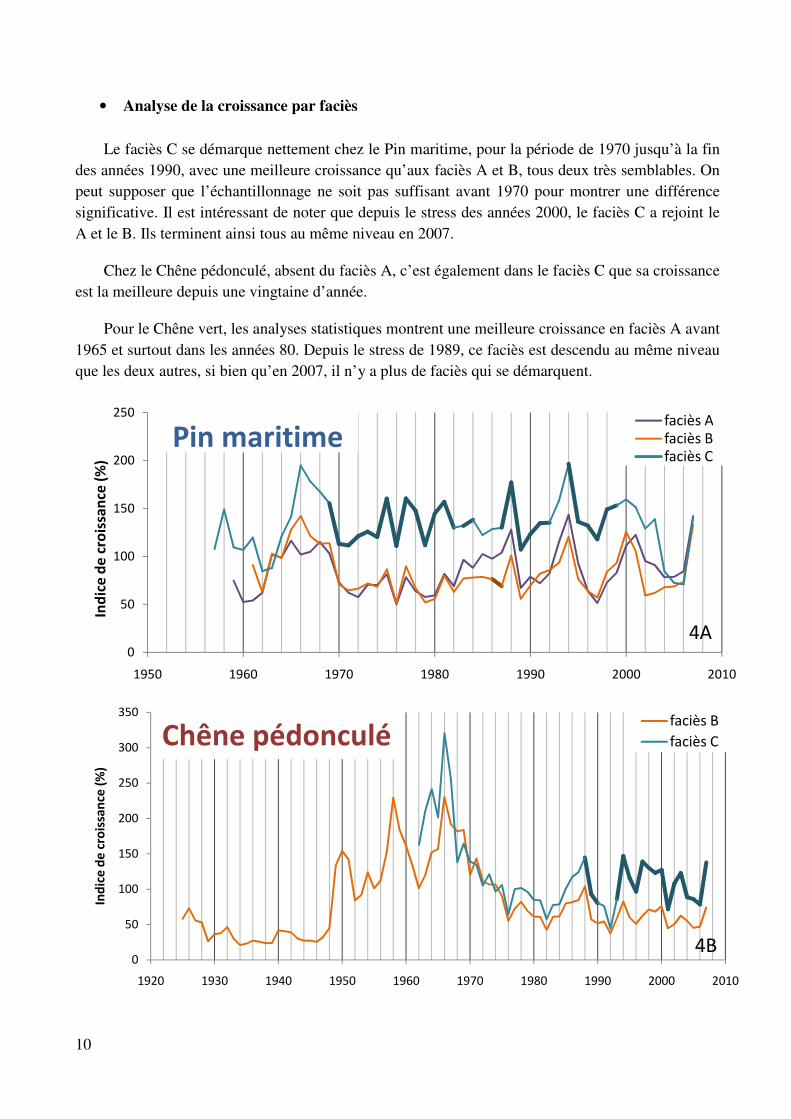
Le faciès C se démarque nettement chez le Pin maritime, pour la période de 1970 jusqu’à la fin des années 1990, avec une meilleure croissance qu’aux faciès A et B, tous deux très semblables. On peut supposer que l’échantillonnage ne soit pas suffisant avant 1970 pour montrer une différence significative. Il est intéressant de noter que depuis le stress des années 2000, le faciès C a rejoint le A et le B. Ils terminent ainsi tous au même niveau en 2007.
Chez le Chêne pédonculé, absent du faciès A, c’est également dans le faciès C que sa croissance est la meilleure depuis une vingtaine d’année.
Pour le Chêne vert, les analyses statistiques montrent une meilleure croissance en faciès A avant 1965 et surtout dans les années 80. Depuis le stress de 1989, ce faciès est descendu au même niveau que les deux autres, si bien qu’en 2007, il n’y a plus de faciès qui se démarquent.
0
50
100
150
200
250
1950 1960 1970 1980 1990 2000 2010
Ind
ice
de
cro
issa
nce
(%
)
faciès A
faciès B
faciès CPin maritime
4A
0
50
100
150
200
250
300
350
1920 1930 1940 1950 1960 1970 1980 1990 2000 2010
Ind
ice
de
cro
issa
nce
(%
)
faciès B
faciès CChêne pédonculé
4B
11
Figure 4 : Variation de la croissance en fonction du faciès chez le Pin maritime (4A), le Chêne pédonculé (4B) et le Chêne vert (4C). Les parties en gras symbolisent les années où un faciès montre une croissance
significativement différente de celle des 2 autres faciès (test de Bonferroni, α=0,05).
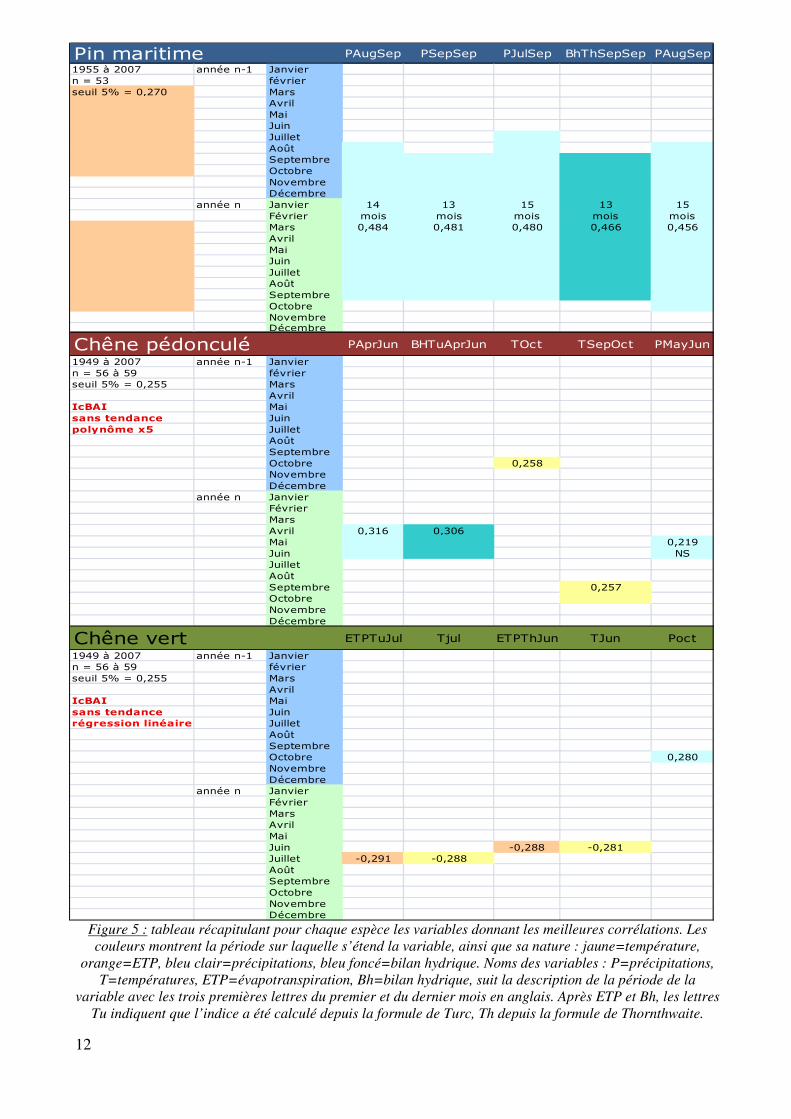
• Rôle du climat (figure 5)
Pour les trois espèces, un ensemble de 219 variables a été testé sur les courbes standardisées de croissance moyenne de chaque espèce. Ces variables comprennent la température, les précipitations, les ETP et bilans hydriques associés. Elles ont été regroupées par tranche de 2 ou 3 mois, sur la saison de croissance, etc. Elles s’étendent de décembre de l’année n (année de formation du cerne, à janvier de l’année n-1 (année précédente).
Chez les deux espèces de chêne, les courbes comprenaient une tendance globale. Il a fallu appliquer chez le chêne pédonculé un polynôme en x5 et chez le chêne vert une régression linéaire pour enlever cette tendance ; voici les équations :
- équation du polynôme : * 26588,77 � 1973,83 I ����� � 1900� � 57,0544 I ����� � 1900�� � 0,79943 I ����� � 1900�V� 0,005441224 I ����� � 1900�R � 0,000014435 I ����� � 1900�Q
- équation de la régression linéaire : * 1513,53914 � 0,7118 I ����
Chez le Pin maritime, la meilleure corrélation, positive, apparaît avec les précipitations cumulées de août (année n-1) à septembre (année n). Les variables suivantes confirment l’importance des précipitations sur cette période. Pour le Chêne pédonculé ce sont les précipitations et le bilan hydrique d’avril à juin qui jouent le rôle le plus important dans la formation du cerne avec une corrélation positive. Quant au Chêne vert, ce sont principalemet les ETP et les températures des mois d’été qui influencent négativement sa croissance.
On peut remarquer que les coefficients de relation sont relativement faibles pour les chênes. Cela peut être dû au fait que les mesures d’autocorrélation avec le logiciel Arstan (version 6.05, Cook, 2009) ont montrées que chez ces deux espèces, les facteurs climatiques peuvent avoir un impact pendant trois ans (annexe 8). Nos relations sont alors en parties masquées par des facteurs plus anciens non pris en compte. Chez le Pin, l’autocorrélation ne dure pas plus d’une année, ce que l’on retrouve dans les résultats car les meilleures corrélation prennent en compte l’année n-1.
0
50
100
150
200
250
300
1950 1960 1970 1980 1990 2000 2010
Ind
ice
de
cro
issa
nce
(%
)faciès A
faciès B
faciès C
Chêne vert
4C
12
Figure 5 : tableau récapitulant pour chaque espèce les variables donnant les meilleures corrélations. Les couleurs montrent la période sur laquelle s’étend la variable, ainsi que sa nature : jaune=température,
orange=ETP, bleu clair=précipitations, bleu foncé=bilan hydrique. Noms des variables : P=précipitations, T=températures, ETP=évapotranspiration, Bh=bilan hydrique, suit la description de la période de la
variable avec les trois premières lettres du premier et du dernier mois en anglais. Après ETP et Bh, les lettres Tu indiquent que l’indice a été calculé depuis la formule de Turc, Th depuis la formule de Thornthwaite.
Saison de croissance du
Pin
Pin maritime PAugSep PSepSep PJulSep BhThSepSep PAugSep
1955 à 2007 année n-1 Janviern = 53 févrierseuil 5% = 0,270 Mars
AvrilMaiJuinJuilletAoûtSeptembreOctobreNovembreDécembre
année n Janvier 14 13 15 13 15Février mois mois mois mois moisMars 0,484 0,481 0,480 0,466 0,456AvrilMaiJuinJuilletAoûtSeptembreOctobreNovembreDécembre
Chêne pédonculé PAprJun BHTuAprJun TOct TSepOct PMayJun
1949 à 2007 année n-1 Janviern = 56 à 59 févrierseuil 5% = 0,255 Mars
AvrilIcBAI Maisans tendance Juinpolynôme x5 Juillet
AoûtSeptembreOctobre 0,258NovembreDécembre
année n JanvierFévrierMarsAvril 0,316 0,306Mai 0,219Juin NSJuilletAoûtSeptembre 0,257OctobreNovembreDécembre
Chêne vert ETPTuJul Tjul ETPThJun TJun Poct
1949 à 2007 année n-1 Janviern = 56 à 59 févrierseuil 5% = 0,255 Mars
AvrilIcBAI Maisans tendance Juinrégression linéaire Juillet
AoûtSeptembreOctobre 0,280NovembreDécembre
année n JanvierFévrierMarsAvrilMaiJuin -0,288 -0,281Juillet -0,291 -0,288AoûtSeptembreOctobreNovembreDécembre

13
Discussion
La gestion sylvicole de la dune boisée peut expliquer plusieurs aspects de la répartition des trois
espèces en classes d’âges. Chez le Pin maritime, la distribution en classes d’âge obtenue correspond à l’âge des parcelles où ont été réalisées les carottes. Le Chêne vert, considéré comme espèce envahissante, a été combattu par l’ONF jusque dans les années 50. Les arbres ont depuis poussés librement, donnant naissance à des individus qui ont aujourd’hui jusqu’à 60 ans, ce qui correspond à ce que l’on retrouve dans l’histogramme des âges (figure 1). À l’inverse, le Chêne pédonculé, présent majoritairement en faciès C, a toujours été favorisé dans la gestion de la forêt. C’est la raison pour laquelle son peuplement est beaucoup plus âgé et compte des arbres qui peuvent atteindre 100 ans.
Les précédents rapports d’étude ont mis en évidence un possible déclin des populations de Chêne pédonculé pouvant conduire à son extirpation malgré sa dominance en faciès C. À la lumière de nos résultats, il n’est aujourd’hui pas possible de confirmer ou d’infirmer cette hypothèse. Il est vrai que depuis 1970, le Chêne pédonculé semble connaître une période difficile avec une croissance inférieure à ce qu’elle était avant (figure 3). Il est vraisemblable que l’espèce a été affaiblie par plusieurs années sécheresse surtout en 1970 et 1976 et que la compétition avec le Chêne vert, libre de pousser depuis une vingtaine d’années, a pu l’empêcher de retrouver son niveau de croissance antérieur. Néanmoins, cette observation se fait aussi chez le Chêne vert avec une période « avant 1970 » à croissance élevée, puis une croissance plus faible jusqu’en 2003. L’espèce subit à cette date une nouvelle importante chute de croissance. Les analyses climatiques ont mis en évidence la sensibilité de l’espèce à la température des mois d’été et elle n’a manifestement pas supporté la canicule de cette année 2003 (figure 5). C’est également suite à cette année que l’on a trouvé le plus de cernes manquants, signe de l’importance du stress subi par cette espèce.
Dans ces circonstances, la bonne croissance observée en 2007 révèle le rôle clé que vont jouer les années suivantes dans l’avenir de la composition de la forêt. Alors que le Chêne pédonculé semble se maintenir à un niveau de croissance équivalent, le Chêne vert va-t-il se remettre de la canicule de 2003 durant les années suivantes ? Ceci est d’autant plus important que sa croissance est belle et bien la plus faible observée (figure 2). Le Pin maritime montre quant à lui une croissance extrêmement stable dans le temps.
Les analyses par faciès montrent que chez le Pin maritime et le Chêne pédonculé, la croissance en faciès A et B est limitée par un facteur que l’on ne trouve pas en faciès C car la croissance y est plus importante (figure 4). S’agit-il de l’eau ? Les mesures d’humidité du sol faites entre février et mai 2008 montrent des différences en faveur du faciès C mais pas au-delà de 20cm de profondeur alors que les racines de chênes vont puiser l’eau bien au-delà de cette profondeur. On peut supposer que les importantes précipitations du début d’année 2008 ont pu saturer le sol en eau, et donc que globalement ce facteur est bien limitant en A et B. Cette hypothèse pourrait se vérifier à l’aide d’une nouvelle campagne de mesure d’humidité du sol en été.

14
Les directives sylvicoles dans la zone étudiée ont aujourd’hui pour but d’orienter la forêt vers des peuplements à feuillus dominant en favorisant le Chêne pédonculé. Cet objectif semble réalisable, en laissant toutefois un point d’interrogation : si cette étude permet de visualiser les grandes tendances dans l’évolution des populations d’arbres adultes, elle est en revanche incapable d’estimer l’avenir des jeunes de la strate de régénération. Et de ce côté-là, on ne sait pas encore si le Chêne pédonculé sera capable de maintenir un niveau de jeunes suffisant pour assurer sa pérennité dans la dune boisée ou si les jeunes chênes vert vont petit à petit étouffer les autres espèces de la strate de régénération, conduisant à plus long terme à l’extirpation non seulement du Chêne pédonculé mais également du Pin maritime.
Il reste encore beaucoup de choses à tirer des résultats obtenus. Tout d’abord, l’autocorrélation à montré des effets du climat pendant trois ans chez les deux espèces de chênes (annexe 8). Il est donc nécessaire pour mieux comprendre les relations espèce-climat de chercher des corrélations en remontant plus loin dans le temps. De plus, les analyses n’ont porté que sur les largeurs de cernes. Il semble intéressant pour aller plus loin de s’intéresser également aux largeurs de bois initial et de bois final chez le Pin maritime et le Chêne pédonculé.
Conclusion
Ce stage aura été l’occasion pour moi de rentrer dans le monde de la recherche auquel je m’intéresse depuis plusieurs années, dans un domaine qui me tient à cœur car c'est celui vers lequel mes études m’ont orienté depuis la terminale : l’écologie. Après les interviews que j'avais effectuées en UPP1 et SVI503 (formation au projet professionnel I et II), ces deux mois ont plus que confirmé mon envie de faire de la recherche. Je vais donc poursuivre mes études comme je le souhaitais vers un master I en écologie puis un master 2 recherche dans un domaine que je ne connais pas encore. En effet, bien que ce stage m'ait ouvert beaucoup d'horizons, je compte encore sur la première année de master pour m'aider à choisir, dans l'écologie, vers quelle discipline poursuivre.

15
Bibliographie
Badeau V., Dupouey J-L., Cluzeau C., Drapier J., Le Bas C., 2004, Modélisation et cartographie de l’aire climatique potentielle des grandes essences forestières françaises, Projet CARBOFOR Tâche D1ECOFOR n°202.17 INRA n°4154 B Rapport final
Cherubini P., Barbara L., Roberto T., Otto U., Werner S., John L., 2002, Identification, measurement and interpretation of tree rings in woody species from Mediterranean climates, Biological reviews of the Cambridge Philosophical Society, 78, p119-148
Favennec J., 1999, Aménagement des forêts littorales : Cas des forêts dunaires du littoral atlantique français, Revue forestière française, n°51, p.217-229
Jolly A., 1993, Estimation par télédétection satellitaire de la récolte annuelle en bois dans la futaie de pin maritime du massif des Landes de gascogne, Thèse Univ. Toulouse, 305 p
Lamy J-B., 2008, Ecophysiologie de Quercus ilex, L. et Quercus robur, L. en limite de leur aire de répartition dans un contexte de changement global récent, MASTER II « Ecologie terrestre », Université de Bordeaux 1-INRA, France
Urli M., 2008, Discrimination isotopique du carbone des espèces arborées dominantes des forêts littorales aquitaines, MASTER I « Ecologie », Université de Bordeaux 1-INRA, France
Université Bordeaux 1-INRA, UMR 1202 BIOGECO, équipe écologie des communautés, 2008, Etude de la colonisation du chêne vert en limite nord de son aire de répartition : fonctionnement de la dune boisée littorale, rapport final-projet ONF Gironde-Réserve naturelle d’Hourtin
UMR BIOGECO, présentation, équipes et thèmes de recherches, consulté le 27/07/09, http://www.pierroton.inra.fr/biogeco

16
Annexes
Annexe 1 : À gauche, représentation schématique du positionnement des trois faciès (A, B et C) sur la dune. À droite, l’emplacement des 4 transects reporté sur une carte (le transect a servi à placer des
stations météo pour obtenir les conditions extérieures au peuplement)
Annexe 2 : Carotte de pin maritime présentant plusieurs faux cernes. Les limites de cernes sont indiquées par les flèches noires. En 1 se trouve un faux cerne particulièrement trompeur. D’autres sont visibles aux
emplacements 2, 3, 4 et même ailleurs.
Annexe 3 : Carotte de Chêne pédonculé mettant en évidence sur la droite (vers l’écorce) que les cernes peuvent être très fins. Les gros vaisseaux au début du bois initial restent visibles, mais on ne voit
pratiquement plus de bois final, ce qui complique la détection des limites de cernes.
1a 1b
17
Annexe 5 : Carotte de Chêne vert sur laquelle la limite entre le cerne de 1984 et celui de 1985 est très caractéristique avec la présence d’une trace rouge le long de la limite ainsi qu’un épaississement des
faisceaux ligneux en 1985. Les traits au crayon à papier symbolisent les limites de cernes, dont la plupart ne sont pas visibles sans eau.
Annexe 6 : photographie aérienne mettant en évidence les chutes exceptionnelles de températures en janvier 1985 (Jolly A., 1993)
18
Annexe 7 : Capture d’écran du logiciel Windendro. Les limites de cernes sont placées directement sur
l’image en haut. Les deux courbes du bas sont la courbe de référence (en noir) et la courbe de la carotte en cours (en rouge). Elles permettent de vérifier les mesures par interdatation, ce qui a ici permis la détection
de plusieurs cernes manquants après 2003. La limite 1984-1985 est elle aussi visible sur la gauche de l’image.
Annexe 8 : Autocorrélations calculées sur 10 ans avec Arstan pour chaque espèce. Les courbes noires représentent le seuil de 5%. On constate que pour le Pin maritime, l’autocorrélation ne remonte pas à plus d’un an, contrairement aux chênes où il y a une autocorrélation sur
trois ans. L’allure caractéristique de l’histogramme du Chêne vert est due aux dents de scie observées sur les courbes de croissance.
-0,8
-0,6
-0,4
-0,2
0
0,2
0,4
0,6
0,8
1 2 3 4 5 6 7 8 9 10
Au
toco
rré
lati
on
Pin maritime
1 2 3 4 5 6 7 8 9 10
Chêne pédonculé
1 2 3 4 5 6 7 8 9 10
Chêne vert















![Page de garde 1 - bu.umc.edu.dz · NOMENCLATURE r coordonnée radiale [m] r* coordonnée radiale adimensionnelle ( ) r Di R rayon du conduit [m] Ra nombre de Rayleigh ([( ) ]3 air](https://static.fdocuments.fr/doc/165x107/5b9ba42409d3f2d06f8d3bc9/page-de-garde-1-buumcedudz-nomenclature-r-coordonnee-radiale-m-r-coordonnee.jpg)

