
Principaux caractères différentiels : Eucaryotes...
16
1 Principaux caractères différentiels : Eucaryotes, procaryotes, virus. - Eucaryotes : cellule à noyau et à compartiments. - Procaryotes : pas de noyau individualisé. On distingue 3 groupes de procaryotes : ¤ archaebactéries. ¤ eubactéries : 3 groupes : ∝ cyanobactéries ( ancien nom : algues bleues ) ∝ bactéries photosynthétiques ∝ bactéries non photosynthétiques. ¤ virus = structure non cellulaire : acaryote. Les archaebactéries : ces bactéries archaïques sont les plus anciennes. Elles vivent dans des milieux inhospitaliers comme la boue des marais ( bactéries méthanogènes : dégagent du méthane ), dans l'eau salée ( elles sont alors halophiles ), à des températures élevées ( 70 à 80 °C : elles sont thermophiles, et vivent donc au niveau des sources chaudes océaniques.) On a aussi des bactéries chimio-synthétiques qui produisent de l'énergie à partir d'H 2 S. Elles sont à la base des chaînes alimentaires ( nourrissent les coquillages). On peut dire que ce sont de nouvelles formes de vie car elles sont à la base d'îlots de vie. Elles ont une particularité génétique qui est le morcellement des gènes. Les archaebactéries ont des introns ( comme les eucaryotes ) alors que les eubactéries n'en ont pas. Cela signifie que les archaebactéries ont de l'information génétique qui ne sert à rien : les eubactéries ont un développement plus rapide ( moins d'information à copier ). Les cellules eucaryotes sont-elles dérivées des archaebactéries ? On peut utiliser l'argument des introns. De plus, les archaebactéries sont apparues il y a 3,5 milliards d'années, alors que les eucaryotes ne sont apparues qu'il y a 1,5 milliards d'années.
Transcript of Principaux caractères différentiels : Eucaryotes...

1
Principaux caractères différentiels : Eucaryotes, procaryotes, virus.
- Eucaryotes : cellule à noyau et à compartiments. - Procaryotes : pas de noyau individualisé. On distingue 3 groupes de procaryotes : ¤ archaebactéries. ¤ eubactéries : 3 groupes : ∝ cyanobactéries ( ancien nom : algues bleues ) ∝ bactéries photosynthétiques ∝ bactéries non photosynthétiques. ¤ virus = structure non cellulaire : acaryote. Les archaebactéries : ces bactéries archaïques sont les plus anciennes. Elles vivent dans des
milieux inhospitaliers comme la boue des marais ( bactéries méthanogènes : dégagent du méthane ), dans l'eau salée ( elles sont alors halophiles ), à des températures élevées ( 70 à 80 °C : elles sont thermophiles, et vivent donc au niveau des sources chaudes océaniques.) On a aussi des bactéries chimio-synthétiques qui produisent de l'énergie à partir d'H2S. Elles sont à la base des chaînes alimentaires ( nourrissent les coquillages). On peut dire que ce sont de nouvelles formes de vie car elles sont à la base d'îlots de vie. Elles ont une particularité génétique qui est le morcellement des gènes.
Les archaebactéries ont des introns ( comme les eucaryotes ) alors que les eubactéries n'en ont pas. Cela signifie que les archaebactéries ont de l'information génétique qui ne sert à rien : les eubactéries ont un développement plus rapide ( moins d'information à copier ). Les cellules eucaryotes sont-elles dérivées des archaebactéries ? On peut utiliser l'argument des introns. De plus, les archaebactéries sont apparues il y a 3,5 milliards d'années, alors que les eucaryotes ne sont apparues qu'il y a 1,5 milliards d'années.
2
I) Caractères distinctifs eucaryotes / procaryotes.
1) Organisation des procaryotes.
Elles peuvent avoir une structure sphérique et sont alors appelées coques. On a par exemple les monocoques, les diplocoques, les streptocoques ( les cellules s'accolent pour former des fibres) et les staphylocoques ( elles forment des amas et ont un pouvoir infectieux important). Elles peuvent avoir également une structure en bâtonnets, et s'appellent des bacilles. Enfin, on peut avoir des bactéries longues et flexueuses ( exemple des spirochètes).
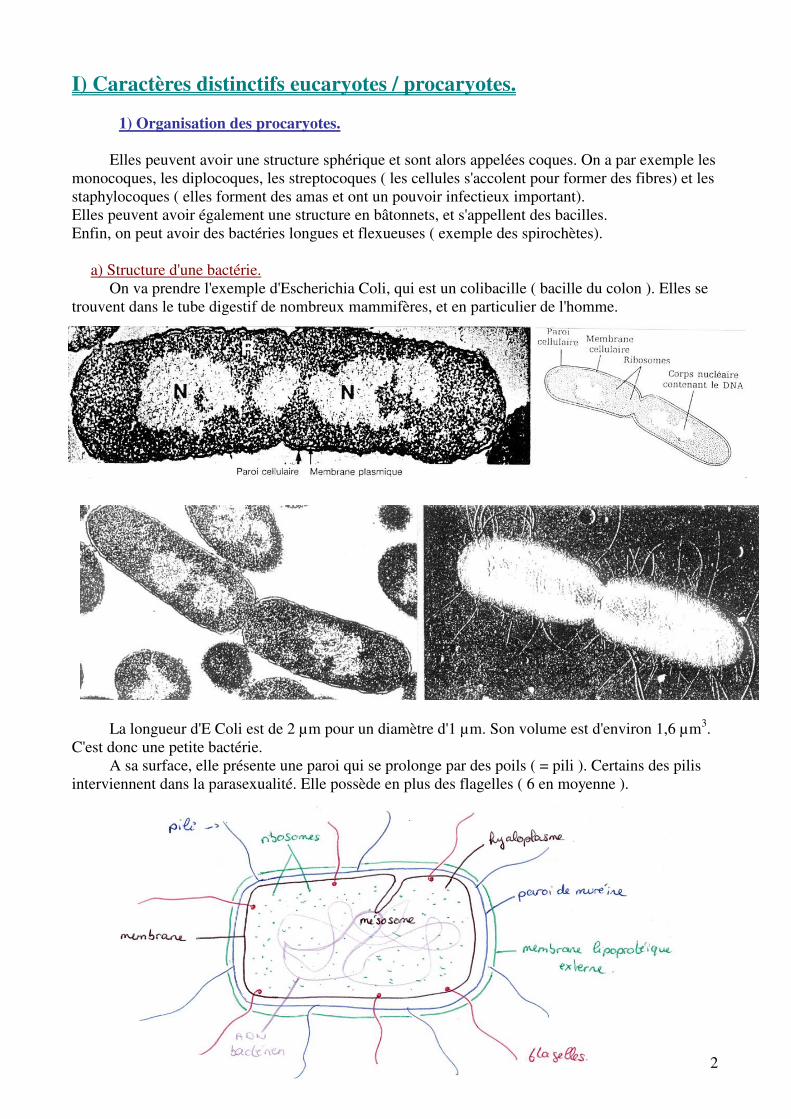
a) Structure d'une bactérie. On va prendre l'exemple d'Escherichia Coli, qui est un colibacille ( bacille du colon ). Elles se
trouvent dans le tube digestif de nombreux mammifères, et en particulier de l'homme.
La longueur d'E Coli est de 2 µm pour un diamètre d'1 µm. Son volume est d'environ 1,6 µm3. C'est donc une petite bactérie.
A sa surface, elle présente une paroi qui se prolonge par des poils ( = pili ). Certains des pilis interviennent dans la parasexualité. Elle possède en plus des flagelles ( 6 en moyenne ).
3
Il n'existe pas de compartiment cellulaire : il n'y a donc pas de mitochondrie ni de RER, mais elle présente un nucléoïde où se trouve l'ADN. Il est donc regroupé dans la cellule. C'est un chromosome circulaire sans histone, fixé au mésosome .On trouve également des ribosomes. Les documents représentent E Coli en division .
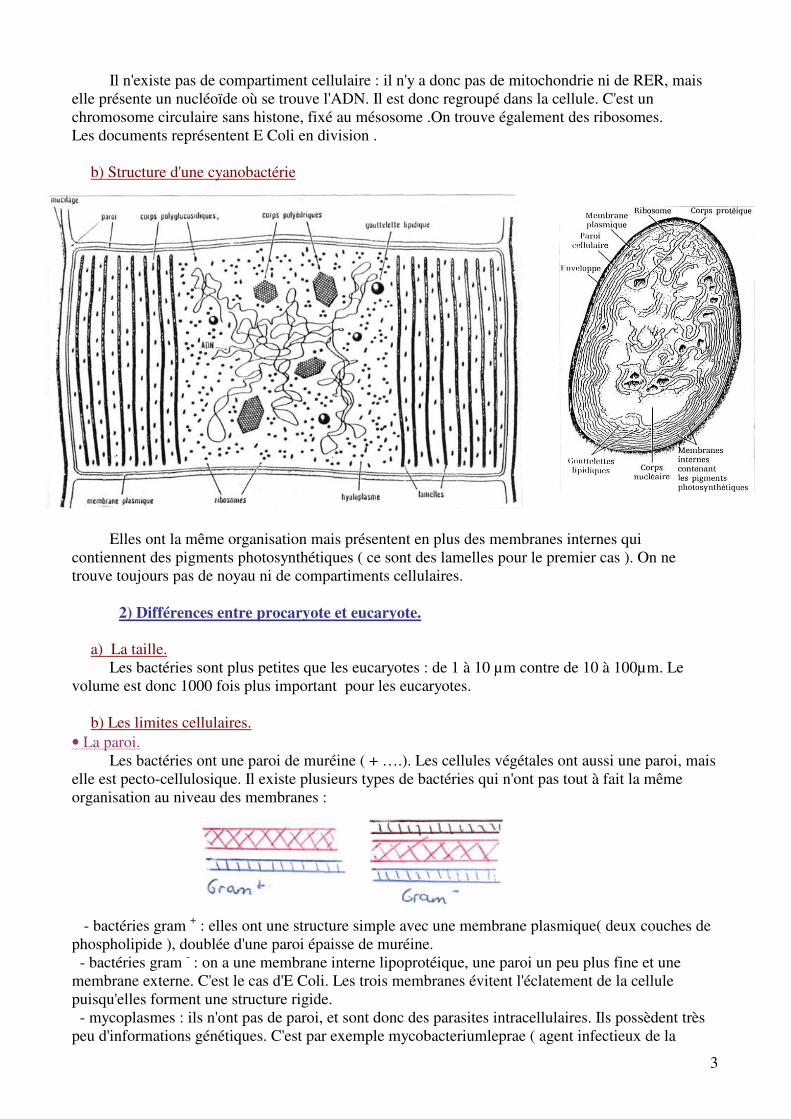
b) Structure d'une cyanobactérie
Elles ont la même organisation mais présentent en plus des membranes internes qui contiennent des pigments photosynthétiques ( ce sont des lamelles pour le premier cas ). On ne trouve toujours pas de noyau ni de compartiments cellulaires.
2) Différences entre procaryote et eucaryote.
a) La taille. Les bactéries sont plus petites que les eucaryotes : de 1 à 10 µm contre de 10 à 100µm. Le
volume est donc 1000 fois plus important pour les eucaryotes.
b) Les limites cellulaires. • La paroi.
Les bactéries ont une paroi de muréine ( + ….). Les cellules végétales ont aussi une paroi, mais elle est pecto-cellulosique. Il existe plusieurs types de bactéries qui n'ont pas tout à fait la même organisation au niveau des membranes :
- bactéries gram + : elles ont une structure simple avec une membrane plasmique( deux couches de phospholipide ), doublée d'une paroi épaisse de muréine. - bactéries gram - : on a une membrane interne lipoprotéique, une paroi un peu plus fine et une membrane externe. C'est le cas d'E Coli. Les trois membranes évitent l'éclatement de la cellule puisqu'elles forment une structure rigide. - mycoplasmes : ils n'ont pas de paroi, et sont donc des parasites intracellulaires. Ils possèdent très peu d'informations génétiques. C'est par exemple mycobacteriumleprae ( agent infectieux de la

4
lèpre) et mycobacterium tuberculosis ( tuberculose ). Ils possèdent à leur surface des molécules résistant à l'hydrolyse des enzymes contenues dans les lysosomes. - archaebactéries : elles ont une grande diversité de nature chimique de paroi. • la capsule.
Elle est constituée de polysaccharides et se situe à l'extérieur de la paroi, mais toutes les cellules n'en ont pas. • la membrane plasmique.
Toutes les bactéries en ont une et elle est interne. Elle est de nature lipoprotéique mais ne possède pas de cholestérol. C'est un repli interne de cette membrane qui fixe le matériel génétique .
c) Particularités cytoplasmiques. • Pas de cytosquelette .
Les bactéries ne possèdent pas de centriole ni de microtubules. Pourtant E Coli possède 6 flagelles. Ici, les flagelles sont constituées de flagelline : elles ont donc une structure moléculaire et une organisation très différente .
Le déplacement est permis par une rotation du flagelle et non pas par un battement de celui-ci ( moteur électrique ). Le flagelle s'appelle d'ailleurs le rotor stator : l'énergie mise en jeu provient d'un gradient de protons ( flux de protons qui fait tourner ) . La rotation peut s'inverser , donc on trouve à la base un inverseur . Cela permet de faire marche arrière . • Pas de compartiment Il n'y a pas de noyau, pas de réticulum, pas de vésicules. Il y a une exception avec les membranes internes des cyanobactéries contenant les molécules photosynthétiques. Dans ce cas, les membranes font des replis mais ne forment pas de compartiment. • Des ribosomes
Ils ne sont pas fixés sur du RE mais sont libres. Ils se fixent sur l'ARN en cas de transcription et on appelle cette association des polysomes .
d) Pas d'organites convertisseurs • Pas de mitochondries.
Les bactéries ne sont pas capables de réaliser des oxydations dans des endroits particuliers : en fait, la bactérie est une grande mitochondrie.
La membrane des bactéries contient des chaînes d'oxydo-réduction comme dans les membranes des mitochondries. Elles sont donc capables de fabriquer de l'ATP. Les mitochondries

5
des eucaryotes sont assimilées à d'anciennes bactéries qui seraient rentrées dans une cellule eucaryote ancestrale ne produisant de l'ATP que par fermentation. C'est la théorie endosymbiotique.
Au départ , la symbiose s'est établie : c'était une association à bénéfices réciproques. La bactérie est protégée, elle peut se nourrir, et en échange, elle livre des molécules d'ATP qu'elle produit en grande quantité. Quels sont les arguments ?
- les mitochondries possèdent de l'information génétique qui a la même forme que celle des bactéries : elle est circulaire et sans histone.
- elles possèdent des mitoribosomes. - elles présentent une double membrane. La membrane interne de la mitochondrie
correspond à la membrane interne de la bactérie, et la membrane externe serait une membrane d'endocytose. Le rôle de la paroi était d'éviter l'éclatement mais il n'y a plus de variation de la pression osmotique de la cellule, donc la paroi a disparu, étant inutile. - le mode de division des mitochondries est identique à celui des bactéries. • Pas de chloroplastes.
C'est le cas des cyanobactéries. En fait, c'est la même chose que pour les mitochondries, et les cyanobactéries seraient de grands chloroplastes. On a la aussi une théorie d'endosymbiose : les chloroplastes des cellules eucaryotes seraient d'anciennes bactéries endocytées par une cellule ancestrale, avec peut-être déjà des mitochondries, et il se serait formé une symbiose. La cyanobactérie protégée permet à la cellule se fabriquer de la matière organique à partir de matière minérale et de lumière. Quels sont les arguments ?
- les chloroplastes possèdent de l'information génétique identique à celle des cyanobactéries ( ADN circulaire sans histones ).
- Présence de ribosomes ( = plastoribosomes ) dans le stroma → ce sont donc des organismes semi-autonomes.
- les chloroplastes présentent une membrane interne, qui serait la membrane de la cyanobactérie et une membrane externe qui serait une membrane d'endocytose. La paroi a régressé puisqu'il n'y a plus de variation du potentiel osmotique.
Actuellement, les mitochondries et les chloroplastes sont dépendant de l'ADN du noyau. Les
protéines de la mitochondrie et du chloroplaste sont codées par l'ADN du noyau et synthétisées dans le hyaloplasme. La situation de symbiose se serait transformée en situation de dépendance. Il n'y a plus de semi-autonomie. Quels sont alors les arguments ?
- les gènes ont migrés de l'ADN mitochondrial vers l'ADN nucléaire. - le rendement des mitochondries est amélioré par des protéines venant de l'ADN du noyau.
De plus, il s'est établit une différence entre les cellules végétales et les cellules animales.

6
e) Le matériel génétique. Les bactéries possèdent ce qu'on peut appeler un chromosome circulaire présent en un seul
exemplaire. Il est fixé sur un cœur protéique, lui-même fixé au mésosome.
Les eucaryotes présentent de longs filaments d'ADN avec des histones. Les chromosomes existent parfois en 2 exemplaires et sont souvent nombreux.
Les eubactéries n'ont pas d'intons. On a donc eu une perte de ces introns lors de l'évolution, si on considère les archaebactéries comme des souches. Elles ont moins d'information génétique mais leur réplication est plus rapide.
On peut également trouver des plasmides ( où se trouvent des gènes de résistance aux antibiotiques) et des épisomes. Les 2 sont du matériel génétique circulaire.
f) Efficacité dans les phénomènes de prolifération. La prolifération est rapide si on place E Coli dans des conditions optimales ( température et
nutrition ) : elles se divise toutes les 20 mn. On peut avoir une reproduction par division : à partir d'un individu, on en obtient 2. On peut
avoir également un phénomène de parasexualité ( échange de gènes ), qui est assez intéressant puisqu'on obtient des souches recombinées au lieu de clones, lors de la division. De plus, la transcription et la traduction sont simultanées : il n'y a pas de compartiments cellulaires donc il n'est pas nécessaire d'avoir une migration pour obtenir les protéines voulues.
Pour les eucaryotes, on avait : transcription → pré ARN → ARN dans le noyau puis traduction → maturation dans le hyaloplasme. Les maturation se font en passant dans le RER, le corps de Golgi… C'est par exemple le phénomène de glycosylation.
Le procaryotes ont un grand pouvoir évolutif car ils sont haploïdes : si un gène est muté, le
phénotype sera modifié tout de suite. Les bactéries sont souvent pathogènes ou toxiques à cause des toxines qu'elles rejettent.

7
Sujets de colle : - différence eucaryote / procaryote. - Particularité des bactéries… II) Les virus.
Leur originalité structurale leur donne une certaine originalité biologique. On a en fait
soupçonné leur existence avant même de pouvoir les observer. Il y a eu tout d'abord la découverte des bactéries : un filtre de porcelaine pouvait les retenir, mais le liquide ( ou filtrat ) avait toujours un pouvoir pathogène. C'est donc qu'il existait des agents pathogènes plus petits que les bactéries, et donc non visibles au microscope optique.
On parlera de virus au sens large, c'est à dire quand on parle d'une population, et d'un virion quand on parle d'un seul individu.
1) Organisation d'un virus.
Les virus sont constitués d'information génétique sous forme d'ADN mono-caténaire ou bi-caténaire, ou d'ARN.
La protection de l'information génétique est permise par une capside protéique, présente chez tous les virus. On peut avoir en plus une enveloppe lipoprotéique ( cas de la grippe et du VIH ). On peut également avoir des enzymes associées ( mais pas dans tous les cas ).
a) Virions hélicoïdaux : le VMT. Le VMT est le virus de la mosaïque du tabac. Il se développe dans les feuilles de tabac et
provoque la mort des cellules qui deviennent alors brunes. Il apparaît donc des tâches brunes sur les feuilles vertes.
Il se présente sous forme de baguette à peu près circulaire, et mesure 300 nm de long pour 17 nm de diamètre.
La capside est constituée de 2200 unités qui sont des protéines à structure tertiaire. Les 2200 unités sont toutes identiques : un seul gène suffit pour coder toutes les protéines de protection. Ces protéines sont organisées en structure hélicoïdale. On peut donc dire que la capside est une protéine à structure quaternaire. Chaque capsomère est constitué de 158 acides aminés, donc moins de 500 nucléotides sont nécessaires, ce qui est assez peu. L'information génétique est de l'ARN, qui possède en tout 6000 nucléotides. Il est enroulé dans un sillon de la capside. → C'est un édifice inerte, déshydraté, et même acellulaire.
8
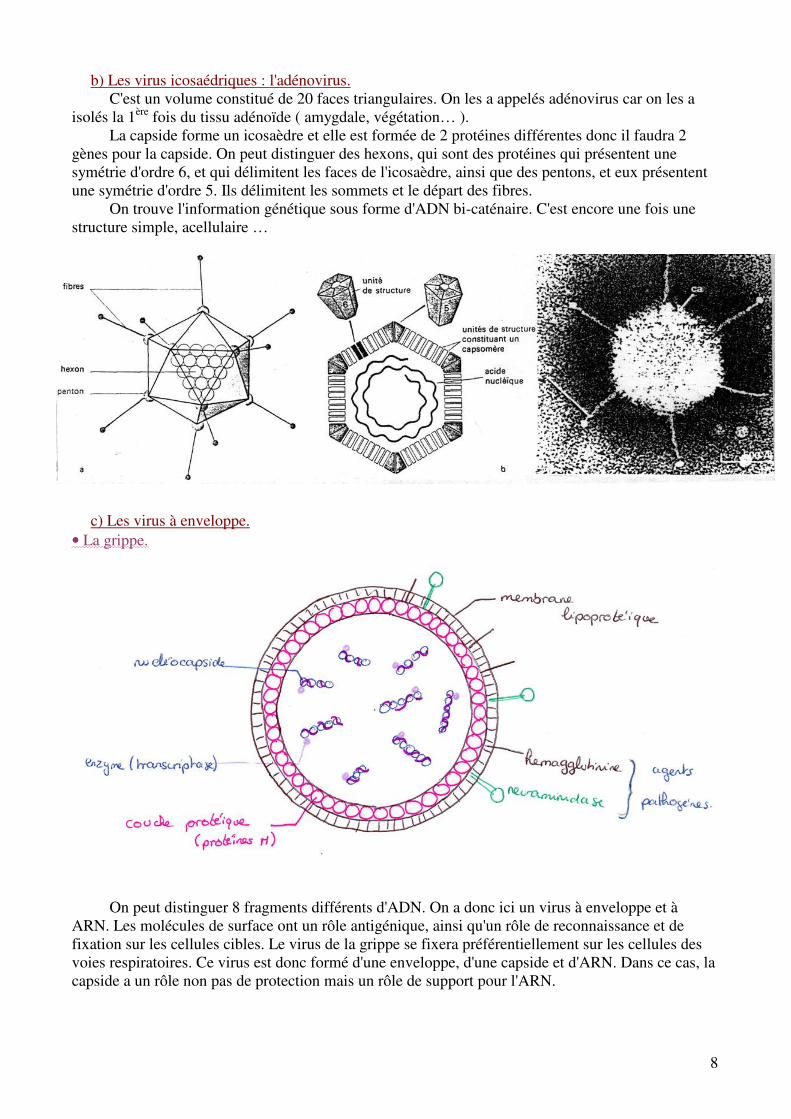
b) Les virus icosaédriques : l'adénovirus. C'est un volume constitué de 20 faces triangulaires. On les a appelés adénovirus car on les a
isolés la 1ère fois du tissu adénoïde ( amygdale, végétation… ). La capside forme un icosaèdre et elle est formée de 2 protéines différentes donc il faudra 2
gènes pour la capside. On peut distinguer des hexons, qui sont des protéines qui présentent une symétrie d'ordre 6, et qui délimitent les faces de l'icosaèdre, ainsi que des pentons, et eux présentent une symétrie d'ordre 5. Ils délimitent les sommets et le départ des fibres.
On trouve l'information génétique sous forme d'ADN bi-caténaire. C'est encore une fois une structure simple, acellulaire …
c) Les virus à enveloppe.
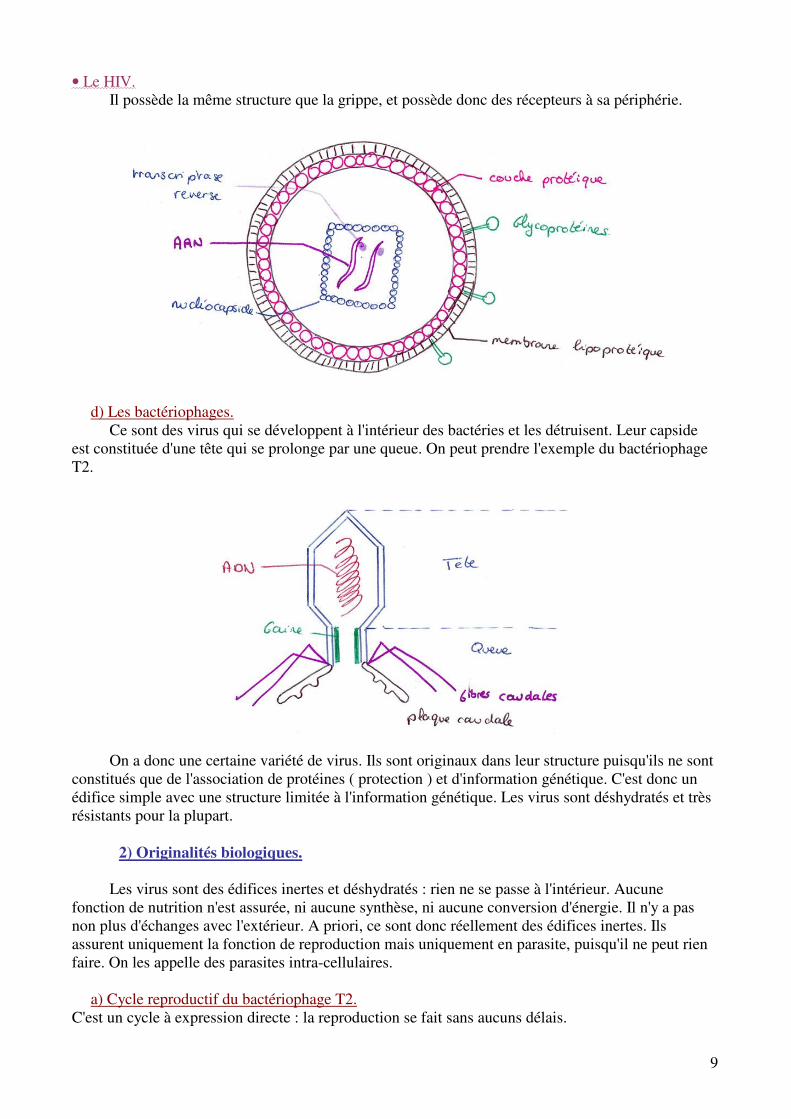
• La grippe.
On peut distinguer 8 fragments différents d'ADN. On a donc ici un virus à enveloppe et à
ARN. Les molécules de surface ont un rôle antigénique, ainsi qu'un rôle de reconnaissance et de fixation sur les cellules cibles. Le virus de la grippe se fixera préférentiellement sur les cellules des voies respiratoires. Ce virus est donc formé d'une enveloppe, d'une capside et d'ARN. Dans ce cas, la capside a un rôle non pas de protection mais un rôle de support pour l'ARN.
9
• Le HIV. Il possède la même structure que la grippe, et possède donc des récepteurs à sa périphérie.
d) Les bactériophages. Ce sont des virus qui se développent à l'intérieur des bactéries et les détruisent. Leur capside
est constituée d'une tête qui se prolonge par une queue. On peut prendre l'exemple du bactériophage T2.
On a donc une certaine variété de virus. Ils sont originaux dans leur structure puisqu'ils ne sont constitués que de l'association de protéines ( protection ) et d'information génétique. C'est donc un édifice simple avec une structure limitée à l'information génétique. Les virus sont déshydratés et très résistants pour la plupart.
2) Originalités biologiques.
Les virus sont des édifices inertes et déshydratés : rien ne se passe à l'intérieur. Aucune fonction de nutrition n'est assurée, ni aucune synthèse, ni aucune conversion d'énergie. Il n'y a pas non plus d'échanges avec l'extérieur. A priori, ce sont donc réellement des édifices inertes. Ils assurent uniquement la fonction de reproduction mais uniquement en parasite, puisqu'il ne peut rien faire. On les appelle des parasites intra-cellulaires.
a) Cycle reproductif du bactériophage T2. C'est un cycle à expression directe : la reproduction se fait sans aucuns délais.

10
• Phase d'infection. - Adsorption ( = fixation ).
Le virus se fixe sur la bactérie au niveau de la plaque caudale. Ce bactériophage ne peut se fixer que sur E Coli : il est donc doué de moyens de reconnaissance de la cellule cible, grâce aux protéines de la plaque caudale. - Hydrolyse de la paroi.
Elles se fait par une enzyme virale, le lysozyme, présente dans la plaque caudale. Elle hydrolyse la paroi de la bactérie, puisque cette dernière est principalement composée de muréine. - Contraction de la queue. La gaine se rétrécit et la gaine peut ainsi passer à travers l'orifice fait par l'action du lysozyme. - Injection de l'information génétique. L'ADN peut passer à l'intérieur de la bactéries ( chemin tout tracé ). • Phase d'éclipse.
Le virus semble avoir disparu : on ne voit plus que la capside vide ( = fantôme ), puisque l'ADN a déjà été injecté.
Le chromosome bactérien est détruit par des endonucléases fabriquées par le virus. Il peut alors y avoir synthèse de protéines virales : c'est l'ADN viral qui dirige tout le métabolisme de la cellule. Le virus fait donc fabriquer ses propres protéines, y compris les protéines de la capside et le lysozyme. On a également réplication de l'ADN viral. Finalement, tout est synthétisé pour faire de nouveaux virus ( ADN + protéines ). • Phase d'auto-assemblage.
Les capsomères s'assemblent entre elles pour former de nouvelles capsides. L'ADN rentre alors dans ces capsides et on a donc de nombreux virions à l'intérieur de la bactérie. Il s'est écoulé seulement 12 mn depuis l'injection de l'ADN. • Libération de nouveaux virions.
La paroi de la cellule est hydrolysée par le lysozyme. Le virus a finalement vraiment pris à son compte la bactérie, puisqu'il a utilisé ses ribosomes… Au total, le cycle aura duré 25 mn, et on appelle ce type de cycle un cycle lytique.
La multiplication des virus s'accélèrent alors si on a une population plus ou moins importante d'E Coli. entre 100 et 200 nouveaux virus sont créés à chaque fois.
11
Il peut y avoir mutation de gène, et puisque les virus sont haploïdes, l'expression de ces mutation est rapide. Il peut exister tout de même des variations pour ce cycle lytique.
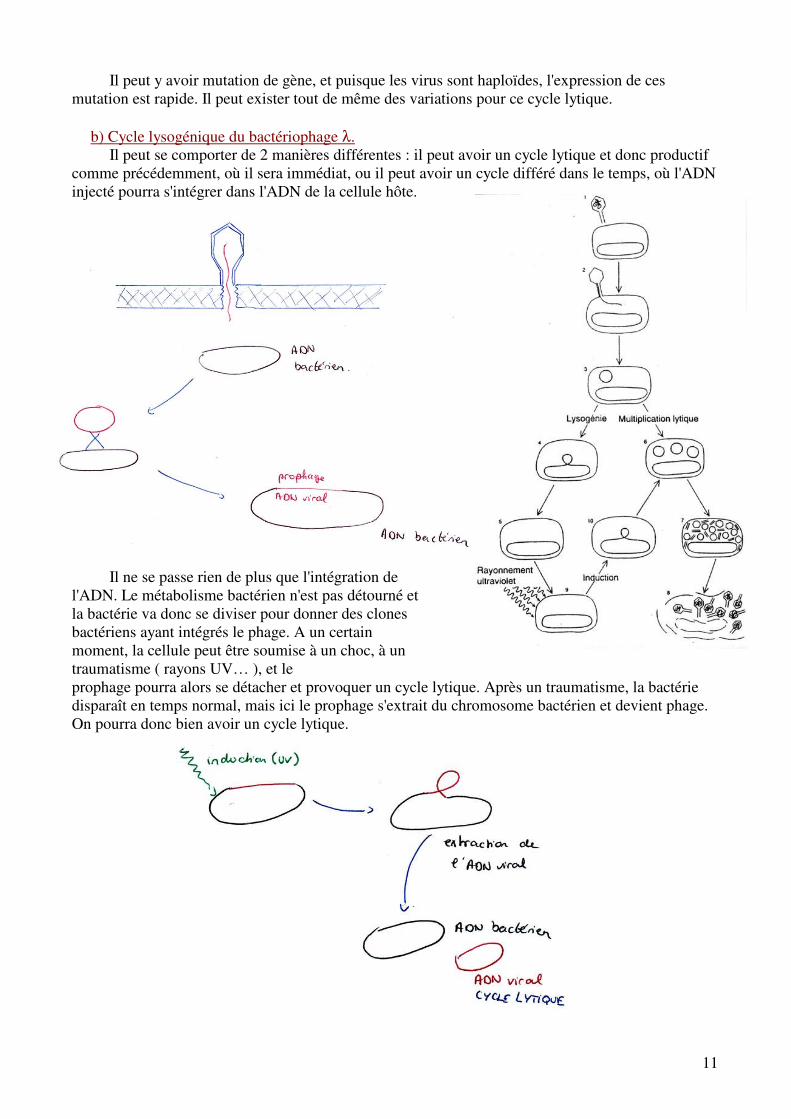
b) Cycle lysogénique du bactériophage λ.
Il peut se comporter de 2 manières différentes : il peut avoir un cycle lytique et donc productif comme précédemment, où il sera immédiat, ou il peut avoir un cycle différé dans le temps, où l'ADN injecté pourra s'intégrer dans l'ADN de la cellule hôte.
Il ne se passe rien de plus que l'intégration de l'ADN. Le métabolisme bactérien n'est pas détourné et la bactérie va donc se diviser pour donner des clones bactériens ayant intégrés le phage. A un certain moment, la cellule peut être soumise à un choc, à un traumatisme ( rayons UV… ), et le prophage pourra alors se détacher et provoquer un cycle lytique. Après un traumatisme, la bactérie disparaît en temps normal, mais ici le prophage s'extrait du chromosome bactérien et devient phage. On pourra donc bien avoir un cycle lytique.
12
Le cycle lytique peut se produire longtemps après, et l'ADN viral aura été dupliqué autant de fois que l'aura fait la bactérie infectée.
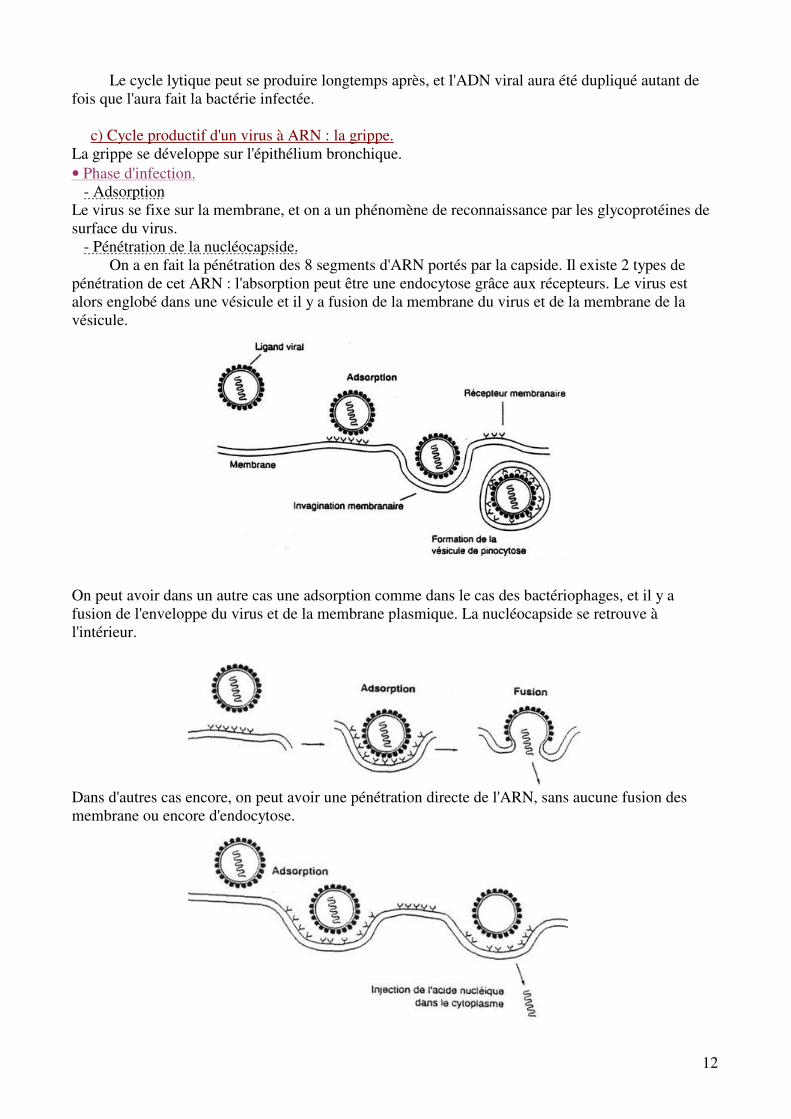
c) Cycle productif d'un virus à ARN : la grippe. La grippe se développe sur l'épithélium bronchique. • Phase d'infection. - Adsorption Le virus se fixe sur la membrane, et on a un phénomène de reconnaissance par les glycoprotéines de surface du virus. - Pénétration de la nucléocapside.
On a en fait la pénétration des 8 segments d'ARN portés par la capside. Il existe 2 types de pénétration de cet ARN : l'absorption peut être une endocytose grâce aux récepteurs. Le virus est alors englobé dans une vésicule et il y a fusion de la membrane du virus et de la membrane de la vésicule.
On peut avoir dans un autre cas une adsorption comme dans le cas des bactériophages, et il y a fusion de l'enveloppe du virus et de la membrane plasmique. La nucléocapside se retrouve à l'intérieur. Dans d'autres cas encore, on peut avoir une pénétration directe de l'ARN, sans aucune fusion des membrane ou encore d'endocytose.

13
• Phase d'éclipse. - Synthèse de protéines virales.
Cette synthèse pose le problème des ARNm. L'ARN du virion n'est pas messager donc il y a nécessité de synthétiser de l'ARNm. L'ARN va en fait servir de matrice à la synthèse des ARNm, transcrit grâce à la transcriptase portée par le virus. Dans le cas de la grippe, la transcriptase est l'ARN polymérase ARN dépendante. C'est une enzyme spécifique qu'on ne trouve pas dans la cellule. L'ARN du virion est appelé ARN- il est le complémentaire de l'ARNm. On pourra alors avoir le phénomène de traduction grâce aux ribosomes, aux acides aminés et à l'ATP de la cellule pour donner des protéines virales. Il n'y a pas destruction de l'ADN de la cellule donc cela signifie que le métabolisme de la cellule est détourné en partie.
Remarque : dans certains cas, l'ARN viral est déjà l'ARNm. On les appellera donc les ARN+.
De toutes façons, il sera nécessaire d'avoir un brin complémentaire pour pouvoir synthétiser d'autres brins d'ARNm.
Les protéines de la capside, la transcriptase spécifique au virus, les protéines spécifiques de l'enveloppe qui sont le plus souvent des glycoprotéines sont les principales protéines synthétisées.
La synthèse de la capside et de la transcriptase se fait dans le hyaloplasme grâce au ribosomes libres. Les protéines de l'enveloppe sont synthétisées sur le RER, puis sont intégrées à la membrane de ce même RER. Le flux membranaire permet d'amener ces protéines vers la membrane plasmique, qui ressemblera donc par endroit à la membrane du virus. - Synthèse de nouveaux ARN.
On a un ARN-. On va avoir la formation d'un ARN complémentaire grâce à la transcriptase. A partir de lui se formera alors un autre ARN complémentaire qui sera identique au premier. Cette action se déroule dans le noyau.
• Phase d'auto-assemblage. - Formation de la nucléocapside.
Elle consiste en fait à l'association capside + ARN-, et se fait dans le noyau. Les protéines synthétisées dans le hyaloplasme ont migré vers le noyau. Se rajoute ensuite la transcriptase.

14
- Incorporation de la nucléocapside à l'enveloppe. La nucléocapside ainsi formée migre vers la périphérie de la cellule et se place contre la
membrane qui est en fait une membrane d'enveloppe virale ( glycoprotéines virale ). • Libération de nouveaux virions.
L'incorporation et la libération sont en réalité liées : elles se font en même temps. La libération se fait par bourgeonnement et ne provoque donc pas de lyse. La nucléocapside s'échappe en s'entourant de la membrane de l'enveloppe. Cela peut expliquer la structure lipoprotéique de l'enveloppe puisqu'elle est intégrée à la membrane plasmique.
C'est un cycle assez rapide : il prend environ 2 à 3 heures mais tous les virions libérés sont très
proches d'autres cellules de l'épithélium. L'infection des répand très rapidement et tout le tissu épithélial est infecté.
Il existe un nouveau médicament, un aérosol de produits chimiques, qui empêcherait l'adsorption des virus. Il se fixerait sur les récepteurs de la membrane. Les antibiotiques sont inutiles mais sont souvent donnés pour prévenir d'autres infections par des bactéries. Le meilleur moyen de prévenir la grippe reste le vaccin mais c'est un virus mutant : il faut donc identifier les souches, mettre en culture pour avoir un vaccin. On doit donc attendre les 1er cas de la maladie.
Pourquoi existe-t-il des mutations ? La transcriptase n'est pas fiable et des changements dans les glycoprotéines de l'enveloppe entraînent l'incapacité pour les anticorps de les reconnaître.
La grippe est un virus mortel pour les personnes âgées et par les complications qu'elle provoque. L'épidémie de la grippe espagnole a causé plusieurs millions de morts en Europe, tout comme celle de la grippe du poulet.
d) Cycle d'un virus à interactions transformantes : le rétrovirus HIV. On l'appelle également virus intégratif, qui a une expression différée comme le bactériophage
λ. Il se développe dans les lymphocytes T4, qui sont des cellules du système immunitaire. Elles sont détruites et disparaissent après l'action du virus. La mort interviendra donc par défaut du système immunitaire : toute maladie bénigne sera alors mortelle.
• Pénétration dans la cellule T4.
C'est un virus à enveloppe donc on retrouve à peu près le même principe que pour la grippe. Le virus reconnaît la cellule cible grâce aux glycoprotéines de surface. • Formation d'un ADN à partir de l'ARN.
C'est un ARN qui est injecté, et qui devient matrice pour créer un ADN viral, grâce à une enzyme portée initialement par le virus. Ici, l'enzyme est une transcriptase reverse, l'ADN polymérase ARN dépendante. Ce phénomène explique le terme de rétrovirus, puisqu'on passe de l'ARN à l'ADN, et de transcriptase reverse. On obtient un ADN mono-caténaire, qui pourra devenir bi-caténaire par l'action d'une ADN polymérase ADN dépendante.

15
• Intégration de l'ADN viral dans l'ADN hôte.
Le virus semble n'exprimer qu'un seul gène au départ, et ce gène bloquerait le fonctionnement des autres gènes. La cellule porte donc le virus mais il ne s'exprime pas : on dit qu'on a un porteur sain. L'intégration de l'ADN viral est permise par une enzyme, l'intégrase. • Production de nouveaux virions.
On peut avoir 3 comportements différents : soit il ne se passe rient et cette situation peut durer des mois voire des années, soit il y a la formation de quelques virions, ce qui n'affecte pas vraiment la cellule, soit encore il peut y avoir formation de nombreux nouveaux virions, ce qui provoque alors la mort de la cellule.
Quand le T4 est sollicité dans une réponse immunitaire, il se met à produire une quantité importante de virions ( puisqu'il se doit de produire beaucoup d'anticorps ). De plus, le T4 se reproduit en temps normal, mais ici, il va libérer des virions en mourant, et ne pourra pas se multiplier : il y aura de moins en moins de T4.
Heureusement, c'est un virus qui ne se transmet pas par voies respiratoires, mais uniquement par contact ( sang, sécrétions génitales… ). De plus, c'est un virus qui a une durée de vie assez courte, ce qui peut être une chance.
L'AZT ( ou azothymine ) est un traitement chimique qui prend la place de la thymine lors de la réplication, ce qui a pour conséquence d'arrêter la synthèse d'ADN. Cela permet de ralentir la progression de la maladie, mais aussi le métabolisme. L'existence d'un vaccin est rendue presque impossible à cause de mutations trop nombreuses. La trithérapie est l'association de 3 molécules, et c'est pour cela que c'est le traitement qui marche le mieux. Conclusion.
Cela montre que la biologie n'est pas une science figée, et la médecine triomphante des années 60 s'est effondrée avec la découverte du HIV. Il apparaît de nouvelles maladies virales…
De plus, la maladie de la vache folle est encore inconnue. On ne sait pas ce qu'est le prion, et on ne le connaît que par la protéine. Mais si c'est la protéine qui est responsable de la maladie, alors elle aurait un pouvoir d'auto-reproduction ( arriverait à remonter à l'ADN ). C'est un phénomène encore totalement inconnu.
Qu'est-ce qu'un virus et d'où viennent-ils ? Est-ce que ce sont des organismes primitifs ? Non, puisqu'ils ont besoin d'autres cellules pour se reproduire. Apparemment, ils proviendraient de petits bouts d'ADN échappés d'une cellule. En effet, dans le cas des virus oncogènes, responsables des cancers, possèdent des gènes que l'on trouve naturellement dans nos cellules, mais on les trouve à l'état muté. Sujets de colle. - Les virus ARN. - Originalité des virus. - Cycles de reproduction des virus.

16
I) Caractères distinctifs eucaryotes / procaryotes.
1) Organisation des procaryotes.
a) Structure d'une bactérie.
b) Structure d'une cyanobactérie
2) Différences entre procaryote et eucaryote.
a) La taille.
b) Les limites cellulaires.
c) Particularités cytoplasmiques.
d) Pas d'organites convertisseurs
e) Le matériel génétique.
f) Efficacité dans les phénomènes de prolifération.
II) Les virus.
1) Organisation d'un virus.
a) Virions hélicoïdaux : le VMT.
b) Les virus icosaédriques : l'adénovirus.
c) Les virus à enveloppe.
d) Les bactériophages.
2) Originalités biologiques.
a) Cycle reproductif du bactériophage T2.
b) Cycle lysogénique du bactériophage λ.
c) Cycle productif d'un virus à ARN : la grippe.
d) Cycle d'un virus à interactions transformantes : le rétrovirus HIV.


















