Physiologie animaleaurelien.chateigner.free.fr/- Physiologie -/Physiologie... · Web viewEx : CO2...
46
1 Physiologie Générale Pr. Tobias Hevor Physiologie animale CH1 : Les cellules nerveuses I. Découverte de la neurobiologie La physiologie = étude du fonctionnement du tissu, des organes, des cellules entières et des organismes entier. 1865 Jan PURKINJE (tchèque) = découverte des cellules de PURKINJE 1865 Otto DIETERS (Bonn) = découverte des dendrites et des axones A. Camillo GOLGI = utilise la technique de golgi (utilisation de nitrate d’argent) pour observer les cellules nerveuses 1888 Santiago RAMON = identifie les neurones 1887 Wilhem HIS 1888 August FOREL 1891 Wilhem WALDEYER Un neurone = une cellule capable d’émettre un message, de propager un message, de communiquer ce message a une autre cellule. Les neurones ont aussi la capacité de sécréter des substances et ont donc une fonction sécrétrice. Plusieurs neurones forment un circuit neuronique. II. Cytologie du neurone A. Aspect général La membrane des neurones est en mosaïque de type SINGER NICHOLSON (lipide + protéine+ protéine particulière). A l’extérieur de cette membrane on retrouve : - des fibronectines - des protéines d’adhésion (Cell Adhesion Molecule = CAM => neural-CAM = N-CAM) A l’intérieur : - de l’actine (tension du neurone) - du NICHOLSON (tension du neurone) - de la spectrine (formation de la forme du neurone) - de la fodrine (formation de la forme du neurone) Le noyau des neurones est « spécial » en effet :
Transcript of Physiologie animaleaurelien.chateigner.free.fr/- Physiologie -/Physiologie... · Web viewEx : CO2...

1
Physiologie Générale Pr. Tobias Hevor
Physiologie animale
CH1 : Les cellules nerveuses
I. Découverte de la neurobiologieLa physiologie = étude du fonctionnement du tissu, des organes, des cellules entières et des organismes entier.
1865 Jan PURKINJE (tchèque) = découverte des cellules de PURKINJE1865 Otto DIETERS (Bonn) = découverte des dendrites et des axonesA. Camillo GOLGI = utilise la technique de golgi (utilisation de nitrate d’argent) pour observer
les cellules nerveuses1888 Santiago RAMON = identifie les neurones1887 Wilhem HIS1888 August FOREL1891 Wilhem WALDEYER
Un neurone = une cellule capable d’émettre un message, de propager un message, de communiquer ce message a une autre cellule. Les neurones ont aussi la capacité de sécréter des substances et ont donc une fonction sécrétrice.Plusieurs neurones forment un circuit neuronique.
II. Cytologie du neurone
A. Aspect généralLa membrane des neurones est en mosaïque de type SINGER NICHOLSON (lipide + protéine+ protéine particulière).A l’extérieur de cette membrane on retrouve : - des fibronectines - des protéines d’adhésion (Cell Adhesion Molecule = CAM => neural-CAM = N-CAM)
A l’intérieur : - de l’actine (tension du neurone) - du NICHOLSON (tension du neurone) - de la spectrine (formation de la forme du neurone) - de la fodrine (formation de la forme du neurone)
Le noyau des neurones est « spécial » en effet :- il n’y a plus de mitose dans les neurones = noyau a phase de repos permanent - la chromatine est uniforme- le nucléole a une taille importante (= fabrication de protéine importante)- présence d’organes circumventriculaire- les pores nucléaires sont en grand nombre- présence de petites molécules = stéroïdes

2
Physiologie Générale Pr. Tobias HevorLe réticulum endoplasmique (RE) est très actif, est s’appelle le corps de NISSL car très développé. De plus les lysosomes sont très fonctionnels, les mitochondries sont très actives et le cytosquelette de la cellule est très développé
Les protéines synthétisées peuvent être classées selon 3 familles :- les protéines qui reste dans le cytoplasme = gel cytoplasmique- les protéines qui sont incorporées dans les différents organites (noyau, mitochondrie,…)- les protéines véhiculées le long des prolongements (dendritiques, récepteur) = servent à
renouveler la membrane des axones.
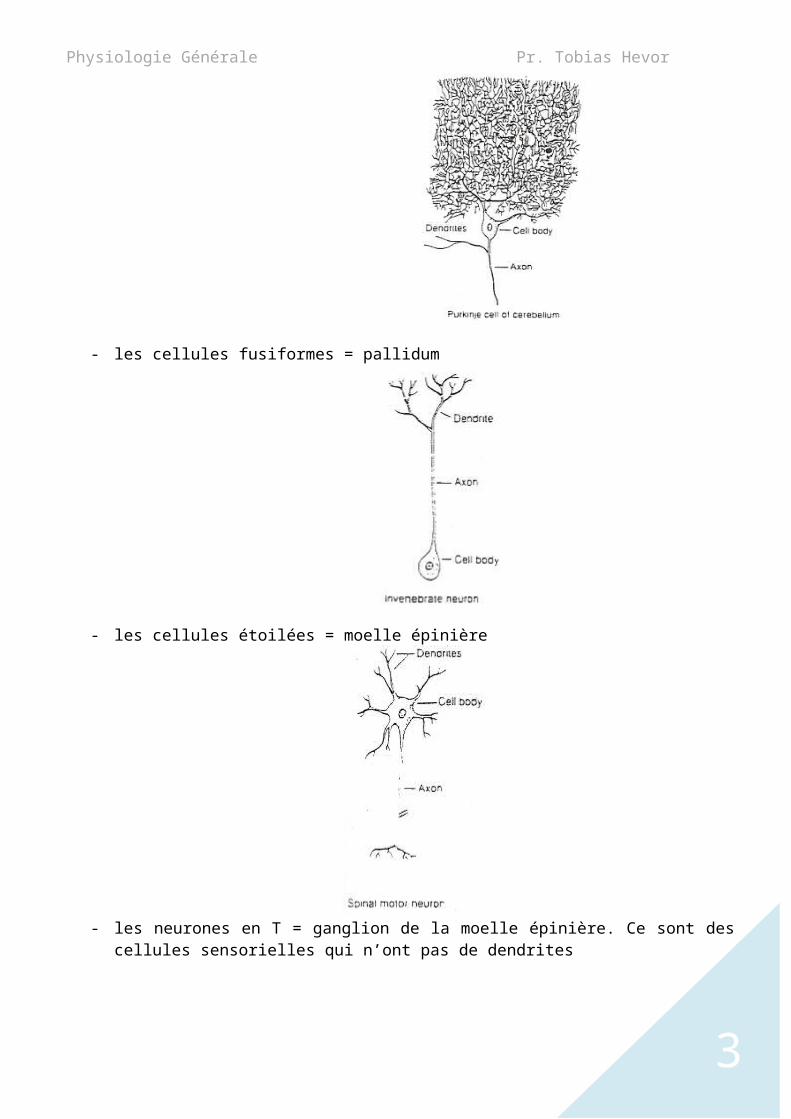
B. Le corps cellulaire et les dendritesCorps cellulaire = SOMA = péricaryon.Le SOMA est associé d’un coté aux dendrites (pole récepteur du neurone ou les informations arrivent) et de l’autre coté a un axone (pole émetteur du neurone qui transmet les informations).Le corps cellulaires est polymorphe c'est-à-dire qu’il a des formes variables :- les cellules pyramidales = hippocampe
- les cellules ovoïdes (en œuf) = cellules de PURKINJE
- les cellules fusiformes = pallidum
3
Physiologie Générale Pr. Tobias Hevor
- les cellules étoilées = moelle épinière
- les neurones en T = ganglion de la moelle épinière. Ce sont des cellules sensorielles qui n’ont
pas de dendrites
- les neurones bipolaires = rétine.

4
Physiologie Générale Pr. Tobias Hevor
C’est le SOMA qui assure l’intégrité de la cellule du neurone et par conséquent qui fabrique les macro-protéines nécessaire à la régénération cellulaire.Les dendrites sont des extensions du cytoplasme, il y en a environ 9 lorsque l’on quitte le corps cellulaire mais elles sont recouvertes d’épines dendritiques (environ 10000 épines par dendrite), cependant certains neurones sont lisse et n’ont pas d’épines dendritiques. Les dendrites ont un diamètre qui diminue au fur et à mesure que l’on s’éloigne du corps cellulaire.
C. L’axone et les collatéralL’axone est toujours lisse et capable d’avoir des ponts collatéraux, ce pont collatéral peut revenir sur son propre SOMA= pont collatéral récurent.
L’axone est recouvert par une gaine isolante = gaine de myéline.Les neurones de type Golgi I = neurone cortico-thalamique (neurones assez long).Les neurones de type Golgi II = neurone permettant la création de circuit locaux (neurone relativement court).
L’axone se ramifie à son extrémité, chaque extrémité va se renfler en bouton (bouton terminal). Un bouton + un récepteur forme une jonction plus communément appelé synapse.Le pole collatéral est considéré comme le pole d’émission. Au niveau du segment initial il y a la création d’un potentiel d’action qui se propage le long de l’axone et permet la libération de substance (neurotransmetteur) qui permet la transmission du message. Ce message peut être modulé au niveau du segment initial.
III. Synthèse et trafic intra neuroniques
A. Maintient de la forme des neuronesLe cytosquelette est le support matériel :- microtubule (25 nm de diamètre)

5
Physiologie Générale Pr. Tobias Hevor- les Microtubule Associated Proteine ou MAPs = charpente de l’axone- l’actine globulaire (actine G)
Il faut deux filaments d’actine F pour faire un filament d’actine- des filaments intermédiaires (7 à 11 nm), ce sont des neurofilaments constitués de 3 protéines
agglomérées.Les microtubules/filaments sont des éléments labiles (qui ne sont pas fixe) qui se polymérisent plus du coté axonique que du coté dendritique. Le transport de ces microtubules est rétrograde et antérograde.
B. Synthèse de macromoléculesLes mitochondries sont allongées. Dans les dendrites se trouvent le RE et les polysomes mais l’appareil de golgi ne se trouve pas dans les dendrites. Dans les dendrites se trouvent des MAPs spécifiques aux dendrites.Les ARN sont transportés du corps cellulaires jusqu’aux dendrites (500μm par jour) ou sont synthétisé des molécules de petites tailles.Dans l’axoplasme il n’y a ni RE ni appareil de golgi, il n’y a donc pas de possibilité de synthétiser des protéines dans l’axone. Les protéines de l’axoplasme sont donc synthétisées dans le SOMA.
IV. Transport axonique
A. Mise en évidencePour mettre en évidence ce transport axonique on utilise des axones géants de cellule de calmar qui font environ 1 mm de diamètre.Expérience : on pose une ligature sur l’axone de calmar et on observe un renflement juste avant la ligature.On peut donc en conclure qu’à l’intérieur de l’axone du matériel a été bloqué par la ligature.
Néanmoins si on met une autre ligature on observe un autre renflement de l’autre coté de la ligature. Il y a donc un transport de matière dans les deux sens :- transport antérograde/extracellulaire = noyau queue

6
Physiologie Générale Pr. Tobias Hevor- transport rétrograde = queue noyau
Si on enlève la membrane plasmique de l’axone de calmar pour introduire des composés fluorescents et effectuer une microcinématographie, on observe des milliards de vésicules de transport circulant le long de l’axone à des vitesses différentes.
B. Transport antérograde rapideIl est effectué par des vésicules de 50nm de diamètre qui ont une vitesse d’environ 100 à 400nm par jour. Ces vésicules sont issues du corps de Golgi jusqu'à l’extrémité terminale de l’axone. Dans ces vésicules on retrouve :- des matériaux permettant de renouveler la membrane cellulaire- des enzymes de fabrication- des neurotransmetteurs- des précurseurs de neurotransmetteurs
On peut bloquer le transport axonique avec de la colchicine et de la vinblastine. En effet ce transport utilise les microtubules comme des rails pour avancer. La kinésine constitue le câble de liaison entre la vésicule et le microtubule activité ATPasique.
C. Transport antérograde lentLa vitesse est beaucoup plus lente : environ 1à 2 nm par jour. Ce transport assure le déplacement des éléments de grande taille comme les éléments du cytosquelette (microtubule, microfilament, neurofilament, protéines cytosolubles utilisées par le métabolisme secondaire).
D. Transport de mitochondriePour le transport de mitochondrie la vitesse est relativement lente car le transport s’effectue de façon saccadée. Cependant au final les mitochondries se retrouvent dans les extrémités des terminaisons axoniques. Une fois que la mitochondrie a fini sa vie, elle revient dans le sens opposé pour être dégradé dans le corps cellulaire.
E. Transport rétrogradeLe transport rétrograde chemine les vésicules de grandes tailles (ensemble de petites vésicules). Le mécanisme de transport est quasi similaire au transport antérograde mais il est effectué par des MAP-

7
Physiologie Générale Pr. Tobias Hevor1C/dynéine qui ont une activité ATPasique également. Ce transport véhicule les molécules vieillies (protéine membranaire) pour le recyclage lysosomal.Grâce à la peroxydase de RAIFORT (= peroxyde qui sont des composés coloré) on peut faire une carte de la sensibilité générale, en effet si on place ces peroxyde a l’extrémité axonique ils vont circuler le long de la voie nerveuse et tant qu’ils sont colorés on peut suivre où ils passent.Nerve Growth Factor (NGF) subit le transport rétrograde et permet la différenciation des neurones. Chez l’adulte cette protéine permet de maintenir la forme des neurones.
V. Connections des neurones en circuits myotatiquesLes réflexes dus aux stimulations mécaniques ou chimiques comme les genoux peuvent se restreindre à un réseau de 2 éléments et s’organisent en deux niveaux :- En effet un neurone en T est un neurone sensoriel à deux ramifications dont l’une part vers un
neurone étoilé qui permet la stimulation du muscle agoniste et permet le réflexe.- Et l’autre ramification part vers un neurone de type Golgi I qui permet une inhibition du muscle
antagoniste ou opposé au muscle agoniste (stimulé).

8
Physiologie Générale Pr. Tobias Hevor
CH2 : Les cellules gliales
I. GénéralitésLes neurones représentent environ un tiers des cellules du cerveau. C’est VIRCHOW qui met en évidence la présence des cellules gliales dans le cerveau et c’est plus tard que Camillo GOLGI mettra en évidence leur rôle.Ces cellules ont eu des noms variés au cours du temps en effet au début on les considérait comme de la colle des neurones (Nerve glue = névroglie = glue), en effet ces cellules comblent les espaces entre les neurones.Elles sont très diversifiées : - La glie interstitielle = Oligodendrocytes, Astrocytes, Microgliocytes se situent entre les
neurones.- La glie épithéliale = Ependymocytes se situent a la surface des cavités encéphales.- La cellule de SCHWANN.
Ces cellules gliales comme les neurones ont une origine ectodermique sauf les microgliocytes qui sont d’origine mésodermique. Ces cellules ne sont pas des cellules excitables elles se divisent encore après la naissance et sont les principales responsable de tumeur du cerveau = gliome.
II. Les Astrocytes
A. MorphologieSe sont des petites cellules de 10μm de diamètre avec des petits prolongements fin. Les prolongements font environ 50µm de longueur et se terminent en se plaçant à coté d’un capillaire = pied terminal. A l’intérieur des astrocytes se trouvent des protéines spécifiques = Glial Fibrillary Acidic Protein (GFAP) qui sont des protéines qui se mettent sous forme de glio-filament, ce qui permet de reconnaître deux catégories d’astrocytes :- les astrocytes avec beaucoup de cytoplasme = protoplasmique- les astrocytes fibreux
Une cellule gliale et sa voisine peuvent établir des jonctions lâches = gap jonction qui permet le passage d’une cellule à l’autre.
B. Astrocytes de type 1, protoplasmiqueCes cellules élaborent les pieds terminaux qui recouvrent les vaisseaux sanguins et vont créer des manchons qui vont s’associer avec une partie de méninge pour créer la glia limitans externa
C. Astrocytes de type 2Ces astrocytes vont au contact des dendrites et des nœuds de RANVIER et aident à la fabrication de la gaine de myéline. Les astrocytes de type2 et les oligodendrocytes viennent d’un progéniteur O-2A commun.
D. Régulation et composition du volume du liquide extracellulaireLes astrocytes ont 3 grands rôles :- Ils servent de filtre entre les neurones et les vaisseaux sanguins/le milieu extracellulaire.- Ils règlent le taux de Potassium dans le milieu extracellulaire (le potassium en grande quantité
dépolarise les neurones et entraîne leur mort) présence de Na-K-ATPase.

9
Physiologie Générale Pr. Tobias Hevor- Ils coopèrent pour la neurotransmission. Le glutamate sert de neurotransmetteur et sera libéré
par le neurone puis capté par les Astrocytes qui le transformeront en glutamine grâce à une enzyme (la glutamine synthétase), plus tard les Astrocytes cèdent la glutamine au neurone qui la transforme en glutamate. Il y a une coopération car l’Astrocyte fait une part du travail et le neurone l’autre part.
III. Les Oligodendrocytes
A. Les Oligodendrocytes inter fasciculaireIls font de 6a 8µm de diamètre et possèdent des petits prolongements qui s’élargissent énormément par rapport au corps cellulaire. Ces Oligodendrocytes s’enroulent autour d’un axone de passage et constituent la gaine de myéline (faite de lipides essentiellement : galactoceramide-C en particulier).
Pour une expérience on introduit de la galactoceramide-C à un lapin qui considère cette protéine comme un antigène. Le lapin secrète des anticorps anti-galactoceramide-C. On récupère le sérum de lapin et on le met sur une coupe de cerveau. Ce marquage immunocytologique permet de reconnaître la galactoceramide-C grâce a l’anti-galactoceramide-C = anti-gal-C
B. Formation de la gaine de myélineCertains axones sont recouverts par une gaine lipidique. Ce sont des manchons crées par les oligodendrocytes qui peuvent atteindre 1mm et pour s’enrouler autour de l’axone = nœud de Ranvier. Un oligodendrocyte peut enrouler quelques dizaines ou centaines de neurone.La myélination dure de la naissance a environ 2ans d’où l’augmentation du volume de l’encéphale chez l’enfant.
La gaine de myéline est composée de lipide dont le cholestérol, des phospholipides et des glycolipides. La myélination permet de conduire les informations plus rapidement.
Chez certain individus le corps cellulaire dégénère du a la dégradation de la gaine de myéline sur toute une région du cerveau : le flux nerveux ralentit = décalage arrivé du flux nerveux ce qui entraîne des problèmes moteurs sclérose en plaque.
IV. Les MicrogliocytesSe sont les cellules gliales qui sont représentées en le plus petit nombre (environ 5 à 10% des cellules gliales. Ces cellules sont très polymorphes et sont positionnées à coté des corps cellulaires des neurones, des vaisseaux sanguins, de la glia limitans externa, des cellules voisines des macrophages. Leur rôle est de phagocyter les débris du système nerveux (neurone mort, cellule morte et les substances inhabituelles qui s’introduisent dans le système nerveux.
V. Les Ependymocytes
A. Localisation Dans l’encéphale on observe des cavités qui contiennent le liquide céphalorachidien : les cavités des ventricules de l’épithélium, ce liquide est réalisé par des cellules épendymères de deux classes différentes :- les cellules épendymères de plexus choroïde = fabrication du liquide céphalorachidien- les cellules épendymères extra choroïdiennes

10
Physiologie Générale Pr. Tobias HevorCes cellules sont polarisées (coté apical, coté basal). Ces cellules sont unies latéralement par des jonctions étanches (pour que le liquide céphalorachidien ne passe pas)
B. Les Ependymocytes choroïdiensLe plexus choroïde (plexus = enchevêtrement) = plexus de capillaire situé en dessous des Ependymocytes qui sont posés sur la lame épendymale = permet l’élaboration du LCR
Les cavités sont constituées par les épendymocytes extra-choroïdiens. Les épendymocytes choroïdes se situent dans les ventricules latéraux.La fonction des choroïdiens est de fabriquer LCR et d’assurer l’étanchéité (barrière entre le sang et le LCR). Le LCR doit rester limpide sinon il entraîne des pathologies
Les tanycytes sont des cellules à morphologie spéciale qui assure une communication entre le LCR et les tissus/vaisseaux sanguins.
VI. Les cellules de SCHWANNLes cellules de SCHWANN peuvent être :- céphalorachidiennes = élaborent la gaine de myéline- Non myélinisantes- Satellites
Dans les axones périphériques on retrouve les cellules de SCHWANN myélinisantes qui élaborent la gaine de myéline des axones. Le cytoplasme de la cellule de SCHWANN entoure l’axone une cellule = un manchon alors que les oligodendrocytes peuvent créer plusieurs manchon avec une seule cellule.
Certains axones n’ont pas de gaine de myéline et donc sont protégés par les cellules de SCHWANN non myélinisantes qui ont un cytoplasme qui se prolonge légèrement puis forment des petits tunnels dans lesquels vont passer des axones (une vingtaine d’axones peuvent passer dans le cytoplasme d’une même cellule de SCHWANN).
Les cellules de SCHWANN satellites (teloglie) sont utilisées car à la l’extrémité de l’axone la gaine de myéline s’arrête. Cependant l’axone a besoin de protection et est recouvert par des cellules satellites.

11
Physiologie Générale Pr. Tobias Hevor
CH3 : Le tissu nerveux
I. Organisation générale
A. Le système nerveux centrale et périphériqueLe système nerveux est divisé en deux éléments :- le système nerveux central (SNC) L’encéphale = télencéphale, diencéphale, tronc cérébrale et le bulbe rachidien Moelle épinière - le système nerveux périphérique (SNP) Le système orthosympathique Le système parasympathique
Le SNP innerve les parties végétatives/autonomes du corps = cœur, poumon, intestin… SN autonome.
Le système orthosympathique = série de ganglion nerveux qui longe la moelle épinière (para vertébrale). A partir de ce S. orthosympathique partent des fibres nerveuses qui innervent les organes de la vie végétative. Ce système nerveux utilise comme dernier neurotransmetteur la noradrénaline.
Le système parasympathique = constitué par un nerf qui part du bulbe rachidien et qui s’oriente aussi vers les organes de la vie végétative plus les organes excréteurs = nerf pneumogastrique. Ce système nerveux utilise comme dernier transmetteur l’acétylcholine.
Le système nerveux crânien est protégé par l’os crânien, mais il est aussi protégé de l’os par des enveloppes/méninges qui sont structuré en 3 parties :- la parti la plus externe, accolée a l’os crânien = la dure-mère- en dessous un espace remplit de fibre organisé en toile d’araignée plus du LCR = arachnoïde - une parti solidaire au tissu nerveux = la pie-mère qui se mélange a la glia limitans externa. C’est
une protection/enveloppe pas très étanche.
De même pour les vertèbres il existe un espace libre entre l’os et les méninges = espace épidural ou l’on retrouve des vaisseaux sanguins, lymphatiques, tissus adipeux qui constitue un tissu spongieux qui va protéger tout la moelle épinière et permettre les mouvements de la colonne vertébrale sans dégrader la moelle épinière.
Dans le cerveau le sang est séparé des cellules par une barrière = la barrière hémato-encéphalique. Cependant le LCR est élaboré par le plexus choroïde à partir du plasma sanguin auquel il retire les protéines et change la concentration ionique. La pression hydrostatique dû à sa formation pousse le LCR et entraîne une circulation. Si la pression du LCR était trop importante il y aurait un risque « d’explosion » ce qui est impossible grâce à un système de villosité qui absorbe le surplus de LCR pour le reverser dans le sang.Grâce a la circulation du LCR on peut anesthésier certaine partit du corps mais a l’origine le LCR sert à amortir les chocs pour ne pas endommager le cerveau.
B. Constitution du tissu nerveuxSi on coupe le tissu nerveux on observe différente couleur :- de la matière grise = corps cellulaire des neurones avec départ des axone et dendrite plus les
cellules gliale

12
Physiologie Générale Pr. Tobias Hevor- de la matière blanche = ce sont les axones de passage (la couleur est du a la myéline)
La matière grise se situe essentiellement dans le cortex = partie périphérique. Néanmoins on peut retrouver de la matière grise dans de la matière blanche dans ce cas on appelle la matière grise = noyau.Le thalamus = matière grise dans la matière blanche = noyau thalamique.Dans la moelle épinière on a la matière blanche à l’extérieur et la matière grise a l’intérieur (= corne antérieur de la moelle épinière)
Une série de neurone = voie nerveuse.
Les neurones peuvent longer la moelle mais peut aussi croiser la ligne médiane
S’il y a une stimulation et que le neurone a une commissure on dit que la réponse est contralaterale.S’il n’y a pas de commissure la réponse est homolatérale/ipsilaterale.
C. Le système nerveux périphériqueLe nerf rachidien = une paire de nerfs rachidiens qui part de la moelle épinière = nerf moteur.Dans la moelle épinière se trouve des ganglions qui sont constitués :- de neurones en T

13
Physiologie Générale Pr. Tobias Hevor- de SOMA de neurone- de cellules de SCHWANN satellite- du tissu endoganglionaire avec de la matière extracellulaire développé par des fibroblastes qui
élabore une gaine autour et donne une impression de boule.Les nerfs sont des ensembles d’axones inclus dans un environnement. Chaque axone aura sa propre protection par des cellules de SCHWANN myélinisantes ou non. L’ensemble est entouré de fibres = basales. L’ensemble s’appelle une fibre nerveuse. Elle est accompagnée par un capillaire sanguin et du tissus conjonctif = endonerve. De plus la matière enveloppe plusieurs fibres nerveuses = faisceaux. L’épinière enveloppe plusieurs faisceaux avec le périnère.L’épinière est constitué de fibroblastes et de vaisseaux sanguins, de vaisseaux lymphatiques et de tissu adipeux
II. La barrière hémato-encéphaliqueLes vaisseaux sanguins sont perméables à l’eau, en effet les cellules des vaisseaux sanguins sont plates et laissent un canal à l’intérieur.
Il y a donc des possibilités d’échanges avec l’extérieur, cependant ces échanges sont très limités dans le cerveau grâce a la présence d’une barrière entre le sang et les neurones = la barrière hémato-encéphalique.
A. Les jonctions serrées de l’endothéliumDans le cerveau les jonctions des capillaires sont de type serré = zonula occludens ce qui rend le système perméable.Cependant dans l’encéphale il y a des zones sans barrière :- l’hypophyse - l’hypothalamus- l’épiphyse- l’area postrema = centre du vomissement
Cette barrière a pour rôle d’empêcher de passer des substances comme :- des enzymes dont les enzymes de dégradation- les monoamines-oxydases => dégrade des monoamines mais sont très réactive avec le système
nerveux peut stimuler le système nerveux
Cette barrière peut cependant être traversée.
B. Le passage de la barrière Cette barrière peut être traversée par plusieurs molécules ou mécanismes :

14
Physiologie Générale Pr. Tobias Hevor- les molécules liposolubles qui peuvent traverser car les membranes des capillaires sont des
membranes lipidiquesEx : l’héroïne et les substances dopantes car liposolubles- Les gaz se diffusent rapidement à travers la membrane
Ex : CO2 O2
C. Le passage inverse de la barrièreL’encéphale crée des déchets, il y a une nécessité de se débarrasser de ces déchets. Ces déchets franchissent la barrière car ils sont soit :- liposolubles- si hydrosolubles présences de transporteurs - besoin de se débarrasser des surplus de K+ présences de nombreux canaux ioniques

15
Physiologie Générale Pr. Tobias Hevor
CH4 : Les canaux ioniques et les protéines membranaires plasmiques
I. Rappel de l’architecture de la membrane plasmiqueLa membrane est une bicouche phospho-lipidique. La structure des lipides = - un acide gras - du glycérol
On observe plusieurs types de lipides :- Les monoglycérol = un glycérol + un acide gras- Les diglycéride = un glicérol + deux acide gras
La membrane est aussi constituée par des protéines : lipoprotéines ou glycoprotéines, qui peuvent etre :- extrinsèques- intrinsèques
Ceci est basé sur le systeme de Singer et Nicholson système en mosaïque.
II. Les protéines canauxLa membrane est imperméable à tous les ions. Cependant à l’interieur de la cellule on observe la présence d’ions. C’est donc qu’il y a une possibilité d’échange d’ions au travers de la membrane, soit des points de passage à l’interieur de la membrane = pore ou perméant.Ces canaux sont sélectifs : un canal laisse passer certain ions. Ex : canal potassique, canaux sodiques, canaux calciques.Les molécules d’eau se collent aux ions : quand l’ion passe, de l’eau passe aussi. C’est ce que l’on appelle de l’eau de salvation.Na 8 H2OK 6 H2O
Le passage d’ions est facile car les protéines sont des ensembles d’acides aminés.

16
Physiologie Générale Pr. Tobias HevorA. Les protéines directement perméables aux ions
Les canaux ioniques peuvent être constitués d’une ou plusieurs protéines identiques ou différentes. Les canaux protéiques voltage dépendants = fonctionnent en fonction de la valeur de voltage
environnant. Ce sont des canaux sélectifs : Ex canaux sodiques voltage dépendant :4 domaines composé de 6 segments homologues (un domaine = 300 acides aminés pour le canal sodique voltage dépendant).
Les canaux chimio-dépendants = fonctionnent si une molécule se fixe sur un site de fixation. Ces molécules peuvent être variées : soit des hormones soit des neurotransmetteurs.Ex : l’acétylcholine se fixe sur la protéine canal canal nicotinique.Si les 5 sous unités sont différentes on parle de pentamère.Par exemple le canal GABAa qui a pour neurotransmetteur la glycine.
B. Les protéines GCette protéine n’est pas un canal, cependant cette protéine est capable de recevoir des protéines étrangères à la cellule. Ces protéines membranaires sont en liaison avec d’autres protéines globulaires sous la membrane dans le cytoplasme. Lors d’une fixation (le premier message), la protéine s’active en changeant de conformation et active la protéine cellulaire qui va créer des réactions en effectuant des transformations à l’intérieur de la cellule, par exemple en synthétisant de l’AMPc (Adénosine Mono Phosphate cytoplasmique). Tout ces changement peuvent provoquer l’ouverture d’un canal et donc permettre le passage d’ions.Les récepteurs muscaridiques (qui se trouvent dans le cœur) sont constitués de segments hydrophobes, 7 segments transmembranaires.
C. Les autres canauxNéanmoins dans la membrane il y a d’autres canaux :- qui dépendent de la concentration
Ex : Les canaux potassiques, chlorures, calciques- les canaux jonctionnel = 2 membranes se rapprochent et créent une conexine. Chaque
membrane crée une connexion qui se rapproche et forme un tunnel. Ce système permet de passer rapidement d’une cellule à l’autre.
III. Pompes ioniques et transporteur d’ionsIl y a une activité enzymatique capable d’hydrolyser l’APT. ATPase = qui a besoin d’énergie pour fonctionner.Par exemple là, on récupère le potassium pour le mettre dans la cellule et on éjecte le sodium dans le milieu extracellulaire. Il y a aussi la présence de Ca-ATPase qui expulse le calcium de la cellule.
IV. Répartition et rôles des protéines membranairesLes canaux sodiques voltage dépendants sont regroupés au niveau du segment initial dans le neurone, au niveau des nœuds de Ranvier pour l’axone myélinisé et pour les axones non myélinisé sur tout le long de l’axone.Les canaux potassiques sont repartit sur les dendrites et le SOMA.Les canaux calciques voltage dépendants se trouvent sur les extrémités post-synaptiques.

17
Physiologie Générale Pr. Tobias Hevor
CH5 : Mécanisme de la perméabilité MembranaireLes cellules sont des êtres élémentaires, par conséquent elles ont besoin de nutriments. Or ces éléments nutritifs se trouvent à l’extérieur de la cellule et doivent passer dans la cellule pour être assimilés par celle-ci nécessité de franchir la membrane.Il y a deux grandes modalités pour franchir la membrane :- sans utiliser d’énergie cellulaire d’origine métabolique => transport passif- besoin de l’énergie de la cellule => transport actif.
I. Le transport passif
A. Diffusion simple
1. Diffusion de l’eauLes molécules sont en agitation permanente dû à l’agitation thermique. Pour stabiliser les molécules il faudrait atteindre la température de 0°K soit -273,15°C.Comme le montre une expérience (expérience de Fick), on observe un flux entrant et un flux sortant.
2. Equation de FickF = -D.S. (C 1 -C 2 ) X
D = coefficient de diffusionS = surface d’échangeC1-C2 = différence de concentrationX = épaisseur de la membraneL’épaisseur de la membrane plasmique varie selon la cellule.Cependant on peut modifier l’équation de Fick et on obtient :
F = -D.S. (C1-C2)
X
=> F= Pd.S. (C1-C2)Avec Pd = coefficient de perméabilité
3. Equilibre de Donan (Gibbs-Donan) Le système tend à rompre l’éléctroneutralité, on observe un gradient de charge électrique gradient électrique entre les deux compartiments.On observe la présence d’un équilibre mais pas d’une égalité. Il y a un déséquilibre de la concentration de chaque coté de la membrane => polarisation de la membrane.Ek = potentiel d’équilibre de l’ion potassium.[K]A.[Cl]A = [K]B.[Cl]B
Où[K] A = [Cl] B [K]B [Cl]A
Le gradient électrochimique établit la polarité des membranes plasmiques

18
Physiologie Générale Pr. Tobias HevorB. Perméabilité osmotique
π = R . T . (C1-C2)π. V = n.R.T
C. Perméabilité aux solutés neutres
1. Diffusion simpleCertains éléments traversent la membrane en suivant le gradient de concentration :- Les gaz se distribuent en fonction de leur concentration- Les substances liposolubles qui suivent le gradient => coefficient de partage des substances
liposolubles I. Expérience : [Voir schéma cours PG]
2. Diffusion facilitéePour franchir la membrane cellulaire, d’autres composés, qui ne sont pas liposolubles mais hydrosolubles, utilisent des protéines membranaires sur lesquelles vont se fixer les composés. La protéine se retrouve de l’autre coté de la cellule et libère le composé qui se retrouve dans la cellule. Ces protéines sont appelées des transporteurs. On peut saturer les protéines de transport si il y a une grande quantité de molécules à faire passer = saturation du systeme de transport.
Les protéines de transport sont :- saturables- spécifiques- peuvent être inhibées de façon compétitive.
Ex : La Phloridzine à une forme géométrique identique a celle du glucose. Il y donc une compétition avec le glucose. C’est un inhibiteur des transporteurs de glucose.- dépendent du pH => charge électrostatique change si le pH de la protéine change- pas besoin d’ATP pour fonctionner.
Le transport peut prendre deux molécules différentes en même temps on dit qu’il y a un co-transport Ex : L’alanine et le sodium, les deux molécules vont dans le même sens et vont dans la cellule = transport symport.
Le transport peut aussi prendre deux molécules mais dans les deux sens on dit qu’il y a co-transport en sens opposé = transport antiport
D. Perméabilité passive aux ionsUn ion existe uniquement en milieu aqueux = hydrosoluble. Les ions ne peuvent pas passer la membrane ils ont besoin d’un passage pour pouvoir pénétrer = canal ionique. Ces canaux permettent aux ions de passer la membrane. Cependant ils nécessitent de l’énergie.
II. Transport actifLe transport actif utilise l’énergie sous forme d’ATP. Le transport a les mêmes propriétés que le transport facilité :- saturable- spécifique- peut être inhibé par un inhibiteur chimique. Par exemple avec du cyanure qui bloque l’énergie
de la chaîne respiratoire et bloque le transport actif, ou le dinitrophénol qui bloque la fabrication d’ATP.

19
Physiologie Générale Pr. Tobias Hevor- pHdépendant- besoin d’énergie (ATP pour fonctionner)- influencé par la température : en effet si la température est faible le transport fonctionne
moins bien.
Il y a la présence de co-transport, par exemple le co-transport effectué par la Na-K-ATPase mais cet enzyme hydrolyse l’ATP pour fonctionner.Grâce à l’ouabaïne extraite de la digitale on peu bloquer la Na-K-ATPase ce qui entraîne une baisse du gradient de concentration puis la dépolarisation de la membrane et enfin la mort de la cellule.
Autre transport actif :- La Ca-ATPase = pour faire sortir le Ca de la cellule- L’ATPase à bicarbonate = pour faire sortir le bicarbonate - L’ATPase à proton/ H-ATPase = éjecte les ions H

20
Physiologie Générale Pr. Tobias Hevor
CH6 : Le potentiel de membranePour les gros éléments transportés la cellule est obligée de former des vésicules.
A. Un ion perméantL’endocytose est une invagination de la membrane.La pinocytose = endocytose de petits éléments.La phagocytose = endocytose des très gros éléments, La phagocytose entraîne une vésicule qui contient le phagosome qui sera détruit par les lysosomes (le systeme endolysosomal).
L’exocytose = permet de faire sortir de la cellule des éléments par exemple les neurotransmetteurs. Ek= -75mVENa= 440 = +55mV 50ECl= -60mV
C= q q= 96500 coulombs C = capacité membranaire V voltage (mV)Grâce a cette formule on trouve que de nombreux ions on traversé la membrane :Si C = q /VNombre d’ions traversant 1 cm2 de membrane ?C : capacité membranaire (1 microfarad / cm2)V : voltage (100 mV)(1 mole d’ions q = 96500 coulombs)C = q / V d’où q = C Vq = 10-6 x 10-1
F x Vq = 10-7 coulombs/cm2
Sachant que : 1 mole = 96500 coulombsX mole = 10-7 coulombs
96500 x X = 1.10-7 ou X = 1.10-7 / 96500X = 10-12 mole / cm2
B. Plusieurs ions perméant [Voir schéma cour PG page 18 et 19]
Ek = -70mV ; Vm = -70mV Ek = Vm = équation de Nernst
Si Vm ≠ Ek Vm – Ek ≠ 0 (-68) – (-70) = 2mV
Vm – Ek = V si V= I.R Vm – Ek = Ik.R
Vm – Ek = Ik R

21
Physiologie Générale Pr. Tobias Hevor
Ik = 1x Vm – Ek Ik = gk.(Vm – Ek) R
Vm – Ek = driving forcegk = conductance de l’ion KINa = gNa. (Vm - ENa)
Ik = - INa soit gk.(Vm – Ek) = -gNa. (Vm – ENa)
Vm = ENa. g Na + EK. g K ( gNa+gK) (gNa+gK)
On divise le numérateur et le dénominateur par gK.
Vm = ENa. (g Na /g K ) + EK. (g K /g K ) (gNa+gK)/gK (gNa+gK)/gK
Soit ά = gNa/gK
Vm = E K + α E Na ou Vm = EK
α + 1L’ion perméant impose son potentiel d’équilibre à la membrane.
On utilise la même formule avec trois ou quatre ions.
C. Le potentiel de membrane au repos. Equation de GoldmangNa ≈ 0 et gK augmente n’est valable que pour les fortes concentrations.
Si les concentrations sont petites l’estimation n’est pas valable et on doit utiliser l’équation de Goldman.
Vm = 58.log P K [K] e + P Na [Na] e + P Cl [Cl] i PK [K]i + PNa [Na] i + PCl [Cl] e
Les ions Cl sont deux fois plus perméant que les ions K qui sont 100 fois plus perméant que les ions Na.La Na-K-ATPase est à l’origine du gradient sodique de potassique. Les ions chlorures ne peuvent polariser une membrane, c’est le gradient qui polarise la membrane.
D. Equilibre électronique de la membrane au repos[Schéma cours page 20 PG]Au repos il n’y a pas de courant.INa = - IK
Dans une brancheVm = ENa + Vrésistance comme U+R.IVm = ENa + R.INa comme R= 1/g
Vm = ENa + 1/gNa.INa
Soit

22
Physiologie Générale Pr. Tobias HevorINa = gNa. (Vm – ENa)
Vm. ENa = driving force
Le canal élémentaire γ :Conductance généralegNa = γNa.NNa.PNa
NNa = nombre de canaux Na par cellulePNa = probabilité d’ouverture
Conductance en général gion = γion. Nion. Pion
g s’exprime ne Siemens (pico Siemens)
Expérience sur un axone géant de calmar
[Schéma cour prise de mesure PG]

23
Physiologie Générale Pr. Tobias Hevor
CH7 : Le potentiel d’action
I. Décours du potentiel d’actionLe PA peut changer de valeurs lorsqu’il est excité. Neurones, cellules musculaires, cellules glandulaires peuvent être excitables.

24
Physiologie Générale Pr. Tobias Hevor
CH8 :Les Synapses

25
Physiologie Générale Pr. Tobias Hevor
Ch9 : La neurotransmission
II. Les acides aminés inhibiteursCe sont des composés dérivés du glutamate par perte de l’amine : gammaaminobutiric acid = GABA.Expérience de microiontophorèse : injection de GABA dans la membrane post synaptiques, qui entraine une hyperpolarisation, inhibant de ce fait la cellule. C’est le premier inhibiteur découvert. On obtient le GAD en enlevant le COO de l’acide glutamique.
A. Le GABALe cofacteur pour la GAD est le pyridoxal. Les récepteurs sont organisés en 2 familles : les protéines canaux, qui sont des récepteurs de GABAA, et perméables aux ions chlorures, et GABAB permet l’ouverture des canaux à K+. Recapture et dégradation dans la fente par une enzyme, la gabatransaminase (GABAT).
B. La glycineAcide aminé le plus, on réalise l’expérience de microiontophorèse = on constate une hyperpolarisation = neurone glycinergique. Obéit au critère des neurotransmetteurs.Enzyme de synthèse = serine hydroxyméthyltransferase, les récepteurs sont des récepteur de type canaux qui laisse passer sélectivement des ions chlorures => hyperpolarisation de la membrane.Système utilisant la glycine : systeme nerveux réflexe au niveau du bras [Voir schéma cours scanner]
Neurone de renshaw = régule et inhibe le fonctionnement des motoneurones
III. les monoamines
A. La catécholamineCrée à partir de catéchol
1. Biosynthèse et régulationCellules cathecholaminergiques et cellules chromaphines (glande médullosurrénale).Point de démarrage est double :- début avec la phénylalanine qui grâce à la phénylalaninehydrolilase subit une oxydation
[Voir schéma cours 1]- ou début avec de la tyrosine
Étape suivante = hydroxyler la structure aromatique puis supprimer la fonction alcool la voie de métabolisation peut s’arrêter a ce niveau et ce stock dans les neurones dopaminergiques, mais cette voie de biosynthèse peut aussi continuer par une hydroxylation sur le carbone bêta et donner de la noradrénaline, stockée dans les neurones noradrénergiques. Mais dans quelques neurones et surtout dans les neurones de la glande médullosurrénale la voie métabolique existe et continue avec une méthylation du N de l’amine pour composer l’adrénaline ou l’épinéphrine => stocke cette adrénaline.
La tyrosine hydroxylase constitue une étape clé car c’est cette tyrosine hydroxylase qui va conditionner la suite de la voie métabolique. Cet enzyme subit une forte régulation => rétro inhibition/feed back (produit synthétisé par enzyme capable de l’inhiber)- tétrahydroptérine est un cofacteur d’enzyme et les produits des enzymes inhibent la réaction.

26
Physiologie Générale Pr. Tobias Hevor- Les kinases vont phosphorer la tyrosine hydroxylase qui aura plus d’affinité avec les tetrahydroptérines.- Si les cellules perdent leur catécholamine. Grâce à la réserpine on observe quasiment une disparition des cathécholamines, si le taux de cathécholamines baisse on observe une grande production d’ARNm de tyrosine hydroxylase.
2. Les récepteurs cathécholaminergiquesHomogénéité des récepteurs = essentiellement des récepteur liés aux protéines G :- Récepteur D1 => utilise l’adénine cycles comme intermédiaire, la stimulation de l’adénine
cyclase qui ouvre les canaux = active- Récepteur D2 => fonctionne pour inhiber adénine cyclase
Les neurones noradrénergiques :- alpha 1 => distingue en utilisant un agoniste (phényléphrine) utiliser pour augmenter la tension
artérielle en cas d’hypotension- alpha 2 => antagoniste la yohimbine- bêta 1 => agoniste : le salbutamol = ventoline (broncho-dilatateur)- bêta 2 => antagoniste : le propranolol = avlocardyl (pour les problèmes cardiaques)
Tout ces produit ciblent les terminaisons noradrénergiques = mime le systeme orthosympathique => sympathomimétique Les sympatholytiques = s’oppose au système orthosympathique
3. L’inhibition Catécholamines a une importance énorme dans l’inhibition mais présence aussi enzymes qui vont détruire les neurotransmetteurs : les monoamines oxydases. Dégrade les neurotransmetteurs par oxydation. Deux sous-ensembles :Monoamine oxydase (MAO) :- forme A = préférence pour la noradrénaline- forme B = agit indifféremment sur noradrénaline et dopamines
IMAO = inhibiteur de monoamines
Méthylation inhibé grâce à la catéchol-oxygéne-méthyl-transferase ou COMT qui inhibe les neurotransmetteurs du groupe catéchilergique. On peut aussi inhiber la recapture grâce a l’imipramine = tofranil => traitement dépression etc.Les amphétamines sont des sympathomimétique indirects qui facilitent la libération des neurotransmetteurs = excellent produit stimulant => dopant et toxique a long termes. Capable de produire un dérivé la 6-OHDOPA mais tue les neurones catécholinergique => utilisé pour détruire expérimentalement les neurones catécholinergique. Les métabolismes de ces catéchol apparaissent dans le liquide céphalorachidien.
B. La sérotonineAu démarrage la synthèse part du tryptophane [Voir schéma cours 2]
Etape clef = tryptophane hydroxylase mais a besoin de tetrahydroptérines Les récepteur = 5-HT plusieurs groupe:- 5-HT1a - 5-HT2- 5-HT3
On a synthétisé des antagonistes de ces récepteurs (les tryptans) parmi ces tryptophanes le sumatriptan (antimigraineux de références) qui sont analgésiques.

27
Physiologie Générale Pr. Tobias HevorL’inhibition se fait par recapture et aussi par dégradation (oxydation grâce aux monoamines et aux 5-hydroxy-indole catécholamines acid (5-HIAA) apparaît dans le liquide céphalorachidien et dans les urines => index du fonctionnement. Défaut dans les troubles psychiques (schizophrénie)
C. Les voie monoaminergiquesLa dopamine : les voies dopaminergiques :- noyau mésencéphalique = la substance substantia nigra - dans le diencéphale = hypothalamus - dans la rétine- dans le bulbe olfactif
La noradrénaline : les voies noradrénergiques :- locus coeruleus
La sérotonine : les voies sérotoninergiques :- noyau du raphé
Quelques neurones adrénergiques mais surtout les cellules chromaphines
IV. Les neuropeptidiquesOn observe dans le tube digestif des peptides qui ont des propriétés qui ressemblent aux propriétés des neurotransmetteurs, certaines substance fonctionnent comme des neurotransmetteurs (une trentaine de répertoriées.
A. Les tachykinines : la substance PDécouvert par Von Euler et Gaddum qui isolent de l’intestin une substance poudreuse qui contracte l’intestin = substance poudreuse = peptides de 12 acides aminées dodecapeptides. Un peptide est incapables de synthétiser des substances donc synthétisés dans le corps cellulaire qui seront transportés par le système orthograde rapide vers les terminaisons ou des enzymes vont hydrolyser ces précurseur en neurotransmetteur actif qui seront concentrés dans les vésicules synaptiques finalement la synthèse se fait dans la terminaison. Si on bloque le transport axonique on bloque la synthèse Ces vésicules libèrent leur contenu par un systeme calcium dépendant, il se fixe sur des récepteur peptidergiques.Les voies peptidergiques = 4 sous ensembles:[Voir schéma cours 3]- Neurone qui arrive à la paroi intestinale possède la substance P comme neurotransmetteur - Les neurones de la sensibilité douloureuse utilise aussi la substance P = neurones nociceptif vont utiliser comme neurotransmetteur la substance P. on a essayé de contourner cette voie pour annuler la douleur dans les viscères.- dans le striatum et dans la substancia nigra on trouve ces neurones.
B. Les neuropeptides opioïdes = les endomorphinesMorphine = extrait végétal de Papaver somniferum grâce a un écoulement de sève exsudat qui une fois séché donne de l’opium, qui sert d’analgésique. Mais progrès chimique permet l’extraction de la morphine (opiacée) si un produit végétal est capable d’etre analgésique il doit se fixer sur les récepteurs c’est qu’il doit exister de façon naturelle des ligands de ces récepteurs (travaux de Guillemin et Schally cherche ce ligand naturel) = découverte de l’endomorphine, fonctionne comme de la morphine = opioïde qui calme aussi la douleur de l’individu.Possibilité d’avoir des agonistes et des antagonistes :- naloxone- naltrexone

28
Physiologie Générale Pr. Tobias Hevor
3 familles d’endomorphine :- enképhalines- beta-endorphine- dynorphine => Neurotransmetteur opioïde
Enképhaline au nombre de 2Une constituée de tyrosine, de glycine(x2) phénylalanine et méthionine = met-enképhalineL’autre constitué de Tyr, Gly, Gly Phe, leu = leu-enképhalineLe précurseur de ces neurotransmetteur sont des protéines synthétisées par le péricaryon et transportées jusqu’aux terminaisons axoniques. 3 catégories de neurotransmetteur :- pro-opiomélanocortine (POMC)- pro-enképhaline - pro-dynorphine
Présence de la substance P au début de la séquence de la pro-opiomélanocortine, puis présence de mélanocyte stimulating hormone (MSH) => très grande protéines qui sera clivéMême logique pour les 2 autres molécules.Les récepteurs sont à 7 domaines transmembranaires (récepteur a protéine G) que l’on classe en 3 sous ensemble :- les récepteurs µ- le récepteur δ- le récepteur kappa
Les anképhaline ont une préférence pour les mus et les deltas les dynorphine ont une préférence pour les kappas. La morphine se fixe sur les récepteurs mu. L’inactivation peut se faire de manière non spécifique ce sont des aminopeptidases (découpe les tyrosines aux extrémités terminal et inactive la protéine) ou des ankephalinases (besoin de résidus métallique pour fonctionner = métallo-endopeptidase découpe peptide au milieu le coupe en deux et donc l’inactive) les voies nerveuses enkalinergiques sont des circuits locaux du cortex cérébrale et de l’hippocampe. Les neurones dynorphinergiques = cortex cérébrale et hippocampe. Pour la bêta endorphine beaucoup plus rare ont les trouve dans le noyau arqué.
C. Coexistence des neuropeptides et d’un autre neurotransmetteurOn utilise la méthode d’immunochimie pour localiser les neurotransmetteurs. On observe que parfois que certains neurotransmetteurs cohabitent dans les voies nerveuses. Un premier neurotransmetteur de petite taille est libéré puis un autre plus gros et plus tardif qui soutient l’autre.

29
Physiologie Générale Pr. Tobias Hevor
Ch10 : Les seconds messagers
I. Les protéines GCertaines protéines membranaires n’ont pas de canal, quand le ligand (neurotransmetteurs) se fixe il n’y a pas de canal pour s’ouvrir. Présence de protéines globulaires (G) dans la cellule qui permettent l’ouverture du canal.Lorsque le neurotransmetteur se fixe :- la protéine G se débrouille pour ouvrir directement le canal récepteur muscarinique de
l’acétylcholine ex cellules atrials du cœur- la protéine G passe par une autre protéine intercellulaire, petites substance qui ouvre le canal
=> second messager. Varié mais les principaux sont : AMPc (adénosine mono phosphate cyclique) ; la GMPc ; le calcium et des derivée d’acide gras comme le phosphatidylinositols, le diacylglycerol (DAG), l’inositol-triphosphates, l’acide arachidonique.
Plusieurs étapes- fixation du ligand sur le récepteur qui s’active (changement de conformation de la protéine
membranaire - activation transmise a la protéine G- agit sur une autre protéine de la cellule (protéine enzymatique qui va fabriquer le second
messager.
Les protéines G sont constituées en 2 familles- les monomériques (1 seule et unique protéine)- les trimériques (3 sous unités alpha bêta gamma qui sont associées a un GDP) lors de
l’activation le GDP s’en va a cause de la déformation. Puis la partie alpha de la protéine se sépare dés qu’elle a récupéré un GTP ensuite le GTP s’hydrolyse pour activer l’enzyme.
Colleras produit une toxine la toxine pertussique, le germe pseudomonas se fixe de façon permanente a la protéine G.
Qu’il s’agisse de monomère ou de trimère ont trouve des sous familles :- les protéines Gs stimulent l’adénine cyclase- les protéines Gi qui inhibe l’adénine cyclase- les protéines G0 qui agisse au sein de calcium extracellulaire- la transducine permet de traduire les systeme lumineux en systeme chimique fonctionne avec
des protéines G
II. Les enzymes des seconds messagers.L’adénine cyclase qui est une enzyme membranaire, associée à la membrane mais du coté interne. Transformer adénosine tri phosphate en AMPc qui est une molécule très réactive, soit l’AMPc va directement au niveau du canal ou l’AMPc passe par un autre intermédiaire PK (protéine kinase) qui était inactive jusqu'à la fixation de l’AMPc qui va activer la protéine et phosphoriler le canal qui va s’ouvrir et laisser passer les ions spécifiques.Sinon le canal s’ouvre par hydrolyse de l’AMPc qui devient de l’AMP et le canal se referme. L’enzyme qui permet cette hydrolyse est la phosphodietérase. Si on souhaite augmente le taux de AMPc on peut inhiber cette phosphodietérase par exemple en utilisant des dérivés de méthylxantine ou l’isobutyl-

30
Physiologie Générale Pr. Tobias Hevorméthylxantine (IBMX). La caféine est une inhibitrice de la phosphodietérase, le thé contient la théophylline qui est une xanthine mais moins efficace, puis la théobromine mais encore moins active.La guanyl-cyclase = le découpe dégradé par une phosphodietérase. Peut etre assimiler a l’AMPc.
Phospholipase c est capable d’agir sur cet inositol est libère l’inositol triphosphate puis il reste un diglycéride qui va etre soumis a l’action de la même phospholipase qui va découper une autre extrémité pour constituer l’acide arachidonique, et du diacide-glycerol. Chacun de ces composés vont constituer des second messagers IP3 pour agir sur le réticulum endoplasmique intracellulaire pour libérer du calcium dans le cytoplasme, responsable de formation de prostaglandine = lipide aux activités très variées (coagulation du sang….).
III. Les protéines kinase et les photos protéines phosphataseSe sont des protéines enzymatiques qui utilise l’ATP comme substrat en lui prenant un phosphate pour le mettre sur un autre composé (acide aminé d’autre protéines (serine, arginine, tyrosine)) la protéine sera phosphorilée et le canal va s’ouvrir. Ont peut classer ces kinase grâce au second messager qui les actives :- les PKA. Activé par l’AMPc - les PKG. Activé par la découpe- les PKC. Activé par le diacide glycérol
Déphosphorylations qui vont hydrolyser le phosphate situé sur l’acide aminé- type 1- type 2

31
Physiologie Générale Pr. Tobias Hevor
CH11 : La contraction musculaire
I. Rappel de la structure du muscle striéChez l’homme 80% de la masse d’un homme sont ces muscles. Plusieurs catégories :- muscle squelettique (striée)/ muscle volontaire- muscle cardiaque- les muscles lisses
A. Organisation Les cellules musculaires sont aussi appelées fibre musculaire ou myocytes. Les myocytes proviennent de cellules embryonnaires les myoblastes, pendant l’embryogenèse il y a fusion de myoblastes mais les noyaux ne fusionnent pas, après plusieurs fusions on a la création d’un syncytium. C’est pourquoi ces cellules sont plurinucléées. Ces fibre musculaire sont regroupées en faisceaux qui sont unis par une matrice extracellulaire d’origine conjonctive, la fibre musculaire est un faisceau très allongé (plusieurs centimètre au maximum) et assez épais (100µm au maximum). Terminologie :- Le cytoplasme = sarcoplasme => conglomérat de fibres parallèles (les myofibrilles) qui sont
réunies en faisceaux avec des organites (RE, le golgi) mais aussi des réserves métaboliques (réserve de glycogène, la myoglobine capable de fixer de l’O2 pour former une réserve)
- La membrane = sarcolemme- RE = réticulum sarcoplasmique qui entoure les faisceaux de myofibrilles disposés tout autour
Les noyaux sont périphériques sur la membrane plasmique.
B. Le sarcolèmeMembrane plasmique traditionnelle/ conventionnelle qui recouvre la lame basale composée de glycoprotéines, beaucoup de fibre élastique. Si on regarde de près la membrane on constate des petites invaginations (des tubes qui rentrent dans la cellule) => système tubulaires transverse = systeme T.
C. Les myofibrillesCes myofibrilles constituent 80% du volume du sarcoplasme, si on fait une coupe transversale d’un muscle il y a un million de myofibrille par cm2. Chaque myofibrille est un cylindre très allongé constitué d’une succession d’éléments sombres (bande/disque A= anisotrope) et d’élément clair (bande/disque I = isotrope) plus une bande H placer au milieu de la bande A et au milieu de la bande H on observe la ligne M. dans la bande I on observe la strie Z. entre deux strie Z on délimite le sarcomère qui est répété de nombreuses fois dans un myofilament. Dans la bande A on trouve des filaments épais (10nm de diamètre) qui vont chevaucher partiellement avec des filaments minces (6nm), et les filaments minces constituent la bande I. en phase de repos les filaments ne se touchent pas mais il y a des zones de proximité de 4nm. Et pourtant ces deux filament on une très forte affinité. Un filament épais est entouré par 6 filaments minces et cette géométrie est répéter n fois.
D. Le système transverse et le réticulum sarcoplasmiqueEntoure des myofilaments et fait des diverticules/expansions des poches qui peuvent recouvrir le tubule transverse mais il n’y a pas de contact physique, le bloc tube et manchon est appelé triade ou diade, sont très proche physiquement les plus privilégiés dans de couplage de l’excitation et la contraction musculaire = plaque electro-physiologique

32
Physiologie Générale Pr. Tobias HevorII. La contraction musculaire
A. Les molécules de la contraction musculaire.Deux familles de molécules qui sont impliquées dans la contraction musculaire :- famille majoritaire (actine myosine)- famille minoritaire
1. La myosineLa myosine = molécule composée d’une partie allongée fibrillaire et une parti plus globulaire au bout. La molécule est un hexamère (2 grandes sous unité sous forme hélicoïdale et qui deviennent globulaire a une extrémité = 200000 Da ; au niveau de la partie globulaire 4 petite sous unité de chaîne légère = 20000 à 40000 Da).La partie linéaire est la queue de la myosine.Les propriétés de la queue ont un rôle de structure alors que la tête a une activité enzymatique ATPasique. Son activité dépend du calcium et son activité est inhibée par le magnésium.La myosine est la matière des filaments épais et par rapport à la ligne M ces myosines sont disposées de part et d’autre de la ligne M, les têtes sont décalés les unes par rapport aux autres ce qui donne une sorte d’hélicoïde tout autour de l’épaisseur de filament épais.
[Schéma myosine PG]
2. L’actineL’actine est une protéine globulaire qui est capable de se polymériser pour constituer un filament (actine F). Dans la nature ces filaments sont sous la forme de dimère entrelacée.
[Schémas actine PG]Très forte affinité entre la tête de myosine et le filament d’actine ce qui donne l’acto-myosine.
3. Protéine régulatriceCes protéines sont indispensables pour la contraction musculaire :- la tropomyosine = protéine filamenteuse de petite taille (dimère). Ce positionne sur la molécule
d’actine[Schéma actine tropomyosine]
- la troponine(TN) = 3 sous unité : la TN-C a une affinité avec le calciumla TN-I inhibiteurla TN-T attachement
la TN-I occupe le site d’affinité = inhibe l’association myosine actine
B. Mécanisme de la contraction musculaire
1. Aspect généralSi on soumet un sarcomère a une contraction on observe une diminution de la taille du sarcomère => emboîtement de l’actine entre les myosines = bande H rétrécit et disparaît en cas de contraction maximale.
2. Mécanisme moléculaireLa TN-C a une forte affinité pour le Calcium, le calcium se fixe sur la TN-C et change sa conformation dans l’espace, les autres sous unité subissent aussi ce changement de conformation et par conséquent la TN-I va changer de place et libérer le site d’affinité actine myosine. Le changement de conformation de la TN-C fait bouger tout le bloc. La myosine va venir se fixer pour créer un pont. L’ATP va être hydrolysé et l’énergie libérée va permettre une rotation de la tête de myosine qui est attachée à l’actine, elle va entraîner l’actine puis en utilisant encore l’énergie de l’ATP cette tête se détache et si le calcium est encore présent le cycle recommence. Transduction de l’énergie chimique sous forme d’énergie mécanique.

33
Physiologie Générale Pr. Tobias HevorC. L’énergie nécessaire a la contraction musculaire
Besoin d’ATP pour déplacer la tête de myosine de plus s’il n’y a pas d’ATP dans la cellule la liaison actine-myosine reste définitive et le muscle devient rigide. Cette énergie vient de l’ATP donc de l’utilisation du glucose, qui sera dégradé par la glycolyse qui donnera du pyruvate qui ira dans le cycle de Krebs ou il y aura la formation d’un certain nombre de molécules d’ATP. Le cycle de Krebs est en relation avec la chaîne respiratoire pour fabriquer beaucoup d’ATP. Les cellules musculaires ont mis en place d’autre systeme pour créer de l’ATP par exemple ADP va etre modifier par la myokinase, cet enzyme va prendre deux ADP pour former un ATP et un AMP. Mais les cellules musculaires ont des phosphagènes qui sont capable de générer du phosphate (créatine phosphate qui peut être phosphorilée) si une créatine phosphate phosphorilée rencontre un ADP elle peut transférer son phosphate a un ADP pour créer un ATP = transphosphorylation. Le glucose utiliser peut provenir du sang mais il peut aussi provenir d’une réserve que la cellule musculaire a fait (le glycogène). Les cellules musculaires sont capables d’utiliser des acides gras (corps cétoniques). La cellule musculaire a beaucoup évolué pour toujours avoir des réserves d’énergie.Plusieurs catégories de cellule musculaires :- blanche = pauvre en myoglobine (réserve d’oxygène)- rouge = riche en myoglobine
III. Couplage de l’excitation contraction
A. La plaque motrice1) la terminaison axonique
Voir antérieurement2) la fente synaptique
Voir antérieurement3) la sarcolemme
B. Chronologie de la contraction musculaire1) libération de l’acétylcholine
Voir antérieurementLorsque l’axone moteur n’est pas stimulé s’il n’y a pas de potentiel d’action de temps en temps il y a une vésicule synaptique qui libère sont contenu en acétylcholine dans la fente synaptique. Dans cette libération aléatoire il y a une certaine quantité d’acétylcholine cette libération est qualifié de libération quantique. Mais elle n’est pas suffisante pour dépolariser la membrane post-synaptique. Lors d’une stimulation il y a environ 200 vésicules qui vont se libérer et on voit un potentiel d’action de la membrane musculaire. Le délai synaptique est de 200 à 300µs qui est majoritairement du a l’éxocytose.
2) action de l’acétylcholine L’acétylcholine se fixe sur des récepteurs nicotiniques, au passage une partie de cette acétylcholine est dégradée. Ces récepteurs nicotiniques sont des pentamères. Ce canal s’ouvre et laisse passer 20000 ions sodium par canal et si on considère un seul canal on observe une dépolarisation locale pour commencer à dépolariser localement = potentiel post synaptique excitateur (PPSE)
Exemple chez la torpille : les cellules musculaires sont empilées en très grand nombre un motoneurone envoie des collatérales pour faire des synapses avec tout la pille. On évalue le potentiel musculaire à 150 mV par cellules. Vu le nombre de cellules musculaires la torpille arrive à déclencher un décharge de 500V.
3) potentiel post synaptique excitateur (PPSE) et potentiel musculaire Les PPSE se comportent comme des potentiels électro-toniques c'est-à-dire qui se propagent de façon décrémentiel. C’est PPSE sont sommable et dés que la somme des PPSE atteint le seuil de

34
Physiologie Générale Pr. Tobias Hevordéclenchement de la cellule musculaire il se transforme en potentiel d’action musculaire qui se propage tout le long de la membrane de la fibre musculaire. Vitesse est environ un dixième de celle d’un axone. On peut arrêter artificiellement ces PPSE en utilisant du curare (D-tubocurarine) qui se positionne au niveau du curare et le bloque lorsqu’il s’ouvre.Le potentiel d’action rentre à l’interieur de la cellule grâce au systeme tubulaire transverse et passe a coté du réticulum sarcoplasmique qui est munie de canaux calciques voltage dépendants qui vont s’ouvrir. Or le réticulum sarcoplasmique contient 1000 fois plus de calcium que le sarcoplasme et donc le gradient fait que le calcium sort rapidement et va sur la TN-C pour initier le mécanisme de contraction.Expérience : calcul de plusieurs paramètres sur une fibre musculaire de balane. Ces paramètres sont :- taux de calcium dans la fibre (avec l’utilisation échorine qui fluore en présence de calcium)- le potentiel d’action- la contraction musculaire
on observe rapidement un potentiel d’action puis une luminescence qui correspond a l’entrée de calcium dans le sarcoplasme puis ensuite la contraction.
Il y a au niveau du réticulum sarcoplasmique des pompes à calcium pour faire rentrer le calcium dans le réticulum sarcoplasmique. Dés qu’il n’y a plus de calcium on revient à l’état de repos.
IV. Autre type de fibres
A. Le muscle cardiaque Le myoblaste est un muscle avec un noyau axial, la cellule est ramifiée par un fuseau simple. Les cellules sont unies par des connexions (disques intercalaires = scalariforme) sinon les sarcomères sont classiques.Le potentiel d’action de la membrane est un plateau puis une pointe. Si on bloque les canaux sodiques grâce a une toxine on remarque que la pointe disparaît et l’on devine que cette pointe est due a l’entré de sodium dans la cellule. Si on remplace du calcium par du manganèse on observe une disparition du plateau qui est dû aux ions calcium. Ainsi on démontre que dans les cellules cardiaques il y a des canaux calco-sodiques.
B. Les fibres lissesFuseaux réduit a une seule cellule les microfibrilles sont décaler les uns au autre = puissance moins forte. Conexine entre les cellules des muscles lisses, la synapse est très large, la fente synaptique est très large. Le potentiel d’action arrive est après la réponse arrive beaucoup plus tard. => utérus vessie vaisseaux sanguin.

35
Physiologie Générale Pr. Tobias Hevor
CH12 Biomécanique de la contraction musculaire
I. La contraction musculaireLa contraction est isotonique dans le cas du raccourcissement d’un muscle, la valeur de la force reste la même. Si on accroche les extrémités, le muscle ne peut pas se contracter et cette fois-ci on a une contraction isométrique (de même longueur), la valeur de la contraction augmente. La fibre musculaire ne se contracte pas seule = sommation.
II. Sommation 2 types de sommation :- somation spatiale- somation temporelle (2 contractions fusionnent si elles sont très proches dans le temps) => tétanos où l’on distingue les différentes contractions unitaires (tétanos imparfait), mais si l’on ne distingue pas les contractions unitaire (tétanos parfait).Modèle de schématisation- composante contractile- composante élastique en série- composante élastique en parallèle
Si entre deux contractions la composante élastique n’a pas encore rendu tout l’énergie elle emmagasine de l’énergie => contraction plus élevée et tétanos parfaits.
Si le muscle est fatigué l’amplitude diminue, la valeur de base augmente.
III. L’électromyographieOn peut analyser le muscle au repos en action etc.…Ceci permet d’avoir une idée de la contraction musculaire et permet de déceler les anomalies musculaires.