
PHYSIOLOGIE DES ANIMAUX DOMESTIQUESDOMESTIQUES · PHYSIOLOGIE DES ANIMAUX DOMESTIQUESDOMESTIQUES...
31
Rosalie Adina BĂ LĂ CEANU Nicolae DOJANĂ Rosalie Adina BĂ LĂ CEANU Nicolae DOJANĂ Rosalie Adina BĂ LĂ CEANU Nicolae DOJANĂ Rosalie Adina BĂ LĂ CEANU Nicolae DOJANĂ PHYSIOLOGIE DES ANIMAUX PHYSIOLOGIE DES ANIMAUX PHYSIOLOGIE DES ANIMAUX PHYSIOLOGIE DES ANIMAUX DOMESTIQUES DOMESTIQUES DOMESTIQUES DOMESTIQUES Physiologie de la respiration I case you want to refer to material in this textbook, the proper citation is: R.A. Balaceanu, N. Dojana. Physiologie des animaux domestiques: Physiologie de la respiration, Printech, 2020, Bucharest, Romania Chapitre élaboré avec la contribution de P. Morizot
Transcript of PHYSIOLOGIE DES ANIMAUX DOMESTIQUESDOMESTIQUES · PHYSIOLOGIE DES ANIMAUX DOMESTIQUESDOMESTIQUES...

Rosalie Adina BĂ LĂ CEANU Nicolae DOJANĂRosalie Adina BĂ LĂ CEANU Nicolae DOJANĂRosalie Adina BĂ LĂ CEANU Nicolae DOJANĂRosalie Adina BĂ LĂ CEANU Nicolae DOJANĂ
PHYSIOLOGIE DES ANIMAUX PHYSIOLOGIE DES ANIMAUX PHYSIOLOGIE DES ANIMAUX PHYSIOLOGIE DES ANIMAUX
DOMESTIQUESDOMESTIQUESDOMESTIQUESDOMESTIQUES Physiologie de la respiration
I case you want to refer to material in this textbook, the proper citation is: R.A. Balaceanu, N. Dojana. Physiologie des animaux domestiques: Physiologie de la respiration, Printech, 2020, Bucharest, Romania Chapitre élaboré avec la contribution de P. Morizot

2 R. Bălăceanu, N. Dojană
Chapitre 3 : Physiologie respiratoire
1- Organisation fonctionnelle du système respiratoire
Le système respiratoire est l’ensemble d’organes permettant la respiration externe, c'est-à-dire les échanges gazeux entre l'organisme et l'environnement.
• Structure générale du système respiratoire L’appareil respiratoire peut être subdivisé en deux parties : - partie « non alvéolaire » à rôle de conduction de l’aire - partie alvéolaire à rôle déchanges gazeux avec l’organisme. Ces deux parties sont en continuité l’une avec l’autre via une suite d’embran-chements de plus en plus nombreux et de calibre de plus en plus petit. Cette structure très ramifiée lui vaut également le nom d’«arbre respiratoire» (Fig. 3.1).
L'arbre respiratoire Voie de conduction de l’air Voies d’échange gazeuse
Voies cartilagineuses Voies membranaires Unitatés respiratoires terminales
Trachée Bronches Bronchioles Bronchiole respiratoires
Canaux alvéolaires
Sacs alvéolaires
Nonalvéolaires Alvéolaires
Génération de branchement
1, 2 …………. 10 11, 13 ……. 16 17 - 19 20, 22, 23, 24 …
Circulation bronchique Circulation pulmonaire
• Fonctions du système respiratoire La fonction primaire du système respiratoire est l'échange de gaz respiratoires (O2 et CO2) entre l'organisme et l'environnement. Autres fonctions: - thermorégulation - métabolisme - participation à l’ajustement du pH sanguin
Fig. 3.1. Schéma d’arbre respiratoire avec la représentation des voies de conduction de l’aire, des voies d’échange et de certains détailles histologiques
(selon W.F. H o f m a n et D.C. M e y e r , Respiratory physiology dans Essentials of human physiology, Georgia, 1996)

Physiologie des animaux 3
- phonation - protection contre des substances toxiques, la poussière et les agents infectieux.
Volumes et capacités pulmonaires
Volume courant (VC): c’est le volume mobilisé à chaque cycle respiratoire pendant une respiration normale (de repos). Automatique et inconsciente, il est une fonction de la masse corporelle (Mc) (selon Schmid t -Nie l sen , 1983):
VC = 0,0062 × Mc1,01
Volume de réserve inspiratoire (VRI): c’est le volume maximum pouvant être inspiré en plus du VC par une inspiration profonde (environ 13 L chez le cheval). Volume de réserve expiratoire (VRE) : volume maximum pouvant être rejeté en plus du volume courant par une expiration profonde (environ 11 L chez le cheval). Volume résiduel: volume d'air se trouvant dans les poumons à la fin d'expiration forcée. Autrement dit qu'il est impossible d'expirer. Il est impossible de mesurer ce volume avec des tests de spirométrie. Le volume de collapse est le volume d'air retiré des poumons en ouvrant l'espace pleural. Le volume minimum est le volume d'air restant dans les poumons après l’élimination du volume de collapse. La capacité inspiratoire est le volume d'air qui peut être inspiré à la fin d'une expiration courant. Il résulte de la somme du volume courant avec le volume de réserve inspiratoire. La capacité résiduelle fonctionnelle (CRF) est le volume d'air dans les poumons à la fin d'une expiration courant. Il résulte de la somme du volume résiduel avec le volume expiratoire de réserve. Le CRF est chiffré à environ 45 ml / kg de poids vif ou, en chiffres absolus: 16 L chez le cheval, 1,5 L chez le mouton, 1 L chez le chien, 0,1 L chez le chat. La capacité vitale est le volume total d'air qui peut être expulsé après une inspiration maximale. Il résulte de la sommation du VRC avec le volume inspiratoire de réserve et le volume expiratoire de réserve. La capacité pulmonaire totale (CPT) est le volume total d'air dans les poumons à la fin de l'inspiration forcée. Le CPT résulte de la somme de tous les volumes pulmonaires. C'est environ 42 L chez le cheval. Les volumes pulmonaires sont mesurés par spirométrie. Le volume résiduel est le seul qu’on ne peut pas le mesurer directement par le spiromètre car on ne peut pas expirer ce volumed’air. Ainsi, la détermination de la capacité pulmonaire totale, qui inclut le volume résiduel, implique l'utilisation de méthodes indirectes telles que la pléthysmographie corporelle totale. Volume expiratoire maximal PAR seconde (VEMS): c’est le volume d’air expiré en une seconde au cours d’une expiration forcée effectuée après une inspiration maximale. Le sujet gonfle au maximum puis expire aussi fort et aussi rapidement qu’il le peut :

4 R. Bălăceanu, N. Dojană
VEMS/Cap. inspir = 0,8 .
2- Mécanique de la respiration
La respiration désigne ici la fonction biologique permettant l'absorption de l'oxygène dans l’organisme et le rejet de gaz carbonique. On ne parle pas ici de la respiration cellulaire.
a- Cycle respiratoire
Le cycle respiratoire est la succession des processus du moment du debut d’une inspiration jusqu’au moment du debut de l’inspiration suivante. On a décrit deux phases : l'inspiration et l’expiration.
• L’inspiration Inspiration c’est le processus pendant/par lequel l'air est aspiré dans les poumons. Il est un mouvement respiratoire actif, initié par la contraction du muscle diaphragme et des muscles intercostaux externes. Les principales étapes de l’inspiration sont : - la contraction des muscles intercostaux permet aux côtes et au sternum d’être tirés vers le haut ; - le diaphragme se contracte, devient plat en s’abaissant et se raidit ; - la cage thoracique augmente ainsi en volume ; - les poumons prennent alors aussi en volume. La plèvre étant collée à la paroi interne de la cage thoracique, elle force les poumons à s'étirer ; - la pression à l’intérieur des poumons diminue à des valeurs sous les valeurs atmosphériques ; - l'air pénètre dans les poumons, en s'écoulant librement le long des voies de conduction, jusqu'à ce que la pression alvéolaire soit égale à la pression atmos-phérique. Volume respiratoire courant (VRC) c’est le volume d'air pénétrant dans les poumons lors d’une inspiration légère. La pression alvéolaire évolue au cours des différentes phases du cycle. Elle vaut ~760 mmHg à la fin de l’expiration.
• L’expiration Expiration est le mouvement respiratoire/processus pendant/par lequel l'air est éliminé des poumons. Les principales étapes de l’expiration sont : - relâchement des muscles intercostaux permettant aux côtes et au sternum de redescendre et du diaphragme qui se courbe et remonte - la cage thoracique perd ainsi en volume et les poumons ont alors un volume plus petit - la pression à l’intérieur des poumons devienne plus grande que celle atmosphérique.

Physiologie des animaux 5
Ainsi, l’air existant dans les poumons en est retiré jusqu’à ce que la pression alvéolaire soit à nouveau égal à pression athmosphérique, ce qui marque la fin de l’expiration, après quoi le cycle reprend. L’expiration est un acte habituellement passif, résultant de la relaxation des muscles respiratoires et de la rétraction élastique des tissus pulmonaires et thoraciques tendu par l’inspiration. La dernière partie de l'expiration, cela dit, peut être assistée par la contraction des muscles expiratoires intercostaux internes et abdominaux.
• Coefficient de ventilation Lors d’un cycle respiratoire, tout l'air présent dans les alvéoles n'est pas remplacé par de l'air «frais». Le pourcentage d'air alvéolaire remplacé par l'air atmosphérique frais à chaque respiration est appelé coefficient de ventilation.
b- Fréquence respiratoire
Fréquence respiratoire (FR) est le nombre de cycles respiratoires par minute (tableau 3.1). Le débit respiratoire est le produit de la FR et le VRC : FR x VRC = débit respiratoire
Tableau 3.1. Fréquence respiratoire chez les animaux au repos (selon Kolb, 1973)
Espèce Moyenne (respirations/min)
Limites de variation
Cheval Vache
Mouton Chèvre
Porc Chien (grande taille)
Poule Chat
12 20 15 15 13 15 45 30
8 - 16 12 - 28 12 - 20 12 - 20 8 - 18
10 (15) – 20 (30) 40 – 50 20 – 40
c- Types de respiration
Selon la façon dont se fait la respiration, les mouvements qui la caractérisent, on différencie plusieurs types respiratoire. Ces types respiratoires peuvent être corrélés avec un type énergétique et peuvent caractériser une espèce. • Type costal Ce type respiratoire se caractérise par une prédominance de l'amplitude des mouvements des côtes sur celle des mouvements abdominaux (ex : chevaux de petite taille et chiens). • Type abdominal Les mouvements abdominaux prédominent sur ceux des côtes (cas des ruminants et des lapins).

6 R. Bălăceanu, N. Dojană
• Type costo-abdominal (mixte) L'amplitude des mouvements costaux est relativement égale à celle des mouvements abdominaux. • Facteurs de modification du type respiratoire Les douleurs abdominales, la compression de la poitrine (par exemple à cause du harnais d'un cheval), la gestation ou l'accumulation de gaz dans le rumen entraîneront une transition vers la respiration costale. NB : Chez les espèces à long palais mou (velum, chez le cheval), la respiration est exclusivement nasale. Chez les espèces à palais mou court, la respiration orale est possible.
3- Les forces impliquées dans la respiration
a- Force active et force de résistance
La force active (Fa) désigne la force qui augmente le volume de la cavité thoracique lors de l’inspiration. Il s’agit donc de la force de contraction des muscles inspirateurs. Les principaux muscles inspirateurs étant : - le diaphragme. - les muscles intercostaux externes. - les muscles sternocléidomastoïdien et scalènes dans une moindre mesure. La force de résistance (Fr) désigne celle qui s'oppose à l'augmentation du volume de la cavité thoracique (tableau 3.2). Elle est représentée par :
Tableau 3.2. Distribution de l'effort respiratoire en fonction de la résistance des composants de l'arbre respiratoire (selon Cunningham, 1992)
Résistance à vaincre:
Composants % de l'effort
respiratoire total
élastique 1. Poumons (surfactant 50 – 80%, tissue 50%)
2. Parois de la cage thoracique 60 – 66
friction 1. Tissus (20%)
2. Voies respiratoires (80%) 30 – 35
inertie 1. Poumons
2. Parois de la cage thoracique 3. Air
2 – 5
- la force de résistance élastique (Fe) due à l’élasticité des poumons et à l’élasticité de la paroi de la cage thoracique. - la force de friction de l'air et des tissus (Ff) - la force d'inertie de l'air et de la masse tissulaire thoracique et pulmonaire (FI) : Fr = Fe + Ff + FI
Pour que l'inspiration ait lieu, Fa doit être supérieure à Fr.
Physiologie des animaux 7
b- Élasticité, élastance et compliance pulmonaires
Élasticité pulmonaire est la propriété des poumons de s'allonger sous l'action d'une force d'étirement et de revenir à leur taille initiale lorsque la force d'étirement cesse d'agir. • Elastance pulmonaire L’élastance pulmonaire (Fe) est un concept en physiologie qui mesure la résistance élastique opposée à la tension pulmonaire. On la calcule selon la formule :
V
PFe
∆
∆=
où ∆P est la différence de pression qui produit un différence de volume pulmonaire, ∆V. L'élastance pulmonaire est due aux fibres élastiques de la structure pulmonaire (50%) et au surfactant (50%). • Compliance pulmonaire Il est préférable d'utiliser la notion de compliance pulmonaire (C) pour caractériser les propriétés des poumons. La compliance pulmonaire définit la facilité avec laquelle un poumon peut être gonflé :
P
V
Fe
1C
∆
∆== .
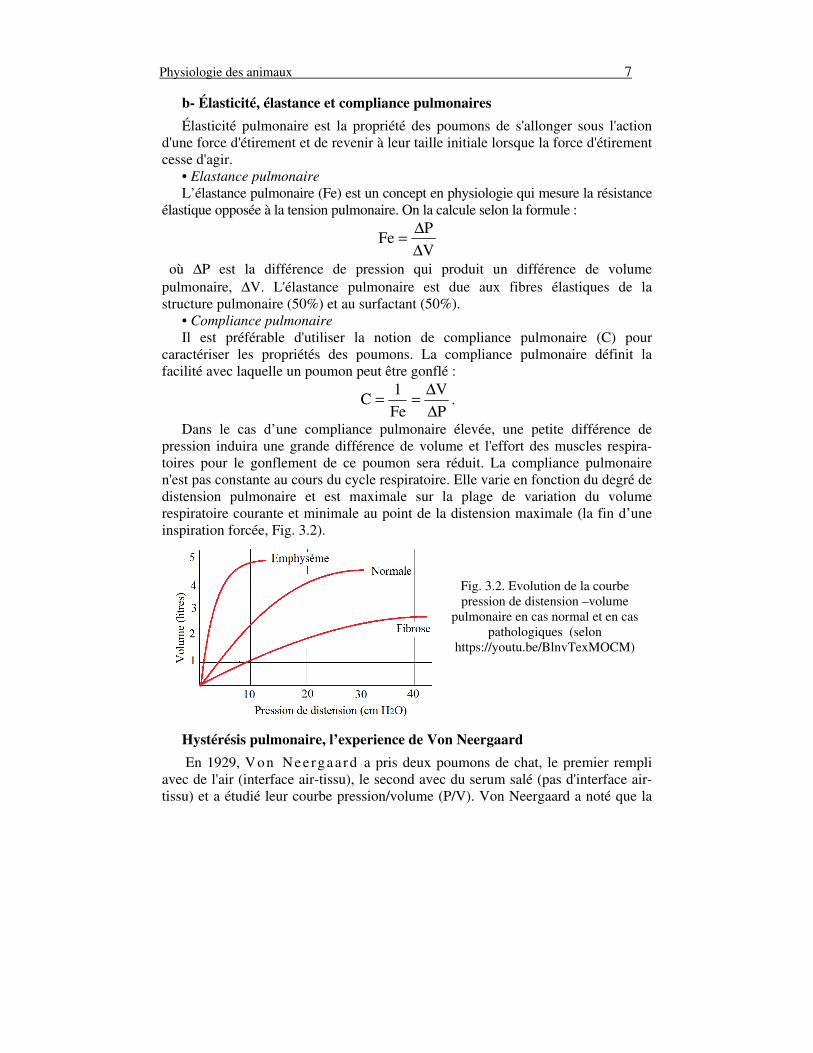
Dans le cas d’une compliance pulmonaire élevée, une petite différence de pression induira une grande différence de volume et l'effort des muscles respira-toires pour le gonflement de ce poumon sera réduit. La compliance pulmonaire n'est pas constante au cours du cycle respiratoire. Elle varie en fonction du degré de distension pulmonaire et est maximale sur la plage de variation du volume respiratoire courante et minimale au point de la distension maximale (la fin d’une inspiration forcée, Fig. 3.2).
Hystérésis pulmonaire, l’experience de Von Neergaard
En 1929, Von Neergaa rd a pris deux poumons de chat, le premier rempli avec de l'air (interface air-tissu), le second avec du serum salé (pas d'interface air-tissu) et a étudié leur courbe pression/volume (P/V). Von Neergaard a noté que la
Fig. 3.2. Evolution de la courbe pression de distension –volume
pulmonaire en cas normal et en cas pathologiques (selon
https://youtu.be/BlnvTexMOCM)

8 R. Bălăceanu, N. Dojană
pression nécessaire pour gonfler / dégonfler les poumons remplis d'air est presque le double de la pression nécessaire pour les poumons remplis d'eau. En revanche, la courbe de déflation du poumon rempli d'air ne suit pas la même courbe pression-volume que celle lors du gonflement. Cette différence entre la pression pendant le gonflement et celle de la déflation (respectivement, entre les propriétés d'élasticité du poumon pendant le gonflage et le dégonflage) est appelée hystérésis ou histérèse. On remarque que la courbe P/V du poumon rempli avec du sérum salé a une pente et donc une compliance beaucoup plus importante et une hystérèse beaucoup moins marqué (moins de différence entre inflation et déflation). On observe une meilleure compliance en l'absence d'interface et donc de tension superficielle (poumon gonflé a l'eau salée). La tension superficielle s'oppose a la compliance, mais cette tension superficielle peut être limitée par le surfactant pulmonaire (Fig. 3.3). L'hystérèse pulmonaire est due au surfactant pulmonaire, absent en cas de gonflement avec une solution saline. L'hystérèse pulmonaire est plus élevée chez le fœtus jusqu'au premier cycle respiratoire, tant que le poumon est complètement essoufflé. Pendant la respiration eupnéique, le gonflage pulmonaire commence de la capacité résiduelle fonctionnelle (environ 40 à 45% de CPT). Il en résulte une forte hystérèse pulmonaire car les alvéoles sont déjà ouvertes, leur pression d'ouverture étant nulle.
c- Surfactant pulmonaire
La pression nécessaire pour détendre un poumon rempli d'eau est plus faible que lorsqu'il est remplie d'air car les alvéoles pulmonaires sont tapissées de surfactant pulmonaire, un mince film de liquide aux propriétés tensioactives (diminue la tension superficielle, TS). Ce film liquide est dissous, disparaissant lors du remplissage des poumons avec de l'eau. Le surfactant pulmonaire est le complexe de substances lipoprotéiques qui couvre la surface des alvéoles pulmonaires sous forme de fine pellicule de liquide. Sa composition est la suivante : - 40% de di-palmito-phosphatidylcholine - 40% d'autres phospholipides (phosphatidylcholine, phosphatidylglycérol)
Fig. 3.3. Démonstration de l’histérèse et de la compliance pulmonaire. Compa-raison des courbes P/V avec des pou-mons de chat contenant soit une solution salée soit de l’air. On remarque qu’an poumon gonflé par une solution salée a une compliance plus élevée et un hystérèse moins marquée que le pou-mon gonflé avec de l’air (d’aprés E.P. Radfort, Tissue Elasticity. Washington, D.C., American Physiology Soc. 1957, p. 177)

Physiologie des animaux 9
- 5% de protéines associées - du cholestérol et des traces d'autres substances Le surfactant est produit par des cellules alvéolaires de type II. Les molécules spécifiques au surfactant sont tensioactives, elles ont la propriété d'abaisser la TS des liquides dans lesquels elles se trouvent. En effet, les molécules de surfactant ont un pôle hydrophile et un pôle hydrophobe. La partie hydrophobe est dirigée vers la surface du film liquide, en contact avec l'air alvéolaire. Le pôle hydrophile est dirigée vers la profondeur du film. Il occupe la surface de contact des molécules d'eau avec l'air, abaissant la force d'attraction entre les molécules d'eau et donc la TS. À l'inspiration, la surface alvéolaire augmente de sorte que les molécules de surfactant sont plus rares, plus dispersées, et celles d'eau plus denses à la surface des alvéoles (Fig. 3.4). Cette situation augmente la TS. À l'expiration, lorsque les alvéo-les pulmonaires diminuent, les mo-lécules de surfactant deviennent plus denses et, par conséquent, réduisent la TS. Les valeurs de TS du film de fluide alvéolaire sont d’environ 5 dynes/cm en début d’inspiration et atteignent ~50 dynes/cm à la fin de l‘inspiration.
• Les rôles du surfactant pulmo-naire - Augmentation de la compliance pulmonaire en réduisant l'effort d'in-spiration. - Contribution à la relaxation pulmonaire en facilitant l'expiration. - Réduction de la pression d'ouverture ou de gonflement dans les alvéoles effondrées ou petites. - Contribution à la stabilité des alvéoles, empêchant leur fusion. En effet, si les alvéoles étaient tapissées d'un film d'eau ou de plasma, la pression de repos transpulmonaire de −5 cmH2O serait insuffisante pour maintenir les alvéoles enflées et elles fusionneraient.
d- La résistance pulmonaire
La résistance pulmonaire s'oppose au passage de l'air et est caractérisée notamment par les forces de friction entre : - les molécules d'air et la paroi des voies respiratoires. - les surfaces de la plèvre pariétale et viscérale. - les tissus de la cavité thoracique.
Fig. 3.4. L’effet de l’inspiration sur le surfactant pulmonaire (iconographie personnelle)

10 R. Bălăceanu, N. Dojană
La résistance pulmonaire à l'écoulement de l'air est influencée par la surface totale de section de l'arbre respiratoire. La surface totale de cette section est la somme des surfaces de chaque voie respiratoire. De faibles changements dans la section transversale ont de grands effets sur la résistance et entraînent des effets sur l'effort respiratoire.
• Loi d’Ohm Selon la loi d'Ohm, la résistance pulmonaire (R) est égale au rapport de la différence de pression entre deux points des voies aériennes (P1− P2 = ∆P) et le débit respiratoire ( ) :
• Loi de Poiseuille Selon la loi de Poiseuille adaptée à l'écoulement des liquides dans des tubes cylindriques, la résistance à l'écoulement (R) est calculée selon la formule : , où : η est le coefficient de viscosité l et r sont, respectivement, la longueur et le rayon du tube. De ces deux relations, on peut calculer le débit respiratoire : L'analyse combinée des deux équations montre que lorsque l'on augmente la longueur du tube, par exemple de deux fois, la ∆P doit augmenter deux fois pour maintenir le constant. Mais pour une diminution de moitié du rayon du tube, la pression devrait augmenter de 16 fois pour maintenir le débit d'air constant.
Fig. 3.5. Distribution de la résistance respiratoire dans l'arbre trachéobronchique (courbe noire): la résistance au passage de l'air est
réduite dans la trachée et les grandes bronches, maximum dans
les bronches moyennes, puis diminue jusqu'à la périférie de l'arbre respiratoire. La surface totale de la coupe augmente progressivement alors que la
vitesse de l'air diminue progressivement
V
PR
&
∆=
4rπ
l8ηR
⋅
⋅=
l8η
r P
R
1P
R
∆PV
4
⋅
⋅π∆=×�∆==&

Physiologie des animaux 11
La trachée et les grandes bronches offrent une faible résistance au passage de l'air. La résistance la plus élevée s’effectue au niveau des bronches de taille moyenne, d’ordre 4 à 8. Les voies respiratoires ayant un diamètre compris entre 2 et 5 mm contribuent à plus de 80% à la résistance pulmonaire, tandis que la contribution des bronchioles est inférieure à 20%. 80% de la résistance pulmonaire totale est due aux voies respiratoires et 20% par la viscosité des tissus (Fig. 3.5).
•Facteurs influençant la résistance pulmonaire Une bronchoconstriction et une sécrétion accrue des glandes de la structure de l'épithélium de l'arbre respiratoire augmentent la résistance alors qu’une broncho-dilatation la diminue.
e- L’inertie pulmonaire
La force d'inertie est directement proportionnelle à la masse (m) des tissus impliqués dans le mouvement et à la masse de l'air inspiré ainsi qu'à l'accélération ou au taux de variation du flux respiratoire (v2). Les poumons et la cage thoracique ont une masse élevée mais une accélération réduite, de sorte que leur inertie est réduite. L'air qui pénètre dans les poumons a une forte accélération, mais une masse faible, donc l'inertie de l'air est également faible.
4- Echanges gazeux respiratoires
Les gaz respiratoires échangés sont l'oxygène et le dioxyde de carbone. Cet échange de gaz s’effectue à deux niveaux : - au niveau alvéolo-capillaire (entre les alvéoles et les capillaires qui les tapissent) - au niveau capillairo-tissulaire, entre les capillaires et les autres tissus.
a- Echanges gazeux pulmonaires (alvéolo-capillaires)
Les échanges gazeux pulmonaires ont lieu entre l'air des alvéoles et le sang des capillaires alvéolaires. Ces échanges consistent au transport de l'oxygène de l'air alvéolaire vers le sang des capillaires alvéolaires et du dioxyde de carbone sanguin vers l'air alvéolaire. Le mécanisme d'échange est celui de la diffusion facilitée. Rappel: le sens de la diffusion est celui du gradient de concentration, c’est-à-dire des zones de concentration les plus fortes aux zones de concentration les plus faibles. Cela, jusqu'à ce que les concentrations soient à l’équilibre entre les deux milieux. • Taux de diffusion Le taux de diffusion (TD) d’un gaz à travers la membrane alvéolaire, selon le loi de Fick dépend de : - la différence de pression partielle ou de concentration (∆P) entre les deux faces de la membrane de diffusion. - la surface de diffusion (A) - la distance c’est-à-dire l’épaisseur de la membrane de diffusion (D)

12 R. Bălăceanu, N. Dojană
- du coefficient de diffusion spécifique à chaque substance, où S est la solubilité et PM est le poids moléculaire: NB : L'ouverture de nouveaux capillaires dans un tissu entraîne une diminution de la distance de diffusion et une augmentation de la surface de diffusion. Les tissus nécessitant une plus grande quantité d'oxygène ont un plus grand nombre de capillaires ouverts, ce qui fournit une surface de diffusion plus élevée et une distance inférieure entre le capillaire et le tissu.
• Echanges alvéolaires d’oxygène La pression partielle d'oxygène dans l'air alvéolaire est d’~100 mmHg tandis que la pression partielle de l'oxygène dans le sang veineux alvéolaire est de ~70 mmHg. La différence de 30 mmHg entraîne l'oxygène de l'air alvéolaire dans le sang où il se lie à l'hémoglobine. L’équilibre de diffusion de l'oxygène entre l'air et le sang alvéolaire est effectué en 0,25 à 0,30 secondes, soit environ un tiers du temps nécessaire pour que le sang se déplace sur toute la longueur du capillaire alvéolaire (~1 s.). La liaison de l'oxygène par l’hémoglobine maintient la différence de concentration et le gradient de diffusion (Fig.3.6).
• Echange alvéolaire de CO2 La pression partielle de CO2 dans l'air alvéolaire est d’~40 mmHg tandis que celle du sang veineux alvéolaire vaut ~46 mmHg. Cette différence de pression est suffisante pour assurer la diffusion du CO2 du sang vers l'air alvéolaire. Bien que le transfert de CO2 capillaire-alvéolaire soit réalisé avec un gradient de diffusion de seulement 6 mmHg, l’équilibre de diffusion est généralement atteint car le CO2 a un coefficient de solubilité supérieur à celui de l’oxygène, ce qui lui permet une diffusion 20 fois plus rapide.
Fig. 3.6. Les valeurs alvéolaires pulmonaires, tissulaires et sanguines des pressions partielles (P) d'oxygène et de dioxyde de carbone et les directions d'échange gazeux
respiratoire. Ppa – pressions partielles alvéolaires; Ppt - pressions partielles tissulaires (iconographie personnelle)
Sang bien oxygéné
Sang faible oxygéné
O2
Tissu Alvéole pulmonaire
Ppa O2 =100 PpaCO2 = 40
Ppt O2 =70 PptCO2 = 46
CO2 CO2
O2
PMD
SAPTD
×
××∆=
PM
S

Physiologie des animaux 13
b- Echanges gazeux tissulaires
L’échange des gaz au niveau des tissus (extra-alvéolaires) consiste au passage de l'oxygène du sang vers les tissus et du CO2 des tissus vers le sang. Cet échange est aussi réalisé par diffusion facilitée. L'oxygène et le dioxyde de carbone diffusent à travers la paroi capillaire dans la direction du gradient de concentration jusqu'à ce que l'équilibre de diffusion soit atteint.
• Échange de l’oxygène tissulaire Le sang artériel pénètre dans les capillaires tissulaires avec une PaO2 d'environ 100 mmHg. La pression partielle d'oxygène dans les tissus varie en fonction de leur taux de consommation dans divers processus métaboliques, avec une moyenne de PO2 à environ 70 mmHg.
• Echange du dioxyde de carbone tissulaire Le sang bien oxygéné pénètre dans les tissus avec une PaCO2 de ~40 mmHg et la pression partielle de CO2 dans les tissus est d'environ 46 mmHg. Par conséquent, un effet de diffusion opposé à l'alvéole se produit.
Tableau 3.3. Pourcentage et pression partielle (mmHg) des gaz dans l’atmosphère sèche (selon W.F. Hofman et D.C. Meyer, Respiratory function in Essentials of human
physiology, 1996)
Nom du gaz Pourcentage Pression partielle du gaz atmosphérique
Pression atmosphérique
totale Azote
Oxygène Dioxyde de carbone Argon + autres gaz
78,0 20,9 0,03 1,07
592,80 158,80
0,23 8,17
760
Pressions partielles et pressions totales des gaz respiratoires dans l'air alvéolaire et les tissus
L'air ambiant inhalé sera saturé de vapeurs d'eau dans les voies nasales et l'arbre trachéobronique. Les vapeurs d'eau créent une pression partielle, tout comme les autres molécules gazeuses dans l'air. À une température corporale de 37°C, la pression partielle des vapeurs d'eau, PH2O, dans l'air inspiré est de 47 mmHg. L'ajout de vapeurs d'eau à l'air inspiré réduit la pression partielle des autres gaz sans modifier la pression totale des gaz. Pour l'air dans l'arbre respiratoire, la pression barométrique (PB) est la somme de la pression des gaz respiratoires et de la pression partielle des vapeurs d'eau:
PB = PN2 + PO2 + PAr2 + PCO2 + PH2O + Pautres gaz

14 R. Bălăceanu, N. Dojană
Les valeurs des pressions partielles des gaz respiratoires dans l'air alvéolaire, le sang veineux, le sang artériel et les tissus animaux sont présentées dans le tableau 3.4.
Tableau 3.4. Pression totale et pression partielle des gaz respiratoires (mmHg) dans le sang, les tissus et l'air alvéolaire chez les mammifères terrestres, au niveau de la mer
(en mmHg, selon W.O. Reece, Physiology of domestic animals. 2nd Edition, Williams & Wilkins, 1996)
Le gaz Dans l'air
atmosphérique Dans l’air alvéolaire
Dans le sang artériel
Dans le sang veineux
O2 CO2 N2 H2O
TOTAL
104 40 569 47 760
40 45 569 47 701
100 40 569 47 756
30 50 569 47 696
L'analyse des données du tableau 3.4 montre ce qui suit: 1. la PO2 alvéolaire (104 mmHg) est inférieure à la PO2 atmosphérique (159 mmHg). Cette différence est due à trois facteurs: a) L’oxygène étant consommé par les tissus, il quitte en permanence de l’alvéole et se diffuse dans le sang; b) l'air alvéolaire est humidifié, l'eau crée un PH2O de 47 mmHg à 37°C, une pression qui dilue l'oxygène, et c) le dioxyde de carbone pénètre toujours l'air alvéolaire, en diluant également l'oxygène. L’augmentation de la ventilation pulmonaire entraîne l’augmen-tation de la PO2 du fait que le remplacement de l’air alvéolaire, relativement appauvri en oxygène, par l’air atmosphérique, s’effectue plus rapidement ; 2. il y a de différence entre la PO2 artérielle et la PO2 alvéolaire causée par des shunts artério-veineux (une partie du sang veineux n'est pas oxygénée); 3. la PCO2 alvéolaire et la PCO2 artérielle sont identiques en raison d'un coefficient de diffusion du dioxyde de carbone plus élevé et d'une différence artério-veineuse plus faible pour le dioxyde de carbone; 4. l'azote est en équilibre dans tout le système car il n’est pas ni produit ni consommé; 8. la pression de vapeur d'eau est la même dans tout le système, les gaz étant saturés avec 100% d'eau; 6. la somme des pressions partielles des gaz sanguins veineux et tissulaires est inférieure à celle de l'air atmosphérique en raison de la consommation d'oxygène, ce qui entraîne la formation du vide observé à l'ouverture chirurgicale de la cavité abdominale. C'est précisément à cause de ce vide que les gaz provenant de différents espaces organiques fermés sont absorbés.
Rapport ventilation-perfusion ( Q/V && )
Le rapport Q/V && peut être défini comme le rapport entre la quantité d'air pénétrant dans les alvéoles (le débit de la ventilation alvéolaire, & , en mL / min)

Physiologie des animaux 15
et la quantité de sang envoyée aux poumons (le débit cardiaque, Q& - également en mL / min). Le calcul du rapport Q/V && est assez simple:
Q/V && = ventilation alvéolaire/débit cardiaque
Q/V && = (≈4 L/min)/( ≈5 L/min) ≈ 0,8
La diminution du rapport (une diminution de la ventilation ou une augmentation de la perfusion) - dans les deux cas, la ventilation ne suit pas le rythme de la perfusion, donc l'oxygène diminuera et le CO2 augmentera dans le sang. Pour produire une augmentation du rapport ventilation-perfusion, deux choses doivent être faites: - augmenter la ventilation (apporter plus d'O2 aux alvéoles, évacuer plus de CO2) ; - diminuez la perfusion (pour que le sang enlève moins d'O2, libère moins de CO2). Cela entraînera une augmentation de la PAO2 (et donc de la PaO2) et une diminution de la PACO2 et de la PaCO2. Chez l'homme, le V& le Q& par gramme de poumon augmentent de l'apex pul-monaire au diaphragme. Comme le taux d'augmentation du Q& est supérieur au taux d'augmentation du V& , le rapport diminue vers la base du poumon, créant un gradient vertical. Il existe peu d'études sur le gradient du rapport Q/V && chez les animaux domestiques. Chez le cheval on a montré un gradient vertical du V& , mais cela s'accompagne d'un gradient similaire du Q& , de sorte que le rapport Q/V && reste inchangé. Dans les situations pathologiques, le rapport peut être modifié, ce qui entraîne une perturbation des échanges pulmonaires de gaz. Le blocage complet des voies respiratoires conduit à un rapport Q/V && égal à zéro et le blocage du flux sanguin pulmonaire conduit à un rapport Q/V && infini. Entre ces limites extrêmes, il peut y avoir des nombreuses possibilités intermédiaires (Fig. 3.7).
Fig. 3.7. Représentation schématique de l’alvéole et du capillaire alvéolaire avec différents effets du rapport Q/V && sur la pression partielle des gaz respiratoires dans le sang quittant
l'alvéole. Le blocage complet des voies respiratoires conduit à un rapport Q/V && égal à zéro (gauche) et le blocage du flux sanguin pulmonaire conduit à un rapport Q/V && infini (droit)
(iconographie personelle)
16 R. Bălăceanu, N. Dojană
5- Transport sanguin des gaz respiratoires
a- Transport sanguin de l’oxygène
L'oxygène est transporté par le sang sous deux formes : libre (dissous dans le plasma) et lié (combiné à l'hémoglobine).
• Forme dissoute du transport d’oxygène La quantité totale d'oxygène dissoute dans le plasma est faible, mais elle augmente proportionnellement à la pression partielle. A PO2 = 100 mmHg, la quantité d'oxygène dissoute dans le plasma est de 0,3 mL/dL de sang. La fraction d’oxygène dissout augmente régulièrement avec l'augmentation de la pression artérielle en oxygène (loi de Henry). Si un homme respire une atmosphère d'oxygène pur, où la PO2 est de 600 mmHg, son sang contiendra 1,8 mL d'oxygène/dL de sang. Bien que petite, cette quantité est importante dans le cas d'une intoxication respiratoire où l'Hb serait bloquée. Environ 98% de l'oxygène est transporté par l'hémoglobine, les 2% restants étant transportés par le plasma.
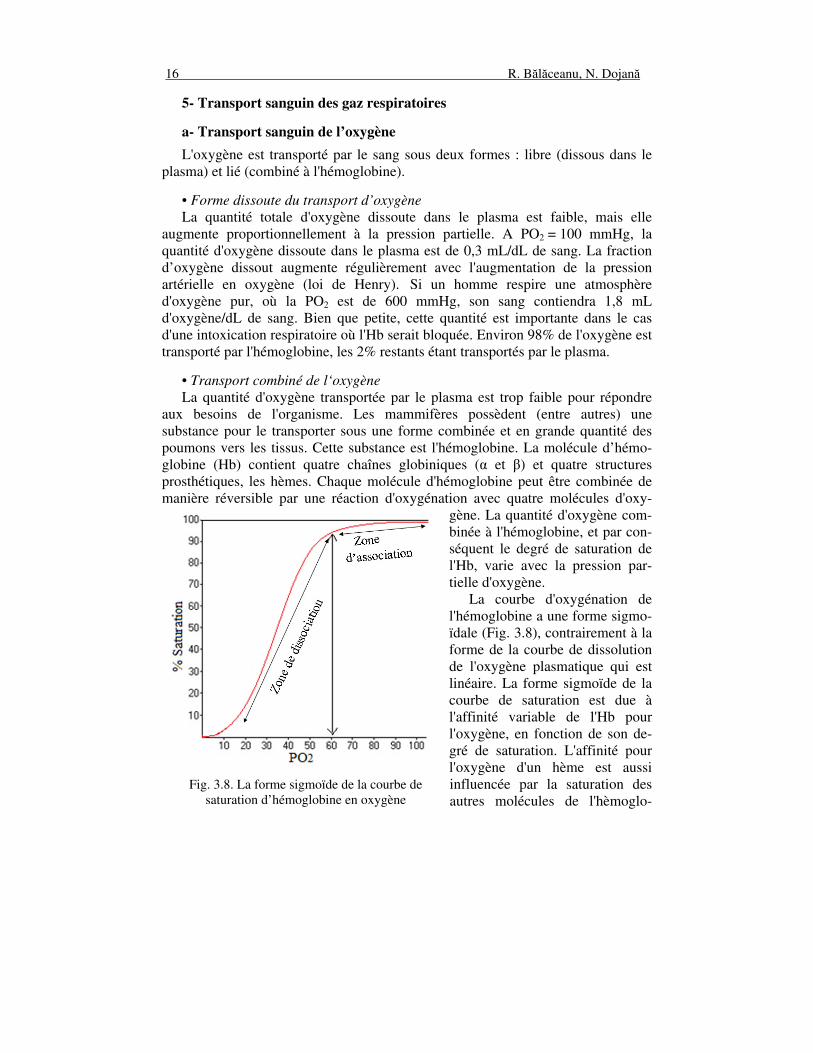
• Transport combiné de l‘oxygène La quantité d'oxygène transportée par le plasma est trop faible pour répondre aux besoins de l'organisme. Les mammifères possèdent (entre autres) une substance pour le transporter sous une forme combinée et en grande quantité des poumons vers les tissus. Cette substance est l'hémoglobine. La molécule d’hémo-globine (Hb) contient quatre chaînes globiniques (α et β) et quatre structures prosthétiques, les hèmes. Chaque molécule d'hémoglobine peut être combinée de manière réversible par une réaction d'oxygénation avec quatre molécules d'oxy-
gène. La quantité d'oxygène com-binée à l'hémoglobine, et par con-séquent le degré de saturation de l'Hb, varie avec la pression par-tielle d'oxygène. La courbe d'oxygénation de l'hémoglobine a une forme sigmo-ïdale (Fig. 3.8), contrairement à la forme de la courbe de dissolution de l'oxygène plasmatique qui est linéaire. La forme sigmoïde de la courbe de saturation est due à l'affinité variable de l'Hb pour l'oxygène, en fonction de son de-gré de saturation. L'affinité pour l'oxygène d'un hème est aussi influencée par la saturation des autres molécules de l'hèmoglo-
Fig. 3.8. La forme sigmoïde de la courbe de saturation d’hémoglobine en oxygène

Physiologie des animaux 17
bine. La liaison de l’oxygène se fait donc en quatre étapes, une par monomère. La saturation en oxygène est progressive, combinant d'abord le premier hème puis le second et ainsi de suite. La combinaison de l'oxygène avec le premier hème augmente l'affinité du second pour l'oxygène et ainsi de suite, de sorte que le quatrième hème a une affinité 125 fois plus grande que le premier. Ces interactions sont responsables de l'aspect sigmoïde de la courbe de dissociation de l'oxyhémoglobine (hémoglobine saturée d’oxygène). La quantité d'oxygène combinée à l'hémoglobine est influencée par la PO2 dans l'air alvéolaire et dans le sang. À des valeurs de PO2 d’environ 70 mmHg, l'hémo-globine est déjà saturée d'oxygène (environ 100%). L'augmentation du PO2 au dessous de 70 mmHg de PO2 détermine la combinaison que d'une faible quantité d'hémoglobine avec l'oxygène. La capacité de saturation à des valeurs d’environ 70 mmHg permet à l'hémoglobine se saturer en oxygène même dans les conditions de déplacement des animaux à des altitudes modérées. Sous 60 mmHg de PO2, la courbe de dissociation de l'hémoglobine présente une forte pente. Une diminution de la PO2 inférieure à 60 mmHg est obtenue dans les tissus, permettant ainsi à l’oxygène se détacher de l’hémoglobine. Un gramme d'hémoglobine peut lier jusqu’à 1,39 mL d'oxygène. Ainsi, 1 dL de sang contenant 15 g d'hémoglobine peut contenir environ 20 mL d'oxygène. La quantité d'oxygène pouvant être liée à l'hémoglobine est appelée capacité d'oxygénation du sang.
b- Facteurs influençant l'affinité de l'hémoglobine pour l'oxygène
L'affinité de l'hémoglobine pour l'oxygène, c'est-à-dire la forme de la courbe de dissociation de l'hémoglobine, est affectée par les principaux facteurs suivants : - la température corporelle - le pH tissulaire - la concentration de dioxyde de carbone dans le sang - les phosphates organiques des érythrocytes - l’espèce - l’altitude géographyque.
• Température L'augmentation de la tempé-rature du sang diminue l'affinité de l'hémoglobine pour l'O2, ce qui augmente la P50 (déplace la courbe de dissociation de l'oxyhémoglobine vers la droite). Une augmentation de la température du sang accompagne généralement un taux métabolique
Fig. 3.8. Effect de différentes concentrations en dioxyde de carbone sur la courbe de saturation
d’hémoglobine en oxygène chez le cheval (selon Bohr, Hasselbalch et Krogh, 1904)
18 R. Bălăceanu, N. Dojană
élevé et une forte demande en oxygène, favorisant ainssi la libération d'oxygène de l'oxyhémoglobine.
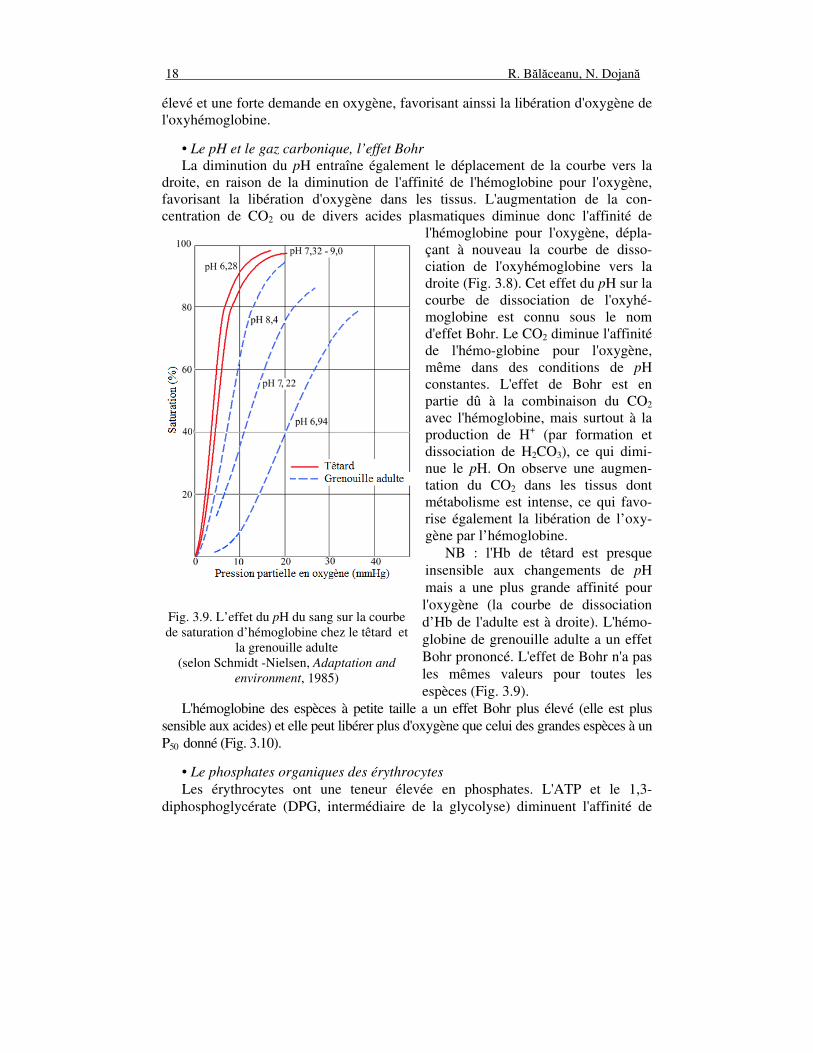
• Le pH et le gaz carbonique, l’effet Bohr La diminution du pH entraîne également le déplacement de la courbe vers la droite, en raison de la diminution de l'affinité de l'hémoglobine pour l'oxygène, favorisant la libération d'oxygène dans les tissus. L'augmentation de la con-centration de CO2 ou de divers acides plasmatiques diminue donc l'affinité de
l'hémoglobine pour l'oxygène, dépla-çant à nouveau la courbe de disso-ciation de l'oxyhémoglobine vers la droite (Fig. 3.8). Cet effet du pH sur la courbe de dissociation de l'oxyhé-moglobine est connu sous le nom d'effet Bohr. Le CO2 diminue l'affinité de l'hémo-globine pour l'oxygène, même dans des conditions de pH constantes. L'effet de Bohr est en partie dû à la combinaison du CO2 avec l'hémoglobine, mais surtout à la production de H+ (par formation et dissociation de H2CO3), ce qui dimi-nue le pH. On observe une augmen-tation du CO2 dans les tissus dont métabolisme est intense, ce qui favo-rise également la libération de l’oxy-gène par l’hémoglobine. NB : l'Hb de têtard est presque insensible aux changements de pH mais a une plus grande affinité pour l'oxygène (la courbe de dissociation d’Hb de l'adulte est à droite). L'hémo-globine de grenouille adulte a un effet Bohr prononcé. L'effet de Bohr n'a pas les mêmes valeurs pour toutes les espèces (Fig. 3.9).
L'hémoglobine des espèces à petite taille a un effet Bohr plus élevé (elle est plus sensible aux acides) et elle peut libérer plus d'oxygène que celui des grandes espèces à un P50 donné (Fig. 3.10).
• Le phosphates organiques des érythrocytes Les érythrocytes ont une teneur élevée en phosphates. L'ATP et le 1,3-diphosphoglycérate (DPG, intermédiaire de la glycolyse) diminuent l'affinité de
Fig. 3.9. L’effet du pH du sang sur la courbe de saturation d’hémoglobine chez le têtard et
la grenouille adulte (selon Schmidt -Nielsen, Adaptation and
environment, 1985)

Physiologie des animaux 19
l'hémoglobine pour l’O2 en dépla-çant la courbe de dissociation de l'hémoglobine vers la droite (aug-mente P50), favorisant la libération d'oxygène. Le DPG régule la combinaison de l'hémoglobine avec l'oxygène. La concentration de DPG est ainsi plus élevée chez les personnes vivant en haute altitude que celles vivant au niveau de la mer. L’hémo-golbine du sang foetal a également une concentration plus élevée de DPG favorisant la libération d'O2 dans les tissus du foetus.
• Taille de l’espèce Chez les petits mammifères, la courbe de dissociation de l'oxyhémoglobine est décalée vers la droite (moins d'affinité pour O2) que chez les espèces plus grandes. Avantage : pour la même pression partielle d'oxygène, une plus grande quantité d'oxygène est libérée que chez les grands mammi-fères. La différence de pression partielle d'O2 est déterminée par le taux métabolique: les petits animaux ont un taux métabolique plus élevé que les grandes espèces. Les petites espèces ont une consommation spéci-fique d'oxygène plus élevée, ce qui est également assurée par ce mécanisme (une affinité inférieure est compensée au niveau pulmonaire par la pression partielle d'oxygène par rapport aux besoins).
• Altitude Une faible pression atmosphérique à haute altitude entraîne une pression d'oxygène atmosphérique inférieure à celle de plus basse altitude comme au
Fig. 3.10. Rélation masse corporelle - effet Bohr (selon Schmidt -Nielsen, Adaptation and
environment, 1985)
Fig. 3.11. Les courbes de dissociation d'oxyhémoglobine de llama (Lama guanico) et de vicuna (Vicogna) sont situées à gauche de
celles d'autres espèces de mammifères (la zone hachurée). L'affinité plus élevée du sang de ces
animaux aide à prendre de l'oxygène à faible pression atmosphérique
(selon Schmidt -Nielsen, Adaptation and environnement, 1985)

20 R. Bălăceanu, N. Dojană
niveau de la mer. Les animaux acclimatés à un environnement pauvre en oxygène possèdent une hémoglobine avec une affinité pour l'oxygène plus élevée (courbe de dissociation décalée vers la gauche, Fig. 3.11). L'affinité plus élevée du sang de ces animaux aide à prélever l'oxygène à faible pression atmosphérique. Le déplacement de l’homme en haute altitude est suivi d’un processus d'adap-tation, mais cette adaptation n'inclut pas l'augmentation de l'affinité de l'hémoglo-ine pour l'oxygène. Paradoxalement, par adaptaion aux hautes altitudes, la courbe de dissociation de l'hémoglobine chez l'homme se déplace vers la droite. L’explication est celle de l’augmentation du contenu en DPG dans les érythrocytes NB: Le DPG diminue l'affinité de l‘Hb pour l'oxygène, favorisant sa libération dans les tissus.
c- Transport sanguin du dioxyde de carbone
Reprise du dioxyde de carbone des tissus
Le dioxyde de carbone diffuse des tissus dans le sang grâce à une pression partielle élevée au niveau des tissus. Il est transporté par le sang sous deux formes: libre et combinée. Environ 5% du dioxyde de carbone qui pénètre dans le sang est transporté dissous dans le plasma (ce qui représente un total de 2,7 mL de dioxyde de carbone transporté par 1 dL de plasma). La plupart du dioxyde de carbone diffuse dans les érythrocytes où environ 63% est combinée avec de l’eau (réaction d’hydratation) pour former de l’acide carbonique (la première forme combinée de transport du dioxyde de carbone), qui se dissocie:
H2O + CO2 →← H2CO3 →← H+ + −
3HCO
Cette réaction se produit également dans le plasma avec 5% du dioxyde de carbone total diffusé, mais dans les érythrocytes, la présence d’anhydrase carbonique accélère l’hydratation du dioxyde de carbone plusieurs centaines de fois. Les ions H+ résultant de la réaction d'hydratation sont tamponnés par l'hémoglobine, H+ remplaçant K+ de sa structure. K+ ions libérés se combinent avec
−
3HCO . La pluart (70%) du −
3HCO diffuse à travers la membrane cellulaire vers
l'extérieur, en échange d'une pénétration de Cl¯ dans la cellule pour maintenir l'équilibre éléctrique de la membrane de Gibbs - Donnan. La pénétration de Cl¯ dans les globules rouges augmente leur osmolarité, entraînant une pénétration de l'eau plasmatique, augmentant leur volume et augmentant de 3% l'hématocrite veineux par rapport au sang artériel. La deuxème forme combinée de transport du dioxyde de carbone résultent de la réaction du dioxyde de carbone avec les groupes −NH2 de protéines, en particulier l'hémoglobine, mais également les protéines plasmatiques. Ces sont l’hémoglobine carbaminée et des protéines carbaminées qui resultent. Les com-posés carbaminés sont responsables du transport de 20 à 30% du dioxyde de carbone total.

Physiologie des animaux 21
Phénomène de Hamburger Dans l'alvéole pulmonaire, Cl¯ migre dans la direction opposée à celle tissulaire, quittant l'hématie. La migration de Cl¯ à travers la membrane de l'hématite vers l'extérieur au niveau des poumons et vers l'intérieur dans les tissus est appelée phénomène de Hamburger.
Libération de dioxyde de carbone dans les poumons, l’effet Haldane Lorsque le sang veineux atteint les capillaires pulmonaires, le dioxyde de carbone diffuse du sang dans l'air alvéolaire, inversant ainsi le sens des réactions, grâce à sa pression partielle diminuée au niveau des alvéoles. L'oxygénation du sang au niveau des alvéoles pulmonaires entraîne une baisse de l'affinité de l'Hb pour le dioxyde de carbone et donc une accélération de sa libération. L'effet Haldane et la propriété d’Hb à perdre l'affinité pour le dioxyde de carbone quand la pression partielle d’oxygène augmente dans le sang (Fig. 3.12). Cette situation se rencontre dans les alvéoles pulmonaires, favorisant la libération de dioxyde de carbone par l'Hb. À mesure que le sang s'oxygène, l'Hb perd son affinité pour le dioxyde de carbone, ce qui favorise sa libération.
6- Contrôle de la respiration Le contrôle de la respiration doit remplir deux tâches importantes :
- etablir/maintenir une rythmicité automatique des mouvements respiratoires, rythmogenèse ;
- ajuster ce rythme aux demandes : métabolique (gaz du sang, pH) /mécanique (posture) /comportements épisodiques non-ventilatoires (parler, manger), adapta-tion physiologique. Les facteurs de régulation respiratoire sont :
- mécaniques : degré d'étirement des voies aériennes, des alvéoles et de la cage thoracique.
- chimiques (humoraux), représentés par la concentration sanguine de dioxyde de carbone, oxygène et ions d’hydrogène (le pH).
Fig. 3.12. L'effet Haldane : l'évolution de la capacité de
transport de dioxyde de carbone par plasma sanguin est une fonction de la pression partielle d'oxygène. Le
sang faiblement oxygéné a une capacité de transport de dioxyde de carbone plus élevée que le sang bien
oxygéné. Entre les limites physiologiques, la courbe a un
aspect rectiligne (selon Cunningham, 1992)
22 R. Bălăceanu, N. Dojană
a- L’automatisme de la respiration
•Récepteurs des voies aériennes Les récepteurs sensitifs des voies aériennes se trouvent dans différentes parties du système respiratoire (tableau 3.5).
Tableau 3.5. Types de récepteurs mécaniques des voies respiratoires
• Les chémorécepteurs Les chémorécepteurs contribuent au contrôle dit humoral de la respiration. Le terme « humoral » se réfère uniquement à la nature des excitateurs physiologiques, représentés par P(O2), P(CO2) et [H+] situés dans le sang ou le liquide interstitiel. Le niveau sanguin ou interstitiel de ces substances est monitorisés par des struc-tures spéciales appelées chémorécepteurs. Ces structures spéciales sont des seg-ments périphériques d'arcs réflexes (réflexes chémorécepteurs). Les chémorécep-teurs ont une localisation périphérique et une localisation centrale. Les chémoré-cepteurs périphériques sont situés dans les corpuscules carotidiens et l'arc aortique.
Récepteurs Localisation Sensibilité à Afférence Effets
Tensiorécepteurs (mécanorécepteur
particulier) à adaptation lente
Muscles lisses bronchique s
L’étirement des voies aériennes
et à la distension
pulmonaire
Nerf vague, tronc cérébral
Allongement du temps inspiratoire
(réflexe de Hering-Breuer)
Tensiorécepteurs à adaptation rapide
Muscles lisses bronchiques
L’étirement des voies aériennes
et à la distension
pulmonaire
Nerf vague, tronc cérébral
Inhibition de l’expiration
Récepteurs à l’irritation
L’épithélium bronchique
Irritation Nerf vague,
tronc cérébral
Hyperpnée, bronchoconstricti
on, toux
Récepteurs alvéolaires
juxtacapillaires (récepteurs J)
Paroi alvéolaire, près des capillaires
Dilatation des capillaires,
augmentation de la pression interstitielle
Nerf vague (fibres C
amyéliniques), tronc
cérébral
Ventilation rapide et superficielle; apnée lors de stimulation importante
Récepteurs proprioceptifs de la
paroi thoracique
Paroi, articulations, tendons
thoraciques, fuseaux
neuromusculaires
Activité musculaire respiratoire
Nerf phrénique, tronc cérébral
Modulation de la ventilation

Physiologie des animaux 23
Ils surveillent la concentration sanguine de l’O2, CO2 et H+. Les chémorécepteurs centraux ont les charactéristiques suivantes : - ils sont situés à la surface ventrale du bulbe - ils sont stimulés par les ions H+ (PCO2) présent dans le LCR qui les baigne - ils ne sont pas sensibles à la PO2 - ils stimulent les neurones inspiratoires et entraînent une hyperventilation Ni l’anémie, ni l’empoisonnement au monoxyde de carbone n’entraînent une hyperventilation. La baisse du pH et la hausse de la concentration de CO2 provoquent une hyperventilation.
• Les centres nerveux des arcs respiratoires Ils sont placés dans le bulbe, dans le pont et au-dessus du pont. Il existe quatre centres respiratoires (Fig. 3.13): - le centre pneumotaxique, situé dans le pont; - le centre apnéustique, également situé dans le pont; - le groupe respiratoire dorsal, situé dans le bulbe rachidien; - le groupe respiratoire ventral, également situé dans le bulbe rachidien.
Les neurones du centre pneumotaxique inhibent les neurones inspiratoires du centre apnéustique et les neurones inspiratoires du GRD et GRV. Ils provoquent la fin de l'inspiration (centre off-switch). Les neurones du centre apnéustique stimulent les neurones inspiratoires en induisant une inspiration prolongée (apnéuse). Le GRV contient deux noyaux distincts, chacun contenant des neurones inspiratoiress et des neurones expiratoires. Les neurones inspiratoires sont projetés sur les motoneurones spinaux du diaphragme et des intercostaux externes (muscles
Fig. 3.13. Centres nerveux respiratoires (gauche) et l’effet des transsections totales sur la respiration (droit), dans des conditions de nerfs vagues intacts et coupés. 1. 2. 3. 4- les
zones où les transsections du tronc cérébral ont été effectuées (selon Cunningham, 1992)

24 R. Bălăceanu, N. Dojană
inspiratoires). Les neurones expiratoires stimulent les motoneurones spinaux des muscles expiratoires, intercostaux internes et abdominaux. Le GRD ne contient que des neurones inspiratoires.
b- Réflexe respiratoire Hering-Breuer
L'act réflexe de respirer (inspiration + expiration) a été décrit par Hering et Breuer en 1868 et porte leurs noms. On parle d’un réflexe inhibiteur de l'inspiration d'Hering-Breuer. Le réflexe est basé sur un automatisme interne des neurones inspiratoires, automatisme qui est influencé par un certain nombre de facteurs. Les neurones inspiratatoires envoient automatiquement des impulsions stimula-trices vers les motoneurones médullaires qui stimulent les muscles inspiratoires (diaphragme et intercostaux externes). Ces neurones déterminent la contraction des muscles inspirateurs et l'expansion de la poitrine. Avec l'expansion de la poitrine, au cours de l'inspiration, les récepteurs du muscle lisse trachéobronchique et les voies intrapulmonaires sont stimulés. L’information sensorielle pulmonaire est envoyée par les nerfs vagues. Celle qui vient de la parois de la cage thoracique est envoyée par les nerfs sensitives somatiques. Elle stimule les neurones des centres pneumotaxiques et inhibe les neurones inspirateurs du GRV et GRD en inhibant l'inspiration. Lorsque l'inspiration est terminée, l'expiration apparaît comme un acte passif, comme résultat de la relaxation des fibres élastiques dans la structure des poumons et de la paroi thoracique. La dernière phase de l'expiration est assistée par la contraction des muscles expiratoires.
7- Particularités de la respiration chez le fœtus
On peut subdiviser le développement intra-utérin en trois étapes: zygote, embryon et foetus1. Le foetus a une circulation sanguine fonctionnelle et une respiration placentaire.
a- Saturation et PO2 du sang foetal
Le sang fœtal a une faible PO2 car le placenta est un échangeur de gaz respiratoire peu efficace, et le sang bien oxygèné et celui à faibe teneur en oxygène
1 Le stade zygote est la période de développement de l'amphimixie à l'implantation. A ce stade, le processus de segmentation a lieu en passant par les stades de deux, quatre, huit, 16 cellules, la morule, le blastocyste et la gastrule. Le stade embryonnaire est la période de développement qui commence par l’esquisse des organes primordiaux et se termine par l’achèvement des caractères morphologiques spécifiques et qui dure de la nidation à la formation du placenta. Au cours de cette période, des processus d’organogenèse et la formation des annexes fœtales se produisent. Le stade fœtal dure de la formation du placenta jusqu’à la parturition. Cette étape est caractérisée par la croissance et la différenciation de tous les tissus et organes du fœtus.
Physiologie des animaux 25
sont mélangés à plusieurs endroits de la circulation fœtale : foie, cœur et poumons. Le fœtus est adapté à cet état d'hypoxie chronique de deux manières. Première-ment, le cœur du fœtus a un débit élevé, ce qui permet un afflux important de sang dans les tissus systémiques. Deuxièmement, le fœtus produit des érythrocytes contenant une Hb dont affinité pour l’oxygène est supérieure à celle de l’animal adulte. La pression partielle de l'oxygène dans le sang fœtal faible oxygénée, à l’arrivée au placenta, est d'environ 22 mmHg (saturation à 56%). À la sortie du placenta (veines ombilicales), le sang bien oxygéné a une PO2 d’environ 30 mmHg (saturation à 80%).
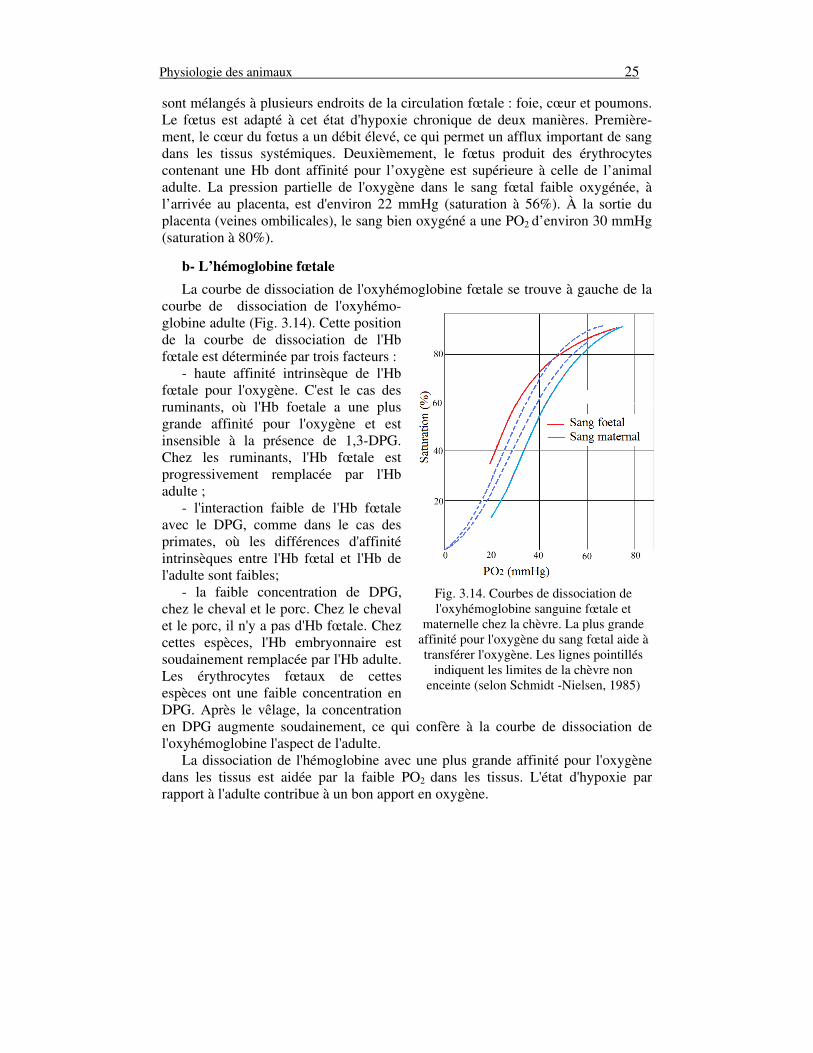
b- L’hémoglobine fœtale
La courbe de dissociation de l'oxyhémoglobine fœtale se trouve à gauche de la courbe de dissociation de l'oxyhémo-globine adulte (Fig. 3.14). Cette position de la courbe de dissociation de l'Hb fœtale est déterminée par trois facteurs : - haute affinité intrinsèque de l'Hb fœtale pour l'oxygène. C'est le cas des ruminants, où l'Hb foetale a une plus grande affinité pour l'oxygène et est insensible à la présence de 1,3-DPG. Chez les ruminants, l'Hb fœtale est progressivement remplacée par l'Hb adulte ; - l'interaction faible de l'Hb fœtale avec le DPG, comme dans le cas des primates, où les différences d'affinité intrinsèques entre l'Hb fœtal et l'Hb de l'adulte sont faibles; - la faible concentration de DPG, chez le cheval et le porc. Chez le cheval et le porc, il n'y a pas d'Hb fœtale. Chez cettes espèces, l'Hb embryonnaire est soudainement remplacée par l'Hb adulte. Les érythrocytes fœtaux de cettes espèces ont une faible concentration en DPG. Après le vêlage, la concentration en DPG augmente soudainement, ce qui confère à la courbe de dissociation de l'oxyhémoglobine l'aspect de l'adulte. La dissociation de l'hémoglobine avec une plus grande affinité pour l'oxygène dans les tissus est aidée par la faible PO2 dans les tissus. L'état d'hypoxie par rapport à l'adulte contribue à un bon apport en oxygène.
Fig. 3.14. Courbes de dissociation de l'oxyhémoglobine sanguine fœtale et
maternelle chez la chèvre. La plus grande affinité pour l'oxygène du sang fœtal aide à transférer l'oxygène. Les lignes pointillés
indiquent les limites de la chèvre non enceinte (selon Schmidt -Nielsen, 1985)

26 R. Bălăceanu, N. Dojană
8. Particularités de la respiration chez les oiseaux
a- Mécanique respiratoire
Comme les oiseaux ne disposent pas du diaphragme et que la cavité thoracique communique avec la cavité abdominale, tout le volume de la cavité thoraco-abdominale change tout au long du cycle respiratoire. Les oiseaux ventilent leurs poumons non pas par l’expansion des poumons eux-mêmes mais par l’expansion des sacs aériens. Les changements de volume sont dus à la contraction des muscles squelettiques. À l'expiration, les muscles se contractent, entraînant une diminution du corps. La diminution du volume corporel augmente la pression dans les sacs aériens, forçant l'air à s'écouler à travers les poumons et, par conséquent, dans l'environnement extérieur. L'inspiration est un effet de la relaxation des muscles du corps, relaxation qui entraîne l'augmentation du volume corporel. L'augmentation du volume corporel est suivie d'une diminution de la pression sur les sacs aériens, de l'expansion des sacs aériens et d'une diminution de leur pression. En consé-quence, l'air atmosphérique passe par les voies aériens vers les sacs aériens postérieurs (thoracique caudal et abdominal) et l’air des poumons passe dans les sacs aériens antérieurs (cervical, claviculaire, thoracique crânial).
b- Cycle respiratoire
Au cours de l'inspiration, la majeure partie de l'air inspiré est dirigée direc-tement dans les sacs aériens caudaux (figure 3.15). Bien que le volume des sacs aériens crâniens augmente en inspiration, ils ne reçoivent pas d’air inspiré; Au lieu de cela, ces sacs reçoivent l'air du poumon. Lors de l'expiration, l'air des sacs caudaux est dirigé dans les poumons au lieu d'être éliminé par la bronche principale. À l'inspiration suivante, l'air des poumons est entraîné dans les sacs
Fig. 3.15. Mouvement d'un bol d'air le long du système respiratoire chez les oiseaux. 1 - la bronche principale; 2 – sacs aériens caudaux; 3 – poumons; 4 - sacs aériens
crâniens (selon S c h m i d t - N i e l s e n , 1983)

Physiologie des animaux 27
crâniens. Dans l’expiration suivante, l'air des sacs crâniens est éliminé à l'extérieur. De cette façon, chez les oiseaux, deux cycles respiratoires sont nécessaires pour déplacer un seul bol d'air dans le système respiratoire. Comme décrit ci-dessus, il y a deux des caractéristiques de la mécanique respiratoire chez les oiseaux: 1) la direction unique de la circulation de l'air à travers des poumons, toujours caudo-crânienne (Fig. 3.16), et 2) le mouvement de l'air dans les poumons à la foie en inspiration et expiration (Schmid t -Nie l sen , 1983).
c- Ventilation pulmonaire
La ventilation pulmonaire chez les oiseaux est plus intense que chez les mammifères, à la fois en inspiration et expiration, ce qui rapproche la composition de l'air pulmonaire de l'air atmosphérique, en créant un gradient de diffusion plus élevé et en favorisant les changements respiratoires. En hypoxie, les poumons sont hyperventilés, ce qui entraîne une diminution de la PCO2 artérielle à des valeurs très basses: inférieures à 8 mmHg. Ainsi, dans des conditions de vol en haute altitude (hypoxie), les oiseaux sont sujets à l'alcalose, une condition qu'elles suppor-tent mieux que les mammifères. Contrairement aux mammifères, où la ventilation est incomplète, alors qu’un volume d’air résiduel reste dans les poumons la fin d’un cycle respiratoire, chez les oiseaux la ventilation est complète et, lors d’un le cycle respiratoire, tout la quantité d'air des poumons est remplacée par l’air frais.
d- Échanges gazeux entre les bronchioles et capillaires sanguins
Le sens du flux sanguin à travers les capillaires sanguins pulmonaires est croisé (perpendiculairement) à celui de l'air, ce qui favorise les échanges gazeux (Fig. 3.17). La sens de la circulation du sang et le mouvement continu de l'air à travers
Fig. 3.16. Illustration de la direction unique du transport aérien à travers des parabronches paléopulmonaires, à la fois dans l'inspiration (→) et expiration (→)

28 R. Bălăceanu, N. Dojană
les poumons (en inspiration et en expiration) permettent au sang qui passe dans les capillaires pulmonaires d'avoir une PO2 croissante et une PCO2 en diminution constante. Le sang sortant du poumon fait encore des échanges gazeux avec l'air qui vient d'entrer dans le poumon, air qui présente une PO2 élevé. A la mesure que cet air traverse les poumons, il échange des gaz avec un sang qui content moins d'oxygène, ce qui permet de continuer à donner plus d'oxygène au sang. Cela trouve son utilité dans des conditions de vol en altitude, où la ventilation est
renforcée par l'hypoxie. Dans ces conditions, la PO2 dans le sang n'a que quelques mmHg de moins que la PO2 atmosphérique. En outre, dans le sang artériel, la pression partielle du dioxyde de carbone étant plus basse et le pH plus élevé que chez les mammifères, les changements respiratoires sont également favorisées (tableau 3.6). Cela favorise la respiration à haute altitude où la pression barométrique est réduite.
Tableau 3.6. Valeurs des pressions partielles de gaz respiratoires et du pH sanguin chez les oiseaux (valeurs chez l'animal au repos, selon Reece, 1996)
Espèce PO2 (mmHg) PCO2 (mmHg) pH
Oie Canard (selon K a w a s h i r o , 1975) Canard (selon C a l d e r , 1968) Poule Leghorn Colombe (selon C a l d e r , 1968) Colombe (selon B e r n s t e i n , 1981)
- 82 -
82 - -
24,5 38 29,9 33 28,5 25,0
7,53 7,49 7,50 7,52 7,52 7,45
e- Transport des gaz respiratoires Le pourcentage de saturation en oxygène de l'hémoglobine dans le sang est plus faible chez les oiseaux que chez les mammifères: environ 90% par rapport de 97,5%. La même situation se retrouve dans le sang veineux: 40% à 72%. La PCO2 dans le sang est aussi plus basse chez les oiseaux que chez les mammifères: 28 – 34 mmHg versus 40 – 45 mmHg (R e e c e , 1996).
Fig. 3.17. Schéma du système d'échange de gaz à contre-
courant du poumon de l'oiseau. V est la direction du mouvement de l'air, et Q c'est la direction du mouvement du sang. Les flèches épaisses indiquent la direction du
mouvement de l'air (selon Reece, 1996)

Physiologie des animaux 29
Bibliographie
Akaevsky, A.I., D.I.A. Krinitsyn – Fizialoghia selîskahaziaistvenîh jîvotnîh c asnovami anatomii. Moscwa, 1958.
Allard, J. – Le générateur spinal de l’éjaculation. Correspondances en pelvi–perineologie, - n° 2, vol. V -avril/mai/juin 2005, université de Cincinnati, Collège de Médecine, 2015.
Bachmanov A.A., Beauchamp G.K. – Taste receptor genes. Annu. Rev. Nutr. 2007;27:389-414.
Barone, R. – Anatomie comparée des mammifères domestiques, Lyon, 1976. Berg, J., M. S. Tymoczko – Biochemistry (5th ed.), New York, 2002. Brugère, H. – Physiologie de l’appareil urinaire. École Nationale Vétérinaire d’Alfort, France, 2001. Brugère, H. – Régulations métaboliques (Cours et travaux dirigés). École Nationale
Vétèrinaire d’Alfort, France, 2001. Brugère, H. – Système nerveux, École Nationale Vétèrinaire d’Alfort, France, 2002. Bullock, T.H., M.V.L. Bennett, D. Johnston, R. Josephson, E. Marder, R. Fields – The
Neuron Doctrine, Science, V.310, p. 791-793, 2005. Chauveau, A., S. Arlong – Traité d’Anatomie Comparé des Animaux Domestiques.
Cinquième Edition, Libraire J.-B. Maillière et Fils, Paris, 1905. Chirent, Mylène. – Les alternatives à la castration chirurgicale chez l’étalon, Thèse,
Université Claude-Bbernard – Lyon, 2015. Chauveau, A., S. Arlong – Traité d’Anatomie Comparé des Animaux Domestiques.
Cinquième Edition, Libraire J.-B. Maillière et Fils, Paris, 1905. Clarkson, T. – Handbook of biological data, 1956. Clermont, Y., M. Lalli et Z. Bencsath-Makkai, Atlas d’histologie en miroscopie optique, http://audilab.bmed.mcgill.ca/HA/html/resp_21_F.html Cotru ţ , M. – Fiziologie, Cours lito, Iaşi, 1992. Cunningham, J.G. – Textbook of veterinary physiology. Editura W. B. Saunders, U.S.A., 1992. Dacheux, F.– La reproduction chez les mammifères et l’homme – I.N.R.A. Editions
Ellipses, Paris, 2001. Dixon, M. – The enzymes, Longmans Ed., London, 1976. Dojană, N. – Tratat de fiziologia animalelor domestice. Editura Academiei, 2009. Dogui, M. – Physiologie humaine. ISBN 979-10-262-4403-5, Librinova. Ginsburg, J.M., J.L. Börke – Renal physiology în Essentials of human physiology, 1996. Ginsburg, J.M., A. Costoff – Gastrointestinal physiology în Essentials of human
Physiology, 1996. Godth, R.E. – Muscle physiology în Essentials of human physiology, 1996. Granciu, I., S. Duica, I. Cureu – Grupele sanguine la animalele domestice (Groupes san-guins chez les animaux domestiques) Editura Ceres, Bucureşti, 1973. Gray, H. – Gray’s Anatomy, John W. Parker & son Ed., Finland, 1997. Guémené, D., N. Kansaku, D. Zadworny – L'expression du comportement d'incubation
chez la dinde et sa maîtrise en élevage. I.N.R.A. Prod. Anim., 14, p. 147-160, 2001. Guyton, C. – Fiziologie. Fiziologie umană şi mecanismele bolilor. 5th Edition. Ediţia în lim-
ba română sub redacţia R. Cârmaciu. Editura Medicală Amaltea, WB Saunders, 1997. Kolb, E. – Physiologie des animaux domestiques, Vigot Frères Èd., Paris, 1973. Leung E, Tasker SL, Atkinson L, Vaillancourt T, Schulkin J, Schmidt LA. – Perceived
maternal stress during pregnancy and its relation to infant stress reactivity at 2 days and

30 R. Bălăceanu, N. Dojană
10 months of postnatal life. Clin Pediatr. 2010;49:158–165. doi: 10.1177 /0009922809346570. [PubMed] [CrossRef] [Google Scholar].
Hăulică, I. – Fiziologie. Editura Medicală, Bucureşti, 1996. Hofman, W.F., D.C. Meyer – Respiratory physiology in Essentials of human physiology,
1999. Houpt, K. – Domestic Animal Behavior for Veterinarians and Animal Scientists. Iowa State
Press, Ames, Iowa, 2005. Jarrige, R. – Ruminant nutrition: recommended allowances and feed tableaus. ISBN 0-
86196-247-8, INRA, Paris, 1989. JoHnson, N., W.T. Everitt – Essentials of Reproduction, Black-well Scientific Publications,
Oxford, 1996. Kandel E.R., J.H. Schwartz, T.M. Jessell – Principles of Neural Science, 4th ed., McGraw-
Hill, New York, 2000. Hafez, E.S.E. –Reproduction and Breeding Techniques for Laboratory Animals, Lea &
Febiger Ed., Philadelphia, 1970. Hafez, E.S.E. – Reproduction in farm animals. 6th Edition, Lea & Febiger Ed., 1993. H o f m a n W.F. et D.C. M e y e r , Respiratory physiology dans Essentials of human
physiology, Georgia, 1996. Jammes, H., J. Djiane – Le développement de la glande mammaire et son contrôle hormonal
dans l’espèce bovine. INRA Productions animales, 1 (5), pp.299-310, 1988. Jausein, 2005 – La glande mammaire et sa cancérisation. I.F.É., http://mammary.nih.gov
/reviews/development/Development001/index.html Kolb, E. Physiologie des animaux domestiques. Vigot Frères Ed., 1965. Knobil et al. – Androgen action and the sex accessory tissues in The physiology of
reproduction, Nez York, Raven, 1988:1349–1377, 1988. Mader, S.S. – Biology, 4th ed, Ed. Wm.C. Brown Publishers, Dubuque, IA, USA, 1993. Mallarmé, J. – Hématologie de laboratoire. Données et techniques. Editions Mont
Chrétien, Paris, 1964. Meyer C., ed. sc., 2019, Dictionnaire des Sciences Animales. [On line]. Montpellier,
France, Cirad. [17/09/2019]. <URL : http://dico-sciences-animales.cirad.fr/. Mombay, B.M., D.M. Bishop – A.C.T.H. in diagnosis of adrenal insufficiency (Thorn test).
British Medical Journal, 3, 1953. Müller, M.J., Seitz, H.J. – Thyroid hormone action on intermediary metabolism. Part III.
Protein metabolism in hyper- and hypothyroidism. Klin. Wochenschr. 1;62(3):97-102, 1984.
Ogle, T.F., A. Costoff – Endocrinology dans Essentials of human physiology, 1996. Ognean, L., N. Dojana, Corina Rosioru – Fiziologia animalelor. Editura Presa Universitară
Clujeană, Cluj-Napoca, 2000. Orgebin, Marie-Claire – Étude du transit épididimaire des spermatosïdes de taureau
marqués a l’aide du 32P. Annales de biologie animale, biochimie, biophysique, 1961, 1 (2), pp. 117-120. Thèse pour le Doctorat vétérinaire, 1987, École Nationale Vétérinaire d’Alfort, 1961.
Rajaofetra, N., j.g. Passagia, l. Marlier – Serotoninergic, noradrenergic and peptidergic innervation of Onuf's nucleus of normal and transected spinal cords of baboons (Papio papio). J. Comp. Neurol. 318 (1): 1–17, 1992.
Ramón y Cajal, S. – Histology, 10th ed., Wood, Baltimore, 1933.

Physiologie des animaux 31
Reece, W.O. – Physiology of domestic animals. 2nd Edition, Williams & Wilkins Ed., 1996.
Reece, W.O. – Physiology of domestic animals. Lea & Febiger, Philadelphia, 1991. Reece, W.O. – Physiology of domestic animals. 2nd Edition, Williams & Wilkins Ed.,
1996. Roberts, A., B.M.H. Bush – Neurons Without Impulses. Cambridge University Press,
Cambridge, 1981. Pavlov, I. P. – Conditioned reflexes. London: Routledge and Kegan Paul, 1927. Rouvière, H. – Précis d’anatomie et de dissection. Collection de précis médicaux, Masson
& Cie Editeurs, 1939. Ruckebuch, Y., L.P. Phanaeuf, R. Dunlop – Physiology of small and large animals. Decker,
Philadelphia, 1991. Rudas, P. – Veterinary physiology. Department of veterinary physiology, University of
Veterinary Science, Budapest, Hungary, 2001. Sánchez-Vizcaíno Rodríguez – Course of introduction in swine immunology. ISBN: 84-
699-5917-4, 2001. Schermer, S., Morphologie du sang des animaux de laboratoire, Ed. Davis Company,
Etats-Unis d'Amérique, 1967 Schmidt Nielsen, K. – Animal Physiology: Adaptation and environment, third edition,
Cambridge University Press, 1983. S h e r r i n g t o n , C.S. – The integrative action of the nervous system., Cambridge University
Press, London, 1906. Silbernagl S., et A. Despopoulos. Médecine-Siences Flammarion - 1995 - 366 p. Nerf et
Muscle p. 37 et 39Singh, G. – Réserves spermatiques épididimaires chez le verrat. Annales de biologie animale, biochimie, biophysique. 2 (1), pp. 47-50. hal-00896163, 1962.
Stoney, S. 1996 - Brain function, in Essentials of human physiology. Strungaru, Gr. 1983 – Fiziologie animală; E.D.P., Bucureşti. Swenson, M., W.O. Reece – Dukes’ Physiology of domestic animals. 11th Ed., Cornell Uni-
versity Press, 1993. Watkins Mullen, P., D. Horn Chang, H. J. Kron – Biology for living. Silver Burdett Co.,
1982. Wiedmeier, V.T. – Cardiac and circulatory physiology, dans Essentials of human
physiology, 1996. Yang, S., Smith, C., Prahl, J.M., Luo, X., DeLuca, H.F. (1993). Vitamin D deficiency suppresses
cell-mediated immunity in vivo. Archives of Biochemistry and Biophysics, 303, 98-106. Transport actif primaire. (2019, juillet 26). Wikipédia, l'encyclopédie libre. Page consultée
le 20:09, juillet 26, 2019 à partir de http://fr.wikipedia.org/w/index.php?title= Transport_ actif_ primaire&oldid=161311053.


















