
PARTIE I : CONTEXTE ET BIBLIOGRAPHIE -...
63
3 PARTIE I : CONTEXTE ET BIBLIOGRAPHIE 3 4 5 1) Présentation historique et socio-économique du paludisme 6 7 1.1) Historique du paludisme, apparition de la chimiorésistance 8 9 Historiquement, le paludisme est une maladie très ancienne. En 3000 avant JC, les Egyptiens 10 en souffraient et en mourraient déjà. Cette certitude est issue de la découverte de Plasmodies 11 dans des momies (Miller, 1994). A peu près à la même époque, soit à partir de 2700 avant JC, 12 des cas d’accès palustres sont décrits en Chine. Cette maladie étant déjà à l’époque un mal 13 familier, le paludisme a sa place dans la mythologie chinoise. Il y est représenté sous la forme 14 de trois démons armés respectivement de marteau, brasero et marmite d’eau froide, illustrant 15 les céphalées, fièvres et frissons (Debacker, 2000). 16 17 Cependant, bien que les hommes souffrent du paludisme depuis si longtemps, il faudra 18 attendre l’année 1630 pour voir apparaître le premier traitement à base d’écorce de Quinquina 19 découvert par Don Francisco Lopez. Deux siècles seront encore nécessaires pour que les 20 pharmaciens J. Pelletier et J.B. Caventou parviennent à isoler l’agent actif : la quinine. A 21 partir de cette époque, les recherches s’accélèrent et l’agent pathogène est découvert par 22 Alphonse Laveran en 1880 (Laveran, 1880). En 1897, les moustiques Anophèles sont 23 reconnus comme étant les vecteurs de la maladie et en 1922, les quatre Plasmodiums (P.) 24 infectant l’homme sont enfin tous décrits : P. falciparum, P. malariae, P. vivax et P. ovale. Il 25 faudra alors près de 60 ans pour comprendre entièrement le cycle parasitaire et ses 26 caractéristiques. La chloroquine et l’amodiaquine, deux antipaludiques de synthèse très 27 utilisés, font leur apparition en 1944. Malheureusement, dès 1960, des souches de P. 28 falciparum résistantes à la chloroquine sont découvertes sur deux continents (Asie et 29 Amérique du Sud). Ces chimiorésistances apparaissent simultanément avec des résistances 30 des anophèles à l’usage du DTT, utilisé depuis 1948 (dès 1953 en Grèce). 31 32 L’ère des chimiorésistances du vecteur et du parasite vient de débuter. Elle ne cessera de 33 s’étendre, au point de rendre la lutte contre le paludisme de plus en plus difficile, les 34 traitements perdant leur efficacité. L’extension rapide de la chimiorésistance s’explique, entre 35
-
Upload
phungthuan -
Category
Documents
-
view
218 -
download
0
Transcript of PARTIE I : CONTEXTE ET BIBLIOGRAPHIE -...

3
PARTIE I : CONTEXTE ET BIBLIOGRAPHIE 3
4
5
1) Présentation historique et socio-économique du paludisme 6
7
1.1) Historique du paludisme, apparition de la chimiorésistance 8
9
Historiquement, le paludisme est une maladie très ancienne. En 3000 avant JC, les Egyptiens 10
en souffraient et en mourraient déjà. Cette certitude est issue de la découverte de Plasmodies 11
dans des momies (Miller, 1994). A peu près à la même époque, soit à partir de 2700 avant JC, 12
des cas d’accès palustres sont décrits en Chine. Cette maladie étant déjà à l’époque un mal 13
familier, le paludisme a sa place dans la mythologie chinoise. Il y est représenté sous la forme 14
de trois démons armés respectivement de marteau, brasero et marmite d’eau froide, illustrant 15
les céphalées, fièvres et frissons (Debacker, 2000). 16
17
Cependant, bien que les hommes souffrent du paludisme depuis si longtemps, il faudra 18
attendre l’année 1630 pour voir apparaître le premier traitement à base d’écorce de Quinquina 19
découvert par Don Francisco Lopez. Deux siècles seront encore nécessaires pour que les 20
pharmaciens J. Pelletier et J.B. Caventou parviennent à isoler l’agent actif : la quinine. A 21
partir de cette époque, les recherches s’accélèrent et l’agent pathogène est découvert par 22
Alphonse Laveran en 1880 (Laveran, 1880). En 1897, les moustiques Anophèles sont 23
reconnus comme étant les vecteurs de la maladie et en 1922, les quatre Plasmodiums (P.) 24
infectant l’homme sont enfin tous décrits : P. falciparum, P. malariae, P. vivax et P. ovale. Il 25
faudra alors près de 60 ans pour comprendre entièrement le cycle parasitaire et ses 26
caractéristiques. La chloroquine et l’amodiaquine, deux antipaludiques de synthèse très 27
utilisés, font leur apparition en 1944. Malheureusement, dès 1960, des souches de P. 28
falciparum résistantes à la chloroquine sont découvertes sur deux continents (Asie et 29
Amérique du Sud). Ces chimiorésistances apparaissent simultanément avec des résistances 30
des anophèles à l’usage du DTT, utilisé depuis 1948 (dès 1953 en Grèce). 31
32
L’ère des chimiorésistances du vecteur et du parasite vient de débuter. Elle ne cessera de 33
s’étendre, au point de rendre la lutte contre le paludisme de plus en plus difficile, les 34
traitements perdant leur efficacité. L’extension rapide de la chimiorésistance s’explique, entre 35

4
autre, par une forte capacité de mutation génétique du parasite (Mercereau-Puijalon, 2000), 36
propriété intrinsèque de P. falciparum, mais également par des traitements mal administrés. 37
Ceux-ci sont bien souvent trop courts - arrêtés par le patient pour diverses raisons - ou sous 38
dosés - mauvaise prescription ou médicaments contrefaits - (White, 1998). 39
40
En 1976, la science progresse grâce à W. Trager et J.B. Jensen qui mettent au point la culture 41
in vitro de P. falciparum (Trager, 1976). Cette importante avancée ouvre la voie aux 42
approches moléculaires et immunologiques. Elle permet ainsi de faciliter l’étude de nouvelles 43
molécules antipaludiques et d’avancer vers la mise au point d’un vaccin. 44
45
Ces dernières années ont vu l’apparition sur le marché d’antipaludiques dérivés de 46
l’artémisine, un médicament « redécouvert » issu de la pharmacopée chinoise. Il apporte un 47
regain d’espoir dans le cadre de la chimiothérapie antipaludique. Cependant, il existe d’ores et 48
déjà une pression de sélection qui tend à l’apparition de chimiorésistance (Jambou, 2005). 49
C’est la raison pour laquelle l’OMS a préconisé en 2005 que les dérivés d’artémisinine soient 50
utilisés uniquement en bithérapie et jamais seuls, dans le but de limiter l’apparition du 51
phénomène (OMS, 2005). 52
53
Actuellement, un volet important dans la lutte antipaludique est l’utilisation de moustiquaires 54
imprégnées d’insecticides. Elles permettent de réduire le taux d’inoculation, donc l’incidence 55
des fièvres, et diminuent ainsi la mortalité et la morbidité (Maxwell, 2002). 56
57
Enfin, des études cliniques en cours au Sénégal montrent l’intérêt d’un traitement intermittent 58
préemptif chez les jeunes enfants pendant la saison de transmission (Cisse, 2006). 59
60
1.2) Le coût socio-économique du paludisme 61
62 Actuellement, on estime à 3 milliards le nombre de personnes vivant dans des zones à risque 63
de paludisme et ce chiffre est malheureusement en constante augmentation ces dernières 64
années. 500 millions de cas de paludisme sont répertoriés annuellement causant 1 à 3 millions 65
de morts (Guinovart, 2006). 90% des cas de paludisme recensés dans le monde le sont en 66
Afrique (Ruxin, 2005). Cela engendre des conséquences désastreuses, aussi bien sociales 67
qu’économiques, pour ce continent. Il est en effet important de noter qu’un accès palustre 68
simple, sans complication aucune, immobilise le malade pendant au moins une semaine, 69

5
expliquant le frein qu’apporte cette maladie au développement. On observe aujourd’hui pour 70
le paludisme une distribution géographique mondiale superposable à la planisphère des pays 71
pauvres (cf figure 1). Mais il est important de souligner que ce n’est pas une maladie des pays 72
pauvres mais un élément constituant de la pauvreté des pays qui en souffrent à grande échelle. 73
En 2001, une étude estimait que le produit intérieur brut moyen des pays impaludés est 5 fois 74
plus faible que celui des pays non impaludés (Gallup, 2001). Le poids du paludisme dans 75
l’économie africaine est estimé, pour ses effets directs, à plus de 1,3% de déficit de croissance 76
annuel (RBM), soit une perte de productivité équivalente à 12 milliards de dollars. Cette 77
maladie a également de nombreux effets collatéraux notables tels que des difficultés de 78
scolarisation des enfants dues à l’absentéisme prolongé et aux conséquences neurologiques 79
d’un accès grave. 80
81
82
Figure 1 : Répartition mondiale des pays souffrant de paludisme et de pauvreté. 83
84
Au Sénégal, le paludisme représente la première cause de morbidité et mortalité avec 35% des 85
motifs de consultations. 80% des accès palustres recensés sont dus à P. falciparum, parasite 86
induisant les formes les plus graves de la maladie. Chaque année, au Sénégal dont la 87
population est de 10 millions d’habitants, on dénombre 1 million d’accès palustres dans les 88
structures sanitaires. Un pourcentage important de ces accès provoque des avortements ou 89
accouchements précoces, les femmes enceintes et les enfants de moins de 5 ans étant les plus 90
touchés (PNLP, 2004). A l’heure actuelle, on estime que chez les populations rurales en 91
Afrique, un enfant sur cinq n’atteindra jamais son cinquième anniversaire. 92
93
94
95
96
97

6
2) Epidémiologie et clinique du paludisme 98
99
2.1) Mode de transmission de la maladie 100
101
Le paludisme est une parasitose due à des hématozoaires du genre Plasmodium transmis par 102
la piqûre des moustiques femelles du genre Anophèles. 103
104
Sur les 400 espèces et sous espèces d’Anophèles (A.) recensées, 35 sont considérées comme 105
des vecteurs potentiels du paludisme humain, dont notamment A. arabiensis, A. gambiae et A. 106
funestus qui sont les plus couramment recueillies dans les villages d’étude de Ndiop et 107
Dielmo (Fontenille, 1997). Les Anophèles ont besoin pour leur développement de chaleur et 108
de surface d’eaux peu profondes, allant des flaques d’eau aux petites rivières, qui leur servent 109
de gîte larvaire. C’est la raison pour laquelle le paludisme est essentiellement concentré vers 110
les tropiques et que les saisons des pluies sont les plus favorables à sa transmission. 111
Soulignons que, la distance de vol des Anophèles n’étant que de deux kilomètres, des villages 112
proches peuvent présenter des profils d’endémie différents. 113
114
Pour ce qui est des Plasmodiums, 123 espèces ont été répertoriées. Elles ont toutes une grande 115
spécificité d’hôte et des cycles biologiques similaires bien que non identiques. Sur les quatre 116
espèces spécifiques à l’homme, certains Africains sont naturellement résistants à l’une, à 117
savoir P. vivax, grâce à leur gène Duffy négatif (Miller, 1976 B). Aussi, ce parasite n’est que 118
très peu, voire pas, observé sur le continent Africain. La seule espèce réellement meurtrière 119
est P. falciparum, espèce la plus présente au Sénégal et dans de nombreuses parties du globe. 120
La majorité des études, la nôtre y compris, ont pour objectif d’analyser les mécanismes et les 121
cibles de l’immunité naturelle que l’on observe pour cette espèce de parasite. 122
123
2.2) La complexité du cycle parasitaire : un obstacle au développement du vaccin 124
125
Le cycle parasitaire se révèle excessivement complexe et multiple. Un cycle sexué est observé 126
chez le moustique, tandis que chez l’homme nous observons un cycle asexué qui se trouve 127
d’abord être pré érythrocytaire (intra-hépatique) puis érythrocytaire. Cette complexité 128
explique grand nombre de difficultés dans l’élaboration et le choix de molécules 129
potentiellement vaccinales et thérapeutiques. 130
131
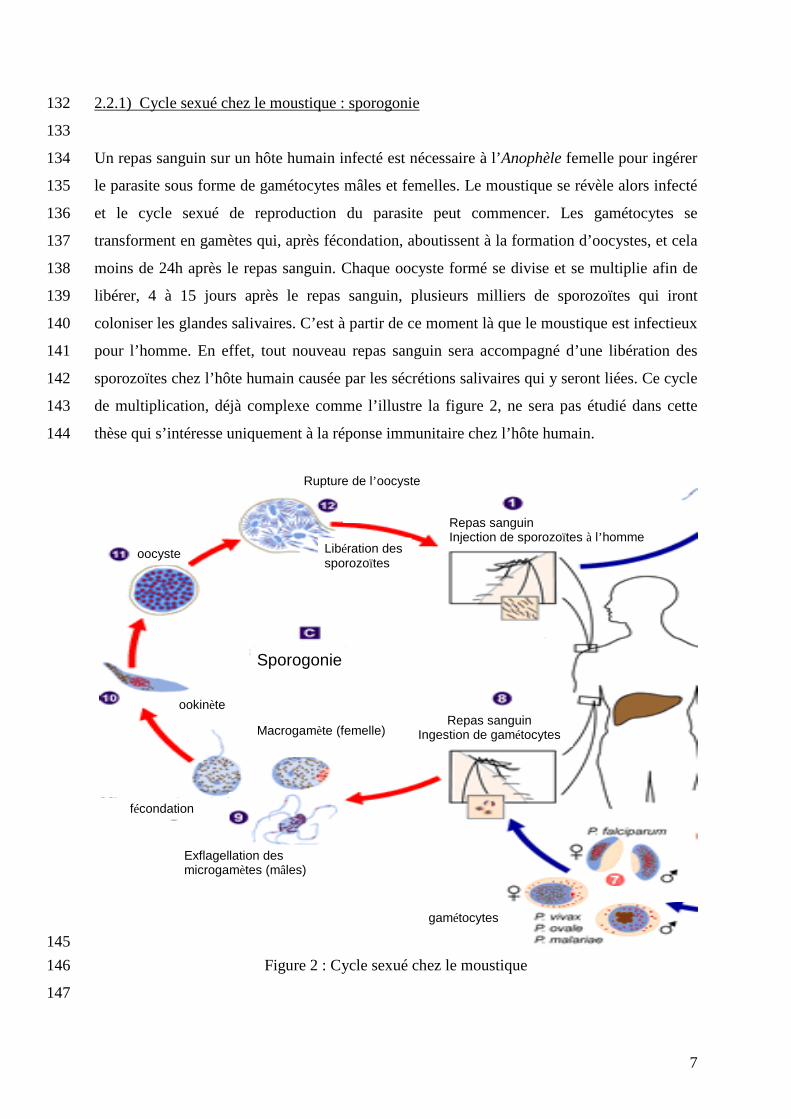
7
2.2.1) Cycle sexué chez le moustique : sporogonie 132
133
Un repas sanguin sur un hôte humain infecté est nécessaire à l’Anophèle femelle pour ingérer 134
le parasite sous forme de gamétocytes mâles et femelles. Le moustique se révèle alors infecté 135
et le cycle sexué de reproduction du parasite peut commencer. Les gamétocytes se 136
transforment en gamètes qui, après fécondation, aboutissent à la formation d’oocystes, et cela 137
moins de 24h après le repas sanguin. Chaque oocyste formé se divise et se multiplie afin de 138
libérer, 4 à 15 jours après le repas sanguin, plusieurs milliers de sporozoïtes qui iront 139
coloniser les glandes salivaires. C’est à partir de ce moment là que le moustique est infectieux 140
pour l’homme. En effet, tout nouveau repas sanguin sera accompagné d’une libération des 141
sporozoïtes chez l’hôte humain causée par les sécrétions salivaires qui y seront liées. Ce cycle 142
de multiplication, déjà complexe comme l’illustre la figure 2, ne sera pas étudié dans cette 143
thèse qui s’intéresse uniquement à la réponse immunitaire chez l’hôte humain. 144
145
Figure 2 : Cycle sexué chez le moustique 146
147
Exflagellation des microgamètes (mâles)
fécondation
Macrogamète (femelle)
ookinète
Sporogonie
oocyste
Rupture de l’oocyste
Libération des sporozoïtes
Repas sanguin Injection de sporozoïtes à l’homme
Repas sanguin Ingestion de gamétocytes
gamétocytes
8
Repas sanguin Injection de sporozoïtes à l’homme Cycle pré-érythrocytaire
Intra-hépatique
Sporozoïtes se dirigeant vers les
hépatocytes
Hépatocytes infectés
Formation de schizontes hépatiques
Rupture et libération des mérozoïtes
Les mérozoïtes se retrouvent dans la circulation sanguine
Maturation
Maturation
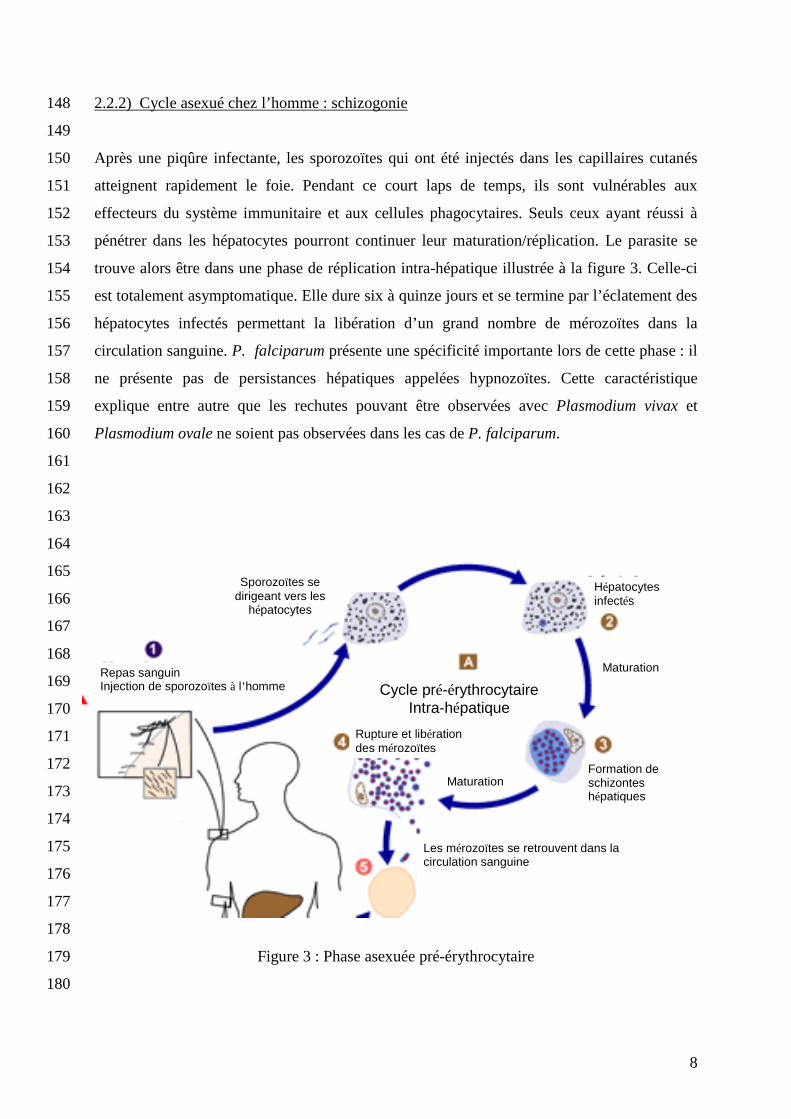
2.2.2) Cycle asexué chez l’homme : schizogonie 148
149
Après une piqûre infectante, les sporozoïtes qui ont été injectés dans les capillaires cutanés 150
atteignent rapidement le foie. Pendant ce court laps de temps, ils sont vulnérables aux 151
effecteurs du système immunitaire et aux cellules phagocytaires. Seuls ceux ayant réussi à 152
pénétrer dans les hépatocytes pourront continuer leur maturation/réplication. Le parasite se 153
trouve alors être dans une phase de réplication intra-hépatique illustrée à la figure 3. Celle-ci 154
est totalement asymptomatique. Elle dure six à quinze jours et se termine par l’éclatement des 155
hépatocytes infectés permettant la libération d’un grand nombre de mérozoïtes dans la 156
circulation sanguine. P. falciparum présente une spécificité importante lors de cette phase : il 157
ne présente pas de persistances hépatiques appelées hypnozoïtes. Cette caractéristique 158
explique entre autre que les rechutes pouvant être observées avec Plasmodium vivax et 159
Plasmodium ovale ne soient pas observées dans les cas de P. falciparum. 160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
Figure 3 : Phase asexuée pré-érythrocytaire 179
180
9
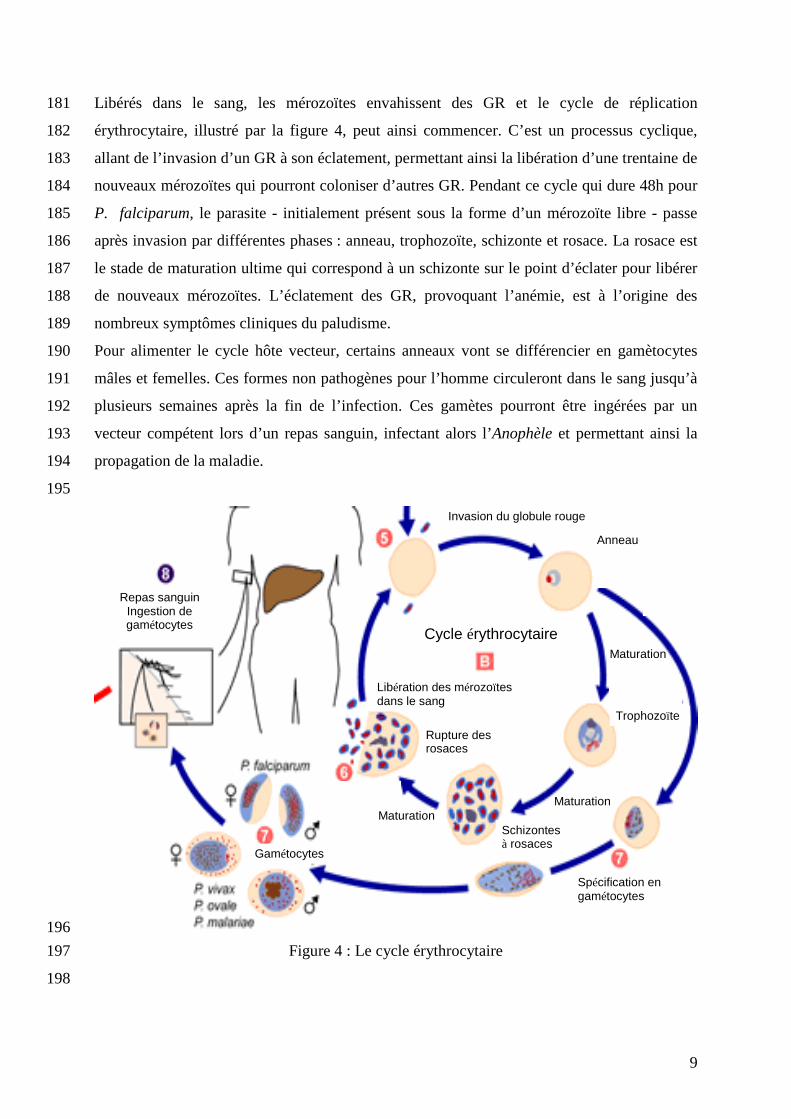
Libérés dans le sang, les mérozoïtes envahissent des GR et le cycle de réplication 181
érythrocytaire, illustré par la figure 4, peut ainsi commencer. C’est un processus cyclique, 182
allant de l’invasion d’un GR à son éclatement, permettant ainsi la libération d’une trentaine de 183
nouveaux mérozoïtes qui pourront coloniser d’autres GR. Pendant ce cycle qui dure 48h pour 184
P. falciparum, le parasite - initialement présent sous la forme d’un mérozoïte libre - passe 185
après invasion par différentes phases : anneau, trophozoïte, schizonte et rosace. La rosace est 186
le stade de maturation ultime qui correspond à un schizonte sur le point d’éclater pour libérer 187
de nouveaux mérozoïtes. L’éclatement des GR, provoquant l’anémie, est à l’origine des 188
nombreux symptômes cliniques du paludisme. 189
Pour alimenter le cycle hôte vecteur, certains anneaux vont se différencier en gamètocytes 190
mâles et femelles. Ces formes non pathogènes pour l’homme circuleront dans le sang jusqu’à 191
plusieurs semaines après la fin de l’infection. Ces gamètes pourront être ingérées par un 192
vecteur compétent lors d’un repas sanguin, infectant alors l’Anophèle et permettant ainsi la 193
propagation de la maladie. 194
195
196
Figure 4 : Le cycle érythrocytaire 197
198
Repas sanguin Ingestion de gamétocytes
Spécification en gamétocytes
Gamétocytes
Schizontes à rosaces
Rupture des rosaces
Anneau
Trophozoïte
Libération des mérozoïtes dans le sang
Invasion du globule rouge
Maturation
Maturation Maturation
Cycle érythrocytaire

10
Les formes asexuées du cycle érythrocytaire sont donc responsables de la morbidité et 199
mortalité. 200
201
2.3) Des accès cliniques d’intensités variables 202
203
L’infection à P. falciparum présente des intensités très variables selon les individus ou les 204
crises. En effet, cela va du portage asymptomatique à des cas de paludismes graves entraînant 205
la mort à la suite d’un coma - neuropaludisme - ou de déficiences d’un ou plusieurs organes. 206
Entre ces deux extrêmes se trouve l’accès simple qui provoque généralement une maladie 207
durant d’environ une semaine pendant lesquels les symptômes sont une très forte fièvre, des 208
sueurs et frissons, des douleurs articulaires et des céphalées. 209
210
La survenue de cas de paludismes graves serait provoquée par une combinaison complexe de 211
multiples facteurs. Ceux-ci sont aussi bien liés au polymorphisme génétique des parasites 212
qu’à la diversité génétique et immunologique de l’hôte humain. L’espèce vectrice et 213
plasmodiale, ainsi que l’intensité de la transmission sont également déterminantes dans la 214
gravité de la maladie. 215
D’un point de vue immunologique, la production de taux élevés de cytokines (TNFα/β, IFNγ, 216
IL6,…) induirait la production de molécules d’adhérence. Celles-ci favoriseraient alors la 217
séquestration d’hématies parasitées au niveau de l’endothélium capillaire de certains 218
vaisseaux profonds, provoquant ainsi des accès pernicieux pouvant entraîner la mort (Salvi-219
Zwetyenga, 1998). 220
221
3) L’immunité anti-palustre 222
223
3.1) L’immunité innée (cas des personnes drépanocytaires) 224
225
A l’inverse des personnes souffrant de paludisme aggravé, certains individus sont moins 226
sensibles, voire même réfractaires, à l’infection primaire. Cela peut s’expliquer par : i) 227
certaines anomalies génétiques altérant le développement du parasite dans le GR - 228
hémoglobinopathies et déficience en glucose-6-phosphate déshydrogénase - ; ii) l’absence de 229
molécule de surface du GR indispensable à la pénétration du parasite, telle que la molécule du 230
groupe Duffy pour P. vivax (Miller, 1976 A). 231
232
11
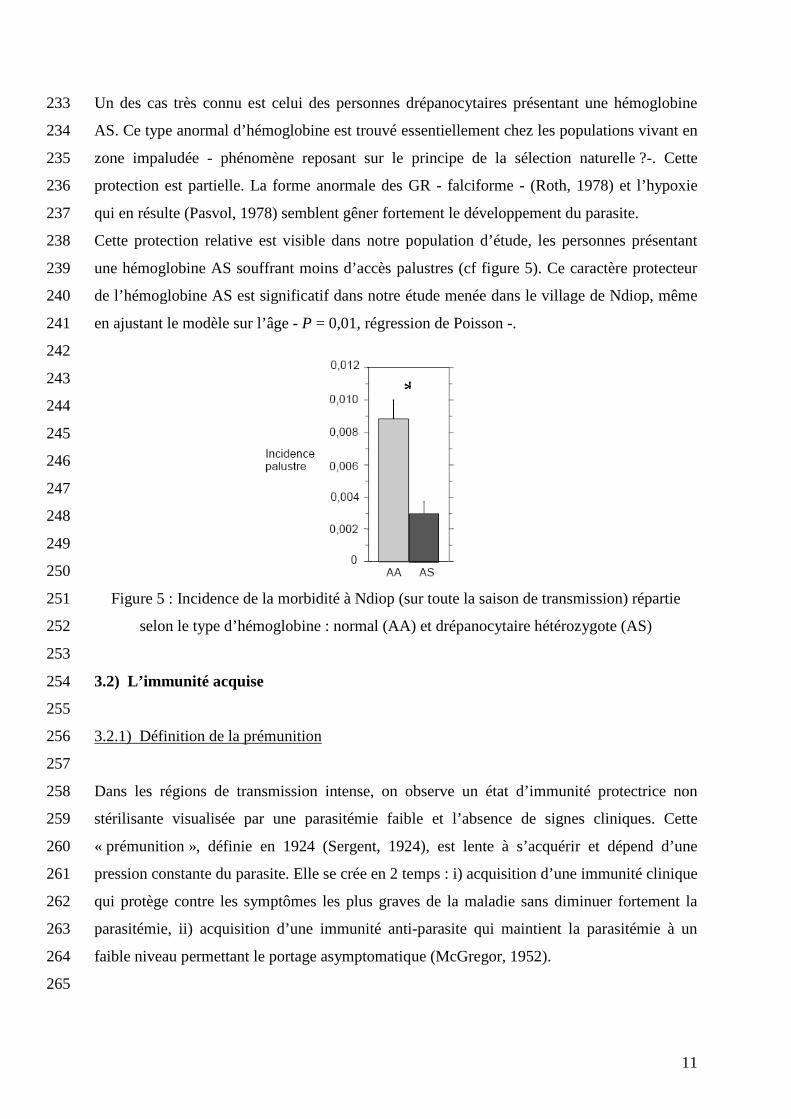
Un des cas très connu est celui des personnes drépanocytaires présentant une hémoglobine 233
AS. Ce type anormal d’hémoglobine est trouvé essentiellement chez les populations vivant en 234
zone impaludée - phénomène reposant sur le principe de la sélection naturelle ?-. Cette 235
protection est partielle. La forme anormale des GR - falciforme - (Roth, 1978) et l’hypoxie 236
qui en résulte (Pasvol, 1978) semblent gêner fortement le développement du parasite. 237
Cette protection relative est visible dans notre population d’étude, les personnes présentant 238
une hémoglobine AS souffrant moins d’accès palustres (cf figure 5). Ce caractère protecteur 239
de l’hémoglobine AS est significatif dans notre étude menée dans le village de Ndiop, même 240
en ajustant le modèle sur l’âge - P = 0,01, régression de Poisson -. 241
242
243
244
245
246
247
248
249
250
Figure 5 : Incidence de la morbidité à Ndiop (sur toute la saison de transmission) répartie 251
selon le type d’hémoglobine : normal (AA) et drépanocytaire hétérozygote (AS) 252
253
3.2) L’immunité acquise 254
255
3.2.1) Définition de la prémunition 256
257
Dans les régions de transmission intense, on observe un état d’immunité protectrice non 258
stérilisante visualisée par une parasitémie faible et l’absence de signes cliniques. Cette 259
« prémunition », définie en 1924 (Sergent, 1924), est lente à s’acquérir et dépend d’une 260
pression constante du parasite. Elle se crée en 2 temps : i) acquisition d’une immunité clinique 261
qui protège contre les symptômes les plus graves de la maladie sans diminuer fortement la 262
parasitémie, ii) acquisition d’une immunité anti-parasite qui maintient la parasitémie à un 263
faible niveau permettant le portage asymptomatique (McGregor, 1952). 264
265
*

12
Une étude parue en 1999 indique que dans les zones de fortes endémies, on observe 266
l’apparition d’une bonne immunité contre les accès graves non cérébraux dès la première ou 267
deuxième infection (Gupta, 1999). Celle-ci s’instaure donc rapidement contre les formes 268
graves mais plus lentement contre les accès simples. 269
En plus de sa lenteur d’acquisition - on compte de 6 à 15 ans selon l’intensité de la 270
transmission pour observer une prémunition satisfaisante -, elle est délétère et nécessite des 271
piqûres infectantes régulières pour être maintenue (Druilhe, 1987). A cela se rajoute une 272
spécificité d’espèce - pas d’immunité croisée entre les différentes espèces plasmodiales - et 273
une spécificité de stade. 274
275
Dans les zones de faible endémie comme à Dakar ce niveau d’immunité n’est pas atteint et les 276
habitants de tout âge sont susceptibles de faire des accès graves. 277
278
3.2.2) Immunité contre les stades sanguins – le cas des mérozoïtes 279
280
L’immunité contre les stades sanguins est complexe car régie par de multiples mécanismes 281
anti-parasites complémentaires. Leurs cibles sont le mérozoïte et les différents stades intra 282
érythrocytaires. 283
Cette thèse s’intéressant au mérozoïte et à des candidats vaccins basés sur ses protéines de 284
surface, nous allons maintenant expliciter plus en détail le rôle majeur du mérozoïte. 285
286
Le mérozoïte est la forme libre et invasive du parasite. C’est une petite cellule polarisée de 287
forme ovoïde de 1,5 µm de long et 1 µm de large. Malgré sa petite taille, c’est une cellule 288
excessivement complexe (cf figure 6) qui contient de nombreux organites apicaux impliqués 289
dans le phénomène d’invasion : ce sont les rhoptries, micronèmes et granules denses. 290

13
291
Figure 6 : Constitution d’un mérozoïte (Cowman 2006) 292
293
Le contact initial entre le mérozoïte et le GR est une étape cruciale, réversible et de faible 294
affinité. Le parasite pourrait ainsi « choisir » le GR le plus apte à l’accueillir. Ses critères de 295
sélection sont à l’heure actuelle encore mal définis mais il semblerait que Plasmodium soit 296
sensible : i) à l’âge des GR, ii) à l’état de phosphorylation du squelette érythrocytaire, iii) au 297
taux d’ATP intra-érythrocytaire, indispensable à l’invasion, iv) à un facteur cytosolique de 298
nature protéique (Grellier, 1990). 299
Dans cette étape de contact initial, le mérozoïte peut se fixer au GR à partir de n’importe quel 300
point de sa surface. Par la suite, il est capable de se redresser afin que sa proéminence apicale 301
soit en contact avec le GR. Ce contact est indispensable à l’invasion et permet une meilleure 302
accroche : la « jonction serrée ». Le parasite est alors partiellement internalisé et le contenu 303
des rhoptries est libéré pour former la vacuole parasitophore. Cette dernière procurera un 304
environnement favorable au développement du parasite. Les granules denses sont ensuite 305
libérées dans la vacuole et le mérozoïte s’internalise totalement (Cowman, 2006). Ce 306
processus d’invasion schématisé à la figure 7, dure moins d’une trentaine de secondes 307

14
. 308
Figure 7 : Processus d’invasion d’un globule rouge par un mérozoïte 309
310
L’internalisation du mérozoïte implique donc un processus complexe : reconnaissance, 311
réorientation, formation de la jonction serrée et de la vacuole parasitophore. Toutes ces étapes 312
impliquent de nombreuses protéines parasitaires. Elles ont été et sont encore beaucoup 313
étudiées afin de sélectionner les plus importantes pour réaliser des candidats vaccins. Dans la 314
revue écrite récemment par A.F. Cowman (Cowman, 2006), 47 protéines du mérozoïtes sont 315
répertoriées (cf tableau I). 316
317
Tableau I : Référencement des différentes protéines du mérozoïte connues 318
(les protéines étudiées dans cette thèse sont surlignées) 319
Protéines accrochées à la surface par un GPI
Protéines de surface périphériques
Protéines des micronèmes
protéines des rhoptries
protéines au col des rhoptries
MSP1 ABRA AMA1 RAMA Rh1 MSP2 Antigène S EBA140/BAEBL RAP1 Rh2a MSP4 GLURP EBA 175 RAP2 Rh2b MSP5 MSP3 EBA181/JESEBL RAP3 Rh3 MSP10 MSP6 EBL1 RhopH1(2) Rh4 Pf12 H101 MITRAP RhopH1(3.1) Rh5 Pf38 H103 ASP RhopH1(3.2) Pf92 MSP7 SUB2 RhopH1(9) Pf113 MSP7 like RhopH2 Pf41 RhopH3 SERA3 SERA4 SERA5 SERA6 320
Parmi celles-ci, MSP1 est la protéine de surface la plus abondamment trouvée sur le 321
mérozoïte. Une étude récente a estimé qu’elle représente à elle seule 30% des protéines de 322

15
surface accrochées par un GPI au mérozoïte. Pour comparaison, MSP4 et MSP5 confondus ne 323
représentent que 5% de ces protéines (Gilson, 2006). Cette forte représentation sur toute la 324
surface du mérozoïte conforte l’hypothèse que MSP1 joue un grand rôle dans les phénomènes 325
de reconnaissance et d’attachement, ceux-ci pouvant se dérouler à partir de n’importe quel 326
point du mérozoïte. 327
328
Parmi ces nombreuses protéines répertoriées, plusieurs subissent des modifications entre le 329
moment où le mérozoïte est libéré et celui où il envahit le GR. Par exemple, AMA1, 330
initialement présente dans les micronèmes, est transportée à la surface du mérozoïte juste 331
avant l’invasion (Cowman, 2006) et MSP1 subit une protéolyse en plusieurs étapes (Holder, 332
1994). 333
334
3.2.3) Le rôle des IgG 335
336
Les anticorps (Ac) sont les effecteurs majeurs de la protection contre les stades sanguins de 337
l’infection chez les individus immuns. Cela a pu être mis en évidence dans les années soixante 338
par transfert passif d’immunoglobulines, purifiées à partir d’un pool de sérums d’individus 339
hyper immuns, à des enfants en phase aiguë d’infection à P. falciparum. Cette injection d’IgG 340
a permis de noter une forte diminution de la parasitémie, indépendamment de l’origine 341
géographique des IgG. Inversement, des sérums déplétés en IgG n’ont aucun effet 342
(McGregor, 1963). Cette expérience à été renouvelée par la suite et les résultats ont été 343
confirmés à l’aide d’IgG purifiées (Sabchareon, 1991). 344
345
Les IgG ont donc une importance primordiale dans la protection anti-palustre et plus 346
particulièrement, les sous classes cytophiles à savoir les IgG1 et IgG3 (Garraud, 2003). Ils 347
permettent le phénomène d’opsonisation, indispensable pour certaines réactions immunitaires. 348
Une corrélation positive entre les IgG1 anti-MSP1p19 et la protection a été observée (Braga, 349
2002). Un résultat similaire a été obtenu avec les IgG3 anti-MSP2 (Metzger, 2003). A 350
l’inverse, les IgG2 et IgG4 agiraient en compétition avec les IgG1 et les IgG3 et bloqueraient 351
l’opsonisation due à ces dernières (Groux, 1990). Depuis, plusieurs études ont fait état de 352
relation entre les taux d’IgG non cytophiles et la gravité de la maladie. Un taux élevé d’IgG4 353
a été montré comme relié à un nombre important d’accès palustres (Soe, 2004). Une autre 354
étude récente a également montré que les personnes souffrant de neuropaludisme ont des taux 355
d’IgG2 et d’IgG4 beaucoup plus élevés que celles atteintes d’accès simples (Schreiber, 2006). 356

16
Cependant, chez l’homme, le titre d’Ac total n’est pas relié à la protection (McGregor, 1972), 357
les IgG non cytophiles n’ayant pas le même caractère protecteur anti-palustre que les IgG 358
cytophiles. Il est donc nécessaire pour analyser les capacités protectrices d’étudier, à travers 359
des suivis longitudinaux et transversaux, les relations entre la réponse spécifique d’un 360
antigène (Ag) et le statut immun de l’individu. Il est également possible, grâce à des tests 361
fonctionnels, de mesurer l’impact réel d’Ac spécifiques dans la réponse immunitaire 362
observable in vitro. Ces deux types d’études ont été réalisés au cours de cette thèse et seront 363
développés par la suite. 364
365
Le long délai d’acquisition de la prémunition serait nécessaire pour produire des Ac de bonne 366
affinité et avidité, capables de reconnaître les différents variants antigéniques et les 367
déterminants antigéniques mineurs, et cela avec un titre suffisant pour leur efficacité (Behr, 368
1992). Ces Ac protecteurs, une fois opérationnels, pourraient agir à différents niveaux : i) en 369
se fixant aux Ag de surface du mérozoïte, empêchant son interaction/pénétration dans le GR 370
(Mercereau-Puijalon, 1991), ii) en bloquant le développement du parasite à l’intérieur du GR 371
grâce à une voie de transport intra globulaire des Ac (Pouvelle, 1991), iii) en coopérant avec 372
d’autres cellules effectrices du système immunitaire (Druilhe, 1987). 373
374
3.2.4) Le rôle des cellules immunitaires (polynucléaires, macrophages,…) 375
376
De nombreuses cellules du système immunitaire ont un impact important dans l’acquisition de 377
l’immunité protectrice : les lymphocytes T, les cellules NK, les neutrophiles, les 378
macrophages/monocytes… Ces cellules peuvent réagir en coopération avec les Ac ou seules 379
par des propriétés fonctionnelles qui leur sont propres. Actuellement, nous connaissons leurs 380
rôles in vitro. En revanche, nous avons peu de preuves de ce qui se passe en réalité in vivo, 381
n’ayant pas de modèle animal réellement fiable. 382
383
Les lymphocytes T et les cellules NK seraient suractivés, permettant ainsi une production de 384
lymphokines et cytokines. Celles-ci seraient à l’origine du contrôle et de l’activation d’autres 385
cellules du système immunitaire. Pour exemple, les cellules NK induiraient entre autre une 386
activation primaire en IFNγ, un des facteurs principaux stimulant les macrophages, tandis que 387
la deuxième vague de production d’IFNγ serait plutôt due aux cellules T (Artavanis-Tsakonas, 388
2003). 389
390

17
Les polynucléaires et les monocytes/macrophages sont des cellules effectrices avec des rôles 391
similaires. Elles permettent divers types de phagocytose : 392
� une phagocytose directe permettant une réponse non spécifique ; 393
� une phagocytose médiée par les Ac accompagnée d’une libération de toxines capable 394
de tuer le parasite dans le GR. C’est le cas de l’ADCI pour les monocytes qui est détaillé au 395
paragraphe 5.3.1.2 ; 396
� une phagocytose médiée par les Ac accompagnée d’une destruction par libération de 397
radicaux oxygénés : la poussée respiratoire. C’est la base du test de chimiluminescence 398
développé dans notre travail et explicité au paragraphe 5.3.2. 399
400
Après avoir fait un rapide tour d’horizon de l’immunité naturelle anti-palustre, nous allons 401
maintenant nous intéresser aux candidats vaccins et à leur développement. 402
403
4) L’état actuel de la recherche vaccinale contre le paludisme 404
405
4.1) Les différentes étapes du développement d’un vaccin : de la recherche à 406
l’autorisation de mise sur le marché 407
408
Le développement d’un vaccin contre le paludisme suppose une stratégie claire sur les 409
objectifs et les résultats attendus. P. falciparum est l’espèce plasmodiale majoritairement 410
choisie pour les études car c’est le parasite responsable des formes mortelles de la maladie. P. 411
vivax. P. malariae et P. ovale sont quant à eux très peu étudiés en recherche vaccinale - 412
parasites non cultivables in vitro -. 413
414
En théorie, plusieurs types de vaccins sont réalisables : 415
� un vaccin empêchant l’infection en détruisant les sporozoïtes avant qu’ils n’atteignent 416
le foie ou en s’attaquant au parasite pendant le stade hépatique; 417
� un vaccin cherchant à réduire le développement de la maladie et ainsi empêcher les 418
formes graves d’apparaître. Mais dans ce dernier cas, quelle cible choisir ? Le mérozoïte ou le 419
GRp ? 420
� un vaccin altruiste cherchant à limiter l’endémicité et la transmission ; 421
� un vaccin multi stades. 422
Tous ces types de vaccins sont actuellement en développement et tous peuvent apporter une 423
solution future dans la lutte contre le paludisme. 424

18
425
Une fois le type de vaccin défini, il faut sélectionner l’Ag ou la construction synthétique qui 426
parait la plus apte à aboutir à un vaccin. De nombreuses études préalables sont nécessaires car 427
le parasite est excessivement polymorphe et ses Ag, et spécialement ceux des stades sanguins, 428
présentent en plus des structures conformationnelles complexes à reproduire. Celles-ci sont 429
cependant indispensables, la fonctionnalité des Ag étant structure - dépendante (Ballou, 430
2005). Il faut donc trouver le bon fragment de l’Ag afin qu’il soit représentatif d’un maximum 431
de variants antigéniques mais également le bon système d’expression qui permettra d’obtenir 432
une structure similaire à la structure native. 433
434
Lorsque le candidat vaccin est défini et que les études de structure des Ag synthétisés sont 435
concluantes, les tests précliniques peuvent commencer afin d’évaluer le potentiel vaccinal. 436
Ces tests sont des tests immuno-épidémiologiques réalisés in vitro à partir de sérums de 437
volontaires humains (ELISA, tests fonctionnels, …) et/ou des tests d’infection d’épreuve chez 438
des modèles animaux. Pour P. falciparum, les animaux utilisés sont essentiellement des 439
singes du Nouveau Monde, comme Aotus ou Saimiri, et certains modèles murins (souris 440
BXN) (Baddel, 1995). Cependant, ce ne sont pas les hôtes naturels de cette espèce de 441
Plasmodium. Aussi, les infections et réactions du système immunitaire ne peuvent pas être 442
parfaitement superposables à ce qui se passe chez l’homme (Mercereau-Puijalon, 2002). Ce 443
sont donc des modèles permettant uniquement de donner une tendance. 444
445
Lorsque les études précliniques sont concluantes, le candidat vaccin peut enfin être produit 446
selon les « Good Manufacturer Practice » (GMP) par des entreprises spécialisées et les essais 447
cliniques en tant que tels peuvent alors commencer. Ils comportent différentes phases qui sont 448
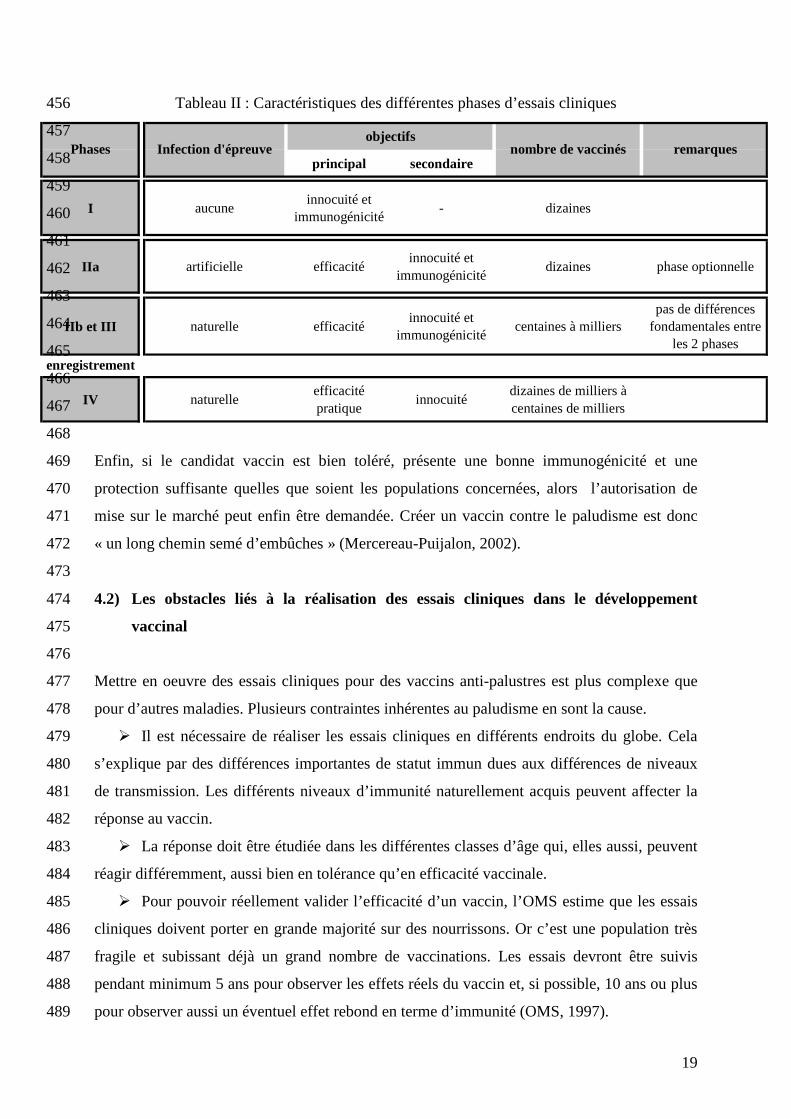
décrites dans le tableau II. Des résultats concluants à chaque phase sont nécessaires pour 449
pouvoir accéder à la suivante. Généralement, les tests de phase I, essentiellement des tests 450
d’innocuité, sont réalisés dans le pays à l’origine du candidat vaccin. Une zone d’endémie 451
n’est pas requise pour cette phase qui vise à prouver la tolérance par l’homme au vaccin et 452
non pas son efficacité. Il en est de même pour la phase IIa, l’infection y étant faite de façon 453
artificielle. A partir de la phase IIb, les tests doivent se dérouler dans des zones d’endémie et 454
de préférence, dans plusieurs zones d’endémie différentes. 455
19
principal secondaire
I aucune innocuité et
immunogénicité- dizaines
IIa artificielle efficacitéinnocuité et
immunogénicitédizaines phase optionnelle
IIb et III naturelle efficacitéinnocuité et
immunogénicitécentaines à milliers
pas de différences fondamentales entre
les 2 phases
IV naturelleefficacité pratique
innocuité dizaines de milliers à centaines de milliers
remarques
enregistrement
Phases Infection d'épreuveobjectifs
nombre de vaccinés
Tableau II : Caractéristiques des différentes phases d’essais cliniques 456
457
458
459
460
461
462
463
464
465
466
467
468
Enfin, si le candidat vaccin est bien toléré, présente une bonne immunogénicité et une 469
protection suffisante quelles que soient les populations concernées, alors l’autorisation de 470
mise sur le marché peut enfin être demandée. Créer un vaccin contre le paludisme est donc 471
« un long chemin semé d’embûches » (Mercereau-Puijalon, 2002). 472
473
4.2) Les obstacles liés à la réalisation des essais cliniques dans le développement 474
vaccinal 475
476
Mettre en oeuvre des essais cliniques pour des vaccins anti-palustres est plus complexe que 477
pour d’autres maladies. Plusieurs contraintes inhérentes au paludisme en sont la cause. 478
� Il est nécessaire de réaliser les essais cliniques en différents endroits du globe. Cela 479
s’explique par des différences importantes de statut immun dues aux différences de niveaux 480
de transmission. Les différents niveaux d’immunité naturellement acquis peuvent affecter la 481
réponse au vaccin. 482
� La réponse doit être étudiée dans les différentes classes d’âge qui, elles aussi, peuvent 483
réagir différemment, aussi bien en tolérance qu’en efficacité vaccinale. 484
� Pour pouvoir réellement valider l’efficacité d’un vaccin, l’OMS estime que les essais 485
cliniques doivent porter en grande majorité sur des nourrissons. Or c’est une population très 486
fragile et subissant déjà un grand nombre de vaccinations. Les essais devront être suivis 487
pendant minimum 5 ans pour observer les effets réels du vaccin et, si possible, 10 ans ou plus 488
pour observer aussi un éventuel effet rebond en terme d’immunité (OMS, 1997). 489
20
Les essais cliniques sont donc très lourds à mettre en place. 490
491
Enfin, à toutes ces difficultés et contraintes, s’ajoute le choix de l’adjuvant utilisé. En effet, 492
l’adjuvant agit directement sur l’efficacité du vaccin via son action sur : i) la configuration de 493
l’Ag, ii) le criblage antigénique, iii) la modulation immune. Son choix est donc d’une 494
importance capitale car un candidat vaccin prometteur qui donnerait des résultats très 495
décevants en essais cliniques pourrait voir ses mauvais résultats s’expliquer par une mauvaise 496
compatibilité avec l’adjuvant choisi (cf § 4.3 avec le cas de RTS,S). 497
498
4.3) Les différents essais vaccinaux actuellement en cours 499
500
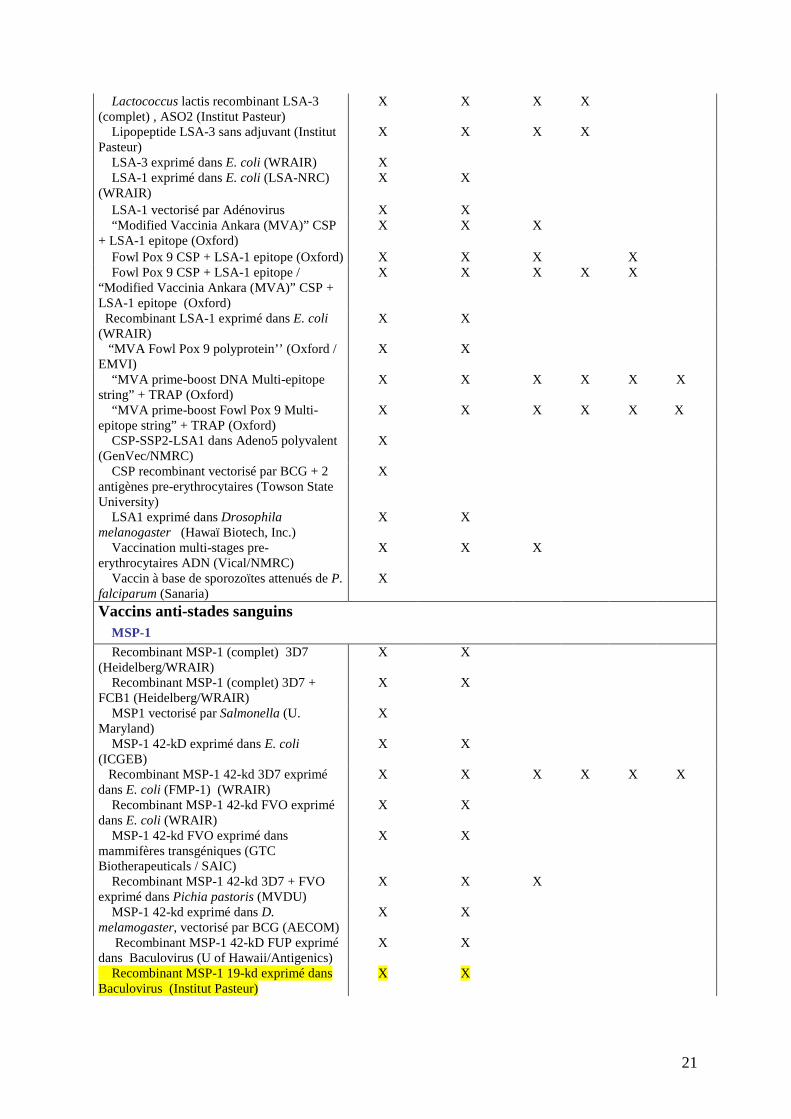
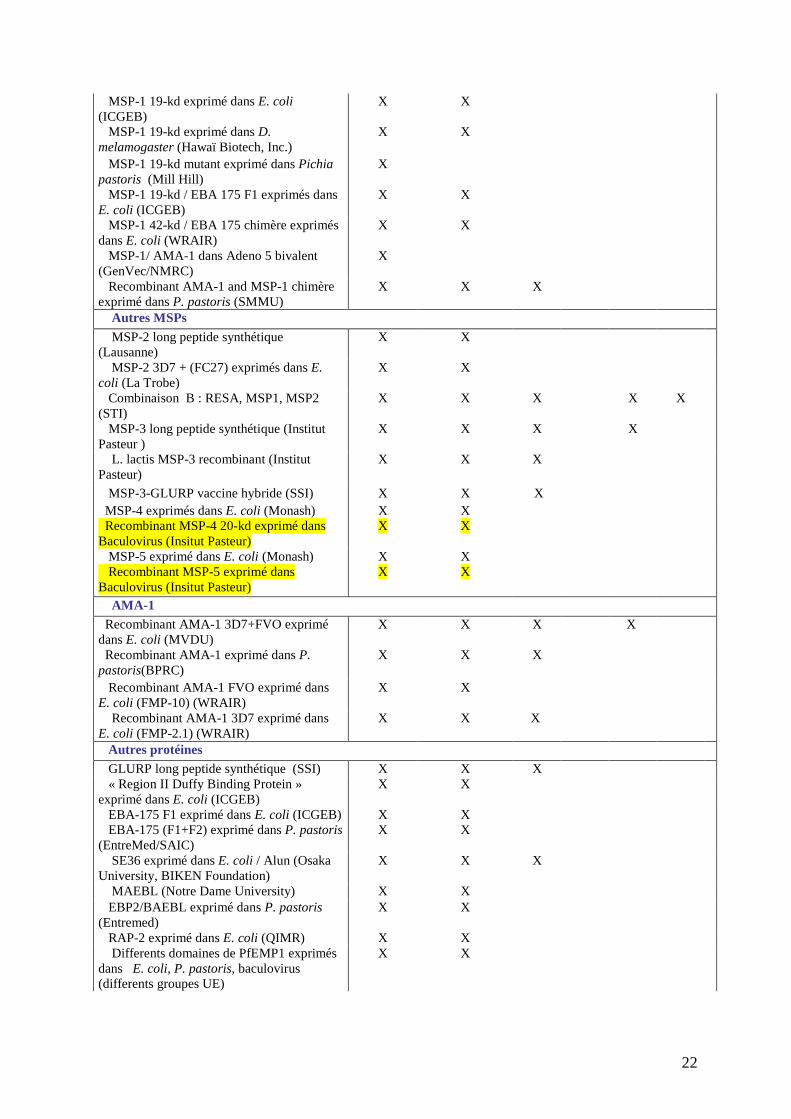
Il est très difficile de chiffrer le nombre réel de candidats vaccins potentiels actuellement en 501
développement ou en essais cliniques. Le tableau III en répertorie déjà un nombre important 502
mais il ne se veut pas exhaustif. Aujourd’hui, on peut donc recenser plus d’une centaine de 503
candidats vaccins en développement. 504
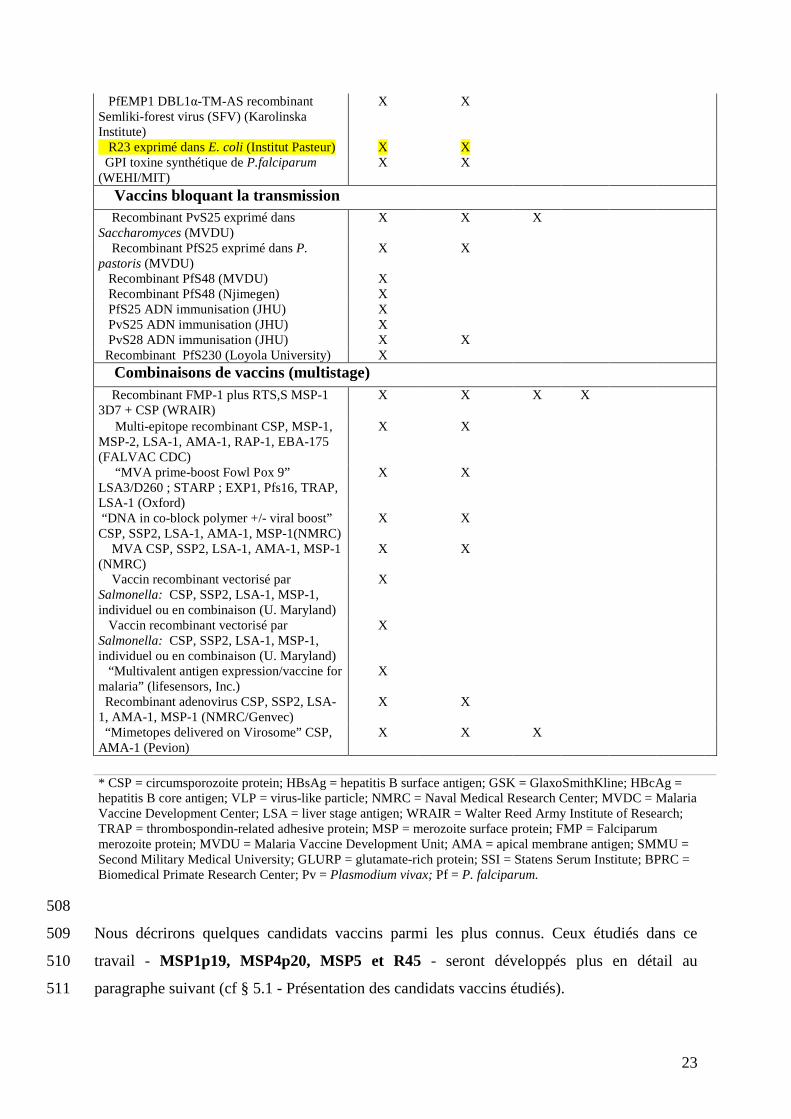
505 Tableau III : Candidats vaccins en développement clinique 506
(les candidats vaccins étudiés dans cette thèse sont surlignés) 507 Recherche Développement
préclinique Phase
1a Phase
2a Phase
1b Phase
2b
Vaccins pré-erythrocytaires CSP RTS,S/AS02A (GSK) X X X X X X RTS,S/ASO1B (GSK) X X X X HBcAg-CSP VLP (Malarivax Apovia) X X X X
“Modified Vaccinia Ankara (MVA)” CSP testé en combinaison avec RTS,S, AS02 (Oxford-GSK)
X X X X
CSP Recombinant adenovirus (NYU) X CSP Recombinant adenovirus (WRAIR/Crucell Holland)
X X
CSP Recombinant influenza (NYU) X CSP Recombinant vaccinia (NYU) X
CSP Recombinant Sindbis virus (NYU) X CSP Recombinant Yellow fever virus (NYU) X CSP long peptide synthétique (Lausanne) X X X X CSP ADN immunisation (Vical/NMRC) X X X X Long Vivax CSP C-terminal 72 acides aminés (MVDC)
X X X
Long Vivax CSP N-terminal 77 acides aminés (MVDC)
X X X
Long Vivax CSP repetition de 48 acides aminés (MVDC)
X X X
Autres antigènes LSA-3 long peptide synthétique (Institut Pasteur)
X X X
21
Lactococcus lactis recombinant LSA-3 (complet) , ASO2 (Institut Pasteur)
X X X X
Lipopeptide LSA-3 sans adjuvant (Institut Pasteur)
X X X X
LSA-3 exprimé dans E. coli (WRAIR) X LSA-1 exprimé dans E. coli (LSA-NRC) (WRAIR)
X X
LSA-1 vectorisé par Adénovirus X X “Modified Vaccinia Ankara (MVA)” CSP + LSA-1 epitope (Oxford)
X X X
Fowl Pox 9 CSP + LSA-1 epitope (Oxford) X X X X Fowl Pox 9 CSP + LSA-1 epitope / “Modified Vaccinia Ankara (MVA)” CSP + LSA-1 epitope (Oxford)
X X X X X
Recombinant LSA-1 exprimé dans E. coli (WRAIR)
X X
“MVA Fowl Pox 9 polyprotein’’ (Oxford / EMVI)
X X
“MVA prime-boost DNA Multi-epitope string” + TRAP (Oxford)
X X X X X X
“MVA prime-boost Fowl Pox 9 Multi-epitope string” + TRAP (Oxford)
X X X X X X
CSP-SSP2-LSA1 dans Adeno5 polyvalent (GenVec/NMRC)
X
CSP recombinant vectorisé par BCG + 2 antigènes pre-erythrocytaires (Towson State University)
X
LSA1 exprimé dans Drosophila melanogaster (Hawaï Biotech, Inc.)
X X
Vaccination multi-stages pre-erythrocytaires ADN (Vical/NMRC)
X X X
Vaccin à base de sporozoïtes attenués de P. falciparum (Sanaria)
X
Vaccins anti-stades sanguins MSP-1
Recombinant MSP-1 (complet) 3D7 (Heidelberg/WRAIR)
X X
Recombinant MSP-1 (complet) 3D7 + FCB1 (Heidelberg/WRAIR)
X X
MSP1 vectorisé par Salmonella (U. Maryland)
X
MSP-1 42-kD exprimé dans E. coli (ICGEB)
X X
Recombinant MSP-1 42-kd 3D7 exprimé dans E. coli (FMP-1) (WRAIR)
X X X X X X
Recombinant MSP-1 42-kd FVO exprimé dans E. coli (WRAIR)
X X
MSP-1 42-kd FVO exprimé dans mammifères transgéniques (GTC Biotherapeuticals / SAIC)
X X
Recombinant MSP-1 42-kd 3D7 + FVO exprimé dans Pichia pastoris (MVDU)
X X X
MSP-1 42-kd exprimé dans D. melamogaster, vectorisé par BCG (AECOM)
X X
Recombinant MSP-1 42-kD FUP exprimé dans Baculovirus (U of Hawaii/Antigenics)
X X
Recombinant MSP-1 19-kd exprimé dans Baculovirus (Institut Pasteur)
X X
22
MSP-1 19-kd exprimé dans E. coli (ICGEB)
X X
MSP-1 19-kd exprimé dans D. melamogaster (Hawaï Biotech, Inc.)
X X
MSP-1 19-kd mutant exprimé dans Pichia pastoris (Mill Hill)
X
MSP-1 19-kd / EBA 175 F1 exprimés dans E. coli (ICGEB)
X X
MSP-1 42-kd / EBA 175 chimère exprimés dans E. coli (WRAIR)
X X
MSP-1/ AMA-1 dans Adeno 5 bivalent (GenVec/NMRC)
X
Recombinant AMA-1 and MSP-1 chimère exprimé dans P. pastoris (SMMU)
X X X
Autres MSPs
MSP-2 long peptide synthétique (Lausanne)
X X
MSP-2 3D7 + (FC27) exprimés dans E. coli (La Trobe)
X X
Combinaison B : RESA, MSP1, MSP2 (STI)
X X X X X
MSP-3 long peptide synthétique (Institut Pasteur )
X X X X
L. lactis MSP-3 recombinant (Institut Pasteur)
X X X
MSP-3-GLURP vaccine hybride (SSI) X X X MSP-4 exprimés dans E. coli (Monash) X X Recombinant MSP-4 20-kd exprimé dans Baculovirus (Insitut Pasteur)
X X
MSP-5 exprimé dans E. coli (Monash) X X Recombinant MSP-5 exprimé dans Baculovirus (Insitut Pasteur)
X X
AMA-1
Recombinant AMA-1 3D7+FVO exprimé dans E. coli (MVDU)
X X X X
Recombinant AMA-1 exprimé dans P. pastoris(BPRC)
X X X
Recombinant AMA-1 FVO exprimé dans E. coli (FMP-10) (WRAIR)
X X
Recombinant AMA-1 3D7 exprimé dans E. coli (FMP-2.1) (WRAIR)
X X X
Autres protéines GLURP long peptide synthétique (SSI) X X X « Region II Duffy Binding Protein » exprimé dans E. coli (ICGEB)
X X
EBA-175 F1 exprimé dans E. coli (ICGEB) X X EBA-175 (F1+F2) exprimé dans P. pastoris (EntreMed/SAIC)
X X
SE36 exprimé dans E. coli / Alun (Osaka University, BIKEN Foundation)
X X X
MAEBL (Notre Dame University) X X EBP2/BAEBL exprimé dans P. pastoris (Entremed)
X X
RAP-2 exprimé dans E. coli (QIMR) X X Differents domaines de PfEMP1 exprimés dans E. coli, P. pastoris, baculovirus (differents groupes UE)
X X
23
PfEMP1 DBL1α-TM-AS recombinant Semliki-forest virus (SFV) (Karolinska Institute)
X X
R23 exprimé dans E. coli (Institut Pasteur) X X GPI toxine synthétique de P.falciparum (WEHI/MIT)
X X
Vaccins bloquant la transmission Recombinant PvS25 exprimé dans Saccharomyces (MVDU)
X X X
Recombinant PfS25 exprimé dans P. pastoris (MVDU)
X X
Recombinant PfS48 (MVDU) X Recombinant PfS48 (Njimegen) X PfS25 ADN immunisation (JHU) X PvS25 ADN immunisation (JHU) X PvS28 ADN immunisation (JHU) X X Recombinant PfS230 (Loyola University) X Combinaisons de vaccins (multistage) Recombinant FMP-1 plus RTS,S MSP-1 3D7 + CSP (WRAIR)
X X X X
Multi-epitope recombinant CSP, MSP-1, MSP-2, LSA-1, AMA-1, RAP-1, EBA-175 (FALVAC CDC)
X X
“MVA prime-boost Fowl Pox 9” LSA3/D260 ; STARP ; EXP1, Pfs16, TRAP, LSA-1 (Oxford)
X X
“DNA in co-block polymer +/- viral boost” CSP, SSP2, LSA-1, AMA-1, MSP-1(NMRC)
X X
MVA CSP, SSP2, LSA-1, AMA-1, MSP-1 (NMRC)
X X
Vaccin recombinant vectorisé par Salmonella: CSP, SSP2, LSA-1, MSP-1, individuel ou en combinaison (U. Maryland)
X
Vaccin recombinant vectorisé par Salmonella: CSP, SSP2, LSA-1, MSP-1, individuel ou en combinaison (U. Maryland)
X
“Multivalent antigen expression/vaccine for malaria” (lifesensors, Inc.)
X
Recombinant adenovirus CSP, SSP2, LSA-1, AMA-1, MSP-1 (NMRC/Genvec)
X X
“Mimetopes delivered on Virosome” CSP, AMA-1 (Pevion)
X X X
* CSP = circumsporozoite protein; HBsAg = hepatitis B surface antigen; GSK = GlaxoSmithKline; HBcAg = hepatitis B core antigen; VLP = virus-like particle; NMRC = Naval Medical Research Center; MVDC = Malaria Vaccine Development Center; LSA = liver stage antigen; WRAIR = Walter Reed Army Institute of Research; TRAP = thrombospondin-related adhesive protein; MSP = merozoite surface protein; FMP = Falciparum merozoite protein; MVDU = Malaria Vaccine Development Unit; AMA = apical membrane antigen; SMMU = Second Military Medical University; GLURP = glutamate-rich protein; SSI = Statens Serum Institute; BPRC = Biomedical Primate Research Center; Pv = Plasmodium vivax; Pf = P. falciparum.
508
Nous décrirons quelques candidats vaccins parmi les plus connus. Ceux étudiés dans ce 509
travail - MSP1p19, MSP4p20, MSP5 et R45 - seront développés plus en détail au 510
paragraphe suivant (cf § 5.1 - Présentation des candidats vaccins étudiés). 511

24
512
Concernant les vaccins pré érythrocytaires, RTS,S/ASO2A a fourni dernièrement des 513
résultats encourageants en phase clinique IIB. Une étude en double aveugle menée chez des 514
enfants en bonne santé de 1 à 4 ans a montré une baisse de la mortalité de 50% et du nombre 515
d’accès graves de 58%. Cependant l’infection n’a été réduite que de 37% et aucun impact 516
significatif sur l’anémie n’a pu être observé. Cette diminution de l’infection est faible pour un 517
vaccin pré érythrocytaire car il n’agit pas directement sur les formes sanguines, son action se 518
faisant uniquement sur les sporozoïtes (Alonso, 2004). Des études ont été reprises, portant 519
cette fois-ci sur le choix de l’adjuvant. Actuellement, c’est l’adjuvant ASO1B qui a été 520
sélectionné à la place de ASO2A car il semblerait permettre une combinaison avec RTS,S 521
plus immunogène. Des essais cliniques de phase I, IIa et IIb sont donc actuellement 522
renouvelés avec ce nouvel adjuvant (Stewart, 2006). 523
524
En ce qui concerne les vaccins contre les stades sanguins, le plus connu et le plus testé est 525
MSP1 avec différentes tailles de l’Ag - MSP1, MSP1 block 2, MSP1p42, MSP1p19 - et 526
différents types de vecteurs d’expression - Escherichia coli, Baculovirus, etc. -. Il est à 527
l’origine de plusieurs dizaines d’études. Ce chiffre, qui peut paraître surprenant, s’explique 528
par la difficulté de trouver : 529
� la partie de l’Ag - totalité ou fragment à déterminer - qui permet la reconnaissance 530
d’un maximum de variants parasitaires ; 531
� le système d’expression à utiliser - similarité avec la conformation native, coût de 532
fabrication, etc. - ; 533
� l’adjuvant avec lequel le combiner. 534
535
Les autres Ag des stades sanguins actuellement très étudiés par différentes équipes sont 536
AMA1 et MSP3, tous deux étant aussi des Ag du mérozoïte. 537
538
AMA1 est un Ag du pôle apical du mérozoïte, impliqué dans le phénomène d’invasion des 539
GR. C’est aussi un Ag présent dans les micronèmes des sporozoïtes, qui pourrait donc 540
également jouer un rôle dans l’invasion des hépatocytes. Cette double localisation et cette 541
possible double fonction en ont fait un des premiers candidats vaccins érythrocytaires, avec 542
MSP1, à avoir intéressé les chercheurs pour les potentialités qu’il représente. Cependant, sa 543
production difficile, due à la complexité de réalisation de sa structure secondaire et de son 544
repliement, deux facteurs capitaux pour sa fonctionnalité, a freiné les recherches pendant un 545

25
certain temps. En 2005, W.R. Ballou (Ballou, 2005) recense 5 constructions vaccinales 546
différentes issues de cet Ag étant, ou rentrant, en phase I d’essais cliniques. 547
548
MSP3 est un Ag dont l’intérêt a été découvert plus tardivement par l’équipe de Pierre Druilhe 549
lors de leurs travaux sur l’ADCI (pour explication de l’ADCI, cf § 5.3.1.2) (Oeuvray, 1994). 550
Depuis, cet Ag du mérozoïte est devenu lui aussi un important Ag d’étude. Des essais 551
cliniques de phase I ont donné des résultats immunologiques très prometteurs : toutes les 552
personnes vaccinées ont produit une forte réponse en lymphocytes T et plus de 70% ont 553
secrété des Ac à fort effet anti-parasite (Druilhe, 2005). Des essais cliniques de phase II et III 554
sont actuellement en cours en Afrique. 555
556
Un autre axe de recherche consiste à valider un vaccin correspondant à plusieurs Ag 557
différents. Cette approche pourrait permettre de limiter les risques d’effets nuls dus aux 558
variations génétiques du parasite. Cette multivalence autoriserait également une efficacité sur 559
plusieurs formes parasitaires, grâce à une combinaison d’Ag pré-érythrocytaires et 560
érythrocytaires. De plus, si les effets de chacun venaient à se révéler additifs, cela permettrait 561
de faire un très bon vaccin avec deux candidats vaccins qui, pris isolement, donnent des 562
résultats insuffisants. Cependant, cela n’est qu’une spéculation, et l’hypothèse d’interférence 563
entre les différents Ag annulant leur réponse individuelle est tout aussi envisageable. La 564
tolérance peut, elle aussi, être modifiée dans une combinaison vaccinale (Mahanty, 2003). 565
Aussi la réalisation d’un vaccin regroupant différentes constructions vaccinales nécessite 566
l’intégralité de nouveaux tests cliniques, même si celles-ci ont déjà toutes été intégralement 567
testées. Actuellement, on peut noter dans les associations vaccinales envisagées les 568
combinaisons « RTS,S + MSP1 » et « AMA1 + MSP1 ». Cette dernière combinaison a 569
d’ailleurs été montrée comme permettant l’obtention d’une meilleure réponse Ac in vitro et 570
des essais d’immunisation chez le singe ont donné des résultats très prometteurs (Pan, 2004). 571
Dans le cadre de l’équipe de Vaccinologie-Parasitaire de l’Institut Pasteur, on envisage une 572
association de MSP1p19 et MSP4p20, deux candidats vaccins ayant tous deux pour cible le 573
mérozoïte. 574
575
576
577
578
579

26
5) Contexte du sujet de travail 580
581
5.1) Présentation des candidats vaccins étudiés 582
583
5.1.1) MSP1p19 et le parasite transgénique d’étude D10-PcMEGF 584
585
MSP1 est une protéine de surface du mérozoïte, auparavant connue sous les noms de gp195, 586
P190, PMMSA ou encore MSA1. Elle est liée au mérozoïte par une ancre glycosyl-587
phosphatidyl-inositol (GPI) et comporte 2 domaines de type facteurs de croissance (EGF) à 588
son extrémité C terminale. C’est une molécule de haut poids moléculaire, environ 190 kDa, 589
qui subit une protéolyse. Celle-ci aboutit à la génération de fragments de 83 kDa, 42 kDa, 38 590
kDa et un quatrième d’une trentaine de kDa. Ils demeurent tous les quatre liés à la surface du 591
mérozoïte extra-érythrocytaire via un complexe non covalent. Au moment de l’invasion, le 592
complexe se rompt et le fragment de 42 kDa, ancré dans le mérozoïte par la GPI, se scinde à 593
nouveau. Il ne reste alors à la surface du mérozoïte qu’un fragment de 19 kDa comportant les 594
2 domaines EGF. Celui-ci, appelé MSP1p19, ne disparaîtra qu’une fois le parasite devenu 595
trophozoïte (Cavanagh, 1998). 596
597
La mise en évidence de cette protéolyse complexe a augmenté l’intérêt des chercheurs pour 598
cette molécule et de nombreuses constructions vaccinales à base de fragments différents de 599
MSP1 sont en cours d’étude. Cet intérêt a été soutenu par des essais de vaccination chez le 600
singe. Ils ont montré que la protéine MSP1p19 de P. cynomologi permettait d’obtenir chez 601
son hôte naturel une très bonne protection pour 8 des 9 singes vaccinés et une apparente 602
immunité stérile pour deux d’entre eux (Perera, 1998). 603
604
Le laboratoire de Vaccinologie Parasitaire de l’Institut Pasteur a concentré ses efforts sur 605
MSP1p19 exprimé par le Baculovirus (Bv) (Bonnet, 2006). Des études de cristallographie ont 606
montré que ce système d’expression permettait de synthétiser parfaitement les ponts 607
disulfures et que les constructions BvMSP1p19 de P. falciparum, P. vivax et P. cynomolgi 608
présentent un repliement et une structure tridimensionnelle similaires bien que leur séquence 609
en acides aminées diffèrent (Chitarra, 1999) (Pizarro, 2003). Ces études de cristallographie 610
qui mettent en avant l’importance de la structure ont été complétées par des études génétiques. 611
Bien qu’un « knock-out » au niveau du gène de MSP1 empêche la survie du parasite (Gaur, 612
2004), les domaines EGF de PfMSP1p19 peuvent être remplacés par ceux d’autres espèces 613

27
plasmodiales sans affecter la survie du parasite (O'Donnell, 2000) (O'Donnell, 2001). De plus, 614
il semblerait que les deux facteurs de croissance riches en cystéine de MSP1p19 soient la 615
première cible de la réponse immunitaire protectrice (John, 2004). 616
La structure des domaines EGF de l’Ag PfMSP1p19 serait primordiale pour sa fonctionnalité 617
et on suppose donc, au vu des études de cristallographie, que BvMSP1p19 a une structure 618
similaire à celle du fragment natif. 619
620
Certains Ac contre MSP1p19 peuvent prévenir la protéolyse et empêcher ainsi l’invasion des 621
GR. Inversement, d’autres types d’Ac anti-MSP1p19 peuvent, en se fixant sur l’Ag, empêcher 622
la fixation des Ac qui inhibent l’invasion des GR (Holder, 1999) (Chitnis, 2000). C’est une 623
des raisons pour laquelle tester la réponse Ac uniquement avec des tests ELISA est 624
insuffisante car cette méthode comptabilise tous les Ac, qu’ils soient fonctionnels ou non, 625
voire même bloquants. 626
En plus de ces capacités d’inhibition de réinvasion, MSP1p19 jouerait un ou plusieurs rôles 627
lors du développement intra-érythrocytaire du parasite (Woehlbier, 2006). 628
629
Dans le but d’analyser la fonctionnalité de cet Ag, nous avons travaillé avec deux types de 630
parasites transgéniques synthétisés par l’équipe de BS Crabb : 631
� D10-PcMEGF. Le plasmide PcMEGF a intégré une séquence ADN codante 632
permettant de remplacer l’intégralité des domaines EGF de PfMSP1p19 par ceux de P. 633
chabaudi, parasite dont l’hôte naturel est la souris. Des parasites de souche D10 ont ensuite 634
été transfectés par le plasmide ainsi obtenu. Cette modification génétique de MSP1p19 ne 635
s’est montrée aucunement délétère, contrairement à un knock-out complet du gène. Le 636
parasite obtenu est parfaitement viable et son cycle d’invasion se déroule normalement. L’Ag 637
MSP1p19 exprimé par ce parasite n’est plus reconnu par les Ac anti-PfMSP1p19 mais sa 638
localisation est correcte et il subit normalement sa protéolyse (O'Donnell, 2001) ; 639
� D10-PfM3’, parasite contrôle. Il a été transfecté par un plasmide contenant une 640
séquence non codante de même taille qui n’affecte pas l’intégrité de PfMSP1p19 (O'Donnell, 641
2000). 642
643
5.1.2) MSP4p20 644
645
MSP4 est une molécule de surface du mérozoïte découverte beaucoup plus récemment (1997) 646
et les premières études ELISA analysant sa réponse Ac lors d’infection naturelle chez 647

28
l’homme ne sont publiées que depuis 2001 (Wang, 2001). Elles montrent que MSP4 est une 648
cible de l’immunité naturelle chez l’homme, reconnue avec une forte prévalence chez les 649
personnes vivant en zone d’endémie - Vietnam -. 650
651
MSP4 est une protéine de surface du mérozoïte très importante pour l’invasion des GR car le 652
knock-out du gène codant pour MSP4 empêche la réinvasion. Mais il se peut également que 653
MSP4 ait d’autres fonctions à d’autres stades du développement du parasite (Polson, 2005 A). 654
Des études ont soutenu que cet Ag était présent à presque tous les stades de développement du 655
parasite, y compris sur des formes non sanguines. Cette hypothèse a été réfutée par l’équipe 656
de L. Wang qui l’a effectivement trouvé exprimée sur les différentes formes sanguines du 657
parasite - quel est son rôle une fois l’invasion réalisée ? - mais pas sur les sporozoïtes et 658
gamétocytes. Les Ac anti-MSP4 reconnaîtraient en réalité des épitopes d’Ag de ces deux 659
formes parasitaires selon un phénomène de réaction croisée des Ac (Wang, 2000). Celui-ci 660
peut s’expliquer par des homologies importantes, telle que celle observée avec la protéine 661
Sporozoite And Liver Stage Antigen (SALSA). Une homologie de 92% a en effet été 662
déterminée entre les résidus 53 à 135 de MSP4 et le fragment antigénique SALSA. Ce dernier 663
est connu pour se lier spécifiquement aux hépatocytes et permettre l’invasion de ces dernières 664
par les sporozoïtes (Puentes, 2004). 665
666
MSP4 est une molécule composée de 272 résidus, d’un domaine EGF et est accrochée au 667
mérozoïte par une ancre GPI. Son poids moléculaire est de 40 kDa. Sa particularité est de 668
présenter un faible polymorphisme. Aucun polymorphisme n’a pu être observé au niveau de 669
l’ancre GPI et de la séquence hydrophobe, un seul nucléotide polymorphe a été identifié dans 670
le domaine EGF (Benet, 2004). Ce polymorphisme très faible explique que MSP4 est étudié 671
généralement dans sa construction entière. 672
673
Au laboratoire de Vaccinologie-Parasitaire de l’Institut Pasteur, deux types de construction 674
ont été exprimés dans le système d’expression Baculovirus/cellules d’insectes : MSP4p40 qui 675
correspond à l’Ag entier et MSP4p30, appelé MSP4modII, dans lequel la seule séquence 676
réellement polymorphe, de 30 acides aminés, a été supprimée. En synthétisant ces deux 677
protéines, il est apparu qu’elles se dégradaient toutes les deux afin d’aboutir à un même 678
produit, MSP4p20. Il correspond au fragment C terminal de 20 kDa contenant le domaine 679
EGF (cf figures 8 et 9) (Polson, 2005 A). 680
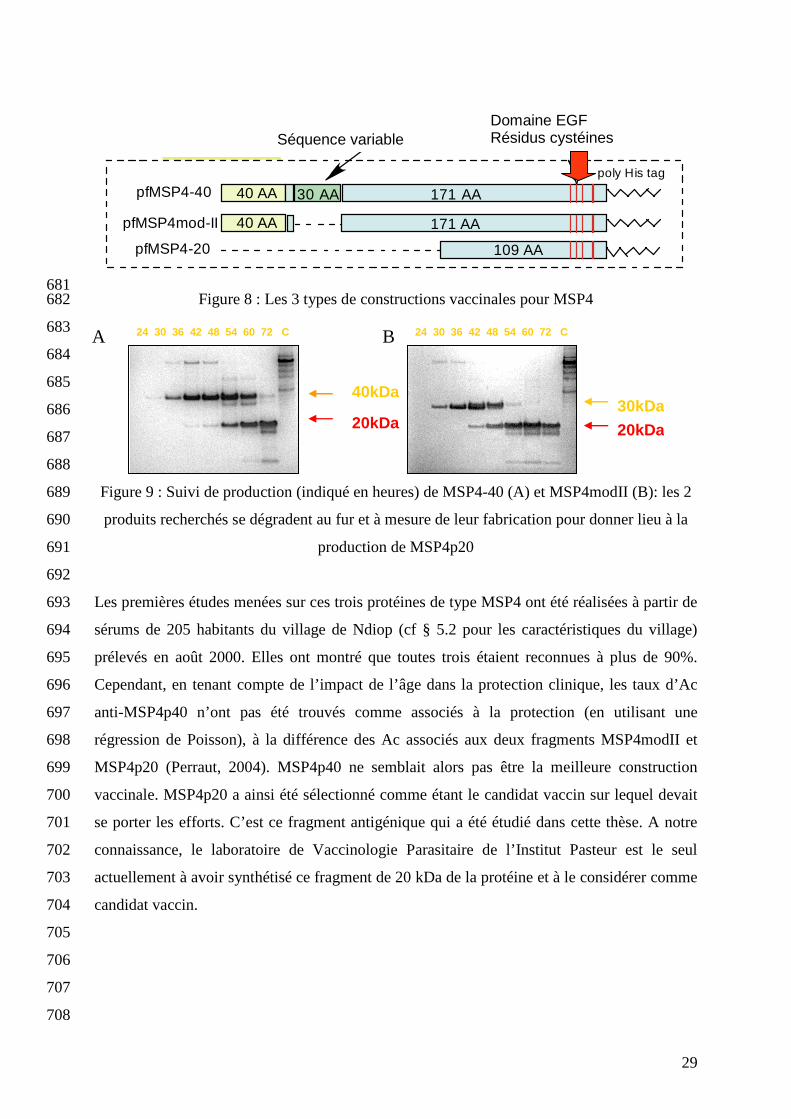
29
681 Figure 8 : Les 3 types de constructions vaccinales pour MSP4 682
683
684
685
686
687
688
Figure 9 : Suivi de production (indiqué en heures) de MSP4-40 (A) et MSP4modII (B): les 2 689
produits recherchés se dégradent au fur et à mesure de leur fabrication pour donner lieu à la 690
production de MSP4p20 691
692
Les premières études menées sur ces trois protéines de type MSP4 ont été réalisées à partir de 693
sérums de 205 habitants du village de Ndiop (cf § 5.2 pour les caractéristiques du village) 694
prélevés en août 2000. Elles ont montré que toutes trois étaient reconnues à plus de 90%. 695
Cependant, en tenant compte de l’impact de l’âge dans la protection clinique, les taux d’Ac 696
anti-MSP4p40 n’ont pas été trouvés comme associés à la protection (en utilisant une 697
régression de Poisson), à la différence des Ac associés aux deux fragments MSP4modII et 698
MSP4p20 (Perraut, 2004). MSP4p40 ne semblait alors pas être la meilleure construction 699
vaccinale. MSP4p20 a ainsi été sélectionné comme étant le candidat vaccin sur lequel devait 700
se porter les efforts. C’est ce fragment antigénique qui a été étudié dans cette thèse. A notre 701
connaissance, le laboratoire de Vaccinologie Parasitaire de l’Institut Pasteur est le seul 702
actuellement à avoir synthétisé ce fragment de 20 kDa de la protéine et à le considérer comme 703
candidat vaccin. 704
705
706
707
708
40 AA 171 AA30 AA
40 AA 171 AA
109 AA
pfMSP4-40
pfMSP4mod-II
pfMSP4-20
signal sequence
Sequence with �majority of variableresidues
poly His tag
EGF Cysteine residuesSéquence variable Domaine EGF Résidus cystéines
24 30 36 42 48 54 60 72 C
40kDa
20kDa
A 24 30 36 42 48 54 60 72 C
30kDa
20kDa
B

30
5.1.3) MSP5 709
710
La protéine MSP5 - Merozoite Surface Protein 5 - est jusqu’à présent très peu étudiée. La 711
plupart des articles traitant de cet Ag sont liés à son homologie avec MSP4. Génétiquement, 712
les locus des deux gènes codant pour les deux protéines se trouvent accolés sur le 713
chromosome 2. Dans trois espèces plasmodiales murines - P. chabaudi, P. yoelii, P. berghei - 714
elles se présentent sous la forme d’un seul gène codant pour une seule protéine appelée 715
MSP4/5 (Polson, 2005 B). 716
La protéine MSP5 présente également de grandes similitudes avec MSP4 : même nombre de 717
résidus protéiques (272), même poids moléculaire (40 kDa) et toutes deux possèdent un 718
domaine EGF à leur extrémité C terminale et sont reliées au mérozoïte par une ancre GPI. 719
Aussi suppose-t-on que ces deux Ag, tellement similaires, ont forcément des différences de 720
fonction et/ou de localisation sur le mérozoïte (Polson, 2005 B). Cette hypothèse est renforcée 721
par le fait que chez P. vivax, une étude a montré que MSP4 est réparti sur l’ensemble de la 722
surface du mérozoïte alors que MSP5 est localisé au niveau du pôle apical (Black, 2002). 723
724
Du point de vue du polymorphisme, MSP5 semble parfaitement conservé, caractéristique 725
plutôt rare chez P. falciparum. 726
727
La construction de MSP5 étudiée ici correspond à l’Ag en entier, synthétisé dans le 728
Baculovirus. Son analyse par spectrométrie de masse et une étude par marquage radioactif ont 729
montré que la protéine est libérée par les cellules d’insectes sous une forme myristilée - ajout 730
à la protéine d’acides gras - (Polson, 2005 A). Ce groupement myristile n’a pas pu être isolé, 731
aussi, les analyses portent sur MSP5 myristilé et non uniquement sur la protéine MSP5. 732
733
Avec cette construction, les études préliminaires ont montré que les sérums des populations 734
de Ndiop (cohorte de l’année 2000), reconnaissaient bien moins cet Ag que MSP1p19 ou 735
MSP4p20. Néanmoins, la réponse Ac qui lui est associée a été trouvée comme 736
significativement reliée à la protection clinique (Perraut et al, article en préparation). Cet Ag 737
est donc pour l’heure étudié dans l’objectif d’une association future avec d’autres Ag, allant 738
dans le sens des études réalisées chez la souris. Une immunisation des souris par MSP4/5 et 739
MSP1p19 est beaucoup plus efficace qu’une immunisation par un seul des deux Ag. Cet effet 740
n’est pas dû à une réaction croisée mais à une addition des réactivités de chaque Ag, ceux-ci 741

31
couvrant, quand ils sont combinés, une plus grande surface du mérozoïte. Il est probable 742
qu’ils empêchent ainsi plus efficacement la réinvasion (Kedzierski, 2002). 743
744
5.1.4) R23 (R45) 745
746
R45 est un Ag de surface du GRp. C’est une molécule de 160 kDa exprimée par les 747
trophozoïtes et toujours présente au stade schizonte. En étudiant sa séquence, l’équipe d’Odile 748
Mercereau-Puijalon, à l’Institut Pasteur, a détecté qu’un fragment de six acides aminés en 749
constitue la grande majorité : His Lys Ser Asp Ser Asn (HKSDSN). Celui-ci est répété, avec 750
de très faibles permutations, quatre-vingt dix fois dans la protéine. Un clone de AgR45, 751
appelé R23, contient quant à lui onze fois le motif HKSDSN (Perraut, 2001). Leur découverte 752
a montré que le fragment R23 était reconnu par les sérums immuns. (Bonnefoy, 1992). 753
754
Des études d’immunisation de singes par R23 - adjuvants alu-gel ou PAO - ont fourni des 755
résultats intéressants mais aussi contradictoires. Une augmentation de la réponse Ac contre P. 756
falciparum a été observée, et les analyses par IFA ont montré que les Ac produits étaient bien 757
R23 spécifiques. Les titres en Ac ont diminué après le test d’infection, laissant suggérer que 758
ces Ac jouent un rôle précoce dans le contrôle de la multiplication du parasite. Cependant, 759
seuls quatre singes vaccinés sur sept ont eu cette réaction positive, les trois autres ayant fourni 760
des résultats négatifs allant de l’absence totale de réaction à une sécrétion d’Ac dont la 761
spécificité n’a pas pu être prouvée (Perraut, 2001). Ces résultats montrent donc qu’un essai 762
vaccinal avec R23 en présence d’alu-gel ou de PAO peut fournir une réponse antigénique 763
spécifique. Cependant, le fait qu’elle n’ait été observée que dans la moitié des cas et de plus 764
sur un modèle animal ne permet pas de prévoir l’impact d’une vaccination chez l’homme. 765
766
Des études in vitro ont également été menées chez les habitants de Ndiop et Dielmo. Une 767
étude réalisée à Ndiop au début de la saison de transmission (cohorte de l’année 2000) a 768
montré que les sérums des habitants reconnaissaient modérément R23. De plus, la réponse 769
observée en ELISA n’était reliée ni à l’âge, ni à la réapparition de parasites (Perraut, 2003). 770
Une autre étude, se déroulant avant et après la saison de transmission (cohorte de l’année 771
1996), a permis de montrer que cette non relation avec l’âge n’était plus vérifiée en fin de 772
saison de transmission, et cela dans les deux villages. Les Ac anti-R23, ont été retrouvés en 773
quantité significativement plus importante chez les adultes au mois de décembre, reflétant une 774
dynamique des Ac due à la transmission (Perraut, 2000). 775

32
5.2) Présentation des villages de Ndiop et Dielmo – Des sites d’étude privilégiés 776
777
5.2.1) Deux villages sénégalais situés dans deux zones d’endémies différentes 778
779
Ndiop et Dielmo sont des villages pilotes pour l’étude de la prémunition anti-palustre en suivi 780
longitudinal depuis l’année 1990. Ils se situent dans la région de Fatick, au Sénégal et ont été 781
sélectionnés pour leur zone d’endémie, la faible, voir nulle, médicalisation des populations et 782
la proximité relative avec les laboratoires à Dakar (280 km). 783
784
785
Figure 10: Situation des villages de Ndiop et Dielmo sur la carte du Sénégal 786
787
Bien que très proches géographiquement - seulement 5 km les séparent - (cf figure 10), ces 788
deux villages présentent des caractéristiques totalement différentes du point de vue du 789
paludisme : 790
� Ndiop est dans une zone de mésoendémie où la transmission est saisonnière et 791
l’immunité met une quinzaine d’années à s’instaurer ; 792
� Dielmo est dans une zone d’holoendémie où la transmission est permanente. La cause 793
en est la présence d’une petite rivière, la Néma, en eau toute l’année et dont les rives 794
marécageuses servent de gîtes larvaires permanents (cf figure 11). L’immunité met alors six 795
ans à s’instaurer. 796
797
798
799
800
15°
14°
13°
16° N
17° W 16° 15° 14° 13° 12°
Domaine Soudano-guinéen
M A L I
G U I N E E
M A U R I T A N I E
1 0 0
k m
DIELMO NDIOP
Domaine Sahélo-soudanien
Domaine Soudanien
Domaine Sahélien
DAKAR
GAMBIE
Isohyète 300 mm
Isohyète 600 mm

33
801
802
803
804
805
806
807
808
Figure 11 : Images satellites « Quickbird » de Dielmo, prises en saison sèche (mars 2004) 809
810
A l’âge de soixante ans, les habitants de ces deux villages n’auront pas du tout le même 811
historique d’accès palustres. Un habitant de Ndiop aura eu en moyenne 62 accès palustres 812
dont 41% à l’âge adulte alors qu’un habitant de Dielmo, beaucoup plus soumis aux piqûres 813
infectantes, n’en aura eu que 43 dont 23% à l’âge adulte (Rogier, 1995). Cet exemple illustre 814
bien les différences d’acquisition de la prémunition en fonction des zones d’endémie et 815
l’intérêt de travailler sur ces deux villages simultanément. 816
De plus, les adultes à Dielmo souffrant encore occasionnellement d’accès palustres, la 817
prémunition n’apporte jamais une immunité totale et stérilisante (Trape, 1994). 818
819
5.2.2) Les différences de transmission 820
821
Ces deux villages sont situés dans deux zones d’endémies différentes. En zone de 822
mésoendémie, cas de Ndiop, l’indice splénique - à savoir le nombre de splénomégalies pour 823
100 personnes - est compris entre 11 et 50% et l’indice plasmodique - soit le pourcentage de 824
personnes avec des hématozoaires dans le sang - est compris entre 26 et 50%. 825
En zone d’holoendémie, cas de Dielmo, ces deux indices sont tous deux supérieurs à 75%. 826
827
D’un point de vue plus concret, la grande différence entre ces deux zones est la durée de la 828
saison de transmission palustre et le nombre de piqûres infectantes par habitant. A Dielmo, la 829
transmission est pérenne - les habitants sont donc soumis au paludisme toute l’année -, alors 830
qu’à Ndiop, la transmission intervient durant la saison de pluie - soit généralement des accès 831
palustres de août à décembre -. Afin de déterminer le nombre de piqûres infectantes, les 832
entomologistes réalisent des captures de moustiques sur l’homme et déterminent ainsi le 833
nombre d’anophèles piquants par 24h (ma) et le taux de vecteurs infectés (IS). Une fois ces 834
Santhié-Mouride
La Néma
Dielmo
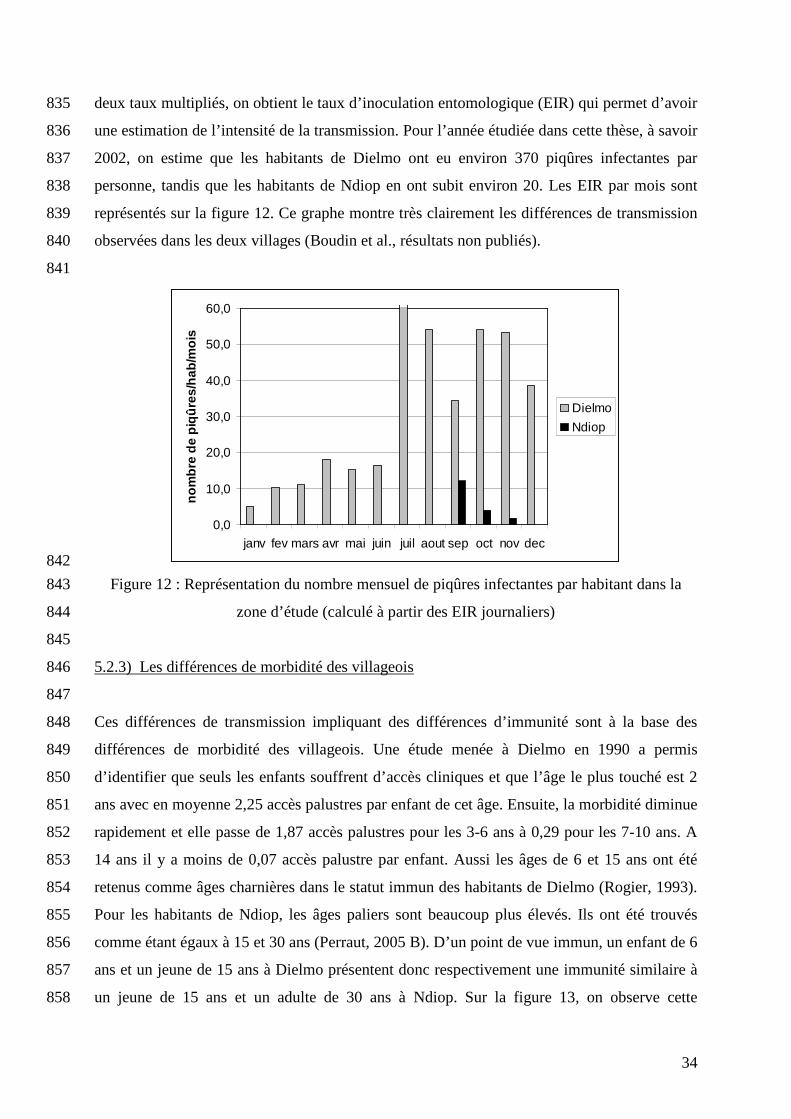
34
deux taux multipliés, on obtient le taux d’inoculation entomologique (EIR) qui permet d’avoir 835
une estimation de l’intensité de la transmission. Pour l’année étudiée dans cette thèse, à savoir 836
2002, on estime que les habitants de Dielmo ont eu environ 370 piqûres infectantes par 837
personne, tandis que les habitants de Ndiop en ont subit environ 20. Les EIR par mois sont 838
représentés sur la figure 12. Ce graphe montre très clairement les différences de transmission 839
observées dans les deux villages (Boudin et al., résultats non publiés). 840
841
0,0
10,0
20,0
30,0
40,0
50,0
60,0
janv fev mars avr mai juin juil aout sep oct nov dec
nom
bre
de p
iqûr
es/h
ab/m
ois
Dielmo
Ndiop
842
Figure 12 : Représentation du nombre mensuel de piqûres infectantes par habitant dans la 843
zone d’étude (calculé à partir des EIR journaliers) 844
845
5.2.3) Les différences de morbidité des villageois 846
847
Ces différences de transmission impliquant des différences d’immunité sont à la base des 848
différences de morbidité des villageois. Une étude menée à Dielmo en 1990 a permis 849
d’identifier que seuls les enfants souffrent d’accès cliniques et que l’âge le plus touché est 2 850
ans avec en moyenne 2,25 accès palustres par enfant de cet âge. Ensuite, la morbidité diminue 851
rapidement et elle passe de 1,87 accès palustres pour les 3-6 ans à 0,29 pour les 7-10 ans. A 852
14 ans il y a moins de 0,07 accès palustre par enfant. Aussi les âges de 6 et 15 ans ont été 853
retenus comme âges charnières dans le statut immun des habitants de Dielmo (Rogier, 1993). 854
Pour les habitants de Ndiop, les âges paliers sont beaucoup plus élevés. Ils ont été trouvés 855
comme étant égaux à 15 et 30 ans (Perraut, 2005 B). D’un point de vue immun, un enfant de 6 856
ans et un jeune de 15 ans à Dielmo présentent donc respectivement une immunité similaire à 857
un jeune de 15 ans et un adulte de 30 ans à Ndiop. Sur la figure 13, on observe cette 858
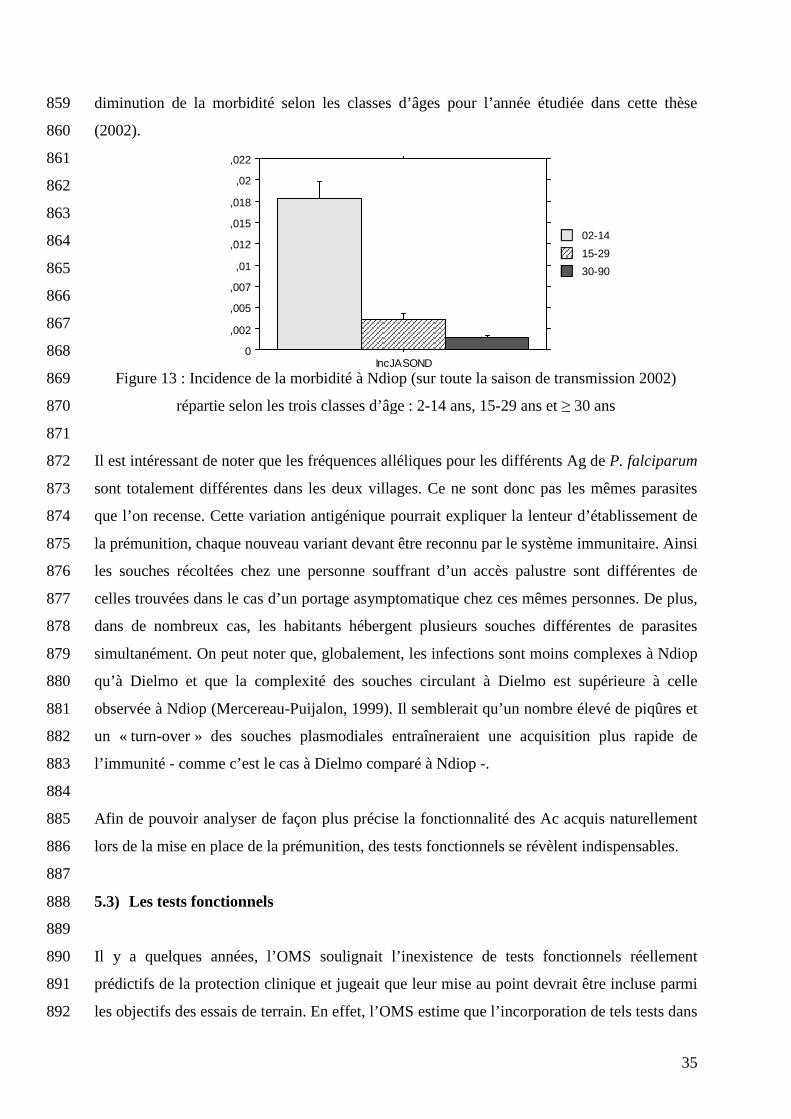
35
0
,002
,005
,007
,01
,012
,015
,018
,02
,022
IncJASOND
30-90
15-29
02-14
diminution de la morbidité selon les classes d’âges pour l’année étudiée dans cette thèse 859
(2002). 860
861
862
863
864
865
866
867
868
Figure 13 : Incidence de la morbidité à Ndiop (sur toute la saison de transmission 2002) 869
répartie selon les trois classes d’âge : 2-14 ans, 15-29 ans et ≥ 30 ans 870
871
Il est intéressant de noter que les fréquences alléliques pour les différents Ag de P. falciparum 872
sont totalement différentes dans les deux villages. Ce ne sont donc pas les mêmes parasites 873
que l’on recense. Cette variation antigénique pourrait expliquer la lenteur d’établissement de 874
la prémunition, chaque nouveau variant devant être reconnu par le système immunitaire. Ainsi 875
les souches récoltées chez une personne souffrant d’un accès palustre sont différentes de 876
celles trouvées dans le cas d’un portage asymptomatique chez ces mêmes personnes. De plus, 877
dans de nombreux cas, les habitants hébergent plusieurs souches différentes de parasites 878
simultanément. On peut noter que, globalement, les infections sont moins complexes à Ndiop 879
qu’à Dielmo et que la complexité des souches circulant à Dielmo est supérieure à celle 880
observée à Ndiop (Mercereau-Puijalon, 1999). Il semblerait qu’un nombre élevé de piqûres et 881
un « turn-over » des souches plasmodiales entraîneraient une acquisition plus rapide de 882
l’immunité - comme c’est le cas à Dielmo comparé à Ndiop -. 883
884
Afin de pouvoir analyser de façon plus précise la fonctionnalité des Ac acquis naturellement 885
lors de la mise en place de la prémunition, des tests fonctionnels se révèlent indispensables. 886
887
5.3) Les tests fonctionnels 888
889
Il y a quelques années, l’OMS soulignait l’inexistence de tests fonctionnels réellement 890
prédictifs de la protection clinique et jugeait que leur mise au point devrait être incluse parmi 891
les objectifs des essais de terrain. En effet, l’OMS estime que l’incorporation de tels tests dans 892

36
les phases I permettrait d’augmenter l’importance de cette phase préliminaire et faciliterait 893
ainsi le suivi des essais cliniques. Ces tests fonctionnels ne remplaceraient en rien les essais 894
cliniques de phases II à IV mais permettraient de renforcer le poids de la phase I et ainsi, de 895
mieux sélectionner les candidats vaccins aptes à passer en phase II (OMS, 1997). C’est dans 896
cette optique d’intégration de tests fonctionnels dans les phases I et éventuellement II d’essais 897
cliniques, que nous avons cherché au cours de cette thèse à développer et/ou valider des tests 898
fonctionnels en zone d’endémie et de les relier à la protection clinique observée in vivo. 899
900
5.3.1) Les tests les plus connus 901
902
Différents tests fonctionnels ont été étudiés afin de modéliser l’immunité anti-palustre et de 903
mieux la comprendre. Il s’agit essentiellement des tests d’inhibition de culture et de 904
réinvasion, de l’ADCI, mais également dans une moindre mesure de la chimiluminescence. 905
906
5.3.1.1) Les inhibitions de culture et de réinvasion (GIA et EIA) 907
908
Les inhibitions de culture parasitaire - 1 cycle, de schizontes à schizontes, de 40H à 48H selon 909
les souches - et de réinvasion - de schizontes à anneaux, environs 18H - sont pratiquées 910
depuis longtemps afin de déterminer l’impact du rôle bloquant des Ac dans la croissance 911
parasitaire. Les techniques de détection de la parasitémie permettant de déterminer le 912
pourcentage d’inhibition n’ont pas cessé d’évoluer. 913
914
i) Les méthodes classiques par lecture sur frottis ou détection par radiométrie 915
916
Initialement, les résultats de l’inhibition étaient lus par détection de la parasitémie sur frottis 917
coloré au Giemsa. Les différentes parasitémies étaient estimées au microscope et comparées 918
par rapport à des contrôles (Bouharoun-Tayoun, 1990). Cette technique est toujours à l’heure 919
actuelle une technique de référence mais n’étant absolument pas automatisée, elle demande 920
beaucoup de temps et de travail. De plus, elle ne permet pas de mesurer facilement la viabilité 921
des parasites, qui est pourtant un facteur fondamental. 922
923
L’autre technique de référence est quand à elle automatisée mais basée sur la radioactivité. Un 924
radio-isotope, l’hypoxanthine tritiée, est rajouté au milieu de culture afin d’être incorporé par 925
le parasite dans son ADN. La lecture de la radioactivité par compteur à scintillation β permet 926

37
d’estimer la parasitémie, la radioactivité détectée étant proportionnelle à celle-ci (Chulay, 927
1983) (Diouf, 2002). Cette technique a été beaucoup utilisée mais l’usage de la radioactivité 928
étant de plus en plus difficile, elle commence à être délaissée au profit de nouvelles méthodes. 929
De plus, l’hypoxanthine ne peut être incorporée que par les formes parasitaires trophozoïtes 930
âgées et jeunes schizontes ce qui crée des contraintes supplémentaires, surtout en inhibition de 931
réinvasion (Van der Heyde, 1995). Cependant elle reste la technique de référence. 932
933
ii) L’usage de la cytométrie en flux 934
935
La cytométrie en flux peut être utilisée comme technique de lecture rapide des parasitémies, 936
permettant l’analyse des résultats des tests d’inhibition de culture et de réinvasion. 937
938
Elle permet l’étude rapide de cellules isolées entraînées par un flux de liquide. Plusieurs 939
centaines de cellules d’une population hétérogène peuvent être analysées par minute. Les 940
populations et sous populations cellulaires sont alors identifiables car triées selon leur taille 941
(FSC), leur granulosité (SSC), et leur marquage par des fluorochromes spécifiques qui 942
fluorescent dans différents canaux. 943
944
On peut ainsi différencier les GRp des leucocytes grâce à leur taille (Van der Heyde, 1995), et 945
les GRp des GR sains grâce à de nombreux fluorochromes tels que le thiazole orange (TO) 946
(Diatta, 2004) (Jouin, 2004), l’hydroéthidine (HE) (Jouin, 2004) (Bergmann-Leitner, 2006), le 947
Hoechst 33 342 (Jouin, 1995), l’iodure de propidium (Contreras, 2004), le Syto 16 948
(Bergmann-Leitner, 2006), etc. Le choix des fluorochromes marqueurs d’ADN est donc vaste 949
et tous ne fluorescent pas dans les mêmes canaux - TO dans le canal FL1, HE dans le canal 950
FL2, etc.-. Le fluorochrome à utiliser peut donc être choisi en fonction des éventuels autres 951
marquages souhaités - double marquage de la population cible, marquage des Ac, marquage 952
de surface, etc. -. De plus, certains fluorochromes comme l’HE sont des marqueurs de 953
viabilité. Ils rendent donc l’interprétation du test encore plus sensible et performante. 954
955
iii) La méthode enzymatique par dosage de la pLDH 956
957
Une autre technique de détermination de l’inhibition réalisée repose sur le dosage de la 958
Lactate Déshydrogénase, LDH, une enzyme permettant le métabolisme du pyruvate en acide 959
lactique par l’intermédiaire de la NADPH pour permettre la production d’ATP. 960

38
961
La pLDH, LDH plasmodiale, est une des enzymes les plus actives du parasite. Son activité 962
augmente significativement quand le parasite est au stade de développement trophozoïte 963
âgé/schizonte. Elle présente la particularité d’utiliser rapidement le 3-acetyl pyridine adénine 964
dinucléotide (APAD) comme coenzyme. Elle est 200 à 300 fois plus réactive que la LDH 965
synthétisée par le GR pour réaliser cette réaction en présence d’APAD, permettant une bonne 966
spécificité de cible (Miura, 2007). Aussi un dosage par fluorométrie de l’APADH créé donne 967
un résultat proportionnel à la parasitémie (Makler, 1993) (OMS, 2004). Ce résultat permet 968
donc ensuite par calcul de déterminer le pourcentage d’inhibition réalisée. 969
970
iv) Remarques générales aux différentes méthodes 971
972
Toutes ces méthodes représentent différentes approches pour arriver au même résultat : 973
déterminer, grâce à la parasitémie ou à un facteur qui lui est lié, le pourcentage d’inhibition 974
parasitaire obtenue grâce aux Ac. Seule la phase de lecture et éventuellement de calcul de 975
l’inhibition varie réellement. Les résultats d’inhibition obtenus avec chacune de ces méthodes 976
sont similaires (Van der Heyde, 1995) (Contreras, 2004) (Bergmann-Leitner, 2006). 977
978
En comparant les différents articles cités dans l’ensemble du chapitre, on note que les tests 979
d’inhibition doivent être initiés avec un hématocrite de 1 à 2 %, une parasitémie de 0,3 à 980
0,5 %, en présence de 10 à 20 % de sérum. En effet, si le taux de parasites initial est trop 981
élevé, le sérum sera consommé trop vite et son action d’inhibition ne pourra pas être observée 982
pendant toute la durée du test, faussant ainsi les résultats. Il faut donc démarrer le test avec 983
une culture de parasitémie basse pour pouvoir quantifier réellement la réponse. 984
985
Il est important de noter que la souche utilisée et les conditions de culture - milieu de culture, 986
% CO2, … - affectent les résultats (Diouf, 2002). 987
988
5.3.1.2) L’ADCI 989
990
L’équipe de Pierre Druilhe à l’Institut Pasteur a développé il y a une vingtaine d’années un 991
test in vitro permettant d’étudier la réponse Ac anti-malarique en présence de monocytes 992
activés. Ce test est appelé ADCI pour Inhibition Cellulaire Dépendante des Ac (sigle en 993
anglais) (Druilhe, 2002). Il consiste à mettre en culture des parasites synchrones - stade 994

39
schizonte - en présence de monocytes activés - dans un ratio de 1 monocyte pour 200 GR - et 995
de milieu de culture supplémenté avec des IgG purifiées ou des sérums. La culture est ensuite 996
laissée à 37°C et dans les conditions de gaz adéquates pour la croissance parasitaire pendant 997
96H, avec ajout de milieu de culture au bout de 48H et 72H. La parasitémie passe alors de 998
0.5% à une valeur comprise entre 5 et 12% suivant l’efficacité des Ac dans le phénomène 999
d’ADCI (Bouharoun-Tayoun, 1995). 1000
1001
Ce test correspond à un mécanisme de protection à médiation cellulaire indirecte. Ce n’est pas 1002
une phagocytose par des monocytes que l’on observe mais le blocage du cycle parasitaire via 1003
une libération de dérivés toxiques. Ces toxines, relarguées par les monocytes, font suite à la 1004
présentation au monocyte par un Ac d’un mérozoïte. Elles sont alors capables de bloquer la 1005
maturation/réplication du parasite à l’intérieur du GR (cf figure 14) (Druilhe, 1997). 1006
1007
1008
Figure 14 : Représentation schématique du phénomène observé en ADCI 1009
1010
Cependant, ce test est difficilement reproductible. Pierre Druilhe lui-même mentionne une 1011
contrainte qui n’est pas négligeable : il est essentiel, pour que l’ADCI puisse être observé, que 1012
l’eau utilisée dans le milieu de culture présente des traces de minéraux. Une eau ultra pure, 1013
pourtant parfaitement adaptée à la culture de parasites et généralement utilisée en laboratoire, 1014
ne permet pas la réalisation du phénomène d’ADCI (Bouharoun-Tayoun, 1995). 1015
L’ADCI est donc une technique avec de nombreux déterminants extrinsèques au phénomène 1016
lui-même. 1017
1018
1019
1020
1021
1022

40
5.3.2) Un test à développer : la chimiluminescence 1023
1024
5.3.2.1) Les polynucléaires neutrophiles et leur action de phagocytose 1025
1026
Les cellules polynucléaires neutrophiles (PNN) sont des leucocytes polynucléés de 10 à 15 1027
microns, pouvant avoir jusqu’à 5 lobes. Ils représentent de 50% à 70% de la totalité des 1028
leucocytes, soit un taux sanguin moyen de 4000/µL. Ils ne circulent que durant 5H à 7H dans 1029
le sang périphérique, avant de se diriger vers les tissus où leur durée de vie n’est que de 1030
quelques jours. Ce sont généralement les premières cellules à arriver sur le lieu d’une 1031
inflammation et donc les premières cellules de défense immunitaire via leurs fonctions de 1032
phagocytose et bactéricidie. 1033
1034
La phagocytose par les PNN implique deux récepteurs fonctionnels différents : les récepteurs 1035
Fcγ de type FcγRIIA (CD32) et les récepteurs du Complément CR3 (CD11b et CD18 1036
intégrine). Ces deux récepteurs fonctionnels sont aidés dans la formation du complexe par 1037
deux co-récepteurs, FcγRIIIB (CD16) et CR1 (CD35) (Witko-Sarsat, 2000). 1038
Une fois la molécule accrochée à la surface du PNN, elle commence à être absorbée par celui-1039
ci et on observe la formation d’un phagosome qui active la NADPH oxydase (cf figure 15). 1040
1041
Figure 15 : Le phagosome 1042
1043
La NADPH oxydase est un complexe enzymatique multi protéique contenant de nombreuses 1044
sous unités qui sont soit des composés membranaires tels que i) le cytochrome b558 contenant 1045
le FAD, 2 hèmes, p22phox et gp91phox, ii) Rap1A, et iii) les protéines Nox ; soit des composés 1046
cytosoliques tels que i) p47phox, ii) p67phox, iii) p40phox, iv) Rac1/2, v) Cdc42 et vi) p29 1047
peroxiredoxine (Sheppard, 2005). Lors de l’activation des PNN et de la création du 1048
phagosome, ces différents composés vont se déplacer et se complexer, afin de permettre le 1049
transfert d’un électron et générer ainsi l’anion superoxyde (O2.-) à partir de l’oxygène (cf 1050
figure 16). 1051

41
1052
Figure 16 : Génération de l’anion superoxyde (réaction simplifiée) 1053
1054
Cette étape est appelée « burst oxydatif » ou « poussée respiratoire » (Lee, 2003). Elle est à 1055
l’origine de la formation d’une multitude de composés oxydants, tel que le peroxyde 1056
d’hydrogène (H2O2) et les oxydants halogénés (HOCl entre autre), qui peuvent tuer le micro-1057
organisme préalablement endocyté. 1058
1059
La phagocytose par les PNN est en réalité un phénomène beaucoup plus complexe que ce qui 1060
vient d’être présenté et qui implique une multitude d’autres enzymes. Les PNN renferment 1061
dans leurs granules primaires et secondaires de nombreuses enzymes lytiques et substances 1062
bactéricides : myélopéroxydases, phosphatases acides, hydrolases acides et alcalines, 1063
aminopeptidases, lysosyme, etc. (Witko-Sarsat, 2000). Les neutrophiles sont donc capables de 1064
détruire les microorganismes par deux voies, l’une dépendante et l’autre indépendante de 1065
l’oxygène. 1066
1067
Si l’on se concentre sur la voie dépendante de l’oxygène, il est intéressant de noter que bien 1068
que ce phénomène de poussée respiratoire existe également chez les macrophages, il est plus 1069
important chez les neutrophiles. Ces derniers libèrent un taux plus important de radicaux 1070
oxygénés dans le sang, auxquels s’ajoute un taux de défensines également plus élevé. Ces 1071
deux observations laissent donc supposer que les micro-organismes ingérés sont tués 1072
majoritairement par la première ligne de défense que sont les neutrophiles (Goldsby, 2000-1073
2001). 1074
1075
Ce phénomène de phagocytose par les PNN a pu être observé dans le cas du paludisme. In 1076
vivo tout d’abord, chez un enfant nigérian hospitalisé pour accès grave en 1985 (Sun, 1985), 1077
puis in vitro lors de nombreuses études. Celles-ci ont montré que les PNN activés avaient une 1078
préférence pour les mérozoïtes par rapport aux autres formes parasitaires sanguines 1079
(Malhotra, 1988). Elles ont aussi montré que des PNN inactivés et ne reconnaissant pas les 1080

42
GRp étaient capables de phagocyter des mérozoïtes; cette phagocytose entraînant une 1081
activation et ainsi une reconnaissance des GRp (Nnalue, 1988). Il semble donc tout à fait 1082
approprié de réaliser dans le domaine du paludisme un test fonctionnel basé sur la 1083
phagocytose des PNN par les mérozoïtes. 1084
1085
5.3.2.2) La poussée respiratoire, reflet de la phagocytose 1086
1087
Comme nous l’avons vu, la phagocytose est un phénomène excessivement complexe et donc 1088
difficile à exploiter. Cependant, il a l’avantage d’être rapide comme l’a montré l’équipe de 1089
W.L. Lee (cf figure 17) (Lee, 2003) : une fois les molécules accrochées aux récepteurs Fc des 1090
PNN, le phagosome se crée en moins de 3,5 minutes. 1091
1092
Figure 17 : Phagocytose par des neutrophiles humains de billes de latex opsonisées 1093
observée par microscopie électronique (temps exprimé en minute) 1094
1095
Observer la phagocytose au microscope demande beaucoup de temps et de travail. Cependant, 1096
elle peut être détectée de façon automatisée si on se limite au phénomène de poussée 1097
respiratoire. 1098
1099
Ce dernier est une étape clé dans le phénomène de phagocytose. Il se déroule quasiment 1100
instantanément, moins d’une minute après l’attachement à la surface des PNN (Dieye, 1986). 1101
Son action est d’autant plus importante que nous sommes en présence de mérozoïtes ou de 1102
formes âgées, car durant sa croissance le parasite consomme 80% de l’hémoglobine. Or, 1103
celle-ci crée une barrière anti-oxydante naturelle qui protége les formes anneaux. Aussi, de 1104
part sa consommation par le parasite, elle est de moins en moins efficace au fur et à mesure 1105
que celui-ci se développe, le rendant ainsi de plus en plus sensible à la poussée respiratoire 1106
(Stocker, 1984). 1107
1108
L’avantage majeur de ce test est la facilité de détection de la poussée respiratoire. Le luminol 1109
(5 amino-2,3-dihydro-1,4-phthalazind-ione) réagit avec les dérivés oxygénés libérés par les 1110

43
PNN. Leur réaction, présentée à la figure 18, crée alors un rayonnement lumineux détectable à 1111
411 nm. 1112
1113
1114
Figure 18 : Oxydation du luminol 1115
1116
De plus, des études menées in vitro ont montré qu’il existait une relation directe entre la 1117
génération de radicaux oxygénés par les PNN et la diminution de la parasitémie (Greve, 1118
1999), mais également entre la réponse fournie par le luminol et la poussée respiratoire 1119
(Dahlgren, 1999). 1120
1121
5.3.2.3) Diversité des protocoles du test de chimiluminescence 1122
1123
L’étude de la poussée respiratoire analysée par une technique de chimiluminescence dans le 1124
cas des études sur le paludisme est rapportée sous de nombreuses variantes. 1125
1126
Du point de vue de l’effecteur cellulaire, certaines équipes ont préféré travailler avec des 1127
monocytes plutôt qu’avec des PNN (Khusmith, 1982). D’autres groupes ont fait le choix 1128
d’utiliser du sang total afin de ne pas fragiliser les effecteurs cellulaires en raison des étapes 1129
de purification (Dieye, 1986). Cependant, certaines études ont montré que les GR présents 1130
dans le test de chimiluminescence influencent la réponse (Magrisso, 1995) et donc qu’une 1131
étape de purification de l’effecteur cellulaire choisi semblait tout de même préférable. 1132
1133
Du point de vue de la forme parasitaire utilisée pour ce type de test, nous avons vu ci-dessus 1134
qu’il pouvait être réalisé avec des schizontes ou des mérozoïtes, bien qu’une préférence soit 1135
observée pour le cas des mérozoïtes (Malhotra, 1988). Les techniques de préparation des 1136
mérozoïtes sont aussi extrêmement variables : une simple centrifugation (Lunel, 1990) ou une 1137

44
technique de flottaison sur Percoll (Kumaratilake, 1992). D’autres études portant sur des 1138
gamétocytes ont également été réalisées (Lensen, 1997). 1139
1140
Enfin, certaines équipes travaillant avec des PNN et des mérozoïtes, rajoutaient des cytokines 1141
pour activer les PNN. Cela augmente le coût financier du test et impliquait une macro 1142
méthode (volume totale de 1mL), alors qu’une micro méthode serait très fortement préférable 1143
pour un test de suivi d’essais cliniques. 1144
1145
Ces dernières années, aucune publication dans le domaine du paludisme n’a pu inclure une 1146
exploitation approfondie du test de chimiluminescence, celui-ci ayant été abandonné par 1147
manque de standardisation et d’homogénéisation des méthodes entre les divers utilisateurs. 1148
1149
5.4) Problématique 1150
1151
De très nombreux candidats vaccins sont actuellement en développement dans les différents 1152
laboratoires. Or l’absence de modèle animal réellement fiable ne permet pas de 1153
présélectionner efficacement ces différents candidats vaccins. Pour connaître cette efficacité, 1154
il faut donc atteindre les phases IIb des essais cliniques. Cela demande des investissements 1155
très lourds dans tous les domaines, créant un véritable frein au développement du vaccin. De 1156
plus, cela implique de tester en zone d’endémie des molécules dont l’efficacité vaccinale n’a 1157
pas encore été prouvée. Dans de telles conditions, il est important de pouvoir mettre en place 1158
des tests fonctionnels véritablement prédictifs de la protection pendant la phase Ia. Ils 1159
renforceraient l’impact des résultats observés dans cette phase et permettraient ainsi de faire 1160
une présélection basée sur la réponse réelle du vaccin et pas seulement sur sa toxicité (OMS, 1161
1997) (Miller, 1997) (Giersing, 2006). 1162
1163
Un laboratoire utilise actuellement des tests d’inhibition de culture avec quantification des 1164
parasites par la pLDH pour suivre les essais cliniques de phase Ia de leur construction AMA 1 1165
(Malkin, 2005). Cependant cette technique n’a pas encore été validée chez l’homme comme 1166
prédictive de la protection clinique, bien qu’elle ait été démontrée comme telle chez le modèle 1167
primate Aotus (Singh, 2006). Aussi cette approche très intéressante n’apporte pour l’instant 1168
que des indications de ce que pourrait être la protection procurée par ce type de vaccin, sans 1169
toutefois être extrapolable de façon certaine à la réponse observée in vivo chez l’homme. 1170
1171

45
Toutes ces approches montrent bien l’intérêt que porte la communauté scientifique et l’OMS 1172
à l’obtention d’un test fonctionnel fiable associé à la protection clinique. De plus, les 1173
différents tests existants et évoqués ci-dessus (cf § 5.3 - les tests fonctionnels) reposent sur 1174
des aspects différents d’une immuno-réaction reproduite in vitro, et nous ne connaissons ni ne 1175
pouvons argumenter actuellement de leur réalité in vivo. 1176
1177
La problématique de base est donc soulevée par les questions suivantes. 1178
� Un mécanisme donné est-il plus particulièrement représentatif de l'immunité de 1179
protection ? Si oui, lequel ? 1180
� Peut-on le modéliser par un test reproductible in vitro qui serait applicable dans un 1181
futur essai clinique ? 1182
Le but de cette thèse est donc d'analyser et de valider une approche fonctionnelle, en terme de 1183
mécanismes de destruction parasitaire modélisés in vitro, de la réponse Ac anti - P. 1184
falciparum. 1185
1186

46
PARTIE II : MATERIEL ET METHODES 1187
1188
1189 1) Les cohortes d’étude 1190
1191
1.1) Les populations des villages de Ndiop et Dielmo 1192
1193
L’Institut Pasteur et l’Institut de Recherche et Développement (IRD) étudient les populations 1194
des villages de Ndiop - depuis 1993, zone de mésoendémie - et Dielmo - depuis 1990, zone 1195
d’holoendémie - afin de mieux comprendre les principes de l’immunité naturelle. Le 1196
consentement éclairé des participants à l’étude est renouvelé deux fois par an et les enfants de 1197
moins de 2 ans ne sont pas éligibles dans le programme. 1198
1199
Depuis le début de l’étude, 735 personnes ont participé au programme à Ndiop et 512 à 1200
Dielmo. Certains de ces individus ont quitté l’étude car ils sont partis du village, ont souhaité 1201
se retirer du programme ou encore, sont malheureusement décédés. A l’inverse, certaines 1202
personnes intégraient régulièrement le programme suite à une installation récente dans le 1203
village, un changement d’opinion quant aux études entreprises, ou des naissances. A Ndiop, 1204
383 personnes faisaient toujours partie de programme en 2002 et à Dielmo, ils étaient au 1205
nombre de 316 cette même année. Ces habitants acceptent de se soumettre à des prélèvements 1206
sanguins et à un suivi médical et épidémiologique poussé. 1207
1208
1.2) Le suivi des populations 1209
1210
Dans les deux villages, après une enquête démographique avec bilan clinique et biologique, 1211
des enquêtes transversales sont menées chaque année et une enquête longitudinale a duré de 1212
1990 à 2003. 1213
1214
L’enquête transversale correspond à un passage unique en une période déterminée de l’année, 1215
à savoir dans notre étude le début du mois de juillet, soit juste avant le début de la saison des 1216
pluies qui est la période de plus forte transmission. Les habitants sont suivis d’un point de vue 1217
épidémiologique et clinique. Des prélèvements veineux sont effectués afin de pouvoir réaliser 1218
les études immunologiques. Des prélèvements sériques et gouttes épaisses sont également 1219
collectés pour permettre de déterminer une parasitémie potentielle. Les gouttes épaisses seront 1220

47
par la suite toutes lues sur 200 champs et classées en groupes de parasitémie (F1, F2, …) 1221
selon une méthode établie par JF Trape (Trape, 1985). 1222
1223
L’enquête longitudinale correspond quant à elle à un suivi coordonné sur plusieurs années de 1224
la population, enregistrant tous les événements cliniques, parasitologiques et entomologiques. 1225
L’acquisition de la prémunition peut ainsi être analysée et des corrélations entre les études 1226
immuno-épidémiologiques et la protection clinique peuvent être établies. En contrepartie, 1227
l’Institut Pasteur et l’IRD se sont engagés à assurer une présence médicale 24H/24 dans 1228
chacun des villages et à prendre en charge médicalement les populations. 1229
1230
Grâce au suivi quotidien par l’équipe médicale, tous les accès palustres des personnes 1231
présentes sur le site lors de ces accès sont recensés de manière active. Un accès palustre est 1232
défini par la présence de fièvre et/ou de symptômes assimilés au paludisme, accompagné 1233
d’une parasitémie vérifiée soigneusement par examen microscopique réalisé par un lecteur 1234
expérimenté. Afin de se placer dans les conditions optimales de l’acquisition naturelle de 1235
l’immunité, des seuils de traitements, spécifiques aux différentes tranches d’âges ont été 1236
définis : ils commencent à une valeur comprise entre 12000 et 15000 parasites par microlitre 1237
de sang chez les enfants et diminuent avec l’âge pour arriver à une valeur comprise entre 2500 1238
et 4000 parasites/µL chez les adultes. Seules les personnes ayant des taux de parasitémie 1239
supérieurs au seuil défini pour leur âge sont traitées par des antipaludéens et l’accès palustre 1240
est comptabilisé comme tel. Si le seuil de traitement n’est pas atteint, le paracétamol est 1241
l’unique prescription et un suivi journalier de la parasitémie est réalisé pour détecter à partir 1242
de quand un traitement devient nécessaire. Cela permet, tout en soulageant le malade et en le 1243
surveillant de près, de détecter les personnes dont la prémunition est suffisante pour « s’auto 1244
soigner » - cas non comptabilisés dans les accès palustres -. Cela évite également le 1245
développement des chimiorésistances (Rogier, 1995) (Rogier, 1996). 1246
Enfin, les épisodes cliniques sont considérés comme indépendants s’ils sont séparés d’au 1247
moins 15 jours. 1248
1249
1.3) La transversale Ndiop - Dielmo 2002 1250
1251
Afin de réaliser la double transversale 2002 qui servira de base à notre étude, nous avons 1252
effectué des prélèvements sur une partie de la population encore enrôlée en juillet 2002. 1253

48
Pour les deux villages, les prélèvements ont eu lieu avant le début de la saison de transmission 1254
à Ndiop et avant le pic de transmission maximal à Dielmo, soit entre le 2 et le 15 juillet 2002. 1255
Nous avons donc à notre disposition les sérums d’une série synchrone de prélèvements. 1256
1257
A Ndiop, la cohorte ainsi prélevée comprend 221 individus, âgés de 3,4 à 76,9 ans, avec une 1258
moyenne d’âge de 24,0 ans. Elle est composée à 54% de femmes et 14% de la population 1259
étudiée présentent une hémoglobine à profil drépanocytaire hétérozygote AS. La transmission 1260
du paludisme était interrompue - non mesurable par les techniques entomologiques utilisées - 1261
depuis huit mois lors des prélèvements. Cependant, malgré cette « non transmission », 17% 1262
de la population d’étude présentent des parasites circulants et seulement 2% de la population 1263
totale - inclus dans les 17% - ont une parasitémie asymptomatique supérieure à 500/µL 1264
correspondant à une catégorie F3 et plus (Trape, 1985). 1265
1266
A Dielmo, la cohorte se compose de 186 individus, âgés de 3,4 à 80 ans, avec une moyenne 1267
d’âge de 28,5 ans. Elle comprend 53% de femmes et 7,5% de la population d’étude présente 1268
une hémoglobine à profil drépanocytaire AS. Le nombre de personnes parasitées y est 1269
nettement plus élevé qu’à Ndiop, soit 46% des individus prélevés et 13% de la population 1270
d’étude présente un portage asymptomatique lié à une parasitémie de type F3 et plus. La 1271
transmission du paludisme y est pérenne et le taux d’inoculation nettement plus élevé. 1272
1273
Ces deux cohortes ont donc des profils très similaires en terme de recrutement mais des 1274
profils immunologiques différents. Tous les sérums de cette cohorte seront étudiés vis-à-vis 1275
de leurs réponses naturelles reconnaissant les candidats vaccins MSP1p19, MSP4p20, MSP5 1276
et R23-GST (études ELISA). Les données épidémiologiques correspondantes à ces différents 1277
prélèvements ont été colligées, grâce au suivi longitudinal des populations d’étude. 1278
1279
Le temps de suivi exact de chaque villageois est connu pour les 5,5 mois d’étude, afin de 1280
pouvoir réaliser les analyses épidémiologiques. 1281
1282
Les caractéristiques principales des différentes cohortes (effectifs, moyennes d’âges) sont 1283
résumées au tableau VI. La sous cohorte pour les tests fonctionnels est explicitée dans le 1284
paragraphe suivant. 1285
1286
1287
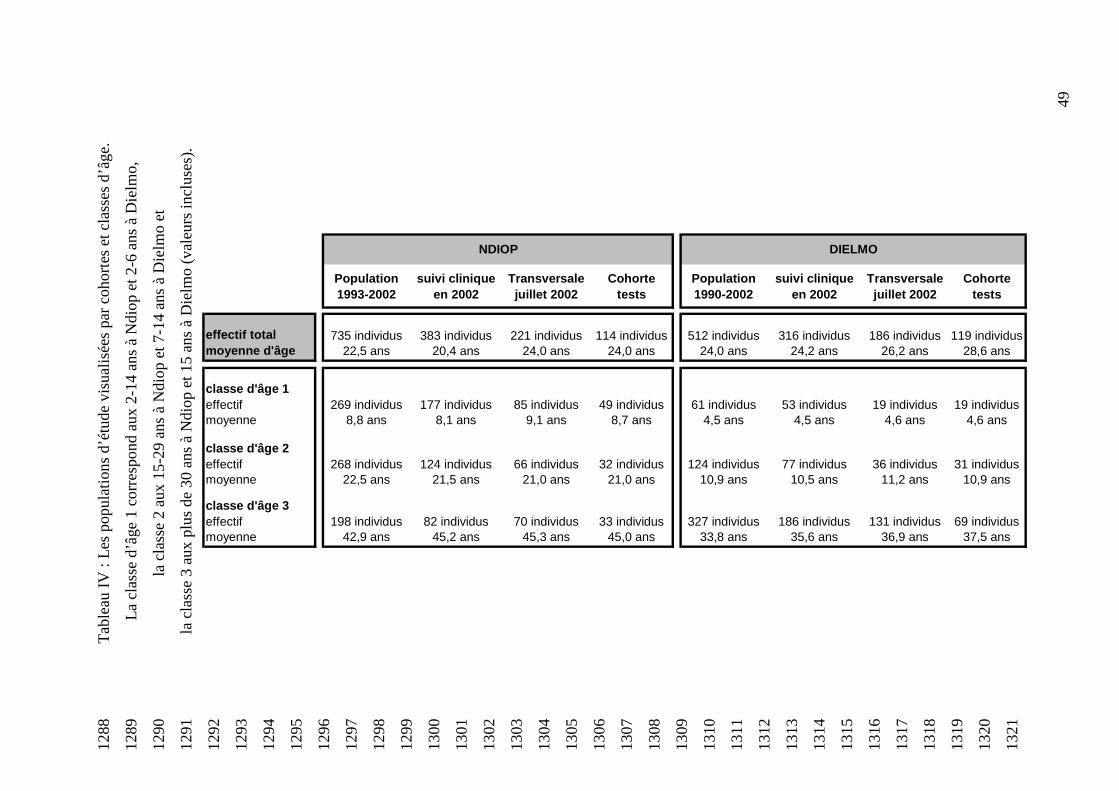
49
Population 1993-2002
suivi clinique en 2002
Transversale juillet 2002
Cohorte tests
Population 1990-2002
suivi clinique en 2002
Transversale juillet 2002
Cohorte tests
effectif total moyenne d'âge
735 individus 22,5 ans
383 individus 20,4 ans
221 individus 24,0 ans
114 individus 24,0 ans
512 individus 24,0 ans
316 individus 24,2 ans
186 individus 26,2 ans
119 individus 28,6 ans
classe d'âge 1 effectif moyenne
269 individus 8,8 ans
177 individus 8,1 ans
85 individus 9,1 ans
49 individus 8,7 ans
61 individus 4,5 ans
53 individus 4,5 ans
19 individus 4,6 ans
19 individus 4,6 ans
classe d'âge 2 effectif moyenne
268 individus 22,5 ans
124 individus 21,5 ans
66 individus 21,0 ans
32 individus 21,0 ans
124 individus 10,9 ans
77 individus 10,5 ans
36 individus 11,2 ans
31 individus 10,9 ans
classe d'âge 3 effectif moyenne
198 individus 42,9 ans
82 individus 45,2 ans
70 individus 45,3 ans
33 individus 45,0 ans
327 individus 33,8 ans
186 individus 35,6 ans
131 individus 36,9 ans
69 individus 37,5 ans
NDIOP DIELMO
Tab
leau
IV :
Les
popu
latio
ns d
’étu
de v
isua
lisée
s pa
r co
hort
es e
t cla
sses
d’â
ge.
1288
La
clas
se d
’âge
1 c
orre
spon
d au
x 2-
14 a
ns à
Ndi
op
et 2
-6 a
ns à
Die
lmo,
12
89
la c
lass
e 2
aux
15-2
9 an
s à
Ndi
op e
t 7-1
4 an
s à
Die
lmo
et
1290
la c
lass
e 3
aux
plus
de
30 a
ns à
Ndi
op e
t 15
ans
à D
ielm
o (v
aleu
rs in
clus
es).
12
91
12
92
12
93
12
94
12
95
12
96
12
97
12
98
12
99
13
00
13
01
13
02
13
03
13
04
13
05
13
06
13
07
13
08
13
09
13
10
13
11
13
12
13
13
13
14
13
15
13
16
13
17
13
18
13
19
13
20
13
21

50
1.4) Les sous cohortes d’étude et leur mode de sélection 1322
1323
1.4.1) Cohortes pour les tests fonctionnels 1324
1325
Tous les individus présents dans la double transversale ne nous permettent pas de réaliser une 1326
analyse statistique prenant en compte la morbidité en raison de leurs absences prolongées au 1327
niveau du site d’étude. Les personnes absentes plus de 30 jours consécutifs sur les 5,5 mois 1328
suivant le prélèvement ont été exclues de la sous cohorte d’analyses fonctionnelles. 1329
1330
Ensuite, la cohorte d’étude a été équilibrée pour les tests fonctionnels de façon à avoir au 1331
minimum 30 individus par tranche d’âge - sauf pour les plus jeunes de Dielmo qui est une 1332
classe naturellement limitante avec uniquement 19 enfants -. Ce nombre de 30 a été retenu 1333
pour fournir aux analyses statistiques la « puissance » nécessaire aux différents modèles. 1334
1335
Nous avons également équilibré les cohortes afin qu’elles soient les plus représentatives 1336
possible de la population. Pour cela, nous avons pris en compte différentes caractéristiques 1337
essentielles : 1338
� la répartition globale de la population sénégalaise. 42% des Sénégalais ont moins de 1339
15 ans selon l’INED (Institut National des Etudes Démographiques). Dans nos sous cohortes 1340
d’étude, 43% ont moins de 15 ans à Ndiop et 43,5% à Dielmo. 1341
� les moyennes d’âge observées dans les villages. Toutes les personnes ayant été inclues 1342
dans l’étude depuis son commencement dans les années 90 ont été prises comme référence. 1343
Le profil d’une population type a été déterminé à partir de leur date de naissance. 1344
* A Ndiop, les moyennes d’âge obtenues sont de 8,8 ans pour la classe 1 (2-14 ans), 22,5 ans 1345
pour la classe 2 (15-29 ans) et 42,9 ans pour la classe 3 (≥ 30 ans). La moyenne d’âge globale 1346
est de 22,5 ans. Dans notre sous cohorte d’étude (114 personnes), nous arrivons à 8,7 ans pour 1347
la classe 1, 21,0 ans pour la classe 2 et 45,0 ans pour la classe 3. Cette dernière se démarque 1348
de la référence choisie. Cela s’explique par le fait que la moyenne d’âge de la classe 3 pour la 1349
population réellement suivie cliniquement en 2002 est de 45,2 ans et que celle pour la 1350
population prélevée lors de la transversale est de 45,3. La moyenne d’âge générale est relevée 1351
à 24,0 ans. 1352
** A Dielmo, nous obtenons une moyenne d’âge de la classe 1 (2-6 ans) égale à 4,5 ans, la 1353
classe 2 (7-14 ans) à 10,9 ans et la classe 3 (≥ 15 ans) à 33,8 ans, soit une moyenne d’âge 1354
globale de 24 ans. Dans notre sous cohorte d’étude (119 personnes), la moyenne d’âge de la 1355
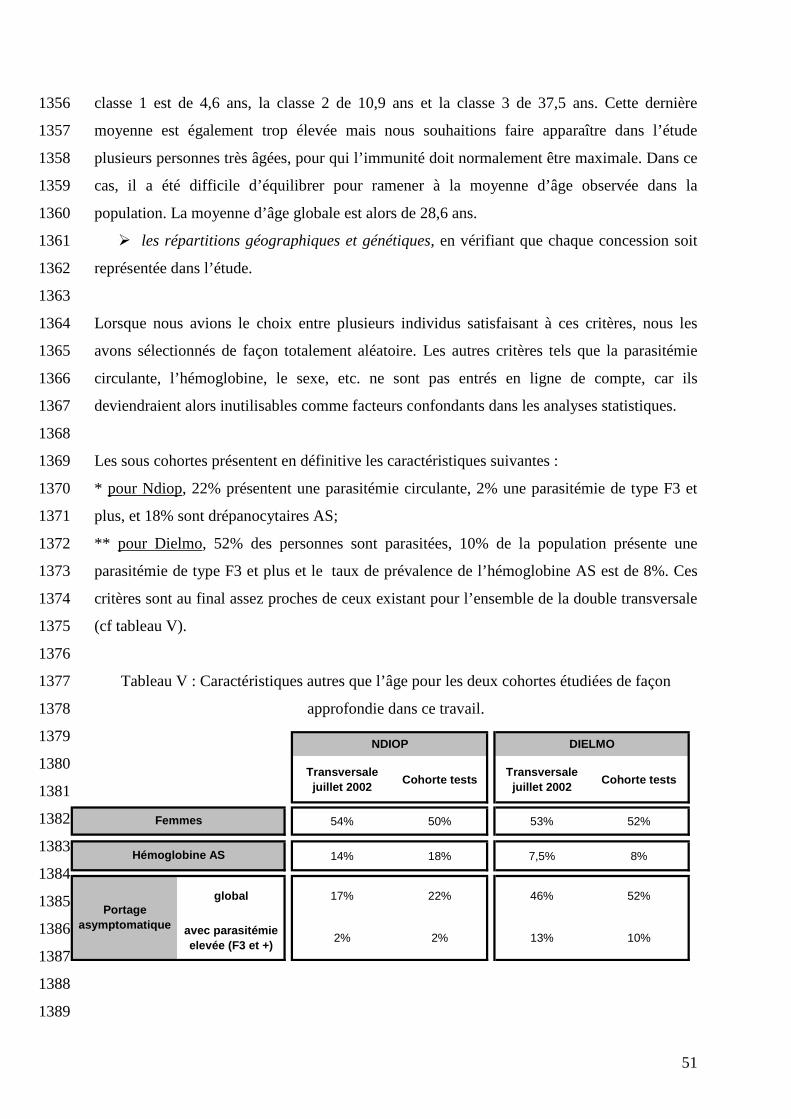
51
Transversale juillet 2002
Cohorte testsTransversale juillet 2002
Cohorte tests
54% 50% 53% 52%
14% 18% 7,5% 8%
global 17% 22% 46% 52%
avec parasitémie elevée (F3 et +)
2% 2% 13% 10%
Portage asymptomatique
Femmes
Hémoglobine AS
NDIOP DIELMO
classe 1 est de 4,6 ans, la classe 2 de 10,9 ans et la classe 3 de 37,5 ans. Cette dernière 1356
moyenne est également trop élevée mais nous souhaitions faire apparaître dans l’étude 1357
plusieurs personnes très âgées, pour qui l’immunité doit normalement être maximale. Dans ce 1358
cas, il a été difficile d’équilibrer pour ramener à la moyenne d’âge observée dans la 1359
population. La moyenne d’âge globale est alors de 28,6 ans. 1360
� les répartitions géographiques et génétiques, en vérifiant que chaque concession soit 1361
représentée dans l’étude. 1362
1363
Lorsque nous avions le choix entre plusieurs individus satisfaisant à ces critères, nous les 1364
avons sélectionnés de façon totalement aléatoire. Les autres critères tels que la parasitémie 1365
circulante, l’hémoglobine, le sexe, etc. ne sont pas entrés en ligne de compte, car ils 1366
deviendraient alors inutilisables comme facteurs confondants dans les analyses statistiques. 1367
1368
Les sous cohortes présentent en définitive les caractéristiques suivantes : 1369
* pour Ndiop, 22% présentent une parasitémie circulante, 2% une parasitémie de type F3 et 1370
plus, et 18% sont drépanocytaires AS; 1371
** pour Dielmo, 52% des personnes sont parasitées, 10% de la population présente une 1372
parasitémie de type F3 et plus et le taux de prévalence de l’hémoglobine AS est de 8%. Ces 1373
critères sont au final assez proches de ceux existant pour l’ensemble de la double transversale 1374
(cf tableau V). 1375
1376
Tableau V : Caractéristiques autres que l’âge pour les deux cohortes étudiées de façon 1377
approfondie dans ce travail. 1378
1379
1380
1381
1382
1383
1384
1385
1386
1387
1388
1389

52
1.4.2) La cohorte pour les études des réponses IgG (classe et sous classes) 1390
1391
Nous devions également réaliser une étude nous permettant de visualiser le profil des 1392
réponses IgG vis à vis des différents Ag envisagés. Cependant, en raison des individus ayant 1393
une parasitémie circulante pouvant traduire le décours d’une infection récente - ou une 1394
infection en cours -, et affectant dynamiquement les taux d’Ac mesurables, nous avons choisi 1395
d’étudier les réponses IgG sur une cohorte plus restreinte. Pour cela, nous avons sélectionné 1396
les 88 individus de la sous cohorte de Ndiop ne présentant pas de parasites circulants. 1397
1398
Cette cohorte de personnes non parasités comporte 16% d’individus AS et est composée 1399
d’individus de tous les âges, avec une moyenne d’âge de 23,0 ans. En dehors de la 1400
parasitémie, il n’y a donc pas eu de biais important pour les caractéristiques d’âge et 1401
d’incidence de porteurs d’Hb AS par rapport à la cohorte initiale. 1402
1403
2) Méthodologie des tests ELISA 1404
1405
2.1) Méthode de travail 1406
1407
La technique ELISA (Enzyme Linked ImmunoSorbent Assay) est une technique 1408
immunométrique qui est appliquée pour le dosage des immunoglobulines - classe et/ou sous-1409
classes - dirigées contre un Ag ou une espèce antigénique prédéfinie. L’Ag est tout d’abord 1410
adsorbé sur la phase solide que représente la plaque de 96 puits. Les Ac spécifiques de cet Ag 1411
vont se fixer dessus pendant la période d’incubation et des anti-immunoglobulines couplées à 1412
la peroxydase permettront ensuite de détecter leur présence et de déterminer par colorimétrie 1413
leur quantité. 1414
1415
2.1.1) Méthodologie ELISA pour l’analyse de prévalence des candidats vaccins 1416
1417
Les tests ELISA permettent d’étudier les prévalences des réponses Ac associées aux candidats 1418
vaccins testés et d’analyser celles-ci en fonction de différents paramètres épidémio-cliniques - 1419
zone d’endémie, âge, parasitémie -, ceci de manière indépendante de leur fonctionnalité. Les 1420
candidats vaccins utilisés ont été synthétisés à l’Institut Pasteur de Paris. Les Ag synthétiques 1421
MSP1p19, MSP4p20 et MSP5 ont été exprimés en Baculovirus par l’équipe du Dr Shirley 1422
Longacre dans le Laboratoire de Vaccinologie Parasitaire. La protéine recombinante R23 a été 1423

53
synthétisée dans E. coli par l’équipe du Dr Odile Mercereau-Puijalon dans l’Unité 1424
d’Immunologie Moléculaire des Parasites. Les analyses immunologiques des sérums ont été 1425
réalisées à l’Institut Pasteur de Dakar selon la méthodologie utilisée sur place et explicitée ci 1426
après. 1427
1428
Les tests sont réalisés sur des plaques Immulon 4HBX (Dynatech). Les Ag MSP1p19, 1429
MSP4p20 et MSP5 sont adsorbés via 100 µL d’une solution antigénique à 0,5 µg/mL dans du 1430
PBS, une nuit à 4°C. Pour R23, le coating est de 1,5 µg/mL avec en parallèle sur une plaque 1431
contrôle de la protéine du vecteur GST à 0,6 µg/mL - les résultats seront soustraits de ceux de 1432
GST-R23 -. La plaque est ensuite lavée avec du PBS Tween 0,1% et saturée pendant 1H à 1433
37°C avec 150 µL d’une solution de PBS Tween 0,1% BSA 5% (BSA fraction V pH7, Acros 1434
Organique) avant de déposer les sérums d’étude. Ceux-ci sont dilués au 1/200 dans du PBS 1435
Tween 0,1% BSA 1% azide 0,05% et déposés en duplicate (100 µL). Trois contrôles positifs 1436
et trois contrôles négatifs sont également déposés systématiquement sur chaque plaque, à la 1437
même dilution. Après 1H d’incubation à 37°C et un nouveau cycle de lavage, 100 µL d’anti-1438
immunoglobulines couplées à la peroxydase (HRP) (Cappel) sont déposées au 1/6000 dans 1439
une solution de PBS Tween 0,1% BSA 1%. Après à nouveau 1H d’incubation à 37°C et un 1440
cycle de lavage intense, 100 µL/puits d’une solution d’ortho-toluidine/eau oxygénée préparée 1441
extemporanément - 12 µL d’H2O2 pour 30mL de solution d’ortho-toluidine - sont ajoutés et 1442
les plaques sont laissées dans l’obscurité, à température ambiante, pendant 15 min avant que 1443
la coloration ne soit stoppée à l’aide de 50 µL d’acide sulfurique 4N. 1444
1445
La lecture s’effectue à 450 nm (référence 620 nm). Les résultats sont exprimés à la fois en 1446
ratio de DO : 1447
rDO = (Moyenne DO sérum) / (Moyenne DO négatifs testés) 1448
1449
et en unités arbitraires : 1450
U = (Moyenne DO sérum) / (Moyenne DO SHI) * 100 1451 1452
Le Sérum Hyper Immun (SHI) est l’un des trois contrôles positifs. Il est constitué d’un 1453
mélange de trente sérums d’habitants de Dielmo et représente le « contrôle standard » dans 1454
toutes nos études. 1455
1456

54
Un sérum est considéré comme répondeur à un Ag donné si son ratio de DO est supérieur ou 1457
égal à 2, ce qui correspond d’une manière générale à : 1458
DO du sérum naïf (témoin négatif) + 2 déviations standard (SD). 1459
. 1460
2.1.2) Méthodologie ELISA pour l’analyse des réponses IgG contre le mérozoïte (classe et 1461
sous classes) – méthode du Dr Remarque 1462
1463
Nous avons également réalisé des tests ELISA afin de déterminer le profil IgG des réponses 1464
contre les Ag « totaux » du mérozoïte. Les préparations mérozoïtaires utilisées sont les 1465
mêmes que celles impliquées dans le test de chimiluminescence (cf § 5.1 - Purification des 1466
mérozoïtes). Les tests ont été réalisés à l’Institut Pasteur de Paris, sur une cohorte restreinte 1467
aux habitants de Ndiop non parasités (cf § 1.2.2 - la cohorte pour les tests IgG) et suivant la 1468
méthodologie utilisée sur place. Elle est différente du point de vue de la révélation, expliquant 1469
des DO brutes plus élevées. 1470
1471
Les merozoites, après 3 cycles de congélation – décongélation pour permettre une bonne lyse 1472
et donc une bonne libération des Ag, sont dosés à l’aide du Kit BCA (Pierce). Ils sont ensuite 1473
dilués à 10 µg de protéines/mL dans du PBS et déposés sur des plaques MaxiSorp (Nunc) - 1474
100 µL par puits, une nuit à 4°C -. Après les cycles de lavage et celui de saturation - 1475
150 µL/puits d’une solution de PBS Tween 0,1% BSA 5% sont ajoutés pendant 1H30 à 1476
température ambiante -, 100 µL de sérums par puits sont mis à incuber pendant 1H30 à 1477
température ambiante aux dilutions 1/200, 1/400 et 1/800 ou 1/5000, 1/10000 et 1/20000 1478
respectivement pour le dosage des IgG1 et IgG3 ou des IgG totales. Sur chaque plaque, le 1479
contrôle positif SHI est titré en duplicate de la dilution 1/200 à 1/12800 pour les IgG1 et IgG3 1480
ou 1/5000 à 1/320000 pour les IgG totales. Les IgG2 et IgG4 sont testées à une seule dilution, 1481
1/100, afin de détecter d’éventuels répondeurs et non pas de quantifier leur réponse. Les 1482
isotypes seront détectés par leur conjugaison avec des Ac monoclonaux de souris spécifiques 1483
des différentes sous classes d’IgG humaines et couplés à la peroxydase (Southern Biotech) - 1484
100 µL, dilution 1/1000, 1H30 à température ambiante -. Pour les IgG totales, elles sont 1485
révélées par des IgG anti-humaines conjuguées à la peroxydase - 50 µL, dilution 1/1000, 1486
1H30 à température ambiante -. La révélation consiste en l’ajout de 100 µL d’une solution 1487
d’OPD préparée extemporanément (10mL d’H2O, 2 tablettes d’OPD, 10µL d’H2O2). Les 1488
plaques sont laissées 10 min dans l’obscurité à température ambiante avant que la coloration 1489

55
ne soit stoppée par ajout de 50 µL d’acide chlorhydrique 3N. Elles sont ensuite lues à la 1490
double longueur d’onde 490 – 650 nm. 1491
Des titres arbitraires liés aux titres en Ac sont ensuite extrapolés par rapport à la courbe de 1492
régression logistique du SHI en utilisant le programme de standardisation OMS mis au point 1493
par le Dr Ed Remarque. Ces unités « titres » sont comparables d’une plaque à l’autre pour le 1494
même type de réponses. Il est important de préciser que dans ces calculs, les valeurs obtenues 1495
pour les IgG1 et les IgG totales ne sont donc pas comparables entre elles, le titre de référence 1496
étant différent. 1497
1498
3) Méthodes de purification des anticorps pour les analyses ciblant les antigènes 1499
1500
3.1) Déplétions de sérums sur résine TALON 1501
1502
Des sérums forts répondeurs en ELISA pour les candidats vaccins étudiés ont été choisis afin 1503
d’être déplétés d’un ou de plusieurs types d’Ac spécifiques. Des déplétions simples ou 1504
multiples contre MSP1p19, MSP4p20 et MSP5 ont été réalisées sur résine TALON (Metal 1505
affinity, Ozyme). Ces déplétions spécifiques ont pour but de déterminer, par comparaison 1506
avec le sérum original, l’effet sur les tests fonctionnels des Ac spécifiques des différents 1507
candidats vaccins. 1508
1509
100 µL de sérum sont dilués au tiers dans du PBS - cette étape n’existe pas si le sérum a déjà 1510
été déplété, la dilution ayant déjà eu lieu lors de la déplétion précédente -. Ils sont ajoutés aux 1511
50 µg de la construction vaccinale choisie. La fixation dure 30 min, via agitation sur robot 1512
roue à température ambiante. Pendant ce temps, 400 µL de résine TALON en solution sont 1513
tassées et lavées avec i) de l’eau, ii) de la solution A1 (20mM TrisHCl, 500mM NaCl) et iii) 1514
du PBS. La solution Ag-Ac est ensuite ajoutée aux 200 µL de résine équilibrée recueillie et 1515
mélangée à cette dernière sur robot roue à température ambiante pendant 3H. Après 5 min de 1516
centrifugation à grande vitesse, le sérum déplété - et ainsi dilué au tiers - est obtenu. Les IgG 1517
spécifiques accrochées sur la colonne ne peuvent pas être isolées en bon état. En effet, elles 1518
sont forcément décrochées avec leur Ag car la liaison TALON – ‘HisTag de la construction 1519
vaccinale’ est très fragile. De plus, une séparation du complexe après décrochage de la résine 1520
entraîne systématiquement une dénaturation des Ac, raison pour laquelle nous n’obtenons 1521
avec cette technique que des sérums déplétés. 1522

56
R45 ne peut pas être déplété avec cette méthode car il posséde une GST et non pas un HisTag. 1523
Cependant, la première étape permet une saturation des Ac avec leur Ag et cette liaison est 1524
suffisamment solide pour qu’ils ne puissent plus exercer leur fonction. Cet effet a été vérifié 1525
par test ELISA. 1526
1527
3.2) Purifications d’IgG sur protéine G 1528
1529
Pour étudier l’impact des sérums déplétés, il est important de valider les tests fonctionnels 1530
utilisés comme étant bien essentiellement - qualitativement et éventuellement 1531
quantitativement - dépendants des IgG. Pour ce faire, il est nécessaire de purifier des IgG 1532
totales et de les tester comparativement aux sérums dans les tests fonctionnels. 1533
1534
Les IgG totales ont été purifiées sur protéine G (résine Ultralink® immobilized, Pierce). La 1535
protéine A n’a pas pu être utilisée car elle ne fixe pas les IgG3 humaines et ne purifie pas la 1536
totalité des IgG1 humaines. 0,4 mL de sérum sont dilués au 1/5 dans Immunopure® G IgG 1537
binding buffer (solution d’équilibration, Pierce) puis passés 3 fois sur 2 mL de résine tassée et 1538
équilibrée. Le sérum déplété est récupéré dilué, puis la colonne contenant les IgG est lavée 1539
avec 15 volumes de solution d’équilibration. Les IgG sont ensuite éluées avec Immunopure® 1540
elution buffer (solution d’élution, Pierce), neutralisées avec 1M TrisHCl pH 8.0, dialysées 1541
dans du PBS, et concentrées à 0,4 mL (Amicon Ultra (5,000 MWCO): Millipore). Les sérums 1542
déplétés sont également ramenés à 0,4 mL par dialyse/concentration. Il n’y a donc au final 1543
aucun effet de dilution consécutif à ce traitement. 1544
1545
4) Culture de Plasmodium falciparum 1546
1547
4.1) Conditions de culture 1548
1549 Pour réaliser les tests, il est impératif d’avoir au préalable des parasites P. falciparum - souche 1550
Palo Alto alias FCR3 - maintenus in vitro en culture continue. Compte tenu des conditions 1551
techniques générales - pas d’approvisionnement en gaz et source de sérum humain peu 1552
accessible de manière continue -, la méthode opérationnelle a été l’utilisation de GR O+, en 1553
présence d’albumax et selon la méthode de la « candle jar » (Jensen, 1977). Les cultures ne 1554
sont donc pas gazées mais incubées dans une enceinte où trois bougies de type « chauffe-1555

57
plat » - sans fumée - sont allumées chaque fois que l’enceinte est ouverte de façon à faire 1556
baisser le taux d’oxygène présent à une valeur inférieure à 17 %. 1557
1558
Le milieu de culture utilisé est du RPMI-Albumax 0,5%, complémenté avec des antibiotiques 1559
(gentamicine) et des compléments nutritionnels. 2L de RPMI 1640 médium (Sigma) sont 1560
complétés avec 4g de glucose, 10g d’albumax II (Gibco), 20mL d’hépès, de L.glutamine, 1561
d’acides aminés non essentiels et de pyruvate de sodium, 4 mL d’hypoxanthine 1562
(C = 500 µg/mL) et d’acide orotique et 1 mL de gentamicine (C = 50 mg/mL). Le milieu de 1563
culture est filtré puis stocké à -20°C, jusqu’à utilisation. 1564
1565
Une culture en sérum présente de nombreuses difficultés à l’Institut Pasteur de Dakar. En 1566
effet, les sérums obtenus sur place peuvent comporter des antipaludéens et un certain nombre 1567
d’individus ne sont pas naïfs vis à vis de P. falciparum. Il est donc préférable d’importer le 1568
sérum de France, ce qui pose de nombreuses autres contraintes supplémentaires. 1569
Dans la méthode préalablement validée impliquant l’Albumax (Diouf, 2002), la croissance 1570
parasitaire, bien que moins efficace, reste correcte et le milieu exempt d’Immunoglobulines 1571
présente alors un gros avantage. 1572
1573
Les cultures sont suivies quotidiennement sur frottis avec coloration par Giemsa : fixation au 1574
méthanol puis coloration au Giemsa diluée au 1/10 dans l’eau, pendant 20 min, et 1575
visualisation sur microscope à immersion objectif 100. Le milieu de culture est renouvelé tous 1576
les jours. Les cultures sont réalisées dans des flasques de 50 ou 250 mL contenant 1577
respectivement 0,5 et 2,5 mL de GRp dans 7 et 27 mL de milieu. Dans ces conditions, des 1578
parasitémies maximales de 10% peuvent être atteintes. Les GR O+, après lavage dans du 1579
RPMI 1640, sont utilisables pendant 1 mois, à condition d’être convenablement stockés à 1580
4°C. 1581
1582
4.2) Conditions particulières de culture pour D10-PcMEGF et pour D10-PfM3’ 1583
1584 Afin d’étudier l’impact de PfMSP1p19, nous avons utilisé les souches transgéniques 1585
D10-PcMEGF et D10-PfM3’. Elles ont été cultivées à l’Institut Pasteur de Paris, selon les 1586
conditions prédéfinies par l’équipe de B.S. Crabb. Les cultures sont gazées avec un mélange 1587
de composition : 2% O2, 5,5% CO2 et 92,5% N2 et cultivées dans un milieu comportant à la 1588
fois du sérum et de l’albumax. 1589

58
1590
Le milieu de culture est préparé en deux temps : i) pour 2L de RPMI 1640 medium, ajout de 1591
25 mM d’hépès, 200 µM d’hypoxanthine et 0,8 mL de gentamicine (C = 50 mg/mL), filtration 1592
puis stockage à 4°C, ii) au moment de l’utilisation, complémentation de 250 µL de milieu 1593
avec 12,5 mL d’albumax et de sérum décomplémenté et 14,5 mL d’une solution de NaHCO3 1594
à 3,6%. Après filtration, 125 ng/mL de pyriméthamine dans le DMSO sont ajoutés au milieu 1595
de culture. Ce dernier est alors stable pendant 4 à 5 jours uniquement. La pyriméthamine est 1596
essentielle pour empêcher les dérives génétiques et conserver la souche transgénique telle 1597
quelle. 1598
1599
Des essais d’adaptation ont montré que les 12,5 mL de sérum pouvaient être remplacés par 1600
12,5 mL d’albumax - soit 25 mL d’albumax au total -, sans affecter la croissance parasitaire. 1601
En revanche, le taux d’oxygène joue un rôle important et la méthode en « candle jar » réduit 1602
fortement la multiplication des parasites transgéniques. 1603
1604
4.3) Décongélation, synchronisation, congélation 1605
1606
Après décongélation de l’aliquot de la souche parasitaire à 37°C, 0,2 volumes de NaCl 12% 1607
sont ajoutés au goutte à goutte en homogénéisant. Après 5 min, 10 volumes de NaCl 1.6% 1608
sont rajoutés, toujours au goutte à goutte, en homogénéisant régulièrement. La solution 1609
parasitaire est ensuite centrifugée pendant 5 min à 400xg puis lavée avec du RPMI 1640 avant 1610
d’être mise en culture dans les conditions habituelles. Les parasites mettent en moyenne une 1611
semaine avant d’arriver à une vitesse de croissance stable et reproductible. 1612
1613
Lorsque la culture présente une très grande majorité d’anneaux, elle peut être synchronisée à 1614
l’aide de d’une solution de D-sorbitol - 5g dans 100mL d’eau -. 5 volumes de D-sorbitol sont 1615
ajoutés au culot parasitaire et la solution est ensuite laissée pendant 5 min à 37°C. Après une 1616
centrifugation de 5 min à 400xg et un cycle de lavage avec du RPMI 1640, le culot parasitaire 1617
est remis en culture. Seules les formes jeunes sont encore viables et la croissance parasitaire 1618
reprendra de façon synchrone. 1619
1620
Il est possible de congeler la culture quand la parasitémie est très élevée et comporte une 1621
grande majorité de formes jeunes. Pour cela, une solution de glycerolyte est utilisée - 57g de 1622
glycerol, 10mL de PBS 10x, 1,6g de lactate de sodium, 0,03g de chloride de potassium, le 1623

59
tout dans 100mL d’eau -. 0,4 volumes de glycérolyte sont ajoutés au goutte à goutte en 1624
homogénéisant. Après 5 min à température ambiante, ajout de 1,2 volumes de glycérolyte 1625
supplémentaire, au goutte à goutte, avec agitation. 5 min plus tard, un nouvel ajout de 2,4 1626
volumes de glycérolyte est réalisé. Les aliquots de 1mL sont stockés dans l’azote liquide. 1627
1628
5) Méthodologie pour le test de chimiluminescence 1629
1630
Le premier test fonctionnel à avoir été développé dans ce travail a été le test de 1631
chimiluminescence. Il repose sur une réaction de phagocytose immune des mérozoïtes 1632
provenant des cultures parasitaires par des PNN préalablement purifiés à partir de sang de 1633
donneurs. 1634
1635
5.1) Purification des mérozoïtes 1636
1637
Plusieurs méthodes de purification de mérozoïtes ont été étudiées : par flottaison sur Percoll 1638
ou par simple ou double centrifugation. Nous avons finalement retenu la méthode la plus 1639
simple, à savoir une centrifugation unique (cf Résultats §B1.1). 1640
1641
Les cultures parasitaires dont le culot fait 2,5 mL et dont la parasitémie est supérieure à 5% 1642
peuvent permettre la récolte de mérozoïtes, surtout si elles sont synchrones et qu’elles sont au 1643
stade de réinvasion ou juste après celui-ci. La culture est alors centrifugée pendant 5 min à 1644
400xg pour séparer les GRp - qui pourront être remis en culture - du milieu de culture 1645
contenant les mérozoïtes libérés n’ayant pas encore réenvahi d’autres GR. Celui-ci est alors 1646
centrifugé pendant 20 min à 1500xg de manière à obtenir un culot de couleur marron qui 1647
correspond à la « préparation mérozoïtaire » - mérozoïtes entiers et fragmentés, hémozoïne et 1648
débris cellulaires -. Ce culot est alors isolé de façon la plus concentrée possible et stocké à -1649
20°C jusqu’à utilisation. Cette technique présente l’énorme avantage d’être non destructrice 1650
pour la culture parasitaire. 1651
1652
5.2) Purification des polynucléaires 1653
1654
Les PNN sont excessivement fragiles et ont une demi-vie très courte en dehors du corps 1655
humain. Il est donc impératif de travailler avec du sang très frais. Les prélèvements veineux 1656
sont réalisés dans des tubes EDTA K3 au moment même du test, sur des volontaires venant 1657

60
effectuer des prises de sang à l’Institut Pasteur de Dakar. 6 tubes, de 6 donneurs différents et 1658
comportant chacun 6 mL de sang sont traités et rassemblés de manière à permettre l’analyse 1659
d’une quarantaine de sérums d’étude. Les PNN sont purifiés par extraction sur gradient ficoll-1660
hystopaque de densité 1077 (Sigma), ce choix étant apparu plus judicieux que la technique 1661
basée sur la purification par centrifugation à l’interface de 2 couches de ficoll de densité 1662
différente 1077 et 1119 (cf Résultats § B1.2). 1663
1664
Le sang frais est centrifugé pendant 10 min à 800xg afin que les leucocytes se déposent à 1665
l’interface GR – sérum. Cette interface est collectée et diluée dans 2 mL de solution saline 1666
tamponnée de Hanks (HBSS, Sigma) et déposée sur 5 mL de ficoll 1077. Une centrifugation 1667
pendant 30 min à 400xg à température ambiante est réalisée de manière à séparer les 1668
monocytes (interface HBSS – ficoll) des PNN (interface ficoll – GR). L’interface contenant 1669
les PNN est ensuite traitée avec une solution de lyse - 8,32 g/L de NH4Cl, 0,8 g/L de 1670
bicarbonate de sodium et 0,043 g/L d’EDTA, pendant 8 min à 4°C puis centrifugation 5 min à 1671
400xg à 4°C - de façon à éliminer les GR contaminants. Les PNN sont ensuite lavés à deux 1672
reprises avec du HBSS et les PNN toujours vivants sont dénombrés à l’aide d’une solution de 1673
bleu trypan diluée au 1/5 dans du PBS. 5 µL de la solution de PNN de volume total 1 mL sont 1674
diluées dans 500 µL de bleu trypan/PBS. Les PNN observables via l’objectif 10 d’un 1675
microscope sont dénombrés pour 9 carreaux distincts de la grille de comptage de la lame. Le 1676
nombre de PNN obtenu, multiplié par 106, fournit la quantité de PNN par mL. 1677
Le culot de PNN est enfin ramené à 5.107 PN/mL. Ils sont immédiatement utilisés dans 1678
l’expérience de chimiluminescence. 1679
1680
5.3) Conditions de réalisation du test de chimiluminescence 1681
1682
La chimiluminescence est mesurée en utilisant des plaques blanches opaques de 96 puits 1683
(Berthold) et un luminomètre passeur de plaque MicroLumat Plus (Berthold). Le processus de 1684
lecture est géré par le logiciel WinGlow. La chimiluminescence est exprimée en unités 1685
relatives de luminescence (rlu) et le bruit de fond se situe généralement à une valeur d’environ 1686
15 rlu. 1687
Pour chaque expérience, une quarantaine de sérums sont déposés avec des contrôles positifs et 1688
négatifs systématiques. Le SHI, contrôle positif standard « fort », est déposé en première et 1689
dernière ligne. Chaque puits sera lu pendant 1 seconde, toutes les minutes, pendant 1h. 1690

61
Les puits contiennent initialement 40 µL de mérozoïtes opsonisés au préalable pendant un 1691
minimum de 30 min avec 10 µL de sérum ou IgG. En dernier lieu, 100 µL de PNN soit 5.106 1692
PN et 100 µL d’une solution stock de luminol (4-aminophtalhydrazide, Sigma) diluée au 1693
1/100 dans du PBS - solution stock à 2 mg/mL dans du DMSO, stockée à -20°C jusqu’à 1694
utilisation - sont ajoutés. La lecture débute juste après l’addition des PNN et du luminol, qui 1695
s’effectue le plus rapidement possible. 1696
1697
Les résultats sont standardisés en fonction de la valeur maximale moyenne du SHI selon le 1698
calcul d’« ADPm » (Phagocytose des mérozoites Dépendante des Ac, sigle en anglais) : 1699
1700 ADPm = (rlu max éch / rlu max SHI) x 1000 1701
1702
6) Méthodologie pour les tests d’inhibition de culture 1703
1704
Nous nous sommes également intéressés à un deuxième test fonctionnel à savoir l’inhibition 1705
de culture : GIA (sigle en anglais). Celui-ci souligne un autre aspect fonctionnel des Ac : la 1706
destruction parasitaire issue de leur action bloquante directe. Aucune autre cellule du système 1707
immunitaire - Complément, cellules phagocytaires - n’est présente. 1708
1709
6.1) Conditions de réalisation du test d’inhibition de culture 1710
1711
Les tests d’inhibition de culture commencent toujours avec des formes âgées synchrones, de 1712
préférence schizontes, et se terminent au même stade de développement, un cycle après. 1713
Cependant les souches parasitaires cultivées en laboratoire n’ont plus des cycles naturels de 1714
48H mais plutôt de 45H pour PA et 42H pour 3D7. La durée du test doit donc être compris 1715
entre 40H et 44H selon la souche utilisée. Les tests se déroulent dans des plaques stériles de 1716
96 puits, avec des conditions similaires à celles utilisées pour la culture parasitaire de routine : 1717
même milieu de culture et méthode en « candle jar ». Pendant toute la durée du test, le 1718
contenu des puits reste inchangé, on ne renouvelle ni le milieu ni le sérum étudié. 1719
1720
Les conditions pour la réalisation d’un test en micro-méthode sont issues des mises au point 1721
réalisées au laboratoire pour un test en macro-méthode (Diouf, 2002). Chaque puits contient 2 1722
µL de GRp de parasitémie 0,3%, 20 µL de sérum ou IgG à tester et 110 µL de RPMI-Alb 1723

62
0,5%. L’hématocrite finale est donc de 1,5% et la quantité de sérum d’étude de 15%. Ces 1724
conditions ont été utilisées pour l’étude en cytofluorométrie 1725
1726
Les conditions standardisées par l’équipe de C.A. Long au NIH lors de la mise au point du 1727
test avec lecture par dosage de la pLDH (Zhou, 2005) sont différentes de celles définies au 1728
laboratoire. Dans cette nouvelle méthode, chaque puits contient 1 µL de GRp de parasitémie 1729
0,3%, 20 µL de sérum ou IgG à tester et 79 µL de milieu de culture. L’hématocrite finale est 1730
donc de 1% et la quantité de sérum d’étude de 20%. Ces conditions ont été utilisées pour 1731
tenter de reproduire la méthode avec détection de la pLDH. 1732
1733
6.2) Lecture par cytofluorométrie 1734
1735
Une des méthodes de lecture du test de GIA étudiée est la cytométrie en flux. Afin de lire les 1736
plaques de GIA au FACScalibur, il est nécessaire d’introduire un fluorochrome en fin 1737
d’expérience. Le fluorochrome choisi après étude comparative a été l’hydroéthidine (HE, 1738
Polysciences) (cf Résultats § C.1.1). 1739
1740
Les plaques sont centrifugées 5 min à 400xg de façon à pouvoir éliminer le milieu de culture 1741
et le sérum d’étude. Le culot de GRp est ensuite incubé pendant 20 min à 37°C avec 200 µL 1742
d’une solution d’HE - 10 mg/mL dans du PBS, stockée à -20°C - diluée extemporanément au 1743
1/1000 dans du PBS. Deux lavages au PBS précèdent la lecture au FACScalibur pour 1744
supprimer tout excédent de fluorochrome. La fluorescence par HE apparaît en FL2. 1745
1746
Les réglages utilisés sont vérifiés avant chaque lecture afin d’être modifiés si besoin - à cause 1747
du vieillissement du laser par exemple -. Les plaques sont lues de façon automatisée grâce au 1748
logiciel Plate Manager - prélèvement automatisé de 50 µL par puits de GRp/PBS - et 1749
analysées en dot-plot et histogramme par le logiciel Cell Quest Pro. 1750
Les GR sains se trouvent compris dans la zone définie par 0 ≤ FL1 < 1.101 et 0 ≤ FL2 < 1751
1.101. Les GRp ont une valeur de FL2 comprise entre 1.101 et 103. 1752
1753
En préréglant ces zones d’étude, le logiciel affiche directement le pourcentage de parasitémie, 1754
à partir duquel il est possible de définir le pourcentage d’inhibition : 1755
1756 % inhib = 100 x (% GRp - % éch) / % GRp 1757

63
et l’index de positivité : 1758
IP = (% éch - % SAB) / (% SHI - % SAB) 1759
1760
6.3) Lecture par dosage de la pLDH 1761
1762
Une autre méthode de lecture des tests de croissance parasitaire a été évaluée, il s’agit d’une 1763
méthode colorimétrique dosant un composant enzymatique du parasite, la Plasmodium lactate 1764
déshydrogénase (pLDH). Pour ce faire, en fin d’expérience de GIA, 50 µL de la solution 1765
sanguine contenue dans chaque puits sont transférés dans 250 µL de PBS froid, préalablement 1766
déposés dans une plaque Immulon 4HBX. Cette plaque est ensuite centrifugée 10 min à 1767
800xg et à 4°C. Après avoir supprimé 240 µL du surnageant, les plaques sont congelées 1768
pendant 1 journée. 1769
1770
Pour réaliser ensuite le dosage et la révélation de celui-ci, il est nécessaire d’avoir au 1771
préalable les solutions stocks suivantes : 1772
* TrisHCl 1M pH 8 : 6,057 g de Trizma Base dans 35 mL d’eau, ajusté à pH 8 avec du HCl 1773
« fumant » (11,3N) et complété à 50 mL avec de l’eau - stockage à température ambiante si 1774
besoin -. 1775
* Tampon LDH : 50 mL de TrisHCl 1M pH 8, 450 mL d’eau, puis 2,8 g de sodium-L-lactate 1776
et enfin 1,25 mL de Triton X-100, sous agitation pendant 30 min et stockage par aliquots de 1777
50 mL à -20°C 1778
* Solution de 3-AcetylPyridine Adenine Dinucleoide (APAD) : 25 mg d’APAD dans 2,5mL 1779
d’eau, stockage par aliquots de 50 µL à -20°C 1780
* Solution de Diaphorase : 300 unités de diaphorase dans 6 mL d’eau, stockage par aliquots 1781
de 200 µL à -20°C 1782
Ces 3 solutions se conservent plusieurs mois à -20°C. Elles servent à la réalisation des deux 1783
solutions nécessaires au dosage de la pLDH : 1784
* la solution de NBT, qui ne se conserve que 3 semaines, à 4°C et à l’abri de la lumière. Elle 1785
est composée de 50 mL de LDH buffer et 1 tablette de NBT - 15 min à température ambiante, 1786
avec homogénéisation douce -, 1787
* la solution de révélation préparée extemporanément composée de 10 mL de NBT, 50 µL 1788
d’APAD et 200 µL de diaphorase par plaque. 1789
1790

64
Quand les plaques et les réactifs se trouvent être à température ambiante, la solution de 1791
révélation est réalisée et est déposée dans chaque puits d’étude (100 µL). Après une 1792
centrifugation rapide à 1500xg pour chasser l’air, les plaques sont laissées 10 min à l’abri de 1793
la lumière sur plateau agitateur. La lecture se réalise ensuite à 650 nm (étude à Paris) ou à 1794
défaut 620 nm (étude à Dakar). Les pourcentages d’inhibitions, calculés à partir de la formule 1795
1796 % inhib = 100% – ((A éch – A GRO+) / (A SAB – A GRO+)*100) 1797
1798 sont similaires à ces 2 longueurs d’ondes (données C.A. Long) - A désignant l’absorbance -. 1799
1800
7) Les analyses statistiques 1801
1802
Les différents résultats sont étudiés d’un point de vue statistique. Les tests statistiques utilisés 1803
sont réalisés à l’aide des logiciels Statview (version 5.0; SAS Institute) pour les analyses 1804
univariées - tests non paramétriques - et Egret (version 3.01; Cytel) pour les régressions de 1805
Poisson. Ces dernières sont applicables à une variable correspondant au dénombrement d’un 1806
nombre « petit » d’événements, tel que le nombre d’accès palustres mensuel ou annuel. 1807
1808
Le caractère significatif des résultats a été choisi avec un seuil de 5%, c'est-à-dire que nous 1809
considérons que les résultats obtenus sont significatifs si P est inférieure à 0,05. En cas de 1810
tests multiples, la correction de Bonferroni est appliquée à ce seuil. Grossièrement, cela 1811
correspond à diviser ce seuil par le nombre de tests réalisés. Cette correction permet d’assurer, 1812
pour l’ensemble de l’analyse, que le risque d’erreur de 1ère espèce reste à 5%. 1813
1814
Les analyses réalisables avec les tests univariés non paramétriques sont : 1815
� analyse d’une différence entre deux groupes appariés à l’aide du test de rang de 1816
Wilcoxon ou non appariés grâce au test de Mann Whitney. 1817
� analyse d’une différence entre plus de 2 groupes - tels que les strates d’âges, de 1818
parasitémie,… - en utilisant le test de Kruskal-Wallis. 1819
� Analyse de la corrélation entre 2 variables quantitatives via le test de Spearman -1820
coefficient de corrélation : rho*100 -. 1821
1822
Des modèles plus complexes peuvent également être utilisés tel que le modèle de régression 1823
multiple de Poisson. Il permet en particulier d’évaluer le risque relatif, par rapport à une 1824

65
classe de la variable étudiée prise comme référence, ajusté sur les autres variables incluses 1825
dans le modèle. Ce test est utilisé pour analyser le rôle de paramètres immunologiques dans la 1826
protection clinique. Le risque de tomber malade est évalué pour une classe immunologique 1827
donnée - dépassement d’un seuil d’Ac …- par rapport à la classe de référence - sujets dont le 1828
taux d’Ac est inférieur au seuil -, en prenant en compte les autres caractéristiques des 1829
individus - âge, type d’hémoglobine …-. Si la valeur du risque est positive, celui-ci est 1830
augmenté par rapport au groupe de référence, et il est au contraire diminué s’il est négatif. Ce 1831
modèle permet d’identifier les variables à inclure dans le modèle - celles qui sont 1832
significativement associées à la protection clinique - et de les traiter en considérant les 1833
influences de chacune. Cela permet par exemple d’obtenir une mesure des risques relatifs de 1834
morbidité en fonction de l’ADPm et indépendamment de l’âge. Pour déterminer le meilleur 1835
modèle parmi les modèles envisagés, il convient d’utiliser le critère d’Akaïké (AIC). Basé sur 1836
la valeur de la déviance - mesure de l’éloignement des données par rapport au modèle - et le 1837
nombre de degrés de liberté du modèle, il permet d’identifier le modèle le mieux adapté aux 1838
données. Celui-ci est caractérisé par la valeur d’AIC la plus faible. 1839
1840
1841
1842


















