
MUSEUM NATIONAL D'HISTOIRE NATURELLE ECOLE...
233
MUSEUM NATIONAL D'HISTOIRE NATURELLE ECOLE DOCTORALE « SCIENCES DE LA NATURE ET DE L’HOMME » (ED 227) Année 2011-12 N° attribué par la Bibliothèque |_| |_| |_| |_| |_| |_| |_| |_| |_| |_| |_| |_| THESE Pour obtenir le grade de DOCTEUR DU MUSÉUM NATIONAL D’HISTOIRE NATURELLE Discipline : Ecologie Présentée et soutenue publiquement par Amandine Gasc Le 3 décembre 2012 SUJET « ANALYSE ET SUIVI DE LA BIODIVERSITE ANIMALE PAR DES TECHNIQUES NOVATRICES DE BIOACOUSTIQUE » Sous la direction de : Jérôme SUEUR, Sandrine PAVOINE et Philippe GRANDCOLAS (HDR) JURY Denis COUVET Professeur Muséum national d'Histoire naturelle (Paris, France) Président Michael GREENFIELD Professeur Institut de Recherche sur la Biologie de l'Insecte UMR 7261, Université François Rabelais (Tours, France) Rapporteur Rafaël MARQUEZ Chargé de recherche Fonoteca Zoólogica, Museo Nacional de Ciencias Naturales (Madrid, Spain) Rapporteur Thierry AUBIN Directeur de recherche CNRS NPS-CNRS UMR 8195, Université Paris-Sud (Orsay, France) Examinateur Almo FARINA Professeur Faculty of Environmental Sciences, Institute of Biomathematic (Urbino, Italia) Examinateur Philippe GRANDCOLAS Directeur de recherche CNRS Muséum national d'Histoire naturelle (Paris, France) Directeur de thèse Sandrine PAVOINE Maître de conférences Muséum national d'Histoire naturelle (Paris, France) Co-directrice de thèse Jérôme SUEUR Maître de conférences Muséum national d'Histoire naturelle (Paris, France) Co-directeur de thèse
Transcript of MUSEUM NATIONAL D'HISTOIRE NATURELLE ECOLE...

MUSEUM NATIONAL D'HISTOIRE NATURELLE
ECOLE DOCTORALE « SCIENCES DE LA NATURE ET DE L’HOMME »
(ED 227)
Année 2011-12 N° attribué par la Bibliothèque |_| |_| |_| |_| |_| |_| |_| |_| |_| |_| |_| |_|
THESE
Pour obtenir le grade de
DOCTEUR DU MUSÉUM NATIONAL D’HISTOIRE NATURELLE
Discipline : Ecologie
Présentée et soutenue publiquement par
Amandine Gasc
Le 3 décembre 2012
SUJET
« ANALYSE ET SUIVI DE LA BIODIVERSITE ANIMALE PAR DES TECHNIQUES NOVATRICES DE BIOACOUSTIQUE » Sous la direction de : Jérôme SUEUR, Sandrine PAVOINE et Philippe GRANDCOLAS (HDR)
JURY
Denis COUVET Professeur Muséum national d'Histoire naturelle (Paris, France)
Président
Michael GREENFIELD
Professeur Institut de Recherche sur la Biologie de l'Insecte UMR 7261, Université François Rabelais (Tours, France)
Rapporteur
Rafaël MARQUEZ
Chargé de recherche Fonoteca Zoólogica, Museo Nacional de Ciencias Naturales (Madrid, Spain)
Rapporteur
Thierry AUBIN Directeur de recherche
CNRS NPS-CNRS UMR 8195, Université Paris-Sud (Orsay, France)
Examinateur
Almo FARINA
Professeur Faculty of Environmental Sciences, Institute of Biomathematic (Urbino, Italia)
Examinateur
Philippe GRANDCOLAS
Directeur de recherche CNRS
Muséum national d'Histoire naturelle (Paris, France)
Directeur de thèse
Sandrine PAVOINE Maître de conférences Muséum national d'Histoire naturelle (Paris,
France) Co-directrice de thèse
Jérôme SUEUR Maître de conférences Muséum national d'Histoire naturelle (Paris,
France) Co-directeur de thèse


Résumé En parallèle des méthodes classiques d'inventaires, qui demandent un haut niveau d'expertise taxonomique, des méthodes dites de mesure rapide de la biodiversité ont été proposées. Parmi ces méthodes, la méthode acoustique globale, ou Rapid Acoustic Survey (RAS), a été développée en 2008. L'objectif général de ce travail de thèse a été de fournir une définition plus précise de l’approche RAS d'un point de vue méthodologique, afin d'en connaître les possibles applications pour l'estimation, le suivi et l'analyse de la distribution spatiale et temporelle de la biodiversité animale. Des indices acoustiques mesurant l'entropie du signal sonore d'une communauté animale et la dissimilarité acoustique entre communautés ont déjà été proposés. La sensibilité de ces indices à différentes causes d'hétérogénéité acoustique sans lien avec la diversité acoustique a été évaluée (e.g. impact du bruit ambiant). Les résultats montrent que les indices ne sont pas sensibles de la même manière aux différentes causes d’hétérogénéité étudiées ; les biais générés par ces sources d’hétérogénéité doivent donc être traités différemment en fonction de l'indice de diversité utilisé. Une fois ces étapes méthodologiques traitées, plusieurs applications de l'approche ont révélé les points suivants. Dans les communautés d'oiseaux, en France continentale métropolitaine, des corrélations positives entre diversités acoustiques et phylogénétiques d'une part et entre diversités acoustiques et fonctionnelles (liées à la reproduction et au poids) d'autre part ont été identifiées. En Nouvelle-Calédonie, le taux de micro-endémisme est élevé sur l'ensemble du territoire et la biodiversité est donc difficile à décrire. L'utilisation de mesures de dissimilarités acoustiques a pu mettre en évidence des différences acoustiques dans trois régions éloignées de l'île, possédant des niveaux de richesse spécifique et des structures végétales semblables mais des compositions de communautés différentes dues au taux élevé de micro-endémisme. Cette différence dans la composition de communautés animales multi-taxons est donc reflétée par les analyses de diversité acoustique par la méthode RAS. En Inde, deux milieux différents de plantations agro-forestières de café ont été étudiées, l'une utilisant des espèces natives d'arbres et l'autre une espèce exotique. Une étude acoustique a permis de mettre en évidence des différences dans la diversité et la composition acoustique des communautés animales vivant dans ces deux types d'agro-forêt, révélant un impact de l'utilisation de l'espèce exotique sur les communautés d'espèces chanteuses. L'acoustique globale RAS semble donc être une méthode prometteuse pour l'évaluation de la diversité animale par les méthodes d'acoustique non invasives, même si certaines limites méthodologiques devront être dépassées pour fournir un outil utilisable dans de nombreuses conditions acoustiques.
Abstract Alongside conventional methods of inventories, which require a high level of taxonomic expertise, several methods known as rapid measurement of biodiversity have been proposed. Among these methods, an acoustic method, designated as Rapid Acoustic Survey (RAS), was developed in 2008. The general objective of my thesis has been to provide a better definition of RAS approach from a methodological point of view, in order to evaluate the possible applications of RAS for estimating, tracking and analysing the spatial and temporal distribution of animal biodiversity. Acoustic indices have already been proposed which measure the overall entropy of the sound signal in an animal community and the acoustic dissimilarity among communities. The sensitivity of these indices to different causes of heterogeneity unrelated acoustic diversity was assessed (e.g. ambient noise effect). The results show that the indices are not sensitive in the same way to the different causes of heterogeneity studied; the generated biased should thus be treated differently depending on the diversity index used. Once these methodological approaches have been treated, several applications of the RAS revealed the following points. In bird communities in Metropolitan, continental France, positive correlations between acoustic and phylogenetic diversity on the one hand and between acoustic and functional (reproduction traits and weight) diversity on the other hand have been identified. In New Caledonia, the level of micro-endemism is high which make the description of biodiversity difficult. The use of acoustic spectral dissimilarity measures highlight acoustic differences in three distant locations in the island with similar species richness and vegetation structure, but different in the composition of animal communities due to micro-endemism. This difference in multi-taxa animal communities is clearly revealed by the acoustic diversity analysis RAS. In India, two different managements of the coffee agroforest system are found, one using native species and the other one using an exotic species of shade trees. An acoustic survey highlighted differences in acoustic diversity and composition between the two managements revealing an impact of the use of exotic shade tree species on singing animal communities. The RAS method seems then a promising tool to assess local animal diversity even if some methodological issues should be considered to produce a reliable tool in several acoustic conditions.


1
Remerciements
Je tiens à remercier Louis Deharveng et Philippe Grandcolas, respectivement directeur et co-
directeur de l'UMR 7205 « Origine, Structure et Evolution de la Biodiversité », pour leur accueil
au sein du laboratoire et au sein de l'équipe « Phylogénie et Evolution ».
Je remercie chaleureusement mes trois directeurs de thèse Philippe Grandcolas, Sandrine
Pavoine et Jérôme Sueur pour m'avoir offert l'opportunité de réaliser ce travail. Leur expérience
scientifique et leurs connaissances très complémentaires m'ont beaucoup apporté. Merci à eux
trois pour leur disponibilité, leur soutien et leurs encouragements.
Je remercie les membres du jury, Thierry Aubin, Denis Couvet, Almo Farina et particulièrement
les deux rapporteurs Michael Greenfield et Rafaël Marquez, d'avoir accepté de juger ce travail de
thèse. Je leur suis très reconnaissante du temps qu'ils m'auront accordé.
J'aimerais également remercier les chercheurs qui ont accepté de faire partie de mon comité de
thèse, Olivier Adam, Fréderic Jiguet, Purificatión López-García et Jean-Yves Rasplus pour leurs
conseils précieux.
Je tiens à remercier toutes les personnes avec qui j'ai eu l'opportunité de collaborer : Coralie
Burrow, Benjamin Dechant, Marion Depraetere, Vincent Devictor, Pierre-Yves Martel, Fréderic
Jiguet, Laurent Lellouch, Ayyappan Narayanan, Roseli Pellens, Raphaël Pélissier, Alexandra
Rodriguez; et toutes les personnes dont les discussions scientifiques m'ont permis d'avancer :
Isabelle Leviol, Anne-Béatrice Dufour, Denis Clos, Tony Robillard, Laure Desutter, Romain
Nattier, et Frédéric Legendre.
Je remercie les personnes qui ont pris le temps de corriger l'anglais des manuscrits : Ryan
Calsbeek, Yoland Savriama, Brent Berger and James Windmill; ceux qui ont pris le temps de
relire une ou plusieurs parties de ce travail : Lorène Marchal, Adrien Perrard et Antonio
Fischetti; et merci à Nicolas Deguines pour m'avoir accompagné sur le terrain en Inde.
J'ai eu la chance de faire des missions de terrain riches scientifiquement, culturellement et
humainement. Merci à l'aide des personnes sur place pour leur connaissance du terrain et leur
soutient technique : Addy (Inde), Philippe Gaucher et Patrick Châtelet (Guyane française) et
Hervé Jourdan et Edouard Bourguet (Nouvelle-Calédonie).

2
Ce travail est issu d'un partenariat entre le Centre National de Recherche Scientifique, qui a
financé mes trois années de thèse par une bourse de l'Institut Ecologie et Environnement, et le
Muséum national d'Histoire naturelle qui a mis à ma disposition un bureau, les accès aux
ressources bibliographiques nécessaires et un contexte de travail unique. Les missions de
terrain et l'achat du matériel n'aurait pas été possible sans les financements de l'ANR
BIONEOCAL et du projet BIOSOUND de la FRB.
Je tiens à remercier les membres de ma famille, mes parents pour croire en moi en toutes
circonstances et mon frère pour nos discussions sur la recherche.
Merci à Gregory pour avoir assurer l'assistance téléphonique 24h sur 24 et pour son amour et
son soutien qui sont toujours intacts après ces trois ans.
Enfin merci à tous les amis qui ont contribué à faire de ces trois ans une belle aventure: Lorène,
le bureau n'est plus le même sans toi et, bientôt sans moi, imagine!; Marion, si je suis heureuse
d'avoir pu faire cette thèse, je le suis d'autant plus que c'est elle qui nous a rapproché à vie; Fred,
mon petit freu-feu, bien sûr que les blattes sont merveilleuses, et que, tant que tu a l'âge de
porter un sweat à capuche, tu n'es pas vieux; Davinia, ma colloque préférée, tu n'auras
finalement pas eu à supporter ces derniers jours fatidiques de fin de thèse; Adrien merci pour les
pauses dansantes et pleines de chocolat; Clément, merci pour les discussions profondes et pour
avoir pris soin de notre Grignotte national; Jérémie, je retiendrais ton charme irrésistible quand
tu parles des séries de ce cher Monsieur Fourier; Merci à Fabrice pour sa constance, Alexandre
pour sa perversion, Paul pour ta future participation à notre groupe de rock, Yoland pour avoir
partagé ton amour du street dance; Alexandra pour ta vision du monde rassurante, Jim pour ta
bonne humeur. Merci à mes bests Lucie et Nathalie, surtout ne changez rien je vous adore.
Merci à tous mes amis d'avant thèse (et oui, il y a un avant et un après) qui ne m'ont pas
beaucoup vu ces derniers mois (n'est ce pas Anton, Alex, Emma, Laura, Coco, Steph et je ne peux
pas tous vous citer mais vous vous reconnaitrez!)
Enfin, merci à toute l'équipe du bâtiment d'entomologie et particulièrement Alice et Gilles.

3
Table des matières Liste des figures ................................................................................................................ 9 Liste des tables ................................................................................................................. 11 ---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- Introduction générale .................................................................................................. 13 1 Biodiversité: définition et estimation ....................................................................... 15 1.1 Définition de la biodiversité .................................................................................. 15
1.1.1 Naissance du terme « biodiversité » ........................................................ 15 1.1.2 Les différentes échelles : une vision espèce centrée ............................... 16 1.1.3 Les différentes facettes de la biodiversité ................................................ 17
1.2 Estimation de la biodiversité ................................................................................. 19 1.2.1 Combien d’espèces sur terre ? ................................................................. 19 1.2.2 L’érosion de la biodiversité, de l'objectif 2010 à l'objectif 2020 ............ 20 1.2.3 Comment estimer la biodiversité ? .......................................................... 23
1.3 Priorités de conservation ....................................................................................... 28
2 L’acoustique passive: un outil pour une méthode globale d’estimation de la biodiversité ................................................................................................................. 30
2.1 Le son ...................................................................................................................... 30 2.1.1 Explication physique d'un phénomène ondulatoire ............................... 30 2.1.2 Les séries et transformées de Fourier ou comment prendre des mesures d'un son naturel par une décomposition en fréquence .............. 32 2.1.3 Numérisation du son ................................................................................ 35 2.1.4 Les représentations du son ...................................................................... 36
2.2 La production sonore animale .............................................................................. 38 2.3 L'acoustique passive .............................................................................................. 41
2.3.1 Un domaine de recherche en plein essor ................................................ 43 2.3.2 Un outil pour estimer la biodiversité ....................................................... 45 2.3.3 La méthode acoustique globale: Rapid Acoustic Survey ....................... 49
3 Objectifs de la thèse et présentation des chapitres ............................................. 52
----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------

4
Chapitre 1 - Les indicateurs de l’hétérogénéité acoustique ................ 55 I.1 Résumé ............................................................................................................................. 57 I.2 Introduction .................................................................................................................... 58 I.3 Matériel et méthodes ................................................................................................... 60
I.3.1 Simulations ........................................................................................................... 60 I.3.1.1 Question 1: Quel est l'impact des intensités relatives des séquences sonores sur les indices acoustiques ? ...................................... 61 I.3.1.2 Question 2: Quel est l’impact de la durée d'un signal par rapport à la durée d'enregistrement sur les indices acoustiques ? .......... 61 I.3.1.3 Question 3 : Quel est l’impact du décalage temporel entre les signaux sur les indices acoustiques ? ......................................................... 62 I.3.1.4 Question 4 : Quel est l’impact du bruit ambiant sur les indices acoustiques ? ................................................................................... 62
I.3.2 Les indices de diversité acoustiques .................................................................. 62
I.3.2.1 L'indice d'entropie acoustique H .......................................................... 64 I.3.2.2 L'indice de richesse acoustique (Acoustic Richness) AR .................... 65 I.3.2.3 L'indice de complexité acoustique (Acoustic Complexity Index) ACI ..................................................................................................................... 66 I.3.2.4 Nombre de pics fréquentiels NP ........................................................... 68 I.3.2.5 L'indice de dissimilarité acoustique D.................................................. 69 I.3.2.6 Les indices de distance de Kullback-Leibler KL et de Kolmogorov-Smirnov KS ......................................................................... 70
I.4 Résultats ........................................................................................................................... 71
I.4.1 Question 1: Quel est l'impact des intensités relatives des séquences sonores sur les indices acoustiques ? .......................................................................... 71 I.4.2 Question 2: Quel est l’impact de la durée d'un signal par rapport au temps d'enregistrement sur les indices acoustiques ? ......................................... 72 I.4.3 Question 3 : Quel est l’impact du décalage temporel entre les signaux sur les indices acoustiques ? ........................................................................................ 73 I.4.3 Question 4 : Quel est l’impact du bruit ambiant sur les indices acoustiques ? .............................................................................................. 75
I.5 Discussion ........................................................................................................................ 76 I.5.1 Variation d'intensités relatives: une détectabilité déséquilibrée des son ....... 76 I.5.2 Variation de la durée de l'enregistrement par rapport à la durée du signal étudié ............................................................................................................. 78

5
I.5.3 Variation du décalage temporel ......................................................................... 78 I.5.4 L'effet du bruit ambiant ..................................................................................... 79 I.5.5 Les indices de dissimilarité : des indices points à points ................................. 80
I.6 Conclusion ....................................................................................................................... 80 ---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- Chapitre 2 - Mesurer la biodiversité par le son : la diversité acoustique peut-elle refléter la diversité phylogénétique et fonctionnelle de communautés d’oiseaux ?. ........ 83 II.1 Abstract/Résumé ......................................................................................................... 86
II.1.1 Abstract ............................................................................................................... 86 II.1.2 Résumé ................................................................................................................ 87
II.2 Introduction .................................................................................................................. 89 II.3 Material and methods ............................................................................................... 91
II.3.1 Data ..................................................................................................................... 93
II.3.1.1 Bird communities ................................................................................. 93 II.3.1.2 Bird songs ............................................................................................. 93 II.3.1.3 Phylogenetic data ................................................................................. 93 II.3.1.4 Functional data .................................................................................... 94
II.3.2 Distance matrices ............................................................................................... 94
II.3.2.1 Acoustic distances ................................................................................ 94 II.3.2.2 Phylogenetic distances ......................................................................... 95 II.3.2.3 Functional distances ............................................................................ 95 II.3.3 Diversity indices ..................................................................................... 96
II.3.4 Statistical analyses ............................................................................................. 96 II.3.4.1 Species level .......................................................................................... 97 II.3.4.2 Community level ................................................................................... 97
II.4 Results ............................................................................................................................. 99
II.4.1 Species level ........................................................................................................ 99 II.4.2 Community level .............................................................................................. 100

6
II.5 Discussion .................................................................................................................... 103 II.6 Acknowledgements .................................................................................................. 107 ---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- Chapitre 3 - Echantillonnage de la biodiversité par une approche globale d'acoustique passive : différences entre sites abritant des micro-endémiques de Nouvelle-Calédonie .......... 109 II.1 Abstract/Résumé ....................................................................................................... 112
II.1.1 Abstract ............................................................................................................. 112 II.1.2 Résumé .............................................................................................................. 112
II.2 Introduction ................................................................................................................ 114 II.3 Material and methods ............................................................................................. 116
III.3.1 Study sites ........................................................................................................ 116 III.3.2 Passive acoustic recording ............................................................................. 117 III.3.3 Acoustic activity level ..................................................................................... 118 III.3.4 Background noise reduction ......................................................................... 118 III.3.5 Acoustic complexity ....................................................................................... 119 III.3.6 Spectral global differences ............................................................................ 119 III.3.7 Spectral differences detailed ......................................................................... 121
III.4 Results ......................................................................................................................... 121
III.4.1 Classical inventories ....................................................................................... 121 III.4.2 Acoustic sampling .......................................................................................... 122
III.5 Discussion and conclusions ................................................................................. 126 III.6 Acknowledgements ................................................................................................. 130 ----------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------

7
Chapitre 4 - Effet des stratégies de gestion des agro-systèmes sur le niveau de biodiversité dans les Western Ghats (Inde) : une étude acoustique et botanique .................................................................... 131 IV.1 Abstract/Résumé ..................................................................................................... 134
IV.1.1 Abstract ........................................................................................................... 134 IV.1.2 Résumé ............................................................................................................. 135
IV.2 Introduction .............................................................................................................. 136 IV.3 Material and methods ........................................................................................... 139
IV.3.1 Sites .................................................................................................................. 139 IV.3.2 Tree diversity .................................................................................................. 140 IV.3.3 Tree density ..................................................................................................... 141 IV.3.4 Passive acoustic recording ............................................................................. 141 IV.3.5 Acoustic data selection ................................................................................... 142 IV.3.6 Acoustic diversity indices ............................................................................... 142 IV.3.7 Spatial correlation .......................................................................................... 143 IV.3.8 Statistical analysis of acoustic data ............................................................... 144
IV.4 Results ......................................................................................................................... 145 IV.4.1 Tree diversity and density ............................................................................. 145 IV.4.2 Acoustic diversity ........................................................................................... 146
IV.5 Discussion and conclusions .................................................................................. 149 IV.6 Acknowledgements ................................................................................................. 151 ---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- Chapitre 5 - Discussion générale ........................................................................ 153 V.1 La diversité acoustique ........................................................................................... 155

8
V.2 Critiques méthodologiques .................................................................................... 157 V.2.1 Dissimilarité temporelle................................................................................... 157 V.2.2 Community Acoustic Diversity ....................................................................... 158 V.2.3 Diversité acoustique/diversité fonctionnelle .................................................. 158 V.2.4 Test sur communautés réelles: validation des indices sur la base d'inventaires ............................................................................................................... 159 V.2.5 L'utilisation du spectre moyen........................................................................ 159 V.2.6 Le traitement du bruit ..................................................................................... 160 V.2.7 Protocole d'échantillonnage acoustique ......................................................... 161
V.3 La méthode acoustique globale : une méta-méthode .................................. 161 V.4 Perspectives pour l'analyse et le suivi de la biodiversité ............................ 165 ---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- Références bibliographiques ................................................................................ 169 ---------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- Annexe 1 : Chapitre du livre Le Gaillard, J-L., Guarini, J-M. and Gaill, F. (2012) Sensors for ecology. CRNS, Paris............... ............................................................................ 188 Annexe 2 : Matériel supplémentaire du Chapitre 2 .......................................................... 211 Annexe 3 : Matériel supplémentaire du Chapitre 3 .......................................................... 219

9
Liste des figures
Introduction générale Fig. 1 : Représentation des concepts de richesse spécifique et d'abondance relative des espèces dans une communauté ............................................................................... 17 Fig 2 : Schéma reprenant les différentes facettes de la biodiversité ......................................... 19 Fig. 3 : Arbre représentant les différents niveaux taxonomiques pour le calcul de l'indice de diversité taxonomique ......................................................................................... 25 Fig. 4 : Arbre représentant les différents niveaux taxonomiques pour le calcul de l'indice de diversité phylogénétique ...................................................................................... 26 Fig. 5 : Schéma des éléments utilisés pour calcul de l'entropie quadratique ............................ 27 Fig. 6 : Schéma de la propagation d'une onde dans la matière .................................................. 31 Fig. 7 : Schéma illustrant différentes caractéristiques d'une onde sinusoïdale ......................... 32 Fig. 8 : Illustrations de l'ouvrage « Théorie analytique de la chaleur » de Joseph Fourier ....... 33 Fig. 9 : Schéma d’une onde périodique non sinusoïdale ........................................................... 34 Fig. 10 : Schéma de la décomposition en série de Fourier d'une onde sinusoïdale .................. 34 Fig. 11 : Schéma d’une onde non périodique, non sinusoïdale ................................................. 35 Fig. 12 : Oscillogramme calculé à partir d'un son de Zonotrichia capensis mâle .................... 37 Fig. 13 : Spectrogramme calculé à partir d'un son de Zonotrichia capensis mâle .................... 38 Fig. 14 : Spectre moyen calculé à partir d'un son de Zonotrichia capensis mâle ..................... 38 Fig. 15 : Evolution du nombre d'articles, dont le sujet possède le mot-clef « Bioacoustic » ... 43 Fig. 16 : Méthode d'estimation de la biodiversité par des techniques d'acoustique passive ..... 46
Chapitre 1 Fig. I.1 : Schéma théorique représentant la diversité acoustique de la communauté ............... 58 Fig. I.2 : Schéma reprenant les étapes permettant le calcul des indices Hf et Ht ...................... 64 Fig. I.3 : Schéma reprenant les étapes permettant le calcul des indices Ht et M ....................... 65 Fig. I.4 : Schéma reprenant les étapes permettant du calcul de l'indice ACI ............................ 67

10
Fig. I.5 : Schéma reprenant les étapes permettant le calcul de l'indice NP ............................... 68 Fig. I.6 : Schéma reprenant les étapes permettant le calcul des indices Df et Dt ..................... 69 Fig. I.7 : Schéma représentant les étapes permettant le calcul des indices KL et KS ................ 70 Fig. I.8 :Boxplot des valeurs des écart-types mesurés pour chaque communauté et chaque indice.......................................................................................................................... 71 Fig. I.9 : Variation de l'écart-type des indices acoustiques en fonction du nombre de séquences sonores ................................................................................................................. 72 Fig. I.10 : Valeurs des indices acoustiques en fonction de la durée d'enregistrement par rapport à la durée du signal d'intérêt.................................................................................... 73 Fig. I.11 : Valeurs des indices acoustiques en fonction du décalage des séquences dans le signal .............................................................................................................................. 74 Fig. I.12 : Valeurs des indices de distances acoustiques en fonction du chevauchement des séquences dans le signal ....................................................................... 75 Fig. I.13 : Valeurs des indices acoustiques sur des signaux soumis à un bruit ambiant faible, moyen ou fort ................................................................................................... 76 Fig. I.14 : Spectre moyen d’un signal composé de deux sons de fréquences différentes ......... 77
Chapitre 2 Fig. II.1: From data to indices: process used for the computation of diversity indices ............ 92 Fig. II.2: Histogram of diversities corrected with the null model ........................................... 102
Chapitre 3 Fig. III.1: Sampled sites in the great island of New Caledonia ............................................... 117 Fig. III.2: Acoustic activity on the three sites.......................................................................... 122 Fig. III.3: Number of frequency peaks .................................................................................... 123 Fig. III.4: Principal Coordinate Analysis (PCoA) on the acoustic dissimilarity matrix ......... 124 Fig. III.5: Distance-based ReDundancy Analysis (dbRDA) ................................................... 125 Fig. III.6: Histogram of the frequency used among all recordings ......................................... 126

11
Liste des Tables
Introduction générale
Table 1 : Transformation appliquée sur les indices pour qu'ils respectent le principe de réplication............................................................................................................ 28 Table 2 : Comparaison des critères utilisés pour la détection d'unités sonores dans les chants d'oiseaux par différentes méthodes de classification ................................................ 48
Chapitre 1 Table I.1 : Résumé des indices acoustiques .............................................................................. 63
Chapitre 2 Table II.1: Mantel correlations, based on Spearman method with randomization, between the different matrices of pair-wise acoustic distances between species ..................... 99 Table II.2: Results of the Mantel test, with randomization using Spearman correlation coefficient, between the different distance matrices ............................ 100 Table II.3: Spearman correlation coefficients between acoustic diversity indices ................. 101

12

13
Introduction générale
« L'ennui naquit un jour de l'uniformité »
Antoine de La Motte-Houdar (1719, Fables nouvelles)

14

15
1 Biodiversité : définition et estimation
1.1 Définition de la biodiversité
1.1.1 Naissance du terme « biodiversité »
La biodiversité est également appelée diversité biologique ou variabilité du monde vivant. Le
terme de « Biodiversité » est proposé en 1985 par Walter G. Rosen, et utilisé lors du
« National Forum on Biological Diversity » organisé par le National Research Council en
1986. C'est à partir de 1988, date à laquelle ce mot apparaît pour la première fois dans une
publication (Wilson and Peter, 1988), qu'il est largement utilisé par la communauté
scientifique et plus tard par le monde politique et plus largement encore par l'ensemble du
grand public.
C'est lors du « Sommet de la terre » à Rio de Janeiro, en 1992, que naît la Convention
sur la diversité biologique (CDD). Ce traité international a pour but de garantir la
« conservation de la diversité biologique », « l'utilisation durable de ses éléments » et « le
partage juste et équitable des avantages découlant de l'exploitation des ressources
génétiques » (article 3, CDD). Une définition de la biodiversité est alors proposée :
« La variabilité des organismes vivants de toute origine y compris, entre autres, les
écosystèmes terrestres, marins et autres écosystèmes aquatiques et les complexes écologiques
dont ils font partie ; cela comprend la diversité au sein des espèces et entre espèces ainsi que
celle des écosystèmes. »
Article 2 de la Convention sur la diversité biologique, 1992, Sommet de la terre à Rio de
Janeiro
Wilson (in Reaka-Kudla et al., 1997) propose une définition plus large dans l'ouvrage
Biodiversity II qui s'ouvre à toutes les échelles de la diversité biologique :
« Biodiversity is defined as all hereditarily based variation at all levels of organization, from
the genes within a single local population or species, to the species composing all or part of a
local community, and finally to the communities themselves that compose the living parts of
multivarious ecosystems of the world. The key of the effective analysis of biodiversity is the
precise definition of each level of organization when it is being addressed. »

16
Ainsi la « Biodiversité » est un concept large qui définit la diversification du vivant
sur les différents niveaux d'organisation. Elle peut être considérée à toutes les échelles
écologiques, du gène à l'écosystème en passant par la diversité au niveau de l'individu, de
l'espèce, de la population, de la communauté ou du paysage. Dépendante de son histoire et de
son environnement, la biodiversité évolue dans le temps et varie dans l'espace. L’estimation
de la biodiversité d’un même site peut aussi varier selon le groupe vivant ou l’échelle étudiés
(Gaston, 2000).
1.1.2 Les différentes échelles : une vision espèce-centrée
Si la biodiversité peut être considérée à de multiples échelles, la plus utilisée est sans doute
l’échelle de l’espèce. La définition de l'espèce a été, et est encore, discutée. La plus largement
utilisée est sans doute la définition biologique de l'espèce, tel qu'elle est énoncée par Wilson
& Peter (1988) :
« Species are regarded conceptually as a population or series of populations within which
free gene flow occurs under natural conditions. This means that all the normal,
physiologically competent individuals at a given time are capable of breeding with all the
other individuals of the opposite sex belonging to the same species or at least that they are
capable of being linked genetically to them through chains of other breeding individuals ».
Le principe de reproduction est admis par la communauté scientifique mais ce critère n’est
pas celui que les taxonomistes utilisent pour décrire les espèces. Ceux-ci admettent le critère
de reproduction comme intrinsèque à la notion d'espèce mais la plupart des espèces sont
décrites sur la base de critères de ressemblance (morphologiques, moléculaires et /ou
comportementaux). Ces critères sont plus utilisés car les espèces définies sur la communauté
de reproduction ne sont pas forcément monophylétiques. Si cette réflexion reste importante et
fondamentale dans les travaux biologiques, nous ne prendrons pas parti ici. Quelque soit la
définition choisie, le fait que les mesures de la biodiversité soient largement espèce-centrée
s'explique certainement par le fait que l'espèce est le plus bas niveau de description du monde
du vivant choisi par les taxonomistes et les systématiciens. L'espèce est une unité
taxonomique, décrite sur des critères de ressemblance et intéressante d'un point de vue
biologique étant donné l'échange de gènes entre individus de la même espèce qui leur
confèrent une histoire évolutive unique (Purvis & Hector, 2000).

17
1.1.3 Les différentes facettes de la biodiversité
Si la notion de variabilité est simple à définir au sens large, concrètement comment la
représenter, comment la mesurer ? Quelque soit l’échelle considérée, il existe divers moyens
d’appréhender cette variabilité. Ces différentes facettes peuvent être classées selon trois types
de mesure : la richesse, l’abondance et la différence. Considérant le niveau spécifique par
exemple, la diversité peut être quantifiée par la richesse spécifique c'est-à-dire le nombre
d’espèces, mais également par l’abondance relative de chacune de ces espèces. Deux
communautés peuvent ainsi être à richesse égale mais avec des niveaux de diversité différents
si on considère l'abondance relative des espèces. La figure 1, issue de l’article de Purvis &
Hector (2000), illustre ces deux aspects de la diversité.
Fig. 1 : Représentation des concepts de richesse spécifique et d'abondance relative des espèces dans une
communauté. Deux ensembles d'insectes issus de deux sites différents varient selon deux critères : la richesse
spécifique et l'abondance relative des espèces. L'échantillon A ("Sample A") pourrait être considéré plus divers
car il contient trois espèces alors que l'échantillon B ("Sample B") n'en contient que deux. Cependant, si on tirait
aléatoirement deux individus dans l'un des échantillons, on aurait moins de chance de tirer deux individus de la
même espèce dans l'échantillon B que dans l'échantillon A. La distribution d'abondance relative des espèces est
un critère de diversité qui n'est pas forcement lié à la richesse spécifique. Issue de l'article de Purvis & Hector
(2000).
Enfin, la diversité spécifique au sein d'une communauté peut être mesurée par la
différence qui existe entre les espèces. Pavoine et al. (2005a) abordent cette approche en
prenant l'exemple de deux ensembles d'espèces, l'un étant composé de trois espèces dont une

18
espèce de Felidae, une de Canidae et une d’Ursidae et l'autre étant composé de trois espèces
différentes appartenant aux Felidae. Pour un même nombre d’espèces et une même abondance
relative des espèces, il existe dans cet exemple des différences plus ou moins fortes entre les
espèces au sein du même ensemble. Différencier les espèces a donné lieux à différents types
de qualifications ; espèces dites ingénieurs (Wright et al., 2002), rares (Patil & Taillie, 1982),
endémiques (Myers et al., 2000), parapluies (Roberge, 2004), d'intérêt économiques (Brehm,
2010), clef de voutes (Simberloff, 1998) ou indicatrices (Fleishman, 2005). Cependant, les
différenciations les plus utilisées aujourd'hui en écologie et biologie de la conservation sont
liées à l’histoire phylogénétique des espèces (diversité phylogénétique, Faith, 1992 ; Vane-
Wright et al., 1991) ou aux traits fonctionnels (diversité fonctionnelle, Petchey & Gaston,
2002). Deux espèces sont d'autant plus différentes phylogénétiquement qu'elles sont éloignées
dans un arbres phylogénétique et donc qu'elles présentent des différences dans les traits de
caractères utilisés pour la construction de l'arbre (Magurran, 2004). Dans ce contexte, plus
une espèce contribue à la diversité totale des espèces d'un arbre phylogénétique, plus cette
espèce est dite « originale » (Vane-Wright et al., 1991). De même, la diversité fonctionnelle
différencie les espèces sur la base de leurs caractères reliés aux processus éco-systémiques.
Ainsi la facette phylogénétique permet de rendre compte de l'histoire évolutive et donc de
l’ensemble des caractères partagés par les espèces. La différence fonctionnelle, quant à elle,
rend compte de la variabilité des rôles de ces espèces dans les processus éco-systémiques
(Petchey & Gaston, 2006). La richesse, l'abondance relative ou les différences entre espèces
sont donc des éléments reflétant la diversité d’une communauté.
Trois types de diversités, α, β et γ ont été dans un premier temps définis au niveau de
l'espèce (Whittaker, 1972). La diversité α permet d’estimer la diversité d'espèces au sein d'un
seul habitat. La diversité β mesure la diversité entre les habitats permettant de comprendre le
patron spatio-temporel de la diversité spécifique. Enfin, la diversité γ mesure la diversité à
l'intérieur d'un paysage (Fig. 2).

19
Fig 2 : Schéma reprenant les différentes facettes de la biodiversité.
Pour mesurer ces différentes facettes, un très grand nombre d'indices mathématiques
ont été proposés. La majorité d’entre eux ont été construits sur l'unité de mesure de l'espèce
mais sont adaptables à d’autres échelles d'étude. Il serait difficile de faire un résumé complet
de tous ces indices, c’est pourquoi seuls les indices utilisés dans ce travail seront présentés
(voir partie 1.2.3)
1.2 Estimation de la biodiversité
Toute mesure exacte d’un niveau de biodiversité est un objectif difficilement atteint. Il est
donc plus correct de parler d’estimation de biodiversité. Avant toute estimation possible, la
biodiversité doit être connue, décrite. C'est le travail des taxonomistes. De la taxonomie est
née la systématique qui cherche à retracer l'évolution des espèces ou groupes d'espèces. Ces
sciences décrivent donc une à une chaque espèce avec pour but commun de rendre compte de
la diversité du monde vivant. Ce travail de description a permis d’aborder une question à
laquelle nous n’avons toujours pas de réponse définitive : combien y a-t-il d’espèces sur la
Terre et quelle est l’évolution de leur maintien ?
1.2.1 Combien d’espèces sur Terre ?
Un inventaire exhaustif des espèces sur Terre pourrait permettre de répondre à cette question.
Cependant un tel travail semble irréalisable. A partir des informations sur les espèces décrites,
de nombreux chercheurs ont proposés des estimations. Lovejoy (1997) considère le nombre
d'espèces vivantes sur terre entre 10 et 100 millions. May (2011) propose un chiffre entre 3 et
50 millions. Mora et al. (2011) prédisent approximativement 8,74 millions d'espèces

20
eucaryotes pour 1,5 millions décrites, ils estiment également à 1200 le nombre d'années
nécessaire pour finir la description des espèces eucaryotes si on dispose de 303 000
taxonomistes et pour un coût de 364 milliards de dollars américains.
Les connaissances en termes d'espèces sont très variables en fonction des lieux
géographiques et des taxons considérés (Lovejoy, 1997). Par exemple, May (2011) rapporte
un même effort de description des espèces pour les vertébrés, les invertébrés et les plantes
alors que les espèces de plantes sont 10 fois plus nombreuses et les espèces d'invertébrés 100
fois plus nombreuses que les espèces de vertébrés. Une estimation globale de 10 millions
d'espèces suggérerait que 86% des espèces sont inconnues, 7% sont uniquement connus d'une
localité, 7% pour plus d'une localité et enfin, le danger d'extinction n'est connu que pour 0.5%
des espèces (Stork, 1997 ; Mora et al., 2011).
Décrire une espèce peut être réalisé sur un seul et unique individu ou quelques
individus collectés sur une seule localité, ce qui ne permet absolument pas de rendre compte
de la biologie, de la distribution ou de l’écologie de l’espèce. Ainsi, beaucoup d’espèces sont
décrites sans qu’il n’y ait aucun moyen de définir leur risque d’extinction ou les pressions
qu’elles subissent. Entre 5 et 15 millions d'espèces décrites seraient dans ce cas, représentant
10% à 30% de toutes les espèces décrites (Stork, 1997).
La description des espèces est donc un premier pas nécessaire vers la connaissance de la
variabilité du vivant mais très vite les scientifiques ont compris que le nombre d’espèces
n’était pas suffisant pour décrire une diversité biologique. Les notions d’abondance et de
composition ont ensuite été intégrées, et avec elles, des indicateurs de biodiversité
complémentaires à la simple richesse spécifique. Avant de s’intéresser aux mesures de la
biodiversité, il est intéressant de comprendre les enjeux de ces mesures.
1.2.2 L’érosion de la biodiversité, de l'objectif 2010 à l'objectif 2020
Si la biodiversité fait l'objet d'autant de réflexions, de conventions et d’assemblées
internationales, c'est qu'une sonnette d'alarme a été tirée. La prise de conscience collective est
essentiellement liée à deux phénomènes : l'accumulation de données sur la déforestation et sur
les extinctions d'espèces et une prise de conscience du lien entre le domaine de l'économie et
celui de la conservation de la biodiversité (Reaka-Kudla, M. L, 1997).
Fragmentation et dégradation des habitats, introduction d’espèces, changement
d’usage des sols, pollution, surexploitation des ressources naturelles, et changement
climatique sont les principaux phénomènes d'origine anthropique identifiés comme

21
responsables, à des échelles différentes, de la perte massive de la biodiversité actuelle (Araújo
et al., 2006, Thuiller et al., 2010). L'érosion de la biodiversité est associée à un taux
d'extinction d'espèces 100 à 1000 fois supérieur au taux d'extinction moyen à l'échelle des
temps géologiques (Mace et al., 2005). Ce niveau de perte de diversité biologique est
considéré comme pouvant provoquer des changement brutaux dans les systèmes naturels
(Rockström et al., 2009). La perte d'espèces a pour conséquence une homogénéisation
biotique avec une disparition des espèces spécialistes au profit des espèces généralistes
(Julliard et al., 2004 ; Kotze & O’Hara, 2003 ; Warren et al., 2001). L'érosion de la
biodiversité est également associée à une perte des services éco-systémiques, du potentiel
évolutif, d'un service alimentaire, et ce, associé à des problèmes éthiques, scientifiques et
sociaux (Chapin III et al., 2000 ; De Groot & Ramakrishnan, 2005 ; Díaz et al., 2005 ;
Lovejoy et al., 1994). C'est un enjeu à la fois écologique, éthique, économique, social et
politique.
2010 Biodiversity Target
En 2002, plus de 190 pays rassemblés par la Convention sur la diversité biologique
s'engageaient à réduire significativement l'érosion de la biodiversité mesurée en 2002, à un
niveau local, régional et national (Balmford et al., 2005). Cet objectif est appelé le « 2010
Biodiversity Target ». Pour savoir si cet objectif a été atteint ou non, un certain nombre
d'indicateurs ont été développés concernant 1) l'état des différents éléments constitutifs de la
diversité biologique, 2) les pressions exercées sur la diversité biologique, 3) les réponses
apportées pour gérer le problème de l'appauvrissement de la diversité biologique
(Perspectives mondiales de la diversité biologique 3). Le principal problème du calcul de ces
indicateurs est que la quantité et la couverture de données sont insuffisantes.
Les indicateurs estimant l'état des différents éléments constitutifs de la diversité
biologique sont concentrés sur trois niveaux de biodiversité :
- les gènes : la diversité génétique a été mesurée dans les écosystèmes naturels, mais
aussi dans les systèmes de production de cultures et d'animaux d'élevage.
- les espèces : l'évaluation de la perte de biodiversité au regard des espèces est
essentiellement basée sur les informations de la liste rouge UICN.
- Les écosystèmes: trois types d'écosystèmes ont été différenciés : les écosystèmes
terrestres, des eaux continentales et marins et côtiers. En ce qui concerne les écosystèmes
terrestres, les indicateurs utilisés sont la déforestation, la disparition des savanes et prairies,
l'utilisation des pratiques agricoles traditionnelles, le morcellement des habitats et la surface

22
des aires protégées. (Perspectives mondiales de la diversité biologique 3). Pour les
écosystèmes des eaux continentales, les indicateurs utilisés sont l'assèchement des zones
humides, la qualité de l'eau et la fragmentation des réseaux par des barrages et des réservoirs.
Enfin les écosystèmes marins et côtiers sont évalués par rapport à l'érosion des habitats côtiers
(mangrove, herbiers marins, marais salants, etc) et des récifs coralliens, l'évolution des stocks
de poissons et la superficie des aires marines protégées.
Les résultats de ces indicateurs démontrent que l'objectif « 2010 Biodiversity Target » n'est
atteint sur aucune des évaluations effectuées. Si un grand nombre de mesures ont été mises en
place, les méthodes de mesures et de conservation de la biodiversité sont trop lentes en regard
de l'avancée des impacts humains sur l’environnement.
L'objectif 2020, le « Grand Plan »
Après cet échec, c'est au cours de la 10e réunion de la Conférence des Parties à la Convention
sur la diversité biologique organisée en 2010 à Nagoya au Japon que de nouveaux objectifs
ont été redéfinis au travers d'un nouveau Grand Plan. Près de 200 gouvernements
s'engageaient à « orienter le processus décisionnel des secteurs publiques et privés sur les dix
prochaines années » (UICN, rapport Bilan de 2010). Ce nouveau texte a été approuvé par les
193 Etats membres des Nation Unies lors de la conférence des Nations Unies sur le
développement durable de Rio qui a eu lieu cette année (2012).
Parmi les objectifs énoncés, on notera un engagement pour :
« Renforcer la mise en œuvre au moyen d’une planification participative, de la gestion des
connaissances et du renforcement des capacités » (But stratégique E)
« D’ici à 2020, les connaissances, la base scientifique et les technologies associées à la
diversité biologique, ses valeurs, son fonctionnement, son état et ses tendances, et les
conséquences de son appauvrissement, sont améliorées, largement partagées et transférées, et
appliquées. » (Objectifs 19)
Tous ces acteurs de la société ont donc en commun le souci de mieux appréhender la
biodiversité. Nous avons besoin de savoir comment elle s'organise spatialement et comment
elle évolue dans le temps afin de comprendre quelles pressions elle subit et si les mesures
prisent sont efficaces.

23
1.2.3 Comment mesurer la biodiversité ?
S'il est impossible de mesurer l'entière variabilité du vivant, il est possible de l'évaluer
grâce à des indicateurs. Un indicateur doit rendre compte des variations et des tendances de ce
qu'il synthétise. Il doit résumer une information complexe qui permet à différents acteurs de
dialoguer (EEA Technical report No 11, 2007). Les indicateurs peuvent être classés selon cinq
groupes (DPSIR: Driver, Pressure, State, Impact, Response), selon qu'ils résument des
informations d'activité (ex. : surpêche), de pression (ex. : menaces sur la biodiversité), d'état
(ex. : état de la biodiversité), d'impact (ex. : impact sur la biodiversité), ou de réponse (ex. :
biodiversité face aux mesures prises). Dans le cas de la mesure de la biodiversité, les indices
sont donc des indicateurs d'état (EEA Technical report No 11, 2007).
Nous avons vu précédemment que la biodiversité se composait de différentes facettes,
le niveau de diversité de chacune d'entre elles pouvant être estimé par des indices. Nous
appellerons indice une fonction mathématique qui permet de mesurer une facette de la
biodiversité. Certains indices prennent en compte différents aspects de la biodiversité afin
d'obtenir une valeur la plus représentative possible de l'état général de celle-ci. Ces valeurs
sont parfois relatives variant entre 0 et 1 (Magurran, 2004).
La diversité se mesure dans une collection d'entités regroupées en catégories. Par
exemple, une communauté (collection) est composée d'espèces (catégories) représentées par
un nombre d'individus (entités). Un très grand nombre d'indices ont été conçus pour obtenir
des mesures de la biodiversité à partir d'un échantillon, nous ne détaillerons que les indices
qui seront abordés dans cette thèse.
Les trois indices principalement utilisés sont la richesse spécifique, l'indice de
Shannon-Wiener et l'indice de Gini-Simpson. Ces trois indices diffèrent par leur sensibilité
vis-à-vis des espèces rares, le plus sensible étant la richesse spécifique et le moins sensible
l'indice de Simpson (Pavoine et al., 2005a).
L'indice de richesse se mesure par nombre d'entités S moins 1, afin qu'une collection ne
contenant qu'une entité ait une richesse nulle :
1rH S= −
Les deux indices suivant tiennent compte de l'abondance relative de chacune des espèces.

24
L'indice de Shannon-Wiener (Shannon 1948) corrige l'indice de richesse par les abondances
des catégories transformées en fréquences :
1( ) ln( )
s
s w k kk
H p p p−=
= −∑
avec p l'ensemble des fréquences associées à chaque catégorie k sachant S le nombre de
catégories.
L'indice de Gini-Simpson (Magurran, 2004, Simpson 1949) mesure la probabilité de tirer
deux entités appartenant à deux catégories différentes:
2
1( ) ( )
s
g s kk
H p p−=
= −∑
avec p l'ensemble des fréquences associées à chaque catégorie k, et S le nombre de catégories.
Enfin il existe des indices qui permettent de rendre compte de la diversité taxonomique,
phylogénétique ou fonctionnelle. Certains sont fondés sur des mesures de distance entre les
espèces représentées par un arbre, celui-ci peut être un arbre phylogénétique, une taxonomie
ou un dendrogramme construit à partir de distances fonctionnelles.
L’indice de diversité taxonomique de Warwick & Clarke (1995) correspond à la
longueur moyenne du chemin dans un arbre taxonomique entre deux catégories choisies
aléatoirement dans la collection.
0. ( 1) / 2
( 1) / 2ij i j i ii j i
i j i ii j i
w x x x x
x x x x<
<
+ −∆ =
+ −∑∑ ∑∑∑ ∑
avec xi l'abondance associée à l'espèce i et wij le poids liant les deux espèces i et j selon la
classification hiérarchique (Fig. 3). L'indication « 0. », peu commune, indique qu'une distance
séparant deux individus de la même espèce est égale à 0 comme on peut le voir entre les
individus e et f de la Fig. 3. Cet indice tient donc compte de tous les niveaux taxonomiques.

25
Fig. 3 : Arbre représentant les différents niveaux taxonomiques pour le calcul de l'indice de diversité
taxonomique. Les valeurs représentent l’indice de diversité taxonomique selon Warwick & Clarke (1995). Le
poids des couples d'individus vaut 1 si on mesure la diversité taxonomique des individus a et b, 2 pour les
individus c et d et 0 pour les individus e et f. Modifiée à partir de Warwick & Clarke (1995)
L'indice de diversité phylogénétique (PD) de Faith (1992) prend appui sur les travaux
de Vane-Wright (1991) et May (1990) qui ont mis en place des indices de diversité
phylogénétique sensibles à l'originalité de l'histoire évolutive dans le but de mettre en place
des listes de priorités de conservation reposant sur cette forme de « rareté » évolutive.
L’indice PD permet de rendre compte de la variabilité des histoires évolutives des catégories
au sein d'une collection de manière globale sans attribuer des poids d'originalité à certaine
catégories. A partir d'un arbre phylogénétique, la diversité phylogénétique d'un sous-groupe
de catégories (feuilles de l'arbre) est égale à la somme des longueurs de branches reliant ces
catégories entre elles (Fig. 4).
Fig. 4 : Arbre représentant les différents niveaux taxonomiques pour le calcul de l'indice de diversité
phylogénétique. Sous-ensemble d'un arbre de 10 taxons qui regroupe ici les taxons 2, 6, 8 et 10. Les marques
noires représentent une unité de longueur de branches représentatives de l'évolution d'état de caractères. Pour le
sous-ensemble des taxons 2, 6, 8 et 10, la diversité phylogénétique (PD) est égale à 28. Modifiée à partir de Faith
(1992).

26
L'indice de diversité fonctionnelle (FD) (Petchey & Gaston, 2002) reflète la diversité
des traits fonctionnels ayant un rôle dans le fonctionnement des écosystèmes. Des méthodes
existent pour obtenir des distances fonctionnelles à partir de plusieurs types de variables, par
exemple de type binaire, ordinale ou nominales (Pavoine et al., 2009). A partir de ces
distances, il est possible de construire un dendrogramme. Il existe plusieurs méthodes de
construction de dendrogramme que nous ne détaillerons pas ici (pour plus d'information voir
l'article de Mouchet et al. (2008). L’indice FD est proposé sur la base de calcul de l'indice de
diversité phylogénétique (PD). Il s’agit de la somme des longueurs de branches sur un
dendrogramme fonctionnel qui contient toutes les espèces d’une communauté.
L'indice de diversité de Rao (1986) appelé entropie quadratique est de plus en plus
utilisé en écologie (Chave et al. 2007, Pavoine et al., 2004 ; Pavoine et al., 2010). Il permet de
mesurer la diversité d'une communauté d'espèce (collection de catégories) à partir des valeurs
d'abondance relative et de différence entre les espèces (catégories) (Fig. 5).
Fig. 5 : Schéma des éléments utilisés pour le calcul de l'entropie quadratique. Les deux tables nécessaires au
calcul de l'entropie quadratique, à gauche, la matrice de distance entre les différentes catégories, à droite la table
des fréquences de chaque catégorie au sein de chaque collection. Comme exemple, identifions les collections
comme des communautés et les catégories comme les espèces dont l'abondance relative varie en fonction des
individus recensés pour chaque espèce. La matrice de distance pourrait être représentée par des distances
phylogénétiques entre les espèces deux à deux et la table des fréquences indiquerait pour chaque communauté k,
les abondances relatives des n espèces. Par le calcul de l'entropie quadratique, une diversité phylogénétique serait
obtenue pour chaque communauté tenant compte à la fois des différences entre espèces et des valeurs
d'abondance relative.

27
L'entropie quadratique de Rao (1986) se calcule selon la formule suivante:
1 1( )
n n
D i j iji j
H p p p d= =
=∑∑
avec dij la matrice de distance, p l'ensemble des fréquences des n catégories d'une collection, i
et j représentant les catégories. Cet indice est une généralisation de l'indice de Gini-Simpson,
les deux indices étant équivalents dans le cas particulier où toutes les valeurs de la matrice D
sont égales à 1 hormis pour les valeurs de la diagonale qui sont égales à 0. Champely &
Chessel (2002) ont démontré l'avantage d'utiliser des distances euclidiennes qui permettent de
quantifier les diversités et de les comparer dans un espace euclidien qui est géométriquement
et graphiquement interprétable. De plus, Pavoine et al. (2005b) utilisent différent types de
matrices euclidiennes de distances entre espèces et démontrent l'intérêt des distances dites
« ultramétriques ». En effet cette propriété mathématique permet d'avoir un indice qui n’est
jamais maximisé en enlevant une espèce d’une communauté. Ainsi selon cet indice, une
communauté idéale avec diversité maximale a toujours la richesse spécifique la plus grande
possible. Les transformations mathématiques amenant à des distances ultramétriques sont
nombreuses. Nous utiliserons dans le travail présenté ici la méthode UPGMA (Unweighted
Pair Group Method with Arithmetic mean ; Petchey et al., 2009).
Chao et al. (2010) remettent en cause les indices de diversité qui ne respectent pas le principe
de réplication à cause de leur relation non linéaire avec la richesse spécifique. Le principe de
réplication est le suivant : considérant N groupes de même diversité (nombre d'espèces,
abondance des espèces et différence entre les espèces), sans aucune espèce en commun, la
diversité de l'ensemble des N groupes doit être N fois la diversité d'un seul groupe. Chao et al.
(2010) montrent qu'un certain nombre d'indices classiquement utilisés comme par exemple
l'indice de Shannon, de Gini-Simpson et l'entropie quadratique de Rao ne respectent pas ce
principe. Les auteurs rappellent qu'il existe des transformations pour les indices de Shannon et
Gini-Simpson et proposent une transformation pour l'indice d'entropie quadratique afin qu'ils
respectent le principe de réplication (Table 1). Le calcul de T dépend de la qualité
ultramétrique ou non ultramétrique du dendrogramme. Dans le cas d'un arbre ultramétrique, T
est la longueur de l'arbre du premier nœud jusqu'à la dernière feuille. Dans le cas d'un arbre
non-ultramétrique, le calcul de T prend en compte les longueurs de branches.

28
Table 1 : Transformation appliquée sur les indices pour qu'ils respectent le principe de réplication
Indice Transformation appliquée
Shannon (S) exp( )S
Gini-Simpson (GS) 1(1 )GS−
Entropie quadratique (Q) Distances ultramétriques :
1
1 ( )QT
−; Distances non ultramétriques :
1
1 ( )QT
−
1.3 Priorité de conservation : l’intérêt des méthodes de Rapid assessment
Face à la crise écologique, l’urgence de la situation et le manque de moyens pour y faire face,
il a été proposé d’identifier des priorités de conservation, de concentrer les efforts de
conservation sur des espèces et/ou des habitats d’intérêt majeur. L’une des méthodes les plus
utilisées est l’identification d’espèces présentes sur la liste rouge éditée par l’UICN (Union
Internationale pour la Conservation de la Nature). Cette liste, basée sur des critères
démographiques et biogéographiques classent les espèces selon leur risque d’extinction à
court et moyen terme. Rendue publique à l’occasion de la Conférence des Nations Unies de
Rio de 2012 sur le développement durable, la nouvelle mise à jour de la Liste rouge de
l’UICN des espèces menacées montre que sur les 63 837 espèces étudiées, 19 817 sont
menacées d’extinction. D’autres critères identifient plus largement des zones d’intérêts, les
« hotspots » ou point chaud de la biodiversité en font partis. Le concept des hotspots est un
concept biogéographique. L’identification de lieux géographiques comme des points chauds
dépend d’une combinaison de valeur de richesse spécifique, de rareté des espèces, de leur
endémisme et des menaces, qu'elles soient naturelles ou anthropiques, qui pèsent sur ces
espèces (Myers et al., 2000). La classification de ces hotspots peut prendre en compte un ou
plusieurs de ces critères (Reid, 1998). Ainsi, pour proposer une classification des hotspots à
travers le monde, les célèbres travaux de Myers et al. (2000) se basent sur deux critères, la
mesure de l'endémisme et celle de la perte des habitats.
Ces deux méthodes sont fondées sur des caractéristiques particulières associées aux espèces.
Ainsi, une zone géographique pourrait être classée comme prioritaire en raison de la présence
d'une seule espèce. Les indices de biodiversité présentés en paragraphe 1.2.3 mesurent des
diversités prenant en compte toute la communauté d'espèces. Cependant, quelque soit les
indicateurs utilisés, les mesures de biodiversité ne sont pas toujours congruentes. Une seule
mesure d'un aspect de la biodiversité ne suffit donc pas pour rendre compte de la biodiversité
globale (Devictor et al., 2010 ; Duelli & Obrist, 2003). Ainsi, les travaux d’Orme et al. (2005)

29
montrent que les différents critères qui permettent de définir des hotspots ne sont pas toujours
convergents et que le choix de l'un ou l'autre peut conduire à des classifications
biogéographiques différentes. D'après cette étude, le critère de l'endémisme apparaît, en ce qui
concerne la diversité biologique de l'avifaune, comme l'indicateur englobant le plus les autres
critères étudiées, à savoir la richesse spécifique et les espèces menacées. Les travaux de
Devictor et al. (2010) comparent la répartition spatiale des diversités taxonomiques
fonctionnelles et phylogénétiques sur le territoire français et mettent en évidence le danger de
prendre un composant de la biodiversité comme représentant des autres. De plus, leurs
résultats révèlent une protection inégale des différentes facettes de la diversité par les aires
protégées. Ces études montrent bien la difficulté de mettre en place des priorités de
conservation et le choix des indices de biodiversité, notamment pour classer des zones
d'intérêt ou pour observer l'évolution d'un site dans le temps (Buckland et al., 2005).
Toutes ces mesures se basent sur un travail d'inventaire d'espèces des zones que l'on
veut évaluer. Face à la difficulté de faire un inventaire exhaustif, des méthodes d'inventaires
partiels, ou échantillonnages, ont été développées. Ces méthodes d'échantillonnage (transect,
quadrat, etc.) permettent de multiplier les répétitions spatiales et temporelles afin d'avoir une
meilleure estimation de l'objet d'étude (Sutherland, 2006). Cependant, ce travail nécessite
toujours une identification précise des échantillons. Face à la rapidité estimée de la
dégradation de la biodiversité, ce sont des résultats rapides d’évaluation qui sont attendus. La
notion d’inventaire d’espèces pour de telles évaluations soulève un problème logistique et
temporel. Les programmes ATBI (All Taxa Biodiversity Inventory) ont pour objectif de faire
des inventaires exhaustifs des espèces, mais ces études demandent la présence d'un grand
nombre de spécialistes pour couvrir la majorité des taxons et, de plus, sont très limités en
dimension spatiale et temporelle. De ce fait, les chercheurs ont élaboré de nouvelles méthodes
d’échantillonnage plus rapides car demandant moins de connaissances pour identifier les
individus échantillonnés.
Parmi ces méthodes rapides, nous distinguerons les méthodes taxonomiques des
méthodes non-taxonomiques. Deux méthodes taxonomiques peuvent être utilisées :
l'utilisation de taxons indicateurs (Jones & Eggleton, 2000 ; Pearson, 1994) et l'identification
taxonomique sur des niveaux supérieurs à celui de l'espèce (genre, famille, etc.) qui demande
moins d'expertise (Groc et al., 2010 ; Mazaris et al., 2008 ; Rosser & Eggleton, 2012). Parmi
les méthodes non-taxonomiques, nous distingueront la para-taxonomie et les méta-méthodes.
Toutes les méthodes non-taxonomiques se basent sur des critères non taxonomiques pour
catégoriser des unités. Ainsi, la para-taxonomie est une méthode de reconnaissance d’unité

30
taxonomique (RTU=Recognizable Taxonomic Units ; Oliver & Beattie, 1996 ; Oliver et al.,
2000) qui propose de ne pas identifier les individus collectés jusqu'à l’espèce mais de s’arrêter
aux morpho-espèces c'est-à-dire à des groupes reconnaissables facilement par la morphologie
(Basset et al., 2000 ; Kerr et al., 2000 ; Krell, 2004). Ainsi une morpho-espèce peut regrouper
plusieurs espèces taxonomiques. Enfin les méta-méthodes seront différenciées des méthodes
de para-taxonomie de par l'unité de description utilisée. En effet, si la para-taxonomie, comme
les méthodes taxonomiques, utilisent l'individu comme unité de description, les méta-
méthodes (méta-génomique, méta-acoustique ou Rapid Acoustic Survey) utilisent la
communauté comme unité de mesure. Dans la partie discussion, nous détaillerons ces
méthodes et les comparerons avec la méthode Rapid Acoustic Survey (RAS) avec pour
objectif de mettre en perspective les limites et les attentes de celle-ci.
Si la bioacoustique est depuis longtemps utilisée comme une méthode indirecte de
mesure de la biodiversité, la méthode RAS a été proposée très récemment (Sueur et al.,
2008b). Dans la section qui va suivre, nous présenterons la bioacoustique de manière générale
puis nous expliquerons l'utilité des méthodes d'acoustique passive pour l'estimation de la
biodiversité. Enfin, nous présenterons la méthode RAS, sur laquelle portent les travaux de
recherche présentés dans les chapitres suivants.
2 L’acoustique passive : un outil pour une méthode globale d’estimation de la biodiversité 2.1 Le son
2.1.1 Explication physique d'un phénomène ondulatoire
Le son est un phénomène ondulatoire. Il résulte de vibrations de molécules. A la source de
tout son, il y a la vibration d'un corps, vibration qui se transmet alors dans le milieu dans
lequel il se trouve, de proche en proche, d'une molécule à l'autre, permettant un transport
d'énergie sans transport de matière (Fischetti, 2003). Ainsi, le son se propage dans l'air mais
également dans tous les milieux comme par exemple l'eau, le sol, le bois et les métaux.
L'exemple des ondes produites par un caillou que l'on jette dans l'eau illustre la formation et la
propagation des ondes au travers de la matière.
Le son est donc du à une variation de pression de la matière. Des molécules, mises en
vibration par une perturbation initiale, se compressent dans un sens puis se décompressent
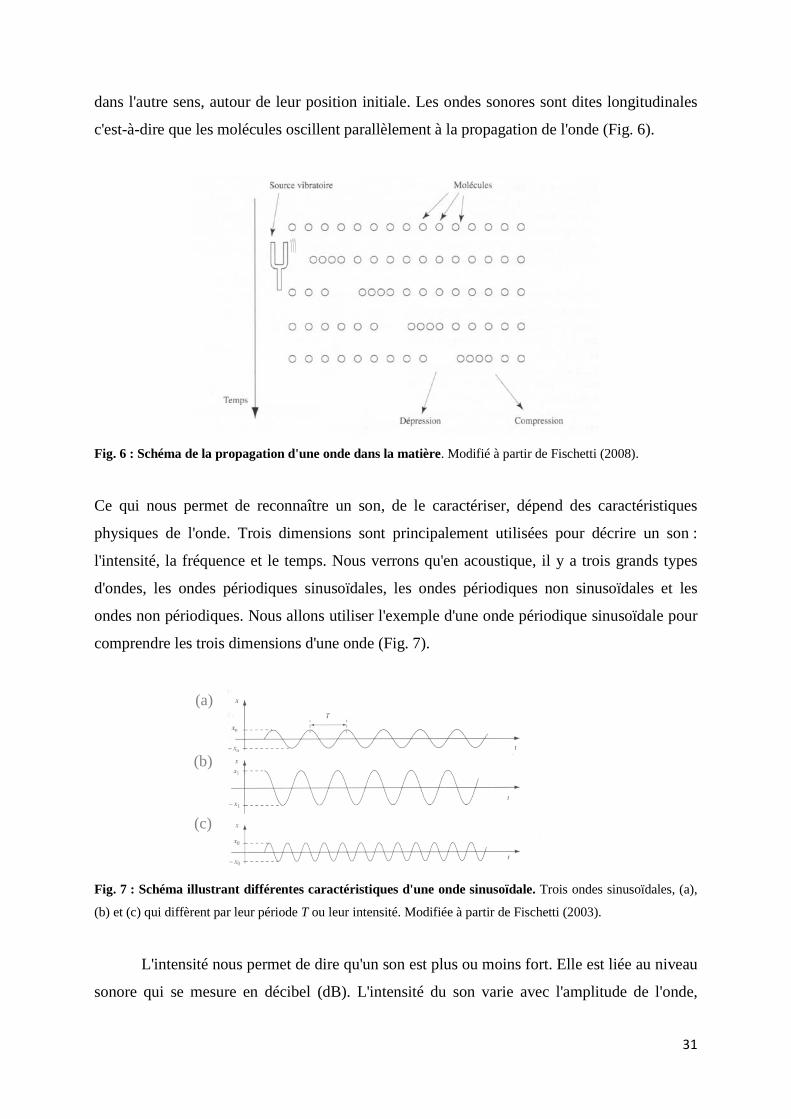
31
dans l'autre sens, autour de leur position initiale. Les ondes sonores sont dites longitudinales
c'est-à-dire que les molécules oscillent parallèlement à la propagation de l'onde (Fig. 6).
Fig. 6 : Schéma de la propagation d'une onde dans la matière. Modifié à partir de Fischetti (2008).
Ce qui nous permet de reconnaître un son, de le caractériser, dépend des caractéristiques
physiques de l'onde. Trois dimensions sont principalement utilisées pour décrire un son :
l'intensité, la fréquence et le temps. Nous verrons qu'en acoustique, il y a trois grands types
d'ondes, les ondes périodiques sinusoïdales, les ondes périodiques non sinusoïdales et les
ondes non périodiques. Nous allons utiliser l'exemple d'une onde périodique sinusoïdale pour
comprendre les trois dimensions d'une onde (Fig. 7).
Fig. 7 : Schéma illustrant différentes caractéristiques d'une onde sinusoïdale. Trois ondes sinusoïdales, (a),
(b) et (c) qui diffèrent par leur période T ou leur intensité. Modifiée à partir de Fischetti (2003).
L'intensité nous permet de dire qu'un son est plus ou moins fort. Elle est liée au niveau
sonore qui se mesure en décibel (dB). L'intensité du son varie avec l'amplitude de l'onde,
(a)
(b)
(c)

32
c'est-à-dire la distance parcourue par la molécule. Par exemple, la distance entre -x0 et x0 est
plus grande pour la Fig. 7 (a) que la distance entre -x1 et x1 pour la Fig. 7 (b). La valeur de 0
dB correspond à la plus petite intensité audible par l'oreille humaine et 120 dB à la limite de
déchirement du tympan.
La fréquence varie avec la vitesse de vibration des molécules. Plus cette vitesse est
rapide, plus le son est aigu. La mesure de cette vitesse s'exprime en Hertz (Hz). Par exemple,
pour une vitesse équivalente à 400 allers-retours d'une molécule en une seconde, la fréquence
est de 400 Hz. Sur une onde sinusoïdale simple, cette fréquence est l'inverse de la période T
(Fig. 7). Ainsi la fréquence sera plus élevée pour l'onde représentée en Fig. 7 (a) et celle en
Fig. 7 (c). L'oreille humaine peut détecter des fréquences de 20 à 20 000 Hz. En dessous de 20
Hz, les vibrations sont trop lentes pour être détectées par l'oreille humaine, ces sons sont
appelés des infrasons. Au dessus de 20 000Hz, ces vitesses de vibrations sont trop élevées, on
appelle alors ces sons des ultrasons.
L'amplitude ou la fréquence peuvent varier dans le temps. Le son dans sa dimension
temporelle peut alors être défini selon sa durée et selon les modulations d'intensité (rythme) et
de fréquence.
2.1.2 Les séries et transformées de Fourier ou comment prendre des mesures d'un son
naturel par une décomposition en fréquence
Dans la nature, les sons sont très rarement purs. Il est donc beaucoup plus fréquent d'entendre
des sons non périodiques (chute d'eau, chœurs d'oiseaux). Comment déterminer la ou les
fréquences de ces sons complexes ? Cette question a été traitée par un certain nombre de
chercheurs dont Joseph Fourier (1768-1830) fut le précurseur.
Fig. 8 : Illustrations de l'ouvrage « Théorie analytique de la chaleur » de Joseph Fourier, à droite une
gravure représentant Joseph Fourier et à gauche la couverture de son ouvrage « Théorie analytique de la
chaleur ». Extrait de Burke (1995).
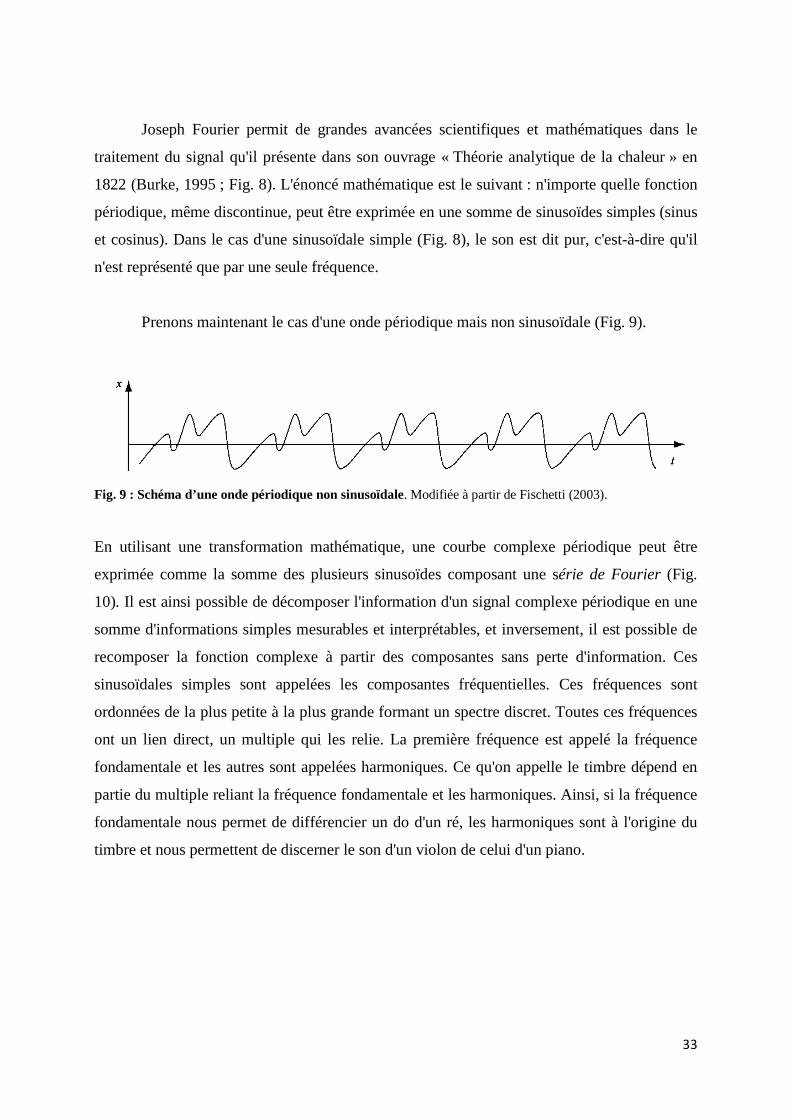
33
Joseph Fourier permit de grandes avancées scientifiques et mathématiques dans le
traitement du signal qu'il présente dans son ouvrage « Théorie analytique de la chaleur » en
1822 (Burke, 1995 ; Fig. 8). L'énoncé mathématique est le suivant : n'importe quelle fonction
périodique, même discontinue, peut être exprimée en une somme de sinusoïdes simples (sinus
et cosinus). Dans le cas d'une sinusoïdale simple (Fig. 8), le son est dit pur, c'est-à-dire qu'il
n'est représenté que par une seule fréquence.
Prenons maintenant le cas d'une onde périodique mais non sinusoïdale (Fig. 9).
Fig. 9 : Schéma d’une onde périodique non sinusoïdale. Modifiée à partir de Fischetti (2003).
En utilisant une transformation mathématique, une courbe complexe périodique peut être
exprimée comme la somme des plusieurs sinusoïdes composant une série de Fourier (Fig.
10). Il est ainsi possible de décomposer l'information d'un signal complexe périodique en une
somme d'informations simples mesurables et interprétables, et inversement, il est possible de
recomposer la fonction complexe à partir des composantes sans perte d'information. Ces
sinusoïdales simples sont appelées les composantes fréquentielles. Ces fréquences sont
ordonnées de la plus petite à la plus grande formant un spectre discret. Toutes ces fréquences
ont un lien direct, un multiple qui les relie. La première fréquence est appelé la fréquence
fondamentale et les autres sont appelées harmoniques. Ce qu'on appelle le timbre dépend en
partie du multiple reliant la fréquence fondamentale et les harmoniques. Ainsi, si la fréquence
fondamentale nous permet de différencier un do d'un ré, les harmoniques sont à l'origine du
timbre et nous permettent de discerner le son d'un violon de celui d'un piano.
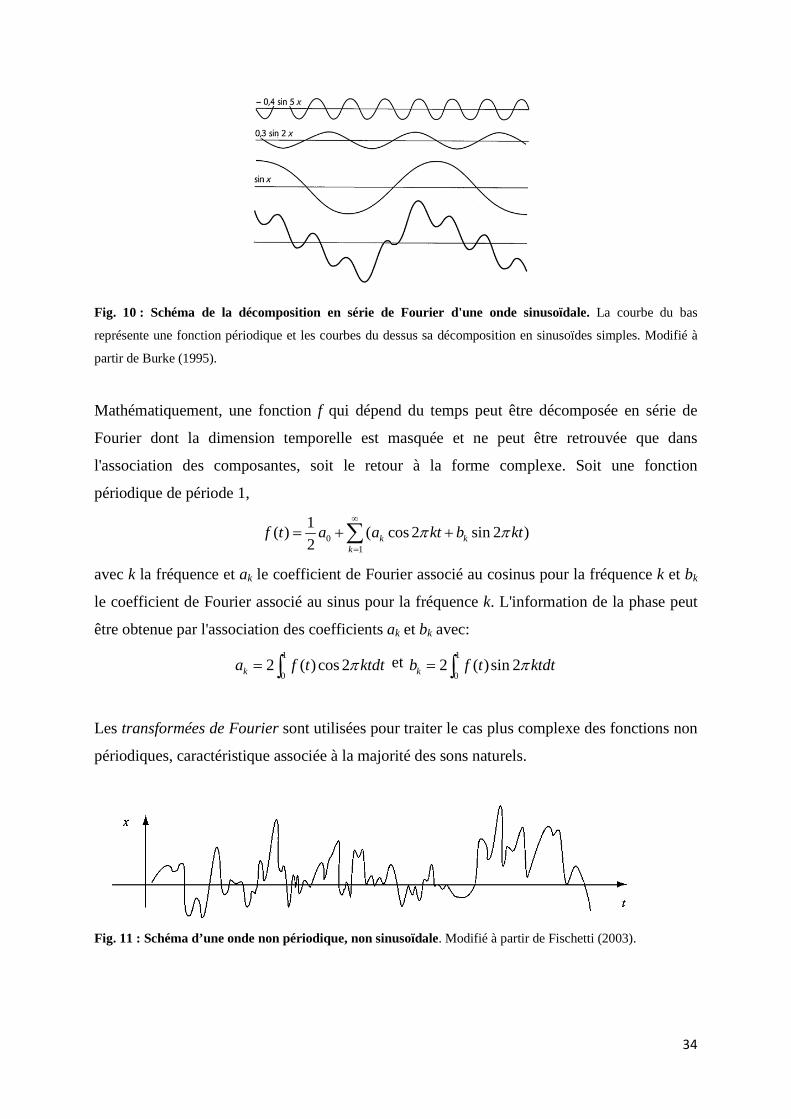
34
Fig. 10 : Schéma de la décomposition en série de Fourier d'une onde sinusoïdale. La courbe du bas
représente une fonction périodique et les courbes du dessus sa décomposition en sinusoïdes simples. Modifié à
partir de Burke (1995).
Mathématiquement, une fonction f qui dépend du temps peut être décomposée en série de
Fourier dont la dimension temporelle est masquée et ne peut être retrouvée que dans
l'association des composantes, soit le retour à la forme complexe. Soit une fonction
périodique de période 1,
01
1( ) ( cos 2 sin 2 )2 k k
kf t a a kt b ktπ π
∞
=
= + +∑
avec k la fréquence et ak le coefficient de Fourier associé au cosinus pour la fréquence k et bk
le coefficient de Fourier associé au sinus pour la fréquence k. L'information de la phase peut
être obtenue par l'association des coefficients ak et bk avec: 1
02 ( ) cos 2ka f t ktdtπ= ∫ et 1
02 ( )sin 2kb f t ktdtπ= ∫
Les transformées de Fourier sont utilisées pour traiter le cas plus complexe des fonctions non
périodiques, caractéristique associée à la majorité des sons naturels.
Fig. 11 : Schéma d’une onde non périodique, non sinusoïdale. Modifié à partir de Fischetti (2003).

35
Les transformées de Fourier sont des dérivées des séries de Fourier. La différence réside dans
le fait que pour une série de Fourier, toutes les fréquences des sinusoïdes sont reliées par un
multiple entier. Dans le cas d'une fonction non périodique (Fig. 11), il est possible de
décomposer le signal en série de sinus et cosinus sur l’ensemble du spectre fréquentiel
(spectre continu). Ainsi on obtient les coefficients pour toutes les fréquences par les formules
suivantes:
( ) ( ) cos 2a f t tdtτ πτ+∞
−∞= ∫ et ( ) ( )sin 2b f t tdtτ πτ
+∞
−∞= ∫
où, τ est l'ensemble des fréquences.
La FFT (Fast Fourier Transform) est un algorithme présenté par James Cooley et John Tukey
en 1965 qui a permis d'accélérer le temps de calcul en passant de n2 à n*log2*n le nombre de
calculs nécessaires pour obtenir la transformée de Fourier d'une fonction, de n valeurs en se
basant sur des calculs matriciels. Le gain de temps de calcul augmente avec le nombre de
valeur n composant le signal sonore (Burke, 1995).
2.1.3 Numérisation du son
Concrètement, le mouvement des molécules d'air se traduit par une variation de pression de
l'air. Cette variation de pression met en mouvement la membrane d'un microphone qui
retranscrit cette variation en signal électrique, signal qui peut ensuite être numérisé. La
numérisation du son revient à relever de petits échantillons de la différence de pression d'air à
intervalle de temps précis. Ce signal numérique dépend de plusieurs paramètres. La fréquence
d'échantillonnage est le nombre d'échantillons par seconde, se mesure en Hz et joue sur la
qualité du signal. La fréquence d'échantillonnage doit être suffisamment grande afin de
préserver la forme du signal. Le théorème de Nyquist-Shannon stipule que la fréquence
d'échantillonnage doit être égale ou supérieure au double de la fréquence maximale contenue
dans ce signal. Notre oreille perçoit les sons environ jusqu'à 20 000 Hz, il faut donc une
fréquence d'échantillonnage au moins de l'ordre de 40 000 Hz pour obtenir une qualité
satisfaisante à l'oreille humaine. Le nombre de bits d'un échantillon peut être codé en 8 bits ou
16 bits. Ainsi, un échantillon ou prise de mesure de la variation de pression de l'air est codé
par un nombre de valeurs plus ou moins grand. Cela correspond à un nombre de valeurs
respectivement de 256 (2^8) ou 65 536 (2^16). Le nombre de voies influence également la
numérisation, une seule correspondant à de la monophonie, deux à de la stéréophonie, et
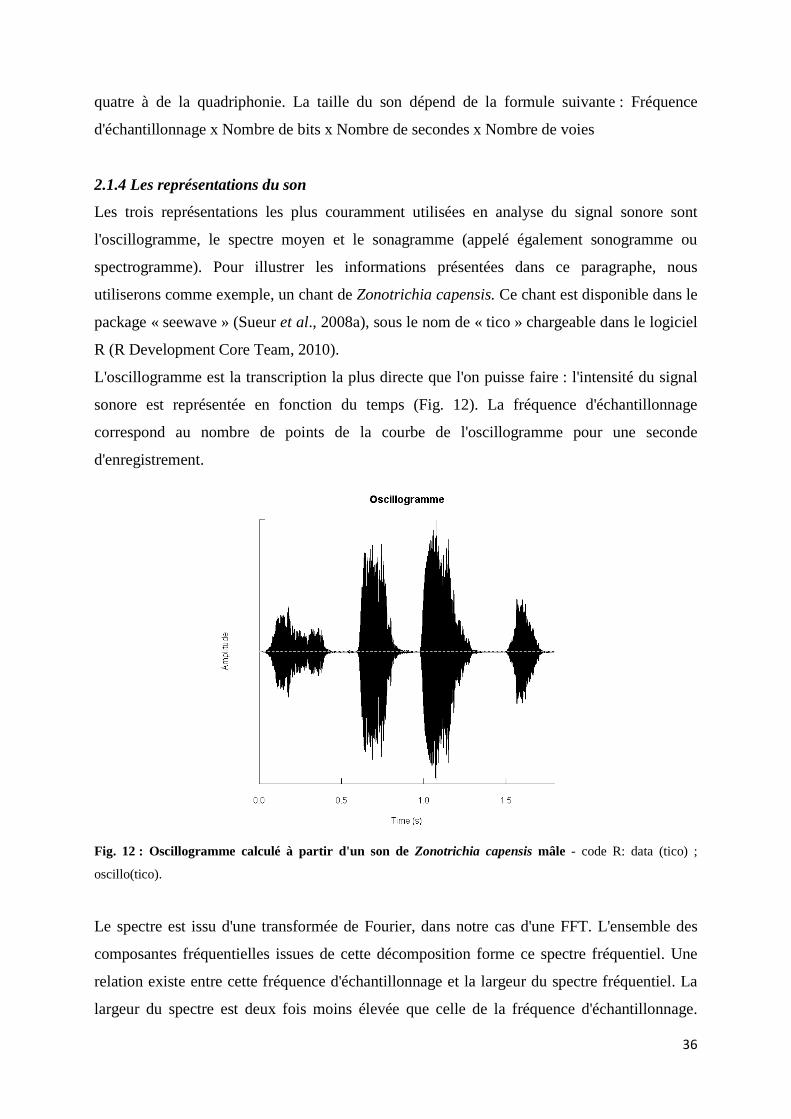
36
quatre à de la quadriphonie. La taille du son dépend de la formule suivante : Fréquence
d'échantillonnage x Nombre de bits x Nombre de secondes x Nombre de voies
2.1.4 Les représentations du son
Les trois représentations les plus couramment utilisées en analyse du signal sonore sont
l'oscillogramme, le spectre moyen et le sonagramme (appelé également sonogramme ou
spectrogramme). Pour illustrer les informations présentées dans ce paragraphe, nous
utiliserons comme exemple, un chant de Zonotrichia capensis. Ce chant est disponible dans le
package « seewave » (Sueur et al., 2008a), sous le nom de « tico » chargeable dans le logiciel
R (R Development Core Team, 2010).
L'oscillogramme est la transcription la plus directe que l'on puisse faire : l'intensité du signal
sonore est représentée en fonction du temps (Fig. 12). La fréquence d'échantillonnage
correspond au nombre de points de la courbe de l'oscillogramme pour une seconde
d'enregistrement.
Fig. 12 : Oscillogramme calculé à partir d'un son de Zonotrichia capensis mâle - code R: data (tico) ;
oscillo(tico).
Le spectre est issu d'une transformée de Fourier, dans notre cas d'une FFT. L'ensemble des
composantes fréquentielles issues de cette décomposition forme ce spectre fréquentiel. Une
relation existe entre cette fréquence d'échantillonnage et la largeur du spectre fréquentiel. La
largeur du spectre est deux fois moins élevée que celle de la fréquence d'échantillonnage.
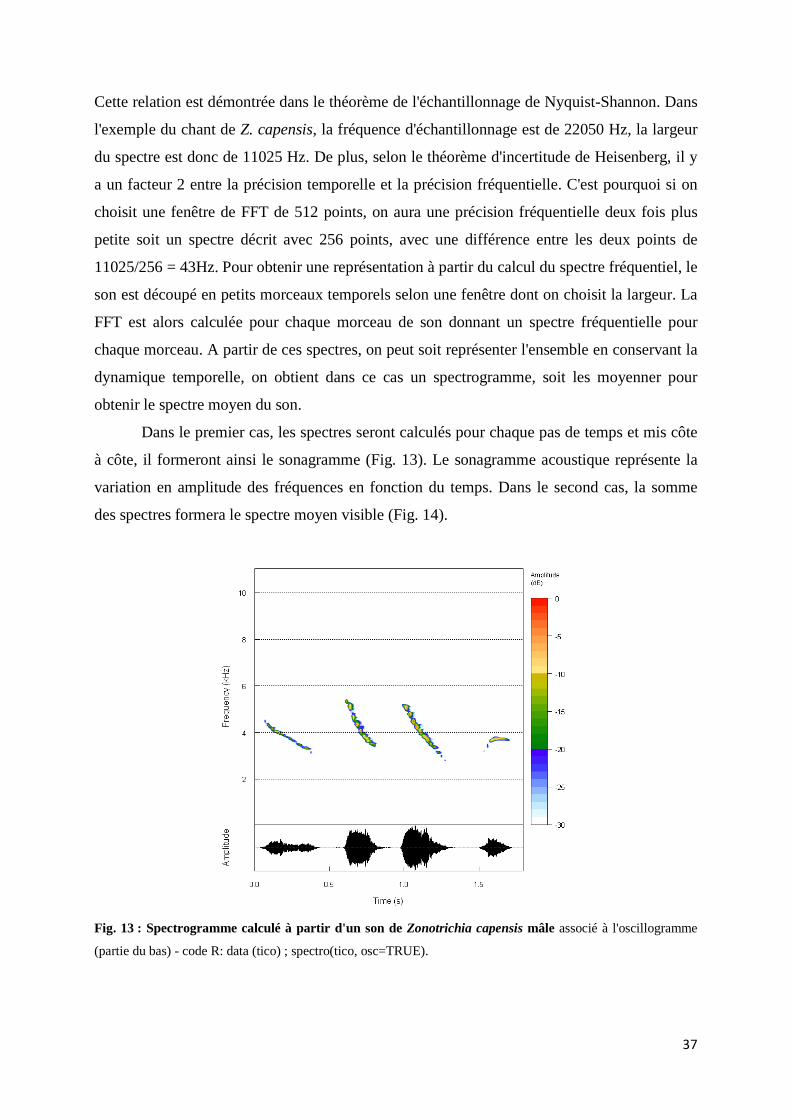
37
Cette relation est démontrée dans le théorème de l'échantillonnage de Nyquist-Shannon. Dans
l'exemple du chant de Z. capensis, la fréquence d'échantillonnage est de 22050 Hz, la largeur
du spectre est donc de 11025 Hz. De plus, selon le théorème d'incertitude de Heisenberg, il y
a un facteur 2 entre la précision temporelle et la précision fréquentielle. C'est pourquoi si on
choisit une fenêtre de FFT de 512 points, on aura une précision fréquentielle deux fois plus
petite soit un spectre décrit avec 256 points, avec une différence entre les deux points de
11025/256 = 43Hz. Pour obtenir une représentation à partir du calcul du spectre fréquentiel, le
son est découpé en petits morceaux temporels selon une fenêtre dont on choisit la largeur. La
FFT est alors calculée pour chaque morceau de son donnant un spectre fréquentielle pour
chaque morceau. A partir de ces spectres, on peut soit représenter l'ensemble en conservant la
dynamique temporelle, on obtient dans ce cas un spectrogramme, soit les moyenner pour
obtenir le spectre moyen du son.
Dans le premier cas, les spectres seront calculés pour chaque pas de temps et mis côte
à côte, il formeront ainsi le sonagramme (Fig. 13). Le sonagramme acoustique représente la
variation en amplitude des fréquences en fonction du temps. Dans le second cas, la somme
des spectres formera le spectre moyen visible (Fig. 14).
Fig. 13 : Spectrogramme calculé à partir d'un son de Zonotrichia capensis mâle associé à l'oscillogramme
(partie du bas) - code R: data (tico) ; spectro(tico, osc=TRUE).

38
Fig. 14 : Spectre moyen calculé à partir d'un son de Zonotrichia capensis mâle. La largeur de la fenêtre de
FFT est de 512 points donnant un spectre moyen formée de 256 points soit autant de fréquences représentées et
une précision fréquentielle de 43 Hz. L'amplitude ici est maximisée à 1 - code R: data (tico) ; meanspec(tico).
2.2 La production sonore animale
La plupart des animaux communiquent entre eux. Pour ce faire, ils envoient des signaux vers
un ou plusieurs récepteurs. Ces signaux peuvent être visuels (couleurs, mouvement),
chimiques (odeurs, phéromones de contact), électriques, tactiles ou sonores (Bradbury &
Vehrencamp, 1998). La communication acoustique permet aux individus de signaler leur
présence facilitant la rencontre avec d'autres individus (notamment pour la reproduction ou le
rassemblement en groupes sociaux) ou la défense de leur territoire. L'acoustique est un mode
de communication très utile en cas de non visibilité, comme par exemple dans le feuillage
pour les oiseaux ou dans les profondeurs de l'océan pour les cétacés, ou en cas d'urgence du
fait de sa rapidité. Les animaux peuvent également utiliser les sons pour localiser une proie ou
se déplacer dans le milieu par l'écholocation.
La majorité des signaux émis par les animaux sont organisés en unités sonores et émises selon
un rythme particulier. La communication acoustique se retrouve chez les oiseaux, les
mammifères terrestres et marins, les amphibiens, les poissons, les reptiles, les insectes et
certains autres arthropodes (Greenfield, 2002 ; Leroy, 1979).

39
Il y a une grande variabilité d'appareils de production et de réception du son, ainsi que
des substrats utilisés pour la transmission du signal (air, eau, végétaux, sol). Cinq types
d'appareil vibratoires ont été recensés (Bradbury & Vehrencamp, 1998). Le son peut être issu
de la vibration d'une membrane ou d'un sac par une action musculaire, ce que font par
exemple les cigales. Il peut également être initié par la trémulation (vibration d'un appendice
ou du corps entier) par une action musculaire. La stridulation est l'action d'une partie du corps
sur une autre produisant un son. Cet appareil vibratoire est présent chez la plupart des
arthropodes qui stridulent par exemple par un contact aile -aile (les grillons) ou aile-patte (les
criquets). La quatrième façon de produire du son est de contraindre les molécules, que ce soit
en milieu aérien ou aquatique, à passer par un orifice. C'est par exemple le cas des vibrations
des cordes vocales par le passage forcé de l'air à travers le larynx, système que l'on retrouve
chez les Anoures et beaucoup de Mammifères. Enfin, des vibrations peuvent être provoquées
par percussion sur un substrat. Cet appareil vibratoire est retrouvé chez certaines araignées et
insectes.
Il existe également différent récepteurs du son. Cette réception peut se faire soit par
des capteurs de mouvement du milieu comme par exemple les antennes du moustique qui
vont détecter les mouvements de l'air, ou des détecteurs de variation de pression du milieu
comme le tympan ou des membranes de fonction similaire. Les spectres fréquentiels
d'émission du son sont variables par rapport aux spectres fréquentiels de réception pour
chaque espèce et est bien sûr différent entre les espèces (Tanzarella, 2006). Si l'oreille
humaine ne capte que des sons de 20Hz à 20kHz, les appareils auditifs des animaux peuvent
être sensibles à des ondes de fréquences plus basses (les infrasons) ou plus hautes (les
ultrasons). Les infrasons ont la qualité de pouvoir parcourir de longues distances, qualité
utilisée par exemple par les éléphants (Langbauer, 2000). Les ultrasons sont utilisés pour
l'écholocation en raison de la rapidité de leur propagation. Ce mode d'utilisation des ondes
sonores n'est retrouvé que chez certains groupes tel que les chiroptères, les oiseaux et les
cétacés (Barataud, 2012 ; Leroy, 1979).
Toutes ces variations entrainent une grande diversité de sons, diversité inter-
spécifique, mais également intra-spécifique, appelée dans ce deuxième cas « répertoire
acoustique de l'espèce ». Cette variabilité des sons émis par les animaux se caractérise par des
différences d'utilisation de fréquences mais également par des différences de rythme. La
variation de ces deux composantes acoustiques, qui sont la composante temporelle et la
composante fréquentielle, permet le codage de l'information.

40
Plusieurs hypothèses évolutives ont été avancées pour essayer d'expliquer cette variabilité.
Pour arriver à communiquer, les espèces doivent échanger un signal clair. Dans le cas d'un
signal reproducteur, les informations sont l'espèce, le sexe (et dans certains cas la qualité du
reproducteur) et la localisation. Cela peut également être un son porteur d'une autre
information comme par exemple le cri d'alarme ou de regroupement. Dans tous les cas, le but
est de se faire entendre par le récepteur. Deux contraintes sont généralement considérées pour
expliquer l'évolution des chants : la compétition acoustique qui contraint à la diversification et
le filtre d'habitat qui lui contraint les paramètres physiques du milieu dans lequel se déplace le
son.
Sous la contrainte de la compétition pour la ressource de l'espace sonore, une grande
diversité de fréquences utilisées, de modulation d'amplitude et/ou de fréquence dans le temps
et de syntaxe ont émergé (Bradbury & Vehrencamp, 1998). Par exemple, la fréquence de
répétition d'un signal dans le temps permet d'augmenter les chances de réception du signal et
des informations qu'il transporte. La séparation de la période d'activité dans le temps, à
l'échelle de la journée (Ficken et al., 1985) ou de la saison (Gottsberger & Gruber, 2004), ou
dans l'espace en choisissant des postes de chants à différentes hauteurs (Diwakar &
Balakrishnan, 2007a) a pour conséquence un évitement de chevauchement des signaux
acoustiques et donc une diminution de la compétition ou un évitement des prédateurs. Il en est
de même pour la séparation des fréquences utilisées pour éviter le chevauchement à un même
temps t (Sueur et al., 2002). Ainsi, certains travaux suggèrent que le succès de communication
pourrait être augmenté avec la séparation acoustique fréquentielle, temporelle et/ou spatiale
de certaines espèces (Garcia-Rutledge & Narins, 2001 ; Chek et al., 2003).
Un certain nombre de caractéristiques de l'habitat influe sur la qualité physique de
l'onde sonore et donc de la transmission du signal vers le récepteur. La température et
l'humidité peuvent faire varier l'absorption atmosphérique. La densité de la végétation peut
atténuer le son et/ou provoquer la réverbération des ondes. Par exemple, la présence de troncs
et de feuilles pour les milieux fermés sont à l'origine de phénomènes de réflexion des ondes
sonores (Slabbekoorn, 2004 ; Wiley & Richards, 1978). En milieu forestier, l'absorption est
plus forte pour les hautes fréquences, les basses fréquences se propageant sur de plus grande
distance (Richards & Wiley, 1980). Le type de milieu (aérien, aquatique, solide) possède des
propriétés différentes quand à la propagation du son (Forest et al., 1993 ; Richards & Wiley,
1980). Enfin, le bruit ambiant, différent selon les types d'habitats, affecte la détectabilité du
signal sonore (Slabbekoorn, 2004).

41
Il existe également une contrainte morphologique issue de l'histoire évolutive des
espèces. Plusieurs études ont montré une corrélation négative entre le poids (ou la taille) et la
hauteur des fréquences utilisées chez les oiseaux (Wallschlager, 1980, Martin et al., 2011)
mais aussi dans d'autres groupes (Fletcher, 2007).
Toutes ces contraintes, liées à l'habitat ou à l'interaction entre les individus inter- ou intra-
spécifique, sont supposées influencer l'évolution des signaux acoustiques des espèces. Il a été
montré à plusieurs reprises que, sous la pression d'un même type d'habitat, les espèces ont
tendance à se ressembler acoustiquement (Boncoraglio & Saino, 2007 ; Bradbury &
Vehrencamp, 1998 ; Wiley, 1991). Si un grand nombre de stratégies visent à se différencier le
plus possible des autres que ce soit par un partitionnement des fréquences, ou de la période
d'activité, comment expliquer les phénomènes de chœurs ? Les chœurs sont définit comme
une convergence d'activité au même moment de la journée qui aurait pour risque une
interférence des signaux acoustiques. Plusieurs hypothèses ont été envisagées pour expliquer
ce phénomène : l'hypothèse de transmission acoustique expliquerait ce phénomène par des
conditions atmosphériques optimales pour la transmission acoustique (Henwood & Fabrick,
1979 ; Brown & Handford, 2003) qui a été testée sans succès en ce qui concerne les chœurs
d'oiseaux qui ont lieu à l'aube (Dadelsteen & Mathevon, 2002), l'hypothèse de protection
contre les prédateurs expliquerait ce phénomène par le fait que chanter en même temps
rendrait la localisation plus difficile pour les prédateurs (Wolda, 1993), enfin l'hypothèse
d'inefficacité de recherche de nourriture explique les chœurs par le fait que l'activité
acoustique est reliée au niveau de luminosité qui étant trop faible, ne permet pas la recherche
de nourriture (Hutchinson, 2002).
2.3 L'acoustique passive
La bioacoustique est une science pluridisciplinaire qui se situe, comme son nom l'indique,
entre l'acoustique physique et la biologie. Elle est utilisée dans différents domaines d'études,
que ce soit en éthologie pour essayer d'expliquer des comportements, en neurophysiologie
afin de comprendre comment l'information portée par le signal acoustique est traitée par le
système nerveux, en biologie évolutive où est étudiée par exemple la sélection sexuelle par le
choix du signal mâle par les femelles et en écologie afin de comprendre les interactions entre
individus chanteurs et le milieu qu'il soit biotique ou abiotique (Ewing, 1989 ; Bradbury &
Vehrencamp, 1998 ; Gerhardt & Huber, 2002).

42
L'acoustique passive est utilisée pour répondre à des questions qui se posent à
différentes échelles d'études écologiques :
Individu. Cette échelle est la plus petite recensée en acoustique passive. Des travaux de
recherche travaillent à la localisation d'individus dans le temps et l'espace (Huetz & Aubin,
2012 ; Mennill et al., 2006 ; Hammer et al., 2001), d'autres sur la signature acoustique
individuelle (Pollard et al., 2010), d'autres encore sur le lien entre comportement et activité
acoustique (Gess et al., 2011) ou encore sur le lien entre acoustique et contrainte
morphologique (Mathias et al, 2009 ; Martin et al., 2011 ; Wallschlager, 1980).
Population. A l'échelle de la population, un grand nombre de questions peuvent être
abordées par l'acoustique passive comme par exemple la taille ou la densité des populations
d'oiseaux (Dawson & Efforf, 2009) ou d'insectes (Mankin et al., 2001 ; Forrest, 1988), la
structure des populations (Laiolo et al., 2008), le suivi temporel et spatial de population à
l'échelle locale ou régionale (Julliard et al., 2004) permettant de décrire les migrations ou la
distribution spatiale des populations.
Espèce. Au niveau spécifique, l'acoustique passive est essentiellement utilisée pour
identifier les espèces, décrire leur biologie et écologie (Obrist et al., 2010).
Communauté. Au niveau des communautés, l'utilisation de l'acoustique passive donne
accès à des informations sur la structure des communautés (Diwakar & Balakrishnan, 2007a,
2007b ; Riede, 1993, 1997). Des méthodes de mesures de diversités acoustiques des
communautés animales se sont développées. Ces Rapid Biodiversity Assessment (RBA)
utilisant l'acoustique passive seront détaillées dans le paragraphe 2.3.3 de cette introduction.
Paysage. Le concept du paysage sonore (soundscape) couple les caractéristiques de
l'échelle du paysage (landscapes) et de l'acoustique (Pijanowski et al., 2011a, 2011b, Farina &
Pieretti, 2012). Le paysage sonore est composé de sons issus de l'activité acoustique des
organismes ou « biophony », de sons issus de l'activité anthropique ou « antropophony » et de
l'activité non-biologique ou « geophony ». Ainsi la « soundscape ecology » est l'étude des
interactions entre les trois différentes parties du paysage sonore et l'impact de ces interactions
sur les communautés humaines et naturelles (Pijanowski et al., 2011b). Une des principales
questions qui se pose à cette échelle est l'impact des sons d'origine anthropique sur la
communication acoustique des espèces animales et les conséquences biologiques de ces
perturbations (Barber et al., 2011 ; Joo et al., 2011).
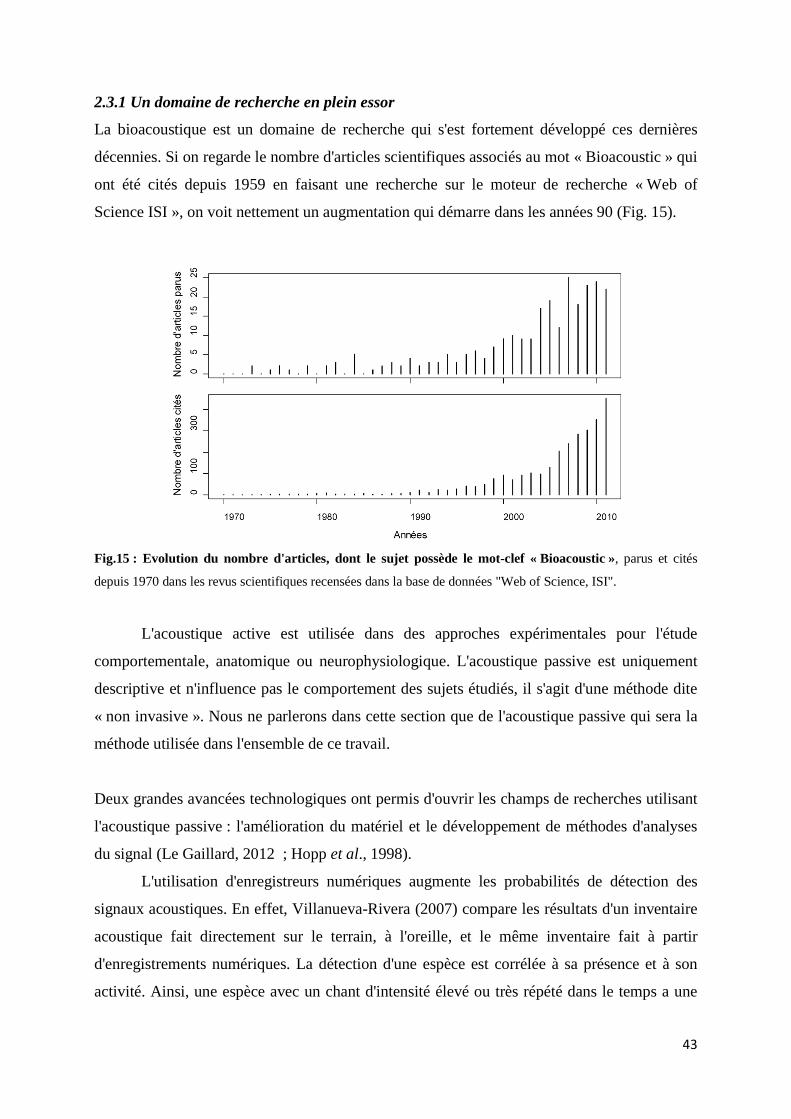
43
2.3.1 Un domaine de recherche en plein essor
La bioacoustique est un domaine de recherche qui s'est fortement développé ces dernières
décennies. Si on regarde le nombre d'articles scientifiques associés au mot « Bioacoustic » qui
ont été cités depuis 1959 en faisant une recherche sur le moteur de recherche « Web of
Science ISI », on voit nettement un augmentation qui démarre dans les années 90 (Fig. 15).
Fig.15 : Evolution du nombre d'articles, dont le sujet possède le mot-clef « Bioacoustic », parus et cités
depuis 1970 dans les revus scientifiques recensées dans la base de données "Web of Science, ISI".
L'acoustique active est utilisée dans des approches expérimentales pour l'étude
comportementale, anatomique ou neurophysiologique. L'acoustique passive est uniquement
descriptive et n'influence pas le comportement des sujets étudiés, il s'agit d'une méthode dite
« non invasive ». Nous ne parlerons dans cette section que de l'acoustique passive qui sera la
méthode utilisée dans l'ensemble de ce travail.
Deux grandes avancées technologiques ont permis d'ouvrir les champs de recherches utilisant
l'acoustique passive : l'amélioration du matériel et le développement de méthodes d'analyses
du signal (Le Gaillard, 2012 ; Hopp et al., 1998).
L'utilisation d'enregistreurs numériques augmente les probabilités de détection des
signaux acoustiques. En effet, Villanueva-Rivera (2007) compare les résultats d'un inventaire
acoustique fait directement sur le terrain, à l'oreille, et le même inventaire fait à partir
d'enregistrements numériques. La détection d'une espèce est corrélée à sa présence et à son
activité. Ainsi, une espèce avec un chant d'intensité élevé ou très répété dans le temps a une

44
probabilité de détection plus forte et peut cacher des signaux d'espèces dont l'intensité est plus
faible. Cette étude démontre que l'utilisation d'un enregistreur digital permet d'augmenter la
détection des signaux puisque l'analyse acoustique peut se faire à posteriori de
l'enregistrement, en ayant la possibilité de réécouter plusieurs fois l'enregistrement et d'utiliser
des outils de filtration qui permettent d'augmenter la probabilité de détection des signaux dans
le cas d'interférences acoustiques. Le fait de conserver les enregistrements permet également
des identifications par plusieurs experts et ainsi une diminution des erreurs d'identification
(Bridges & Dorcas, 2000).
La miniaturisation et l'automatisation des systèmes d'enregistrements (ARS,
Automatic Recording System) permettent un échantillonnage acoustique sur de larges
échelles temporelles et spatiales. L'utilisation de ces appareils minimise considérablement le
temps passé sur le terrain et permet de mettre en évidence des patrons jusque là impossible à
connaître en réalisant des points d'écoute sur de petites périodes de temps (Bridges & Dorcas,
2000). Que ce soit des enregistrements continus ou programmés, ces équipements
automatiques offrent de nouvelles perspectives de recherche et semblent plus performants que
les échantillonnages acoustiques par écoute directe (point d'écoute ou transects)
traditionnellement utilisés pour des études des communautés d'oiseaux et d'amphibiens
(Acevedo & Villanueava-Riviera, 2006).
L'amélioration du matériel passe également par le développement des microphones,
dont la capacité de détection est améliorée, et son utilisation dans d'autres milieux comme le
milieu aquatique est aujourd'hui possible grâce aux hydrophones (Hopp et al., 1998). De plus,
les microphones sont de plus en plus résistants que ce soit à la manipulation ou aux variations
de conditions climatiques qu'imposent le travail de terrain (température, humidité).
Les méthodes d'analyses du signal sont aujourd'hui facilitées par l'utilisation de
logiciels de traitement du signal développés par des entreprises privées ou des groupes de
recherche. Filtres, mesures automatiques, outils pour la transformation des données ou pour
l'analyse graphique sont autant de moyens qui facilitent le traitement des enregistrements et la
prise de mesure (Obrist et al., 2010).
De plus, ces méthodes possèdent des avantages par rapport aux inventaires classiques. Elles
permettent d'explorer des habitats dont l'accès est limité. C'est le cas de la canopée (Riede,
1997), des habitats aquatiques marins et terrestres (Luczkovich et al., 2008), du sol ou des
structures vivantes que sont les plantes (Mankin et al., 2000) et enfin, des environnements où
la lumière est faible ou absente (Meyer et al., 2011 ; Obrist et al., 2004). Cette méthode est

45
non-invasive et permet donc des échantillonnages dans des zones sensibles telles que les
réserves et ne nécessite pas de permis de capture. Grâce aux enregistreurs automatiques, le
temps passé sur le terrain est réduit ce qui permet d'entreprendre des échantillonnages à larges
échelles temporelle et spatiale. Enfin, l'échantillonnage est répétable et automatisable ce qui
permet des prises de mesure simultanées sur différents sites.
L'amélioration technique des équipements de plus en plus accessibles financièrement et de
plus en plus simples d'utilisation est une des explications du succès de la bioacoustique. Cette
barrière matérielle tombée, la bioacoustique s'est vue attribuer de nouveaux champs de
recherche.
2.3.2 Un outil pour estimer la biodiversité
L'importance de l'estimation de la biodiversité a été abordée dans la partie 1 de cette
introduction. La détection de signaux acoustiques peut fournir des informations sur l'âge, le
sexe, la localisation géographique de l'individu mais avant tout, des informations de la
présence et l'appartenance spécifique de l'individu chanteur. Il est donc possible de faire des
inventaires d'espèces par des techniques d'acoustique passive (Obrist et al., 2010).
L'utilisation de techniques d'acoustique passives ne permet qu'un accès limité à une partie de
la biodiversité, la diversité acoustique. De plus, cet accès à la diversité acoustique est lui-
même limité par la détection des signaux acoustiques et donc par le matériel utilisé (Fig. 16).
Fig. 16 : Méthode d'estimation de la biodiversité par des techniques d'acoustique passive. De gauche à
droite, cette figure présente (i) la restriction de l'accès à la biodiversité par l'acoustique, restriction qui concerne
la diversité animale chanteuse détectée par l'enregistreur ou l'observateur, (ii) les différentes méthodes
d'échantillonnage acoustique permettant d'obtenir des informations sur la biodiversité (listées dans le cadre rose)
et enfin (iii) les avantages (+) et les limites (-) associées à chacune de ces méthodes. Modifé de l'article de Obrist
et al. (2010).

46
Trois techniques d'acoustique passive, permettant d'identifier les espèces, sont à différencier.
La technique la plus simple est l'écoute directe, où l'observateur va déterminer, à l'oreille,
l'espèce et le nombre d'individus appartenant à l'espèce qu'il entend. C'est le cas des points
d'écoute du programme de Suivi Temporel de Oiseaux Commun (STOC) qui depuis 2001 a
permis de suivre les populations de 255 espèces d'oiseaux sur le territoire français. Une des
limites majeures de l'identification directe est un problème de répétabilité et de subjectivité
(Anderson et al., 1996).
Pour le cas où l'identification directe est plus difficile, il existe des protocoles d'identification
passant par l'utilisation d'outil d'analyse du signal comme ceux que nous avons détaillés en
partie 2.1. L'avantage de cette méthode est qu'elle est très généralement couplée avec
l'enregistrement du signal. Cela permet une répétabilité et une vérification de l'identification.
Le programme de suivi des Chiroptères du Muséum national d'Histoire naturelle se base sur
l'enregistrement des cris de chauve-souris qui permettent une identification spécifique
ultérieure par l'observateur, en utilisant le spectrogramme. Les différences entre les cris de
Chiroptères sont difficiles à percevoir en écoute directe car pour beaucoup d'espèces la
différence acoustique se caractérise par une faible variation des fréquences. L'arrivée des
enregistreurs automatiques a engendré d'énormes quantités de données acoustiques (voir
section 2.3.1). L'identification de tels jeux de données devient un travail extrêmement lourd
pour l'expert qui doit traiter un à un les sons en les écoutant et/ou les visualisant.
C'est pourquoi, une troisième technique a vu le jour, celle de l'identification automatique ou
ACR (Automatique Calls Recognition). Les méthodes pour l'identification automatique des
espèces sont encore en plein développement. La détection automatique se fait majoritairement
sur un taxon donné. On retrouve des études qui se sont intéressées à l'identification des
espèces d'oiseaux (Brandes, 2008 ; Anderson et al., 1996), de mammifères marins (Marques
et al., 2009 ; Brown et al., 2006, Mercado III et al., 1998) ou terrestres (Clemins et al., 2005,
Skowronski & Harris, 2006 ; Parsons & Jones, 2000), d'amphibiens (Han et al., 2010) et
d'insectes (Ganchev et al., 2007). Quelques études portent sur une détection automatique
multi-taxons, c'est le cas des études d’Acevedo et al. (2009) qui travaillent sur les oiseaux et
les amphibiens, de Brandes et al. (2006) sur les amphibiens et les grillons, de Chesmore
(2001) sur, à la fois les oiseaux, les amphibiens, les orthoptères et les mammifères, et de
Towsey et al. (2012) sur, à la fois les oiseaux, les mammifères terrestres et les amphibiens.

47
A partir d'un enregistrement acoustique fait sur le terrain, deux étapes sont nécessaires à
l'ACR : la détection des évènements acoustiques d'intérêt au sein de l'enregistrement et la
classification de ces évènements acoustiques. Pour chacune de ces deux étapes, il existe un
grand nombre de méthodes que nous ne pourrons pas toutes aborder ici.
Détecter. Erbe & King (2008) comparent trois méthodes de détection : (i) le détecteur
de pics d'énergie (peak energy detector) qui sélectionne l'évènement par rapport à sa valeur
d'énergie, valeur qui doit être supérieure à la valeur d'énergie associée au bruit de fond, (ii) le
détecteur d'énergie multi-bande (Mutliband energy detector) qui utilise le même principe que
le détecteur de pics mais suppose que les chants d'origine animale ont une énergie concentrée
sur plusieurs bandes de fréquence et (iii) le détecteur d'entropie (entropy detector) qui est une
mesure plus globale de l'énergie prise sur le spectre entier. La mesure d'entropie considère
donc l'ensemble des fréquences et se base sur le fait que beaucoup de bruits se retrouvent sur
une large bande de fréquence tandis que les sons d'origine animale ont un spectre étroit. En ce
qui concerne la détection de chants de mammifères marins, le détecteur d'entropie semble le
plus performant (Erbe & King, 2008).
Classer. En 2008, Brandes et al. publient un article reprenant l'ensemble de la
littérature concernant l'identification automatique des chants d'oiseaux. Au travers de cette
synthèse, les auteurs abordent les difficultés d'identification acoustique qui varient en fonction
de la diversité des sons au sein d'une espèce, à savoir la variabilité intra-spécifique des chants,
la complexité des arrangements des motifs, la largeur du répertoire, les dialectes et le
mimétisme acoustique (Briefer et al., 2011; Catchpole & Slater, 1995). Même si leurs chants
sont moins complexes que pour les oiseaux, les difficultés sont valables pour les chants des
autres groupes d'espèces chanteuses. Par exemple, les orques produisent des « clicks », des
« sifflements » et des « pulsations » (Brown et al., 2006). La variabilité intra-spécifique des
sons est une difficulté supplémentaire à l'identification automatique. Trois types de méthodes
d'identification acoustiques ACR sont utilisés. Certaines méthodes se basent sur la description
de paramètres du chant, comme la fréquence dominante, la largeur de fréquence utilisée, la
durée du chant et bien d'autres encore. C'est le cas des méthodes de classification Bayesiennes
(Brandes et al., 2006), des distances euclidiennes et des réseaux neuroniques (Mercado III et
al., 1998). Un deuxième type de méthode permet de classer des chants en fonction de leur
forme générale, c'est le cas des méthodes DTW (Dynamic Time Wraping ; Anderson et al.,
1996 ; Brown et al., 2006) et de la morphométrie géométrique (Lundy et al., 2011) qui se
basent sur le fait que la forme d'un son sur le spectrogramme semble être une information très
utile à l'identification. Enfin certaines méthodes tentent de modéliser le chant, c'est le cas des

48
HMM (Hidden Markov Models), et des méthodes GMM (Gaussian mixture models) (Table
2).
Table 2 : Comparaison des critères utilisés pour la détection d'unités sonores dans les chants d'oiseaux par
différentes méthodes de classification. Cette liste résume les méthodes référencées dans l'article de Brandes et
al. (2008). Modifiée à partir de Brandes et al. (2008).
Critère extrait Méthodes de classification
Mesures directes temporelles et fréquentielles Classification bayésienne, distance Euclidenne
Durée entre les impulsions acoustiques (pulse-to-
pulse duration)
Réseau de neurones
Forme du son Déformation temporelle dynamique (DTW pour
dynamic time warping), seuil minimum de corrélation
croisée
Estimation multi-spectral Statistique multi-variée
Contour des pics de fréquence Classification bayésienne, Déformation temporelle
dynamique (DTW pour dynamic time warping),
model Markov caché, Réseau de neurones
(cepstral coefficients) Déformation temporelle dynamique (DTW pour
dynamic time warping), model mixte Gaussien,
model Markov caché
Ce champ de recherche est en plein essor et d'autres méthodes de classification sont
encore développées (Bardeli et al., 2010 ; Connor et al., 2012)
Une seule méthode d'identification automatique ne peut pas être adaptée à
l'identification automatique de tous les chants d'origine animale à cause d'une trop grande
diversité. Towsey et al. (2012) montrent que le succès des méthodes d'identification
automatique dépend du type de son à reconnaitre et propose un ensemble de méthodes ou
« Tool box » adapté à l'identification de différents chants.
Toutes ces méthodes se confrontent à deux problèmes principaux en ce qui concerne
l'identification de signaux acoustiques enregistrés en milieu naturel : le problème du bruit de
fond et celui du chevauchement en temps et en fréquence des signaux. En effet, la présence de
bruit engendre une mauvaise détection du signal recherché et des problèmes d'identification.
Certains auteurs mettent en place une phase de dé-bruitage avant les étapes de détection et
classification (Bardeli et al., 2010). Selon le milieu, les types de bruits qui peuvent être
rencontrés diffèrent. En milieu aquatique marin par exemple, les bruits peuvent avoir pour
origine le vent, les vagues, les bateaux, le bruit de la glace, de bulles d'air, ou de la circulation

49
de l'eau (Erbe & King, 2008). En milieu terrestre, il s'agira de bruit de la pluie, du vent, et
beaucoup de bruit issus de l'activité humaine que sont les machines (voiture, avion) ou les
animaux domestiques. Pour réduire l'effet du bruit, il existe des méthodes de traitement du
signal. Le plus souvent, ce sont les filtres qui sont utilisés en fréquence ou en amplitude
(Bardeli et al., 2010). Il est également possible de caractériser le bruit de fond si celui-ci est
constant et de le soustraire au signal. Pour les problèmes de chevauchement, Bardeli et al.
(2010) recommandent de coupler les méthodes avec des détections de sources.
Une autre limitation de ces méthodes est de connaître a priori le signal acoustique de
l'espèce dans toute sa variabilité. Détecter et classer les chants nécessite des bases de données
de référence représentatives de la variabilité inter- et intra-espèces qui sont loin d'être faciles à
renseigner. Bien que la recherche sur l'identification automatique des espèces fasse de grandes
avancées, les difficultés techniques d'analyse du signal qui viennent d'être énoncées, ainsi que
l'absence de description d'un grand nombre de chants d'espèces, limitent fortement l'utilisation
de telles méthodes sur le terrain. Face au besoin d'analyser automatiquement de grandes
quantités d'enregistrements acoustiques, l'identification par un expert est un travail énorme et
n'est pas envisageable pour des suivis à long terme et l'identification automatique est encore
en cours de développement pour être utilisable en milieu naturel. Face aux difficultés des
inventaires classiques, des méthodes de « Rapid Biodiversity assessment » (RBA) ont été
développées (voir paragraphe 1.3). Il en est de même pour les méthodes acoustiques.
2.3.3 La méthode acoustique globale: Rapid Acoustic Survey
Une méthode globale d'analyse d'enregistrement acoustique a été proposée par Sueur
et al. (2008b), « the Rapid Acoustic Survey » (RAS). Cette méthode permet d'analyser le signal
acoustique émis par une communauté animale sans identification des espèces. C'est donc une
mesure globale d'une diversité acoustique locale. L'objectif est d'extraire directement les
informations du signal afin d'avoir une mesure de la diversité acoustique de la communauté
animale. Cette approche a été rapprochée des méthodes de RBA puisqu'elle ne cherche pas à
identifier précisément les espèces mais permet d'obtenir rapidement une information plus
large. Dans le cas de la RAS, l'échelle de mesure est celle de la communauté acoustique
caractérisant cette méthode en tant que « méta-méthode » (voir paragraphe 1.3 et Discussion
générale).
L'hypothèse de départ de cette méthode est la suivante : les espèces chantant au même
moment et au même endroit entrent potentiellement en compétition pour occuper l'espace
acoustique. Supposant cette compétition pour les niches écologiques acoustiques, plus une

50
communauté est composée d'espèces différentes, plus les chants issus de cette communauté
seront différents pour un temps donné. Ainsi, un signal hétérogène en fréquence et/ou en
temps devrait donc être reflété par une grande diversité d'espèces chanteuses (Obrist et al.,
2010 ; Sueur et al., 2008b).
Deux types d'indices acoustiques ont été proposés (Sueur et al., 2008b). Un indice de
diversité α, appelé indice d'entropie acoustique noté H, et un indice de diversité β, appelé
indice de dissimilarité acoustique noté D. L'indice H permet d'obtenir une diversité acoustique
basée sur un enregistrement acoustique. Cet indice reflète l'hétérogénéité spectrale et
temporelle de la partie active d'une communauté animale chanteuse. L'indice D permet
d'obtenir une différence acoustique temporelle et spectrale entre deux enregistrements et
reflète ainsi la différence entre les parties actives de deux communautés animales chanteuses.
Même si toutes les espèces chanteuses d'une communauté ne se manifestent pas en même
temps et donc ne sont pas présentes sur tous les enregistrements, l'obtention d'un très grand
nombre d'enregistrements permet d'avoir un échantillon représentatif de la majeure partie de
la communauté animale chanteuse. Ces indices ont été d'abord testés sur des communautés
simulées en laboratoire. Des chants de 15 espèces d'oiseaux, de 15 espèces d'amphibiens et de
15 espèces d'insectes ont été assemblés aléatoirement. L'intérêt de travailler sur des
communautés simulées est la capacité de contrôler leur composition en espèces et les
paramètres des chants (intensité, répétition, etc). Selon l'étude, l'indice H augmente en
moyenne avec le nombre d'espèces présentes dans la communauté simulée. L'indice D
diminue en moyenne avec le nombre d'espèces partagées par deux communautés simulées.
Les mêmes indices ont ensuite été testés sur deux forêts tropicales de Tanzanie distantes de 50
kilomètres et différentes en termes de pression anthropique (déforestation). Des
enregistrements au lever et coucher du soleil, qui sont des périodes de forte activité
acoustique, ont été comparés. Les analyses montrent une différence significative entre les
deux forêts avec une valeur de H plus faible pour la forêt subissant une forte pression
anthropique.
Cette méthode a été testée également en milieu tempéré par Depraetere et al. (2012) où
les auteurs proposent un nouvel indice de diversité α (Acoustic Richness, AR). Dans cette
étude, des communautés d'oiseaux ont été enregistrées dans trois types d'habitat différents. Un
expert a ensuite identifié les espèces à l'origine des chants présents dans les enregistrements
ce qui a permis de comparer ces résultats (nombre d'espèces d'oiseaux identifiées sur
l'enregistrement) avec les valeurs obtenues par le calcul des indices acoustiques. La même
tendance a été observée entre les résultats de la diversité α issue de l'identification à l'oreille et

51
les résultats de l'indice AR. De même, l'indice de diversité β (D) présente la même tendance
que les résultats obtenus par l'identification à l'oreille. Il semble donc y avoir congruence
entre la richesse spécifique des espèces chanteuses et les indices acoustiques présentés dans
cette étude.
Une équipe de recherche italienne a développé en même temps un indice acoustique
appelé ACI (Acoustic Complexity Index) basé sur la même idée d'un indice globale de
diversité acoustique (Pieretti et al., 2011 ; Farina et al., 2011). Les auteurs démontrent la
capacité de cet indice à représenter l'abondance des vocalisations d'oiseaux dans un
enregistrement. Le lien n'est donc pas directement fait avec le nombre d'espèces à l'origine de
ces chants. C'est un indice permettant d'identifier des variations d'activité acoustique.
Les indices présentés ici seront détaillés, comparés et critiqués dans le chapitre 2 de la
thèse. La méthode acoustique globale (RAS) a été présentée dans un chapitre d'ouvrage
intitulé « Sensors in Ecology » (Sueur et al., 2012). Ce chapitre est disponible en annexe 1.
Pour conclure, les scientifiques s'accordent à dire que l'acoustique passive est une méthode
qui permet l'étude et le suivi de la biodiversité (Obrist et al., 2010 ; Sueur et al., 2008b) et
présente un certain nombre d'avantages : elle est non invasive et permet l'exploration de zones
difficiles d'accès. De plus, l'arrivée d'enregistreurs automatiques permet des études à larges
échelles spatiales et temporelles ce qui induit une grande quantité de données.
Augmenter l'échantillonnage c'est augmenter la détectabilité des espèces qui est un
biais important des protocoles de comptages (voir Meyer et al., 2011). Cependant, analyser
ces grands jeux de données acoustiques à l'oreille est un travail considérable et très coûteux en
temps. Les méthodes d'identification automatique sont prometteuses mais encore loin d'être
applicables sur le son d'une communauté acoustique complexe.
La méthode RAS n'apporte pas de solutions à ces difficultés. Elle prend un chemin
différent, contourne les difficultés et apporte des informations complémentaires aux méthodes
d'échantillonnage acoustique classiques. Les avantages de cette méthode sont liés à son
automatisation, de l'échantillonnage au calcul des mesures acoustiques. De plus, son
utilisation ne nécessite pas de connaissances scientifiques particulières ce qui la rend
accessible. Elle pourrait permettre de suivre l'évolution des communautés à moindre coût.
Avant de pouvoir utiliser les potentialités de cette méthode, des études doivent être menées.
Nous tenterons de répondre à certaines de ces questions.

52
3-Objectifs et présentation des chapitres
Estimer la biodiversité est actuellement un enjeu majeur. Si un certain nombre de mesures ont
été prises pour essayer d'enrayer la perte de biodiversité actuelle, elles n'ont pas abouti à la
hauteur espérée et le problème de l'évaluation et du suivi de la biodiversité a été mis en avant.
En parallèle des méthodes classiques d'inventaires, qui demandent un haut niveau d'expertise
et du temps, des méthodes dites de mesure rapide de la biodiversité ont été proposées, dont la
méthode acoustique globale fait partie.
L'objectif général de cette thèse est de mieux appréhender cette approche d’acoustique globale
d'un point de vue méthodologique afin de connaître les possibles applications de cette
méthode pour estimer, suivre et mieux comprendre la distribution spatiale et temporelle de la
biodiversité.
Des indices acoustiques mesurant globalement l'hétérogénéité d'un signal sonore d'une
communauté animale ont déjà été proposés. L'objectif de ces indices est de rendre compte de
la diversité acoustique (de la diversité des sons animaux) dans un signal sonore. Cependant,
l'hétérogénéité d'un signal sonore peut avoir d'autres causes que la diversité des sons. Le
premier chapitre présentera quelques indices acoustiques. La sensibilité de ces indices à
différentes causes d'hétérogénéité acoustique sans lien avec la diversité acoustique sera
évaluée dans le Chapitre 1.
Nous avons vu dans la première partie de l'introduction que le terme biodiversité était
large et se définissait au travers d'un certain nombre de facettes complémentaires et dont les
valeurs ne sont pas toujours corrélées entre elles. Quelle facette de la diversité biologique, la
diversité acoustique permet-elle de refléter ? Pour répondre à cette question, les relations entre
la diversité acoustique et les diversités phylogénétique et fonctionnelle de communautés
d'oiseaux ont été analysées. Les résultats de cette étude sont présentés sous forme d'un article
dans le Chapitre 2.
Le Chapitre 3 évaluera la capacité d'un indice de dissimilarité acoustique à révéler
des différences de compositions de communautés. La Nouvelle-Calédonie présente un patron
particulier de micro-endémisme. L’indice de dissimilarité a pu être testé sur trois sites dont la
richesse spécifique est équivalente mais la composition spécifique est très différente du fait de
multiples évènements de spéciations allopatriques locales dues à l’isolement géographique.

53
Enfin le Chapitre 4 est une application de l'utilisation de ces indices pour tester
l'impact de différentes méthodes de gestion des agro-forêts. Trois types d'habitat ont été
échantillonnés selon le gradient suivant : forêt non perturbée, agro-forêt contenant des arbres
natifs, agro-forêt contenant des arbres non natifs. Les résultats acoustiques des différents
habitats, obtenus avec la méthode globale ont été comparés.
En Chapitre 5, une discussion générale présentera une réflexion autour du concept des
« méta-méthodes ». Un certain nombre de méthodes d'estimation de la biodiversité n'ont pas
pour objectif d'obtenir des identifications jusqu'à l'espèce. Deux grands types de méthodes
seront présentés. 1- Les méthodes prenant directement les mesures sur une échelle supérieure
à celle de l'individu, c'est le cas de la méta-génomique et de la méthode acoustique globale. 2-
Les méthodes dont la prise de mesure se fait à l'échelle de l'individu mais qui procèdent à une
identification plus globale et rapide que l'identification à l'espèce. Cette réflexion sera
accompagnée d’une discussion des différents résultats obtenus pendant ce travail de thèse.

54

55
Chapitre 1 Les indices de l’hétérogénéité
acoustique

56

57
I.1 Résumé La diversité acoustique est une composante de la biodiversité animale. L'idée d'évaluer cette
diversité par l'hétérogénéité acoustique d'un signal sonore a été proposée par différents
chercheurs. Plusieurs indices de mesure de l'hétérogénéité acoustique existent. Si la diversité
des sons joue un rôle dans l'hétérogénéité d'un enregistrement acoustique, d'autres causes sont
possibles. L'objectif de ce chapitre est de tester la sensibilité de neuf indices à l'hétérogénéité
acoustique générée par quatre causes différentes qui ne sont pas liées à la diversité des sons.
Ces quatre causes sont : la variation d'intensité relative des sons, le rapport entre la durée du
signal d'intérêt et la durée de l'enregistrement, le décalage temporel des sons, et le bruit de
fond. A partir de quatre-vingt-dix descriptions de communautés réelles d'oiseaux et du chant
des espèces disponibles dans la sonothèque du Muséum national d'Histoire naturelle, des
signaux acoustiques de communautés animales ont pu être simulés. Deux types d'indices ont
été calculés. Les indices de diversité α qui mesurent une valeur d'hétérogénéité pour chaque
enregistrement et les indices de diversité β qui mesurent des différences entre deux
enregistrements. Les résultats montrent que chaque indice est sensible à au moins une des
quatre causes d'hétérogénéité. Cependant ils ne possèdent pas les mêmes sensibilités. Les
indices de dissimilarité par exemple sont tous sensibles au bruit de fond contrairement à deux
des indices α. Selon l'étude envisagée, l'utilisation des indices, qu'ils soient basés sur des
caractéristiques spectrales et/ou temporelles du son, est à considérer selon leur sensibilité aux
différentes causes possibles d'hétérogénéité acoustique.

58
I.2 Introduction
La définition d'une communauté acoustique animale, proposée ici, est une application au
domaine acoustique de la définition de la communauté proposée par Diamond et Case (1986).
Selon cette définition, la communauté est une entité qui comprend les populations des espèces
coexistantes sur un même site ou dans une même région. Ainsi, la communauté acoustique
animale est définie comme une entité qui comprend les populations des espèces chanteuses
coexistantes sur un même site ou dans une même région. Une nette distinction doit être faite
entre la diversité acoustique d'une communauté (CAD pour Community Acoustique
Diversity) et la diversité d'une communauté acoustique. Dans le premier cas, il s'agit bien de
la diversité mesurée à partir de la collection de sons. Dans le second, cas, la définition est plus
large et rassemble à la fois la diversité acoustique de la communauté, mais également toutes
les autres diversités biologiques pouvant être associées à cette sous-partie de la communauté.
Une mesure de la diversité acoustique est faite à partir d'une collection de sons, comme la
diversité spécifique est une mesure faire à partir d'une collection d'espèces et la diversité
fonctionnelle à partir d'une collection de traits fonctionnels. Ainsi, les diversités spécifique,
fonctionnelle, phylogénétique et acoustique peuvent être calculées en tant que diversité de la
communauté acoustique (Fig. I.1).
Fig. I.1 : Schéma théorique représentant la diversité acoustique de la communauté
comme composante de la diversité de la communauté acoustique.

59
La méthode RAS, définie en introduction, propose de mesurer la diversité acoustique par des
indices estimant l’hétérogénéité globale d’un enregistrement acoustique d’une communauté
animale. L’objectif de ces indices est donc de refléter une ou plusieurs facettes de la diversité
acoustique d’une communauté, généralement, le nombre total de sons (quantitatif) ou le
nombre de types de sons différents (qualitatif).
Plusieurs paramètres acoustiques sont susceptibles de biaiser les valeurs des indices de
diversité acoustique. Nous considérerons quatre d’entre eux : l’intensité relative des espèces,
l'alignement temporel du démarrage des signaux, la durée de l’enregistrement par rapport à la
celle du signal d'intérêt et, enfin, le bruit ambiant. Nous tenterons donc de répondre aux quatre
questions suivantes :
Question 1 : L’intensité plus ou moins forte d'un son résulte soit des caractéristiques
énergétiques intrinsèques de l'espèce, soit de l'éloignement de la source par rapport au
microphone. Cette perte d’intensité avec la distance peut être accentuée ou diminuée selon les
caractéristiques du milieu. Pour une collection donnée de sons, ceux-ci peuvent donc être
d’intensités relatives différentes. Il est donc légitime de se demander comment cette variation
influe sur les indices acoustiques. Précisément nous tenterons de répondre à la question
suivante : Quels est l’impact des intensités relatives des sons sur les indices acoustiques ?
Question 2 : Le principe des indices acoustiques, présentés ici, est de prendre une
mesure sur l'ensemble d'un enregistrement sonore. Quelle est l'importance relative d'un signal
dans un enregistrement bref ou long ? Autrement dit, quel est l’impact de la durée d'un signal
par rapport au temps d'enregistrement sur les indices acoustiques ?
Question 3 : Même en supposant que la communauté animale acoustique entière est
détectée et enregistrée, les individus ne chantent pas tous en même temps. L'objectif de ce test
est de savoir si l'organisation temporelle des signaux au sein d'un enregistrement influe sur la
valeur des indices. Autrement dit, quel est l’impact du décalage temporel entre les signaux
d’une communauté sur les indices acoustiques ?
Question 4 : Un des principaux problèmes des méthodes d'acoustique passive est très
certainement le bruit qui vient brouiller le signal acoustique d’intérêt. Nous distinguerons
deux types de bruit : les bruit filtrables (tels que la plupart des bruits issus de l'activité
anthropique que sont, par exemple, les bruits d'avions ou de voitures) et les bruits intraitables
en analyse du signal tel que le bruit du vent et de la pluie. Ces derniers rendent, à la lumière
des techniques acoustiques actuelles, l'enregistrement inutilisable. Le bruit ambiant occupe
toutes les fréquences du spectre mais est en général d’intensité plus faible que le signal

60
d’intérêt. Si des techniques de filtres existent, elles sont à prendre avec précautions. Quel est
l’impact du bruit ambiant sur les indices acoustiques ?
Ce chapitre sera organisé en deux parties. La première partie présentera mathématiquement
les indices de diversité acoustique. La deuxième partie sera consacrée à la présentation des
tests permettant de répondre aux quatre questions posées.
I.3 Matériel et méthodes
I.3.1 Simulations
Afin de répondre à ces questions, nous utiliserons des simulations de signaux acoustiques de
communautés d’oiseaux sur la base de compositions réelles. Nous utiliserons le terme
« signal » pour désigner le signal composé de plusieurs chants d'une communauté d'oiseaux.
L’utilisation de simulations nous permet de maîtriser les variations des caractéristiques
étudiées. Les sons utilisés ont été réduits à ceux des oiseaux. En effet, leur comportement est
très bien connu et documenté grâce à la sonothèque du Muséum national d’Histoire naturelle
qui contient des enregistrements de bonne qualité pour des centaines d’espèces françaises. De
plus, grâce aux travaux présentés dans l’article de Depraetere et al. (2012), nous disposons
d’une collection d’enregistrements réels de signaux de communautés d’oiseaux faite en Haute
Vallée de Chevreuse du 24 Mars au 5 Juin 2010. Parmi cette collection d’enregistrements, 90
fichiers ont été sélectionnés pour avoir un bruit ambiant négligeable. Ces enregistrements ont
été écoutés et les signaux ont été identifiés par Frédéric Jiguet, responsable du programme de
Suivie Temporel de Oiseaux Commun (STOC) sur le territoire Français. Au total, vingt-cinq
espèces ont été identifiées. Les signaux de communauté d'oiseaux sont composés au minimum
de deux espèces et au maximum huit espèces. Une « séquence sonore » (ou « son » ou
« chant ») est ici définie par un enregistrement de 30 secondes de l’activité acoustique d’un
individu (fréquence d'échantillonnage = 44100, monophonie, nombre de bits= 16). Pour
chaque espèce, un chant typique a été sélectionné à partir de la sonothèque du Muséum
national d’Histoire naturelle (Paris). Nous pouvons recréer artificiellement le signal
acoustique d’une communauté par la simple superposition de ces séquences. La méthodologie
choisie pour simuler ces signaux acoustiques utilise une seule séquence de trente secondes de
vocalisation par espèce. Cela implique que la variabilité intra-spécifique n'est pas prise en
compte. De plus, si la diversité peut être appréhendée de deux façons différentes via le

61
nombre de types de sons différents ou le nombre total de sons, la méthodologie employée
dans ce chapitre ne nous permettra pas de différencier ces deux aspects. L'impact de
l'abondance des sons dans le signal acoustique ne sera pas étudié ici.
I.3.1.1 Question 1: Quel est l'impact des intensités relatives des séquences sonores sur les
indices acoustiques ?
Chaque espèce décrite est représentée par une seule séquence sonore. Pour chacune des
communautés, nous avons fait varier l’intensité relative de chaque chant. Un coefficient
d’intensité relative tiré aléatoirement parmi des valeurs allant de 0 à 1 avec un pas de 0,1 a été
attribué à chaque séquence sonore préalablement mise à l’échelle entre +1 et -1. Pour chacune
des 90 communautés, 99 simulations ont été réalisées en faisant varier à chaque fois les
coefficients d'intensité, générant un total de 8910 simulations par indice.
Pour chaque communauté, nous avons calculé les indices acoustiques α. Les indices
ont été mis à l’échelle en les divisant par leur maximum observé. Pour chaque communauté et
chaque indice, nous avons calculé l'écart-type de ces valeurs maximisées obtenues pour les 99
simulations. Une plus forte valeur de l'écart-type a été attribuée à une plus forte sensibilité de
l'indice aux variations d'intensités relatives des signaux.
Les dissimilarités fréquentielles ont été calculées entre chaque paire des 99 répétitions
de communautés simulées. Ainsi, pour chaque indice et chaque communauté, une matrice de
taille 99x99 a été obtenue. De la même façon que pour les indices de diversité α, la valeur
d'écart-type ainsi que la variation de cette valeur en fonction du nombre de chants présents
dans le signal de la communauté ont été estimées.
I.3.1.2 Question 2: Quel est l’impact de la durée d'un signal par rapport au temps
d'enregistrement sur les indices acoustiques ?
Chaque séquence sonore de 30 secondes a été prise individuellement et a été allongée par un
son correspondant à du bruit ambiant de 30, 60, 90 puis 120 secondes générant 100 signaux.
Ce bruit ambiant, issu de Depraetere et al. (2012), a été enregistré dans la forêt tempérée de
Rambouillet. Les indices de diversité acoustique α seront calculés sur ces signaux. Les indices
β de dissimilarité spectrale ont été mesurés sur chaque paire de signaux de même durée
générant quatre matrices symétriques de dissimilarité de taille 25x25 par indice. Nous
regarderons l'évolution des indices de diversité α avec la taille de l'enregistrement
indépendamment pour les 25 séquences sonores. De même, les matrices étant symétriques,
nous regarderons l'évolution des 300 valeurs ((25x25-25)/2) de dissimilarité par matrice.

62
I.3.1.3 Question 3 : Quel est l’impact du décalage temporel entre les signaux sur les indices
acoustiques ?
Pour chacune des 90 communautés, trois décalages temporels entre les séquences sonores ont
été appliqués : i) aucun décalage (synchrone), la superposition des séquences est complète, ii)
un décalage de la moitié du son, le début de la séquence suivant étant décalé de 15 secondes
par rapport à la séquence précédente, les superposant de moitié (décalé), iii) un décalage
maximum, aucune des séquences ne se superposent (alterné). Nous comparerons les valeurs
de chaque indice en fonction de ce degré de recouvrement temporel.
I.3.1.4 Question 4 : Quel est l’impact du bruit ambiant sur les indices acoustiques ?
Un bruit ambiant a été ajouté à chacune des simulations acoustiques de 90 communautés.
Pour chaque simulation, trois niveaux de bruit ont été considérés : bruit faible pour lequel
aucune modification du son n'a été faite, bruit moyen pour lequel a été appliqué un coefficient
d'intensité 50 fois plus élevé que celui du bruit ambiant initial, et bruit fort pour lequel a été
appliqué un coefficient d'intensité 100 fois plus élevé que celui du bruit de ambiant initial.
Nous comparerons l’évolution de chaque indice en fonction de l'augmentation de l'intensité du
bruit ambiant relative à celle du signal acoustique d'une communauté.
I.3.2 Les indices de diversité acoustique
Deux types d'indices de diversité acoustique ont été développés, les indices de diversité α et
les indices de diversités β.
Les indices de diversité α sont calculés par simulation. Ces mesures considèrent la
complexité du signal en se basant sur l'hypothèse que plus un signal est complexe, plus la
diversité des sons qui le composent est forte. Ces indices traitent la composante fréquentielle
ou/et temporelle du signal sonore. Trois indices de diversité acoustique α ont été proposés
(Table I.1): H (Sueur et al., 2008b), ACI (Pieretti et al., 2011) et AR (Depraetere et al., 2012).
Pour chacun d'eux, les auteurs ont tenté de relier la valeur de leurs indices à une diversité de
la communauté acoustique, soit le nombre de vocalisations (Sueur et al., 2008b; Pieretti et al.,
2011) et ont, pour certains, essayé d'établir un lien entre diversité acoustique et une autre
facette de la diversité de la communauté acoustique en regardant la corrélation entre indice
acoustique et nombre d’espèces chanteuses (Depraetere et al., 2012). Nous avons développé
un nouvel indice, NP, comme mesure de la complexité spectrale.

63
Les indices de diversité β mesurent la dissimilarité entre deux signaux acoustiques
différents (Table I.1). Un seul indice, l'indice de dissimilarité acoustique D, a été proposé dans
l'article de Sueur et al. (2008b). Cependant de nombreux autres indices de dissimilarité de
distributions existent. Les indices de Kullback-Leibler (KL) (Kullback et al., 1951) et de
Kolmogorov-Smirnov (KS) (Rachev et Svetlozar T., 1991) ont été ajoutés à cette analyse.
Certains indices sont calculés à partir du sonogramme ou du spectre moyen faisant appel à
une transformée de Fourier à court terme (STFT, Short-time Fourier Transform, taille de
fenêtre = 512 points, précision temporelle = 0.012 s, précision fréquentielle= 86 Hz, pas de
superpositions des fenêtres).
Table I.1 : Résumé des indices acoustiques.
Indice Référence Temporel/spectral
Diversités alpha
H Sueur et al. (2008b) Temporel et spectral ACI Farina et Morri (2008); Pieretti
et al. (2011) Temporel et spectral
AR Depraetere et al. (2012) Temporel NP Chapitre 3 et 4 Spectral
Diversités béta
D Sueur et al. (2008b) Temporel et spectral KL Kullback et al. (1951) Spectral KS Rachev et Svetlozar T. (1991) Spectral
I.3.2.1 L'indice d'entropie acoustique H
H est le produit de deux indices, l'entropie temporelle Ht et l'entropie spectrale Hf (Fig. I.2) :
H H Ht f= × avec 0 ≤ H ≤ 1
Hf est calculé à partir de l'enveloppe du spectre moyen. Il analyse la complexité du signal sur
sa composante fréquentielle. A chaque point de l'enveloppe est associée une probabilité liée à
l'intensité du point par rapport aux autres points (Fig. I.2) :
1( ) log ( ) log ( )2 21
NH S f S f Nf
f−= − × ×∑
= avec 0 ≤ Hf ≤ 1
Ht est calculé de la même façon mais sur la composante temporelle du signal.
1( ) log ( ) log ( )2 21
nH A t A t nt
t−= − × ×∑
= avec 0 ≤ Ht ≤ 1

64
où, N est le nombre de points composant le spectre moyen, S(f) est la fonction de masse
(probabilités) des valeurs d'intensités du spectre moyen du signal, n est le nombre de points
composant l'enveloppe, A(t) est la fonction de masse (probabilités) des valeurs d'intensités des
points de l'enveloppe.
Fig. I.2 : Schéma reprenant les étapes permettant le calcul des indices Hf et Ht.
I.3.2.2 L'indice de richesse acoustique (Acoustic Richness) AR
L'indice AR ne prend pas en compte Hf qui semble être très sensible au bruit ambiant mais
intègre la médiane de l'enveloppe d'intensité des signaux, M (Fig. I.3) avec :
(1 )( ( )) 2 depthM median A t −= × avec 0 ≤ AR ≤ 1
où A(t) est ici différent de celui de la fonction de Ht puisqu'il s'agit ici de la fonction décrivant
les valeurs d'intensité des points de l'enveloppe (valeurs bruts, sans transformation en
probabilités) et depth est la valeur de digitalisation du signal (8 or 16 bits)
[ ]( ) ( )2
rank H rank MtARn
×= avec 0 ≤ AR ≤ 1
où n est le nombre de fichiers analysés.
Cet indice est donc calculé sur les rangs des valeurs. Cette caractéristique le rend relatif au jeu
de données utilisé, les valeurs de cet indice ne sont donc pas comparable d'une étude à l'autre.

65
Fig. I.3 : Schéma reprenant les étapes permettant le calcul des indices Ht et M.
I.3.2.3 L'indice de complexité acoustique (Acoustic Complexity Index) ACI
Cet indice, développé par Farina et Morri (2008) mesure la variabilité d'une matrice de
valeurs correspondantes à la matrice d'un sonogramme pour laquelle, pour une fréquence f et
un temps donné t, correspond une valeur d'intensité Ik (Fig. I.4). L'indice ACI augmente avec
la variabilité temporelle des sons, se basant sur l'hypothèse que les variations d’intensités sont
associables à une activité acoustique animale contrairement à un son d'origine anthropique
dont l’intensité varie peu dans le temps.
Le spectrogramme est découpé une première fois en plages temporelles de 5 secondes. Pour
une plage temporelle j, le sonogramme est découpé une deuxième fois en n plages temporelles
(Δtk) et en q plages fréquentielles (Δfl).
Soit une seule plage de fréquence Δfl, dk est la différence absolue entre deux valeurs
d'intensités adjacentes Ik et I(k+1) :
1d I Ik k k= − +

66
L'indice D est la somme de ces différences :
1
nD dk
k=
=∑
où n est le nombre de Δtk dans la plage temporelle j.
Cette valeur D est ensuite divisée par la somme totale des intensités de j dans le but de réduire
la différence des intensités entre vocalisations qui pourrait être due à un éloignement de la
source par rapport au microphone :
1
DACI j nIk
k
=
=∑
L'indice appelé ACI ici est donc calculé sur une seule plage de fréquence et sur une plage
temporelle j (issue de la première découpe temporelle). La somme des ACI sur toutes les
plages temporelles issues de la première découpe est calculée :
( )1
mACI ACIft j
j=∆
=∑
m est le nombre de plages temporelles issues de la première découpe.
Enfin, la somme de ce nouvel indice sur toutes les plages fréquentielles Δfl est calculée:
( )1
qACI ACItot fl
l= ∆
=∑
où q est le nombre de plages temporelles Δfl. Ainsi, l'indice ACItot est la valeur de l'indice sur
l'enregistrement entier.
Dans notre étude, la valeur de j sera fixée à 5 secondes. L'indice ACItot sera nommé
simplement ACI.

67
Fig. I.4 : Schéma reprenant les étapes permettant du calcul de l'indice ACI.
I.3.2.4 Nombre de pics fréquentiels NP
Cet indice a été développé ici dans le but de pallier le problème de sensibilité de bruit ambiant
de l'indice Hf, problème énoncé dans l'article de Depraetere et al. (2012). Le spectre moyen du
bruit ambiant comme défini dans l'introduction, ne présente pas de pics majeur. La détection
de pic est donc un moyen de ne prendre en compte que les valeurs du spectre moyen issu
d'une activité animale. Cet indice se calcule en deux étapes. La première étape est une étape
de détection des pics. Un point du spectre moyen est déterminé comme pic si et seulement si
les points précédents et suivants ont une valeur plus faible. La seconde étape est une étape de
sélection des pics. En effet, des pics très proches peuvent être considérés comme un seul pic.
Le critère de séparation des pics selon un seuil fréquentiel est dans ce cas utilisé. Dans le
cadre de cette étude, une valeur de séparation de 200 Hz minimum est requise pour considérer
deux pics comme différents. Des pics de pentes trop faibles peuvent également se trouver
exclus de la sélection. Dans ce cas, ce sont les critères de pentes précédant, Pei et suivant le
pic, Pei+1, qui seront considérés (Fig. I.5). Les seuils de pentes ont été fixés à 0,01.
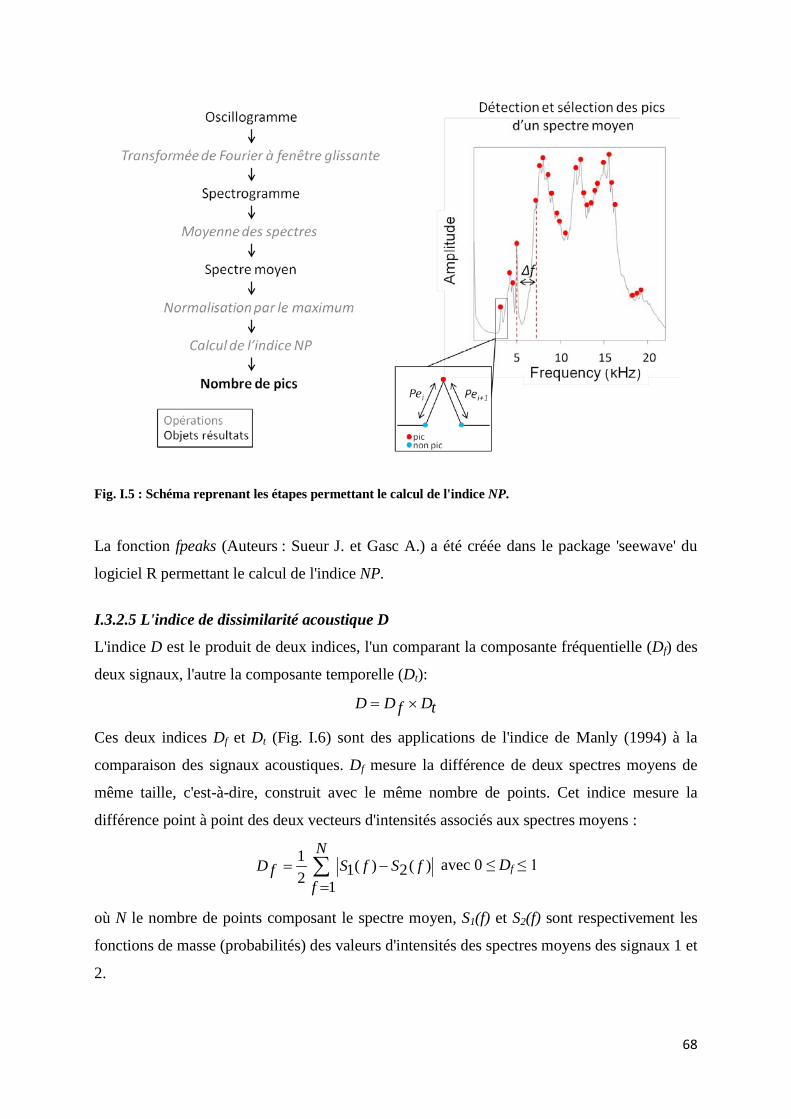
68
Fig. I.5 : Schéma reprenant les étapes permettant le calcul de l'indice NP.
La fonction fpeaks (Auteurs : Sueur J. et Gasc A.) a été créée dans le package 'seewave' du
logiciel R permettant le calcul de l'indice NP.
I.3.2.5 L'indice de dissimilarité acoustique D
L'indice D est le produit de deux indices, l'un comparant la composante fréquentielle (Df) des
deux signaux, l'autre la composante temporelle (Dt):
D D Df t= ×
Ces deux indices Df et Dt (Fig. I.6) sont des applications de l'indice de Manly (1994) à la
comparaison des signaux acoustiques. Df mesure la différence de deux spectres moyens de
même taille, c'est-à-dire, construit avec le même nombre de points. Cet indice mesure la
différence point à point des deux vecteurs d'intensités associés aux spectres moyens :
1 ( ) ( )1 22 1
ND S f S ff
f= −
=∑ avec 0 ≤ Df ≤ 1
où N le nombre de points composant le spectre moyen, S1(f) et S2(f) sont respectivement les
fonctions de masse (probabilités) des valeurs d'intensités des spectres moyens des signaux 1 et
2.

69
Il en est de même pour la composante temporelle. Dt mesure de la même façon la différence
entre les enveloppes temporelles de deux signaux. Étant une comparaison point à point, les
deux signaux doivent avoir la même longueur.
1 ( ) ( )1 22 1
nD A t A tt
f= −
=∑ avec 0 ≤ Dt ≤ 1
où n le nombre de points composant l'enveloppe temporelle des signaux, A1(t) et A2(t) sont
respectivement les fonctions de masse (probabilités) des valeurs d'intensités des enveloppes
temporelles des signaux 1 et 2.
Fig. I.6 : Schéma reprenant les étapes permettant le calcul des indices Df (à gauche) et Dt (à droite).
I.3.2.6 L'indice de distance symétrique de Kullback-Leibler KL et l'indice de Kolmogorov-
Smirnov KS
Ces deux indices ne diffèrent de l'indice Df que dans l'opération faite entre les points. Df est
une différence absolue alors que KL est un ratio et KS une différence absolue d'une somme
cumulée.
L'indice KL se calcule selon la formule suivante :
( ( ) || ( )) ( ( ) || ( ))1 2 2 12
kl S f S f kl S f S fKL +=
sachant que:
( )1( ( ) || ( )) ( ( ) log( ))1 2 1 ( )2
S fkl S f S f sum S fS f
= ×
où S1(f) et S2(f), respectivement les fonctions de masse (probabilités) des valeurs d'intensités
des spectres moyens des signaux 1 et 2.

70
L'indice KS se calcule selon la formule suivante :
( ( )) ( ( ))1 2max( )max( ( ( ))) max( ( ( )))1 2
cumsum S f cumsum S fKScumsum S f cumsum S f
= − avec 0 ≤ KS ≤ 1
avec cumsum la somme cumulée des valeurs et S1(f) et S2(f), respectivement, les fonctions de
masse (probabilités) des valeurs d'intensités des spectres moyens des signaux 1 et 2 (Fig. I.7).
Fig. I.7 : Schéma représentant les étapes permettant le calcul des indices KL et KS.
La durée d'enregistrement a un impact très fort sur l'indice ACI pour des raisons
mathématiques, d'où l'importance de ne comparer que des enregistrements de même longueur
pour cet indice. Ainsi les résultats de l'indice ACI n’ont pas pu être interprétés pour les
questions 2 et 3.
L'ensemble des analyses a été effectuée à l'aide du logiciel R (R Development Core Team,
2010) et des packages 'seewave' (Sueur et al., 2008a) et 'tuneR' (Ligges, 2011).

71
I.4 RESULTATS
I.4.1 Question 1: Quel est l'impact des intensités relatives des séquences sonores sur les
indices acoustiques ?
Pour chaque communauté, nous avons calculé les écart-types des 99 valeurs issues des
variations d’intensités relatives. Ainsi, 90 valeurs d’écart-types sont présentées pour chaque
indice en Fig. I.8. AR présente les écart-types les plus forts. Les indices NP et M présentent
des valeurs élevées et une variabilité forte qui indique que la composition de la communauté
pourrait influer sur la sensibilité de l’indice aux variations des intensités relatives des
séquences sonores. Les indices Hf, ACI et Ht présentent les écart-types les plus faibles. Les
indices Df et Ks ont des écart-types proches, tous deux supérieurs à celui de Kl.
Fig. I.8 :Boxplot des valeurs des écart-types mesurés pour chaque communauté et chaque indice.
Les écart-types de Ht et Hf diminuent avec le nombre de séquences et les écart-types de M
augmentent fortement avec le nombre de séquences (Fig. I.9). Les écart-types des indices NP,
AR et ACI ne présentent pas de tendance claire de leur sensibilité au nombre de séquences. La
sensibilité des indices de dissimilarité Df, Kl et Ks à la variation des intensités relatives des
séquences diminue avec le nombre de séquences sonores espèce-spécifiques composant le
signal de la communauté (Fig. I.9). Notons que la variance des valeurs d’écart-type diminue
avec l'augmentation du nombre de séquences dans le signal acoustique de la communauté.
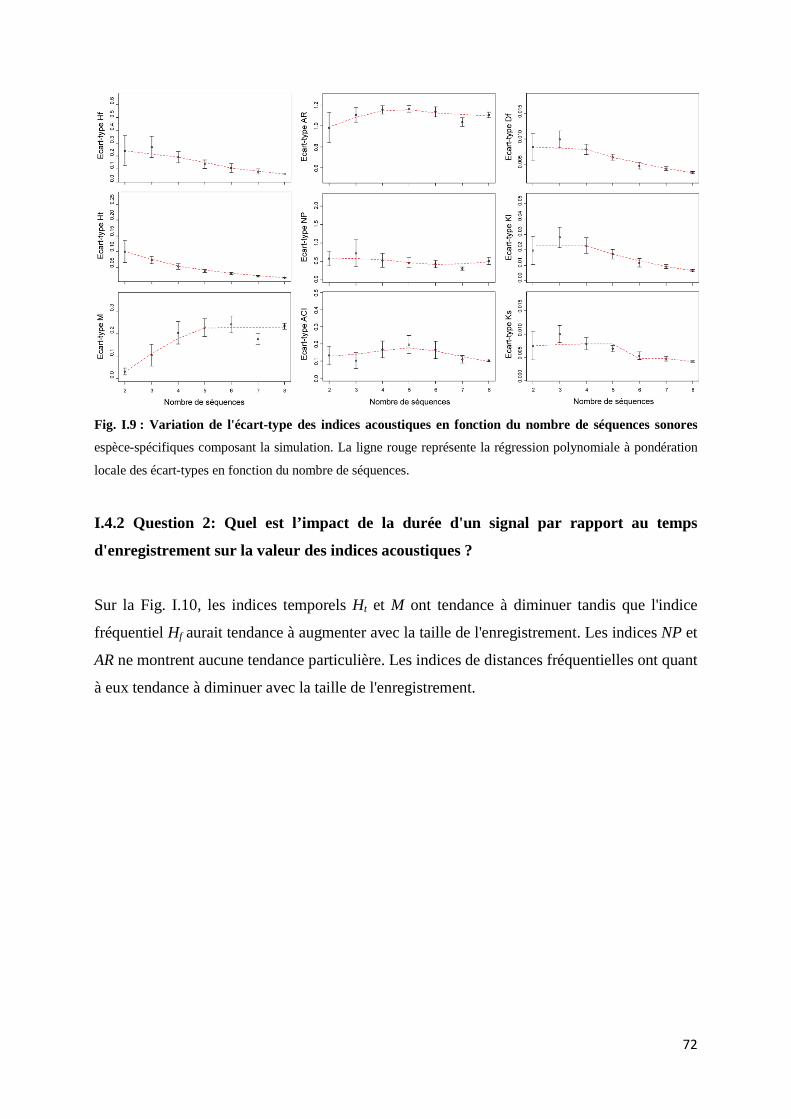
72
Fig. I.9 : Variation de l'écart-type des indices acoustiques en fonction du nombre de séquences sonores
espèce-spécifiques composant la simulation. La ligne rouge représente la régression polynomiale à pondération
locale des écart-types en fonction du nombre de séquences.
I.4.2 Question 2: Quel est l’impact de la durée d'un signal par rapport au temps
d'enregistrement sur la valeur des indices acoustiques ?
Sur la Fig. I.10, les indices temporels Ht et M ont tendance à diminuer tandis que l'indice
fréquentiel Hf aurait tendance à augmenter avec la taille de l'enregistrement. Les indices NP et
AR ne montrent aucune tendance particulière. Les indices de distances fréquentielles ont quant
à eux tendance à diminuer avec la taille de l'enregistrement.
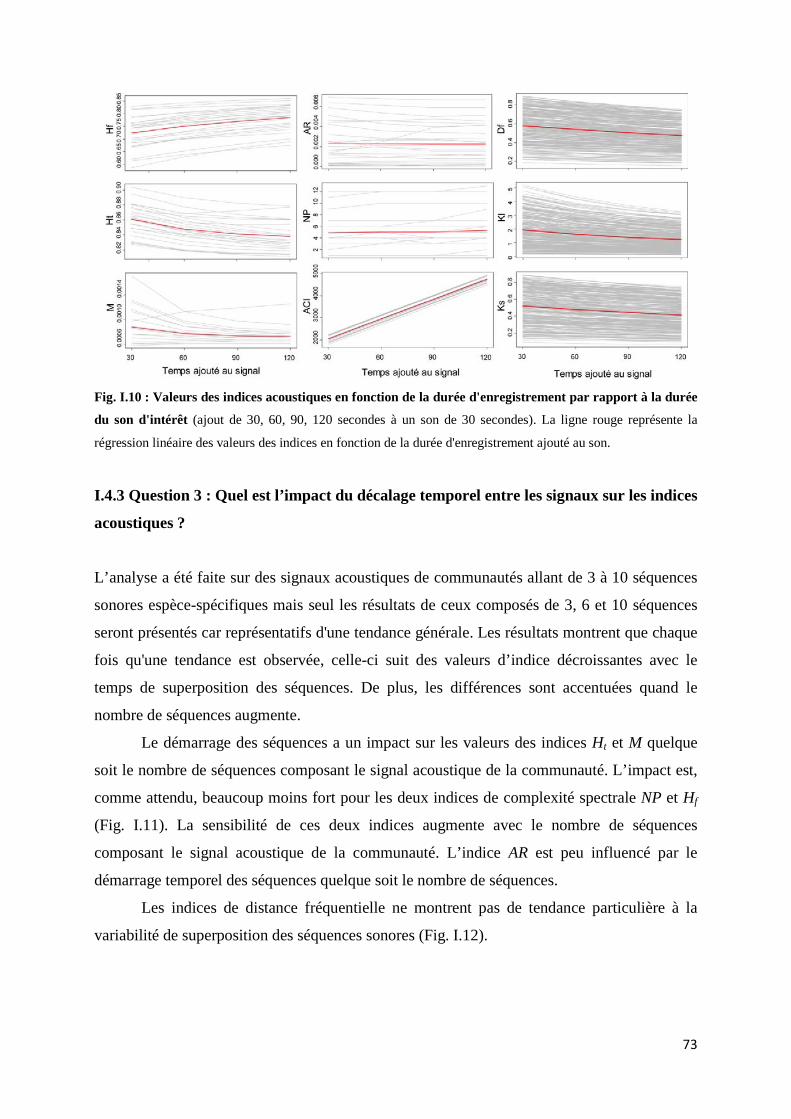
73
Fig. I.10 : Valeurs des indices acoustiques en fonction de la durée d'enregistrement par rapport à la durée
du son d'intérêt (ajout de 30, 60, 90, 120 secondes à un son de 30 secondes). La ligne rouge représente la
régression linéaire des valeurs des indices en fonction de la durée d'enregistrement ajouté au son.
I.4.3 Question 3 : Quel est l’impact du décalage temporel entre les signaux sur les indices
acoustiques ?
L’analyse a été faite sur des signaux acoustiques de communautés allant de 3 à 10 séquences
sonores espèce-spécifiques mais seul les résultats de ceux composés de 3, 6 et 10 séquences
seront présentés car représentatifs d'une tendance générale. Les résultats montrent que chaque
fois qu'une tendance est observée, celle-ci suit des valeurs d’indice décroissantes avec le
temps de superposition des séquences. De plus, les différences sont accentuées quand le
nombre de séquences augmente.
Le démarrage des séquences a un impact sur les valeurs des indices Ht et M quelque
soit le nombre de séquences composant le signal acoustique de la communauté. L’impact est,
comme attendu, beaucoup moins fort pour les deux indices de complexité spectrale NP et Hf
(Fig. I.11). La sensibilité de ces deux indices augmente avec le nombre de séquences
composant le signal acoustique de la communauté. L’indice AR est peu influencé par le
démarrage temporel des séquences quelque soit le nombre de séquences.
Les indices de distance fréquentielle ne montrent pas de tendance particulière à la
variabilité de superposition des séquences sonores (Fig. I.12).

74
Fig. I.11 : Valeurs des indices acoustiques en fonction du décalage des séquences dans le signal. De gauche
à droite sont présentés les résultats pour les signaux composés de 3, 6 et 10 séquences. Les résultats pour les
signaux composés de 4, 5, 7, 8 et 9 séquences sont intermédiaires, leurs tendances suivant les résultats présentés
ici. Les étiquettes Syn Dec et Alt indiquent respectivement les positions synchrones, décalées ou alternées des
séquences sonores.
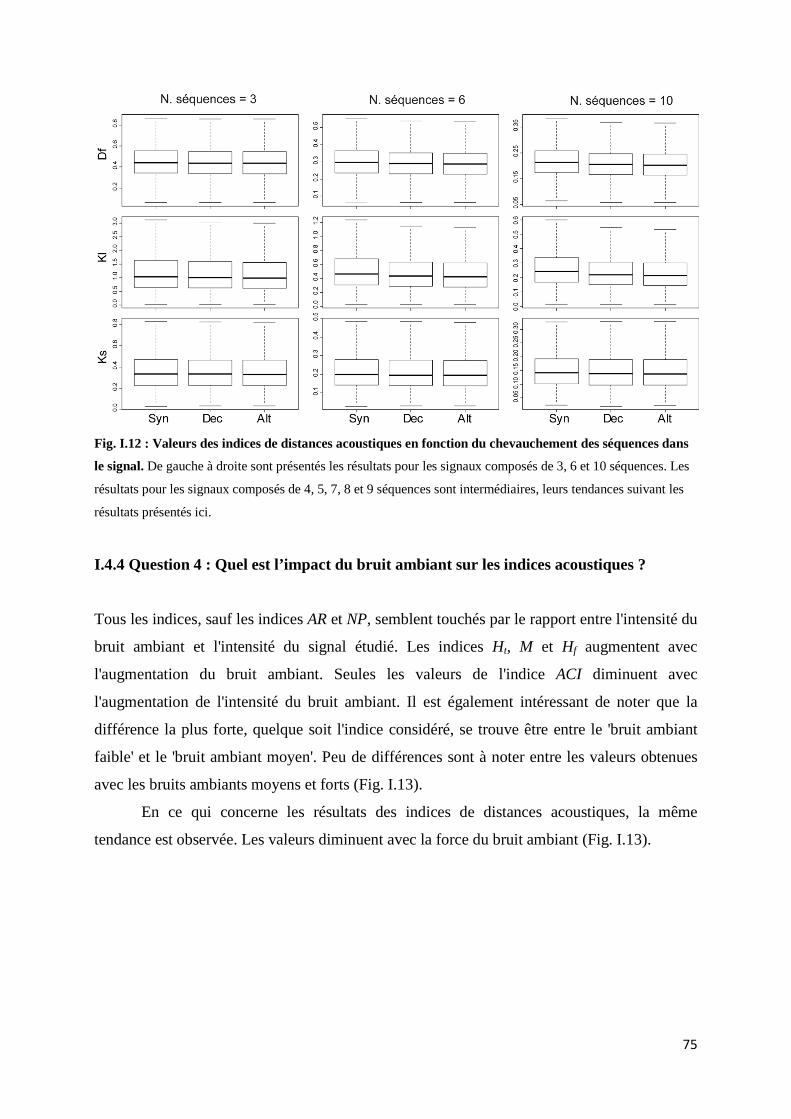
75
Fig. I.12 : Valeurs des indices de distances acoustiques en fonction du chevauchement des séquences dans
le signal. De gauche à droite sont présentés les résultats pour les signaux composés de 3, 6 et 10 séquences. Les
résultats pour les signaux composés de 4, 5, 7, 8 et 9 séquences sont intermédiaires, leurs tendances suivant les
résultats présentés ici.
I.4.4 Question 4 : Quel est l’impact du bruit ambiant sur les indices acoustiques ?
Tous les indices, sauf les indices AR et NP, semblent touchés par le rapport entre l'intensité du
bruit ambiant et l'intensité du signal étudié. Les indices Ht, M et Hf augmentent avec
l'augmentation du bruit ambiant. Seules les valeurs de l'indice ACI diminuent avec
l'augmentation de l'intensité du bruit ambiant. Il est également intéressant de noter que la
différence la plus forte, quelque soit l'indice considéré, se trouve être entre le 'bruit ambiant
faible' et le 'bruit ambiant moyen'. Peu de différences sont à noter entre les valeurs obtenues
avec les bruits ambiants moyens et forts (Fig. I.13).
En ce qui concerne les résultats des indices de distances acoustiques, la même
tendance est observée. Les valeurs diminuent avec la force du bruit ambiant (Fig. I.13).

76
Fig. I.13 : Valeurs des indices acoustiques sur des signaux soumis à un bruit ambiant faible, moyen ou fort
I.5 Discussion
Neuf indices mesurant l'hétérogénéité acoustique d'un signal ont été considérés. Au delà des
caractères acoustiques intrinsèques des signaux qui composent le signal, il existe d'autres
causes à l'origine de l'hétérogénéité acoustique que l’on peut mesurer à partir d’un
enregistrement acoustique d'une communauté. Dans ce chapitre quatre de ces causes ont été
considérées : la variation des intensités relatives des signaux, la variation de la durée de
l'enregistrement par rapport à la durée du signal d'intérêt, la variation temporelle de démarrage
des signaux et enfin l’intensité du bruit ambiant relative à celle du signal acoustique d'une
communauté. L'objectif de ce chapitre était de comparer le comportement des indices
acoustiques face à ces quatre causes d'hétérogénéité. Les résultats montrent que tous ces
indices sont sensibles à au moins une des quatre causes d’hétérogénéité considérées.
I.5.1 Variation d’intensité relative: une détectabilité déséquilibrée des sons
Les indices ont été calculés sur des objets maximisés. Cela signifie que les valeurs de
l'enveloppe temporelle ou du spectre moyen on été divisées par leur maximum. Cette
manipulation permet d’avoir des valeurs entre 0 et 1 qui sont alors comparables. L'intensité

77
relative des signaux a un impact sur l'enveloppe temporelle et le spectre moyen. Par exemple,
supposons un signal composé de deux sons concentrés sur deux bandes de fréquences
différentes. Considérons que l’intensité du premier son est deux fois plus élevée que celle du
second. Le spectre moyen reflétera cette différence par des intensités relatives différentes
associées aux deux fréquences (Fig. I.14). Ainsi, si la différence d’intensité relative est très
forte, la participation du premier son au spectre moyen peut aller jusqu'à disparaître.
Fig. I.14 : Spectre moyen d’un signal composé de deux sons de fréquences différentes. A gauche, les deux
sons ont la même intensité et à droite, le deuxième son a une intensité deux fois plus forte.
Deux points de réflexion en découlent. Le premier concerne la différence de détectabilité des
signaux qui est, dans le cas des indices sur objet maximisé (spectre moyen, sonogramme,
enveloppe temporelle), relative aux autres signaux. La seconde concerne le fait de prendre en
compte cette différence d’intensité relative. Reprenons l’exemple en Fig. I.14, l’intensité
relative des points varie, mais pas le nombre de pics. L’indice NP est un indice de comptage,
il ne prend donc pas en compte la différence des intensités des pics. Les indices Hf et Ht, par
contre, mesurent l'hétérogénéité des intensités relatives des points de l'enveloppe temporelle et
du spectre moyen. Nous aurions pu donc nous attendre à voir l’indice NP moins sensibles aux
variations d’intensités que les indices Hf et Ht. Ce n’est pas ce que montrent les résultats. En
effet, l’indice NP est plus sensible aux variations d’intensité que les indices Hf et Ht. Dans
notre exemple, la variation d’intensité relative des signaux ne modifie pas le nombre de pics
or ce n’est pas le cas sur des signaux d’oiseaux composés de sons complexes. En effet, un
chant d’oiseau s’étale sur différentes fréquences. Modifier l’intensité relative de ces chants
dans un signal peut faire varier le nombre de pics (NP) sur le spectre. La forme globale de
l'objet, enveloppe ou spectre moyen, varie donc moins que le nombre de pics sélectionnés
avec la variation des intensités relatives des signaux.

78
L'indice AR montre une sensibilité très élevée à la variation d’intensité relative. L'indice ACI
semble assez robuste aux variations d’intensités relatives. Cette caractéristique était attendue
de par la correction mathématique proposée par les auteurs pour réduire cet effet (Pieretti et
al., 2011). Parmi les trois indices de dissimilarité proposés KL semble moins sensible que les
deux autres à la variation des intensités relatives des signaux. Notons que KL est le seul indice
de dissimilarité qui n’est pas une comparaison stricte des valeurs point à point du spectre
moyen. Cela pourrait être à l’origine de cette différence de sensibilité.
Ces effets d'intensité relative sont une cause d'hétérogénéité acoustique qui se reflète,
plus ou moins, dans les indices acoustiques. Si un indice est sensible aux variations
d’intensités relatives des signaux, il est alors légitime de se demander si cette hétérogénéité
acoustique est associable à une mesure de diversité acoustique de la communauté animale. Il
est assez difficile de pouvoir corriger ce biais de détectabilité des signaux sur les indices
présents ici. Le seul recours qui pourrait atténuer cet effet serait d'avoir un très grand
échantillonnage (spatial et temporel) de signaux issus de l'activité acoustique de la
communauté animale en supposant que, de ce fait, l’ensemble des enregistrements prendra en
compte les variations d’intensités relatives des chants.
1.5.2 Variation de la durée de l'enregistrement par rapport à la durée du signal étudié
Le rapport du temps d'enregistrement sur le temps du signal acoustique d'intérêt a un impact
sur les valeurs de tous les indices sauf des indices NP et AR. Ces résultats correspondent avec
les résultats de l’effet du bruit de fond. En effet, le bruit de fond ne semble pas avoir un effet
fort sur les valeurs de ces deux indices. L’ajout donc d’un signal correspondant à un bruit de
fond n’a pas un impact fort sur la valeur de ces deux indices.
1.5.3 Variation du décalage temporel
L'impact du démarrage temporel des signaux est assez fort pour tous les indices sauf pour
l’indice NP, dont l'effet négatif ne semble visible que pour des signaux composés d'un nombre
important de sons et sur l'indice AR, qui ne semble pas du tout affecté par cette variation. Pour
les indices M et Ht, ce résultat n'est pas surprenant. La complexité de l'enveloppe temporelle
diminue quand la répartition des sons le long du signal est plus hétérogène. Par contre, ce n'est
pas attendu pour l'indice Hf. Ce résultats révèle peut être un effet du décalage temporel des

79
sons sur le spectre moyen. En effet, prenons le cas des séquences synchrones (cas 1) et
alternées (cas 2) pour le cas extrême de 10 espèces. Dans le premier cas, la même information
est moyennée sur un nombre de points beaucoup plus faible que dans le cas 2 dont le signal a
une durée dix fois plus élevée. Cet effet sur le spectre moyen est révélé par l’indice Hf.
Les différences dans le spectre moyen ne sont pas mises en évidence par les indices de
dissimilarité. Ils sont donc peu sensibles au décalage de chant dans le signal.
1.5.4 L'effet du bruit ambiant
Deux études ont été conduites de manière indépendante : Paisley-Jones (2011) et Depraetere
et al. (2012). Ces deux études ont choisi de corriger les indices proposés par Sueur et al.
(2008b) par l'énergie du signal afin de mettre un poids négatif aux signaux ne présentant
qu’un bruit ambiant, qui présentent des valeurs d’indices très fortes. Si cette approche est
intéressante, elle suppose que deux signaux composés des mêmes sons mais d'intensité
différente pourraient être dans le cas de la pondération par l'énergie, associés à deux niveaux
de diversité acoustique différents.
L'indice NP est né de la réflexion du problème de sensibilité au bruit ambiant de
l'indice Hf soulevé par Depraetere et al. (2012). Les résultats confirment que NP est beaucoup
moins sensible au bruit ambiant que l'indice spectral Hf. L'effet du bruit ambiant est donc, à
part pour les indices NP et AR, une source indéniable de variation. Pour l'instant, aucun indice
de distance acoustique ne permet d'éviter totalement ce problème. Leur utilisation devra donc
tenir compte de cette limite.
Traiter le bruit ambiant d'un signal sonore est un sujet très sensible. Il existe des
moyens pour limiter ce bruit comme l'utilisation de filtres utilisant des seuils d’intensité ou de
fréquence. Dans le cas de la méthode RAS, les filtres fréquentiels ne sont pas les plus
recommandés. En effet, toutes les fréquences peuvent être potentiellement utilisées par un
individu chanteur. De plus, le spectre du bruit ambiant peut être représenté sur toutes les
fréquences. Pour terminer avec les spectres fréquentiels, si le matériel impose ses propres
limites de détection fréquentielle, l'utilisation de filtres peut entrainer une perte d'information
non négligeable. Ils seront utilisés dans nos travaux uniquement si l'enregistrement présente
des bruits d'origine anthropique limités à de basses fréquences (en dessous de 1kHz).
Concernant les filtres d’intensité, ils peuvent être très grossiers. L’intensité du bruit varie
généralement en fonction de la fréquence considérée. Une méthode a été mise au point dans le

80
chapitre 2 pour limiter le bruit ambiant en considérant cette variation le long des bandes de
fréquences.
En ce qui concerne les indices de dissimilarité, les résultats montrent que les différences entre
les signaux sont moins fortes lorsque le bruit ambiant est élevé. Ce résultat est attendu
puisque le bruit ambiant étant commun entre les enregistrements, il aura un impact négatif sur
leurs différences spectrales. Pour un bruit ambiant équivalent dans deux enregistrements, il est
attendu que la valeur que celui-ci prend dans l’hétérogénéité du spectre moyen du premier
enregistrement s’annule avec la valeur du bruit ambiant dans le spectre moyen du deuxième
enregistrement. Ainsi, le bruit ambiant n’est pas pris en compte dans le résultat de l’indice.
1.5.5 Les indices de dissimilarité : des indices point à point
Les indices Df et Ks sont dit point à point car ils reposent sur une différence deux à deux des
points qui composent le spectre moyen. De très récentes études ont mis en évidence les
limites des indices points à points (Lellouch, 2012) qui impliquent notamment des réflexions
sur le problème d'alignement des séquences pour les rendre comparables. C'est face à cette
difficulté que l'indice Dt a pour l'instant été mis de côté. En effet, il nous a semblé difficile de
mesurer une distance acoustique sur la base d'une enveloppe temporelle dont les points sont
difficilement comparables. Au contraire, aligner les points du spectre moyen a un sens
acoustique, chaque point représentant une fréquence identique au même point d'un autre
spectre moyen. Cette difficulté de l’alignement temporel est valable pour l'indice 1-RV
(Robert & Escoufier, 1976) qui se base sur une corrélation des points de deux matrices
correspondantes à deux sonagrammes. Cet indice a été utilisé dans les travaux présentés en
chapitre 2 sur la base de comparaison non pas de signaux, mais de sons espèce-spécifiques. Si
cet indice mesure à la fois les distances spectrales et temporelles de deux signaux, son
utilisation reste discutable sur la partie temporelle.
1.6 Conclusion
Les indices acoustiques présentés dans ce chapitre capturent l'hétérogénéité acoustique d'un
signal sonore, or toute hétérogénéité sonore n'est pas liée à la diversité acoustique d'une
communauté animale. L'objet d'étude de ce chapitre n'est pas de démontrer la pertinence de

81
ces indices à mesurer la diversité acoustique mais de comparer leur sensibilité aux autres
causes d'hétérogénéité.
Si les sons sont des causes d'hétérogénéité du signal, les caractéristiques physiques du
milieu dans lequel se déplacent les ondes jouent un rôle important. Ainsi, un milieu ouvert ou
fermé n'aura pas le même impact sur la propagation des ondes. Les phénomènes de
diffraction, réverbération, absorption dépendront des conditions météorologiques et des
obstacles présents dans le milieu de propagation (Richards & Wiley, 1980; Wiley & Richards,
1978). Ces conditions, tout aussi intéressantes que les quatre causes d'hétérogénéité que nous
avons considérées, sont plus difficiles à contrôler en laboratoire et leur impact sur le
comportement de nos indices est donc plus difficile à tester. Des effets croisés sont possibles
entre les différentes causes d'hétérogénéité. Par exemple, il peut y avoir un lien entre l'activité
biotique et les conditions physiques du milieu (Wiley, 1991).
Considérant tous les tests effectués dans ce chapitre, l'indice NP semble globalement
le moins sensible aux causes d'hétérogénéité non désirées, c'est pourquoi nous avons choisi
d'utiliser cet indice dans la suite de nos travaux. En ce qui concerne les indices de distance
acoustique, peu de différences dans leur comportement ont été notées. Si d’autres indices de
dissimilarité existent, leurs calculs reposent sur une comparaison point à point. Bien que
consciente des limites et sensibilités de ces indices de distance, l'indice Df a été choisi pour la
suite nos travaux. Cet indice est notamment beaucoup moins coûteux en temps de calcul, ce
qui n’est pas une caractéristique négligeable en regard du traitement de milliers
d’enregistrements sonores.
L’étude théorique du chapitre 2 soulève la question du lien entre diversité acoustique
et diversité spécifique, phylogénétique et fonctionnelle à partir de l'indice Df. La
méthodologie qui a été employée a permis d'éviter les effets de bruit ambiant, de variations
d’intensités relatives, de décalage temporel des signaux ou de différence de rapport entre la
taille de l'enregistrement et la taille de la séquence sonore.
Les chapitres 3 et 4 reposent sur des enregistrements in natura incluant toutes les
causes possibles d'hétérogénéité énoncées. Tous les sites du chapitre 3 se trouvent dans des
conditions les plus similaires possibles en termes de formation végétale, l'échantillonnage est
très large et les effets de bruits ambiants ont été limités par une méthode de filtre du spectre
moyen qui prend en compte la variabilité du bruit de fond dans les fréquences. Le cas du
chapitre 4 est beaucoup plus complexe puisqu'il utilise des enregistrements venant de sites
dont la formation végétale diffère.

82

83
Chapitre 2 Mesurer la biodiversité par le son : la
diversité acoustique peut-elle refléter la
diversité phylogénétique et fonctionnelle de
communautés d’oiseaux ?
Gasc A, Sueur J, Jiguet F, Devictor V, Grandcolas P, Burrow C, Depraetere M Pavoine S
(2013) - Assessing biodiversity with sound: do acoustic diversity indices reflect phylogenetic
and functional diversities of bird communities? Ecological Indicators, 25: 279-287
Communications :
Gasc A., J. Sueur, V. Devictor, P. Grandcolas, F. Jiguet, C. Burrow and S. Pavoine (2011)
Assessing biodiversity with sound: what do acoustic diversity indices measure?. International
Bioacoustic congress (IBAC) congress, presentation 15 minutes.
Gasc A., J. Sueur, V. Devictor, P. Grandcolas, F. Jiguet, C. Burrow and S. Pavoine (2011)
Assessing biodiversity with sound: what do acoustic diversity indices measure? European
Ecology Federation (EEF) congress, flash presentation 3 minutes.

84

85
Assessing biodiversity with sound: do acoustic diversity indices reflect phylogenetic and
functional diversities of bird communities?
Gasc A.1,2, J. Sueur1, F. Jiguet2, V. Devictor3, P. Grandcolas1, C. Burrow2, M. Depraetere1 and
S. Pavoine2, 4
1Muséum national d'Histoire naturelle, Département Systématique et Évolution, UMR 7205
CNRS OSEB, 45 rue Buffon, Paris, France
2Muséum National d’Histoire Naturelle, Département Ecologie et Gestion de la Biodiversité,
UMR 7204-CNRS-UPMC, CP 51, 55-61 rue Buffon, 75005 Paris, France
3Université Montpellier 2, Institut des Sciences de l’Evolution, UMR CNRS-UM2 5554,
Place Eugène Bataillon, 34095 Montpellier Cedex 05, France
4Mathematical Ecology Research Group, Department of Zoology, University of Oxford,
South Parks Road, Oxford, OX1 3PS, UK
*Author for correspondence
Amandine GASC
Muséum national d’Histoire naturelle
Département Systématique et Évolution
UMR CNRS 7205 OSEB
CP 50, 45 rue Buffon
F-75005 Paris, France
Tel. + 33 1 40 79 31 34
Fax. + 33 1 40 79 56 79
Email. [email protected]

86
II.1 Abstract/Résumé
II.1.1 Abstract
Measuring biodiversity is a challenging task for research in taxonomy, ecology and
conservation. Biodiversity is commonly measured using metrics related to species richness,
phylogenetic-, or functional-trait diversity of species assemblages. Because these metrics are
not always correlated with each other, they have to be considered separately. A descriptor of
animal diversity based on the diversity of sounds produced by animal communities, named
here the Community Acoustic Diversity (CAD), was recently proposed. In many cases, the
CAD could be easier to measure than other metrics. Although previous analyses have
revealed that acoustic diversity might increase as species richness increases, the ability of
CAD to reflect other components of biodiversity has not been formally investigated. The aim
of this study is to test theoretically whether functional and phylogenetic diversities could be
reflected by acoustic diversity indices in bird communities. Data on species assemblages were
collated by the French Breeding Bird Survey describing spatial and temporal variation in
community structure and composition across France since 2001. Phylogenetic and functional
data were compiled from literature. Acoustic data were obtained from sound libraries. For
each of the 19420 communities described, indices of phylogenetic, functional and acoustic
diversity of bird communities were calculated based on species’ pair-wise distance matrices
and species’ abundances. The different aspects of biodiversity were compared through
correlation analyses. The results showed that acoustic diversity was correlated with
phylogenetic diversity, when the branch lengths of the tree were considered, and to functional
diversity, especially body mass and reproduction. Correlations between phylogenetic,
functional and acoustic distances among species did not entirely explain the correlations
between phylogenetic, functional and acoustic diversity within communities. This result was
interpreted as an effect of local ecological processes underpinning how bird communities
assemble. Comparing the diversity patterns with a null model, phylogenetic and functional
diversities were significantly clustered whereas acoustic diversity was not different from what
was expected by chance. A comparison between acoustic indices showed that spectral
component of acoustic diversity seems more appropriate to reveal bird phylogenetic diversity
whereas temporal component seems more adapted to reveal functional diversity of a bird
community. Overall, even if the processes at the origin of the different facets of biodiversity

87
are different, CAD reveals part of phylogenetic diversity and to some extent of functional
diversity.
Keywords: Acoustic diversity, Biodiversity assessment, Bird community, Biodiversity
indices.
Highlights
Acoustics is a developing tool for animal diversity assessment
Acoustic diversity of 19,420 French bird communities was theoretically analysed
Correlations between acoustic, phylogenetic, functional diversities were computed
Acoustic and phylogenetic diversities were correlated when considering branch lengths
Acoustic diversity was correlated with body mass and reproduction diversity
II.1.2 Résumé
Mesurer la biodiversité est un enjeu pour les recherches en taxonomie, écologie et biologie de
la conservation. La biodiversité est couramment estimée par la richesse spécifique, la diversité
des traits phylogénétiques ou fonctionnels d'assemblages d'espèces. Ces mesures ne sont pas
toujours corrélées entre elles et doivent donc être considérées séparément. Récemment, un
indice de mesure de la diversité acoustique basé sur la diversité des sons produits par les
espèces animales d'une communauté, qui sera nommé CAD pour « Community acoustic
diversity », a été proposé. Bien souvent, la CAD est beaucoup plus simple a obtenir que les
autres mesures. Bien que de précédentes analyses ont mis en évidence que la diversité
acoustique pouvait augmenter avec le nombre d'espèces chanteuses, la capacité du CAD à
refléter d'autres composants de la biodiversité n'a pas été évaluée. L'objectif de cette étude et
de tester, de manière théorique, si les diversités phylogénétique et fonctionnelle de
communautés d'oiseaux peuvent être reflétées par des indices de diversité acoustique. Des
données d'assemblages d'espèces d'oiseaux en communautés ont été collectées par programme
de Suivi Temporel de Oiseaux Communs (STOC) qui décrit les variation spatiale et
temporelle de la composition des communautés d'oiseaux à travers la France depuis 2001. Les
données phylogénétiques et fonctionnelles ont été rassemblées à partir de la littérature
scientifique disponible. Pour les 19420 communautés décrites, des indices des diversités
phylogénétique, fonctionnelle et acoustique on été calculés à partir de matrices de distances

88
espèce-espèce et des relevés d'abondances des espèces. Les corrélations entre les différents
aspects de diversités ont ensuite été étudiées. Les résultats montrent que la diversité
acoustique est positivement corrélée avec la diversité phylogénétique lorsque les longueurs
des branches de l'arbre phylogénétique sont prises en compte et à la diversité fonctionnelle
considérant les traits de poids et de reproduction. Les corrélations entre les distances
phylogénétiques, fonctionnelles et acoustiques entre les espèces deux à deux n'expliquent pas
entièrement les résultats des diversités des assemblages d'espèces. Ce résultat a été interprété
comme un effet local du à l'assemblage des espèces au sein de la communauté. La
comparaison des valeurs de diversités par rapport à un modèle nul révèle que les diversités
phylogénétique et fonctionnelle sont significativement groupées selon leur proximité
phylogénétique et fonctionnelle. La diversité acoustique ne semble pas différente de celle que
l'on pourrait trouver par chance, sans assemblage particulier d'espèces. Une comparaison des
indices de diversité acoustique montre que la composante spectrale des chants semblent plus
appropriée pour révéler la diversité phylogénétique tandis que la composante temporelle
semble plus appropriée pour révéler la diversité fonctionnelle des communauté d'oiseaux.
Dans l'ensemble, si les processus à l'origine des différentes facettes de la biodiversité ne
semblent pas les mêmes, le CAD révèle, dans une certaine mesure, une part de la diversité
phylogénétique et fonctionnelle.
Mots-clefs: Diversité acoustique, mesure de biodiversité, communauté d'oiseaux, indice de
biodiversité.
Points importants
L'étude du son est un outil en développement pour l'évaluation de la diversité animale
La diversité acoustique de 19 420 communautés d'oiseaux françaises a été analysée de
manière théorique
Les corrélations entre diversités acoustique, phylogénétique et fonctionnelle ont été étudiées
Les diversités acoustique et phylogénétique sont corrélées lorsque la longueur des branches de
l'arbres est considérée
La diversité acoustique et les diversités fonctionnelles (poids et traits liés à la reproduction)
sont corrélées

89
II.2 Introduction
Diversity indices have been growing in number, but have been developed separately
by different research fields related to evolution or ecology (Pavoine & Bonsall, 2011). At the
community level, indices mostly used to describe biodiversity include species richness,
species relative abundance, and rarity. However, several recent studies have shown that these
traditional metrics are insufficient to capture all facets of diversity and have proposed to focus
instead on species functional and phylogenetic distinctiveness. Functional diversity is
measured from functional traits that describe a variety of roles that different organisms play in
their ecosystem (Petchey and Gaston, 2002). Depending on the ecosystem process analyzed,
these traits might depend, for instance, on physiological, life-history, morphological,
ecological or behavioural characteristics. In the last two decades, there has been an increasing
number of functional diversity studies focusing on the local biotic and abiotic interactions that
could explain the processes of species assemblages (Pavoine & Bonsall, 2011; Pillar et al.,
2009). Phylogenetic diversity measures the dispersion of species belonging to a community
over a phylogenetic tree (Faith, 1992; Pavoine et al., 2005a; Vane Wright et al., 1991). In
theory, phylogenetic diversity can reflect functional diversity if functional traits have a strong
phylogenetic signal and thus if the phylogenetic distance between any two species reflects
their difference in terms of a combination of functional traits (Webb et al., 2002). In practice,
few studies have compared results collected from different aspects of biodiversity and some
have shown that these were only partially correlated (Devictor et al., 2010; Pavoine et al.,
2005a; Petchey et al., 2002).
Working on phylogenetic and functional diversities can shed light on ecological
processes. Indeed, functional traits could be affected by local ecological processes including
competition and environmental filters. Competition leads to functional diversification
contrary to environmental filters. Therefore, low functional diversity might be obtained with
high species richness and abundance if environmental constraints filter species with similar
traits (Holdaway & Sparrow, 2006; Petchey et al., 2007). Moreover, an evolutionary
convergence of the functional traits of distantly related lineages can lead communities to
include many lineages (high phylogenetic diversity) but which are functionally similar (low
functional diversity) (e.g.; Cavender et al., 2004; Grandcolas, 1993, 1998). In addition, in
regions where species recently emerged by rapid radiation, high species richness might be
attained with low phylogenetic diversity as species have diverged only recently (Slingsby &

90
Verboom, 2006). Overall, although phylogeny and functional trait variations are becoming
frequently used to assess biodiversity, they do not necessarily correlate due to evolutionary
and ecological processes and have to be considered separately.
Beyond these components of biodiversity, acoustic diversity recently emerged as a
possible relevant indicator of biodiversity for several reasons (Obrist et al., 2010; Sueur et al.,
2008b). First, there are practical advantages to using passive acoustic methods in conservation
surveys. Acoustics allows exploring habitats that are difficult to access such as canopy (Riede,
1997), marine and freshwater habitats (Luczkovich et al., 2008), soil and interior structures of
plant (Mankin et al., 2000), and dark environments (Meyer et al., 2011; Obrist et al., 2004).
Moreover, compared to classic methods of biodiversity assessment based on field inventories,
passive acoustic censuses are less costly especially when automatic recordings are used which
can ensure large temporal and spatial scale surveys. The description of sound provides
information on acoustic diversity at individual (Pollard et al., 2010), population (Dawson &
Efford, 2009; Forrest, 1988), species (Brandes et al., 2006; Skowronski & Harris, 2006),
community (Cadoso & Price, 2010; Diwakar & Balakrishnan, 2007a, 2007b; Riede, 1997,
1993) and landscape levels (Pijanowski et al, 2011a, 2011b). At the landscape scale, a
soundscape ecological approach distinguishes three components in the soundscape, namely
the biophony, the geophony and the anthrophony. Biophony is mainly defined as “a collection
of sounds produced by all living organisms in a given habitat over a specified time”
(Pijanowski et al, 2011a). The present paper considers the diversity of the acoustic
community here named for the first time Community Acoustic Diversity (CAD). Biophony is
very closely related to acoustic community. However, the concept of community includes not
only the species assemblage but also the interactions between species that are supposed to
compete for the acoustic resource. The CAD considers the competitive interaction for the
sound space, structured by the sound resource. Contrary to biophony, community acoustic
diversity intends to reflect all ecological processes that determine how many diverse biotic
sounds co-occur in a community. Acoustic signal of an animal community is generally
analysed using three approaches: species identification by an expert, species identification
using automatic recognition and global acoustic measure without species identification. As
species identification of singing individuals is necessary and requires a high level of expertise,
identification has to be achieved by trained experts (Dickinson et al., 2010) or by automatic
classification processes (e.g. Acevedo et al., 2009; Han et al., 2010). Both approaches are
difficult to apply due to song overlap observed in rich acoustic communities. Consequently,
new indices have been recently developed to obtain a global measure of acoustic diversity

91
without any species identification. For instance, an entropy-like index based only on the
frequencies recorded was proved to increase with the number of species. Likewise, a
dissimilarity index based on the variations of frequencies collected was shown to be inversely
correlated with the number of shared species between two focal communities (Depraetere et
al., 2012; Sueur et al., 2008b). An acoustic variability index was also shown to evolve with
the dynamics of bird communities (Farina et al., 2011; Pieretti et al., 2011).
These approaches generally support a correlation between the CAD and species
richness, and the CAD and abundance, but whether and how CAD can be related to other
biodiversity components has never been investigated. Rather than working on pure simulated
communities as Sueur et al. (2008) did it first, we chose to test the index and to try to
understand the information it can provide by working on bird communities that were
described through data gathered by local field workers. A theoretical analysis was performed
on data extracted from different sources: (i) bird community composition from the French
breeding bird survey, (ii) phylogenetic and functional data corresponding to each species from
literature and (iii) acoustic data corresponding to each species from different sound libraries to
answer the following questions: i) Do spectral and temporal parts of an acoustic signal have
different impacts on acoustic diversity calculation? ii) Does the acoustic diversity reflect
phylogenetic and/or functional diversities of bird communities? iii) Do the acoustic,
phylogenetic and functional diversities result from the same ecological and evolutionary
processes?
II.3 Material and methods
Acoustic, phylogenetic and functional diversity indices were computed using the quadratic
entropy index (Chave et al., 2007; Pavoine et al., 2005b, 2004; Pavoine & Bonsall, 2011;
Rao, 1986). The quadratic entropy index was applied to abundance data describing bird
communities and a pair-wise distance matrix among species for each diversity type (i.e.
acoustic, phylogenetic, and functional diversity) (Fig. II.1). Two types of data were necessary
to calculate diversity indices: the detail of bird community assemblage in terms of species’
abundances and acoustic, phylogenetic and functional data used to calculated the distances
between species.
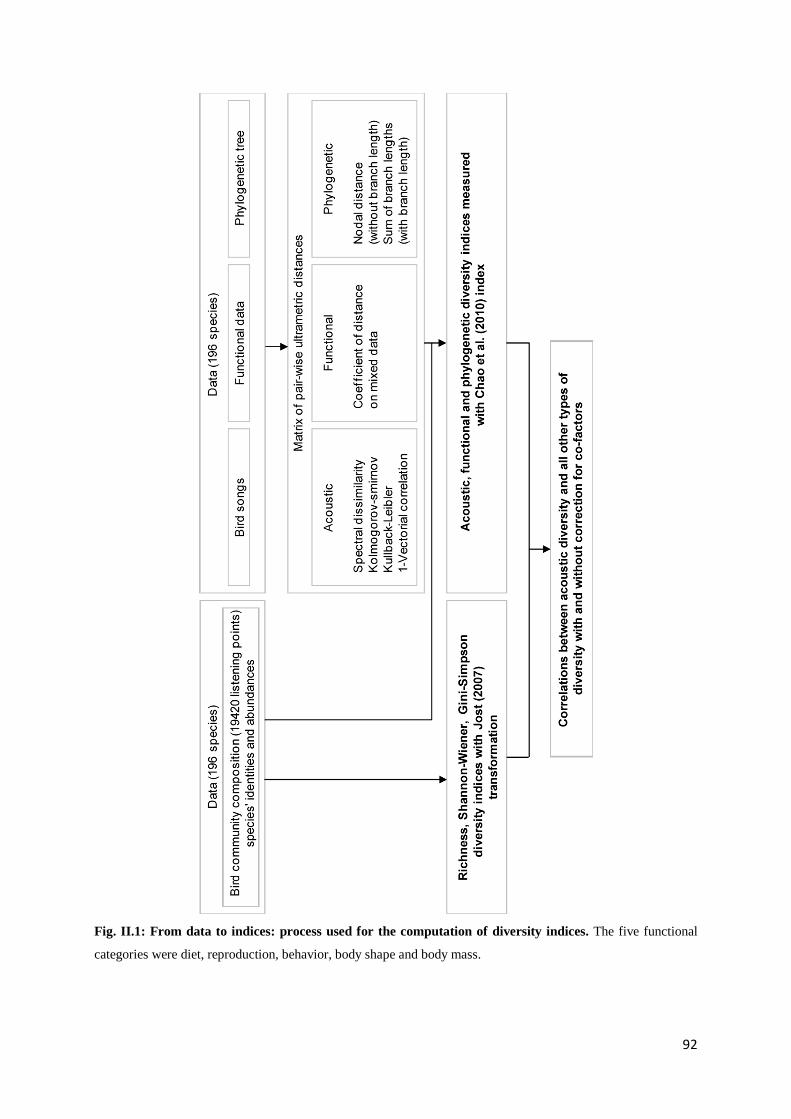
92
Fig. II.1: From data to indices: process used for the computation of diversity indices. The five functional
categories were diet, reproduction, behavior, body shape and body mass.

93
II.3.1 Data
II.3.1.1 Bird communities
Bird community compositions were extracted from data collected by the French breeding bird
survey (FBBS; Jiguet et al., 2011). This survey is based on a standardized protocol conducted
by a volunteer network coordinated by the Muséum national d'Histoire naturelle (Paris,
France, Jiguet et al., 2011; Julliard et al., 2006). A sampling quadrat of 2x2 km is randomly
localized in a 10 kilometers circle around a location chosen by a volunteer. Ten points are
then located homogeneously and proportionately to the habitats occurring in the quadrat.
Points have to be separated by at least 300 meters. Every spring, each volunteer spends five
minutes on each point at two dates before and after May 8th. For each species, the volunteer
counts the number of detected individuals, which is considered as an index of species
abundance. For each point visited at least one year between 2001 and 2010, average species
abundances over all available years were calculated, for a total of 19420 points. Among the
species detected in at least two points, 196 species were selected based on song recording
availability (see 3.1.2. and list of species in supplementary information on Table S.II.1,
supplementary information available in "Annexe 2").
II.3.1.2 Bird songs
A species-specific territorial song of each bird species was selected (Bossu & Charron, 2004,
1998; Deroussen, 2011, 2006, 2001; Roché, 1990; Xeno-canto, http://www.xeno-canto.org)
according to two conditions: (i) the song had to be emitted by an isolated individual, and (ii)
the Signal to Noise Ratio (SNR) of the recording had to exceed 10. This ratio was estimated
following: /SNR As An= , where As and An are the probability mass functions of the
amplitude envelope of 0.5 second of noise and of 0.5 second of signal both randomly chosen
along the recording considered, respectively. This recording selection constituted our acoustic
data.
II.3.1.3 Phylogenetic data
The phylogenetic tree came from a mega-phylogeny approach performed by Thuiller et al.
(2011) who combined supertree and supermatrix approaches with sequences obtained from
GenBank. This tree has branch lengths that can be defined in terms of time since speciation.
Within the 340 bird species composing the tree, a tree of 192 species was sub-sampled. This

94
tree was completed with four missing species (Lanius meridionalis, Anas strepera, Ardea
alba, Larus fuscus) using literature data (Davis et al., 2008). The final tree was composed of
the 196 bird species selected for acoustics.
II.3.1.4 Functional data
Forty-eight functional traits were extracted from Devictor et al. (2010, n=17), Cramp et al.
(1998, n=17), Davies et al. (2004, n=13) and BirdLife International (2004, n=1) (see
supplementary information Table S.II.2, supplementary information available in "Annexe 2").
The traits were organized in five functional categories: behavior (social, migration,
nocturnal), diet (type, foraging, substrate), morphology (body size, tarsus size, tail size, span),
reproduction (habitat type, nest construction, parental behavior), and body mass. Morphology
variables were highly correlated to body mass. In order to have an idea of body shape,
independently to body mass, the morphology variables were transformed as follow:
3
VmVmcVp
=
with Vmc the morphological variable corrected, Vm the morphological variable and Vp the
body mass variable.
In the following text, the transformed morphology variable will be referred to "body shape".
A logarithmic transformation was applied on "body shape" and "body mass" variables.
II.3.2 Distance matrices
From the data described in the previous subsection distinct matrices of distances among
species were calculated and used to obtain measures of acoustic, phylogenetic, and functional
diversities.
II.3.2.1 Acoustic distances
Evolution might not have been constant for all song features (Robillard & Desutter-
Grandcolas, 2011). For this reason, the acoustic indices used in this paper are not based on
classical acoustic parameters but on a single distance measure embedding temporal and/or
spectral characteristics. Four types of pair-wise acoustic distances between species were
computed:

95
i) A frequency distance index (Df), as described in Sueur et al. (2008b), was based on
the difference between a pair of mean spectra. Each spectrum was computed as the average of
a Short Time Fourier Transform with a non-overlapping sliding window (length = 512 points,
time precision = 0.012 s, frequency precision = 86 Hz). Each spectrum was transformed into a
probability mass function Si(f) and vectorial difference was calculated between the two
spectra to be compared.
ii) Kolmogorov-Smirnov distance (KS), which was defined as "the maximum value of
the absolute difference between two cumulative distribution functions" (Rachev, 1991), was
computed between cumulated spectra.
iii) Symmetric Kullback-Leibler (KL) distance was estimated by computing the
relative entropy between two probability frequency spectra (Kullback et al., 1951).
iv) The similarity RV correlation coefficient was used as developed by Escoufier
(1973) and Robert and Escoufier (1976). RV correlation measures the correlation between two
matrices. Short-Term Fourier transform with a non-overlapping sliding window was
computed for each recording leading to a data matrix. The columns of each matrix were the
successive spectra computed along the time scale with a time precision of 0.012 s and a
frequency precision of 86 Hz. RV was computed according to Kazi-Aoual et al. (1995) and
Josse et al. (2007). The complement 1-RV was computed to obtain a dissimilarity measure.
II.3.2.2 Phylogenetic distances
Phylogenetic distances were calculated as the number of branches on the shortest path that
connects two species in the phylogenetic tree without branch length. A phylogenic distance
including branch lengths was also calculated as the sum of branch lengths on the shortest path
that connects two species.
II.3.2.3 Functional distances
Functional distance matrices were computed using Pavoine et al.'s coefficient of distance
calculated from species-specific traits (Pavoine et al., 2009). This index, based on Gower
distance (Gower, 1971), allows measuring functional distances from any type of variables
(e.g. quantitative, nominal, ordinal, proportional, binary, circular). The types of the traits here
considered are given in Table S.II.2. Details on these types of traits can be found in Pavoine et
al. (2009). Quantitative traits and (rank-transformed) ordinal traits were treated by formula 3
in Pavoine et al. (2009, Euclidean metric). The distance between two species based on a
nominal trait was 1 if the two species had similar attributes and 0 elsewhere. Proportional

96
(fuzzy) variables were treated by formula 11 in Pavoine et al. (2009). Multichoice variables
were treated by Jaccard (1901) index. Five distance matrices were separately calculated using
the five functional groups defined a priori (diet, behavior, body shape, weight, reproduction,
Table S.II.2). These functional groups contained several traits each (see Table S.II.2). An
average distance over all traits per group was obtained as defined in Pavoine et al. (2009).
Traits were weighted to take account of their pair-wise correlations as defined by Petchey
(2009). All functional distance matrices were transformed into ultrametric distances using the
UPGMA (Unweighted Pair Group Method with Arithmetic Mean) method (Legendre &
Legendre, 1998).
II.3.3 Diversity indices
Species richness (number of species - 1), Shannon and Gini-Simpson indices were calculated
for each community (Magurran, 2004). In order to respect the replication principle, Shannon
and Gini-Simpson indices were corrected using the exponential of Shannon and the inverse of
Gini-Simpson index as 1/(1-Gini-Simpson values) (Chao et al., 2010). These indices equal
zero for a community with a single species. Acoustic, phylogenetic and functional diversity
indices were computed using the quadratic entropy from the distances previously calculated
and species’ abundances within bird communities (Fig. II.1). In Chao et al. (2010), the
authors proposed a new correction for the quadratic entropy to comply the replication
principle. This correction was applied and the resulting index will be called Chao et al.' index
in the following text. The resulting diversity estimates had symmetric distributions. A part of
the correlation observed between the acoustic, phylogenetic and functional diversity indices
could possibly be driven by the intrinsic mathematic property of these indices associated with
the inverse Gini-Simpson index. All diversities calculated following Chao et al. (2010) index
shared this co-variable due to the mathematical formula. To correct for this co-variable, the
three diversities values (acoustic, phylogenetic and functional) were divided by 1/(1-Gini-
Simpson values), as recommended by Jost (2007).
II.3.4 Statistical analyses
Acoustic, phylogenetic and functional diversities were computed with the same mathematical
methodology that depends on pair-wise distances between species and species’ abundances.
This led to a dependency between the three diversities. Indeed, if two pair-wise distance

97
matrices were strongly correlated (i.e. pair-wise acoustic and phylogenetic distances between
species were strongly correlated), and the corresponding values of the diversity index were
correlated as well (i.e. the acoustic and phylogenetic diversities of the bird communities were
also strongly correlated), the similarity between two aspects of diversity (i.e. phylogenetic and
acoustic diversity) could be explained only by the similarity of the distances between species
and not by the assemblage processes that underpin bird communities. By contrast, a difference
in the correlations calculated between distances at species level and those calculated between
diversities at community level would indicate that diversity at the community level could not
be considered just as the random addition of pair-wise species distances but that the
community structure, in terms of how species assemble, had an influence on the diversity of
the community. Finally, if two matrices of pair-wise distances between species were not (or
moderately) correlated, then the corresponding values of the diversity index within
communities might still be correlated if similar stochastic, or deterministic ecologically-based
processes, influence the community composition. To counteract this issue, statistical analyses
were achieved at species level based on the distance matrices and at community level based
on the calculation of the diversity indices (see below).
II.3.4.1 Species level
A preliminary Mantel test with Spearman correlation and 999 permutations (Hardy &
Pavoine, 2012; Mantel, 1967) was used to compare the four acoustic dissimilarity indices.
The Mantel test was also applied to compare acoustic distances with phylogenetic and
functional distances. The equality between phylogenetic distances and acoustic distances
between species would imply evolutionary-based mechanisms such as the Brownian model of
evolution if the phylogeny has branch lengths or punctuated evolution of species’ acoustic
properties if the phylogeny has no branch lengths (Brooks & McLennan, 1991; Legendre et
al., 2008; Robillard & Desutter-Grandcolas, 2011). Similarly, the equality between functional
distances and acoustic distances between species would imply that species’ acoustics could
depend directly or indirectly via a correlation with a co-factor, on the trait(s) analysed.
II.3.4.2. Community level
Spearman correlation coefficients were calculated between the different aspects of
biodiversity at the community level (see the description of the diversity indices above). No
test was performed because the high number of communities (19420) is known to lead to

98
significant tests (low p-values) even with a correlation coefficient close to zero. In that case,
analysing the correlation coefficient was more informative than tests.
To reveal if ecological processes can explain the observed composition of bird
assemblages, Webb et al. (2002) suggested to compare the observed diversity within each
community with the one expected according to a null model. The following null model was
used: for each community, the distribution of species abundances, and the number of species
were fixed but species identities were randomly extracted from the 196 species pool (model
1p in Hardy, 2008). For each aspect of biodiversity (functional, phylogenetic and acoustic
diversity), Chao et al. (2010) index was calculated for the corresponding artificial
communities. This process was repeated 999 times. At the end for each aspect of diversity and
each community, 1000 theoretical diversity values were calculated (one obtained from the real
communities and 999 obtained based on the null model). Then a new diversity value within
the community considered was defined by: * ( ) /obs sim DsimD D D SD= − where Dsim is the
theoretical diversity values, Dobs is the observed diversity value and SDDsim the standard
deviation of the theoretical diversity values. The index D* was calculated for each of our
19420 communities. The null model chosen assumed that the distribution of species within
communities was not dependent on species phylogenies, traits or acoustics. Negative values
of the corrected index were expected if the species with close characteristics tended to co-
occur in local communities. In contrast, large positive values were expected if species co-
occurring within local assemblages tended to have distinct characteristics. A value close to
zero indicated that the composition of the community supported the null model. These trends
were visualized with histograms.
All calculations were achieved with the R software (R Development Core Team, 2010)
including the packages ‘seewave’ (Sueur et al., 2008a), ‘ade4’ (Dray & Dufour, 2007),
‘vegan’ (Oksanen et al., 2011), and ‘FactoMineR’ (Husson et al., 2011).

99
II.4 Results
II.4.1 Species level
Correlations between the acoustic distances were all significant (Table II.1). All distances
associated with the spectral component only (i.e. Df, KL and KS) were highly correlated. Even
if high, the lowest correlations were between 1-RV and the other indices. Due to high
redundancy between Df, KS and KL and because these three indices measure the same sound
component, only Df and 1-RV indices were considered in the following analyses.
Correlations between the acoustic distances and the functional distances were all
significant except between reproduction category and 1-RV. The significant values were low
to moderate ranging from 0.061 to 0.307 (Table II.2). The highest values were observed
between acoustic distances and distances among species defined by differences in their body
mass and in their body shape. The correlations between acoustic distances and phylogenetic
distances were significant but moderate (ranging from 0.124 to 0.296). The highest correlation
was observed with phylogenetic diversity including branch lengths.
Table II.1: Mantel correlations, based on Spearman method with randomization, between the different
matrices of pair-wise acoustic distances between species: frequency distance (Df), Kolmogorov-Smirnov
distance (KS), Symmetric Kullback-Leibler (KL) and dissimilarity RV correlation (1-RV).
Acoustic distances Df KL KS
KL 0.902, p= 0.001
KS 0.864, p= 0.001 0.912, p= 0.001
1-RV 0.785, p= 0.001 0.838, p= 0.001 0.841, p= 0.001
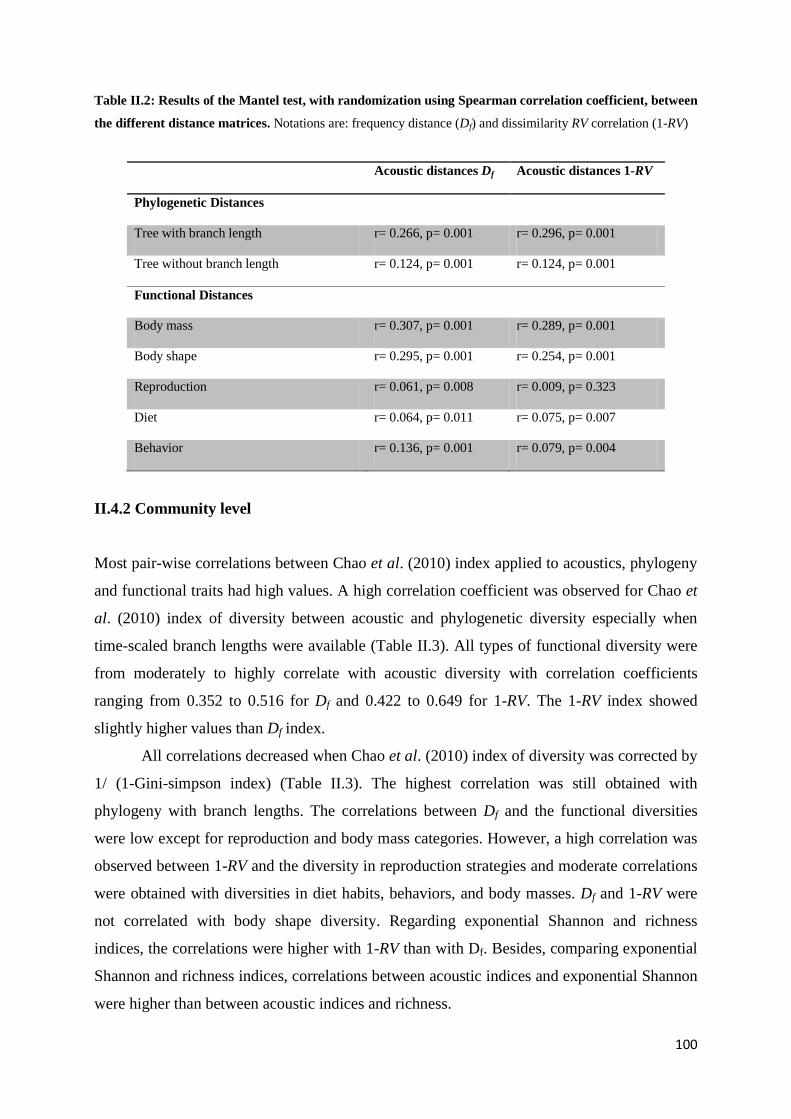
100
Table II.2: Results of the Mantel test, with randomization using Spearman correlation coefficient, between
the different distance matrices. Notations are: frequency distance (Df) and dissimilarity RV correlation (1-RV)
Acoustic distances Df Acoustic distances 1-RV
Phylogenetic Distances
Tree with branch length r= 0.266, p= 0.001 r= 0.296, p= 0.001
Tree without branch length r= 0.124, p= 0.001 r= 0.124, p= 0.001
Functional Distances
Body mass r= 0.307, p= 0.001 r= 0.289, p= 0.001
Body shape r= 0.295, p= 0.001 r= 0.254, p= 0.001
Reproduction r= 0.061, p= 0.008 r= 0.009, p= 0.323
Diet r= 0.064, p= 0.011 r= 0.075, p= 0.007
Behavior r= 0.136, p= 0.001 r= 0.079, p= 0.004
II.4.2 Community level
Most pair-wise correlations between Chao et al. (2010) index applied to acoustics, phylogeny
and functional traits had high values. A high correlation coefficient was observed for Chao et
al. (2010) index of diversity between acoustic and phylogenetic diversity especially when
time-scaled branch lengths were available (Table II.3). All types of functional diversity were
from moderately to highly correlate with acoustic diversity with correlation coefficients
ranging from 0.352 to 0.516 for Df and 0.422 to 0.649 for 1-RV. The 1-RV index showed
slightly higher values than Df index.
All correlations decreased when Chao et al. (2010) index of diversity was corrected by
1/ (1-Gini-simpson index) (Table II.3). The highest correlation was still obtained with
phylogeny with branch lengths. The correlations between Df and the functional diversities
were low except for reproduction and body mass categories. However, a high correlation was
observed between 1-RV and the diversity in reproduction strategies and moderate correlations
were obtained with diversities in diet habits, behaviors, and body masses. Df and 1-RV were
not correlated with body shape diversity. Regarding exponential Shannon and richness
indices, the correlations were higher with 1-RV than with Df. Besides, comparing exponential
Shannon and richness indices, correlations between acoustic indices and exponential Shannon
were higher than between acoustic indices and richness.
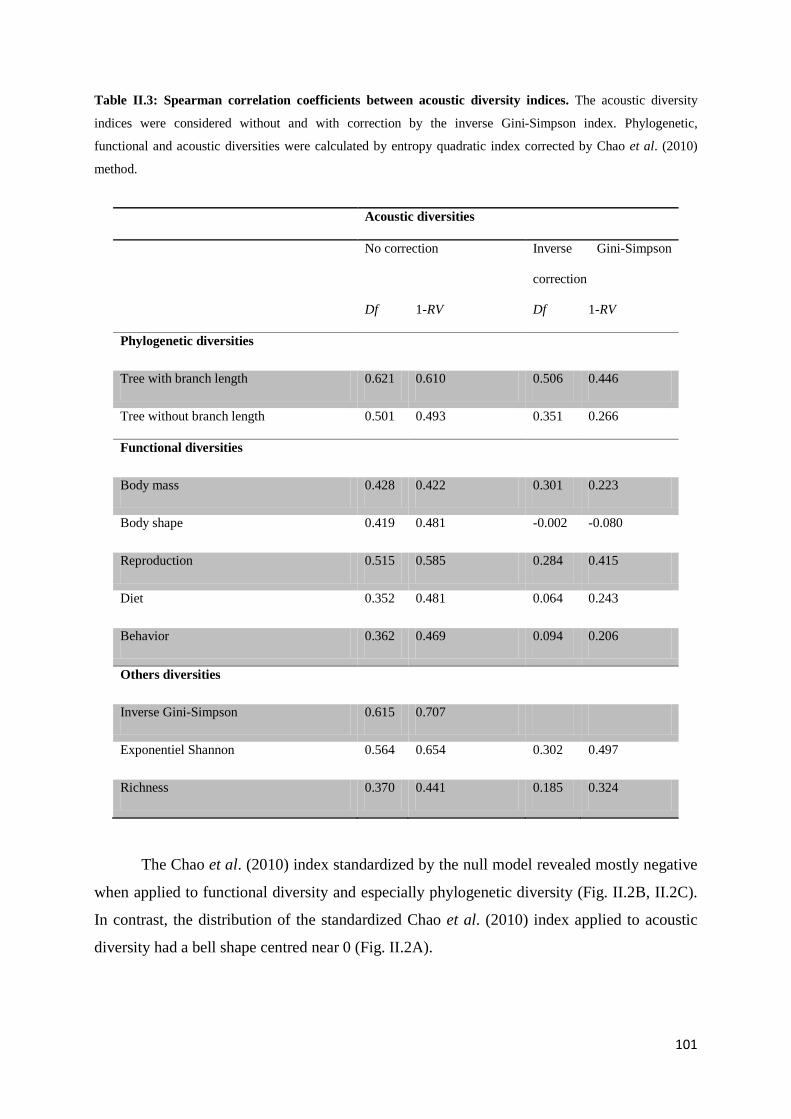
101
Table II.3: Spearman correlation coefficients between acoustic diversity indices. The acoustic diversity
indices were considered without and with correction by the inverse Gini-Simpson index. Phylogenetic,
functional and acoustic diversities were calculated by entropy quadratic index corrected by Chao et al. (2010)
method.
Acoustic diversities
No correction Inverse Gini-Simpson
correction
Df 1-RV Df 1-RV
Phylogenetic diversities
Tree with branch length 0.621 0.610 0.506 0.446
Tree without branch length 0.501 0.493 0.351 0.266
Functional diversities
Body mass 0.428 0.422 0.301 0.223
Body shape 0.419 0.481 -0.002 -0.080
Reproduction 0.515 0.585 0.284 0.415
Diet 0.352 0.481 0.064 0.243
Behavior 0.362 0.469 0.094 0.206
Others diversities
Inverse Gini-Simpson 0.615 0.707
Exponentiel Shannon 0.564 0.654 0.302 0.497
Richness 0.370 0.441 0.185 0.324
The Chao et al. (2010) index standardized by the null model revealed mostly negative
when applied to functional diversity and especially phylogenetic diversity (Fig. II.2B, II.2C).
In contrast, the distribution of the standardized Chao et al. (2010) index applied to acoustic
diversity had a bell shape centred near 0 (Fig. II.2A).
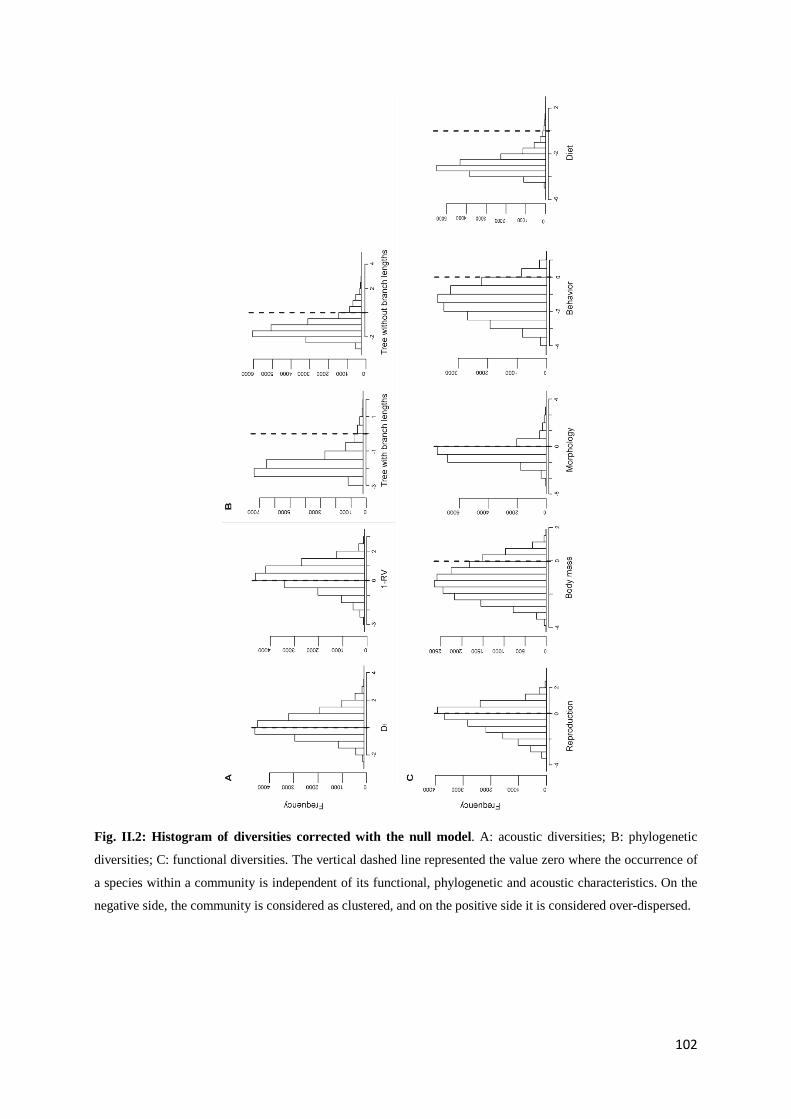
102
Fig. II.2: Histogram of diversities corrected with the null model. A: acoustic diversities; B: phylogenetic
diversities; C: functional diversities. The vertical dashed line represented the value zero where the occurrence of
a species within a community is independent of its functional, phylogenetic and acoustic characteristics. On the
negative side, the community is considered as clustered, and on the positive side it is considered over-dispersed.

103
II.5 Discussion Interest in using acoustic methods as a new tool for the identification of singing species, the
monitoring of species or populations, or the evaluation of global acoustic diversity is rapidly
increasing (Acevedo et al., 2009; Blumstein et al., 2011; Obrist et al., 2010). In particular, the
evaluation of global acoustic diversity (e.g. Sueur et al., 2008b) appears as an innovative
method in conservation biology as it can be deployed over large spatial and temporal scales
and can facilitate the assessment of numerous communities simultaneously. The previously
observed correlations between acoustic diversity and number of species (Depraetere et al.,
2012; Sueur et al., 2008b) reinforce the interest of acoustic diversity studies to evaluate the
biodiversity. Before applying passive acoustic diversity analyses at large scales, there is a
critical need to know whether acoustic aspects reflect phylogenetic (Faith, 1992; Isaac et al.,
2007; Pavoine et al., 2005a; Vane-Wright et al., 1991) and/or functional diversity (Dalerum et
al., 2009) levels. An important database on bird community was therefore used to
theoretically assess the relative parts of phylogenetic and functional diversities embedded in
the measures of acoustic diversity.
To complete the first acoustic distance produced by Sueur et al. (2008b, Df), three
other dissimilarity measurements based on spectral (KS, KL) or time and spectral (1-RV)
profiles were first used. It appears that the mathematical method used to calculate spectral
dissimilarity between two sounds does not significantly impact the result as shown by the
high correlation between Df, KL and KS. We found differences between 1-RV and Df leading
to a better correlation between 1-RV and the others diversities. However, 1-RV is quite more
costly in terms of computational time and might be difficult to obtain when handling large
datasets. In practice, the choice of the index will result in a trade-off between the processing
time and the accuracy of the expected results.
At the species level, the moderate correlations between acoustic distances and body
shape and body mass distances among species indicate that the more different species are in
their body mass and body shape the more different their songs are likely to be. This implies
that species acoustics could depend directly or indirectly via a correlation with a co-factor on
these traits. This could be explained by morphological constraints known to occur on the
frequency range produced by birds. Body weight is negatively correlated to the mean song
frequency in passerine birds (Wallschlager, 1980). This relationship can be extended to

104
several bird taxa (Martin et al., 2011), and even to vertebrates for which the optimal
communication frequency is inversely proportional to body mass (Fletcher, 2004).
High correlations between acoustic and phylogenetic distances based on the tree with branch
length available suggest the occurrence of evolutionary-based mechanisms that have led to a
phylogenetic signal embedded in the songs of extant species. Overall, branch lengths seem to
have a decisive impact on the correlation between phylogenetic and acoustic distances. Given
the assumption that the phylogenetic tree with branch length is clockwise, acoustic behavior
evolution may be associated to clockwise evolution; although this hypothesis should be tested
on larger data sets. Another explanation might be that the data set included a large number of
Passerine birds (n=105). Consequently, the consideration of branch lengths in the calculation
of phylogenetic diversity increased the importance given to the phylogenetic distinctiveness
of the non-passerine species, which also contained several of the most acoustically distinct
species. This greater differentiation among bird orders might be responsible for the increased
detection of phylogenetic signal in acoustic distances among species.
At the community level, the expected correlation between acoustic diversity and richness
(Depraetere et al., 2012; Sueur et al., 2008b) was confirmed when considering the diversities
calculated by Chao et al. index without any corrections. The strong correlation between
acoustic diversity and the inverse Gini-Simpson index was expected as Chao et al. index is
mainly based on inverse Gini-Simpson index. The high correlation between acoustic,
functional and phylogenetic diversity indices suggest that acoustic diversity index, as
calculated here, could be used as an acceptable surrogate of phylogenetic and functional
diversities indices.
As expected, the correlations among different aspects of biodiversity differed with and
without correction by inverse Gini-Simpson index. This difference was due to the important
co-variable role of species’ abundance distribution within all diversity indices computed with
Chao et al. (2010) index. However, the high correlation between tree with branch length and
acoustic diversity was conserved, which indicates that the evolution time information is
included in the acoustic diversity of the bird community. Our results also suggested that
spectral information may be sufficient to reflect phylogenetic diversity even if temporal
variation participates to the diversity and encoding processes of bird songs (Catchpole &
Slater, 1995 for a review). Considering the functional diversities, the corrections also pointed
out a difference between Df and 1-RV. The temporal information supported by 1-RV appears
to have higher links with functional information, especially with diversity in reproduction

105
strategies. This result supports the fact that sexual selection of several bird species is based on
temporal features of male song (Catchpole & Slater, 1995 for a review). It was in particular
demonstrated that male song rate is associated to reproductive success and territory quality
(Alatalo et al., 1990; Greig-Smith, 1982). The diversity of frequencies supported by Df, was
nevertheless also related to diversity in reproduction. A moderate correlation between Df and
1-RV and body mass was maintained after the correction for the co-factor. The spectral
diversity seems thus to better reflect phylogenetic diversity whereas temporal diversity would
better reflect functional diversity.
Acoustic diversity was correlated with species richness, exponential Shannon index
and inverse Gini-Simpson index. When corrected for the inverse Gini-Simpson index, it was
still correlated with the exponential Shannon index and to a lesser extent with species
richness. Inverse Gini-Simpson, exponential Shannon and the richness indices differ in the
importance they give to rare vs. abundant species (Patil & Tallie, 1982). A correlation
between acoustic diversity, exponential Shannon index and species richness, even after a
correction by the inverse Gini-Simpson suggest that less abundant species are important in
explaining the level of acoustic diversity in a community. This might be due to the high
abundance of passerine species in French bird communities and to the acoustic distinctiveness
of less abundant non-passerine species whose occurrence in a community impacts the overall
level of acoustic diversity despite their lower abundance.
The correlations found between different aspects of distances among species
(functional, phylogenetic vs. acoustic distances) were different from the correlations found
between different aspects of diversity within communities. Several researches focused on the
environment filtering and the competitive interaction as two contrasting processes underlying
community assembly, as reviewed in Emerson and Gillespie (2008). Our results highlight that
the community structure, in term of how species assemble within communities, is likely to
have an influence on the diversity of the community. Bird communities are not just random
assemblages of species but depend on ecological processes. According to the FBBS protocol,
each local community was investigated within a homogeneous habitat. Environmental filters
within each habitat are therefore likely to filter species based on their functional traits
(Lebrija-Trejos et al., 2010).
In our case study, phylogenetic and functional clustering was indeed found in local
communities, which means that co-occurring species tend to be closely related and to share
similar traits. In other words, communities tend to have closely related species with similar
traits and similar phylogenetic history. Phylogenetic and functional clustering might simply

106
be explained by limits in the geographic distribution range of related species with similar
traits (such as Devictor et al., 2010) and also by local ecological processes, especially habitat
filtering. Species found in the same habitat type are expected to share functional traits
(Cornwell et al., 2006) and phylogenetic positions (Emerson & Gillespie, 2008). However, in
contrast to the observed phylogenetic and functional clustering, bird communities
continuously ranged from acoustically clustered, through random to over-dispersed patterns.
Within a homogeneous habitat, acoustic overdispersion is expected because specificity
ensures an efficient communication between congeners (Chek et al., 2003). However,
acoustic clustering is expected if the pressure of habitat type leads to acoustic adaptation of
the species. In that case acoustic features are dependent on particular habitats (Boncoraglio &
Saino, 2007). For instance, closed habitats have greater reflections than tree trunks and leaves
(Slabbekoorn, 2004; Wiley & Richards, 1978). These opposed processes could lead to
different community acoustic assemblies, from clustered to over-dispersed. Even if the
acoustic diversity seems to be positively correlated with phylogenetic and body mass
diversities, the processes that structure the acoustic diversity are thus expected to be partly
different than those that structure phylogenetic and functional diversity.
In conclusion, a means of estimating a finer definition of acoustic diversity at the community
level was suggested. The Community Acoustic Diversity (CAD) contains all the variability
that can be extracted out of a sound produced by an animal community. Traditionally, two
components of the sound, frequency and time structure are used to describe this variability.
The number of species (Depraetere et al., 2012; Sueur et al, 2008b) and the number of signals
(Farina et al., 2011; Pieretti et al., 2011) can potentially be assessed by the CAD. The present
work showed that, in bird communities, the CAD contains a phylogenetic signal with
evolution time information. Still in regard to the data used here, the CAD contains also a
functional signal essentially linked to body mass and reproduction and more slightly to diet
and behavior. This preliminary work is theoretical and it raised new hypotheses that now have
to be tested on real community songs. However, it supports the idea that biodiversity
assessment through acoustics could then provide valuable information on different
biodiversity facets. The CAD could be used on real animal community acoustic signal
through data collected by automatic devices settled in the fields. To applied the CAD index on
the field, this theoretical work needs to be complete by tests on real acoustic community
signal.

107
II.6 Acknowledgements
This work was supported by the CNRS INEE PhD grant and the FRB BIOSOUND program.
The authors would like to thank Pierre-Yves Henry, Marsha Schlee, Grégoire Lois, Philippe
Clergeau, Camila Andrade for their help when we collecting bird functional traits. We greatly
thank the hundreds of volunteers who have been taking part in the national breeding bird
survey (FBBS program). We thank Ryan Calsbeek for improving the language of the
manuscript and the two anonymous referees for their helpful comments on the manuscript.

108

109
Chapitre 3
Echantillonnage de la biodiversité par une
approche globale d'acoustique passive :
différences entre sites abritant des
microendemics de Nouvelle-Calédonie
PLoS ONE, en révision.
Communication :
Gasc, A., S. Pavoine, J. Sueur, P. Grandcolas (2010) Biodiversity sampling using a meta-
acoustic method: micro-endemsim detection in New-Caledonia. Ecologie 2010 congress,
poster.

110

111
Biodiversity sampling using a global acoustic approach: contrasting sites with
microendemics in New Caledonia
Authors: Gasc A.1*, J. Sueur1, S. Pavoine2, 3, R. Pellens1 and P. Grandcolas1.
1Muséum national d'Histoire naturelle, Département Systématique et Évolution, UMR 7205-
CNRS-OSEB, 45 rue Buffon, Paris, France
2Muséum national d’Histoire naturelle, Département Ecologie et Gestion de la Biodiversité,
UMR 7204-CNRS-UPMC, CP 51, 55-61 rue Buffon, 75005 Paris, France
3Mathematical Ecology Research Group, Department of Zoology, University of Oxford,
South Parks Road, Oxford, OX1 3PS, UK
*Author for correspondence
Amandine GASC
Muséum national d’Histoire naturelle
Département Systématique et Évolution
UMR CNRS 7205 OSEB
CP 50, 45 rue Buffon
F-75005 Paris, France
Tel. + 33 1 40 79 31 34
Fax. + 33 1 40 79 56 79
Email. [email protected]

112
III.1 Abstract/Résumé
III.1.1 Abstract
New Caledonia is a Pacific island with a high conservation value due to its unique
biodiversity and micro-endemism. The microendemic distributions observed on this island are
extremely restricted, localized to mountains or rivers. Evaluation of biodiversity is so a hard
task. Could global analyse of acoustic diversity reveal this unique spatial structure? Three
sites (Mandjélia, Koghis and Aoupinié mounts) were selected because of their ecological and
alpha diversity similarity, but high beta diversity, with different microendemic assemblages at
each site. Local acoustic communities were sampled with audio recorders placed at each site.
Acoustic activity of animal communities was analysed without any species identification. A
spectral complexity index (NP) was used as an estimate of the level of acoustic activity and a
spectral dissimilarity index (Df) assessed differences between pairs of recordings. As
expected, the results suggested significant differences in acoustic activity between sites.
Acoustic variability could first be explained by circadian variation and second by the
difference between the three sites. The results support the hypothesis that global acoustic
analyses can detect acoustic differences between ecologically similar sites with different
species assemblages. This study demonstrates that global acoustic methods applied at broad
spatial and temporal scales could help for local biodiversity assessment. The method could be
deployed over large areas, and could help compare different sites and determine conservation
priorities.
Keywords: Biodiversity, broad spatial scale, dissimilarity, microendemism, New Caledonia,
passive acoustics, remote sensing.
III.1.2 Résumé
La Nouvelle-Calédonie est une île du Pacifique qui possède une biodiversité unique associée à
un taux élevé de micro-endémisme. Les distributions des espèces micro-endémiques sont très
localisées, restreintes à une montagne ou une rivière rendant le travail d'évaluation de la
biodiversité très difficile. Une analyse acoustique globale permet-elle de révéler cette
structure spatiale unique ? Trois sites (les monts Mandjélia, Koghis et Aoupinié) ont été

113
sélectionnés. Ces trois sites présentent des similarités écologiques et des diversité α proches,
mais également des diversités β différentes. Celles -ci sont notamment associées à des
compositions d'espèces micro-endémiques qui varient d'un site à l'autre. Les communautés
acoustiques locales ont été échantillonnées à l'aide d'enregistreurs automatiques placés sur
chaque site. L'activité acoustique des communautés animales a été analysée sans identification
des espèces chanteuses à l'aide d'un indice de complexité spectrale (NP), utilisé pour estimer
le niveau d'activité acoustique d'un enregistrement, et un indice de dissimilarité spectrale (Df),
utilisé pour mesurer les différences entre les enregistrements deux à deux. Comme attendu, les
résultats des analyses suggèrent que des différences acoustiques significatives existent entre
les sites. La variabilité acoustique contenue dans les données pourrait d'abord s'expliquer par
des variations acoustiques entre les heures le long d'un cycle circadien, puis, par des
variations acoustique entre les trois sites. Les résultats supportent l'hypothèse selon laquelle
l'analyse acoustique globale permet de détecter des différences entre des sites abritant des
communautés d'un niveau de biodiversité similaire mais dont les assemblages spécifiques
diffèrent. Cette étude suggère que, appliquée à une large échelle temporelle et spatiale, la
méthode acoustique globale pourrait être utile pour comparer la biodiversité de différents sites
et pour aider à déterminer des priorités de conservation.

114
III.2 Introduction
New Caledonia has been classified as one of the 25 most important hotspots of biodiversity
conservation regarding the number of endemic species and degree of threat (Myers et al.,
2000). Kier et al. (2009) also ranked New Caledonia as the terrestrial region with the highest
level of endemism. As an example, 19% of bird, 67% of mammal, 86% of reptile and 76% of
plant species are endemic to this Pacific island at the regional scale (Myers et al., 2000). The
emphasis put on regional endemism and biogeography hides another remarkable feature of
New Caledonia, the extremely high level of local endemism, hereafter called microendemism.
The so-called microendemic species of plants, lizards or insects show a very short
distributional range, limited to small mountains or rivers of New Caledonia. According to
recent studies, this local endemism mainly originated through recent allopatric speciation with
little ecological differentiation (Grandcolas et al., 2008), with the notable exception of
adaptation to metalliferous soils derived from ultramafic rocks covering one third of the island
(Pillon et al., 2010). In this paradigm, the island can be considered as a wonderful natural
laboratory for evolution, where many questions can be studied regarding speciation and
endemism through a large time window of 37 million years (e.g., Espeland & Johanson, 2010;
Grandcolas et al., 2008; Murienne et al., 2005, 2008, 2009).
Microendemism also bears strong consequences for conservation since very short
distributional ranges increase the risk of species extinction, facing three principal threats that
are nickel mining, fire and invasive species (reviewed in Pellens & Grandcolas, 2010).
The origin and composition of this threatened biodiversity have to be quickly described and
deciphered, both to allow theoretical studies of speciation and evolution to be conducted and
to permit the establishment of sound conservation policies. It is therefore necessary to rapidly
characterize sites and their communities. However, such a task is made complex by the fact
that sites differ not only by distinct species combinations within the same local pool, but also
by the fact that each mountain or river usually shows local endemics formed by allopatric
speciation (e.g., Grandcolas, 2009). Inventorying in New Caledonia has proved to be a
fastidious and slow process due to a complex landscape, a small local scientific community,
and long distance isolation from most academic centers.
Therefore, the inventorying process needs to be enhanced and sped up by using other possible
approaches. In this respect, inventories based on passive acoustic methods have demonstrated

115
convincing advantages as they are non invasive, allow large automatic sampling, can be
simultaneously used on several taxa, and provide very large temporal and spatial data sets
(Acevedo & Villanueva-Rivera, 2006; Wimmer et al., 2012). Similar to classical inventory
methods, passive acoustics can potentially be used to identify species directly by ear (Dawson
& Efford, 2009; Riede, 1993) or by automatic identification methods (Acevedo et al., 2009;
Brandes, 2008, Hans et al., 2011). However, identification of species by human observers
have its limits (i.e. discrepancies among observers, difficulty to handle large samples sizes).
Complex classification methods are used to automatic recognition (e.g., Towsey et al., 2012).
These processes characterized species songs by a set of signal parameters that require an
important sound reference database that need a critical preliminary description of species’
songs. Such an approach seems very complex to undertake in New Caledonia where species
richness is particularly high, and where species acoustic diversity has not been fully registered
and described yet. Faced with this difficulty a global acoustic passive method might be
helpful and provide new data on local animal diversity (Sueur et al., 2008a).
The global acoustic method consists in analysing the acoustic output of animal communities
by measuring how diverse or complex a community signal is in terms of spectral and/or
temporal acoustic properties, without conducting species identification. Such measures of
acoustic complexity have been linked previously to the number of singing species (Depraetere
et al., 2012; Sueur et al., 2008a) and to the number of vocalizations (Pieretti et al., 2011).
Following Diamond and Case (Diamonds & Case, 1986), who defined a community as an
entity that "comprises the populations of some or all species coexisting at a site or in a
region", an acoustic community could be seen as a collection of sounds produced by all living
organisms in a given habitat over a specified time, considering the competitive interaction for
the sound space, structured by the sound resource. Two complementary approaches are
currently developed, one to evaluate the acoustic diversity level of each community based on
a measure of acoustic signal complexity (α diversity), and one to compute acoustic distances
between two communities (β diversity) (Sueur et al., 2008a).
In addition, the level of species richness in New Caledonia is high throughout the island.
Compare species richness of sites is not likely to be very informative (Pellens & Grandcolas,
2010). Establishing complementarities of different sites in terms of species composition
should help in making conservation priorities for a New Caledonia diversity evaluation and
conservation (Faith & Walker, 1986; Margules & Pressey, 2000).

116
The aim of this study is to evaluate the capacity of the global acoustic method to detect
dissimilarity between sites that are otherwise known to be similar in species richness and
ecologically similar but be distinctive in species composition and microendemics. Automatic
recorders were used to collect acoustic data on the three sites. The computation of distance
based acoustic indices and statistical analysis were used to compare the acoustic
dissimilarities among three different sites. The three sites located on the main island possess a
high level of microendemism that guaranteed differences in species assemblages, according to
a census based on all published phylogeographic studies to the present day (see Table S1 for
references, supplementary information available in "Annexe 3"). Global Biodiversity
Inventory Facility (GBIF) records from collection databases have also been used to evaluate
the biodiversity knowledge available for these three sites.
The study can be seen as a very stringent test of the capacity of the global acoustic method to
detect dissimilarity of biodiversity, given that the importance of allopatric speciation with
niche conservatism in New Caledonia is expected to maximise ecological similarity between
sites. The global acoustic method could be an effective new approach to contrast sites.
III.3 Material and methods
III.3.1 Study sites
The three sites, Mandjélia (20°24,098'S, 164°31,458'E), Aoupinié (21°10,034'S,
165°19,017'E) and Koghis (22°10.833'S, 166°30.632'E) mounts, were chosen as reference
sites for their distinct species occurrence and specific microendemism (Fig. III.1).
All sites are otherwise comparable in climate (tropical humid), soil (non metalliferous
soils) and vegetation (tropical humid forest). Within each site, three recorders were placed
with a distance of 200 meters between each of them. All recorders were placed at a similar
altitude of 600 m (+/- 84 m).

117
Fig. III.1: Sampled sites in the great island of New Caledonia. There were three audio recorders at each site
settled at a similar altitude of 600 m a.s.l. and separated by 200 m.
The Global Biodiversity Information Facility (GBIF, 2012) provided a dataset of all
specimens collected on a square of 80 km2 around the three sites, belonging to both animal
and vegetal taxa. The number of species was dependent on the collection effort, which was
not comparable from one site to another. In order to correct this bias, the number of species
for each site was divided by the number of collectors. Other potential hidden bias, such as
different sampling efforts and techniques could not be controlled.
III.3.2 Passive acoustic recording
The recordings were made from April 10th to June 30th 2010, with Song Meter SM2 digital
audio field recorders (Wildlife Acoustics, 2009). These offline and weatherproof recorders
were equipped with a single omnidirectional microphone (frequency response: -35±4dB
between 20 Hz and 20 kHz) oriented horizontally at a height of 1.5 m. The signals were
digitized at 44.1 kHz and 16 bits digitization. The files were saved in the lossless compressed
format .wac and then transformed into the format .wav with the software WAC to WAV
Converter Utility version 1.1 (Wildlife Acoustics, 2009). One minute was recorded every hour
leading to a total of 13,602 sound files.
All recordings were examined by A.G. to remove biases due to the occurrence of rain,
wind or anthropogenic noise. This resulted in a selection of 6,571 files (53% for Aoupinié,
48% for Mandjélia and 49% for Koghis, see Table S2, supplementary information available in
"Annexe 3").

118
No specific permits were required for the described field studies because passive acoustic
method was used in a location that was not privately-owned or protected in any way. This also
implies that the study did not involve endangered or protected species. The acoustic
recordings are deposited in the sound library of the Muséum national d'Histoire naturelle
(Paris, France).
III.3.3 Acoustic activity level
A level of acoustic activity was assessed by ear by (A.G.) for every minute of recording Three
levels were used: (1) "Moderate to high activity" when more than two different song types
were identified, (2) "Low activity" when one or two different song types occurred, and (3)
"Null activity" when no animal songs could be detected (see Table S3, supplementary
information available in "Annexe 3"). This scale was designed to fulfil aural discrimination
constraints on a very large data set (6 571 files). In particular, it was not possible to clearly
distinguish more than two song types in complex recordings. This characterisation of acoustic
activity level was also built for an easy use by potential other observers. The acoustic activity
detected was mainly due to bird, Orthoptera and cicada species, but no species identification
was achieved.
III.3.4 Background noise reduction
Considering that differences between two recordings could be solely due to different
background noises, a filtering process was applied to all files to remove noise as much as
possible. For each site, all recordings with null activity were selected and averaged to obtain
the mean spectrum of ambient noise that was computed using a short term Fourier transform
(STFT) based on a non overlapping sliding function window of 512 samples (= 11ms). The
mean spectrum of each file was subtracted by the mean spectrum of ambient noise of the local
site. This subtraction was weighted by the amplitude level of each recording as follows:
( ) ( ) ( )c i iS f M S f M Sn f= × −
where Sc(f) is the probability mass function of the mean spectrum corrected, S(f) is the
probability mass function of the original mean spectrum, Sni(f) is the probability mass
function of the mean spectrum of the ambient noise of the site i, a the amplitude level of the

119
recording and bi the mean of amplitude levels of files containing ambient noise for the site i.
The amplitude level parameters M and iM were obtained following:
( ) (1 )( ) 2 depthM median A t −= × with 0 < M < 1,
where A(t) is the amplitude envelope and depth is signal digitization depth (here, 16 bits).
Only the files with "Moderate to high activity" and "Low activity" were kept for subsequent
analyses.
III.3.5 Acoustic complexity
A new index was developed to assess acoustic complexity. This index, named NP, counts the
number of major frequency peaks obtained on a mean spectrum scaled between 0 and 1. The
mean spectrum was obtained with a STFT (non overlapping window size = 512 samples = 11
ms).
A program was developed within the R environment (R Development Core Team,
2010) in the ‘seewave’ package (Sueur et al., 2008b) to allow automatic spectral peaks
detection. All peaks of the spectrum were first detected and then selected using amplitude and
frequency thresholds. The first selection factor was based on the amplitude slopes of each
peak. Only peaks with slopes higher than 0.01 were kept. The second selection factor was
based on frequency. If consecutive peaks were less than 200 Hz apart then, only the highest
peak in amplitude was kept. NP was simply defined as the number of peaks after detection
and selection of peaks. In order to evaluate the reliability of NP in revealing the acoustic
activity detected by ear, the distribution of NP data measured on a spectrum before ambient
noise reduction was visualized by boxplot depending on the factor of activity level (high,
medium or null activity) and then, a pairwise Mann-Whitney test was applied with a Holm
correction to test differences between the pairwise groups of activity level. The same analysis
(boxplot on values following by Mann-Whitney test with a Holm correction) was carried out
to test the differences between NP values measured on spectra without ambient noise among
sites.
III.3.6 Spectral global differences
Acoustic spectral dissimilarity Df, as defined by Sueur et al. (2008a), has been shown to be
informative, embedding compositional diversity information. Df was computed according to:

120
0.5 1( ) 2( )fD sum S f S f= × −
where S1(f) and S2(f) are the probability mass functions of the mean spectra of the two
recordings to be compared. Each function was the result of a STFT with a non overlapping
window of 512 samples (= 11ms). Df was computed on mean spectra after ambient noise
reduction.
Df was assessed between all pairs of recording files leading to a first dissimilarity
matrix. Each recording was associated with a recording time, a day, a recorder, and a site. To
factor out the differences among days which could reflect meteorological variations, the
differences between each recording was averaged over all available days. This second
dissimilarity matrix with dimension 216*216 (= (3 sites * 3 recorders * 24 time periods ) * (3
sites * 3 recorders * 24 time periods)) was transformed by the Lingoes approach to obtain
Euclidean properties (Lingoes, 1971). Then, the second dissimilarity matrix was analyzed
with a Principal Coordinate Analysis (PCoA) (Gower, 1966) in order to visualize whether the
factors "Site" and "Time" could explain the acoustic variability, and which factor is the most
important in explaining the acoustic differences observed. Each point projected in the
resulting multidimensional space represents the acoustic position of one site, one recorder, at
one hour. The ability of the "Site" and "Time" factors to explain the acoustic differences was
evaluated using a distance-based ReDundancy Analysis (dbRDA) (Legendre & Anderson,
1999). Associated to the dbRDA, a permutation test was applied with 1000 permutations
considering each factor independently, which means that to test the factor "Site" the
permutations were constrained by the factor "Time" and vice-versa.
A last step was made to validate the PCoA and dbRDA results considering the factor
“Site”. To be sure that these results could not be biased by an unbalanced number of
recordings per hour for each site, due to recordings being discarded because of bad weather
conditions, the analyses were done a second time on a balanced subsample of hours from the
averaged dissimilarity matrix. Six hours with balanced samples were selected: 1, 3, 4, 18, 20
and 21h. The dimension of this submatrix was 54*54 (= (3 sites * 3 recorders * 6 time
periods) * (3 sites * 3 recorders * 6 time periods)). This submatrix was analysed with a
dbRDA with site as a factor. The results were compared with the 216*216 matrix with all
hours included. In order to quantify these differences between each pair of sites, the Euclidean
distances of the barycentre of the points associated to the three levels of the factor "Site" were
calculated from differences of the 216*216 matrix (Rao’s disc coefficient of dissimilarity
between sites; Rao, 1982; Pavoine et al., 2004).

121
To validate the noise reduction step, the dbRDA analyses were applied to the original
data before noise reduction.
III.3.7 Spectral differences detailed
The dbRDA can highlight acoustic differences between sites due to the factor "Site", but
cannot provide details regarding these differences. To identify these differences, an analysis
was developed to identify at which frequencies the differences occur when computing Df. As
already described, the mean spectrum was composed of 256 frequency bins and the Df index
was the sum of these 256 differences between two mean spectra. Here, the 256 frequency bin
differences calculated were not summed but analysed independently. Instead of a single Df
matrix, 256 matrices were analysed, one for each frequency. For each frequency bin, a
between sites distance value was computed with Rao's disc coefficient from the 256 matrices.
The index Df computes a simple difference between each frequency bin of the two spectra to
be compared. The index Df will then be greater than 0 when comparing two spectra with the
similar profiles but slightly shifted in amplitude. Such a difference is then not informative in
terms of frequency content. To check for such difference, the relative importance of each
frequency bin was assessed by estimating the distribution of the frequency peaks detected
when computing the index NP. A peak detected several times would then indicate that the
corresponding peak is relevant in the difference identified by Df . The distribution of the
frequency peaks and Df results were compared on a single plot.
All analyses were performed with the R software (R Development Core Team, 2010)
including the packages ‘seewave’ (Sueur et al., 2008b) and ‘ade4’ (Dray & Dufour, 2004).
III.4 Results
III.4.1 Classical inventories
From the 17 phylogenetic datasets analysed, the percentage of microendemics calculated was
12.50% for Mandjélia, 15.38% for Aoupinié and 23.73% for Koghis. From the GBIF data
(GBIF, 2012), the ratio of endemic species to the number of collectors was 4.89 for
Mandjélia, 3.76 for Aoupinié and 4.71 for Koghis.
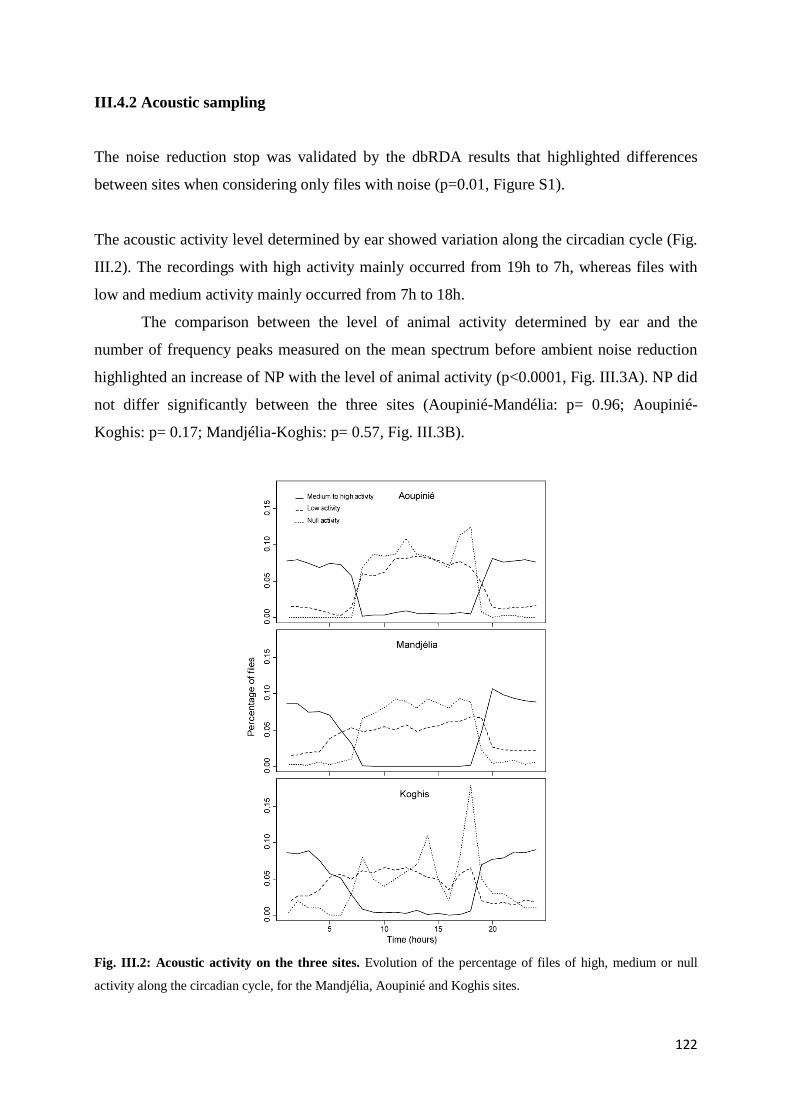
122
III.4.2 Acoustic sampling
The noise reduction stop was validated by the dbRDA results that highlighted differences
between sites when considering only files with noise (p=0.01, Figure S1).
The acoustic activity level determined by ear showed variation along the circadian cycle (Fig.
III.2). The recordings with high activity mainly occurred from 19h to 7h, whereas files with
low and medium activity mainly occurred from 7h to 18h.
The comparison between the level of animal activity determined by ear and the
number of frequency peaks measured on the mean spectrum before ambient noise reduction
highlighted an increase of NP with the level of animal activity (p<0.0001, Fig. III.3A). NP did
not differ significantly between the three sites (Aoupinié-Mandélia: p= 0.96; Aoupinié-
Koghis: p= 0.17; Mandjélia-Koghis: p= 0.57, Fig. III.3B).
Fig. III.2: Acoustic activity on the three sites. Evolution of the percentage of files of high, medium or null
activity along the circadian cycle, for the Mandjélia, Aoupinié and Koghis sites.
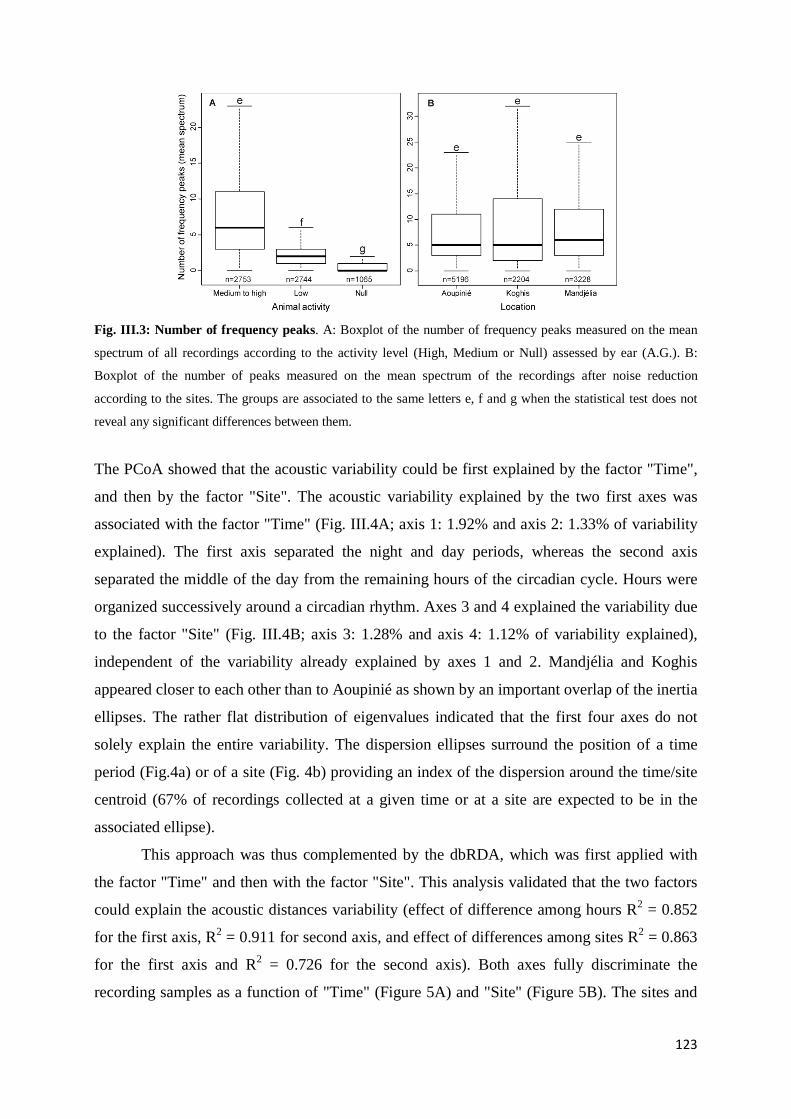
123
Fig. III.3: Number of frequency peaks. A: Boxplot of the number of frequency peaks measured on the mean
spectrum of all recordings according to the activity level (High, Medium or Null) assessed by ear (A.G.). B:
Boxplot of the number of peaks measured on the mean spectrum of the recordings after noise reduction
according to the sites. The groups are associated to the same letters e, f and g when the statistical test does not
reveal any significant differences between them.
The PCoA showed that the acoustic variability could be first explained by the factor "Time",
and then by the factor "Site". The acoustic variability explained by the two first axes was
associated with the factor "Time" (Fig. III.4A; axis 1: 1.92% and axis 2: 1.33% of variability
explained). The first axis separated the night and day periods, whereas the second axis
separated the middle of the day from the remaining hours of the circadian cycle. Hours were
organized successively around a circadian rhythm. Axes 3 and 4 explained the variability due
to the factor "Site" (Fig. III.4B; axis 3: 1.28% and axis 4: 1.12% of variability explained),
independent of the variability already explained by axes 1 and 2. Mandjélia and Koghis
appeared closer to each other than to Aoupinié as shown by an important overlap of the inertia
ellipses. The rather flat distribution of eigenvalues indicated that the first four axes do not
solely explain the entire variability. The dispersion ellipses surround the position of a time
period (Fig.4a) or of a site (Fig. 4b) providing an index of the dispersion around the time/site
centroid (67% of recordings collected at a given time or at a site are expected to be in the
associated ellipse).
This approach was thus complemented by the dbRDA, which was first applied with
the factor "Time" and then with the factor "Site". This analysis validated that the two factors
could explain the acoustic distances variability (effect of difference among hours R2 = 0.852
for the first axis, R2 = 0.911 for second axis, and effect of differences among sites R2 = 0.863
for the first axis and R2 = 0.726 for the second axis). Both axes fully discriminate the
recording samples as a function of "Time" (Figure 5A) and "Site" (Figure 5B). The sites and
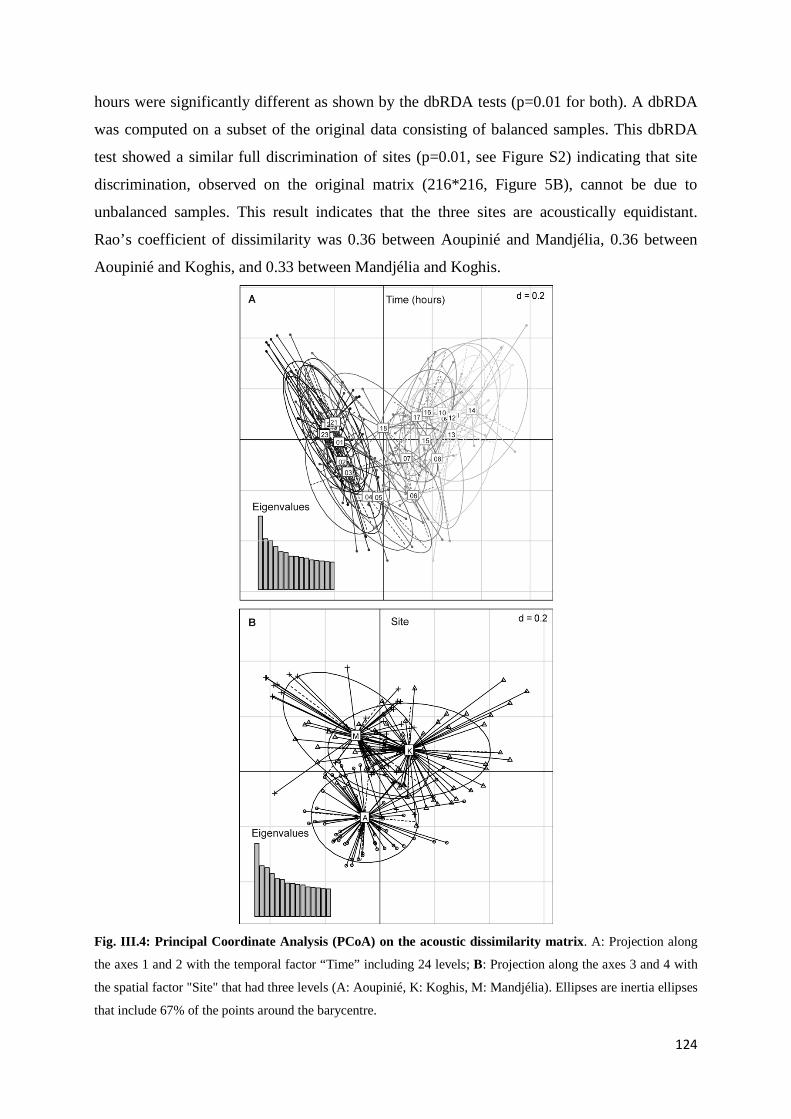
124
hours were significantly different as shown by the dbRDA tests (p=0.01 for both). A dbRDA
was computed on a subset of the original data consisting of balanced samples. This dbRDA
test showed a similar full discrimination of sites (p=0.01, see Figure S2) indicating that site
discrimination, observed on the original matrix (216*216, Figure 5B), cannot be due to
unbalanced samples. This result indicates that the three sites are acoustically equidistant.
Rao’s coefficient of dissimilarity was 0.36 between Aoupinié and Mandjélia, 0.36 between
Aoupinié and Koghis, and 0.33 between Mandjélia and Koghis.
Fig. III.4: Principal Coordinate Analysis (PCoA) on the acoustic dissimilarity matrix. A: Projection along
the axes 1 and 2 with the temporal factor “Time” including 24 levels; B: Projection along the axes 3 and 4 with
the spatial factor "Site" that had three levels (A: Aoupinié, K: Koghis, M: Mandjélia). Ellipses are inertia ellipses
that include 67% of the points around the barycentre.
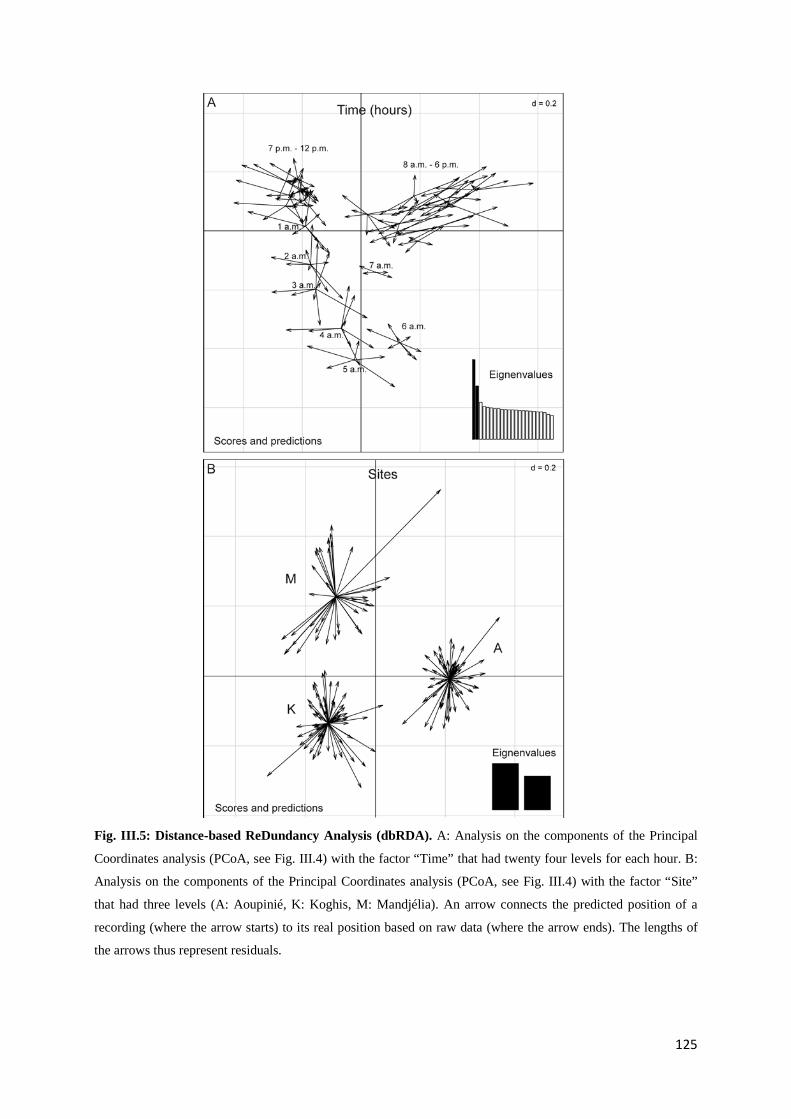
125
Fig. III.5: Distance-based ReDundancy Analysis (dbRDA). A: Analysis on the components of the Principal
Coordinates analysis (PCoA, see Fig. III.4) with the factor “Time” that had twenty four levels for each hour. B:
Analysis on the components of the Principal Coordinates analysis (PCoA, see Fig. III.4) with the factor “Site”
that had three levels (A: Aoupinié, K: Koghis, M: Mandjélia). An arrow connects the predicted position of a
recording (where the arrow starts) to its real position based on raw data (where the arrow ends). The lengths of
the arrows thus represent residuals.
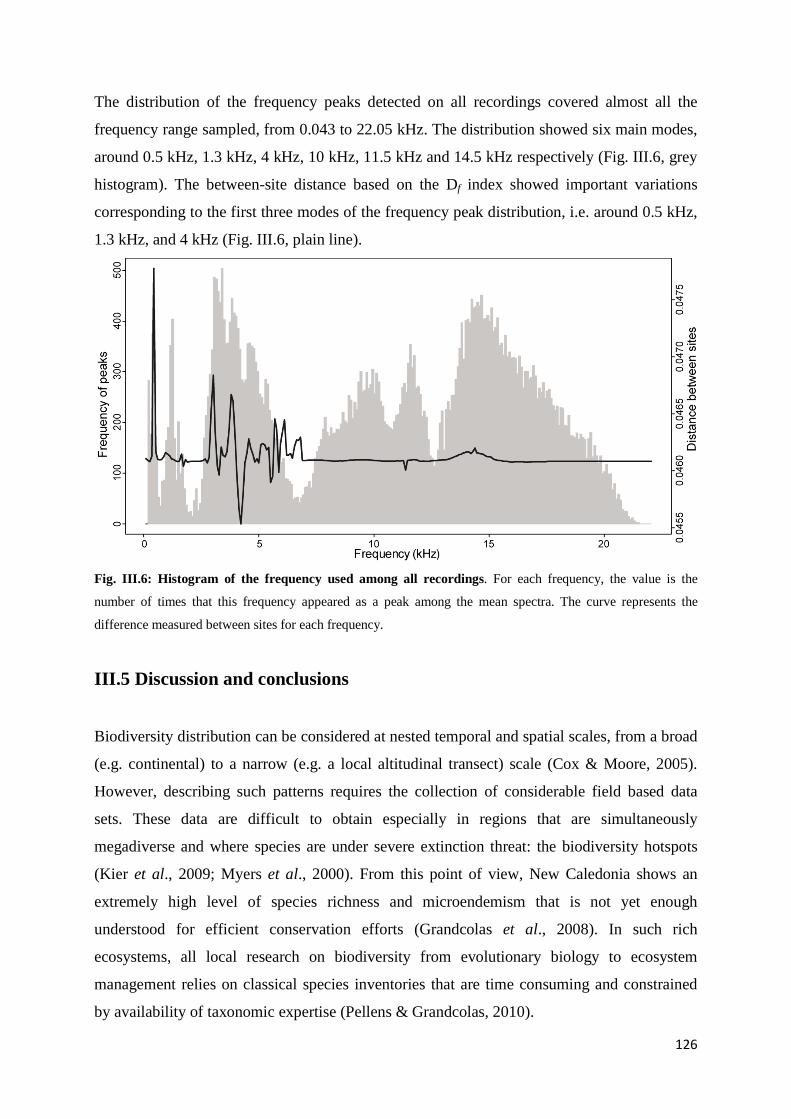
126
The distribution of the frequency peaks detected on all recordings covered almost all the
frequency range sampled, from 0.043 to 22.05 kHz. The distribution showed six main modes,
around 0.5 kHz, 1.3 kHz, 4 kHz, 10 kHz, 11.5 kHz and 14.5 kHz respectively (Fig. III.6, grey
histogram). The between-site distance based on the Df index showed important variations
corresponding to the first three modes of the frequency peak distribution, i.e. around 0.5 kHz,
1.3 kHz, and 4 kHz (Fig. III.6, plain line).
Fig. III.6: Histogram of the frequency used among all recordings. For each frequency, the value is the
number of times that this frequency appeared as a peak among the mean spectra. The curve represents the
difference measured between sites for each frequency.
III.5 Discussion and conclusions
Biodiversity distribution can be considered at nested temporal and spatial scales, from a broad
(e.g. continental) to a narrow (e.g. a local altitudinal transect) scale (Cox & Moore, 2005).
However, describing such patterns requires the collection of considerable field based data
sets. These data are difficult to obtain especially in regions that are simultaneously
megadiverse and where species are under severe extinction threat: the biodiversity hotspots
(Kier et al., 2009; Myers et al., 2000). From this point of view, New Caledonia shows an
extremely high level of species richness and microendemism that is not yet enough
understood for efficient conservation efforts (Grandcolas et al., 2008). In such rich
ecosystems, all local research on biodiversity from evolutionary biology to ecosystem
management relies on classical species inventories that are time consuming and constrained
by availability of taxonomic expertise (Pellens & Grandcolas, 2010).

127
Recent ecological sensing methods should help in drawing such general diversity patterns by
relaxing the constraint of taxonomic inventorying. In particular, non-invasive global acoustic
approaches may be considered as an alternative or a complementary approach to classical
biodiversity surveys (Sueur et al, 2008a, 2012). Comparing the sound properties of animal
assemblages through the computation of acoustic dissimilarities may help in pointing out
differences between sites that have distinct species composition, at least partly related to
different evolutionary histories and microendemism. A preliminary test for the efficiency of
such an approach was run on three major diversity sites in New Caledonia. Global acoustic
analyse discriminated sites that are known for their high and comparable level of species
richness, according to records in biodiversity meta-databases GBIF (2012), and their high
level of microendemism, according to all reported phylogeographic studies.
Acoustics is an emerging route for broad scale diversity estimation and as such has produced
promising indices: a temporal and spectral entropy index (index H, Sueur et al., 2008a), a
ranked index combining signal amplitude and temporal entropy (index AR, Depraetere et al.,
2012), and a spectrogram complexity index (index ACI, Pieretti et al., 2011). The H index is
easy to compute, but might be biased by unwanted background noise, the AR index probably
misses important spectral information by considering only amplitude and time scales, and the
ACI is not sensitive to constant acoustic signals that may occur in tropical environments
where cicadas (Boulard, 2006) produce sustained sounds. A research effort is therefore
needed to compare and improve these indices. A new index, NP, based on the spectral
properties was developed here as a simple measure of spectral complexity. NP is less sensitive
to ambient noise and seems to be linked to animal sound activity. This index behaved as
expected when applied to the three sites as it returned comparable values for these sites,
which are known to have comparable species richness level. Such similar results in terms of
peak richness indicate that subsequent dissimilarity analyses through acoustic distances could
be interpreted in terms of differences in acoustic spectral profile, but not as a simple scale
effect related to acoustic activity level.
Considering the temporal scale of acoustic activity on the three sites, acoustic activity level
profiles along day time and multivariate analyses based on pairwise spectral distances clearly
showed that acoustic variability among sites could be first explained by a circadian pattern.
The distribution of the recordings within multivariate space (PCoA, dbRDA) suggests a
structured succession of different acoustic communities. Such temporal organization is well

128
known in both temperate and tropical environments with clear temporal windows occupied by
specific groups (Mann et al., 2009; Riede, 1997), and the best-studied periods being the dawn
and dusk choruses of birds (Henwood & Fabrick, 1979). Such temporal separation could be
the outcome of an acoustic inter-specific interference (Ficken et al., 1985) or the result of
different predator pressures according to the time of day (Heller & Von Helversen, 1993). As
no attempt was made here to identify the singing species, the details of the groups following
along the day and night cycle could not be estimated, but it was obvious that night soundscape
was dominated by insects, mainly Orthoptera, whereas the day soundscape was essentially
due to birds and insects such as cicadas. Even if the acoustic level was higher during the
night, sampling should not be limited to this period as information during low activity periods
could also be significant and provide information regarding dissimilarity between sites.
Temporal variability was so high that it overcame the geographical variability that was
first targeted. However, the three sites undoubtedly appeared distinct when projecting the
samples on the subsequent axes of the multivariate analysis (dbRDA results), which are
independent of the axes displaying temporal tendencies, and when running the distance based
redundancy analysis. In particular the Rapid global acoustic surveying seems to be sensitive
enough to highlight differences among three sites with similar habitats. These differences
were not due to residual background noise or unbalanced time sampling but were associated
with animal community activity. Besides, similar NP values for each site ensured that
differences were not due to different activity levels, but to acoustic composition. The
accompanying data reporting the presence/absence of species in the three sites (phylogenetic
data and GBIF) were not the result of standardized sampling protocols, but still support the
fact that there are important differences between the species assemblages, a strong part of
which is related to allopatric speciation between mountains and the establishment of
subsequent microendemism. The global acoustic approach returns a single distance value, but
does not provide accurate information on the acoustic differences. To understand the patterns
observed, it is important to identify which part of the spectrum differs among sites. An
original analysis comparing the frequency peaks detected and the distance values between
sites for each frequency revealed that most of the acoustic differences were occurring over a
sharp frequency band below 7 kHz. Are there any endemics known to produce sound in this
frequency window? The lack of an inventory identifying calling species unfortunately limits
the interpretation regarding these acoustic differences. Among microendemic species listed in
phylogeographic studies (see Table S2), only the Agnotecous cricket genus, microendemic to
this island, can produce sound. Agnotecous are known to produce sound above 9 kHz with a

129
dominant frequency between 12 and 19 kHz well above 7 kHz (Desutter-Grandcolas &
Robillard, 2006; Robillard et al., 2010). The global acoustic method does not seem to be able
to detect the differences potentially due to Agnotecous species as the distance value between
sites was flat at this frequency range. The lack of an obvious difference might be due to a
frequency turnover from one species to another that used a similar frequency range. Most
birds, Orthoptera and cicada species sing below 8 kHz, leading to the definition of biophony
(Pijanowski et al., 2011a), and so could be at the origin of the between sites acoustic
differences. New Caledonia counts 183 bird species including 23 regional endemics
(Desmoulins & Barré, 2005). No microendemic species of birds are expected at this local
scale because of the capacity of the birds to disperse. Even if birds are not narrow
microendemics such as the insects, lizards or plants species limited to one small mountain,
some bird species were described to be restricted to a part of the island. Of the 40 species
inventoried in New Caledonia 11 are not found all over the main island (Desmoulins & Barré,
2005). Unfortunately, no inventory for either Orthoptera or cicadas, have been undertaken in
the three sites sampled. This lack of complete inventory data prevents fine conclusions about
the differences observed in this study. The global acoustic method could highlight acoustic
differences between sites. The method was unable to identify all differences due to
microendemic species but the hypotheses that the evolutionary history could have led to
different acoustic assemblages in terms of species composition, species interaction and
species dynamics is not excluded. Furthermore, other hypotheses as dispersal or community
interactions could lead to acoustic differentiation between sites. It is still also quite difficult to
pinpoint a factor that could explain the occurrence of acoustic differences between sites. An
acoustic adaptation to the habitat, as formulated by the acoustic adaptation hypothesis
(Boncoraglio & Saino, 2007; Ey & Fischer, 2009), seems unlikely as all sites were
characterized by the same habitat (tropical rainforest) at a similar altitude. However, this
needs to be confirmed with experiments testing the filtering acoustic properties of each site.
Even if the global acoustic approach was limited to three sites with three points per site, it
seems to be a powerful tool to reveal compositional differences between sites and could
therefore help in conservation management. Such a clear message from acoustics is not trivial
as the sites showed a low level of ecological differentiation, and produced an equivalent
acoustic activity level, under similar environmental parameters. Rapid global acoustic surveys
might not always be able to detect differences in species assemblages, but they open new
possibilities for biodiversity estimation and monitoring of endangered species at large scales.

130
Numerous recorders could be deployed over a large territory covering different sites and
habitats. Such remote sampling could be achieved quite quickly and could provide
standardized data without an intense effort or a high level of expertise. This is especially
useful in hotspots places where exploring biodiversity is an urgent and challenging task.
Microendemism is a well-identified challenge for local conservation policies, which requires
an improvement in the number, size and quality of protected areas (Jaffré et al., 2010; Pellens
& Grandcolas, 2010). Detecting dissimilarity and complementarity between sites could help
in delimiting areas to conserve. In future, recordings collected using the global acoustic
method could be re-examined based on these results, to then identify individual species and
monitor populations.
III.6 Acknowledgements
We would like to thank Dr. Hervé Jourdan and Edouard Bourguet for their help when
collection the acoustic data. We thank Dr.James Windmill and Dr. Yoland Savriama for
improving the language of the manuscript.

131
Chapitre 4
Effet des stratégies de gestion des agro-
systèmes sur le niveau de biodiversité
dans les Western Ghats (Inde) : une étude
acoustique et botanique
Communication :
Gasc A., Dechant B., Ayyappan N., Pélissier R, Pavoine S., Grandcolas P. and Sueur J.
(2012) Impact of agroforest management on animal diversity : application of an acoustic
method. Biodiversity Asia congress. poster.

132

133
Effect of agrosystem management on biodiversity level in Western Ghats, India: an
acoustical and botanical survey.
Authors: Gasc A.1*, B. Dechant1, R. Pelissier2, 3, N. Ayyappan3, S. Pavoine4, 5, P.
Grandcolas1, J. Sueur1.
1Muséum national d'Histoire naturelle, Département Systématique et Évolution, UMR 7205
CNRS OSEB, 45 rue Buffon, Paris, France
2Institut de Recherche pour le Développement (IRD), UMR AMAP, Boulevard de la Lironde,
TA A-51/PS2, 34398 Montpellier, France
3Departement of Ecology and Environmental Sciences, Pondicherry University, Pondicherry,
605 014, India
4Muséum National d’Histoire Naturelle, Département Ecologie et Gestion de la Biodiversité,
UMR 7204-CNRS-UPMC, CP 51, 55-61 rue Buffon, 75005 Paris, France
5Mathematical Ecology Research Group, Department of Zoology, University of Oxford,
South Parks Road, Oxford, OX1 3PS, UK
*Author for correspondence: Amandine GASC
Muséum national d’Histoire naturelle
Département Systématique et Évolution
UMR CNRS 7205 OSEB
CP 50, 45 rue Buffon
F-75005 Paris, France
Tel. + 33 1 40 79 31 34
Fax. + 33 1 40 79 56 79
Email. [email protected]

134
IV.1 Abstract/Résumé
IV.1.1 Abstract
Coffee plantations in the Western Ghats (India), like in any other tropical region, are grown
traditionally under the shade trees. Such plantations ensure ecological continuity providing
habitat for many species outside protected reserves. Because of difficulties in obtaining
permits for felling, transport and selling of native timbers, farmers often choose to plant
exotic species. This pressure has led to two strategies of agrosystem management: 1) plant
native species as shade trees; or 2) regularly replace the dead trees with exotic tree species.
The objective of our study was to evaluate the impact of these two agrosystem management
strategies in comparison with the forests on tree diversity and in local singing animal diversity
by employing non-invasive Rapid Biodiversity Assessments (RBA). The present study was
conducted in the Kodagu district of Karnatka in three habitat types: preserved forest, coffee
plantation with native trees, and coffee plantation with the exotic tree Grevillea robusta. Four
locations were sampled per habitat type for a total of 12 study sites. For each site, tree
diversity was assessed using a variable sampling area protocol and by calculating two α
diversity indices (the number of species and the Shannon index) and two β diversity indices
(the Euclidian and the Sørensen distances). The local acoustic community was recorded one
minute every hour during 19 days using automatic acoustic recorders. Acoustic data were
analysed using two indices without species identification. The first index, based on the
heterogeneity of the mean frequency spectrum, measured the animal activity level for an
estimation of an α singing-species diversity. The second index measured the distances
between pair-wise recordings calculating their (point-wise) differences in the acoustic
frequency domain for an estimation of β singing-species diversity. The differences in
diversity values were tested with non-parametric tests and by including acoustical or tree
composition distances into multivariate analyses. The exploration of tree data demonstrated a
clear difference between agroforest with exotic trees and agroforest with native trees. As
expected, agroforests with exotic tree species were associated with the lowest tree species
richness, lowest tree species evenness and lowest number of tree species shared with both
'forest' and the 'agroforests with native trees' sites. ‘Agroforests with native trees’ were close
to ‘forests’ tree diversity. Differences in animal acoustic diversity were found between the
three habitat types, especially at night. The acoustic α diversity index showed lower values for

135
‘forests’ than for ‘agroforests’ types (with native trees and with exotic trees). The acoustic β
diversity index showed significant differences between the three habitat types. Moreover,
these differences were organized along the habitat type gradient from forest to agroforest with
exotic species. This work revealed a negative impact of the use of exotic trees on tree
diversity and support the idea of a positive impact of agroforest on acoustic animal diversity
that could be explained by a positive ecotone effect and by the fact that coffee plantation can
be a food reserve for some animal species. This hypothesis will to be confirmed by further
analyses as soon as the acoustic identification of the local singing species is possible.
Keywords: Passive acoustics, agroecosystems, animal diversity, coffee plantations, native
and exotic trees, tree diversity.
IV.1.2 Résumé
Traditionnellement, les plants de café des Western Ghats (Inde), comme dans beaucoup de
régions tropicales, sont cultivés à l’ombre des arbres. De telles plantations assurent une
continuité écologique procurant un habitat pour de nombreuses espèces en dehors des sites
préservés. En raison de la difficulté d’obtenir des permis pour couper, transporter et vendre
des essences d’arbre natives, les agro-forestiers choisissent souvent de planter des espèces
d’arbres exotiques. Cette pression économique a fait donc naître deux stratégie de gestion qui
diffèrent par le fait de remplacer les arbres morts exclusivement par des espèces natives ou
par des espèces exotiques. L’objectif de cette étude est d’évaluer l’impact de ces deux
stratégies de gestion des agro-systèmes sur la diversité des arbres et des espèces chanteuses en
utilisant des méthodes non-invasives de Rapid Biodiversity Assessments (RBA). L’étude a été
réalisée à Kodagu dans le district de Karnatka dans trois types d’habitats : des forêts
préservées, des plantations de café avec des arbres natifs et des plantations de café avec
l’espèce d’arbre exotique Grevillea robusta. Pour chaque type d’habitat, quatre sites ont été
échantillonnés. Un total de 12 sites a donc été considéré. Pour chaque site, la diversité des
arbres a été évaluée à partir d’un protocole à aire d’échantillonnage variable et par le calcul de
deux indices de diversité α, à savoir le nombre d’espèce et l’indice de Shannon, et deux
indices de diversité β, à savoir les distances euclidiennes et de Sørensen.
La communauté acoustique locale a été enregistrée pendant une minute toutes les minutes
durant 19 jours à l’aide d’enregistreurs automatiques. Les données acoustiques ont été

136
analysées à l'aide de deux indices, sans identification des chants à l’espèce. Le premier indice,
fondé sur l’hétérogénéité du spectre fréquentiel moyen, a permis de mesurer le niveau
d’activité animale donnant une estimation de la diversité α des espèces-chanteuses. Le
deuxième indice a permis de mesurer la distance entre les enregistrements deux à deux en
mesurant les différences entre les spectres moyens donnant une estimation de la diversité β
des espèces chanteuses. Comme attendu, les agro-forêts avec les arbres exotiques présentent
la plus faible richesse spécifique (species richness), une uniformité des abondances relatives
(evenness) et un faible nombre d’espèces d'arbre partagées avec les deux autres types
d’habitat. Les agro-forêts avec des espèces d’arbres natives ont une diversité d’arbre proche
de celle des forêts. Les diversités acoustiques d’origine animale semblent différentes entre les
trois types d’habitat pour les horaires de nuit. L’indice de diversité acoustique α présente des
valeurs un peu plus faibles en comparaison des agro-forêts. L’indice de diversité acoustique β
présente des différences significatives entre les trois types d’habitat, différences qui sont
organisées le long d’un gradient suivant les types d’habitat, de la forêt aux agro-forêts avec
arbres natifs. Ce travail révèle un impact négatif de l’utilisation des espèces d’arbres
exotiques et supporte l’idée d’un impact positif des agro-forêts sur la diversité acoustique qui
pourrait s’expliquer par un effet d’écotone des agro-forêts et par le fait que les plantation de
café puissent être des réservoirs de nourriture pour certaines espèces. Cette hypothèse doit
être testée par des analyses complémentaires dès que l’identification des chants composants
les enregistrements sera possible.
IV.2 Introduction
Agroforestry is a promising approach to slow down the current trends toward deforestation by
combining shade trees with agricultural crops. Based on the observation that biological
diversity decreases with the conversion from shade to sun agroforestry systems and on the
fact that shade agroforestry systems with high canopy cover and less intensive management
share a higher proportion of species with the remnants of intact forest, shade agroforestry
systems are considered to be refuges for biodiversity (Bhagwat et al., 2008, Perfecto et al.,
1996). Trees not only provide food for animals in the form of living or dead vegetative tissue,
but they also provide a variety of stable microhabitats with lower ambient temperature and
slower ambient air currents (Campanha, 2004). Compared to monoculture agricultural
systems, agroforestry systems show an optimized ecosystem functioning as far as water

137
quality, nutrient cycling, soil erosion, high wind and pest impacts are concerned (Altieri,
1999, Malézieux et al., 2009; Perfecto et al., 1996). Due to all these beneficial properties,
agroforestry systems can provide habitat for many species outside protected reserves in
otherwise highly human dominated landscapes. Thus, agroforestry plantations can help to
maintain or recreate ecological continuity between nature reserves or remnants of natural
habitats; thereby, avoiding isolation of these protected areas and allowing movement of
species supporting migratory species (Bhagwat et al., 2008; Perfecto et al., 1996).
In the Western Ghats (India), coffee plantations are grown traditionally under the
shade of native or exotic trees. Even if shade can have a negative impact on coffee plant
productivity due to a lack of sunlight as a factor of plant development (Campanha et al.,
2004), farmers are aware of all advantages of shade trees on ecosystem functioning leading to
the good health of the plantation. Besides, the timber exploitation of dead trees is an
additional resource. Because of difficulties in obtaining permits for felling, transport and
selling of native timbers, farmers often choose to plant exotic species. In Nath et al. (2011),
the authors illustrated the use of Grevillea robusta by the fact that this is the most dominant
species in coffee plantation of Kodagu area, representing 20% of a census of over 20 000 trees
in 2008.
The facts that Kodagu belong to the Western Ghats identified as a biodiversity hotspot
(Myers et al., 2000) and that already 33% of the surface area of the district is used for coffee
cultivation (Garcia et al., 2010) raise the question of the impact of alien species on the
biodiversity of agrosystem (McNeely, 2004). The objective of the present work was to
evaluate the impact on alpha and beta local diversities (singing animals and trees) of two
different agrosystem management strategies that replace a dead tree by either a native species
or the exotic species G. robusta. The hypothesis that 'Agroforest with exotic trees' would have
lower biodiversity level than 'Agroforest with native trees' was tested. Field work was
conducted in three different habitat types in Kodagu district of Karnatka area: preserved
forest, coffee plantation with native trees and coffee plantation with G. robusta, by using trees
inventory (Nath et al., 2010; Sheil et al., 2003) and bioacoustical methods (Sueur et al.,
2008b). The term 'tree' is exclusively used for shade trees excluding coffee plants. Exotic trees
were all planted by farmers. No tree colonisation has to be considered. The habitat type called
'Agroforests with exotic trees' embeds agroforests with both native and exotic trees in
comparison to 'Agroforests with native trees' that considers only agroforests with native
species.

138
Methods for tree inventory or bioacoustical sampling were both non-invasive and
followed the principle of Rapid Biodiversity Assessment (RBA). Different sampling methods
have been developed to accelerate tree sampling in tropical forests. This is the case of the
modified Variable Area Transect (VAT) proposed by Sheil et al. (2003). This easy-to-apply
method has the advantage of rapidly assessing a tropical forest site with a comparative
efficiency and accuracy of a square plot sampling method for vegetation expected to be
randomly dispersed (Nath et al., 2010). In bioacoustical surveys, there seems to be a tendency
to focus either on a single species or a particular taxonomic group as far as automated pattern
recognition methods are concerned. This focus may be very useful to detect the presence of a
certain species, but the gap between methods tailored to a single species or taxonomic group
and those needed for monitoring an entire acoustically active animal community appears to be
considerable. Technical difficulties include not only the creation of comprehensive animal
sound libraries with high quality recordings, but also the parallel implementation of a variety
of methods, each adapted to a certain type of song (Brandes, 2008, Towsey et al., 2012). In
order to avoid these difficulties, it has been suggested to analyse animal acoustic information
on the scale of the community rather than at species level. In this approach, species are not
identified but the global properties of the recordings were extracted and analysed under the
hypothesis of a statistical link between the resulting indices and indices commonly used to
evaluate the biodiversity level, such as number of vocalizations (Sueur et al., 2008b) or
singing species richness (Depraetere et al., 2012). In this paper we assume that the song type
diversity is positively correlated with the singing species diversity. Such a non-invasive
method aims at providing a rapid assessment over large time and spatial scales and allows
simultaneous access to several taxa (mammals, birds, insects, and amphibians).
As expected, the results highlighted differences in trees diversities (α and β) between
on one hand, the agroforests with exotic trees and, on the other hand, the agroforests with
native trees and the forests. The acoustic diversity indices revealed differences between sites
in acoustic activity level that could be linked to tree diversity. Moreover, there were acoustic
differences between habitat types with an intermediate position of agroforests with native
trees situated between agroforests with exotic trees and forests.

139
IV.3 Materiel and methods
IV.3.1 Sites
Twelve sites in Kodagu district were selected with a mean altitude of 981 ±120 m (Fig. IV.1).
Four sites were chosen for each of the three habitat types, namely 'forest' (Fo), 'agroforest
with native trees' (An) and 'agroforest with exotic trees' (Ae). A number from 1 to 4 indicating
habitat type repetition was added after the annotations (An, Fo or Ae) to indicate the sites.
The sites were all in private area; as a consequence, the choice of the sites depended mainly
on the approval of the farmer. In agroforest, the two habitat types were defined without any
idea of the tree composition on the sites. The presence of G. robusta was the only criteria used
to define the habitat type: 'Agroforest with exotic trees'.
Fig. IV.1: Sampled sites in the Kodagu disctrict (India). There were four audio recorders at each habitat type
separated by a minimum of 200 m. The GPS points were located between 12°05,862'N-12°22,803'N and
075°35,733'E-075°52,792'E.

140
IV.3.2 Tree diversity
Local tree diversity was assessed once from February 12th to 18th 2011 using a modified
standard variable sampling area protocol (Sheil et al., 2003; Nath et al., 2010). For each site, a
40 m transect-line delineated the sample unit. The transect was divided into four parts of 10 m
each composing the width of eight cells (four cells on each side of the transect). If the width
of the cell was fixed, its length was variable. Each cell captures the first five trees. The length
of each cell was determined by the position of the fifth-most distant tree from the baseline.
Sampling was stopped at a length of 20 m even if less than five trees were found or at a length
of 15 m if no trees were found at all. Thus, the eight cells together per transect provide data on
up to 40 trees, and the maximum area sampled is 40 m by 40 m. All trees were identified by
one of us (A.N.) referring to Ramesh et al. (2010) and Keshawa Murthy and Yoganarasimhan
(1990).
Tree diversity was assessed with four indices, two α diversity indices and two β
diversity indices. For each diversity facet (α and β), one considered the number of species and
the other one considered the relative abundances of the species. Alpha diversity was measured
as species richness (the number of species - 1) and Shannon diversity index calculated
following:
1log( )
i
j i iShannon p p= − ×∑
with pi the proportion of the species i for the site j. Species richness was standardized by the
number of trees counted on the site (i.e. normalized species richness per stem). The
exponential of Shannon diversity index was used in order to comply the replication principle
that supposes that "if we have N equally large, equally diverse groups with no species in
common, the diversity of the pooled groups must be N times the diversity of a single group"
(Chao et al., 2010).
Beta diversity was measured with two indices. The Euclidian distance (Deza and
Deza, 2009) and the Sørensen distance (Sørensen, 1948) measured the pair-wise distances
between sites in species composition. The euclidian distance considered the relative
abundance of the species whereas Sørensen distance considered only the presence / absence of
tree species. Considering the inventory data, the Euclidian distances were calculated as
following:

141
2
1( , ) ( )
n
i ii
d p q q p=
= −∑
where p and q are the two vectors, one for each site, representing the relative abundances of
each species i.
The Sørensen distances were calculated following:
1 2
21 CdS S
= −+
where C is the number of species shared between the two sites, S1 the number of species
found in the first site and S2 the number of species found in the second site.
IV.3.3 Tree density
Tree density was estimated for each site following Sheil et al. (2003). Considering the
sampling protocol detailed above, there were three different types of cells: (i) a complete 10-
m-wide and X-m-length cells containing 5 trees, (ii) a 10-m-wide and 20-m-length cells
containing less than 5 trees and (iii) 10-m-wide and 15-m-length cells containing no trees.
Tree density was therefore calculated in relation to the length of sampled cells. The density of
the first type of cell was calculated following:
1
45 10
N
cell ii
density nX=
=× ×∑
where X is the length of the cell depending of the most distant tree, N the number of species
and n the number of individual of the species i. The density of the second type of cell was
calculated following:
21
120 10
N
cell ii
density n=
=×∑
and the last type of cell containing no trees had a density of 0.
IV.3.4 Passive acoustic recording
The recordings were made from February 12th to March 29th 2011. The recording devices
were programmed to make one-minute long recordings every hour. SM2 autonomous digital
recording devices (Wildlife Acoustics, 2009) equipped with weatherproof omnidirectional
microphones with a flat frequency response in the range of 20Hz to 20 kHz were used for all

142
acoustic recordings. At each site, a microphone was placed approximately 2 m above ground.
All recordings were sampled at a 44.1 kHz rate and stored in the form of Wildlife Acoustics
Audio Compression (WAC) files, which were converted into Waveform Audio File Format
(WAV) with the software 'WAC to WAV Converter Utility' (Wildlife Acoustics, 2009).
IV.3.5 Acoustic data selection
Out of the 33 days during which all recorders have worked simultaneously, a sample of 19
days was drawn randomly and kept for the subsequent analysis in order to reduce the amount
of data to an acceptable level for listening. No particular focus on the 'dawn chorus' or periods
of high acoustic activity during the night was intended. All recording files were listened to by
B.D to prepare the files for acoustic measures. Using a spreadsheet, information about each
recording obtained from the listening and visual inspection of the spectrograms was listed to
systematically. Recording files with considerable noise above frequencies of 1 kHz were
excluded; all the other recording files were filtered in order to disregard everything below 1
kHz since most of the noise independent of the recording equipment was in this range. Most
excluded recording files were affected by rain (and/or wind noise) or noise in some way
related to human activity such as human voices, domestic animals' vocalizations and
machinery noise. From this selection, the acoustic indices (see below) were calculated
automatically on each recording file. On one of the agroforest sites with native trees (site
'An3') the microphone had ceased working properly on March 3. This reduced the number of
days from 19 to 9 days for this site, which might be considered a 'sampling bias' compared to
the other sites. One of the other four sites of this habitat type, site 'An2', had very regular
hours of loud machinery noise in the evenings leading to complete exclusion of the hours
from 19h to 21h and a reduction of the hour repetition for 22h to 1h.
From the theoretically expected 19×12× 24 = 5472 recordings, only 3925 could be used for
further analysis, which translates to a loss of 28% of the recording files.
IV.3.6 Acoustic diversity indices
Two acoustic indices were automatically calculated on the mean frequency spectrum obtained
with a Short Term Fourier Transform (STFT, non-overlapping Hamming window size = 512
samples = 11 ms) applied to each audio file.

143
The number of peaks (NP) of each mean frequency spectrum was previously
associated with an acoustic activity level, in terms of number of different song types, in a
tropical environment (unpublished results; see Chapter 2 of the thesis). The mean frequency
spectrum was scaled by its maximum leading to values ranging from 0 to 1. All frequency
peaks were first detected. A selection according to amplitude and frequency thresholds was
then applied to remove residuals peaks due to background noise. The first selection factor was
based on the amplitude slopes of the peaks, i.e. on the first derivative of the frequency
spectrum. Only peaks with slopes above 0.01 were kept. The second selection factor was
based on frequency with a minimum frequency interval between successive peaks of 200 Hz.
NP was then simply defined as the number of selected peaks on a mean spectrum after
detection and selection.
Acoustic spectral distance Df, as defined by Sueur et al. (2008b), measures the
acoustic distance between two recordings. Df was computed according to:
10.5 1( ) 2( )
N
ff
D S f S f=
= × −∑
where S1(f) and S2(f) are the probability mass functions of length N of the mean spectra of the
two recordings to be compared.
NP and Df values measured on each recording were averaged over all available days.
This average will decrease variations due to meteorological fluctuations.
IV.3.7 Spatial correlation
The geographic distances between each pair-wise site were calculated using the latitude and
longitude coordinates (see appendix 1) following the Sinnott (1984) formula that assumes a
spherical Earth and ignores an ellipsoidal effect:
2 21 2 1 22 arcsin sin ( ) cos( 1) cos( 2) sin ( )2 2geo
lat lat lon lond R lat lat− −= × × × × ×
where R is Earth radius, lat1 and lat2 the latitude of the two sites and lon1 and lon2 the
longitude of the two sites.
The correlation between geographic distances (11*11, sites-sites) and acoustic
distances ((11*24)*(11*24), Sites-Hours*Sites-Hours) were tested. Rao’s (1982) DISC index
was used to compute a between-site distance matrix; this coefficient enhances differences
among sites, removing differences observed among hours. A Mantel test with 999 replications
(Mantel, 1967) was applied on the resulting two matrices (11*11, sites-sites). This test

144
revealed a positive, moderate and significant correlation between the geographical and
acoustic matrices (r=0.266; p=0.037). This result was expected due to the geographic
closeness of 'Fo3' and 'Fo4' sites. The same test was run after the exclusion of 'Fo3' site
returning a non-significant p-value (r=0.210; p-value=0.191). Fo3 site was therefore excluded
from the analyses.
IV.3.8 Statistical analysis of acoustic data
Acoustic data were analysed separately for night time (8 p.m. - 5 a.m.) and for day time (6
a.m. - 7 p.m.), 6 a.m. corresponding to the sunrise and 8 p.m to the sunset. NP values were
first log-transformed to fit a normal distribution and then used as a dependent variable of a
linear model. The explanatory variables were the habitat type, the tree diversity (species
richness) and the tree density. Due to temporal autocorrelation among NP values, the temporal
variable (hours) was decomposed into trigonometric coordinates and included into the linear
model. A 5% significance level was used as a criterion for rejecting the null hypotheses.
A Principal Coordinate Analysis (PCoA; Gower, 1966) was used to visualize whether
the type of habitat could explain differences in tree compositions.
The Df distance matrix was first subjected to a Principal Coordinate Analysis. However, the
first and second axes explained a low percentage of variability (14.24 % and 4.86 %
respectively for night time and 2.97% and 2.23% respectively for day time). The Df distance
matrix was then analyzed with a partial distance-based ReDundancy Analysis (dbRDA) with
the habitat type factor as a constraint and the time (hours) as a conditional variable (Anderson,
M.J. & Willis, 2003; Legendre & Anderson, 1999). Associated to the partial dbRDA, a
permutation test was applied with 199 permutations as defined by the function capscale of the
package vegan (Oksanen et al., 2011).
All calculations were achieved using R software (R Development Core Team, 2010) including
the packages ‘seewave’ (Sueur et al., 2008a), ‘ade4’ (Dray & Dufour, 2007), ‘geosphere’
(Hijmans et al., 2011) and ‘vegan’ (Oksanen et al., 2011).
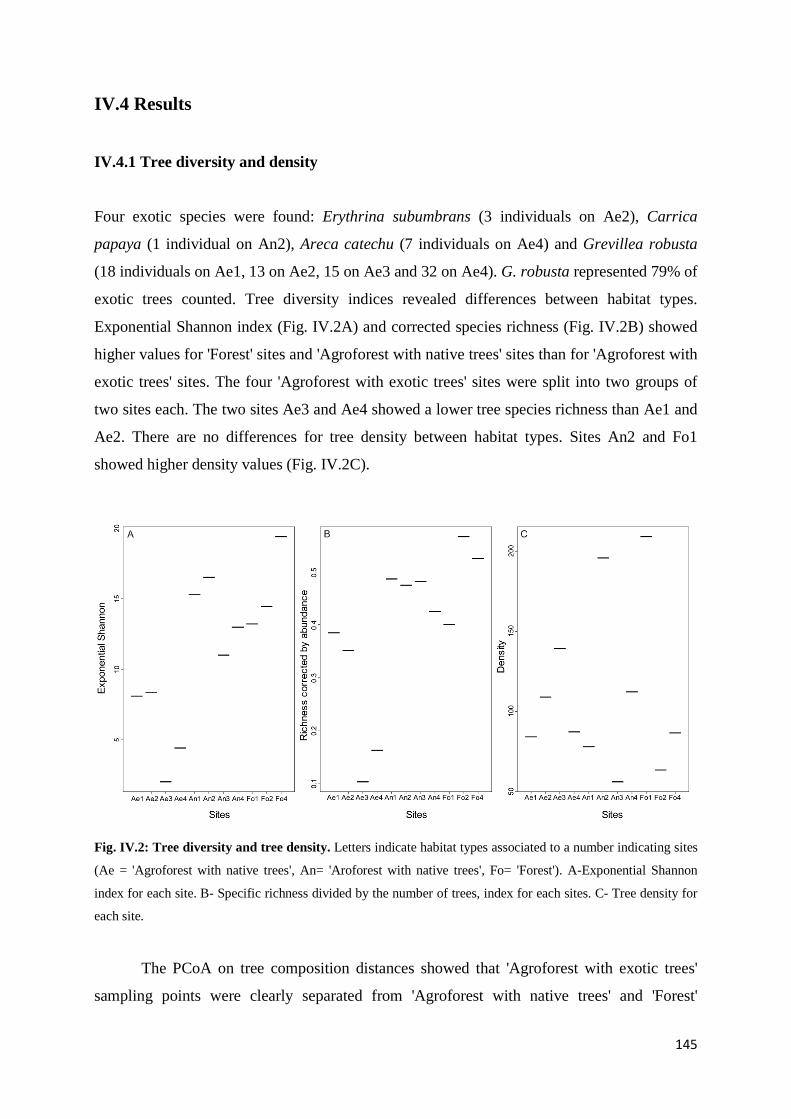
145
IV.4 Results
IV.4.1 Tree diversity and density
Four exotic species were found: Erythrina subumbrans (3 individuals on Ae2), Carrica
papaya (1 individual on An2), Areca catechu (7 individuals on Ae4) and Grevillea robusta
(18 individuals on Ae1, 13 on Ae2, 15 on Ae3 and 32 on Ae4). G. robusta represented 79% of
exotic trees counted. Tree diversity indices revealed differences between habitat types.
Exponential Shannon index (Fig. IV.2A) and corrected species richness (Fig. IV.2B) showed
higher values for 'Forest' sites and 'Agroforest with native trees' sites than for 'Agroforest with
exotic trees' sites. The four 'Agroforest with exotic trees' sites were split into two groups of
two sites each. The two sites Ae3 and Ae4 showed a lower tree species richness than Ae1 and
Ae2. There are no differences for tree density between habitat types. Sites An2 and Fo1
showed higher density values (Fig. IV.2C).
Fig. IV.2: Tree diversity and tree density. Letters indicate habitat types associated to a number indicating sites
(Ae = 'Agroforest with native trees', An= 'Aroforest with native trees', Fo= 'Forest'). A-Exponential Shannon
index for each site. B- Specific richness divided by the number of trees, index for each sites. C- Tree density for
each site.
The PCoA on tree composition distances showed that 'Agroforest with exotic trees'
sampling points were clearly separated from 'Agroforest with native trees' and 'Forest'
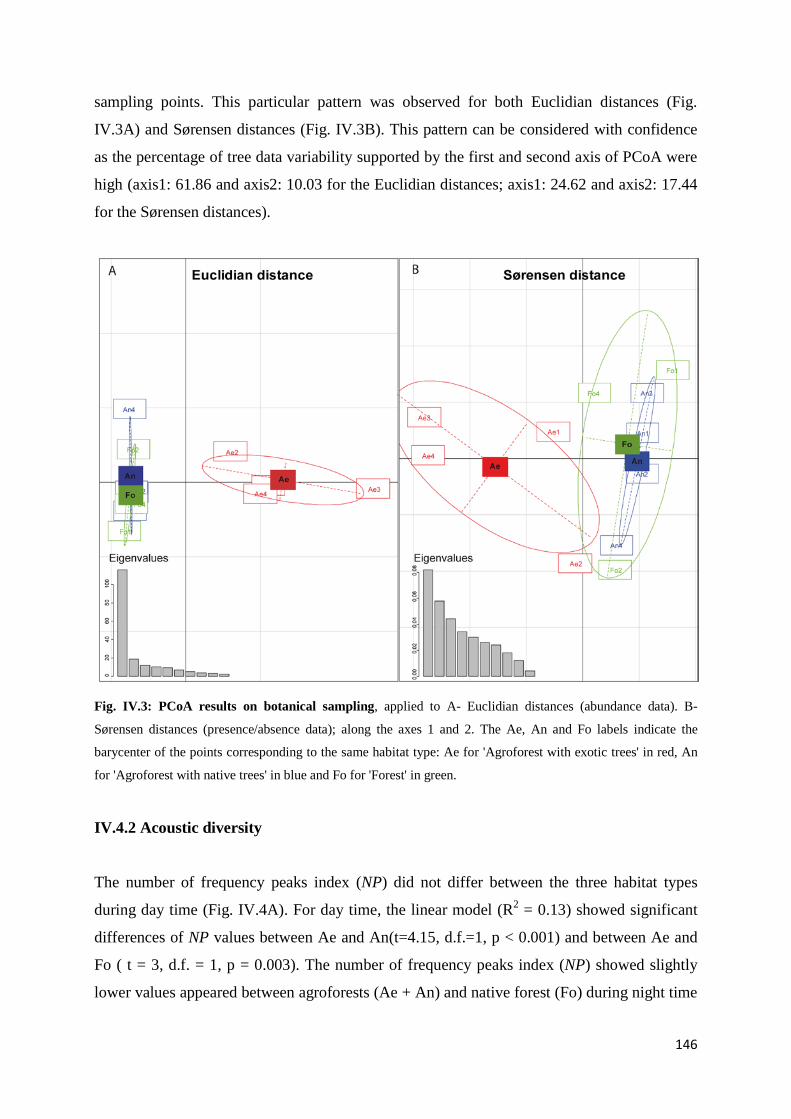
146
sampling points. This particular pattern was observed for both Euclidian distances (Fig.
IV.3A) and Sørensen distances (Fig. IV.3B). This pattern can be considered with confidence
as the percentage of tree data variability supported by the first and second axis of PCoA were
high (axis1: 61.86 and axis2: 10.03 for the Euclidian distances; axis1: 24.62 and axis2: 17.44
for the Sørensen distances).
Fig. IV.3: PCoA results on botanical sampling, applied to A- Euclidian distances (abundance data). B-
Sørensen distances (presence/absence data); along the axes 1 and 2. The Ae, An and Fo labels indicate the
barycenter of the points corresponding to the same habitat type: Ae for 'Agroforest with exotic trees' in red, An
for 'Agroforest with native trees' in blue and Fo for 'Forest' in green.
IV.4.2 Acoustic diversity
The number of frequency peaks index (NP) did not differ between the three habitat types
during day time (Fig. IV.4A). For day time, the linear model (R2 = 0.13) showed significant
differences of NP values between Ae and An(t=4.15, d.f.=1, p < 0.001) and between Ae and
Fo ( t = 3, d.f. = 1, p = 0.003). The number of frequency peaks index (NP) showed slightly
lower values appeared between agroforests (Ae + An) and native forest (Fo) during night time
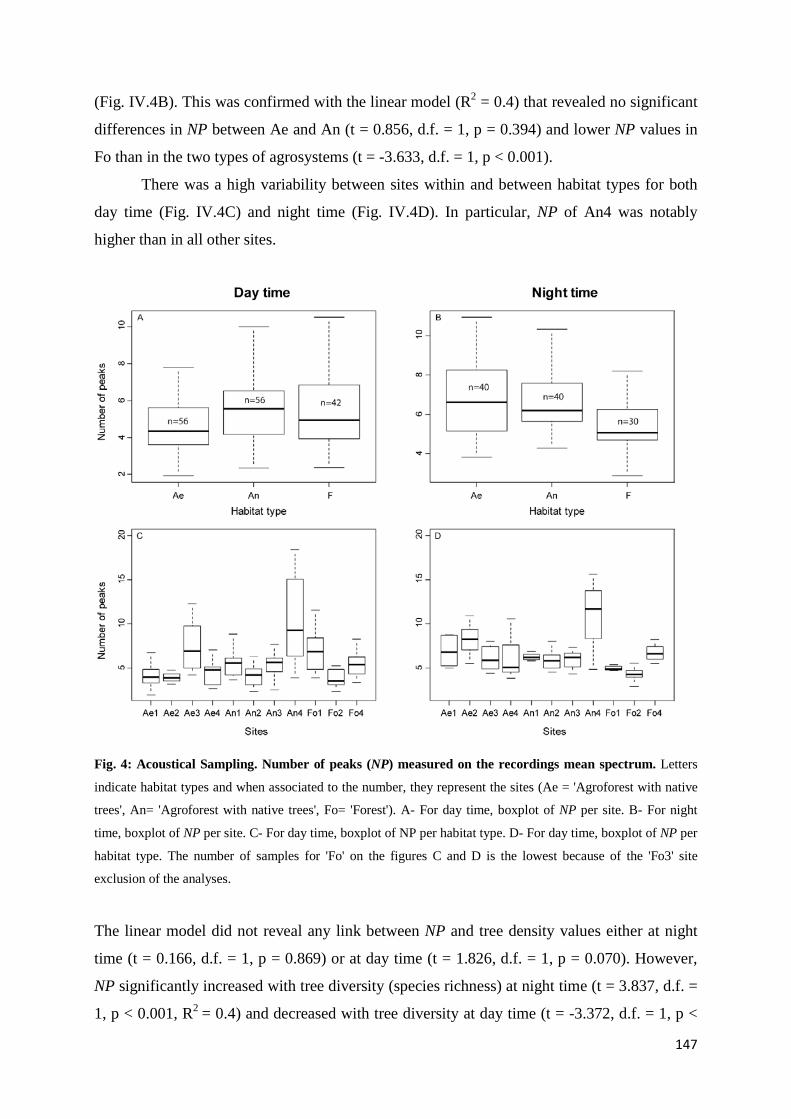
147
(Fig. IV.4B). This was confirmed with the linear model (R2 = 0.4) that revealed no significant
differences in NP between Ae and An (t = 0.856, d.f. = 1, p = 0.394) and lower NP values in
Fo than in the two types of agrosystems (t = -3.633, d.f. = 1, p < 0.001).
There was a high variability between sites within and between habitat types for both
day time (Fig. IV.4C) and night time (Fig. IV.4D). In particular, NP of An4 was notably
higher than in all other sites.
Fig. 4: Acoustical Sampling. Number of peaks (NP) measured on the recordings mean spectrum. Letters
indicate habitat types and when associated to the number, they represent the sites (Ae = 'Agroforest with native
trees', An= 'Agroforest with native trees', Fo= 'Forest'). A- For day time, boxplot of NP per site. B- For night
time, boxplot of NP per site. C- For day time, boxplot of NP per habitat type. D- For day time, boxplot of NP per
habitat type. The number of samples for 'Fo' on the figures C and D is the lowest because of the 'Fo3' site
exclusion of the analyses.
The linear model did not reveal any link between NP and tree density values either at night
time (t = 0.166, d.f. = 1, p = 0.869) or at day time (t = 1.826, d.f. = 1, p = 0.070). However,
NP significantly increased with tree diversity (species richness) at night time (t = 3.837, d.f. =
1, p < 0.001, R2 = 0.4) and decreased with tree diversity at day time (t = -3.372, d.f. = 1, p <
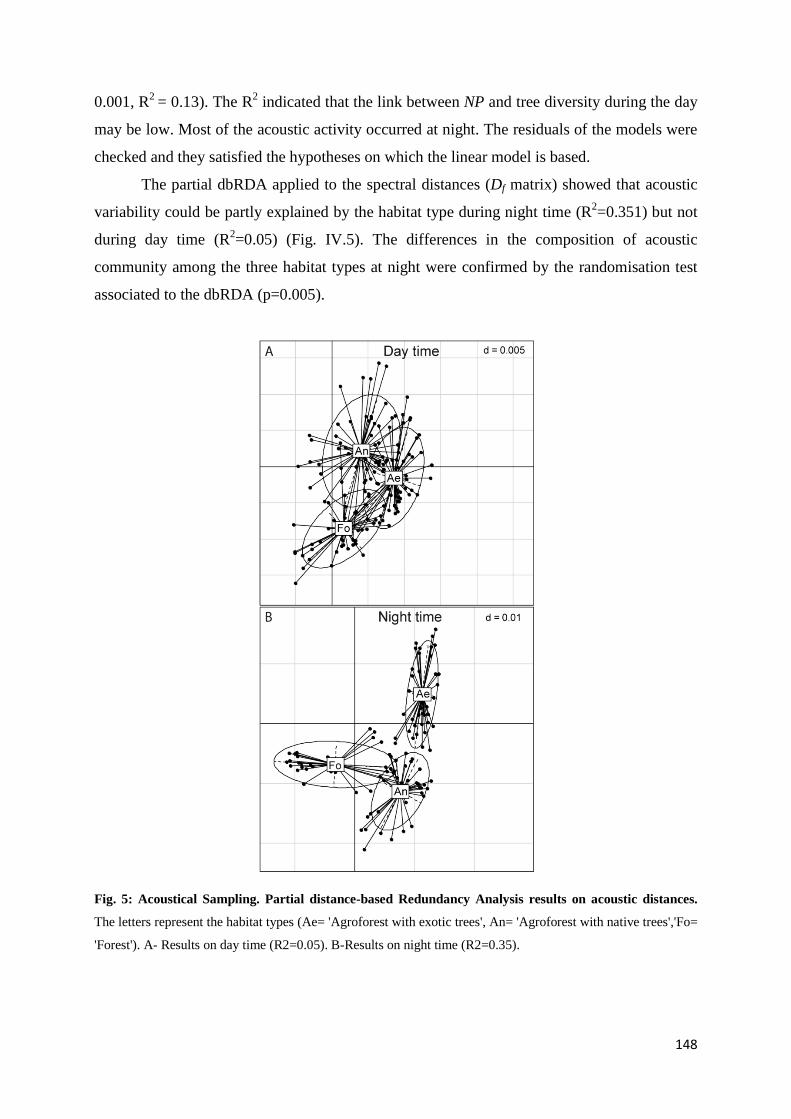
148
0.001, R2 = 0.13). The R2 indicated that the link between NP and tree diversity during the day
may be low. Most of the acoustic activity occurred at night. The residuals of the models were
checked and they satisfied the hypotheses on which the linear model is based.
The partial dbRDA applied to the spectral distances (Df matrix) showed that acoustic
variability could be partly explained by the habitat type during night time (R2=0.351) but not
during day time (R2=0.05) (Fig. IV.5). The differences in the composition of acoustic
community among the three habitat types at night were confirmed by the randomisation test
associated to the dbRDA (p=0.005).
Fig. 5: Acoustical Sampling. Partial distance-based Redundancy Analysis results on acoustic distances.
The letters represent the habitat types (Ae= 'Agroforest with exotic trees', An= 'Agroforest with native trees','Fo=
'Forest'). A- Results on day time (R2=0.05). B-Results on night time (R2=0.35).

149
IV.5 DISCUSSION
Face to the coffee market crisis, economical policy of many tropical countries leads to the
replacement of native species by exotic shade tree species in coffee plantation. G. robusta is
the exotic species mainly used in coffee plantation in Kodagu district (Nath et al., 2011). The
present paper analyses the impact of two agroforest management strategies using two Rapid
Biodiversity Assessment techniques to estimate tree and acoustic diversities. The hypothesis
that 'Agroforest with exotic trees' would have lower biodiversity level than 'Agroforest with
native trees' was supported regarding α and β tree diversity and for β diversities but rejected
for acoustic α diversity.
Recent efforts in bioacoustics have been undertaken to move from species to community
levels for biodiversity assessment (Pieretti, 2011; Villanueva-Rivera, 2011; Sueur et al.,
2008b; Joo et al., 2011). The sampling done in the Western Ghats aimed at using this under
developed approach with two complementary indices, namely the number of frequency peaks
(NP) and spectral dissimilarity (Df). Based on aural tests (unpublished results, see Chapter 2
of the thesis), the NP index was assumed to increase with the number of song types revealing
a level of acoustic activity.
Assuming that NP is correlated with the number of different song types, acoustic
activity would be higher during night than during day, an expected result as sound is known
as an efficient communication modality in darkness (Bradbury and Vehrencamp, 1998).
Nocturnal acoustic songs assemblages in agroforests were different from those occurring in
the forest habitat. Contrary to night time, the R-squared of this linear model for day time was
low indicating that the model results were not conclusive.
NP values showed a high variability between sites. This variability could be explained
by species composition but also site-specific physical characteristics related to degree of tree
density. Sound propagation is indeed severely constrained by obstacles as trees and bushes
(Richards & Wiley, 1980; Wiley & Richards, 1978). This can induce important deviation in
sound properties leading to acoustic differences between sites. Here tree density did not differ
between sites and therefore did not imply different acoustic propagation rules. Tree density
could not be linked to NP variability between sites. However, other vegetation parameters as
understory density, which were unfortunately not estimated, might differ between sites.

150
At the habitat level, the number of song types was higher in agroforest than in forest
sites. Two possible explanations could be considered. First, the number of song types could be
linked to a number of singing species, or the number of song types is linked to species with
high acoustic activity. In these two cases, there is an effect of agroforest on species richness
and/or species composition. The beta diversity results support that the species assemblage
could be affected by habitat type. Birds and Orthoptera are mainly responsible of songs
recorded. Agroforests provide food resources for bird species that could lead to an increase in
bird diversity (Wunderle & Latta, 1998). On the contrary, the bird diversity could be
negatively impacted by the presence of G. robusta (Bali et al., 2006).
The agroforest sites with native shade trees showed a high tree diversity level similar
to forest sites in both α diversity and β diversity. This advocates the role of shade tree
agrosystem in biodiversity conservation (Bhagwat et al., 2008, Perfecto et al., 1996). In
contrast, agroforest sites with exotic trees had tree richness lower than the agroforests that use
native shade trees due to the massive use of a single exotic tree species in the area of Kodagu
(Nath et al., 2011). The acoustic distances between the three habitat types suggested that the
song compositions changed from forest to agroforest with exotic tree species with an
intermediate position of agroforest with native tree species. An additional acoustic analysis
would permit the exact number of song types in each recording and the identificaty of singing
species to be determined. This additional analysis would allow further discussion about which
species are at the origin of the acoustic differentiation between habitat types and if
homogenisation occurs for singing species. Nevertheless, such an analysis is not
straightforward, as it requires reliable aural identifications conducted by several experts
whose are difficult to find for tropical habitats where species-specific acoustics is far to be
described.
To favour the conservation of native shade trees in agrosystems, some authors proposed to
establish a quality label to compensate the yield loss of farmers that provide biodiversity
service by their agroforest management (Gobbi, 2000; Perfecto, 2005). The assumption that a
trade off exists between yield and biodiversity was recently disputed. A study monitored nine
taxonomic groups during two years in 43 smallholder cacao forestry systems and
demonstrated that with appropriate management, high biodiversity and high yield are
compatible (Clough et al., 2010). Even if the acoustic interpretation has to be confirmed,
these results may have implications on the coffee agroforest management strategy and support

151
the use of native species in agroforest management as tree diversity and acoustic composition
are the closest to natural systems.
IV.6 ACKNOWLEDGEMENTS
This work was supported by a CNRS INEE PhD grant allocated to AG, the FRB (Fondation
pour la Recherche sur la Biodiversité) grant BIOSOUND. We would like to thank Nicolas
Deguines and Addy for their help for collecting the acoustic data and Dr. Cheryl Nath for
sharing missing botanical data. We thank Dr Brent Berger for improving the language of the
manuscript.

152

153
DISCUSSION
GENERALE

154

155
Les travaux présentés dans les quatre chapitres précédents apportent un nouveau regard
critique sur, d'une part, la diversité acoustique en tant qu'aspect de la biodiversité et, d'autre
part, sur les outils disponibles permettant d'y avoir accès. Nous nous sommes concentrés sur
la méthode de Rapid Acoustic Survey (RAS) qui nous a semblé d'un grand intérêt puisqu'elle
tente de répondre aux demandes actuelles de mesure de la biodiversité, à savoir, analyser et
suivre la biodiversité de manière rapide et en ayant le moins d’impact possible sur les objets
d’étude.
Ce dernier volet présentera dans un premier temps une réflexion globale sur la notion
de diversité acoustique. La deuxième partie présentera une discussion sur certains points
méthodologiques. La troisième partie présentera une réflexion autour de la notion de méta-
méthode dont la RAS fait partie, du potentiel de ces méthodes et des perspectives globales qui
peuvent être attendues pour la RAS en particulier. Enfin nous conclurons par une discussion
plus générale sur les perspectives de recherches à venir pour une estimation et un suivi de la
distribution spatiale et temporelle de la biodiversité.
Pendant le travail effectué, de nombreuses limites méthodologiques ont été soulevées.
La discussion méthodologique qui va suivre en est le reflet.
V.1 La diversité acoustique
La diversité acoustique est ici définie comme la diversité des sons issus de l’activité
acoustique animale. Elle est donc dépendante des individus produisant ces sons. Cette
diversité acoustique peut être le résultat d'une variabilité inter- et intra-spécifique (dialectes,
répertoires).
Est-il légitime de réduire la diversité acoustique à la variabilité interspécifique ? Dans
l'étude de Sueur et al. (2008), les auteurs ont utilisé des simulations de signaux acoustiques
produit par une communauté pour tester l'effet du nombre d'espèces chanteuses sur l'indice
d'entropie acoustique (H). L'hypothèse suivante a été testée : plus le nombre d'espèces d’une
communauté animale est élevé, plus le nombre de signaux différents est élevé et plus
l'hétérogénéité du signal est forte. Les résultats des mesures faites sur les simulations
montrent que l'hétérogénéité du signal augmente avec le nombre d'espèces. Remarquons que
la communauté est simulée en ne considérant qu'un seul son par espèce. Le nombre d'espèces
est ainsi directement associé au nombre de types de sons différents ce qui ne permet pas de
prendre en compte la variabilité intraspécifique. Il en est de même pour l'étude menée sur la

156
diversité acoustique des communautés d'oiseaux présentée dans le chapitre 2. Dans ces deux
études, seule la variabilité interspécifique est considérée. Il serait intéressant de tester si ce
choix est légitime en comparant l'effet de la variabilité interspécifique par rapport à l'effet de
la variabilité intraspécifique sur la diversité acoustique. Il est attendu que les espèces avec un
grand répertoire de chants aient un plus fort impact sur la diversité acoustique d'une
communauté. Si on ajoute une espèce dans une communauté de n espèces animales, quelle
que soit l'espèce, la richesse spécifique sera n+1. Cette observation n'est pas transposable à la
diversité acoustique. Si on considère uniquement la variabilité interspécifique, les sons de
deux espèces peuvent être plus ou moins différents selon les espèces considérées. L'ajout d'un
chant d'une espèce dans un signal d'une communauté n'apportera pas la même variabilité
(Lellouch, 2012, Chapitre 1). La première conclusion qui peut être faite ici est que si la
diversité acoustique (diversité α) d'une communauté augmente avec le nombre d'espèces
chanteuses, elle ne le fait pas de la même façon pour chaque espèce, et n'augmente pas
linéairement (Sueur et al., 2008). Cela implique qu'il n'est pas nécessairement simple d'utiliser
la diversité acoustique comme indicateur du nombre d'espèces.
Qu'en est-il des autres facettes de la biodiversité ? Les résultats présentés dans le
chapitre 2 montrent que la diversité acoustique peut être corrélée à d’autres types de diversités
comme la diversité phylogénétique ou fonctionnelle (dans le cas de cette étude, pour les traits
fonctionnels du poids du corps et de reproduction). De plus, cette étude révèle que les
mécanismes à la base de la diversité acoustique semblent différents des mécanismes à
l'origine de la diversité phylogénétique et fonctionnelle. Ces résultats montrent que si certains
aspects de la diversité biologique semblent converger avec la diversité acoustique, une partie
de cette diversité pourrait être considérée comme un aspect de la biodiversité à part entière.
Dire que la diversité acoustique est liée en partie à d'autres aspects de la biodiversité
est une affirmation acceptable. Cependant, dire que la diversité acoustique pourrait permettre
d'estimer d'autres aspects de la biodiversité ne serait pas correct. Chaque aspect de la
biodiversité doit être mesuré par son indicateur (Duelli & Obrist, 2003).
Sans pouvoir affirmer le nombre exact d'espèces ni leur identité, les indices de diversité
acoustique α pourraient-ils être utilisés pour identifier des différences dans des niveaux de
biodiversité ? L’utilisation des indices acoustiques se base sur l’hypothèse que l'hétérogénéité
d'un chœur augmente avec la diversité des sons composant le chœur (Depraetere et al., 2012;
Sueur et al., 2008). Nous avons vu dans le chapitre 1 qu’il existe beaucoup de sources
d’hétérogénéité, et que, bien que plutôt robuste au bruit ambiant, NP est assez sensible aux

157
variations d’amplitude des chants. Pourtant l’indice NP semble positivement lié au nombre de
types de sons présents dans un enregistrement dans le cas de l'étude menée en Nouvelle-
Calédonie (Chapitre 3), suggérant que cette source d'hétérogénéité joue un rôle important
dans le calcul de NP. Cependant, la classification qui a été faite pour caractériser le nombre de
types de sons est assez grossière et nécessite une étude plus fine de la capacité de cet indice à
refléter le nombre de sons composant le signal sonore d'une communauté.
La diversité acoustique peut-elle être utilisée pour identifier des différences dans la
composition des communautés ? Les chapitres 3 et 4 montrent que l'utilisation de la
dissimilarité spectrale sur un large échantillon d'enregistrements peut mettre en évidence des
différences acoustiques par site (Chapitre 3) ou par type d'habitat (Chapitre 4). Les différences
acoustiques inter-sites ou inter-habitats sont plus faibles que les différences intra-sites ou
intra-habitats. Beaucoup de précautions ont été prises pour vérifier que la différence
acoustique observée entre sites pouvait être attribuée à la seule activité acoustique animale
dans le chapitre 3. En ce qui concerne les résultats du chapitre 4, nous n'avons pas pu vérifier
si la différence entre types d'habitat pouvait être influencée par la seule différence de bruit
ambiant et/ou par la caractéristique physique des habitats (Slabbekoorn, 2004). Si l'indice
permet de révéler des différences acoustiques et de les relier à des différences de composition
de communauté, il reste difficile d'identifier biologiquement ces différences. D'ailleurs, il est
très intéressant de remarquer que, pour le cas du chapitre 4, une différence acoustique n'est
observée que durant les heures de nuit. Cela veut-il dire que les communautés changent
pendant la journée ? Non bien entendu, les individus chanteurs peuvent être présents sans
pour autant émettre un signal sonore et ne sont donc pas détectés. Si l'échantillonnage
acoustique n'avait été fait qu'en journée, nous n'aurions détecté aucune différence
significative.
V.2 Critiques méthodologiques
V.2.1 Dissimilarité temporelle
Les indices utilisés dans les chapitres 3 et 4 sont des indices mesurant uniquement
l'hétérogénéité spectrale. Pourtant l'hétérogénéité temporelle reste une mesure qu'il serait
intéressant de prendre en compte. Les indices qui restent les plus pertinents pour l'analyse de

158
la diversité acoustique sont les indices de dissimilarité. Or, si mesurer des différences entre
deux fonctions en faisant la différence des points deux à deux, ce que nous appelons « point à
point », peut avoir un sens sur un spectre, cela reste difficilement interprétable sur une
enveloppe temporelle. En effet les points sont homologues d'un spectre à l'autre tandis qu'ils
ne le sont pas du tout entre deux enveloppes temporelles. C'est pour cette raison que l'indice
Dt a été écarté des analyses. L'indice 1-RV est pour cette même raison discutable. En effet, cet
indice mesure à la fois la différence temporelle et spectrale de deux spectrogrammes (Robert
et Escoufier, 1976). La mesure de dissimilarité temporelle entre deux enregistrements est
donc limitée par le fait que les indices utilisés sont des indices de mesure point à point. Une
piste de réflexion sur l'élaboration de nouveaux indices ne se basant pas sur ce type de mesure
a été proposée (Lellouch, 2012).
V.2.2 Community Acoustic Diversity
La mesure de la diversité acoustique a été nommée Community Acoustic Diversity « CAD »
et non simplement « Biophony » (Krause, 1987; Pijanowski, 2011a). Il est vrai que cette
mesure aurait pu être appelée « Biophony diversity » mais il nous a semblé pertinent
d'introduire ici la notion de communauté. Si, dans notre cas la diversité acoustique de la
communauté nous intéresse essentiellement du point de vue de l'assemblage des espèces, les
applications de cet indice peuvent être plus larges. En effet, notre étude a démontré que les
corrélations observées en regardant les distances entre les espèces ne correspondent pas
forcement aux corrélations des diversités observées au niveau des communautés. La CAD
n'est donc pas simplement une addition aléatoire des chants. La CAD met en évidence un
assemblage de chants où l'activité des individus chanteurs ne se fait pas au hasard, peut être
du fait de l'interaction entre les individus et l’habitat.
V.2.3 Diversité acoustique/diversité fonctionnelle
Mettre en parallèle diversité acoustique et diversité fonctionnelle est un choix discutable. En
effet, la diversité acoustique pourrait être associée à une diversité fonctionnelle, puisque la
production de sons par les espèces animales relève d'une fonction de communication (Leroy,
1979). Dans le cas de notre étude (Chapitre 2), nous avons considéré la diversité acoustique
d'une part et la diversité fonctionnelle de l'autre. La diversité fonctionnelle est un aspect de la
diversité très difficile à appréhender. En effet, elle peut inclure un très grand nombre de traits

159
fonctionnels (Tilman, 2001; Violle et al., 2007). Dans le cas présent, nous garderons à l'esprit
que les résultats ont été obtenus à partir d’une sélection de quelques traits pour lesquels des
données étaient disponibles.
V.2.4 Test sur communautés réelles : validation des indices sur la base d'inventaires
Nous avons abordé le problème des différentes sources d'hétérogénéité qui peuvent affecter la
mesure des indices acoustiques (Chapitre 1). Si les simulations en laboratoire nous permettent
de tester un à un les effets de ces sources d'hétérogénéité, il serait intéressant de pouvoir tester
ces effets sur des communautés réelles. Cela pourrait peut-être mettre en évidence des effets
croisés de ces sources, ou des sources qui n'ont qu'un faible impact sur le calcul des indices
par rapport aux autres. De plus, il serait intéressant d'étendre l'étude théorique présentée au
chapitre 2 à des signaux acoustiques réels de communautés. Cela implique d'avoir les données
fonctionnelles et phylogénétiques des espèces identifiées sur les enregistrements. En ce qui
concerne les oiseaux, ces données sont accessibles pour la plupart des espèces; mais pour
élargir l'étude à des communautés multi-taxons ces données sont peu ou pas disponibles.
Un travail d'inventaire doit être envisagé pour une étude finalisée afin de savoir si le
nombre de types de sons différents suffit à renseigner sur la diversité spécifique, fonctionnelle
et/ou phylogénétique des espèces. Il serait utile d'avoir accès aux programmes ATBI (All
Taxa Biodiversity Inventory) en cours et d'y associer un échantillonnage acoustique, qui
pourrait précéder la perturbation du site par des échantillonnages classiques. Cela permettrait
de savoir si la diversité acoustique mesurée est représentative de la diversité des espèces
chanteuses et/ou de la diversité de l'ensemble des espèces.
V.2.5 L'utilisation du spectre moyen
La mesure de l'hétérogénéité du spectre moyen comme mesure de la diversité acoustique a
fortement été remise en question (Chapitre 1; Dechant, 2012). Le spectre moyen représente
les fréquences utilisées avec des valeurs d'amplitude relatives. En effet, un chant répété
prendra plus de poids dans le spectre moyen qu’un chant bref (Dechant, 2012). Ce biais de
détection des chants est difficile à évaluer. Le problème de l’utilisation du spectre moyen
réside dans le fait que non seulement les espèces n’apportent pas le même niveau de
variabilité (spectre plus ou moins complexe) mais en plus, les communautés acoustiques ne se
font pas aléatoirement. Si les indices augmentent avec le nombre de types de chants ajoutés au

160
hasard dans les signaux de communautés au laboratoire(Sueur et al., 2008), l’assemblage non
aléatoire peut engendrer des résultats différents en milieu naturel. Par exemple, il est tout a
fait possible d’avoir des espèces dont le chant est très complexe en fréquence mais qui ne
chantent que pendant la nuit. En forêt tropicale, il y a une forte activité acoustique nocturne.
Ces chants complexes pourrait donc toujours être associés à un signal de communauté dans
lequel on trouve un grand nombre de type de sons (c'est souvent le cas de certains
Orthoptères). Ce phénomène peut entraîner une interprétation biaisée de l'indice NP. Il est
intéressant de noter qu'augmenter le nombre d'échantillons ne permettra pas de réduire cet
effet. Ainsi l’utilisation du spectre moyen pour obtenir une mesure représentant la diversité
acoustique, comme par exemple le nombre de types de sons, n’est peut être pas la méthode la
mieux adaptée. L’indice ACI par exemple travaille sur le sonagramme (Farina & Morri, 2008;
Pieretti et al., 2011). Cet indice possède des propriétés intéressantes comme le fait d'être peu
sensible aux variations relatives d'amplitude, et de prendre en compte à la fois les
composantes spectrales et temporelles (Chapitre 1). Les tests présentés en chapitre 1 ne nous
ont permis d'étudier ni l'effet du démarrage temporel des chants ni l'impact du rapport de la
durée du signal sur la durée de l'enregistrement. De nouveaux tests pourraient permettre de
connaitre ces effets. Il serait également intéressant de réfléchir au développement d'un indice
de diversité acoustique β sur la base du sonagramme.
V.2.6 Le traitement du bruit
Le bruit est un problème récurrent en analyse du signal sonore (Sueur et al., 2012). Il reste un
élément très limitant de nos analyses faites sur le terrain. En effet, avant toutes analyses, nous
avons du faire une écoute des enregistrements afin de sélectionner les fichiers non-bruités par
la pluie, le vent ou des bruits d'origine anthropiques (voitures, paroles, chiens, coups de feux,
etc.). Cette sélection est nécessaire mais reste la partie la plus longue de l'analyse. Ce temps
d'écoute a été limitant et nous a conduit à faire une sous-sélection de fichiers, en ce qui
concerne l'étude réalisée en Inde. De plus, nos enregistrements ont été faits sur une durée
variant entre 1 et 2 mois, mais le temps qui pourrait être beaucoup plus long pour d'autres
analyses et le tri des données encore plus conséquents. Nous avons essayé de faire une
sélection des fichiers en nous appuyant sur des données météorologiques de bonne qualité
(Météo France pour la Nouvelle-Calédonie) mais beaucoup trop de ces données étaient
manquantes. La solution la plus adaptée pour obtenir des données météorologiques précises
est d'associer une petite station météorologique à chaque enregistreur et d'utiliser des seuils au

161
delà desquels l'utilisation de l'enregistrement est exclue (Sueur et al., 2012), ou d'utiliser des
analyses multi-variées appropriées pour identifier les fichiers acceptables pour l'analyse
(Depraetere et al., 2012).
Le traitement du bruit ambiant intrinsèque au site a pu être traité dans le cas de l'étude
en Nouvelle-Calédonie sur la base des fichiers sonore vides d'activité animale. Cela n'a pas pu
être fait sur les données collectées en Inde parce qu'aucun fichier dénué de chants animaux n'a
été relevé.
V.2.7 Protocole d'échantillonnage acoustique
Il a été montré qu'il existe une très grande variabilité entre les enregistrements d'un
même site. La plus grande part de la variabilité est due à une variabilité horaire (Chapitre 3,
4). En effet, les espèces peuvent signaler leur présence à différents moments de la journée et
de la saison (Ficken et al., 1985; Gottsberger & Gruber, 2004). L'assemblage des sons s'en
trouve modifié. Cette variabilité est également vraie en ce qui concerne les inventaires
d'espèces. Dans les inventaires acoustiques classiques, la détection des espèces varie en
fonction de leur activité. Il y a donc une différence de détection entre les espèces (espèce plus
visible qu'une autre, espèce plus chanteuse qu'une autre) et pour une même espèce le long de
la journée et de la saison. L'échantillonnage acoustique fait donc face aux mêmes biais que les
autres méthodes d'échantillonnage, mais l'utilisation d'enregistreurs automatiques permet
d'avoir un échantillonnage précis et mieux réparti dans le temps, augmentant les chances de
détecter toutes les espèces chanteuses. Les programmes de suivi de la biodiversité supposent
en général que la détectabilité des espèces est la même, or la détectabilité des espèces n'est
constante ni dans le temps ni dans l'espace, ce qui entraîne des biais potentiellement
importants. Afin de diminuer ce biais, Meyer et al. (2011) proposent d'augmenter le nombre
de relevés par an sur le même point d'échantillonnage, ce qui peut être facilité par la pose
d'enregistreurs automatiques.
V.3 La méthode acoustique globale : une méta-méthode
L’inquiétude montante au sujet de la perte de la biodiversité a pour conséquence une demande
sociétale et scientifique forte d'une meilleure connaissance de l’état de la biodiversité. Face à
la rapidité estimée de cette dégradation, ce sont des résultats rapides d’évaluation du niveau
de biodiversité qui sont attendus. Mesurer la diversité spécifique, phylogénétique et/ou

162
fonctionnelle d’un site nécessite une identification à l’espèce, ce qui est un travail très lourd.
Traditionnellement, ces mesures se basent sur des échantillonnages à l’échelle de l’individu.
Face à ces besoins et difficultés, les chercheurs ont donc élaboré de nouvelles
méthodes globales et rapides d’échantillonnage. La rapidité de ces méthodes est due à leur
imprécision, du fait qu'elles ne nécessitent pas une identification à l'espèce. Deux types de
méthodes sont à différencier, les méthodes d’échantillonnage dites « taxonomiques », et celles
dites « non taxonomiques ». Ainsi, alors que les méthodes taxonomiques identifient les
espèces vivantes et ont pour but d'en faire un inventaire complet, certaines d'entre elles
s’arrêtent à l'identification d’un ordre supérieur à celui de l’espèce, comme par exemple le
genre ou la famille. Au sein des méthodes dites « non taxonomiques », nous distinguerons les
méthodes « para-taxonomiques » des « méta-méthodes ». La méthode RAS fait partie du
groupe des méta-méthodes. Cette distinction des méta-méthodes se fait sur le critère de la
prise de mesure qui, au lieu de se faire à l'échelle de l'individu, se fait à l'échelle de la
collection d’individus.
Les méthodes taxonomiques : Classiquement, évaluer l’état de la biodiversité d’un site se fait
en identifiant toutes les espèces présentes sur ce site, espèces précédemment décrites sur des
critères taxonomiques. Identifier toutes les espèces nécessite un temps et une expertise
considérable. La plupart des études se concentrent sur un inventaire partiel d’espèces qui
peuvent être d’un intérêt particulier. C’est pour cette raison que l'utilisation des espèces
indicatrices est un sujet largement traité par la communauté scientifique (Jones & Eggleton,
2000; Pearson, 1994). Ces espèces peuvent être indicatrices d'un groupe plus large. Kerr et al.
(2000) montrent que les espèces des papillons semblent être de bonnes indicatrices de la
richesse spécifique des Hyménoptères en général. Cependant, trouver une espèce ou un
groupe d’espèces représentatif de la biodiversité en général (le plus souvent considérée
comme le nombre de toutes les espèces présentes sur le site) est une idée très controversée, du
fait de l'absence de corrélations entre différents groupes taxonomiques (Lawton et al., 1998).
Une autre méthode permettant un accès plus rapide, demandant des efforts moins
importants d’identification, consiste à identifier l’individu collecté jusqu’à un groupe
taxonomique supérieur à celui de l’espèce. Certaines études montrent une corrélation positive
entre la richesse spécifique et la richesse des genres ou familles (Gaston & Blackburn,1995,
Groc et al., 2010; Rosser & Eggleton, 2012). C'est par exemple le cas de l'étude de Mazaris et
al. (2008) qui montrent que la diversité au niveau des genres et des familles de groupes
d'oiseaux, d'amphibiens, de mammifères et de reptiles est un bon indicateur de la richesse

163
spécifique de ces mêmes groupes pour la région étudiée. Dans une autre étude, Mazaris et al.
(2010) montrent qu'il en est de même pour l'estimation de la richesse spécifique d'espèces
végétales. Cette analyse montre que cette estimation est également valide en ne considérant
qu'un certain nombre d'espèces dites « communes », réduisant encore l'effort
d'échantillonnage et le coût de l'inventaire. Cette méthode est très utile sur certains groupes
dont l’identification à l’espèce nécessite un travail complexe, comme par exemple un travail
de dissection en laboratoire.
Les méthodes « non-taxonomiques » : Comme leur nom l’indique, elles se basent sur
l'absence de connaissances taxonomiques. Deux types de méthodes, ayant le point commun de
ne pas identifier les individus, seront discutés ici : les méthodes para-taxonomiques dont la
prise de mesure se fait au niveau de l’individu (comme c'est la cas pour les méthodes
d’échantillonnage traditionnelles), et les méta-méthodes (méta-acoustique, méta-génomique et
méta-imagerie) dont la prise de mesure se fait au niveau d'une collection d'individus. C’est
donc l’unité de description qui définira une méthode en tant que méta-méthode.
La méthode para-taxonomique est une méthode de classement des individus collectés
selon des critères morphologiques qui ne suivent pas toujours les critères taxonomiques. Les
individus sont donc classés selon des RTUs (Recognisable Taxonomic Units) appelées
également « morpho-species ». Les résultats de cette méthode ont été comparés aux résultats
des méthodes d’identification taxonomique à l’espèce par, entre autres, Oliver & Beattie
(1996) et Ward & Stanley (2004). Les auteurs s’accordent pour dire que les résultats
dépendent du groupe considéré mais que cette méthode semble acceptable pour comparer des
niveaux de diversité entre sites. Krell (2004) est très critique sur l'utilisation de cette méthode
et met en avant la difficulté de mettre en place une nomenclature fiable de morpho-espèce.
Selon Krell (2004), la para-taxonomie n'est pas une méthode scientifique puisqu'elle ne
respecte pas les principes de vérifiabilité et de reproductibilité. En effet, le classement des
individus dans tel ou tel morpho-groupe ne suit pas des critères définis au préalable. Ainsi la
vérification est impossible car aucune référence n'existe et les critères de classement,
dépendants du chercheur, ne permettent pas la reproductibilité du travail. Face à ce problème,
l'auteur prévient du danger de prendre cette méthode comme un estimateur du nombre
d'espèces taxonomiques, et n'admet l'utilité de cette méthode que comme une étude
préliminaire dont les résultats doivent être systématiquement confirmés ou corrigés par une
étude taxonomique classique. Ainsi, malgré les limites de cette méthode, la para-taxonomie

164
peut être envisagée pour une étude générale de comparaison de niveau de diversité, mais doit
être complétée par une étude taxonomique pour une estimation du nombre d'espèces.
Les méta-méthodes sont celles qui nous intéressent particulièrement puisque la
méthode RAS en fait partie. Prenons l'exemple de trois méta-méthodes : la méta-imagerie, la
méta-génomique et la méta-acoustique. Ces trois méthodes ont en commun de prendre une
mesure sur une collection d'individus en même temps, individus appartenant potentiellement à
différentes espèces. A différencier des méta-analyses, les méta-méthodes se placent au niveau
écologique de la communauté. La méta-imagerie permet d'extraire des informations telles que
la classe des sols et végétations, des mesures d'hétérogénéité végétale et de productivité
primaire; variables pouvant être utilisées dans l'évaluation de biodiversité (Gillespie et al.,
2008). Cependant, même s'il est en déclin, le coût reste élevé pour l'obtention et le traitement
des images. Les outils informatiques spécialisés nécessaires au traitement des images ne sont
pas toujours faciles d'accès, et des données précises, nécessaires à la validation les
informations issues de l'analyse des images, sont manquantes (Turner et al., 2003). Sur ce
point, un parallèle peut être fait avec la méta-acoustique. Si le traitement des données n'est pas
une limite en termes de coût, les approches méta-acoustiques nécessiteraient d'être validées
par un grand nombre de données issues d'inventaires taxonomiques. La méta-génomique a du
faire face aux problématiques de l'étude des microorganismes que sont les virus et les
bactéries. Ces organismes sont généralement non cultivables, soit parce que les conditions de
culture ne sont pas connues, soit parce qu’ils résistent aux tentatives de culture (Baussand &
Carbone, 2006). Face à cette difficulté, la méta-génomique propose de regarder directement
l’ensemble des séquences d’ADN extraites de communautés prélevées dans l’environnement,
appelé le méta-génome. Les nouveaux outils de code barre à ADN (DNA-barcoding)
permettent d'aller jusqu'à l'identification spécifique des organismes dont les molécules d'ADN
sont extraites (Valentini et al., 2008). Cette identification nécessite une base de données de
référence. Ainsi la méta-génomique permet, à partir d'une collection d'individus, d'avoir accès
à une collection d'ADN que l'on peut isoler, puis à une liste d'espèces correspondante à cette
collection. De la même façon, la méta-acoustique permet à partir d'une collection d'individus
d'avoir accès à une collection de sons; mais ces sons sont difficiles à isoler avec les méthodes
actuelles. La difficulté d'isolation des séquences acoustiques et la limitation en terme de
banque de données de références ne permettent pas, à ce jour, l'accès à l'identification des
espèces à l'origine de sons issus d'un signal acoustique de communauté animale.

165
V.4 Perspectives pour l'analyse et le suivi de la biodiversité
La méthode RAS est une méthode passive, multi-taxons, dont l'échantillonnage est
automatisable et répétable, rapide et permettant un traitement large de données et sur des
échelles d'études temporelles et spatiales larges. Les perspectives de recherche seront
associées d'une part aux limites méthodologiques énoncées plus en amont, et d'autre part à la
possibilité d'utiliser la dissimilarité acoustique pour mettre en évidence des différences de
composition de communautés.
Les limitations majeures qui ont été présentées sont de deux types: la capacité de la mesure
d'hétérogénéité à représenter une diversité des sons (détectabilités des sons, indices
mathématiques) et la partie non automatique de sélections des fichiers utilisables pour
l'analyse. Il serait intéressant de travailler à la recherche d'indices mathématiques travaillant
sur le spectrogramme et qui ne seraient pas des indices de dissimilarité mesurant les
différences deux à deux entre les points des spectres, différence « point à point ». Les futures
recherches devront donc, en plus de l'automatisation de l'échantillonnage déjà acquise,
apporter des solutions tendant vers une automatisation de la mesure. Comme nous l'avons dit
précédemment, il parait impératif d'équiper les enregistreurs automatiques de petites stations
météorologiques autonomes afin de faciliter la sélection des fichiers non bruités. Le traitement
du bruit ambiant reste une des principales perspectives de recherche. Enfin, la caractérisation
acoustique du milieu dans lequel les enregistrements sont faits doit être une priorité de
recherche. Une étude est en cours au laboratoire LRPC (Laboratoire régional des ponts et
chaussées) de Strasbourg sous la direction de Guillaume Dutilleux. L'objectif de cette étude
est de caractériser l'impact des troncs d'arbres sur la propagation d'un signal sonore. Ces
études sont nécessaires pour envisager de maîtriser les biais d'échantillonnage de différents
types d'habitats comme c'est le cas pour l'étude menée dans le chapitre 4.
En comparaison des autres méta-méthodes, il est apparu que la RAS, ou méta-
acoustique, partageait quelques limitations avec la méta-imagerie, comme celle d'avoir des
données fiables de terrain pour une validation des mesures. Il serait intéressant de coupler un
échantillonnage acoustique à des données de terrain relevées de manière indépendante de
l'acoustique. De manière pratique et afin d'avoir le plus de données possible, il serait
intéressant de s'associer avec des projets d'inventaire déjà mis en place. Un projet se
développe dans ce sens en Nouvelle-Calédonie (Thèse de Jérémy Anso, IRD-MNHN, dirigée

166
par Laure Desutter-Grandcolas). Celui-ci a pour objectif de comparer des données d'inventaire
de grillons avec les données acoustiques dans le but d'étudier la mise en place d'un suivi des
populations de grillons par des méthodes acoustiques. L'étude est donc centrée sur un groupe
taxonomique particulier. Ce projet nécessitera donc de s'interroger sur la caractérisation des
sons et l'isolement des sons. L'isolement des sons dans un enregistrement peut être
problématique surtout en ce qui concerne les superpositions temporelles et spectrales. Une
piste de recherche consisterait à ajouter une 4ème dimension dans l'étude acoustique, l'espace.
Des travaux de localisation de source dans l'espace utilisant une série de microphones existent
(Huetz & Aubin, 2012) et pourraient être couplés avec une analyse méta-acoustique. Ce
premier pas vers l'isolation des sons dans un enregistrement pourrait être suivi d'une mesure
de dissimilarité entre les sons pour obtenir une mesure globale de la diversité acoustique,
pondérant les sons répétés.
La méta-génomique et le barcoding sont un exemple des potentialités des méta-
méthodes utilisant des bases de données pour séparer et identifier les données issues de ces
méta-méthodes. L'identification à l'espèce à partir de l'enregistrement d'un signal nécessiterait
l'utilisation de bases de données acoustiques très riches. Cependant, bien que les sonothèques
se développent, la couverture pour un grand nombre d'espèces, et surtout pour les espèces
tropicales, est pauvre (Ranft, 2004). L'enrichissement des sonothèques, dont celle du Muséum
national d'Histoire naturelle de Paris, pourra donc ouvrir de nouvelles possibilités dans le
traitement des enregistrements acoustiques.
Une partie des travaux présentés ici montre que la diversité acoustique pourrait être utilisée
pour identifier des différences dans la composition des communautés. L’étude menée en
Nouvelle-Calédonie a mis en évidence la potentialité de l’utilisation de la méthode globale,
non pas pour mesurer le niveau de biodiversité, mais pour mettre en évidence des différences
acoustiques entre des sites qui ont le même niveau de biodiversité α . Cet exemple des points
chauds de biodiversité est particulièrement approprié pour mettre en évidence l’importance
d’une telle étude. Ces zones sont très sensibles et ont un niveau de richesse spécifique très
élevé. Des mesures sont prises pour mener des missions de conservation, mais concrètement
ces régions ne peuvent pas être mises sous cloche et des choix de sites prioritaires pour la
conservation doivent être faits à petite échelle. Dans ce cas, la complémentarité des sites est
un critère décisif. Ces résultats restent néanmoins préliminaires et devront être confirmés sur
un plus grand nombre de sites. A terme, il faudra savoir si cette méthode peut mettre en
évidence des sites plus proches que d’autres sur le plan de la biodiversité, afin de mettre en

167
évidence de potentielles complémentarités de sites. L'objectif final serait de pouvoir utiliser la
méta-acoustique pour identifier des différences acoustiques entre des sites sans a priori.
En plus d'une caractérisation des sites, la dissimilarité temporelle d'un même site
pourrait être utilisée dans des programmes de suivi de la biodiversité. En effet, l'utilisation de
la méta-acoustique pourrait permettre d'identifier des changements d'activité acoustique dans
le temps sur un site. Ces changements peuvent être de nature biologique (comportement
saisonnier) mais également être le reflet de modifications de la fréquentation du site par les
espèces chanteuses (moins de fréquentation, renouvellement des espèces fréquentant le site).
De plus, le suivi d'un même point permet d'éliminer certain biais de détectabilité dus à la
différence d'habitats (Buckland et al., 2005). Enfin, l'explorations de milieu difficile d'accès
ou fragile est un avantage certain de la méthode RAS. Deux projets sont actuellement en
cours, pour évaluer la potentialité de cette méthode pour des études en canopée et dans les
milieux aquatiques terrestres.
Pour conclure, l'objectif général de cette thèse était de « mieux appréhender cette approche
d’acoustique globale d'un point de vue méthodologique afin de connaître les possibles
applications de cette méthode pour estimer, suivre et mieux comprendre la distribution
spatiale et temporelle de la biodiversité ». Le travail effectué a permis d'apporter des éléments
de réflexion sur la définition de la diversité acoustique d'une communauté (CAD), en tant
qu'aspect de la diversité biologique, et contenant des informations sur d'autres aspects de
biodiversité. La mesure de cette diversité par la méthode RAS est une approche innovante. Le
potentiel de l'indice de dissimilarité pour identifier des différences de compositions de
communauté a été mis en avant permettant d'ouvrir des possibilités d'exploration de milieux
fragiles comme les points chauds de biodiversité, avec l'exemple de l'étude faite en Nouvelle-
Calédonie, ou de l'impact de l'utilisation d'espèces exotiques dans les agro-forêts, avec
l'exemple de l'étude en Inde. Des pistes d'améliorations méthodologiques ont été proposées et
leur exploration devrait permettre, à terme, de fournir un outil utilisable dans de nombreuses
conditions acoustiques. Il existe deux approches complémentaires de méta-acoustique : la
RAS et les méthodes espèces-centrées qui cherchent à isoler et identifier les sons dans le
signal acoustique d'une communauté. Le choix de l'utilisation de l'une ou l'autre de ces
méthodes doit dépendre de la question scientifique posée et de la prise en compte des
avantages et inconvénients de chacune des méthodes. Le parallèle fait avec les avancées de la
méta-génomique ouvre des perspectives sur l'utilisation de la méta-acoustique pour une
estimation passive de la biodiversité.

168

169
REFERENCES BIBLIOGRAPHIQUES
Acevedo, M. A. & Villanueva-Rivera, L. J. (2006) Using automated digital recording systems as effective tools for the monitoring of birds and amphibians. Wildlife Society Bulletin, 34, 211-214.
Acevedo, M. A., Corrada-Bravo, C. J., Corrada-Bravo, H., Villanueva-Rivera, L. J. & Aide,
T. M. (2009) Automated classification of bird and amphibian calls using machine learning: a comparison of methods. Ecological Informatics, 4, 206-214.
Alatalo, R. V., Glynn, C. & Lundberg, A. (1990) Singing rate and female attraction in the
pied flycatcher - an experiment. Animal Behaviour, 39, 601-603. Altieri, M.A., 1999. The ecological role of biodiversity in agroecosystems. Agriculture
Ecosystems & Environment, 74, 19-31. Anderson, S. E., Dave, A. S. & Margoliash, D. (1996) Template-based automatic recognition
of birdsong syllables from continuous recordings. Journal of the Acoustical Society of America, 100, 1209-1219.
Anderson, M.J. & Willis, T.J. (2003) Canonical analysis of principal coordinates: a useful
method of constrained ordination for ecology. Ecology, 84, 511–525. Araujo, M. B., Thuiller, W. & Pearson, R. G. (2006) Climate warming and the decline of
amphibians and reptiles in Europe. Journal of Biogeography, 34, 192-192. Bali, A., Dolia, J. & Anand, M. O. (2006) An assessment of biodiversity in coffee plantations:
A report on a tri-taxa study in coffee plantations in Chikmagalur. WCS-India. Disponible en ligne: http://msc.wcsindia.org/COFFEE%20REPORT%20Final.pdf
Balmford, A., Crane, P., Dobson, A., Green, R. E. & Mace, G. M. (2005) The 2010 challenge:
data availability, information needs and extraterrestrial insights. Philosophical Transactions of the Royal Society B-Biological Sciences, 360, 221-228.
Bardeli, R., Wolff, D., Kurth, F., Koch, M., Tauchert, K. H. & Frommolt, K. H. (2010).
Detecting bird sounds in a complex acoustic environment and application to bioacoustic monitoring. Pattern Recognition Letters, 31, 1524-1534.
Barataud, M. (2012) Ecologie acoustique des Chiroptères d'Europe. Biotope édition et
publications du Muséum national d'Histoire naturelle, Paris. Barber, J. R., Burdett, C. L., Reed, S. E., Warner, K. A., Formichella, C., Crooks, K. R.,
Theobald, D. M. & Fristrup, K. M. (2011) Anthropogenic noise exposure in protected natural areas: estimating the scale of ecological consequences. Landscape Ecology, 26, 1281-1295.
Basset, Y., Novotny, V., Miller, S. E. & Pyle, R. (2000) Quantifying biodiversity: experience
with parataxonomists and digital photography in Papua New Guinea and Guyana. Bioscience, 50, 899-908.

170
Bhagwat, S. A., Willis, K. J., Birks, H. J. B. & Whittaker, R. J. (2008) Agroforestry: a refuge
for tropical biodiversity? Trends in Ecology & Evolution, 23, 261-267. Blumstein, D. T., Mennill, D. J., Clemins, P., Girod, L., Yao, K., Patricelli, G., Deppe, J. L.,
Krakauer, A. H., Clark, C., Cortopassi, K. A., Hanser, S. F., McCowan, B., Ali, A. M., Kirschel, A. N. G. (2011) Acoustic monitoring in terrestrial environments using microphone arrays: applications, technological considerations and prospectus. Journal of Applied Ecology, 48, 758-767.
Boncoraglio, G. & Saino, N. (2007) Habitat structure and the evolution of bird song: a meta-
analysis of the evidence for the acoustic adaptation hypothesis. Functional Ecology, 21, 134-142.
Bossu, A. & Charron, F. (1998) Le chant des oiseaux. Sang de la terre, Paris, 1CD. Bossu, A. & Charron, F. (2004) Guide des chants d'oiseaux d'Europe occidentale :
Description et comparaison des chants et des cris. Delachaux et Niestlé, Neuchâtel, Switzerland, 2CDs.
Boulard, M. (2006) Acoustic Signals, Diversity and Behaviour of cicadas (Cicadae,
Hemiptera).In: Drosopoulos S, Claridge MF, editors. Insect sounds and communication, physiology, behaviour, ecology and evolution. Boca raton: Taylor and Francis. pp. 331-350.
Bradbury, J. W. & Vehrencamp, S. L. (1998) Principles of Animal communication. Sinauer
Associates, Inc., Sunderland. Brandes, T. S. (2008) Automated sound recording and analysis techniques for bird surveys
and conservation. Bird Conservation International, 18, 163-173. Brandes, T. S., Naskrecki, P. & Figueroa, H. K. (2006) Using image processing to detect and
classify narrow-band cricket and frog calls. Journal of the Acoustical Society of America, 120, 2950-2957.
Brehm, J. M., Maxted, N., Martins-Loucao, M. A. & Ford-Lloyd, B.V. (2010) New
approaches for establishing conservation priorities for socio-economically important plant species. Biodiversity and Conservation, 19, 2715-2740.
Bridges, A. S. & Dorcas, M. E. (2000) Temporal variation in anuran calling behavior:
Implications for surveys and monitoring programs. Copeia, 2 , 587-592. Briefer, E., Rybak, F. & Aubin, T. (2011) Microdialect and group signature in the song of the
skylark alauda arvensis. Bioacoustics, 20, 219-233. Brooks, D.R. & McLennan, D.A. (1991) Phylogeny, ecology, and behavior: a research
program in comparative biology. University of Chicago Press, Chicago. Brown, T. J., & Handford, P. (2003) Why birds sing at dawn: the role of consistent song
transmission. Ibis, 145, 120-129.

171
Brown, J. C., Hodgins-Davis, A. & Miller, P. J. O. (2006) Classification of vocalizations of
killer whales using dynamic time warping. Journal of the Acoustical Society of America, 119, l34-l40.
Buckland, S. T., Magurran, A. E., Green, R. E. & Fewster, R. M. (2005) Monitoring change
in biodiversity through composite indices. Philosophical Transactions of the Royal Society B-Biological Sciences, 360, 243-254.
Burke, B. B. (1995) Ondes et ondelettes. Belin, Paris. Campanha, M. M., Santos, R. H. S., de Freitas, G. B., Martinez, H. E. P., Garcia, S. L. R. &
Finger, F. L. (2005) Growth and yield of coffee plants in agroforestry and monoculture systems in Minas Gerais, Brazil. Agroforestry Systems, 63, 75-82.
Cardoso, G.C. & Price, T.D. (2010) Community convergence in bird song. Evolutionary
Ecology, 24, 447-461. Catchpole, C. K. & Slater, P. J. B. (1995) Bird song: biological themes and variations.
Cambridge University Press, Cambridge, UK. Cavender-Bares, J., Ackerly, D. D., Baum, D. A., Bazzaz, F. A. (2004) Phylogenetic
overdispersion in Floridian oak communities. The American Naturalist 163, 823-843. Champely, S. & Chessel, D. (2002) Measuring biological diversity using Euclidean metrics.
Environmental and Ecological Statistics, 9, 167-177. Chao, A., Chiu, C. H. & Jost, L. (2010) Phylogenetic diversity measures based on Hill
numbers. Philosophical Transactions of the Royal Society B-Biological Sciences, 365, 3599-3609.
Chapin III, F. S., Zavaleta, E. S., Eviner, V. T., Naylor, R. L., Vitousek, P. M., Reynolds, H.
L., Hooper, D. U., Lavorel, S., Sala, O. E., Hobbie, S. E., Mack, M. C. & Diaz, S. (2000) Consequences of changing biodiversity. Nature, 405, 234-242.
Chave, J., Chust, G. & Thébaud C. (2007) The importance of phylogenetic structure in
biodiversity studies. In Scaling Biodiversity (eds. D. Storch, P. Marquet, J.H. Brown), pp. 151-167. Santa Fe Institute Editions, Santa Fe.
Chek, A. A., Bogart, J. P. & Lougheed, S. C. (2003) Mating signal partitioning in multi-
species assemblages: a null model test using frogs. Ecology Letters, 6, 235-247. Chesmore, E. D. (2001) Application of time domain signal coding and artificial neural
networks to passive acoustical identification of animals. Applied Acoustics, 62, 1359-1374.
Clemins, P. J., Johnson, M. T., Leong, K. M. & Savage, A. (2005) Automatic classification
and speaker identification of African elephant (Loxodonta africana) vocalizations. Journal of the Acoustical Society of America, 117, 956-963.

172
Clough, Y., Barkmann, J., Juhrbandt, J., Kessler, M., Wanger, T. C., Anshary, A., Buchori, D., Cicuzza, D., Darras, K., Putra, D. D., Erasmi, S., Pitopang, R., Schmidt, C., Schulze, C. H., Seidel, D., Steffan-Dewenter, I., Stenchly, K., Vidal, S., Weist, M., Wielgoss, A. C. & Tscharntke, T. (2011) Combining high biodiversity with high yields in tropical agroforests. Proceedings of the National Academy of Sciences of the United States of America, 108, 8311-8316.
Connor, E. F., Li, St. & Li, Se (2012) Automating identification of avian vocalizations using
time–frequency information extracted from the Gabor transform. Journal of the Acoustical Society of America,132, 507-517.
Cornwell, W. K., Schwilk, D. W. & Ackerly, D. D. (2006) A trait-based test for habitat
filtering: Convex hull volume. Ecology, 87, 1465-1471. Cox, C. B. & Moore, P. D. (2005) Biogeography: an ecological and evolutionary approach.
Oxford: Blackwell Science Ltd. Cramp, S., Simmons, K. E. L., Snow, D. W. & Perrins, C. M. (2004) The Birds of the Western
Palearctic; interactive. BirdGuides, Sheffield, UK. Dabelsteen T. & Mathevon N. (2002) Why do songbirds sing intensively at dawn? A test of
the acoustic transmission hypothesis. Acta Ethologica, 4, 65-72. Dalerum, F., Cameron, E. Z., Kunkel, K. & Somers, M. J. (2009) Diversity and depletions in
continental carnivore guilds: implications for prioritizing global carnivore conservation. Biology Letters, 5, 35-38.
Davies, C. E, Moss, D. & O'Hill, M. (2004) EUNIS habitat classification revised 2004;
Report to the European Topic Centre on Nature Protection and Biodiversity. European Environment Agency, Paris.
Davis, K.E. (2008) Reweaving the tapestry: a supertree of birds. PhD thesis, University of
Glasgow. URL: http://linnaeus.zoology.gla.ac.uk/~rpage/birdsupertree/index.php. Dawson, D. K. & Efford, M. G. (2009) Bird population density estimated from acoustic
signals. Journal of Applied Ecology, 46, 1201-1209. Dechant, B. (2012) Analyse et suivi de la biodiversité animale tropicale par la bioacoustique:
le cas des agro-forêts des Western Ghats (Inde). Rapport de stage Master 2 (OSEB), Muséum national d'Histoire naturelle, Paris.
De Groot, D., Ramakrishnan, P. S., van de Berg, A., Kulenthran, T., Muller et al. (2005)
Cultural and Amenity Services. In: Conditions and Trends Assessment. Millennium Ecosystem Assessment ( eds R. Hassan, R. Scholes, N. Ash), pp. 455-476. Island Press, Washington.
Depraetere, M., Pavoine, S., Jiguet, F., Gasc, A., Duvail, S. & Sueur, J. (2012) Monitoring
animal diversity using acoustic indices: implementation in a temperate woodland. Ecological Indicators, 13, 46-54.

173
Deroussen, F. (2001) Oiseaux des jardins de France. Nashvert Production, Charenton, France, 1CD.
Deroussen, F. & Jiguet, F. (2006) La sonothèque du Muséum: Oiseaux de France, les
passereaux. Nashvert production, Charenton, France. Deroussen, F. (2011) Sound Library of the national Museum of natural History of Paris.
National Museum of natural History, Paris. http://museumedia.mnhn.fr/content/sons. Desmoulins, F. & Barré, N. (2005) Oiseaux des forêts sèches de Nouvelle-Calédonie. Païta:
IAC-Programme forêt sèche. Desutter-Grandcolas, L. & Robillard, T. (2006) Phylogenetic systematics and evolution of
Agnotecous in New Caledonia (Orthoptera : Grylloidea, Eneopteridae). Systematic Entomology, 31, 65-92.
Devictor, V., Mouillot, D., Meynard, C., Jiguet, F., Thuiller, W. & Mouquet, N. (2010)
Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world. Ecology Letters, 13, 1030-1040.
Deza E. & Deza M. M. (2009) Encyclopedia of distances. Springer. Diamond, J., & Case T. J. (1986) Community Ecology. Harper and Row, New York. Díaz, S., D. Tilman, J. Fargione, F. S. Chapin, I., R. Dirzo, et al. (2005) Biodiversity
Regulation of Ecosystem Services. In Ecosystems and human well-being: current state and trends (eds H. Hassan, R. Scholes, N. Ash), pp. 297–329. Island Press, Washington.
Dickinson, J. L., Zuckerberg, B. & Bonter, D. N. (2010) Citizen science as an ecological
research tool: challenges and benefits. Annual Review of Ecology, Evolution, and Systematics, 41, 149-172.
Diwakar, S. & Balakrishnan, R. (2007a) Vertical stratification in an acoustically
communicating ensiferan assemblage of a tropical evergreen forest in southern India. Journal of Tropical Ecology, 23, 479-486.
Diwakar, S. & Balakrishnan, R. (2007b) The assemblage of acoustically communicating
crickets of a tropical evergreen forest in southern India: call diversity and diel calling patterns. Bioacoustics, 16, 113-135.
Dray, S. & Dufour, A. B (2007) The ade4 package: implementing the duality diagram for
ecologists. Journal of Statistical Software, 22, 1-20. Duelli, P. & Obrist, M. K. (2003) Biodiversity indicators: the choice of values and measures.
Agriculture Ecosystems & Environment, 98, 87-98. Emerson, B. C. & Gillespie, R. G. (2008) Phylogenetic analysis of community assembly and
structure over space and time. Trends in Ecology and Evolution, 23, 619-630.

174
Erbe, C. & King, A. R. (2008) Automatic detection of marine mammals using information entropy. Journal of the Acoustical Society of America, 124, 2833-2840.
Escoufier, P. (1973) Le traitement des variables vectorielles. Biometrics, 29, 751-760. Espeland, M. & Johanson, K. A. (2010) The effect of environmental diversification on species
diversification in New Caledonian caddisflies (Insecta: Trichoptera: Hydropsychidae). Journal of Biogeography, 37, 879-890.
Ey, E. & Fischer, J. (2009) The "Acoustic Adaptation Hypothesis" - a Review of the Evidence
from Birds, Anurans and Mammals. Bioacoustics, 19, 21-48. Ewing, A. E. (1989) Arthropod bioacoustics, neurobiology and behaviour. Edinburgh
University Press, Edinburgh. Faith, D. P. (1992) Conservation Evaluation and Phylogenetic Diversity. Biological
Conservation, 61, 1-10. Faith, D. P. & Walker, P. A. (1996) How do indicator groups provide information about the
relative biodiversity of different sets of areas? On hotspots, complementarity and pattern-based approaches. Biodiversity Letters, 3, 18-25.
Farina, A. & Morri, D. (2008) Source-sink e eco-field: ipotesi ed evidenze sperimen- tali. Atti
del X congresso nazionale della SIEP-IALE. Ecologia e governance del paesaggio: esperienze e prospettive. Bari, 365–372.
Farina, A &Pieretti, N. (2012) The soundscape ecology: A new frontier of landscape research
and its application to islands and coastal systems. Journal of Marine and Island Culture, 1, 21-26
Farina, A., Pieretti, N. & Piccioli, L. (2011) The soundscape methodology for long-term bird
monitoring: A Mediterranean Europe case-study. Ecological Informatics, 6, 354-363. Ficken, R. W., Popp, J. W. & Matthiae, P. E. (1985) Avoidance of acoustic interference by
ovenbirds. Wilson Bulletin, 97, 569-571. Fischetti, A. (2003) Initiation à l'acoustique, cours et exercices, 2nde édition. Belin, Paris. Fletcher, N. H. (2004) A simple frequency-scaling rule for animal communication. Journal of
the Acoustical Society of America, 115, 2334-2338. Fletcher, N. H. (2007) Animal Bioacoustics. Springer Handbook of Acoustics (ed. by T.D.
Rossing), pp. 473-490. Springer, Stanford. Fleishman, E., Thomson, J. R., Mac Nally, R., Murphy, D. D. & Fay, J. P. (2005) Using
indicator species to predict species richness of multiple taxonomic groups. Conservation Biology, 19, 1125-1137.

175
Forrest, T. G., Miller, G. L. & Zagar, J. R. (1993) Sound propagation in shallow water: Iimplications for acoustic communication by aquatic animals. Bioacoustics, 4, 259-270.
Forrest, T. G., (1988) Using insect sounds to estimate and monitor their populations. Florida
Entomologist, 71, 416-426. Ganchev, T., Potamitis, I. & Fakotakis, N. (2007) Acoustic monitoring of singing insects.
IEEE International Conference on Acoustics, Speech, and Signal Processing, 4, 721-724.
Garcia, C. A., Bhagwat, S. A., Ghazoul, J., Nath, C. D., Nanaya, K. M., Kushalappa, C. G.,
Raghuramulu, Y., Nasi, R. & Vaast, P. (2010) Biodiversity Conservation in Agricultural Landscapes: Challenges and Opportunities of Coffee Agroforests in the Western Ghats, India. Conservation Biology, 24, 479-488.
Garcia-Rutledge, E. J. & Narins, P. M. (2001) Shared acoustic resources in an old world frog
community. Herpetologica, 57, 104-116. Gaston, K. J. (2000) Global patterns in biodiversity. Nature, 405, 220-227. Gaston, K. J. & Blackburn, T. M. (1995) Mapping biodiversity using surrogates for species
richness: Macro-scales and New World birds. Proceedings of the Royal Society B-Biological Sciences, 262, 335-341.
GBIF (2012) portail de données en ligne : http://data.gbif.org/welcome.htm. Gerhardt, H. C. & Huber, F. (2002) Acoustic communication in insects and anurans, common
problems and diverse solutions. University of Chicago, Chicago. Gess, A., Schneider, D. M., Vyas, A. & Woolley, S. M. N. (2011) Automated auditory
recognition training and testing. Animal Behaviour, 82, 285-293. Gillespie, T. W., Foody, G. M., Rocchini, D., Giorgi, A. P. & Saatchi, S. (2008) Measuring
and modelling biodiversity from space. Progress in Physical Geography, 32, 203-221. Gobbi, J. A. (2000) Is biodiversity-friendly coffee financially viable? An analysis of five
different coffee production systems in western El Salvador. Ecological Economics, 33, 267-281.
Gottsberger, B. & Gruber, E. (2004) Temporal partitioning of reproductive activity in a
neotropical anuran community. Journal of Tropical Ecology, 20, 271-280. Gower, J. C. (1971) A general coefficient of similarity and some of its properties. Biometrics,
27, 857-871. Grandcolas, P. (1993) The origin of biological diversity in a tropical cockroach lineage - a
phylogenetic analysis of habitat choice and biome occupancy. Acta Oecologica, 14, 259-270.

176
Grandcolas, P. (1998) Phylogenetic analysis and the study of community structure. Oikos, 82, 397-400.
Grandcolas, P. (2009) Zoologia Neocaledonica 7: Biodiversity studies in New Caledonia.
Mémoires du Muséum national d'Histoire naturelle, 198, 1-440. Grandcolas, P., Murienne, J., Robillard, T., Desutter-Grandcolas, L., Jourdan, H., Guilbert, E.
& Deharveng, L. (2008) New Caledonia: A very old Darwinian island? Philosophical Transactions of the Royal Society B: Biological Sciences, 363, 3309-3317.
Greenfield, M. D (2002) Signalers and receivers. Mechanisms and evolution of arthropod
communication. Oxford University Press, New York. Greig-smith, P. W. (1982) Song rates and parental care by individual male stonechats
(Saxicola torquata). Animal Behaviour, 30, 245-252. Groc, S., Delabie, J. H. C., Longino, J. T., Drivel, J., Majer, J. D., Vasconcelos, H. L. &
Dejean, A. (2010) A new method based on taxonomic sufficiency to simplify studies on Neotropical ant assemblages. Biological Conservation, 143, 2832-2839.
Han, N. C., Muniandy, S. V. & Dayou, J. (2011) Acoustic classification of Australian anurans
based on hybrid spectral-entropy approach. Applied Acoustics, 72, 639-645. Hammer, O. & Barrett, N. (2001) Techniques for studying the spatio-temporal distribution of
animal vocalizations in tropical wet forests. Bioacoustics, 12, 21-35. Hardy, O. J. (2008) Testing the spatial phylogenetic structure of local communities: statistical
performances of different null models and test statistics on a locally neutral community. Journal of Ecology, 96, 914-926.
Hardy, O. & Pavoine, S. (2012) Assessing phylogenetic signal with measurement error: a
comparison of Mantel tests, Blomberg et al.’s K and phylogenetic distograms. Evolution. In press.
Heller, K. G. & Von Helversen, D. (1993) Calling Behavior in Bush-Crickets of the Genus
Poecilimon with Differing Communication-Systems (Orthoptera, Tettigonioidea, Phaneropteridae). Journal of Insect Behavior, 6, 361-377.
Henwood, K., & Fabrick, A. (1979) A quantitative analysis of the dawn chorus: temporal
selection for communicatory optimization. American Naturalist, 114, 260-274. Hijmans, R. J., Williams, E. & Vennes, C. (2011) Geosphere: Spherical Trigonometry. R
package version 1.2-26. http://CRAN.R-project.org/package=geosphere. Holdaway, R. J. & Sparrow, A. D. (2006) Assembly rules operating along a primary riverbed-
grassland successional sequence. Journal of Ecology, 94, 1092-1102. Hopp, S. L. , Owren, M. J. & Evans, C. S. (1998) Animal acoustic communication, sound
analysis and research methods. Springer-Verlag, Berlin.

177
Huetz, C. & Aubin, T. (2012) Bioacoustics approaches to locate and identify aniamals in terrestrial environments. In Sensors for ecology (eds by J-F. Le Gaillard, J-M. Guarini & F. Gaill), pp. 83-96. CNRS, Paris.
Husson, F., Josse, J., Le, S. & Mazet, J. (2011) FactoMineR: multivariate exploratory data
analysis and data mining with R. R package version 1.16. Hutchinson, J. M. C. (2002) Two explanations of the dawn chorus compared : how
monotonically changing light levels favour a short break from singing. Animal Behaviour, 64, 527-539.
Isaac, N. J. B., Turvey, S. T., Collen, B., Waterman, C. & Baillie, J. E. M. (2007) Mammals
on the EDGE: Conservation Priorities Based on Threat and Phylogeny. PLoS ONE, 2, e296.
Jaccard, P. (1901) Étude comparative de la distribution florale dans une portion des Alpes et
des Jura. Bulletin de la Société Vaudoise des Sciences Naturelles, 37, 547-579. Jaffré, T., Munzinger, J. & Lowry, P. P. (2010) Threats to the conifer species found on New
Caledonia’s ultramafic massifs and proposals for urgently needed measures to improve their protection. Biodiversity Conservation, 19, 1485-1502.
Jiguet, F., Devictor, V., Julliard, R. & Couvet, D. (2012) French citizens monitoring ordinary
birds provide tools for conservation and ecological sciences. Acta Oecologica, 44, 58-66. doi:10.1016/j.actao.2011.05.003.
Jones, D. T. & Eggleton, P. (2000) Sampling termite assemblages in tropical forests: testing a
rapid biodiversity assessment protocol. Journal of Applied Ecology, 37, 191-203. Joo, W., Gage, S. H. & Kasten, E. P. (2011) Analysis and interpretation of variability in
soundscapes along an urban-rural gradient. Landscape and Urban Planning, 103, 259-276.
Josse, J., Pages, J. & Husson, F. (2008) Testing the significance of the RV coefficient.
Computational Statistics & Data Analysis, 53, 82-91. Jost, L. (2007) Partitioning diversity into independent alpha and beta components. Ecology,
88, 2427-2439. Julliard, R., Clavel, J., Devictor, V., Jiguet, F. & Couvet, D. (2006) Spatial segregation of
specialists and generalists in bird communities. Ecology Letters, 9, 1237-1244. Julliard, R., Jiguet, F. & Couvet, D. (2004) Common birds facing global changes: what makes
a species at risk? Global Change Biology, 10, 148-154. Kazi-aoual, F., Hitier, S., Sabatier, R. & Lebreton, J.D. (1995) Refined approximations to
permutation tests for multivariate inference. Computational Statistics & Data Analysis, 20, 643-656.

178
Kerr, J. T., Sugar, A. & Packer, L. (2000) Indicator taxa, rapid biodiversity assessment, and nestedness in an endangered ecosystem. Conservation Biology, 14, 1726-1734.
Keshawa Murthy, K. R. & Yoganarasimhan, S. N. (1990) Flora of Coorg (Kodagu)
Karnataka, India. Vimsat Publishers, Bangalore. Kier, G., Kreft, H., Lee, T. M., Jetz, W., Ibisch, P. L. et al. (2009) A global assessment of
endemism and species richness across island and mainland regions. Proceedings of the National Academy of Sciences, 23, 9322-9327.
Kotze, D. J. & O'Hara, R. B. (2003) Species decline-but why? Explanations of carabid beetle
(Coleoptera, Carabidae) declines in Europe. Oecologia, 135, 138-48. Krause B. (1987) Bioacoustics, habitat ambience in ecological balance. Whole Earth Review,
57, 14–18. Krell, F. T. (2004) Parataxonomy vs. taxonomy in biodiversity studies - pitfalls and
applicability of 'morphospecies' sorting. Biodiversity and Conservation, 13, 795-812. Kullback, S. & Leibler, R. A. (1951) On information and sufficiency. Annals of Mathematical
Statistics, 22, 79-86. Laiolo, P., Vögeli, M., Serrano, D. & Tella, J. L. (2008) Song diversity predicts the viability
of fragmented bird populations. PloS ONE, 3, e1822. Langbauer, W. R. (2000) Elephant communication. Zoo Biology, 19, 425-445. Lawton, J. H., Bignell, D. E., Bolton, B., Bloemers, G. F., Eggleton, P., Hammond, P. M.,
Hodda, M., Holt, R. D., Larsen, T. B., Mawdsley, N. A., Stork, N. E., Srivastava, D. S. & Watt, A. D. (1998) Biodiversity inventories, indicator taxa and effects of habitat modification in tropical forest. Nature, 391, 72-76.
Lebrija-Trejos, E., Perez-Garcia, E. A., Meave, J. A., Bongers, F. & Poorter, L. (2010)
Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology, 91, 386-398.
Le Gaillard, J-F., Guarini, J-M. & Gaill, F. (2012) Sensors for ecology. CNRS, Paris. Legendre, P. & Anderson, M. J. (1999) Distance-based redundancy analysis: Testing
multispecies responses in multifactorial ecological experiments. Ecological Monographs, 69, 512-512.
Legendre, F., Robillard, T., Desutter-Grandcolas, L., Whiting, M.F. & Grandcolas, P. (2008)
Phylogenetic analysis of non-stereotyped behavioural sequences with a successive event-pairing method. Biological Journal of the Linnean Society, 94, 853-867.
Legendre, P., & Legendre, L. (1998) Numerical ecology (2nd ed.). Elsevier, Amsterdam.

179
Lellouch, L. (2012) Etude des liens entre diversités spécifique et acoustique de communautés d'oiseaux. Rapport de Stage M2 (CERSP-OSEB), Muséum national d'Histoire naturelle, Paris.
Leroy, Y. (1979) L'univers sonore animal. Gauthier-villars, Paris. Ligges, U. (2011) tuneR: Analysis of music. URL: http://r-forge.r-project.org/projects/tuner/. Lingoes, J. C. (1971) Some Boundary Conditions for a Monotone Analysis of Symmetric
Matrices. Psychometrika, 36, 195-203. Lovejoy, T. E. (1994) The quantification of biodiversity - an esoteric quest or a vital
component of sustainable development. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 345, 81-87.
Lovejoy, T. E. (1997) Biodiversity: What is it ? In Biodiversity II: understanding and
protecting our biological resources (eds. by ML Reaka-Kudla, DE Wilson & EO Wilson), pp. 15-24. Joseph Henry Press, Washington.
Luczkovich, J. J., Mann D. A. & Rountree R. A. (2008) Passive acoustic as a tool in fisheries
Science. Transactions of the American Fisheries Society, 137, 533-541. Lundy, M., Teeling, E., Boston, E., Scott, D., Buckley, D., Prodohl, P., Marneli, F. &
Montgomery, I. (2011) The shape of sound: elliptic fourier descriptors (Efd) discriminate the echolocation calls of Myotis Bats (M. Daubentonii, M. Nattereri and M. Mystacinus). Bioacoustics-the International Journal of Animal Sound and Its Recording, 20, 101-115.
Mace, G., Masundire, H. & Baillie, J. (2005) Biodiversity. In Ecosystems and human
wellbeing: current state and trends (eds by H. Hassan, R. Scholes, and N. J. Ash), pp. 79-115. Island Press, Washington, D.C., USA.
Magurran, A. (2004) Measuring biological diversity. Blackwell, Oxford. Malézieux E., Crozat Y., Dupraz C., Laurans M., Makowski D., Ozier-Lafontaine H., Rapidel
B., de Tourdonnet S. & Valantin-Morison M. (2009) Mixing plant species in cropping systems: concepts, tools and models. A review. Agronomy for Sustainable Development, 29, 43-62.
Mankin, R. W., Brandhorst-Hubbard, J., Flanders, K. L., Zhang, M., Crocker, R. L., Lapointe,
S. L., McCoy, C. W., Fisher, J. R. & Weaver, D. K. (2000) Eavesdropping on insects hidden in soil and interior structures of plants. Journal of Economic Entomology, 93, 1173-1182.
Mankin, R. W., Lapointe, S. L., & Franqui, R. A. (2001) Acoustic surveying of subterranean
insect populations in citrus groves. Journal of Economic Entomology, 94, 853-859. Manly, B. F. (1994) Multivariate Statistical Methods. A primer., Second edition. Chapman &
Hall, London.

180
Mann, D., Locascio, J., Coleman, F. & Koenig, C. (2009) Goliath grouper Epinephelus itajara sound production and movement patterns on aggregation sites. Endanger Species Research, 7, 229–236.
Mantel, N. (1967) The detection of disease clustering and a generalized regression approach.
Cancer Research, 27, 209–220. Margules, C. R. & Pressey, R. L. (2000) Systematic conservation planning. Nature, 405, 243-
253. Marques, T. A., Thomas, L., Ward, J., DiMarzio, N. & Tyack, P. L. (2009) Estimating
cetacean population density using fixed passive acoustic sensors: An example with Blainville's beaked whales. Journal of the Acoustical Society of America, 125, 1982-1994.
Martens, P., Rotmans, J. & de Groot, D. (2003) Biodiversity: luxury or necessity? Global
Environmental Change-Human and Policy Dimensions, 13, 75-81. Martin, J. P., Doucet, S. M., Knox, R. C. & Mennill, D. J. (2011) Body size correlates
negatively with the frequency of distress calls and songs of Neotropical birds. Journal of Field Ornithology, 82, 259-268.
Mathias, D., Thode, A., Straley, J. & Folkert, K. (2009) Relationship between sperm whale
(Physeter macrocephalus) click structure and size derived from videocamera images of a depredating whale (sperm whale prey acquisition). Journal of the Acoustical Society of America, 125, 3444-3453.
May, R. M. (1990) Taxonomy as Destiny. Nature, 347, 129-130. May, R. M. (2010) Tropical arthropod species, more or less? Science, 329, 41-42. May, R. M. (2011) Why worry about how many species and their loss? PLoS Biology, 9,
e1001130. Mazaris, A. D., Kallimanis, A. S., Sgardelis, S. P. & Pantis, J. D. (2008) Does higher taxon
diversity reflect richness of conservation interest species? The case for birds, mammals, amphibians, and reptiles in Greek protected areas. Ecological Indicators, 8, 664-671.
Mazaris, A. D., Kallimanis, A. S., Tzanopoulos, J., Sgardelis, S. P. & Pantis, J. D. (2010) Can
we predict the number of plant species from the richness of a few common genera, families or orders? Journal of Applied Ecology, 47, 662-670.
McNeely, J. A. (2004) Nature vs. nurture: managing relationships between forests,
agroforestry and wild biodiversity. Agroforestry Systems, 61, 155-165. Mennill, D. J., Burt, J. M., Fristrup, K. M. & Vehrencamp, S. L. (2006) Accuracy of an
acoustic location system for monitoring the position of duetting songbirds in tropical forest. Journal of the Acoustical Society of America, 119, 2832-2839.

181
Mercado III, E. & Kuh, A. (1998) Classification of humpback whale vocalizations using a self-organizing neural network. Neural Networks for Signal Processing Xii, 2, 1584-1589.
Mora, C., Tittensor, D. P., Adl, S., Simpson, A. G. B. & Worm, B. (2011) How many species
are there on earth and in the ocean? PloS Biology, 9, e100127. Mouchet, M., Guilhaumon, F., Villeger, S., Mason, N. W. H., Tomasini, J. A. & Mouillot, D.
(2008) Towards a consensus for calculating dendrogram-based functional diversity indices. Oikos, 117, 794-800.
Meyer, C. F. J., Aguiar, L. M. S., Aguirre, L. F., Baumgarten, J., Clarke, F. M. et al. (2011)
Accounting for detectability improves estimates of species richness in tropical bat surveys. Journal of Applied Ecology, 48, 777-787.
Murienne, J., Grandcolas, P., Piulachs, M. D., Belles, X., D'Haese, C., Legendre, F., Pellens,
R. & Guilbert, E. (2005) Evolution on a shaky piece of Gondwana: is local endemism recent in New Caledonia? Cladistics, 21, 2-7.
Murienne, J., Pellens, R., Budinoff, R. B., Wheeler, W. C. & Grandcolas, P. (2008)
Phylogenetic analysis of the endemic New Caledonian cockroach Lauraesilpha. Testing competing hypotheses of diversification. Cladistics, 24, 802-812.
Murienne, J., Guilbert, E. & Grandcolas, P. (2009) Species diversity in the New Caledonian
endemic genera Cephalidiosus and Nobarnus (Insecta: Heteroptera: Tingidae), an approach using phylogeny and species distribution modeling. Biological Journal of the Linnean Society, 97, 177-184.
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. & Kent, J. (2000)
Biodiversity hotspots for conservation priorities. Nature, 403, 853-8. Nath, C. D., Pelissier, R. & Garcia, C. (2010) Comparative efficiency and accuracy of
variable area transects versus square plots for sampling tree diversity and density. Agroforestry Systems, 79, 223-236.
Nath, C. D., Pelissier, R., Ramesh, B. R. & Garcia, C. (2011) Promoting native trees in shade
coffee plantations of southern India: comparison of growth rates with the exotic G. robusta. Agroforestry Systems, 83, 107-119.
Obrist, M. K., Boesch, R. & Fluckiger, P. F. (2004) Variability in echolocation call design of
26 Swiss bat species: consequences, limits and options for automated field identification with a synergetic pattern recognition approach. Mammalia, 68, 307-322.
Obrist, M.K., Pavan, G., Sueur, J., Riede, K., Llusia, D. & Marquez, R. (2010). Bioacoustics
approaches in biodiversity inventories. Abc Taxa, 8, 68-99. Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., Simpson,
G. L., Solymos, P., Henry, M., Stevens H. & Wagner, H. (2011) Vegan: community ecology package. R package version 2.0-0.

182
Olden, J. D., Poff, N. L., Douglas, M. R., Douglas, M. E.& Fausch, K. D. (2004) Ecological and evolutionary consequences of biotic homogenization. Trends in Ecology & Evolution, 19, 18-24.
Olden, J. D. & Rooney, T. P. (2006) On defining and quantifying biotic homogenization.
Global Ecology and Biogeography, 15, 113-120. Oliver, I. & Beattie, A. J. (1996) Invertebrate morphospecies as surrogates for species: A case
study. Conservation Biology, 10, 99-109. Oliver, I., Pik, S., Britton, D., Dangerfield, J. M., Colwell, R. K. & Beattie, A. J. (2000)
Virtual biodiversity assessment systems. Bioscience, 50, 441-450. Orme, C. D. L., Davies, R. G., Burgess, M., Eigenbrod, F., Pickup, N., Olson et al. (2005)
Global hotspots of species richness are not congruent with endemism or threat. Nature, 436, 1016-1019.
Paisley-Jones, C. E. (2011) Monitoring agroecosystem biodiversity using bioacoustics and
remote recording units. Master of Science report, Ohio State University Parsons, S. & Jones, G. (2000) Acoustic identification of twelve species of echolocating bat
by discriminant function analysis and artificial neural networks. Journal of Experimental Biology, 203, 2641-56.
Patil, G. P. & Taillie, C. (1982) Diversity as a concept and its measurement. Journal of the
American Statistical Association, 77, 548-561. Pavoine, S., Baguette, M. & Bonsall, M. B. (2010) Decomposition of trait diversity among the
nodes of a phylogenetic tree. Ecological Monographs, 80, 485-507. Pavoine, S. & Bonsall, M.B. (2011) Measuring biodiversity to explain community assembly:
a unified approach. Biological Reviews, 86, 792-812. Pavoine, S., Dufour, A. B. & Chessel, D. (2004) From dissimilarities among species to
dissimilarities among communities: a double principal coordinate analysis. Journal of Theoretical Biology, 228, 523-537.
Pavoine, S., Ollier, S. & Dufour, A.B. (2005a). Is the originality of a species measurable?
Ecology Letters, 8, 579-586. Pavoine, S., Ollier, S. & Pontier, D. (2005b) Measuring, diversity from dissimilarities with
Rao's quadratic entropy: Are any dissimilarities suitable? Theoretical Population Biology, 67, 231-239.
Pavoine, S., Vallet, J., Dufour, A. B., Gachet, S. & Daniel, H. (2009) On the challenge of
treating various types of variables: application for improving the measurement of functional diversity. Oikos, 118, 391-402

183
Pearson, D. L. (1994) Selecting Indicator Taxa for the Quantitative Assessment of Biodiversity. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 345, 75-79.
Pellens, R. & Grandcolas P. (2010) Conservation and management of the biodiversity in a
hotspot characterized by short range endemism and rarity: the challenge of New Caledonia. In: Rescigno V, Maletta S, editors. Biodiversity Hotspots. New York: Nova Science Publishers. pp. 139-151.
Perfecto, I., Rice, R. A., Greenberg, R. & VanderVoort, M. E. (1996) Shade coffee: a
disappearing refuge for biodiversity. Bioscience, 46, 598-608. Perspectives mondiales de la diversité biologique 3, Rapport disponible à l'adresse
www.cbd.int/doc/publications/gbo/gbo3-final-fr.pdf. Petchey, O. L., Evans, K. L., Fishburn, I. S. & Gaston, K. J. (2007) Low functional diversity
and no redundancy in British avian assemblages. Journal of Animal Ecology, 76, 977-985.
Petchey, O. L. & Gaston, K. J. (2002) Functional diversity (FD), species richness and
community composition. Ecology Letters, 5, 402-411. Petchey, O. L. & Gaston, K. J. (2006) Functional diversity: back to basics and looking
forward. Ecology Letters, 9, 741-758. Petchey O.L., O'Gorman E. & Flynn, D.F.B. (2009) A functional guide to functional diversity
measures. In: Biodiversity and Human Impacts (eds. Naeem S., Bunker D.E., Hector A., Loreau M., Perrings C.) pp. 49-60, Oxford University Press, Oxford.
Pieretti, N., Farina, A. & Morri, D. (2011) A new methodology to infer the singing activity of
an avian community: the Acoustic Complexity Index (ACI). Ecological Indicators. 11, 868-873.
Pijanowski, B. C., Farina, A., Gage, S. H., Dumyahn, S. L. & Krause, B. L. (2011a) What is
soundscape ecology? An introduction and overview of an emerging new science. Landscape Ecology, 26, 1213-1232.
Pijanowski, B. C., Villanueva-Rivera, L. J., Dumyahn, S. L., Farina, A., Krause, B. L. et al.
(2011b). Soundscape Ecology: The science of Sound in the Landscape. Bioscience, 61, 203-216.
Pillar, V. D., Duarte, L. D., Sosinski, E. E. & Joner, F. (2009) Discriminating trait-
convergence and trait-divergence assembly patterns in ecological community gradients. Journal of Vegetation Science, 20, 334-348.
Pillon, Y., Munzinger, J., Amir, H. & Lebrun, M. (2010) Ultramafic soils and species sorting
in the flora of New Caledonia. Journal of Ecology, 98, 1108-1116.

184
Pollard, K. A., Blumstein, D. T. & Griffin, S. C. (2010) Pre-screening acoustic and other natural signatures for use in noninvasive individual identification. Journal of Applied Ecology, 47, 1103-1109.
Purvis, A. & Hector, A. (2000). Getting the measure of biodiversity. Nature, 405, 212-219. R Development Core Team R (2010) A language and environment for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/.
Rachev, S.T. (1991) Probability metrics and stability of stochastic models, JohnWiley &
Sons. New York. Rao, C. R. (1982) Diversity and dissimilarity coefficients: a unified approach. Theoretical
Population Biology, 21, 24–43. Rao, C.R. (1986) Rao's axiomization of diversity measures. Encyclopedia of statistical
sciences, 7, 614-617. Ramesh, B. R., Ayyappan, N., Grard, P., Prospering, J., Aravajy, S. & Pascal, J.-P. (2010)
Western Ghats v.1.0 - A multimedia identification system of evergreen species of the Western Ghats, India. http://www.biotik.org/.
Reaka-Kudla, M. L., Wilson, D. E., & Wilson, E. O. (1997). Biodiversity II: understanding
and protecting our biological resources. Joseph Henry Press, Washington. Reid, W. V. (1998) Biodiversity hotspots. Trends in Ecology & Evolution, 13, 275-280. Richards, D. G.& Wiley, R. H. (1980) Reverberations and Amplitude Fluctuations in the
Propagation of Sound in a Forest - Implications for Animal Communication. The American Naturalist, 115, 381-399.
Riede, K. (1993) Monitoring Biodiversity: analysis of Amazonian rainforest sounds. Ambio,
22, 546-548. Riede, K. (1997) Bioacoustic monitoring of insect communities in a Bornean rainforest
canopy. In Canopy Arthropods (eds. by N.E. Stork and J.A. Adis), pp. 442-452. Chapman & Hall, London
Roché, J. C (1990) Oiseaux d'Europe, Sitelle. Mens, 4CDs. Rockström, J., Steffen, W., Noone, K., Persson, Å., Chapin, S.III.F. et al. (2009) Planetary
boundaries : exploring the safe operating space for humanity. Ecology and Society, 14, 32.
Roberge, J. M. & Angelstam, P. (2004) Usefulness of the umbrella species concept as a
conservation tool. Conservation Biology, 18, 76-85. Robert, P. & Escoufier, Y. (1976) A unifying tools for linear multivariate statistical methods.
Journal of the royal Statistical Society, 25, 257-265.

185
Robillard, T. & Desutter-Grandcolas, L. (2011) Evolution of calling songs as multicomponent
signals in crickets (Orthoptera: Grylloidea: Eneopterinae). Behaviour, 148, 627-672. Robillard, T., Nattier, R. & Desutter-Grandcolas, L. (2010) New species of the New
Caledonian endemic genus Agnotecous (Orthoptera, Grylloidea, Eneopterinae, Lebinthini). Zootaxa, 2559, 17-35.
Rosser, N. & Eggleton, P. (2012) Can higher taxa be used as a surrogate for species-level data
in biodiversity surveys of litter/soil insects? Journal of Insect Conservation, 16, 87-92. Sheil, D., Ducey, M. J., Sidiyasa, K. & Samsoedin, I. (2003) A new type of sample unit for
the efficient assessment of diverse tree communities in complex forest landscapes. Journal of Tropical Forest Science, 15, 117-135.
Simberloff, D. (1998) Flagships, umbrellas, and keystones: is single-species management
passe in the landscape era? Biological Conservation, 83, 247-257. Simpson, E.H., 1949. Measurement of diversity. Nature. 688. Sinnott, R. W (1984) Virtues of the Haversine. Sky and Telescope, 68, 159. Skowronski, M. D. & Harris, J. G. (2006) Acoustic detection and classification of
microchiroptera using machine learning: lessons learned from automatic speech recognition. Journal of the Acoustical Society of America, 119, 1817-1833.
Slabbekoorn, H. (2004) Habitat-dependent ambient noise: consistent spectral profiles in two
African forest types. Journal of the Acoustical Society of America, 116, 3727-3733. Slingsby, J.A. & Verboom, G.A. (2006) Phylogenetic relatedness limits co-occurrence at fine
spatial scales: Evidence from the schoenoid sedges (Cyperaceae : Schoeneae) of the Cape Floristic Region, South Africa. The American Naturalist, 168, 14-27.
Sørensen, T. A. (1948) A method of establishing groups of equal amplitude in plant sociology
based on similarity of species content, and its application to analyses of the vegetation on Danish commons. Kongelige Danske Videnskabernes Selskabs Biologiske Skrifter, 5, 1–34
Stork, N. E. (1997) Measuring global biodiversity and its decline. In Biodiversity II:
understanding and protecting our biological resources (eds. by ML Reaka-Kudla, DE Wilson & EO Wilson), pp41-68. Joseph Henry Press, Washington.
Sueur, J. (2002) Cicada acoustic communication: potential sound partitioning in a
multispecies community from Mexico (Hemiptera: Cicadomorpha: Cicadidae). Biological Journal of the Linnean Society, 75, 379-394.
Sueur J., Aubin T. & Simonis C. (2008a). Seewave: a free modular tool for sound analysis
and synthesis. Bioacoustics, 18: 213-226.

186
Sueur, J., Pavoine, S., Hamerlynck, O. & Duvail, S. (2008b) Rapid Acoustic Survey for Biodiversity Appraisal. PLoS ONE, 3, e4065.
Sueur, J., Gasc, A., Grandcolas, P. & Pavoine, S. (2012) Global estimation of animal diversity
using automatic acoustic sensors. In: Le Gaillard JF, Guarini JM, Gaill F, editors. Sensors for ecology. Paris: CNRS. pp. 99-117.
Sutherland, W. J. (2006) Ecological census techniques, 2nd edition. Cambridge University
Press, Cambridge. Tanzarella, S. (2006) Perception et communication chez les animaux. De boeck & Larcier,
Bruxelles. Thuiller, W., Lavergne, S., Roquet, C., Boulangeat, I., Lafourcade, B. & Araujo, M. B. (2010)
Consequences of climate change on the tree of life in Europe. Nature, 470, 531-534. Tilman, D. (2001) Functional diversity. Encyclopedia of Biodiversity (ed. Levin, S.A.), pp.
109-120. Academic Press, San Diego, CA. Towsey, M., Planitz, B., Nantes, A., Wimmer, J. & Roe, P. (2012) A tool box for animal call
recognition. Bioacoustics, 21, 107-125. Turner, W., Spector, S., Gardiner, N., Fladeland, M., Sterling, E. & Steininger, M. (2003)
Remote sensing for biodiversity science and conservation. Trends in Ecology & Evolution, 18, 306-314.
UICN, rapport Bilan de 2010. Disponible en ligne : http://www.uicn.fr/IMG/pdf/Bilan_SNB_04-10_UICN_France.pdf Vane-Wright, R. I., Humphries, C. J. & Williams, P. H. (1991) What to protect? Systematics
and the agony of choice. Biological Conservation, 55, 235-254. Villanueva-Rivera, L. J. (2007) Digital recorders increase detection of Eleutherodactylus
frogs. Herpetological Review, 38, 59-63. Violle, C., Navas, M. L., Vile, D., Kazakou, E., Fortunel, C., Hummel, I. & Garnier, E. (2007)
Let the concept of trait be functional! Oikos, 116, 882-892. Wallschlager, D. (1980) Correlation of song frequency and body-weight in Passerine birds.
Experientia, 36, 412-412. Ward, D. F. & Stanley, M. C. (2004) The value of RTUs and parataxonomy versus taxonomic
species. New Zealand Entomologist, 27, 3-9. Warren, M. S., Hill, J. K., Thomas, J. A., Asher, J., Fox, R., Huntley, B., Roy, D. B., Telfer,
M. G., Jeffcoate, S., Harding, P., Jeffcoate, G., Willis, S. G., Greatorex-Davies, J. N., Moss, D. & Thomas, C. D. (2001) Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature, 414, 65-69.

187
Warwick, R. M. & Clarke, K. R. (1995) New 'biodiversity' measures reveal a decrease in taxonomic distinctness with increasing stress. Marine Ecology-Progress Series, 129, 301-305.
Webb, C. O., Ackerly, D. D., McPeek, M. A., Donoghue & M. J. (2002) Phylogenies and
community ecology. Annual Review of Ecology and Systematics, 33, 475-505. Whittaker, R. H. (1972) Evolution and measurement of species diversity. Taxon, 21, 213-251. Wildlife Acoustics (2009) Wildlife Acoustics, Inc. Bioacoustics Software and Field
Recording Equipment. Disponible en ligne : http://www.wildlifeacoustics.com/. Wiley H. R. & Richards, D. G. (1978) Physical constraints on acoustic communication in the
atmosphere: implications for the evolution of animal vocalizations. Behavioral Ecology and Sociobiology, 3, 69-94.
Wiley, R. H. (1991) Associations of song properties with habitats for territorial Oscine birds
of Eastern North-America. American Naturalist, 138, 973-993. Wilson, E. O & Peter, F. M. (1988) Biodiversity. National Academy Press, Washington. Wimmer, J., Towsey, M., Planitz, B., Williamson, I. & Roe, P. (2012) Analysing
environmental acoustic data through collaboration and automation. Future Generation Computer Systems, sous presse.
Wolda, H. (1993) Diel and seasonal patterns of mating calls in some neotropical cicadas.
Acoustic interference. Proceedings, Koninklijke Nederlandse Akademie van Wetenschappen, 96, 369-381.
Wright, J. P., Jones, C. G. & Flecker, A. S. (2002) An ecosystem engineer, the beaver,
increases species richness at the landscape scale. Oecologia, 132, 96-101. Wunderle, J. M. & Latta, S. C. (1998) Avian resource use in Dominican shade coffee
plantations. Wilson Bulletin, 110, 271-281.

188

189
Annexe 1 Chapitre du livre
Le Gaillard, J-L., Guarini, J-M.
and Gaill, F. (2012) Sensors for
ecology. CNRS, Paris.

190

191
Chapter 1: Global estimation of animal diversity using
automatic acoustic sensors
Jérôme Sueur, Amandine Gasc, Philippe Grandcolas, Sandrine Pavoine
1. Introduction
The estimation of biodiversity can be considered as one of the main challenges in modern
biology. When dealing with ecology, evolutionary biology and conservation biology, there is
an inescapable need to describe the composition and dynamics of biological diversity
(Magurran, 2004). In ecology, the concept of biological diversity is mainly species-oriented,
even if other evolutionary units or traits can also be used. In this context, biodiversity
potentially refers to all species encountered in a given area at a specified time, including every
potential species from underground bacteria to giant trees. Therefore, biodiversity assessment
can turn out to be a time-consuming and complex task, as it relies on species inventory that
may involve very different taxonomic groups.
Exhaustive approaches such as the all taxa biodiversity inventory (ATBI) programs aim at
inventorying the whole biodiversity mainly in tropical habitats (Gewin, 2002), but these
programs are highly sensitive to the logistic and time-constraints of most inventory studies.
An alternative to these approaches is to focus on one or a few taxa and consider them as
biodiversity indicators (Pearson, 1994), but the choice of representative taxa is not trivial
(Lawton et al., 1998). In addition, it is well known that patterns of species diversity for
different taxa are sensitive to the observation scale. More precisely, there is a general
congruence for species diversity between different taxa at a large area scale (more than 1km2)
but not at a fine scale (less than 1km2, Weaver, 1995). This renders difficult the definition of
an indicator taxon or even of several indicator taxa supposedly representative of the diversity
in other forms of organisms (Ricketts et al., 1999). Irrespective of the taxonomic breadth of
any biodiversity assessment, the estimation of species biodiversity relies on inventories and
species examination by one or several taxonomic experts that can be supported with genetic
barcoding techniques (see chapter II, 3). Sampling in the field and identification in museum
collections can require a considerable effort when the objective is to sample a large region for
a long time period. To improve the rate of specimen collection, non-specialist taxonomic

192
workers – or para-taxonomists – can separate morpho-species instead of identifying valid
species. This solution is advocated by the rapid biodiversity assessment (RBA) programs that
have been especially developed for the rapid exploration of biodiversity in tropical habitats
(Oliver and Beattie, 1993; 1996; Oliver et al., 2000).
Biodiversity assessment is often restricted to species richness, i.e. to the counting of the total
number of species. However, a collection of species cannot be described solely by the number
of items it includes. The abundance of each species has to be assessed to provide an
estimation of species evenness. Evolutionary, ecological and life history characters of the
species also describe facets of biodiversity (Brooks and McLennan, 1991; Vane-Wright et al.,
1991; Grandcolas, 1998; Pavoine et al., 2009; Petchey et al., 2009). Species turnover along
time and/or spatial scales is also required to take into account biodiversity dynamics. All of
these requirements led to a plethora of biodiversity indices that have been developed for
decades (Magurran, 2004; B uckland et al., 2005; Pavoine and Bonsall, 2010).
In practice, a measure of biodiversity can be achieved with direct or indirect sampling. In the
latter case, the use of a sensor should be ideally considered by employing a simple tool that
returns an index of biodiversity. A human observer or a network of human observers might be
considered as a “biodiversity sensor”, but may be biased by the experience of the observers
and cannot be “deployed” in rough terrains for long periods of time. Another possibility is to
work with local image, video capture instruments or with global satellite imagery. Satellite-
based earth observations, or remote sensing, can produce environmental parameters from
biophysical characteristics that can be indirectly used to assess species ranges and species
richness patterns (Kerr et al., 2003, Turner et al., 2003; Wang et al., 2010; see also IV, 2).
These methods are undoubtedly very attractive, but they rely on extremely expensive
equipment and are difficult to adapt to small spatial scales. Like other methods, remote
sensing often requires a time-consuming validation step. For instance, vegetation mapping
with satellite images is based on a colorimetric calibration of pixels with a large set of direct
vegetation samples.
The use of a given sensor should be made according to a sampling strategy designed and
evaluated carefully with respect to the type of data to be collected. It is particularly important
to identify the precision of the measurement system (data quality) and the level of accuracy
that has to be reached along both time and space scales (data quantity). In most sampling
strategies, there is a basic trade-off between precision and accuracy. In this context, we are
currently developing an acoustic sensor that would produce a biodiversity index by analysing
the sound produced by local animal communities. This approach could provide a portable,

193
cheap, reasonably accurate and non-invasive animal diversity sensor that could be used at
different space and time scales.
2. Sensing diversity through bioacoustics
Some animal species, including taxa often used in biodiversity studies, produce active sounds
during their social interactions or in other contexts. For example, some fish and reptiles, most
amphibians, birds, mammals, insects and other arthropods use sound for communication,
navigation or predation acts. These acoustic signals generally produce a species-specific
signature and several techniques in bioacoustics were developed to exploit these signals as an
indication of species occurrence and as a tool for biodiversity studies (Obrist et al., 2010). The
most elementary application is sensing by observers. This is usually achieved when following
animal populations through aural listening and identification (e.g., Cano-Santana et al., 2008,
for crickets). When based on a massive network of listeners, such a census method can
generate large datasets of strong interest to ecologists (e.g. Devictor et al., 2008, for birds).
Nonetheless, volunteer based call surveys tend to be replaced by the automated digital
recording system (ADRS), which is an electronic equipment that allows automatic data
collection and generates a large amount of high-quality information about species biodiversity
(e.g. Acevedo and Villanueva-Rivera, 2006, for amphibians).
The problems alluded above for species identification with museum specimens is also true for
species identification with sound. Acoustic identification is based on the experience of the
observers, which can be biased due to sensory or training differences. As any other
identification, it also relies on a taxonomic database providing information on the
correspondence between every species and its acoustic signature. Automatic identification of
the different songs embedded in the recording is rather complex and still suffers errors (e.g.
Skowronski and Harris, 2006, for bats). These approaches are also difficult to deploy in
complex acoustic environments like tropical forest soundscapes, where tens of signals mix up
and many species still remain unknown (Riede, 1993). Reliable results can be obtained only
when focusing on a single species with a rather simple and loud call as demonstrated with the
neotropical bird Lipaugus vociferans (see I, 4) and the Blue Whale Balaenoptera musculus in
a marine context (see I, 3).
Keeping in mind these constraints, we applied the concept of RBA to sounds produced by
animals and even pushed the concept one step further. We recently suggested tackling the

194
problem of diversity assessment at the community level by using bioacoustic methods (Sueur
et al., 2008a). In the case of bioacoustics, the unit to work with is the acoustic community,
which is defined as the sum of all sounds produced by animals at given location and time. The
signals produced by different species can overlap, interfere and consequently reduce signal
transmission between the emitter and the receiver of a focal species. Sound produced by other
species is indeed considered as noise for the focal species and acts as a severe constraint on
the evolution of conspecific signals (Brumm and Slabbekoorn, 2005). Consequently, species
sharing the same acoustic space are supposed to show an over-dispersion of the frequency and
time amplitude parameters of their songs reducing the risk of interference. This has been
reported in several acoustic communities (e.g., Lüddecke et al., 2000, for amphibians; Sueur,
2002, for cicadas; Luther, 2009, for birds). A measure of sound complexity could then work
as a proxy of community richness and composition. The acoustic indices we are developing
are mainly based on this concept of acoustic partitioning. We hereafter review the recording
equipment and analysis we used to try and build an animal diversity acoustic sensor.
3. Listening and measuring acoustic diversity
A biodiversity sensor provides a measure of a single or a set of variables characterising
biodiversity. Even if a sensor is composed of several probes and data analysers, it is often
viewed as a all-in-one equipment that senses and analyses the environment concomitantly.
Our method currently relies on two different equipments that are not used at the same time.
However, we here consider that these sub-units constitute together a single sensor (figure 1).
The first sub-unit is a digital sound-recorder that can be settled outdoor. The second sub-unit
is a computer installed with software specifically developed to analyse sound diversity.
Further statistical analyses on the acoustic indices, i.e. the biodiversity variables measured by
the sensor, are not considered as part of the sensor but as part of data analysis processes. We
hereafter detail the sampling protocols based on a single recorder or an array of recorders, the
properties of the autonomous recorder currently in use and, eventually, the algorithm
developed to compute the diversity indices from sound files.

195
Figure 1: Diagram showing the successive steps of the global estimation of animal diversity.
Here, the biodiversity sensor is considered as the combination of different processes:
recording, audio file conservation, signal analysis, and indices computation. These processes
– in situ recording step with autonomous equipment and ex situ calculation of indices from
stored acoustic data – are currently separated from each other. However, a portable all-in-one
system might be developed in the future.
3.1. From a single manual recording spot to a network of autonomous recorders
The sampling protocol is mainly constrained by the recording equipment available. Our
method was first tested with a comparison between two closely spaced dry lowland coastal
forests in Tanzania. The recording of the animal communities inhabiting these forests was
achieved with a digital recorder (Edirol© R09) equipped with an omnidirectional microphone
(Sennheiser© K6/ME62). Recordings were done by a single person at three times of the day
and successively in the two forests. This procedure limited the sampling to a few days and to
only two sampling sites. Such digital recorders also provide internal microphones that can be
used to reduce costs. In this case, several items can be purchased to cover a wider area and a
longer period of time. However, the recorders still have to be triggered and stopped manually,
a condition that makes field work rather challenging.
Unattended recorders were not available since the North American company Wildlife
Acoustics© provided an autonomous digital field recorder (see details about this recording
package in section 3.2). An autonomous system was absolutely necessary to design sampling
protocols with synchronised units such as regular, cluster, multi-level, or stratified protocols.
We first used three of these recorders to assess animal diversity within temperate woodlands
by simultaneously recording a mature forest, a young forest and an edge forest (figure 2A,
Depraetere et al., 2012). We then increased the number of recorders to estimate biodiversity
endemism of three New-Caledonian sites. We planned a stratified sampling with four
recorders set in each site. This ensured a repetition per site and allowed comparisons within
and between study sites (figure 2B). Later, we tracked acoustic diversity of a typical tropical

196
forest by deploying a network of 12 recorders regularly spaced on a 100 ×100m grid in
French Guiana (see IV, 2). Each recorder was equipped with a microphone settled 1.5m high
and a second microphone placed 20m high in the canopy (figure 2C). This 3D regular
sampling covered 12ha of forest for more than 40 days. Eventually, we tried to transfer our
method to freshwater habitats like forest ponds. This was achieved by adapting the
autonomous recorder with a Reson© hydrophone and an Avisoft© pre-amplifier (figure 2D).
This high-quality equipment is expensive (about 2700€ per unit) and sampling was therefore
limited to three recording units. We therefore designed a rotating sampling by regularly
moving the hydrophone position along transects.
Figure 2: The autonomous Wildlife Acoustics© recorder installed for outdoor studies. A.
First version of the recorder (SM1) settled in a temperate woodland to estimate local bird
acoustic community (Rambouillet, France). B. Second version of the recorder (SM2) with a
single microphone in action (Mandjelia, New- Caledonia). C. The same recorder with two
microphones, one 2 m high and the other one ready to be set 20 m up in the canopy
(Nouragues experimental station, French Guiana). D. Recorder connected to an hydrophone to
record underwater sound of a pond (Rambouillet, France).
3.2. The Song Meter: an autonomous acoustic sensor
Wildlife Acoustics© developed two generations of autonomous digital recorders, namely the
Song Meter SM1 and SM2 (figure 3). These stereo recorders, which weigh 1.6kg each and
measure 20.3 × 20.3 × 6.4cm, possess a stereo recording system with omnidirectional

197
microphones that have a f lat frequency response between 0.02 and 20kHz. These
microphones can be directly connected to the main box, where data are stored, or can be
settled up to 50m away from it. Given that terrestrial animals produce sound with an intensity
of ca. 80dB at 1m re. 2×10-5 μPa (Sueur, unpublished data) and given that the microphones
have a sensitivity of -36 ± 4dB, we can estimate that in a closed habitat, such as a forest, the
microphone detects sounds up to around 100m from the source. A SM2 platform would then
cover an area of approximatively 3,1ha.
The recording sampling rate can be set from 4 to 48kHz with the standard SM2 and up to
384kHz with the ultrasonic SM2 option. The SM2 recorders are currently working with a
lossless compression format (.wac) that can be written on four secure digital (SD) cards. The
four SD slots provide 128Go storage space. Choosing an adequate sampling rate is not an
easy task as it results from a trade-off between cost, data storage and the sound frequency
used by animals. Increasing the sampling rate to high frequency requires a specific and
expensive motherboard and, above all, generates very large sound files that are difficult to
handle and to analyse. However, this is the only solution to record the acoustic activity of
some insects and bats that emit ultrasound signals for communication or navigation. Up to
now, we sampled the animal acoustic communities at a 44.1kHz sampling rate. A network of
recorders generates thousands of files that need to be stored and analysed (see section 4.2).
Using a higher sampling rate will certainly preclude the estimation of acoustic diversity by
generating too high an amount of data.
Electrical power is provided by four alkaline or LR20 batteries ensuring a maximum of 240
hours of recordings. Energy can also come from an external 12V battery potentially connected
to a solar panel. Eventually, the SM2 platform provides also an internal temperature sensor
and a connection for an external sensor. The additional data are written on the SD cards
together with sound files. The main advantage of the Song Meter is that it can be easily
programmed to record on simple time-of-day schedules or to implement complex monitoring
protocols, even scheduling recordings relative to local sunrise, sunset and twilight. For
instance, a schedule can be programmed to record regularly all day and night long, but also to
record more intensively around sunrise and sunset, when dawn and dusk choruses of birds,
insects and amphibians occur.

198
Figure 3: The second version of the recorder (SM2) opened to show the main characteristics.
A cable can be used to set the microphones away from the main box. Detailed characteristics
can be obtained at http://www.wildlifeacoustics.com. © Wildlife Acoustics.
3.3. Computing the acoustic indices
Biodiversity is traditionally decomposed into two levels, the average diversity within
communities, or α diversity, and the diversity between communities, or β diversity. We
therefore developed two acoustic indices aiming at estimating these two components of
biodiversity (Sueur et al., 2008a). Both indices can be computed with the package seewave
(Sueur et al., 2008b) of the free R environment (R Development Core Team, 2012). The first
index, named H, is a Shannon-like index. The index H gives a measure of the entropy of the
acoustic community by considering both temporal and frequency entropy. H is computed
according to:
H = Ht × Hf with 0 ≤ H ≤ 1, and
Ht = - Σ (A(t) × log(A(t)) / log (n)), and
Hf = - Σ (S( f ) × log(S( f )) / log (N)),
where n = length of the signal in number of digitized points, A(t) = probability mass function
of the amplitude envelope, S( f ) = probability mass function of the mean spectrum calculated
using a short term Fourier transform (STFT) along the signal with a non-overlapping
Hamming window of N = 512 points (figure 4).
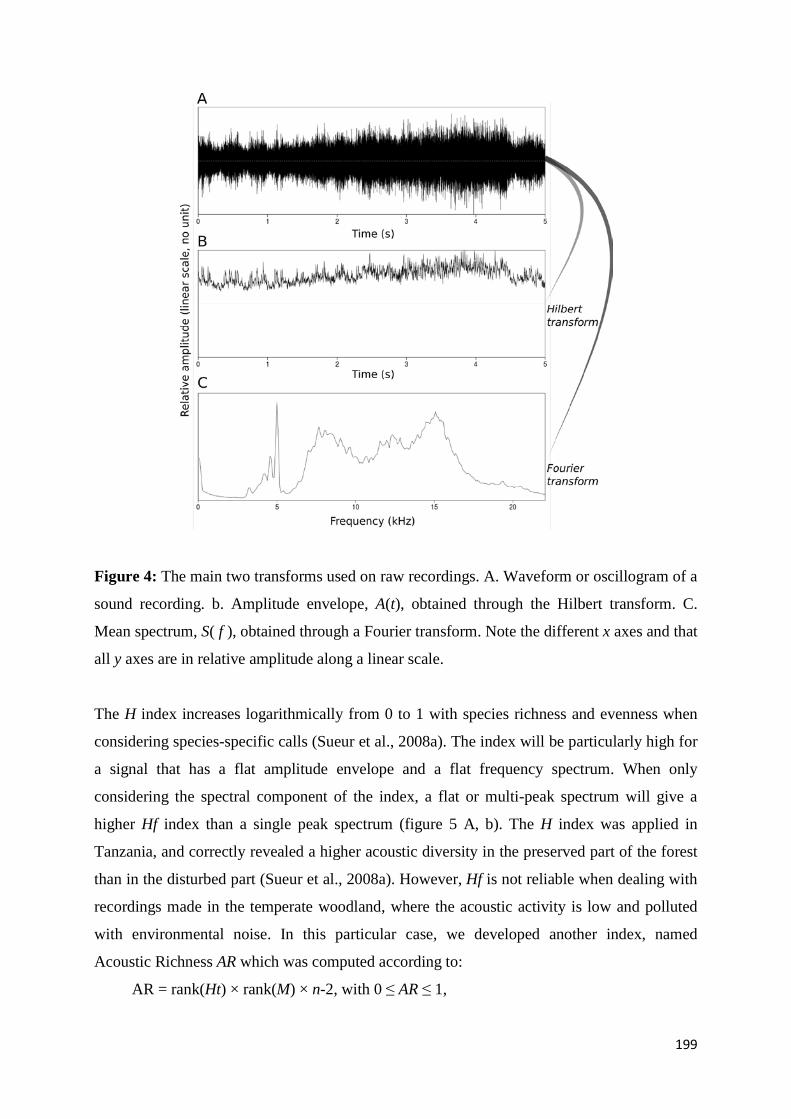
199
Figure 4: The main two transforms used on raw recordings. A. Waveform or oscillogram of a
sound recording. b. Amplitude envelope, A(t), obtained through the Hilbert transform. C.
Mean spectrum, S( f ), obtained through a Fourier transform. Note the different x axes and that
all y axes are in relative amplitude along a linear scale.
The H index increases logarithmically from 0 to 1 with species richness and evenness when
considering species-specific calls (Sueur et al., 2008a). The index will be particularly high for
a signal that has a flat amplitude envelope and a flat frequency spectrum. When only
considering the spectral component of the index, a flat or multi-peak spectrum will give a
higher Hf index than a single peak spectrum (figure 5 A, b). The H index was applied in
Tanzania, and correctly revealed a higher acoustic diversity in the preserved part of the forest
than in the disturbed part (Sueur et al., 2008a). However, Hf is not reliable when dealing with
recordings made in the temperate woodland, where the acoustic activity is low and polluted
with environmental noise. In this particular case, we developed another index, named
Acoustic Richness AR which was computed according to:
AR = rank(Ht) × rank(M) × n-2, with 0 ≤ AR ≤ 1,

200
where rank is the value position along the ordered samples, M is the median of the amplitude
envelope and n the number of recordings (Depraetere et al., 2012).
The second index, named D, is a simple acoustic dissimilarity measure. D is similarly
composed of two sub-indices based on a difference between amplitude envelopes and
frequency spectra respectively (figure 5 C). D is calculated like following:
D = Dt × Df with 0 ≤ D ≤ 1, and
Dt = 0.5 × Σ |A1 (t) – A2 (t)|, and
Df = 0.5 × Σ |S1 ( f ) – S2 ( f )|,
where A1(t), A2(t) are probability mass functions of the amplitude envelope for the two
recordings under comparison, and S1( f ), S2( f ) are probability mass functions of the mean
spectrum for the two recordings to be compared. The D index increases linearly with the
number of unshared species between the two recordings, or communities (Sueur et al., 2008a).
Both indices may suffer a bias as some species naturally produce signals with high temporal
and/or spectral entropy. This is particularly the case of cicadas whose noise-like sound can be
mistakenly interpreted as a high local diversity. Such bias can be buffered with a large
sampling including a high number of time and space repetitions. The indices can also produce
false values when background noise overlaps with the sound produced by the animal
community (see section 4.1). Both indices are currently tested in different temperate and
tropical habitats in this respect.
Other acoustic indices have been developed elsewhere to monitor habitat state or community
activity. Qi et al. (2008) divided the soundscape of an ecosystem following three frequency
bands: the anthrophony, between 0.2 and 1.5kHz, the biophony, which starts at 2kHz with a
peak at 8kHz, and the geophony, which can cover the entire spectrum with dominant low
frequency. By computing a ratio between biological and anthropogenic signals, they coined
an ecological estimator of ecosystem health. This original procedure does not give an
estimation of local diversity but assesses the level of biological sound activity relative to
anthropogenic activities. Pierreti et al. (2010) and Farina et al. (2011) designed an acoustic
complexity index (ACI). This index computes time and frequency variability of a sound
extrapolated from a spectrogram. The ACI appears to be correlated with the number of
vocalisations produced by a bird community. However, this index assesses neither species
diversity nor community turnover. The ACI index proved to be poorly sensitive to invariant
noise, such as continuous noise from cars or aircrafts, but can be impactedby unpredictable
noise such as wind, running water or irregular human activity. All these acoustic indices,
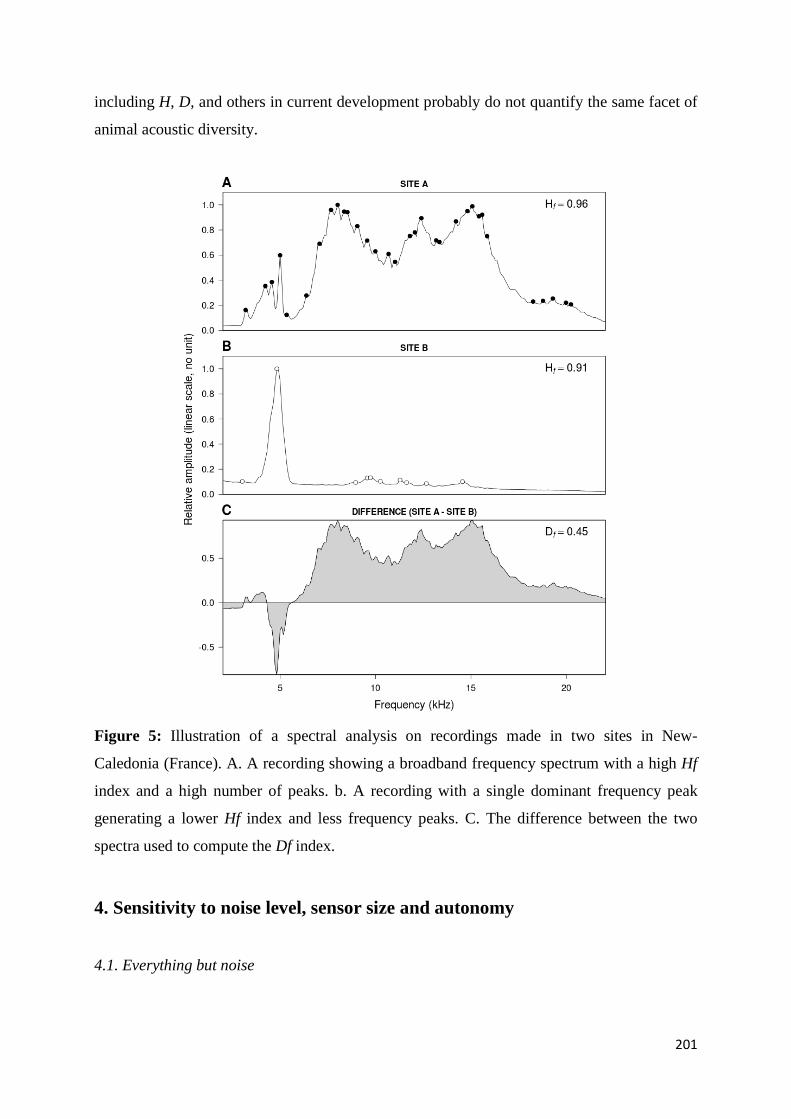
201
including H, D, and others in current development probably do not quantify the same facet of
animal acoustic diversity.
Figure 5: Illustration of a spectral analysis on recordings made in two sites in New-
Caledonia (France). A. A recording showing a broadband frequency spectrum with a high Hf
index and a high number of peaks. b. A recording with a single dominant frequency peak
generating a lower Hf index and less frequency peaks. C. The difference between the two
spectra used to compute the Df index.
4. Sensitivity to noise level, sensor size and autonomy
4.1. Everything but noise

202
Background noise is probably the primary issue in bioacoustics. Noise can significantly
impairs acoustic observations and experiments by masking or distorting both time-amplitude
and frequency parameters (Hartmann, 1998; Vaseghi, 2000). There are three main sources of
noise to consider when recording outdoor: i) anthropogenic noise due to machinery, car, boat,
plane, train traffic, or any other human activity, ii) biotic noise due to the activity of
surrounding species, and iii) environmental noise due to rain, wind, river stream, waterfall, or
sea wave (Brumm and Slabbekoorn, 2005; Laiolo, 2010). The estimation of animal diversity
through acoustics is based on the recording of a whole community and as such does not face
the classical problems encountered when trying to record a single species in the background
noise generated by surrounding active species. However, anthropogenic and environmental
noise can have negative effects on the results. In a few instances, anthropogenic noise can be
removed by applying classical frequency filters (Stoddard, 1998). Recordings made close to
an airport or a road with a regular traffic can be cleaned with a high-pass filter that will
remove the low frequency band generated by plane or car engines. Such filters might exclude
low pitch animal calling songs, but this can be accounted for when computing diversity
indices. The main difficulty arises when recordings are polluted with unpredictable and/or
broadband noise that can be interpreted erroneously as animal sounds. Removing such chaotic
sound is a challenge to be solved in bioacoustics as well as in other acoustic disciplines
(Rumsey and McCormick, 1992; Hartmann, 1998; Stoddard, 1998; Vaseghi, 2000). Usual
frequency filters cannot be used as noise may overlap the frequency band used by the animal
community. Other noise reduction algorithms use noise spectrum as a reference to be
convoluted with the original signal. This solution might appear elegant but still suffers
important limitations. First, the noise has to be constant in its frequency content, a condition
rarely met in a natural acoustic environment. Second, it is necessary to identify accurately a
time window where only noise occurs. This latter condition is very difficult to meet when
faced with hundreds or thousands of recordings.
Fortunately, some upstream solutions can be considered to reduce the anthropogenic and
environmental noise (Obrist et al., 2010). When using an outdoor acoustic sensor, the most
important parameter to consider is the direction and the protection of the microphone. The
microphone can be oriented in a horizontal or vertical position as soon as its directivity
pattern is omnidirectional. A vertical upward position should be avoided when possible, as
rain drops might directly strike the microphone membrane. A vertical upside-down
orientation might be the best solution in avoiding rain and lateral wind effects. More
generally, adapting the orientation of the microphone to the local main sources of noises is

203
usually advocated. For instance, the noise of running water or passing-by cars can be reduced
by orienting the microphone perpendicularly to the source, and windscreens should be used to
attenuate wind noise. Another upstream solution is to exclude data potentially corrupted with
environmental noise. This can be achieved in three ways. The first option consists in cutting
off the recording session when weather conditions are too bad. It is not yet available but could
certainly be implemented quickly, given the availability of climate sensors in sound meter
devices. The second option is to apply a signal-to-noise algorithm that indicates the
occurrence of an important background noise. A threshold could be used as a reference to
keep or to remove the files from the dataset. This solution is under development in our group.
The third and last option, which is currently in use, is to gather climatic parameters from a
local station and identify the time periods when the weather was too bad to allow a correct
estimation of the acoustic diversity. This identification can be achieved automatically with a
threshold applied on the climatic parameters or by running a redundancy analysis (RDA, Rao,
1964) to the acoustic indices with the climatic parameters as factors (Depraetere et al., 2012).
4.2. Optimal size of recorders
As described above, the SM2 recorder weighs around 1.6kg and can be fitted with two
microphones (figure 3). Hence, handling several of these units in a hard-to-reach environment
requires a significant effort. A reduced size and weight would make field work easier and
could also allow settling more units in the habitat. However, this has to be traded off against
the size of the data that needs to be stored and analysed. A typical .wav file, which is the most
popular uncompressed audio format, has a size of around 690kb/s (= 84ko/s) when sampled at
a 44.01kHz rate. This means that one minute of recording is roughly equivalent to 5Mo for a
single channel (mono) or around 10Mo for two channels (stereo). Sampling quickly generates
x×102 hours of recording in x×103 files for a total x×102 Go data. As detailed above, the
recorders have storage capacity of 128Go, which is enough for most applications sampled at
44.01kHz, but might appear limited for an over-month or over-year survey or for a long
ultrasound monitoring. The next step of data transfer onto a hard disk for storage and
conservation can take a significant time as writing speed is usually slow (around 6Mb/s =
0.7Mo/s). Eventually, the longterm storage of teraoctets of data can encounter some limits
with a standard hard disk or server capacity.
Regarding the calculation of indices, the larger the file, the slower the analysis process. Even
if automated with R scripts, the analysis of thousands sound files is time-consuming. This is
due to three main factors: i) the number of files to be analysed, ii) the size of each file, and iii)

204
the time taken by R to work with large files. There is no easy way yet to counteract these
three caveats. The number of files will increase as samples will be larger. The size of each file
cannot be reduced. Compressed formats in particular, such as .mp3, cannot be used for
obvious reasons of signal quality. The platform R is very convenient as it is free and open-
source. It makes it a perfect tool for sharing our research and transferring our techniques to
other laboratories. However, it may be relevant to look for other software solutions (see
section 5.2).
4.3. Energy
The SM2 recorder was developed to consume as less energy as possible, but current batteries
ensure 240 hours of recording and therefore put a strong limit on the duration of sampling. A
solution is to connect the recorder with a 12V battery fuelled with solar energy. However, if
such autonomous energy system properly works in sunny areas, it is not adapted to cloudy or
shaded areas like the understory of a tropical forest where a very low percentage of solar
radiation reaches the ground.
5. What’s next?
5.1. Sampling
Our method needs to be tested, validated and eventually applied in several acoustic conditions
from different habitats. So far, we have tested it with both simulated and field acoustic
communities (Sueur et al., 2008a; Depraetere et al., 2012). Tests on field communities
concerned African tropical coast forest and temperate forest habitats. The latter test implied a
modification of the indices to take into account the background noise and the low activity of
the acoustic community. We are currently sampling several other places including mountain
tropical forests in New Caledonia, neotropical evergreen forest in French Guiana, and
evergreen monsoon forest in India. We are also transferring the technique to freshwater
habitats by using hydrophones immerged in ponds.
One of the aims of our method is to provide a long-term and large-scale sampling. We are
currently sampling species diversity with a network of 10-16 sensors working about 40 days
long over approximately 16 ha of tropical forest in French Guiana and India. This time period
is too short to track seasonal variations of species diversity. We would like to extend it to at
least one year or even longer periods. Moreover, we plan to increase the number of sensors to

205
monitor a larger area. Increasing the sampling time and network size will generate serious
storage issues. A cut-off system that stops recordings when the meteorological conditions are
not good enough could constitute a nice and cheap solution to overcome this difficulty.
Another way could consist in sending directly the data from the recorder to a server through a
satellite connection, as wireless connection to a base radio may be too slow for heavy sound
files (see IV, 2 section 2.3). However such technological improvement mainly depends on the
industry and may take some time to emerge.
5.2. Improving the indices
As explained earlier, background noise is a central issue, and our indices, especially the index
H, are particularly sensitive to noise. It is therefore necessary to develop new indices that are
noise-resistant. Current research is ongoing in our laboratory to develop a new measurement
of the richness based on the frequency peaks of the Fourier spectrum (figure 5). The spectrum
can be smoothed or residual peaks due to noise can be filtered out so as to improve the
measure in case of rain or wind noise. Amplitude or frequency threshold will be also applied
on the envelope and the frequency spectrum respectively, to try to increase the signal-to-noise
ratio. Whatever the index in use is, we also need to exactly identify which biodiversity
information is collected by using the acoustic community as a proxy of animal diversity. Does
the H index only embed a richness-evenness value or does it include phylogenetic and/or
functional diversity information? Eventually, as outlined above, the signal analysis can be
slow due to R process. Software directly written in C language will be developed on the next
years to significantly speed up the analysis process.
5.3 Sharing the method with other scientists and citizens
There is an important ethical requirement for making available the bioacoustic sensor and
primary biodiversity data for later uses in terms of knowledge, engineering or conservation
(e.g. Graham et al., 2004; Suarez and Tsutsui, 2004). The recording equipment we used so far
can be purchased to the company Wildlife Acoustics©. The H and D index can be computed
with the free R package seewave. The sensor and integrated bioacoustic system is therefore
available to anyone. However, R does not have a user-friendly interface and we plan therefore
to share the method soon through an interactive website. Any user will be able to upload
recording files for analysis. The acoustic indices will be returned to the user together with an
optional graphical representation of the sound analysed (e.g., waveform, envelope,
spectrogram, spectrum). On a long-term scale, the recorder and the signal analysis would not

206
be separated but associated in a small and light all-in-one system. This system could be a
‘smartphone’ including a free application that computes the indices. Smartphones were
proved to work as nice sensors for mapping the noise level of European cities (Maisonneuve
et al., 2010). A similar citizen-science experience could be undertaken to assess animal
acoustic diversity inside or around cities.
Authors’ references
Jérôme Sueur, Amandine Gasc, Philippe Grandcolas:
Muséum national d’Histoire naturelle, Département Systématique et
Évolution, UMR 7205 Paris, France
Sandrine Pavoine:
Muséum national d’Histoire naturelle, Département Écologie et Gestion
de la B iodiversité, UMR 7204 Paris, France
Corresponding author: Jérôme Sueur, [email protected]
Aknowledgement
This research has been supported by the INEE (CNRS) with a PEPS program
and a PhD grant awarded to AG. Sampling in French Guiana was
achieved thanks to a CNRS Nouragues 2010 grant. Sampling in New
Caledonia was realised thanks to the ANR B IONEOCAL grant to PG.
Main part of research was financed with the FRB BIOSOUND grant
(Fondation pour la Recherche sur le Biodiversité). Marion Depraetere,
Vincent Devictor, Stéphanie Duvail, Olivier Hamerlynck, Frédéric Jiguet,
Isabelle Leviol, Pierre-Yves Martel, and Raphaël Pélissier participated at
different steps to the development of this new sensing method or provided
field data with which to compare our acoustic indices.
References
Acevedo M.A., Villanueva-Rivera L.J., 2006. Using automated digital recording systems as
effective tools for the monitoring of birds and amphibians. Wildlife Society Bulletin,
34, pp. 211-214.

207
Brooks D. R., McLennan D. A., 1991. Phylogeny, ecology, and behavior: a research program
in comparative biology. University of Chicago Press, Chicago, USA.
Brumm H., Slabbekoorn H., 2005. Acoustic communication in noise. Advances in the Study
of Behavior, 35, pp. 151-209.
Buckland S. T., Magurran A. E., Green R. E., Fewster R. M., 2005. Monitoring change in
biodiversity through composite indices. Philosophical Transactions of the Royal Society
of London B, 360, pp. 243-254.
Cano-Santana Z., Romero-Mata, A., Perez-Escobedo H. M., 2008. Modified Petraborg index
applied to the sampling of male crickets by aural detection. Journal of Orthoptera
Research, 17, pp. 111-112.
Depraetere M., Pavoine S., Jiguet F., Gasc A., Duvail S., Sueur, J., 2012. Monitoring animal
diversity using acoustic indices: implementation in a temperate woodland. Ecological
Indicators, 13, pp. 46-54.
Devictor V., Julliard R., Couvet D., Jiguet F., 2008. Birds are tracking climate warning, but
not fast enough. Proceedings of the Royal Society of London B, 275, pp. 2743-2748.
Farina A., Pieretti N., Piccioli L., 2011. The soundscape methodology for longterm bird
monitoring: a Mediterranean Europe case-study. Ecological Informatics, 6, pp. 354-363.
Gewin V., 2002. All living things, online. Nature, 418, pp. 362-363.
Graham C. H., Ferrier S., Huettman F., Moritz C., Peterson A. T., 2004. New developments in
museum-based informatics and applications in biodiversity analysis. Trends in Ecology
and Evolution, 19, pp. 497-503.
Grandcolas P., 1998. Phylogenetic analysis and the study of community structure. Oikos, 82,
pp. 397-400.
Hartmann W.M., 1998. Signals, sound, and sensation. Springer, Berlin, Germany.
Kerr J.T., Ostrovsky M., 2003. From space to species: ecological applications for remote
sensing. Trends in Ecology and Evolution, 18, pp. 299-305.
Laiolo P., 2010. The emerging significance of bioacoustics in animal species conservation.
Biological Conservation, 7, pp. 1635-1645.
Lawton J. H., Bignell D. E., Bolton B ., Bloemers G. F., Eggleton P., Hammond P. M., Hodda
M., Holt R. D., Larsenk T. B ., Mawdsley N. A., Stork N. E., Srivastava D. S., Watt A.
D., 1998. B iodiversity inventories, indicator taxa and effects of habitat modification in
tropical forest. Nature, 391, pp. 72-76.

208
Lüddecke H., Amézquita A., Bernal X., Guzmàn F., 2000. Partitioning of vocal activity in a
neotropical highland-frog community. Studies on Neotropical Fauna and Environment,
35, pp. 185-194.
Luther D., 2009. The influence of the acoustic community on songs of birds in a neotropical
rain forest. Behavioral Ecology, 20, pp. 864-871.
Magurran A. E., 2004. Measuring biological diversity. Blackwell Science Ltd, Malden, USA.
Maisonneuve N., Stevens M., O chab B ., 2010. Participatory noise pollution monitoring
using mobile phones. Information Polity, 15, pp. 51-71.
Obrist M. K., Pavan G., Sueur J., Riede K., Llusia D., Márquez R., 2010. Bioacoustics
approaches in biodiversity inventories. Abc Taxa, 8, pp. 68-99.
Oliver I., Beattie A.J., 1993. A possible method for the rapid assessment of biodiversity.
Conservation Biology, 7, pp. 562-568.
Oliver I., Beattie A.J., 1996. Invertebrate morphospecies as surrogates for species: a case
study. Conservation Biology, 10, pp. 99-109.
Oliver I., Pik A., B ritton D., Dangerfield J. M., Colwel, R. K., B eattie A. J., 2000. Virtual
biodiversity assessment systems. BioScience, 50, pp. 441-450.
Pavoine S., Love M. S., B onsall M. B ., 2009. Hierarchical partitioning of evolutionary and
ecological patterns in the organization of phylogeneticallystructured species
assemblages: application to rockfish (genus: Sebastes) in the Southern California Bight.
Ecology letters, 12, pp. 898-908.
Pavoine S., B onsall M. B ., 2010. Measuring biodiversity to explain community assembly: a
unified approach. Biological Reviews, 86, pp. 792-812.
Pearson D. L., 1994. Selecting indicator taxa for the quantitative assessment of biodiversity.
Philosophical Transactions of the Royal Society of London B, 345, pp. 75-79.
Petchey O . L., O ’Gorman E., Flynn D. F. B ., 2009. A functional guide to functional
diversity measures, in: Naeem S., B unker D., Hector A., Loreau M., Perrings, C. (Eds)
Biodiversity, Ecosystem functioning, and Human wellbeing: An Ecological and
economic perspective. O xford University Press, O xford, pp. 49-60.
Pieretti N., Farina A., Morri F.D., 2010. A new methodology to infer the singing activity of an
avian community: the Acoustic Complexity Index (ACI). Ecological Indicators, 11, pp.
868-873.
Qi J., Gage S. H., Wooyeong J., Napoletano B ., Biswas, S., 2008. Soundscape characteristics
of an environnement. A new ecological indicator of ecosystem health, in: Wei J. (Ed)

209
Wetland and water resource modeling and assessment, a watershed perspective. CRC
Press, B oca Raton, pp. 201-211.
R Development Core Team, 2012. The R Project for Statistical Computing. Accessed on
February 14, 2012, http://www.r-project.org/.
Rao C. R., 1964. The use and interpretation of principal component analysis in applied
research. Sankhya A, 26, pp. 329-359.
Ricketts T. H., Dinerstein E., Olson D. M., Loucks C., 1999. Who’s where in North America?
BioScience, 49, pp. 369-381.
Riede K., 1993. Monitoring biodiversity: analysis of Amazonian rainforest sounds. Ambio,
22, pp. 546-548.
Rumsey F., McCormick T., 1992. Sound and recording: an introduction. Focal Press, B oston,
USA.
Skowronski M. D., Harris J. G., 2006. Acoustic detection and classification of
Microchiroptera using machine learning: lessons learned from automatic speech
recognition. Journal of the Acoustical Society of America, 119, pp. 1817-1833.
Stoddard P. K., 1998. Application of filters in bioacoustics, in: Hopp. S. L., Owren M. J.,
Evans C. S. (Eds) Animal acoustic communication. Springer, Berlin, pp. 105-127.
Suarez A. V., Tsutsui N. D., 2004. The value of Museum collections for research and society.
Bioscience, 54, pp. 66-74.
Sueur J., 2002. Cicada acoustic communication: potential sound partitioning in a multispecies
community from Mexico (Hemiptera: Cicadomorpha: Cicadidae). Biological Journal of
the Linnean Society, 75, pp. 379-394.
Sueur J., Pavoine S., Hamerlynck O ., Duvail S., 2008a. Rapid acoustic survey for
biodiversity appraisal. PloS O ne, 3, e4065. Sueur J., Aubin T., Simonis C., 2008b.
Seewave: a free modular tool for sound analysis and synthesis. Bioacoustics, 18, pp.
213-226.
Turner W., Spector S., Gardiner N., Fladeland M., Sterling E., Steininger M., 2003. Remote
sensing for biodiversity science and conservation. Trends in Ecology and Evolution, 18,
pp. 306-314.
Vane-Wright R. I., Humphries C. J., Williams P. H., 1991. What to protect? – Systematics
and the agony of choice. Biological Conservation, 55, pp. 235-254.
Vaseghi S.V., 2000. Advanced digital signal processing and noise reduction, second ed.
Wiley, Chichester, UK.

210
Wang K., Franklin S.E., Guo X., Cattet M. 2010. Remote sensing of ecology, biodiversity and
conservation: a review from the perspective of remote sensing specialists. Sensors, 10,
pp. 9647-9667.
Weaver J.C., 1995. Indicator species and scale of observation. Conservation Biology, 9, pp.
939-942.

211
Annexe 2 Matériel supplémentaire du
chapitre 2

212
213
Table S.II.1: Liste des 196 espèces d'oiseaux considérées pour l'étude.
Species Accipiter gentilis Accipiter nisus Acrocephalus arundinaceus Acrocephalus schoenobaenus Acrocephalus scirpaceus Aegithalos caudatus Alauda arvensis Alcedo atthis Alectoris graeca Alectoris rufa Anas crecca Anas platyrhynchos Anas querquedula Anas strepera Anser anser Anthus campestris Anthus trivialis Anthus pratensis Anthus spinoletta Apus apus Apus melba Apus pallidus Aquila chrysaetos Ardea alba Ardea purpurea Ardeola ralloides Asio otus Athene noctua Aythya ferina Aythya fuligula Bonasa bonasia Botaurus stellaris Bubo bubo Bubulcus ibis Burhinus oedicnemus Buteo buteo Calandrella brachydactyla Caprimulgus europaeus Carduelis cannabina Carduelis carduelis Carduelis chloris Carduelis flammea Carduelis spinus Certhia brachydactyla Certhia familiaris Cettia cetti Charadrius alexandrinus Charadrius dubius Chlidonias hybridus Cinclus cinclus Circaetus gallicus Circus aeruginosus Circus cyaneus Circus pygargus Cisticola juncidis
214
Clamator glandarius Columba oenas Columba palumbus Coracias garrulus Corvus corax Corvus corone Corvus frugilegus Corvus monedula Coturnix coturnix Crex crex Cuculus canorus Cygnus olor Delichon urbica Dendrocopos major Dendrocopos medius Dendrocopos minor Dryocopus martius Egretta garzetta Emberiza cia Emberiza cirlus Emberiza citrinella Emberiza hortulana Emberiza schoeniclus Erithacus rubecula Falco naumanni Falco peregrinus Falco tinnunculus Ficedula hypoleuca Fringilla coelebs Galerida cristata Gallinago gallinago Gallinula chloropus Garrulus glandarius Gelochelidon nilotica Haematopus ostralegus Hieraaetus pennatus Himantopus himantopus Hippolais icterina Hippolais polyglotta Ixobrychus minutus Jynx torquilla Lagopus mutus Lanius collurio Lanius excubitor Lanius meridionalis Lanius senator Larus fuscus Larus marinus Larus ridibundus Locustella luscinioides Locustella naevia Lullula arborea Luscinia megarhynchos Luscinia svecica Melanocorypha calandra Merops apiaster Miliaria calandra Milvus migrans
215
Milvus milvus Monticola saxatilis Monticola solitarius Motacilla alba Motacilla cinerea Motacilla flava Muscicapa striata Numenius arquata Oenanthe hispanica Oenanthe oenanthe Oriolus oriolus Otus scops Pandion haliaetus Panurus biarmicus Parus ater Parus caeruleus Parus cristatus Parus major Parus montanus Parus palustris Passer domesticus Passer montanus Perdix perdix Pernis apivorus Petronia petronia Phalacrocorax aristotelis Phalacrocorax carbo Phoenicurus ochruros Phoenicurus phoenicurus Phylloscopus bonelli Phylloscopus collybita Phylloscopus sibilatrix Phylloscopus trochilus Pica pica Picus canus Picus viridis Podiceps cristatus Podiceps nigricollis Porzana porzana Prunella collaris Prunella modularis Ptyonoprogne rupestris Pyrrhocorax graculus Pyrrhocorax pyrrhocorax Pyrrhula pyrrhula Rallus aquaticus Recurvirostra avosetta Regulus ignicapillus Regulus regulus Riparia riparia Saxicola rubetra Saxicola torquata Scolopax rusticola Serinus citrinella Serinus serinus Sitta europaea Sterna albifrons Sterna hirundo
216
Sterna sandvicensis Streptopelia decaocto Streptopelia turtur Strix aluco Sturnus vulgaris Sylvia atricapilla Sylvia borin Sylvia cantillans Sylvia communis Sylvia conspicillata Sylvia curruca Sylvia hortensis Sylvia melanocephala Sylvia undata Tetrax tetrax Tichodroma muraria Troglodytes troglodytes Turdus merula Turdus philomelos Turdus pilaris Turdus torquatus Turdus viscivorus Tyto alba Upupa epops Vanellus vanellus
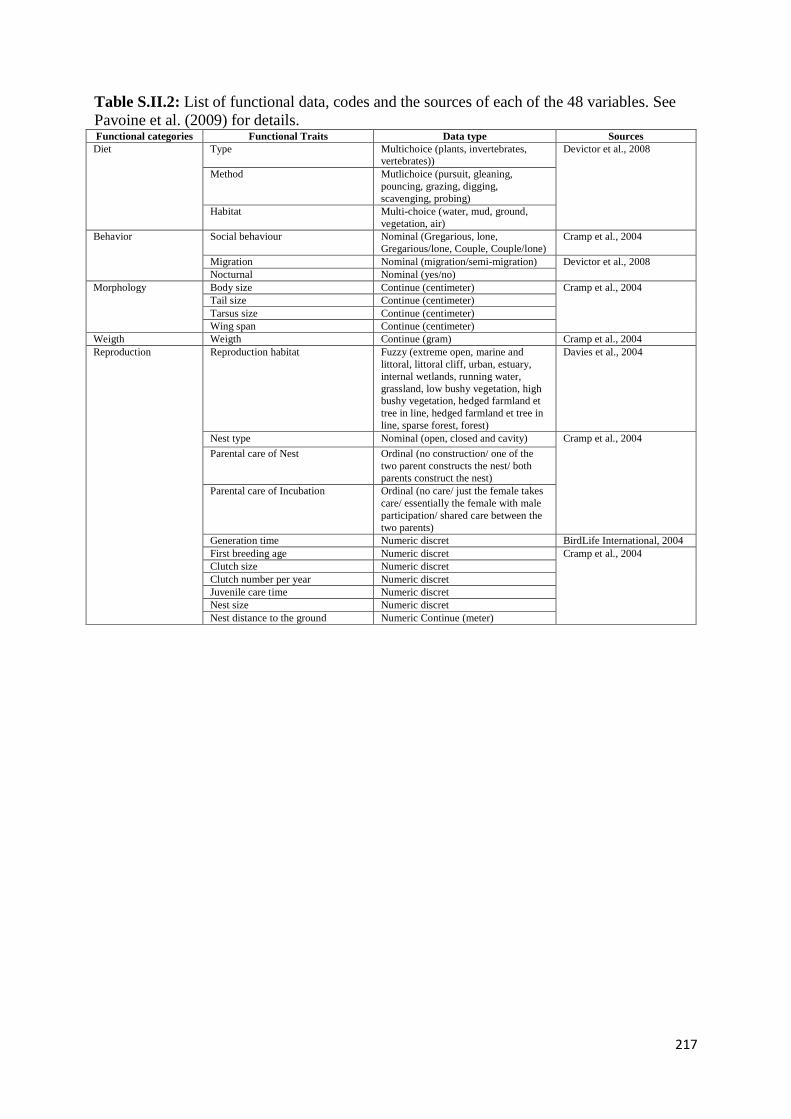
217
Table S.II.2: List of functional data, codes and the sources of each of the 48 variables. See Pavoine et al. (2009) for details. Functional categories Functional Traits Data type Sources Diet Type Multichoice (plants, invertebrates,
vertebrates)) Devictor et al., 2008
Method Mutlichoice (pursuit, gleaning, pouncing, grazing, digging, scavenging, probing)
Habitat Multi-choice (water, mud, ground, vegetation, air)
Behavior Social behaviour Nominal (Gregarious, lone, Gregarious/lone, Couple, Couple/lone)
Cramp et al., 2004
Migration Nominal (migration/semi-migration) Devictor et al., 2008 Nocturnal Nominal (yes/no)
Morphology Body size Continue (centimeter) Cramp et al., 2004 Tail size Continue (centimeter) Tarsus size Continue (centimeter) Wing span Continue (centimeter)
Weigth Weigth Continue (gram) Cramp et al., 2004 Reproduction Reproduction habitat Fuzzy (extreme open, marine and
littoral, littoral cliff, urban, estuary, internal wetlands, running water, grassland, low bushy vegetation, high bushy vegetation, hedged farmland et tree in line, hedged farmland et tree in line, sparse forest, forest)
Davies et al., 2004
Nest type Nominal (open, closed and cavity) Cramp et al., 2004 Parental care of Nest Ordinal (no construction/ one of the
two parent constructs the nest/ both parents construct the nest)
Parental care of Incubation Ordinal (no care/ just the female takes care/ essentially the female with male participation/ shared care between the two parents)
Generation time Numeric discret BirdLife International, 2004 First breeding age Numeric discret Cramp et al., 2004 Clutch size Numeric discret Clutch number per year Numeric discret Juvenile care time Numeric discret Nest size Numeric discret Nest distance to the ground Numeric Continue (meter)

218

219
Annexe 3 Matériel supplémentaire du
chapitre 3

220
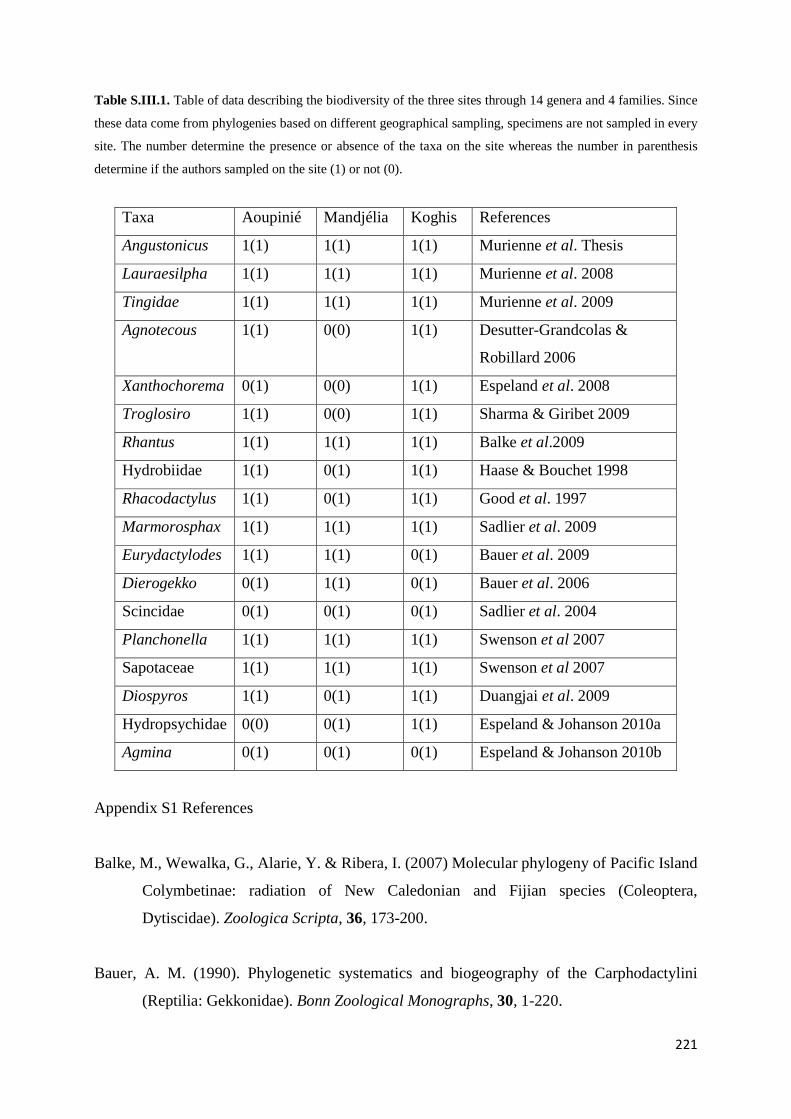
221
Table S.III.1. Table of data describing the biodiversity of the three sites through 14 genera and 4 families. Since
these data come from phylogenies based on different geographical sampling, specimens are not sampled in every
site. The number determine the presence or absence of the taxa on the site whereas the number in parenthesis
determine if the authors sampled on the site (1) or not (0).
Taxa Aoupinié Mandjélia Koghis References
Angustonicus 1(1) 1(1) 1(1) Murienne et al. Thesis
Lauraesilpha 1(1) 1(1) 1(1) Murienne et al. 2008
Tingidae 1(1) 1(1) 1(1) Murienne et al. 2009
Agnotecous 1(1) 0(0) 1(1) Desutter-Grandcolas &
Robillard 2006
Xanthochorema 0(1) 0(0) 1(1) Espeland et al. 2008
Troglosiro 1(1) 0(0) 1(1) Sharma & Giribet 2009
Rhantus 1(1) 1(1) 1(1) Balke et al.2009
Hydrobiidae 1(1) 0(1) 1(1) Haase & Bouchet 1998
Rhacodactylus 1(1) 0(1) 1(1) Good et al. 1997
Marmorosphax 1(1) 1(1) 1(1) Sadlier et al. 2009
Eurydactylodes 1(1) 1(1) 0(1) Bauer et al. 2009
Dierogekko 0(1) 1(1) 0(1) Bauer et al. 2006
Scincidae 0(1) 0(1) 0(1) Sadlier et al. 2004
Planchonella 1(1) 1(1) 1(1) Swenson et al 2007
Sapotaceae 1(1) 1(1) 1(1) Swenson et al 2007
Diospyros 1(1) 0(1) 1(1) Duangjai et al. 2009
Hydropsychidae 0(0) 0(1) 1(1) Espeland & Johanson 2010a
Agmina 0(1) 0(1) 0(1) Espeland & Johanson 2010b
Appendix S1 References
Balke, M., Wewalka, G., Alarie, Y. & Ribera, I. (2007) Molecular phylogeny of Pacific Island
Colymbetinae: radiation of New Caledonian and Fijian species (Coleoptera,
Dytiscidae). Zoologica Scripta, 36, 173-200.
Bauer, A. M. (1990). Phylogenetic systematics and biogeography of the Carphodactylini
(Reptilia: Gekkonidae). Bonn Zoological Monographs, 30, 1-220.

222
Bauer, A. M., Jackman, T., Sadlier, R. A. & Whitaker, A. H. (2006) A Revision of the
Bavayia validiclavis group (Squamata: Gekkota: Diplodactylidae), a Clade of New
Caledonian Geckos Exhibiting Microendemism. Proceedings of the California
Academy of Sciences, 57, 503-547.
Bauer, A. M., Jackman, T., Sadlier, R. A. & Whitaker, A. H. ( 2009) Review and phylogeny
of the New Caledonian diplodactylid gekkotan genus Eurydactylodes Wermuth, 1965,
with the description of a new species. In Zoologia Neocaledonica 7, Systematics and
Biodiversity in New Caledonia (ed. Grandcolas, P.), pp. 13-36, Mémoires du Muséum
National d'Histoire Naturelle.
Desutter-Grandcolas, L. & Robillard, T. (2006) Phylogenetic systematics and evolution of
Agnotecous in New Caledonia (Orthoptera: Grylloidea, Eneopteridae). Systematic
Biology, 31, 65-92.
Duangjai, S. Samuel, R, Munzinger, J, Forest, F, Wallnöfer, B, Barfuss, M. H, Fischer G, &
Chase, M.W. (2009) A multi-locus plastid phylogenetic analysis of the pantropical
genus Diospyros (Ebenaceae), with an emphasis on the radiation and biogeographic
origins of the New Caledonian endemic species. Molecular Phylogenetics and
Evolution, 52, 602–620.
Espeland, M., Johanson, K. A. & Hovmöller, R. (2008) Early Xanthochorema (Trichoptera,
Insecta) radiations in New Caledonia originated on ultrabasic rocks. Molecular
Phylogenetics and Evolution, 48, 904-917.
Espeland, M. & Johanson, K. A. (2010a) The effect of environmental diversification on
species diversification in New Caledonian caddisflies (Insecta: Trichoptera:
Hydropsychidae). Journal of Biogeography, 37, 879-890.
Espeland, M. & Johanson, K. A. (2010b) The diversity and radiation of the largest
monophyletic animal group on New Caledonia (Trichoptera: Ecnomidae: Agmina).
Journal of Evolutionary Biology, 23, 2112-2122.

223
Good, D. A., Bauer, A. M. & Sadlier, R. A. (1997) Allozyme evidence for the phylogeny of
the giant New Caledonian geckos (Squamata: Diplodactylidae: Rhacodactylus), with
comments on the status of R. leachianus henkeli. Australian Journal of Zoology, 45,
317-330.
Haase, M. & Bouchet, P. (1998) Radiation of crenobiontic gastropods on an ancient
continental island: the Hemistomia-clade in New Caledonia (Gastropoda:
Hydrobiidae). Hydrobiologia, 367, 43-129.
Mann, D. A, Locascio J. V., Coleman, F. C. and Koening, C. C (2008) Goliath grouper
Epinephelus itajara sound production and movement patterns on aggregation sites.
Endangered species research, 7, 229–236.
Munzinger, J. & Swenson, U. (2009) Three new species of Planchonella (Sapotaceae) with a
dichotomous and an online key to the genus in New Caledonia. Adansonia, 31, 175-
189.
Murienne, J. (2006) Origine de la biodiversité en Nouvelle-Calédonie: Analyse
phylogénétique de l’endémisme chez les Insectes Dictyoptères, PhD Thesis,
Université Pierre et Marie Curie - Paris 6.
Murienne, J., Pellens, R., Budinoff, R. B., Wheeler, W. & Grandcolas, P. (2008) Phylogenetic
analysis of the endemic New Caledonian cockroach Lauraesilpha. Testing competing
hypothesis of diversification. Cladistics, 24, 802-812.
Murienne, J., Guilbert, E. & Grandcolas, P. (2009) Species’ diversity in the New Caledonian
endemic genera Cephalidiosus and Nobarnus (Insecta: Heteroptera: Tingidae), an
approach using phylogeny and species’ distribution modelling. Biological Journal of
the Linnean Society, 97, 177–184.
Sadlier, R. A., Smith, S. A., Bauer, A. M. & Whitaker, A. H. (2009) Three new species of
skink in the genus Marmorosphax Sadlier (Squamata: Sincidae) from New Caledonia.
In Zoologia Neocaledonica 7, Systematics and Biodiversity in New Caledonia (ed.
Grandcolas, P.), pp. 247-263. Mémoires du Muséum National d'Histoire Naturelle.

224
Sadlier, R. A., Smith, S. A., Bauer, A. M. & Whitaker, A. H. (2004) A new genus and species
of live-bearing Scincid lizard (Reptilia: Scincidae) from New Caledonia. Journal of
Herpetology, 38, 320-330.
Sharma, P. & Giribet, G. (2009) A relict in New Caledonia: phylogenetic relationships of the
family Troglosironidae (Opiliones: Cyphophthalmi). Cladistics, 25, 1-16.
Swenson, U., Munzinger, J. & Bartish, I. V. (2007) Molecular phylogeny of Planchonella
(Sapotaceae) and eight new species from New Caledonia. Taxon, 56, 329-354.
Swenson, U., Lowry II, P. P., Munzinger, J., Rydin, C. & Bartish, I. V. (2008) Phylogeny and
generic limits in the Niemeyera complex of New Caledonian Sapotaceae: evidence of
multiple origins of the anisomerous flower. Molecular Phylogenetics and Evolution,
49, 909-929.
Swenson, U. & Munzinger, J. (2009) Revision of Pycnandra subgenus Pycnandra
(Sapotaceae), a genus endemic to New Caledonia. Australian Systematic Botany, 22,
437–465.
Swenson, U. & Munzinger, J. (2010) Revision of Pycnandra subgenus Achradotypus
(Sapotaceae) with five new species from New Caledonia. Australian Systematic
Botany, 23, 185-216.
Swenson, U. & Munzinger, J. (2010) Revision of Pycnandra subgenus Sebertia (Sapotaceae)
and a generic key to the family in New Caledonia. Adansonia, 32, 239-249.
Swenson, U. & Munzinger, J. (2010) Taxonomic revision of Pycnandra subgenus Trouettia
(Sapotaceae) with six new species from New Caledonia. Australian Systematic
Botany, 23, 333-370.
Table S.III.2. Table of number and percentage of files associated to different noise types and number of files
after exclusion of the noisy files for each site.
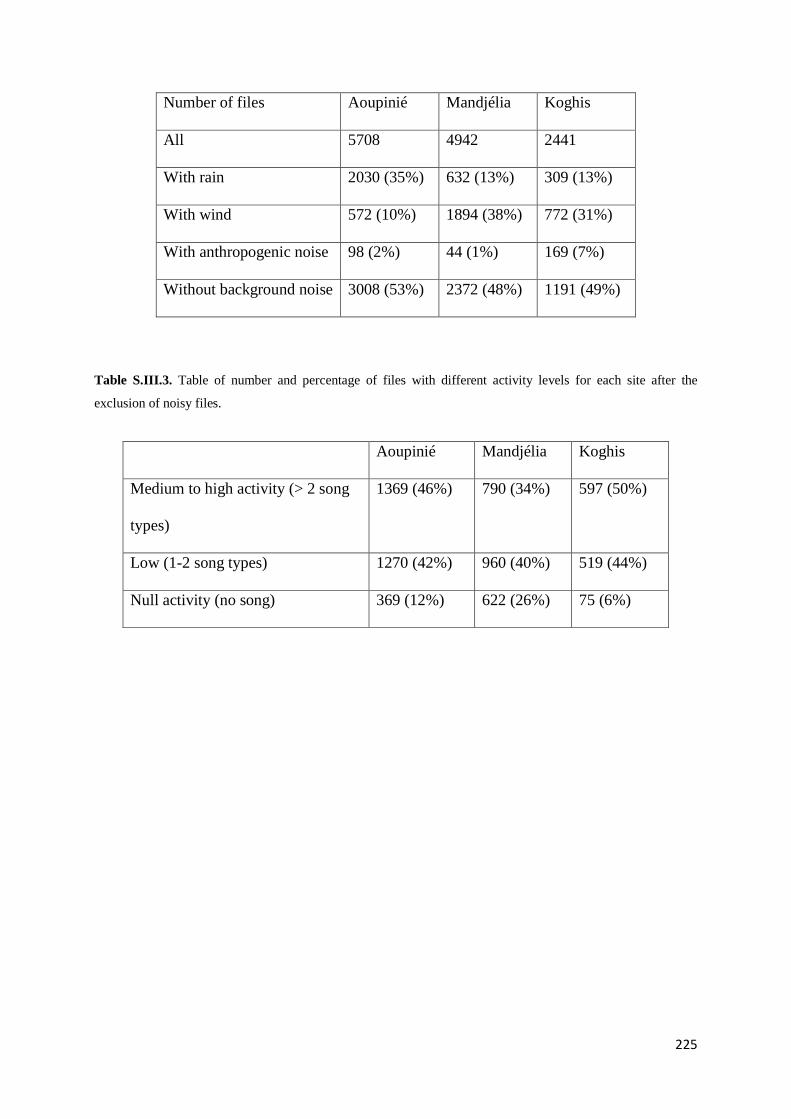
225
Number of files Aoupinié Mandjélia Koghis
All 5708 4942 2441
With rain 2030 (35%) 632 (13%) 309 (13%)
With wind 572 (10%) 1894 (38%) 772 (31%)
With anthropogenic noise 98 (2%) 44 (1%) 169 (7%)
Without background noise 3008 (53%) 2372 (48%) 1191 (49%)
Table S.III.3. Table of number and percentage of files with different activity levels for each site after the
exclusion of noisy files.
Aoupinié Mandjélia Koghis
Medium to high activity (> 2 song
types)
1369 (46%) 790 (34%) 597 (50%)
Low (1-2 song types) 1270 (42%) 960 (40%) 519 (44%)
Null activity (no song) 369 (12%) 622 (26%) 75 (6%)
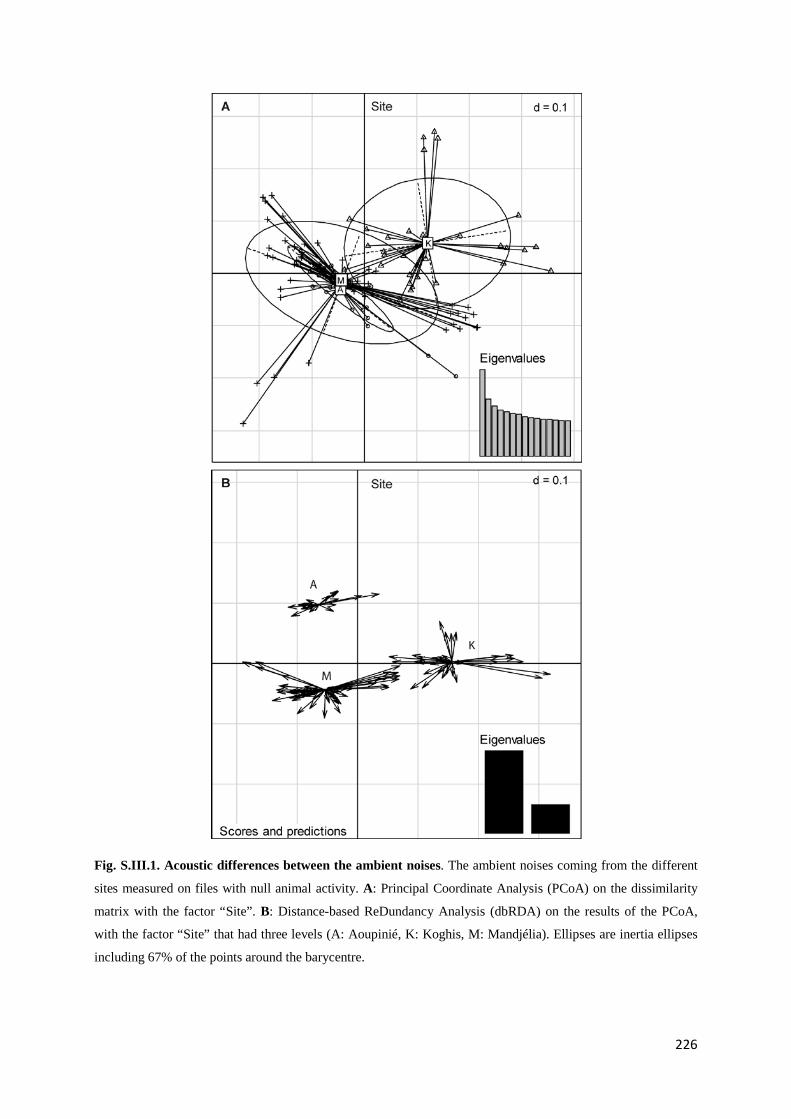
226
Fig. S.III.1. Acoustic differences between the ambient noises. The ambient noises coming from the different
sites measured on files with null animal activity. A: Principal Coordinate Analysis (PCoA) on the dissimilarity
matrix with the factor “Site”. B: Distance-based ReDundancy Analysis (dbRDA) on the results of the PCoA,
with the factor “Site” that had three levels (A: Aoupinié, K: Koghis, M: Mandjélia). Ellipses are inertia ellipses
including 67% of the points around the barycentre.
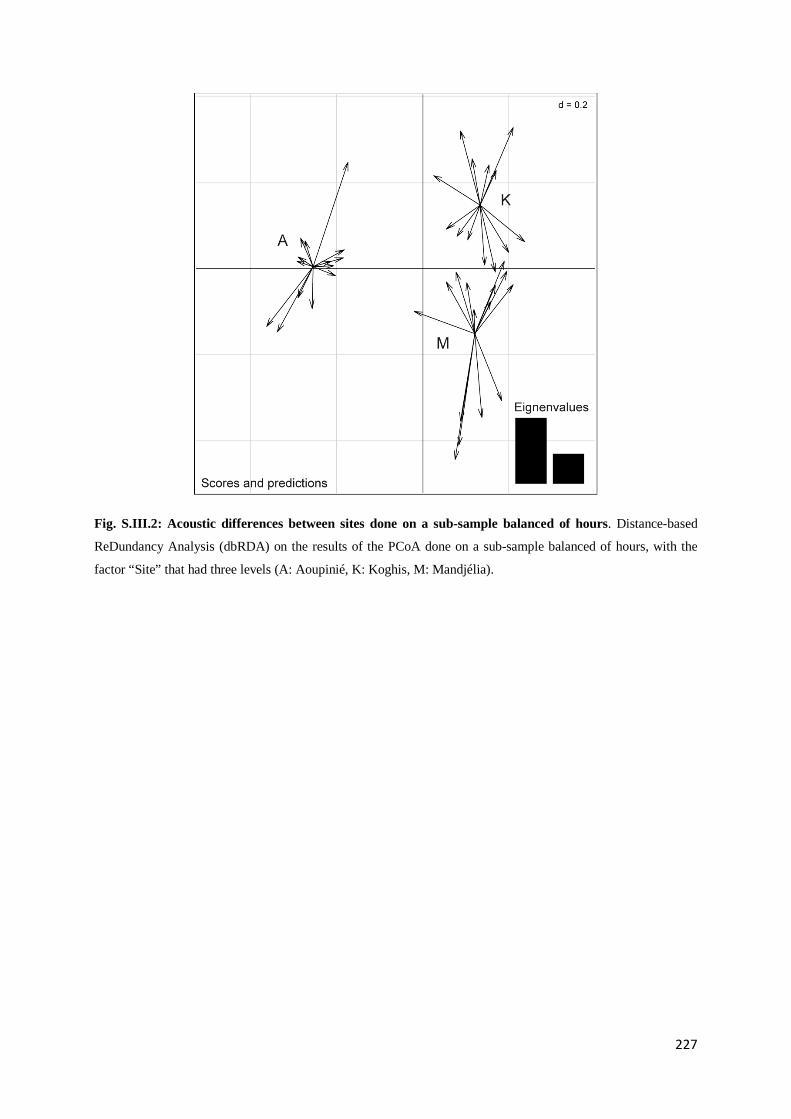
227
Fig. S.III.2: Acoustic differences between sites done on a sub-sample balanced of hours. Distance-based
ReDundancy Analysis (dbRDA) on the results of the PCoA done on a sub-sample balanced of hours, with the
factor “Site” that had three levels (A: Aoupinié, K: Koghis, M: Mandjélia).

228

229
Résumé En parallèle des méthodes classiques d'inventaires, qui demandent un haut niveau d'expertise taxonomique, des méthodes dites de mesure rapide de la biodiversité ont été proposées. Parmi ces méthodes, la méthode acoustique globale, ou Rapid Acoustic Survey (RAS), a été développée en 2008. L'objectif général de ce travail de thèse a été de fournir une définition plus précise de l’approche RAS d'un point de vue méthodologique, afin d'en connaître les possibles applications pour l'estimation, le suivi et l'analyse de la distribution spatiale et temporelle de la biodiversité animale. Des indices acoustiques mesurant l'entropie du signal sonore d'une communauté animale et la dissimilarité acoustique entre communautés ont déjà été proposés. La sensibilité de ces indices à différentes causes d'hétérogénéité acoustique sans lien avec la diversité acoustique a été évaluée (e.g. impact du bruit ambiant). Les résultats montrent que les indices ne sont pas sensibles de la même manière aux différentes causes d’hétérogénéité étudiées ; les biais générés par ces sources d’hétérogénéité doivent donc être traités différemment en fonction de l'indice de diversité utilisé. Une fois ces étapes méthodologiques traitées, plusieurs applications de l'approche ont révélé les points suivants. Dans les communautés d'oiseaux, en France continentale métropolitaine, des corrélations positives entre diversités acoustiques et phylogénétiques d'une part et entre diversités acoustiques et fonctionnelles (liées à la reproduction et au poids) d'autre part ont été identifiées. En Nouvelle-Calédonie, le taux de micro-endémisme est élevé sur l'ensemble du territoire et la biodiversité est donc difficile à décrire. L'utilisation de mesures de dissimilarités acoustiques a pu mettre en évidence des différences acoustiques dans trois régions éloignées de l'île, possédant des niveaux de richesse spécifique et des structures végétales semblables mais des compositions de communautés différentes dues au taux élevé de micro-endémisme. Cette différence dans la composition de communautés animales multi-taxons est donc reflétée par les analyses de diversité acoustique par la méthode RAS. En Inde, deux milieux différents de plantations agro-forestières de café ont été étudiées, l'une utilisant des espèces natives d'arbres et l'autre une espèce exotique. Une étude acoustique a permis de mettre en évidence des différences dans la diversité et la composition acoustique des communautés animales vivant dans ces deux types d'agro-forêt, révélant un impact de l'utilisation de l'espèce exotique sur les communautés d'espèces chanteuses. L'acoustique globale RAS semble donc être une méthode prometteuse pour l'évaluation de la diversité animale par les méthodes d'acoustique non invasives, même si certaines limites méthodologiques devront être dépassées pour fournir un outil utilisable dans de nombreuses conditions acoustiques.
Abstract Alongside conventional methods of inventories, which require a high level of taxonomic expertise, several methods known as rapid measurement of biodiversity have been proposed. Among these methods, an acoustic method, designated as Rapid Acoustic Survey (RAS), was developed in 2008. The general objective of my thesis has been to provide a better definition of RAS approach from a methodological point of view, in order to evaluate the possible applications of RAS for estimating, tracking and analysing the spatial and temporal distribution of animal biodiversity. Acoustic indices have already been proposed which measure the overall entropy of the sound signal in an animal community and the acoustic dissimilarity among communities. The sensitivity of these indices to different causes of heterogeneity unrelated acoustic diversity was assessed (e.g. ambient noise effect). The results show that the indices are not sensitive in the same way to the different causes of heterogeneity studied; the generated biased should thus be treated differently depending on the diversity index used. Once these methodological approaches have been treated, several applications of the RAS revealed the following points. In bird communities in Metropolitan, continental France, positive correlations between acoustic and phylogenetic diversity on the one hand and between acoustic and functional (reproduction traits and weight) diversity on the other hand have been identified. In New Caledonia, the level of micro-endemism is high which make the description of biodiversity difficult. The use of acoustic spectral dissimilarity measures highlight acoustic differences in three distant locations in the island with similar species richness and vegetation structure, but different in the composition of animal communities due to micro-endemism. This difference in multi-taxa animal communities is clearly revealed by the acoustic diversity analysis RAS. In India, two different managements of the coffee agroforest system are found, one using native species and the other one using an exotic species of shade trees. An acoustic survey highlighted differences in acoustic diversity and composition between the two managements revealing an impact of the use of exotic shade tree species on singing animal communities. The RAS method seems then a promising tool to assess local animal diversity even if some methodological issues should be considered to produce a reliable tool in several acoustic conditions.


















