
Modulation Implicite de la Dynamique de la Rivalité...
72
Modulation Implicite de la Dynamique de la Rivalité Binoculaire par une Tâche de Recherche Visuelle Adrien Chopin sous la direction de Pascal Mamassian (Laboratoire de Psychologie de la Perception) 2007 - 2008 2 e année de Master de Recherche en Sciences Cognitives (CogMaster)
Transcript of Modulation Implicite de la Dynamique de la Rivalité...

Modulation Implicite de la Dynamique de la
Rivalité Binoculaire par une Tâche de
Recherche Visuelle
Adrien Chopin
sous la direction de Pascal Mamassian
(Laboratoire de Psychologie de la Perception)
2007 - 2008
2e année de Master de Recherche en Sciences Cognitives (CogMaster)

2

3
Remerciements
Je tiens à remercier Pascal Mamassian pour sa compétence, sa sagacité et tout le temps qu’il a pu me
consacrer. Je remercie également Jean Lorenceau et Andrei Gorea pour leurs conseils, mais aussi Antoine
Barbot, pour ses commentaires et ses critiques, Jennifer Lalanne et ma famille pour leur patience infinie.
J’ai aussi une pensée reconnaissante pour les participants de l’expérience, pour Mohammed Ghezal et
Virginie Poilièvre.
Résumé
Quand deux stimuli très différents sont présentés à chacun des yeux, la rivalité
binoculaire s’installe : la perception alterne alors entre les deux stimuli. Dans cette
expérience, il est demandé au participant de rechercher une cible parmi des
distracteurs en rivalité binoculaire sur l’orientation. La cible apparaît plus
fréquemment dans l’œil droit et du côté orienté vers la droite. Les résultats
montrent que les stimuli de l’œil droit et d’orientation droite deviennent
dominants. Nous interprétons ces modulations implicites comme reflétant
l’utilisation de fonction de gain (théorie bayésienne). Les caractéristiques
temporelles des effets observés démontrent l’existence et l’indépendance de
compétitions inter-oculaire et inter-orientation.
Mots-Clés: Bistabilité, perception visuelle, apprentissage implicite, apprentissage
statistique, théorie bayésienne, effets attentionnels, compétition neuronale.

4
INTRODUCTION ....................................................................................................5
QU'EST-CE QUI EST EN COMPETITION PENDANT LA RIVALITE BINOCULAIRE? .....................7 Compétition interoculaire ..........................................................................................................................................7 Compétition entre attributs .......................................................................................................................................8 Alternatives ...........................................................................................................................................................10
LES EFFETS ATTENTIONNELS SUR LA DYNAMIQUE DE LA RIVALITE BINOCULAIRE ............12 APPLICATION DU CADRE BAYESIEN A LA RIVALITE BINOCULAIRE ...........................................15 BUT DE L'EXPERIENCE .......................................................................................................................17
METHODE ............................................................................................................. 21
PARTICIPANTS.......................................................................................................................................21 MATERIEL ..............................................................................................................................................21 STIMULI ..................................................................................................................................................22 PROCEDURE ..........................................................................................................................................23
Calibrage ...........................................................................................................................................................23 Tâche..................................................................................................................................................................24 Etape 1 : neutre................................................................................................................................................26 Etapes 2 et 3 : biaisées ..................................................................................................................................28 Etape 4 : neutre ...............................................................................................................................................30
HYPOTHESES ........................................................................................................................................30
RESULTATS............................................................................................................32
DOMINANCE..........................................................................................................................................32 ANALYSE AVANCEE DE LA DOMINANCE..........................................................................................37
Normalisation par un nombre fixé de renversements.........................................................................37 Normalisation par le premier maximum d’effet ...................................................................................40
RELATIONS ENTRE EFFET OCULAIRE ET EFFET D'ORIENTATION ..........................................42 DOMINANCE DE LA CIBLE..................................................................................................................44 DUREE DES PHASES .............................................................................................................................46 CONTROLE DE L'INDEPENDANCE ŒIL-ORIENTATION ..............................................................47 CONTROLE DU POURCENTAGE DE REPONSES CORRECTES .......................................................48
DISCUSSION...........................................................................................................50
MODULATION DE LA DOMINANCE DE L’ORIENTATION ET DE L’ŒIL BIAISES ......................50 Hypothèses alternatives écartées...............................................................................................................52
Hypothèses portant sur la force du stimulus..............................................................................................................52 Hypothèses portant sur l'effet de l'attention endogène............................................................................................53 Hypothèse concernant l'attention exogène ................................................................................................................54
EFFET PRECOCE ANTI-CIBLE ............................................................................................................54 Hypothèse du prétraitement..........................................................................................................................................55 Apprentissage statistique et hypothèse de facilitation transitoire ...........................................................................56
CONSEQUENCES SUR LA NATURE DES COMPETITIONS SOUS-JACENTES.................................57 Indépendance temporelle des modulations de la rivalité.............................................................................................58
CONCLUSION........................................................................................................60
BIBLIOGRAPHIE...................................................................................................62
ANNEXES ...............................................................................................................66
ANNEXE 1 : CALIBRAGE, PREFERENCES ROUGE/VERT ET DOMINANCE OCULAIRE............66 ANNEXE 2 : NORMALISATION PAR LE MAXIMUM D'EFFET ........................................................69 ANNEXE 3 : DONNEES INDIVIDUELLES, LISSAGES ET MAXIMA.................................................71 ANNEXE 4 : NORMALISATION PAR LE MOMENT DU MAXIMUM D'EFFET DE LA COULEUR
VERTE......................................................................................................................................................72

5
Introduction
Toute image est profondément ambiguë. Le système visuel est confronté en permanence
à de l'incertitude et il doit prendre en continu des décisions rapides sur l'interprétation
qu'il fait de la scène. Cette construction du monde visuel se fait la plupart du temps à
couvert de notre vie consciente (von Helmholtz, 1867/1962), sans effort, et sans que
nous réalisions vraiment la complexité des opérations nécessaires. Cette situation est
particulièrement évidente lors d'un phénomène qui est appelé bistabilité. Face à une
stimulation physique constante, lorsque nous regardons une image par exemple, la
perception peut alterner sans cesse entre deux interprétations. De nombreuses figures
suffisamment ambiguës pour générer de la bistabilité ont été découvertes, comme le cube
de Necker (1832), les mouvements de plaids (Hupé & Rubin, 2003; Wallach, 1935) ou le
« slant rivalry » (van Ee, 2005; van Ee, van Dam, & Erkelens, 2002) dans lequel une
interprétation utilisant la disparité est en compétition avec une autre s'appuyant sur la
forme géométrique. C'est pourquoi depuis sa découverte (DuTour, 1760) et son étude
systématique il y a un siècle (Breese, 1899; Diaz-Caneja, 1928), la bistabilité ne cesse de
susciter une littérature foisonnante et de nourrir les débats philosophiques, de
Wittgenstein (1953) à son utilisation récente pour comprendre la conscience (Block, 2005;
Crick & Koch, 2003). La bistabilité est ainsi mise à profit en tant que paradigme
expérimental pour comprendre le système perceptif (Haijiang, Saunders, Stone, & Backus,
2006; Lorenceau & Shiffrar, 1992) et ses mécanismes neuronaux (Fries, Roelfsema, Engel,
Konig, & Singer, 1997; Fries, Schroder, Roelfsema, Singer, & Engel, 2002; Srinivasan,
Russell, Edelman, & Tononi, 1999). L'un des cas les plus étudiés de la bistabilité est la
rivalité binoculaire, qui a lieu lorsque deux images très différentes sont présentées à

6
chaque œil de manière prolongée. Pourtant, de nombreuses interrogations sur la nature
même de la rivalité ne connaissent que des réponses partielles (Blake & Logothetis, 2002).
Notamment, il y a toujours eu une opposition entre les théories cherchant la cause dans
un mécanisme central et celles impliquant une compétition périphérique (Walker, 1978).
Les premières étaient fréquemment favorisées pour expliquer la bistabilité en général
tandis que les secondes semblaient rendre mieux compte du cas particulier de la rivalité
binoculaire. De récents travaux remettent en cause cette vision en mettant en lumière le
caractère actif du sujet dans la rivalité binoculaire par l'intermédiaire d'effets attentionnels
(Ooi & He, 1999; van Ee, van Dam, & Brouwer, 2005).
D'autre part, le cadre théorique bayésien a été développé précisément pour résoudre les
problèmes de décision sous incertitude. Il a été appliqué avec succès pour comprendre
comment le système visuel choisit l'interprétation de la scène la plus probable parmi les
différentes possibilités (Kersten, Mamassian, & Yuille, 2004; Mamassian, 2006). Il apparaît
donc comme un outil idéal pour comprendre le mécanisme bistable (Mamassian & Landy,
1998; Sundareswara & Schrater, 2008).
Cette étude s'inscrit dans cette double problématique. Nous souhaitons savoir si d'une
part la dynamique de la rivalité binoculaire peut être influencée par une forme d'attention
inconsciente basée sur des régularités statistiques et si, d'autre part le système visuel
cherche l'interprétation la plus probable de la scène, ou alors la plus utile étant donné la
tâche que le sujet est en train d'accomplir.

7
Qu'est-ce qui est en compétition pendant la rivalité binoculaire?
Poser cette question revient en fait à se demander quelle est la nature du câblage neuronal
qui permet la bistabilité ou la rivalité binoculaire: quelles structures cérébrales s'opposent
ou quels sont les attributs pour lesquels codent les structures neuronales en compétition?
Trois ensembles de théories répondent à cette question : les théories interoculaires, les
théories de compétitions entre représentations de plus haut-niveau et les théories
intermédiaires.
Compétition interoculaire
Pour Levelt (1966), la compétition oppose les représentations des images présentées à
chaque œil. Il en est de même pour Blake (1989), qui propose la théorie neuronale de la
rivalité binoculaire et un réseau neuronal minimum possible pour en rendre compte
(Figure 1). Dans celui-ci, les neurones monoculaires orientés tentent de supprimer
l'activité des neurones monoculaires de l'autre œil (dans un espace donné, e.g. une colonne
corticale) quels que soient les autres attributs codés (orientation, fréquence spatiale).
L'adaptation et l'inhibition réciproque seraient donc les principales causes de la rivalité
binoculaire.
Parmi, les dernières preuves en faveur de ce modèle, Leopold et Logothetis (1996) ont
trouvé par des mesures électro-physiologiques que 18% des neurones de V1-V2
répondaient en accord avec le percept, ce qui est faible. Mais une autre lecture des
données indique que 21% des neurones binoculaires orientés et 33% des monoculaires

8
orientés s'activent avec le percept. D'autre part, Tong et Engel (2001) étudient l'activité
hémodynamique (IRMf) dans la région de la tâche aveugle pendant la rivalité binoculaire
et trouvent des activités corrélées avec la perception bistable. Comme cette zone d'intérêt
est strictement monoculaire, ils en déduisent que la compétition est interoculaire.
Figure 1 : Modèle proposé par Blake (1989), dans le cadre de la théorie neuronale de la rivalité
binoculaire. Il simule un XOR (OU exclusif). Lorsque le signal est fort, la bistabilité s'installe. Lorsqu'il est
faible, un système de fusion prend le relais. L'inhibition est symbolisée par un trait terminé par un cercle
plein. Chaque inter-neurone monoculaire inhibe tous les neurones orientés de l'autre œil, ce qui est une
compétition interoculaire.
Compétition entre attributs
Outre l'œil d'origine, les neurones peuvent coder et s'opposer sur des informations
comme l'orientation ou la fréquence spatiale. Ainsi, dans les mesures de Leopold et
Logothetis (1996), aucun des neurones monoculaires non-orientés ne modulait son
activité en accord avec la perception. Il semble donc nécessaire que les neurones codent

9
pour une orientation dans la rivalité binoculaire, ce qui est en faveur d'une compétition
entre orientations. L'expérience de Tong et Engel (2001) peut elle-même être
réinterprétée comme une compétition entre neurones orientés (Andrews, 2001). Deux
expériences ont fortement jeté le doute sur l'hypothèse interoculaire. Celle de Logothetis,
Leopold et Sheinberg (1996), dite de rivalité permutée, présente aux observateurs des
réseaux de fréquence spatiale papillotants à une fréquence de 18 Hz tandis que les stimuli
sont échangés à une fréquence de 3 Hz. Dans ces conditions, une compétition
interoculaire donnerait des phases très courtes, tandis que ce qui est observé est une
rivalité dont les phases sont normales. L'autre expérience est celle de Kovacs,
Papathomas, Yang et Feher (1996) dans laquelle deux stimuli-puzzles sont formés en
échangeant certaines pièces de deux stimuli cohérents. La perception est alors multi-
stable, alternant entre les deux puzzles et les deux images reconstituées, ce qu'une
compétition interoculaire est incapable d'expliquer. L'hypothèse de compétition
interoculaire est aussi mise en difficulté par la rivalité monoculaire (Andrews & Purves,
1997) pendant laquelle des réseaux de couleur et d'orientation différentes sont présentés
superposés monoculairement, ce qui déclenche une bistabilité. L'hypothèse ne peut bien-
sûr pas non plus rendre compte de toutes les figures ambiguës.
Ainsi, il est clair qu'une compétition entre attributs existe dans la bistabilité et la rivalité
binoculaire. Par contre, il n'existe que très peu de preuves en faveur de l'hypothèse
interoculaire qui ne puissent être réinterprétées. La difficulté vient du fait que les neurones
codant pour l'œil d'origine sont pour la plupart orientés. Des expériences supplémentaires
sont donc clairement nécessaires pour séparer les effets potentiels de chaque compétition.

10
Alternatives
Il existe des théories hybrides comme le modèle computationnel de Wilson (2003). Celui-
ci implique une hiérarchie entre une première couche de neurones monoculaires orientés
envoyant des connexions antagonistes sélectives à l'orientation sur les neurones codant
pour l'autre œil, et une seconde couche binoculaire instaurant une compétition inter-
orientation pure (Figure 2). Ce modèle permet ainsi de rendre compte de la rivalité
permutée (Logothetis, Leopold, & Sheinberg, 1996) sans pour autant expliquer les
groupements interoculaires (Kovacs, Papathomas, Yang, & Feher, 1996).
Enfin, il est intéressant de signaler que d'autres hypothèses de compétitions sont étudiées.
Par exemple, Pettigrew (2001) a proposé une compétition inter-hémisphérique. Une
expérience d'Oshea (2003) lui a porté un coup fatal en démontrant que la rivalité était
normale chez les patients au corps calleux sectionné. D'autre part, Mitchell, Stoner et
Reynolds (2004) propose une compétition inter-objet, donc entre représentations issues
d'un liage perceptif. Il en apporte une preuve grâce à un subtil paradigme. Celui-ci
implique deux surfaces de points en mouvement. L'attention basée sur l'objet est orientée
sur l'une d'elle : un renversement a alors lieu sur cette surface, phénomène qui nécessite
qu'une compétition inter-objet existe afin d'être biaisée.
Il existe de nombreux facteurs qui peuvent influencer la bistabilité en général et la rivalité
binoculaire en particulier (Leopold & Logothetis, 1999; Walker, 1978). Levelt (1966)
s'aperçoit que plus le stimulus présenté dans un œil contient de contours, plus il est perçu
longtemps. Il définit donc la force d'un stimulus par sa quantité de contours et son
contraste. Il élabore ensuite le deuxième principe selon lequel plus la force d'un stimulus

11
est augmentée dans un œil, plus la durée des phases de l'autre œil est diminuée (une phase
est le temps qui sépare deux renversements). Cela veut dire que la force d'un stimulus
correspond à son potentiel de suppression du stimulus en compétition (Levelt, 1966;
Mamassian & Goutcher, 2005). Ce principe est considéré comme un des principaux
résultats sur les aspects de la dominance dirigés par le stimulus. Il a été récemment corrigé
par de récents travaux (Brascamp, van Ee, Noest, Jacobs, & van den Berg, 2006), qui
montrent que le principe est seulement partiellement validé dans des conditions
différentes. La règle serait plutôt qu'une modification de la force (i.e. du contraste) d'un
stimulus affecte principalement les durées de phase du stimulus de plus fort contraste.
Cette étude isole aussi le bruit neuronal comme un des principaux facteurs de la rivalité
binoculaire.
Figure 2 : Modèle neuronal hybride de Wilson (2003) impliquant deux niveaux. Le niveau du bas
comprend des neurones monoculaires orientés inhibant les neurones monoculaires orientés de l'autre œil
et d'une orientation différente (lignes terminées par un cercle plein, inclut un inter-neurone). Ils excitent
une seconde couche de neurone recevant des signaux binoculairement des neurones de même orientation

12
(flèche simple, qui correspondent aussi à un feedback vers le niveau inférieur). Ces neurones sont eux-
mêmes en compétition.
Pourtant, la rivalité n'est pas uniquement influencée par les mécanismes d'adaptation,
d'inhibition réciproque et de bruit neuronal. L'attention peut aussi influencer la
dynamique de la rivalité. Nous allons donc passer en revue les principales expériences sur
le sujet.
Les effets attentionnels sur la dynamique de la rivalité binoculaire
L'attention peut être définie sur deux aspects: comme une ressource (Kahneman, 1973) et
comme un mécanisme de sélection (Broadbent, 1958). En tant que mécanisme sélectif,
l'attention dirigée vers des attributs permet de sélectionner tous les éléments ayant
certains attributs (comme une même couleur) et pas les autres. Broadbent (1958)
développe la théorie du filtre précoce de l'attention. Selon lui, seule l'information
structurale grossière est analysée précocement, l'attention bloquant la plupart de celle-ci,
et n'en laissant passer qu'une petite partie pour un traitement de haut niveau. Certaines
expériences montrent pourtant qu'un traitement de haut niveau est possible là où
l'attention ne se porte pas (Moray, 1959), menant aux théories de filtre tardif, ou à une
version allégée du filtre précoce, qui est la théorie de l'atténuation (Treisman, 1960). Selon
cette dernière, le filtre ne fait qu'atténuer l'information non-sélectionnée, la rendant tout
de même disponible en cas de besoin, par l'intermédiaire de mémoires sensorielles.
L'attention spatiale consiste à porter son attention sur une zone de l'espace visuel en y

13
sélectionnant tous les attributs. Posner, Nissen et Ogden (1978) développent le paradigme
d'indiçage attentionnel qui permet de différencier les attentions endogène et exogène. Le
paradigme est une tâche de détection de cible d'un côté ou d'un autre. Dans le cas de
l'attention exogène, un indice est flashé avant la cible soit du même côté (valide) soit de
l'autre côté (invalide). Les temps de réaction étant plus courts dans les essais valides, c'est
que l'attention a été involontairement attirée vers l'indice. Avec un indice central, une
flèche montrant où va être flashé la cible dans 80% des cas, il est possible de mettre en
évidence l'attention endogène. Dans ce cas, le sujet a intérêt à volontairement détourner son
attention du côté indicé. Il en résulte encore des temps de réaction plus courts pour les
cas valides que pour les cas invalides, ce qui n'est pas lieu si l'indice central ne prédit pas la
position de la cible. Par extension, l'attention endogène regroupe tous les phénomènes
attentionnels dans lequel le sujet a conscience de volontairement déplacer son attention
tandis que l'attention exogène est inconsciemment dirigée (comme dans le pop-out ou
lors d'un mouvement inattendu).
L'attention peut aussi influencer la bistabilité (Paffen, Alais, & Verstraten, 2006). Elle fut
d'ailleurs historiquement considérée comme le moteur de la rivalité binoculaire (von
Helmholtz, 1867/1962), mais l'hypothèse fut aussi combattue (Pastukhov & Braun, 2007).
Un observateur peut par exemple forcer dans une certaine mesure un stimulus à être
dominant. Ooi et He (1999) ont ainsi mis en évidence grâce à un paradigme de Posner en
rivalité que le renversement d'un percept pouvait être volontairement empêché. Van Ee,
van Dam et Brouwer (2005) ont confirmé ces résultats en les étendant à d'autres stimuli
bistables (Visage-maison; cube de Necker et 'Slant-rivalry'). D'autre part, ils montrent que

14
les stimuli peuvent être non seulement maintenus mais aussi volontairement renversés et
le nombre de renversements augmenté.
Meng et Tong (2004) ont de plus réussi à séparer les effets dirigés par le stimulus des
effets de l'attention endogène. Grâce à un ingénieux procédé, ils montrent que la rivalité
binoculaire diffère des autres bistabilités par le fait que les forces dirigées par le stimulus y
sont bien plus efficaces que celles de l'attention endogène.
Enfin, l'effet de l'attention exogène a aussi été étudié par Ooi et He (1999). Cet effet était
déjà connu par les paradigmes de perturbation. Ainsi, un mouvement dans un stimulus
supprimé le rend automatiquement dominant. L'intérêt des travaux de Ooi et He est que
seul l'effet pop-out est utilisé pour rendre le stimulus dominant, c'est-à-dire une attention
exogène basée sur un attribut (ici l'orientation).
Un troisième type d’attention a été mis en évidence. Lorsque des régularités existent entre
la position d’une cible et une configuration de distracteurs, une forme d’attention
implicite peut être rapidement déployée vers les positions les plus probables, ce qui
diminue les temps de réaction (Chun, 2000; Chun & Jiang, 1998, 1999; Jiang & Chun,
2001). Comment cette attention basée sur des statistiques peut influencer la bistabilité est
une question, qui, à notre connaissance, n’a pas encore été étudiée.
Les études sur l'attention et la rivalité binoculaire ont montré que le sujet pouvait être
acteur de cette bistabilité et que la rivalité était influencée par un mécanisme central. Cela
change le point de vue sur la fonction de la rivalité. Selon la théorie neuronale de la
rivalité binoculaire (Blake, 1989), la rivalité est simplement le résultat par défaut d'une
fusion impossible entre les deux stimuli (la fusion étant elle-même un processus différent

15
de celui qui extrait l'information stéréoscopique ; Wolfe, 1986). A l'opposé, pour Leopold
et Logothetis (1999), la bistabilité fait partie d'un processus actif général qui est retrouvé
dans la perception normale pendant les comportements exploratoires spontanés. Ainsi la
bistabilité partage la même dynamique moyenne que les saccades exploratoires
(distribution gamma des durées de phase, indépendance des durées de phases successives)
et implique des structures cérébrales similaires selon certaines études d'imageries
(notamment le cortex préfrontal: Lumer, Friston, & Rees, 1998; Lumer & Rees, 1999). La
bistabilité ne serait donc que le cas limite de l'exploration perceptive où les interprétations
possibles sont d'égale force. Choisir parmi différentes interprétations est le problème type
dans lequel l'inférence bayésienne peut être utile.
Application du cadre bayésien à la rivalité binoculaire
Le principe de l'inférence bayésienne est de calculer une distribution a posteriori d’une
variable à estimer qui représente la probabilité des différentes possibilités de cette variable,
étant donnée une image. Une distribution de probabilités (dite vraisemblance) de la
variable à estimer est d’abord calculée à partir des règles de projection (par exemple) : elle
correspond à la probabilité d’obtenir cette image pour chaque valeur de la variable. Puis il
faut calculer une distribution a priori de la variable qui représente les connaissances du
système. Certains auteurs utilisent des a priori uniformes (Ernst & Banks, 2002). Pour
obtenir une distribution a posteriori, il suffit de multiplier la distribution de vraisemblance
par la distribution a priori. Dans une expérience contrôlée, les expérimentateurs peuvent
ainsi calculer comment un observateur idéal se comporterait. Il a été montré à plusieurs

16
reprises que le système visuel dans certaines conditions était comparable à un système
bayésien idéal (Kersten, Mamassian, & Yuille, 2004; Mamassian, 2006; Mamassian, Landy,
& Maloney, 2002), de même que le système d'apprentissage sensori-moteur (Kording &
Wolpert, 2004). Il existe en fait plusieurs règles possibles sur la manière de choisir une
interprétation unique à partir de la distribution a posteriori. Le système peut par exemple
choisir le maximum a posteriori. Une autre possibilité est d'appliquer une règle de
correspondance des probabilités, c'est-à-dire que la probabilité qu'une interprétation soit
perçue correspond à sa probabilité a posteriori. Mamassian et Landy (1998) utilisent cette
règle dans une expérience qui teste à la base la validité de certaines connaissances a priori
pour expliquer la manière dont les surfaces sont perceptivement construites en utilisant le
cadre théorique bayésien. Les stimuli utilisés étant bistables (ils pouvaient en effet être
interprétés comme concave ou convexe), les auteurs trouvent en effet que la probabilité
de la réponse concave (par exemple) sur le premier percept correspond à la probabilité a
posteriori de l'interprétation concave. Dans les scènes complexes de la vie réelle, la
redondance de l'information fait qu'une interprétation est souvent plus que majoritaire. La
bistabilité serait alors effectivement le cas limite dans lequel plusieurs interprétations ont
de fortes probabilités a posteriori.
Mais les théories de l'inférence bayésienne préconisent une étape supplémentaire après
l'obtention de la distribution a posteriori. Il s'agit d'ajouter une fonction de gain dépendante
de la tâche (Mamassian, Landy, & Maloney, 2002), qui pondère les probabilités a posteriori
par le gain (ou la perte) attendu à statuer en faveur d'une interprétation. Certains auteurs
ont montré la validité des fonctions de gain dans la sélection motrice (Trommershäuser,
Maloney, & Landy, 2003). Nous pensons donc que le système visuel tire bénéfice de ces

17
fonctions de gain et que le paradigme bistable peut permettre de les mettre en évidence
qualitativement.
But de l'expérience
L’expérience s’inscrit dans un projet dont le but est de montrer que le système visuel
utilise des fonctions de gain. Autrement dit, nous proposons que le système visuel
cherche l'interprétation la plus utile, c'est-à-dire un compromis entre l'interprétation la
plus probable et celle qui apporterait le plus de gains.
Dans l’expérience présente, nous cherchons à biaiser la dynamique de la rivalité
binoculaire, c'est-à-dire faire en sorte que l'un des percepts soit vu plus souvent que l'autre
au nom d'une fonction de gain. Mais ce percept doit à la base être aussi probable que le
percept en compétition. C'est le cas dans la rivalité binoculaire de Gabors orientés de
même contraste: l'observateur perçoit naturellement autant celui qui est orienté vers la
droite que celui qui est orienté vers la gauche. Pour biaiser la perception, il faut créer des
conditions dans lesquelles le sujet a plus intérêt à voir l'un des percepts que l'autre. Le
protocole expérimental consiste donc à imposer une tâche de bistabilité prolongée (le
sujet rapporte sa perception) en même temps qu'une tâche de recherche de cible. La cible
est en rivalité et n'est présente que pour un œil. Elle est cachée parmi des distracteurs eux-
mêmes en rivalité (selon l'orientation) et le sujet doit la localiser. L'observateur ne peut

18
donc la trouver que si le percept conscient est celui qui contient la cible. Il a dans ce cas
tout intérêt à voir plus souvent le côté qui contient la cible.
Par défaut, l’observateur n’a aucune raison de favoriser un côté plutôt que l’autre. Par
contre, nous pouvons introduire un biais vers l’œil droit dans la répartition de la cible,
c'est-à-dire que la cible est plus fréquente dans l'œil droit que dans l'œil gauche. A l’insu de
l’observateur, le système visuel peut alors détecter ce biais par apprentissage statistique, et
en tenir compte pour modifier le percept conscient.
Comme il était possible que l'effet ait lieu à différents niveaux de la hiérarchie visuelle, un
biais de la répartition de la cible vers les percepts orientés vers la droite a aussi été ajouté.
Dans ce cas, l’observateur a aussi tout intérêt à percevoir plus longtemps le côté
présentant les orientations droites pour y trouver la cible. Deux expériences séparées
auraient pu être conduites pour cela, mais un simple remaniement des répartitions a
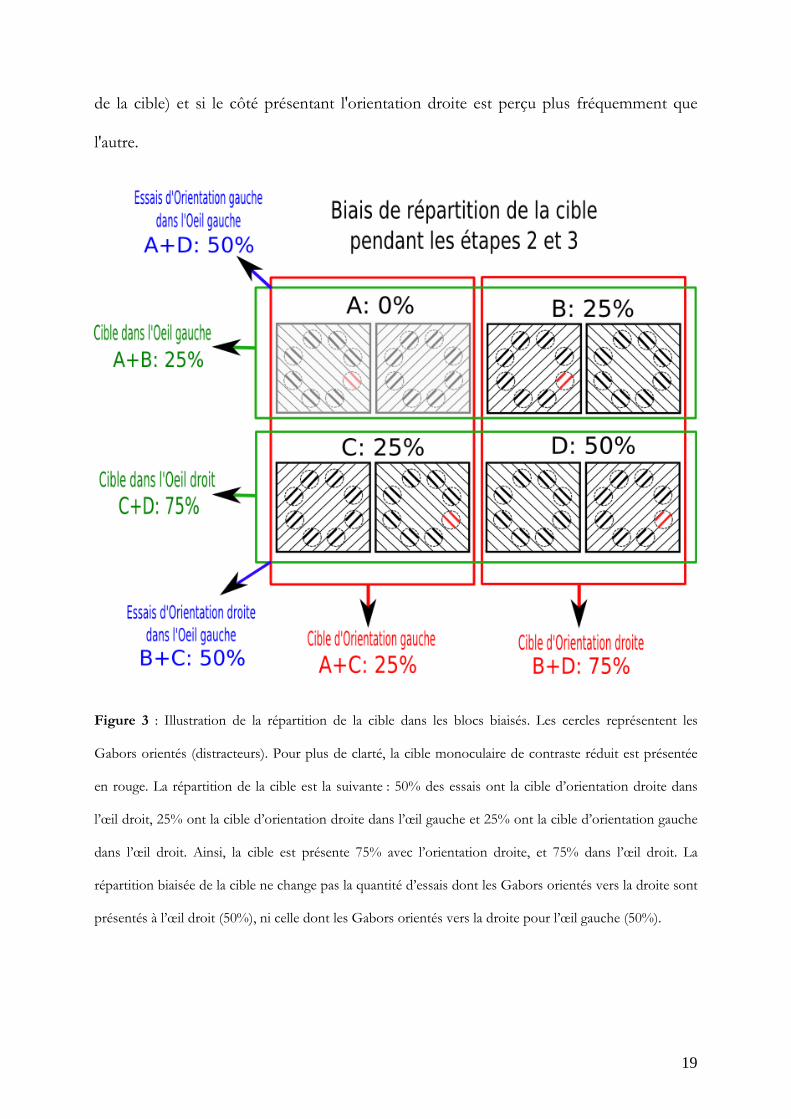
permis de tester les deux hypothèses en une seule expérience (Figure 3). Ce remaniement
consiste à présenter la cible 75% des essais dans l'œil droit, et 75% du côté qui contient
les orientations droites. Pour résumer la manipulation, prenons quatre essais. Deux essais
auront la cible présentée dans l'œil droit avec l'orientation droite, un essai contiendra la
cible dans l'œil droit mais avec l'orientation gauche et le dernier essai aura la cible dans
l'œil gauche avec l'orientation droite. De cette manière, trois essais sur quatre ont la cible
dans l'œil droit et trois essais sur quatre ont la cible avec l'orientation droite. Par contre,
pour la moitié des essais, les Gabors sont orientés à droite dans l'œil droit et pour l'autre
moitié, ils sont orientés à gauche dans l'œil droit. Cela garantit l'indépendance entre œil et
orientation. Cette indépendance nous permet d’analyser séparément si le stimulus de l'œil
droit est vu plus fréquemment que celui de l'œil gauche (indépendamment de la position
19
de la cible) et si le côté présentant l'orientation droite est perçu plus fréquemment que
l'autre.
Figure 3 : Illustration de la répartition de la cible dans les blocs biaisés. Les cercles représentent les
Gabors orientés (distracteurs). Pour plus de clarté, la cible monoculaire de contraste réduit est présentée
en rouge. La répartition de la cible est la suivante : 50% des essais ont la cible d’orientation droite dans
l’œil droit, 25% ont la cible d’orientation droite dans l’œil gauche et 25% ont la cible d’orientation gauche
dans l’œil droit. Ainsi, la cible est présente 75% avec l’orientation droite, et 75% dans l’œil droit. La
répartition biaisée de la cible ne change pas la quantité d’essais dont les Gabors orientés vers la droite sont
présentés à l’œil droit (50%), ni celle dont les Gabors orientés vers la droite pour l’œil gauche (50%).

20
Nous testons donc l'hypothèse que la rivalité binoculaire peut être influencée par un
processus qui ne dépend ni de la force du stimulus, ni de l'attention endogène ou exogène,
mais plutôt du gain attendu dans une tâche simultanée.

21
Méthode
Participants
Les participants étaient 14 étudiants consentants, en master ou thèse, d'acuité visuelle
normale ou corrigée. Cinq autres participants ont été exclus par échec de la procédure de
calibrage (dominance oculaire trop forte, ou trop forte préférence pour le vert ou le
rouge: la procédure est décrite plus bas).
Matériel
Les stimuli étaient présentés à l'aide d'un stéréoscope de Wheatstone modifié. Cet appareil
est formé de 4 miroirs sur un banc optique. Placés parallèlement deux à deux, les miroirs
permettent de présenter la moitié droite de l'écran à l'œil droit et la moitié gauche à l'œil
gauche. Les informations de convergence et d'accommodation pouvant être manipulées
indépendamment les unes des autres, ces dernières ont été égalisées. Les stimuli
apparaissaient à une distance de 89 cm à l'aide d'un programme écrit en Matlab et utilisant
les fonctions de la PsychToolBox (Brainard, 1997; Pelli, 1997). Celui-ci opérait sur un
Apple Mac G3 connecté à un écran CRT de 21 pouces (300 x 400 mm) à une résolution
de 1280 x 960 pixels et un taux de rafraîchissement de 75 Hz. L'installation était
complétée par un repose-front/menton.

22
Stimuli
Les stimuli étaient des réseaux sinusoïdaux de fréquence spatiale 3 cycles par degré d'angle
visuel, orientés de manière perpendiculaire dans chaque œil. Huit enveloppes gaussiennes
par image (de 0.2 degré d'angle visuel à demi-hauteur) ont été appliquées, disposées en
cercle (d'un rayon de 1.32 degré d'angle visuel), créant ainsi seize Gabors. Les enveloppes
gaussiennes ont été ajustées de manière à ce que le réseau sinusoïdal soit toujours visible,
même au centre du cercle. Ainsi, le contraste de Michelson au centre d'un Gabor était de
50% (la luminance était calibrée pour chaque participant) et il restait à 5% au centre du
cercle. Cela permettait de diminuer le morcellement pendant la bistabilité. Pour la même
raison, le fond de l'image présentée dans un œil était rouge et l'autre vert (Figure 4).
Ces précautions étaient importantes car les stimuli mesuraient plus de 1 degré d'angle
visuel, taille à partir de laquelle le morcellement apparaît. Les propriétés des stimuli ont
aussi été choisies de manière à maximiser le nombre de renversements (Andrews &
Purves, 1997). Une cible était un Gabor (donc monoculaire) de contraste réduit. Des
carrés noir et blanc non-bistables formaient des cadres autour des stimuli pour aider la
fusion.

23
Figure 4 : Stimuli utilisés pendant l'expérience. L'un est présenté à l'œil gauche et l'autre à l'œil droit. Les
cadres de carrés non-bistables facilitent la fusion. Le réseau reste visible même au centre. Le contraste a
été augmenté pour plus de clarté. La cible de contraste réduit a été entourée et fléchée pour la figure.
Procédure
L'expérience consiste à introduire un protocole original de recherche visuelle sur des
cibles et distracteurs en rivalité binoculaire, tandis que les répartitions de la cible sont
modifiées. Nous observons alors comment la dynamique de la rivalité est influencée de
manière implicite.
Calibrage
Pour la suite de l'expérience, il était important d'annuler toute dominance oculaire (Handa
et al., 2004; Mapp, Ono, & Barbeito, 2003) ainsi que toute préférence rouge/vert,

24
autrement dit, que le rouge soit perçu aussi souvent que le vert en rivalité binoculaire.
Cela permettait notamment d'assurer l'indépendance entre œil et orientation tel que
discuté dans les résultats.
Le sujet commençait donc par passer un calibrage utilisant la méthode des stimuli
constants. Le stimulus de l'expérience (donc bistable) était présenté côté rouge pour l'œil
droit (1er cas) avec une luminance qui variait d'un essai sur l'autre. L'essai suivant, le côté
vert était affiché pour l'œil droit (2e cas) de manière à éviter les effets d'adaptations. La
tâche du sujet était de rapporter la première orientation perçue. Pour chacun des deux cas,
une courbe psychométrique était calculée et les valeurs de luminance amenant à une
rivalité à hauteur de 50% furent sélectionnées pour l'expérience. Nous avons choisi de
faire varier la luminance en laissant intacte le contraste parce que la tâche de recherche
visuelle portait précisément sur ce contraste. Les résultats et calculs de ce calibrage sont
disponibles en annexe 1.
Tâche
Avant un essai, le participant avait le temps de fusionner correctement grâce aux cadres
de carrés et à une croix de fixation. Placé ensuite devant les stimuli de manière prolongée,
il devait reporter sa perception quand elle changeait en appuyant sur une touche qui
indiquait l'orientation perçue des réseaux (droite ou gauche). De plus, il devait rechercher
activement une cible de contraste réduit, et une autre touche lui permettait d'annoncer
qu'il avait trouvé la cible (Figure 5). Un écran lui proposait alors de sélectionner la
position de la cible. Pour avoir des phases entières, la consigne préconisait de n'appuyer

25
Figure 5 : Illustration du déroulement d'un essai. La croix de fixation apparaît. L'essai commence en
appuyant sur une touche. La rivalité binoculaire est alors introduite pendant une durée maximale de 20
secondes. Pour plus de clarté, la cible monoculaire de contraste réduit est présentée en rouge. L'essai
s'arrête lorsque le participant appuie sur la touche CTRL. Les Gabors sont alors remplacés par des
chiffres. A l'aide des flèches, il peut déplacer le cercle sur le chiffre qui est à la position où la cible a été
localisée.

26
sur les touches qu'au moment des renversements, même pour indiquer une cible trouvée.
L'essai s'arrêtait après 20 secondes où dès qu'une réponse sur la cible était donnée. Un
feedback était affiché après chaque essai.
La répartition de la cible parmi les huit positions possibles de chaque œil était aléatoirisée
et n'est donc pas le sujet de cette étude. La couleur (rouge ou vert) du fond d'une image
était aussi aléatoirisée. Pour l’analyse, nous utiliserons des indicateurs du décours temporel
de la dynamique bistable qui sont dérivés de précédents travaux (Mamassian & Goutcher,
2005). Il est donc prévu d’analyser cette dynamique à différents temps au cours d’un essai,
les effets attendus pouvant se manifester précocement ou tardivement. Les essais ayant
une durée variable, une durée minimale englobant suffisamment d’essais sera déterminée.
Le participant était familiarisé avec la tâche avant de commencer l'expérience proprement
dite. Les participants passaient alors les quatre étapes suivantes.
Etape 1 : neutre
La première étape consistait en trois blocs identiques de 16 essais de 20 secondes
maximum (soit 48 essais). Pour la moitié des essais, les Gabors orientés étaient vers la
droite pour l'œil droit. Pour l'autre moitié, ils étaient orientés vers la droite pour l'œil
gauche.
Répartition de la cible
Au sein d'un bloc, 4 essais (25%) avaient la cible dans l'œil droit avec l'orientation droite,
4 essais avaient la cible dans l'œil droit avec l'orientation gauche (25%), 4 essais avaient la

27
cible dans l'œil gauche avec l'orientation droite et 4 essais présentaient la cible dans l'œil
gauche avec l'orientation gauche (Figure 6).
Figure 6 : Illustration de la répartition de la cible dans les blocs neutres. Les cercles représentent les
Gabors orientés (distracteurs). Pour plus de clarté, la cible monoculaire de contraste réduit est présentée
en rouge. La répartition de la cible équilibrée : 25% des essais ont la cible d’orientation droite dans l’œil
droit, 25% ont la cible d’orientation droite dans l’œil gauche, 25% ont la cible d’orientation gauche dans
l’œil droit et 25% des essais ont la cible d’orientation gauche dans l’œil gauche. Ainsi, la cible est présente
50% avec l’orientation droite, et 50% dans l’œil droit.

28
Etapes 2 et 3 : biaisées
Le participant passait ensuite trois blocs identiques de 16 essais (étape 2: 48 essais) et
encore trois blocs identiques de 16 essais (étape 3: 48 essais). La moitié des essais
présentaient les Gabors orientés vers la droite dans l'œil droit et l'autre moitié avaient les
Gabors orientés vers la droite dans l'œil gauche.
Répartition de la cible
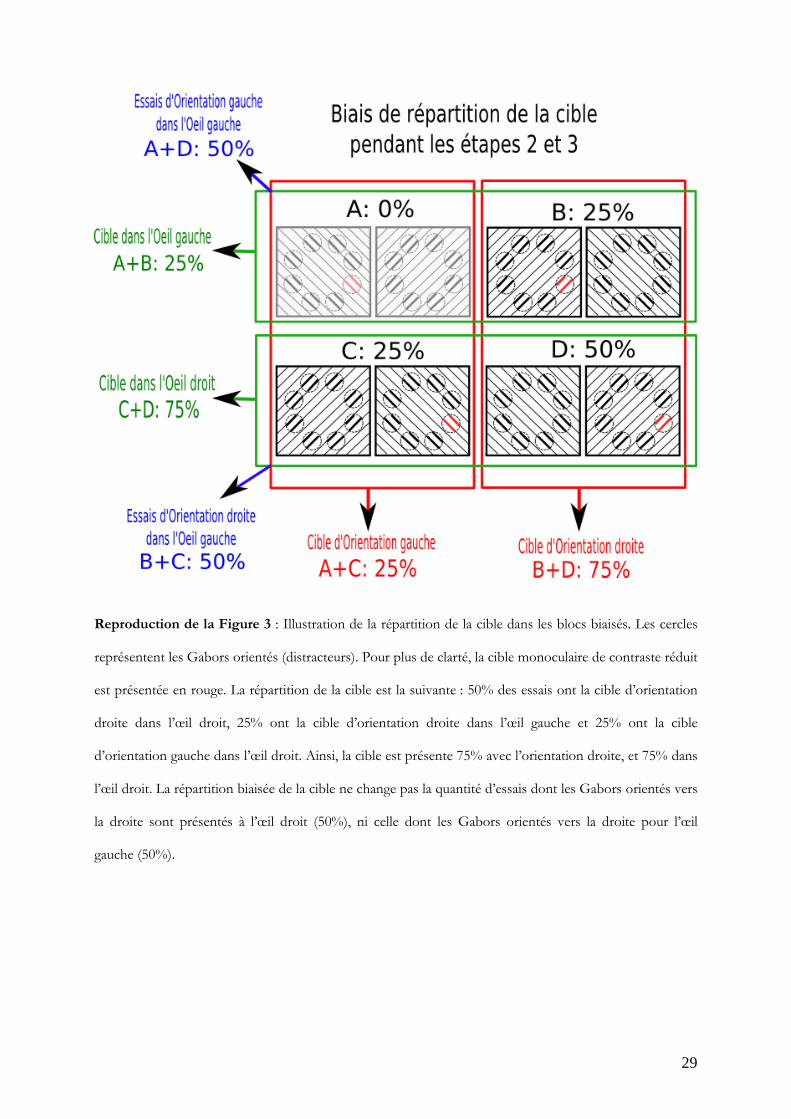
La répartition de la cible dans ces blocs était modifiée comme expliqué dans
l’introduction. Il s'agissait de biaiser arbitrairement la répartition de la cible vers l'œil droit
(présence dans 75% des essais) et vers l'orientation droite (présence dans 75% des essais).
Pour cela, dans chaque bloc, lors de 8 essais (50%), la cible était dans l'œil droit avec
l'orientation droite, 4 essais avaient la cible dans l'œil droit avec l'orientation gauche
(25%), 4 essais avaient la cible dans l'œil gauche avec l'orientation droite et aucun essai ne
présentait la cible dans l'œil gauche avec l'orientation gauche (Figure 3).
29
Reproduction de la Figure 3 : Illustration de la répartition de la cible dans les blocs biaisés. Les cercles
représentent les Gabors orientés (distracteurs). Pour plus de clarté, la cible monoculaire de contraste réduit
est présentée en rouge. La répartition de la cible est la suivante : 50% des essais ont la cible d’orientation
droite dans l’œil droit, 25% ont la cible d’orientation droite dans l’œil gauche et 25% ont la cible
d’orientation gauche dans l’œil droit. Ainsi, la cible est présente 75% avec l’orientation droite, et 75% dans
l’œil droit. La répartition biaisée de la cible ne change pas la quantité d’essais dont les Gabors orientés vers
la droite sont présentés à l’œil droit (50%), ni celle dont les Gabors orientés vers la droite pour l’œil
gauche (50%).

30
Etape 4 : neutre
Enfin, le participant repassait trois blocs non-biaisés de 16 essais (soit 48 essais). Tout
était ici identique à l'étape 1.
Lors du débriefing, on s'assurait que le participant n'avait pas détecté la répartition biaisée
de la cible. Pendant toute l'expérience, le contraste de la cible était ajusté dynamiquement
afin de maintenir la difficulté constante à 84% de réponses correctes en compensant les
effets d'apprentissage. Pour cela, une nouvelle procédure d'escalier psychométrique a été
conçue afin de converger rapidement étant donné le faible nombre d'essais. Le contraste
augmentait donc à chaque essai en cas de bonne réponse, et diminuait en cas d'erreur.
Pour converger vers un taux de réponses correctes fixe, le pas en augmentant était
différent du pas en diminuant. Cette différence était fonction de la distance entre le
pourcentage de réponses correctes atteint et la valeur à atteindre. Un faux pourcentage de
réponses correctes était présenté à la fin de chaque bloc, mimant une augmentation
plausible des performances au vue de l'apprentissage réel. Toute l'expérience avait lieu
dans l'obscurité.
Hypothèses
L’hypothèse nulle que nous souhaitons rejeter est:
- que le percept qui contient plus fréquemment la cible (œil droit et orientation droite)
n'est pas perçu plus fréquemment pendant les étapes biaisées (2 et 3) que pendant l'étape
1 (niveau de base, neutre).

31
Les hypothèses s'étendent à l'étape 4, car pour constituer un effet d'apprentissage
statistique de la répartition biaisée de la cible, celui-ci doit perdurer un certain temps après
la suppression du biais de répartition.
Le plan d’analyse est donc :
S14*E4*T(variable) avec:
-S le facteur sujet systématique
-E le facteur croisé « Etape » à 4 modalités
-T le facteur temps à nombre de modalités variable en fonction de la durée disponible des
essais.

32
Résultats
Dominance
L'analyse principale de la dynamique bistable consiste à calculer la dominance, c'est-à-dire
la proportion du temps entre t et t+1, où le participant répond voir un certain percept
moyennée sur tous les essais. Pour le percept A (par exemple, le côté de l’œil droit), la
dominance DA dans le temps sur les N essais d’une étape, s’écrit :
Le pas d’analyse (le temps entre tj et tj+1) qui a été choisi est 0.3 secondes. Les dominances
de l'orientation et de l'œil biaisés ont été calculées pour chacune des étapes. Une durée de
8.5 secondes a été sélectionnée, garantissant que suffisamment d'essais par sujet sont
disponibles pour l’analyse. La valeur N en (1) est donc le nombre d’essais restants à chaque
temps.
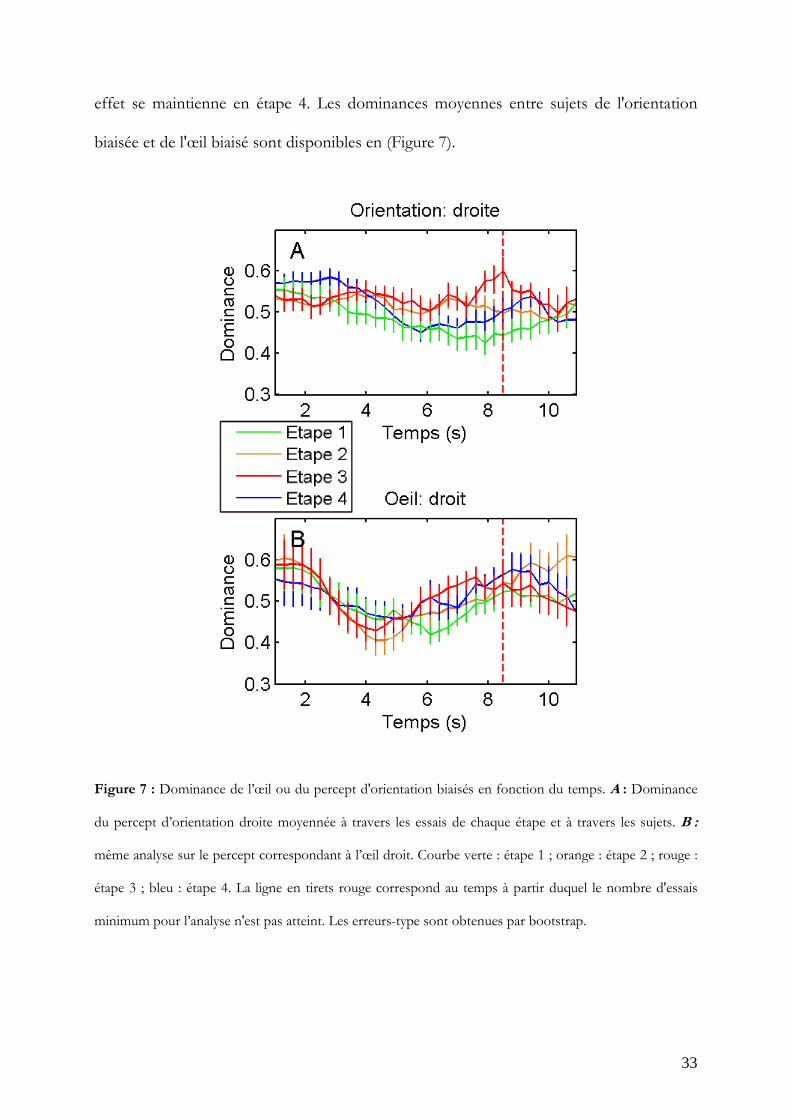
La dominance en étape 1 est utilisée comme niveau de base. Si le gain attendu a influencé
la dynamique de la bistabilité, il est prévu que les dominances des percepts biaisés en
étape 2 et 3 soient supérieures aux dominances en étape 1 et proches de 0.75, et que cet
33
effet se maintienne en étape 4. Les dominances moyennes entre sujets de l'orientation
biaisée et de l'œil biaisé sont disponibles en (Figure 7).
Figure 7 : Dominance de l’œil ou du percept d'orientation biaisés en fonction du temps. A : Dominance
du percept d’orientation droite moyennée à travers les essais de chaque étape et à travers les sujets. B :
même analyse sur le percept correspondant à l’œil droit. Courbe verte : étape 1 ; orange : étape 2 ; rouge :
étape 3 ; bleu : étape 4. La ligne en tirets rouge correspond au temps à partir duquel le nombre d'essais
minimum pour l’analyse n'est pas atteint. Les erreurs-type sont obtenues par bootstrap.

34
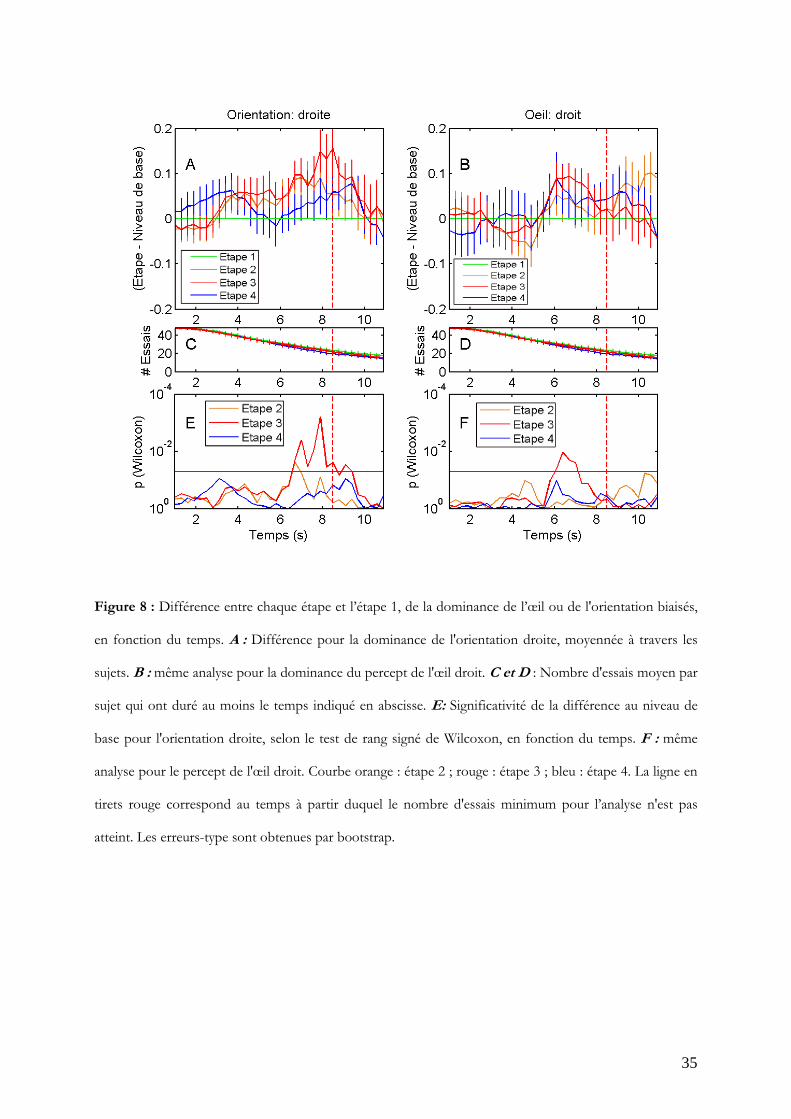
Pour plus de clarté, la différence de ces dominances par rapport à celle en étape 1 (neutre)
est présentée en figure 8, ainsi que le nombre d'essais moyen par sujet disponibles pour
l’analyse, et le degré de significativité de cette différence selon le test de rang signé de
Wilcoxon. Comme attendu grâce au calibrage, la dominance en étape 1 est très proche de
0.5. Les tracés des étapes biaisées (2 et 3) sont très similaires entre eux (Figures 8, A et B).
L'effet attendu est présent pour l'orientation biaisée dès 6.7 s en étape 2 et 3 (W = 71 et
W = 73; p<0.05) mais ne se maintient vraiment qu'en étape 3, avec un maximum de
significativité à 7.9 s (W = 99; p<0.001). Par contre, l'effet n'est pas mis en évidence en
étape 4 (Figure 8 E).
La dominance de l'œil biaisé n'est significativement différente de l'étape 1 qu'en étape 3
(Figure 8 F), entre 6.1 s et 7 s (W = 65 à 79; p<0.05).
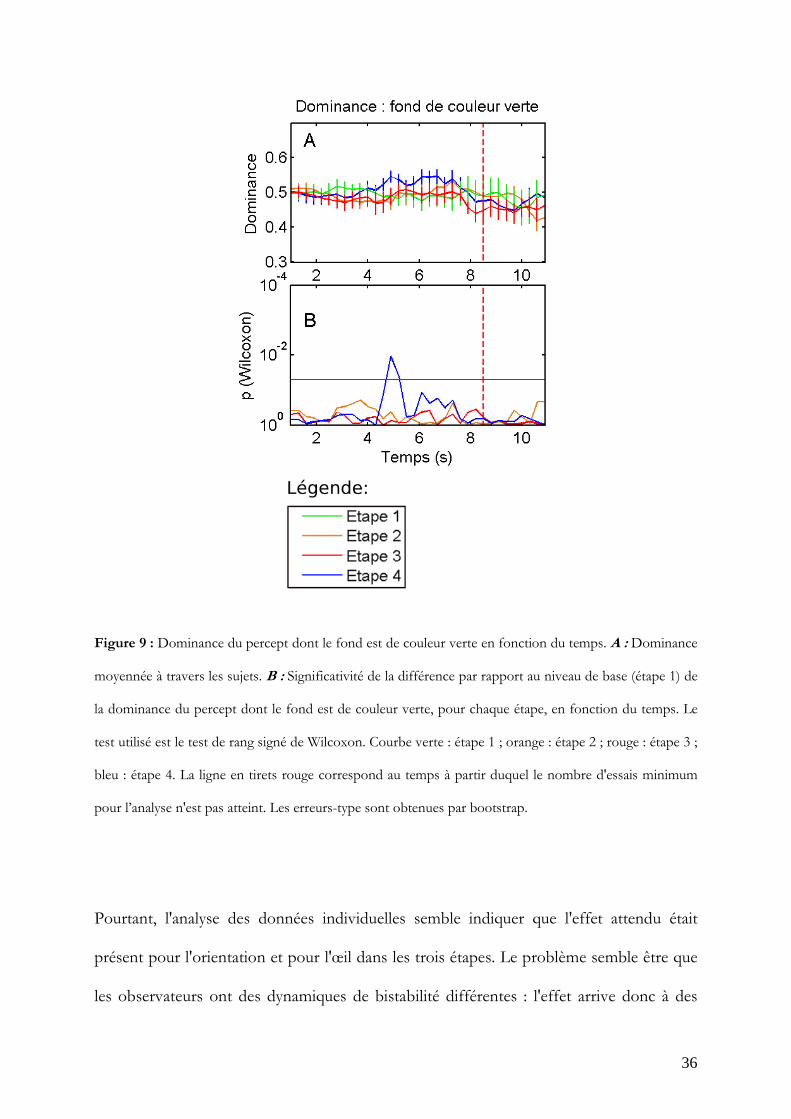
Pour comparaison, la même analyse est conduite sur la dominance du fond de couleur
verte qui était aléatoirisé pendant toute l'expérience et ne devrait donc pas présenter
d'effet (Figure 9). Aucune différence par rapport à l'étape 1 n'est significative ou
n'approche la significativité pour les étapes 2 et 3. Un pic de significativité à 4.9 s est
constaté pour l'étape 4 (W = 79 ; p<0.05).
35
Figure 8 : Différence entre chaque étape et l’étape 1, de la dominance de l’œil ou de l'orientation biaisés,
en fonction du temps. A : Différence pour la dominance de l'orientation droite, moyennée à travers les
sujets. B : même analyse pour la dominance du percept de l'œil droit. C et D : Nombre d'essais moyen par
sujet qui ont duré au moins le temps indiqué en abscisse. E: Significativité de la différence au niveau de
base pour l'orientation droite, selon le test de rang signé de Wilcoxon, en fonction du temps. F : même
analyse pour le percept de l'œil droit. Courbe orange : étape 2 ; rouge : étape 3 ; bleu : étape 4. La ligne en
tirets rouge correspond au temps à partir duquel le nombre d'essais minimum pour l’analyse n'est pas
atteint. Les erreurs-type sont obtenues par bootstrap.
36
Figure 9 : Dominance du percept dont le fond est de couleur verte en fonction du temps. A : Dominance
moyennée à travers les sujets. B : Significativité de la différence par rapport au niveau de base (étape 1) de
la dominance du percept dont le fond est de couleur verte, pour chaque étape, en fonction du temps. Le
test utilisé est le test de rang signé de Wilcoxon. Courbe verte : étape 1 ; orange : étape 2 ; rouge : étape 3 ;
bleu : étape 4. La ligne en tirets rouge correspond au temps à partir duquel le nombre d'essais minimum
pour l’analyse n'est pas atteint. Les erreurs-type sont obtenues par bootstrap.
Pourtant, l'analyse des données individuelles semble indiquer que l'effet attendu était
présent pour l'orientation et pour l'œil dans les trois étapes. Le problème semble être que
les observateurs ont des dynamiques de bistabilité différentes : l'effet arrive donc à des

37
moments différents entre sujets. En moyennant, l'effet est fortement amoindri voire
perdu. Une normalisation sur un critère objectif signant cette dynamique individuelle a été
entreprise.
Analyse avancée de la dominance
Normalisation par un nombre fixé de renversements
Le critère choisi est un nombre fixé de renversements. Pour chaque sujet, une durée est
déterminée pour laquelle il y a suffisamment d'essais pour faire l’analyse. Puis les
renversements sont cumulés sur tous les essais de l'étape 3 à chaque temps. Le temps
pour lequel le nombre arbitraire de 30 renversements cumulés est atteint, est choisi
comme la constante de normalisation et les dominances sont recalculées (ce temps
correspondra donc à une unité normalisée). Les données obtenues sont ensuite
moyennées entre sujets après avoir soustrait la dominance en étape 1. Les constantes de
normalisation utilisées sont disponibles en figure 10E. L'effet sur la dominance de
l’orientation biaisée apparaît beaucoup moins significatif suite à cette normalisation
(Figure 10, A et C): l'effet en étape 2 est significatif uniquement à 0.9 unité de temps
normalisé (W = 63 ; p<0.05) et celui en étape 3 uniquement à 1.7 unité de temps

38
Figure 10 : Différence par rapport à l’étape 1 de la dominance de l'orientation ou de l’œil biaisé en
fonction du temps après normalisation sur 30 renversements. A : Différence de la dominance de
l'orientation biaisée, moyennée à travers les sujets, pour chaque étape, après normalisation. Une unité de
temps correspond donc au moment où 30 renversements en étape 3 ont eu lieu. B : Même analyse pour la
dominance du percept de l'œil droit. C et D: Significativité de la différence pour l'orientation droite (C )
ou l’œil droit (D ) selon le test de rang signé de Wilcoxon, en fonction du temps. Courbe orange : étape 2 ;
rouge : étape 3 ; bleu : étape 4. E : Distribution des constantes de normalisation utilisées ci-dessus. Ligne
rouge verticale: moyenne des constantes de normalisation à travers les sujets. Les erreurs-type sont
obtenues par bootstrap.

39
normalisé (W = 69 ; p<0.05). L'effet de la dominance de l'œil biaisé est par contre
favorisé par cette analyse (Figure 10, B et D): il est significatif en étapes 2 et 3 à partir de
2.2 unités de temps normalisé, ce qui correspond en moyenne à 9 secondes. Le maximum
de significativité en étape 2 se trouve à 2.7 unités normalisées (soit en moyenne 11.3 s ; W
= 45; p<0.005) et à 2.5 unités normalisées en étape 3 (soit en moyenne 10.5 s ; W = 47;
p<0.05).
Toutefois, il est possible que la constante de normalisation choisie ne soit pas en rapport
avec les dynamiques des effets recherchés. Une normalisation selon un autre indicateur de
la dynamique individuelle (la durée de phase moyenne) a donc été conduite par sujet (non
présenté) mais n'a pas été concluante car les durées de phase moyennes étaient très
similaires entre participants. C'est pourquoi une nouvelle normalisation, cette fois
directement sur les maxima d’effet (maximum de [dominance en étape 3 – dominance en
étape 1]) a été calculée. Après avoir déterminé ces maxima, ils étaient utilisés pour aligner
les courbes de tous les sujets entre elles (normalisation). Cette analyse est présentée en
annexe 2 : les résultats étaient largement significatifs pour toutes les étapes, à l’exception
de l’étape 4 pour l’analyse de l’œil biaisé. Toutefois, la distribution des constantes de
normalisation utilisées pour l’orientation (maxima) apparaissait bimodale. Cela laisse
penser que les participants ont plusieurs maxima pour l'orientation, ce qui est confirmé
par les données individuelles pour l'orientation mais aussi pour l'œil. Pour analyser
correctement les dynamiques et surtout afin de pouvoir étudier la corrélation entre le
moment du maximum de la dominance de l'orientation biaisée et celui de l'œil biaisé, il est
nécessaire de bien séparer ces différents maxima, ce qui est l'objet de l’analyse suivante.

40
Elle est identique à la précédente, excepté que la constante de normalisation est le premier
maximum des effets d’orientation et d’œil biaisés, et non le maximum absolu.
Normalisation par le premier maximum d’effet
Pour chaque participant, une durée d est déterminée pour laquelle il y a suffisamment
d'essais pour faire l’analyse. Les courbes de différences de dominance entre l'étape 3 et
l'étape 1 ont été lissées en utilisant une moyenne mobile dont la fenêtre est
proportionnelle aux durées d. Les maxima sont ensuite extraits de ces données par sujet.
Les données individuelles, les lissages et les moments des maxima extraits sont
disponibles en annexe 3.
Enfin, les dominances de l'orientation et de l'œil biaisés sont normalisées en utilisant
comme constante respectivement le moment du premier maximum obtenu pour
l'orientation biaisée et celui du premier maximum obtenu pour l'œil biaisé. En
conséquence, tous les premiers maxima d'effets sont alignés et moyennés ensemble à une
unité normalisée, évitant un artefact de moyennage. Les constantes utilisées sont
présentées en figure 11 E et F: les distributions sont maintenant uni-modales de même
moyenne bien que de formes différentes. Suite à la normalisation, toutes les étapes
deviennent significativement différentes de la dominance en étape 1 (Figure 11, C et D).
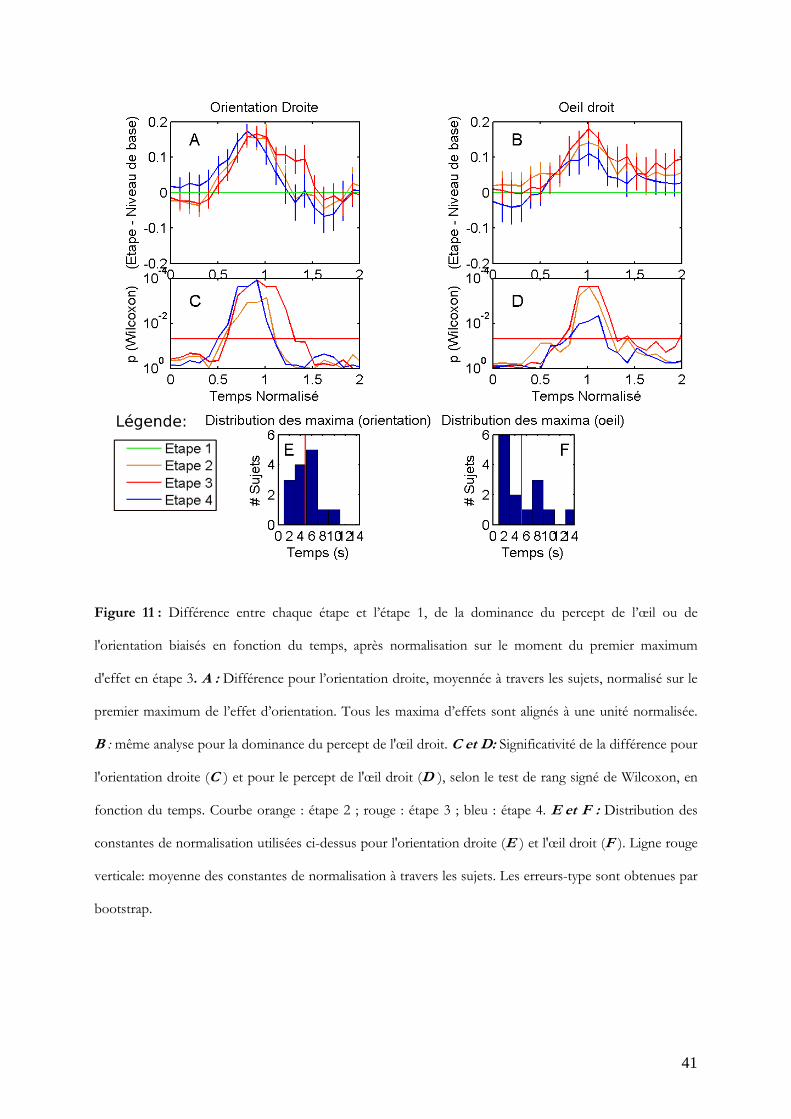
41
Figure 11 : Différence entre chaque étape et l’étape 1, de la dominance du percept de l’œil ou de
l'orientation biaisés en fonction du temps, après normalisation sur le moment du premier maximum
d'effet en étape 3. A : Différence pour l’orientation droite, moyennée à travers les sujets, normalisé sur le
premier maximum de l’effet d’orientation. Tous les maxima d’effets sont alignés à une unité normalisée.
B : même analyse pour la dominance du percept de l'œil droit. C et D: Significativité de la différence pour
l'orientation droite (C ) et pour le percept de l'œil droit (D ), selon le test de rang signé de Wilcoxon, en
fonction du temps. Courbe orange : étape 2 ; rouge : étape 3 ; bleu : étape 4. E et F : Distribution des
constantes de normalisation utilisées ci-dessus pour l'orientation droite (E ) et l'œil droit (F ). Ligne rouge
verticale: moyenne des constantes de normalisation à travers les sujets. Les erreurs-type sont obtenues par
bootstrap.

42
Les effets pour l'orientation biaisée et l'œil biaisé se maintiennent significatifs pendant 0.4
à 0.7 unités normalisées (soit de 2 à 3.5 s en moyenne, la durée d'une phase, environ)
autour d'une unité normalisée (à 5 s en moyenne). Le maximum de significativité pour
l'orientation biaisée (Figure 11, A et C) est atteint à une unité en étape 3 et 4 (W = 105;
p<0.0001). Quant à l'œil biaisé, le maximum de significativité (Figure 11, B et D) est
atteint à 1.1 unité en étape 2 et 3 (W = 99 et 103 respectivement; p<0.0005).
En comparaison, la même normalisation a été opérée sur la dominance du percept dont le
fond est de couleur verte. Le maximum de la différence de dominance entre étape 3 et
étape 1 a été extrait individuellement de la manière. Les moments de ces maxima ont
ensuite été utilisés pour la normalisation. Aucune différence significative avec l’étape 1 n’a
pu être mise en évidence sur cette analyse (annexe 4).
Relations entre effet oculaire et effet d'orientation
Les corrélations entre les moments des maxima de l'œil biaisé et de l'orientation biaisée
étaient toutes faibles (entre premiers maxima: -0.06, Figure 12; entre seconds maxima:
0.16; entre premier maximum de l'œil biaisé et second maximum de l'orientation biaisée:
0.09; entre premier maximum de l'orientation biaisée et second maximum de l'œil biaisé: -
0.31). Le même calcul de corrélation sur les maxima divisés par les durées D donnait des
résultats similaires. Une dernière analyse (non-présentée) consistait à conduire d'une part
la même normalisation, mais en utilisant les différentes combinaisons de maxima
possibles ci-dessus, et d'autre part à normaliser les données de l'orientation biaisée avec
les constantes de normalisation de l'œil biaisé et vice versa. La première partie n'a pas

43
donné de résultats qualitativement différents de ceux déjà présentés (toutes les étapes
étaient significativement différentes de l'étape 1). La seconde partie n'a donné aucun
résultat significativement différent de l'étape 1. Toutes ces données prises ensemble
Figure 12 : Moment du premier maximum d'effet de l'œil droit en étape 3 en fonction du moment du
premier maximum d'effet de l'orientation droite en étape 3. Chaque point correspond à un sujet. Les deux
valeurs ne sont pas corrélées.
amènent à conclure à l'indépendance entre les maxima d'effets sur l'orientation et de l'œil
biaisé.
Enfin, la stabilité intra-individuelle des premiers maxima a été étudiée. Les corrélations
entre ces maxima trouvés en étape 2 et ces maxima calculés en étape 3 et 4 étaient toutes
comprises entre 0.40 et 0.67 pour l'orientation biaisée et entre 0.72 et 0.90 pour l'œil
(Table 1). Des étendues de corrélation similaires ont été trouvées en refaisant ces calculs
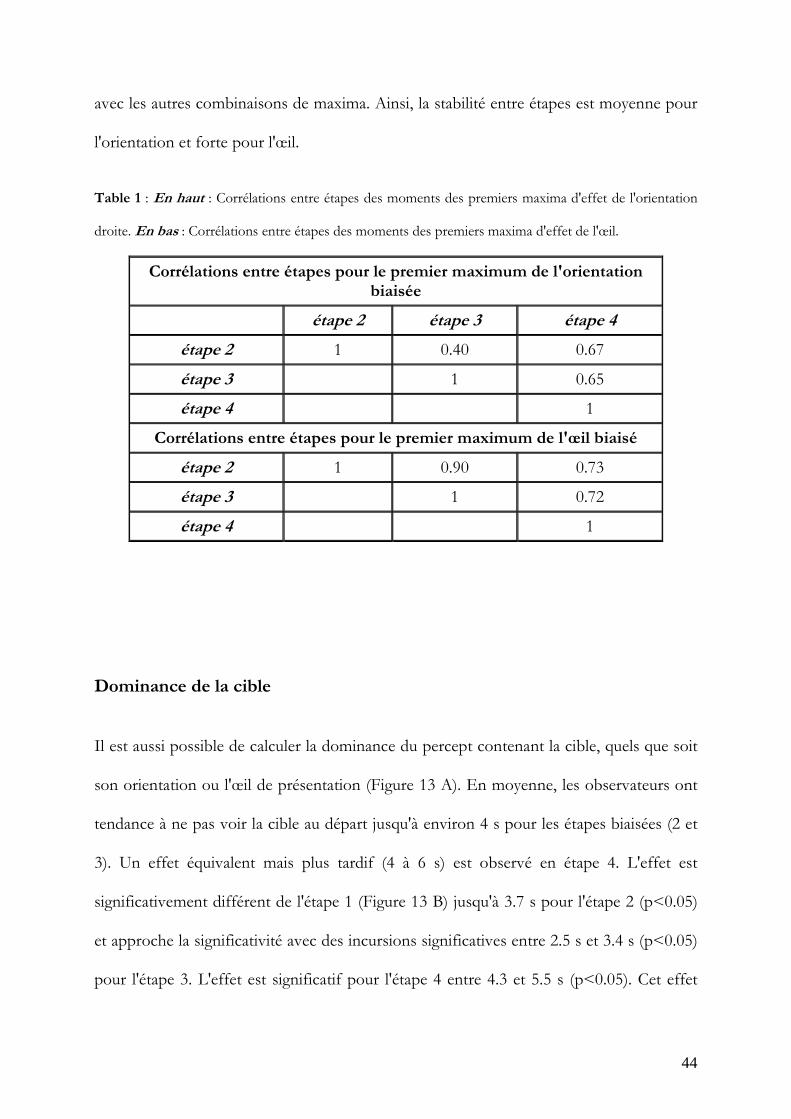
44
avec les autres combinaisons de maxima. Ainsi, la stabilité entre étapes est moyenne pour
l'orientation et forte pour l'œil.
Table 1 : En haut : Corrélations entre étapes des moments des premiers maxima d'effet de l'orientation
droite. En bas : Corrélations entre étapes des moments des premiers maxima d'effet de l'œil.
Corrélations entre étapes pour le premier maximum de l'orientation biaisée
étape 2 étape 3 étape 4
étape 2 1 0.40 0.67
étape 3 1 0.65
étape 4 1
Corrélations entre étapes pour le premier maximum de l'œil biaisé
étape 2 1 0.90 0.73
étape 3 1 0.72
étape 4 1
Dominance de la cible
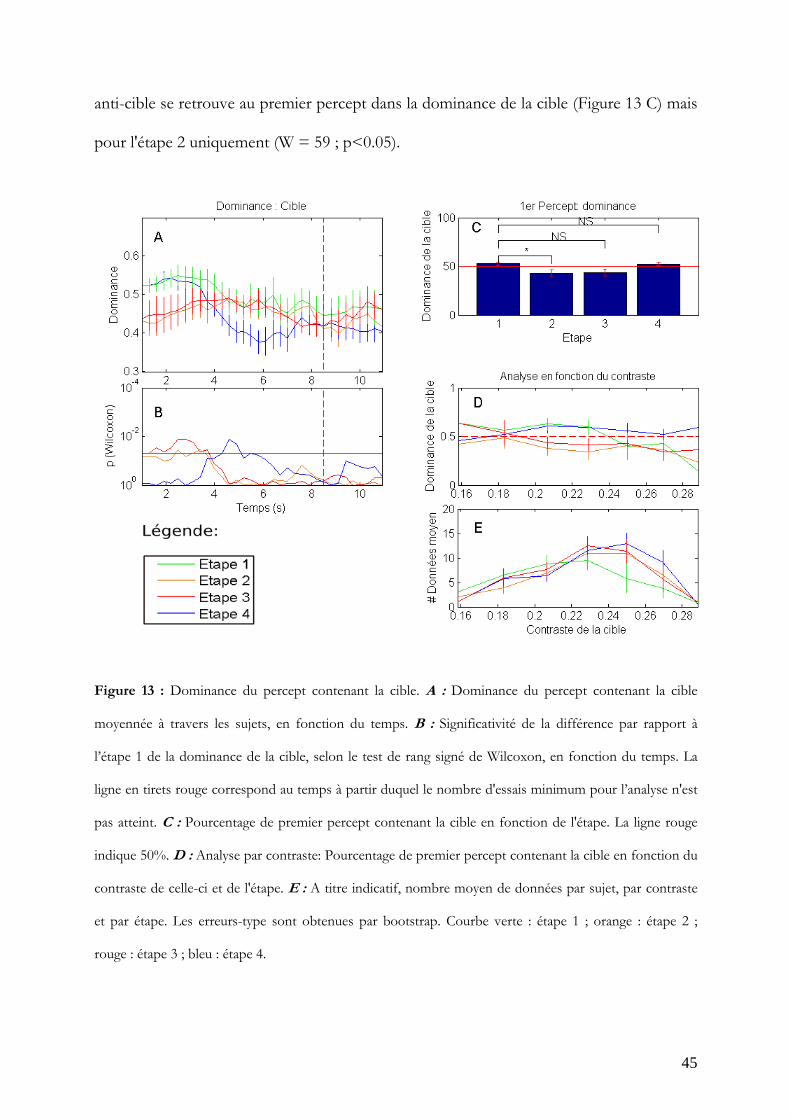
Il est aussi possible de calculer la dominance du percept contenant la cible, quels que soit
son orientation ou l'œil de présentation (Figure 13 A). En moyenne, les observateurs ont
tendance à ne pas voir la cible au départ jusqu'à environ 4 s pour les étapes biaisées (2 et
3). Un effet équivalent mais plus tardif (4 à 6 s) est observé en étape 4. L'effet est
significativement différent de l'étape 1 (Figure 13 B) jusqu'à 3.7 s pour l'étape 2 (p<0.05)
et approche la significativité avec des incursions significatives entre 2.5 s et 3.4 s (p<0.05)
pour l'étape 3. L'effet est significatif pour l'étape 4 entre 4.3 et 5.5 s (p<0.05). Cet effet
45
anti-cible se retrouve au premier percept dans la dominance de la cible (Figure 13 C) mais
pour l'étape 2 uniquement (W = 59 ; p<0.05).
Figure 13 : Dominance du percept contenant la cible. A : Dominance du percept contenant la cible
moyennée à travers les sujets, en fonction du temps. B : Significativité de la différence par rapport à
l’étape 1 de la dominance de la cible, selon le test de rang signé de Wilcoxon, en fonction du temps. La
ligne en tirets rouge correspond au temps à partir duquel le nombre d'essais minimum pour l’analyse n'est
pas atteint. C : Pourcentage de premier percept contenant la cible en fonction de l'étape. La ligne rouge
indique 50%. D : Analyse par contraste: Pourcentage de premier percept contenant la cible en fonction du
contraste de celle-ci et de l'étape. E : A titre indicatif, nombre moyen de données par sujet, par contraste
et par étape. Les erreurs-type sont obtenues par bootstrap. Courbe verte : étape 1 ; orange : étape 2 ;
rouge : étape 3 ; bleu : étape 4.

46
Cet effet non-prévu par les hypothèses s'avère assez difficile à expliquer par le protocole
expérimental. Une hypothèse est que le contraste de la cible étant réduit, la force du
stimulus s'est trouvée suffisamment réduite pour créer une aversion précoce pour la cible.
Pour le vérifier, la dominance de la cible au premier percept a été calculée en fonction des
contrastes de la cible présentés. Si le contraste est responsable de cet effet, la dominance
de la cible devrait augmenter avec le contraste de la cible. Ce n'est le cas dans aucune des
étapes (Figure 13 D), et une tendance inverse est même présente.
Durée des phases
Puisque les dominances de l'orientation et de l'œil biaisés augmentent par rapport à l'étape
neutre 1, nous avons voulu savoir si cela avait lieu à l'aide d'une augmentation des phases
du percept biaisé ou au contraire, par une diminution du percept non-biaisé comme
l'impose le second principe de Levelt (1966), voire par une combinaison des deux causes.
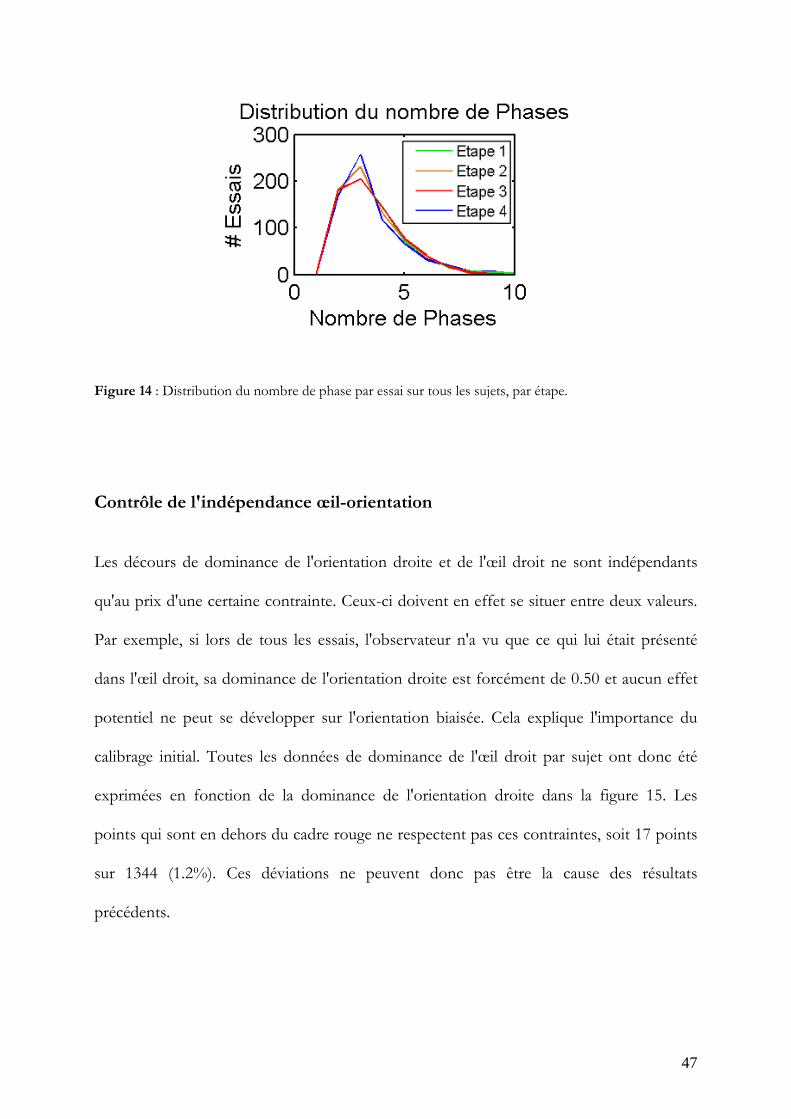
La distribution du nombre de phases par essai sur l'ensemble des sujets (Figure 14)
indiquait que la plupart des essais impliquaient de deux à quatre phases. La durée
moyenne des phases pour chaque orientation, chaque œil et chaque étape a donc été
calculée de la deuxième à la quatrième phase. Celles-ci ne diffèrent significativement de la
durée moyenne de l'étape 1 que pour le deuxième percept de l'œil gauche en étape 3 et 4
(W = 69 ; p< 0.05 et W = 71 ; p<0.05).
47
Figure 14 : Distribution du nombre de phase par essai sur tous les sujets, par étape.
Contrôle de l'indépendance œil-orientation
Les décours de dominance de l'orientation droite et de l'œil droit ne sont indépendants
qu'au prix d'une certaine contrainte. Ceux-ci doivent en effet se situer entre deux valeurs.
Par exemple, si lors de tous les essais, l'observateur n'a vu que ce qui lui était présenté
dans l'œil droit, sa dominance de l'orientation droite est forcément de 0.50 et aucun effet
potentiel ne peut se développer sur l'orientation biaisée. Cela explique l'importance du
calibrage initial. Toutes les données de dominance de l'œil droit par sujet ont donc été
exprimées en fonction de la dominance de l'orientation droite dans la figure 15. Les
points qui sont en dehors du cadre rouge ne respectent pas ces contraintes, soit 17 points
sur 1344 (1.2%). Ces déviations ne peuvent donc pas être la cause des résultats
précédents.

48
Figure 15 : Indépendance Œil-Orientation. Dominance de l'œil droit en fonction de la dominance du
percept d'orientation droite. Chaque point est la donnée d'un sujet à un temps. Carré rouge : zone à
l'intérieur de laquelle l'indépendance est respectée.
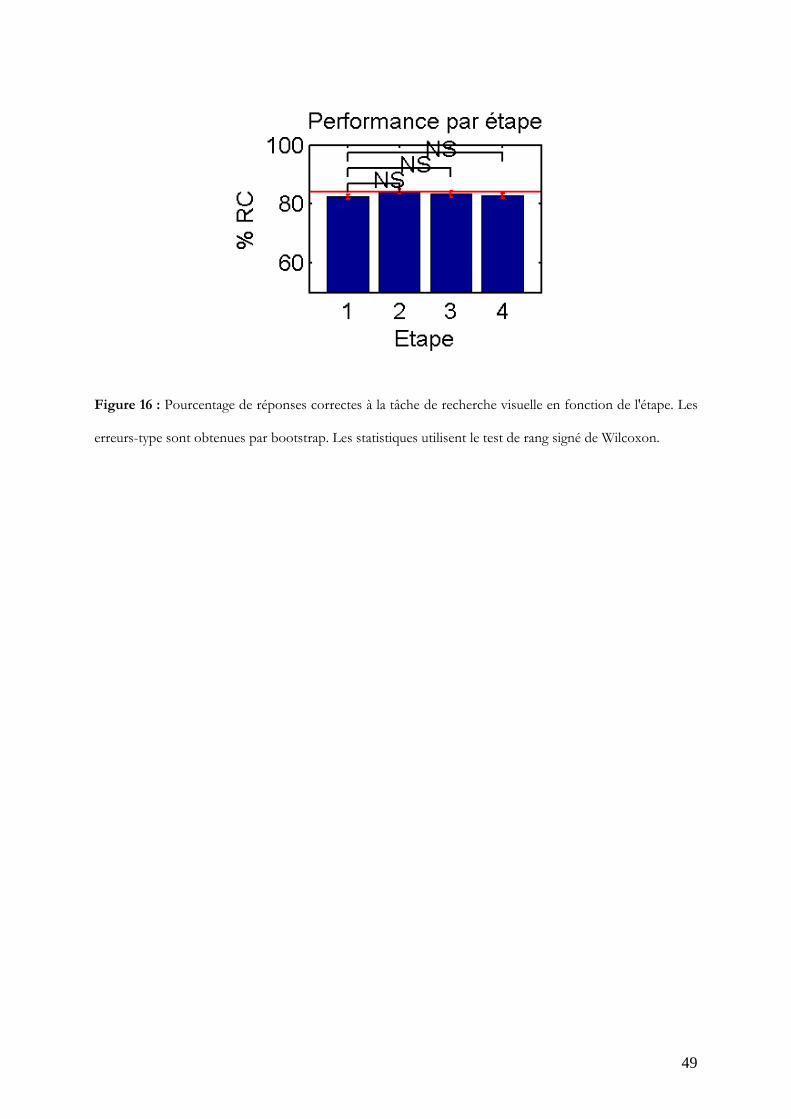
Contrôle du pourcentage de réponses correctes
Une procédure dynamique réglant le contraste de la cible avait pour but de maintenir le
taux de réponses correctes constant malgré les effets d'apprentissage. La procédure a
fonctionné correctement puisqu'aucune différence significative n'existe entre le taux en
étape 1 et les autres étapes (Figure 16) et puisque les taux sont très proches des 84% de
réponses correctes visés.
49
Figure 16 : Pourcentage de réponses correctes à la tâche de recherche visuelle en fonction de l'étape. Les
erreurs-type sont obtenues par bootstrap. Les statistiques utilisent le test de rang signé de Wilcoxon.

50
Discussion
Les trois résultats principaux de ce travail sont donc une augmentation importante de la
dominance de l'orientation et de l'œil biaisés, une indépendance entre les moments de ces
effets et un effet anti-cible précoce.
Modulation de la dominance de l’orientation et de l’œil biaisés
Le large effet sur la dominance en rivalité binoculaire est attribuable au protocole
expérimental, c'est-à-dire à l'introduction d'une répartition biaisée de la cible dans une
tâche simultanée, plus fréquente dans une orientation et dans un œil.
Pour l'orientation, cet effet est mis en évidence facilement en étape 3 malgré le
moyennage. La normalisation sur un nombre fixe de renversements diminue les effets
visibles. Les normalisations par maximum font apparaître les effets forts présents en étape
2 et 4. L'amplitude de ces effets (jusqu'à +0.23 lorsque normalisé sur les moments des
maximum : [annexe]) est alors très proche du maximum attendu si une fonction de gain
était utilisée (+0.25). Par contre, les analyses effectuées ne permettent pas de conclure que
l'effet en étape 3 est supérieur à ceux en étape 2 et 4 (notamment parce que les
normalisations ont été faites en utilisant le maximum de l'étape 3 mais aussi parce que sa
présence dans l'analyse de la dominance simple pourrait ne refléter qu'une plus forte
homogénéité des dynamiques des sujets dans cette étape).

51
Pour l'œil, l'effet est aussi facilement découvert en étape 3. Il apparaît en étape 2 après la
normalisation sur un nombre fixe de renversements. La différence de ces résultats avec
ceux de l'orientation indique que la dynamique de l'effet sur l'œil est plus en rapport avec
celle exprimée dans les renversements. Autrement dit, l'effet de l'œil arrive après un
certain nombre de renversement alors que celui de l'orientation n'est pas lié aux
renversements. L'effet apparaît ensuite aussi en étape 4, suite à la normalisation par
maximum. Là encore, l'amplitude de l'effet est celle attendue.
En comparaison, l'effet ponctuel obtenu pour le percept de couleur verte apparaît fortuit,
ce qui est confirmé par l’absence d’effet significatif après normalisation par premier
maximum.
Nous interprétons formellement les effets de l'œil et de l'orientation biaisés comme une
utilisation par le système visuel de fonction de gain dans un cadre bayésien. Celui-ci a été
utile pour la bistabilité de figures ambiguës (Mamassian & Landy, 1998). Dans le cas de la
rivalité binoculaire, il n'y a pas à l'heure actuelle de travaux montrant le passage de la force
du stimulus aux probabilités bayésiennes. Néanmoins, la distribution discrète a posteriori
serait uniforme pour les deux possibilités qu'elle contient, par exemple, l'œil droit et l'œil
gauche, puisqu’en étape 1, la dominance est équilibrée. De même, les préférences pour
l'œil et la couleur étant annulées par le calibrage, les a priori potentiels sont compensés.
Nous proposons d’étendre la règle de correspondance des probabilités aux gains attendus.
Ce genre de règle de correspondance se rapporte à un comportement général bien connu
des travaux d'éthologie. Par exemple, la fréquence et le temps de butinage dans différentes
zones chez les abeilles sont proportionnels à la quantité de gain délivrés dans ces zones
(Greggers & Menzel, 1993). Greggers et Mauelshagen (1997) ont montré que les abeilles

52
avaient besoin d'une covariation entre les attributs (couleur et odeur) de ces zones et le
gain associé pour mettre en place la règle de correspondance. Celle-ci fut d'abord
découverte par Herrnstein (1961) chez les pigeons, espèce qui est capable de bistabilité
(Vetter, Haynes, & Pfaff, 2000).
Il est important de noter l'effet probablement facilitateur dans cette expérience du
feedback à chaque essai et du score (faux pourcentage de réponses correctes) à la fin de
chaque bloc d'essais, pour motiver les participants.
Hypothèses alternatives écartées
Pour expliquer les effets obtenus, un certain nombre d'alternatives ont été étudiées.
Hypothèses portant sur la force du stimulus
Du fait du contraste plus faible de la cible, et bien que cette différence soit très faible
quand elle est divisée par la surface angulaire du stimulus complet, il est possible que la
force réduite du stimulus contenant la cible telle que définie par Levelt (1966), ait
influencé la dominance. Néanmoins, cette force réduite va contre nos hypothèses
puisqu'elle aurait eu pour effet de rendre le côté ne contenant pas la cible plus dominant.
Cette hypothèse est donc écartée.

53
Hypothèses portant sur l'effet de l'attention endogène
Ces hypothèses ont pour point commun de placer l'action volontaire du sujet comme
cause de l'effet observé. D'une part, les observateurs n'étaient pas avertis de la répartition
biaisée de la cible. A la fin de l'expérience, aucun participant n'a rapporté avoir détecté de
biais dans les répartitions vers un œil ou une orientation. Tous les participants étaient
naïfs concernant le but de l'expérience. Il était demandé aux participants de ne pas tenter
d'influencer leur bistabilité. Ainsi, même si les participants n'avaient pas respecté cette
dernière consigne, il y aurait autant de chance que leur volonté se porte sur le côté biaisé
que sur le non-biaisé. Concernant l'effet sur l'œil biaisé, la plupart des sujets est d'ailleurs
incapable de déterminer consciemment l'œil d'origine du stimulus (tâche utroculaire :
Wolfe & Franzel, 1988). Enfin, l'amplitude de l'effet est bien supérieure à ce qui est
observé dans la littérature pour le contrôle volontaire de la dominance en rivalité
binoculaire. Van Ee, van Dam et Brouwer (2005) par exemple trouve des effets d'une
ampleur de +/-0.07. Une variante de cette hypothèse est de considérer que les
participants détectent la cible mais gardent un doute. Pour lever ce doute, ils vont alors
tenter de maintenir le percept qui contient la cible jusqu'à la prise de décision. Comme la
cible est présente dans 75% des essais dans l'œil droit et dans l'orientation droite, il est
alors normal d'observer une tendance tardive de la dominance du percept biaisé à
augmenter. Si les observateurs avaient utilisé une telle stratégie, l'effet ne devrait pas se
maintenir en étape 4, qui ne présente plus de répartition biaisée de la cible, ce qui n'est pas
le cas. Ce dernier élément montre plutôt qu'un apprentissage implicite des répartitions de
la cible a eu lieu dans les étapes 2 et 3, et que celui-ci perdure en étape 4. Ce genre

54
d'apprentissage est une condition nécessaire (et une preuve en faveur) de l'utilisation des
fonctions de gain.
Hypothèse concernant l'attention exogène
Suite aux travaux de Ooi et He (1999) qui démontre un effet de l'attention exogène de
type pop-out sur la rivalité binoculaire, il est naturel d'essayer d'attribuer l'effet observé
dans cette étude au fait que le côté contenant la cible de contraste réduit pouvait
constituer un stimulus pop-out comparé à celui ne contenant que des distracteurs de
même contraste. Ici, la dynamique de l'effet observé invalide cette hypothèse. En effet,
l'effet de l'attention exogène mesuré par Ooi et He (1999) est nul à partir de 3 s. Or, celui
présenté ici n'est significatif qu'à partir de 3 s environ (sur les données normalisées sur les
maxima après lissage). L'attention exogène ne semble donc pas impliquée.
Effet précoce anti-cible
Le second résultat important de ce travail est l'effet précoce anti-cible obtenu sur la
dominance de la cible du premier percept en étape 2 et sur les dominances de la cible
entre 1 et 4 secondes pour les étapes 2 et 3 et entre 4 et 6 secondes en étape 4. L'analyse
par contrastes a écarté la possibilité que cet effet soit dû au contraste réduit de la cible.
Une autre explication possible s'inspire des travaux de Leopold, Wilke, Maier et
Logothetis (2002). Ces auteurs ont montré que lorsque le stimulus bistable était
interrompu, le dernier percept avait tendance à survivre au moment de la réapparition du
stimulus. Cet effet dépendait en fait de la durée qui séparait l'interruption du

55
renversement précédent. Pour des durées inférieures à deux secondes, le percept ne
survivait pas pour plus de 80% des essais. Dans notre expérience, les participants avaient
pour consigne de ne répondre qu'aux renversements. Or, le dernier percept avant le
dernier renversement était la cible seulement entre 26.2% et 32.1% des essais (selon les
étapes). Ainsi, juste avant la fin des essais, entre 67.9% et 73.8% des percepts contenaient
la cible. Comme les sujets venaient alors juste de renverser, ces percepts n'ont
probablement pas survécus, conduisant à l'effet anti-cible précoce. Malheureusement, si
cette explication était juste, l'effet serait présent dans les quatre étapes, ce qui n’est pas le
cas.
Hypothèse du prétraitement
La diminution de dominance dans les étapes biaisées reste donc inexpliquée. Elle permet
par contre de confirmer un élément à propos de l'effet sur la dominance de l'orientation
et de l'œil biaisés. Une hypothèse alternative à la nôtre est que le système visuel prétraite
l'information visuelle. La cible serait inconsciemment détectée (une situation comparable
à la détection en vision aveugle ; Kentridge, Heywood, & Weiskrantz, 2004), résultant en
un détournement de l'attention exogène vers le côté contenant la cible. Cela aurait pour
conséquence de rendre ce côté dominant, expliquant ainsi les deux effets (œil et
orientation). Pourtant, la cible n'est pas plus dominante dans les étapes 2 et 3 par rapport
à l'étape 1 et cet effet anti-cible est même observé. L'hypothèse de prétraitement ne peut
donc tenir. Cela signifie que ce sont les attributs biaisés (œil d'origine et orientation) qui
ont été favorisées indépendamment de la présence réelle de la cible.

56
Apprentissage statistique et hypothèse de facilitation transitoire
L'application de fonction de gain nécessite en effet que le système visuel soit capable d'un
apprentissage statistique, c'est-à-dire de former des relations à moyen ou long-terme entre
des caractéristiques (œil d'origine et orientation) et la présence d'une cible. Certains
travaux ont montré que ce type d'apprentissage implicite était possible (Chun, 2000; Jiang
& Chun, 2001; Perruchet & Pacton, 2006). Chun et Jiang (1998) par exemple, utilisent un
paradigme de recherche contextuelle. Les sujets sont capables d'apprendre implicitement
des covariations entre la position de la cible et une configuration spatiale particulière de
distracteurs, indépendamment de la forme et de la couleur de la cible. Le même type
d'apprentissage statistique peut avoir lieu sur la base de la forme des distracteurs
indépendamment de leur configuration spatiale, mais aussi sur la base du type de
déplacement (Chun & Jiang, 1999). Une association statistique entre position et présence
de la cible semble pouvoir être apprise. Hoffmann et Kunde (1999) compare les temps de
réaction pour trouver une cible A qui est plus probable dans une position comparée à une
cible B équiprobable. Les temps sont plus courts pour A autour de la position favorisée et
plus longs à distance de cette position. Geng et Behrmann (2002) obtient des résultats
similaires avec une cible répartie à 80% d'un côté, et ce, même chez des patients hémi-
négligents du côté biaisé. Mais une étude récente affirme que ces résultats sont
artefactuels (Walthew & Gilchrist, 2006) et ne reflètent qu'une facilitation transitoire de
l'essai précédent (en particulier les travaux de Hoffmann & Kunde, 1999 et Geng &
Behrmann, 2002). D’autres travaux de Geng et Behrmann (2005) souffrent partiellement
de ce biais: ils reproduisent leur résultats de 2002 avec un paradigme différent et trouvent
surtout que leur effet (qu'il constitue un apprentissage statistique ou une facilitation

57
transitoire) était additif (et donc indépendant) avec ceux de l'attention endogène et de
l'attention exogène. Ces deux dernières ont aussi été récemment définies comme
indépendantes entre elles (Berger, Henik, & Rafal, 2005). La facilitation transitoire est le
fait qu'une forme d'attention reste bloquée à l'endroit où la cible vient d'être détectée
pendant un court laps de temps (Maljkovic & Nakayama, 1996). Si une cible est présente
fréquemment à un endroit, le nombre d'essais où la cible bénéficie de la facilitation
transitoire augmente, puisqu'il y a plus d'essais contenant la cible à la même position qui
se suivent. Dans notre expérience, la position de la cible (dans le cercle de distracteurs)
n'était pas un facteur pertinent puisqu'il était aléatoire. Mais des effets de facilitation
transitoire cumulatifs sur plusieurs attributs définissant la cible ont aussi été trouvés
(Maljkovic & Nakayama, 1994, 1996). Pourtant, ce biais ne peut expliquer nos résultats
puisque la facilitation est à très court terme et qu'encore une fois, un effet anti-cible est
observé précocement. Il est donc plus probable qu'un réel apprentissage statistique ait eu
lieu, d'autant plus que la modulation de la rivalité se maintient en étape 4.
Conséquences sur la nature des compétitions sous-jacentes
Outre la nature de l'effet en lui-même, ce que montre aussi la présence d'effet sur l'œil
biaisé et l'orientation biaisée est qu'il existe au moins une compétition entre les yeux ou
entre les orientations. Qu'il existe une compétition entre représentations de plus haut
niveau que l'œil d'origine n'est pas un fait nouveau, comme nous l'avons présenté en
introduction. Si seul un effet de l'œil avait été observé, il aurait été possible de conclure
que la compétition interoculaire existait forcément, sans quoi il ne serait pas possible de la

58
biaiser. Ce qui est intéressant ici est la présence de deux effets (œil et orientation) :
l'existence de modèle hybrides comme celui de wilson03 empêche de trancher sur la
nature réelle des compétitions sous-jacentes à partir de ces simples résultats puisque les
feedbacks du niveau supérieur (compétition inter-orientation pure) vers le niveau inférieur
(compétition interoculaire orientée) peuvent rendre compte de la présence de l'effet de
l'œil biaisé (Wilson, communication personnelle). Le point clé est que dans ce cas, les
effets résultants seraient simultanés ou séparés par une durée relativement constante.
Indépendance temporelle des modulations de la rivalité
Le troisième élément important est donc que les moments des maxima des effets obtenus
sont indépendants entre l'œil et l'orientation. Les analyses entreprises n'ont en effet pas
permis de trouver une relation qui puisse permettre d'associer les deux effets. La
corrélation la plus importante entre les moments des maxima d'œil et d'orientation biaisés
est de -0.31 pour le 1er maximum de l'orientation biaisée et le 2e maximum de l'œil. Une
corrélation positive importante aurait indiqué que la durée qui sépare ces moments est
relativement fixe entre participants. Ici, une corrélation négative indique au contraire une
tendance pour les maxima de l'œil et de l'orientation à faire pivot autour d'une certaine
valeur. Cela signifie que lorsque le maximum de l'orientation est précoce, celui de l'œil a
tendance à être tardif et vice versa, mais aussi que lorsque le moment du maximum de
l'orientation se rapproche de la valeur pivot, celui de l'œil aussi, jusqu'à devenir égaux.
Tous les cas de figures sont donc envisageables et il n'est pas possible d'affirmer par
exemple que l'un des effets se produit toujours avant ou après l'autre, ou qu'ils ont lieu

59
simultanément. Les faibles corrélations observées entre moments des maxima d’effets
contrastent avec leur stabilité intra-sujet, mesurée par les fortes corrélations de ces
moments entre étapes. D'autre part, d'autres différences sont observées entre les analyses
selon l'œil biaisé ou l'orientation biaisée, comme le fait que la normalisation par nombre
fixe de renversements donne des résultats plus significatifs pour l'œil que pour
l'orientation, de même que la durée des phases, qui ne semble respecter le second principe
de Levelt (1966), que pour les phases de l'œil biaisé et non-biaisé.
Tous ces résultats nous amènent à conclure que les deux types de compétitions existent et
que la compétition interoculaire a lieu séparément de la compétition inter-orientation.
Ces données ne sont donc pas compatibles avec le modèle de Wilson (2003), les modèles
qui ne postulent qu'une compétition interoculaire comme la théorie neuronale de la
rivalité binoculaire de Blake (1989) et les modèles qui ne postulent pas de compétition
interoculaire.

60
Conclusion
Nous avons donc introduit un paradigme original pour étudier les effets d’une tâche de
recherche visuelle sur la dynamique de la rivalité binoculaire. Cette tâche consciente porte
sur la sortie de la rivalité et ajoute aux mécanismes de la perception une notion de gain.
Nous avons montré ici que ce gain était exploité par le système visuel pour choisir parmi
plusieurs interprétations celle qui devait être perçue pendant la bistabilité. Ce phénomène
est interprété comme l’utilisation d’une fonction de gain dans un cadre bayésien. La
modulation de la bistabilité et l’interprétation que nous en faisons, ne sont pas
incompatibles avec une autre version selon laquelle une forme d’attention est déployée sur
la base de régularités statistiques, rendant dominant le côté qui a le plus de chances de
contenir la cible. Nous prévoyons une extension de notre expérience, dans laquelle, si
l’hypothèse des fonctions de gain est juste, la bistabilité serait arrêtée par une répartition
de la cible biaisée entièrement vers une orientation (100% des essais), ce qui semble
impossible en cas d’effet purement attentionnel (Pastukhov & Braun, 2007).
Nous avons mis en évidence un apprentissage statistique sur un temps très court (moins
de 48 essais de 10 secondes en moyenne). Cet effet est apparu grâce à une analyse précise
du décours temporel de la dynamique bistable, plutôt que par les indicateurs moyens
traditionnels (dominance moyenne, durée de phase moyenne), démontrant l’utilité d’une
telle procédure.
Il est néanmoins peu probable que le système visuel implémente des fonctions de gain
uniquement pour appréhender des situations de rivalité binoculaire, assez rares
écologiquement. La modulation de la bistabilité mesurée dans cette étude reflète

61
vraisemblablement un mécanisme plus général qui serait actif dans la perception normale
pour guider la résolution des ambiguïtés. Tout au moins, nous prédisons que le même
effet peut être enregistré en rivalité monoculaire, dite normale (Andrews & Purves, 1997).
Enfin, nous avons apporté de nouvelles données en faveur d’une indépendance entre au
moins deux niveaux représentationnels en compétition pendant la rivalité binoculaire, qui
n’est pas rendue compte par les modèles actuels.

62
Bibliographie
Andrews, T. J. (2001). Binocular rivalry and visual awareness. Trends in Cognitive Sciences, 5(10), 407-409.
Andrews, T. J., & Purves, D. (1997). Similarities in normal and binocularly rivalrous viewing. Proceedings of the National Academy of Sciences of the United States of America, 94(18), 9905.
Berger, A., Henik, A., & Rafal, R. (2005). Competition between endogenous and exogenous orienting of visual attention. Journal of Experimental Psychology: General, 134(2), 207-221.
Blake, R. (1989). A neural theory of binocular rivalry. Psychological Review, 96(1), 145–167. Blake, R., & Logothetis, N. K. (2002). Visual competition. Nature Reviews Neuroscience, 3(1),
13-21. Block, N. (2005). Two neural correlates of consciousness. Trends in Cognitive Sciences, 9(2),
46-52. Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial Vision, 10(4), 433-436. Brascamp, J. W., van Ee, R., Noest, A. J., Jacobs, R. H., & van den Berg, A. V. (2006).
The time course of binocular rivalry reveals a fundamental role of noise. Journal of Vision, 6(11), 8.
Breese, B. B. (1899). On Inhibition. Psychological Revue (Monographs Supplements) 3, 1-65. Broadbent, D. E. (1958). Perception and Communication: Pergamon. Chun, M. M. (2000). Contextual cueing of visual attention. Trends in Cognitive Sciences, 4(5),
170-178. Chun, M. M., & Jiang, Y. (1998). Contextual Cueing: Implicit Learning and Memory of
Visual Context Guides Spatial Attention. Cognitive Psychology, 36(1), 28-71. Chun, M. M., & Jiang, Y. (1999). Top-Down Attentional Guidance Based On Implicit
Learning of Visual Covariation. Psychological Science, 10(4), 360-365. Crick, F., & Koch, C. (2003). A framework for consciousness. Nature Neuroscience, 6(2),
119-126. Diaz-Caneja, E. (1928). Sur l’alternance binoculaire. Annales D’Oculistique, 165(October),
721–731. DuTour, M. (1760). Discussion d’une question d’optique (discussion on a question of
optics). Academie des Sciences: Memoires de Mathematique et de Physique Presentes par Divers Savants, 3, 514-530.
Ernst, M. O., & Banks, M. S. (2002). Humans integrate visual and haptic information in a statistically optimal fashion. Nature, 415(6870), 429-433.
Fries, P., Roelfsema, P. R., Engel, A. K., Konig, P., & Singer, W. (1997). Synchronization of oscillatory responses in visual cortex correlates with perception in interocular rivalry. Proceedings of the National Academy of Sciences, 94(23), 12699.
Fries, P., Schroder, J. H., Roelfsema, P. R., Singer, W., & Engel, A. K. (2002). Oscillatory Neuronal Synchronization in Primary Visual Cortex as a Correlate of Stimulus Selection. Journal of Neuroscience, 22(9), 3739.
Geng, J. J., & Behrmann, M. (2002). Probability Cuing of Target Location Facilitates Visual Search Implicitly in Normal Participants and Patients with Hemispatial Neglect. Psychological Science, 13(6), 520-525.

63
Geng, J. J., & Behrmann, M. (2005). Spatial probability as an attentional cue in visual search. Perception & Psychophysics, 67(7), 1252-1268.
Greggers, U., & Mauelshagen, J. (1997). Matching behavior of honeybees in a multiple-choice situation: the differential effect of environmental stimuli on the choice process. Animal Learning Behavior, 25, 458-472.
Greggers, U., & Menzel, R. (1993). Memory dynamics and foraging strategies of honeybees. Behavioral Ecology and Sociobiology, 32(1), 17-29.
Haijiang, Q., Saunders, J. A., Stone, R. W., & Backus, B. T. (2006). Demonstration of cue recruitment: Change in visual appearance by means of Pavlovian conditioning. Proceedings of the National Academy of Sciences, 103(2), 483-488.
Handa, T., Mukuno, K., Uozato, H., Niida, T., Shoji, N., & Shimizu, K. (2004). Effects of Dominant and Nondominant Eyes in Binocular Rivalry. Optometry and Vision Science, 81(5), 377.
Herrnstein, R. J. (1961). Relative and absolute strength of response as a function of frequency of reinforcement. Journal of the Experimental Analysis of Behavior, 4(3), 267.
Hoffmann, J., & Kunde, W. (1999). Location-specific target expectancies in visual search. Journal of experimental psychology. Human perception and performance, 25(4), 1127-1141.
Hupé, J. M., & Rubin, N. (2003). The dynamics of bi-stable alternation in ambiguous motion displays: a fresh look at plaids. Vision Research, 43(5), 531-548.
Jiang, Y., & Chun, M. M. (2001). Selective attention modulates implicit learning. The Quarterly Journal of Experimental Psychology Section A, 54(4), 1105-1124.
Kahneman, D. (1973). Attention and effort: Prentice-Hall Englewood Cliffs, NJ. Kentridge, R. W., Heywood, C. A., & Weiskrantz, L. (2004). Spatial attention speeds
discrimination without awareness in blindsight. Neuropsychologia, 42(6), 831-835. Kersten, D., Mamassian, P., & Yuille, A. (2004). Object Perception as Bayesian Inference.
Annual Review of Psychology, 55, 271-304. Kording, K. P., & Wolpert, D. M. (2004). Bayesian integration in sensorimotor learning.
Nature, 427(6971), 244-247. Kovacs, I., Papathomas, T. V., Yang, M., & Feher, A. (1996). When the brain changes its
mind: Interocular grouping during binocular rivalry. Proceedings of the National Academy of Sciences, 93(26), 15508.
Leopold, D. A., & Logothetis, N. K. (1996). Activity changes in early visual cortex reflect monkeys' percepts during binocular rivalry. Nature, 379(6565), 549-553.
Leopold, D. A., & Logothetis, N. K. (1999). Multistable phenomena: changing views in perception. Trends in Cognitive Sciences, 3(7), 254-264.
Leopold, D. A., Wilke, M., Maier, A., & Logothetis, N. K. (2002). Stable perception of visually ambiguous patterns. Nature Neuroscience, 5(6), 605-609.
Levelt, W. J. M. (1966). The alternation process in binocular rivalry. British Journal of Psychology, 57(3), 225-238.
Logothetis, N. K., Leopold, D. A., & Sheinberg, D. L. (1996). What is rivalling during binocular rivalry? Nature, 380(6575), 621-624.
Lorenceau, J., & Shiffrar, M. (1992). The Influence of Terminators on Motion Integration Across Space. Vision Research, 32(2), 263-273.
Lumer, E. D., Friston, K. J., & Rees, G. (1998). Neural Correlates of Perceptual Rivalry in the Human Brain. Science, 280(5371), 1930.

64
Lumer, E. D., & Rees, G. (1999). Covariation of activity in visual and prefrontal cortex associated with subjective visual perception. Proceedings of the National Academy of Sciences, 96(4), 1669.
Maljkovic, V., & Nakayama, K. (1994). Priming of pop-out: I. Role of features. Memory and Cognition, 22(6), 657-672.
Maljkovic, V., & Nakayama, K. (1996). Priming of pop-out: II. The role of position. Perception & Psychophysics, 58(7), 977-991.
Mamassian, P. (2006). Bayesian inference of form and shape. In S. Martinez-Conde, S. L. Macknik, L. M. Martinez, J.-M. Alonso & P. U. Tse (Eds.), Progress in Brain Research (Vol. 154): Elsevier B.V.
Mamassian, P., & Goutcher, R. (2005). Temporal dynamics in bistable perception. Journal of Vision, 5(4), 7.
Mamassian, P., & Landy, M. S. (1998). Observer biases in the 3D interpretation of line drawings. Vision Research, 38(18), 2817-2832.
Mamassian, P., Landy, M. S., & Maloney, L. T. (2002). Bayesian Modelling of Visual Perception. In R. Rao, B. Olshausen & M. Lewicki (Eds.), Probabilistic Models of the Brain: Perception and Neural Function (pp. 13-36). Cambridge, MA: MIT Press.
Mapp, A. P., Ono, H., & Barbeito, R. (2003). What does the dominant eye dominate? A brief and somewhat contentious review. Perception & Psychophysics, 65(2), 310-317.
Meng, M., & Tong, F. (2004). Can attention selectively bias bistable perception? Differences between binocular rivalry and ambiguous figures. Journal of vision, 4(7), 539.
Mitchell, J. F., Stoner, G. R., & Reynolds, J. H. (2004). Object-based attention determines dominance in binocular rivalry. Nature, 429(6990), 410-413.
Moray, N. (1959). Attention in dichotic listening: Affective cues and the influence of instructions. The Quarterly Journal of Experimental Psychology, 11(1), 56-60.
Necker, L. A. (1832). Observations on some remarkable optical phaenomena seen in Switzerland; and on an optical phaenomenon which occurs on viewing a figure of a crystal or geometical solid. London and Edinburgh Philosophical Magazine and Journal of Science, 1, 329–337.
O Shea, R. P. (2003). Binocular rivalry in split-brain observers. Journal of Vision, 3(10), 610-615.
Ooi, T. L., & He, Z. J. (1999). Binocular rivalry and visual awareness: the role of attention. Perception, 28(5), 551-574.
Paffen, C. L. E., Alais, D., & Verstraten, F. A. J. (2006). Attention Speeds Binocular Rivalry. Psychological Science, 17(9), 752-756.
Pastukhov, A., & Braun, J. (2007). Perceptual reversals need no prompting by attention. Journal of Vision, 7(10), 5.
Pelli, D. G. (1997). The VideoToolbox software for visual psychophysics: transforming numbers into movies. Spatial Vision, 10(4), 437-442.
Perruchet, P., & Pacton, S. (2006). Implicit learning and statistical learning: one phenomenon, two approaches. Trends in Cognitive Sciences, 10(5), 233-238.
Pettigrew, J. D. (2001). Searching for the Switch: Neural Bases for Perceptual Rivalry Alternations. Brain and Mind, 2(1), 85-118.
Posner, M. I., Nissen, M. J., & Ogden, W. C. (1978). Attended and unattended processing modes: The role of set for spatial location. Modes of perceiving and processing information, 137-157.

65
Srinivasan, R., Russell, D. P., Edelman, G. M., & Tononi, G. (1999). Increased Synchronization of Neuromagnetic Responses during Conscious Perception. Journal of Neuroscience, 19(13), 5435.
Sundareswara, R., & Schrater, P. R. (2008). Perceptual multistability predicted by search model for Bayesian decisions. Journal of Vision, 8(5), 1-19.
Tong, F., & Engel, S. A. (2001). Interocular rivalry revealed in the human cortical blind-spot representation. Nature, 411(6834), 195-199.
Treisman, A. M. (1960). Contextual cues in selective listening. The Quarterly Journal of Experimental Psychology, 12(4), 242-248.
Trommershäuser, J., Maloney, L. T., & Landy, M. S. (2003). Statistical decision theory and the selection of rapid, goal-directed movements. Journal of the Optical Society of America A, 20(7), 1419-1433.
van Ee, R. (2005). Dynamics of perceptual bi-stability for stereoscopic slant rivalry and a comparison with grating, house-face, and Necker cube rivalry. Vision Research, 45(1), 29-40.
van Ee, R., van Dam, L. C. J., & Brouwer, G. J. (2005). Voluntary control and the dynamics of perceptual bi-stability. Vision Research, 45(1), 41-55.
van Ee, R., van Dam, L. C. J., & Erkelens, C. J. (2002). Bi-stability in perceived slant when binocular disparity and monocular perspective specify different slants. Journal of Vision, 2(9), 597-607.
Vetter, G., Haynes, J. D., & Pfaff, S. (2000). Evidence for multistability in the visual perception of pigeons. Vision Research, 40(16), 2177-2186.
von Helmholtz, H. (1867/1962). Treatise on Physiological Optics. New York: Dover. (First published in 1867, Leipzig: Voss).
Walker, P. (1978). Binocular rivalry: Central or peripheral selective processes. Psychological Bulletin, 85(2), 376–389.
Wallach, H. (1935). Über visuell wahrgenommene Bewegungsrichtung. Psychological Research, 20(1), 325-380.
Walthew, C., & Gilchrist, I. D. (2006). Target location probability effects in visual search: an effect of sequential dependencies. J Exp Psychol Hum Percept Perform, 32(5), 1294-1301.
Wilson, H. R. (2003). Computational evidence for a rivalry hierarchy in vision. Proceedings of the National Academy of Sciences, USA, 100, 14499–14503.
Wittgenstein, L. (1953). Philosophical Investigations. New York: GEM Anscombe. Wolfe, J. M. (1986). Stereopsis and binocular rivalry. Psychological Review, 93(3), 269–282. Wolfe, J. M., & Franzel, S. L. (1988). Binocularity andvisual search. Perception &
Psychophysics, 44(1), 81-93.

66
Annexes
Annexe 1 : Calibrage, préférences rouge/vert et dominance oculaire
Lors du calibrage, deux PSE (luminance pour laquelle la bistabilité est équilibrée) étaient
déterminés : le PSE lorsque le fond vert est présenté à l’œil gauche (cas 1 : PSE1) et celui
lorsque le fond rouge est présenté à l’œil gauche (cas 2 : PSE2). L’unité du PSE est un
Préférences rouge/vert individuelles : Les ordonnées correspondent au facteur par lequel il faut
multiplier la luminance appliquée à l'autre côté pour obtenir une rivalité binoculaire équilibrée. Le signe de
l'ordonnée n'est à prendre en compte que pour déterminer le sens des préférences.

67
Dominances oculaires individuelles : Les ordonnées correspondent au facteur par lequel il faut
multiplier la luminance appliquée à l'autre côté pour obtenir une rivalité binoculaire équilibrée. Le signe de
l'ordonnée n'est à prendre en compte que pour déterminer le sens des préférences.
facteur de luminance à appliquer au stimulus. A partir des PSEs, la préférence rouge/vert
F et la dominance oculaire O peuvent être déterminé simplement par :
Si F >1, la préférence est en faveur du rouge. F correspond au facteur de luminance par lequel il
faut multiplier le côté vert pour obtenir 50% de premières réponses ‘vert’ dans la tâche de
bistabilité. Si O >1, la dominance est en faveur de l’œil gauche. O correspond au facteur de
luminance par lequel il faut multiplier le côté gauche pour obtenir 50% de premières réponses
‘gauche’ dans la tâche de bistabilité. Les calibrages ont donné les résultats ci-contre pour la
préférence rouge/vert (F est présenté si rouge, 1/F si vert).
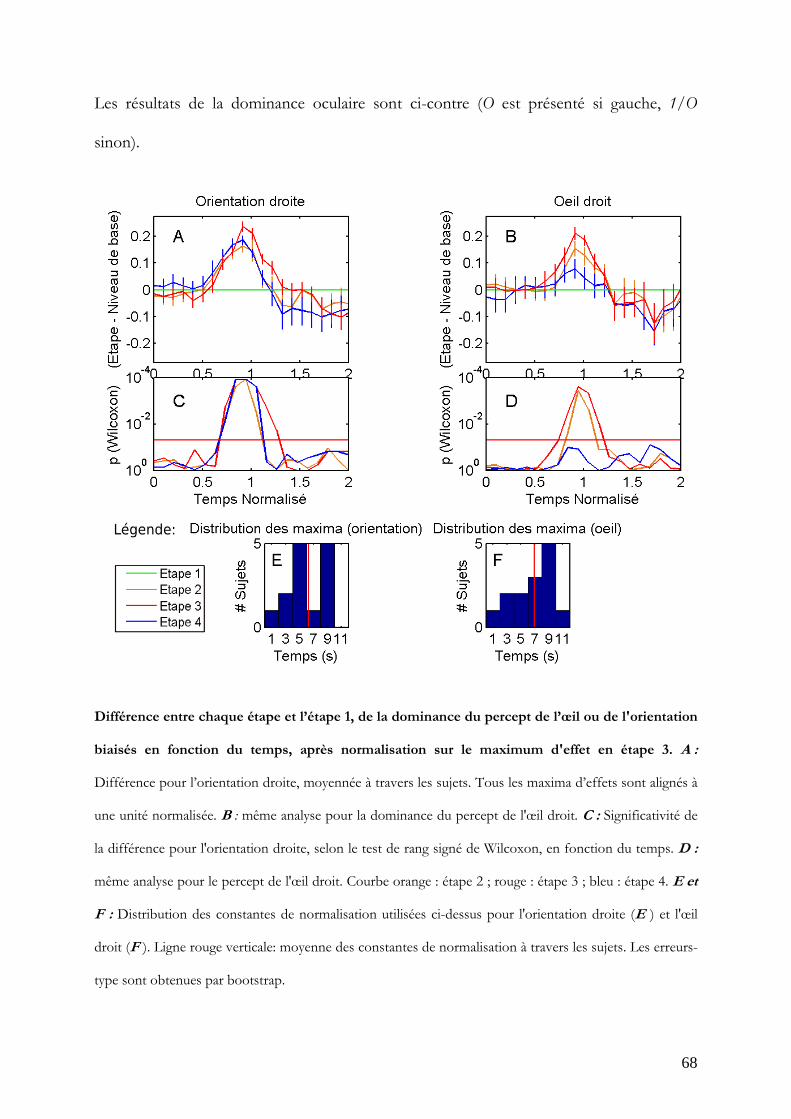
68
Les résultats de la dominance oculaire sont ci-contre (O est présenté si gauche, 1/O
sinon).
Différence entre chaque étape et l’étape 1, de la dominance du percept de l’œil ou de l'orientation
biaisés en fonction du temps, après normalisation sur le maximum d'effet en étape 3. A :
Différence pour l’orientation droite, moyennée à travers les sujets. Tous les maxima d’effets sont alignés à
une unité normalisée. B : même analyse pour la dominance du percept de l'œil droit. C : Significativité de
la différence pour l'orientation droite, selon le test de rang signé de Wilcoxon, en fonction du temps. D :
même analyse pour le percept de l'œil droit. Courbe orange : étape 2 ; rouge : étape 3 ; bleu : étape 4. E et
F : Distribution des constantes de normalisation utilisées ci-dessus pour l'orientation droite (E ) et l'œil
droit (F ). Ligne rouge verticale: moyenne des constantes de normalisation à travers les sujets. Les erreurs-
type sont obtenues par bootstrap.

69
Annexe 2 : Normalisation par le maximum d'effet
Pour chaque participant, une durée est déterminée pour laquelle il y a suffisamment
d'essais pour faire l’analyse. Puis, le maximum de la différence entre dominance dans
l'étape 3 et dominance dans l'étape 1 est extrait. Le temps correspondant à ce maximum
est utilisé comme constante de normalisation pour recalculer les dominances du sujet dans
toutes les étapes. La différence entre chaque étape et l'étape 1 est ensuite moyennée entre
sujets. De cette manière, toutes les étapes sont significatives pour l'effet sur la dominance
de l'orientation biaisée. Les étapes 2 et 4 sont significatives entre 0.8 et 1.1 unité
normalisée (avec des maxima de significativité à 1 unité: W = 105; p<0.0001), l'étape 3
entre 0.8 et 1.3 unité normalisée (avec un maximum de significativité à 1 unité: W = 105;
p<0.0001). Pour la dominance de l'œil biaisé, les étapes 2 et 3 sont significativement
différentes de l'étape 1 (respectivement entre 0.9 et 1.1 unité normalisée; W = 104,
p<0.0005; et entre 0.9 et 1.2 unité normalisée; W = 103, p<0.0005) mais pas l'étape 4.
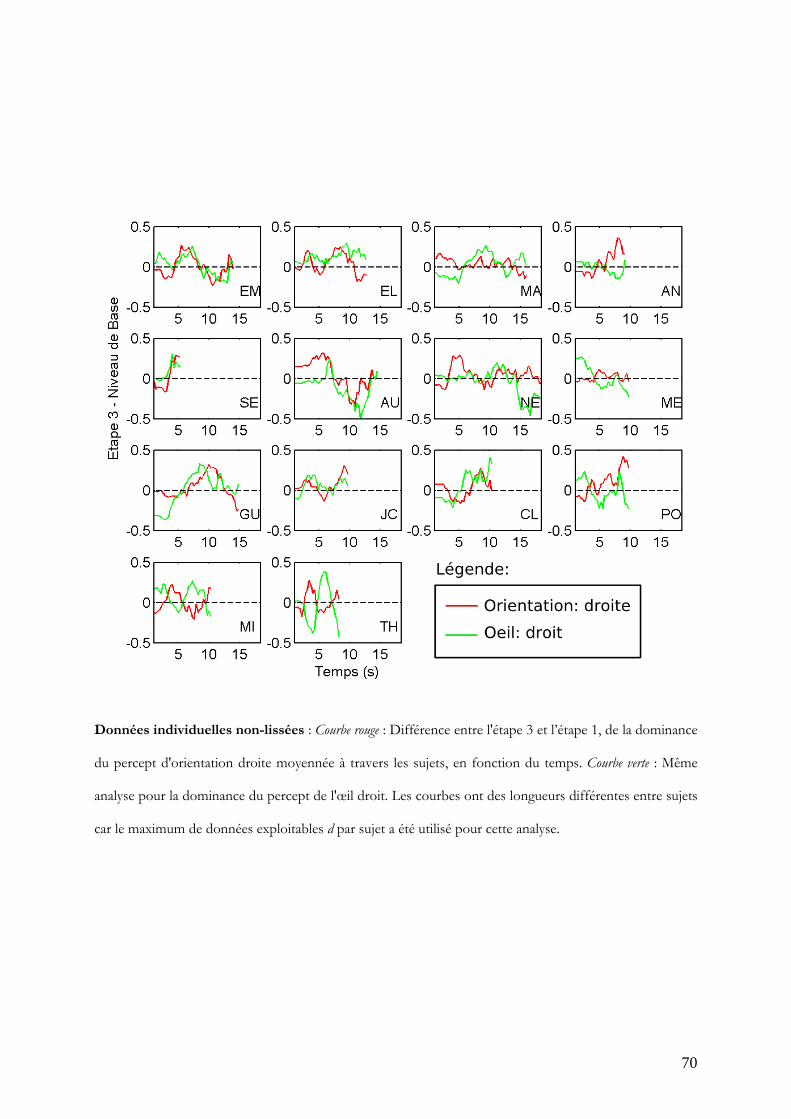
70
Données individuelles non-lissées : Courbe rouge : Différence entre l'étape 3 et l’étape 1, de la dominance
du percept d'orientation droite moyennée à travers les sujets, en fonction du temps. Courbe verte : Même
analyse pour la dominance du percept de l'œil droit. Les courbes ont des longueurs différentes entre sujets
car le maximum de données exploitables d par sujet a été utilisé pour cette analyse.
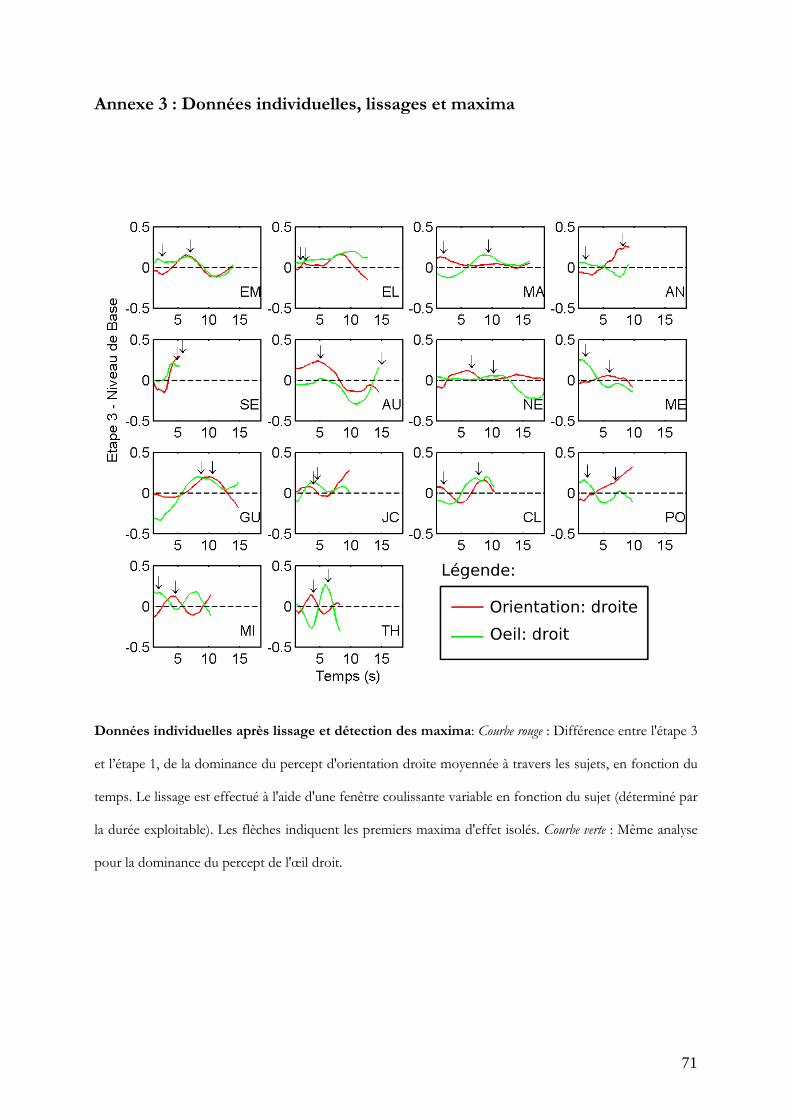
71
Annexe 3 : Données individuelles, lissages et maxima
Données individuelles après lissage et détection des maxima: Courbe rouge : Différence entre l'étape 3
et l’étape 1, de la dominance du percept d'orientation droite moyennée à travers les sujets, en fonction du
temps. Le lissage est effectué à l'aide d'une fenêtre coulissante variable en fonction du sujet (déterminé par
la durée exploitable). Les flèches indiquent les premiers maxima d'effet isolés. Courbe verte : Même analyse
pour la dominance du percept de l'œil droit.

72
Annexe 4 : Normalisation par le moment du maximum d'effet de la couleur verte
Différence entre chaque étape et l’étape 1, de la dominance du percept dont le fond est de couleur
verte en fonction du temps, après normalisation sur le maximum d'effet en étape 3 (A). Tous les
maxima d’effets sont alignés à une unité normalisée. B : Significativité de la différence selon le test de rang
signé de Wilcoxon, en fonction du temps.








