
MEMOIRE GEMINI 20 - 193.54.228.31193.54.228.31/pdf0609/curot_j_12_M2.pdf · comprendre et...
59
1 Mémoire de master 2 recherche Neuropsychologie et neurosciences cliniques _ Année 2011-2012 Jonathan CUROT Travail mené sous la direction d’Emmanuel Barbeau Directeur de recherche CNRS UMR 5549
Transcript of MEMOIRE GEMINI 20 - 193.54.228.31193.54.228.31/pdf0609/curot_j_12_M2.pdf · comprendre et...

1
Mémoire de master 2 recherche
Neuropsychologie et neurosciences cliniques
_
Année 2011-2012
Jonathan CUROT
Travail mené sous la direction d’Emmanuel Barbeau
Directeur de recherche CNRS UMR 5549

2
REMERCIEMENTS
Je tiens à remercier sincèrement toutes les personnes qui m’ont aidé et soutenu pour
l’élaboration de ce mémoire tout au long de cette année 2011-2012, et plus particulièrement :
Emmanuel Barbeau, sans qui je n’aurais pu faire ce master 2 et qui m’a ouvert les portes de
la recherche en neurosciences et donné l’envie de poursuivre dans cette voie ;
Michèle Fabre-Thorpe pour m’avoir accueilli dans son laboratoire et permis d’être associé à
ce projet ;
Les Dr. Marie Denuelle et Luc Valton, sans qui également je n’aurais pu faire ce master 2 et
poursuivre ma spécialisation en épileptologie, pour leur confiance et pour m’avoir donné
accès tout au long de cette année à leur service pour la réalisation du protocole expérimental ;
Claire Braboszcz, Damien Matéo, Roger Koenig et surtout Maxime Cauchoix et Rama
Chakravarthi pour leur aide et leurs compétences techniques, pour m’avoir permis de
comprendre et d’avancer dans le traitement du signal EEG ou dans l’utilisation de Matlab.
Une grande partie des résultats de ce mémoire n’aurait pas été possible sans eux ;
Gladys Barragan-Jason, Marlène Poncet, Thomas Busigny et Aïcha Lyoubi pour m’avoir
aidé et supporté tout long de l’année, chacun pour différentes raisons...
Les étudiants du Master 2 dont Maïa et Pauline pour leurs passages réguliers et animés
dans le bureau, et les autres thésards du CERCO ;
Et bien sûr toute l’équipe des explorations fonctionnelles du CHU Rangueil, tous mes
amis, mes co-internes, ma famille et surtout Marie.
Je tiens enfin à remercier la Fondation pour la Recherche Médicale, pour son soutien
financier tout au long de cette année.

3
SOMMAIRE
RESUME……………………………………………………………………………………………………….. 5 INTRODUCTION………………………………………………………………………………………………. 6
I- La mémoire autobiographique………………………………………………………………………. ..9
1. Définition…………………………………………………………………………………….. 9
2. Le rappel en mémoire autobiographique ………………………………………………..10
A- Le modèle constructiviste de Conway …………………………………………………....10
B- Deux modes d’accès aux souvenirs…………………………………………………….....11
C- Les corrélats neuronaux du rappel…………………………………………………………12
D- Méthodes d’évaluation de la mémoire autobiographique…………………………….....13
E- Mémoire autobiographique et épilepsie……………………………………………………14
II- Le cortex cingulaire postérieur………………………………………………………………………..15 III- Electrophysiologie de la mémoire……………………………………………………………………19
1. Les états cérébraux…………………………………………………………………………19
2. Oscillations cérébrales et mémoire autobiographique……………………………….....19
PROBLEMATIQUE ET HYPOTHESES DE TRAVAIL…………………………………………………….24 METHODES…………………………………………………………………………………………………….25
I- Population…………………………………………………………………………………………25
II- Paradigme expérimental………………………………………………………………………...26
III- Electrophysiologie……………………………………………………………………………......28
1. Stéréo-électroencéphalographie et bilan préchirurgical d’épilepsie……………………28
2. Localisation des électrodes…………………………………………………………………29
3. Traitement du signal………………………………………………………………………...30
a- enregistrement des données et pré-processing………………………………………31

4
b- analyse spectrale et décomposition temps-fréquence……………………………....33
RESULTATS……………………………………………………………………………………………………34
I. Caractéristiques des patients et résultats comportementaux……………………………………...34
II. reproductibilité des résultats entre les patients au sein du CCP…………………………………..36
III. Dynamique du signal au moment de la bascule attentionnelle au sein du CCP…………………38
IV. Dynamique du signal avant la bascule attentionnelle et pendant le rappel………………………39
V. Spécificité de ce pattern électrophysiologique au CCP gauche……………………………………42
DISCUSSION………………………………………………………………………………………………… 44
I. Des résultats reproductibles…………………………………………………………………………44
II. Activité électrophysiologique pendant le rappel…………………………………………………...45
III. Activité électrophysiologique pendant la bascule attentionnelle…………………………………46
IV. Un pattern d’activité électrophysiologique spécifique au CCP…………………………………..49
V. Une activation gamma mais sans activité thêta au sein de la BA 23 gauche………………….49
VI. Des résultats compatibles avec une hétérogénéité fonctionnelle du CCP ?............................50
VII. Une ligne de base valide ?.......................................................................................................52
VIII. Limites de l’étude……………………………………………………………………………………...52
CONCLUSION………………………………………………………………………………………………….54 REFERENCES BIBLIOGRAPHIQUES…………………………………………………………………..….55

5
RESUME
Nous nous intéressons à la mémoire autobiographique épisodique, et plus précisément à l’un
ses phénomènes: celui qui concerne les mécanismes de bascule entre l’attention portée vers le monde
extérieur et l’attention portée vers notre monde interne (ou mental). L’idée générale est que pour
rappeler des informations épisodiques, il faut créer un espace mental temporaire vers lequel doit porter
notre attention. Or l’attention étant une ressource limitée, elle ne peut porter à la fois sur l’extérieur et
l’intérieur, d’autant plus que les processus de rappel d’un souvenir requièrent beaucoup de ressources.
Ainsi, un phénomène de bascule est nécessaire.
Le cortex cingulaire postérieur (CCP) est impliqué dans le rappel autobiographique mais
également dans toutes les tâches d’introspection puisqu’il s’agit d’un nœud central du réseau par
défaut. Mais il possède une grande hétérogénéité fonctionnelle avec plusieurs sous-régions et sa
fonction reste imprécise. Des études récentes ont montré que par des connexions avec les réseaux
fronto-pariétaux, certaines de ces sous-régions pourraient contrôler une balance entre des activités
cognitives dirigées vers l’environnement ou vers le monde mental. Ce rôle de « régulateur cognitif »
en fait probablement une région clé pour la bascule attentionnelle nécessaire au rappel en mémoire
autobiographique.
Nous avons étudié la puissance spectrale du signal EEG intracrânien chez des sujets
épileptiques, au sein du CCP lors du rappel d’épisodes en mémoire autobiographique. Nous l’avons
comparé au signal EEG obtenu lors de tâches de calcul et de mémoire sémantique, avec l’hypothèse
que la dynamique des activités au sein du CCP devait être évocatrice d’une bascule entre états
attentionnels différents, spécifique à la mémoire autobiographique.
Nous avons pu exploiter les données de 4 sujets. La méthode apparait reproductible entre les
patients et entre les tâches. Nous retrouvons une augmentation significative de puissance gamma dans
la bande 40-100 Hz uniquement le temps de la bascule question/rappel lors du rappel
autobiographique au sein du CCP gauche (aire de Brodmann 23). Cette activité gamma est absente
lors du calcul et de la tâche sémantique. Nous montrons un rôle du CCP dans la bascule attentionnelle
nécessaire au rappel en mémoire autobiographique. Transitoire et non associé à une activation dans
d’autres bandes de fréquence, ce pic d’activité gamma n’est probablement pas à relier à un contenu
cognitif comme la manipulation de représentations mnésiques. Il pourrait correspondre à une forme de
signal d’engagement dans un processus, ici le rappel autobiographique, vis à vis d’autres régions
cérébrales.
Nos travaux doivent être poursuivis afin d’acquérir d’autres données, notamment dans le CCP
droit, afin d’étayer l’hypothèse que l’activité électrophysiologique diffère entre sous-régions du CCP
et d’étudier les interactions du CCP avec des aires cérébrales plus distantes.

6
INTRODUCTION
Faisons une expérience qui permettra de bien comprendre le travail qui suit. Je vais vous
poser deux questions, et vous demander de vous prêter au jeu en observant bien ce qui se
passe dans votre esprit pendant que vous tentez d’y répondre. Voici la première question :
« Quel président de la république a succédé à Georges Pompidou ? ». Voici maintenant la
seconde question : « Qui était situé à votre droite pendant le repas de Noël ? ».
La réponse à la première question fait appel à la mémoire sémantique, dont le contenu
couvre l’ensemble des connaissances générales décontextualisées que possède un individu sur
le monde. La réponse à la seconde question nécessite de faire appel à la mémoire épisodique,
un type de mémoire défini plus bas dans ce travail. Que vous ayez réussi à répondre ou non à
cette seconde question, ce qui se passe en général sont les phénomènes suivants : alors que
vous tentiez d’y répondre, le monde extérieur a soudainement quasiment cessé d’exister, vous
vous êtes concentré sur votre monde interne, vous avez voyagé mentalement dans le temps,
vous avez essayé de retrouver des indices vous permettant de vous remettre dans la situation
la plus proche possible de la situation initiale. Vous avez ainsi procédé par étapes et
éventuellement vous avez réussi à répondre. Parallèlement, vous ne l’avez sûrement pas
remarqué, mais la direction de votre regard a très probablement changé alors que vous
commenciez à effectuer la recherche du souvenir. Ce cheminement semble spécifique à la
mémoire autobiographique, vous n’avez en effet pas réalisé les mêmes étapes pour répondre à
la question sémantique.
Dans mon travail de master 2, je m’intéresse à la mémoire autobiographique
épisodique, et plus précisément au phénomène de rappel des souvenirs au sein de cette
mémoire. Le rappel d’épisodes est un processus complexe comme nous venons de l’illustrer.
L’idée générale est que pour rappeler des informations épisodiques, il faut créer un espace
mental temporaire vers lequel doit porter notre attention. Or l’attention étant une ressource
limitée, elle ne peut porter à la fois sur le monde extérieur et sur le monde intérieur (notre
monde mental), d’autant plus que la mise en place du contenu de l’espace mental requièrent
beaucoup de ressources. Ainsi, un phénomène de bascule attentionnelle est nécessaire.

7
Nous émettons l’hypothèse que cette bascule permet d’opérer un passage entre deux
états cérébraux, deux états qui pourraient être radicalement différents, voire anti-corrélés,
comme certaines études en IRMf le suggèrent (Fox et al. 2005, Fransson et al. 2005) et que
nous détaillerons plus loin. Parallèlement, les connaissances sur les corrélats neuronaux de la
recherche en mémoire autobiographique sont parcellaires. Certaines régions nécessaires au
rappel sont très étudiées comme l’hippocampe ou le cortex préfrontal. D’autres le sont
beaucoup moins, comme le cortex cingulaire postérieur (CCP).
Le CCP est une des régions les plus précocement atteintes dans la maladie
d’Alzheimer (Minoshima et al. 1997, Chételat et al. 2003, Choo et al. 2010). Il est désormais
démontré qu’il s’agit aussi de l’un des deux sous-systèmes pivots, avec le cortex préfrontal
médial (CPFM), du réseau par défaut (Default Mode Network ou DMN ; Raichle et al. 2001,
Buckner et al. 2008, Fransson et al. 2008). Le DMN est un large réseau constitué de plusieurs
sous systèmes (la formation hippocampique, le CCP, le cortex rétrosplénial, le CPFM et le
lobule pariétal inférieur ou IPL) dont l’activité augmente durant les tâches d’introspection en
lien avec le self d’un individu comme se souvenir, raisonner sur des interactions sociales et se
projeter dans le futur. Ce réseau se désactive lorsque le sujet réalise une tâche nécessitant
qu’il porte son attention vers l’environnement. Le DMN semble donc en compétition
permanente avec un réseau attentionnel dorsal ou externe (EAS pour « external attentional
system ») qui s’active lorsque l’on porte son attention vers des stimuli externes (figure 1).
Cette anti-corrélation suggère que le cerveau doit fréquemment effectuer une bascule entre
deux états correspondant à différents modes de traitement des informations, pour une
adaptation et une alternance efficace entre l’introspection et des comportements dirigés vers
l’environnement.
Figure 1 : L’activité intrinsèque
suggère que le « default mode network »
(DMN ou réseau par défaut) est anti-corrélé
avec les réseaux cérébraux impliqués dans
l’attention focalisée sur un stimulus de
l’environnement. Ici sont représentées en
bleu les structures négativement corrélées
avec le réseau par défaut et en orange les
structures positivement corrélées (Buckner et
al. 2008).

8
Svoboda et al. en 2006 confirmeront à travers une méta-analyse de 24 études
d’imagerie fonctionnelle la correspondance entre le DMN et les zones activées lors du rappel
en mémoire autobiographique (figure 4). Le CCP fait partie de ces deux systèmes. Cette
dynamique compétitive entre EAS et DMN se reflète également dans la mémoire : durant la
réalisation d’une tâche mnésique, une activité importante au sein du DMN est associée avec
un mauvais encodage (qui requiert de l’attention envers les stimuli externes), mais avec un
rappel mnésique réussi (qui requiert de l’attention vers les processus mentaux internes), tandis
que les régions du réseau attentionnel externe montrent un pattern tout à fait opposé (Kim et
al. 2010). Le CCP pourrait donc être un territoire clé pour cette « bascule attentionnelle »
du monde extérieur vers le monde mental pour rappeler un souvenir personnel.
La majorité des études sur la mémoire autobiographique et le DMN utilise de
l’imagerie fonctionnelle. Si chaque neurone et chaque synapse a un rôle dans le traitement
d’une information, c’est la synchronisation de populations de neurones qui permet une
intégration à grande échelle de tous les traitements et qui permet l’émergence d’une réponse
cohérente entre les différentes régions cérébrales (Varela et al. 2001). Pour étudier une
activité cognitive, comme la mémoire, il est donc nécessaire de se pencher sur l’analyse de la
synchronisation cérébrale et donc sur une analyse quantitative des oscillations codant le
transfert de l’information d’une population neuronale à une autre, ce que ne permet pas l’IRM
fonctionnelle. L’électrophysiologie apparaît donc être une méthode complémentaire à
l’imagerie fonctionnelle, et nécessaire par sa plus forte résolution temporelle. Des données
intracérébrales peuvent être obtenues chez des patients épileptiques implantés en cours de
bilan préchirurgical, accroissant ainsi la résolution spatiale et la qualité du signal
électroencéphalographique.
Le but de ce travail est donc une analyse de l’activité électrophysiologique
intracérébrale pendant et après la bascule afin d’identifier les différences entre les états
cérébraux correspondant aux phases de question puis de recherche d’un souvenir, pour
apporter une meilleure compréhension des mécanismes de rappel des souvenirs
autobiographiques. Mes travaux cette année se sont principalement concentrés sur le
cortex cingulaire postérieur, où aucune étude en EEG intracérébrale chez l’homme n’est
rapportée au cours d’une tâche de rappel en mémoire autobiographique. Les résultats au
sein de cette région seront intégrés avec ceux obtenus dans d’autres aires cérébrales. Il

9
s’agit d’un travail préliminaire qui s’inscrit au sein d’un projet plus global dont les objectifs
sont de mieux comprendre le rappel autobiographique et de caractériser à la fois sur un plan
comportemental et électrophysiologique cette bascule attentionnelle nécessaire au rappel.
Nous commencerons par décrire les modèles définissant la mémoire autobiographique
et son rappel, puis l’état des connaissances actuelles sur le CCP au sein du réseau de la
mémoire autobiographique. Enfin nous introduirons dans une troisième partie, les principaux
travaux d’électrophysiologie sur la mémoire.
I- LA MEMOIRE AUTOBIOGRAPHIQUE
1- DEFINITION
La mémoire autobiographique (MA) concerne les traces mnésiques du passé propre à
chaque individu, accumulées depuis son plus jeune âge. Par opposition à la mémoire
sémantique, il s’agit d’une mémoire strictement personnelle. Elle est à la base de la
construction d’un sentiment d’identité et de continuité (Conway, 2001), et donc participe à la
construction du self.
La mémoire autobiographique est dichotomique. On distingue en son sein, une
composante épisodique (concernant des souvenirs spécifiques contextualisés) et une
composante sémantique (regroupant les connaissances générales sur soi). Cette dichotomie
est décrite pour la première fois par Tulving (1988), suite à l’étude du patient amnésique KC.
KC conservait des connaissances personnelles décontextualisées comme des adresses ou des
noms de personnes de son entourage mais se trouvait dépourvu de souvenirs strictement
épisodiques. Il était incapable de se projeter dans son passé.
Mon travail porte sur la composante épisodique de cette mémoire du passé, celle qui
correspond aux événements spécifiques. Il s’agit de souvenirs personnellement vécus, situés
dans le temps et l’espace avec des caractéristiques précises :
- un souvenir épisodique fait nécessairement référence à un événement unique par définition
car il est référencé précisément dans le temps (le jour et l’heure où l’événement s’est produit)
et dans l’espace (l’endroit où l’événement a eu lieu) ;
- son évocation s’accompagne d’un état de conscience particulier permettant de revivre
mentalement les détails de l’événement, sous forme d’images mentales correspondant à la

10
« réinstantiation » la plus fidèle possible de l’expérience phénoménologique qui a eu lieu lors
de l’événement en question (Brewer, 1996) ;
- cet état de conscience est qualifié d’autonoétique (Tulving, 1985) car c’est soi-même qui
refait l’expérience d’un événement qui est arrivé à soi-même antérieurement. Par conséquent
il faut une sorte de « mind’s eye » ou d’œil interne pour percevoir la réinstantiation
lorsqu’elle a lieu ;
- le rappel de ces souvenirs s’accompagne d’un « voyage mental dans le temps », structuré le
long d’une flèche du temps interne, pendant lequel le sujet tente de se remettre dans la
situation initiale ;
- le souvenir fait référence à un épisode d’une durée brève (quelques minutes en général) par
définition, car il est précisément situé dans le temps et qu’un souvenir de durée plus longue ne
s’accompagne pas de ce phénomène de « réinstantiation » ;
- les souvenirs épisodiques sont souvent riches sur le plan émotionnel, ce qui permet
d’augmenter la vivacité des détails rapportés.
2- LE RAPPEL EN MEMOIRE AUTOBIOGRAPHIQUE
Des études sur le rappel en mémoire épisodique ont montré que le cortex auditif et le
cortex visuel étaient sélectivement activés pendant la recollection de sons et d’objets visuels
(Nyberg et al. 2000, Wheeler et al. 2000). Parallèlement, Polyn et al. (2005) dans une étude en
IRM fonctionnelle ont montré que les mêmes patterns d’activité, spécifiques d’une catégorie
(célébrité, objet, paysage), mis en jeu lors de l’encodage étaient réactivés lors du rappel. Bien
que basé sur des stimuli de laboratoire, on peut légitimement supposer que ces mêmes
processus aient cours lors du rappel d’épisodes autobiographiques.
Mais un souvenir autobiographique complet requiert un système mnésique intégratif
avec au moins un type d’imagerie mentale spécifique d’une modalité (principalement visuelle
et à des degrés variables de l’imagerie spatiale ou des autres sens), un raisonnement narratif et
des émotions. Les mécanismes de reconstruction du souvenir autobiographiques sont donc
complexes et différents modèles tentent les expliquer.
A - Le modèle constructiviste de Conway
Selon le modèle constructiviste de Conway et Pleydell-Pearce (2000), la mémoire
autobiographique fait intervenir trois types de représentations emboitées : les périodes de vie,
les évènements généraux et les événements spécifiques (figure 2). Au niveau le plus général,

11
les périodes de vie rassemblent plusieurs années de vie et sont habituellement définies par un
thème. Les évènements généraux comprennent à la fois des évènements répétés et des
évènements étendus dans le temps. Les évènements spécifiques quant à eux correspondent au
registre phénoménologique des souvenirs. Il s’agit d’épisodes précis, uniques, comportant des
attributs perceptivo-sensoriels, souvent des images visuelles (Brewer, 1996). L’accès aux
événements spécifiques se fait selon un ordre chronologique (Conway et Pleydell-Pearce,
2000) et commence souvent par une recherche d’évènements généraux (Conway, 2001) par
une sorte de stratégie « en entonnoir ».
Figure 2 : Le modèle constructiviste de Conway
(adapté par P. Piolino).
B- Deux modes d’accès aux souvenirs
Il existe deux modes d’accès aux souvenirs (Moscovitch 1989, Moscovitch et Winocur
2002) : l’accès direct et l’accès indirect. Le processus de rappel peut être direct et spontané
(associatif), impliquant un accès à des détails spécifiques évoqués de manière incontrôlée et
passive par un indice extérieur dont les attributs perceptifs et sensoriels vont conduire
directement à la réactivation du souvenir (par exemple, je passe devant un pont et tout à coup
je me souviens d’un épisode antérieur en relation avec un pont, ou encore la fameuse
madeleine de Proust). L’accès indirect (stratégique) est le plus fréquent et correspond à un
rappel génératif et intentionnel, impliquant alors la recherche active d’informations
nécessaires à la construction du souvenir. Il s’obtient par exemple par des questions orientant
le sujet vers la recherche d’un épisode précis. Il nécessite la réalisation de plusieurs étapes :
trouver un contexte, rechercher dans ce contexte et vérifier. Selon Conway, ces différentes
étapes sont gérées par un processus exécutif, le «self de travail », qui participe à la

12
reconstruction des souvenirs selon les principes de cohérence (que le souvenir garde sens avec
son self, son identité) et de correspondance (avec la réalité). Il existe donc un phénomène de
rétroaction entre le type de souvenir auquel on accède et le self (Wilson et Ross 2003).
On comprend donc que ce rappel génératif est coûteux sur le plan cognitif, long et
progressif. Il impliquerait donc une concentration importante du sujet sur ces différentes
étapes mentales et la nécessité de détacher (au moins partiellement) son attention des
stimuli de l’environnement. Une bascule attentionnelle apparaît alors nécessaire pour
permettre ce voyage mental dans le temps. C’est ce mode d’accès au souvenir qui nous
intéresse.
Le rappel de souvenirs autobiographiques épisodiques est donc une expérience très
personnelle, indissociable des notions de self, de conscience autonoétique et de temps
subjectif. Il requiert vraisemblablement une cascade de processus dont les corrélats neuronaux
sont encore mal connus.
C- Corrélats neuronaux du rappel en mémoire autobiographique
Dans une revue récente des différents résultats obtenus en imagerie fonctionnelle,
Cabeza et Saint Jacques (2007) proposent un modèle de corrélats neuronaux permettant la
récupération des souvenirs autobiographiques (figure 3).
Figure 3 : Modèle du rappel en mémoire
autobiographique selon Cabeza et Saint-Jacques
(2007) Ce modèle décrit les composants principaux du réseau de
rappel en mémoire autobiographique. 1- les processus de
recherche (search) et de contrôle impliquent le cortex
préfrontal gauche 2- les processus en lien avec le self sont
contrôlés par le cortex préfrontal médial 3- la recollection
implique l’hippocampe et le cortex rétrosplénial 4-les
émotions sont traitées dans l’amygdale 5- l’imagerie
visuelle a lieu dans le cortex occipital et dans les régions du
précunéus et du cunéus 6- les processus d’exactitude (FOR
pour feeling of rightness) dans le cortex préfrontral
ventromédial.
Selon ces auteurs, le rappel indirect d’un souvenir, en suivant un indice de récupération,
ferait d’abord intervenir des processus stratégiques de recherche qui sont médiés par le cortex
préfrontal latéral gauche (recherche et contrôle des informations récupérées dans les régions
postérieures par le cortex préfrontal ventrolatéral, et manipulation de l’information en

13
mémoire de travail par le cortex préfrontal dorsolatéral). Ils interagissent avec des processus
de référence à soi via le cortex préfrontal médial. L’interaction entre ces deux types de
processus aboutit à la récupération d’un évènement spécifique localisé dans le temps et
l’espace.
La recollection qui est médiée par l’hippocampe et par le cortex rétrosplénial serait
augmentée par des processus émotionnels qui surviendraient dans l’amygdale et une imagerie
visuelle qui prendrait naissance dans les régions occipitales du précunéus et du cunéus. Très
fréquemment ces processus de récupération peuvent ne pas atteindre la bonne « cible », c’est
à dire le bon souvenir et être erronés. Les souvenirs épisodiques impliqueraient donc des
processus d’exactitude et de cohérence de soi (FOR, feeling of rightness, Moscovitch 2002)
qui seraient dépendants du cortex préfrontal ventromédian. Bien que certaines autres régions
soient très probablement impliquées, les auteurs présentent les régions précédentes comme les
composants principaux de ce réseau de recherche et de récupération en mémoire épisodique
autobiographique.
Il est important de noter que ce modèle n’insiste pas sur le rôle du cortex cingulaire
postérieur. Seul le cortex rétrosplénial est relié à la recollection selon Cabeza et Saint Jacques.
Mais comme nous l’avons déjà cité plus haut, plusieurs études convergent vers une activation
du cortex cingulaire postérieur lors du rappel en mémoire autobiographique (Andreasen et al.
1995, Svoboda et al. 2006). La figure 4 extraite de la méta-analyse de Svoboda et collègues
affiche les régions s’activant lors du rappel autobiographique sur 24 études utilisant le PET ou
l’IRMf. Le cortex rétrosplénial mais également le CCP apparaissent activés avec un fort
niveau de convergence entre ces différentes études. Nous reviendrons plus précisément sur le
rôle du CCP dans la seconde partie de l’introduction de ce travail.
Figure 4 : Méta-analyse de tâches de
mémoire autobiographique sur 24 études en
imagerie fonctionnelle (PET et IRM) de Svoboda et
al. 2006. Sont représentées successivement de haut en bas la
face latérale, la face médiale du cerveau puis la formation
hippocampique. Les couleurs indiquent les régions s’activant
avec un niveau de convergence entre fort (rouge), moyen (vert) et
faible (bleu) entre les études. Notez que le cortex cingulaire
postérieur (et les autres nœuds du DMN) apparaissent avec un
fort niveau de convergence.
CPP=cortex cingulaire postérieur, Rsp=Cortex
rétrosplénial, vMPFC=cortex préfrontal médial ventral,
dMPFC= cortex préfrontal médial dorsal, HF= formation
hippocampique.

14
D- Méthodes d’évaluation de la mémoire autobiographique
Plusieurs outils ont été élaborés afin d’étudier la mémoire autobiographique rétrograde ;
par exemple : la méthode des mots indices (Crovitz et Schiffman, 1974), le questionnaire
autobiographique de Kopelman (1989), le test de fluence verbale autobiographique (Drischel
et al.1992), le TEMPau (Piolino, 2003), l’EAMI (Irish et al. 2008). Ces épreuves permettent
d’obtenir des indices qualitatifs et quantitatifs sur la capacité à rappeler des souvenirs
épisodiques. Elles sont essentiellement basées sur l’analyse du contenu des épisodes
rapportés. Plus le contenu est détaillé et fait référence à un épisode précis dans lequel le sujet
a été impliqué, meilleur est le score en mémoire épisodique. L’évaluation de la mémoire
épisodique repose donc en général sur ce que rapporte verbalement le sujet et ainsi sur ses
capacités narratives. Certains scores vont toutefois un peu plus loin en y incluant une analyse
phénoménologique et de l’état de conscience associé au revécu du souvenir : le TEMPau
intègre un paradigme RKG (Remember/Know/Guess) et la perspective acteur ou spectateur ;
Irish et al.(2008) dédient une partie de l’EAMI à l’évaluation des marqueurs
comportementaux de conscience autonoétique, en cotant en plus de la perspective spectateur
ou acteur, le degré de mouvement dans le souvenir, une échelle de « vivacité » du souvenir,
l’impression subjective de revivre ou non le souvenir ainsi que la connexion émotionnelle.
Cependant, aucune de ces épreuves ne renseigne sur les processus mis en œuvre lors
du rappel d’épisodes. En réalité, ces processus sont globalement peu étudiés.
E- Mémoire autobiographique et épilepsie
La mémoire autobiographique est atteinte dans de nombreuses pathologies, comme la
maladie d’Alzheimer, mais aussi dans certaines formes d’épilepsie. L’épilepsie est une des
affections neurologiques chroniques les plus fréquentes. D’après les données de
l’Organisation Mondiale de la Santé (O.M.S), elle concerne de 0,5 à 0,8% de la population.
L’épilepsie du lobe temporal est la plus fréquente des épilepsies partielles (80% des cas). Elle
est la plus importante des épilepsies focales du fait de sa haute prévalence, de l’importance
des pharmaco-résistances et généralement de par ses effets néfastes sur les fonctions
mnésiques (Engel, 1996). La zone épileptogène concerne fréquemment les structures de la
région temporale interne (l’hippocampe, le cortex entorhinal ou l’amygdale) dont on connaît
l’importance dans la mémoire épisodique.

15
S’il n’est pas rare de relever une plainte concernant les souvenirs personnels chez les
patients présentant une épilepsie du lobe temporal, ce fait clinique est aujourd’hui largement
objectivé par plusieurs études qui ont mis en évidence un déficit du rappel des évènements
épisodiques chez ces patients (Viskontas et al., 2000 ; Voltzenlogel et al., 2006; Lah et al.,
2006; Addis et al., 2007 ; Noulhiane et al., 2007; St Laurent et al., 2009). Viskontas et al.
(2000) ont ainsi rapporté un déficit de rappel des épisodes autobiographiques s’étendant
jusqu’à la petite enfance avec une préservation de la sémantique personnelle. L’étude menée
par Voltzenlogel et al. (2006) montre également que ce déficit touche toutes les périodes de
vie en l’absence de gradient temporel. En 2006, l’étude de Lah et al. rapporte un déficit de la
fluence autobiographique chez les patients atteints d’épilepsie du lobe temporal, quelque que
soit la latéralisation du foyer épileptogène. Récemment, une atteinte spécifique et isolée de la
mémoire autobiographique a été rapportée chez certains patients épileptiques dans le cadre de
l’indentification d’un nouveau tableau nosologique : le Transient Epileptic Amnesia (TEA,
Butler et al., 2007; Tramoni et al., 2011).
Il existe donc un réel enjeu à mieux connaître les processus de rappel de la mémoire
autobiographique pour en comprendre les dysfonctionnements dans ces pathologies.
II- LE CORTEX CINGULAIRE POSTERIEUR
Le gyrus cingulaire appartient au système limbique et forme une structure continue à la
partie médiale du cerveau. En 1937, Papez l’intègre au sein d’un circuit cortico-sous-cortical
gérant les émotions : le circuit hippocampo-mamillo-thalamo-cingulaire ; avant que ce dernier
soit rattaché également à la mémoire à long terme. Actuellement, le modèle prévalent
d’organisation du gyrus cingulaire est celui d’une subdivision en 4 régions aux rôles,
connexions et cyto-architectures différentes : le cortex cingulaire antérieur, le cortex
cingulaire moyen, le CCP et le CRS (Palomero-gallagher et al. 2009).
Nous l’avons vu plus haut, plusieurs études montrent une activation de la région
cingulaire postérieure lors du rappel en mémoire autobiographique. Le CCP forme avec le
cortex rétrosplénial (CRS) la partie postérieure du gyrus cingulaire (figure 5). Dans
l’organisation cérébrale, le CPP occupe une place spécifique en raison de son haut niveau de
connectivité avec les cortex frontal, pariétal et temporal et les régions sous corticales :
Buckner et al. (2008) et Hagmann et al. (2008) en font un des nœuds structurels principaux du
cerveau. Il a notamment d’importantes connexions réciproques avec le lobe temporal médian
et plus particulièrement avec les cortex entorhinaux et parahippocampiques (Morris et al.
16
1999). C’est la région du cerveau avec métabolisme le plus important (Minoshima et al. 1997,
Raichle et al. 2001). Actuellement, de plus en plus de travaux convergent vers l’idée que les
nombreuses connexions hétérogènes du CCP sont incompatibles avec une fonction cognitive
unique et donc que le CCP jouerait des rôles multiples. Pourtant ces fonctions restent
incertaines.
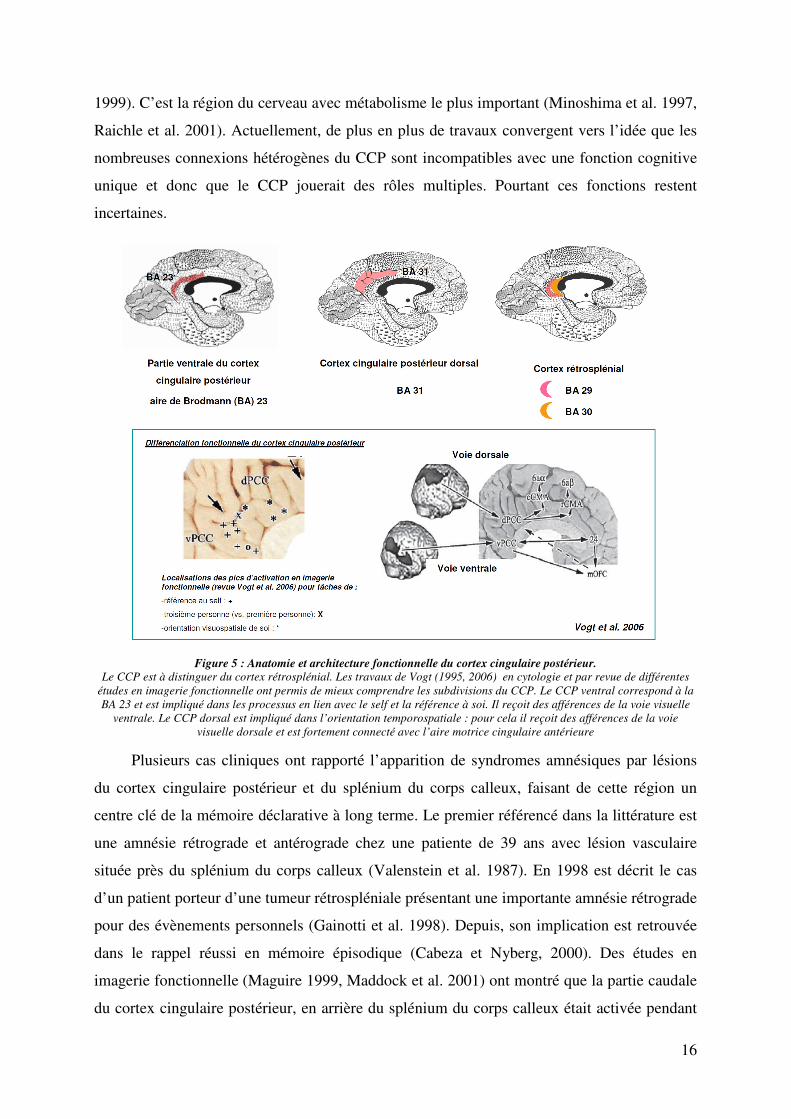
Figure 5 : Anatomie et architecture fonctionnelle du cortex cingulaire postérieur. Le CCP est à distinguer du cortex rétrosplénial. Les travaux de Vogt (1995, 2006) en cytologie et par revue de différentes
études en imagerie fonctionnelle ont permis de mieux comprendre les subdivisions du CCP. Le CCP ventral correspond à la
BA 23 et est impliqué dans les processus en lien avec le self et la référence à soi. Il reçoit des afférences de la voie visuelle
ventrale. Le CCP dorsal est impliqué dans l’orientation temporospatiale : pour cela il reçoit des afférences de la voie
visuelle dorsale et est fortement connecté avec l’aire motrice cingulaire antérieure
Plusieurs cas cliniques ont rapporté l’apparition de syndromes amnésiques par lésions
du cortex cingulaire postérieur et du splénium du corps calleux, faisant de cette région un
centre clé de la mémoire déclarative à long terme. Le premier référencé dans la littérature est
une amnésie rétrograde et antérograde chez une patiente de 39 ans avec lésion vasculaire
située près du splénium du corps calleux (Valenstein et al. 1987). En 1998 est décrit le cas
d’un patient porteur d’une tumeur rétrospléniale présentant une importante amnésie rétrograde
pour des évènements personnels (Gainotti et al. 1998). Depuis, son implication est retrouvée
dans le rappel réussi en mémoire épisodique (Cabeza et Nyberg, 2000). Des études en
imagerie fonctionnelle (Maguire 1999, Maddock et al. 2001) ont montré que la partie caudale
du cortex cingulaire postérieur, en arrière du splénium du corps calleux était activée pendant

17
la récupération réussie de souvenirs autobiographiques. De façon cohérente, le cortex
cingulaire postérieur est la première région montrant un hypométabolisme avant une atrophie
significative dans la maladie d’Alzheimer (Minoshima et al. 1997, Chételat et al. 2003, Choo
et al. 2010). Mais les points de vue sur l’implication du CCP dans la mémoire
autobiographique restent divergents. Par exemple, certains auteurs associent la recollection
avec le CCP (Skinner et Fernandes 2007, Summerfield et al. 2009) ; d’autres ne l’attribuent
qu’au CRS (Steinvorth et al. 2006, Cabeza et Saint Jacques 2007).
Parallèlement, il s’agirait de l’un des principaux nœuds du DMN (Buckner et al. 2009,
Mevel et al. 2010). La figure 6 issue de la revue de Buckner (2009) illustre l’activation du
DMN par 4 types de tâches dans 3 études: la mémoire autobiographique, se projeter dans le
futur, la théorie de l’esprit et les jugements moraux. Toutes aboutissent à une activation forte
du CCP et du CPFM. Le lien entre ces 4 tâches est l’introspection et la référence à soi. Ainsi,
si dans leur modèle Cabeza et Saint Jacques (2007) placent les processus de référence au self
dans le cortex préfrontal médial, l’implication du CCP est également désormais démontrée
dans le self (Northoff et Bermpohl, 2004 ; Svoboda et al. 2006). Andrews-Hanna et al. (2010)
insistent sur une organisation centrale du DMN autour de deux nœuds : le CCP et le CPFM
antérieur (figure 7). Ce réseau central s’active lorsque le sujet prend des décisions affectives
ou en lien avec le self. Il interagit avec deux sous-systèmes. Un sous-système est centré sur le
lobe temporal médial avec la formation hippocampique, l’IPL, le cortex parahippocampique,
le CPFM ventral et le cortex rétrosplénial. Ce dernier serait spécifiquement engagé dans la
construction de scènes mentales basées sur la mémoire. L’autre sous-système comprend le
pôle temporal, la jonction pariéto-temporale, le CPFM dorsal et le cortex temporal latéral ;
pour l’instant il n’a pas été apporté par l’auteur d’activité spécifique à ce second sous-
systèmes.
Figure 6 : Le DMN est active par diverses formes de tâches
qui requièrent la simulation mentale de scènes imaginées, de
perspectives alternatives ou de scènes passées. Ici sont
représentés quatre exemples de la littérature en imagerie
fonctionnelle (Buckner et al. 2008). (A) La mémoire autobiographique. Addis et al. (2007). (B) Se projeter
dans le future. Addis et al. (2007).(C) La théorie de l’esprit. Les sujets
devaient répondre à une tâche comme s’ils percevaient la perspective
d’une autre personne. Saxe and Kanwisher (2003). (D) Prises de
decisions morales (lors d’un dilemme). Greene et al. (2001).Toutes les
études activent fortement le CCP et le CPFM. En raison de leur
superposition, il est très probable que ces cartes représentent plusieurs
sous-sytèmes interagissant entre eux.

18
Figure 7 : La connectivité
fonctionnelle intrinsèque du DMN :
un « nœud médian » représenté par
le cortex cingulaire postérieur et le
cortex préfrontal médian antérieur ;
et deux sous systèmes (un sous-
système centré sur le lobe temporal
médial et un sous-système centré sur
le cortex préfrontal médial dorsal
(Andrews-Hanna et al. 2010).
Si l’activation de ces régions médiales que sont le CPFM et le CPP pendant la
mémoire autobiographique semblent en lien avec les processus de référence au self, les
mécanismes modulant l’activité de ces régions médianes pendant le rappel autobiographique
sont mal connus et les liens entre self et mémoire autobiographique au sein du CCP restent
flous. Summerfield et al. (2009) ont montré que la recollection d’évènements
autobiographiques réels impliquait spécifiquement le cortex préfrontal ventromédial et le
CCP. Tandis que d’autres régions du cortex retrosplénial et du cortex préfrontal médial
ventral et dorsal seraient spécifiquement impliquées dans des évènements liés au self qu’ils
soient réels ou imaginés. De façon contradictoire, il a été suggéré que le CCP serait
essentiellement impliqué dans les processus de référence à sa propre personne plutôt que
directement dans la mémoire épisodique autobiographique en soi. Cette dissociation a pu être
montrée en IRMf (Sajonz et al. 2010).
Cette dernière étude va dans le sens des travaux cytologiques et en imagerie
fonctionnelle de Vogt et al. (1995, 2006). Ils ont permis d’avancer dans la connaissance de
l’architecture du CCP qui s’avère inclure deux subdivisions aboutissant à une dualité
fonctionnelle (figure 5). On distingue ainsi un CPP ventral (vCPP, aire 23 de Brodmann) et
un CPP dorsal (aire 31 de Brodmann). Les deux subdvisions reçoivent des afférences
différentes des voies visuelles respectivement ventrales et dorsales. Le vCPP possède
principalement des interactions avec le cortex cingulaire subgénual afin de permettre le
traitement des émotions, des informations et des objets, tous en lien avec le self d’un individu.
Tandis que le dCPP possède des interactions préférentielles avec la région motrice cingulaire

19
antérieure et participe à l’orientation du corps dans l’espace. Le CRS (aires 29 et 30 de
Brodmann) a lui des liens étroits avec le cortex péricalleux et le thalamus. Ces afférences
thalamiques expliquent la fonction du CRS dans l’accès aux souvenirs de la mémoire à long
terme, rejoignant ainsi le modèle de Cabeza et Saint Jacques qui lui attribuent un rôle dans la
recollection.
Leech et al. (2012) confirment en IRMf cette architecture fonctionnelle complexe du
CPP avec de multiples sous-régions. Mais cette équipe va plus loin en attribuant au CCP un
rôle actif dans la régulation de la cognition. Ils retrouvent une région prédominante ventrale
avec une forte connectivité fonctionnelle avec le reste du DMN. Ils montrent l’existence de
deux sous-régions au sein du CCP dorsal avec une haute connectivité avec un réseau fronto-
pariétal. Or ces dernières sous-régions sont les seules à afficher un pattern d’activité BOLD
distinct du reste du CCP avec une hypoactivation durant la réalisation de tâches coûteuses sur
le plan attentionnel et corrélée à l’activation du réseau fronto-pariétal. Alors que les régions
ventrales liées au DMN ne s’inactivent pas. Ces résultats suggèrent qu’une partie importante
du CCP continue d’être active lors d’une demande attentionnelle forte. Le CCP dorsal
pourrait donc réguler une sorte de balance entre la cognition dirigée vers le monde interne
(introspection) et celle dirigée vers le monde externe (environnement), (Leech et al. 2012).
Ces résultats vont dans le sens de la haute activité métabolique de cette région et pourrait
unifier deux hypothèses actuelles (Buckner et al. 2008) quant au rôle exact du réseau par
défaut : une fonction introspective tournée vers soi ou une fonction de sentinelle de
l’environnement sensée réagir à des évènements potentiels quand l’attention est relâchée vis à
vis des stimuli de l’environnement extérieur.
Enfin, Vogt et al. (2005) associent le CCP, le CRS et le précunéus et les intègrent au
sein du réseau neuronal de la conscience (neural network correlates of consciousness). Ils leur
attribuent même un rôle pivot (Vogt et al. 2005). Chez des sujets souffrant de la variante
comportementale de démence frontotemporale avec altération de la conscience autonoétique,
était retrouvé un hypométabolisme en PET FDG (tomographie par émission de positons au
fluorodésoxyglucose) dans le cortex frontal antérieur et moyen médial, le cortex pariétal
inférieur gauche mais aussi dans le cortex cingulaire postérieur qui n’est pas une région
habituellement connue pour être atteinte dans la démence frontotemporale (Bastin et al.
2012), soulignant son implication dans les mécanismes de métaconscience.

20
On comprend qu’il persiste toujours de grandes incertitudes quant aux fonctions
précises du CCP. Son implication démontrée dans les processus cognitifs très élaborés que
sont le self, la conscience, la mémoire autobiographique, et ce rôle de régulateur cognitif
(approché par Leech et al.) en fait très probablement une région clé pour la bascule
attentionnelle nécessaire au rappel en mémoire autobiographique. Mais à ce jour la plupart
des études se concentrent sur de l’imagerie fonctionnelle, de rares analyses
électrophysiologiques intracérébrales dans le CCP ont été rapportées chez l’homme, et
aucune n’a cherché à mieux comprendre le décours temporel de l’activité du CCP lors
d’une tâche autobiographique. Ces hypo/hyperactivations observées en IRMf pourraient
refléter des patterns d’activités électrophysiologiques beaucoup plus complexes qu’un
simple antagonisme de signaux BOLD.
III - ELECTROPHYSIOLOGIE DE LA MEMOIRE AUTOBIOGRAPHIQUE
1- L’étude des états cérébraux
Les états cérébraux se définissent comme des patterns d’activité cérébrale qui
impliquent la coactivation ou la connectivité de multiples réseaux neuronaux à large échelle.
Une revue récente de l’équipe de Posner (Tang et al. 2012) explore les corrélats neuronaux de
maintenance d’un état ou de bascule entre deux états cérébraux. Parmi ces états cérébraux on
retrouve les états d’éveil (dont le default mode network) et l’état de sommeil. Les états
cérébraux semblent également jouer le rôle de prédicteur de performance dans une variété de
tâches perceptuelles, mnésiques ou de résolution de problèmes. Les états d’éveil ont été
beaucoup moins étudiés que le sommeil, que ce soit chez l’homme et chez l’animal. Et
souvent la focalisation des recherches ne se fait que sur un seul état cérébral. Les mécanismes
mêmes à l’origine du maintien d’un état cérébral ou la bascule entre deux états cérébraux
différents ont également été peu étudiés. C’est donc dans le champ des états cérébraux d’éveil
que nous inscrivons notre étude électrophysiologique. Il n’est pas possible de parler de rappel
en mémoire autobiographique sans évoquer une bascule et une transition entre deux
états cérébraux. Il a déjà été démontré en EEG de surface que le passage entre deux états
attentionnels différents (par exemple se perdre dans ses pensées et se concentrer sur sa
respiration) correspondait à deux états cérébraux distincts sur le plan électrophysiologique
(Braboszcz et al. 2011).
2- Oscillations cérébrales et mémoire autobiographique
La construction d’un souvenir épisodique est coûteuse sur le plan cognitif, et demande
un temps de recherche variable, parfois long. Bien qu’ils soient performants sur le plan

21
spatial, l’IRM fonctionnelle et le PET ont une résolution temporelle insuffisante pour bien
comprendre les différentes phases du rappel, notamment les phases les plus précoces comme
celles impliquées dans la bascule attentionnelle et l’initiation de la recherche du souvenir. Par
sa forte résolution temporelle l’électrophysiologie trouve ici toute sa place. La résolution
spatiale s’en trouve améliorée par le biais d’enregistrements intracérébraux. Les explorations
intracérébrales peuvent apparaître suffisantes chez l’animal pour certains types de mémoire
comme l’apprentissage associatif. Mais la mémoire autobiographique, en lien avec le self de
chaque individu, requiert l’introspection ; elle ne peut donc s’étudier que chez l’homme. Les
sujets épileptiques en cours de bilan pré-chirurgicaux, apportent l’opportunité unique
d’approcher les corrélats neuronaux de la mémoire au plus près, d’autant plus que beaucoup
d’implantations concernent des structures dont on connaît l’importance dans l’encodage et la
récupération mnésiques comme le lobe temporal interne.
Il existe une littérature florissante sur les oscillations cérébrales et leur association aux
processus cognitifs, notamment la mémoire, avec des rôles différents attribués à chaque bande
de fréquence. Une des hypothèses principales est que tout acte cognitif correspond à
l’émergence d’une assemblée neuronale distribuée spécifique transitoire et synchrone
caractérisée par le niveau de synchronie de ses constituants (Varela et al. 2001). Les
interactions, dont la synchronisation, se font à la fois à un niveau local et à large échelle entre
différentes régions cérébrales. Les oscillations constituent ainsi un des mécanismes neuronaux
clés dans le stockage et la récupération au sein de la mémoire à long terme (Fell et Axmacher
2011 pour revue). Mais peu de travaux ont étudié spécifiquement le champ de la mémoire
autobiographique. En détaillant les connaissances actuelles attribuées au différentes
oscillations, nous insisterons seulement sur les travaux exploitant la mémoire à long terme et
uniquement sur ceux étudiant les mécanismes de rappel et de récupération des souvenirs
(memory retrieval) qui correspond au thème de notre travail.
L’une des premières études électrophysiologiques à s’intéresser spécifiquement à la
mémoire autobiographique est celle de l’équipe de Conway en 2003. Les auteurs ont étudié en
EEG de surface les changements de potentiels corticaux lors de la génération d’un souvenir
sur mot-indice. Ils ont réussi à montrer une dynamique du rappel autobiographique avec une
activation prédominante initiale dans le cortex frontal gauche pour aboutir à la formation du
souvenir, puis l’activation augmente dans les réseaux postérieurs de l’hémisphère
controlatéral (cortex temporal et occipital droits) et se maintient pendant que l’événement est
« visualisé mentalement » et maintenu à la conscience (Conway et al. 2003).

22
Les oscillations dans les bandes thêta et gamma sont sûrement celles qui ont été le plus
observées et étudiées dans le champ de la mémoire à long terme. Une augmentation de la
synchronisation dans la bande thêta (4-8 Hz) et dans la bande gamma (30-100 Hz) jouerait un
rôle primordial dans l’encodage et la récupération via des effets sur la plasticité synaptique et
sur la coordination d’assemblées neuronales aboutissant à la réactivation des souvenirs.
Une augmentation de la puissance et une synchronisation dans la bande thêta
(définie de 4 à 8 Hz par Kahana et al. 2001) a été enregistrée dans plusieurs études en EEG de
scalp durant l’encodage et la recollection. La bande thêta est impliquée dans l’encodage de
nouvelles informations en facilitant la potentialisation à long terme. Elle serait par ailleurs
plus importante durant le rappel épisodique que pendant l’encodage (Klimesh et al. 1999,
2001). Généralement ces oscillations thêtas sont retrouvées dans des régions cérébrales
étendues.
Elle interviennent également dans les fonctions exécutives liées à la mémoire de
travail et participent à la maintenance d’une information en mémoire à court terme avec une
puissance augmentant de façon parallèle à l’augmentation de la quantité d’information à
maintenir. Cette activité thêta est alors maintenue pendant toute la période de rétention
(Jensen et Tesche, 2002), principalement dans les régions frontales et serait la conséquence
d’une activité neuronale soutenue reflétant la maintenance de représentations mnésiques. Son
rôle a aussi été démontré dans l’attention soutenue au niveau de sites frontaux médian où l’on
observe une activité thêta rythmique (‘frontal midline thêta’) pendant que les sujets effectuent
une tâche exigeante, cette activité augmente avec la charge d’information (Gevins et al. 1997,
Onton et al. 2005).
Les synchronies dans la bande gamma (30-100 Hz) sont le reflet de l’activité de
petits réseaux neuronaux locaux dans des aires spécialisées et seraient importantes pour la
communication neuronale (Fries et al. 2005). Les réponses gamma reflètent ainsi des
connexions corticocorticales au sein d’aires hautement associatives et perceptuelles. Ces
dernières sont par exemple impliquées dans l’activation d’une représentation corticale
perceptuelle et sémantique d’un objet (Tallon Baudry et al. 1999). Il est difficile d’associer les
activités gamma avec un processus perceptuel ou cognitif spécifique mais elles tiendraient un
rôle central dans la mémoire (à long terme et de travail) et l’attention dirigée (Jensen, Kaiser
et Lachaux, pour revue, 2007, Jerbi et al. 2009).

23
Les enregistrements intracrâniens ont permis de progresser dans la connaissance du
rôle de ces bandes de fréquence et de leurs interrelations.
Il a déjà été proposé que le transfert d’informations entre les systèmes hippocampiques
et corticaux associatifs était médié par les oscillations thêtas (Kirk et Mackay 2003). Ainsi
lors de stimulation intracérébrale de la région périrhinale produisant la réminiscence de
souvenirs sémantiques autobiographiques chez un sujet en bilan pré-chirurgical d’une
épilepsie, une synchronisation dans la bande thêta est retrouvée entre les structures du lobe
temporal médian, ainsi qu’entre ces structures et une aire visuelle primaire, cette
synchronisation entre régions distantes étant nécessaire pour produire le phénomène
expérientiel (Barbeau et al. 2005) .
Une augmentation de l’activité gamma locale dans l’hippocampe et une
synchronisation entre les aires rhinales et hippocampiques ont été observées durant un
encodage épisodique réussi en EEG intracrânien (Fell et al. 2003). Mais une augmentation de
l’activité gamma reflète aussi un rappel mnésique réussi. Il est désormais acquis que les
oscillations thêta et gamma dans des régions cérébrales étendues varient en amplitude si un
stimulus est rappelé avec succès (Sederberg et al. 2003). De plus pendant le rappel mnésique,
l’activité thêta et gamma apparaît dans les mêmes régions que lors de l’encodage (Sederberg
et al. 2007) : le même pattern dans les hautes fréquences gamma « 44-100 Hz » était activé à
la fois lors de l’encodage pour les items qui seraient rappelés, et lors du rappel pour les items
correctement rappelés. Cela va dans le sens de résultats qui ont montré que la récupération
mnésique impliquait le même pattern d’activité cérébrale que durant l’apprentissage (Polyn et
al. 2007). Enfin, une augmentation de puissance gamma et une synchronisation de phase entre
sites distants correspondrait à l’activation récurrente d’assemblées cellulaires reflétant l’accès
à des traces mnésiques existantes (Gruber et al. 2002) mais également la formation de
nouvelles représentations corticales (Gruber et al. 2005). Durant le rappel l’activité gamma
témoignerait ainsi de la réactivation de représentations « encodées sur le plan synaptique ».
Les modèles cognitifs de la mémoire de reconnaissance font la distinction entre une
reconnaissance par recollection et une reconnaissance par familiarité, qui seraient supportées
par des réseaux neuronaux différents. La recollection s’accompagne de différentes
informations contextuelles donc d’informations sensorielles et spatiales qui ont besoin d’être
« associées » entre elles pour obtenir un souvenir et ce en dépit d’une distribution disparate de
ces informations dans les différentes régions du cerveau. De larges interconnexions avec
différentes régions cérébrales font du lobe temporal médian l’une des principales zones de
convergence de ces informations. Les oscillations thêta et gamma jouent très probablement un

24
rôle dans la coordination de ces interactions à longue distance nécessaires à une recollection.
En effet l’amplitude des oscillations thêta est plus importante pendant la recollection que
pendant la familiarité (Guderian et Duzel 2005) ; elles seraient mêmes spécifiquement
associées à la recollection (Gruber et al. 2008). Les oscillations gamma (dans les bandes 35-
80 Hz) seraient quant à elles associées au processus de familiarité (Gruber et al. 2008).
Néanmoins il existe peu de travaux en EEG intracrânien dans le cadre spécifique du
rappel en mémoire autobiographique. Une étude récente illustre les interrelations entre ces
deux bandes de fréquence et montre qu’elles peuvent refléter des interactions dynamiques
entre le système hippocampique et le néocortex lors du rappel autobiographique. Des
enregistrements intracrâniens par microélectrodes au sein du cortex entorhinal humain ont été
réalisées pendant une recherche en mémoire autobiographique lors de la présentation d’un
indice aboutissant à la recollection de souvenirs remontant à plus de dix ans. Les oscillations
gamma prédominaient dans les couches superficielles à projections hippocampiques et
recevant des afférences néocorticales. Cette augmentation de puissance dans la bande gamma
apparaissait très précocement lors de la présentation de l’indice et se prolongeait tout au long
du rappel. Pour les auteurs elles reflèteraient la correspondance entre la phrase indice et les
représentations de la mémoire à long terme, pour que celles-ci soient réexploitées pour
produire la reminiscence du souvenir. L’augmentation de la puissance thêta prolongée tout au
long du rappel prédominait dans les couches à projection néocorticales. Ces oscillations
thêtas, enregistrées simultanément dans les cortex préfrontaux et temporaux, reflèteraient
donc les processus de transfert d’informations entre la formation hippocampique et les aires
corticales associatives quand le sujet essaie de se rappeler le souvenir autobiographique, en
synchronisant l’activité d’aires cérébrales éloignées (Steinvorth et al. 2009).
En 2011, l’équipe de Dastjerdi explorent en EEG intracrânien, une large région
cérébrale postéro-médiale, incluant CPP et CRS. Une majorité des sites d’enregistrement
montrait une hyperactivation gamma au repos (30-180 Hz), tandis que d’autres sites, proches
du CRS, ne répondaient qu’à des stimuli autobiographiques. Sans étudier spécifiquement le
rappel autobiographique, mais principalement la référence au self, ils confirment sur le plan
électrophysiologique, l’hétérogénéité de cette région. Nous reviendrons plus longuement sur
les résultats de cette étude dans notre discussion.

25
PROBLEMATIQUE ET HYPOTHESES DE TRAVAIL
Notre étude porte sur les processus en œuvre pendant le rappel d’épisodes
autobiographiques. Elle a été menée chez des sujets épileptiques implantés dans le cadre
d’une stéréo-électroencéphalographie (SEEG) pour un bilan préchirurgical.
Les travaux de l’équipe de Steinvorth ont montré l’importance des informations
apportées par les enregistrements intracrâniens et leur application pour comprendre le rappel
en mémoire autobiographique. Ils ne se sont focalisés que sur une seule région cérébrale
connue pour être impliquée dans le réseau de la mémoire autobiographique : le cortex
entorrhinal. Ils ont ainsi pu préciser le rôle fonctionnel de cette région et ses interrelations
hippocampiques et frontales.
Au sein du réseau de rappel de la mémoire autobiographique et des processus du self,
une structure clé comme nous l’avons montré plus haut n’a pour l’instant toujours fait l’objet
que de rares études électrophysiologiques intracérébrales : le cortex cingulaire postérieur.
L’étude de Leech insiste sur l’architecture complexe du CCP et sur son rôle de régulateur
cognitif d’une balance entre des activités cognitives orienté vers l’environnement et des
activités d’introspection. Il pourrait donc participer à cette bascule attentionnelle définie plus
haut et nécessaire au rappel en mémoire autobiographique.
Le but principal de ce travail de master 2 est d’approcher les phénomènes
électrophysiologiques au sein de cortex cingulaire postérieur lors du rappel d’épisodes en
mémoire autobiographique, avec les hypothèses suivantes :
- la dynamique des activités au sein du CCP devrait être évocatrice d’une bascule entre
états attentionnels différents et donc entre états cérébraux différents lors du rappel
autobiographique.
- les patterns électrophysiologiques obtenus devraient être spécifiques à la mémoire
autobiographique, tant lors de la bascule que lors du rappel.
- nous devrions également observer dans cette région cérébrale les mêmes gammes de
fréquence que l’on peut observer au sein du cortex entorhinal : une activation dans les
bandes thêta et gamma puisque celles-ci permettent des interrelations dynamiques entre
régions cérébrales distantes comme le lobe temporal interne et le lobe frontal.
- l’activité électrophysiologique devrait être différente en fonction des sous-régions du
CCP.
Les activités électrophysiologiques enregistrées devraient nous aider à préciser le rôle
fonctionnel du gyrus cingulaire postérieur dans le rappel autobiographique.

26
METHODES
I. Population
Les enregistrements électrophysiologiques intracrâniens ont pu être obtenus à partir de
6 patients en cours de bilan préchirurgical d’une épilepsie partielle pharmacorésistante avec
une implication temporale interne, dans l’unité de chirurgie de l’épilepsie du CHU Rangueil
de Toulouse entre février 2012 et Juin 2012. Ils sont porteurs d’une épilepsie partielle
symptomatique pharmacorésistante nécessitant la réalisation d’une SEEG pour mieux
localiser la zone épileptogène. L’épilepsie de ces patients est invalidante, avec persistance de
nombreuses crises et impact majeur sur leur qualité de vie. Le bilan préchirurgical de phase I
comprenant une étude précise de la clinique (vidéoEEG) et de l’EEG des crises et intercrises,
de l’IRM morphologique et d’une imagerie nucléaire (PET métabolique ou SPECT critique),
n’a pas permis de conclure avec certitude sur la localisation de la zone épileptogène. Tous les
patients ont signé un consentement écrit avant la participation à cette étude.
II. Paradigme Expérimental
Trois tâches ont été proposées aléatoirement aux sujets (figure 8) :
- une tâche de mémoire autobiographique inspirée de la méthode des mots indices. Il est
demandé au sujet de se remémorer un souvenir sur un item précis : par exemple, « votre
examen du permis de conduire », « une dispute », « un moment où vous avez eu honte »…
- une tâche de mémoire sémantique, où le sujet doit répondre à des questions sur des
personnalités célèbres « Comment est décédé Claude Francois ? ». Ce sont des questions
difficiles concernant des personnalités historiques ou impliquées dans différents domaines de
la vie publique.
- une tâche de calcul mental, consistant en une suite de soustraction par le même chiffre
pendant un temps de douze secondes. Par exemple « 575-5-5…. »
L’expérience est organisée par blocs de quinze questions, 5 de chaque tâche. Chaque question
est précédée d’une période de cinq secondes pendant laquelle le sujet doit fixer une croix
centrale sur un écran d’ordinateur. Cette période de cinq secondes correspond à la ligne de
base utilisée pour les analyses spectrales de l’EEG.
A la fin de la question autobiographique posée oralement par l’examinateur, un trigger
est déclenché. Les sujets sont entrainés à reconnaître les deux principales phases du rappel

27
d’un souvenir : à savoir un temps de recherche nécessaire pour accéder au bon souvenir, et un
temps de réminiscence où le souvenir est retrouvé et revécu intérieurement par le sujet. A la
fin de la phase de recherche puis de la phase de réminiscence les sujets appuient sur les
touches d’un boitier qui envoie un trigger sur l’enregistrement électro-encéphalographique.
La même structure de réponse est obtenue pour les questions sémantiques. A la fin de la
phase de réminiscence, un débriefing est effectué avec le sujet pour coter le souvenir et
s’assurer de son épisodicité.
Figure 8 : design expérimental. Trois tâches sont proposées aléatoirement par blocs de 15 questions, 5 pour chacune des tâches : une tâche
autobiographique, une tâche sémantique et une tâche de calcul mental. Cette dernière tâche fait appel à des ressources
attentionnelles différentes des tâches mnésiques et n’implique pas la mémoire à long terme. Elles sont toutes précédées d’une
période de silence de 5 secondes, pendant laquelle on demande au sujet de fixer une croix sur un écran d’ordinateur. Cette
période de référence correspond à la ligne de base utilisée pour l’analyse du signal EEG. Les tâches sémantiques et
autobiographiques épisodiques ont une structure similaire. Les sujets sont entrainés à repérer deux phases pour aboutir à la
réponse. Tout d’abord une phase souvent longue de recherche avant d’aboutir à la réponse. Puis dans le cadre de la
mémoire autobiographique, une phase où le souvenir retrouvé est revécu : la réminiscence. Dans le cadre de la tâche
sémantique, cette seconde phase consiste en la visualisation des différents éléments de la réponse.

28
Sont considérés comme épisodiques, tous les souvenirs avec localisation précise dans
le temps, l’espace, de durée brève inférieure à 24 heures, pour lesquelles peuvent être
rapportés les personnes présentes et un discours narratif avec plusieurs des éléments suivants :
émotions, objets précis, caractéristiques sensorielles, odeurs, mouvement, importance dans la
vie du sujet, point de vue acteur. Chacune de ces caractéristiques est cotée : les souvenirs ne
remplissant pas l’ensemble de ces critères sont rejetés pour l’analyse spectrale. En raison des
conditions expérimentales (patients hospitalisés et fatigables, tâches expérimentales réalisées
dans des créneaux horaires restreints) et du besoin de nombreux essais, il apparaît difficile
d’obtenir un débriefing plus long avec l’ajout par exemple d’une tâche RKG proposée dans le
TEMPau et une cotation complète de l’autonoéticité proposée par Irish (2008). Pour les tâches
contrôles, les réponses aux questions sémantiques sont retenues si le bon concept culturel est
retrouvé avec plusieurs précisions correctes apportées sur la personne célèbre, et pour le
calcul seuls les essais aboutissant à un résultat exact sont retenus. Seront rejetés lors de
l’analyse du signal, tous les essais où le sujet a parlé pendant la question, le rappel ou s’il n’a
pas respecté la ligne de base.
III. Electrophysiologie
1. Stéréo-électroencéphalographie et bilan pré-chirurgical d’épilepsie
Le but de l’exploration SEEG est de retenir ou réfuter une éventuelle intervention
neurochirurgicale en précisant le réseau épileptique en étudiant son origine et la zone de
propagation. Elle consiste en l’implantation chirurgicale stéréotaxique de macroélectrodes-
aiguilles multiplots en platine-iridium de 0.8 mm de diamètre (Dixi Medical) comportant de 4
à 16 plots en fonction de la structure cérébrale cible. La zone épileptogène suspectée et la
cartographie d’implantation des électrodes sont guidées par le neurologue en concertation
avec le neurochirurgien, les neuroradiologues et les médecins nucléaire sur la base du bilan de
phase I. Un repérage précis des électrodes, implantées orthogonalement ou non (l’électrode
orbitorolandique est oblique verticale par exemple) est effectué grâce à une IRM (imagerie
par résonance magnétique) stéréotaxique et une angiographie (angio-IRM 3 Tesla sur le CHU
de Toulouse) préopératoires. La topographie d’implantation des électrodes se base
uniquement sur des critères cliniques ou paracliniques en rapport avec le réseau épileptogène
supposé et déduit à partir de la sémiologie clinique des crises. Le schéma d’implantation n’est
donc en rien influencé par notre protocole de recherche.

29
2. Localisation des électrodes
Les contacts des électrodes sont anatomiquement localisés dans l’Atlas de Talairach et
Tournoux (1993). Les images (DICOM) de l’IRM morphologique préopératoire
T1+gadolinium et du scanner avec électrodes postopératoire ont été normalisées avec un
template T1 SPM (template utilisé habituellement par le logiciel Statistical Parametric
Mapping, dans référentiel MNI Montreal neurological institute), à l’aide de la boite à outil
Sysiphe développée dans notre institut de recherche. Les coordonnées sont mesurées par
rapport à la référence CA (placée à l’aide de la commissure antérieure) grâce au logiciel
Sysiphe et sont ensuite converties du référentiel MNI vers le référentiel de Talairach. La
localisation cérébrale s’est ensuite faite via le logiciel Talairach Client. Un contrôle visuel des
différents contacts est également effectué sur une superposition du schéma d’implantation
stéréotaxique de chaque patient avec l’IRM morphologique préopératoire (séquence T1), pour
vérifier que les contacts d’intérêt sont en substance grise. Nous avons exclu les contacts en
substance blanche.
3. Traitement du signal
Les oscillations cérébrales, enregistrées au moyen de l’EEG, sont des évènements
transitoires, bruités, non stationnaires et non linéaires, statistiquement imprévisibles et sujets à
des discontinuités temporelles lors des états de transitions. Une analyse des variations
fréquentielles permet donc de décrire ces évènements.
L’électroencéphalogramme standard (de scalp) enregistre ces variations de potentiels
électriques à la surface du crâne et est constitué de la somme de potentiels neuronaux
postsynaptiques synchronisés. Il est sujet à de multiples artéfacts (environnement, musculaire,
cardiaques) et offre une résolution spatiale faible, notamment pour connaître la source précise
de l’activité oscillatoire. Il ne permet pas d’accéder à l’activité électrique des structures
profondes (comme les structures temporales internes ou rolandiques). Si elle est effectivement
invasive et ne permet pas d’étudier des surfaces corticales très étendues avec un hémisphère
souvent privilégié par rapport à l’autre, la technique de SEEG est la seule technique offrant un
accès vers ces populations neuronales très profondes et difficilement accessibles qui ne sont
pas ou que partiellement explorées par l’EEG de surface. Régulièrement, pour les besoins
diagnostiques, des électrodes sont implantées dans des régions qui ont montré leur implication
dans des processus mnésiques : l’amygdale, l’hippocampe, les cortex entorhinal et perirhinal,
le cortex préfrontal et le cortex cingulaire postérieur. L’avantage de ce type d’enregistrement
est d’apporter une source d’information unique dans le domaine des neurosciences :

30
l’enregistrement de local fields potentials (LFP), c’est à dire de la somme des potentiels des
activités synaptiques et dendritiques dans des populations neuronales limitées, donc d’offrir
des enregistrements électrophysiologiques humains in vivo avec une forte résolution spatiale.
4. Enregistrement des données et pré-processing
Les données électroencéphalographiques numériques ont toutes été enregistrées avec
une fréquence d’échantillonnage de 256 Hz ou 512 Hz (Système Micromed). Les données
enregistrées à 512 Hz ont été sous-échantillonées a posteriori à 256 Hz. Le traitement des
signaux électroencéphalographiques s’est effectué sous Matlab avec la boîte à outil EEGlab
10.2.2.4b. Les données ont été filtrées à l’aide d’un filtre passe haut de 0.5 Hz et d’un filtre
passe bas de 100 Hz. L’objectif étant d’obtenir des résultats dans des gammes fréquentielles
suffisamment étendues : des fréquences les plus basses dans la bande delta (1-4Hz) aux
fréquences les plus hautes gamma (30 à 100 Hz) qui ont toutes montré une implication et des
rôles différents dans les processus cognitifs. Les fréquences les plus hautes, au-delà de 100 Hz
n’ont pu être étudiées en raison de la fréquence d’échantillonnage (insuffisamment haute) des
données stéréo-électroencéphalographiques. Un filtre analogique coupe-bande entre 45 et 55
Hz a été appliqué pour limiter le bruit et les perturbations du signal à 50 Hz liés au courant de
secteur. Par un montage référentiel, utilisant pour référence une seule électrode ou un
moyennage commun d’électrodes, on assiste à une amplification des artefacts et des
anomalies intercritiques sur les électrodes à distance de la source du paroxysme. Pour cette
raison nous avons réalisé l’analyse des données sur un montage bipolaire qui annule ce
problème et qui offre une plus haute spécificité spatiale de l’ordre de 3 mm (Lachaux et al.
2003, Jerbi et al. 2009). Chaque contact a donc été re-référencé avec son contact voisin sous
Matlab.
L’ensemble des données de l’étude a été acquis à distance de toute crise épileptique
avec le respect d’au moins 12 heures après une crise comitiale, afin de limiter l’impact de
l’activité épileptique sur le tracé EEG et les fonctions cognitives du patient. Le protocole se
déroule les premiers jours de l’exploration vidéo-SEEG également dans le but d’obtenir des
données avant une dégradation du signal EEG au fur et à mesure du sevrage en traitement
anti-épileptique et de la répétition des crises. Une analyse visuelle permet ensuite le rejet des
artefacts d’électrodes et des anomalies épileptiques. La fenêtre de lecture choisie est similaire
à celle utilisée en clinique par l’équipe médicale pour analyser les tracées EEG et en déduire
le réseau critique et la localisation des activités paroxystiques intercritiques. La fenêtre de
lecture comprenait donc 30 secondes de signal en abscisse, entre 30 et 40 « contacts

31
bipolaires » en ordonnée et une amplitude moyenne de 400 microvolts/cm. Ont été rejetés
avant l’analyse des données tous les contacts montrant des artefacts (artefacts d’électrodes
principalement) et tous ceux avec présence d’au moins une anomalie paroxystique évocatrice
d’une anomalie épileptique. Ce choix entraine évidemment le rejet de nombreux contacts,
souvent en zone d’intérêt comme les zones temporales internes, mais limite le risque
d’analyse d’un signal obtenu en tissu malade.
5. Analyse spectrale et décomposition temps-fréquence
Nous avons utilisé deux techniques d’analyse spectrale des enregistrements
neuronaux :
1- d’abord une décomposition temps fréquence par ondelettes pour obtenir les Event
related spectral perturbations (ERSP) et ainsi avoir une vision globale de la
dynamique sur la fenêtre temporelle d’intérêt au travers des différents essais et sur
chaque contact d’enregistrement.
En électroencéphalographie, le modèle classique de potentiels évoqués (event related
potential ou ERP) a souvent montré ses limites pour étudier des dynamiques cérébrales
complexes. Les ERP sont une mesure de l’amplitude et de la latence (non constante et
dépendante de l’activité EEG en cours) de pics post-stimulus sur une moyenne d’essais EEG
par rapport à un stimulus ; ils représentent une combinaison complexe de la synchronisation
de phase du signal EEG provoquée par un stimulus et de l’augmentation de l’amplitude
spectrale. C’est pour ces différentes raisons que les techniques de décomposition spectrale du
signal comme les event related spectral perturbations (ERSP) sont désormais développées
depuis deux décennies (Makeig et al. 1993, 2004).
Elles consistent en la représentation des changements moyens de puissance spectrale
en réponse à un stimulus, comparativement à une ligne de base (c'est-à-dire par rapport à un
état de référence). Là où les potentiels évoqués vont se focaliser uniquement sur le domaine
temporel, l’approche par ERSP va permettre de caractériser des dynamiques cérébrales
complexes à la fois dans le domaine fréquentiel et dans le domaine temporel. L’ERSP
consiste en un moyennage du spectre de puissance dans des fenêtres temporelles, qui
s’effectue sur de multiples essais « centrés » par rapport à un stimulus. Les ERSP sont donc
des représentations à trois dimensions : le temps, la fréquence et la puissance (figure 9). Pour
des raisons de lecture, ils sont généralement représentés sous 2 dimensions avec des cartes
représentant en ordonnée la fréquence et en abscisse le décours temporel. La variation de la

32
puissance du signal après décomposition spectrale est alors représentée par une échelle de
couleur à différents points temps- fréquence.
Figure 9 : exemple de carte ERSP à partir du signal EEG de l’électrode T8-T9 d’un sujet.
L’image du haut représente l’enregistrement électro-encéphalographique numérisé d’un sujet. En ordonnée sont
représentées les différentes électrodes, référencées par rapport à leur contact voisin (montage bipolaire). L’amplitude du
signal est de 400 microvolts/cm. L’abscisse correspond au temps. Des triggers (barres verticales colorées) apparaissent au
cours de la tâche. Est représentée une époque avec 5 secondes de fin de question et 11.5 secondes de rappel. Le 0 correspond
à la fin de question et au tout début du rappel.
L’encadré rouge correspond au signal EEG du contact T8-T9. Sur ce contact est effectuée une décomposition
temps-fréquence du signal puis une division par rapport à une ligne de base pour obtenir un ERSP (figures du bas). Les
ERSP sont initialement des représentations à 3 dimensions. Ils sont convertis en cartes à 2 dimensions pour plus de facilité
de lecture. La puissance du signal apparait alors selon une échelle de couleur (unité Décibel).
La décomposition spectrale du signal s’est faite à partir du découpage du signal EEG
en fenêtres temporelles (ou époques), centrées sur un trigger qui dans notre travail est la fin de
la question. Les époques choisies sont de durée assez longue : 16.5 secondes (avec 5 secondes
de question suivies de 11.5 de recherche). Le temps de recherche du souvenir pouvait être
parfois plus court. Mais des époques longues apportent l’avantage de nous renseigner sur les

33
corrélats même du rappel, en permettant d’avoir toute une dynamique de récupération des
traces mnésiques. Certains essais épisodiques réussis comprenaient un temps de recherche
beaucoup plus long, mais ce temps de 11.5 secondes a été retenu pour permettre de retenir
suffisamment d’essais. On obtient donc trois ensembles d’époques après un découpage
(manuel via Matlab EEglab Toolbox) : un pool pour les essais épisodiques autobiographiques
retenus, un pour les essais sémantiques réussis, un pour les calculs mentaux réussis.
Nous avons effectué une décomposition du signal par ondelettes de Morlet avec un
nombre de cycles allant de 1 pour la plus basse fréquence et augmentant linéairement jusqu’ à
15 pour la plus haute fréquence (à l’aide de la fonction timefreq de Matlab, Eeglab toolbox).
Les ondelettes de Morlet sont fréquemment utilisées pour l’analyse de signaux EEG (Tallon
Baudry et al. 1997, Braboszcz et al. 2009). Nous avons utilisé 256 points temps (par seconde)
espacés linéairement et une série de 50 fréquences espacées de 1 à 100 Hz.
Pour obtenir des variations de puissance du signal dans les différentes gammes de
fréquence lors de la question et du rappel, le signal doit être comparé par rapport à un état de
référence où le sujet effectue une activité cognitive réduite. La ligne de base a été choisie sur
une période de cinq secondes avant chaque question (décrite plus haut). Différentes méthodes
de normalisation par rapport à la ligne de base existent (Grandchamps et Delorme 2011).
Nous avons choisi le gain model (détaillé dans Delorme and Makeig 2004) qui est l’un des
plus utilisé. Suivant ce modèle pour chaque bande de fréquence, la puissance de chaque point
temps fréquence pendant les temps question + rappel est divisée par la puissance spectrale
moyenne dans la période de ligne de base à la même fréquence. Deux mesures sont ensuite
obtenues : une mesure ERSP absolue et une mesure ERSP après transformation
logarithmique (ERSPlog). Pour des signaux asymétriques comme les signaux
électroencéphalographiques, la distribution logarithmique du signal apparait plus normale que
le signal original. Elle permet la visualisation de plus larges bandes de variations de puissance
et l’affichage des petites amplitudes. Nous avons choisi ce type de représentation. Par
définition, l’unité des ERSPlog est le décibel. Cette mesure est régulièrement utilisée dans la
littérature (Delorme et al. 2007).
2- puis une décomposition par transformée de Hilbert sur des bandes de fréquence
d’intérêt choisies au préalable en fonction des réponses ERSP.
Nous avons donc calculé par transformée de Hilbert, l’amplitude instantanée (l’enveloppe)
du signal EEG dans différentes bandes de fréquence d’intérêt moyennée sur l’ensemble des

34
essais. L’intérêt de la transformée de Hilbert est de pouvoir s’intéresser à l’amplitude du
signal en extrayant et dissociant d’un signal EEG, composite et sinusoïdal, la phase et
l’amplitude. Les différences de puissance du signal lors des étapes question et rappel ont pu
être comparées entre conditions (sémantique, calcul et autobiographique) à l’aide d’un test
des rangs signés de Wilcoxon (test utilisé par d’autres équipes en EEG intracrânien, Ossandon
et al. 2012) dans des bandes de fréquence limitées et choisies à partir des résultats des ERSP
afin de diminuer la complexité des données et donc d’obtenir une plus haute puissance
statistique. Le test de Wilcoxon était suivi d’une correction par False Discovery Rate (taux de
fausses découvertes) afin de limiter le nombre de faux positifs dans ces longues séries
temporelles de données nécessitant de multiples tests (Genovese et al. 2002). La transformée
de Hilbert et les calculs statistiques ont été effectués à l’aide du logiciel Matlab.
L’équivalence des deux méthodes, ondelettes et transformée de Hilbert, dans l’étude de
signaux EEG intracrânien a déjà été démontrée (Le Van Quyen et al. 2001). Multiplier les
méthodes de décomposition du signal est donc un moyen de valider nos résultats, comme des
études en SEEG l’ont déjà appliqué (Ossandon et al. 2012).
RESULTATS
I. Caractéristiques des patients et résultats comportementaux
Le traitement du signal EEG a pu s’effectuer sur 5 des 6 patients (3 hommes et 2
femmes, âge moyen=38.6 +/- 7.2 ans). Leurs caractéristiques cliniques et leurs performances
aux trois tâches sont résumées dans le tableau 1. La première patiente a été exclue pour
plusieurs raisons : une activité épileptique intercritique intense avec un signal comportant de
nombreux paroxysmes épileptiques et un ralentissement sur la majorité des électrodes ; un
quotient intellectuel trop bas et des troubles du caractère empêchant la réalisation des tâches
contrôles (calcul mental et mémoire sémantique) ; un nombre trop faible d’essais épisodiques
pour l’analyse spectrale des données. Chacun des 5 sujets restants a pu réaliser entre 4 et 7
blocs de quinze questions. Dans la mesure où les questions autobiographiques portent sur eux-
mêmes, la tâche est bien acceptée par les patients. Elle est également bien comprise, en
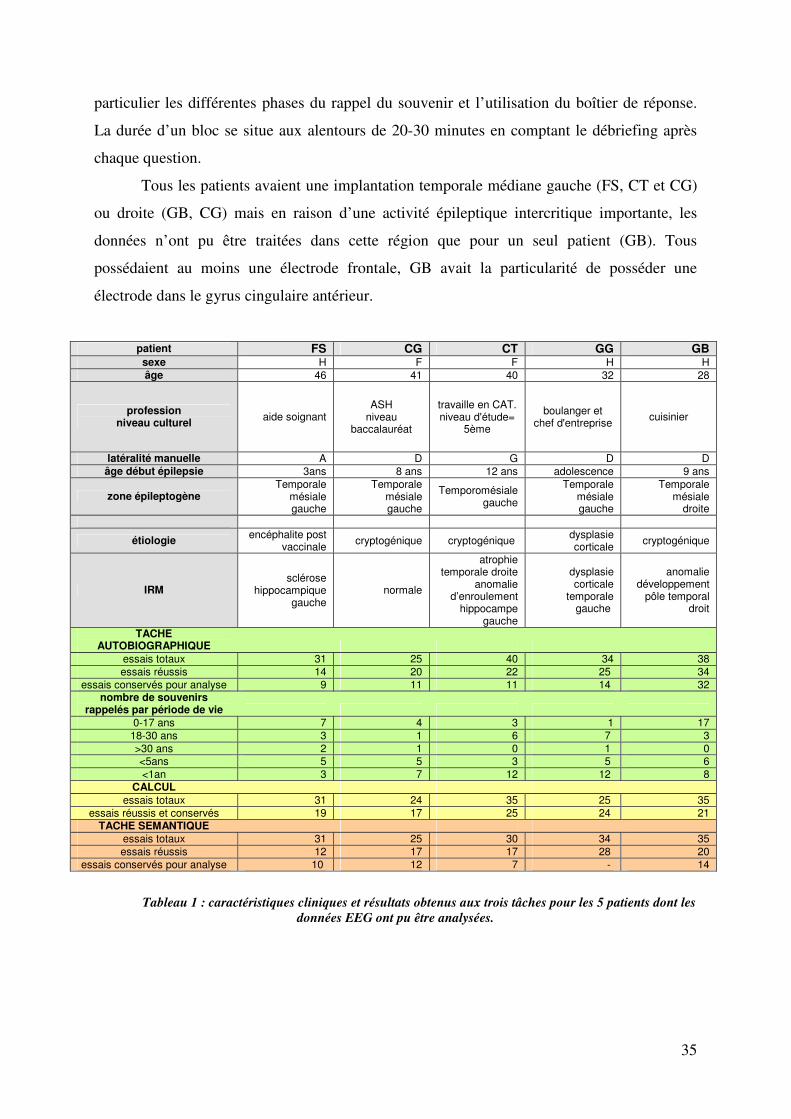
35
particulier les différentes phases du rappel du souvenir et l’utilisation du boîtier de réponse.
La durée d’un bloc se situe aux alentours de 20-30 minutes en comptant le débriefing après
chaque question.
Tous les patients avaient une implantation temporale médiane gauche (FS, CT et CG)
ou droite (GB, CG) mais en raison d’une activité épileptique intercritique importante, les
données n’ont pu être traitées dans cette région que pour un seul patient (GB). Tous
possédaient au moins une électrode frontale, GB avait la particularité de posséder une
électrode dans le gyrus cingulaire antérieur.
patient FS CG CT GG GB sexe H F F H H âge 46 41 40 32 28
profession niveau culturel
aide soignant ASH
niveau baccalauréat
travaille en CAT. niveau d'étude=
5ème
boulanger et chef d'entreprise
cuisinier
latéralité manuelle A D G D D âge début épilepsie 3ans 8 ans 12 ans adolescence 9 ans
zone épileptogène Temporale
mésiale gauche
Temporale mésiale gauche
Temporomésiale gauche
Temporale mésiale gauche
Temporale mésiale
droite
étiologie encéphalite post
vaccinale cryptogénique cryptogénique
dysplasie corticale
cryptogénique
IRM sclérose
hippocampique gauche
normale
atrophie temporale droite
anomalie d’enroulement
hippocampe gauche
dysplasie corticale
temporale gauche
anomalie développement
pôle temporal droit
TACHE AUTOBIOGRAPHIQUE
essais totaux 31 25 40 34 38
essais réussis 14 20 22 25 34
essais conservés pour analyse 9 11 11 14 32 nombre de souvenirs
rappelés par période de vie
0-17 ans 7 4 3 1 17
18-30 ans 3 1 6 7 3
>30 ans 2 1 0 1 0 <5ans 5 5 3 5 6
<1an 3 7 12 12 8 CALCUL
essais totaux 31 24 35 25 35
essais réussis et conservés 19 17 25 24 21 TACHE SEMANTIQUE
essais totaux 31 25 30 34 35
essais réussis 12 17 17 28 20
essais conservés pour analyse 10 12 7 - 14
Tableau 1 : caractéristiques cliniques et résultats obtenus aux trois tâches pour les 5 patients dont les
données EEG ont pu être analysées.

36
II. Reproductibilité des résultats entre les patients au sein du cortex
cingulaire postérieur
Trois patients (FS, CG, CT) possédaient une électrode traversant le cortex pariétal
latéral gauche (gyrus supramarginal ou gyrus angulaire) et se terminant au sein du gyrus
cingulaire postérieur gauche (figure 10). L’utilisation du référentiel de Talairach a permis de
confirmer que les contacts les plus internes de ces électrodes se trouvaient en substance grise
au sein du cortex cingulaire postérieur gauche dans l’aire de Brodmann 23 (BA 23): FS (-4, -
29, 22, BA 23) , CT (-5, -31, 22, BA 23) et CG (-4, -30, 18, BA 23).
Figure 10 : localisation des plots d’intérêt au sein cortex cingulaire postérieur pour les 4 patients.
Les plots des trois patients implantés sur l’hémisphère gauche sont au sein de l’aire 23 de Brodmann.
FS (-4, -29, 22, BA 23), CT (-5, -31, 22, BA 23) et CG (-4, -30, 18, BA 23). Le plot de GB (2, -27, 15) est positionné un
peu plus bas dans l’espace de Talairach.
A gauche, les tâches de mémoire à long terme sémantique et autobiographique, sont
responsables d’une activité électrophysiologique différente de la tâche de calcul, mais dont le
pattern apparait nettement reproductible entre les patients pour chaque tâche. Pour les trois
patients CT, FS et CG, nous obtenons des ERSP semblables. Notre méthode fonctionne et
apparait reproductible. Nous avons donc pu cumuler les différents essais réussis de chaque
patient dans un pool d’essais correspondant au cortex cingulaire postérieur gauche pour
chaque tâche. Une moyenne pondérée des ERSP a été réalisée en affectant un coefficient
correspondant au nombre d’essais réussis par patients pour chacune des trois tâches pour le
cortex cingulaire postérieur gauche. GB possédait une électrode proche du cortex cingulaire
postérieur droit mais plus basses que les électrodes gauches (2, -27, 15), il a donc été analysé
individuellement.

37
Figure 11 : Cartes ERSP obtenues lors des trois tâches pour les quatre patients au sein du CCP.
En haut sont représentées les cartes ERSP pour les patients implantés sur le CCP gauche. Il existe une nette reproductibilité des
résultats entre les trois patients pour chacune des trois tâches. Le pattern lors du calcul apparait nettement différent de celui des tâches
mnésiques avec une hypoactivation dans la bande gamma et une hyperactivation dans les bandes alpha (8-12 Hz) et thêta (4-8 Hz) pendant
les temps de question et de calcul. Le pattern apparait opposé lors de la tâche autobiographique, avec une hypoactivation dans les basses
fréquences thêtas et dans la bande alpha ; on note une hyperactivation gamma lors de la bascule attentionnelle.
Une moyenne pondérée est réalisée pour chacune des trois tâches, au sein du CCP gauche. Cette hyperactivité gamma lors de la
bascule attentionnelle apparait alors nettement et uniquement pour la tâche autobiographique, dans la bande gamma 30-100 Hz.
En bas sont représenté les résultats pour le patient GB, où il n’est objectivé aucune hyperactivité gamma, notamment pendant la
bascule attentionnelle autobiographique. Il faut souligner que ce patient présente un plot plus bas que les plots précédents, en dehors de la
BA 23.

38
III. Dynamique du signal au moment de la bascule attentionnelle au sein du
cortex cingulaire postérieur
Définie plus haut, nous nommerons « bascule attentionnelle », cette courte phase de
transition entre la fin de la phase de question et le début du rappel d’une seconde, allant
d’environ 500 ms avant la fin de la question et 500 ms seconde après.
Nous avons appliqué un test des rangs signés de Wilcoxon sur la puissance du signal
EEG dans différentes bandes de fréquence de 0.5 à 100 Hz sur l’ensemble des essais des
patients et dans chaque tâche. Les décompositions suivantes ont été appliquées : 1-4 Hz pour
la bande delta, 4-8 Hz pour la bande thêta, 8-12 Hz pour la bande alpha, 13-30 Hz pour la
bande bêta, 30-100Hz pour la bande gamma et plusieurs sous décompositions au sein de la
bande gamma (figure 12). Les principales modifications significatives du signal par
rapport à la ligne de base apparaissent dans la bande gamma, lors de cette bascule
attentionnelle.
Sur les ERSP, il apparait nettement une augmentation d’activité dans la bande gamma
et plus précisément dans deux sous-bandes aux alentours de 40-50 Hz et 60-100 Hz. Après
transformée de Hilbert et application d’un test des rangs signés de Wilcoxon, on note une
augmentation de puissance gamma significative lors de la bascule dans les sous-bandes 40-60
Hz, 60-80 Hz et 80-100 Hz dans la tâche autobiographique par rapport au calcul.
Il n’y a toutefois pas de différence significative dans la bande gamma entre les
conditions sémantiques et autobiographiques après transformée de Hilbert et application d’un
test des rangs signés Wilcoxon. Mais si on manque probablement de puissance statistique, il
apparait visuellement sur les ERSP une nette différence entre les tâches sémantique et
autobiographique : cette augmentation d’activité dans la bande gamma n’apparait que pour la
tâche autobiographique.
Il semble également exister une hypoactivation dans les fréquences plus basses (thêta
et alpha notamment) au moment de cette bascule attentionnelle lors du rappel
autobiographique. Elle est donc associée à une augmentation d’activité dans les plus hautes
fréquences. Cette tendance à une diminution de puissance du signal dans les fréquences alpha
et thêta est retrouvée après transformée de Hilbert, mais n’apparait pas statistiquement
significative.

39
Figure 12 : Dynamique du signal lors de la bascule attentionnelle dans le CCP gauche. Au milieu sont représentées les cartes ERSP correspondant aux moyennes pondérées pour les trois tâches, pour les patients CT +
CG + FS. Une décomposition du signal est effectuée par transformée de Hilbert, dans une fenêtre de -500 ms (question) à +500 ms (début
du rappel) correspondant à la bascule attentionnelle, dans les bandes thêta (4-8 Hz), alpha (8-12 Hz) et gamma ( sous-bandes 40-60 Hz, 60-
80 Hz et 80 Hz). Les tâches sont comparées entre elles (test des rangs signes de Wicoxon) et les différences significatives de signal entre
conditions apparaissent sur l’axe des abscisses sous forme d’un trait rouge (- = p<0.05).
Lors de la bascule attentionnelle, il existe une activité gamma significativement plus élevée pendant la tâche autobiographique
par rapport au calcul. Il n’est pas retrouvé de différence significative entre les conditions sémantique et autobiographique dans la bande
gamma.
Les cartes ERSP affichent une hypoactivation dans les bandes thêta et alpha par rapport au calcul, avec un pattern qui semble
anti-corrélé. Toutefois ces différences ne sont pas significatives après application d’une autre méthode de décomposition du signal par
transformée de Hilbert.

40
IV. Dynamique du signal avant la bascule attentionnelle et après lors du
rappel
Les ERSP affichent une baisse de puissance du signal dans les bandes thêta et alpha
pendant l’ensemble de la période couvrant les 5 dernières secondes de question et le début du
rappel lors de la tâche autobiographique. Les ERSP ne montrent pas de diminution de
puissance dans les bandes thêta et alpha lors du calcul. Toutefois ces modifications du signal
n’apparaissent pas significatives après transformée de Hilbert et application d’un test des
rangs signés de Wilcoxon (risque alpha à 0.05, correction par FDR). En revanche il existe lors
du rappel et de la question des différences significatives dans la bande gamma entre les
conditions autobiographique/sémantiques et le calcul mental.

41
Figure 13 : activités électrophysiologiques au sein du CCP gauche lors du rappel en mémoire autobiographique, comparé
par rapport au temps de calcul et au temps de recherche en mémoire sémantique. L’illustration représente les analyses de la puissance du signal lors d’un temps de rappel pris entre 2 et 5 secondes après la fin de
la question. Cette puissance du signal est obtenue après transformée de Hilbert. Les conditions mémoire autobiographique (vert) et calcul
(orange), et les conditions mémoire sémantique (rouge) et mémoire autobiographiques (vert) sont comparées entre elles. Ces comparaisons
sont effectuées dans différentes bandes de fréquence (gamma 40-60 Hz, 60-80 Hz, 80-100 Hz ; thêta 4-8 Hz, alpha 8-12 Hz). Un test des
rangs signés de Wilcoxon est réalisé. Les différences de puissance du signal significative apparaissent sur l’axe des abscisses en rouge
(p<0.05). L’activité dans la bande gamma 40-100 Hz reste significativement plus importante pendant tout le temps de rappel
autobiographique par rapport à la tâche mathématique où il existe une hypoactivité gamma persistante. Bien qu’un pattern d’aspect
anticorrelé appraissent sur les ERSP entre le calcul et la mémoire autobiographique, il n’y pas de différence dans les basses fréquences
thêta et alpha.

42
V. Spécificité de ce pattern électrophysiologique au CPP gauche
Afin de prouver que cette augmentation de puissance gamma pendant la bascule
attentionnelle lors de la tâche autobiographique était spécifique au CCP, nous avons analysé
les patterns électrophysiologiques des régions proches, comme le cortex pariétal externe
(analysé par les plots externes de la même électrode) et des régions plus distantes.
L’augmentation significative de la puissance dans la bande gamma lors de la bascule
semble spécifique au CPP puisqu’aucune différence significative n’apparait dans le cortex
pariétal externe (figure 14). Il n’y a pas de différence significative de puissance du signal
entre la tâche autobiographique et le calcul mental au sein du cortex pariétal externe. Il n’a
également pas été retrouvé de différence significative dans les autres gammes de fréquence
entre les trois conditions.
Figure 14 : Activités électrophysiologiques au sein du cortex pariétal externe gauche.
Aucune hyperactivité gamma n’est observée lors de la bascule attentionnelle lors du rappel autobiographique
sur ces cartes ERSP (moyenne pondérée des résultats des patients FS + CG + CT).
De plus les patterns sont différents par rapport au CCP gauche pour les trois tâches. Ces résultats prouvent une
validité de notre méthode d’analyse du signal : des régions cérébrales différentes connues pour des fonctions
différentes (cortex pariétal externe et CPP) affichent des activités différentes. De plus ils prouvent une spécificité
de cette hyperactivité gamma lors de la bascule attentionnelle au sein du CCP.
VI. Activités électrophysiologiques des autres régions cérébrales : exemple du lobe
temporal externe et de l’hippocampe
Les autres régions cérébrales affichent une dynamique des oscillations différentes du
CCP. Une large zone temporale externe montre une augmentation de puissance dans la bande
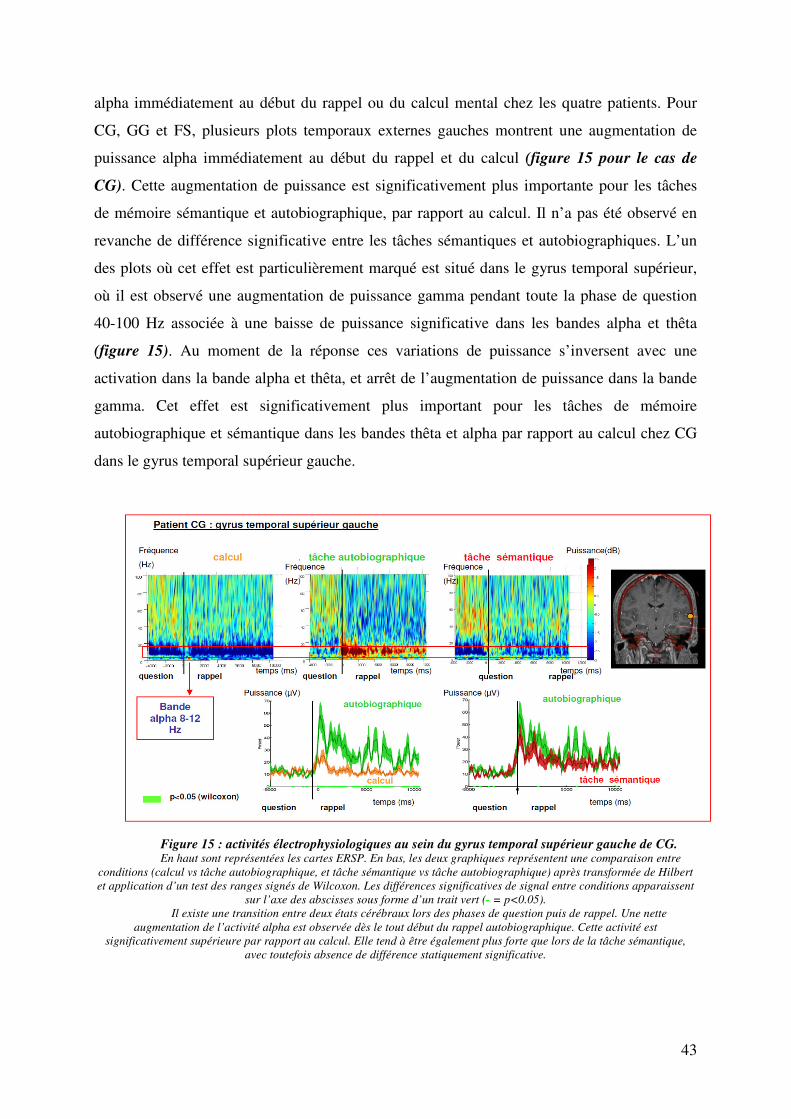
43
alpha immédiatement au début du rappel ou du calcul mental chez les quatre patients. Pour
CG, GG et FS, plusieurs plots temporaux externes gauches montrent une augmentation de
puissance alpha immédiatement au début du rappel et du calcul (figure 15 pour le cas de
CG). Cette augmentation de puissance est significativement plus importante pour les tâches
de mémoire sémantique et autobiographique, par rapport au calcul. Il n’a pas été observé en
revanche de différence significative entre les tâches sémantiques et autobiographiques. L’un
des plots où cet effet est particulièrement marqué est situé dans le gyrus temporal supérieur,
où il est observé une augmentation de puissance gamma pendant toute la phase de question
40-100 Hz associée à une baisse de puissance significative dans les bandes alpha et thêta
(figure 15). Au moment de la réponse ces variations de puissance s’inversent avec une
activation dans la bande alpha et thêta, et arrêt de l’augmentation de puissance dans la bande
gamma. Cet effet est significativement plus important pour les tâches de mémoire
autobiographique et sémantique dans les bandes thêta et alpha par rapport au calcul chez CG
dans le gyrus temporal supérieur gauche.
Figure 15 : activités électrophysiologiques au sein du gyrus temporal supérieur gauche de CG. En haut sont représentées les cartes ERSP. En bas, les deux graphiques représentent une comparaison entre
conditions (calcul vs tâche autobiographique, et tâche sémantique vs tâche autobiographique) après transformée de Hilbert
et application d’un test des ranges signés de Wilcoxon. Les différences significatives de signal entre conditions apparaissent
sur l’axe des abscisses sous forme d’un trait vert (- = p<0.05).
Il existe une transition entre deux états cérébraux lors des phases de question puis de rappel. Une nette
augmentation de l’activité alpha est observée dès le tout début du rappel autobiographique. Cette activité est
significativement supérieure par rapport au calcul. Elle tend à être également plus forte que lors de la tâche sémantique,
avec toutefois absence de différence statiquement significative.
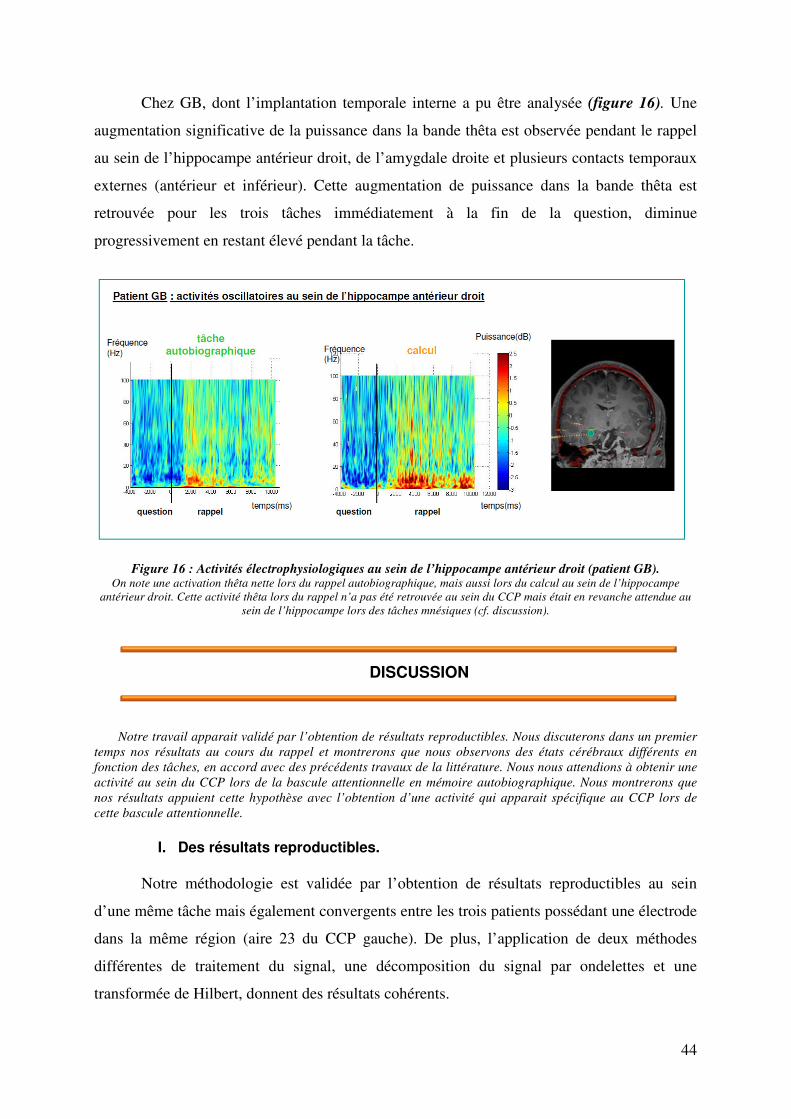
44
Chez GB, dont l’implantation temporale interne a pu être analysée (figure 16). Une
augmentation significative de la puissance dans la bande thêta est observée pendant le rappel
au sein de l’hippocampe antérieur droit, de l’amygdale droite et plusieurs contacts temporaux
externes (antérieur et inférieur). Cette augmentation de puissance dans la bande thêta est
retrouvée pour les trois tâches immédiatement à la fin de la question, diminue
progressivement en restant élevé pendant la tâche.
Figure 16 : Activités électrophysiologiques au sein de l’hippocampe antérieur droit (patient GB). On note une activation thêta nette lors du rappel autobiographique, mais aussi lors du calcul au sein de l’hippocampe
antérieur droit. Cette activité thêta lors du rappel n’a pas été retrouvée au sein du CCP mais était en revanche attendue au
sein de l’hippocampe lors des tâches mnésiques (cf. discussion).
DISCUSSION
Notre travail apparait validé par l’obtention de résultats reproductibles. Nous discuterons dans un premier
temps nos résultats au cours du rappel et montrerons que nous observons des états cérébraux différents en
fonction des tâches, en accord avec des précédents travaux de la littérature. Nous nous attendions à obtenir une
activité au sein du CCP lors de la bascule attentionnelle en mémoire autobiographique. Nous montrerons que
nos résultats appuient cette hypothèse avec l’obtention d’une activité qui apparait spécifique au CCP lors de
cette bascule attentionnelle.
I. Des résultats reproductibles.
Notre méthodologie est validée par l’obtention de résultats reproductibles au sein
d’une même tâche mais également convergents entre les trois patients possédant une électrode
dans la même région (aire 23 du CCP gauche). De plus, l’application de deux méthodes
différentes de traitement du signal, une décomposition du signal par ondelettes et une
transformée de Hilbert, donnent des résultats cohérents.

45
II. Activité électrophysiologique pendant le rappel autobiographique
Nous observons les principales modifications significatives du signal dans le CCP
gauche au sein de la bande de fréquence gamma. Plusieurs études ont démontré que l’activité
dans une large bande gamma, induite par une tâche, était le reflet d’une augmentation de
l’activité neuronale locale (Lachaux et al. 2005, Jensen et al. 2007, Manning et al. 2009).
Tout d’abord, nos résultats vont dans le sens des nombreux articles sur le DMN en
montrant qu’au sein du CCP, l’activité neuronale est significativement plus élevée pendant les
tâches de repos et les tâches d’introspection, que lors de tâches avec une demande
attentionnelle forte (comme le calcul mental) qui montrent une désactivation. Une
désactivation lors du calcul a en effet déjà été observée dans une étude en IRMf où l’activité
BOLD était plus élevée pendant les tâches de repos et de référence au self dans le cortex
postéro-médial (Kennedy et al. 2008). Une anti-corrélation entre ces deux réseaux, cohérente
avec les données en IRMf (Fox et al. 2005), a déjà été montrée sur des analyses
électrophysiologiques intracrâniennes. Ossandon et al. (2011) confirmaient la présence de
deux réseaux anti-corrélés d’augmentation (repos) ou de décroissance (durant une tâche
attentionnelle visuelle) d’activité dans une large bande gamma.
Une étude par électrodes subdurales dans le cortex postéromédial (incluant CPP et
CRS) chez 4 patients épileptiques, a ajouté à l’étude du calcul et du repos, une tâche dite
autobiographique (Dastjerdi et al. 2011). Les sujets devaient lire des phrases du type « j’ai
travaillé sur un ordinateur aujourd’hui » et répondre vrai ou faux. Les auteurs retrouvent un
antagonisme entre une hyperactivation gamma lors de cette tâche « autobiographique » (chez
2 de leurs patients seulement pour 2 électrodes proximales par rapport au CRS) et une
hypoactivation gamma lors du calcul. Nous obtenons le même antagonisme dans notre étude
entre notre condition autobiographique et notre condition calcul.
Toutefois, si on peut admettre que la tâche de Dastjerdi et al.(2011) fait référence au
self de l’individu, ils n’ont pas étudié le processus de rappel en mémoire autobiographique ou
même apporté des preuves de réminiscence d’un souvenir par les sujets. Cette tâche n’était
pas comparée à une autre tâche de mémoire déclarative à long terme, comme la mémoire
sémantique. Peu d’éléments sont donc apportés sur le rôle du CCP au sein même du réseau de
la mémoire.

46
III. Activité électrophysiologique pendant la bascule attentionnelle
a. Une augmentation significative d’activité au sein d’une large bande gamma
Nous avions émis l’hypothèse d’un rôle du CCP pendant la bascule attentionnelle lors
du rappel en mémoire autobiographique. Or dans ce travail, nous observons les principales
modifications significatives du signal dans le CCP gauche dans la bande de fréquence gamma
lors de la tâche autobiographique pendant cette bascule. Notre étude apporte donc la
précision que le rappel en mémoire autobiographique s’accompagne d’une hyperactivation
significative gamma dans la bande 40-100 Hz au moment de la bascule question/rappel au
sein du CCP. L’activité électrophysiologique affiche par ailleurs un pattern différent de la
tâche de calcul dès le stade de la question. Cet état initial ne diffère pas de l’état de repos. Il
semble donc que les sujets soient dans un « état préparatoire », avant même le rappel
autobiographique, qui s’apparente au DMN. Mais le rappel autobiographique semble
surimposer une activité gamma au moment de la bascule question/rappel sur « cet état
électrophysiologique préparatoire ».
b. Profil temporel et spécificité fonctionnelle de l’augmentation d’activité dans une
large bande gamma.
La modification la plus significative du signal au sein de la large bande gamma
apparaît au moment de la bascule question/rappel, environ 500 millisecondes avant le début
du rappel avec une activité gamma plus importante que pour le reste du temps de question et
de recherche du souvenir. Notons que notre ligne de base est équivalente à la tache de repos
de l’étude de Dastjerdi et al. Il est difficile d’être plus précis sur l’intervalle temporel, le
trigger de fin de question est déclenché manuellement par l’examinateur, et le sujet a très
probablement déjà débuté sa recherche du souvenir avant le tout dernier mot de la question.
Dastjerdi et al. (2011) ont montré une hyperactivité gamma au sein du cortex
postéromédial lors d’une tâche de repos. Mais l’un des éléments les plus intéressants est la
présence d’un pic d’activité gamma apparaissant avec une très courte latence après la
présentation de l’indice annonçant le début de cette tâche. Ce pic se répète d’une façon
stéréotypée entre les différents essais et les différentes électrodes répondant au repos. Ils font
donc l’hypothèse que plutôt qu’être associée à un contenu cognitif, cette activité gamma
pourrait être le signal d’un désengagement attentionnel.
Dans notre travail l’activité gamma ne diffère pas significativement de l’état de repos
(ou ligne de base) au cours du rappel suivant cette phase de bascule attentionnelle. On peut
donc également émettre l’hypothèse que ce pic d’activité gamma apparaissant uniquement

47
lors de la bascule n’est probablement pas à relier à un contenu cognitif comme la
manipulation de représentations mnésiques. Mais il pourrait correspondre à une forme de
signal d’engagement dans un processus, ici le rappel autobiographique, vis à vis d’autres
régions cérébrales. Ce caractère très transitoire de l’augmentation d’activité gamma
illustrerait la nature dynamique du réseau pour faciliter une bascule rapide entre différents
états attentionnels. Ce caractère hautement transitoire des modulations dans la bande gamma a
déjà été souligné en EEG intracrânien au sein du DMN (Ossandon et al. 2011).
L’idée est compatible avec les conclusions avancées par l’étude de Fornito et al.
(2012) en IRM fonctionnelle. Une opposition binaire permanente entre le DMN et ce vaste
réseau attentionnel externe (EAS) vis à vis des stimuli environnementaux apparaît aujourd’hui
trop simpliste. On l’a vu précédemment, le DMN et l’EAS ne fonctionnent pas comme des
entités homogènes mais se décomposent en subrégions distinctes qui s’activent en fonction de
la tâche (Vogt et al. 2005, Leech et al. 2012 dans le cas du CCP). Fornito et al. (2012)
montrent ainsi qu’un antagonisme important entre l’activité du DMN et l’EAS n’est pas
toujours associé avec une meilleure performance et qu’il existe une coopération entre les deux
réseaux pour la réalisation de certaines tâches. Ce passage d’un antagonisme à une
coopération entre les deux systèmes deviendrait possible grâce à une ségrégation
fonctionnelle du DMN et EAS en plusieurs sous régions, avec par exemple au sein de l’EAS :
le réseau cingulo-operculaire, le réseau fronto-pariétal gauche et le réseau fronto-pariétal
droit. Ainsi certaines tâches provoqueraient le passage d’un état intrinsèque d’antagonisme
entre les deux systèmes à un état d’interaction coopérative entre certaines de leurs sous
régions pour améliorer les performances. Le CCP droit serait une sous-région pivot dans la
recollection. Il permettrait cette bascule entre un état antagoniste de base (DMN vs EAS), à
une intégration fonctionnelle entre une sous-région de l’EAS, le réseau frontopariétal droit
(pour les processus de contrôle et d’évaluation du rappel) et le DMN (pour le stockage et
recollection des associations contextuelles). Cette bascule aboutirait à une recollection plus
rapide.
Le CCP participerait donc à la réorganisation des interactions à grande échelle entre
ces deux grands systèmes en fonction de la tâche. Ce rôle de régulateur cognitif du CCP a
aussi été proposé par Leech (2012) avec un CCP dorsal qui pourrait réguler une sorte de
balance entre la cognition dirigée vers le monde interne (introspection) et celle dirigée vers le
monde externe (environnement).

48
Toutefois il reste certains points à approfondir nous incitant à poursuivre nos travaux.
Tout d’abord nous observons ce pic d’activité gamma à gauche lors de la bascule entre
question et rappel, là où l’équipe de Fortino attribue un rôle de régulateur de la recollection au
CCP droit. Leur tâche épisodique n’était toutefois pas autobiographique et il s’agit de données
d’IRMf avec une moins bonne résolution spatiale. Nous avons également insuffisamment de
données au sein du CCP droit.
c. Absence de différence significative entre les tâches sémantique et
autobiographique
D’autre part, bien qu’une tendance à une plus forte augmentation gamma du signal
apparaisse en faveur de la mémoire autobiographique sur les ERSP et que l’activité gamma
haute n’était pas significative par rapport à la ligne de base au cours du rappel sémantique, les
différences n’apparaissent pas statistiquement significatives entre les tâches sémantique et
autobiographique. Ce qui au premier abord apparait contradictoire avec le fait que le CCP, et
notamment l’aire 23, appartient au réseau mis en jeu lors des activités cognitives en lien avec
le self d’un individu. Nous pouvons émettre différentes hypothèses. Est-ce lié à un nombre
trop faible d’essais pour suffisamment de puissance statistique? Est-ce lié à une analyse
restreinte de cette large bande gamma à 40-100 Hz en raison d’une limitation par notre
matériel d’échantillonnage? Les effets observés dans l’étude de Dastjerdi et al. (2011)
apparaissent en effet au sein d’une bande de 30-180 Hz. Il en est de même dans l’étude
d’Ossandon et al. (2011) où les modulations significatives de l’activité gamma étaient
observées au sein d’une bande gamma haute 40-160 Hz. Une autre hypothèse est que notre
tâche sémantique n’est pas adaptée. C’est en tout cas la plus difficile à réaliser par les
patients, celle qui donne le moins d’essais réussis. Peut-être utilisent-ils des moyens de
recollection faisant intervenir leur self pour trouver les bonnes réponses. S’assurer au moment
du débriefing que le patient ne s’est pas aidé de souvenirs personnels permettrait de ne garder
que les essais sans lien avec le self du sujet. Il faudrait donc augmenter le nombre d’essais.
L’autre hypothèse est celle proposée par Baddley en 1984 et récemment montrée par
Buraniova et al. (2010) en IRMf : l’existence d’un large réseau commun au rappel en
mémoire déclarative, à savoir l’activation de régions communes pour les mémoires
sémantique, épisodique et autobiographique. Les gyri cingulaires antérieur et postérieur
appartiendraient à ce large réseau nécessaire à la recherche en mémoire déclarative.
Cependant il apparaît important de revoir notre tâche sémantique avec de nouveaux
essais avant de s’orienter vers cette dernière hypothèse.

49
IV. Un pattern d’activité électrophysiologique spécifique au CPP
Les électrodes hors du cortex cingulaire postérieur affichent un pattern d’activité
électrophysiologique différent. C’est en particulier le cas au niveau du cortex pariétal latéral
où il n’est plus observé de désactivation gamma lors du calcul ni de pic d’activité gamma
pendant la bascule pendant la tâche autobiographique.
Sur quatre patients, plusieurs électrodes temporales externes affichent un pic d’activité
alpha et thêta immédiatement au début du rappel et de la tâche de calcul qui décroit
progressivement au cours de la recherche mentale. Pour trois patients dont l’implantation était
gauche, le pic d’activité alpha est significativement plus important pour les tâches de mémoire
que pour la tâche mathématique. Pour GB, dont l’implantation était droite, cette augmentation
de l’activité alpha ne diffère pas entre les conditions mnésiques et de calcul. Peu d’études se
sont focalisées sur les relations entre mémoire et ondes alpha, que l’on sait impliquées dans
les synchronisations longue distance et les processus top-down (Von Stein et al. 2000).
Klimesh et al. (2007, 2010) proposent qu’une augmentation de puissance alpha reflète en
réalité une inhibition. Il s’agirait d’une « inhibition active » et d’un processus sélectif requis
pour inhiber et rendre silencieuses des activités neuronales inutiles à la tâche. Une
augmentation de puissance alpha était également observée lors de la transition entre deux
états attentionnels différents, lors du passage d’un état du sujet dit « perdu dans ses pensées »
à un état où le sujet était concentré sur sa respiration, dans l’étude de Braboszcz et Delorme
(2011). Cette augmentation d’activité alpha pourrait donc être un marqueur de bascule
attentionnelle entre deux états requérant un niveau de concentration différent. Ainsi,
Angelakis et al. (2004) suggéraient que l’augmentation d’activité alpha reflétait un état de
« préparation cognitive ». Par exemple, on peut émettre l’hypothèse que cette augmentation
d’activité alpha sur l’électrode du gyrus temporal supérieur de CG correspond à une inhibition
des zones auditives (actives durant la question) afin de permettre au sujet de se concentrer sur
son « monde interne » pour le rappel et d’être moins gêné par l’environnement.
V. Une activation gamma mais sans activation thêta dans la BA 23 gauche
Le caractère transitoire de l’activation gamma que nous observons se distingue du
pattern de l’activité gamma retrouvée par l’étude de Steinvorth et al. (2009) lors du rappel
autobiographique, au sein d’une autre région cérébrale, le cortex entorhinal. Dans leur étude,
une augmentation prononcée de l’activité gamma apparaissait dans les couches à projection

50
hippocampique environ 200 ms après la présentation de l’indice pour le rappel du souvenir et
se poursuivait environ 12 secondes. Ils émettent l’hypothèse que cette augmentation de
puissance gamma reflèterait les processus permettant d’obtenir la correspondance entre la
phrase indice et les représentations de la mémoire à long terme, pour que celles-ci soient
réexploitées pour la reminiscence. Nous ne pouvons attribuer le même rôle à cette
augmentation de puissance gamma au sein du CCP, puisqu’elle ne dure que le temps de la
transition question-rappel.
Nous nous attendions à retrouver une augmentation d’activité dans la bande thêta lors
du rappel mnésique au sein du CCP, qui aurait pu illustrer des liens dynamiques du CCP avec
les cortex associatifs et le lobe temporal médial. Nous l’avons exposé plus haut : le rôle des
activités thêtas est désormais bien caractérisé pour coordonner l’ensemble des activités
hippocampiques (Buzsaki 2002). Les activités thêtas permettent la synchronisation d’aires
cérébrales distantes pour des processus top down (Von Stein et al. 2000). Et l’association
d’une activation gamma et thêta a été démontrée par Steinvorth et al. (2009) dans le cortex
entorhinal lors du rappel autobiographique, avec un rôle différent attribué à chaque bande de
fréquence. Nous retrouvons d’ailleurs de façon cohérente une augmentation de puissance dans
la bande thêta 4-8 Hz pendant le rappel autobiographique au sein de l’hippocampe et de
l’amygdale droits de GB.
L’étude de Foster et al. (2012) en EEG intracrânien avec électrodes subdurales dans le
cortex postéromédial révèle que l’activité au sein de cette région prédomine dans la bande
thêta aux alentours de 4-5 Hz lors d’une tâche de repos spontanée (cette tâche de repos diffère
de l’étude de Dastjerdi dans laquelle les patients fixaient une croix pendant 5 secondes). Cette
activité thêta modulerait l’activité de la bande gamma par un phénomène de couplage phase-
amplitude. Elle serait même la signature électrophysiologique de la région par rapport aux
régions adjacentes (puisqu’une augmentation de l’activité alpha est prédominante au sein des
régions occipitales proches du CCP par exemple).
Mais nous n’observons pas d’augmentation d’activité thêta dans les zones très proches
ou au sein de l’aire 23. Certes, Foster et al. n’ont pas étudié la mémoire autobiographique. Ils
se sont focalisés sur un état relatif à l’activation du DMN dont on a vu précédemment la
superposition avec le réseau de la mémoire autobiographique. Mais ils ont enregistré
l’ensemble d’une aire postéro-médiale sans tenir compte des subdivisions fonctionnelles de
cette région. Nous revenons donc à notre hypothèse que ce pic d’activité gamma apparaissant
uniquement lors de la bascule n’est pas à relier à un contenu cognitif comme la manipulation

51
de représentations mnésiques. Mais il correspondrait à une forme de signal d’engagement
dans le processus de rappel autobiographique, vis à vis d’autres régions cérébrales. Il est donc
possible qu’une activité thêta soit prédominante dans d’autres subrégions qui interagissent
directement avec le lobe temporal médial, comme le CRS (Andrews-Hanna et al. 2010, leur
modèle est cité plus haut).
VI. Des résultats compatibles avec une hétérogénéité fonctionnelle du CCP ?
Les travaux cités précédemment (Vogt et al. 2005, Dastjerbi et al. 2011, Leech et al.
2012) ont montré la ségrégation fonctionnelle et anatomique de plusieurs sous régions du
CCP. En faisant une analyse sur l’ensemble d’une large région postéro-médiale, Dastjerdi et
al. (2011) ne retrouvaient pas de différence significative d’activité gamma entre la tâche
autobiographique et les autres conditions. En revanche en prenant les électrodes séparément,
pour 2 patients, 2 électrodes montraient une augmentation significative de l’activité gamma
pendant la tâche autobiographique. Ces dernières étaient proches du cortex rétrosplénial.
Donc des électrodes répondaient sélectivement aux stimuli autobiographiques, d’autres au
repos mais jamais aux deux conditions.
Nous manquons de données pour obtenir une véritable cartographie fonctionnelle du
CCP et étayer l’hypothèse que l’activité électrophysiologique lors du rappel autobiographique
doit être différente en fonction des sous régions du CCP. Nos résultats ne vont toutefois pas à
l’encontre de cette hétérogénéité fonctionnelle : nous observons le même pattern
d’hyperactivation gamma pour les trois patients dont les électrodes sont très proches de l’aire
23, dans le cortex cingulaire postérieur gauche. En analysant les coordonnées stéréotaxiques
du plot interne droit de GB, celui-ci apparaît en revanche plus bas que cette aire 23. Et aucune
hyperactivité gamma ni différence entre conditions au sein de cette large bande de fréquence
n’est enregistrée dans cette région.
Il apparaît également difficile de faire des extrapolations à une éventuelle latéralisation
hémisphérique des fonctions du CCP : nous avons trop peu de patients avec surtout une
hétérogénéité des groupes (3 implantations gauches pour une implantation droite) et les
coordonnées stéréotaxiques ne sont pas symétriques. De plus, plusieurs données de la
littérature sont contradictoires. Dans l’étude en IRM fonctionnelle de Maddock (2001), une
distinction est faite entre les deux hémisphères puisque c’est la partie caudale du cortex
cingulaire postérieur gauche qui était la région cérébrale montrant l’activation la plus
importante et la plus constante lors du rappel réussi de souvenirs autobiographiques entre les
sujets. Tandis que Fornito et al. (2012) attribuent un rôle pivot au cortex cingulaire postérieur

52
droit dans la bascule d’un état intrinsèque correspondant au DMN à un état correspondant à la
recollection. Mais il pourrait être intéressant de s’intéresser à cette possible différence
interhémisphérique sur le plan électrophysiologique en multipliant les données sur d’autres
sujets à l’avenir.
VII. Une ligne de base valide ?
On peut discuter le choix de notre ligne de base. Un des débats récurrents dans les
recherches sur le DMN en imagerie fonctionnelle a longtemps été la meilleure définition de la
ligne de base (Buckner et al. 2008). Richard Frackowiak en 1991 disait “to call a free
wheeling state or even a state where you are fixating on a cross and dreaming about anything
you like, a control state is too my mind quite wrong“.
Nous voulions un état neutre n’obligeant pas à l’introspection ni à une attention
soutenue vis à vis de stimuli externe. Nous avons choisi une ligne de base qui s’est avérée a
posteriori être l’équivalent d’une tâche considérée comme du repos par d’autres études
cherchant à étudier le resting-state (Dastjerdi et al. 2011). Cette condition est discutable dans
notre cas comme dans l’étude de Dastjerdi : dans les deux cas nous cherchons à comparer une
activité autobiographique à un état où il est difficile de contrôler les pensées du sujet qui peut
malgré tout se laisser aller à une activité introspective. Mais l’obtention de résultats
reproductibles entre sujets et de données significativement différentes entre conditions
apportent une validité à ce choix méthodologique.
Ce choix apparaît acceptable quand on considère les résultats et hypothèses de l’étude
de l’étude de Fornito et al. (2012). Bien que le DMN et le réseau de la mémoire
autobiographique soient superposés, c’est une réorganisation des interactions entre sous-
modules du DMN et de l’EAS qui favorise une recollection rapide. Notre choix
méthodologique souligne les modifications du signal propres à la tâche autobiographique par
rapport au fonctionnement intrinsèque du DMN. Cette limite pourra de plus être contournée à
l’avenir en utilisant différentes lignes de base (comme par exemple une moyenne de
l’ensemble des activités sur l’ensemble des conditions) puis en comparant les données.
VIII. Limites de l’étude
Nos analyses ont été effectuées sur un faible nombre de patients et d’essais limitant la
puissance de nos statistiques. Mais les données en EEG intracrânien in vivo sont des données
rares voire exceptionnelles et la plupart des publications se font sur peu de patients
(Steinvorth et al. 2009 : 1 patient, Dastjerdi et al. 2011 : 4 patients etc.). Un rapport

53
signal/bruit élevé dans cette technique intracérébrale autorise des analyses sur un faible
nombre d’essais. Braboszcz et Delorme (2011) ont réussi à démontrer des états
électrophysiologiques différents en fonction du niveau attentionnel du sujet en analysant
seulement 13 à 52 époques par sujets en EEG de surface, pourtant plus artéfacté que l’EEG
intracrânien.
Notre tâche comportementale ne peut aboutir à plus d’une trentaine d’essais réussis
autobiographiques en raison de la fatigabilité des sujets et des conditions de recueil des
données en milieu hospitalier. Valider un souvenir sur le plan de l’épisodicité nécessite que le
rappel soit allé à son terme pour obtenir suffisamment de données au débriefing. Nous
souhaitons étudier la dynamique d’un rappel génératif et ses contraintes attentionnelles. Ce
qui n’a pas été fait dans l’étude de Steinvorth (2009) où, bien que le design expérimental
s’approche du nôtre, le rappel ne durait que 6 secondes par patients pour des indices sur
lesquels avait déjà été interrogé préalablement le sujet. Leur étude explorait donc surtout la
réminiscence du souvenir.
Enfin étudier le rappel autobiographique sans étudier les interactions entre différentes
régions notamment avec le lobe temporal interne, et la dynamique des oscillations sur de
larges échelles apparaît insuffisant. Nous avons pour objectif de poursuivre nos analyses et
d’y apporter d’autres méthodes comme du couplage de phase entre électrodes de régions
différentes ou du couplage phase amplitude entre différentes gammes fréquentielles au sein
d’une même région. Le couplage phase amplitude pourra en effet être intéressant pour étudier
cette hypoactivité thêta et alpha observée lors de la bascule attentionnelle, qui n’existe pas
lors du calcul. Bien qu’elle n’apparaisse pas significative par rapport au calcul (après
transformée de Hilbert et application d’un test de Wilcoxon), il sera intéressant de rechercher
d’éventuels liens avec l’activité gamma lors de la bascule.
Dans cette perspective, des données ont été enregistrées dans de multiples autres
régions cérébrales chez nos différents sujets, afin d’approfondir les interactions dynamiques
du CCP avec d’autres régions cérébrales.

54
CONCLUSION
Notre étude montre une augmentation significative d’activité gamma pendant la bascule
question/rappel lors d’une tâche autobiographique au sein du CCP, région impliquée dans le réseau par
défaut. Absente lors d’une tâche de calcul, nous confirmons un rôle du CCP dans la bascule
attentionnelle lors du rappel en mémoire autobiographique. Cette activité gamma n’apparaît que le
temps de cette bascule entre deux états et se surimpose à un pattern électrophysiologique présent avant
le début du rappel, dont les caractéristiques (opposées à une tâche demandant des efforts attentionnels
comme le calcul) sont compatibles avec le DMN. Ce pattern apparaît différent des activités gamma
observés dans d’autres régions du réseau de la mémoire autobiographique : il n’est pas prolongé
comme dans le cortex entorhinal (Steinvorth et al. 2009) et n’est pas associé à une activité thêta
prolongée. Survenant au moment de la transition entre deux états différents avec un caractère très
transitoire, ce pic d’activité gamma n’est probablement pas à relier à un contenu cognitif comme la
manipulation de représentations mnésiques. Il pourrait correspondre à une forme de signal
d’engagement dans un processus, ici le rappel autobiographique, vis à vis d’autres régions cérébrales.
Cette hypothèse est compatible avec une étude récente qui a montré une reconfiguration dynamique et
transitionnelle de l’architecture fonctionnelle du DMN pour faciliter la recollection, faisant du CCP
droit un nœud central dans la commande de cette reconfiguration (Fornito et al. 2012).
Cette étude reste un travail préliminaire. Le CCP est une région faite de multiples sous
régions. Nous devons améliorer notre tâche contrôle sémantique et acquérir une meilleure appréciation
des liens avec le self. D’autres données doivent être obtenues, notamment au sein du CCP droit et dans
d’autres sous régions du CCP pour une meilleure cartographie de la région, car nos principaux
résultats sont dans l’hémisphère gauche. Des données ont été enregistrées dans plusieurs autres aires
cérébrales chez nos différents sujets au cours de cette année, afin de pouvoir analyser les interactions
dynamiques du CCP avec d’autres régions cérébrales.
Ce travail en électrophysiologie s’inscrit dans le cadre d’un projet global cherchant à
caractériser cette bascule attentionnelle permettant le rappel autobiographique, y compris sur un plan
comportemental. Les sujets ont tendance à dévier leur regard immédiatement au début de la recherche
du souvenir. Dans l’hypothèse que cette bascule du regard puisse être un marqueur comportemental de
la bascule attentionnelle pour le rappel d’épisodes en mémoire autobiographique, nous souhaitons
appliquer notre tâche chez des sujets sains avec oculométrie et EEG de surface. Un script a été mis au
point au cours de cette année de master 2 pour les paramétrages de l’eye-tracker. Des sujets pilotes ont
déjà pu passer la tâche.

55
REFERENCES BIBLIOGRAPHIQUES
Addis DR, Moscovitch M, McAndrews MP.Consequences of hippocampal damage across the autobiographical memory network in left temporal lobe epilepsy. Brain. 2007 Sep; 130(Pt 9):2327-42.
Andreasen NC, O'Leary DS, Cizadlo T, Arndt S, Rezai K, Watkins GL, Ponto LL, Hichwa RD.Remembering the past: two facets of
episodic memory explored with positron emission tomography.Am J Psychiatry. 1995 Nov;152(11):1576-85.
Andrews-Hanna JR, Reidler JS, Sepulcre J, Poulin R, Buckner RL. Functional-anatomic fractionation of the brain's default network.
Neuron. 2010 Feb 25;65(4):550-62.
Angelakis E, Lubar JF, Stathopoulou S, Kounios J. Peak alpha frequency: an electroencephalographic measure of cognitive
preparedness. Clin Neurophysiol. 2004 Apr;115(4):887-97.
Barbeau E, Wendling F, Régis J, Duncan R, Poncet M, Chauvel P, Bartolomei F. Recollection of vivid memories after perirhinal region
stimulations: synchronization in the theta range of spatially distributed brain areas.Neuropsychologia. 2005;43(9):1329-37. Epub 2005
Jan 16.
Bastin C, Feyers D, Souchay C, Guillaume B, Pepin JL, Lemaire C, Degueldre C, Collette F, Salmon E. Frontal and posterior cingulate
metabolic impairment in the behavioral variant of frontotemporal dementia with impaired autonoetic consciousness.Hum Brain
Mapp. 2012 Jun;33(6):1268-78. doi: 10.1002/hbm.21282.
Braboszcz C, Delorme A. Lost in thoughts: neural markers of low alertness during mind wandering. Neuroimage. 2011 Feb
14;54(4):3040-7..
Brewer WF, Pani JR. Reports of mental imagery in retrieval from long-term memory. Conscious Cogn. 1996 Sep;5(3):265-87.
Buckner RL, Andrews-Hanna JR, Schacter DL.The brain's default network: anatomy, function, and relevance to disease. Ann N Y Acad
Sci. 2008 Mar;1124:1-38. Review.
Burianova H, McIntosh AR, Grady CL. A common functional brain network for autobiographical, episodic, and semantic memory
retrieval.Neuroimage. 2010 Jan 1;49(1):865-74.
Butler CR, Graham KS, Hodges JR, Kapur N, Wardlaw JM, Zeman AZ.The syndrome of transient epileptic amnesia. Ann Neurol. 2007
Jun;61(6):587-98.
Buzsáki G. Theta oscillations in the hippocampus.Neuron. 2002 Jan 31;33(3):325-40. Review.
Cabeza R, Nyberg L. Neural bases of learning and memory: functional neuroimaging evidence.Curr Opin Neurol. 2000 Aug;13(4):415-
21. Review.
Cabeza R, St Jacques P. Functional neuroimaging of autobiographical memory.Trends Cogn Sci. 2007 May;11(5):219-27. Epub 2007
Mar 26.
Chetelat G, Desgranges B, de la Sayette V, Viader F, Berkouk K, Landeau B, Lalevée C, Le Doze F, Dupuy B, Hannequin D, Baron JC,
Eustache F.Dissociating atrophy and hypometabolism impact on episodic memory in mild cognitive impairment. Brain. 2003
Sep;126(Pt 9):1955-67.
Choo IH, Lee DY, Oh JS, Lee JS, Lee DS, Song IC, Youn JC, Kim SG, Kim KW, Jhoo JH, Woo JI.Posterior cingulate cortex atrophy and
regional cingulum disruption in mild cognitive impairment and Alzheimer's disease. Neurobiol Aging. 2010 May;31(5):772-9.
Conway MA, Pleydell-Pearce CW, Whitecross SE, Sharpe H. Neurophysiological correlates of memory for experienced and imagined
events.Neuropsychologia. 2003;41(3):334-40.
Conway MA, Pleydell-Pearce CW. The construction of autobiographical memories in the self-memory system. Psychol Rev. 2000
Apr;107(2):261-88. Review
Conway MA. Sensory-perceptual episodic memory and its context: autobiographical memory.Philos Trans R Soc Lond B Biol Sci.
2001 Sep 29;356(1413):1375-84. Review.
Dastjerdi M, Foster BL, Nasrullah S, Rauschecker AM, Dougherty RF, Townsend JD, Chang C, Greicius MD, Menon V, Kennedy DP,
Parvizi J. Differential electrophysiological response during rest, self-referential, and non-self-referential tasks in human posteromedial cortex.Proc Natl Acad Sci U S A. 2011 Feb 15;108(7):3023-8.
Delorme A, Makeig S. EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis.J Neurosci Methods. 2004 Mar 15;134(1):9-21.
Delorme A, Westerfield M, Makeig S. Medial prefrontal theta bursts precede rapid motor responses during visual selective attention. J Neurosci. 2007 Oct 31;27(44):11949-59.
Engel J Jr. Introduction to temporal lobe epilepsy. Epilepsy Res. 1996 Dec;26(1):141-50. Review.

56
Fell J, Axmacher N.The role of phase synchronization in memory processes. Nat Rev Neurosci. 2011 Feb;12(2):105-18. Review.
Fell J, Klaver P, Elfadil H, Schaller C, Elger CE, Fernández G. Rhinal-hippocampal theta coherence during declarative memory
formation: interaction with gamma synchronization?Eur J Neurosci. 2003 Mar;17(5):1082-8.
Fornito A, Harrison BJ, Zalesky A, Simons JS. Competitive and cooperative dynamics of large-scale brain functional networks
supporting recollection. Proc Natl Acad Sci U S A. 2012 Jul 17.
Foster BL, Parvizi J. Resting oscillations and cross-frequency coupling in the human posteromedial cortex. Neuroimage. 2012
Mar;60(1):384-91.
Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, Raichle ME. The human brain is intrinsically organized into dynamic,
anticorrelated functional networks. Proc Natl Acad Sci U S A. 2005 Jul 5;102(27):9673-8.
Fransson P. Spontaneous low-frequency BOLD signal fluctuations: an fMRI investigation of the resting-state default mode of brain
function hypothesis. Hum Brain Mapp. 2005 Sep;26(1):15-29.
Fries P.A mechanism for cognitive dynamics: neuronal communication through neuronal coherence.Trends Cogn Sci. 2005
Oct;9(10):474-80. Review.
Genovese CR, Lazar NA, Nichols T. Thresholding of statistical maps in functional neuroimaging using the false discovery
rate.Neuroimage. 2002 Apr;15(4):870-8.
Gevins A, Smith ME, McEvoy L, Yu D. High-resolution EEG mapping of cortical activation related to working memory: effects of
task difficulty, type of processing, and practice. Cereb Cortex. 1997 Jun;7(4):374-85.
Grandchamp R, Delorme A. Single-trial normalization for event-related spectral decomposition reduces sensitivity to noisy trials.
Front Psychol. 2011;2:236. Epub 2011 Sep 30.
Gruber T, Müller MM. Effects of picture repetition on induced gamma band responses, evoked potentials, and phase synchrony in the
human EEG.Brain Res Cogn Brain Res. 2002 May;13(3):377-92.
Gruber T, Müller MM. Oscillatory brain activity dissociates between associative stimulus content in a repetition priming task in the
human EEG.Cereb Cortex. 2005 Jan;15(1):109-16. Epub 2004 Jul 6.
Gruber T, Tsivilis D, Giabbiconi CM, Müller MM. Induced electroencephalogram oscillations during source memory: familiarity is
reflected in the gamma band, recollection in the theta band.J Cogn Neurosci. 2008 Jun;20(6):1043-53.
Guderian S, Düzel E. Induced theta oscillations mediate large-scale synchrony with mediotemporal areas during recollection in
humans.Hippocampus. 2005;15(7):901-12.
Guido Gainotti, Susanna Almonti, Anna Maria Di Betta & Maria Caterina Silveri. Retrograde amnesia in a patient with retrosplenial
tumour. Neurocase: The Neural Basis of Cognition Volume 4, Issue 6, 1998,pages 519-526
Irish M, Lawlor BA, O'Mara SM, Coen RF. Assessment of behavioural markers of autonoetic consciousness during episodic
autobiographical memory retrieval: a preliminary analysis. Behav Neurol. 2008;19(1-2):3-6.
Jensen O, Kaiser J, Lachaux JP. Human gamma-frequency oscillations associated with attention and memory. Trends Neurosci. 2007
Jul;30(7):317-24. Epub 2007 May 17.
Jensen O, Tesche CD. Frontal theta activity in humans increases with memory load in a working memory task. Eur J Neurosci. 2002
Apr;15(8):1395-9.
Jerbi K, Ossandón T, Hamamé CM, Senova S, Dalal SS, Jung J, Minotti L, Bertrand O, Berthoz A, Kahane P, Lachaux JP. Task-related
gamma-band dynamics from an intracerebral perspective: review and implications for surface EEG and MEG.Hum Brain Mapp.
2009 Jun;30(6):1758-71. Review.
Kahana MJ, Seelig D, Madsen JR. Theta returns.Curr Opin Neurobiol. 2001 Dec;11(6):739-44.
Kennedy DP, Courchesne E. Functional abnormalities of the default network during self- and other-reflection in autism.Soc Cogn
Affect Neurosci. 2008 Jun;3(2):177-90. Epub 2008 Apr 28.
Kim H. Dissociating the roles of the default-mode, dorsal, and ventral networks in episodic memory retrieval. Neuroimage. 2010 May
1;50(4):1648-57.
Kirk IJ, Mackay JC. The role of theta-range oscillations in synchronising and integrating activity in distributed mnemonic
networks.Cortex. 2003 Sep-Dec;39(4-5):993-1008. Review.
Klimesch W, Doppelmayr M, Yonelinas A, Kroll NE, Lazzara M, Röhm D, Gruber W.Theta synchronization during episodic retrieval:
neural correlates of conscious awareness.Brain Res Cogn Brain Res. 2001 Aug;12(1):33-8.
Klimesch W. EEG alpha and theta oscillations reflect cognitive and memory performance: a review and analysis.Brain Res Brain Res
Rev. 1999 Apr;29(2-3):169-95. Review.
Kopelman MD, Wilson BA, Baddeley AD. The autobiographical memory interview: a new assessment of autobiographical and personal semantic memory in amnesic patients.J Clin Exp Neuropsychol. 1989 Oct;11(5):724-44.

57
Kopelman MD, Wilson BA, Baddeley AD. The autobiographical memory interview: a new assessment of autobiographical and
personal semantic memory in amnesic patients. J Clin Exp Neuropsychol. 1989 Oct;11(5):724-44
Lachaux JP, George N, Tallon-Baudry C, Martinerie J, Hugueville L, Minotti L, Kahane P, Renault B.The many faces of the gamma band
response to complex visual stimuli.Neuroimage. 2005 Apr 1;25(2):491-501
Lachaux JP, Rudrauf D, Kahane P.Intracranial EEG and human brain mapping.J Physiol Paris. 2003 Jul-Nov;97(4-6):613-28. Review.
Lah S, Lee T, Grayson S, Miller L.Effects of temporal lobe epilepsy on retrograde memory. Epilepsia. 2006 Mar; 47(3):615-25.
Le Van Quyen M, Foucher J, Lachaux J, Rodriguez E, Lutz A, Martinerie J, Varela FJ.Comparison of Hilbert transform and wavelet methods for the analysis of neuronal synchrony.J Neurosci Methods. 2001 Oct 30. 111(2):83-98. Review.
Leech R, Braga R, Sharp DJ. Echoes of the brain within the posterior cingulate cortex.J Neurosci. 2012 Jan 4;32(1):215-22.
Maddock RJ, Garrett AS, Buonocore MH. Remembering familiar people: the posterior cingulate cortex and autobiographical memory
retrieval. Neuroscience. 2001;104(3):667-76
Maguire EA, Mummery CJ. Differential modulation of a common memory retrieval network revealed by positron emission
tomography. Hippocampus. 1999;9(1):54-61.
Makeig S, Debener S, Onton J, Delorme A. Mining event-related brain dynamics.Trends Cogn Sci. 2004 May;8(5):204-10. Review.
Makeig S. Auditory event-related dynamics of the EEG spectrum and effects of exposure to tones. Electroencephalogr Clin
Neurophysiol. 1993 Apr;86(4):283-93.
Manning JR, Jacobs J, Fried I, Kahana MJ. Broadband shifts in local field potential power spectra are correlated with single-neuron
spiking in humans. J Neurosci. 2009 Oct 28;29(43):13613-20.
Mevel K, Grassiot B, Chételat G, Defer G, Desgranges B, Eustache F. The default mode network: cognitive role and pathological
disturbances. Rev Neurol (Paris). 2010 Nov;166(11):859-72.
Michael Ross and Anne E. Wilson . Autobiographical Memory and Conceptions of Self: Getting Better All the Time .Current Directions
in Psychological Science April 2003 12: 66-69,
Minoshima S, Giordani B, Berent S, Frey KA, Foster NL, Kuhl DE. Metabolic reduction in the posterior cingulate cortex in very early
Alzheimer's disease. Ann Neurol. 1997 Jul;42(1):85-94.
Moscovitch M. & Winocur G. 2002. The frontal cortex and working with memory. In (Eds. Stuss D.T. & Knight R.T.), The Frontal Lobes
(pp. 188-209). Oxford: Oxford University Press.
Moscovitch, Morris; Winocur, Gordon Stuss, Donald T. (Ed); Knight, Robert T. (Ed), (2002). The frontal cortex and working with
memory. Principles of frontal lobe function. (pp. 188-209). New York, NY, US: Oxford University Press, xxi, 616 pp.
Northoff G, Bermpohl F.Cortical midline structures and the self.Trends Cogn Sci. 2004 Mar;8(3):102-7.
Noulhiane M, Piolino P, Hasboun D, Clemenceau S, Baulac M, Samson S.Autobiographical memory after temporal lobe resection:
neuropsychological and MRI volumetric findings. Brain. 2007 Dec; 130(Pt 12):3184-99.
Nyberg L, Habib R, McIntosh AR, Tulving E. Reactivation of encoding-related brain activity during memory retrieval
Proceedings of the National Academy of Sciences 97 (20), 98. 2000 Sep.
Onton J, Delorme A, Makeig S. Frontal midline EEG dynamics during working memory. Neuroimage. 2005 Aug 15;27(2):341-56.
Ossandón T, Jerbi K, Vidal JR, Bayle DJ, Henaff MA, Jung J, Minotti L, Bertrand O, Kahane P, Lachaux JP. Transient suppression of
broadband gamma power in the default-mode network is correlated with task complexity and subject performance.J Neurosci. 2011
Oct 12;31(41):14521-30.
Ossandón T, Vidal JR, Ciumas C, Jerbi K, Hamamé CM, Dalal SS, Bertrand O, Minotti L, Kahane P, Lachaux JP. Efficient "pop-out" visual search elicits sustained broadband γ activity in the dorsal attention network.J Neurosci. 2012 Mar 7;32(10):3414-21.
Palomero-Gallagher N, Vogt BA, Schleicher A, Mayberg HS, Zilles K.Receptor architecture of human cingulate cortex: evaluation of the four-region neurobiological model. Hum Brain Mapp. 2009 Aug;30(8):2336-55.
Piolino P, Desgranges B, Belliard S, Matuszewski V, Lalevée C, De la Sayette V, Eustache F. Autobiographical memory and autonoetic consciousness: triple dissociation in neurodegenerative diseases. Brain. 2003 Oct; 126(Pt 10):2203-19.
Piolino P, Desgranges B, Eustache F. Episodic autobiographical memories over the course of time: cognitive, neuropsychological and neuroimaging findings.Neuropsychologia. 2009 Sep;47(11):2314-29. Epub 2009 Jan 19. Review.
Piolino P.Autobiographical memory in aging. Psychol Neuropsychiatr Vieil. 2003 Mar;1(1):25-35. Review.
Polyn SM, Kahana MJ. Memory search and the neural representation of context. Trends Cogn Sci. 2008 Jan;12(1):24-30.
Polyn SM, Natu VS, Cohen JD, Norman KA. Category-specific cortical activity precedes retrieval during memory search. Science.
2005 Dec 23;310(5756):1963-6.

58
Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL.A default mode of brain function. Proc Natl Acad Sci U S
A. 2001 Jan 16;98(2):676-82.
Sajonz B, Kahnt T, Margulies DS, Park SQ, Wittmann A, Stoy M, Ströhle A, Heinz A, Northoff G, Bermpohl F. Delineating self-
referential processing from episodic memory retrieval: common and dissociable networks. Neuroimage. 2010 May 1;50(4):1606-17.
Sederberg PB, Kahana MJ, Howard MW, Donner EJ, Madsen JR. Theta and gamma oscillations during encoding predict subsequent
recall.J Neurosci. 2003 Nov 26;23(34):10809-14.
Sederberg PB, Schulze-Bonhage A, Madsen JR, Bromfield EB, McCarthy DC, Brandt A, Tully MS, Kahana MJ. Hippocampal and
neocortical gamma oscillations predict memory formation in humans.Cereb Cortex. 2007 May;17(5):1190-6.
Skinner EI, Fernandes MA. Neural correlates of recollection and familiarity: a review of neuroimaging and patient
data.Neuropsychologia. 2007 Jun 11;45(10):2163-79. Review.
Steinvorth S, Corkin S, Halgren E. Ecphory of autobiographical memories: an fMRI study of recent and remote memory
retrieval.Neuroimage. 2006 Mar;30(1):285-98.
Steinvorth S, Wang C, Ulbert I, Schomer D, Halgren E.Human entorhinal gamma and theta oscillations selective for remote autobiographical
memory.Hippocampus. 2010 Jan;20(1):166-73.
St-Laurent M, Moscovitch M, Levine B, McAndrews MP. Determinants of autobiographical memory in patients with unilateral
temporal lobe epilepsy or excisions. Neuropsychologia. 2009 Sep; 47(11):2211-21.
Summerfield JJ, Hassabis D, Maguire EA. Cortical midline involvement in autobiographical memory.Neuroimage. 2009 Feb
1;44(3):1188-200.
Svoboda E, McKinnon MC, Levine B. The functional neuroanatomy of autobiographical memory: a meta-analysis.Neuropsychologia.
2006;44(12):2189-208.
Tallon-Baudry C, Bertrand O, Delpuech C, Permier J.Oscillatory gamma-band (30-70 Hz) activity induced by a visual search task in
humans. J Neurosci. 1997 Jan 15;17(2):722-34.
Tallon-Baudry C, Bertrand O. Oscillatory gamma activity in humans and its role in object representation.Trends Cogn Sci. 1999
Apr;3(4):151-162.
Tang YY, Rothbart MK, Posner MI. Neural correlates of establishing, maintaining, and switching brain states.Trends Cogn Sci. 2012
Jun;16(6):330-7.
Tramoni E, Felician O, Barbeau EJ, Guedj E, Guye M, Bartolomei F, Ceccaldi M. Long-term consolidation of declarative memory: insight from temporal lobe epilepsy. Brain. 2011 Mar;134(Pt 3):816-31.
Tulving E, Schacter DL, McLachlan DR, Moscovitch M. Priming of semantic autobiographical knowledge: a case study of retrograde amnesia. Brain Cogn. 1988 Aug;8(1):3-20.
Tulving E. Episodic memory: from mind to brain.Annu Rev Psychol. 2002; 53:1-25
Tulving, E. (1985). How many memory systems are there? American Psychologist, 40, 385-3
Valenstein E, Bowers D, Verfaellie M, Heilman KM, Day A, Watson RT. Retrosplenial amnesia.Brain. 1987 Dec;110 ( Pt 6):1631-46.
Varela F, Lachaux JP, Rodriguez E, Martinerie J.The brainweb: phase synchronization and large-scale integration. Nat Rev Neurosci.
2001 Apr;2(4):229-39.
Viskontas IV, McAndrews MP, Moscovitch M. Remote episodic memory deficits in patients with unilatéral temporal lobe epilepsy and excisions.J Neurosci. 2000 Aug 1; 20(15):5853-7.
Vogt BA, Laureys S. Posterior cingulate, precuneal and retrosplenial cortices: cytology and components of the neural network correlates of consciousness.Prog Brain Res. 2005;150:205-17. Review.
Vogt BA, Nimchinsky EA, Vogt LJ, Hof PR.Human cingulate cortex: surface features, flat maps, and cytoarchitecture.J Comp Neurol.
1995 Aug 28;359(3):490-506.
Vogt BA, Vogt L, Laureys S. Cytology and functionally correlated circuits of human posterior cingulate areas.Neuroimage. 2006 Jan
15;29(2):452-66.
Voltzenlogel V, Després O, Vignal JP, Steinhoff BJ, Kehrli P, Manning L. Remote memory in temporal lobe epilepsy. Epilepsia. 2006
Aug;47(8):1329-36
Von Stein A, Sarnthein J. Different frequencies for different scales of cortical integration: from local gamma to long range alpha/theta synchronization. Int J Psychophysiol. 2000 Dec 1;38(3):301-13.
Wheeler ME, Petersen SE, Buckner RL. Memory's echo: vivid remembering reactivates sensory-specific cortex.Proc Natl Acad Sci U S
A. 2000 Sep 26;97(20):11125-9. Erratum in: Proc Natl Acad Sci U S A. 2004 Apr 6;101(14):5181.

59


















