
Lignée germinale et gamétogenèse -...
43
1 Lignée germinale et gamétogenèse I) Introduction 1) Sexualité et reproduction On va étudier les cellules germinales et la gamétogenèse, mais on n'abordera pas les aspects de sexualité, à savoir la différenciation des organes de la sexualité. La reproduction sexuée s'oppose à la reproduction asexuée, que l'on retrouve notamment chez les bactéries, et au cours de laquelle on a formation de clones ou de paquets de cellules restant associées ou non pour créer un nouvel organisme. A l'opposé, la reproduction sexuée fait intervenir des facteurs particuliers, appelés gamètes, et le gamète mâle vient toujours féconder le gamète femelle. Deux partenaires sont la plupart du temps nécessaires, et le nouvel organisme sera issu de la fusion de 2 cellules. Chez les organismes même les plus simples, on peut retrouver des individus femelles dits F - et des individus mâles dits HFR. Chez les bactéries, la souche HFR injecte un épisome dans le cytoplasme de la souche F - , et par un phénomène de recombinaison, on a inclusion de cet épisome dans le génome femelle. Ce n'est pas le processus normal de la reproduction des bactéries, mais cela apporte une grande variabilité. A partir de là, on peut donner une 1 e ébauche de la définition de la reproduction, à savoir la formation de cellules capables de recombiner leur génome.
Transcript of Lignée germinale et gamétogenèse -...

1
Lignée germinale et gamétogenèse
I) Introduction
1) Sexualité et reproduction On va étudier les cellules germinales et la gamétogenèse, mais on n'abordera pas les aspects de
sexualité, à savoir la différenciation des organes de la sexualité. La reproduction sexuée s'oppose à la reproduction asexuée, que l'on retrouve notamment chez les bactéries, et au cours de laquelle on a formation de clones ou de paquets de cellules restant associées ou non pour créer un nouvel organisme. A l'opposé, la reproduction sexuée fait intervenir des facteurs particuliers, appelés gamètes, et le gamète mâle vient toujours féconder le gamète femelle. Deux partenaires sont la plupart du temps nécessaires, et le nouvel organisme sera issu de la fusion de 2 cellules.
Chez les organismes même les plus simples, on peut retrouver des individus femelles dits F- et des individus mâles dits HFR. Chez les bactéries, la souche HFR injecte un épisome dans le cytoplasme de la souche F-, et par un phénomène de recombinaison, on a inclusion de cet épisome dans le génome femelle. Ce n'est pas le processus normal de la reproduction des bactéries, mais cela apporte une grande variabilité. A partir de là, on peut donner une 1e ébauche de la définition de la reproduction, à savoir la formation de cellules capables de recombiner leur génome.
2
Par rapport à la "re-production" asexuée, la reproduction sexuée présente un grand avantage : il peut arriver qu'une bactérie subisse une mutation favorable, puis produise des clones incluant cette mutation. Si 2 bactéries ont 2 mutations favorables différentes, alors dans la majorité des cas on ne pourra jamais avoir association de ces 2 mutations. La reproduction sexuée favorise le transfert d'une mutation favorable vers un autre organisme, et elle pourra alors s'additionner ou contre-balancer une autre mutation déjà présente dans l'individu receveur. La capacité de déployer des mutations et de recombiner le génome est donc un phénomène essentiel.
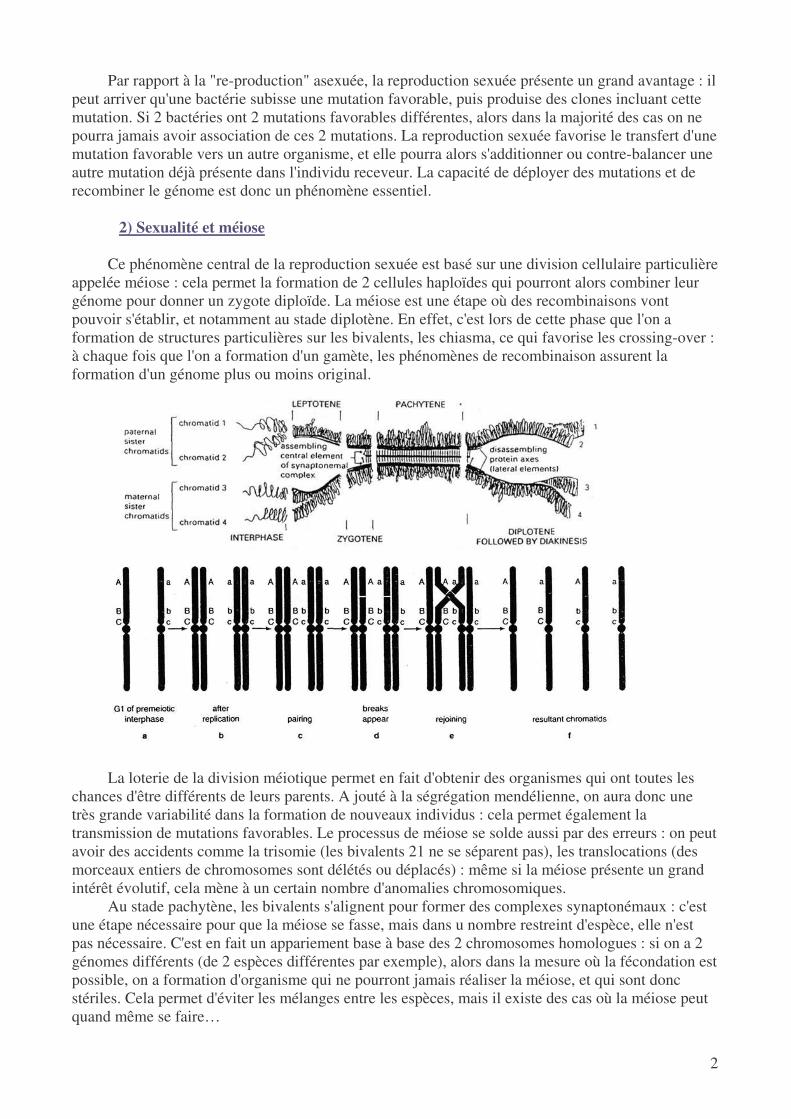
2) Sexualité et méiose
Ce phénomène central de la reproduction sexuée est basé sur une division cellulaire particulière
appelée méiose : cela permet la formation de 2 cellules haploïdes qui pourront alors combiner leur génome pour donner un zygote diploïde. La méiose est une étape où des recombinaisons vont pouvoir s'établir, et notamment au stade diplotène. En effet, c'est lors de cette phase que l'on a formation de structures particulières sur les bivalents, les chiasma, ce qui favorise les crossing-over : à chaque fois que l'on a formation d'un gamète, les phénomènes de recombinaison assurent la formation d'un génome plus ou moins original.
La loterie de la division méiotique permet en fait d'obtenir des organismes qui ont toutes les
chances d'être différents de leurs parents. A jouté à la ségrégation mendélienne, on aura donc une très grande variabilité dans la formation de nouveaux individus : cela permet également la transmission de mutations favorables. Le processus de méiose se solde aussi par des erreurs : on peut avoir des accidents comme la trisomie (les bivalents 21 ne se séparent pas), les translocations (des morceaux entiers de chromosomes sont délétés ou déplacés) : même si la méiose présente un grand intérêt évolutif, cela mène à un certain nombre d'anomalies chromosomiques.
Au stade pachytène, les bivalents s'alignent pour former des complexes synaptonémaux : c'est une étape nécessaire pour que la méiose se fasse, mais dans u nombre restreint d'espèce, elle n'est pas nécessaire. C'est en fait un appariement base à base des 2 chromosomes homologues : si on a 2 génomes différents (de 2 espèces différentes par exemple), alors dans la mesure où la fécondation est possible, on a formation d'organisme qui ne pourront jamais réaliser la méiose, et qui sont donc stériles. Cela permet d'éviter les mélanges entre les espèces, mais il existe des cas où la méiose peut quand même se faire…

3
3) Sexualité et système reproducteur On a vu que la reproduction sexuée s'accompagne du transfert du génome mâle vers le génome
femelle, puis d'un phénomène de conjugaison. Pour en arriver là, plusieurs étapes sont nécessaires et elles vont différencier selon les organismes : en effet, les phases diploïdes et haploïdes vont durer plus ou moins longtemps, et auront des longueurs similaires ou non. La formation des cellules haploïdes va dépendre le plus souvent d'un organe en particulier, alors que le reste de l'organisme restera toujours diploïde.
Les cellules qui se divisent par méiose descendent de lignées cellulaires particulières, appelées
cellules germinales primordiales : elles apparaissent dans des zones particulières de l'organisme lors de l'embryogenèse, mais leur zone de différenciation sera différente de leur zone d'apparition. On va essayer de comprendre ce qui différencie les précurseurs de la lignées germinale des précurseurs dits somatiques, qui organisent l'ensemble de l'organisme qui va se reproduire.
De manière très précoce, on apparition des gonocytes primordiaux qui restent cependant en nombre restreint. Ces cellules vont ensuite migrer vers des organes en cours de développement, qui formeront les futures ovaires et testicules : à ce stade les gonocytes primordiaux forment une population particulière de cellules mais pas un tissu. Selon l'organe d'arrivée (mêle ou femelle), ces cellules germinales deviendront des gamètes mâles ou femelles, avec chacun des spécificités. Une fois dans l'organe définitif, les gonocytes vont commencer à proliférer : on aboutit à la formation d'un très grand nombre d'ovogonies ou de spermatogonies . Cependant, on assiste également à de très nombreux phénomènes de dégénération, mais un très grand nombre de cellules restent tout de même en activité.
4
A l'issu du processus de différenciation, on va obtenir des ovules et des spermatozoïdes qui sont radicalement différents : chez l'homme, on a une anisogamie car les 2 gamètes ont une apparence très différente, et on parle également de gonochorie car on a séparation des sexes.
Lors de la gamétogenèse, on peut retrouver un phénomène de récapitulation comme dans l'organogenèse : au départ, les cellules germinales mâles et femelles apparaissent au même endroit et ne sont que très peu différenciées. On assiste ensuite à une migration des 2 types de gamètes, et au sein d'un même organisme, on peut alors avoir une différenciation des 2 types de gamètes. On a ensuite eu séparation des 2 sexes : cette étape a nécessité des adaptations beaucoup plus complexes. En effet, le gonochorisme nécessite le transfert de l'un des 2 gamètes dans un autre organisme, et pour cela on a du mettre en place tous les organes et les tubulures nécessaires (mais les gonades ont déjà été différenciées lors de l'étape précédente).
Le véritable problème de l sexualité est la rencontre des gamètes : 2 partenaires de sexe
différent sont toujours nécessaires. Le gamète mâle est une cellule mobile, de petit taille et avec u cytoplasme dense, et il va essayer de rejoindre le gamète femelle de grande taille et immobile. Chez les hommes, la phase de croissance et d'accumulation des réserves dans le gamète femelle est assez faible mais chez les poules, l'ovule peut atteindre 5 cm de diamètre ! A l'issu de cela, on voit donc que l'on a un grand partage des tâches entre les 2 gamètes.
Les manifestations extérieures du sexe sont importantes puisque cela permet de différencier les individus et d'assurer une fécondation correcte. La recherche du partenaire coûte cher énergétiquement : ainsi les organismes hermaphrodites sont assez économes, mais la loterie de la reproduction n'est alors plus favorisée du tout. Si la reproduction hermaphrodite est la règle, alors rapidement on va observer un déclin de la population : c'est en fait un phénomène qui assure la survie d'une lignée en cas de conditions défavorables du milieu, mais la reproduction croisée reste possible et obligatoire pour la pérennité de l'espèce.
Il existe de très nombreuses modalités de la sexualité : on a vu que l'hermaphrodisme est le
plus simple mais il mène à une moins grande variabilité. On distingue également la reproduction croisée, qui assure de très bonnes adaptations au milieu. On a enfin la reproduction parthénogénétique (chez les abeilles par exemple): l'activation du gamète femelle ne nécessite pas l'entrée du gamète mâle et une femelle seule sera capable d'engendrer u certain nombre d'autres individus.
La reproduction sexuée a donc un intérêt évident mais elle coûte cher pour la mise en place des
organes, des cellules… D'un autre côté, elle assure une adaptation rapide des organismes, et cela compense le plus souvent le coût élevé. Les adaptations permettant la multiplication de l'espèce seront ensuite très variables…

5
II) Les cellules germinales et leur saga
1) Introduction Les gamètes haploïdes sont produits dans des organes particuliers à partir de précurseurs
primordiaux possédant des caractéristiques propres. Ce sont en général de grosses cellules qui contiennent dans leur cytoplasme des éléments typiques : des granules ribonucléoprotéiques, des mitochondries en grand nombre et des enzymes plus ou moins spécifiques (par exemple on trouve beaucoup de phosphatase alcaline, ce qui permet une coloration spécifique de ce type de cellule).
Les cellules germinales apparaissent très tôt et on pense pour cela qu'elle appartiennent à un lignage particulier. Pendant les phases de la segmentation, elles se divisent assez peu, et ne forment ni épithélium ni tissu organisé. Elles vont ensuite migrer vers les crêtes génitales qui les attirent et qui formeront des organes somatiques. Là, les cellules germinales y subiront leur différenciation : à partir de cellules plus ou moins mésenchymateuses on va avoir formation d'une masse critique. Ce seront ensuite les seules cellules capables de subir la méiose et c'est d'ailleurs une de leurs caractéristiques principales. On ne sait pas encore pourquoi les cellules germinales subissent ce type de différenciation, mais on sait que l'environnement joue un rôle fondamental.
Il existe tout de même une autre caractéristique : les gonocytes primordiaux sont immortels. En effet, pendant leur migration puis leur différenciation, elles ne subissent aucune mort par sénescence. Dans les cellules classiques, les séquences télomériques à l'extrémité des chromosomes est grignotée à chaque division cellulaire : dans le cas des cellules germinales, des télomérases corrigent cette suppression de bases et cela permet de compenser les pertes. Cela confère donc aux gonocytes une certaine immortalité, mais ce n'est pas spécifique à ce type cellulaire : on pourra retrouver des télomérases dans certaines cellules embryonnaires et dans certaines lignées de cellules cancéreuses. Cependant, à la différence de ces dernières, les gonocytes sont capables de s'engager dans une voie de différenciation…
2) La ségrégation soma/germen
La différence entre lignée germinale et lignée somatique est visible assez rapidement au cours
de l'embryogenèse, et pour cela, certains chercheurs, dont Weissman et Nussbaum, ont essayé de prouver que ces 2 types de cellules étaient radicalement différentes. Pour eux, les cellules germinales sont l'essence de notre espèce, alors que les cellules somatiques ne permettent la formation que d'un "récipient" dans lequel pourront se développer les cellules germinales. Dans cette vue attachée fortement à la religion, l'organisme ne sert que de réceptacle protecteur aux gamètes, qui assurent la fonction essentielle de l'organisme, à savoir la reproduction.
Selon eux, la lignée germinale est déterminée dès la cellule œuf, et elle est représentée par le noyau (appelé alors plasme germinal): cela signifie que les informations contenues dans les noyaux des cellules germinales ou somatiques sont complètement différentes… Il existe chez certains organismes comme les Nématodes, des éléments qui tendent à confirmer cette hypothèse. Chez ces individus, la 1e division mitotique qui suit l'amphimixie donne 2 cellules dont l'une donnera la lignée germinale, l'autre la lignée somatique.

6
Chez la souris, les blastomères de la morula sont peu cohésifs et forment une structure particulière. Si on isole l'une de ces cellule, alors on peut la cultiver et réimplanter une des cellules fille dans une nouvelle morula (expérience de Millman) : selon la zone d'insertion, cette cellule fera partie soit du trophectoderme, soit du bouton embryonnaire. Cela signifie que jusqu'au stade morula les cellules sont capables de donner à la fois des cellules somatiques et des cellules germinales : elles ont toutes la même potentialité. Seule leur position dans l'embryon déterminera leur destinée, et la différenciation des cellules germinales sera tout à fait conforme à toute différenciation cellulaire.
Dans les processus de clonage, on prélève un noyau somatique et on peut tout à fait obtenir un organisme viable contenant donc une lignée germinale (expérience de Briggs et King) : cela signifie donc que la séparation entre soma et germen est une vue de l'esprit, mais on est quand même obligé de constater que la lignée germinale est la 1e que l'on peut caractériser. C'est simplement une différenciation comme les autres, avec ses modalités particulières.
Selon les espèces, la lignée germinale n'apparaît pas au même moment : elle commence à se
différencier vers 7,5 jours chez la souris, cad au moment de la gastrulation. En réalité, selon le phylum évolutif de l'espèce concernée, mais aussi la proportion entre héritage maternel et interactions cellulaires dans l'embryogenèse , la différenciation de la lignée germinale surviendra plus ou moins tôt.
3) Plasme germinal et déterminants germinaux.
a) Importance du plasme germinal
Chez Parascaris equorum, le zygote diploïde va subir des phases de segmentation : la position des globules polaires permet de définir le pôle animal et la 1e division se fait de manière équatoriale par rapport à l'axe animal/végétatif. On obtient alors une cellule animal un peu plus grande que la cellule végétative.
Boveri a réalisé une étude sur la segmentation de ce zygote particulier : en effet, on remarque la cellule S1 animale va subir une dégradation de ses chromosomes (= diminution chromosomique), que l'on peut matérialiser par des marquages. La cellule S1 ainsi que tous ses descendants auront donc un génome altéré.
La cellule P1 végétative se divise de manière perpendiculaire à l'axe animal/végétatif : la cellule qui se retrouve au pôle le plus animal, appelée S2, va là encore subir une diminution chromosomique, ainsi que toute la lignée qui en est issue. La cellule P2 du pôle végétatif va de nouveau se diviser en donnant une cellule normal et une cellule "diminuée…
Finalement, les lignées issues des cellules P1, P2… ont un génome entier alors que les lignées issues des cellules S1, S2… auront un génome altéré. Or, ce sont les cellules de la lignées P qui sont à l'origine des gonocytes primordiaux, car seules ces cellules permettent la transmission d'un génome normal. Cet exemple a donc été très utilisée pour étayer les hypothèses de Weissman et Nussbaum, mais Boveri s'est demandé pourquoi ce phénomène de réduction chromosomique avait lieu, et surtout pourquoi il n'atteignait pas les cellules P.
On place un œuf de Nématode dans une centrifugeuse afin de réorganiser le cytoplasme : on
peut par exemple réorienter le fuseau mitotique, et les 2 cellules issues de la 1e division héritent donc d'un morceau de pôle animal et d'un morceau de pôle végétatif. On se rend alors compte qu'aucune de ces 2 cellules ne subit une réduction chromosomique, mais si on laisse les divisions se faire
7
normalement ensuite, alors on va de nouveau avoir formation de cellules au génome réduit si elles sont au pôle animal…
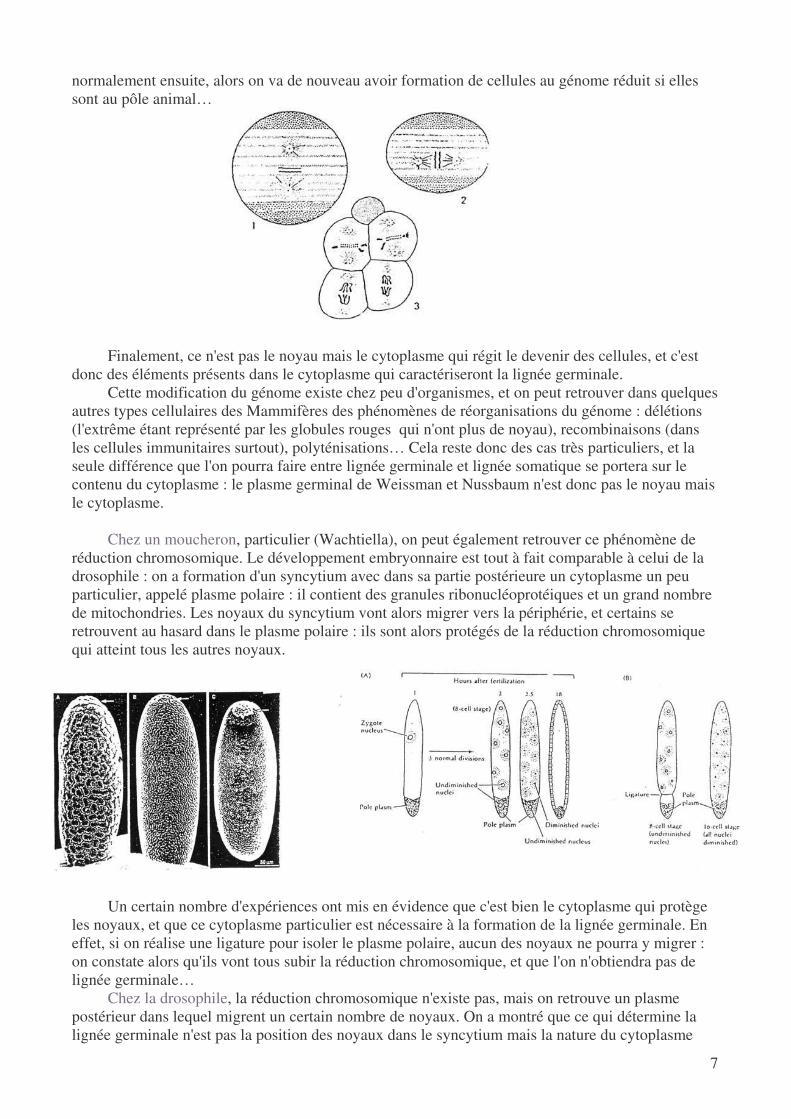
Finalement, ce n'est pas le noyau mais le cytoplasme qui régit le devenir des cellules, et c'est
donc des éléments présents dans le cytoplasme qui caractériseront la lignée germinale. Cette modification du génome existe chez peu d'organismes, et on peut retrouver dans quelques
autres types cellulaires des Mammifères des phénomènes de réorganisations du génome : délétions (l'extrême étant représenté par les globules rouges qui n'ont plus de noyau), recombinaisons (dans les cellules immunitaires surtout), polyténisations… Cela reste donc des cas très particuliers, et la seule différence que l'on pourra faire entre lignée germinale et lignée somatique se portera sur le contenu du cytoplasme : le plasme germinal de Weissman et Nussbaum n'est donc pas le noyau mais le cytoplasme.
Chez un moucheron, particulier (Wachtiella), on peut également retrouver ce phénomène de
réduction chromosomique. Le développement embryonnaire est tout à fait comparable à celui de la drosophile : on a formation d'un syncytium avec dans sa partie postérieure un cytoplasme un peu particulier, appelé plasme polaire : il contient des granules ribonucléoprotéiques et un grand nombre de mitochondries. Les noyaux du syncytium vont alors migrer vers la périphérie, et certains se retrouvent au hasard dans le plasme polaire : ils sont alors protégés de la réduction chromosomique qui atteint tous les autres noyaux.
Un certain nombre d'expériences ont mis en évidence que c'est bien le cytoplasme qui protège
les noyaux, et que ce cytoplasme particulier est nécessaire à la formation de la lignée germinale. En effet, si on réalise une ligature pour isoler le plasme polaire, aucun des noyaux ne pourra y migrer : on constate alors qu'ils vont tous subir la réduction chromosomique, et que l'on n'obtiendra pas de lignée germinale…
Chez la drosophile, la réduction chromosomique n'existe pas, mais on retrouve un plasme postérieur dans lequel migrent un certain nombre de noyaux. On a montré que ce qui détermine la lignée germinale n'est pas la position des noyaux dans le syncytium mais la nature du cytoplasme

8
dont ils héritent. En effet, on prend des noyaux qui devraient se positionner dans le plasme polaire et on les place dans la partie antérieure : ces noyaux seront à l'origine de cellules somatiques, ce qui prouve bien que là encore c'est le cytoplasme qui est nécessaire (expérience à regarder… complexe).
Chez les Amphibiens, on peut retrouver le même genre de phénomène car il existe chez tous
les organisme un cytoplasme particulier par sa composition… Dans la partie végétative de l'embryon, on retrouve ainsi un cytoplasme avec les mêmes caractéristiques que celles vues précédemment (granules et mitochondries). Ce sera à partir des cellules héritant de ce cytoplasme que l'on aura développement de la lignée germinale.
On prélève les cellules de la zone végétative qui auraient dû former les cellules germinales : la
grenouille que l'on obtient alors par développement de l'embryon modifié est stérile. Si on injecte les cellules prélevées dans un autre embryon, alors on constate que les cellules de la lignée germinale de l'adulte ainsi obtenu auront 2 origines différentes… de la même manière, si on irradie au stade blastula les cellules du pôle végétatif, alors on obtient des adultes stériles. Finalement, on retrouve bien chez les Amphibiens des cellules se divisant peu, contenant un cytoplasme particulier, et à l'origine de la lignée germinale…
9
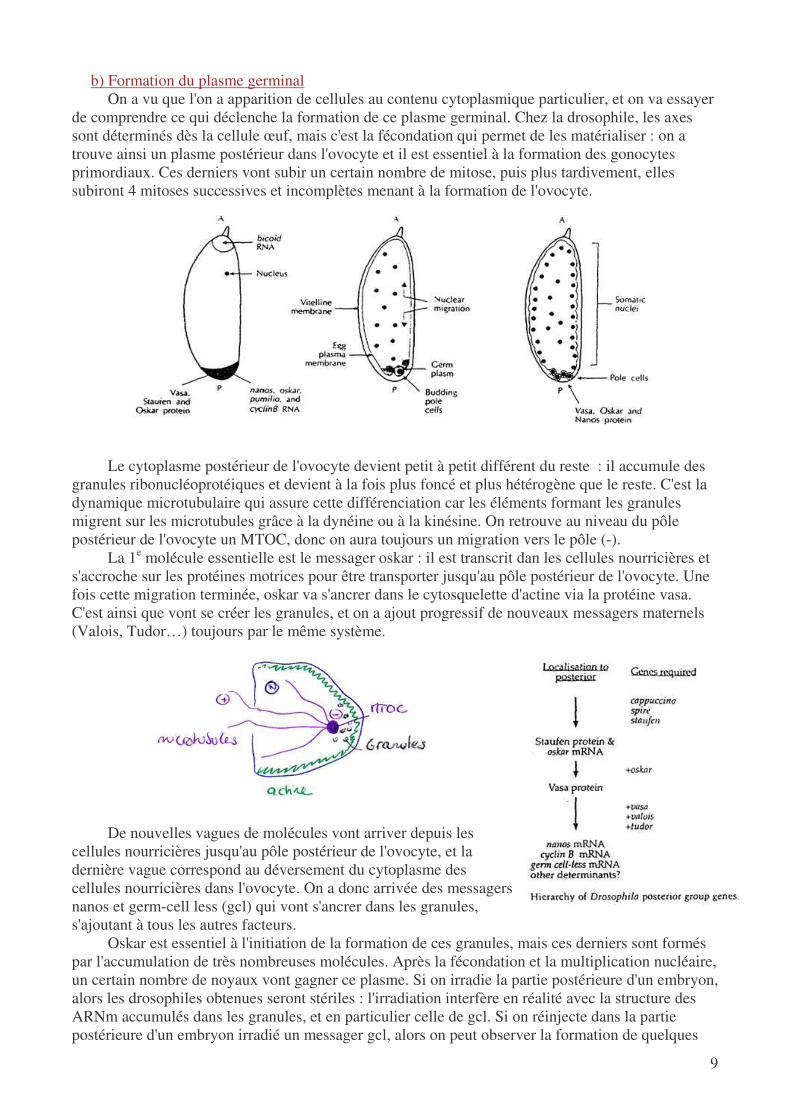
b) Formation du plasme germinal On a vu que l'on a apparition de cellules au contenu cytoplasmique particulier, et on va essayer
de comprendre ce qui déclenche la formation de ce plasme germinal. Chez la drosophile, les axes sont déterminés dès la cellule œuf, mais c'est la fécondation qui permet de les matérialiser : on a trouve ainsi un plasme postérieur dans l'ovocyte et il est essentiel à la formation des gonocytes primordiaux. Ces derniers vont subir un certain nombre de mitose, puis plus tardivement, elles subiront 4 mitoses successives et incomplètes menant à la formation de l'ovocyte.
Le cytoplasme postérieur de l'ovocyte devient petit à petit différent du reste : il accumule des
granules ribonucléoprotéiques et devient à la fois plus foncé et plus hétérogène que le reste. C'est la dynamique microtubulaire qui assure cette différenciation car les éléments formant les granules migrent sur les microtubules grâce à la dynéine ou à la kinésine. On retrouve au niveau du pôle postérieur de l'ovocyte un MTOC, donc on aura toujours un migration vers le pôle (-).
La 1e molécule essentielle est le messager oskar : il est transcrit dan les cellules nourricières et s'accroche sur les protéines motrices pour être transporter jusqu'au pôle postérieur de l'ovocyte. Une fois cette migration terminée, oskar va s'ancrer dans le cytosquelette d'actine via la protéine vasa. C'est ainsi que vont se créer les granules, et on a ajout progressif de nouveaux messagers maternels (Valois, Tudor…) toujours par le même système.
De nouvelles vagues de molécules vont arriver depuis les
cellules nourricières jusqu'au pôle postérieur de l'ovocyte, et la dernière vague correspond au déversement du cytoplasme des cellules nourricières dans l'ovocyte. On a donc arrivée des messagers nanos et germ-cell less (gcl) qui vont s'ancrer dans les granules, s'ajoutant à tous les autres facteurs.
Oskar est essentiel à l'initiation de la formation de ces granules, mais ces derniers sont formés par l'accumulation de très nombreuses molécules. Après la fécondation et la multiplication nucléaire, un certain nombre de noyaux vont gagner ce plasme. Si on irradie la partie postérieure d'un embryon, alors les drosophiles obtenues seront stériles : l'irradiation interfère en réalité avec la structure des ARNm accumulés dans les granules, et en particulier celle de gcl. Si on réinjecte dans la partie postérieure d'un embryon irradié un messager gcl, alors on peut observer la formation de quelques

10
cellules germinales : c'est donc que cet ARNm est essentiel à la différenciation de la lignée. Cependant, il n'intervient pas seul car la restauration n'est que partielle, et c'est tout un contexte qui est nécessaire à la formation de la lignée germinale.
Dans les granules, on retrouve non seulement des ARNm, mais aussi beaucoup de
mitochondries, et c'est vrai chez beaucoup d'espèces. Elles sont capables de synthétiser des ARN ribosomaux de grande taille, que l'on appelle mtlrRNA : ils sont toujours associés à la formation des cellules germinales. En effet, si on irradie la partie postérieure d'un embryon et qu'on ajoute des mtlrRNA, alors on peut voir apparaître des cellules polaires mais elles ne formeront jamais de cellules germinales, puisqu'il manque gcl.
Par contre, si on injecte à la fois ces ARNr et l'ARNm de gcl, alors on obtient des cellules
germinales, et cela signifie finalement que ce sont tous 2 des éléments nécessaires soit à la ségrégation des cellules soit à leur différenciation.
On a pu montrer que les ARNr s'associent avec l'un des 2 centrosomes lors de la division cellulaire, et cela provoque une division asymétrique. Seule la cellule héritant des ARNr, et donc issue du centrosome modifié deviendra une cellule germinale, si elle contient les déterminants nécessaires. On se retrouve dans la problématique de toute différenciation cellulaire, puisqu'elles s'accompagnent toutes d'une division asymétrique, mais ici on aurait trouvé l'une des causes de cette asymétrie…
Chez les Amphibiens, on retrouve des prémices du plasme germinal dans les cellules du pôle végétatif, et elles ségrègent de manière asymétrique pendant la segmentation. Ce sont des cellules qui se multiplient assez peu par rapport aux autres, et dans tous les cas on a une division asymétrique. Cela permet de conserver un nombre fixe et assez faible de cellules germinales.
Pendant l'ovogenèse des Amphibiens, un certain nombre d'ARNm ainsi que des mitochondries
sont stockés dans la partie végétative, dans la partie animale ou bien de manière aléatoire et donc symétrique. On retrouve par exemple des équivalents de vasa… On pense que la détermination des cellules germinales des Amphibiens est très proche de la mécanistique de celle des drosophiles.
Une fois dans les gonades, les divisions deviendront complètement symétriques, mais les cellules seront déjà déterminées dans leur lignée. On ne connaît pas encore les facteurs qui assurent la différenciation des cellules germinales, ni pourquoi on a apparition de ces cellules à un moment donné.
4) La migration des cellules germinales
Les mécanismes de migration qui vont amener les cellules germinales jusqu'aux gonades sont
plus ou moins divers. Chez la drosophile, on sait que l'embryogenèse est le siège de l'invagination d'un sillon dorsal, formant ainsi la bandelette germinative. Cette dernière va s'étaler postérieurement puis dorsalement, et les cellules polaires sont donc entraînées dans ce mouvement. Or, cet étalement

11
s'accompagne de l'invagination proctodéale, et les cellules polaires se retrouvent alors dans cette invagination.
Les cellules polaires sont associées à l'intestin postérieur, mais avant la différenciation de ce
dernier, elles vont migrer à travers la paroi de l'intestin pour gagner les futures gonades. On a donc une étape passive pendant la migration de la bandelette germinative, puis active. Pour que la migration soit possible et surtout efficace, les crêtes génitales sécrètent des substances chimiotactiques.
Les mécanismes morphogénétiques lors de la gastrulation chez les embryons de toutes les
espèces favorisent très souvent le rapprochement des cellules germinales et des gonades, sauf chez les Oiseaux, où les précurseurs devront emprunter la voie sanguine pour gagner les organes appropriés.
Chez les Amphibiens, les cellules germinales apparaissent dans la partie végétative de l'embryon, et restent en petit nombre dans la future paroi du tube digestif. Au moment de la gastrulation, les cellules qui se trouvent dans le plancher de l'intestin vont s'engager dans un processus de migration vers les gonades, mais en même temps elles recommencent à se diviser.
La migration se fait à travers le mésentère, et les cellules germinales gagnent ainsi les crêtes
génitales qui sont en train de se creuser de chaque côté, et se divisent un peu plus. La migration a été active : on peut observer des émissions de filopodes et de lamellipodes qui, en présence d'une matrice extracellulaire riche en intégrine notamment, permettent le déplacement.
12
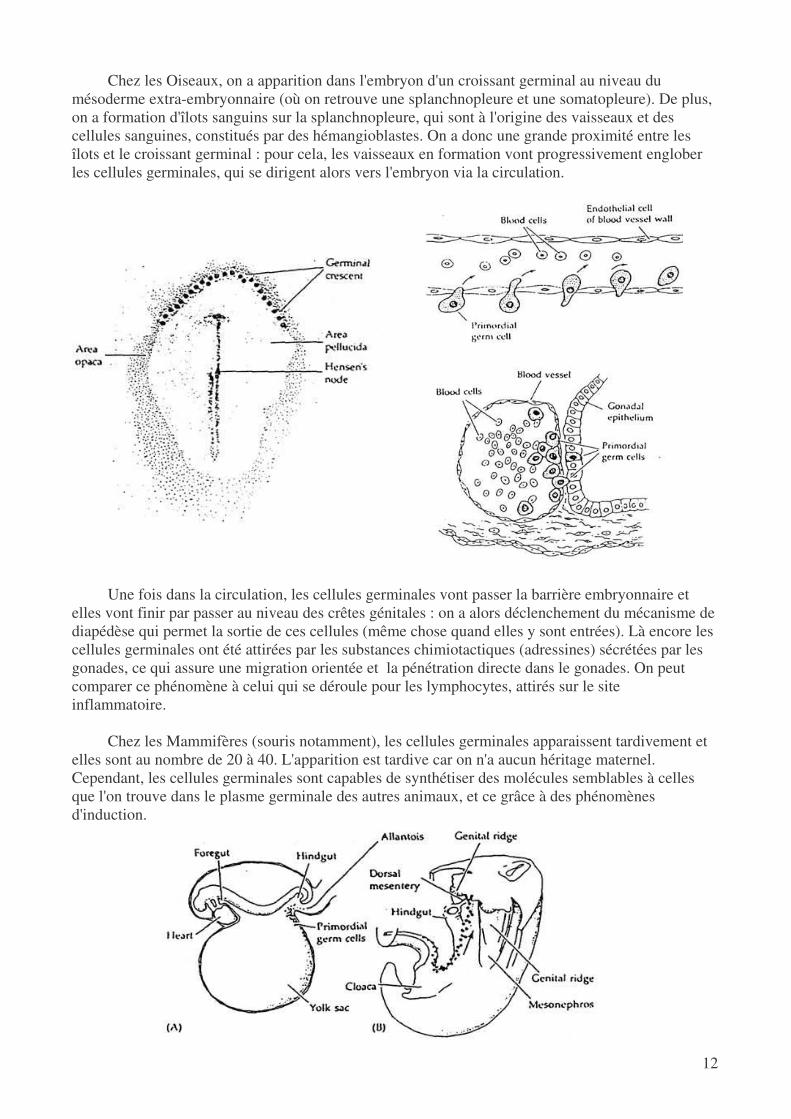
Chez les Oiseaux, on a apparition dans l'embryon d'un croissant germinal au niveau du mésoderme extra-embryonnaire (où on retrouve une splanchnopleure et une somatopleure). De plus, on a formation d'îlots sanguins sur la splanchnopleure, qui sont à l'origine des vaisseaux et des cellules sanguines, constitués par des hémangioblastes. On a donc une grande proximité entre les îlots et le croissant germinal : pour cela, les vaisseaux en formation vont progressivement englober les cellules germinales, qui se dirigent alors vers l'embryon via la circulation.
Une fois dans la circulation, les cellules germinales vont passer la barrière embryonnaire et
elles vont finir par passer au niveau des crêtes génitales : on a alors déclenchement du mécanisme de diapédèse qui permet la sortie de ces cellules (même chose quand elles y sont entrées). Là encore les cellules germinales ont été attirées par les substances chimiotactiques (adressines) sécrétées par les gonades, ce qui assure une migration orientée et la pénétration directe dans le gonades. On peut comparer ce phénomène à celui qui se déroule pour les lymphocytes, attirés sur le site inflammatoire.
Chez les Mammifères (souris notamment), les cellules germinales apparaissent tardivement et
elles sont au nombre de 20 à 40. L'apparition est tardive car on n'a aucun héritage maternel. Cependant, les cellules germinales sont capables de synthétiser des molécules semblables à celles que l'on trouve dans le plasme germinale des autres animaux, et ce grâce à des phénomènes d'induction.

13
Les 1e cellules germinales apparaissent au moment de la gastrulation (7 jours), et elles se situent à la limite de l'ectoderme extra-embryonnaire et de l'épiblaste embryonnaire, et donc à proximité du mésoblaste (à chaque fois le devenir des cellules germinales semble lié à l'intestin…).
Ces cellules sont déterminées dans la voie génitale car elles ont reçu des signaux adéquats. En
effet, l'endoblaste envoie des signaux de la famille des BMP vers l'épiblaste, et ce dernier répond par la différenciation d'une partie de ses cellules en cellules germinales. Ce phénomène est permis par des voies de signalisation de type phosphorylation, qui aboutissent à l'expression de gènes permettant l'accumulation de substances comparables à celles de la drosophile.
Quand l'allantoïde se forme, les cellules germinales se rassemblent, et via le cordon ombilical, elles migrent vers les crêtes génitales en passant par le TD puis le mésentère. Cette migration est facilitée par des mouvements morphogénétiques et par libération de substances chimiotactiques, mais ce mécanisme reste quand même actif. Les 1e cellules qui s'engagent dans la migration sont les plus proches des gonades : lors de leur déplacement, elles vont laisser derrière elles des expansions cytoplasmiques. On a donc formation d'un véritable fil d'Ariane qui servira de guide pour les suivantes : de très nombreux éléments interviennent donc pour rendre cette migration des gonocytes la plus fiable possible.
Lors de la migration, les cellules germinales commencent à se diviser de manière symétrique :
leur nombre va donc augmenter considérablement. Une fois dans les gonades, la prolifération devient alors beaucoup plus importante, et on peut obtenir jusqu'à plusieurs millions de cellules… Ce nombre sera cependant modifié par des phénomènes d'apoptose, ce qui provoque la destruction d'une plus ou moins grande partie des cellules ainsi obtenues : la perte s'élève à 70% pour les spermatogonies et à 30% pour les ovogonies (mais on aura de nouvelles pertes lors de la différenciation folliculaire par la phénomène d'atrésie). On n'observe part contre jamais de mort par sénescence et l'ajustement du stock se fera donc par la seule apoptose (et ce processus est valable aussi pour de très nombreux organes).
5) Aspects généraux de la gamétogenèse. La gamétogenèse suit des modalités différentes selon que l'on se trouve chez le mâle ou chez la
femelle : si dans les 2 cas on a un processus de différenciation, il est continu dans le cas de la spermatogenèse et discontinu dans le cas de l'ovogenèse. De plus, on peut obtenir un très grand nombre de spermatozoïdes par multiplication tardive des spermatogonies, alors que les ovogonies sont stockées dans l'ovules et sont bloquées en méiose (donc elles ne peuvent pas se multiplier).
La différenciation de ces cellules va également se faire à des moments différents : avant la méiose pour les ovogonies, pendant et après pour les spermatogonies. Cependant, dans les 2 cas, on a une régulation assez fine de ces processus grâce à l'axe hypothalamo-hypophysaire qui sécrète différentes hormones comme le LH et le FSH. De plus, les cellules cibles de ces hormones sont dans les 2 cas des cellules somatiques très semblables, mais on les trouve soit dans les testicules soit dans les ovaires.

14
III) La spermatogenèse
1) Description des testicules
La différenciation des cellules germinales primordiales va débuter quand elle auront atteint une
gonade mâle (testicule en formation). Les testicules ont un développement complexe, et leur organisation est très particulière. C'est un organe pair constitué par des lobules séparés entre eux par des septa testis et dans chaque lobule on retrouve les tube séminifères condensés et contournés dont la quantité et la densité sont très importante. La spermatogenèse se fera justement dans la paroi de ces tubes séminifères, et le reste de l'organe sera responsable de la régulation de ce processus.
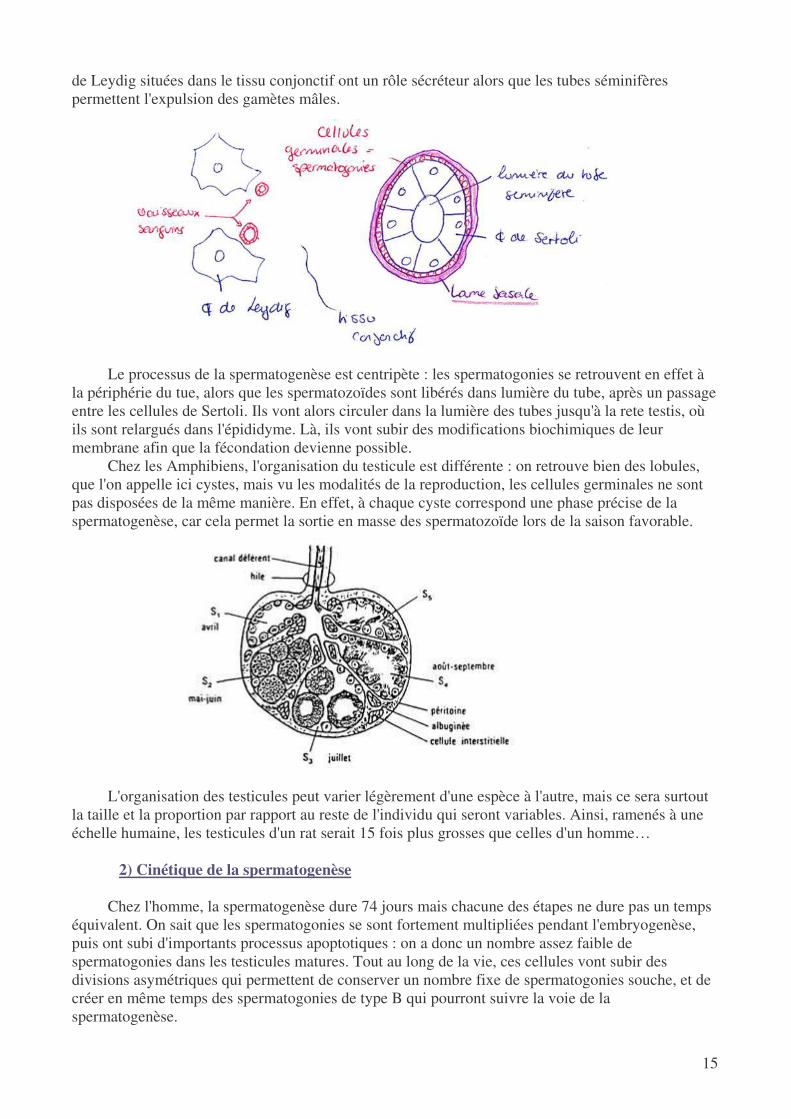
Les testicules sont entourés par la tunique albuginée, qui joue un rôle de protection physique, et
ils sont coiffés par la tête de l'épididyme où sont stockés et modifiés les spermatozoïdes. A l'abri dans l'albuginée, on retrouve en fait un organe à la fois endocrine et exocrine : en effet, les cellules
15
de Leydig situées dans le tissu conjonctif ont un rôle sécréteur alors que les tubes séminifères permettent l'expulsion des gamètes mâles.
Le processus de la spermatogenèse est centripète : les spermatogonies se retrouvent en effet à
la périphérie du tue, alors que les spermatozoïdes sont libérés dans lumière du tube, après un passage entre les cellules de Sertoli. Ils vont alors circuler dans la lumière des tubes jusqu'à la rete testis, où ils sont relargués dans l'épididyme. Là, ils vont subir des modifications biochimiques de leur membrane afin que la fécondation devienne possible.
Chez les Amphibiens, l'organisation du testicule est différente : on retrouve bien des lobules, que l'on appelle ici cystes, mais vu les modalités de la reproduction, les cellules germinales ne sont pas disposées de la même manière. En effet, à chaque cyste correspond une phase précise de la spermatogenèse, car cela permet la sortie en masse des spermatozoïde lors de la saison favorable.
L'organisation des testicules peut varier légèrement d'une espèce à l'autre, mais ce sera surtout
la taille et la proportion par rapport au reste de l'individu qui seront variables. Ainsi, ramenés à une échelle humaine, les testicules d'un rat serait 15 fois plus grosses que celles d'un homme…
2) Cinétique de la spermatogenèse
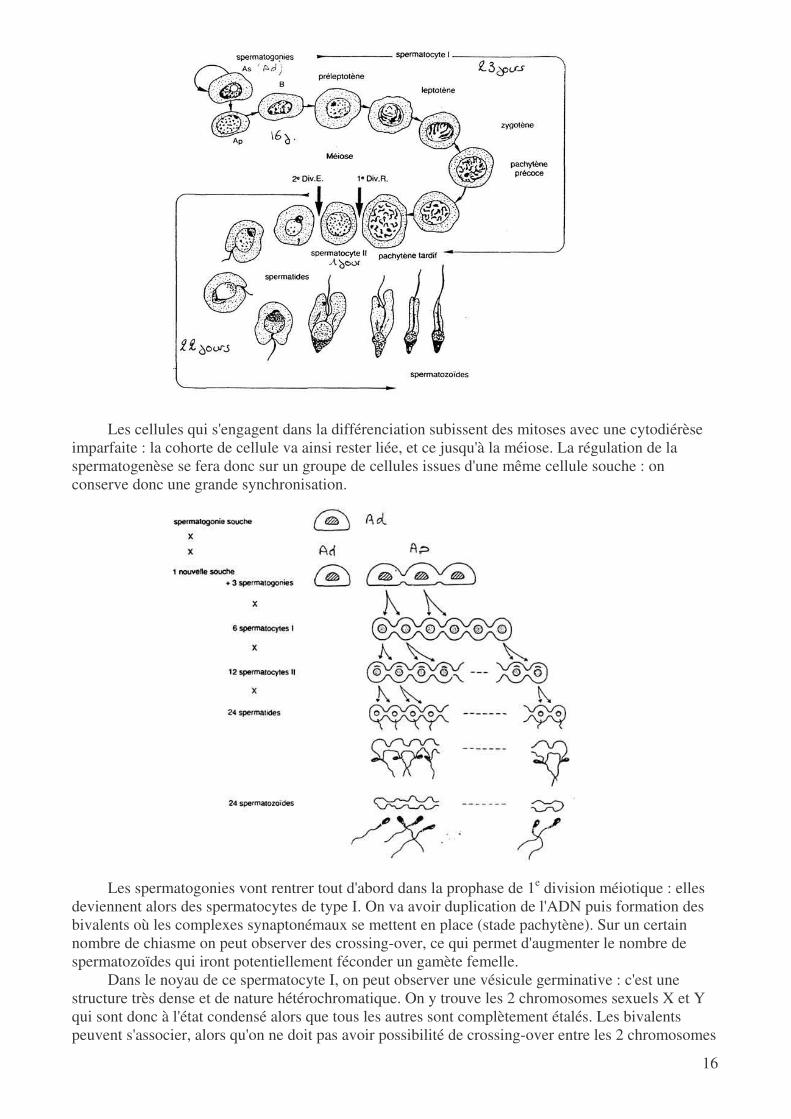
Chez l'homme, la spermatogenèse dure 74 jours mais chacune des étapes ne dure pas un temps
équivalent. On sait que les spermatogonies se sont fortement multipliées pendant l'embryogenèse, puis ont subi d'importants processus apoptotiques : on a donc un nombre assez faible de spermatogonies dans les testicules matures. Tout au long de la vie, ces cellules vont subir des divisions asymétriques qui permettent de conserver un nombre fixe de spermatogonies souche, et de créer en même temps des spermatogonies de type B qui pourront suivre la voie de la spermatogenèse.
16
Les cellules qui s'engagent dans la différenciation subissent des mitoses avec une cytodiérèse
imparfaite : la cohorte de cellule va ainsi rester liée, et ce jusqu'à la méiose. La régulation de la spermatogenèse se fera donc sur un groupe de cellules issues d'une même cellule souche : on conserve donc une grande synchronisation.
Les spermatogonies vont rentrer tout d'abord dans la prophase de 1e division méiotique : elles
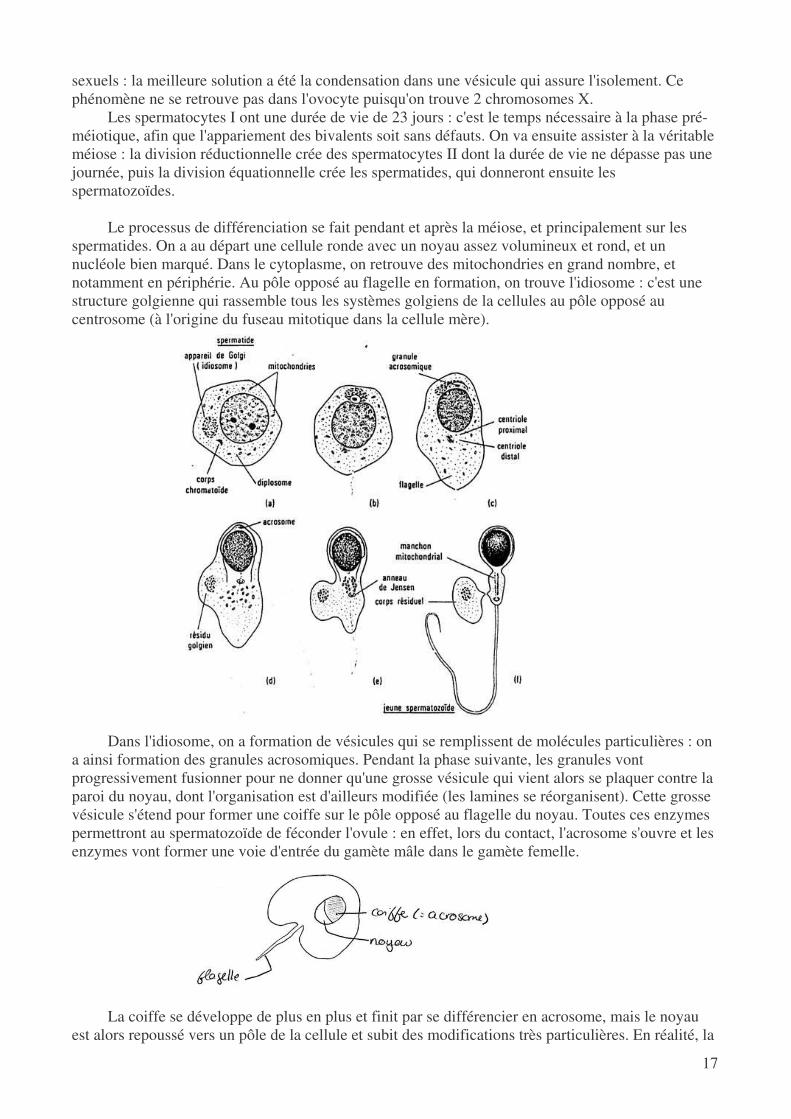
deviennent alors des spermatocytes de type I. On va avoir duplication de l'ADN puis formation des bivalents où les complexes synaptonémaux se mettent en place (stade pachytène). Sur un certain nombre de chiasme on peut observer des crossing-over, ce qui permet d'augmenter le nombre de spermatozoïdes qui iront potentiellement féconder un gamète femelle.
Dans le noyau de ce spermatocyte I, on peut observer une vésicule germinative : c'est une structure très dense et de nature hétérochromatique. On y trouve les 2 chromosomes sexuels X et Y qui sont donc à l'état condensé alors que tous les autres sont complètement étalés. Les bivalents peuvent s'associer, alors qu'on ne doit pas avoir possibilité de crossing-over entre les 2 chromosomes
17
sexuels : la meilleure solution a été la condensation dans une vésicule qui assure l'isolement. Ce phénomène ne se retrouve pas dans l'ovocyte puisqu'on trouve 2 chromosomes X.
Les spermatocytes I ont une durée de vie de 23 jours : c'est le temps nécessaire à la phase pré-méiotique, afin que l'appariement des bivalents soit sans défauts. On va ensuite assister à la véritable méiose : la division réductionnelle crée des spermatocytes II dont la durée de vie ne dépasse pas une journée, puis la division équationnelle crée les spermatides, qui donneront ensuite les spermatozoïdes.
Le processus de différenciation se fait pendant et après la méiose, et principalement sur les
spermatides. On a au départ une cellule ronde avec un noyau assez volumineux et rond, et un nucléole bien marqué. Dans le cytoplasme, on retrouve des mitochondries en grand nombre, et notamment en périphérie. Au pôle opposé au flagelle en formation, on trouve l'idiosome : c'est une structure golgienne qui rassemble tous les systèmes golgiens de la cellules au pôle opposé au centrosome (à l'origine du fuseau mitotique dans la cellule mère).
Dans l'idiosome, on a formation de vésicules qui se remplissent de molécules particulières : on
a ainsi formation des granules acrosomiques. Pendant la phase suivante, les granules vont progressivement fusionner pour ne donner qu'une grosse vésicule qui vient alors se plaquer contre la paroi du noyau, dont l'organisation est d'ailleurs modifiée (les lamines se réorganisent). Cette grosse vésicule s'étend pour former une coiffe sur le pôle opposé au flagelle du noyau. Toutes ces enzymes permettront au spermatozoïde de féconder l'ovule : en effet, lors du contact, l'acrosome s'ouvre et les enzymes vont former une voie d'entrée du gamète mâle dans le gamète femelle.
La coiffe se développe de plus en plus et finit par se différencier en acrosome, mais le noyau
est alors repoussé vers un pôle de la cellule et subit des modifications très particulières. En réalité, la

18
cellule se polarise dans son ensemble : elle s'allonge, les mitochondries se rassemblent et le flagelle continue à se développer activement. Le cytoplasme de la spermatide va alors être condensé dans un corps résiduel qui sera éliminé.
On aboutit ainsi au spermatozoïde classique où on distingue classiquement une tête avec le
noyau et l'acrosome, un cou avec les mitochondries formant un fourreau autour de la naissance du flagelle et un flagelle. Les granules pro-acrosomiques correspondent à toute une batterie d'enzymes typiques : on retrouve ainsi des enzymes de la catégories de trypsines (= acrosine), des enzymes dégradant la matrice extracellulaires…
Le noyau du spermatozoïde va exprimer un certain nombre de gènes qui permet la synthèse de
protéines spécifiques : elles vont assurer un niveau de condensation de la chromatine que seul le spermatozoïde peu atteindre. Cela mène à une structure quasi-cristalline et presque complètement déshydratée : toute activité transcriptionnelle devient alors impossible. Cet état est permis par le remplacement des histones classiques par des protéines type protamines qui assurent une "ultra-condensation".
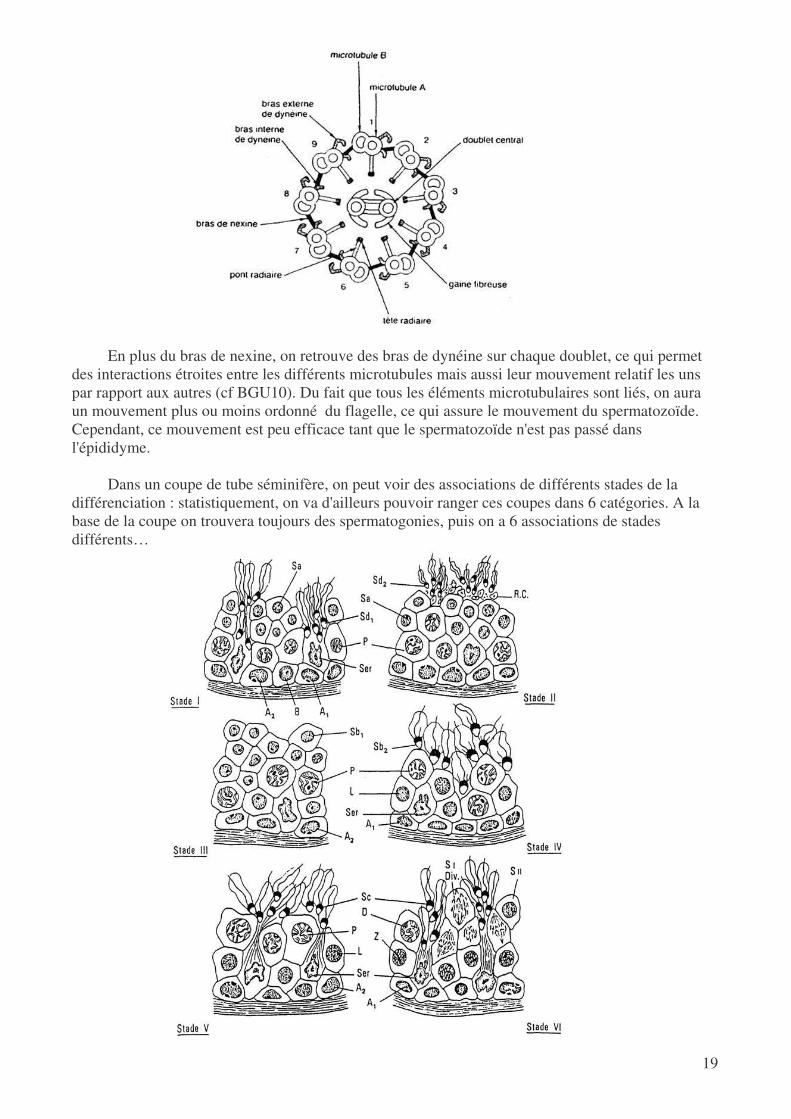
E centrosome peut se diviser en 2 et l'un sera à l'origine du flagelle, qui s'entoure d'une gaine mitochondriale. On y trouve 9 doublets de microtubules (microtubule B à 10 protofilaments et microtubule A à 13 protofilaments) qui représente la partie motrice du flagelles. On a établissement de ponts entre les différents doublets ainsi que des protéines radiaires qui permettent l'interaction avec le doublet central.
19
En plus du bras de nexine, on retrouve des bras de dynéine sur chaque doublet, ce qui permet
des interactions étroites entre les différents microtubules mais aussi leur mouvement relatif les uns par rapport aux autres (cf BGU10). Du fait que tous les éléments microtubulaires sont liés, on aura un mouvement plus ou moins ordonné du flagelle, ce qui assure le mouvement du spermatozoïde. Cependant, ce mouvement est peu efficace tant que le spermatozoïde n'est pas passé dans l'épididyme.
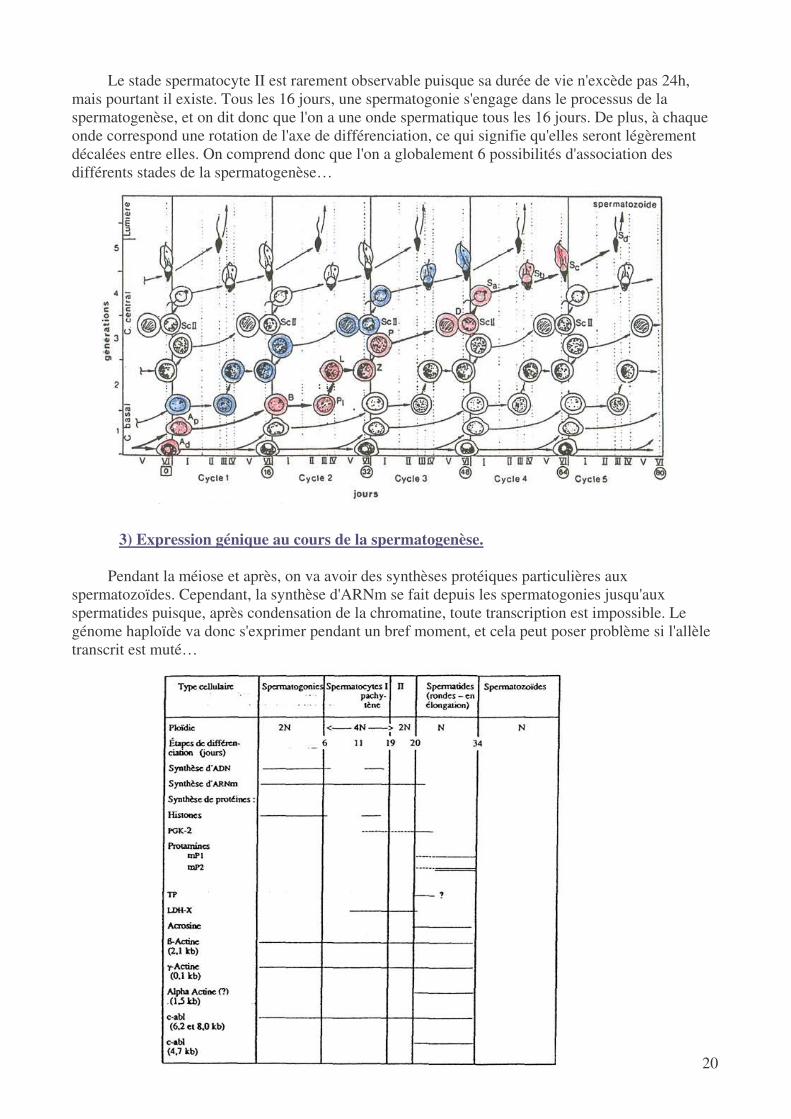
Dans un coupe de tube séminifère, on peut voir des associations de différents stades de la
différenciation : statistiquement, on va d'ailleurs pouvoir ranger ces coupes dans 6 catégories. A la base de la coupe on trouvera toujours des spermatogonies, puis on a 6 associations de stades différents…
20
Le stade spermatocyte II est rarement observable puisque sa durée de vie n'excède pas 24h, mais pourtant il existe. Tous les 16 jours, une spermatogonie s'engage dans le processus de la spermatogenèse, et on dit donc que l'on a une onde spermatique tous les 16 jours. De plus, à chaque onde correspond une rotation de l'axe de différenciation, ce qui signifie qu'elles seront légèrement décalées entre elles. On comprend donc que l'on a globalement 6 possibilités d'association des différents stades de la spermatogenèse…
3) Expression génique au cours de la spermatogenèse. Pendant la méiose et après, on va avoir des synthèses protéiques particulières aux
spermatozoïdes. Cependant, la synthèse d'ARNm se fait depuis les spermatogonies jusqu'aux spermatides puisque, après condensation de la chromatine, toute transcription est impossible. Le génome haploïde va donc s'exprimer pendant un bref moment, et cela peut poser problème si l'allèle transcrit est muté…

21
Les messagers "histones" classiques se retrouvent dans la spermatogonie et la spermatide I, mais à partir de la spermatide II, ils sont remplacés et éliminés par les protamines. On va également avoir synthèse d'hormones, mais ce sont des isoformes qui s'expriment : cela signifie qu'une seule et même molécule existe sous plusieurs formes, chacune caractéristique d'une lignée cellulaire. On a également synthèse de protéines spécifiques dont les protamines…
On a donc toute une régulation de l'expression génique et chaque gène s'exprimera à différents moments de la spermatogenèse. On pourra observer des synthèses de protéines non spécifiques (donc les promoteurs ne sont pas tissus-spécifiques), de protéines non spécifiques mais dans une forme particulière, et de protéines spécifiques. De plus, on pourra avoir des expressions alternatives d'un même gène selon le promoteur utilisé, ainsi que des phénomènes d'épissage alternatif. Tout cela pendant les phases diploïde et le début de la phase haploïde.
4) Contrôle de la fonction testiculaire
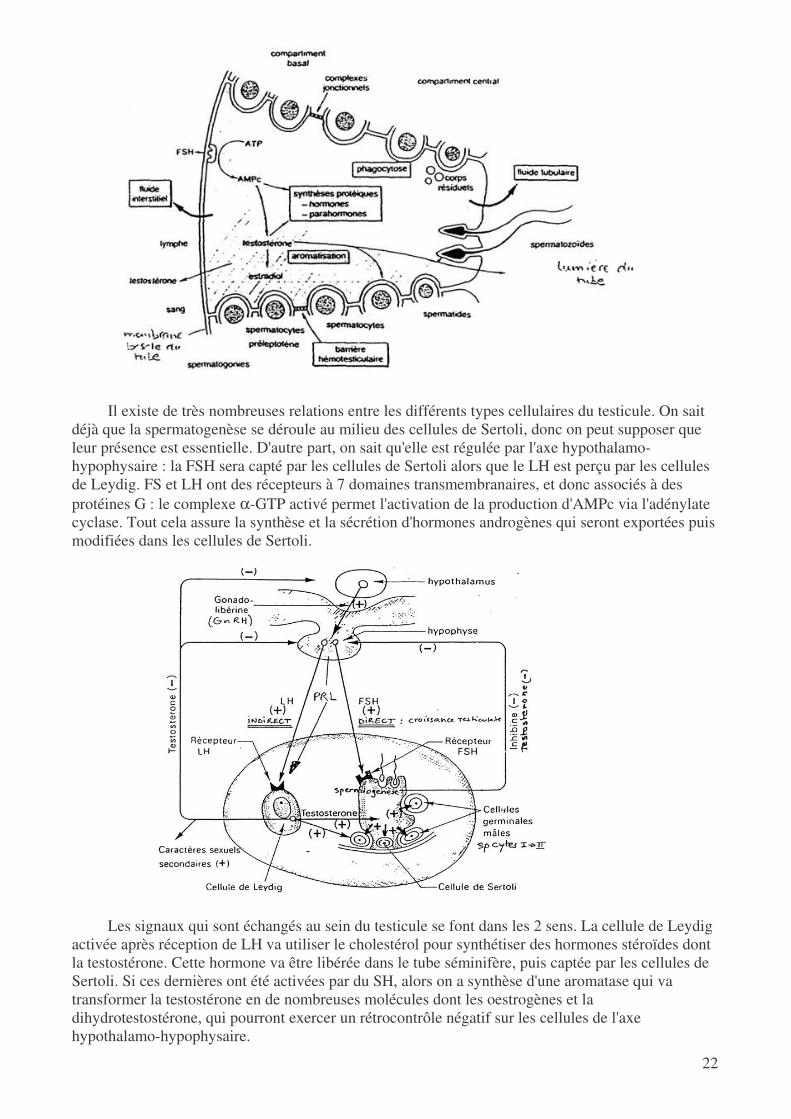
Les cellules de Sertoli forment des compartiments entre elles et c'est dans ces espaces que se
déroulera la spermatogenèse. Il existe une véritable barrière au niveau de ces compartiments : en effet, on peut observer dans une zone particulière la formation de jonction étanches (occludines). Cela crée une barrière hémato-testiculaire qui peut cependant s'ouvrir pour laisser les cellules qui progressent dans leur différenciation : on a ainsi pu remarquer que seuls les spermatocytes préleptotène peuvent traverser cette barrière.
Le compartiment "hémato" renferme donc des cellules où on a des divisions de type mitotique (duplication de l'ADN) alors que le compartiment testiculaire renferme des cellules qui subissent une véritable division méiotique. Au fur et à mesure de leur développement, les spermatozoïdes vont donc être isolés du reste de l'organisme.
22
Il existe de très nombreuses relations entre les différents types cellulaires du testicule. On sait
déjà que la spermatogenèse se déroule au milieu des cellules de Sertoli, donc on peut supposer que leur présence est essentielle. D'autre part, on sait qu'elle est régulée par l'axe hypothalamo-hypophysaire : la FSH sera capté par les cellules de Sertoli alors que le LH est perçu par les cellules de Leydig. FS et LH ont des récepteurs à 7 domaines transmembranaires, et donc associés à des protéines G : le complexe α-GTP activé permet l'activation de la production d'AMPc via l'adénylate cyclase. Tout cela assure la synthèse et la sécrétion d'hormones androgènes qui seront exportées puis modifiées dans les cellules de Sertoli.
Les signaux qui sont échangés au sein du testicule se font dans les 2 sens. La cellule de Leydig
activée après réception de LH va utiliser le cholestérol pour synthétiser des hormones stéroïdes dont la testostérone. Cette hormone va être libérée dans le tube séminifère, puis captée par les cellules de Sertoli. Si ces dernières ont été activées par du SH, alors on a synthèse d'une aromatase qui va transformer la testostérone en de nombreuses molécules dont les oestrogènes et la dihydrotestostérone, qui pourront exercer un rétrocontrôle négatif sur les cellules de l'axe hypothalamo-hypophysaire.
23
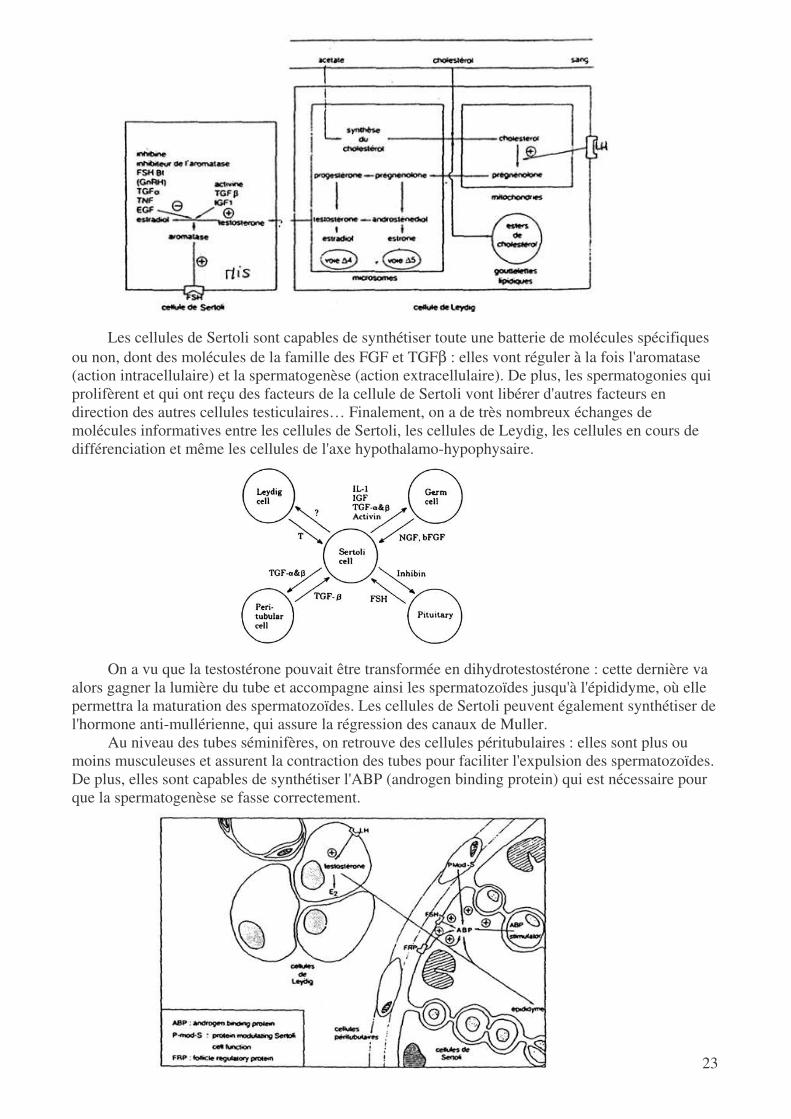
Les cellules de Sertoli sont capables de synthétiser toute une batterie de molécules spécifiques
ou non, dont des molécules de la famille des FGF et TGFβ : elles vont réguler à la fois l'aromatase (action intracellulaire) et la spermatogenèse (action extracellulaire). De plus, les spermatogonies qui prolifèrent et qui ont reçu des facteurs de la cellule de Sertoli vont libérer d'autres facteurs en direction des autres cellules testiculaires… Finalement, on a de très nombreux échanges de molécules informatives entre les cellules de Sertoli, les cellules de Leydig, les cellules en cours de différenciation et même les cellules de l'axe hypothalamo-hypophysaire.
On a vu que la testostérone pouvait être transformée en dihydrotestostérone : cette dernière va
alors gagner la lumière du tube et accompagne ainsi les spermatozoïdes jusqu'à l'épididyme, où elle permettra la maturation des spermatozoïdes. Les cellules de Sertoli peuvent également synthétiser de l'hormone anti-mullérienne, qui assure la régression des canaux de Muller.
Au niveau des tubes séminifères, on retrouve des cellules péritubulaires : elles sont plus ou moins musculeuses et assurent la contraction des tubes pour faciliter l'expulsion des spermatozoïdes. De plus, elles sont capables de synthétiser l'ABP (androgen binding protein) qui est nécessaire pour que la spermatogenèse se fasse correctement.
24
IV) L'ovogenèse
1) Ovaires, follicules et contrôle hormonal de l'ovogenèse
C'est la différenciation des gamètes femelles : elle est particulière puisque c'est un processus méiotique, mais le reste de la différenciation sera un processus tout à fait classique. La majeure partie de la différenciation va se faire pendant le stade diplotène, cad avant la méiose, et c'est une des différences majeures avec la spermatogenèse.
De plus, l'ovogenèse est un processus discontinu : à partir des ovogonies, qui se divisent intensément dans les ovaires, on va obtenir un stock assez important de cellules. On va alors observer une phase d'accroissement en taille, qui sera très variable selon les espèces : on parle de cytodifférenciation, et elle se termine par un blocage des ovocytes. A partir de la puberté, les ovogonies vont rentrer dans un processus de maturation, qui revient en fait à la fin de la méiose (elles étaient bloquées au stade diplotène de la prophase de 1e division méiotique). On va ensuite avoir libération d'ovocytes bloqués en métaphase de 2e division méiotique, et seule la fécondation permettra de terminer le cycle.
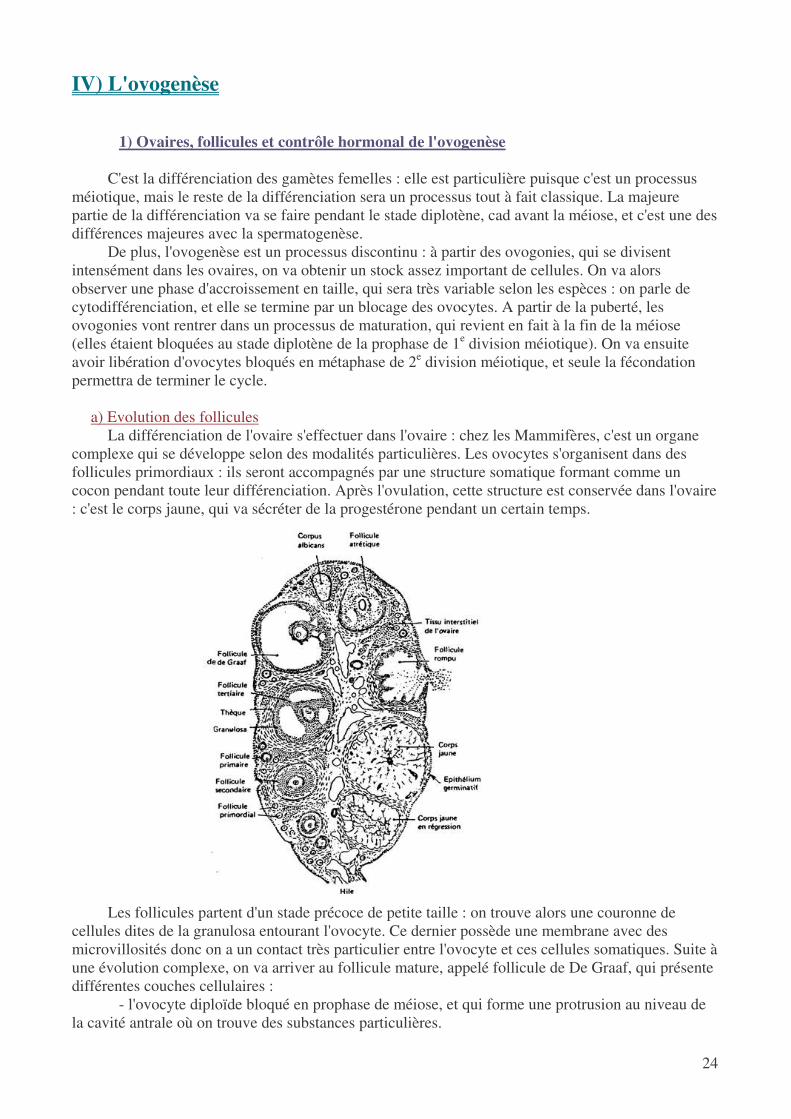
a) Evolution des follicules
La différenciation de l'ovaire s'effectuer dans l'ovaire : chez les Mammifères, c'est un organe complexe qui se développe selon des modalités particulières. Les ovocytes s'organisent dans des follicules primordiaux : ils seront accompagnés par une structure somatique formant comme un cocon pendant toute leur différenciation. Après l'ovulation, cette structure est conservée dans l'ovaire : c'est le corps jaune, qui va sécréter de la progestérone pendant un certain temps.
Les follicules partent d'un stade précoce de petite taille : on trouve alors une couronne de
cellules dites de la granulosa entourant l'ovocyte. Ce dernier possède une membrane avec des microvillosités donc on a un contact très particulier entre l'ovocyte et ces cellules somatiques. Suite à une évolution complexe, on va arriver au follicule mature, appelé follicule de De Graaf, qui présente différentes couches cellulaires :
- l'ovocyte diploïde bloqué en prophase de méiose, et qui forme une protrusion au niveau de la cavité antrale où on trouve des substances particulières.

25
- la granulosa en contact avec le cumulus oophorus qui constitue comme un épithélium - un tissu conjonctif séparé du reste par la membrane de Slavjanski : on y distingue la thèque
interne et la thèque externe, fortement irriguée On voit donc à travers l'étude de toutes ces couches que l'on aura établissement d'interactions
épithélio-mésenchymateuses, et l'accompagnement de l'ovocyte va assurer le bon déroulement de la phase d'accroissement (cytodifférenciation) et de maturation (méiose).
b) Structure des ovaires
Quand on observe la différenciation des cellules germinales, on peut généralement retrouver une certaine récapitulation : en effet chez les Mammifères, les ovogonies ne gagnent que secondairement les gonades qui, au cours de leur développement, montrent des structures de plus en plus complexes.
Chez les Annélides, les cellules germinales sont produites par simple délamination de cellules dans la cavité cœlomique, et on peut voir dans certain cas une association de ces ovocytes avec des cellules germinales. Chez les Echinodermes, on peut distinguer un ovaire qui commence à se différencier…
Chez la Drosophile, les ovaires sont constitués par des grappes d'ovarioles
où on trouve l'ovocyte associé à 15 cellules nourricières d'origine germinale (mais elles ne subissent pas la méiose). Chez les Amphibiens enfin, les ovaires renferment des ovocytes entourés par une couronne de cellules : globalement, ils ont la même structure que chez les Mammifères, mais en beaucoup moins complexe. De plus, la reproduction est saisonnière, et le nombre d'œufs pondus en même temps devra être très important : pour cela, les ovogonies se multiplient chaque années (à l'opposé des Mammifères).
26
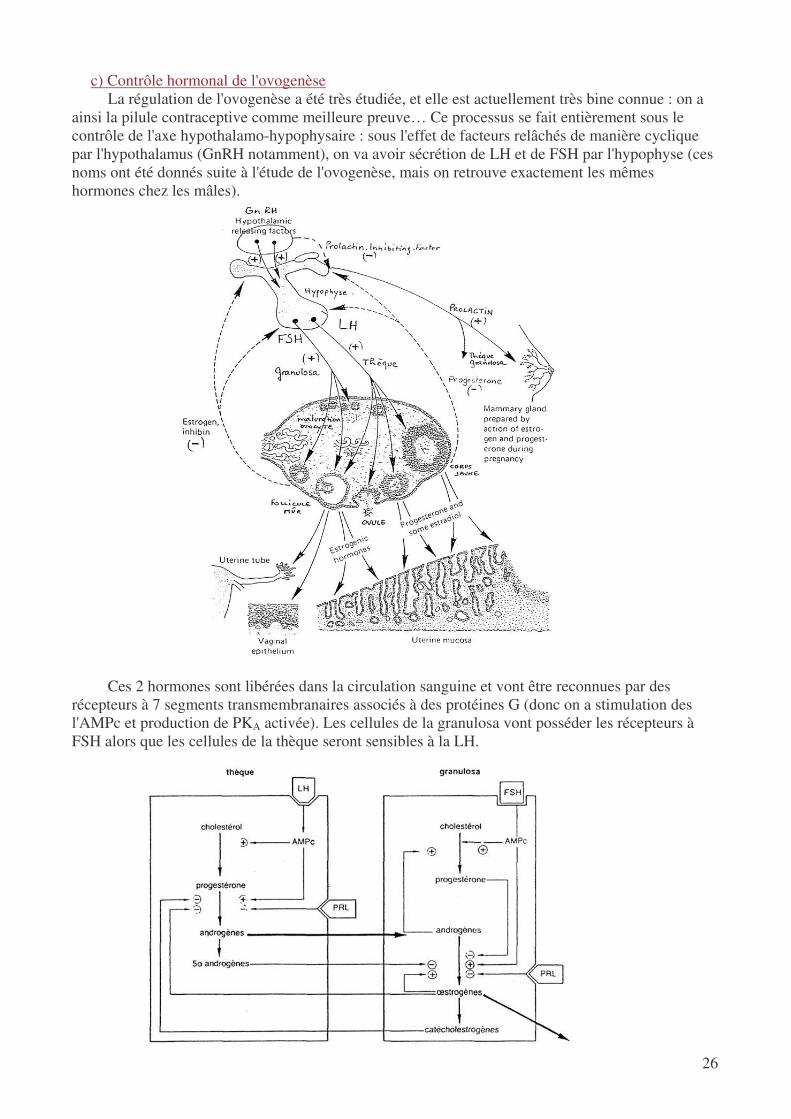
c) Contrôle hormonal de l'ovogenèse La régulation de l'ovogenèse a été très étudiée, et elle est actuellement très bine connue : on a
ainsi la pilule contraceptive comme meilleure preuve… Ce processus se fait entièrement sous le contrôle de l'axe hypothalamo-hypophysaire : sous l'effet de facteurs relâchés de manière cyclique par l'hypothalamus (GnRH notamment), on va avoir sécrétion de LH et de FSH par l'hypophyse (ces noms ont été donnés suite à l'étude de l'ovogenèse, mais on retrouve exactement les mêmes hormones chez les mâles).
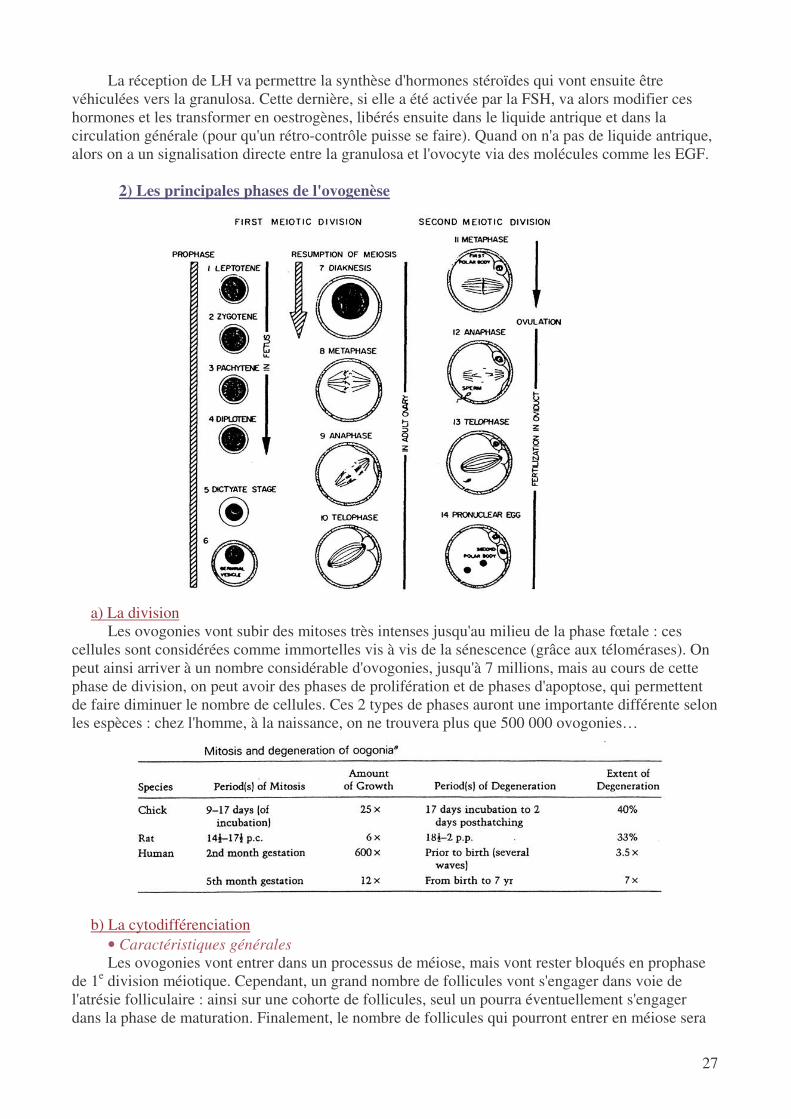
Ces 2 hormones sont libérées dans la circulation sanguine et vont être reconnues par des
récepteurs à 7 segments transmembranaires associés à des protéines G (donc on a stimulation des l'AMPc et production de PKA activée). Les cellules de la granulosa vont posséder les récepteurs à FSH alors que les cellules de la thèque seront sensibles à la LH.
27
La réception de LH va permettre la synthèse d'hormones stéroïdes qui vont ensuite être véhiculées vers la granulosa. Cette dernière, si elle a été activée par la FSH, va alors modifier ces hormones et les transformer en oestrogènes, libérés ensuite dans le liquide antrique et dans la circulation générale (pour qu'un rétro-contrôle puisse se faire). Quand on n'a pas de liquide antrique, alors on a un signalisation directe entre la granulosa et l'ovocyte via des molécules comme les EGF.
2) Les principales phases de l'ovogenèse
a) La division Les ovogonies vont subir des mitoses très intenses jusqu'au milieu de la phase fœtale : ces
cellules sont considérées comme immortelles vis à vis de la sénescence (grâce aux télomérases). On peut ainsi arriver à un nombre considérable d'ovogonies, jusqu'à 7 millions, mais au cours de cette phase de division, on peut avoir des phases de prolifération et de phases d'apoptose, qui permettent de faire diminuer le nombre de cellules. Ces 2 types de phases auront une importante différente selon les espèces : chez l'homme, à la naissance, on ne trouvera plus que 500 000 ovogonies…
b) La cytodifférenciation • Caractéristiques générales Les ovogonies vont entrer dans un processus de méiose, mais vont rester bloqués en prophase
de 1e division méiotique. Cependant, un grand nombre de follicules vont s'engager dans voie de l'atrésie folliculaire : ainsi sur une cohorte de follicules, seul un pourra éventuellement s'engager dans la phase de maturation. Finalement, le nombre de follicules qui pourront entrer en méiose sera

28
assez faible, et au cours d'une vie, le stock d'ovocytes diminue sans arrêt à la fois par les phénomènes d'atrésie et par les ovulations, qui constituent des prélèvements dans le stock…
Il existe un certain nombre de molécules qui empêchent l'atrésie, et elles seront différentes
selon de stade du follicule. Ce sont de manière générale des signaux de survie qui n'atteignent qu'un seul follicule par cohorte. Tout cela fait que le nombre de follicules varie de manière considérable selon qu'on se place chez le fœtus, chez l'embryon une période post-natale.
Les ovocytes qui entrent dans la cytodifférenciation seront toujours bloqués au stade diplotène,
et ce blocage peut durer un temps relativement long : ainsi on a des ovulations qui se manifestent jusqu'à la ménopause, et les ovocytes seront donc restés plus de 50 ans en quiescence. Cepdnant, pendant cette phase, on va avoir accumulation de réserves, mais l'importance de ce stockage va varier selon les espèces.
Chez les Amphibiens, on peut distinguer 2 phases successives d'accroissement de vitesse différentes, mais cela va durer 3 ans et adoptera un aspect saisonnier. Lors de cette croissance, l'œuf va passer de 150µm à 1,5mm de diamètre… On peut remarquer que lorsque l'ovocyte se retrouve au stade pachytène, les chromosomes dans la vésicule germinative (= noyau) sont organisés de manière très particulière : les 2 extrémités de chaque chromosome est ancrée au pôle du noyau tourné vers le centrosome. Cela définit également une zone du cytoplasme très riche en mitochondries : on voit donc que cette organisation très typique confère à l'ovocyte les 1e traces de sa polarité.

29
• La 1e phase d'accroissement L'ovocyte est accroché à l'ovaire et il a un aspect très particulier : en effet, la chromatine des
bivalents forme des écouvillons et à la périphérie du noyau, on retrouve des tâches noires, qui sont en fait des nucléoles. Une cellule quelconque ne contient qu'un seul nucléole mais ici on en retrouve plusieurs dizaines : cela reflète un processus d'amplification de l'ADN ribosomique ADNr.
En effet, chez l'homme, les chromosomes acrocentriques contiennent de l'ADNr qui est en fait
un véritable organisateur nucléolaire : il est composé par des cistrons, cad par des unités de transcriptions. Ces unités sont traduits en un ARNr unique de type 45S et on aura ensuite des phénomènes d'épissage qui permettent la formation des différents ARNr. Chez les Amphibiens, les organisateurs nucléolaires ont subi une phase de multiplication et cela a entraîné du même coup la multiplication des nucléoles. Grâce à ce phénomène, on pourra avoir formation de très nombreux ribosomes, et la traduction pourra donc être très active.

30
L'ARN 5S n'est pas amplifié car il ne se retrouve pas au niveau de ces cistrons : il est en réalité traduit simplement de manière très efficace. Tout cela permet donc une amplification des ARNr tout à fait caractéristique des Amphibiens. Ceci dit, chez toutes les espèces on retrouve un grand nombre de cistrons : en temps normal ils ne sont pas tous utilisés de manière efficace, mais lors de la phase d'accroissement, ils sont utilisés à leur maximum…
L'existence des structures en écouvillon au niveau de la chromatine est reliée elle aussi à une
transcription très importante : en effet, les 2 bivalents se déploient en boucles (jusqu'à 5000 de 200µm chez les Amphibiens), et ce seront alors des zones de transcription très active. Selon le gène, on aura ensuite traduction ou non, selon que l'on a une protéine de réserve ou bien un déterminant maternel. Les gènes codant pour les histones sont transcrits de manière très intensive car la segmentation nécessitera une grande quantité de ces protéines : on a donc formation d'un stock prévisionnel.
Chez les Mammifères, on ne peut pas retrouver ces chromosomes en
écouvillons mais on peut observer une organisation très particulière de la chromatine : on a ainsi apparition de boules qui sont là encore des sites de transcription intense. Finalement, dans les 2 cas, on aura augme,ntation de la transcription, mais c'est un processus qui est poussé à l'extrême chez les Amphibiens.
• La 2e phase d'accroissement Pendant cette phase, on va avoir accumulation du contenu nutritif : ces molécules sont en
général synthétisées par le foie maternel. En effet, les oestrogènes libérés par les cellules folliculaires dans la circulation générale peuvent être captées par le foie : ce dernier va alors synthétiser en réponse de la vitellogénine sérique qui se dimérise.
Cette protéine va alors pénétrer dans l'ovocyte par micropinocytose, et elle sera ensuite clivée en lipovitelline et phosvitine. Ces 2 composés ne restent jamais à l'état libre : ils vont s'amalgamer avec des résidus mitochondriaux pour donner les plaquettes vitellines. Tout cela permet une augmentation très importante de la taille de l'ovocyte.
-Rm- Le vitellus contient toujours de termes lipidiques, glucidiques et protéiques, mais globalement, on ne va retrouver que les 2 molécules citées ci-dessus.
Chez la drosophile, la molécule nutritive principale est également une vitellogénine qui est très semblable à celle de l'Amphibien. Elle est synthétisée là encore par le corps gras, qui est l'équivalent du foie : elle est libérée dans l'hémolymphe puis acheminée et accumulée dans les ovocytes.

31
Finalement, la 1e phase de croissance permet de stocker de très nombreux éléments dont une
grande quantité de messagers maternels, alors que la 2e phase assure le stockage des réserbes. Chez les Mammifères, on retrouve globalement ces 2 mêmes étapes, mais la phase d'accroissement rapide, toute proportion gardée, se fait après la puberté.
c) La maturation
C'est simplement l'achèvement de la méiose : cette division particulière caractérise les cellules germinales, mais dans l'ovogenèse, l'organisation de l'ovule est définitif avant même que la méiose ne commence.
3) L'origine de l'asymétrie de l'ovocyte
On a vu que la phase d'accroissement correspond à une accumulation de très nombreuses
molécules, mais les plaquettes vitellines se retrouvent surtout en partie végétative (au pôle animal, on trouve moins de plaquettes, et elles sont plus petites). Par contre, on retrouve sous la membrane plasmique du pôle animal une accumulation de pigments, et c'est à ce pôle que la vésicule germinative se place. On a donc clairement un œuf asymétrique.
On a longtemps dit que l'on a un gradient de vitellus et un gradient inverse d'ARNm : en réalité, on retrouve une grande quantité d'ARNm au pôle végétatif (quand même moins qu'au pôle animal car on a moins de cytoplasme), et il va d'ailleurs jouer un rôle très important dans l'embryogenèse : ce sont en effet des déterminants maternels.
La surface de l'ovocyte est complexe : en effet, la membrane est garnie de microvillosités, qui ménagent ainsi un espace entre les cellules folliculaires et les ovocyte : c'est un espace où auront lieu de nombreux échanges, et on l'appelle souvent zone pellucide ou espace péri-vitellin. Outre des mitochondries en très grand nombre on peut noter la présence de granules corticaux à la périphérie de l'œuf (moins chez les Amphibiens) : ce sont en fait des vésicules golgiennes contenant une grande quantité d'enzymes, et elles seront exocytées dans l'espace péri-vitellin pendant la fécondation.
32
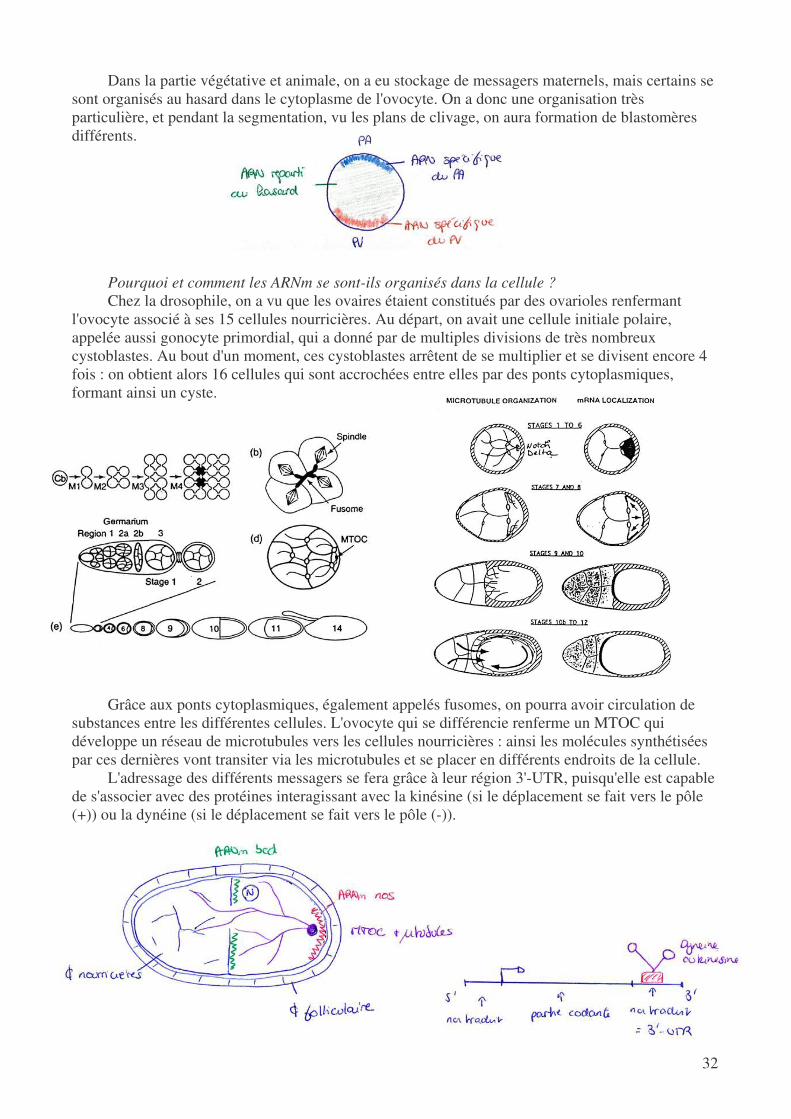
Dans la partie végétative et animale, on a eu stockage de messagers maternels, mais certains se sont organisés au hasard dans le cytoplasme de l'ovocyte. On a donc une organisation très particulière, et pendant la segmentation, vu les plans de clivage, on aura formation de blastomères différents.
Pourquoi et comment les ARNm se sont-ils organisés dans la cellule ? Chez la drosophile, on a vu que les ovaires étaient constitués par des ovarioles renfermant
l'ovocyte associé à ses 15 cellules nourricières. Au départ, on avait une cellule initiale polaire, appelée aussi gonocyte primordial, qui a donné par de multiples divisions de très nombreux cystoblastes. Au bout d'un moment, ces cystoblastes arrêtent de se multiplier et se divisent encore 4 fois : on obtient alors 16 cellules qui sont accrochées entre elles par des ponts cytoplasmiques, formant ainsi un cyste.
Grâce aux ponts cytoplasmiques, également appelés fusomes, on pourra avoir circulation de
substances entre les différentes cellules. L'ovocyte qui se différencie renferme un MTOC qui développe un réseau de microtubules vers les cellules nourricières : ainsi les molécules synthétisées par ces dernières vont transiter via les microtubules et se placer en différents endroits de la cellule.
L'adressage des différents messagers se fera grâce à leur région 3'-UTR, puisqu'elle est capable de s'associer avec des protéines interagissant avec la kinésine (si le déplacement se fait vers le pôle (+)) ou la dynéine (si le déplacement se fait vers le pôle (-)).
33
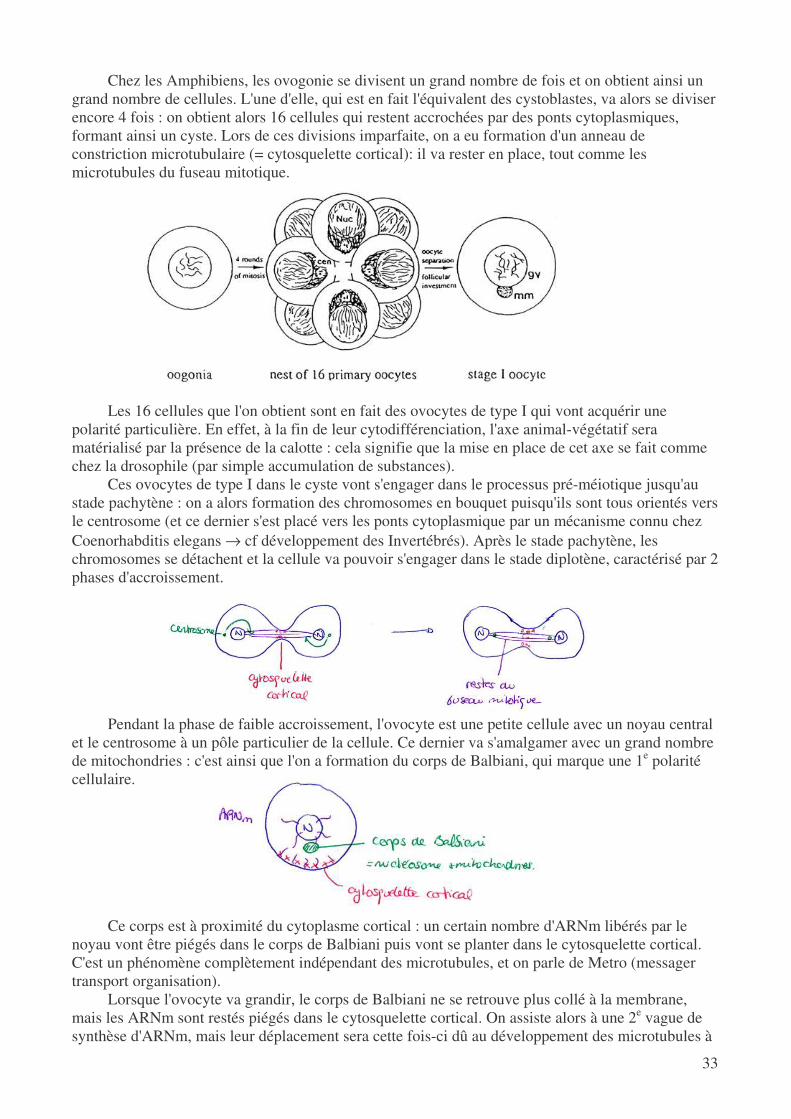
Chez les Amphibiens, les ovogonie se divisent un grand nombre de fois et on obtient ainsi un grand nombre de cellules. L'une d'elle, qui est en fait l'équivalent des cystoblastes, va alors se diviser encore 4 fois : on obtient alors 16 cellules qui restent accrochées par des ponts cytoplasmiques, formant ainsi un cyste. Lors de ces divisions imparfaite, on a eu formation d'un anneau de constriction microtubulaire (= cytosquelette cortical): il va rester en place, tout comme les microtubules du fuseau mitotique.
Les 16 cellules que l'on obtient sont en fait des ovocytes de type I qui vont acquérir une
polarité particulière. En effet, à la fin de leur cytodifférenciation, l'axe animal-végétatif sera matérialisé par la présence de la calotte : cela signifie que la mise en place de cet axe se fait comme chez la drosophile (par simple accumulation de substances).
Ces ovocytes de type I dans le cyste vont s'engager dans le processus pré-méiotique jusqu'au stade pachytène : on a alors formation des chromosomes en bouquet puisqu'ils sont tous orientés vers le centrosome (et ce dernier s'est placé vers les ponts cytoplasmique par un mécanisme connu chez Coenorhabditis elegans → cf développement des Invertébrés). Après le stade pachytène, les chromosomes se détachent et la cellule va pouvoir s'engager dans le stade diplotène, caractérisé par 2 phases d'accroissement.
Pendant la phase de faible accroissement, l'ovocyte est une petite cellule avec un noyau central
et le centrosome à un pôle particulier de la cellule. Ce dernier va s'amalgamer avec un grand nombre de mitochondries : c'est ainsi que l'on a formation du corps de Balbiani, qui marque une 1e polarité cellulaire.
Ce corps est à proximité du cytoplasme cortical : un certain nombre d'ARNm libérés par le
noyau vont être piégés dans le corps de Balbiani puis vont se planter dans le cytosquelette cortical. C'est un phénomène complètement indépendant des microtubules, et on parle de Metro (messager transport organisation).
Lorsque l'ovocyte va grandir, le corps de Balbiani ne se retrouve plus collé à la membrane, mais les ARNm sont restés piégés dans le cytosquelette cortical. On assiste alors à une 2e vague de synthèse d'ARNm, mais leur déplacement sera cette fois-ci dû au développement des microtubules à

34
partir du centrosome. Un certain nombre d'ARNm pourra donc migrer vers le pôle végétatif et le cytosquelette cortical, mais le système est très différent (à partir des microtubules et de la séquence 3'-UTR des ARNm).
Pendant la croissance de l'ovocyte, le corps de Balbiani s'est un peu dissout : un certain nombre
de mitochondries sont restés accrochées au cytosquelette cortical : on a donc formation du futur plasme germinal et la détermination de la lignée germinale est donc déjà commencée.
Pendant la division méiotique, la division sera complètement asymétrique : on a ainsi formation d'une très grosse cellule et d'un globule polaire de très petite taille. Cela permet de conserver la structure et l'organisation de l'ovocyte établies pendant la cytodifférenciation, et c'est l'une des différences majeures avec la spermatogenèse…
Chez la souris, on a pu mettre en évidence des structures qui ressemblent beaucoup au cyste
des drosophiles et des Amphibiens : en effet les ovogonies arrêtent de proliférer à un moment donné, puis se divisent encore 4 fois. On a donc ici un mécanisme très conservé.
On peut se demander pourquoi cette organisation en cyste a été conservée : on ne connaît pas encore la réponse, mais on a pu formuler plusieurs hypothèses. L'une des observations primordiales est que la formation des cystes donne une asymétrie et cela pourrait provoquer une régulation précoce des futurs cellules qui dégénèreront… Cependant chez la souris, on peut se demander quelle est la raison de cette conservation puisque l'ovocyte ne semble pas polarisé…
Les mécanismes d'adressages sont très variables, mais il faut que le messager puisse migrer soit seul, soit par les microtubules soit encore par un flux cytoplasmique. C'est ainsi que certains ARNm se retrouveront à différents pôles de la cellules, mais on peut retrouver ce phénomène dans les cellules somatiques…
4) Déroulement du cycle sexuel chez les femmes
Au cours de la vie d'une femme, on va avoir environ 400 ovulations, donc cela signifie que
seuls 400 ovocytes arriveront à maturité. A partir d'une certain stock d'ovocytes bloqués en prophase de 1e division méiotique, des cohortes vont s'engager dans le processus de maturation et du fort accroissement à partir de la puberté. Cependant, ces différents ovocytes n'achèveront la méiose qu'un an après : la préparation est donc assez lente, et les cellules folliculaires vont y jouer un rôle fondamental.
Le cycle sexuel est toujours contrôlé par l'axe hypothalamo-hypophysaire, mais on peut distinguer plusieurs phases : ainsi on a une phase longue de 300 jours environ (= phase pré-antrique), puis une phase courte de 65 jours (= phase antrique).
a) La phase pré-antrique
Pendant cette phase, l'ovocyte est entouré de plusieurs assises de cellules folliculaires, et tout son développement sera complètement indépendant des gonadotrophines : cette phase n'est donc pas sous le contrôle de l'axe hypothalamo-hypophysaire. Par contre, l'ovocyte lui-même va jouer un rôle important dans la régulation de cette phase : il va en effet préparer les cellules folliculaires à répondre aux gonadotrophines sécrétées pendant la 2e phase du cycle.
On a pu caractériser 2 molécules appartenant à la même famille qui intervenaient dans ce phénomène : ce sont GDF9 et BMP15, toutes 2 de la sous-famille des TGFβ. C'est l'ovocyte qui va

35
sécréter ces molécules en direction des cellules folliculaires, qui, e,n réponse, vont développer et se multiplier. Ainsi, les follicules des souris KO pour les gènes GDF9 ou BMP15 ne se développent jamais.
Quand les cellules de la granulosa et des thèques sont suffisamment nombreuses, ce seront les cellules folliculaires qui prendront le relais : en réponse à la sécrétion de FSH et de LH par l'axe hypothalamo-hypophysaire, de nouvelles molécules seront synthétisées et envoyées vers l'ovocyte…
b) La phase antrique
Tous les ans, une cohorte entière d'ovocytes s'engage dans la phase pré-antrique puis antrique : parmi ce groupe d'ovocyte, seul un pourra entrer dans la phase de maturation. Une partie des autres ovocytes entreront dans un processus apoptotique, mais les autres dégénèreront selon un autre processus. Dans la phase pré-antrique, on distingue souvent différents processus : recrutement, sélection puis dominance, et c'est l'ensemble de ces processus qui permet de ne conserver qu'un seul ovocyte qui s'engagera dans la méiose.
Pendant la phase antrique, on peut voir que la FSH devient très abondante : elle va alors intervenir dans les processus de sélection et de dominance. De plus, cette sécrétion s'accompagne de plusieurs autres phénomènes : en réponse à la FSH, les cellules de la granulosa peuvent sécréter des oestrogènes, qui exerceront un rétro-contrôle négatif sur l'axe. De plus, on aura une signalisation en direction de la thèque, ce qui stimule la transformation du cholestérol en androgènes.
D'autre part, lorsque l'on peut observer le pic de FSH, on voit apparaître des récepteurs à LH sur les cellules de la granulosa, et les récepteurs à la FSH deviennent plus abondants. Finalement, pendant toute cette 1e partie du cycle, on a synthèse de plus en plus d'œstrogènes qui exercent un rétro-contrôle négatif, mais on a aussi de plus en plus de récepteurs aux hormones.
Cela signifie que le rapport récepteur/FSH augmente de manière importante : à un moment donné, on n'aura plus assez de FSH pour tous les follicules de la cohorte. Par hasard, l'un des follicules exprimait suffisamment de récepteurs pour le peu de FSH : c'est alors que se met en place la dominance de ce follicule sur les autres, et ces derniers rentreront en atrésie.
De manière un peu décalée, on observe un pic de LH : or; les cellules de la granulosa ont acquis des récepteurs à la LH, et elles vont alors sécréter en réponse des substances vers l'ovocyte, qui pourra terminer sa maturation et entrer en méiose.
5) Déclenchement de la maturation
a) Blocage et reprise de la 1e division méiotique
• Situation pendant la phase pré-antrique On peut se demander pourquoi les ovocytes sont bloqués en prophase de 1e division méiotique,
mais aussi comment ils sont débloqués et peuvent terminer leur maturation. Pincus a pris des follicules de De Graaf qu'il a débarrassé des assises folliculaires : l'ovocyte
entre alors immédiatement en méiose. C'est donc qu'a priori, les cellules folliculaires exercent un contrôle négatif sur l'entré en méiose, mais si c'est évident chez les Mammifères, le processus ne semble pas si simple chez les Amphibiens. En effet chez ces derniers, même si on enlève les cellules folliculaires, l'ovocyte ne peut pas entrer en méiose.
On sait que les cellules folliculaires ne sont pas en contact étroit avec l'ovocyte puisque ce dernier possède des micro-villosités. Entre ces 2 types de cellules, on retrouve ainsi la zone pellucide, qui présente à la fois les micro-villosités de l'ovocyte et des pieds émis par les cellules folliculaires. Les cellules folliculaire sont reliées à l'ovocyte par des GAP jonctions abondantes dans leurs pieds : les molécules qui pourront passer librement d'une cellule à l'autre ne pourront pas dépasser 1kDa.
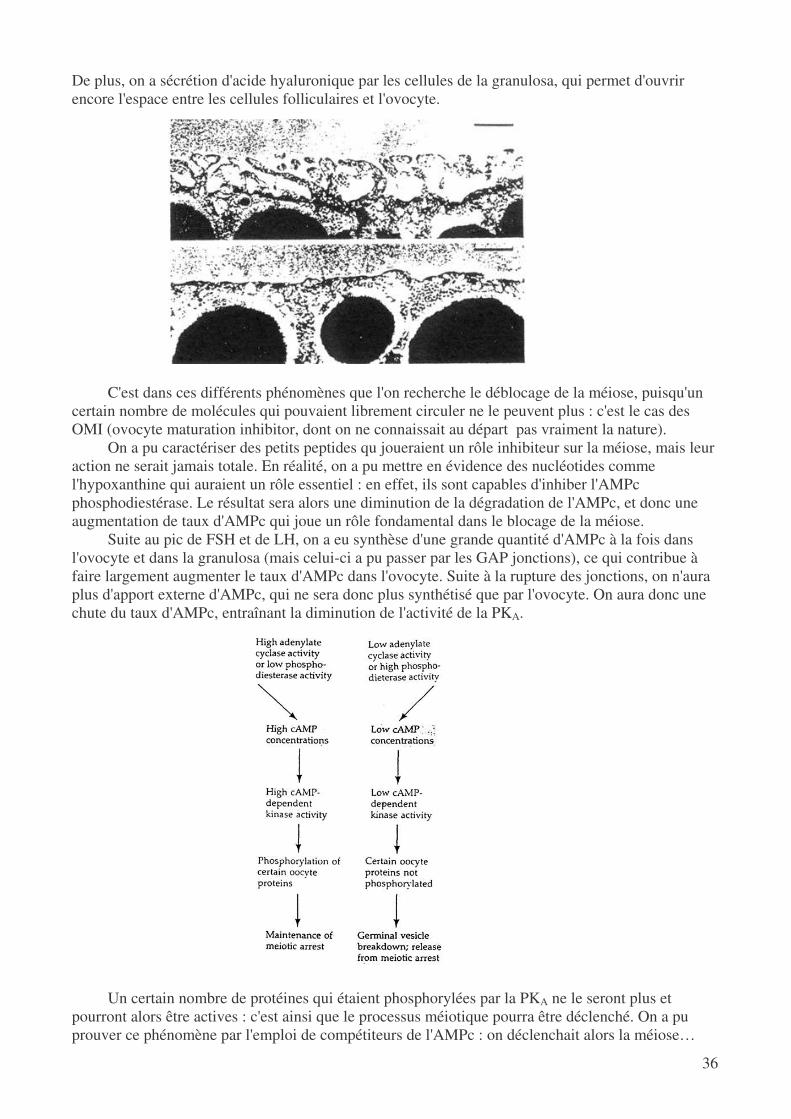
• Situation pendant la phase antrique Quand on assiste au pic de LH, à la fois les cellules de la thèque et de la granulosa vont réagir:
on a alors rupture des GAP jonctions avec l'ovocyte et disparition des micro-villosités de l'ovocyte.
36
De plus, on a sécrétion d'acide hyaluronique par les cellules de la granulosa, qui permet d'ouvrir encore l'espace entre les cellules folliculaires et l'ovocyte.
C'est dans ces différents phénomènes que l'on recherche le déblocage de la méiose, puisqu'un
certain nombre de molécules qui pouvaient librement circuler ne le peuvent plus : c'est le cas des OMI (ovocyte maturation inhibitor, dont on ne connaissait au départ pas vraiment la nature).
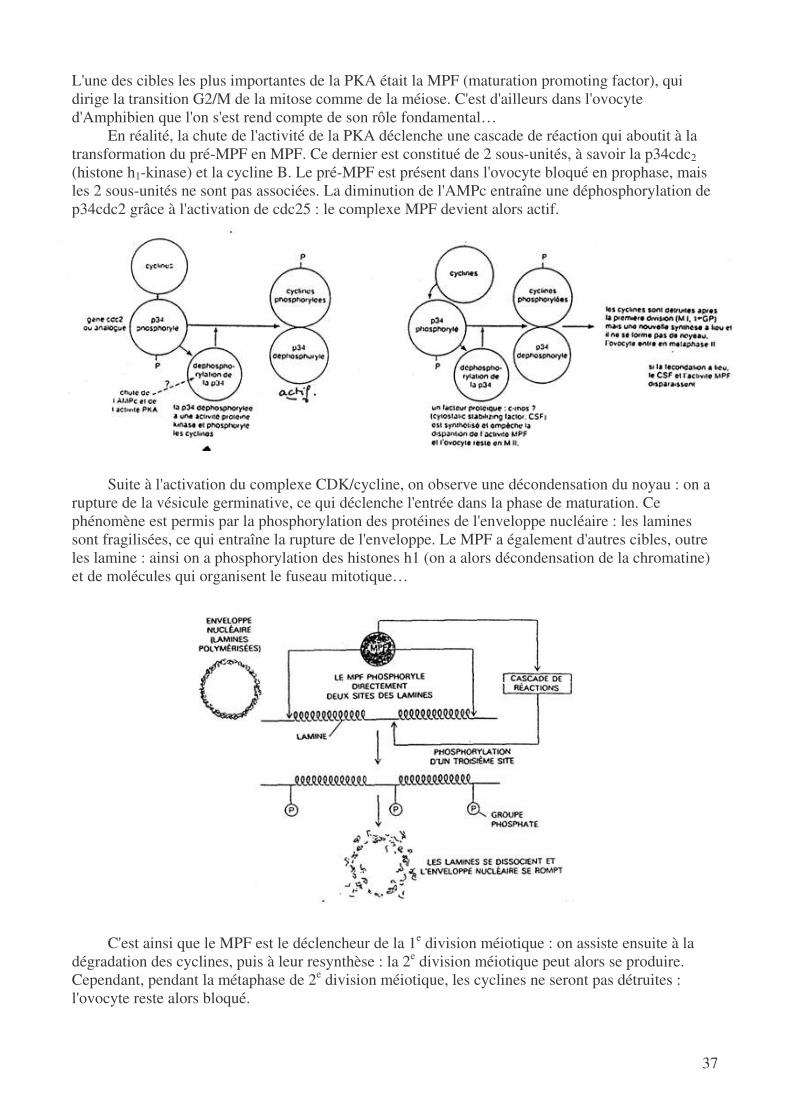
On a pu caractériser des petits peptides qu joueraient un rôle inhibiteur sur la méiose, mais leur action ne serait jamais totale. En réalité, on a pu mettre en évidence des nucléotides comme l'hypoxanthine qui auraient un rôle essentiel : en effet, ils sont capables d'inhiber l'AMPc phosphodiestérase. Le résultat sera alors une diminution de la dégradation de l'AMPc, et donc une augmentation de taux d'AMPc qui joue un rôle fondamental dans le blocage de la méiose.
Suite au pic de FSH et de LH, on a eu synthèse d'une grande quantité d'AMPc à la fois dans l'ovocyte et dans la granulosa (mais celui-ci a pu passer par les GAP jonctions), ce qui contribue à faire largement augmenter le taux d'AMPc dans l'ovocyte. Suite à la rupture des jonctions, on n'aura plus d'apport externe d'AMPc, qui ne sera donc plus synthétisé que par l'ovocyte. On aura donc une chute du taux d'AMPc, entraînant la diminution de l'activité de la PKA.
Un certain nombre de protéines qui étaient phosphorylées par la PKA ne le seront plus et
pourront alors être actives : c'est ainsi que le processus méiotique pourra être déclenché. On a pu prouver ce phénomène par l'emploi de compétiteurs de l'AMPc : on déclenchait alors la méiose…
37
L'une des cibles les plus importantes de la PKA était la MPF (maturation promoting factor), qui dirige la transition G2/M de la mitose comme de la méiose. C'est d'ailleurs dans l'ovocyte d'Amphibien que l'on s'est rend compte de son rôle fondamental…
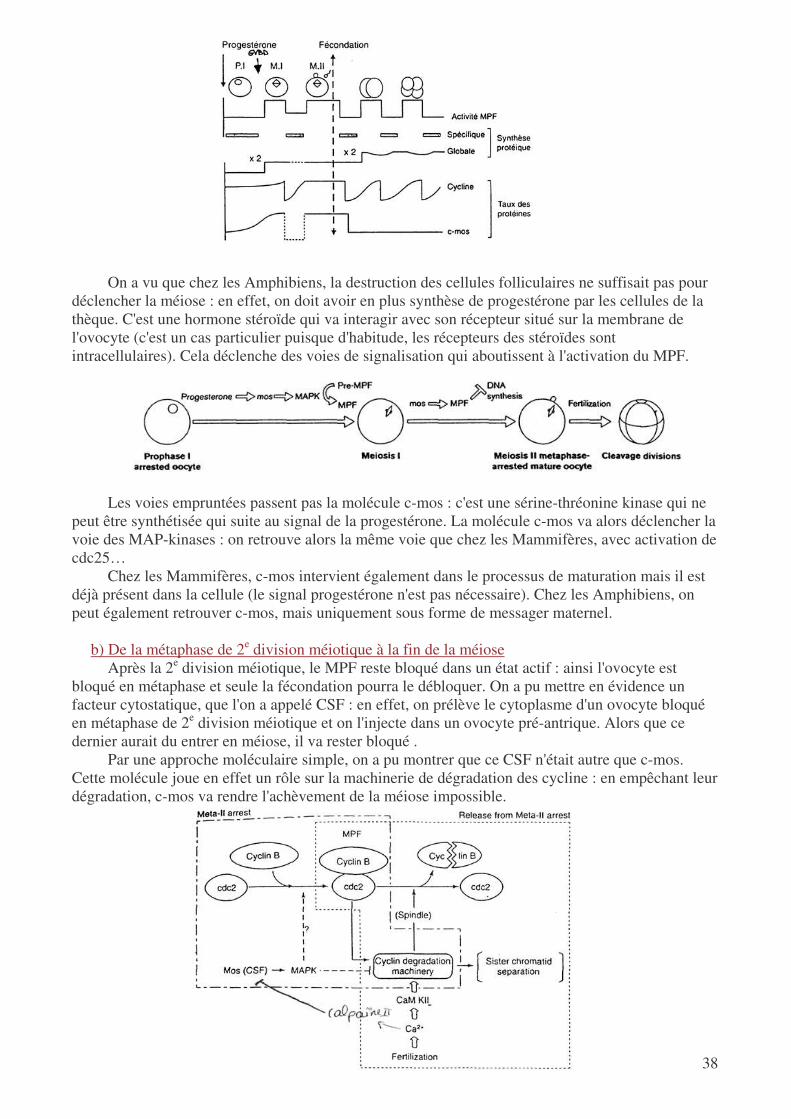
En réalité, la chute de l'activité de la PKA déclenche une cascade de réaction qui aboutit à la transformation du pré-MPF en MPF. Ce dernier est constitué de 2 sous-unités, à savoir la p34cdc2 (histone h1-kinase) et la cycline B. Le pré-MPF est présent dans l'ovocyte bloqué en prophase, mais les 2 sous-unités ne sont pas associées. La diminution de l'AMPc entraîne une déphosphorylation de p34cdc2 grâce à l'activation de cdc25 : le complexe MPF devient alors actif.
Suite à l'activation du complexe CDK/cycline, on observe une décondensation du noyau : on a
rupture de la vésicule germinative, ce qui déclenche l'entrée dans la phase de maturation. Ce phénomène est permis par la phosphorylation des protéines de l'enveloppe nucléaire : les lamines sont fragilisées, ce qui entraîne la rupture de l'enveloppe. Le MPF a également d'autres cibles, outre les lamine : ainsi on a phosphorylation des histones h1 (on a alors décondensation de la chromatine) et de molécules qui organisent le fuseau mitotique…
C'est ainsi que le MPF est le déclencheur de la 1e division méiotique : on assiste ensuite à la
dégradation des cyclines, puis à leur resynthèse : la 2e division méiotique peut alors se produire. Cependant, pendant la métaphase de 2e division méiotique, les cyclines ne seront pas détruites : l'ovocyte reste alors bloqué.
38
On a vu que chez les Amphibiens, la destruction des cellules folliculaires ne suffisait pas pour
déclencher la méiose : en effet, on doit avoir en plus synthèse de progestérone par les cellules de la thèque. C'est une hormone stéroïde qui va interagir avec son récepteur situé sur la membrane de l'ovocyte (c'est un cas particulier puisque d'habitude, les récepteurs des stéroïdes sont intracellulaires). Cela déclenche des voies de signalisation qui aboutissent à l'activation du MPF.
Les voies empruntées passent pas la molécule c-mos : c'est une sérine-thréonine kinase qui ne
peut être synthétisée qui suite au signal de la progestérone. La molécule c-mos va alors déclencher la voie des MAP-kinases : on retrouve alors la même voie que chez les Mammifères, avec activation de cdc25…
Chez les Mammifères, c-mos intervient également dans le processus de maturation mais il est déjà présent dans la cellule (le signal progestérone n'est pas nécessaire). Chez les Amphibiens, on peut également retrouver c-mos, mais uniquement sous forme de messager maternel.
b) De la métaphase de 2e division méiotique à la fin de la méiose
Après la 2e division méiotique, le MPF reste bloqué dans un état actif : ainsi l'ovocyte est bloqué en métaphase et seule la fécondation pourra le débloquer. On a pu mettre en évidence un facteur cytostatique, que l'on a appelé CSF : en effet, on prélève le cytoplasme d'un ovocyte bloqué en métaphase de 2e division méiotique et on l'injecte dans un ovocyte pré-antrique. Alors que ce dernier aurait du entrer en méiose, il va rester bloqué .
Par une approche moléculaire simple, on a pu montrer que ce CSF n'était autre que c-mos. Cette molécule joue en effet un rôle sur la machinerie de dégradation des cycline : en empêchant leur dégradation, c-mos va rendre l'achèvement de la méiose impossible.

39
Une fois pondu, l'ovocyte se retrouve dans les trompes et va être éventuellement fécondé. Les spermatozoïdes vont passer à travers la zone pellucide grâce à leur acrosome et vont fusionner avec l'ovule : cela provoque une vague calcique et l'ovocyte est activé. L'augmentation de la concentration intracellulaire en Ca2+ aboutit entre autre à la dégradation des cyclines. En effet, le calcium active une calmoduline (la CaM-KII) qui active elle-même la voie de dégradation des cyclines et qui permet la dégradation de c-mos. Finalement, on aura donc une activation et plus d'inhibition, et les cyclines pourront être dégradées et le cycle cellulaire pourra se terminer.
Finalement, l'ovocyte ne pourra terminer sa maturation que s'il est fécondé. Cependant, il peut
également s'activer tout seul de manière anomale, et on parle alors de parthénogenèse : on obtient alors un embryon normal… La molécule c-mos permet de bloquer l'activation des ovocyte, et pour cela on le considère comme un garde-fou qui empêche la parthénogenèse. Certains auteurs pensent même que nos ancêtres n'effectuaient que la parthénogenèse, mais qu'au cours de l'évolution, la fécondation a été favorisée pour le brassage génétique.
On peut reprendre en détail l'expulsion de l'ovocyte depuis son follicule. Quand les cellules de
la granulosa réagissent à la LH, elles envoient des signaux vers l'ovocyte et synthétisent de l'acide hyaluronique. Le tout est stocké dans l'antrum, et cela va provoquer un fort gonflement de cette cavité. Les cellules de la granulosa vont même finir par perdre le contact avec l'ovocyte, et le cumulus oophorus va se rompre. L'ovulation intervient quand le follicule éclate suite à la trop grande augmentation de volume. Cela pourra se faire à différents moments et à des tailles différentes, selon l'espèce envisagée.
La granulosa va sécréter plusieurs types de molécules, dont des prostaglandines : elles assurent
alors la synthèse d'un activateur de plasminogène. Pendant l'ovulation, ce plasminogène va se transformer en plasmine, qui est une métalloprotéase. Elle est capable de dégrader le collagène de type IV, que l'on retrouve au niveau de la membrane de Slavjanski, cad au niveau de la lame basale

40
qui sépare la thèque de la granulosa. Finalement, on aura fragilisation de l'enveloppe folliculaire de l'ovocyte, et elle ne pourra pas résister à l'augmentation de volume de l'antrum : c'est ainsi que l'ovulation peut se faire.
On vient donc de voir que l'ovogenèse est un phénomène complexe puisque c'est un processus
discontinu avec des phases de prolifération très périodiques, des apoptoses, des atrésies folliculaires et des arrêts dans le cycle de l'ovocyte. D'autre part, tous les processus que l'on peut observer pendant l'ovogenèse contribuent à la survie d'une des follicules, qui sera le seul à arriver à maturité.
De plus, l'ovogenèse s'accompagne de l'accumulation d'une quantité plus ou moins grande de réserves, et ce de manière très organisée. Lors de la méiose, on aura formation de globules polaires suite à des divisions très asymétriques, afin de conserver l'organisation des réserves dans l'ovocyte. Certains ARNm stockés seront traduits très rapidement, comme c'est le cas pour c-mos, qui est traduit dès la méiose. Par contre, d'autres ne seront traduits que pendant l'embryogenèse : on a donc une machinerie de contrôle très complexe.
Enfin, on peut observer des alternances de très nombreux facteurs moléculaires, et les cycline,s qui assurent en général le bon déroulement de la division, vont ici provoquer des blocages.
6) Quelques remarques
On vient de voir le déroulement classique de l'ovogenèse, mais il faut savoir que de nombreux
paramètres vont varier selon l'espèce envisagée… Par exemple, l'ovocyte est capable de synthétiser des récepteurs à LH : dans ce cas, le déroulement de l'ovogenèse est beaucoup plus complexe que ce qu'on vient d'étudier. Ainsi, le pic de LH déclencherait directement des variation du taux d'AMPc, mais tous les éléments ne sont pas encore connus.
D'autre part, l'ovulation ne se fait pas forcément au même moment du cycle de l'ovocyte chez toutes les espèces. Pour cela, les mécanismes vus pour les Vertébrés ne sont pas forcément applicables chez tous les groupes.

41
De plus, la taille de l'œuf va varier selon la quantité de réserves mais aussi la localisation du vitellus. En plus, l'ovocyte est souvent protégé par des cellules folliculaires, qui ne sont pas directement en contact. Par exemple, chez la souris, on retrouve entre les 2 types cellulaires une zone pellucide avec des protéines glycosylées.
Chez les Amphibiens, la membrane de l'ovocyte est
protégée par le chorion, appelée aussi membrane vitelline, et l'œuf va encore s'entourer d'une gangue polysaccharidique sèche qui va s'hydraté une fois dans l'eau. La constitution de cette gangue est spécifique et permet de limiter la fécondation inter-espèce.
Chez la drosophile, l'ovocyte est entouré par la membrane vitelline et le chorion sera cette fois-
ci une couille supplémentaire synthétisée par l'ovocyte et les cellules folliculaires. Chez les Mammifères, le chorion correspond à la partie ectodermique du placenta (= trophectoderme). On trouvera en plus une enveloppe plasmique, la zone pellucide et la corona radiata.
Chez les Oiseaux, l'ovocyte est également bien
protégé : on trouve ainsi une membrane vitelline, le blanc d'œuf qui se met en place pendant le trajet dans les oviductes, 2 membranes coquillières et la coquille calcaire en dernier lieu.
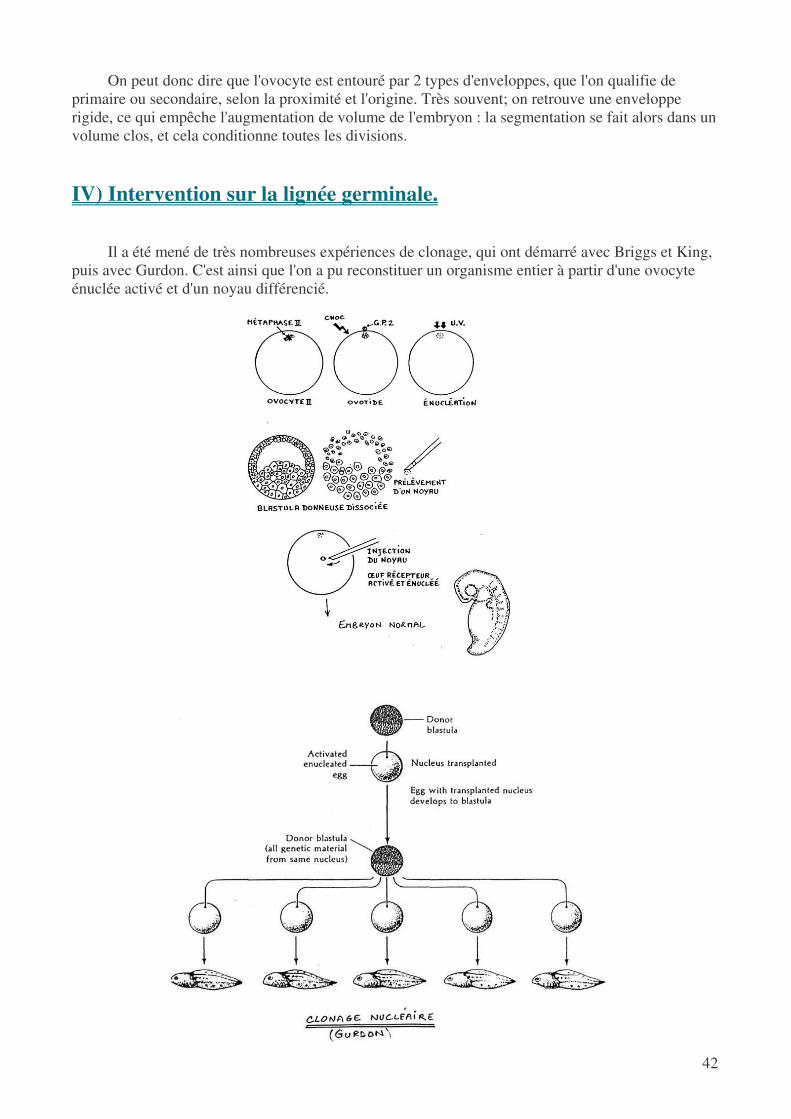
42
On peut donc dire que l'ovocyte est entouré par 2 types d'enveloppes, que l'on qualifie de primaire ou secondaire, selon la proximité et l'origine. Très souvent; on retrouve une enveloppe rigide, ce qui empêche l'augmentation de volume de l'embryon : la segmentation se fait alors dans un volume clos, et cela conditionne toutes les divisions.
IV) Intervention sur la lignée germinale. Il a été mené de très nombreuses expériences de clonage, qui ont démarré avec Briggs et King,
puis avec Gurdon. C'est ainsi que l'on a pu reconstituer un organisme entier à partir d'une ovocyte énuclée activé et d'un noyau différencié.

43
D'autre part, les manipulation sur la lignée germinales ont pu mettre en évidence l'empreinte génomique parentale (EGP). En effet, MacGrath et Solter ont formé des œufs à partir de différentes combinaisons de pronuclei (1 maternel et un paternel ou 2 pronuclei de la même provenance), mais seuls les ovocytes "maternel+paternel" ont pu se développer. C'est ainsi qu'ils en ont déduit que certains phénotypes, contrôlés par un gène ou un groupe de gène, se manifestent en fonction de l'origine parentale du gène (ou du groupe de gène) en question alors que le génotype est identique.
Par exemple, dans l'espèce humaine, certaines maladies sont dues à ce phénomène : certains hétérozygotes peuvent présenter une maladie même si elle est récessive. Les génomes parentaux sont donc fonctionnellement différents et on dit alors qu'ils ont reçu une empreinte : on pense que l'EGP est due à des méthylations.
Cette EGP est transmissible, n'intervient que sur une dizaine de zones chez l'homme mais elle est aussi temporaire. Pendant la gamétogenèse, l'empreinte génomique est détruite pour être remplacée : ainsi c'est l'empreinte maternelle qui est détruite chez les mâles, afin que les gamètes ne portent qu'une empreinte paternelle. Tout cela prouve que certains caractères peuvent être transmis suite à une modification épigénétique.
La méthylation intervient également dans l'inhibition de l'un des 2 chromosomes X : chez les femelles, on voit les 2 X s'associer en bivalents puis subir la même ségrégation que les autosomes, alors que chez les mêles, les chromosomes sexuels se placent dans une vésicule à l'état condensé, ce qui évite d'éventuels crossing-over… Au moment de l'implantation de l'œuf, si on a 2X, l'un des 2 chromosomes X sera inhibé de manière aléatoire, selon les lignées cellulaires : c'est ce phénomène qui est à l'origine des tâches des chats…
On peut tout de même se demander quel est le rôle de l'EGP et comment elle est véritablement remise en place à chaque gamétogenèse. On sait que ce sont des méthylases qui jnterviennent, mais on ne connaît pas tous les phénomènes.
Penser à faire le parallèle entre spermatogenèse et gamétogenèse :
- points communs : cellules mises à part dès le début de l'embryogenèse, rapport étroit entre ovocyte/cellules folliculaires et spermatocyte/cellule de Sertoli…
- différences : méiose symétrique ou asymétrique, processus continu ou discontinu…


















