L’endothéline: pharmacologie cardiovasculaire et ...
12
L es mécanismes qui régissent les propriétés paracrines et endocrines de l’endothélium vasculaire ont été analysés dans plusieurs milliers de publications scientifiques depuis le début des années 1980. Il y a dix ans, qui aurait cru que cette simple monocouche cellulaire qui tapisse tous les vaisseaux sanguins joue un rôle si important dans la phy- siopathologie du système circulatoi- re ? En fait, sa grande surface de dis- tribution, sa localisation anatomique et les facteurs humoraux qu’elle sécrète confèrent à cette couche cel- lulaire un rôle de premier plan dans les mécanismes d’homéostasie du tonus vasculaire et dans certaines anomalies du système circulatoire. Le premier facteur endothélial connu a été la prostacycline, un inhi- biteur de l’agrégation des plaquettes sanguines qui est en outre un puis- sant modulateur du tonus vasculaire systémique et pulmonaire [1, 2]. L’étude classique de Robert Furch- gott a démontré par la suite que l’endothélium sécrète un facteur relaxant autre que les prostanoïdes, l’EDRF (endothelium derived relaxing fac- tor) qui s’est révélé être le monoxyde 563 L’endothéline: pharmacologie cardiovasculaire et considérations physiopathologiques Les progrès dans la connaissance du système des endothé- lines, puissants peptides vasomoteurs produits par l’endo- thélium vasculaire, ont été spectaculaires, depuis leur bio- synthèse grâce à l’enzyme de conversion de l’endothéline (ECE), jusqu’à la pharmacologie de leurs récepteurs, ET A et ET B . La présence d’activités enzymatiques distinctes de l’ECE spécifiques de certains organes fait envisager le développement d’inhibiteurs sélectifs. Les récepteurs sur les cellules musculaires lisses vasculaires et sur les cellules endothéliales relaient, respectivement, des actions vaso- constrictrices (surtout ET A ) et vasodilatatrices (essentielle- ment ET B ). L’étude de la contribution des endothélines à la régulation du système cardiovasculaire et à l’étiologie de certaines affections vasculaires est l’objet d’une forte com- pétition car ces mises à jour pourraient conduire au déve- loppement d’agents thérapeutiques efficaces. ADRESSE P. D’Orléans-Juste, R. Leduc : chercheurs boursiers du Fonds de Recherche en Santé du Québec. J-P. Gratton, A. Claing : étudiants en thèse. Département de pharmacologie. G. Bkaily : chercheur boursier du Fonds de Recherche en Santé du Québec . Département de physio- logie et biophysique, Faculté de médecine, Université de Sherbrooke, Sherbrooke, Québec, J1H 5N4 Canada. SYNTHÈSE médecine/sciences 1996 ; 12 : 563-74 m/s n° 5, vol. 12, mai 96 Pedro D’Orléans-Juste Jean-Philippe Gratton Richard Leduc Ghassan Bkaily Audrey Claing
Transcript of L’endothéline: pharmacologie cardiovasculaire et ...

Les mécanismes qui régissentles propriétés paracrines etendocrines de l’endothéliumvasculaire ont été analysésdans plusieurs milliers de
publications scientifiques depuis ledébut des années 1980.Il y a dix ans, qui aurait cru que cettesimple monocouche cellulaire quitapisse tous les vaisseaux sanguinsjoue un rôle si important dans la phy-siopathologie du système circulatoi-re ? En fait, sa grande surface de dis-tribution, sa localisation anatomiqueet les facteurs humoraux qu’ellesécrète confèrent à cette couche cel-
lulaire un rôle de premier plan dansles mécanismes d’homéostasie dutonus vasculaire et dans certainesanomalies du système circulatoire.Le premier facteur endothélialconnu a été la prostacycline, un inhi-biteur de l’agrégation des plaquettessanguines qui est en outre un puis-sant modulateur du tonus vasculairesystémique et pulmonaire [1, 2].L’étude classique de Robert Furch-gott a démontré par la suite quel’endothélium sécrète un facteurrelaxant autre que les prostanoïdes,l’EDRF (endothelium derived relaxing fac-tor) qui s’est révélé être le monoxyde
563
L’endothéline :pharmacologiecardiovasculaireet considérationsphysiopathologiques
Les progrès dans la connaissance du système des endothé-lines, puissants peptides vasomoteurs produits par l’endo-thélium vasculaire, ont été spectaculaires, depuis leur bio-synthèse grâce à l’enzyme de conversion de l’endothéline(ECE), jusqu’à la pharmacologie de leurs récepteurs, ETAet ETB. La présence d’activités enzymatiques distinctes del’ECE spécifiques de certains organes fait envisager ledéveloppement d’inhibiteurs sélectifs. Les récepteurs surles cellules musculaires lisses vasculaires et sur les cellulesendothéliales relaient, respectivement, des actions vaso-constrictrices (surtout ETA) et vasodilatatrices (essentielle-ment ETB). L’étude de la contribution des endothélines à larégulation du système cardiovasculaire et à l’étiologie decertaines affections vasculaires est l’objet d’une forte com-pétition car ces mises à jour pourraient conduire au déve-loppement d’agents thérapeutiques efficaces.
ADRESSEP. D’Orléans-Juste, R. Leduc : chercheursboursiers du Fonds de Recherche en Santé duQuébec. J-P. Gratton, A. Claing : étudiants enthèse. Département de pharmacologie. G.Bkaily : chercheur boursier du Fonds de Rechercheen Santé du Québec . Département de physio-logie et biophysique, Faculté de médecine,Université de Sherbrooke, Sherbrooke,Québec, J1H 5N4 Canada.
SYNTHÈSEmédecine/sciences 1996 ; 12 : 563-74
m/s n° 5, vol. 12, mai 96
Pedro D’Orléans-JusteJean-Philippe GrattonRichard LeducGhassan BkailyAudrey Claing

d’azote (NO). Le rôle essentiel de cefacteur labile dans les propriétésvasodilatatrices de plusieurs agents,tels l’acétylcholine, la substance P etla bradykinine, a été démontré dansdes vaisseaux isolés de plusieursespèces [3, 4].Toutefois, il ne s’agit là que de deuxdes autacoïdes qui sont sécrétés defaçon majoritairement constitutivepar l’endothélium. Plusieurs autresfacteurs ont été identifiés à partir descellules endothéliales en culture(PAF (platelet activating factor), throm-boxane, prostaglandine E2, interleu-kines, facteurs de croissance) (figure1). L’influence de tous ces facteurssur les fonctions sécrétoires del’endothélium est complexe etimportante à la fois puisqu’elle sug-gère que cette couche cellulaire pos-sède ses propres mécanismes d’auto-régulation.
La découvertede l’endothéline
Après la découverte de la prostacycli-ne et de l’EDRF, l’endothélium nor-mal fut longtemps perçu comme unensemble homogène de cellulessécrétrices impliquées dans des phé-nomènes de vasodilatation.En 1988, Masashi Yanagisawa extra-yait, à partir de 40 litres de surna-geants de cellules endothélialesd’aorte porcine en culture, un fac-teur thermolabile d’origine pepti-dique qui avait pour propriétéd’induire une vasoconstriction trèsimportante de l’artère coronaire por-cine [5]. Intéressés par les puissantespropriétés vasoconstrictrices de cefacteur, Yanagisawa et al. ont poursui-vi l’analyse biochimique moléculaireet génétique de ce peptide alorsappelé endothéline-1 (ET-1) [5]. Parla suite, deux isoformes de l’ET-1furent identifiées, l’ET-2 et l’ET-3[6]. Il fut démontré que les troisendothélines sont codées par troisgènes distincts et qu’elles sont engen-drées par un processus enzymatiqueidentique [6, 7].
Dès 1988, nous avons rapporté quel’ET-1 induit une hypotension transi-toire suivie d’un effet vasopresseurde longue durée lorsqu’elle estadministrée par voie intraveineusechez le rat anesthésié. Dans le cadrede cette première étude faite dans lelaboratoire de Sir John Vane auWilliam Harvey Research Institute(Londres), nous avions, en effet, rap-porté que l’endothéline agissait surl’endothélium lui-même en stimu-lant la libération d’autacoïdes, telsque le NO et/ou la prostacycline,qui agissent comme antagonistesphysiologiques et autocrines deseffets vasoconstricteurs puissants dupeptide [8].Contrairement aux cas d’autres pep-tides vasoactifs, tels les kinines, lesneurokinines et l’angiotensine, lagénétique de l’endothéline et de sesrécepteurs s’est développée deconcert avec les études fonctionnelles(pharmacologique, biochimique etimmunologique), ce qui a permis decorréler entre eux plusieurs conceptspréalablement démontrés dans dessystèmes pharmacologiques ou bio-chimiques à des études de biologiemoléculaire.
Voies de synthèsede l’endothéline
Rôle de la furine dans les voies desynthèse des endothélines
Nous avons récemment démontréqu’une convertase de type subtilisi-ne, la furine, pouvait être respon-sable du clivage de la pré-proET-1 enbig-ET-1 [9] (figure 2A). Puis, uneenzyme de conversion de l’endothéli-ne engendre l’ET-1 et un fragmentcarboxy-terminal inactif, la big-ET-1(22-38). Comme il est montré dans lafigure 2B, la protéine synthétisée dansdes cellules BSC-40 à partir del’ADNc de la pré-proET-1, induit deseffets pharmacologiques sur l’artèrecarotide de lapin après incubationavec la furine et la chymosine. Dansce dernier système, la protéine pro-duite voit son effet vasoconstricteuraboli par un antagoniste sélectif desrécepteurs ETA, le BQ-123 [10] (voirTableau I). Fait intéressant, la furineest constitutivement synthétisée dansles cellules endothéliales d’aortebovine [11].
564 m/s n° 5, vol. 12, mai 96
RÉFÉRENCES1. Vane JR, Botting RM. Control of the cir-culation by vasoactive mediators from endo-thelial cells. J Lip Med Cell Signal 1994 ; 9 : 1-8.
2. Corvol P, Alhenc-Gélas F, Soubrier F.L’endothélium, site de production et demétabolisme des peptides vaso-actifs. médeci-ne/sciences 1993 ; 9 : 1050-61.
3. Furchgott RF, Zawadzky JV. The obligato-ry role of endothelial cells in the relaxationof arterial smooth muscle by acetylcholine.Nature 1980 ; 288 : 373-6.
4. D’Orléans-Juste P, Dion S, Mizrahi J, Re-goli D. Effects of peptides and non-peptideson isolated arterial smooth muscles : role ofendothelium. Eur J Pharmacol 1985 ; 114 : 9-21.
5. Yanagisawa M, Kurihara H, Kimura S, To-mobe Y, Kobayashi M, Mitsui Y, Yazaki Y,Goto K, Masaki T. A novel potent vasocons-trictor peptide produced by vascular endo-thelial cells. Nature 1988 ; 332 : 411-5.
6. Inoue A, Yanagisawa M, Kimura S, KasuyaY, Miyauchi T, Goto K, Masaki T. The hu-man endothelin family : three structurallyand pharmacologically distinct isopeptidespredicted by three separate genes. Proc NatlAcad Sci USA 1989 ; 86 : 2863-7.
7. Lotersztajn S. Les endothélines. médeci-ne/sciences 1993 ; 9 : 1084-93.
8. De Nucci G, Thomas R, D’Orléans-JusteP, Antunes E, Walder C, Warner TD, VaneJR. Pressor effects of circulating endothelinare limited by its removal in the pulmonarycirculation and by the release of prostacy-clin and endothelium-derived relaxing fac-tor. Proc Natl Acad Sci USA 1988 ; 85 : 9797-800.
9. Denault JB, Claing A, D’Orléans-Juste P,Sawamura T, Kido T, Masaki T, Leduc R.Processing of proendothelin-1 by human fu-rin convertase. FEBS Lett 1995 ; 362 : 276-80.
10. Ihara M, Noguchi K, Saeki T, FukurodaT, Tsuchida S, Kimura S, Fukami T, Ishika-wa K, Nishikibe M, Yano M. Biological pro-files of highly potent novel endothelin anta-gonists selective for the ETA receptor. LifeSci 1992 ; 50 : 247-55.
11. Laporte S, Denault JB, D’Orléans-JusteP, Leduc R. Presence of furin mRNA in cul-tured bovine endothelial cells and possibleinvolvement of furin in the processing ofthe endothelin precursor. J Cardiovasc Phar-macol 1993 ; 22 : S7-10.
pré-proET → big-ET → ET$ $
endoprotéases (?) enzyme de+ conversion de
exoprotéase l’endothéline (ECE)

Enzymes de conversion del’endothéline
Chez l’homme, les endothélines-1,-2et -3 sont dérivées de précurseurs de38 (pour la big-ET-1 et 2) et 41 (pourla big-ET-3) acides aminés. Le groupede Kashiwabara et al. a montré que labig-ET-1 et l’ET-1, aux mêmes doses,induisent in vivo des effets équivalentssur la pression artérielle [12]. En
revanche, la big-ET-1 est environ 250 à1 000 fois moins puissante que l’ET-1comme agent vasoconstricteur sur lesvaisseaux isolés. Cette observation apermis de suggérer qu’il existait invivo une conversion dynamique duprécurseur en son métabolite actifl’ET-1 [12].En 1990, nous avons rapporté que labig-ET-1 est quasiment aussi activeque l’ET-1 chez le lapin anesthésié et
que l’administration intraventriculai-re gauche de la big-ET-1 induisait uneaugmentation de la concentrationd’ET-1 circulante, probablement parconversion systémique du précurseur[13]. Ces observations ont été confir-mées par Hemsen et Lundberg chezle porc et, plus récemment, chezl’homme [14]. L’utilisation d’uninhibiteur de l’enzyme de conversionde l’endothéline (ECE), le phospho-
565m/s n° 5, vol. 12, mai 96
Figure 1. Représentation schématique des interactions proposées et/ou connues entre les cytokines, les cellules en-dothéliales (CE) et du muscle lisse vasculaire (CML) avec plusieurs autres cellules dérivées du système immunitaire,telles que les polymorphonucléaires (PN) et autres leucocytes. Les cellules stimulées du système immunitaire libè-rent des leucotriènes, plusieurs cytokines (IL-1 et IL-6) et facteurs de croissance (facteur G), tels que le facteur decroissance dérivé de plaquettes (PDGF), le tumor necrosis factor α (TNFα), le facteur activateur des plaquettes (PAF),le facteur stimulateur de colonies granulocytaire et macrophage (GM-CSF) et le transforming growth factor β (TGFβ).Ces facteurs sécrétés agissent sur les différentes composantes du muscle lisse vasculaire qui, à leur tour, sécrètentdes facteurs tels que l’EDRF et l’endothéline (ET). Quelques unes de ces cellules du système immunitaire peuvent selier aux cellules endothéliales via des molécules d’adhérence telles la GMP-140 et ICAM-1 situées sur la membraneplasmique de l’endothélium activé lors de processus inflammatoires [64]. LTA4, LTB4, LTC4 : leucotriènes A4, B4, C4 ;NO : monoxyde d’azote.

ramidon [15], a permis d’identifierl’activité de l’ECE dans plusieursorganes : le rein, le poumon, le trac-tus gastrointestinal, le système ner-veux central, l’appareil urogénital, lamicrocirculation, etc. [16-21].Une étude récente de Corder etVane [22] montre que, chez le rat,après administration intraventriculai-re de big-ET-1, l’ET-1 n’est pas détec-tée dans la circulation sanguine. Laconversion de la big-ET-1 en ET-1serait donc un phénomène local,relayé par une enzyme située proba-blement sur la partie basale de
l’endothélium, ce qui permet uneaugmentation polarisée de la concen-tration d’ET-1 dans le tissu interstitielsous-endothélial. Toutefois, cesobservations chez le rat ne prennentpas en considération le fait que la big-ET-1 exogène puisse être convertienon seulement par l’ECE mais aussipar l’endopeptidase neutre 24.11[23] dans cette espèce animale.L’absence d’élévation de l’ET-1 plas-matique chez le rat après administra-tion de big-ET-1 pourrait donc êtredue à l’action locale d’autres entitésenzymatiques que l’ECE. En dépit de
566 m/s n° 5, vol. 12, mai 96
Figure 2. A. Représentation schématique des trois précurseurs de la familledes endothélines (h-ET) humaines. Les trois pré-pro ET contiennent un sitede reconnaissance de la furine RXK/RR (Arg-X-Lys/Arg-Arg) encadrant lesportions big-ET. ECE indique l’enzyme de conversion de l’endothéline qui cli-ve le polypeptide entre les résidus tryptophane (W) et valine (V) (ou isoleuci-ne (I) pour la big-ET-3). Les séquences signal sont indiquées en noir et lesportions rouges et roses indiquent les régions correspondant aux big-ET. Lenombre total d’acides aminés est indiqué à droite. R, Arg ; L, Leu ; S, Ser ; K,Lys ; G, Gly [10]. Les chiffres situés au-dessus et au-dessous des sites de cli-vage indiquent la position du résidu immédiatement en amont du site clivé.B. Effet contractile de la pré-pro-ET-1 digérée par la furine et la chymosine(échantillon). Contraction de l’artère carotide de lapin par l’échantillon enprésence (droite) ou en l’absence (gauche) de l’antagoniste sélectif des ré-cepteurs ETA, le BQ-123. Les échantillons témoins (tampon de digestion seul,la chymosine et la furine seulement), la pré-pro ET-1 seule ou clivée par la fu-rine seulement n’ont pas provoqué de réponse sur cette préparation [11].
RÉFÉRENCES12. Kashiwabara T, Inagaki Y, Ohta H, Iwa-matsu A, Nomizu M, Morita A, Nishikori K.Putative precursors of endothelin have lessvasoconstrictor activity in vitro but a potentpressor effect in vivo. FEBS Lett 1989 ; 247 :73-6.
13. D’Orléans-Juste P, Lidbury PS, WarnerTD, Vane JR. Intravascular big endothelinincreases circulating levels of endothelin-1and prostanoids in the rabbit. Biochem Phar-macol 1990 ; 39 : R21-2.
14. Hemsen A, Ahlborg G, Ottosson-Seeber-ger A, Lundberg JM. Metabolism of big-en-dothelin-1 (1-38) and (22-38) in the humancirculation in relation to production of en-dothelin-1 (1-21). Regul Pept 1995 ; 55 : 287-97.
15. McMahon EG, Palomo MA, Moore WM,McDonald JF, Stern MK. Phosphoramidonblocks the pressor activity of porcine big-en-dothelin-1 (1-39) in vivo and conversion ofbig-endothelin-1 (1-39) to endothelin-1 (1-21) in vitro. Proc Natl Acad Sci USA 1991 ; 88 :703-7.
16. Télémaque S, Lemaire D, Claing A,D’Orléans-Juste P. Phosphoramidon-sensiti-ve effect of big-endothelins in the perfusedrabbit kidney. Hypertension 1992 ; 20 : 518-23.
17. D’Orléans-Juste P, Lidbury PS, Télé-maque S, Warner TD, Vane JR. Human big-endothelin releases prostacyclin in vivo andin vitro through a phosphoramidon-sensitiveconversion to endothelin-1. J CardiovascPharmacol 1991 ; 17 : S251-5.
18. Rae GA, Calixto JB, D’Orléans-Juste P.Effects and mechanisms of action of endo-thelins on non-vascular smooth muscle ofthe respiratory, gastrointestinal and uroge-nital tracts. Regul Pept 1995 ; 55 : 1-46.
19. Poulat P, D’Orléans-Juste P, de Cham-plain J, Yano M, Couture R. Cardiovasculareffects of intrathecally administered endo-thelins and big endothelin-1 in consciousrats : receptor characterization and mecha-nism of action. Brain Res 1994 ; 648 : 239-48.
20. Télémaque S, D’Orléans-Juste P. Presen-ce of a phosphoramidon-sensitive endothe-lin-converting enzyme which converts big-endothelin-1, but not big-endothelin-3, inthe rat vas deferens. Naunyn-Schmiedeberg’sArch Pharmacol 1991 ; 344 : 505-7.
21. Lehoux S, Plante GE, Sirois MG, SiroisP, D’Orléans-Juste P. Phosphoramidonblocks big-endothelin-1 but not endothelin-1 enhancement of vascular permeability inthe rat. Br J Pharmacol 1992 ; 107 : 996-1000.

cette exception, dans plusieursespèces animales (autres que le rat)et chez l’homme [14], l’administra-tion intraveineuse de la big-ET-1 pro-voque généralement une augmenta-tion des concentrations sanguinesd’ET-1.Récemment, les groupes de Shimadaet de Yanagisawa [24, 25] ont identi-fié l’enzyme de conversion de l’endo-théline comme une entité biochi-
mique distincte de l’endopeptidaseneutre 24-11. En particulier, Shimadaa rapporté l’étude de l’expressionfonctionnelle de l’ECE dérivée descellules endothéliales de la circula-tion pulmonaire de rat [24]. Cetteenzyme possède une plus grande affi-nité pour la big-ET-1 que pour la big-ET-2 ou la big-ET-3, et a été trèsrécemment identifiée chez l’homme[26]. Récemment, trois isoformes de
l’ECE (ECE-1a, ECE-1b, ECE-2) ontété caractérisées par leur structuremoléculaire, leur localisation cellulai-re et leur affinité pour les trois iso-peptides, les big-ET-1, 2 et 3 [25, 27,28]. L’ECE-1a est localisée au niveau de lamembrane plasmique et son domai-ne catalytique disposé sur la faceexterne de la cellule endothéliale[29]. Cette enzyme serait considéréecomme l’entité physiologique prédo-minante impliquée dans la produc-tion d’endothéline-1. La localisationet la fonction des isoformes ECE-1bet ECE-2 demandent encore à êtredéfinies.
L’ECE pulmonaire possède peud’affinité pour la big-ET-2
Lorsqu’elle est administrée par voieintraveineuse chez le cobaye, la big-ET-1 induit une augmentation de lapression artérielle et de la résistancepulmonaire (pression d’insufflationpulmonaire). Ces deux effets de labig -ET-1 sont fortement réduitslorsque l’animal est prétraité par lephosphoramidon. Le précurseur del’ET-2, la big-ET-2, est aussi un puis-sant agent hypertenseur sensible auphosphoramidon mais, contraire-ment à la big-ET-1, la big-ET-2 n’aug-mente pas la résistance pulmonaire[31] à des doses vasopressives supra-maximales chez le cobaye anesthésié.En outre, la big-ET-1, mais non la big-ET-2, stimule la libération de throm-boxane bronchonconstricteur par lepoumon perfusé de cobaye . A la sui-te de ces expériences, nous avonssuggéré que, chez cette espèce ani-male, l’activité de l’enzyme deconversion de l’endothéline dansl’arbre vasculaire pulmonaire étaitdifférente de celle observée dans lecircuit systémique. La faible affinitéde l’ECE pulmonaire du cobaye pourla big-ET-2 peut donc expliquer soninaptitude à induire des effets bron-choconstricteurs en relation avec sonincapacité de stimuler la libérationd’eicosanoïdes [31]. Cette caractéris-tique de l’ECE pulmonaire (faibleaffinité pour la big-ET-2) a égalementété retrouvée chez le rat [32]. Dans la figure 3, sont présentées lesactivités relatives de la big-ET-1 et dela big-ET-2 à celle de leur métaboliteactif dans le rein et le poumon perfu-sé de lapin : les deux précurseurs
567m/s n° 5, vol. 12, mai 96
Tableau I
CARACTÉRISTIQUES BIOCHIMIQUES ET PHARMACOLOGIQUESDES RÉCEPTEURS DES ENDOTHÉLINES
Récepteur ETA ETB
Nombre d'acides aminés 427 (homme) 442 (homme)427 (bœuf) 441 (bœuf)
426 (rat) 441 (rat)
Ordre de puissance ET-1 ≥ ET-2 >> ET-3 ET-1 = ET-2 = ET-3des endothélines
Effecteurs Gq/11 Gq/11
Mécanisme de transduction IP3/Ca2+/DG IP3/Ca2+/DGdu signal
Agonistes sélectifs [Trp(For)21]-ET-1 IRL 1620peptidiques BQ-3020
Sarafotoxin 6C (?)
Antagonistes sélectifs BQ-123 BQ-788peptidiques FR139317
Antagonistes non sélectifs PD147452peptidiques PD142893
Agonistes sélectifs – RO 46-8443non peptidiques
Antagonistes sélectifs BMS-182874 –non peptidiques
Antagonistes non sélectifs Bosentannon peptidiques SB 209670
IP3 : inositol triphosphate ; DG : diacylglycérol.

sont aussi efficaces pour induire unevasoconstriction rénale chez le lapin.Par ailleurs, la big-ET-2 est significati-vement moins puissante que la big-ET-1 pour induire des effets vaso-constricteurs pulmonaires. La faibleaffinité de l’ECE pulmonaire pour labig-ET-2 se retrouve donc chez troisespèces différentes, le cobaye, le ratet le lapin [31-33].Cette mise en évidence de caractéris-tiques distinctes des ECE pulmonaireet rénale devrait permettre le déve-loppement d’inhibiteurs interférantavec la synthèse tissulaire de l’ET-1de façon sélective. Certaines de cesmolécules sont en cours de dévelop-pement [34].
Rôle de l’ECE vasculaire chez l’homme
Le groupe de Haynes et Webb a mon-tré que, chez l’homme, l’administra-tion de phosphoramidon dans l’artè-re brachiale induit une chute derésistance artérielle locale [35]. Cesobservations illustrent bien l’impor-tance de l’ECE dans la genèse del’ET-1 et du rôle de cette dernière entant que vasomodulateur endogène.Le phosphoramidon, partiellementconstitué d’hydrates de carbone luiconférant des caractéristiques hydro-philes, pénètre difficilement la mem-brane plasmique et ne peut inhiberl’ECE intracellulaire (ECE-2), obser-vation rapportée par Emoto et al.[27]. Le phosphoramidon sembleagir de façon prédominante surl’ECE-1a localisée à la surface exter-ne de la membrane cellulaire. Celaest d’intérêt puisque cet inhibiteursemble avoir des propriétés vasodila-tatrices dans les vaisseaux humainsdues non pas à l’inhibition de la pro-duction d’endothéline intra-cellulai-re (par blocage de l’ECE intracellu-laire), mais à sa capacité d’inhiber laconversion extra-cellulaire en ET-1de la big-ET-1, sécrétée par l’endothé-lium [35]. De façon similaire, unautre inhibiteur de l’ECE, leFR901533, a une faible capacité depénétration intracellulaire et agitdonc également de façon plus mar-quée sur l’ectoenzyme localisée sur lasurface externe des membranes cel-lulaires de l’endothélium [27].L’inhibition de l’ECE intracellulairepar le phosphoramidon ne peut tou-tefois être totalement exclue puisquecet inhibiteur des métalloendopepti-
dases réduit la production d’ET-1 descellules endothéliales en culture[36]. Il est à noter que ces cellules enculture sécrètent peu de big-ET-1 parrapport à l’ET-1.
Récepteurs des endothélines
Les gènes codant pour les récepteursETA et ETB humains ont été clônés[37, 38]. Ces récepteurs possèdentsept domaines transmembranaires etsont couplés aux protéines G, plusspécifiquement Gq/11 [30]. Les gènesETAR et ETBR possèdent au moins85 % d’identité structurale avec ceuxidentifiés dans d’autres espèces demammifères, telles que le rat, le porcet le bœuf. En revanche, il n’y a que59 % de similitude entre les récep-teurs ETA et ETB humains.Lorsqu’ils sont activés, par le biais deprotéines G (sensibles à la toxinepertussique dans le muscle lisse vas-culaire), ces récepteurs activent laphospholipase C qui, à son tour, pro-duit deux seconds messagers, l’inosi-tide-triphosphate (IP3) et le diacylglycerol (DAG). Les IP3 mobilisent lecalcium cytoplasmique et le DAG estresponsable de l’activation de la pro-téine kinase C. Cette cascade d’évé-nements intracellulaires mène audéveloppement des mécanismescontractiles (muscle) ou sécrétoires(cellules endothéliales) [39, 40].Classiquement, les préparations vas-culaires exprimant le récepteur detype ETA répondent aux trois endo-thélines selon l’ordre de puissancesuivant : ET-1 ≥ ET-2 >> ET-3, alorsque dans le cas du récepteur ETB lestrois isoformes sont équipotentes. Unrésumé des caractéristiques biochi-miques et pharmacologiques desrécepteurs ETA et ETB est présentédans le Tableau 1.Plusieurs outils pharmacologiques(agonistes et antagonistes) sont dis-ponibles actuellement pour la carac-térisation des récepteurs ETA et ETB(Tableau I). Parmi ces composés, lesantagonistes sélectifs ETA (BMS-182874) et non sélectifs (bosentan etSB 209670) retiennent l’attentionsurtout pour leur caractéristiquesnon-peptidiques qui accentuent leurbiodisponibilité et permettent leuradministration per os [41-43].
568 m/s n° 5, vol. 12, mai 96
RÉFÉRENCES22. Corder R, Vane JR. Radioimmunoassayevidence that the pressor effect of big endo-thelin-1 is due to local conversion to endo-thelin-1. Biochem Pharmacol 1995 ; 49 : 375-80.
23. Gardiner SM, Kemp PA, Bennett T. Ef-fects of the neutral endopeptidase inhibi-tor, SQ 28,603 on regional haemodynamicresponses to atrial natriurectic peptide orproendothelin-1 [1-38] in conscious rats. BrJ Pharmacol 1992 ; 106 : 180-6.
24. Shimada K, Takahashi M, Tanzawa K.Cloning and functional expression of endo-thelin-converting enzyme from rat endothe-lial cells. J Biol Chem 1994 ; 269 : 18275-8.
25. Xu D, Emoto N, Giaid A, Slaughter C,Kaw S, DeWit D, Yanagisawa M. ECE-1 : amembrane-bound metalloprotease that ca-talyses the protolytic activation of big-endo-thelin-1. Cell 1994 ; 78 : 473-85.
26. Shimada K, Matsushita Y, WakabayashiK, Takahashi M, Matsubara A, Iijima Y, Tan-zawa K. Cloning and fonctional expressionof human endothelin-converting enzymecDNA. Biochem Biophys Res Commun 1995 ;207 : 807-12.
27. Emoto N, Yanagisawa M. Endothelin-converting enzyme-2 is a membrane-bound,phosphoramidon-sensitive metalloproteasewith acidic pH optimum. J Biol Chem 1995 ;270 : 15262-8.
28. Yorimitsu K, Moroi K, Inagaki N, SaitoT, Masuda Y, Masaki T, Seino S, KimuraS.Cloning and sequencing of a human en-dothelin converting enzyme in renal adeno-carcinoma (ACHN) cells producing endo-thelin-2. Biochem Biophys Res Commun 1995 ;208 : 721-7.
29. Takahashi M, Fukuda K, Shimada K,Barnes K, Turner AJ, Ikeda M, Koike H, Ya-mamoto Y, Tanzawa K. Localization of ratendothelin-converting enzyme to vascularendothelial cells and some secretory cells.Biochem J 1995 ; 311 : 657-65.
30. Kelly RA, Eid H, Kramer BK, O’Neill M,Liang BT, Reers M, Smith TW. Endothelinenhances the contractile responsiveness ofadult rat ventricular myocytes to calcium bya pertussis toxin-sensitive pathway. J Clin In-vest 1990 ; 86 : 1164-71.
31. Gratton JP, Rae GA, Claing A, Télé-maque S, D’Orléans-Juste P. Different pres-sor and bronchoconstrictor properties ofhuman big-endothelin-1,2 (1-38) and 3 inketamine/xylazine-anaesthetized guineapigs. Br J Pharmacol 1995 ; 114 : 720-6.
32. Gratton JP, Maurice MC, Rae GA, D’Or-léans-Juste P. Pharmacological properties ofendothelins and big-endothelins in ketami-ne/xylazine or urethane-anesthetized rats.Am J Hypertens 1995 ; 8 : 1121-7.

Pharmacologie des récepteurs ETBsur l’endothélium et sur le muscle lisse vasculaire
Le concept selon lequel les récep-teurs de type ETB sont localisés uni-quement sur l’endothélium et yrelaient les signaux de vasodilatationet les récepteurs de type ETA unique-ment au niveau du muscle lisse vascu-laire et y relaient la vasoconstriction,a été mis en cause par le groupe deMoreland qui a démontré que, danscertains modèles animaux, une pro-portion importante de récepteurs detype ETB au niveau du muscle lissevasculaire sont localisés dans certainsvaisseaux d’origine veineuse [44]. Bien que Davenport et al. [45] aientsuggéré une prédominance desrécepteurs ETA au niveau du musclelisse vasculaire de l’homme, la parti-cipation possible des récepteurs ETBaux effets vasoconstricteurs del’endothéline dans certains modèlesanimaux a incité plusieurs compa-gnies pharmaceutiques à développerdes antagonistes mixtes ETA/ETB [42,46]. Cependant, les agonistes sélectifs desrécepteurs ETB (BQ-3020 et IRL-1620) n’ont pas d’action vasocons-trictrice sur les vaisseaux d’origine
humaine dont l’endothélium est éli-miné mécaniquement. En outre,l’IRL-1620 stimule la libérationd’EDRF des cellules endothéliales dela veine ombilicale humaine [47]alors qu’on ne retrouve sur celles-cique des récepteurs de type ETB.L’identification de récepteurs ETBfonctionnels dans le muscle lisse vas-culaire humain s’est faite à l’aide dela sarafotoxine 6C, une toxine possé-dant une forte homologie structuraleavec les endothélines et utilisée com-me agoniste sélectif des récepteursETB [7, 48]. Or, il apparaît que cettetoxine peut activer des mécanismesintracellulaires au niveau du cerveletde rat via une voie qui n’impliquepas le couplage du récepteur à uneprotéine G sensible à la toxine per-tussique [49]. Il est donc possibleque les effets vasoconstricteurs obser-vés dans des tissus humains à l’aidede la sarafotoxine 6C impliquentl’activation de sites distincts de ceuxproposés pour les récepteurs de typeETB comme l’a récemment suggéréFlynn et al. [50] chez le rat décéré-bré.Des résultats récents de nos labora-toires aux niveaux rénal et pulmonai-re [51] ainsi que in vivo [52] souli-gnent toutefois l’importance de
préserver la possibilité d’activer lesrécepteurs de type ETB lors de l’admi-nistration d’antagonistes de l’endo-théline. En effet, dans les affectionsd’origine vasculaire comportant unehypersécrétion d’endothéline, blo-quer les capacités vasodilatatricesdépendantes de l’endothélium desrécepteurs ETB ne ferait qu’accentuerles propriétés vasoconstrictrices dupeptide. Ces considérations n’ont puémerger qu’après l’avènementd’antagonistes sélectifs des récepteursETB, tel le BQ-788 [52].A titre d’exemple, il a été démontrédans plusieurs études que l’ET-1,administrée par voie systémique,induit des effets presseurs vasocons-tricteurs très puissants au niveaurénal avant que n’apparaissent deseffets presseurs systémiques impor-tants. Ces observations illustrentl’extrême sensibilité de la vascularisa-tion rénale à l’ET-1 chez plusieursmammifères dont l’homme. Enoutre, le groupe d’Ishikawa a montréque le BQ-788 (antagoniste sélectifdu récepteur ETB) potentialise leseffets hypertenseurs de l’ET-1 chez lerat [52]. Contrairement à l’homme,il avait été montré chez le rat, que lesrécepteurs de type ETB vasoconstric-teurs participent à près de 40 %, avec
569m/s n° 5, vol. 12, mai 96
Figure 3. Effet de la big-ET-1 ou de la big-ET-2, exprimé en pourcentage de la réponse à l’ET-1 ou à l’ET- 2 (10 nMdans le rein ; 50 nM dans le poumon), sur la pression de perfusion du rein (A) ou du poumon (B) isolé de lapin. *P < 0,05 lorsque comparé aux effets de big-ET-1, n = 4.

les récepteurs ETA, à l’effet hyperten-seur provoqué par l’administrationd’endothéline. Il apparaît donc quele récepteur ETB localisé sur l’endo-thélium, responsable de la vasodilata-tion, agit de façon prépondérantemalgré la présence, dans cette espè-ce, de récepteurs de type ETB sur lemuscle lisse des vaisseaux résistifspuisque la réponse hypertensive del’endothéline est fortement augmen-tée chez les rats traités au BQ-788[52]. Nous avons montré récemmentque le BQ-788 bloque les propriétésvasodilatatrices de l’EDRF au niveaude la circulation rénale, un phéno-mène sensible à un inhibiteur de laNO synthase [51]. Cet effet potentia-teur du BQ-788 in vivo et in vitro chezle lapin est illustré sur la figure 4.Nous suggérons donc que, dans desconditions dans lesquelles lesconcentrations d’endothéline circu-lante sont élevées (après les trans-plantations, par exemple), l’adminis-tration d’antagonistes sélectifs desrécepteurs ETB ou mixtes ETA/ETBdevrait induire une hyperréactivité àl’endothéline endogène et exogène[52, 53].
Sous-types de récepteurs ETB :entités pharmacologiques distinctes ?
Bien que les récepteurs de type ETBde rat et de l’homme aient une trèsgrande similitude de séquence (plusde 95 % d’identité), l’antagonistepeptidique mixte ETA/ETB, lePD147452, ou l’antagoniste sélectifETB, le BQ-788, ont une affinité diffé-rente pour le récepteur ETB humainet pour celui du rat [54]. Ces diffé-rences d’affinité semblent liées aufait que ces antagonistes se fixent surdes portions protéiques distinctes deces deux types de récepteurs ETBdans ces deux espèces. De plus, desétudes de Warner et al. [55] ont sug-géré l’existence de deux sous-typesdistincts de récepteurs ETB, le récep-teur ETB1 retrouvé sur l’endothéliumet qui est responsable de la libérationde l’EDRF, et le récepteur ETB2 loca-lisé sur le muscle lisse, vasculaire etnon-vasculaire, et qui induit deseffets constricteurs lorsqu’il est acti-vé. Cette classification n’est toutefoispas justifiée par des études de carac-térisation rigoureuse des récepteurs :sur les deux récepteurs ETB, l’ordrede puissance des agonistes naturels
570 m/s n° 5, vol. 12, mai 96
33. Gratton JP, Maurice MC, D’Orléans-JusteP. Characterization of endothelin receptorsand endothelin-converting enzyme activityin the rabbit lung. J Cardiovasc Pharmacol1995 ; 26 : S88-90.
34. Claing A, Neugebauer W, Yano M, RaeGA, D’Orléans-Juste P. [Phe22]-big-endothe-lin-1[19-37] : a new and potent inhibitor ofthe endothelin-converting enzyme. J Cardio-vasc Pharmacol 1995 ; 26 : S72-4.
35. Haynes WG, Webb DJ. Contribution ofendogenous generation of endothelin-1 tobasal vascular tone. Lancet 1994 ; 344 : 852-4.
36. Sawamura T, Kasuya Y, Matsushita Y, Su-zuki N, Shinmi O, Kishi N, Sugita Y, Yanagi-sawa M, Goto K, Masaki T, Kimura S. Phos-phoramidon inhibits the intracellularconversion of big-endothelin-1 to endothe-lin-1 in cultured endothelial cells. BiochemBiophys Res Commun 1991 ; 174 : 779-84.
37. Arai H, Hori S, Aramori I, Ohkubo H,Nakanishi S. Cloning and expression of acDNA encoding an endothelin receptor.Nature 1990 ; 348 : 730-2.
38. Sakurai T, Yanagisawa M, Takuwa Y,Miyazaki H, Kimura S, Goto K, Masaki T.Cloning of a cDNA encoding a non-isopep-tide-selective subtype of the endothelin re-ceptor. Nature 1990 ; 348 : 732-5.
39. Highsmith RF, Blackburn K, SchmidtDJ. Endothelin and calcium dynamics invascular smooth muscle. Annu Rev Physiol1992 ; 54 : 257-77.
40. Rembold CM. Regulation of contractionand relaxation in arterial smooth muscle.Hypertension 1992 ; 20 : 129-37.
41. Stein PD, Hunt J, Floyd D, Moreland S,Dickinson KE, Mitchell C, Liv EC, WebbML, Murugesan N, Dickey J, et al. The dis-covery of sulfonamide endothelin antago-nists and the development of the orally acti-ve ETA selective antagonist S-(dimethylamino)-N-(3,4-dimethyl-S-isoxazoil)-1-naph-talenesulfonamide. J Med Chem 1994 ; 37 :329-31.
42. Clozel M, Breu V, Gray GA, Kalina B,Löffler BM, Burri K, Cassal JM, Hirth G,Müller M, Neidhart W, Ramuz H. Pharma-cological characterization of bosentan, anew potent orally active nonpeptide endo-thelin receptor antagonist. J Pharmacol ExpTher 1994 ; 270 : 228-35.
43. Elliott JD, Amparo Lago M, Consins RD,Gao A, Leber JD, Erhard K, Eggleston D,Nambi P, Lee JA, Brooks DP, Feverstein G,Ruffdo RR, Weinstock J, Gleason JG, Pei-shoff CE, Ohlstein EH. 1,3-Diarylindan-2-Carboxylic acids, potent and selective non-peptide endothelin receptor antagonists. JMed Chem 1994 ; 37 : 1553-7.
44. Moreland S, McMullen DM, DelaneyCL, Lee VG, Hunt JT. Venous smoothmuscle contains vasoconstrictor ETB-like re-ceptors. Biochem Biophys Res Commun 1992 ;184 : 100-6.
45. Davenport AP, Maguire JJ. Is endothe-lin-induced vasoconstriction mediated onlyby ETA receptors in humans ? Trends Phar-macol Sci 1994 ; 15 : 9-11.
46. Ohlstein EH, Nambi P, Douglas SA,Edwards RM, Gellai M, Lago A, Leber JD,Cousins RD, Gao A, Frazee, JS, Peishoff, CE,Bean JW, Eggleston DS, Elshourbagy NA,Kumar C, Lee JA, Yue TL, Louden C,Brooks DP, Weinstock J, Feuerstein G, PosteG, Ruffolo RR Jr, Gleason JG, Elliott JD. SB 209670, a rationally designed potentnonpeptide endothelin receptor antagonist.Proc Natl Acad Sci USA 1994 ; 91 : 8052-6.
47. Tsukahara H, Ende H, Magazine HI, Ba-hou WF, Goligorsky MS. Molecular andfunctional characterization of the non-iso-peptide selective ETB receptor in endothe-lial cells. Receptor coupling to nitric oxidesynthase. J Biol Chem 1994 ;269 : 21778-85.
48. William DL, Jones KL, Pettibone DV, LisEV, Clineschmidt BV. Sarafotoxin S6C : anagonist which distinguishes between endo-thelin receptor subtypes. Biochem Biophys ResCommun 1991 ; 175 : 556-61.
49. Shraga-Levine Z, Galron R, SokolovskyM. Cyclic GMP formation in rat cerebellarslices is stimulated by endothelins via nitricoxide formation and by sarafotoxins via for-mation of carbon monoxide. Biochemistry1994 ; 33 : 14656-9.
50. Flynn, DA, Sargent, CA, Brazdil R,Brown TJ, Roach AG. Sarafotoxin S6C Eli-cits a non-ETA or non-ETB-mediated pressorresponse in the pithed rat. J Cardiovasc Phar-macol 1995 ; 26(suppl 3) : S219-21.
51. D’Orléans-Juste P, Claing A, TélémaqueS, Maurice MC, Yano M, Gratton JP. Blockof endothelin-1-induced release of throm-boxane A2 from the guinea pig lung and ni-tric oxide from the rabbit kidney by a selec-tive ETB receptor antagonist, BQ-788. Br JPharmacol 1994 ; 113 : 1257-62.
52. Ishikawa K, Ihara M, Noguchi K, MaseT, Mino N, Saeki T, Fukuroda T, Fukami T,Ozaki S, Nagase T, Nishikibe M, Yano M.Biochemical and pharmacological profile ofa potent and selective endothelin B recep-tor antagonist, BQ-788. Proc Natl Acad SciUSA 1994 ; 91 : 4892-6.
53. D’Orléans-Juste P, Yano M, Maurice MC,Gratton JP. Hyperresponsiveness to ET-1following treatment with a mixture of ETAand ETB receptor antagonists in the rabbitin vivo and in vitro. J Cardiovasc Pharmacol1995 ; 26 : S369-72.
RÉFÉRENCES

est identique, les agonistes sélectifsdes récepteurs ETB sont actifs et lesantagonistes sélectifs des récepteursde type ETB, tel le BQ-788, interfè-rent efficacement avec l’effet del’ET-1. De plus, Clozel et Gray ontrécemment démontré, à l’aide del’antagoniste ETB RO 46-8443, queles récepteurs ETB, responsables deseffets opposés de constriction et dedilatation, ne représentaient pas desentités pharmacologiques distinctes[56].Par conséquent, les caractéristiquesdistinctes de ces deux sous-types derécepteurs ETB1 et ETB2 devront êtrevalidées par des études de biologiemoléculaire permettant de produiredes récepteurs recombinants quel’on pourra caractériser au point devue pharmacologique. Actuellement,seule la nomenclature ETA et ETB desrécepteurs de l’ET-1 est retenue parIUPHAR (union internationale depharmacologie).
Implication des récepteurs de l’endothéline dans le contrôle de lapression artérielle
Des études récentes de Kurihara et al.et de Yanagisawa et al. ont montréque des souris transgéniques homo-zygotes déficientes en endothéline[57] ou en récepteurs ETA [58] déve-loppent des malformations physiquessévères. Ces observations ont permisà ces groupes de suggérer que l’ET-1et ses récepteurs ETA sont impliquésde façon essentielle dans les méca-nismes de croissance et de dévelop-pement (m/s n° 6-7, vol. 10, p. 740).Chez les souris transgéniques chezlesquelles un allèle du gène de l’ET-1était délété (hétérozygotes), unelégère augmentation de la pressionsanguine a toutefois été notée malgréune diminution des concentrationsplasmatiques d’ET-1 [57]. Il s’agit làde mécanismes fort complexes et dif-ficiles à expliquer puisqu’on a peu
d’information sur la pharmacologiecardiovasculaire de l’endothélinechez cette espèce animale. Notonsqu’il a été récemment démontré que,chez ces animaux hétérozygotes, lafonction du système sympathique etdes baroréflexes est altérée [59].L’importance physiologique desrécepteurs de type ETA dans les vais-seaux humains intacts a été démon-trée par Haynes et Webb [35]. Pourla première fois, ce groupe a rappor-té que l’endothéline endogène joueun rôle important dans le maintiendu tonus vasculaire des vaisseauxd’individus sains. En effet, l’adminis-tration par voie intra-artérielle deBQ-123 (un antagoniste sélectif durécepteur ETA) induit une vasodilata-tion marquée des vaisseaux sanguinsintacts dans l’avant-bras chez l’hom-me (figure 5). Il faut aussi soulignerque le BQ-123 réduit de façon trèsprononcée la pression artériellemoyenne chez le cobaye [60], suggé-
571m/s n° 5, vol. 12, mai 96
Figure 4. (A) Effet vasodilatateur suivi de l’effet vasoconstricteur produit par l’injection intra-artérielle de0,25 nmol/kg d’endothéline-1 chez le lapin anesthésié en absence et en présence de l’antagoniste ETB, le BQ-788(0,25 mg/kg). (B) Effet de l’ET-1 sur la pression de perfusion du rein de lapin perfusé en absence ou en présence deBQ-788 (10 nM), de L-NAME (100 µM) et de BQ-788 (10 nM) + L-NAME (100 µM). *P < 0,05 ; †P < 0,01 lorsque com-paré aux valeurs témoins ; n.s. : non-significatif.

rant qu’au moins chez ce mammifè-re, l’endothéline joue un rôle impor-tant dans le maintien de la pressionartérielle.Une implication de l’endothélinecomme stimulateur de l’hypertro-phie vasculaire a aussi été suggéréedans certains modèles d’hyperten-sion artérielle ; pour plus de détails,le lecteur est prié de se référer à larevue de Schiffrin [61] ainsi qu’àl’article du même auteur dans cenuméro de médecine/sciences [62].
Effet de l’endothéline sur la modulation calcique cytosolique et nucléique
Bien qu’agissant sur des récepteursqui ont été clonés et qui sont caracté-risés au plan pharmacologique [37,
38], l’ET-1 peut également agir enouvrant des canaux calciques de typeL (sensibles aux composés dihydro-pyridines) sur le muscle lisse vasculai-re [63], ou de type R (resting potentialcalcium channels) sur le muscle lissevasculaire ou les cellules endothé-liales [64]. La figure 6 illustre parmicroscopie confocale l’augmenta-tion de [Ca]i et sa distribution spéci-fique au niveau cytosolique etnucléaire dans une cellule de musclelisse vasculaire, au repos ou en pré-sence d’ET-1.
Conclusion
Les récepteurs de type ETA, l’enzymede conversion de l’endothéline etpar conséquent l’ET-1 elle-même,semblent tous être impliqués dans le
572 m/s n° 5, vol. 12, mai 96
Figure 5. Débit sanguin de l’avant-bras chez l’homme après une perfusion in-tra-artérielle d’ET-1 et de BQ-123. La vasoconstriction de l’ET-1 est abolie parle BQ-123 (P=0,006) et le BQ-123 seul induit une vasodilatation progressivede l’avant-bras (P = 0,007) (Tiré de [35] avec autorisation des auteurs).
RÉFÉRENCES54. Reynolds EE, Hwang O, Flynn MA, Wel-ch KM, Cody WL, Steinbaugh B, He JX,Chung FZ, Doherty AM. Pharmacologicaldifferences between rat and human endo-thelin B receptors. Biochem Biophys Res Com-mun 1995 ; 209 : 506-12.
55. Warner TD, Allcock GH, Corder R,Vane JR. Use of the endothelin antagonistsBQ-123 and PD142893 to reveal three endo-thelin receptors mediating smooth musclecontraction and the release of EDRF. Br JPharmacol 1993 ; 110 : 777-82.
56. Clozel M. and Gray G. Are there diffe-rent ETB receptors mediating constrictionand relaxation? J Cardiovasc Pharmacol 1995;26 : S262-4.
57. Kurihara Y, Kurihara H, Suzuki H, Ko-dama T, Maemura K, Nagai R, Oda H, Ku-waki T, Cao WH, Kamada N, Jishage K, Ou-chi Y, Azuma S, Toyoda Y, Ishikawa T,Kumada M, Yazaki Y. Elevated blood pressu-re and craniofacial abnormalities in micedeficient in endothelin-1. Nature 1994 ; 368 :703-10.
58. Goto K, Warner TD. Endothelin versati-lity. Nature 1995 ; 375 : 539-40.
59. Kuwaki T, Kurihara Y, Kurhara H, CaoWH, Ju KH, Ling GY, Onoder M, UnekawaM, Yazaki Y, Kumada M. Cardiorespiratoryabnormalities in mice heterozygous for ET-1 mutation. Fourth International Conference onEndothelin, London, UK, 1995.
60. Veniant M, Clozel JP, Hess P, Clozel M.Endothelin plays a role in the maintenanceof blood pressure in normotensive guineapigs. Life Sci 1994 ; 44 : 445-54.
61. Schiffrin EL. Endothelin in hyperten-sion. Curr Op Cardiol 1995 ; 10 : 485-94.
62. Schiffrin EL. Les endothélines sont-ellesimpliquées dans l’hypertension ? médecine/sciences 1996 ; 12.
63. Goto K, Kasuya Y, Matsuki N, Takuwa Y,Kurihara H, Ishikawa T, Kimura S, Yanagisa-wa M, Masaki T. Endothelin activates the di-hydropyridine-sensitive, voltage-dependentCa2+ channel in vascular smooth muscle.Proc Natl Acad Sci USA 1989 ; 86 : 3915-8.
64. Bkaily G. Regulation of R-type Ca2+
channels by insulin and ET-1 in vascularsmooth muscle. In : Bkaily G, ed. Ion chan-nels in vascular smooth muscle. Austin : Mole-cular Biology Intelligence Unit, RG LandesCompany, 199 : 41-52.
65. Bkaily G, Pothier P, Simaan M, Belzile Fet Jaalouk D. The use of confocal microsco-py in the investigation of cell structure andfunction in the cardiovascular system. MolCell Biochem 1996 (sous presse).

maintien du tonus vasculaire chezl’homme. Qui plus est, l’hormoneelle-même et ses récepteurs ETA(mais non ETB) sont essentiels audéveloppement. Il apparait doncque, chez le sujet sain, il existe unjuste équilibre entre la productiond’ET-1 et l’activation de ses récep-teurs ETA (musculaires) et ETB(endothéliaux). Dans des conditionspathologiques, un choix judicieuxdevra être fait dans l’utilisationd’antagonistes mixtes ou sélectifspour les récepteurs ETA ; nos résultatset les différents aspects discutés icitendent à favoriser la deuxième pro-position.Bien que non exclusive, la figure 7illustre les sites où les récepteurs etles activités enzymatiques de conver-sion de l’endothéline sont les plusabondants et jouent un rôle prédo-minant dans les effets vasculaires desendothélines chez l’homme. Ces sitespourraient être considérés commedes cibles de choix pour le dévelop-pement d’outils thérapeutiques.Chacune des quatre conférencesinternationales sur l’endothéline amarqué un tournant important dansl’étude de cette famille de peptidesvasoactifs. La dernière conférence,tenue à Londres en avril 1995, a sou-ligné l’ère des antagonistes desrécepteurs de l’endothéline et deleur utilisation dans certaines condi-tions physiopathologiques dans les-quelles ce peptide pourrait êtreimpliqué. La prochaine conférencequi se tiendra à Kyoto à l’automne1997 devrait marquer l’ère desenzymes de conversion de l’endothé-line et de leur rôle possible dansl’homéostasie et dans l’étiologie desmaladies cardiovasculaires ■
573m/s n° 5, vol. 12, mai 96
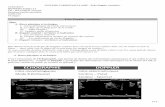
Figure 6. Augmentation de calcium intracellulaire cytosolique ([Ca]c) et nu-cléaire ([Ca]n) dans une cellule musculaire lisse d’aorte humaine au repos oustimulée avec l’ET-1 (10–7 M). A : Vue en coupe tranversale de la fluorescencedu Ca2+ libre au niveau cytosolique et nucléaire. B : L’ET-1 à la concentrationde 10–7 M augmente à la fois le [Ca]c et le [Ca]n. Les panneaux de droite sontdes représentations tridimensionnelles illustrant à la fois l’intensité et la dis-tribution spatiale du signal donné par les ions calciques des cellules degauches (A et B). L’échelle colorimétrique à droite de la figure représente leniveau d’intensité en pseudocouleurs des niveaux de la sonde calcique fluo-rescente (Fluo-3) (expérience # aoSMC17Flo10) [65].
Figure 7. Schéma illustrant les sites essentiels des effets de la big-endothé-line-1 (big-ET-1) et de l’endothéline-1 (ET-1) chez l’homme. CE : cellules en-dothéliales ; CML : cellule musculaire lisse ; ECE : enzyme de conversion del’endothéline ; ETA : récepteur de type ETA ; ETB : récepteur de type ETB.
Remerciements
Les auteurs sont reconnaissants à madameLouise Dubois pour son aide secrétariale et laFondation des Maladies du Cœur du Québecet le Conseil Médical de la Recherche du Ca-nada pour leur soutien financier. Les docteursPedro D’Orléans-Juste, Richard Leduc et Ghas-san Bkaily sont chercheurs boursiers du Fondsde Recherche en Santé du Québec. MadameAudrey Claing et monsieur Jean-Philippe Grat-ton sont détenteurs de bourses d’études docto-rales de la Fondation Canadienne des mala-dies du coeur et du fond FCAR/FRSQ santérespectivement.

574 m/s n° 5, vol. 12, mai 96
SummaryEndothelin cardiovascularpharmacology andpathophysiology
Multidisciplinary approaches inclu-ding pharmacology, biochemistry,immunology and rapidly develo-ping molecular biology techniqueshave been instrumental in the gi-gantic leaps observed in the fieldof endothelins in recent years.With the receptors cloned as wellas proposed biosynthetic pathwaysbeing rapidly elucidated, the roleof endothelin in cardiovascular ho-meostasis as well as in the ethiolo-gy of vascular pathologies may leadto the development of therapeuti-cally useful agents. This review suc-cinctly describes some of the im-portant aspects of the biology ofendothelins from the processing ofpre-pro-endothelin-1 to the activemoiety of endothelin-1. Further-more, physiopathological conside-rations are presented concerningthe usefulness of mixed ETA/ETBreceptors as opposed to selectiveETA receptors. Moreover, distinctendothelin-converting enzyme acti-vity specific to particular organs isoutlined. Finally, the nuclear andcytosolic calcium-mobilizing pro-perties of ET-1 in the vascularsmooth muscle are illustrated. Asfar as the contribution of endothe-lin in cardiovascular regulation isconcerned, a correlation attemptwill be made between the resultsobtained in animal models and
TIRÉS À PARTP. D’Orléans-Juste.