Le rôle des Cellules « Natural Killer » (NK) dans l ... Mavoungou 3.pdf · Le rôle des Cellules...
28
III Le rôle des Cellules « Natural Killer » (NK) dans l’Immunité anti-pathogènes Elie Mavoungou Professeur d’Immunologie Résumé Les cellules “tueuses naturelles” ou natural killer (NK) sont des cellules lymphoïdes qui catalysent une importante activité cytotoxique et produisent de forts taux de cytokines pro-inflammatoires en réponse aux infections. Au cours d’une infection virale, il est admis que la cytotoxicité des cellules NK et la production de cytokines sont induites principalement par les monocytes/macrophages et par les cellules dendritiques. Les ligands codant virtuellement pour les cellules NK sont actuellement en cours de description. La production d’interféron gamma (IFNγ) provenant des cellules NK est en outre essentielle pour le contrôle de plusieurs infections par les protozoaires responsables de la toxoplasmose, la trypanosomiase, la leishmaniose et du paludisme. L’activation des cellules NK par les pathogènes protozoaires est également mobilisée par les cytokines, bien que certaines études récentes suggèrent qu’une reconnaissance directe des parasites par les cellules NK dérivant des cellules accessoires ainsi que la signalisation directe, vraisemblablement par l’intermédiaire des récepteurs NK, soient nécessaires dans le cas des parasites du paludisme (Plasmodium spp.) à stimuler efficacement ces cellules. 1. Introduction La résistance de l’hôte contre les virus, les bactéries et les protozoaires pathogènes dépend d’une interrelation complexe entre les mécanismes de l’immunité innée et ceux de l’immunité spécifique ou acquise. En intervenant en première ligne de la défense de l’organisme, le système de l’immunité innée contribue au contrôle des infections aiguës par l’augmentation des réponses protectrices contre les parasites qui
Transcript of Le rôle des Cellules « Natural Killer » (NK) dans l ... Mavoungou 3.pdf · Le rôle des Cellules...

III
Le rôle des Cellules « Natural Killer » (NK) dans
l’Immunité anti-pathogènes
Elie Mavoungou Professeur d’Immunologie
Résumé
Les cellules “tueuses naturelles” ou natural killer (NK) sont des cellules lymphoïdes qui
catalysent une importante activité cytotoxique et produisent de forts taux de cytokines
pro-inflammatoires en réponse aux infections. Au cours d’une infection virale, il est
admis que la cytotoxicité des cellules NK et la production de cytokines sont induites
principalement par les monocytes/macrophages et par les cellules dendritiques. Les
ligands codant virtuellement pour les cellules NK sont actuellement en cours de
description. La production d’interféron gamma (IFNγ) provenant des cellules NK est en
outre essentielle pour le contrôle de plusieurs infections par les protozoaires responsables
de la toxoplasmose, la trypanosomiase, la leishmaniose et du paludisme.
L’activation des cellules NK par les pathogènes protozoaires est également
mobilisée par les cytokines, bien que certaines études récentes suggèrent qu’une
reconnaissance directe des parasites par les cellules NK dérivant des cellules accessoires
ainsi que la signalisation directe, vraisemblablement par l’intermédiaire des récepteurs
NK, soient nécessaires dans le cas des parasites du paludisme (Plasmodium spp.) à
stimuler efficacement ces cellules.
1. Introduction
La résistance de l’hôte contre les virus, les bactéries et les protozoaires
pathogènes dépend d’une interrelation complexe entre les mécanismes de l’immunité
innée et ceux de l’immunité spécifique ou acquise. En intervenant en première ligne de la
défense de l’organisme, le système de l’immunité innée contribue au contrôle des
infections aiguës par l’augmentation des réponses protectrices contre les parasites qui

2
envahissent l’organisme, avant la mise en place de la riposte de l’immunité à médiation
cellulaire par les lymphocytes T et B, communément appelée : l’immunité spécifique. Les
réponses innées stimulent et modulent aussi les réponses de l’immunité spécifique. Les
cellules NK sont bien connues pour jouer un rôle clé dans ces réponses innées rapides [1].
Dans le présent article, nous relatons brièvement la biologie des cellules NK, la
structure de leurs récepteurs, et nous discutons les connaissances actuelles concernant
leur fonction, dans la mise en œuvre de l’immunité innée contre les infections pathogènes
en nous focalisons plus particulièrement sur l’infection palustre due à Plasmodium
falciparum.
2. Biologie des cellules NK
L’identification de la souche lymphocytaire ayant la capacité d’exercer une
activité cytolytique contre certaines cellules tumorales sans aucune stimulation préalable
a conduit à l’utilisation du terme « cellule tueuse naturelle » (en anglais natural killer
cells, ou cellules NK ) [2, 3].
La capacité des cellules NK à percevoir les modifications dans l’expression des
molécules du complexe majeur d’histocompatibilité de classe I (CMH I) à la surface des
cellules infectées avait été rapportée il y a plusieurs décennies déjà, par Klaus Kärre et
ses collaborateurs à l’Institut Karolisnka de Stockholm. Dans leur étude, ces auteurs
avaient remarqué que des cellules tumorales de souris n’exprimant pas d’antigènes du
CMH I étaient plus facilement détruites par les cellules NK que d’autres cellules
tumorales qui exprimaient un niveau normal d’antigènes de classe I.
Il est maintenant bien connu qu’en plus de la destruction des cellules transformées
encore dites lignées cellulaires transformées [4], les cellules NK peuvent aussi jouer un
rôle dans le rejet d’allogreffes [5] et dans le contrôle d’une grande variété de pathogènes,
et plus spécialement ceux qui infectent directement les cellules hôtes [6].

3
Les cellules NK sont des cellules dites grands lymphocytes granulaires ou LGL de
l’anglais « Large Granular Lymphocytes » qui dérivent de la moelle osseuse et qui chez
l’homme, sont classiquement définies par l’expression à leur surface membranaire de
l’antigène CD56 (une isoforme de la molécule d’adhésion neurale [N-CAM]). Les
cellules NK sont caractérisées également par l’absence de l’expression du marqueur des
thymocytes, l’antigène CD3, et celle des sous unités αβ et γδ du récepteur T (TCR) des
lymphocytes. L’expression d’autres molécules de surface tels le marqueur 7 des
leucocytes (Leu7 encore dénommé CD57), la chaîne β de l’interleukine 2 (IL-2 ou
CD122), et le récepteur B1 des cellules « Killer lectin like » (KLRB1 ou CD61) a été
également utilisé comme marqueur de certaines souches de cellules NK. Toutefois, ces
différentes molécules sont aussi exprimées sur certaines autres sous populations de
cellules T et des cellules NK-T. La caractérisation phénotypique des cellules NK de
souris est plus complexe. Les marqueurs les plus communément utilisés sont, la sous
unité α2 de l’intégrine (DX5 ou CD49b), l’asialo-ganglioside M1 (ASGM1) et le
récepteur P1C des cellules NK (NKR-P1C encore appelé NK1.1). Cependant, les
lymphocytes T CD4+ et T CD8+ peuvent aussi exprimer ces marqueurs [7, 8]. L’ASGM1
par exemple est exprimé par 20 à 30 % de cellules lymphocytaires naïves et par environ
90% de cellules activées [7]. Les anticorps anti-ASGM1 et anti-NK1.1 sont typiquement
utilisés pour la déplétion des cellules NK chez la souris. Cela peut aboutir par mégarde, à
la déplétion simultanée de lymphocytes T exprimant ces marqueurs. Les données des
études testant la déplétion des cellules NK chez la souris nécessitent à cet effet, une
interprétation très prudente.
Les cellules NK représentent approximativement 10 % de cellules mononuclées
du sang périphérique (PBMC), et de 0,4 à 5% de cellules mononuclées des organes
lymphoïdes secondaires [9].
Les mécanismes de cytotoxicité naturelle sont semblables à ceux utilisés par les
lymphocytes T cytotoxiques (CTL) i.e. sécrétion de la protéine formant des pores à la
surface des cellules cibles : la perforine et les protéines induisant l’apoptose : les
granzymes, et/ou l’obligation des récepteurs de mort des cellules cibles [10]. La lyse

4
catalysée par les cellules NK a été démontrée dans plusieurs infections virales humaines
et chez les rongeurs [11], cependant, la signification physiologique de la cytotoxicité
naturelle au cours des infections bactériennes et parasitaires est encore matière à débat
[12, 13]. La seconde fonction effectrice majeure des cellules NK est la sécrétion de
cytokines pro-inflammatoires et de chémokines, tels le facteur activateur des
macrophages : l’IFN-γ, le facteur α de nécrose des tumeurs (TNF-α), la lymphotoxine
(LT-α), le facteur stimulant les colonies de granulocyte-monocytes (GM-CSF) et la
protéine inflammatoire des macrophages CCL3 [14, 15].
L’IFN-γ a un rôle crucial dans la résistance de l’hôte contre plusieurs infections.
Il régule plusieurs centaines de gènes associés aux fonctions du système immunitaire et
draine d’autres cellules effectrices vers le site de l’infection [16]. L’IFN-γ active les
macrophages et les neutrophiles, stimule la différenciation des lymphocytes Th1 CD4+,
augmente la présentation des antigènes par les cellules présentatrices en régulant
positivement le CMH et les molécules associées au CMH ; il induit aussi la commutation
des sous classes d’immunoglobuline G (IgG) dans les lymphocytes B, et de ce fait établit
le lien entre les deux principaux acteurs du système immunitaire notamment, l’immunité
innée et de l’immunité spécifique [16]. Désormais, la sécrétion d’IFN-γ par les cellules
NK est considérée comme ayant un rôle important dans la mobilisation de puissantes
réponses immunes contre des classes variées de pathogènes [13, 17].
Les pathogènes envahissants ou les antigènes viraux, et des protozoaires d’origine
bactérienne sont capturés par les cellules dendritiques (DCs) ou les
monocytes/macrophages qui libèrent ensuite des cytokines activatrices des cellules NK
[18-22]. L’IL-2, l’IL-12, l’IL-15, l’IL-18, le TNF-α et l’IFN-α/β contribuent tous à
l’activation des cellules NK alors que l’IL-4, l’IL-10 et le TGF-β suppriment la fonction
cellulaire de ces mêmes cellules [18, 23]. L’IL-12, qui est une cytokine pro-
inflammatoire principalement produite par des cellules présentatrices d’antigènes
professionnelles, i.e. les macrophages et les cellules dendritiques, est un puissant
inducteur de l’activité cytotoxique des cellules NK, et de la sécrétion d’IFN-γ qui, avec

5
l’IL-18 jouent un rôle crucial dans la mobilisation des réponses immunes catalysées par
les cellules NK [24, 25].
3. Les récepteurs des cellules NK
Alors que la stimulation par les cytokines contribue de façon significative à leur
activation, les cellules NK ont aussi la capacité de détecter directement les cellules
transformées ou les cellules infectées et de répondre immédiatement à de tels stimuli.
L’engagement d’un répertoire élaboré de ligands via un répertoire complexe similaire de
récepteurs de surface permet aux cellules NK de différencier les cellules hôtes normales
des cellules anormales potentiellement dangereuses et cela sans immunisation préalable.
Une fine balance des signaux inhibiteurs et stimulateurs est maintenue en association
avec l’état d’activation des cellules NK [26].
Un aspect de ce processus est la reconnaissance de la perte de soi, « missing-
self », proposé pour la première fois par Klaus Kärre dans les années 1980. Ce processus
par lequel les cellules NK scrutent la surface cellulaire des cellules cibles potentielles, et
s’activent si les cellules cibles ne peuvent pas fournir des signaux suffisamment forts en
retour [27]. Il est maintenant bien compris que cela serait due au fait que, dans les
conditions physiologiques normales, les cellules NK ont besoin de recevoir un signal
négatif fourni par l’engagement des récepteurs inhibiteurs spécifiques au CMH de classe
I en vue de prévenir leur activation [26, 29]. La régulation négative ou la perte des
molécules du CMH de classe I à la surface des cellules cibles peut rompre la balance des
signaux inhibiteurs et activateurs en faveur de la mobilisation de l’activation des cellules
NK.
Plus récemment, il a été reconnu que les cellules NK reconnaîtraient aussi les
modifications du soi, « altered-self » pour lesquelles les cellules NK peuvent aussi être
activées par les cellules exprimant pourtant un complément de CMH de classe I normal
s’il est suffisamment bien engagé dans les récepteurs d’activation NK par les néo
antigènes. Ces modifications peuvent être induites par le stress, la transformation

6
tumorale ou l’infection [29]. Il est maintenant bien connu que les récepteurs activateurs et
inhibiteurs des cellules NK (Voir Tableau 1) reconnaissent un large éventail de molécules
du soi et du non soi. A l’inverse des lymphocytes du système immunitaire spécifique, les
cellules NK n’utilisent pas de réarrangements des gènes somatiques pour générer des
clones ayant la capacité de reconnaissance de divers antigènes ; elles réalisent plutôt leurs
potentialités variées par l’expression simultanée de multiple récepteurs, inhibiteurs et
activateurs ayant différentes spécificités [30, 31].
Il y a trois grandes familles de récepteurs des cellules NK. Les récepteurs
membres de la superfamille des immunoglobulines (G) [(e.g. les récepteurs killer Ig-like)
(KIR), et les récepteurs LIR/ILT, Siglec-7], les récepteurs membres de la famille des
lectines de type C qui sont représentés par les molécules hétéro dimériques
(CD94/NKG2) et les récepteurs de cytotoxicité naturelle (NCR) [31, 32, 33]. Alors que la
plupart des récepteurs présente très peu de polymorphisme, une remarquable diversité a
été trouvée dans le gène de la famille de KIR chez les primates et parmi les gènes du
récepteur équivalent, LY49 chez les rongeurs. Certains récepteurs KIR sont activateurs,
c’est le cas par exemple de KIR2DS et KIR3DS. D’autres comme KIR2DL, KIR2DL4 et
KIR3DL sont des récepteurs inhibiteurs. Les récepteurs KIR inhibiteurs contiennent dans
leur domaine intracytoplasmique des motifs inhibiteurs appelés ITIMs (pour immuno
receptor tyrosine-based inhibitory motifs) tandis que les récepteurs activateurs
contiennent eux des motifs activateurs ITAMs. A chaque ligand du CMH I correspond un
récepteur KIR exprimé à la surface des cellules NK.
Le locus du gène KIR humain sur le chromosome 19q13.4 s’étend sur
approximativement 150 KB codant pour plus de 15 gènes KIR [34]. Les variabilités
haplotypique et allélique sont responsables de la grande hétérogénéité trouvée dans le
génotype de KIR dans une population où chaque individu exprime un ensemble de
caractéristiques de KIR inhibiteurs et activateurs. En outre, des variations dans les loci de
KIR exprimés par différents clones de cellules NK entraînent un répertoire polyclonal
NK au sein d’un même individu. La diversité des génotypes KIR individuel au sein d’une
population est comparable à la diversité trouvée dans les génotypes des antigènes de

7
leucocytes humains (HLA) [35]. Cela suggère que des pressions sélectives similaires de
diversification pourraient être activées dans l’ensemble des loci. Il n’est pas surprenant
que la reconnaissance spécifique des molécules CMH de classe I par des KIR inhibiteurs
forme une part essentielle du mécanisme de discrimination soi/non soi des cellules NK.
A l’inverse, l’identité des ligands physiologiques de KIR activateurs demeure
encore matière à intense investigation. Malgré le fait que les interactions des molécules
HLA avec les KIR activateurs aient été décrites [36], les interactions fonctionnelles avec
des molécules non CMH ne peuvent être exclues.
Les récepteurs polymorphiques lectine de type C de la famille multigène de Ly49
sont des analogues murins du récepteur KIR humain puisqu’ils reconnaissent les
molécules de CMH de classe I. De plus, les molécules de CMH de classe I-like et des
haplotypes individuels de Ly49 varient en nombre de gènes des récepteurs activateurs et
inhibiteurs [33].
La famille des récepteurs de type leucocytaire immunoglobulin like
(LIR/ILT) présente des similarités structurales avec certaines molécules KIR mais
seulement une faible variation allélique et génique n’est conservée à travers les
haplotypes [37, 38]. C’est ainsi par exemple qu’un minimum de récepteurs inhibiteurs
polymorphiques LIR-1/ILT-2 (LILRB1) n’est exprimé sur les cellules NK et fixe une
large gamme de molécules du CMH de classe I et de molécules CMH-I like [38, 39]. La
molécule multigénique NKG2 de la famille des lectines de type C est présente aussi bien
chez l’homme que chez les rongeurs. Les hétérodimères de NKG2A, -B, -C ou –E avec
CD94 sont connus pour reconnaître les molécules CMH-I non classiques (HLA-E/Qa-1)
et contribuent ainsi à la discrimination entre le soi et le non soi [40, 41].
8
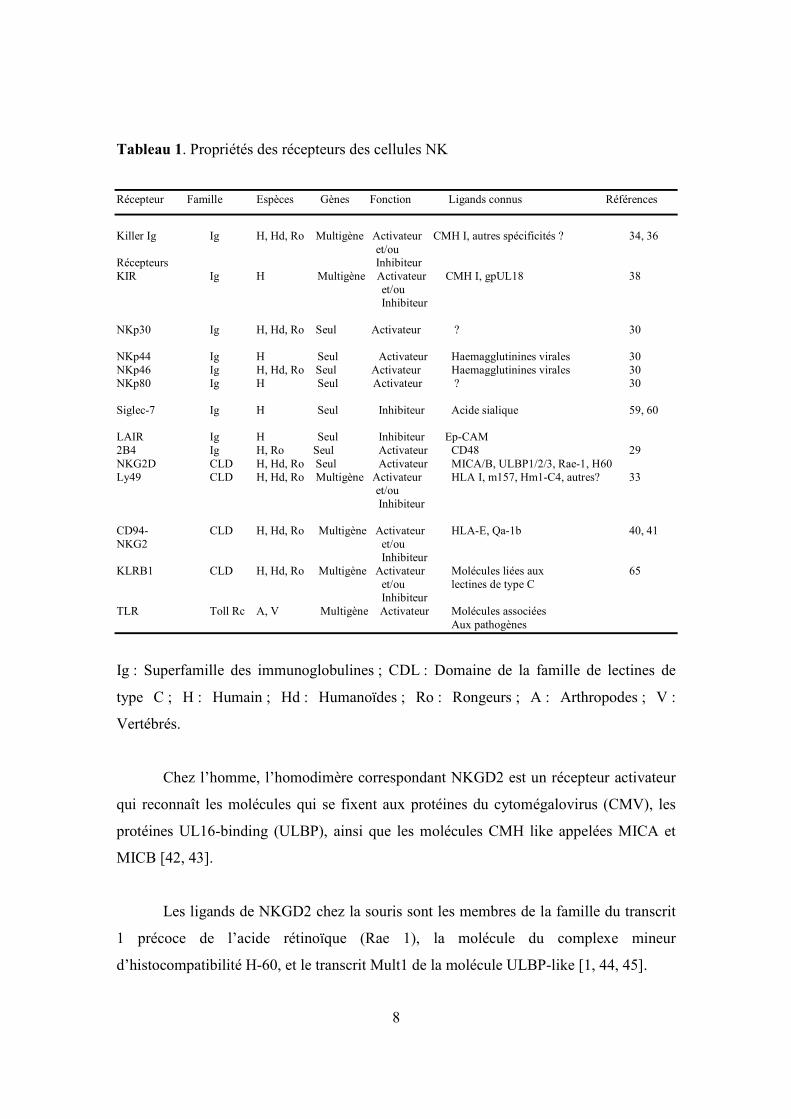
Tableau 1. Propriétés des récepteurs des cellules NK
Récepteur Famille Espèces Gènes Fonction Ligands connus Références
Killer Ig Ig H, Hd, Ro Multigène Activateur CMH I, autres spécificités ? 34, 36 et/ou Récepteurs Inhibiteur KIR Ig H Multigène Activateur CMH I, gpUL18 38 et/ou Inhibiteur NKp30 Ig H, Hd, Ro Seul Activateur ? 30 NKp44 Ig H Seul Activateur Haemagglutinines virales 30 NKp46 Ig H, Hd, Ro Seul Activateur Haemagglutinines virales 30 NKp80 Ig H Seul Activateur ? 30 Siglec-7 Ig H Seul Inhibiteur Acide sialique 59, 60 LAIR Ig H Seul Inhibiteur Ep-CAM 2B4 Ig H, Ro Seul Activateur CD48 29 NKG2D CLD H, Hd, Ro Seul Activateur MICA/B, ULBP1/2/3, Rae-1, H60 Ly49 CLD H, Hd, Ro Multigène Activateur HLA I, m157, Hm1-C4, autres? 33 et/ou Inhibiteur CD94- CLD H, Hd, Ro Multigène Activateur HLA-E, Qa-1b 40, 41 NKG2 et/ou Inhibiteur KLRB1 CLD H, Hd, Ro Multigène Activateur Molécules liées aux 65 et/ou lectines de type C Inhibiteur TLR Toll Rc A, V Multigène Activateur Molécules associées Aux pathogènes
Ig : Superfamille des immunoglobulines ; CDL : Domaine de la famille de lectines de
type C ; H : Humain ; Hd : Humanoïdes ; Ro : Rongeurs ; A : Arthropodes ; V :
Vertébrés.
Chez l’homme, l’homodimère correspondant NKGD2 est un récepteur activateur
qui reconnaît les molécules qui se fixent aux protéines du cytomégalovirus (CMV), les
protéines UL16-binding (ULBP), ainsi que les molécules CMH like appelées MICA et
MICB [42, 43].
Les ligands de NKGD2 chez la souris sont les membres de la famille du transcrit
1 précoce de l’acide rétinoïque (Rae 1), la molécule du complexe mineur
d’histocompatibilité H-60, et le transcrit Mult1 de la molécule ULBP-like [1, 44, 45].

9
Les ligands de NKGD2 ne sont pas généralement exprimés par les tissus
normaux, mais l’expression de MICA, MICB, Rae 1, ULBP et des protéines ULBP-like
est positivement régulée en réponse au stress cellulaire dans les cellules transformées
[44,45, 46-48] et les cellules infectées [49-52] ; c’est ainsi que NKGD2 a été considéré
comme étant impliqué dans l’immunité contre les tumeurs et dans la défense anti-virale
[29]. Le récepteur inhibiteur killer lectine like KLRG1 a été récemment montré comme
pouvant être positivement régulé par l’activation des cellules NK in vivo et pouvant
supprimer la sécrétion d’IFN-γ de la cytotoxicité des cellules NK in vitro, agissant ainsi
comme un avaliseur de l’activité NK [53]. Toutefois, le rôle exact de KLRG1 et l’identité
de ses ligands ne sont pas encore très clairs.
Jusqu’ici, la famille non polymorphique des NCR activateurs est composée de
quatre membres chez la souris et chez l’homme, NKp30, NKp44, NKp46 et NKp80 qui
sont encore tous des récepteurs « orphelins », dans la mesure où leurs ligands ne sont pas
encore connus. Toutefois, Mandelboim et ses collaborateurs ont récemment rapporté
l’engagement de NKp46 par les protéines d’hémagglutinines virales [54], et que NKp30
semblerait être impliqué dans l’activation des cellules NK par les cellules dendritiques ;
le ligand restant cependant à identifier [55]. Les récepteurs NCR sont capables de
déclencher l’activité cytotoxique en dehors de tout contexte de molécule du CMH. Ils ont
été décrits comme étant responsables de la reconnaissance de cellules tumorales.
L’importance des récepteurs de reconnaissance de motif (PRR) dans la régulation
de l’activité des cellules NK est encore très peu comprise. Les récepteurs Toll-like (TLR)
fixent les motifs CpG, les lipopolysaccharides et plus fréquemment des molécules
associées aux pathogènes [56]. Ils sont particulièrement impliqués dans l’activation des
macrophages et des cellules dendritiques et participent dans les réponses immunes
précoces aux infections virales, bactériennes, parasitaires [57]. Néanmoins, puisque les
TLRs exprimés à la surface des cellules NK et sur les lipophosphoglycanes purifiés à
partir des parasites protozoaires ont été trouvés engagés avec des cellules infectées [58],
il est difficile d’écarter la possibilité que l’engagement NK-TLRs ne soit relevant dans

10
l’immunité innée. L’inhibiteur spécifique du récepteur Siglec-7, l’α-2,8-acide di-sialique
a été impliqué dans la reconnaissance des molécules de glycoconjugués non CMH I [59,
60]. La protection de cellules hôtes déficientes en molécules CMH I ou n’en exprimant
que très faiblement (e.g. les neurones ganglionnaires de la racine dorsale et les
érythrocytes) [61, 62], contre la cytotoxicité des cellules NK par la fixation des molécules
du soi au Siglec-7 pourrait être une partie d’un nouveau mécanisme important de la
discrimination entre le soi et le non soi. Il est probable que l’interaction de Siglec-7 avec
les glycoprotéines de surface contenant de l’acide sialique permette à certains parasites
extracellulaires d’échapper à la cytotoxicité NK alors que les pathogènes n’exprimant
pas d’acide sialique (e.g. Trichophyton spp., [63]) seraient sensibles à l’attaque médiée
par les cellules NK.
4. L’activation des cellules NK par les virus et les parasites
pathogènes
Des avancées significatives ont été réalisées au cours de ces deux dernières
décennies dans la compréhension de la fonction des cellules NK dans différentes
maladies infectieuses variées [13, 64]. Bien que la grande majorité des données
proviennent d’études in vitro et in vivo sur les infections virales [18, 65], il y a une
accumulation rapide de preuves soutenant le rôle des cellules NK dans le contrôle des
maladies causées par les protozoaires.
4-1/ Les infections virales
Il est maintenant bien établi que la cytotoxicité des cellules NK et la production
d’IFN-γ joue un rôle crucial dans la résolution des infections causées par le virus de
l’herpès, de papillomavirus et par le virus de l’influenza [18]. De plus, des interactions
des cellules NK avec le virus de l’immunodéficience humaine de type 1 (VIH-1) et le
virus de la leucémie des cellules humaines T (HTLV) ont été démontrées [66, 67].
L’activation des cellules NK au cours des infections virales semble généralement être
médiée de manière indirecte par des cytokines, le plus souvent par l’IL-12 et l’IFN-α/β

11
sécrétées par les monocytes/macrophages [18]. Toutefois, un grand nombre de récentes
études rapportent l’engagement des récepteurs des cellules NK avec les antigènes viraux
in vitro. Par exemple, les protéines d’hémagglutinine des virus de l’influenza et le
complexe hémagglutinine-neuraminidase du virus para influenza sont démontrés comme
étant capables de mobiliser directement la cytotoxicité. Cette cytotoxicité est catalysée
par les cellules NK in vitro par l’interaction spécifique avec le récepteur activateur
NKp46 [54]. Toutefois, la relevance physiologique des interactions directes cellules NK
et virus dans le contrôle de l’infection n’est pas encore complètement démontrée de façon
convaincante in vivo. Plus récemment le groupe de Vidal, Lanier et Yokohama a apporté
une preuve irréfutable que la résistance au cytomégalovirus (CMV) murin dépend de la
stimulation par une glycoprotéine viral CMH like [68-70]. Cela donne une explication
très plausible de la sensibilité différentielles in vivo de différentes souches de souris au
CMV [71].
4-2/ La Leishmaniose
Les cellules NK fournissent aussi la base de la résistance précoce contre la
leishmaniose comme le suggère le fait qu’au cours de l’infection par Leishmania major,
la maladie est plus sévère chez les souris ayant subi une déplétion des cellules NK, et que
la parasitémie de Leishmania amazonensis ne peut pas être efficacement limitée en
absence de cellules NK [72-74].
Bien que la cytotoxicité contre les lignées cellulaires tumorales soit augmentée
chez les animaux résistants, comparé aux animaux sensibles dans l’infection précoce, et
que les macrophages infectés par L. major et L. amazonensis soient détruits par les
cellules NK in vitro, la lyse catalysée par les cellules NK ne semble pas être essentielle à
la résistance, puisque la capacité de contrôler l’infection est seulement faiblement réduite
chez les souris beiges infectées par Leishmania tropica [73, 75-77]. La mutation beige
perturbe sélectivement la voie lytique des cellules NK mais n’a pas modifié la voie de la
sécrétion de cytokines [78]. Pris ensemble, ces données indiquent que la fonction
effectrice des cellules NK dans la leishmaniose est catalysée par la voie à médiation

12
cytokinique plutôt que par celle à médiation cytolytique. Compatible avec cette idée, il a
été montré qu’une production rapide d’IFN-γ au cours des premières heures et des
premiers jours de l’infection par L. major était cruciale pour la survie. De plus les cellules
NK sont la première source de production de cette cytokine [73, 79]. En outre, les souris
sévèrement immunodéprimées (SCID) qui n’ont pas de cellules T mais présentent une
fonction NK normale, sont capables de contenir les parasites de L. major dans le drainage
de ganglions lymphatiques, prouvant l’existence d’un mécanisme indépendant de cellules
T qui limite la propagation des parasites ; la neutralisation de l’IFN-γ ou la déplétion des
cellules NK avant l’infection des souris SCID abolit leur capacité à contrôler la
propagation des parasites [80]. De même, la guérison spontanée et la protection contre la
leishmaniose chez l’homme semblent être associées à la capacité de répondre rapidement
à l’infection à Leishmania aethiopica par la prolifération cellulaire des cellules NK et la
sécrétion de cytokine [81]. La richesse de ces données est le fait que l’activation des
cellules NK au stimulus de la leishmaniose est généralement un phénomène indirect
médié par les cytokines/chémokines, plutôt que par un phénomène direct. L’IL-12 et l’IL-
18 sont les régulateurs majeurs des réponses immunes innée et spécifique contre
l’infection par L. major [25, 82].
Les souris sensibles aux infections par Leishmania major et Leishmania donovani
sont efficacement traitées avec un apport exogène d’IL-12 [83, 84]. L’absence
d’activation des cellules NK par les chémokines conduit en une défense sub-optimale par
les cellules NK [85]. Des études supplémentaires chez les rongeurs ont permis de
suggérer que les cellules dendritiques seraient importantes dans la protection catalysée
par les cellules NK. Elles constitueraient la principale source d’IL-12 précoce au cours de
la leishmaniose, puisqu’une production transitoire de cette cytokine par les cellules
dendritiques dans les premières 24 heures semblerait être l’événement initial requis pour
la mobilisation de l’activation des cellules NK [21, 86, 87].
Il a été récemment démontré que les promastigotes vivants de L. donovani et L.
aethiopiaca stimulent les cellules NK purifiées à secréter l’IFN-γ en absence d’autres
cellules présentatrices d’antigènes [88]. On sait aussi qu’une stimulation directe de TLR-

13
2 de la surface des cellules NK par un lipophosphoglycan (LPG) de L. major entraîne une
régulation positive de TLR-2. Cela augmente la production d’IFN-γ et de TNF-α [58]
suggérant l’existence d’une activation supplémentaire des cellules NK par une voie
indépendante de celle des cellules accessoires. Toutefois, le niveau de production d’IFN-γ
par les cellules NK pures dans cette étude était faible (détectable par ELISPOT mais pas
par ELISA ni par marquage intracellulaire [88]), suggérant que les cytokines provenant
des cellules accessoires pourraient être requises à l’amplification de la réponse directe.
4-3/ La trypanosomiase
L’activité précoce des cellules NK influence aussi le cours de la maladie dans les
trypanosomiases américaines et africaines. La déplétion des cellules NK des souches de
souris résistant à Trypanosoma cruzi provoque une forte parasitémie, une augmentation
de la mortalité très précoce dans l’infection et un décalage dans la production d’IFN-γ par
les cellules T [89-91]. L’analyse ex-vivo des phénotypes de lymphocytes dans la maladie
de Chagas aiguë chez l’homme soutient l’idée que les cellules NK sont activées par T.
cruzi avant que l’immunité des cellules T ne se développe [92].
L’importance des mécanismes de cytotoxicité dans la résistance aux parasites
Trypanosoma est encore discutée. Au début des années 1980, des études ex vivo chez la
souris ont révélé une augmentation de l’activité cytolytique des cellules NK, dans les 24
heures suivant l’infection à T. cruzi, et une destruction directe à médiation NK des formes
épimastigotes et trypomastigotes extracellulaires de T. cruzi et Trypanosoma lewisi in
vitro [93-95]. Curieusement, les parasites de Trypanosoma musculi ne sont pas sensibles
à la lyse par les cellules NK et n’augmentent pas non plus la lyse des lignées cellulaires
tumorales par les cellules NK [95]. Cependant, comme observé pour l’activité des
cellules NK in vivo au cours de l’infection par Leishmania, la mutation beige n’a pas
d’effet majeur sur l’issue de l’infection de T. cruzi [93]. En accord avec cette observation,
une autre étude plus récente indique que la cytotoxicité des cellules NK n’est pas
essentielle au contrôle de l’infection par T. cruzi ni à la survie des souris déficientes en
IFN-α/β [96]. Bien que les souris déficientes, aussi bien pour la voie perforine/granzyme

14
que celle de Fas/sFasL succombent très précocement à l’infection par T. cruzi,
témoignant d’un rôle crucial de la cytotoxicité dans le contrôle de l’infection, il n’est pas
possible, à partir des expériences rapportées dans la littérature, de déterminer si la
cytotoxicité est catalysée par les cellules cytotoxiques T ou par les cellules NK [97].
Comparables aux données sur les mécanismes de résistance dans la leishmaniose précoce,
plusieurs études in vitro et in vivo ont montré que les cellules NK stimulées par l’IL-12
peuvent contrôler la parasitémie en sécrétant l’IFN-γ dans les premiers jours de
l’infection par T. cruzi [89, 91, 98-100]. D’une part, les cellules NK semblent être
activées principalement de façon indirecte par les cytokines produites par les
macrophages et les cellules dendritiques. Le glycosylphosphatidylinositol (GPI) de T.
cruzi et de Trypanosoma brucei active les macrophages de souris in vitro [101, 102, 103]
et la protéine Tc52 libérée par T. cruzi a été montrée comme étant capable d’activer les
cellules dendritiques humaines via TLR-2 [104]. D’autre part, les glycolipides de T. cruzi
stimulent directement l’activité des cellules NK [105]. Il reste à établir quelles sont les
voies de reconnaissance et d’activation qui sont physiologiquement relevant in vivo.
4-4/ La Toxoplasmose
Il est maintenant bien établi que les cellules NK participent aussi dans la
résistance précoce à l’infection par Toxoplasma gondii [106]. La cytotoxicité des cellules
NK contre les cellules tumorales YAC-1 induite par T. gondii est dépendante des
macrophages et les cellules NK présentent une augmentation de la cytotoxicité contre les
macrophages infectés par T. gondii et contre les formes extracellulaires des tachyzoites
[107-110]. Cependant, encore une fois, la cytotoxicité semble être d’une importance
mineure dans la résistance puisque le défaut sélectif de la cytotoxicité des cellules NK
des souris beiges n’aurait pas un effet apparent sur leur survie [111, 112]. A l’inverse,
l’issue sévère de la maladie chez les souris infectées par T. gondii dont les cellules NK
ont été déplétées, laisse supposer une importante fonction protectrice des cellules NK
autre que la fonction cytotoxique [113]. En conséquence, il a été démontré que les
tachyzoites intactes aussi bien que les extraits parasitaires stimulent la production d’IFN-
γ par les cellules NK et que la survie des animaux est entièrement dépendante de cette

15
cytokine [114-117]. Dans les travaux initiaux, il avait été observé qu’au cours de
l’activation indirecte des cellules NK in vitro les étapes de développement extracellulaire
et intracellulaire de T. gondii étaient totalement dépendantes de l’IL-12 produite par les
macrophages [115]. Des travaux plus récents suggèrent aussi un rôle physiologique des
cellules dendritiques dans la reconnaissance de T. gondii et dans la stimulation initiale
des cellules NK [106, 118-120]. De nouveau, les protéines GPI dérivant du parasite
semblent être parmi les facteurs responsables de l’induction de l’IL-2 comme suggéré par
l’activation de la voie de NF-κB dans les macrophages [121].
4-5/ Le Paludisme
Une grande perspective du rôle des cellules NK dans l’immunité contre le
paludisme a été donnée par les études sur les mécanismes liés à l’IL-12 au cours des
stades précoces de la maladie [122]. L’infection des souris A/J normalement sensibles à
Plasmodium chabaudi chabaudi AS (P.chabaudi AS) peut être résorbée de façon
significative par l’administration d’IL-12 exogène très précocement au cours de
l’infection, entraînant le développement de l’immunité dépendant de l’IFN-γ , de TNF-α
et de NO [123]. En outre, la déplétion in vivo des cellules NK, après un traitement avec
un anticorps anti-AGM1 chez la souris C57BL/6 résistant à l’infection par P. chabaudi
AS aboutit en une augmentation de la sévérité de la pathologie. De plus les souris A/J
traitées à l’IL-12 mais déplétées en cellules NK sont totalement incapables de contrôler
l’infection [124]. De même, l’auto-contrôle des infections par P. chabaudi AS et
Plasmodium yoelii est caractérisé par la capacité de l’hôte à mobiliser une forte réponse
IFN-γ dès les 24 heures suivant le challenge [125]. Les animaux privés de leurs cellules
NK ont une diminution significative de la réponse IFN-γ et sont incapables de contrôler
l’infection par P. chabaudi AS. Ils sont de même, incapables de contrôler l’infection par
une souche non létale de P. yoelii. Cela suggère une contribution majeure de l’IFN-γ
dérivé des cellules NK à la résolution de l’infection dans les premiers stades de la
pathologie [125, 126]. Bien que l’interprétation de ces résultats pourrait être compliquée
par l’éventuelle déplétion des souches de cellules T exprimant les marqueurs identiques à
ceux exprimés par les cellules NK (comme expliqué dans l’article « Le paludisme et le

16
système immunitaire humain »), le fait que les souris SCID ayant subi une déplétion de
leurs cellules NK succombent à l’infection non létale par P. yoelii même bien avant les
souris SCID n’ayant subi aucune déplétion, ni les souris traitées avec l’anticorps anti-Thy
1.1 [126], confirme que le rôle des cellules NK ne dépendrait pas des cellules T.
Le rôle in vivo de la cytotoxicité des cellules NK, comme opposé à la production
d’IFN-γ, dans la résistance au paludisme de la souris n’est pas encore très clair. Toutefois
les souris beiges présentent une parasitémie significativement forte dans les stades
précoces de l’infection par Plasmodium berghei, et la lyse catalysée par les cellules NK
des érythrocytes parasités par P. berghei rend les parasites incapables d’infecter
efficacement les souris naïves [12, 122, 127]. Toutes les études citées ci-dessus étaient
réalisées au cours des stades sanguins de l’infection palustre. Toutefois, les sporozoïtes
irradiés par les rayonnements γ sont aussi capables d’induire la production d’IFN-γ in
vitro et in vivo. Les sporozoïtes peuvent aussi entraîner la lyse des cellules tumorales
murines par les cellules NK de la rate dans les 24 heures qui suivent leur inoculation
[128].
Il a été montré plus récemment que l’immunité protectrice dépendant de l’IFN-γ
et de NO et catalysée par les lymphocytes T CD8+ et par la vaccination avec des
sporozoïtes irradiés de P. yoelii ou des constructions d’ADN, requiert à la fois, l’IL-12 et
les cellules NK [129]. Cette étude est un excellent exemple de l’interrelation qui existe
entre l’immunité innée et l’immunité acquise. Au cours de ces dernières années, le rôle
des cellules NK dans le contrôle de l’infection palustre chez l’homme a commencé à être
exploré. Des travaux ont révélé que chez des sujets non immuns, les cellules NK sont
parmi les premières cellules du sang périphérique à produire l’IFN-γ en réponse à
l’infection des érythrocytes par P. falciparum [130].
Dans le cadre de cette étude in vitro, il a été observé que les cellules NK sont
activées au cours des 18 heures d’exposition aux érythrocytes parasités, alors que les
cellules γ/δ T et les cellules NK-T, deux autres populations cellulaires productrices
d’IFN-γ, répondaient plus tard, après 24 ou 48 heures d’exposition au pathogène. Cette

17
activation était dépendant de l’IL-12, et à une moindre mesure de l’IL-18 produits par les
cellules accessoires [130]. Les résultats d’une étude réalisée chez des enfants ayant une
infection aigue à P. falciparum a suggéré une corrélation positive entre l’activité lytique
des cellules NK envers les cellules de la lignée leucémique humaine K562 ex vivo, et le
niveau de parasitémie [131]. Orago et Facer avaient donné la preuve que les cellules NK
de sujets sains et celles des sujets infectés par P. falciparum détruisent directement les
érythrocytes parasités par P. falciparum in vitro [132]. Cette étude avait cependant été
réalisée avec, non pas une population homogène de cellules NK mais avec un mélange
de cellules mononucléaires du sang périphérique encore dénommé PBMC [132].
Nous avons, pour la première fois, clairement démontré le rôle des cellules NK
dans la cytotoxicité des érythrocytes humains parasités par P. falciparum. Nous avons
testé la capacité de lyse des cellules NK purifiées sur des érythrocytes parasités in vitro
par P. falciparum [133]. Dans cette étude, nous avons apporté la preuve que les cellules
NK purifiées inhibaient la croissance des parasites dans des cultures in vitro, et que
l’activité de lyse des cellules NK était probablement réalisée via le système Fas/sFasL et
celui de perforine/granzymes. De plus, nous avons montré que les cellules NK purifiées
stimulées par des érythrocytes parasités par P. falciparum produisent non seulement de
l’IFN-γ, mais aussi du TNF-α et pour la première fois de l’IL-12 [133]. (Ces résultats
seront détaillés dans notre prochain article).
Dans les infections expérimentales de P. falciparum chez des sujets non immuns
réalisées au cours d’une évaluation de vaccin, des taux élevés d’IFN-γ et de granzyme A
soluble ont été détectés au moment de la libération des parasites du foie vers la
circulation générale périphérique, indiquant un possible rôle des cytokines
inflammatoires et de la cytotoxicité, dans la défense initiale de l’organisme contre le
stade sanguin de l’infection in vivo [134]. Les érythrocytes parasités élaborent de grandes
modifications structurales au cours du développement de trophozoïtes et de schizontes
[135] ; ces modifications sont caractérisées par une exposition anormale à la surface de la
membrane des érythrocytes, de protéines telles que la spectrine et la bande 3 [136], et des
néo antigènes exprimés à la surface des parasites [137]. Il est ainsi intéressant de noter

18
que la lyse par les cellules NK, des érythrocytes exprimant à leur surface des antigènes du
stade schizonte de P. falciparum fût significativement plus efficiente que celle des
érythrocytes non parasités. Cela suggère une éventuelle reconnaissance spécifique de la
surface modifiée des érythrocytes par les cellules NK activées [132].
En soutien à cette idée, il a été démontré qu’une activation optimale de la
production d’IFN-γ dérivée des cellules NK in vitro requiert le contact entre les cellules
NK et les érythrocytes parasités par P. falciparum [130, 138]. Ainsi, l’activation des
cellules NK par les stades sanguins asexués de P. falciparum semble dépendre au moins
de deux signaux. La production de cytokines par des cellules « spectatrices » telles les
monocytes/macrophages ou les cellules dendritiques, et la reconnaissance directe des
érythrocytes parasités par les récepteurs des cellules NK (Figure 1).
La capacité de reconnaissance spécifique des érythrocytes pourrait être expliquée
par une expression anormale de ligands de récepteurs activateurs des cellules NK, e.g. les
KIR activateurs, les récepteurs de cytotoxicité naturelle (NCRs), ou des récepteurs Toll-
like, ou alternativement par la régulation négative ou la perte complète d’un ligand des
récepteurs inhibiteurs tel que Siglec-7. De façon très intéressante, la capacité des cellules
NK à répondre aux érythrocytes parasités in vitro par la production d’IFN-γ est un
phénotype stable au niveau individuel de donneurs mais qui varie significativement entre
les individus [130]. Les différences observées dans la réactivité des cellules NK ne sont
pas le résultat de niveau variable de production d’IL-12 et d’IL-18 par les cellules dites
spectatrices puisque l’administration de cytokines exogènes n’augmente pas la réponse
aux érythrocytes parasités par P. falciparum par les cellules NK de sujets non répondeurs
[130].
19
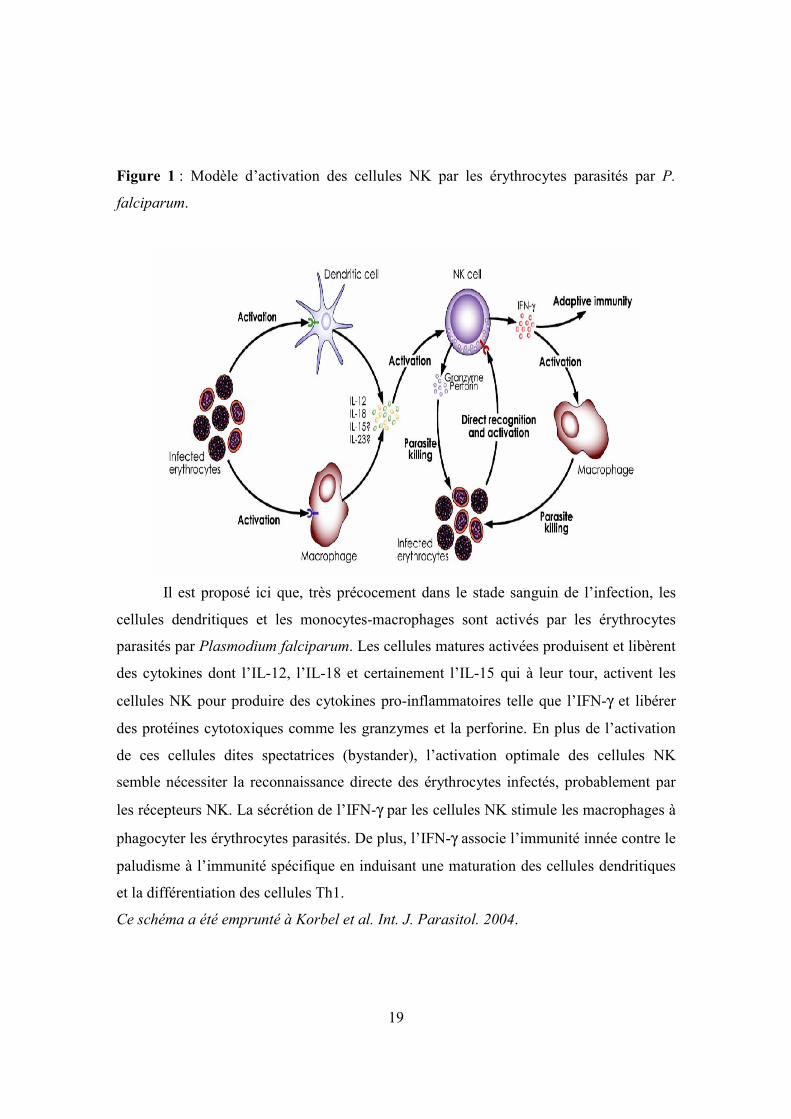
Figure 1 : Modèle d’activation des cellules NK par les érythrocytes parasités par P.
falciparum.
Il est proposé ici que, très précocement dans le stade sanguin de l’infection, les
cellules dendritiques et les monocytes-macrophages sont activés par les érythrocytes
parasités par Plasmodium falciparum. Les cellules matures activées produisent et libèrent
des cytokines dont l’IL-12, l’IL-18 et certainement l’IL-15 qui à leur tour, activent les
cellules NK pour produire des cytokines pro-inflammatoires telle que l’IFN-γ et libérer
des protéines cytotoxiques comme les granzymes et la perforine. En plus de l’activation
de ces cellules dites spectatrices (bystander), l’activation optimale des cellules NK
semble nécessiter la reconnaissance directe des érythrocytes infectés, probablement par
les récepteurs NK. La sécrétion de l’IFN-γ par les cellules NK stimule les macrophages à
phagocyter les érythrocytes parasités. De plus, l’IFN-γ associe l’immunité innée contre le
paludisme à l’immunité spécifique en induisant une maturation des cellules dendritiques
et la différentiation des cellules Th1.
Ce schéma a été emprunté à Korbel et al. Int. J. Parasitol. 2004.

20
Les cellules NK de sujets semi immuns tels des adultes Africains vivant depuis
toujours en zone d’endémie palustre, produisent significativement moins d’IFN-γ en
réponse aux érythrocytes parasités par P.falciparum, que les cellules de sujets caucasiens,
non immuns [130]. L’hypothèse que l’hétérogénéité dans la réponse aux érythrocytes
parasités par P. falciparum serait due aux différences des répertoires exprimés des
récepteurs inhibiteurs ou activateurs des cellules NK a été avancée. Selon cette
hypothèse, cela pourrait refléter une variation haplotypique du génotype de KIR au
niveau individuel, ou d’importants polymorphismes alléliques fonctionnels de KIR et de
TLR [139-141]. Dans ce contexte, il est intéressant de noter que l’analyse génétique d’un
faible nombre d’individus ait révélé une association significative entre les réponses des
cellules NK aux érythrocytes parasités par P. falciparum et le génotype de KIR [138],
suggérant, comme récemment démontré dans la susceptibilité au VIH et au virus de
l’hépatite C [142, 143], qu’il y aurait des différences intrinsèques, génétiquement
déterminées, dans la susceptibilité au paludisme, ce qui pourrait expliquer la variation de
la réponse immune innée. A la suite de la stimulation des PBMC, par des érythrocytes
parasités par P. falciparum, il a aussi été observé une augmentation significative de
l’expression de CD94 et de NKG2A sur les cellules NK des sujets répondeurs, suggérant
l’existence d’une fonction régulatrice pour le récepteur inhibiteur hétérodimérique
CD94 /NKG2A dans la restriction de la pathologie potentielle causée par une activation
continue des cellules NK [138].
Si ces résultats sont confirmés, cela laisserait supposer que l’activation des
cellules NK par les érythrocytes parasités par P. falciparum, et par d’autres protozoaires
pathogènes, est le résultat de l’intégration par les cellules l’ensemble complexe des
signaux émanant à la fois des récepteurs activateurs et inhibiteurs. À cet égard, les stades
finaux de l’activation des cellules NK provoquée par les pathogènes pourraient être
semblables à ceux déjà décrits pour les cellules transformées et les cellules n’exprimant
pas ou peu d’antigènes du CMH I. Toutefois, la confirmation de la fonction protectrice
des cellules NK au cours du paludisme chez l’homme, et la détermination du rôle de KIR

21
ou d’autres polymorphismes dans l’influence de la sensibilité au paludisme nécessite une
évaluation plus importante de populations exposées à l’infection par P. falciparum.
5. Conclusion
Les cellules NK semblent jouer un rôle important dans la réponse immune
précoce contre une grande variété de pathogènes, notamment contre un grand nombre
d’infections parasitaires. Dans tous les cas, l’activation des cellules NK dépend de la
libération de cytokines par les cellules accessoires telles que les macrophages et les
cellules dendritiques et des preuves encourageantes commencent à émerger des
interactions directes entre les protozoaires, ou les cellules infectées par les protozoaires et
les récepteurs exprimées à la surface cellules NK activées. Il est vraisemblable que des
ligands dérivés des pathogènes activateurs des récepteurs des cellules NK existent.
Plusieurs travaux de recherches dans le domaine sont actuellement en cours. Cela
permettra de les identifier dans les prochaines années. L’éventualité que les
polymorphismes, au niveau des récepteurs des cellules NK, et peut être aussi au niveau
des ligands exprimés par les parasites, influenceraient l’issue des interactions cellules
NK-pathogènes, intervenant ainsi dans le cours de l’infection, demeure une possibilité
intéressante.
Références bibliographiques
1. Carayannopoulos LN, Yokoyama WM. Recognition of infected cells by natural killer cells.
Curr. Opin. Immunol. 16, 26-33, 2004. 2. Herberman R, Nunn M, Lavrin D. Natural cytotoxicity reactivity of mouse lymphoid cells
against syngeneic acid allogeneic tumors. I. Distribution of reactivity and specificity. Int. J. Cancer 16:216-229, 1975.
3. Kiessling R, Klein E, Wigzell H. Natural killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur. J. Immunol. 5:112-117, 1975.
4. Wu J, Lanier LL. Natural killer cells and cancer. Adv. Cancer Res. 90:127-156, 2003. 5. Ruggeri L, Capanni M, Tosti A, Urbani E, Posati S, Aversa F, Martelli MF, Velardi A.
Innate immunity against hematological malignancies. Cytotherapy 4:343-346, 2002. 6. Robbins SH, Brossay L. NK cell receptors: emerging roles in host defense against
infectious agents. Microbes Infect. 4:1523-1530, 2002. 7. Slifka MK, Pagarigan RR, Whitton JL, NK markers are expressed on a high percentage of
virus-specific CD8+ and CD4+ T cells. J. Immunol. 164:2009-2015, 2000.

22
8. Assarsson E, Kambayashi T, Sandberg JK, Hong S, Taniguguchi M, Van Kaer L, Ljunggren HG, Chambers BJ. CD8+ T cells rapidly acquire NK1.1 and NK cell-associated molecules upon stimulation in vitro and in vivo. J. Immunol. 165:3673-3679, 2000.
9. Robertson MJ, Ritz J. Biology and clinical relevance of human natural killer cells. Blood 76: 2421-2438, 1990.
10. Lieberman J. The ABCs of granule-mediated cytotoxicity: new weapons in the arsenal. Nat. Rev. Immunol. 3:361-370, 2003.
11. French A, Yokohama W. Natural killer cells and viral infections. Curr. Opin. Immunol. 15: 45-51, 2003.
12. Mohan K, Moulin P, Stevenson M. Natural killer cell cytokine production, not cytotoxicity, contributes to resistance against blood-stage Plasmodium chabaudi AS infection. J. Immunol. 159:4990-4998, 1997.
13. Scharton-Kersten TM, Sher A. Role of natural killer cells in innate resistance to protozoan infections. Curr. Opin. Immunol. 9:44-51, 1997.
14. Trinchieri G. Biology of natural killer cells. Adv. Immunol. 47:187-376, 1989. 15. Robertson MJ. Role of chemokines in the biology of natural killer cells. J. Leukoc. Blood
76:2421-2438, 2002. 16. Boehm U, Klamp T, Groot M, Howard JC. Cellular responses to interferon-gamma. Annu.
Rev. Immunol. 15:749-795, 1997. 17. Shtrichman R, Samuel C. The role of gamma interferon in antimicrobial immunity. Curr.
Opin. Microbiol. 4:251-259, 2001. 18. Biron CA, Nguyen KB, Pien GC, Cousens LP, Salazazar-Mather TP. Natural killer cells in
antiviral defense: function and regulation by innate cytokines. Annu. Rev. Immunol. 17:189-220, 1999.
19. Ferlazzo G, Morandi B, D’Agostino A, Meazza R, Melioli G, Moretta A, Moretta L. The interaction between NK cells and dendritic cells in bacterial infections results in rapid induction of NK cell activation and in the lysis of uninfected dendritic cells. Eur. J. Immunol. 33:306-313, 2003.
20. Lande R, Giacomini E, Grassi T, Remoli ME, Iona E, Miettinen M, Julkunen I, Coccia EM. IFN-alpha beta released by Mycobaterium tuberculosis-infected human dendritic cells induces the expression of CXCL10: selective recruitment of NK and activated T cells. J. Immunol. 170:1174-1182, 2003.
21. Sher A, Pearce E, Kaye P. Shaping the immune response to parasites: role of dendritic cells. Curr. Opin. Immunol. 15:421-429, 2003.
22. Cooper MA, Fehniger TA, Fuchs A, Colonna M, Caligiuri MA. NK cell and DC interactions. Trends Immunol. 25:47-52, 2004.
23. Colucci F, Caligiuri M, Di SJ. What does it take to make a natural killer? Nat. Rev. Immunol. 3:413-425, 2003.
24. Trinchieri G. Interleukin-12: a cytokine at the interface of inflammation and immunity. Adv. Immunol. 70:83-243, 1998.
25. Wei XQ, Leung BP, Niedbala W, Piedrafita D, Feng GJ, Sweet M, Dobbie L, Smith AJ, Liew FY. Altered immune responses and susceptibility to Leishmania major and Staphylococcus aureus infection in IL-18-deficient mice. J. Immunol. 163:2821-2828, 1999.
26. Lanier LL. Natural killer cell receptor signaling. Curr. Opin. Immunol. 15:308-314, 2003. 27. Kärre K. Role of target histocompatibility in regulation of natural killer activity: a
reevaluation and a hypothesis, in: Callewaert D, Herbeman RB. (Eds), Mechanisms of cytotoxicity by NK cells. Academic Press, San Diego, pp. 81-91, 1985.
28. Moretta L, Moretta A. Unravelling natural killer cell function: triggering and inhibitory human NK receptors. Eur. Mol. Biol. J. 23:255-259, 2004.
29. Raulet D. Roles of the NKG2D immunoreceptor and its ligands. Nat. Rev. Immunol. 3:781-790, 2003.
30. Moretta A, Bottino C, Vitale M, Pende D, Cantoni C, Mingari M, Biassoni R, Moretta L. Activating receptors and coreceptors involved in human natural killer cell-mediated cytolysis. Annu. Rev. Immunol. 19:197-223, 2001.

23
31. McQueen K, Parham P: Variable receptors controlling activation and inhibition of NK cells. Curr. Opin. Immunol. 14:615-621, 2002.
32. Shilling HG, Young N, Guethlein LA, Cheng NW, Gardiner CM, Tyan D, Parham P. Genetic control of human NK cell repertoire. J. Immunol. 169: 239-247, 2002.
33. Yokohama W, Plougastel B. Immune functions encoded by the natural killer gene complex. Nat. Rev. Immunol. 3:304-316, 2003.
34. Carrington M, Norman P. The KIR gene cluster. National Library of Medicine, Bethesda, MD.
35. Hsu K, Chida S, Geraghty D, Dupont B. The killer cell immunoglobulin-like receptor (KIR) genomic region: gene-order, haplotypes and allelic polymorphism. Immunol. Rev. 190:40-52, 2002.
36. Winter CC, Gumperz JE, Parham P, Long EO, Wagtmann N. Direct binding and functional transfer of NK cell inhibitory receptors reveal novel patterns of HLA-C allotype recognition. J. Immunol. 161:571-577, 1998.
37. Young N, Canavez F, Uhrberg M, Shum B, Parham P. Conserved organization of the ILT/LIR gene family within the polymorphic human leukocyte receptor complex. Immunogenetics 53:270-278, 2001.
38. Martin AM, Kulski JK, Witt C, Pontarotti P, Christiansen FT. Leukocyte Ig-like receptor complex (LRC) in mice and men. Trends Immunol. 23:81-88, 2002.
39. Natarajan K, Dimasi N, Wang J, Mariuzza RA, Margulies DH. Structure and function of natural killer cell receptors: multiple molecular solutions to self, nonself discrimination. Annu. Rev. Immunol. 20:853-885, 2002.
40. Braud VM, Allan DS, O’Callaghan CA, Soderstrom K, D’Andrea A, Ogg GS, Lazetic S, Young NT, Bell JI, Phillips JH, Lanier LL, McMichael AJ. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 391:795-799, 1998.
41. Vance RE, Kraft JR, Altman JD, Jensen PE, Raulet DH. Mouse CD94/NKG2A is a natural killer cell receptor for the nonclassical major histocompatibility complex (MHC) class I molecule Qa-1(b). J. Exp. Med. 188:1841-1848, 1998.
42. Bauer S, Groh V, Wu J, Steinle A, Philips JH, Lanier LL, Spies T. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 285:727-729, 1999.
43. Cosman D, Mullberg J, Sutherland CL, Chin W, Armitage R, Fanslow W, Kubin M, Chalupny NJ. ULBPs, novel MHC class I-related molecules, bind to CMV glycoprotein UL16 and stimulate NK cytotoxicity through the NKG2D receptor. Immunity 14:123-133, 2001.
44. Cerwenka A, Bakker AB, McClanahan T, Wagner J, Wu J, Philipps JH, Lanier LL. Retinoic acid early inducible genes define a ligand family for the activating NKG2D receptor in mice. Immunity 12:721-727, 2000.
45. Dienfach A, Jamieson AM, Liu SD, Shastri N, Raulet DH. Ligands for the murine NKG2D receptor: expression by tumor cells and activation of NK cells and macrophages. Nat. Immunol. 1:119-126, 2000.
46. Groh V, Rhinehart R, Secrist H, Bauer S, Grabstein K, Spies T. Broad tumor-associated expression and recognition by tumor-derived gamma delta T cells of MICA and MICB. Proc. Natl. Acad. Sci. USA 96:6879-6884, 1999.
47. Girardi M, Oppenheim D, Steele C, Lewis J, Glusac E, Filler R, Hobby P, Sutton B, Tigelaar R, Hayday A. Regulation of cutaneous malignancy by gamma delta T cells. Science 294:605-609, 294, 2001.
48. Jinushi M, Takehara T, Tatsumi T, Kanto T, Groh V, Spies T, Kimura R, Miyagi T, Mochizuki K, Sasaki Y, Hayashi N. Expression and role of MICA and MICB in human hepatocellular carcinomas and their regulation by retinoic acid. Int. J. Cancer 104:354-361, 2003.
49. Das H, Groh V, Kuijl C, Sugita M, Morita C, Spies T, Bukowski J. MICA engagement by human V gamma 2Vdelta 2T cells enhances their antigen-dependent effector function. Immunity 15:83-93, 2001.
50. Groh V, Rhinehart R, Randolph-Habecker J, Topp M, Riddell S, Spies T. Costimulation of CD8 alpha beta T cells by NKG2D via engagement by MIC induced on virus-infected cells. Nat. Immunol. 2:255-260, 2001.

24
51. Tieng V, Le BC, Du ML, Bertheau P, Desreumaux P, Janin A, Charron D, Toubert A. Binding of Escherichia coli adhesin AfaE to CD55 triggers cell-surface expression of the MHC class I-related molecule MICA. Proc. Natl. Acad. Sci. USA 99:2977-2982.
52. Welte S, Sinzger C, Lutz S, Singh-Jasuja H, Sampaio K, Eknigk U, Rammensee H, Steinle A. Selective intracellular retention of virally induced NKG2D ligands by the human cytomegalovirus UL16 glycoprotein. Eur. J. Immunol. 33:194-203, 2003.
53. Robbins SH, Nguyen KB, Takahashi N, Mikayama T, Biron CA, Brossay L. Cutting edge: inhibitory functions of the killer cell lectin-like receptor G1 molecule during the activation of mouse NK cells. J. Immunol. 168:2585-2589, 2002.
54. Mandelboim O, Lieberman N, Lev M, Paul L, Arnon TI, Bushkin Y, Davis DM, Strominger JL, Yewdell JW, Porgador A. Recognition of haemagglutinins on virus-infected cells by NKp46 activates lysis by human NK cells. Nature 409, 1055-1060, 2001.
55. Ferlazzo G, Tsang ML, Moretta L, Melioli G, Steinman RM, Münz C. Human dendritic cells activates resting natural killer (NK) cells and are recognized via the NKp30 receptor by activated NK cells. J. Exp. Med. 195:343-351, 2002.
56. Kopp E, Medzhitov R. Recognition of microbial infection by toll-like receptors. Curr. Opin. Immunol. 15:396-401, 2003.
57. Aderem A, Ulevitch RJ. Toll-like receptors in the induction of the innate immune response. Nature 406:782-787, 2000.
58. Becker I, Salaiza N, Aguirre M, Delgado J, Carrillo-Carrasco N, Kobeh LG, Ruiz A, Cervantes R, Torres AP, Cabrera N, Gonzalez A, Maldonado C, Isibasi A. Leishmania lipophosphoglycan (LPG) activates NK cells through toll-like receptor-2. Mol. Biochem. Parasitol. 130:65-74, 2003.
59. Crocker P. Siglecs: sialic-acid-binding immunoglobulin-like lectins in cell-cell interactions and signaling. Curr. Opin. Struct. Biol. 12:609-615, 2002.
60. Nicoll G, Avril T, Lock K, Furukawa K, Bovin N, Crocker P. Ganglioside GD3 expression on target cells can modulate NK cell cytotoxicity via siglec-7-dependent and –independent mechanisms. Eur. J. Immunol. 33:1642-1648, 2003.
61. Botto M, So A, Giles C, Mason P, Walport M. HLA class I expression on erythrocytes and platelets from patients with systemic lupus erythromatosus, rheumatoid arthritis and from normal subjects. Br. J. Haematol. 75:106-111, 1990.
62. Backström E, Chambers BJ, Ho EL, Naidenko OV, Mariotti R, Fremont DH, Yokoyama WM, Kristensson K, Ljunggren HG. Natural killer cell-mediated lysis of dorsal root ganglia neurons via RAE1/NKG2D interactions. Eur. J. Immunol. 33:92-100, 2003.
63. Esquenazi D, Rozental S, Alviano CS, Travassos LR, Schauer R. Sialic acid are absent from the dermatophytes Trichophyton mentagrophytes and Trichophyton rubrum. Mycoses 46:197-202, 2003.
64. Bancroft GJ. The role of natural killer cells in innate resistance to infection. Curr. Opin. Immunol. 5:503-510, 1993.
65. Yokoyama WM, Seaman WE. The Ly-49 and NKR-P1 gene families encoding lectin-like receptors on natural killer cells: the NK gene complex. Annu. Rev. Immunol. 11:613-635, 1993.
66. Ruscetti FW, Mikovits JA, Kalyanaraman VS, Overton R, Stevenson H, Stromberg K, Herberman RB, Farrar WL, Ortaldo JR. Analysis of effector mechanism against HTLV-I and HTLV-III/LAV-infected lymphoid cells. J. Immunol. 136:3619-3624.
67. Tasca S, Tambussi G, Nozza S, Capiluppi B, Zocchi MR, Soldini L, Veglia F, Poli G, Lazzarin A, Fortis C. Escape of monocyte-derived dendritic cells of HIV-1 infected individuals from natural killer cell-mediated lysis. AIDS 17:2291-2298, 2003.
68. Lee SH, Girard S, Macina D, Busa M, Zafer A, Belouchi A, Gros P, Vidal SM. Susceptibility to mouse cytomegalovirus is associated with deletion of an activating natural killer cell receptor of the C-type lectin superfamily. Nat. Genet. 28:42-45, 2001.
69. Arase H, Mocarski ES, Campbell AE, Hill AB, Lanier LL. Direct recognition of cytomegalovirus by activating and inhibitory NK cell receptors. Science 296:1323-1326, 2002.

25
70. Voigt V, Forbes CA, Tonkin JN, Degli-Esposito MA, Smith HR, Yokoyama WM, Scalzo AA. Murine cytomegalovirus m157 mutation and variation leads to immune evasion of natural killer cells. Proc. Natl. Acad. Sci. USA 100:13483-13488, 2003.
71. Lee SH, Webb JR, Vidal SM. Innate immunity to cytomegalovirus: the Cmv1 locus and its role in natural killer cell function. Microbes Infect. 4:1491-1503, 2002.
72. Lasky T, Rollinghoff M, Solbach W. Natural killer cells participate in the early defense against Leishmania major infection in mice. Eur. J. Immunol. 23:2237-2241, 1993.
73. Scharton TM, Scott P. Natural killer cells are a source of interferon gamma that drives differentiation of CD4+ T cell subsets and induces early resistance to Leishmania major in mice. J. Exp. Med. 178:567-577, 1993.
74. Laurenti MD, Gilund M, Ura DM, Sinhorini IL, Corbett CE, Goto H. The role of natural killer cells in the early period of infection in murine cutaneous leishmaniasis. Braz. J. Med. Biol. Res. 32:323-325, 1999.
75. Kirkpatrick CE, Farrell JP. Leishmaniasis in beige mice. Infect. Immun. 38:1208-1216, 1982.
76. Resnick M, Roguel N, Bercovier H, Enk C, Frankenburg S, Kedar E. Lysis of murine macrophage infected with intracellular pathogens by interleukin 2-activated killer (LAK) cells in vitro. Cell. Immunol. 113:214-219, 1988.
77. Brodskyn CI, Barral A, Boaventura V, Carvalho E, Barral-Netto M. Parasite-driven in vitro human lymphocyte cytotoxicity against autologous infected macrophages from mucosal leishmaniasis. J. Immunol.. 159:4467-4473, 1997.
78. Roder J, Duwe A. The beige mutation in the mouse selectively impairs natural killer cell function. Nature 278:451-453, 1979.
79. Scharton-Kersten T, Scott P. The role of the innate immune response in Th1 cell development following Leishmania major infection. J. Leukoc. Biol. 57:515-522, 1995.
80. Lasky T, Rollinghoff M, Solbach W. Early parasite containment is decisive for resistance to Leishmania major infection. Eur. J. Immunol. 25:2220-2227, 1995.
81. Maasho K, Sanchez F, Schurr E, Hailu A, Akuffo H. Indications of the protective role of natural killer cells in human cutaneous leishmaniasis in an area of endemicity. Infect. Immun. 66:2698-2704, 1998.
82. Scharton-Kersten T, Afonso LC, Wysocka M, Trinchieri G, Scott P. IL-12 is required for natural killer cell activation and subsequent T helper 1 cell development in experimental leishmaniasis. J. Immunol. 154:5320-5330, 1995.
83. Heinzel FP, Schoenhaut DS, Rerko RM, Rosser LE, Gately MK. Recombinant interleukin 12 cures mice infected with Leishmania major. J. Exp. Med. 177:1505-1509, 1993.
84. Murray HW, Hariprashad J. Interleukin 12 is effective treatment for an established systemic intracellular infection: experimental visceral leishmanias. J. Exp. Med. 181:387-391, 1995.
85. Vester B, Müller K, Solbach W, Lasky T. Early gene expression of NK cell-activating chemokines in mice resistant to Leishmania major. Infect. Immun. 67:3155-3159, 1999.
86. Gorak PM, Engwerda CR, Kaye PM. Dendritic cells, but not macrophages, produce IL-12 immediately following Leishmania donovani infection. Eur. J. Immunol. 28:687-695, 1998.
87. Berberich C, Ramirez-Pineda JR, Hambrecht C, Albert G, Skeiky YA, Moll H. Dendritic cell (DC)-based protection against an intracellular pathogen is dependent upon DC-derived IL-12 and can be induced by molecularly defined antigens. J. Immunol. 170:3171-3179, 2003.
88. Nylen S, Maasho K, Soderstrom K, Ilg T, Akuffo H. Live Leishmania promastigotes can directly activate primary human natural killer cells to produce interferon-gamma. Clin. Exp. Immunol. 131:457-467, 2003.
89. Rottenberg M, Cardoni RL, Andersson R, Segura EL, Orn A. Role of T helper/inducer cells as well as natural killer cells in resistance to Trypanosoma cruzi infection. Scand. J. Immunol. 28:573-582, 1988.
90. Sakai T, Hisaeda H, Ishikawa H, Maekawa Y, Zhang M, Nakao Y, Takeuchi T, Matsumoto K, Good RA, Himeno K. Expression and role of heat-shot protein 65 (HSP65) in macrophages during Trypanosoma cruzi infection: involvement of HSP65 in prevention of apoptosis of macrophages. Microbes Infect. 1:419-427, 1999.

26
91. Une C, Andersson J, Eloranta ML, Sunnemark D, Harris RA, Orn A. Enhancement of natural killer (NK) cell cytotoxicity and induction of NK cell-derived interferon-gamma (IFN-gamma) display different kinetics during experimental infection with Trypanosoma cruzi. Clin. Exp. Immunol. 121:499-505, 2000.
92. Sathler-Avelar R, Lemos EM, Reis DD, Medrano-Mercado N, Araujo-Jorge TC, Antas PR, Correa-Oliveira R, Teixeira-Carvalho A, Eloi-Santos SM, Favato D, Martin-Filho OA. Phenotypic features of peripheral blood leucocytes during early stages of human infection with Trypanosoma cruzi. Scand. J. Immunol. 58:655-663, 2003.
93. Hatcher FM, Kuhn RE Cerrone MC, Burton RC. Increased natural killer cell activity in experimental American trypanosomiasis. J. Immunol. 136:313-319, 1981.
94. Hatcher F, Kuhn R. Destruction of Trypanosoma cruzi by natural killer cells. Science 218:295-296, 1982.
95. Albright JW, Hatcher FM, Albright JF. Interaction between murine natural killer cells and trypanosomes of different species. Infect. Immun. 44: 315-319, 1984.
96. Une C, Andersson J, Orn A. Role of IFN-alpha/beta and IL-12 in the activation of natural killer cells and interferon-gamma production during experimental infection with Trypanosoma cruzi. Clin. Exp. Immunol. 134:195-201, 2003.
97. Müller U, Sobeck V, Balkow S, Hölscher C, Müllbacher A, Museteanu C, Mossmann H, Simon MM. Concerted action of perforin and granzymes is crucial for the elimination of Trypanosoma cruzi from mouse tissues, but prevention of early host death is an addition dependent on the FasL/Fas pathway. Eur. J. Immunol. 33:70-78, 2003.
98. Aliberti JC, Cardoso MA Martins GA, Gazzinelli RT, Vieira LQ, Silsa JS. Interleukin-12 mediates resistance to Trypanosoma cruzi in mice and is produced by murine macrophages in response to live trypomastigotes. Infect. Immun. 64:1961-1967, 1996.
99. Cardillo F, Voltarelli JC, Reed SG, Silva JS. Regulation of Trypanosoma cruzi infection in mice by gamma interferon and interleukin 10: role of NK cells. Infect. Immun. 64:128-134, 1996.
100. Hunter CA, Slifer T, araujo F. Interleukin-12-mediated resistance to Trypanosoma cruzi is dependent on tumor necrosis factor alpha and gamma interferon. Infect. Immun. 64:2381-2386, 1996.
101. Tachado SD, Schofield L. Glycosylphosphatidylinositol toxin of Trypanosoma brucei regulates IL-1 alpha and TNF-alpha expression in macrophages by protein tyrosine kinase mediated signal transduction. Biochem. Biophys. Res. Commun. 205:984-991, 1994.
102. Carmago M, Almeida I, Pereira M, Fergusson M, Travassos L, Gazzinelli R. Glycosylphosphatidylinositol-anchored mucin-like glycoproteins isolated from Trypanosoma cruzi trypomastigotes initiate the synthesis of proinflammatory cytokines by macrophages. J. Immunol. 158:5890-5901, 1997.
103. Carmago M, Andrade A, Almeida I, Travassos L, Gazzinelli R. Glycoconjugates isolated from Trypanosoma cruzi but not from Leishmania species membranes trigger nitric oxide synthesis as well as microbicidal activity in IFN-gamma-primed macrophages. J. Immunol. 159:613-6139, 1997.
104. Ouassi A, Guilvard E, Delneste Y, Caron G, Magistrelli G, Herbault N, Thieblemont N, Jeannin P. The Trypanosoma cruzi Tc52-released protein induces human dendritic cell maturation, signals via Toll-like receptor 2, and confers protection against lethal infection. J. Immunol. 168:6366-6374, 2002.
105. De Arruda Hinds LB, Previato LM, Previato JO, Vos Q, Mond JJ, Pecanha LM. Modulation of B-lymphocyte and NK cell activities by glycoinositolphospholipid purified from Trypanosoma cruzi. Infect. Immun. 67:6177-6180, 1999.
106. Sher A, Collazo C, Scanga C, Jankovic D, Yap G, Aliberti J. Induction and regulation of IL-12-dependent host resistance to Toxoplasma gondii. Immunol. Res. 27:521-528, 2003.
107. Hauser Jr. WE, Tsai V. Acute Toxoplasma infection of mice induces spleen NK cells that are cytotoxic for T. gondii in vitro. J. Immunol. 136:313-319, 1986.

27
108. Kamiyama T, Hagiwara T. Augmented followed by suppressed levels of natural cell-mediated cytotoxicity in mice infected with Toxoplasma gondii. Infect. Immun. 36:628-636, 1982.
109. Hauser Jr. WE, Sharma SD, Remington JS. Natural killer cells induced by acute and chronic Toxoplasma infection. Cell. Immunol. 69:330-346, 1982.
110. Subauste CS, Dawson L, Remington JS. Human lymphokine-activated killer cells are cytotoxic against cells infected with Toxoplasma gondii. J. Exp. Med. 176:1511-1519, 1992.
111. Hughes HP, Kasper LH, Little J, Dubey JP. Absence of a role for natural killer cells in the control of acute infection by Toxoplasma gondii oocysts. Clin. Exp. Immunol. 72:394-399, 1988.
112. Jonhson LL, Sayles PC. Strong cytolytic activity of natural killer cells is neither necessary nor sufficient for preimmune resistance to Toxoplasma gondii infection. Nat. Immun. 14:209-215, 1995.
113. Hunter CA, Candolfi E, Subauste C, Van Cleave V, Remington JS. Studies on the role of interleukin-12 in acute murine toxoplasmosis. Immunology 84:16-20, 1995.
114. Denkers EY, Gazzinelli RT, Martin D, Sher A. Emergence of NK1.1+ cells as effectors of IFN-gamma dependent immunity to Toxoplasma gondii in MHC class I-deficient mice. J: Exp. Med. 178:1465-1472, 1993.
115. Gazzinelli RT, Hieny S, Wynn TA, Wolf S, Sher A. Interleukin 12 is required for the T-lymphocyte-independent induction of interferon gamma by an intracellular parasite and induces resistance in T-cell-deficient hosts. Proc. Natl. Acad. Sci. USA 90:6115-6119, 1993.
116. Sher A, Oswald IP, Hieny S, Gazzinelli RT. Toxoplasma gondii induces a T-independent IFN-gamma response in natural killer cells that requires both adherent accessory cells and tumor necrosis factor-alpha. J. Immunol. 150:3982-3989, 1993.
117. Scharton-Kersten TM, Wynn TA, Denkers EY, Bala S, Grunvald E, Hieny S, Gazzinelli RT, Sher A. In the absence of endogenous IFN-gamma, mice develop unimpaired IL-12 responses to Toxoplasma gondii while failing to control acute infection. J. Immunol. 157:4045-4054, 1996.
118. Ries e Sousa C, Sher A, Kaye P. The role of dendritic cells in the induction and regulation of immunity to microbial infection. Curr. Opin. Immunol. 113:214-219, 1999.
119. Aliberti J, Reis e Sousa C, Schito M, Hieny S, Wells T, Huffnagle GB, Sher A. CCR5 provides a signal for microbial induced production of IL-12 by CD8 alpha+dendritic cells. Nat. Immunol. 1:83-87, 2000.
120. Scanga CA, Aliberti J, Jankovic D, Tilloy F, Bennouna S, Denkers EY, Medzhitov R, Sher A. Cutting edge: MyD88 is required for resistance to Toxoplasma gondii infection and regulates parasite-induced IL-12 production by dendritic cells. J. Immunol. 168:5997-60011, 2002.
121. Debierre-Grockiego F, Azzouz N, Schmidt J, Dubremetz JF, Geyer H, Geyer R, Weingart R, Schmidt RR, Schwarz RT. Roles of glycosylphosphatidylinositols of Toxoplasma gondii: induction of tumor necrosis factor alpha production in macrophages. J. Biol. Chem. 278:32987-32993, 2003.
122. Stevenson MM, Su Z, Sam H, Mohan K. Modulation of host responses to blood-stage malaria by interleukin-12: from therapy to adjuvant activity. Microbes Infect. 3:49-59, 2001.
123. Stevenson MM, Tam MF, Wolf SF, Sher A. IL-12-induced protection against blood-stage Plasmodium chabaudi AS requires IFN-gamma and TNF alpha and occurs via a nitric oxide-dependent mechanism. J. Immunol. 155:2545-2556, 1995.
124. Mohan K, Moulin P, Stevenson M. Natural killer cell cytokine production and not cytotoxicity, contributes to resistance against blood-stage Plasmodium chabaudi AS infection. J. Immunol. 159:4990-4998.
125. De Souza JB, Williamson KH, Otani T, Playfair JH. Early gamma interferon responses in lethal and nonlethal murine blood-stage malaria. Infect. Immun. 65:1593-1598, 1997.

28
126. Choudhury H, Sheikh N, Bancroft G, Katz D, De Souza JB. Early nonspecific immune responses and immunity to blood-stage nonlethal Plasmodium yoelii malaria. Infect. Immun. 68:6127-6132, 2000.
127. Solomon J. Natural cytotoxicity for Plasmodium berghei in vitro by spleen cells from susceptible and resistant rats. Immunology 59:277-281, 1986.
128. Ojo-Amaize E, Vilcek J, Cochrane A, Nussenzweig R. Plasmodium berghei sporozoites are mitogenic for murine T cells, induce interferon and activate natural killer cells. J. Immunol. 133:1005-1009, 1984.
129. Doolan D, Hoffman S. IL-12 and NK cells are required for antigen-specific adaptive immunity against malaria initiated by CD8+ T cells in the Plasmodium yoelii model. J. Immunol. 163:884-892, 1999.
130. Artavanis-Tsakonas K, Riley E. Innate immune response to malaria: rapid induction of IFN-gamma from human NK cells by live Plasmodium falciparum-infected erythrocytes. J. Immunol. 169:2956-2963, 2002.
131. Ojo-Amaize E, Salimonu L, Williams A, Akinwolere O, Shabo R, Alm G, Wigzell H. Positive correlation between degree of parasitémie interferon titers, and natural killer cell activity in Plasmodium falciparum-infected children. J. Immunol. 127:2296-2300, 1981.
132. Orago A, facer C. Cytotoxicity of human natural killer (NK) cell subsets for Plasmodium falciparum erythrocytic schizonts: stimulation by cytokines and inhibition by neomycin. Clin. Exp. Immunol. 86:22-29, 1991.
133. Mavoungou E, Luty A, Kremsner PG. Natural killer (NK) cell-mediated cytolysis of Plasmodium falciparum-infected human red blood cells in vitro. Eur. Cytokine. Netw. 14:134-142, 2003.
134. Hermesen CC, Konijnenberg Y, Mulder L, Loe C, van Deuren M, van der Meer JW, van Mierlo GJ, Eling WM, Hack CE, Sauerwein RW. Circulating concentrations of soluble granzyme A and B increase during natural and experimental Plasmodium falciparum infections. Clin. Exp. Immunol. 132:467-472, 2003.
135. Cooke BM, Mohandas N, Coppel RL. The malaria-infected red blood cell: structural and functional changes. Adv. Parasitol. 50:1-86, 2001.
136. Crandall I, Sherman IW. Cytoadherence-related neoantigens on Plasmodium falciparum (human malaria)-infected human erythrocytes result from the exposure of normally cryptic regions of the band 3 protein. Parasitology 108 (Pt 3), 257-267, 1994.
137. Craig A, Scherf A. Molecules on the surface of the Plasmodium falciparum infected erythrocyte and their role in malaria pathogenesis and immune evasion. Mol. Biochem. Parasitol. 115:129-143, 2001.
138. Artavanis-Tsakonis K, Riley E. Innate immune response to malaria: rapid induction of IFN-gamma from human NK cells by live Plasmodium falciparum-infected erythrocytes. J. Immunol. 169:2956-2963, 2003.
139. Uhrberg M, Valiante N, Shum B, Shilling H, Lienert-Wiedenback K, Corliss B, Tyan D, Lanier LL, Parham P. Human diversity in killer cell inhibitory receptor genes. Immunity. 7:753-763, 1997.
140. Arbour N, Lorenz E, Schutte B, Zabner J, Kline J, Jones M, Frees K, Watt J, Schwartz D. TLR4 mutations are associated with endotoxin hyporesponsiveness in humans. Nat. Genet. 25:187-191, 2000.
141. Rajalingam R, gardiner CM, Canavez F, Vilches C, Parham P. Identification of seventeen novel KIR variants : fourteen of them from two non-Caucasian donors. Tissue antigens 57:22-31, 2001.
142. Martin M, Gao X, Lee J, Nelson G, Detels R, Goedert J, Buchbinder S, Hoots K, Vlahov D, Trowsdale J, Wilson M, O’Brien S, Carrington M. Epistatic interaction between KIR3DS1 and HLA-B delays the progression to AIDS. Nat. Genet. 31:429-434, 2002.
143. Khakoo SI, Thio CL, Martin MP, Brooks CR, Gao X, Cheng J, Goedert JJ, Vlahov D, Hilgartner M, Cox S, Little AM, Alexander GJ, Cramp ME, O’Brien SJ, Rosenberg WMC, Thomas DL, Carrington M. HLA and NK cell inhibitory genes in resolving genes in resolving hepatitis C virus infection. Science 305:872-874, 2004.