Le ribosome à l’échelle atomique - Accueil...
6
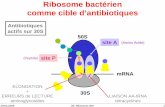
771 Le ribosome à l’échelle atomique Q ui se souvient de l’histoire de la découverte du ribosome [1] et regarde la structure de sa sous-unité 50S à une résolution de 2,4 angströms (Å) peut mesurer le chemin parcouru (figure 1). Après la découverte de particules ribonucléo- protéiques dans le cytoplasme des cellules par Palade [2], il a fallu démontrer que les ribosomes sont le siège de la synthèse protéique [3], puis connaître tous les composants – plus de 50 protéines et trois molé- cules d’ARN – de ce complexe de 2,5 millions de daltons, et enfin com- prendre comment ce complexe décode l’information génétique. La combinaison d’analyses fonction- nelles et structurales a révélé les dif- férentes étapes du processus de tra- duction ainsi que le rôle central qu’y joue l’ARN ribosomique (ARNr). Une fois la séquence des ARN riboso- miques connue, la biochimie structu- rale et la génétique ont permis d’éta- blir la structure secondaire fine de ces ARN et de mettre à jour des inter- actions fonctionnelles à plus grande distance. En même temps, l’accumu- lation des données biochimiques et les progrès de la biologie moléculaire sont venus à l’appui des modèles structuraux, liant ainsi la complexité des structures à l’inventaire des fonc- tions du ribosome. Le ribosome est composé de deux sous-unités, les sous-unités 30S et 50S chez les bacté- ries. De façon schématique, une fonction principale peut être attri- buée à chaque sous-unité : le choix de l’ARN de transfert (ARNt) et le décodage de l’information portée par l’ARN messager (ARNm) pour la sous-unité 30S, la formation de la liai- son peptidique entre les acides ami- nés portés par les ARNt pour la sous- unité 50S (figure 2). La cryo-microscopie électronique et l’analyse cristallographique à haute résolution révèlent aujourd’hui les moindres détails de la structure du ribosome bactérien, apportant une vision précise des bases moléculaires m/s n° 6-7, vol. 17, juin-juillet 2001 MINI-SYNTHÈSE médecine/sciences 2001 ; 17 : 771-6 En moins de trois ans, une dizaine d’études fondées sur la diffraction de cristaux aux rayons X, sur la résonance magnétique nucléaire et sur la cryo- microscopie électronique ont permis d’obtenir une résolution à l’échelle atomique de la structure du ribosome bactérien et de comprendre les bases moléculaires de la synthèse des protéines. Il s’en dégage que l’ARN ribosomique joue un rôle prépondérant dans la structure du ribosome ainsi que dans la formation de la liaison peptidique et le décodage du code génétique. Ces résultats ouvrent la voie à la conception de nouveaux antibiotiques dirigés contre le ribosome bactérien et à d’autres thérapeutiques ayant pour cible la synthèse protéique. Figure 1. Structure à une résolution de 2,4 Å de la sous-unité 50S du ribo- some de l’archaébactérie Haloarcula marismortui. Vue de la face interne, l’ARNr 23S est en bleu clair.
Transcript of Le ribosome à l’échelle atomique - Accueil...

771
Le ribosome à l’échelle atomique
Qui se souvient de l’histoire dela découverte du ribosome [1]et regarde la structure de sa
sous-unité 50S à une résolution de2,4 angströms (Å) peut mesurer lechemin parcouru (figure 1). Après ladécouverte de particules ribonucléo-protéiques dans le cytoplasme descellules par Palade [2], il a falludémontrer que les ribosomes sont lesiège de la synthèse protéique [3],puis connaître tous les composants – plus de 50 protéines et trois molé-cules d’ARN – de ce complexe de2,5 millions de daltons, et enfin com-prendre comment ce complexedécode l’information génétique. Lacombinaison d’analyses fonction-nelles et structurales a révélé les dif-férentes étapes du processus de tra-duction ainsi que le rôle central qu’yjoue l’ARN ribosomique (ARNr).Une fois la séquence des ARN riboso-miques connue, la biochimie structu-rale et la génétique ont permis d’éta-blir la structure secondaire fine deces ARN et de mettre à jour des inter-actions fonctionnelles à plus grandedistance. En même temps, l’accumu-lation des données biochimiques etles progrès de la biologie moléculairesont venus à l’appui des modèlesstructuraux, liant ainsi la complexitédes structures à l’inventaire des fonc-tions du ribosome. Le ribosome estcomposé de deux sous-unités, lessous-unités 30S et 50S chez les bacté-ries. De façon schématique, unefonction principale peut être attri-buée à chaque sous-unité : le choixde l’ARN de transfert (ARNt) et le
décodage de l’information portéepar l’ARN messager (ARNm) pour lasous-unité 30S, la formation de la liai-son peptidique entre les acides ami-nés portés par les ARNt pour la sous-unité 50S (figure 2).
La cryo-microscopie électronique etl’analyse cristallographique à hauterésolution révèlent aujourd’hui lesmoindres détails de la structure duribosome bactérien, apportant unevision précise des bases moléculaires
m/s n° 6-7, vol. 17, juin-juillet 2001
MINI-SYNTHÈSEmédecine/sciences 2001 ; 17 : 771-6
En moins de trois ans, une dizaine d’études fondées surla diffraction de cristaux aux rayons X, sur larésonance magnétique nucléaire et sur la cryo-microscopie électronique ont permis d’obtenir unerésolution à l’échelle atomique de la structure duribosome bactérien et de comprendre les basesmoléculaires de la synthèse des protéines. Il s’en dégage
que l’ARN ribosomique joue un rôle prépondérant dansla structure du ribosome ainsi que dans la formation dela liaison peptidique et le décodage du code génétique.Ces résultats ouvrent la voie à la conception denouveaux antibiotiques dirigés contre le ribosomebactérien et à d’autres thérapeutiques ayant pour cible lasynthèse protéique.
Figure 1. Structure à une résolution de 2,4 Å de la sous-unité 50S du ribo-some de l’archaébactérie Haloarcula marismortui. Vue de la face interne,l’ARNr 23S est en bleu clair.

du mécanisme de traduction etconfirmant le rôle prépondérant del’ARNr dans l’activité du ribosome.Dans cette revue, nous décrirons suc-cessivement la structure de la sous-unité 50S et le mécanisme de forma-tion de la liaison peptidique, lastructure de la sous-unité 30S et sesrelations avec le décodage et les anti-biotiques, puis, à partir de la struc-ture à 7,8 et 5,5 Å de résolution duribosome entier (70S) associé auxARNt et à l’ARNm, nous présente-rons le modèle actuel décrivant lesinteractions entre ces trois parte-naires. Nous essayerons, pourconclure, de lire l’avenir de la tra-duction dans cette boule de cristal.
La structure de la sous-unité 50Set la liaison peptidique
La vision initiale de la sous-unité 50Sétait celle d’un squelette d’ARNorganisant l’arrangement de pro-téines possédant soit une activitécatalytique, soit une fonction de liai-son des différents facteurs de traduc-tion. La résolution à l’échelle ato-mique (2,4 Å) de la structure de lasous-unité 50S [4] permet mainte-nant de localiser 2 711 des 2 923nucléotides de l’ARNr 23S, les122 nucléotides de l’ARNr 5S et 27des 30 protéines. Deux résultats sedégagent de cette structure tridi-mensionnelle : l’un est la confirma-tion éblouissante de la place prépon-dérante de l’ARNr, et l’autre,
attendu, est la démonstration del’activité ribozyme de l’ARNr 23S.La structure secondaire de l’ARNr23S réalisée par Noller et al. [5] mon-trait une organisation en sixdomaines connectés par des régionssouples. La structure tridimension-nelle révèle un grand nombred’interactions entre les hélices desdifférents domaines donnant àchaque domaine une structure ter-tiaire irrégulière. De plus, cesdomaines irréguliers sont imbriquésen une masse d’ARN compacte for-mant un unique domaine monoli-thique (figure 1). Tout aussi éton-nante est la structure des protéinesribosomales ainsi que leur répartitionà la surface de cette masse d’ARN.Parmi les 30 protéines de la sous-unité 50S, 17 ont des structures glo-bulaires que l’on pourrait qualifierde classiques et 13 ont, soit une struc-ture globulaire prolongée par desextensions, soit une forme entière-ment allongée. Le plus remarquableest que les domaines allongés de cesprotéines manquent souvent d’unestructure tertiaire définissable et par-fois même de structure secondaire.L’étrange manque de structure deces extensions pourrait être expliquépar leurs nombreuses interactionsavec l’ARN. Toutes ces protéines sontréparties de façon uniforme à la sur-face de l’ARNr, comblant les trous etles crevasses, comme un mortierentre des briques d’ARN, pourreprendre l’expression des auteurs
[4]. Elles couvrent la surface externe(le dos et le bas) de la sous-unité 50S,tapissant en particulier la sortie dutunnel où passe la chaîne polypepti-dique néosynthétisée. Les six pro-téines qui entourent ce tunnel pour-raient jouer un rôle important dansle mécanisme de sécrétion assurantl’interface avec la membrane du réti-culum et l’appareil de translocationau moment de la synthèse des pro-téines sécrétées. Les extensions desprotéines ribosomiques pénètrentprofondément entre les hélices del’ARNr engageant de nombreuxcontacts avec ses différents domaines.La protéine L22, par exemple, entreen contact avec les six domaines del’ARNr 23S. De même, la plupart desdomaines de l’ARNr 23S interagis-sent avec plusieurs des protéinesribosomiques, le domaine V inter-agissant à lui seul avec 15 protéines.Au total, plus d’un tiers des nucléo-tides de l’ARNr 23S a un contact,sous forme de liaison de van derWaals, avec une protéine riboso-mique. Toutes ces interactionscontribuent à former une structuredense et rigide. Si le rôle des pro-téines qui établissent de nombreuxcontacts avec l’ARNr semble être destabiliser la structure tertiaire des dif-férents domaines, les protéines quin’ont de contact qu’avec un seuldomaine de l’ARNr, ou qui interagis-sent avec d’autres protéines riboso-miques, pourraient participer direc-tement au mécanisme de la synthèseprotéique, ou assurer la fixation desdifférents facteurs de traduction. Detoute la surface de la sous-unité 50S,seuls restent découverts la région decontact avec la sous-unité 30S et lesillon contenant le centre peptidyltransférase, véritable site actif de lasous-unité 50S.Le second résultat remarquable lié àl’élucidation de la structure ato-mique de la sous-unité 50S est que ledomaine V de l’ARNr 23S est respon-sable de l’activité peptidyl transférasedu ribosome (figure 3), en d’autrestermes, que l’ARNr 23S est un ribo-zyme dont l’activité catalytique per-met la synthèse de la liaison pepti-dique [6]. Plusieurs découvertes ontamené ce résultat : la conservation del’activité peptidyl transférase du ribo-some après déprotéinisation exten-
772 m/s n° 6-7, vol. 17, juin-juillet 2001
Figure 2. Représentation schématique du ribosome. L’aminoacyl-ARNt estlocalisé au site A (site aminoacyl), le peptidyl-ARNt qui porte le polypeptideen cours de synthèse au site P (site peptidyl) et l’ARNt désacylé au site E(pour exit).

sive [7], l’activité aminoacylestérasedu ribozyme de Tetrahymena qui équi-
vaut à la formation d’une liaison pep-tidique [8], et l’appariement de
l’extrémité CCA de l’ARNt avec lesnucléotides 2252 et 2253 du domaineV de l’ARNr 23S qui est situé sur laface interne de la grande sous-unité àl’entrée du tunnel [9]. La confirma-tion de l’activité peptidyl transférasede l’ARNr 23S a été obtenue par cris-tallisation de sous-unités 50S com-plexées avec des analogues d’aminoa-cyl-ARNt tels que le CCdA-phosphate-puromycine [6]. Cetteétude montre, d’une part, l’environ-nement nucléotidique autour desanalogues d’aminoacyl-ARNt com-plexés et d’autre part la relativementgrande distance (environ 18 Å) desprotéines ribosomiques avoisinantes.Le nucléotide le plus proche del’extrémité réactive de ces analoguesest l’adénine 2451, située au cœur dudomaine V. Cette base est conservéedans les ribosomes de tous les orga-nismes comme la plupart des basesqui vont jouer un rôle dans le sitepeptidyl transférase. Le mécanismede formation de la liaison peptidiquedécrit sur la figure 3 repose sur leprincipe d’une catalyse de type acide-base. L’environnement de l’azote N3de l’adénine 2451 fait que son pKaest de plus de 7 alors qu’il est ordi-nairement inférieur à 3. Cette éléva-tion locale du pKa permet à l’azoteN3 de fonctionner comme une baseet d’attirer un proton du groupe NH2de l’acide aminé porté par l’ARNtfixé au site A du ribosome. L’azotede ce groupe attaque alors la fonc-tion ester du peptidyl-ARNt fixé ausite P ce qui aboutit à la formation dela liaison peptidique. La résolutionatomique de la structure de la sous-unité 50S a permis de montrer quetous ces constituants, azote N3 del’adénine 2451, oxygène du groupecarbonyle du peptidyl-ARNt et fonc-tion amine de l’aminoacyl-ARNt, setrouvent à 3 Å environ les uns desautres, autorisant ainsi le transfert deprotons. Le polypeptide néosynthé-tisé s’engage dans un tunnel pourressortir sur la face externe du ribo-some (figure 3). Ce tunnel de 100 Åde long a une largeur moyenne de15 Å qui interdit tout autre replie-ment de la chaîne polypeptidiqueque celui en hélice α.On voit à travers cette très brève des-cription combien la connaissance dela structure atomique de la sous-unité
773m/s n° 6-7, vol. 17, juin-juillet 2001
Figure 3. Le centre peptidyl transférase de la sous-unité 50S. (A) Formationde la liaison peptidique par l’activité ribozyme de l’ARNr 23S. L’environne-ment de l’azote N3 de l’adénine 2451 fait que son pKa est de plus de 7. Cetteélévation du pKa permet à l’azote N3 de fonctionner comme une base etd’attirer un proton du groupe NH2 de l’acide aminé porté par l’ARNt fixé ausite A. L’azote de ce groupe attaque alors la fonction carbonyle du peptidyl-ARNt fixé au site P et se lie à l’atome de carbone. Le proton de l’azote N3 sta-bilise la structure en formant une liaison hydrogène avec l’oxyanion portépar le carbone. Le proton est transféré de l’azote N3 sur l’oxygène en 3’ del’ARNt et permet la formation de la liaison peptidique. (B) Structure secon-daire du domaine V de l’ARNr 23S montrant la région de l’adénine 2451(entourée en rouge) et la région de liaison des deux C de l’extrémité 3’ CCAdu peptidyl-ARNt fixé au site P. (C) Coupe de la sous-unité 50S montrant lepassage du peptide néosynthétisé (en bleu) à travers le tunnel. À l’entrée dutunnel se trouve le centre peptidyl transférase (PT) [6].

50S contribue à la compréhensiondu mécanisme de la synthèse pro-téique. Le rôle joué par l’ARNr 23Sdans cette structure aussi bien quedans la synthèse protéique est unargument de poids en faveur del’idée d’un monde primitif à ARN.
La sous-unité 30Set la lecture de l’ARN messager
L’une des fonctions essentielles duribosome est la lecture précise et effi-cace du message inscrit sur l’ARNm.Le décryptage du code génétique estassuré par la sous-unité 30S. Récem-ment, deux équipes de cristallo-graphes ont déterminé par diffrac-tion aux rayons X la structure de lasous-unité 30S de Thermus thermophi-lus à 3 Å de résolution [10-13].La sous-unité 30S est composéed’une molécule d’ARNr 16S(1 518 nucléotides) et de 20 pro-téines. Contrairement à la sous-unité50S qui est essentiellement ronde etrigide, la sous-unité 30S est plate etflexible. Elle est subdivisée en quatredomaines, la tête, le corps, une plate-forme, et un quatrième domaineformé par une longue hélice située àl’interface avec la sous-unité 50S.L’organisation générale de l’ARNr etdes protéines dans la sous-unité 30Sest similaire à celle de la sous-unité50S. La structure tertiaire de l’ARNr16S est stabilisée non seulement pardes interactions ARN-ARN mais éga-lement par des interactions avec lesprotéines. Les protéines sont locali-sées sur la surface exposée au solvantplutôt que dans la région qui formel’interface avec la sous-unité 50S.Comme dans le cas de la sous-unité50S, certaines protéines possèdentdes régions allongées qui s’étendenthors du domaine globulaire et plon-gent dans le cœur ribonucléotidiquede la sous-unité. Ces extensions,dépourvues des motifs habituels derepliement des protéines, interagis-sent avec l’ARNr 16S de façon à lemaintenir dans une conformationqu’une molécule d’ARN pourrait dif-ficilement adopter seule. Les pro-téines ribosomiques semblent doncservir au réglage fin de la structurede l’ARNr, pour lui permettred’accomplir ses fonctions plus rapi-dement et plus efficacement.
Cette vue à l’échelle atomique de lasous-unité 30S permet maintenantd’entrevoir le mécanisme moléculairedu décodage du code génétique. Cedécodage se fait dans le site A parappariement entre l’anticodon del’aminoacyl-ARNt et le codon del’ARNm. On sait que l’appariementdes trois paires de base du codon et del’anticodon n’est pas suffisant pourexpliquer la fidélité de la synthèse pro-téique. Une des fonctions majeures dela sous-unité 30S serait de fournir unenvironnement favorable à un appa-riement correct entre les codons del’ARNm et les anticodons des ARNt.En fait, par des contacts avec le com-plexe ARNt-ARNm, la sous-unité 30Sparticipe activement à la sélection del’aminoacyl-ARNt. Même en l’absencedes ligands ARNt et ARNm, les don-nées cristallographiques montrentque le site P est formé d’éléments del’ARNr 16S appartenant à plusieursdomaines structuraux et que la sur-face de contact entre le site P etl’ARNt est importante, témoignant dela nécessité d’une forte affinité pourmaintenir l’ARNt dans le site P pen-dant que s’opère, au site A, la sélec-tion du nouvel aminoacyl-ARNt. Dansla structure cristalline, la surface dusite A est plus réduite que celle du siteP, en accord avec une affinité plusfaible du ribosome pour l’aminoacyl-ARNt. Comme pour le site P, le site Aest essentiellement formé par l’ARNr16S ce qui souligne à nouveau le rôleessentiel de l’ARNr dans la synthèsedes protéines. Seule la protéine S12,dont des données biochimiques etgénétiques suggéraient un rôle dans lafidélité de la lecture du code géné-tique, serait en mesure d’interagiravec le complexe ARNt-ARNm.Le ribosome est la cible de nombreuxantibiotiques utilisés en médecine[14]. Certains de ces antibiotiques sefixent au ribosome et interfèrent avecses fonctions vitales. Ils constituentdonc une voie importante pour éclai-rer les bases moléculaires de la syn-thèse des protéines. Les aminoglyco-sides ont pour cible la sous-unité 30Set perturbent la lecture du codegénétique. Jusqu’à la déterminationde la structure cristalline de la sous-unités 30S complexée à différentsantibiotiques, nous n’avions à notredisposition qu’une seule structure de
complexe entre un fragment del’ARNr formant le site A et un amino-glycoside, déterminée par résonancemagnétique nucléaire (RMN) [15].Grâce à ces données et à cellesd’études biochimiques, Yoshizawa etal. [16] ont proposé un modèle molé-culaire de lecture du code génétiquepar le ribosome. Le groupe de Rama-krishnan propose un modèle simi-laire sur la base de la structure cristal-line de la sous-unité 30S complexéeau même antibiotique [17]. Deuxrésidus conservés dans toutes lesséquences d’ARNr 16S connues à cejour seraient capables de détecter laprésence de l’ARNt correct lié au siteA et assureraient ainsi, en partie dumoins, la fidélité de la lecture ducode génétique. Cependant, pourl’instant, les structures cristallogra-phiques n’ont pas révélé le méca-nisme de sélection de l’ARNt.Les sites actifs de fixation de l’ARNtet de l’ARNm sont localisés dans unecrevasse à l’intersection de plusieursdomaines structuraux de la sous-unité 30S. Comme nous allons levoir, les mouvements relatifs de cesdomaines sont supposés jouer unrôle important pendant la lecture ducode génétique et lors de la translo-cation du site A vers le site P du pep-tidyl-ARNt auquel vient d’être atta-ché un nouvel acide aminé. Cesmouvements du ribosome seraientassistés par les facteurs d’élongationet par l’hydrolyse du GTP.
La structure du ribosome 70Set le mouvement des ARNt
La structure du ribosome entier(70S) associé aux ARNt a été déter-minée avec une résolution dequelques angströms seulement grâceà la cristallographie (7,8 Å d’abordpuis très récemment 5,5 Å) et à lacryo-microscopie électronique à troisdimensions (11,5 Å) [18-20]. Si l’ana-lyse cristallographique donne uneplus grande précision, la cryo-micro-scopie électronique qui consiste en lacongélation des complexes en moinsd’une seconde, apporte un aspectplus dynamique en fixant les struc-tures dans des conditions physiolo-giques. De plus, elle ne fait pas inter-venir d’atomes lourds qui pourraientmodifier certains détails structuraux.
774 m/s n° 6-7, vol. 17, juin-juillet 2001

Comparées à la taille du ribosome,les régions de contact entre les sous-unités ribosomales sont relativementrestreintes – la résolution à 5,5 Å dela structure du ribosome permet derecenser une douzaine de ponts seu-lement. Les contacts entre ARNr 16Set 23S s’établissent à proximité dessites de liaison des ARNt, alors queles contacts de l’ARNr avec les pro-téines sont périphériques, à distancedes sites fonctionnels. Un des élé-ments qui lie les deux sous-unités estune longue hélice de l’ARNr 16S quis’étend sur toute la longueur del’interface. Cette hélice entre encontact avec des protéines de la sous-unité 50S et avec l’ARNr 23S. Desmodifications dans l’appariement decertains nucléotides de l’ARN 16Sont été observées lorsque la sous-unité 30S passe de l’état où ellecontient l’aminoacyl-ARNt au site A àcelui où elle ne le contient pas [21].Ces modifications entraînent deschangements de conformation quiaffectent la topographie de la sous-unité 30S, celle de l’espace entre lessous-unités, et le réseau complexed’interactions le long de l’hélice del’ARN 16S située à l’interface entreles sous-unités. Ces changements deconformation permettraient le dépla-cement de l’ARNm. Par ailleurs, des expériences dereconstitution de ribosomes et demutations dirigées ont montré que laprotéine L2, qui interagit en deuxsites distincts de l’ARNr 16S, est nonseulement indispensable à l’interac-tion entre les deux sous-unités maisintervient aussi dans l’activité pepti-dyl transférase [22]. La protéine L2jouerait le rôle de relais entre larégion d’interaction des ARNt sur lasous-unité 30S et le centre peptidyltransférase sur la sous-unité 50S. Les trois ARNt qui occupent respecti-vement les sites A, P et E (site de sortiede l’ARNt), chevauchent les deuxsous-unités du ribosome (figure 4). Ilsse situent dans un espace créé entreles sous-unités, espace à travers lequelse glisse également l’ARNm. Les extré-mités acceptrices des ARNt position-nés sur les sites A et P de la sous-unité50S sont proches du tunnel de sortiedu polypeptide. Sur la sous-unité 30S,au site P, six doigts agrippent ferme-ment la boucle anticodon de l’ARNt et
l’ARNm. Ces contacts empêchent ledépart du peptidyl-ARNt et stabilisentl’appariement codon-anticodon, main-tenant ainsi la phase de lecture quandle site A est vacant. Au site A, enrevanche, la boucle anticodon del’aminoacyl-ARNt est beaucoup plusexposée, ce qui lui donne une flexibi-lité suffisante pour orienter l’extré-mité acceptrice de l’ARNt vers lecentre peptidyl transférase, après ledépart du facteur d’élongation EF-Tu(figure 4). Quant au site E, les contactsles plus importants se font avec l’extré-mité CCA libre de l’ARNt désacylé.
La translocation des ARNt du site Avers le site P et du site P vers le site Eimplique un mouvement de l’inter-face entre les sous-unités. La résolu-tion à 5,5 Å de la structure du ribo-some met en évidence de façon nettela proximité entre les sites de fixationdes ARNt et les régions de contactentre les sous-unités [19]. Ellemontre même, pour la première fois,que des contacts s’établissent entreles ARNt et les ponts qui lient lessous-unités. La flexibilité de ces pontssemble donc être la clé du mouve-ment des ARNt.
775m/s n° 6-7, vol. 17, juin-juillet 2001
Figure 4. Les centres fonctionnels du ribosome. (En haut) Représentationschématique du ribosome 70S montrant le canal où passent les ARNt etl’ARNm et le tunnel de sortie du polypeptide. Au-dessous, la séparation desdeux sous-unités montre la position de l’ARNm et des trois ARNt fixés auxsites A, P et E, au niveau du site de décodage sur la sous-unité 30S (àgauche) et au niveau du centre peptidyl transférase de la sous-unité 50S (àdroite) (adapté de Liljas A., Science 1999 ; 285 : 2077-8).

Tout ce système de relais permettraitde lier les événements qui se dérou-lent au site de décodage de la sous-unité 30S à ceux qui ont lieu sur lasous-unité 50S. Ainsi passerait, dansun sens, le signal d’un appariementcorrect entre codon et anticodon et,dans l’autre, celui de la formation dela liaison peptidique.
Conclusions
Que peut-on espérer dans le futur ?Avoir une meilleure vision du pas-sage de l’ARNm à travers le ribo-some, comprendre comment inter-viennent les facteurs de traductionou encore obtenir la structure ato-mique du ribosome eucaryote. Dès àprésent, la connaissance de la struc-ture à l’échelle atomique du ribo-some ouvre tout un monde de nou-velles possibilités non seulementpour la conception rationnelled’antibiotiques dirigés contre le ribo-some bactérien mais aussi pour cellede drogues agissant sur la synthèseprotéique. Il a été montré qu’en pro-voquant des erreurs de décodage, lesaminoglycosides permettent le pas-sage de codons stop, corrigeant lorsde la traduction l’effet de mutationsnon sens responsables de certainesmaladies héréditaires telles que lamaladie de Duchenne [23].Les travaux que nous venons dedécrire ne constituent donc pas unpoint final, mais au contraire le débutd’une nouvelle ère dans l’explorationdu ribosome. Sur le plan fondamen-tal, ils apportent enfin la réponse àune question qui taraudait les spécia-listes depuis des décennies : quel com-posant, ARN ou protéine, catalyse laformation de la liaison peptidique ? Acôté de l’argument évolutionniste, onpeut trouver des raisons biochimiqueconvaincantes à l’utilisation d’ARNcatalytiques pour la synthèse des pro-téines. Les substrats des ribosomessont en partie des ARN, or, via la for-mation de paires de bases ou d’empi-lements de triplets de bases, l’ARN estparticulièrement adapté à la recon-naissance spécifique d’autres ARN.De plus, l’ARN accomplit efficace-ment les changements de conforma-tion à très grande échelle, qui sontrequis pour les mouvements del’énorme structure du ribosome. On
doit se souvenir que le ribosome estune machine, dont la dynamique n’apas encore été capturée par lesétudes cristallographiques. Il nousfaut passer à présent des instantanésau film entier de la synthèse pro-téique. Film à voir au ralenti, car nel’oublions pas, chaque seconde, leribosome bactérien est capable de lire17 codons et donc de synthétiser17 liaisons peptidiques ! ■
RÉFÉRENCES1. Tissières A. Ribosome research : historicalbackground. In : Nomura M, Tissières A,Lengyel P eds, Ribosomes. Cold Spring Har-bor Monograph Series, Cold Spring Har-bor, NY 1974.
2. Palade GE. A small particulate compo-nent of the cytoplasm. J Biophys Biochem Cytol1955 ; 1 : 59.
3. Littlefield JW, Keller EB, Gros J, Zamec-nick PC. Studies on cytoplasmic ribonucleo-proteins particles from the liver of the rat. JBiol Chem 1955 ; 217 : 111.
4. Ban N, Nissen P, Hansen J, Moore PB,Steitz TA. The complete atomic structure ofthe large ribosomal subunit at 2.4 Å resolu-tion. Science 2000 ; 289 : 905-20.
5. Noller HF, Kop J, Wheaton V, et al.Secondary structure model for 23S riboso-mal RNA. Nucleic Acids Res 1981 ; 9 : 6167-89.
6. Nissen P, Hansen J, Ban N, Moore PB,Steitz TA. The structural basis of ribosomeactivity in peptide bond synthesis. Science2000 ; 289 : 920-30.
7. Noller HF, Hoffarth V, Zimniak L. Unu-sual resistance of peptidyl transferase toprotein extraction procedures. Science 1992 ;256 : 1416-9.
8. Piccirilli JA, McConnell TS, Zaug AJ, Nol-ler HF, Cech TR. Aminoacyl esterase activityof the Tetrahymena ribozyme. Science 1992 ;256 : 1420-4.
9. Samaha RR, Green R, Noller HF. A basepair between tRNA and 23S rRNA in thepeptidyl transferase centre of the ribosome.Nature 1995 ; 377 : 309-14.
10. Wimberly BT, Brodersen DE, ClemonsWM, et al. Structure of the 30S ribosomalsubunit. Nature 2000 ; 407 : 327-39.
11. Brodersen DE, Clemons WM, Carter AP,Morgan-Warren RJ, Wimberly BT, Ramakri-shnan V. The structural basis for the actionof the antibiotics tetracycline, pactamycin,and hygromycin B on the 30S ribosomalsubunit. Cell 2000 ; 103 : 1143-54.
12. Carter AP, Clemons WM, Brodersen DE,et al. Crystal structure of an initiation factorbound to the 30S ribosomal subunit. Science2001 ; 291 : 498-501.
13. Schluenzen F, Tocilj A, Zarivach R, et al.Structure of functionally activated smallribosomal subunit at 3.3 Å resolution. Cell2000 ; 102 : 615-23.
14. Gale EF, Cundliffe E, Reynolds PE, Rich-mond MH, Waring MJ. The molecular basis ofantibiotic action. John Wiley & Sons, London1981.
15. Fourmy D, Recht M, Blanchard S,Puglisi J. Structure of the A site of E. coli 16SrRNA complexed with an aminoglycosideantibiotic. Science 1996 ; 274 : 1367-71.
16.Yoshizawa S, Fourmy D, Puglisi JD. Reco-gnition of the codon-anticodon helix byribosomal RNA. Science 1999 ; 285 : 1722-5.
17. Carter AP, Clemons WM, Brodersen DE,Morgan-Warren RJ, Wimberly BT, Ramakri-shnan V. Functional insights from the struc-ture of the 30S ribosomal subunit and itsinteractions with antibiotics. Nature 2000 ;407 : 340-8.
18. Cate JHD, Yusupov MM, Yusupova GZ,Earnest TN, Noller HF. X-ray crystal struc-tures of 70S ribosome functional com-plexes. Science 1999 ; 285 : 2095-104.
19. Yusupov MM, Yusupova GZ, Baucom A,et al. Crystal structure of the ribosome at 5.5Å resolution. Science 2001 ; 292 : 883-96.
20. Gabashvili IS, Agrawal RK, Spahn CMT,et al. Solution structure of the E. coli 70Sribosome at 11.5 Å resolution. Cell 2000 ;100 : 537-49.
21. Gabashvili IS, Agrawal RK, Grassucci RA,Squires CL, Dahlberg AE, Frank J. Majorrearrangements in the 70S ribosomal 3Dstructure caused by a conformational switchin the 16S ribosomal RNA. EMBO J 1999 ;18 : 6501-7.
22. Diedrich G, Spahn CM, Stelzl U, et al.Ribosomal protein L2 is involved in theassociation of the ribosomal subunits, tRNAbinding to A and P sites and peptidyl trans-fer. EMBO J 2000 ; 19 : 5241-50.
23. Howard MT, Shirts BH, Petros LM, Flani-gan KM, Gesteland RF, Atkins JF. Sequencespecificity of aminoglycoside-induced stopcodon readthrough : potential implicationsfor treatment of Duchenne muscular dystro-phy. Ann Neurol 2000 ; 48 : 164-9.
776 m/s n° 6-7, vol. 17, juin-juillet 2001
Olivier Jean-Jean
Unité de biochimie cellulaire, Cnrs FRE2219, Université Paris-6, 9, quai Saint-Bernard, 75005 Paris, France.
Dominique Fourmy
Laboratoire de RMN à Haut-Champ,ICSN Cnrs, 1, avenue de la Terrasse,91190 Gif-sur-Yvette, France.
Anne-Lise Haenni
Institut Jacques-Monod, 2, place Jussieu,75005 Paris, France.
TIRÉS À PARTO. Jean-Jean.