le-coeur
23
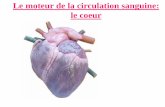
G - LE CŒUR 1 GENERALITES Le cœur résulte embryologiquement du repli du tube cardiaque primitif dans la cavité péricardique. Les deux cavités principales initiales, une oreillette et un ventricule, se subdivisent par la suite chacune en deux par la formation des septums inter-auriculaire et inter-ventriculaire. Le coeur pèse 250 à 350 g chez l'adulte. Sa contraction périodique automatique permet la circulation du sang grâce à la présence d'un ensemble de valvules unidirectionnelles. Il est fait d'un muscle strié particulier, le myocarde. Le sarcoplasme de ces cellules myocardique est riche en mitochondries et le réseau capillaire myocardique est dense, témoignant d'un métabolisme aérobie important. La nappe musculaire et les valvules délimitent quatre cavités disposées en série avec les lits vasculaires pulmonaire et périphérique (Fig 4 et 5).
description
Cours cardiologie
Transcript of le-coeur

G - LE CŒUR 1 GENERALITES Le cœur résulte embryologiquement du repli du tube cardiaque primitif dans la cavité péricardique. Les deux cavités principales initiales, une oreillette et un ventricule, se subdivisent par la suite chacune en deux par la formation des septums inter-auriculaire et inter-ventriculaire. Le coeur pèse 250 à 350 g chez l'adulte. Sa contraction périodique automatique permet la circulation du sang grâce à la présence d'un ensemble de valvules unidirectionnelles. Il est fait d'un muscle strié particulier, le myocarde. Le sarcoplasme de ces cellules myocardique est riche en mitochondries et le réseau capillaire myocardique est dense, témoignant d'un métabolisme aérobie important. La nappe musculaire et les valvules délimitent quatre cavités disposées en série avec les lits vasculaires pulmonaire et périphérique (Fig 4 et 5).

Les cavités et les valves sont tapissées par un endothélium, l’endocarde qui continue celui qui recouvre l’intérieur des vaisseaux sanguins. La surface externe du coeur est recouverte par une membrane à deux feuillets ; l’un, l’épicarde, colle au muscle cardiaque; l’autre, le péricarde, remontant à la racine des vaisseaux de la base, est séparé du premier par une couche de liquide d’environ 10 à 15 ml. Ces feuillets péricardiques facilitent les mouvements de la masse cardiaque au cours de son cycle. L’oreillette et le ventricule droits constituent le « cœur droit », qui reçoit, par les veines caves inférieure et supérieure et le sinus veineux coronaire, le sang veineux déssaturé en provenance des tissus pour l’envoyer, par l’artère pulmonaire, dans les capillaires pulmonaires. A ce niveau s’effectue pour le contact avec le gaz alvéolaire. L’oreillette et le ventricule gauches constituent le « coeur gauche » qui recueille le sang oxygéné sortant des poumons par les veines pulmonaires pour l’envoyer à la périphérie par l’aorte et ses divisions. La paroi ventriculaire gauche est, comme le septum inter-ventriculaire, nettement plus épaisse que la paroi ventriculaire droite. La vascularisation du myocarde est assurée par les deux premières branches collatérales de l’aorte, les artères coronaires droite et gauche qui naissent de la toute première partie de la racine de l’aorte. Leurs ramifications sont intra-myocardiques. Le sang veineux est drainé par un système de vaisseaux dont le collecteur principal est le sinus veineux coronaire.

Les orifices de communication entre oreillettes et ventricules et entre ventricules et vaisseaux de sortie sont munis de valvules qui s'ouvrent toutes dans le même sens dit sens « antérograde » ; elles empêchent, quand elles sont fermées, tout reflux. Si la pression en aval d'une valvule dépasse la pression d'amont, celle-ci se ferme hermétiquement, de purement passive. A l’inverse, si la pression est plus élevée en amont, la valvule s'ouvre, également passivement, permettant la progression de la colonne sanguine qui chemine des hautes vers les basses pressions (Fig 6).
Ces valves sont des structures fibreuses très souples, pratiquement avasculaires, implantées sur des anneaux fibreux qui constituent les orifices qu'elles viennent obstruer. Les valvules auriculoventriculaires, à trois valves ou tricuspide à droite, à deux valves ou mitrale à gauche, sont amarrées aux parois ventriculaires par des cordages qui partent de puissantes excroissances pariétales, les piliers du coeur. L'appareil sigmoïdien sépare chaque ventricule du vaisseau artériel qui lui fait suite. Il est formé de trois valves en forme de cupules dont la convexité est tournée vers le ventricule. Les valves sont insérées par un bord sur l'anneau fibreux, alors que le bord libre, mobile, s'efface lors de l'éjection. A gauche, le bord libre reste à distance de la paroi aortique lors de l'éjection: en effet, les premiers centimètres de l'aorte ascendante sont dilatés en « ampoule », formant les sinus de Valsalva. Cette disposition anatomique évite, au cours de la systole, l'occlusion des orifices des artères coronaires.

2 MECANIQUE CARDIAQUE 2-a Cycle cardiaque L'activité cardiaque est cyclique: la séquence de tous les événements qui se succèdent pour constituer une révolution ou cycle cardiaque ne dure environ qu'une seconde. Elle se répètera durant toute la vie, assurant la progression incessante de la colonne sanguine et le maintien de la pression intravasculaire. Au cours d'un cycle donné, chaque partie du myocarde se trouve successivement en activité, puis au repos (Fig 10).
Le déroulement d'une contraction au niveau d'un fragment isolé du myocarde est plus simple à étudier que celui de la systole enregistrée au niveau du coeur entier car on s'affranchit du phénomène de désynchronisation physiologique de la contraction du coeur qui dépend du temps nécessaire à la conduction de l'excitation de l'étage auriculaire aux ventricules. Si on stimule électriquement un fragment du tissu myocardique, l'enregistrement simultané de l'activité mécanique et de l'activité électrique cellulaire montre que la durée du potentiel d'action est en général au moins égale à celle de la phase ascendante du mécanogramme (Fig 9).

Ceci est en étroite relation avec l'impossibilité de réaliser la fusion des contractions cardiaques successives, autrement dit l'impossibilité de tétaniser le coeur . La phase du cycle pendant laquelle le myocarde se contracte est appelée systole, celle pendant laquelle il se relâche, diastole. Il existe ainsi des systoles (suivies de diastoles) auriculaires droite et gauche, pratiquement simultanées, et des systoles (suivies de diastoles) ventriculaires droite et gauche, également quasi simultanées (fig 10).

En revanche, les activités auriculaires et ventriculaires ne sont pas simultanées : les activités auriculaires précèdent de quelques fractions de seconde les activités électriques ventriculaires. En pratique courante, on parle de « systole » ou de «diastole » ou, parfois, de systole ou diastole « cardiaques » pour désigner la systole ou la diastole ventriculaire, en englobant la contraction ou le relâchement des deux ventricules). 2-b Tissu nodal et automatisme cardiaque La succession des événements et le synchronisme cœur droit/coeur gauche sont assurés par la distribution anatomique de cellules myocardiques particulières, dénuées d'activité mécanique mais produisent spontanément à intervalles réguliers un signal électrique qui stimule les cellules voisines (Fig 11).

L'ensemble de ces cellules constitue le tissu nodal qui comprend deux amas macroscopiquement séparés: le noeud sinusal (ou de Keith et Flack) situé dans la paroi postérieure de l'oreillette droite, contre le septum inter-auriculaire, et le noeud atrio-ventriculaire, dans la partie supérieure de la cloison inter-ventriculaire. Il et qui se prolonge par le faisceau de His qui se divise en deux branches, droite et gauche pour le ventricule correspondant, dans lequel elles se ramifient en un réseau de Purkinje sous-endocardique. Le potentiel de repos de ces différentes cellules n'est pas constant: il augmente progressivement et, lorsqu'il atteint une valeur critique (seuil), la membrane se dépolarise. il se produit alors (loi du tout ou rien) un potentiel d'action qui se propage de proche en proche et déclenche l’activité électrique et mécanique des cellules musculaires; il stimule également, quand il les atteint, les cellules du tissu nodal qui se trouveraient au repos. La vitesse avec laquelle le potentiel de repos de ces cellules évolue vers le potentiel seuil est modifiée par l'action de l’acétylcholine qui l’abaisse, réduisant la fréquence des potentiels d'action, et des catécholamines qui l'augmentent, accroissant la fréquence de ces derniers. Le processus normal de stimulation du coeur naît dans le noeud sinusal, parce que sa fréquence de décharge spontanée est plus élevée 70 à 80.min-1 que celle du noeud auriculo-ventriculaire (40 à 60.min-1) et du tissu nodal sous- jacent Ce noeud sinusal ( « pacemaker », pour les Anglo-Saxons) atteint plus rapidement son potentiel critique et impose, de ce fait, son rythme à tout le coeur : c'est le rythme sinusal, rythme normal du coeur. La stimulation se propage d'emblée à

toute la masse des oreillettes qui se contractent en bloc, puis bute sur l'anneau auriculo- ventriculaire, non conducteur (Fig 12).
Elle est relayée par le nœud auriculo-ventriculaire qui transmet, avec un court temps de latence, une onde de dépolarisation qui atteint, par le faisceau de His, ses branches et le réseau de Purkinje, l'ensemble des deux ventricules. La contraction des ventricules se produit quelques fractions de seconde après celle des oreillettes, compte tenu du temps de propagation de l’onde de dépolarisation. Après chaque activation, les cellules cardiaques demandent un certain temps avant de pouvoir être à nouveau stimulées; c'est la période réfractaire. Celle-ci est d'abord absolue, aucun stimulus, quelle que soit son intensité, ne pouvant alors entraîner de nouvelle dépolarisation. Elle devient ensuite relative, la dépolarisation pouvant alors être déclenchée par un stimulus de plus en plus faible au fur et à mesure que le temps s'écoule. Celle-ci est liée à une élévation du seuil de dépolarisation rapide.

2-c Couplage excitation-contraction : rôle du calcium * Muscle cardiaque Les myocytes cardiaques sont disposés en couches étroitement connectées dans le myocarde. Quand les parois se contractent, comme un poing qui se serre, et compriment le sang qu’elles contiennent. Le muscle cardiaque combine des propriétés du muscle squelettique et du muscle lisse (fig 13).
Ces cellules sont en raison d’une disposition des filaments épais de myosine et des filaments fins d’actine similaire à celle du muscle squelettique. Mais les fibres musculaires cardiaques sont beaucoup plus courtes que les fibres du muscle squelettique et elles présentent de nombreuses ramifications ; les cellules adjacentes sont unies à leurs extrémités par des structures appelées disques intercalaires. Dans ces structures, on trouve des desmosomes qui maintiennent les cellules les unes contre les autres et sur lesquels viennent s’ancrer les myofibrilles. A côté des disques intercalaires, on trouve des jonctions communicantes similaires à celles rencontrées dans de nombreux muscles lisses. Environ 1% des cellules cardiaques n’interviennent pas dans la contraction, mais portent des structures spécialisées qui sont indispensables à l’excitation cardiaque normale.

Ces cellules constituent un réseau appelé système de conduction cardiaque et elles entrent au contact des cellules musculaires cardiaques par l’intermédiaire des jonctions communicantes. Le système de conduction initie le battement cardiaque et assure la propagation rapide de l’influx dans tout le cœur. *Innervation Le cœur reçoit une riche innervation sympathique et parasympathique. Cette dernière est véhiculée par les nerfs vagues. Les fibres post ganglionnaires sympathiques libèrent essentiellement de la noradrénaline et les parasympathiques surtout de l’acétylcholine. Les récepteurs du muscle cardiaque à l’adrénaline sont essentiellement béta-adrénergiques. L’adrénaline, sécrétée par les médullosurrénales, se fixe sur les mêmes récepteurs que ceux de la noradrénaline et a des effets cardiaques identiques. Les récepteurs à l’acétylcholine sont de type muscariniques. *Propagation du PA : Le potentiel d’action qui naît dans le nœud sinusal se propage ensuite à tout le myocarde en passant d’une cellule à une autre par des jonctions communicantes. La propagation dans l’OD puis dans l’OG n’utilise pas de fibre de système de conduction. La conduction au niveau des oreillettes est suffisamment rapide pour que les deux oreillettes se contractent pratiquement en même temps. La propagation de l’influx à travers les ventricules est plus complexe, faisant intervenir le reste du système de conduction : La liaison entre dépolarisation des oreillettes et des ventricules est assuré par le nœud atrio-ventriculaire, localisé à la base de l’OD. Les potentiels d’action qui se propage au sein de l’OD induisent une dépolarisation du nœud atrio-ventriculaire. Par contre, les potentiels d’action traversent relativement lentement ce nœud (0,1 s environ). Ce délai permet à la contraction auriculaire de s’achever avant que ne débute la contraction ventriculaire. Après avoir passé le nœud atrio-ventriculaire, l’influx pénètre dans la paroi qui sépare les deux ventricules (le septum interventriculaire) en empruntant un système de fibres de conduction appelé faisceau de His. Le nœud atrio-ventriculaire et le faisceau de His constituent le seul lien électrique entre oreillettes et ventricules. En dehors de ces voies, les oreillettes sont complètement isolées des ventricules par une couche de tissu conjonctif non conducteur. Dans le septum interventriculaire, le faisceau de His se divise en branches droite et gauche qui pénètrent la paroi de chacun des deux ventricules. A leur tour, ces fibres entrent en contact avec les fibres de Purkinje, qui sont des cellules conductrices volumineuses au travers desquelles l’influx se propage à tous les ventricules.

La conduction rapide de ces fibres et leur distribution diffuses expliquent que la dépolarisation ventriculaire droite et gauche soit quasi simultanée permettant une contraction unique coordonnée. En fait, la dépolarisation et la contraction sont légèrement plus précoce à l’apex des ventricules et se propage vers le haut. *Potentiels d’action cardiaques et excitation du nœud sinusal Le mécanisme de propagation de l’influx le long des membranes des cellules myocardiques ressemble à ce qui se passe dans d’autres cellules de tissus excitables comme les neurones ou les muscles striés squelettiques. Toutefois, des types différents de cellules cardiaques expriment des combinaisons particulières de canaux ioniques, les potentiels d’action ayant alors des formes différentes et des fonctions particulière dans la propagation de l’excitation dans tout le cœur. Un potentiel d’action caractéristique d’une cellule myocardique a pour origine des modifications de perméabilité membranaire (fig 14).
Comme dans les cellules musculaires squelettiques et les neurones, la membrane au repos est beaucoup plus perméable au K+ qu’au Na+. Ainsi, le potentiel membranaire de repos est-il plus proche du potentiel d’équilibre du K+ (-90mV) que du Na+ (-60 mV). De même, la phase de dépolarisation est due principalement à l’ouverture des canaux sodiques voltages dépendants. L’entrée du Na+ dépolarise la cellule et maintient par un rétrocontrôle positif, l’ouverture d’un plus grand nombre de canaux sodiques. Simultanément, la perméabilité au K+

diminue, par fermeture de certains canaux potassiques, ce qui contribue également à la dépolarisation. Là encore, comme dans les cellules des muscles squelettiques ou les neurones, l’augmentation de la perméabilité au Na+ et très transitoire car les canaux sodiques s’inactivent assez rapidement. Mais contrairement aux autres cellules excitables, dans le muscle cardiaque, la baisse de la perméabilité au Na+ ne s’accompagne pas d’une repolarisation membranaire. La membrane reste dépolarisée en plateau, à 0 mV environ. La persistance de cette dépolarisation s’explique par : - une perméabilité au K+ qui reste inférieur à la valeur de repos (c’est-à-dire que ces canaux restent fermés) - une augmentation marquée de la perméabilité au Ca++. Ce dernier mécanisme est le plus important et fonctionne de la de la façon suivante : Dans les cellules myocardiques, la dépolarisation membranaire initiale induit l’ouverture des canaux calciques voltage dépendants, ce qui fait apparaître un flux d’ions Ca++ dans la cellule. Ces canaux s’ouvrent beaucoup lentement que les canaux sodiques et restent plus longtemps ouverts. Ils sont appelés canaux de type « L » (Long lasting, ou, long terme). Le flux de Ca++ entrant dans la cellule équilibre exactement le flux de K+ sortant, ce qui maintient le plateau de dépolarisation membranaire. Enfin, la cellule se repolarise quand les perméabilités au K+ et au Ca++ regagnent leur valeur d’origine. Les potentiels d’action des cellules des oreillettes ont la même forme que ceux ces myocytes ventriculaires mais la durée du plateau est moindre. Il existe par contre de grandes différences entre potentiels d’action des cellules myocardiques et ceux du système de conduction. Le potentiel d’action d’une cellule du nœud sinusal est représenté figure 17.

La cellule du nœud sinusal n’a pas de potentiel de repos stable mais est le siège d’une lente dépolarisation. Cette dépolarisation graduelle est appelée potentiel de pace-maker. Elle amène le potentiel membranaire au seuil de déclenchement du PA. Le potentiel de pace-maker est déclenché par trois mécanismes au niveau des canaux ioniques : 1- diminution progressive de la perméabilité au K+. Les canaux K+ qui se sont ouverts au cours de la dépolarisation du PA précédent se ferment progressivement. 2- Les cellules pace-maker portent une catégorie unique de canaux qui, contrairement aux autres canaux potentiels dépendants, s’ouvrent quand le potentiel membranaire est négatif. Ces canaux entraînement un courant dépolarisant entrant de NA+ qu’il ne faut pas confondre avec les canaux qui engendrent l’ascension du PA dans les cellules neuronales, musculaires squelettiques et les myocytes cardiaques. 3- Le troisième canal pace-maker est un canal calcique qui ne s’ouvre que brièvement (Ca++T) mais contribue au courant calcique entrant dû aux canaux lents (Ca++L). Après un certain délai, l’ouverture des canaux K+ repolarise la membrane. Le retour de potentiels négatifs active de nouveau les mécanismes pace-maker et le cycle se reproduit.

2-d L’électrocradiogramme (Fig 18, 20)

Les myocytes activés se dépolarisent et induisent, de ce fait, une variation du champ électrique intra-thoracique : des électrodes cutanées, reliées à un galvanomètre. détectent une variation de potentiel. L'électrocardiogramme (ECG) est l'enregistrement, en fonction du temps, des fluctuations de potentiel de l'ensemble du coeur produites par sa stimulation. L'asynchronisme de la stimulation des différentes parties (oreillettes puis ventricules), les différences de masse des parois et les directions variables des voies de conduction et de la propagation de l'activation (de l'endocarde vers l'épicarde) se somment pour donner un aspect caractéristique au tracé obtenu. En plaçant les électrodes aux deux poignets (fig 18) ( électrode R pour le droit et L pour le gauche, de l'anglais right et left) et à la cheville gauche (électrode F de foot, pied), on enregistre trois dérivations bipolaires des membres: DI entre R et L, DII entre R et F et DIII entre L et F. Une combinaison précablée de ces dérivations, faite automatiquement au sein de l'appareil d'enregistrement ou électrocardiographe, permet d'obtenir le potentiel local unipolaire amplifié en R, L et F : dérivations unipolaires amplifiées des membres aVR, aVL, aVF. Enfin, le même montage permet d'obtenir les dérivations unipolaires précordiales en déplaçant une électrode réceptrice à la surface du thorax, autour de la projection du cœur . Le tracé de l'électrocardiogramme (Fig 21) comporte une onde P qui traduit l'activation auriculaire puis, après un court instant (0,12 à 0,21s selon la

fréquence cardiaque), un complexe d'ondes dites rapides, étiquetées Q, R et S, témoins de l'activation des ventricules et enfin, une onde lente, l'onde T, produite par la repolarisation de ces derniers et survenant quelques instants après le complexe rapide.
En pratique clinique, il est de règle de disposer les électrodes en des endroits du corps bien déterminés et d'obtenir ainsi, en général, douze dérivations : - trois dérivations bipolaires: DI, DII, DIII. - trois dérivations monopolaires des membres : aVR, aVL, aVF . - six dérivations unipolaires précordiales : VI, V2, V3, V4,V5,V6. L'étude comparative de ces ondes enregistrées dans les différentes dérivations et de leurs rapports chronologiques permet une interprétation clinique de l'électrocardiogramme, en particulier, le calcul de l’axe électrique du cœur qui se fait habituellement sur les dérivations bipolaires d’Einthoven. 2-e Cycle des événements cardiaques Le coeur gauche (oreillette et surtout ventricule) qui dessert la totalité des tissus de l'organisme est la principale source d'énergie mécanique produite par la pompe cardiaque. C’est lui que nous décrirons, étant entendu que les mêmes événements se produisent dans le coeur droit de façon pratiquement synchrone.

Seule diffère la pression intra-ventriculaire qui est cinq fois plus faible à droite qu'à gauche au cours de la systole. *Protodiastole ventriculaire : relâchement et phase de remplissage rapide Juste après la fermeture des valves sigmoïdes, le sang ne peut ni sortir, ni entrer dans le ventricule dans lequel la pression, rapidement décroissante, reste pendant quelques centièmes de seconde trop élevée pour que la valvule mitrale puisse s'ouvrir : c'est la phase de relaxation isovolumétrique, premier temps de la diastole ventriculaire. L'onde T de l'ECG se prolonge sur une partie de cette phase. Quand la pression intra-ventriculaire devient inférieure à la pression auriculaire, la valvule mitrale s'ouvre et le sang, accumulé jusque là dans l'oreillette, passe dans le ventricule dont le volume augmente, rapidement d'abord, puis de plus en plus lentement, jusqu'à la contraction auriculaire. L'ouverture de la mitrale et le mouvement de sang qui s'ensuit se traduisent par une chute de la pression auriculaire qui dessine, de ce fait, une onde en « v » (Fig 24). Celle-ci se trouve située au point de croisement des courbes de pressions ventriculaire et auriculaire.

*télédiastole ventriculaire: phase de remplissage lent Pendant la diastole ventriculaire, oreillette et ventricule gauches sont relâchés et communiquent librement, les valves de la mitrale n'étant pas accolées. En revanche, les sigmoïdes aortiques, fermées du fait de la forte pression régnant dans l'aorte, empêchent tout reflux de sang dans le ventricule gauche. Après une phase de remplissage rapide du ventricule durant la première moitié de la diastole (protodiastole), le sang des veines pulmonaires continue à remplir la cavité auriculo-ventriculaire dont la pression n'augmente alors que de 3 mmHg en moyenne, les parois cardiaques se laissant facilement distendre. Dans les conditions le repos, le ventricule a déjà emmagasiné, pendant la protodiastole, la plus grande quantité du sang qu'il doit recevoir avant de se contracter, le volume ventriculaire n'augmentant que très peu pendant la télédiastole. Tout à la fin de la diastole ventriculaire, le noeud sinusal décharge, déclenchant la dépolarisation du myocarde auriculaire (onde p sur le tracé ECG), et donc sa contraction : c'est la systole auriculaire. Un supplément de sang est alors brusquement déplacé de l'oreillette vers le ventricule, entraînant une élévation transitoire de la pression auriculoventriculaire qui culmine à 6 à 7 mmHg environ (onde a du tracé de pression auriculaire, fig 25).
Cet ultime apport de sang au ventricule est souvent de faible volume. Il peut devenir important en cas de tachycardie (jusqu'à 40 % le l'apport diastolique), en effet, en cas de tachycardie, le raccourcissement de la diastole ne permet

pas un remplissage préalable satisfaisant. Le volume ventriculaire est maximal en fin de diastole (volume télédiastolique), entre 100 et 150 ml, et la pression intra-ventriculaire qui lui correspond (pression télé- diastolique), entre 0 et 7 mmHg, jouent un rôle important en physiologie et en physiopathologie cardiaques. Pendant tout ce temps diastolique, la pression n'a fait que baisser régulièrement dans l'aorte, isolée du ventricule gauche par les sigmoïdes fermées et dont le sang s'écoule en permanence vers la périphérie. Elle atteint sa valeur la plus basse en fin de diastole : c'est la pression artérielle diastolique. *Systole ventriculaire et phase d'éjection Juste après la fin de la dépolarisation auriculaire, l'onde de stimulation myocardique franchit le noeud auriculo-ventriculaire et se propage rapidement dans la masse ventriculaire qu'elle dépolarise (complexe QRS de l'ECG). Celle-ci, de ce fait, se contracte : c'est le début de la systole ventriculaire. Le sang intraventriculaire se trouve alors comprimé, la pression intra-ventriculaire augmente rapidement et, presque immédiatement, dépasse la pression qui règne dans l'oreillette ; elle ferme alors la valvule mitrale, empêchant tout reflux. La fermeture des valvules auriculoventriculaires droite et gauche et les phénomènes mécaniques et hémodynamiques qui l'accompagnent, entraînent des vibrations pariétales et intracardiaques perçues à l'auscultation, sous forme de premier bruit du coeur, B1, bruit sourd, grave qui va se prolonger jusqu'à l'ouverture des valvules sigmoïdes. Il faut un certain temps pour que la pression intraventriculaire gauche atteigne puis dépasse la pression aortique et ouvre les sigmoïdes; ainsi, pendant les premiers centièmes de seconde de la systole ventriculaire, le volume de sang emprisonné dans le ventricule par la fermeture de la mitrale ne peut pas sortir par l'aorte. C'est la phase de contraction isovolumétrique qui précède la phase d'éjection. La variation de pression intraventriculaire gauche (dP/dt), très rapide pendant la phase isovolumétrique (environ 1900 mmHg.s-l), sert parfois d'index de la contraction myocardique. Les piliers du coeur et les cordages empêchent les valves de s'éverser sous la poussée de la pression intraventriculaire ; la valve mitrale bombe cependant à l'intérieur de l'oreillette gauche, y provoquant une petite élévation de pression (onde c). Puis, les sigmoïdes s'ouvrent. Le début de l'éjection fait diminuer le volume ventriculaire. La courbe de volume ventriculaire montre que l'éjection est d'abord rapide; elle ralentit nettement à partir du moment où la pression ventriculaire a atteint sa valeur maximale. L'augmentation de la pression aortique, induite par l'arrivée brutale de l'onde systolique dans l'artère, distend les parois du vaisseau et permet à celui-ci de stocker momentanément une fraction du volume éjecté.

Pendant toute la phase d'éjection, le sang continue d'arriver dans l'oreillette gauche, alors isolée du ventricule par la valvule mitrale fermée ; la pression auriculaire remontera donc progressivement jusqu'à un sommet v. C'est pendant la phase d'éjection lente que débute l'onde T de l'ECG, témoin de la repolarisation myocardique. Le volume de sang éjecté à chaque contraction ventriculaire appelé volume d'éjection ou volume d'éjection systolique (VES), ne constitue qu'une partie (fraction d'éjection) du volume télédiastolique (80 ml, par exemple, sur 120 ou 150 ml, soit entre les deux-tiers et la moitié). Le résidu post-systolique ou volume télésystolique (VTS) ou encore volume résiduel, constitue un volume de réserve utilisable pour augmenter le volume d'éjection suivant. La fraction d'éjection augmente à l'exercice ou en cas de stimulation sympathique importante : elle peut atteindre les trois-quarts du volume télédiastolique. A l'inverse, elle peut s'abaisser à moins de la moitié en cas de défaillance myocardique, c'est-à-dire d'insuffisance cardiaque. La fin de la systole ventriculaire est marquée par la fermeture des valvules sigmoïdes et par la perception acoustique du deuxième bruit du coeur (B2) : ce dernier est contemporain de l'interruption de la colonne sanguine dans l’aorte. En effet, comme la pression intraventriculaire baisse rapidement, le sang aortique tend à refluer et ferme les sigmoïdes. Sur un tracé de pression intra-aortique, l'incisure dicrote, marque la fin de la systole ventriculaire qui se trouve ainsi, pour le clinicien, encadrée par le premier et le second bruit du coeur. *Contraction auriculaire Elle a un triple effet. Survenant juste avant la systole ventriculaire, elle contribue, par l'augmentation de la pression intraventriculaire qu'elle détermine, à fermer la valvule auriculoventriculaire, empêchant un reflux de sang au début de la systole ventriculaire. Elle déplace une dernière quantité de sang qui va se trouver piégée dans le ventricule: elle sert donc ainsi à augmenter d'autant la réserve diastolique. Enfin, elle participe à la détermination du volume télédiastolique ventriculaire, facteur intervenant dans la contraction myocardique. Les variations de pression auriculaire sont transmises de façon rétrograde aux veines de la base du coeur, où elles peuvent être captées. A droite, elles sont parfaitement visibles sur la jugulaire externe et enregistrables à l'aide d'une capsule manométrique : on obtient un sphygmogramme jugulaire sur lequel on repère les ondes a, c et v, témoins de la dynamique auriculoventriculaire droite. A gauche, on peut obtenir un tracé analogue par enregistrement intra-oesophagien des variations de pression à hauteur des veines pulmonaires et de l'oreillette gauche.

3 IRRIGATION DU MYOCARDE PENDANT LE CYCLE CARDIAQUE Les artères coronaires droite et gauche qui naissent à la racine de l'aorte, en regard des valves sigmoïdes, fournissent la totalité du sang qui irrigue le myocarde. La circulation coronaire est influencée par plusieurs types de facteurs mécaniques, nerveux et métaboliques. 3-a Facteurs mécaniques : La pression sanguine dans l'aorte, générée par le coeur lui-même, est un des facteurs déterminants de la perfusion myocardique. Une élévation de la pression artérielle entraîne une augmentation parallèle immédiate du débit coronaire. Cependant, si, expérimentalement, sur cœur isolé, on augmente brutalement la pression artérielle coronaire, l'augmentation du débit ainsi provoquée est brève. On observe dans les 10 à 15 secondes qui suivent la variation induite de débit, un retour progressif de celui-ci vers sa valeur basale. Ce phénomène est un exemple d'autorégulation du débit sanguin (fig 26).
Dans les conditions physiologiques, la pression artérielle est maintenue à des valeurs relativement constantes par les baroréflexes. Ces mécanismes d'autorégulation sont donc d'une importance limitée pour le coeur « in vivo ». Les artères coronaires ont une partie de leur trajet à la surface du coeur et donnent naissance à des artérioles qui pénètrent dans le myocarde perpendiculairement à la surface épicardique. Les artérioles intramyocardiques

sont soumises à des contraintes mécaniques importantes pendant la systole. Dans la paroi du ventricule gauche, ces contraintes sont si élevées que le sang artériel reflue: pendant la systole, le débit artériolaire coronaire s'inverse et le sang circule alors des capillaires vers les artères. Ce phénomène de compression extravasculaire prend toute son importance lorsque la fréquence cardiaque est élevée, c'est-à-dire lorsque la durée de la systole est proportionnellement plus longue dans le cycle cardiaque. Au début de la diastole, lorsque les forces de compression extravasculaires s'annulent, le débit coronaire gauche s'accroît brusquement pour atteindre sa valeur maximale; il décroît ensuite parallèlement à la pression aortique diastolique. Le débit coronaire gauche est typiquement à prédominance diastolique avec même un reflux artériel au tout début de la systole. Le débit coronaire droit a une évolution très proche de celle de la pression aortique avec des valeurs de débit plus importantes pendant la systole. Les deux coronaires, droite et gauche, étant soumises à la même pression artérielle, il est clair que les différences enregistrées entre les courbes de débits sanguin sont dues à des différences entre les forces de compressions extravasculaires dans les parois des ventricules gauche (où elles sont très élevées) et droit (où elles sont minimes). La tachycardie et la bradycardie ont des effets inverses sur le débit coronaire du ventricule gauche. Une modification du cycle cardiaque est essentiellement due à une modification de la durée de la diastole, le temps systolique restant à peu près constant quelle que soit la fréquence cardiaque. Une tachycardie s'accompagne donc d'une diminution du temps diastolique pendant lequel le débit coronaire gauche est élevé. Cela entraîne une réduction du débit coronaire gauche qui ne peut être compensée que par une vasodilatation importante. 3-b Facteurs neuro-humoraux Contrairement à ce que l'on observe dans les artères irriguant le muscle squelettique, il n'y a pas de fibres parasympathiques cholinergiques innervant les vaisseaux coronaires. On a toutefois pu démontrer l'existence de récepteurs a. (constricteurs) et b (dilatateurs) au sein des artères coronaires. Néanmoins, il semble que les résistances coronaires soient contrôlées essentiellement par des mécanismes humoraux. 3-c Facteurs métaboliques Une des caractéristiques de la circulation coronaire est le parallélisme étroit que l'on retrouve entre le niveau du métabolisme du myocarde et le débit coronaire. Cela traduit la capacité du myocarde d'ajuster son débit d'irrigation à son besoin en énergie. De nombreuses substances sont probablement impliquées dans cette régulation: pressions partielles en O2 et CO2, concentration en acide lactique, ions H+, histamine, osmolarité, prostaglandine et adénosine.

L'adénosine est probablement un vasodilatateur métabolique important dans la circulation coronaire. On a démontré qu'une réduction de la pression partielle en O2 du tissu myocardique entraînait une formation d'adénosine par les myocytes. L'adénosine traverserait les membranes cellulaires et, par l'intermédiaire du liquide interstitiel, atteindrait les parois des vaisseaux coronaires. Des récepteurs spécifiques de l'adénosine localisés dans les artérioles coronaires résistives sont activés par l'adénosine du liquide interstitiel et provoquent une vasodilatation coronaire, donc une augmentation du débit d'irrigation myocardique. 3-d Consommation d'oxygène et travail cardiaque Dans les conditions de repos, le myocarde consomme environ 8 à 10 ml d'02.min-1.100g-l de muscle. Cette consommation peut considérablement augmenter (5 à 6 fois) pendant l'exercice musculaire. La pression partielle en oxygène dans le sang veineux coronaire est très faible (environ 5 vol %) et l'extraction de l'oxygène par le myocarde est pratiquement maximale. Une augmentation de l'apport d'oxygène, si elle ne peut se faire par une augmentation de son extraction par les tissus