
La Nutrition Carbonée. -...
44
La Nutrition Carbonée. 1. Introduction La photosynthèse est un mécanisme fondamental qui permet la conversion de l’énergie lumineuse en énergie chimique. Il existe un angle écologique car c’est un processus à l’origine de la fabrication de la biomasse terrestre. On a conversion d’une forme de carbone la plus oxydée en forme plus réduite : CO 2 sucres, protéines, lipides. S’il n’y a pas de photosynthèse, il n’y a pas de biomasse et, la photosynthèse produit de l’O 2 qui représente 20% des composants de l’atmosphère. Les végétaux sont autotrophes. Ils se nourrissent à partir de formes non réduites de carbones, eau, oxygène… Leur capacité hétérotrophique est moins importante, ils utilisent des carbones organiques tel que l’urée mais, pendant la germination, les plantes sont hétérotrophes. Les plantes peuvent assimiler le CO 2 lors de la transformation du pyruvate en oxaloacétate grâce à la pyruvate carboxylase, c’est la β-carboxylation. On a une simple fixation du CO 2 sans lumière.
Transcript of La Nutrition Carbonée. -...

La Nutrition Carbonée.
1. Introduction
La photosynthèse est un mécanisme fondamental qui permet la conversion de l’énergie lumineuse en énergie chimique.
Il existe un angle écologique car c’est un processus à l’origine de la fabrication de la biomasse terrestre. On a conversion d’une forme de carbone la plus oxydée en forme plus réduite : CO2 � sucres, protéines, lipides.
S’il n’y a pas de photosynthèse, il n’y a pas de biomasse et, la photosynthèse produit de l’O2 qui représente 20% des composants de l’atmosphère.
Les végétaux sont autotrophes. Ils se nourrissent à partir de formes non réduites de carbones, eau, oxygène… Leur capacité hétérotrophique est moins importante, ils utilisent des carbones organiques tel que l’urée mais, pendant la germination, les plantes sont hétérotrophes. Les plantes peuvent assimiler le CO2 lors de la transformation du pyruvate en oxaloacétate grâce à la pyruvate carboxylase, c’est la β-carboxylation. On a une simple fixation du CO2 sans lumière.

2. Historique
• Rejet d’oxygène Priestley a montré qu’une plante peut vivre sous une cloche en verre sans renouvellement d’air contrairement à un animal. � Les plantes produisent de l’O2 (1771-1773)
• Lumière indispensable (1779) • CO2 absorbé en même temps que l’émission d’O2 (Senestien 1782-1783)
• La lumière agit comme source d’énergie (Mayer 1845)
• Chloroplastes importants (Sachs 1864)
CO2 + H2O + Energie Lumineuse � Glucides + O2
• Dégagement d’O2 indépendant de la fixation du carbone (Hill 1937)
Réaction de Hill : Chloroplastes isolés + agents oxydants (accepteurs d’électrons) � production d’O2 L’intégrité des chloroplastes n’est pas nécessaire. Seuls les thylakoïdes sont nécessaires.

La réaction globale est une réaction d’oxydo réduction. D’où viennent les électrons ? Les atomes d’oxygènes ? Hill suggère que H2O libère des électrons et de l’ O2 en se dissociant.
• Ruben et Komen (1940) démontrent que l’O2 provient de l’H2O de même que les électrons libérés.
• Van Niel (1941) démontre avec des bactéries photosynthétiques sulfureuses que le
donneur d’électron peut ne pas être de H2O :
CO2 + 2H2S � HCHO + H2O + 2S
On a une différence entre les photosynthèses oxygéniques (production d’O2) et les photosynthèses anoxygéniques (pas de production d’O2). Principe thermodynamique : tout système évolue spontanément d’un état moins probable à un état plus probable. C � CO2 : forme la plus oxydée, la plus stable.
3. Structure et ultrastructure des chloroplastes
Ce sont des organes allongés. Il en existe plusieurs dizaines par cellules chez les végétaux supérieurs. Leur nombre et leur taille varient en fonction de l’âge de la cellule. On les trouve dans les parties aériennes des plantes : tiges et feuilles. Les stomates possèdent des chloroplastes.
Energie Lumineuse
Réaction Lumineuse
(thylakoïdes)
H2O CO2
Fixation Carbone (Stroma)
NADPH ATP
H2O CO2

Les chloroplastes sont issus de la différenciation des proplastes. Ils sont d’origine maternelle (hérédité cytoplasmique). Toute la machinerie de transcription de l’ADN existe chez les plastes qui contiennent de l’amidon, des globules lipidiques. Les chloroplastes sont le lieu de synthèse des acides gras, des caroténoïdes. Ils sont délimités par une enveloppe constituée par une double membrane. Cette enveloppe limite l’espace intérieur, le stroma où se trouvent les enzymes nécessaires aux diverses réactions. Dans le stroma on trouve les thylakoïdes qui sont des empilements de saccules aplaties appelées grana où on trouve les réactions de Hill. La lumière du thylakoïdes est appelée espace inter membranaire.

Il existe des thylakoïdes courts qui forment les grana et des thylakoïdes longs qui relient les grana entre eux. Ils sont parallèles à l’axe du chloroplaste. Ces thylakoïdes contiennent la chaîne de transport d’électrons, les pigments chlorophylliens et le complexe ATP synthétase.
Plus il y a d’empilements et mieux la récupération d’énergie lumineuse se fait. Une plante à l’ombre à plus de grana qu’une plante en lumière forte.
4. Molécules impliquées dans la photosynthèse
a. Pigments On distingue deux pigments :
- Les caroténoïdes
- Les chlorophylliens associés à des protéines. On parle de chloroprotéines. -
La chlorophylle est constituée d’un noyau porphyrine : 4 pyroles en cercle et un atome de magnésium chélaté au centre. On a associé à ce noyau un alcool à large chaîne appelé phytol.
Chlorophylle a : pigment bleu vert présent chez tous les végétaux Chlorophylle b : pigment vert jaune chez les plantes et les algues vertes Chlorophylle c/d : chez les algues brunes et rouges.
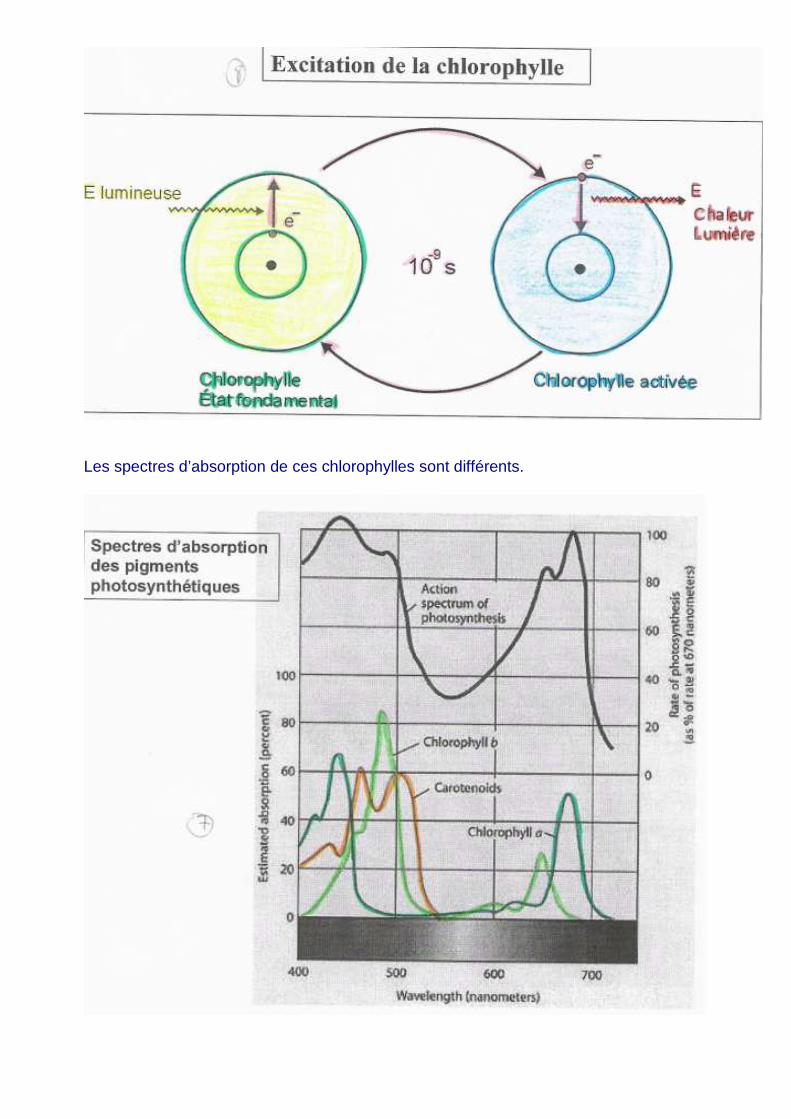
Les spectres d’absorption de ces chlorophylles sont différents.
Les propriétés d’absorption de la lumière varient en fonction des doubles liaisons. Les queues phytols sont hydrophobes, les têtes porphyrines sont hydrophiles. Les complexes chlorophylliens forment des photorécepteurs P680/P700
Les caroténoïdes sont exclusivement lipophiles et appartiennent aux isoprénoïdes (leur synthèse démarre avec le précurseur isoprène). Il existe plusieurs catégories de caroténoïdes tels que les xanthophylles et les carotènes.
Les caroténoïdes apparaissent en orange.
On différencie
- les pigments primaires (chlorophylle a) qui participent aux processus photochimiques
- les pigments accessoires (chlorophylle b et caroténoïdes) qui ne sont pas des composants de la chaîne de réactions mais ils sont organisés dans des antennes collectrices. Ils transfèrent de l’énergie lumineuse vers les pigments primaires.

b. Protéines • Protéines impliquées dans la réception de l’énergie lumineuse Au niveau des photosystèmes :
- LHC-II � Light Harvesting Complex • Codé par les gènes CAB • Représente un tiers des protéines de la membrane thylacoïdale • Stabilise les pigments chlorophylle a et b et les caroténoïdes. • Stabilisation de la moitié de la chlorophylle totale.
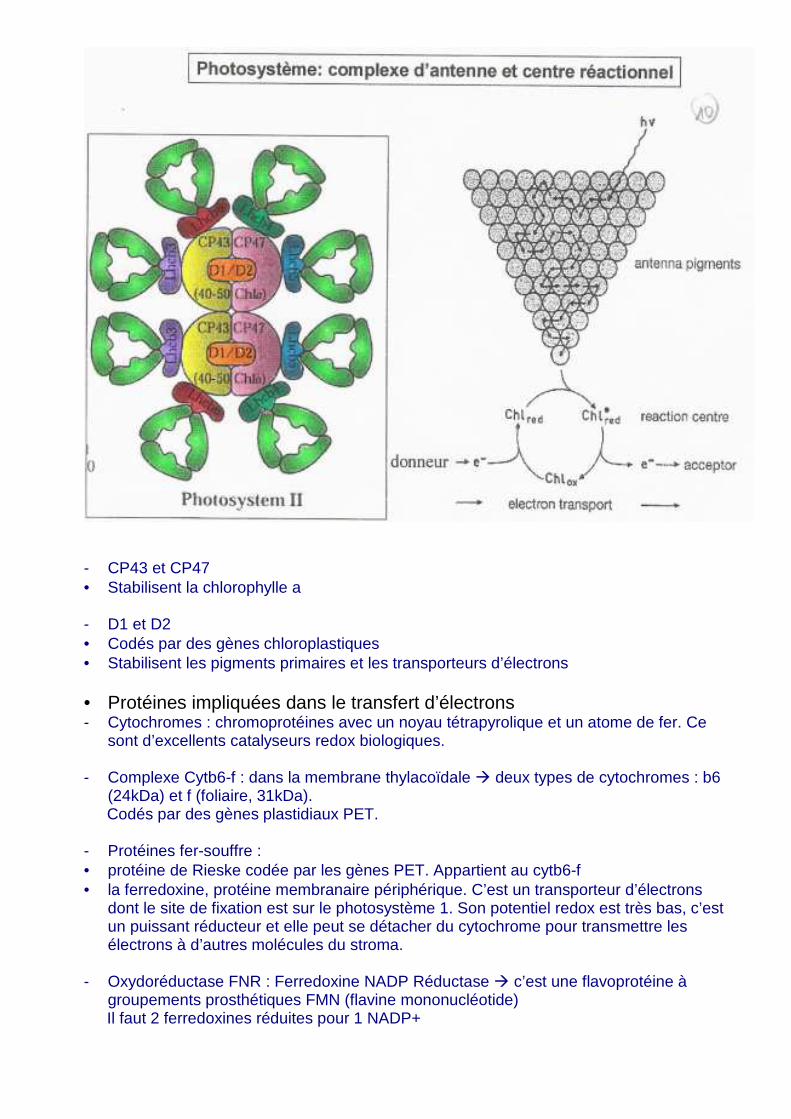
- CP43 et CP47 • Stabilisent la chlorophylle a - D1 et D2 • Codés par des gènes chloroplastiques • Stabilisent les pigments primaires et les transporteurs d’électrons • Protéines impliquées dans le transfert d’électrons - Cytochromes : chromoprotéines avec un noyau tétrapyrolique et un atome de fer. Ce
sont d’excellents catalyseurs redox biologiques. - Complexe Cytb6-f : dans la membrane thylacoïdale � deux types de cytochromes : b6
(24kDa) et f (foliaire, 31kDa). Codés par des gènes plastidiaux PET.
- Protéines fer-souffre : • protéine de Rieske codée par les gènes PET. Appartient au cytb6-f • la ferredoxine, protéine membranaire périphérique. C’est un transporteur d’électrons
dont le site de fixation est sur le photosystème 1. Son potentiel redox est très bas, c’est un puissant réducteur et elle peut se détacher du cytochrome pour transmettre les électrons à d’autres molécules du stroma.
- Oxydoréductase FNR : Ferredoxine NADP Réductase � c’est une flavoprotéine à
groupements prosthétiques FMN (flavine mononucléotide) Il faut 2 ferredoxines réduites pour 1 NADP+

- Plastocyanine : petite protéine renfermant du cuivre. Contient un site de fixation sur Cytb6-f et le PS1. C’est une protéine mobile périphérique située sur la face interne du thylakoïde.
• Protéines impliquées dans la photodissociation de l’eau - Complexe producteur de dioxygène MSP : le centre réactionnel est situé au niveau du
lumen. On a 4 atomes de manganèse liés par des atomes d’oxygène. - Couplage spatial et fonctionnel entre PSII et MSP : le CP47 pourrait stabiliser le
complexe. D1 serait lié au centre réactionnel Mn (histidine) et la chaîne Z (tyrosine de D1) accepte les électrons venant de l’eau.
• Protéines impliquées dans la synthèse d’ATP - Complexe ATP synthase : • CF : canal proton inséré dans la membrane thylacoïdale (imperméable aux protons) • CFI : ensemble catalytique facteur couplant la synthèse d’ATP à la sortie de protons
c. Quinones
Les quinones possèdent un noyau benzénique ou naphtalénique et une chaîne isoprénoïque qui permet la mobilité dans la membrane. Les quinones sont des transporteurs de protons et d’électrons.

On distingue 3 types de quinones : - Qa : plastoquinone primaire très liée à D2 - Qb : plastoquinone secondaire faiblement liée à D1 - PQ : collectif des plastoquinones libres dans la membrane
Photosystème II
HYDROPLASTOQUINONE
1e-
Qa Qa- Qa Qa- Qa
Qb Qb- PQ H2
2e-
2H+ (stroma)

5. Les réactions de la photosynthèse
a. Réception et transformation de l’énergie lumineuse
- Un photosystème est un ensemble de protéines et de pigments. - L’antenne collectrice est composée de pigments accessoires chlorophylles a et b,
caroténoïdes et protéines LHC-II
- Le complexe central est composé de la chlorophylle a et des CP43 et CP47 - Le centre réactionnel contient une paire de chlorophylle à 680-D1-D2-Phéophytine-Qa-
Qb-Complexe producteur d’oxygène Mécanisme :
1. Excitation des pigments par les photons 2. Transfert par résonance de l’énergie d’excitation dans l’antenne. 3. Transfert successif d’électrons dans le centre réactionnel
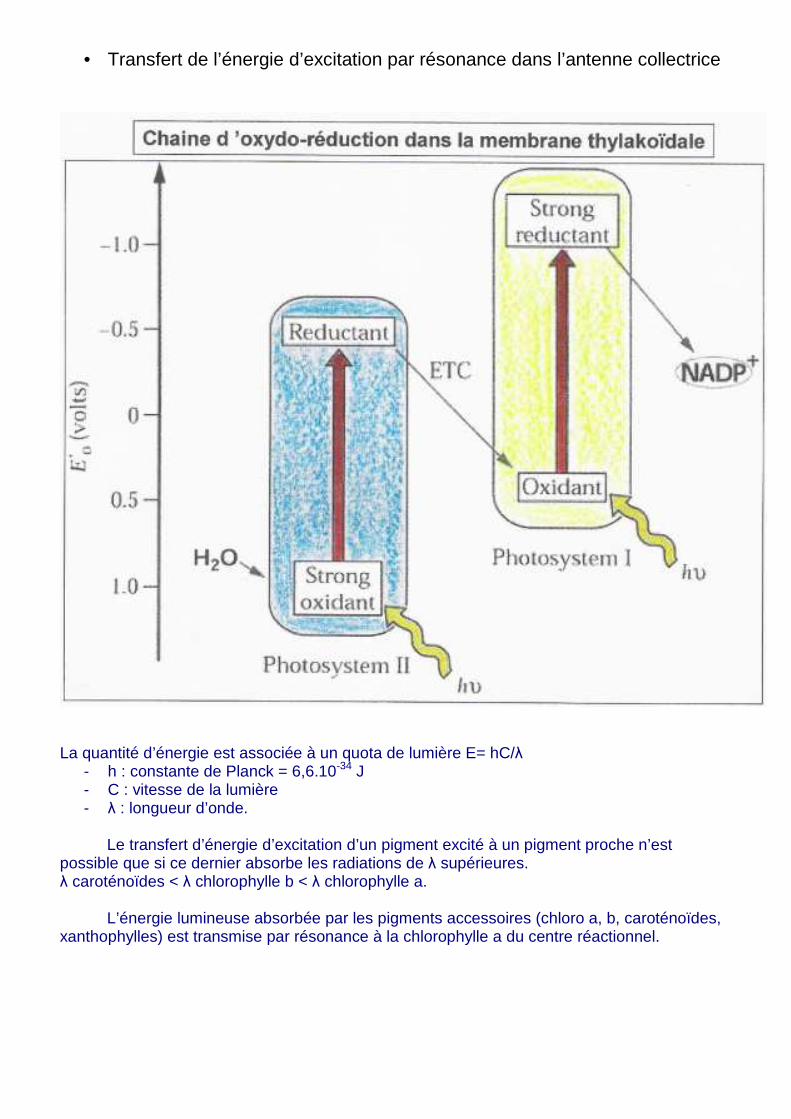
• Transfert de l’énergie d’excitation par résonance dans l’antenne collectrice
La quantité d’énergie est associée à un quota de lumière E= hC/λ
- h : constante de Planck = 6,6.10-34 J - C : vitesse de la lumière - λ : longueur d’onde.
Le transfert d’énergie d’excitation d’un pigment excité à un pigment proche n’est
possible que si ce dernier absorbe les radiations de λ supérieures. λ caroténoïdes < λ chlorophylle b < λ chlorophylle a. L’énergie lumineuse absorbée par les pigments accessoires (chloro a, b, caroténoïdes, xanthophylles) est transmise par résonance à la chlorophylle a du centre réactionnel.

• Transfert successif d’électron dans le centre réactionnel
La chlorophylle a devient un réducteur fort donnant facilement un électron. Elle devient oxydante et l’accepteur devient réduit.
b. Trajet des électrons au cours des réactions photochimiques
Séparation de charges dans le photosystème ACTE PHOTOCHIMIQUE. En présence de lumière, le P680 est excité. C’est un réducteur fort qui donne un électron à la phéophitine qui devient réduite et le P680 est oxydé. La phéophitine transmet son électron à Qa qui transmettra ensuite à Qb.
Le Photosystème II est un condensateur qui stocke l’énergie en séparant et en stabilisant les charges de part et d’autre de la membrane des thylacoïdes.
Les électrons réduisent la chlorophylle et proviennent de l’eau. L’oxydation de 2 molécules d’eau nécessite 4 photons, fournit 4 électrons et 4 protons grâce au complexe producteur d’oxygène du PSII.
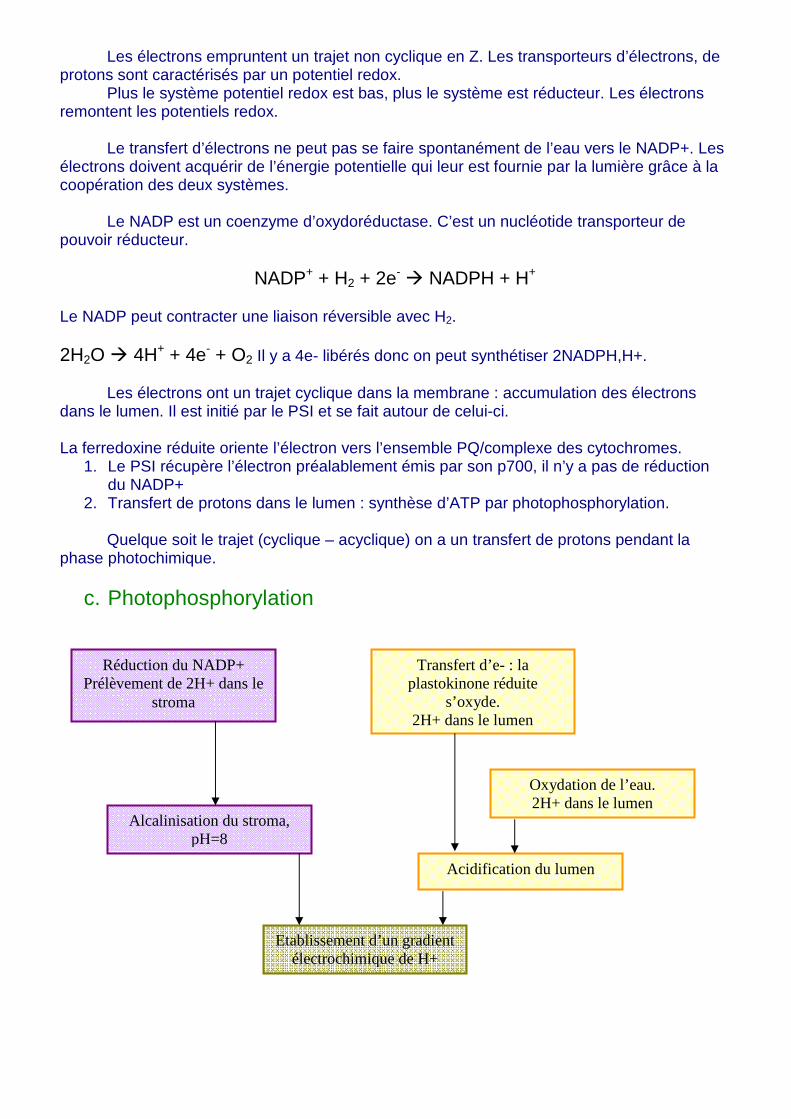
Les électrons empruntent un trajet non cyclique en Z. Les transporteurs d’électrons, de protons sont caractérisés par un potentiel redox. Plus le système potentiel redox est bas, plus le système est réducteur. Les électrons remontent les potentiels redox. Le transfert d’électrons ne peut pas se faire spontanément de l’eau vers le NADP+. Les électrons doivent acquérir de l’énergie potentielle qui leur est fournie par la lumière grâce à la coopération des deux systèmes. Le NADP est un coenzyme d’oxydoréductase. C’est un nucléotide transporteur de pouvoir réducteur.
NADP+ + H2 + 2e- � NADPH + H+ Le NADP peut contracter une liaison réversible avec H2. 2H2O � 4H+ + 4e- + O2 Il y a 4e- libérés donc on peut synthétiser 2NADPH,H+.
Les électrons ont un trajet cyclique dans la membrane : accumulation des électrons dans le lumen. Il est initié par le PSI et se fait autour de celui-ci. La ferredoxine réduite oriente l’électron vers l’ensemble PQ/complexe des cytochromes.
1. Le PSI récupère l’électron préalablement émis par son p700, il n’y a pas de réduction du NADP+
2. Transfert de protons dans le lumen : synthèse d’ATP par photophosphorylation.
Quelque soit le trajet (cyclique – acyclique) on a un transfert de protons pendant la phase photochimique.
c. Photophosphorylation
Réduction du NADP+ Prélèvement de 2H+ dans le
stroma
Alcalinisation du stroma, pH=8
Etablissement d’un gradient électrochimique de H+
Transfert d’e- : la plastokinone réduite
s’oxyde. 2H+ dans le lumen
Oxydation de l’eau. 2H+ dans le lumen
Acidification du lumen
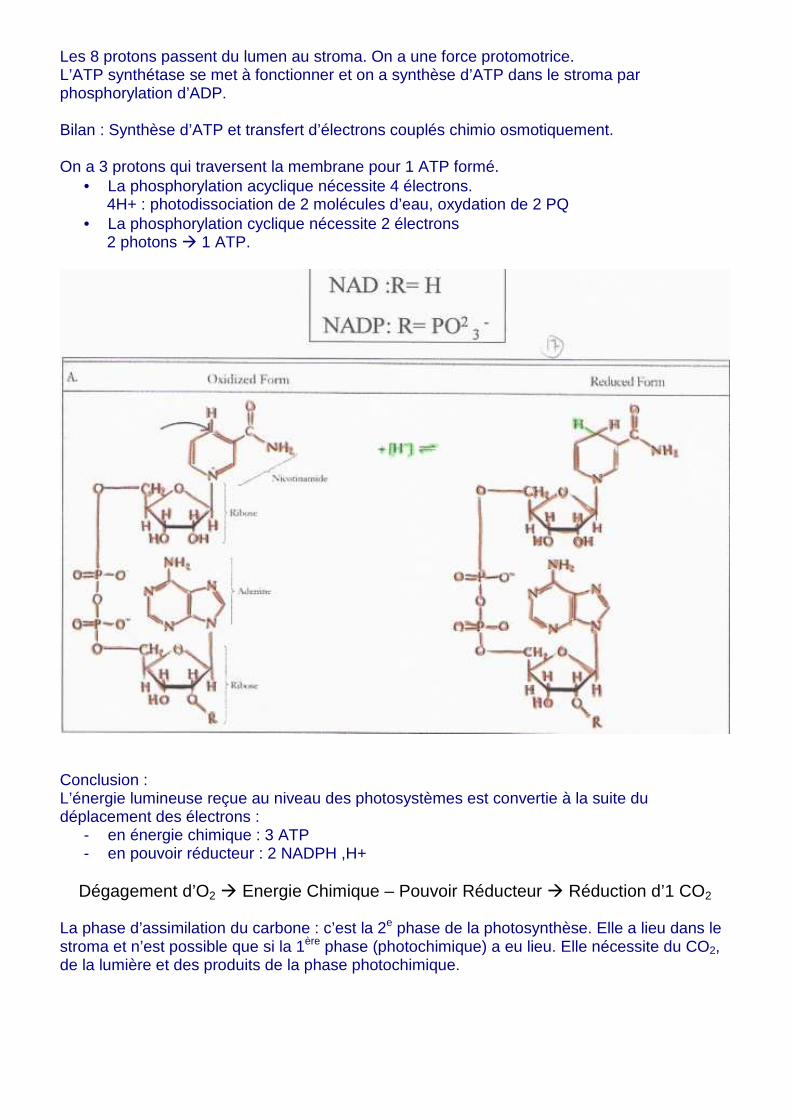
Les 8 protons passent du lumen au stroma. On a une force protomotrice. L’ATP synthétase se met à fonctionner et on a synthèse d’ATP dans le stroma par phosphorylation d’ADP. Bilan : Synthèse d’ATP et transfert d’électrons couplés chimio osmotiquement. On a 3 protons qui traversent la membrane pour 1 ATP formé.
• La phosphorylation acyclique nécessite 4 électrons. 4H+ : photodissociation de 2 molécules d’eau, oxydation de 2 PQ
• La phosphorylation cyclique nécessite 2 électrons 2 photons � 1 ATP.
Conclusion : L’énergie lumineuse reçue au niveau des photosystèmes est convertie à la suite du déplacement des électrons :
- en énergie chimique : 3 ATP - en pouvoir réducteur : 2 NADPH ,H+
Dégagement d’O2 � Energie Chimique – Pouvoir Réducteur � Réduction d’1 CO2
La phase d’assimilation du carbone : c’est la 2e phase de la photosynthèse. Elle a lieu dans le stroma et n’est possible que si la 1ère phase (photochimique) a eu lieu. Elle nécessite du CO2, de la lumière et des produits de la phase photochimique.
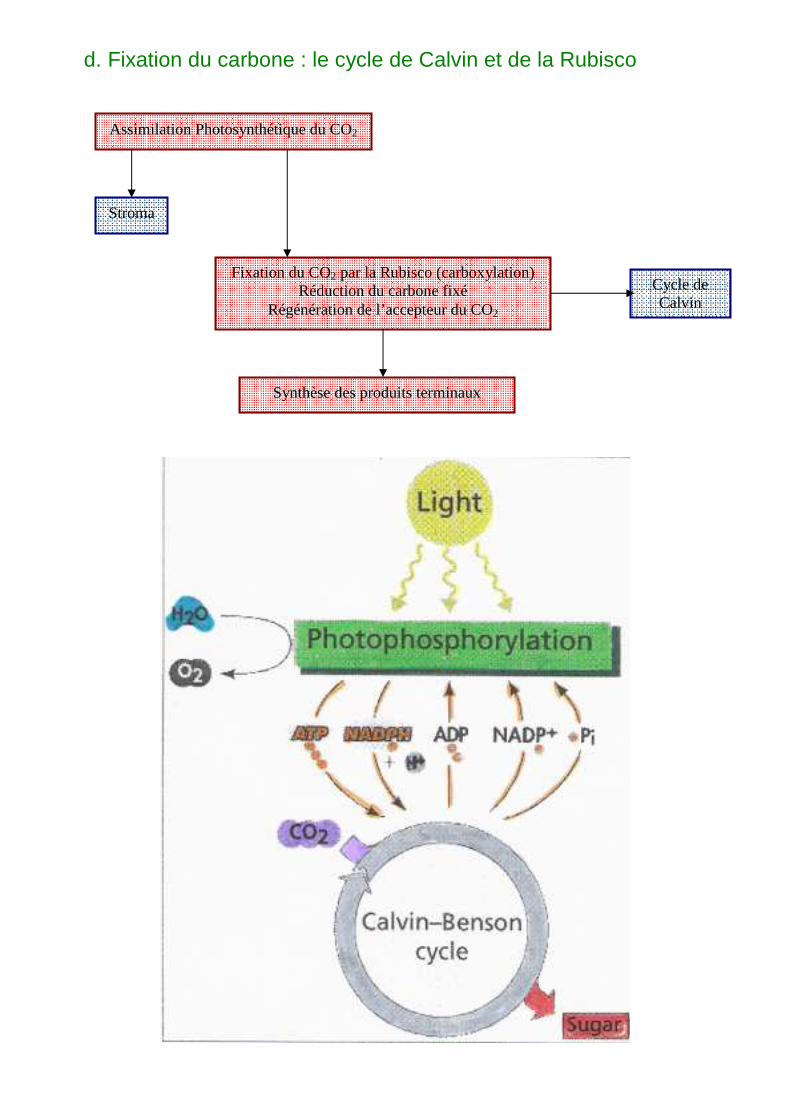
d. Fixation du carbone : le cycle de Calvin et de la Rubisco
Assimilation Photosynthétique du CO2
Stroma
Fixation du CO2 par la Rubisco (carboxylation) Réduction du carbone fixé
Régénération de l’accepteur du CO2
Cycle de Calvin
Synthèse des produits terminaux

Auto radiogrammes : 1ers produits carbonés de la photosynthèse : - Composés radioactifs formés par les algues - Séparés par chromatographie papier - Visualisé par autoradiographie

70% du carbone asymétrique se retrouvent au niveau de la molécule de tri-phospho-glycérate en 5s. Il a fallu a Calvin plus de 10ans pour identifier tous les composés du chromatogramme.
1. Le premier produit formé est le P-OCH2-CHOH-COOH APG : Acide phosphoglycérique 3PGA : tri phosphoglycérate –OOC-HCOH-CH2O-P
2. Augmentation de la concentration en APG : Diminution de la concentration en Rub-P et
Ribulose 1,5bi-P Substrat P-OCH2-CHOH-CHOH-CO-CH2O-P La Rubisco est une enzyme qui permet la fixation du CO2 sur le Rub-P.
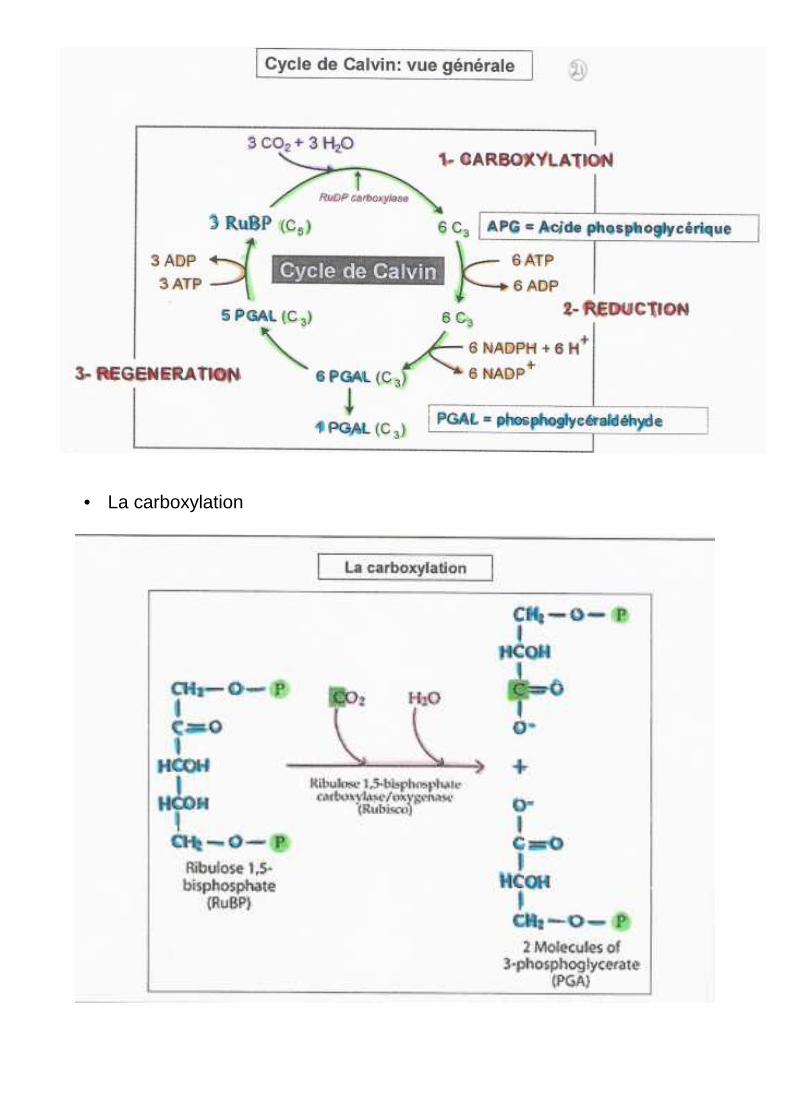
• La carboxylation

Assurée par la rubisco : ribulose 1,5 bi phosphate carboxylase oxygénase. C’est la protéine soluble la plus abondante du chloroplaste et sans doute de la biosphère. Elle a besoin de lumière pour fonctionner. Il y a 16 sous unités 8A-8B (Poids Moléculaire : 560KDa) L8S8 (Large/Small).
- La sous unité B (S : small) de 14kDa joue un rôle dans l’activation de l’enzyme. Elle est codée par un gène nucléaire RBCS
- La sous unité A (L : large) de 56kDa porte le cycle catalytique. Elle est synthétisée dans le chloroplaste.
- A et B s’associent dans le chloroplaste et, les bactéries photosynthétiques n’ont pas de petites sous unités.
Les deux fonctions de la Rubisco
1. Activité Carboxylasique _ Fixation du CO2, Réduction 2. Activité Oxygénasique _ Fixation de l’O2, Oxydation.
Les deux gaz, CO2 (0,03%) et O2 (21%) sont présents simultanément dans l’atmosphère donc la rubisco fixe le CO2 en présence de l’ O2. Il y a compétition des deux gaz pour le site actif et, la rubisco a plus d’affinité pour le CO2 que pour l’O2. Il y a eu une adaptation des plantes à l’enrichissement de l’atmosphère en O2
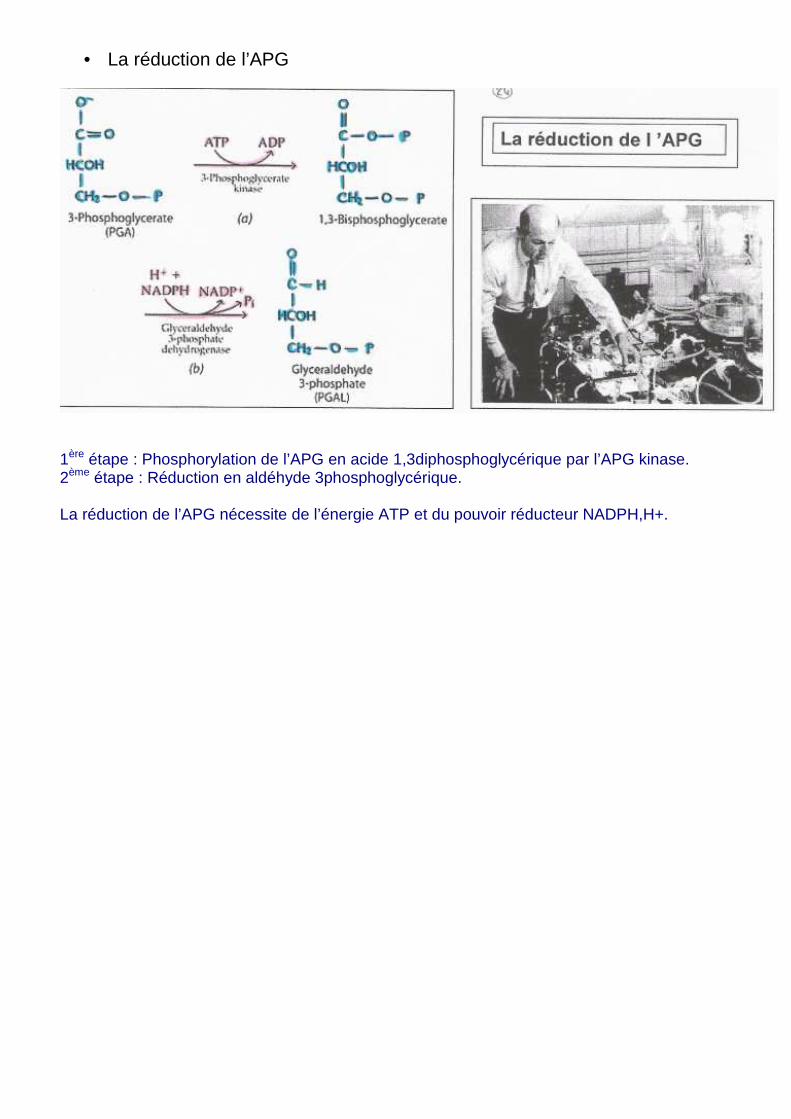
• La réduction de l’APG
1ère étape : Phosphorylation de l’APG en acide 1,3diphosphoglycérique par l’APG kinase. 2ème étape : Réduction en aldéhyde 3phosphoglycérique. La réduction de l’APG nécessite de l’énergie ATP et du pouvoir réducteur NADPH,H+.

• La régénération du RubP

Nécessite 3 ATP. Un triose phosphate nécessite en tout 9 ATP. Bilan : Equation générale de la photosynthèse
CO2 + H2O � CH2O + O2 � 6 CO2 + 6 H2O � 1 C6H12O6 + 6 O2
� 6 CO2 + 12 H2O � 1 C6H12O6 + 6 O2 + 6 H2O Cycle de Calvin Photolyse 2H2O : 2NADPH,H+ et 2ATP Synthèse d’1C2 : fixation de 3CO2 sur 3RubP, 6ATP et 6NADPH,H+ pour la réduction des trioses, 3ATP pour la régénération des 3RubP, 6H2O oxydées et 30 photons nécessaires.

6. La photorespiration

1. Synthèse de P-Glycolate 2. Déphosphorylation et obtention du glycolate qui va être amené au péroxysome. Il réagit
avec l’oxygène pour donner du glyoxylate et de l’H2O2 qui sera éliminé par une catalase.
3. Le glyoxylate va subir une amilation. L’aminotransférase transfert le groupement amine du glutamate au glyoxylate et on obtient la glycine. Elle quitte le péroxysome pour aller dans la mitochondrie. Il faut 2 glycines pour poursuivre les réactions.
4. La première glycine est décarboxylée et on a synthèse de NADH avec libération de NH3. Ce qui reste se combine avec une deuxième glycine pour former la sérine qui va dans le péroxysome.
5. Désamination et obtention d’hydroxypyruvate qui va être réduit par le NADH pour former du glycérate. Il migre dans le chloroplaste pour être phosphorylé par une kinase et donner du phosphoglycérate.

a. Caractéristiques On a quatre compartiments : chloroplaste, péroxysome, mitochondrie et cytoplasme. La respiration utilise de l’oxygène : RubP + O2 � PGA + 2 P-glycolate. On a un rejet de CO2 dans la mitochondrie et il faut de la lumière pour les réactions.
b. Bilan de la photorespiration
Il n’y a pas de forme active d’O2. L’acide P-glycolique est éliminé. On a synthèse :
- de NADH,H+ dans la mitochondrie qui sera ensuite utilisé dans le péroxysome.
- D’APG dans le chloroplaste et perte de CO2 dans la mitochondrie � intérêts des stomates fermés
- De glycine et sérine qui permettent la synthèse de protéines - De NH3 qui permet la synthèse de glutamine.
Contrairement à la respiration, la photorespiration ne libère pas d’énergie mais
consomme de l’ATP et ses équivalents réduits.
7. Le métabolisme photosynthétique des plantes en C3 � Orge, blé, tournesol, betterave, légumineuse, arbres. L’APG est un C3. C’est le premier produit stable.
Rubisco
CO2 Activité Carboxylasique
Photosynthèse
O2 Activité Oxygénasique
Photorespiration
Cycle de Calvin Réductif
Dégagement d’O2 Fixation du CO2
Cycle de Tolbert Oxydatif
Dégagement de CO2 Fixation de l’O2

PHOTOSYNTHESE NETTE = PHOTOSYNTHESE BRUTE – PHOTORESPIRATION
La photosynthèse nette représente le taux d’assimilation et la photosynthèse brute représente la capacité d’assimilation du CO2.
Les réactions d’oxygénation ne peuvent être évitées � 50% du CO2 fixé par photosynthèse est perdu par la photorespiration. Si on a [CO2] > 1% ou [O2] < 2% on a annulation de la photorespiration.
8. Le métabolisme photosynthétique des plantes en C4
a. Caractéristiques Ce sont des plantes monocotylédones : maïs, canne à sucre, graminées tropicales. On a formation d’un sucre en C4 : l’oxaloacétate qui donne rapidement le malate.
Le rendement photosynthétique est très élevé. Le CO2 est utilisé très efficacement donc son taux d’assimilation est proche de la capacité d’assimilation. Il n’y a pas de photorespiration.
Les plantes présentent une anatomie feuillaire particulière de même que leur
métabolisme.
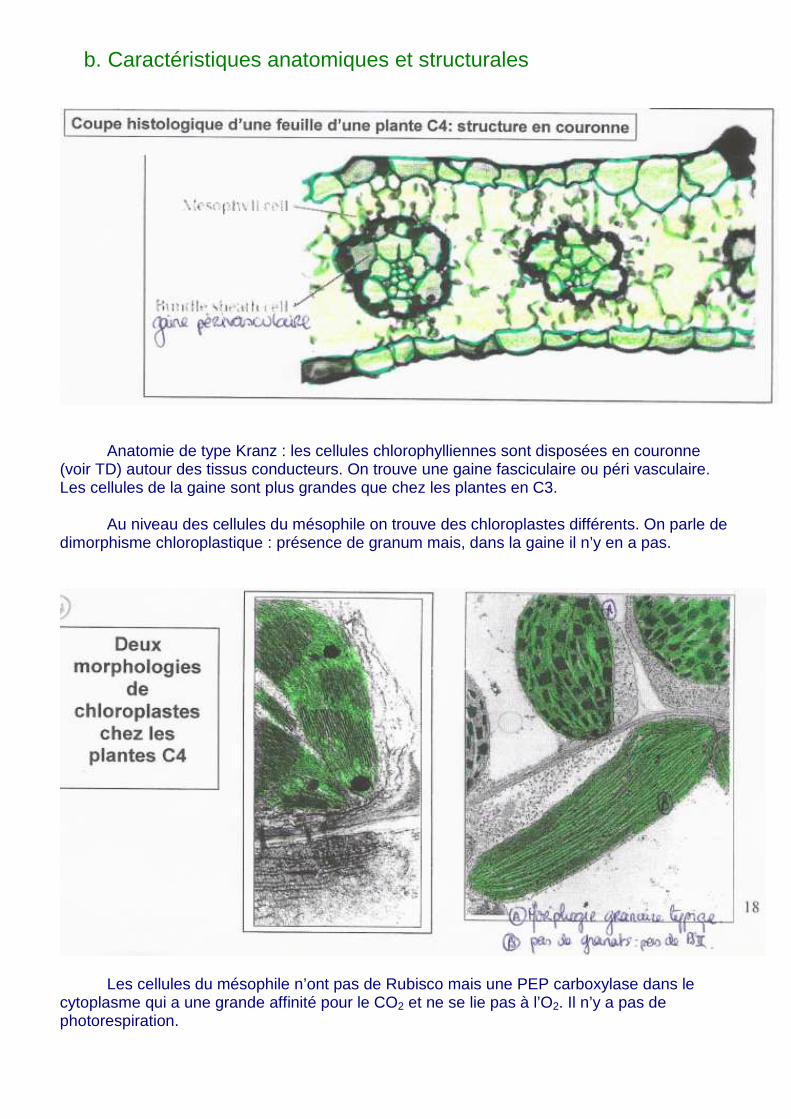
b. Caractéristiques anatomiques et structurales
Anatomie de type Kranz : les cellules chlorophylliennes sont disposées en couronne (voir TD) autour des tissus conducteurs. On trouve une gaine fasciculaire ou péri vasculaire. Les cellules de la gaine sont plus grandes que chez les plantes en C3.
Au niveau des cellules du mésophile on trouve des chloroplastes différents. On parle de
dimorphisme chloroplastique : présence de granum mais, dans la gaine il n’y en a pas.
Les cellules du mésophile n’ont pas de Rubisco mais une PEP carboxylase dans le cytoplasme qui a une grande affinité pour le CO2 et ne se lie pas à l’O2. Il n’y a pas de photorespiration.

Dans le cytoplasme des cellules mésophilliennes le CO2 se combinent à un C3 (PhosphoEnolPyruvate) pour former un C4 (oxaloacétate) et, la réaction est catalysée par la PEP carboxylase.
Dans les cellules de la gaine péri vasculaire on a accumulation de CO2. Elles possèdent la Rubisco. Il va y avoir formation d’un C3 qui va permettre le cycle de Calvin et formation de sucres

La rubisco utilise son activité carboxylasique. Il n’y a pas de PSII donc pas de photolyse de l’eau donc pas d’O2 et pas de photorespiration. Les plantes en C4 se sont adaptées aux fortes insolations et à la sécheresse.
Il existe environ 260 000 espèces de plantes : 5% sont des C4 (plus productives) et 95% des plantes en C3. C3 : il faut 18 ATP pour produire un glucose C4 : il faut 30 ATP.
Adaptation à l’abaissement de la teneur en CO2 au cours de l’évolution. Le photosystème des C4 est devenu insensible à l’O2 Adaptation aux fortes températures. Les C4 peuvent maintenir leurs stomates fermés. Colonisation des milieux très éclairés et à faible teneur en eau. Productivité élevée si les plantes sont en plein soleil avec de l’eau.
Fortes insolations, Sécheresse
Faible ouverture des stomates.
� Peu de CO2 disponible.
Double carboxylation séparée dans l’espace
Décarboxylation du malade : |CO2| x 10
Fixation par PEPc cytoplasmique
Fixation par Rubisco chloroplastique
Vitesse de carboxylation de la Rubisco élevée. Pas ou Peu de PHOTORESPIRATION
PRODUCTIVITE MAXIMALE
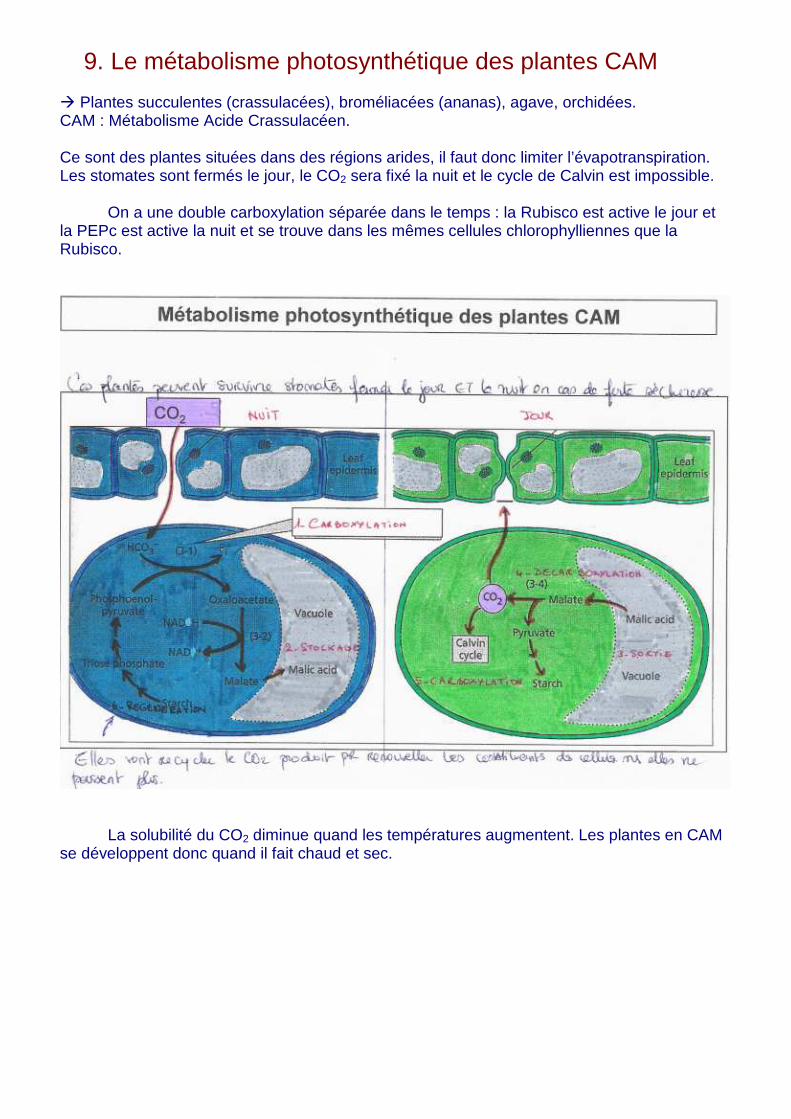
9. Le métabolisme photosynthétique des plantes CAM � Plantes succulentes (crassulacées), broméliacées (ananas), agave, orchidées. CAM : Métabolisme Acide Crassulacéen. Ce sont des plantes situées dans des régions arides, il faut donc limiter l’évapotranspiration. Les stomates sont fermés le jour, le CO2 sera fixé la nuit et le cycle de Calvin est impossible.
On a une double carboxylation séparée dans le temps : la Rubisco est active le jour et la PEPc est active la nuit et se trouve dans les mêmes cellules chlorophylliennes que la Rubisco.
La solubilité du CO2 diminue quand les températures augmentent. Les plantes en CAM se développent donc quand il fait chaud et sec.

10. Biosynthèse de l’amidon et du saccharose : les produits de la photosynthèse
Dans le cytosol, nécessite de l’UTP pour convertir le glu-1P et de l’ATP dans le chloroplaste. Le Saccharose est un dimère de fructose et de glucose, il est soluble et très mobile et, il constitue une forme de transport des sucres dans la plante. Il est synthétisé dans le cytosol à partir de trioses-P et d’UTP. Il est distribué aux organes puits (consommateurs) par le phloème. Le saccharose est dégradé par la saccharose synthase (ou invertase). Il existe de nombreuses isoformes de ces deux types d’enzymes et elles sont tissus spécifique.
C3, C4, CAM � Carbone du CO2
Trioses-P
Acides Gras
Acides Aminés
Acides Nucléiques
Photosynthètats majeurs : Glucides
Amidon Saccharose
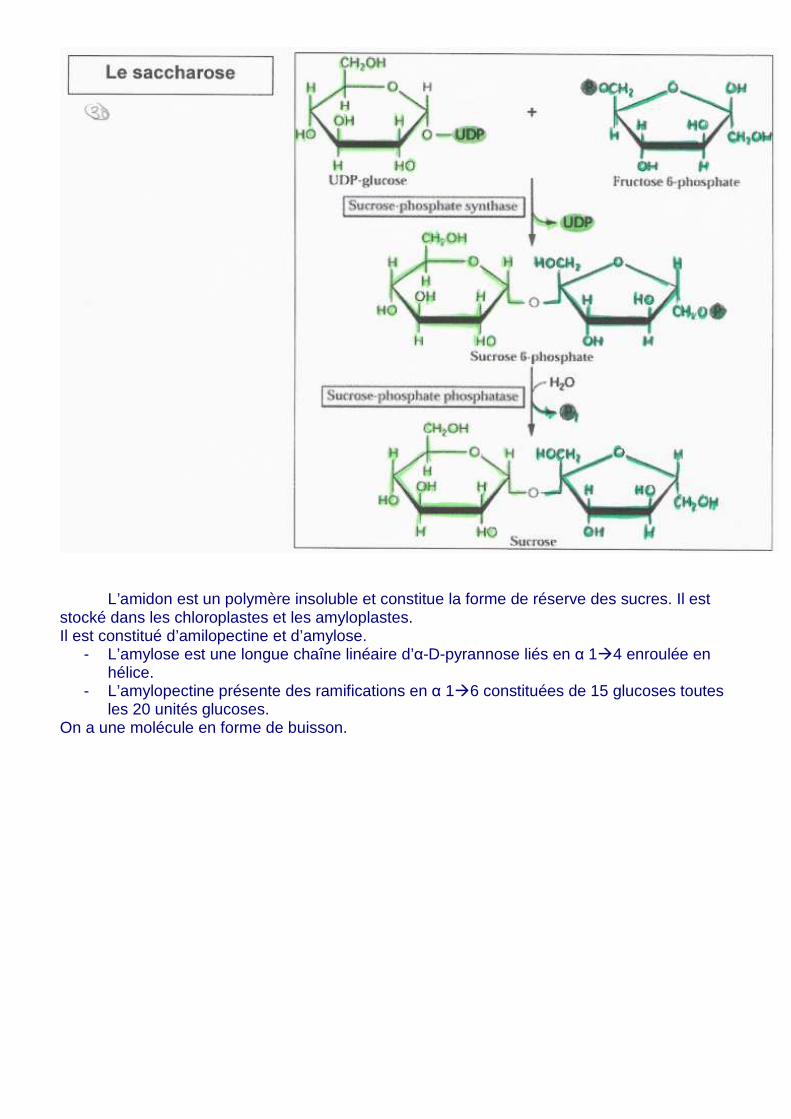
L’amidon est un polymère insoluble et constitue la forme de réserve des sucres. Il est stocké dans les chloroplastes et les amyloplastes. Il est constitué d’amilopectine et d’amylose.
- L’amylose est une longue chaîne linéaire d’α-D-pyrannose liés en α 1�4 enroulée en hélice.
- L’amylopectine présente des ramifications en α 1�6 constituées de 15 glucoses toutes les 20 unités glucoses.
On a une molécule en forme de buisson.

L’amidon est synthétisé dans les chloroplastes pendant la journée et nécessite de l’ATP par l’ADP glucosepyrophosphorylase. Il est hydrolysé la nuit en triose-P par le complexe amylasique et l’amidon phosphorylase + enzyme D. Régulation : interconvertion entre l’amidon et le saccharose
• Jour : synthèse de triose-P en quantité importante puis transfert vers le cytoplasme par transporteur pour synthèse de saccharose.
• Mi journée : trop de saccharose par rapport aux besoins de la plante donc inhibition de la synthèse et début de la synthèse d’amidon dans le chloroplaste pour le stockage des sucres
• Soir : plus de lumière donc plus de phosphorylation donc accumulation des Pi dans le chloroplastes et inhibition de la synthèse d’amidon qui finira par être dégradé.


La plupart des carbones réduits va permettre la synthèse de nombreux métabolites.
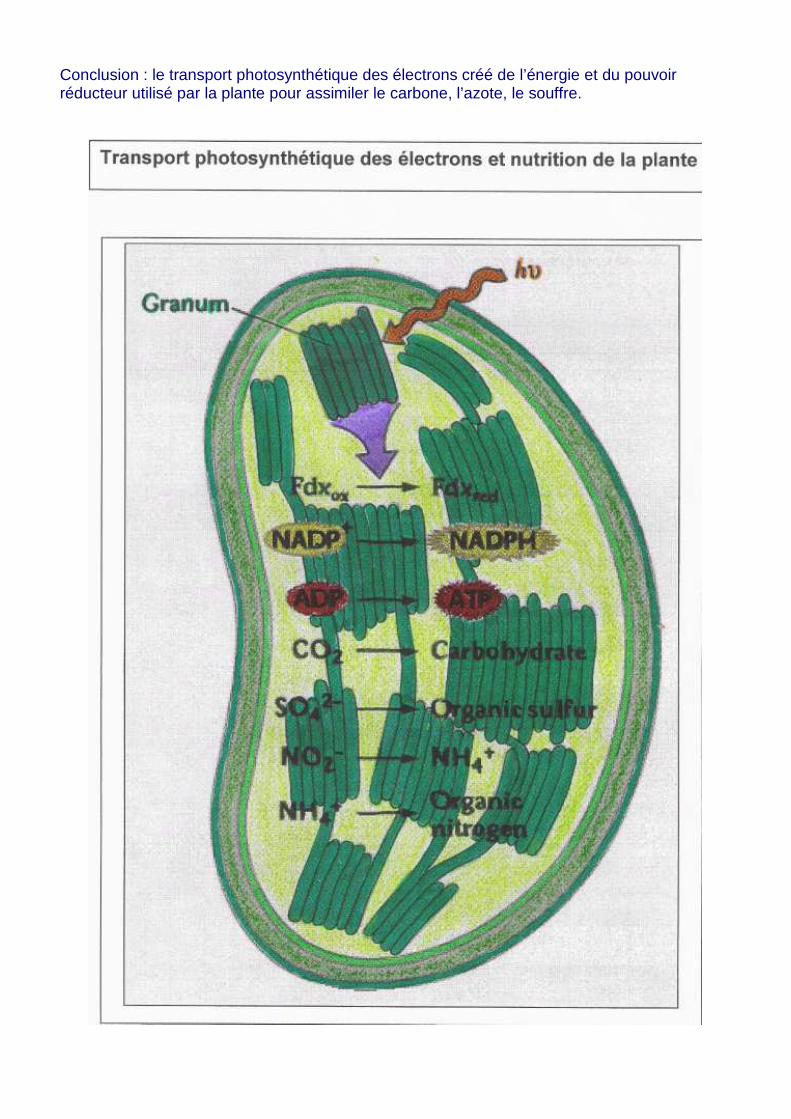
Conclusion : le transport photosynthétique des électrons créé de l’énergie et du pouvoir réducteur utilisé par la plante pour assimiler le carbone, l’azote, le souffre.

11. Photosynthèse nette des plantes d’ombre et de lumière en fonction de l’éclairement.
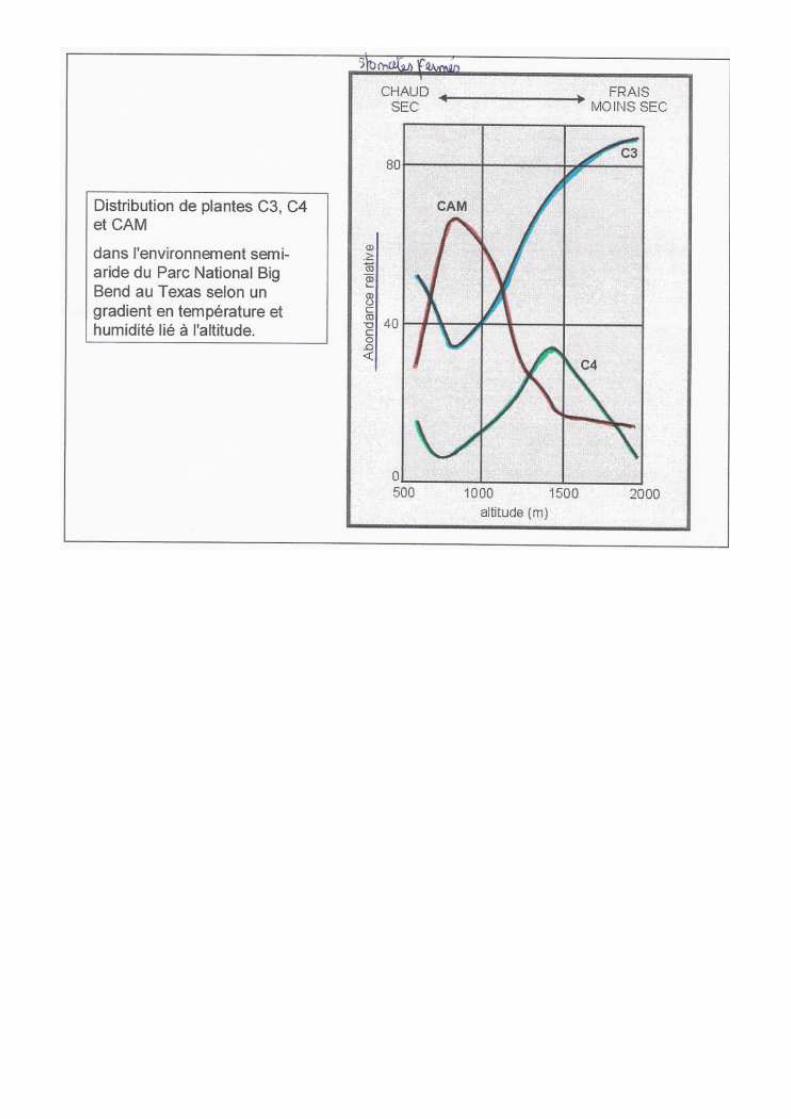
Les plantes d’ombre ont plus de pigments d’antenne et de grana et peuvent donc réaliser la photosynthèse en faible présence de lumière. Les capacités photosynthétiques des plantes de plein soleil sont plus élevées car le plateau est atteint plus tôt. Métabolisme photosynthétique : plateau de saturation atteint pour des intensités lumineuses différentes : C3 : 1/3 de la valeur solaire max donc le facteur limitant n’est pas la lumière contrairement aux C4. Pour les C3, plus on augmente la lumière et plus on augmente la photosynthèse mais aussi la photorespiration par la Rubisco donc le facteur limitant est le CO2.

La teneur en CO2 : [CO2]atm = 0,03% Cette concentration est doublée au dessus d’un sol riche en humus. Elle augmente près d’une route à grande circulation C’est le principal facteur limitant de la croissance des plantes. Les C4 fixent le CO2 même en très faible concentration grâce à la PEPc. Dans l’atmosphère il n’y aura jamais assez de CO2 pour qu’elles puissent atteindre le plateau de saturation. Le CO2 est le facteur limitant des C3.

La teneur en O2 : [O2]atm = 21% Si on se place en atmosphère artificielle de 2 à 5% d’O2 il n’y a pas de photorespiration et on augmente le taux d’assimilation du CO2 Une concentration élevée en O2 limite les rendements photosynthétiques d’une plante en C3.

La température de la feuille : Plus on augmente l’intensité d’éclairement et plus la température augmente mais, la température ambiante ne change pas. La température de la feuille dépend de l’éclairement, de la température ambiante, de la turbulence de l’air et de l’évapotranspiration
Si on enlève l’O2, la courbe des plantes en C3 se rapproche de celle des plantes en C4. Les plantes en C4 présentent un optimum de température plus élevé que les plantes en C3 et, de faibles concentrations en O2 augmentent l’optimum des plantes en C3.

La température agit sur les activités enzymatiques 1. Optimum thermique
C3 : 25°C - 35°C C4 : 40°C – 50°C Si on augmente la température, on augmente l’activité oxygénasique de la rubisco et on diminue la solubilité du CO2 dans l’eau � plus de photorespiration, moins de photosynthèse.
2. Sol froid Photosynthèse limitée par le métabolisme racinaire.
3. Température foliaire faible mais grand soleil Cycle de Calvin non opérationnel, photosystème engorgés � photoinhibition.
L’eau
- Effets stomatiques : déshydratation modérée des feuilles et dessèchement du sol, stress hydrique du système racinaire � fermeture des stomates et baisse de la photosynthèse.
- Effets non stomatiques : déficit hydrique important � déshydratation des tissus, pertes de protéines enzymatiques, chute des feuilles, photosynthèse limitée à long terme.














