
LA - LPO Coordination Auvergne-Rhône-Alpes · lll - Commentaire par espèce - MERLE NOIR lurdus et...
111
Transcript of LA - LPO Coordination Auvergne-Rhône-Alpes · lll - Commentaire par espèce - MERLE NOIR lurdus et...


Yves THONNERIEUX
MIGRATION ET H¡VERNAGE DANSDES ESPECES APPARTENANT
SYNTHESE ET ANALYSE DES
LA REGION RHONE.ALPES
AU GENRE TURDUS.REPRISES DE BAGUES
Référence: THoNNERIEUX (y.) - l981 - Migration et h¡vernage dans la région Rhône-Alpes des espèces appartenant au genre Turdus; synthèse et analyse des reprises de ba-gugs- Bièvre,3 (1), 1 - 54 - cenrre Bégionar de Baguage d'oiseaux de LyoN (c,o.R.A.),Université LYON I -69622 VTLLEURBANNE CEDEX.
lé1u.7é;.Lesfichesdereprisesrégionalesde486oiseauxappartenantaugenre Turduson1été dépouillées. Les conclusions majeures qui se dégagent d'e'cette anarvieäni les suivantes :
.Draines migratrices qu i Rhône-Alpeslemagne orientale. Leur incipalement lamais des reprises s'échel er. Les nicheursartie sédentaires, même

2
Turdus musicus.'comme pour la Grive Litorne, la population f inlandaise (et d'Estonie)transite en nombre par la région Rhône-Alpes. Lds pays scandinaves sont par contrefaiblement représentés. Les reprises régionales s'étendent régulièrement de la mi-octobreà la mi-février avec toutefois une hausse accentuée ma¡s limitée à la première quinzainede décembre. Les HautesAlpes (ainsi que la Drôme et la Haute-Savoie) paraissent parti-culièrement propices à cette Grive. Comme pour l'espèce précédente, des individus (duNord de l'Europe ? ) passent par l'ltalie avant d'aboutir dans la région RhôneAlpes.
Les changementsde zones d'hivernage d'une année à l'autre sont man¡festementcourants chez la plupart des espèces.
D'autres détails comme la longévité, certains records de distance/temps, l'age-ratio,... sont abordés dans ce travail.
Summary.' Migration and wintering in the RhôneAlpes region of species belonging tothe genus Turdus ; a synthesis and analysis of ring recoveries,
The regional recovery record cards of 486 birds belonging to the genus furd¿rshave been processed. The major conclusions merging from this analysis are the follo-wrng :
Turdus merula.'the populations f rom Central Europe (from Czecoslovak¡a in particular)and from the eastern shore of the Baltic migrate and winter in the Rhône-Alpes region.The stronge between id-november.They involv sentially. migrating birdscomes in de s of indiv tre of EuroPe.The stronge in the de e and Ardèche.Some of the Rhône-Alpes breeders are sedentary, the others winter in the Mediterraneandepartments, but also in Corsica and Spain,
Turdus torquatus.'some individuals probably coming from Norway frequent the Rhône-Alpes regions d migrat êr,but Ring Ouzel late asregion. The onl countsmigration of th to be clater recovered in the Baleanic lslands and in ltaly.Turdus phílomelos ,'two populations unquestionatly f requent the Flhône-Alpes regionone coming from inating from the North-East of Europe(very likely from , some British birds may be involved.The post-nuptial ulations takes place in october andnovember, with a A small, later arrival sames to occur inJanuary, essentially from Central Europe, The Song Thrush is found wintering in twodepartments of the southern Rhône-Alpes region (Drôme and Ardtàche), therby for-ming the northern limit of the w¡nter distribution of the migrating populations of thisspecies. Other individuals have been found in this season in Mediterranean France,Spain, the Balearic lslands, Sardinian and Italy.
Turdus viscivorus.'the migrating Mistle Thrushes visiting the RhôneAlpes region comefrom Sw¡tzerland and East Germany, Their passage occurs mainly in the second fort-night of october but recoveries spread out until january. The regional breeders are atleast partially sedentary, even at moderate altitude.
Turdus pitaris.'two distinct breeding nuclei regularly frequent the Rhône-Alpes region :
they are, on the one hand, Finnish birds (and to a lesser extent birds from southernScandinavia) and on theother hand populationsfrom Central Europe (from Poland to
eofma
cterthe mnern
the Rhône-Alpes region via an indirect route across ltaly,

Turdus musicus.'like the Fieldfare, the Finnish (and Esthonian) population crosses theRhône-Alpes region in large numbers, On the other hand, Scandinavian countries arepoorly represented. Fìegional recoveries regularly spread out from mid-october to mid-february, with a marked increase restricted, however, to the f irst fortnight of ciecember.The Hautes-Alpes (together with Drôme and Savoie) seem to be particular favourable tothis thrush. As with the prev¡ous species, some individuals (from Northern Europe ? ) gothrough ltaly before reaching the Bhône-Alpes reg¡on,
Shifts in wintering ranges from one year to another are obviously common inmost species.
Other details such as longevity, some records in distance/time, the age-rat¡on,are dealt with in this work.
| - lntroduction
Les 306 espèces classées dans la grande famille des Turdidés se divisent en 45genres dont le plus répandu est le genre Turdus regroupant une soixantaine d'espècesdans le monde. Une seule habite l'Amérique du Nord alors qu'une vingtaine occupe le
Centre et le Sud de ce continent. Mais c'est surtout dans l'Ancien Monde que le genre
Turdus est le plus solidement implanté : l'Asie est peuplée par plusieurs espèces, parti-culièrement la Sibérie orientale (AUSTIN et SINGER, 1973). L'Europe, pour sa part,est concernée par 6 grands Turdidés qui occupent une place très importante áu æin de
son avifaune : pas une seule région de notre continent n'est en effet totalement vide de
Merles et (ou) de Grives. Des facultés d'adaptation importantes et un tempérament er-rat¡que aigu ont d'ailleurs récemment permis la colonisation de milieux nouveaux (jar-
dins, vergers, squares) et de terres lointaines (lles de l'Atlantique Nord) par plusieursreprésentants du genre. Les aires de reproduction et d'hivernage des 6 Turdidés euro-péens se chevauchent largement: aucune espèce ne remplace entièrement une autregéographiquement. La concurrence inter-spécifique qui pourrait en résulter est limitéepar un ordre de préférence alimentaire variable (ASHMOLE, 1962).Les représentants européens du genre Turdus (à l'exception du Merle à plastron) sontmigrateurs partiels, ce qui signifie que certaines populations sont sédentaires alors que
d'autres appartenant à la même espèce se déplacent (parfois considérablement) à cer-
tains moments de l'année.Compte tenu de ce qui précède, on comprendra aisément que les Grives et les Merles
se situent en bonne place parmi les oiseaux bagués en Europe. Leur statut français degibier explique quant à lui l'abondance des repriæs de bagues enregistrées chez ces
espèces. Le Centre de Baguage rhônalpin en détenant globalement près de 500 (tabl. 1 ),
il nous a paru opportun de nous livrer à une synthèse régionale de toutes ces reprises,
allant en cela dans le sens des travaux du C.R.B.P.O. qui a récemment inscrit les Grives
et les Merles sur la liste du Programme National de Recherche.

4
Tableau 1 : Répartition des reprises par pays d'origine et par espèce
Symbolisation employée pour les états politiques: N = Norvège ; S = Suàle ;
SF = Finlande ; DK = Danemark ; NL = Pays-Bas i DW = Allemagne de l'OuestB = Belgique ; L = Luxembourg ; GB = Grande-Bretagne ; URSS = Russie ;
PL = Pologne ; CS = Tchécoslovaquie ; DE = Allemagne de l'Est ; A = AutricheCH = Suisse ; F = France (hors Rhône-Alpes) ; SP = Espagne ; I = ltalie ;
YU = Yougoslavie.
il 1li N s SF NI I L ,Llcs ol A F
IIERLE NOIR I t5 3 12 t8 3 50 t U
M. A PLASTRd 1 I 7 IGRIVE MUSICIENNI t0 5 7 t9 1 I 3 7 I 6 5 50 t 1 0
G. ORAIN€ I z IG. LITORNE I E 29 5 1a t3 5 6 3 4 E 48 5 I l5¿
G. MAUVIS 3 I t9 I { 3 3 t0 2 I I 5 { 5l
ï0Tlt /PAYS
& (%l7
.t20 5t t:
11I 60
I ?,3
2 12
2,â
26 25 38 r9(t)
1 1 I 1t I
ll - Méthode de travail
Nous avons procédé dans un premier temps à un dépouillement par espèce des
données disponibles sur les fiches de reprises.L'abondante littérature existante, anglaise en particulier avec les travaux de ASHMOLE(1962) et de SIMMS (1978\, nous a permis de replacer nos résultats régionaux dans uncontexte plus général. En outre, l'étude semblable menée par JONES en 1961 à partirdes reprises obtenues sur les mêmes espèces en France méditerranéenne, nous a fournil'occasion de comparer les situations respectives de nos deux régions voisines.En cours d'étude, s'est imposée I'idée d'une enquête à la <source> et au <but> des dif-férentes reprises :
- à la sourcê, c'est-à-dire auprès des organismes étrangers de baguage; car ilparaissait intéressant de situer nos reprises dans les effectifs annuels de Grives et deMerles bagués par pays. Ce premier contact n'a pas débouché sur des résultats vraimentexploitables ; certaines stations ornithologiques n'assurant pas toujours une publicationrégulière de leurs travaux annuels.
- au but, en second lieu, en contactant les Fédérations Départementales deChasæurs, puisque plus de 99 % des reprises rhônalpines de grands Turdidés provien-nent de l'activité cynégétique. Nous avons obtenu de leur part certains renseignementstels que les effectifs de chasseurs par département depuis vingt ans et les dates de fer-meture annuelle du tir aux Grives et Merles dans le même laps de temps. A l'aide decæ chiffres, il nous a été possible d'évaluer. d'après les reprises enregistrées dans cha-

que départemênt (tabl. 2l et la pression de chasse à laquelle chacun d'eux était soumis(fig. 1),destauxdefréquentationdépartementauxparespèce (fig.4,B, 12,16,20,24) ,
beaucoup plus parlants que læ données brutes dont nous disposions initialement (1). uncalcul non paramétrique de statistique de tendance nous a permis d'éprouver la valeurdes résultats ainsi obtenus. Chaque fois que ceux-ci auront été jugés signif icatifs, nous enferons référence dans le texte.
Figure 1 : Pression de chasse par département rhônalpin - Les chiffres gras repréæn-tent la pression de chasse par département (exprimée en pourcentages) d'après le nom-bre de chasseurs (moyenne depuis 20 ans) ; les chiffres fins correspondent à une formedifférente de pression (également exprimée en pourcentages), basée sur la durée annuel-le par département du tir légal aux Grives et aux Merles. On remarque que c'est essen-tiel lement le nombre de chaseurs qui varie d'un département à l'autre ; les écartsétant moindres entre les valeurs fondées sur les jours de chasse. La conclusion logiqueest que les reprises de bagues par département sont surtout tributaires des effectifs deporteurs de fusils.
(1) Ce calcul se fonde sur le postulat selon lequel le nombre des oiseaux repris bagués est propor-tionnel au nombre des oiseaux porteurs d'une bague, effectif lui-même proportionnel au nombredes individus globalement présents par espèce et par département ; restant entendu que nous n'a-vons pas la certitude que certaines populations de Turdidés originaires de pays ou de régions où lebaguage est faible (voire ìnexistant) ne fréquentent pas la zone couverte par les départements rhô-nalp ins.
sAVO I E
AROECHE13,9
O ROME
t{
UTES ALPES

6
Tableau 2 : Répartition des reprises d'origine extérieure par département rhônalpin etpar espèce
(\¡rtl¡'lEõJ
o)(ol¡¡zo¡E
5z;
ñl¡JIølrl,I
(I,Ngoan
utß
$(t,!'I
øÉ,l¡,1.El¡¡
2
(o(\¡ul
=oÉo
ou¡tou¡oG
rc.9EèØg<.et
M ERLE NOIR l0 E I 4 2 3 3l 2Ã t5 t0t
M. A PLASTBON I 0 2 3 3 4 0 2
GRIVE ÀIUSICIENNI I { I 0 2 f4 54 t8 t4f
G. DRAINE 2 0 I 0 0 3 4 I I
G. L ITORNE r3 I 5 I a 22 35 36 21 l5¡
G. MA UV IS 6 3 2 3 0 5 l2 I 7 5:
TOTA I. / OIPART.
& (%)40 22 28
(¡r.IEiU,
t4,'4 36
3,ft2!25)
t4(rtf
63(t3
t8t0l
lll - Commentaire par espèce
- MERLE NOIR lurdus merula
Cette espèce est à la fois commune (7 000 000 de couples dans les lles britan-niques par exemple :SHARROCK, 1975) et largement représentée dans toute l'Europe,
au moins jusqu'au 65e degré de latitude N (ce qui exclut une grande partie des pays
fenno-scandinaves). Des populations orientales englobent une étroite bande comprise
entre les bords de la Mer Noire et les rivages chinois du Pacifique. Par ailleurs, la tota-
lité de l'Afrique du Nord est occupée.
L'examen de la figure 2 montre bien que ce sont les populations du centre de
l'Europe et, dans une moindre mesure celles de la rive orientale de la Baltique, qui tran-
sitent par la région Rhône-Alpes. Onze reprises d'individus bagués poussins désignent
l'Estonie (1 cas), l'Allemagne de l'Ouest (2 cas), la Suisse (2 cas) et surtout la Tchécos-
lovaquie (avec 6 cas) , comme patries d'origine des mi(¡rateurs et hivernants rhônalpins.
D'autres oiseaux bagués en période de nidification en Finlande (1 cas), Lithuanie (1
cas), Pologne (1 cas) , Allemagne orientale (2 cas), Allemagne de l'Ouest (2 cas) , Tché-
coslovaquie (3 cas) et Suisse (1 cas) constituent vraisemblablement des éléments sup-
plémentaires appartenant aux populations reproductrices de ces pays.

7
Le tableau 3 montre que les Merles noirs qui fréquentent la région Rhône-Alpes transitent par l'Allemagne de l'Ouest et la Pologne en septembre-octobre. EnSuisse, les 4 captures de la æconde quinzaine d'août laisænt entendre que certains dé-placements commencent déjà à devenir perceptibles. Sur un plan plus général, d'aprèsSIMMS (1978), ces premiers mouvements (essentiellement juvéniles :voir paragraphe lV),sont très désordonnés, ne trouvant leur orientation définitive qu'à partir de septembre.De fait, la migration devient surtout sensible chez nos voisins suisses à compter de sep-tembre (14 captures) , puis22 en octobre, avec un pic de 14 individus dans les quinzepremiers jours de ce mois. Cette situation est d'ailleurs confirmée par les observations aucol de Cou-Bretolet (DE CBOUSAZ,1960l..ll est remarquable de noter l'absence de captures (ayant donné lieu à une repriæ ulté-rieure) en Europe centrale au cours des mois de novembre -ASHMOLE (1962) associepourtant ce mois à celui d'octobre pour en faire la période de pointe dans le pasage del'espèce- et décembre. llest tentant de rattacher ce phénomène à une baisse numéri-que saisonnière dæ populations de Merles de ces pays, même si I'on peut penser quel'activité de marquage s'infléchit sans doute un peu au cours de la mauvaise saison.D'ailleurs, il est depuis longtemps admis que le tempérament migrateur du Merle noirest directement lié aux degrés de latitude qu'il occupe pendant l'été. Ainsi, à l'extrêmeNord de son aire, l'espèce est entièrement migratrice (ASHMOLE, 19621 ; alors qu'enSuède, d'après RENDHAL, seulement 65 % désertent les lieux; il en serait de mêmeen Tchécoslovaquie (FORMANEK). Mais la proportion tombe à 33 % en Allemagne(DROST),28o/oen Suisæ (BRUNNER) et 14o/oen Belgique (VERHEYEN) (Chiffres,n ASHMOLE,1962l.
A l'échelle régionale, une bonne fréquentation de l'espèce apparaît à la lecturedu tableau 4 et de la figure 3a en octobre, novembre et décembre. Manifestement, lamigration æ concrétise surtout à partir du 16 octobre (seulement 4 captures les joursprécédents et 20 les deux dernières semaines de ce mois). La baisse constatée la secondequinzaine de novembre semble tout à fait correspondre à un creux entre deux vagues;l'une d'automne, principalement constituée d'individus appartenant à la population duCentre pour qui la æconde moitié d'octobre repréænte le pic du passage dans notrerégion (fig. 3b) ; l'autre plus tardive, formée d'oiæaux sans doute chassés par le froidet l'enneigement en Europe du Nord. Sur la figure 3c, leur arrivée massive couvre latotalité du mois de décembre, ainsi que dans une moindre mesure la première quinzainede janvier. De tels mouvements hivernaux en relation avec le froid sont mentionnés parSIMMS (1978) : en janvier et février, les oiæaux qui en automne avaient gagné laSuàCe et le Danemark à partir de contrées situées plus à l'Est, traversent dans un second'temps la Mer du Nord et s'installent dans les lles Britanniques. Egalement au cæur deI'hiver, les populations locales de GrandeBretagne qui étaient restées sur place en au-tomne sont, au moins en partie, chassées par les intempéries. ll s'agirait, d'après ASH-MOLE ('1962) d'une stratégie intermédiaire entre l'état de sédentarisation et le phéno-mène de migration post-nuptiale. PERRINS (1974) écrit même que l'observation parradar des déplacements d'oiæaux au dessus de la Mer du Nord révèle l'existence d'unvaet-vient continu d'oiseaux entre le continent européen et la Grande-Bretagne ælon

I
le temps qui règne au milieu de l'hiver. Pourtant, cette hypothèse des mouvements dif-férés est mise en doute par SPENCER ,n sIMMS (1978) qui pense au contraire quebeaucoup de Merles n'ayant pas migré en début de saison se laissent surprendre par lesintempéries sans chercher à déserter les lieux et meurent en nombre.Dans la région Rhône-Alpes, la baisse de janvier doit être perçue avec prudence, étantcerta¡nement (au moins en partie) provoquée par l'intervention des dates légales defermeture de la chasse au Merle dans plusieurs départements. Aussi la désertion deshivernants rhônalpins est-elle impossible à jalonner dans le temps.
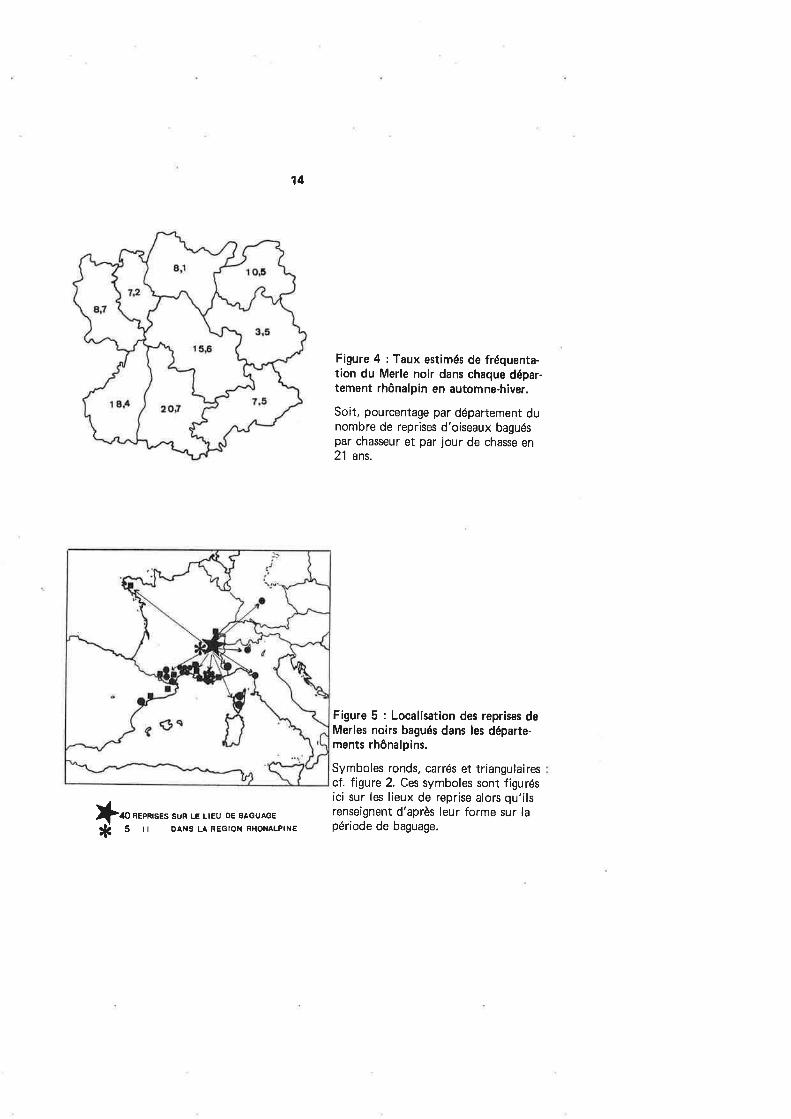
Sur la figure 4, la Drôme et l'Ardèche semblent particulièrement bien fréquen-tées par le Merle noir;ce qui, compte tenu de la localisation de ces deux départementsau sud de notre région, est en accord avec les conclusions de FoRMANEK (voir plusbas).
Les 80 reprises d'oiseaux bagués dans les départements rhônalpins appellent uncertain nombre de réflexions (tabl. 5 et fig. 5). Quarante contrôles faits sur les lieuxde naissance ou à quelques kilomètres de celui-ci se répartissent comme suit dans letemps : 14 dans le mois suivant la date de baguage; 10 dans le deuxième et le troi-sième mois; 16 au-delà du troisième mois. Tous les oiæaux appartenant à cette der-nière catégorie (ainsi sans doute que certains autres des deux premières) peuvent êtreconsidérés comme sédentaires, leur présence ayant été relevée au même lieu au coursdes deux grandes périodes ornithologiques gue constituent I'hivernage et la reproduc-tion. La latitude de notre région æ situant sous régime tempéré, il est normal, d'aprèsce que nous avons dit précedemment, qu'une proportion sans doute importante de nosMerles ne déserte pas en hiver æs lieux de reproduction.Les autres contrôles, effectués à distance du lieu de baguage, éclairent quelque peu lesdéplacements auxquels se livrent les populations régionales migratrices de ce Turdidé :
5 jeunes oiseaux bagués en juillet-août (donc sur leur lieu de naissance ou en erratismeà peu de distance de celui-ci) furent repris la même année dans des départements dusud rhônalpin, en octobre, novembre, décembre et février; 12 autres juvéniles, captu-rés les mêmes mois d'été furent retrouvés dans des départements méridionaux depuisle Vaucluse jusqu'à la Corse, d'octobre à mars ; 2 autres sujets immatures bagués aussien été, effectuèrent un déplacement qui les conduisit en Espagne. Ainsi, l'hivernage desMerles rhônalpins semble-t-il essentiellement concentré sur les départements du midiméditerranéen, avec parfois quelques débordements plus au Sud (Corse, péninsule lbé-rique).outre les reprises d'oiseaux bagués en été, nous disposons par ailleurs de 1b fichès serapportant à des individus capturés chez nous en septembre et octobre (donc ne fai-sant pas forcément partie de nos reproducteurs) et également tués sur le pourtour médi-terranéen (au sens large du terme), de l'Espagne à l'ltalie en passant par divers dépar-tements du Sud de la France.
La population d'Europe centrale qui traverse notre région essentiellemedt enautomne hiverne surtout, hormis dans nos départements, dans le Midi de la France

(FORMANEK,n JONES, 1961). D'ailleurs, les cartes de JONES monrrent clairementque seules ces populations d'Europe centrale (principalement de Suisse, d'Allemagneet de Tchécoslovaquie) fréquentent en hiver les départements méditerranéens. Selon cemême auteur, la Camargue est ralliée par deux voies: I'une passant par la vallée duRhône (donc nous intéressant d¡rectement), l'autre atteignant l'ltalie par la traverséedes Alpes à l'Est des départements rhônalpins. Notons pour notre part à ce propos quecontrairement aux cas des Grives litornes et mauvis (cf. plus loin), la région Rhône-Alpes n'enregistre aucune reprise de Merle noir au cours de la même saison que la datede baguage en ltalie.Ouant à [a population de l'Ouest de la Baltique qui se manifeste chez nous principale-ment en décembre, il semble bien qu'elle passe I'hiver à la latitude de notre région, ouqu'elle bifurque par la suite à l'ouest pour se rendre audelà. car les départements duMidi de la France ne sont pas touchés par des oiseaux de cette provenance. Précisonstoutefois que des Merles de la Baltique ont été tués pour certains dans la Drôme etI'Ardèche en décembre et janvier, ce qui tend à prouver leur présence aux portes duMid¡.D'un point de vue plus général, ASHMOLE (1962) montre que I'hivernage des popula-tions nordiques (Norvège, Suède, Finlande) est séparé des zones où les populationsd'Europe centrale se rendent à la mauvaise saison. C'est en effet en Grande-Bretagneque les oiseaux fenno-scandinaves hivernent pour la plupart; alors que ceux de Grande-Bretagne glissent surtout vers l'lrlande. Compte tenu de cela, il n'est pas surprenantqu'aucune reprise rhônalpine soit relative aux lles Britanniques et à la Scandinavie. llest par contre intéressant de noter qu'un oiseau finnois capturé en période de repro-duction ainsi que trois autres bagués dans ce même pays en migration ont été tuéschez nous. On sait que le Merle noir est en expansion vers le Nord, particulièrement enFinlande où il ne nichait que dans l'extrême Sud dans un passé encore récent. SPENCER(1975) prétend que cette colonisation de la Finlande se fait en provenance d'Estonieet/oude Suède. ASHMOLE (1962) pense que le gain de territoires de nidification plusnordiques est rendu possible par un réchauffement récent du climat qui a pour effetde limiter la mortalité hivernale des Merles sédentaires des régions scandinaves, provo-quant ainsi une sorte de surpopulation locale forcée de s'expatrier de ses contrées origi-nelles. Selon le même auteur, leurs habitudes récemment devenues anthropophiles joue-raient également le rôle d'un facteur favorisant au cours de la période hivernale et ex-pliqueraient en partie I'explosion démographique de l'espèce dans certaines contréesnordiques. ouoi qu'il en soit, la présence dans la région Rhône-Alpes, d'individus fin-nois permet de penær que cette nouvelle population de Merles noirs qui, dans un pre-mier temps, s'est rendue pour hiverner dans la moitié Est de la Grande-Bretagne (1),cherche depuis de nouvelles zones d'hivernage, peut-être en raison d'une concurrenceintrasþécifique avec les autres populations hivernant outre Manche, mieux implantéesqu'elle. ll serait intéressant de savoir si des incursions hivernales de Merles finnoisexistent dans d'autres régions de France, dans la moitié Ouest en particulier, et le caséchéant, s'il s'agit d'un fait récent. Les 4 cas rhônalpins ont été obtenus les années sui-
(1)Depuis1958d'aprèsSPENCEB(1975) reprisparSlMMS(1978) qui seréfèreauxreprisesdeba-gues en provenance de Finlande.

10
vantes : 1965, 1972, 1974 et 1979. En l'absence de données supplémentaires, il nousest, pour notre part, impossible d'en tirer des conclusions.
Quelques cas particuliers :
- oiseau bagué le'l 0 janvier 1963 dans l'Ain; repris le 20 octobre suivant prèsde Bergamo (ltalie). Un changement de zone d'hivernage a peut-être eu lieu d,une annéeà l'autre ; à moins qu'étant originaire d'Europe centrale, ce sujet ait emprunté les deuxvoies possibles suggérées par JONES (1961) (voir précédemment) pour gagner le Midiméditerranéen au cours de deux hivers succesifs.
- adulte bagué le 18 juin 1972dans l'lsère; repris le 11 juin 1g74 dans leFinistère. Ce sujet semble s'être radicalement éloigné de son lieu de reproduction ini-tial, ce qui paraît peu banal pour l'espèce.
Ouelques longévités intéressantes :
- oiseau bagué sans précisicÍn d'âge le 10 octobre 1g67 dans la Loire;tué le2 février 1973 en Espagne (donc âgé de 6 ans 1l2 au moins).
- mâle bagué le 4 janvier 1964 en Tchécoslovaquie; tué le 14 novembre 1g71dans l'Ain (à l'âge minimum de I ans 1/2).
Tableau 3 : Périodes de baguage du Merle noir par pays d'origine
Symbolisation des états politiques : cf. tableau 1.
La mentíon * 1 chevauchant les deux quinzaines d'un même mois signifie que seul lemois de baguage (et non la date précise ou à défaut la quinzaine) figire surcertainesfiches.
\msPTYS-\
2 3 4 5 6 7 I 9 to 11 't2 t0It L
/ tNJ¡-tlç .r- l9
CH I 2 I 2 I 4 r0 14l t I 2 50
,n r, t líJ ¡0 0 I tt I 2 4 I 12 2 3 3 5 8 r6 22 I 0 0 2 107/ffi 3 I 5 I t 6 5 I 24 35 2

11
Tableau 4 l Périodes de reprise du Merle noir par département rhônalpin
La mention * 1 chevauchant les deux quinzaines d'un même mois signifie que seul lemois de reprise (et non la date précise ou à défaut la quinzaine) figure sur certainesfiches.
Le total des reprises utilisées est inférieur à l'effectif réel (cf, tableau 1) car certainesdonnées sont absentes ou incomplètes, ce qui rend partiellement inutilisables les fichescorræpondantes.
------\ t0lS0tPAIltrrlls--.-
9 10 11 't2 1 2 3 4 T0Til.rrPml
AIN 01 3 2 I z Ir{AUTESAVOIE 7 I I I 1 I
,|
ISERE 38 2 3 2 29
DROME 26 1 3 4 4 3 {{ I I 25
ARDECHE O? 3. 2 z t4/t5 J 00 { 20 l0 5 l2 il s 5 0 3 I 0 0 0
103/ tnt\ I 2t 25 t? 5 5 0
Tableau 5 : Merles noirs bagués dans la region Rhône'Alpes : départements debaguaç et lieux de reprise
Symbolisation des états politiques : cf. tableau 1.
Les chiffres soulignés d'un point noir correspondent à des contrôles sur place.
LrtllI0tUt
0t
8t0l,lt8t
R HOI{ E.ALPES FRANCE HORS RHONE.ALPES 4èÉ
E
Þ
ô¡Ë
o@ o
rt Cr' lno
ao
ct(o
ôl o
t¡Jtnfo
êat
o
zG Ð
.El!
7-Éto
GUItGoo
t¡,¡olrJ(,gIE
G
a
Ht¡¡
Izq-
À¡t,
:=ê
savorE 73 ! 3 I I 3 2 22
ï0Iil. / utll 0tRIPIISI
t5 5 3 3 i I 5 I 2 I I 2 5 5 ? I I I 3
4 I

12
Figure 2 :
Origine des Merles noirs-Turdus merula- repris dans larégion Rhône-Alpes
- cartes miniaturisées :
A gauche, aire de reproductionnoircie ; les flèches indiquentle sens d'une expansion récentequi n'est pas achevée.
A droite, répartition en hivernageà I'intérieur des gros pointillés ;
les flèches indiquent ici quel'espèce est susceptible d'élargirparfois sa distribution en hiver.
(d'après HEINZEL et al., 1972 ;
PETERSON et a|.,1976 ; ainsique diverses informations plusrécentes obtenues dans la litté-rature consultée : cf. bibliographie).
- grande carte :
Codification utilisée :
Pour faciliter la lecture de cettecarte, nous avons limité à trois signes la représentation des origines de bagues. Cettesymbolisation ne permet donc pas de différencier les reprises ayant eu lieu l'année debaguage de celles obtenues ultérieurement. Toutefois, les cas pour lesquels une telleprécision s'imposait ont été citées au fil du texte.
. un rond noir pour les oiseaux bagués en dehors de la période de reproduction (d'aoûtà avril), donc très souvent à l'écart des lieux de nidification et de naissance;
. un carré noir pour les oiseaux (adultes et juvéniles) bagués pendant la nidification(mai, juin, juillet) :probabilité de reproduction ou de naissance;
. untriangle noir pour les oiseaux bagués au nid (poussins ou adultes couvant).
Les traits fléchés pleins montrent les trajets présumés suivis par les individus apparte-nant aux différentes populations qui fréquentent la région Rhône-Alpes.

13
50
30
20
10
o
30
20
10
o
121110
,e,, J?, J1' 11, ,1,, ,2t, ,3,, ,4,,5 l0 r5 3r r5 30 l5 3t 15 3r r5 2l 15 ll 1530
Figure 3 : Taux de fréquentation mensuels et sem¡-mensuels du Merle noir (en pourcen'
tagesl pour I'ensemble des départements rhônalp¡ns
Certainæ fiches oil figurait une date incomplète n'ont pu être utilisées que pour l'évo-lution mensuelle (et pas au niveau des quinzaìnes).a) Evolution mensuelle (et par quinzaine) globale, soit toutes populations confondues ;
b) Population du centre de l'Europe ;
c) Population nordique.
%
a
2432q2
I4
2L 223
13'T&
165¿r
56
0'1
!_1q
4,9 4,9
3
o r0ts
14
f,* ^.r",r.s su. LE L'EU DE BA.,AGE
* 5 I I oals LA REclor{ RHoNALPINE
Figure 4 : Taux estimés de fréquenta-tion du Merle noir dans chaque dépar-tement rhônalpin en automne-hiver.
Soit, pourcentage par département dunombre de reprises d'oiseaux baguéspar chasseur et par jour de chasse en21 ans.
Figure 5 : Localisation des reprises deMerles noirs bagués dans les départe-ments rhônalpins.
Symboles ronds, carrés et triangulairescf. figure 2. Ces symboles sont figurésici sur læ lieux de reprise alors qu'ilsrenseignent d'après leur forme sur lapériode de baguage.

15
- MEBLE A PLASTRON Turdus torquatus
Cet autre Turdidé (le seul à se montrer entièrement migrateur) est aussi l'uni-que à ne fournir aucune donnée concernant les lieux de naisance des populations quitraversent la région Rhône-Alpes au double passage. Le Merle à plastron est en effetun oiseau peu bagué en Europe en raison de sa niche écologique très spécialisée et desa distribution géographique en conséquence très discontinue.Rappelons que I'espèce peuple d'une part sous la raceTurdus t. torquatus une bonnepartie des lles Britanniques ainsi que la totalité de la Norvège; et que plus au Sud, lasous-espèce Turdus torquatus alpestris s'est confinée après les glaciations aux <systèmesmontagneux d'Europe moyenne et méridionale> (GEROUDET, 1963), soit esentielle-ment les Alpes, le Jura, les Balkans, les Carpathes;mais aussi les Vosges, le MassifCentral, les Apennins, les Pyrénées et la Sierra Cantabrique, avec une couverture moinshomogène. A l'extérieur de l'Europe, une autre population, orientale, s'est fixée entreles rivages de la Mer Noire et de la Caspienne. Mais l'aire globale de répartition de cetteespèce reste la plus restreinte des 6 représentants européens du genre.
Les 16 reprises étrangères obtenues chez nous (fig.6) concernentdes oiseauxbagués pour la plupart à la migration d'automne. Les individus marqués sur l'île d'Hé-ligoland en Mer du Nord provenaient très certainement de la population nicheuse nor-végienne. L'oiseau capturé au mois d'octobre sur la côte Est de l'Angleterre devaitégalement appartenir à la population scandinave car d'après WITHERBY //, ASHMOLE(1962], , c'æt à cette période de l'année que les lles Britanniques sont fréquentées pardes individus de cette provenance (les mois d'août et septembre étant pour leur partréservés aux mouvements de la population insulaire). Plusieurs espèces de Turdidésscandinaves effectuent ainsi un (crochet) par la Grande Bretagne à la migration d'au-tomne. ASHMOLE (1962) qui s'inspire de LACK et SIMMS (1978) qui se réfère àdes travaux ultérieurs de ce même auteur, écrivent que de telles incursions sont la plu-part du temps causées par la force des vents saisonniers soufflant de l'Est qui font dé-river au dæsus de la Mer du Nord des individus voyageant de nuit. Ceux-ci, dès le petitjour, rectifient leur trajectoire vers I'Est eR apercevant la mer qu'ils survolent. Maispour beaucoup d'autres, les côtes britanniques sont atteintes. Le Merle à plastron entredans cette catégorie d'espèces détournées à l'Ouest de leurs directions régulières.Sept autres reprises concernent des sujets capturés en Suisse à la migration post-nuptiale.Mais en l'absence de mention précisant la sous-espèce à laquelle appartenait chacun decæ oiseaux, il s'avère impossible de dire s'il s'agisait de locaux ou de migrateurs nordi-ques (les deux formes fréquentant les cols alpins : DE CROUSAZ,1960l.
Le tableau 6 nous enseigne que le passage d'automne sur l'lle d'Héligolands'opère en septembre-octobre; il en est d'ailleurs de même en Suisse, ce qu¡ est confir-mé par les données de l'Atlas des Oiseaux Nicheurs de ce pays (SCHIFFERLI et al.,1980).

16
En ce qui concerne la lat¡tude rhônalpine (tabl. 7 et fig. 7),les repriæs s'éta-lent de septembre à la première quinzaine de décembre (encore 3, soit 19 % en cedernier mois de l'année, il èst vrai dans deux départements du Sud, où les tentativesd'hivernage sont peut-être plus fréquentes que ne le précise l'Atlas des Oiseaux NicheursRhônalpins). Le <maximum> de la migration post-nuptiale paraît atteint en octobreavec 7 reprises (44 o/o de l'ensemble) ; résultats avancés sous toutes réserves, en regarddu nombre restreint des données sur lesquelles nous fondons cette supposition.
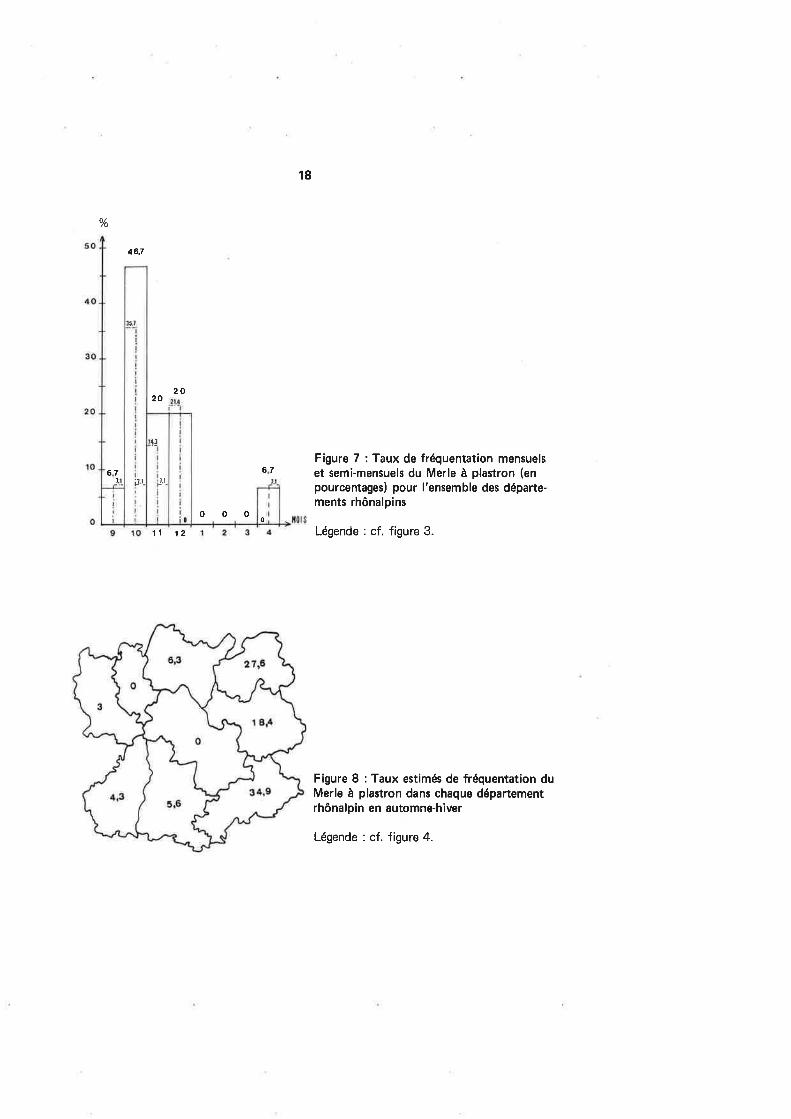
La fréquentation des départements rhônalpins par le Merle à plastron n'est pasdu tout homogène : il apparaît clairement sur la figure I que les trois départementsalpins (les deux Savoie et les Hautes-Alpes) concentrent une part prépondérante desmigrateurs (au total plus de 80 % des reprises), d'ailleurs à la fois en provenance deSuisse et d'Héligoland. Les tests d'indice de tendance appliquée dans un premier tempsau nombre de chasseurs par département puis à la localisation géographique, apportentune très nette confirmation. De toute évidence, la migration du Merle à plastron sefait de façon préférentielle le long de l'Arc alpin depuis le Nord jusqu'au Sud (en
dépit de la présence des reliefs du Massif Central qui longent l'autre rive du Rhône, àl'Ouest), L'éclatement multidirectionnel des populations migratrices doit donc s'opérertrès bas, aux derniers contreforts annonciateurs de la Méditerranée. Cette déductionest à rapprocher d'une remarque formulée par DROST /n JONES (1961), qui écrivaitque le Mede à plastron restait tributaire des reliefs au cours de æs migrations.
L'absence de reprises régionales au delà de décembre est en accord avec lesdonnées fournies par l'observation (espèce rare en janvier d'après l'Atlas Rhône-Alpes).L'hivernage d'un bout à l'autre de la saison est donc tout au plus accidentel chez nous.
On sait que les quartiers d'hiver les plus importants du Merle à plastron setrouvent, outre dans l'Est et le Centre de la Péninsule lbérique, sur les sommets d'Afriquedu Nord : Monts d'Oranie, Moyen-Atlas, Rif, Atlas tellien oùl les oiseaux arriveraient sur-tout en masse au milieu de l'hiver, à la suite de vagues de froid dans le Sud de l'Europe(ASHMOLE, 1962). Les départements méditerranéens seraient fréquentés par les popu-lations scandinaves; alors que les nicheurs britanniques (8 000 à 16 000 couples d'aprèsSHARROCK, 1975) hiverneraient plutôt en Espagne, via l'Ouest de la France (DURMAN,15761.Ainsi, d'une façon générale, l'orientation de la migration de ce Turdidé est-elle forte-ment accentuée plein Sud ; ce caractère ayant déjà été m¡s en évidence par DROST/n JONES (1961). En conséquence, une double raison semble pousser le Merle à plas-tron à emprunter le Massif alpin sur toute sa longueur: d'une part sa dépendance del'altitude, d'autre part l'axe général de ses déplacements automnaux.L'ltalie accueille également quetques hivernants (PETERSON et al., 1976], comme cefut sans doute le cas pour I'individu qui, bagué au col de la Golèze (Savoie) en sep-tembre 1974, fut tué près de Chieti en mars 1979 (fig. I et tabl. 8) ; à moins que cetoiseau ait opté pour la voie orientale Cap Bon - Sicile - ltalie au moment de regagnerl'Europe après un séjour dans le Nord tunisien. Citons encore le cas de cet autre oiseau

17
bagué en ltalie du Nord le 11 septembre 1972 et tué dans la Drôme le 7 décembre del'année suivante, qui fournit l'indice d'éventuels changements de lieux d'hivernage (oude passage) d'une année à l'autre.
Figure 6 : Origine des Merles à plastron -Turdus torquatus- repris dans la région Rhône-. Alpes.
Lfuende : cf . ligure 2.Sur la carte miniaturisée montrant l'aire de nidif ication,le pointillé figure la séparationentre les deux races géographiques. Sur la grande carte, les iignes fléchées pointillées sontporteuses d'une incertitude quant à l'origine réelle de l'individu bagué dans les lles britan-nrques.
o/o
46:l
67671.
5.
i7l
2020
{¡
1,1
ooo0
18
12tl
Figure 7 : Taux de fréquentation mensuelset semi-mensuels du Merle à plastron (enpourcentagesl pour l'ensemble des départe-ments rhônalpins
Légende : cf. figure 3.
Figure 8 : Taux estimés de fréquentation duMerle à plastron dans chaque départementrhônalpin en automne-h¡ver
Légende : cf. figure 4.

19
Figuré 9 : Localités de reprise des Merles à plastron bagués dans les départementsrhônalpins
Légende des symboles : cf. figuræ 2 et 5.
Tableau 6 : Périodes de baguage du Merle à plastron par pays d'origine
Légende : cf. tableaux 1 et 3.
\r¡lOlSPAYS \\
1 2 3 4 5 6 7 I 9 10 12 l0ltt/ 0rv(
/ ts.l 0 0 00 0 0 0 0 2 0 0 0 0 0 2 0 2 5 2 0 0 00 16- /tî 0 0 0 0 2 0 0 2 5 I 0
20
Tableau 7 : Périodes de reprise du Merle à plastron par département rhônalpin
Légende : cf. tableaux 3 et 4.
9 10 11 12 1 2 3 4 l0TÀtII PARI
42
74 2 3
73 I 2
oõ 3 I 4
rñr¡r '/ 1s J 0 5 2 3 0 0 0 0 0 0 0 0 I 15rn s 3 3 0 0 0 I
Légende
Tableau 8 : Merles à plastron bagués dans la région Rhône-Alpes :
départements de baguage et lieux de reprise
: cf. tableau 1.
r. t t lJI
0tIPflI$
ABlJAEI
e¡é.CÊr<c¡
{

21
- GRIVE MUSICIENNE lurdus philomelos
Bien que moins commun que le Merle noir (3 500 000 couples nicheurs dansles lles Britanniques, par exemple : SHARRocK, 1g7b), cet autre représentant dugenre Turdus a néanmoins une distribution géographique englobant une grande partiede l'Europe, hormis quelques contrées nordiques d'u.R.s.s. et de scandinavie ainsi quela moitié Sud de la Péninsule lbérique et certaines parties de la frange côtière méditer-ranéenne de France, d'ltalie et des Balkans. Plus à l'Est, existe une autre populationentre la Mer Noire et la Caspienne et des Musiciennes atteignent même le 110e degréde longitude aux confins de la Mongolie et de la Chine.
139 reprises étrangères illustrent la présence de cette Grive dans notre région.La figure 10 montre que le secteur RhôneAlpes reçoit des populations aux originestrès semblables au Merle noir. Nos départements sont non seulement concernés par desindividus d'Europe centrale (qui forment plus de la moitié des données), mais égalementpar des oiæaux du Nord-Est de l'Europe. ll s'agit des points figurant sur notre carte(fig. 10) de chaque côté de la Baltique oùr les oiseaux ont été capturés à la migrationd'automne. Leur origine présumée doit se situer en Finlande et à l'extrême Nord de laRussie. Notons qu'aucune reprise ne concerne la population de Norvège, ni celle deSuàle septentrionale dont ASHMOLE (1962) précise qu'elles hivernent principalementdans le sud-ouest de la France et en Espagne, en suivant donc une voie qui ne passepas par la région Rhône-Alpes. Par contre, les départements rhônalpins accueillent vrai-semblablement quelques sujets britanniques bien que nous manquions de preuvesréelles (baguage au nid) pour confirmer cette origine [1]. Les oiseaux bagués pouss¡nsle furent en effet au Sud de la Finlande (2 cas), en Pologne (3), Allemagne de l'Est (1),Allemagne de l'ouest (4), Tchécoslovaquie (2) et suisse (2). Les aurres indices de re-production ou de naisance (oiseaux, rappelons-le, marqués en mai, juin, juillet) concer-nent les lles Britanniques (2 cas). la Belgique (1), à nouveau la Pologne (2), la Tdrécos-lovaquie (4), l'Allemagne de l'Est (1), l'Allemagne de l'Ouest (b) et la Suisse (6).
La migration à travers I'Europe semble pouvoir se résumer de la sorte (tabl.9) :
5 oiæaux finnois ont été bagués antérieurement au 30 septembre, signe d'une désertionprécoce des contrées nordiques. Le Sud de la Suede est traversé en septembre et surtoutoctobre, dates en accord avec celles données par ASHMoLE (1962) ; le Danemark à lamême période, ainsi que l'Allemagne de l'Ouest (dans ces trois pays, c'est la premièrequinzaine d'octobre qui est la plus marquée par les passages : 11 cas sur les 1b de l'en-semble du mois). La Suisse confirme cette tendance avec 16 captures comprises entrele 1er et le 15 octobre (contre 5 dans la æconde partie du mois), mais déjà 10 au cours

22
des quinze derniers jours de septembre. Ce type de schéma migratoire est confirmé parle déroulement des passages au col alpin de Bretolet (DE CROUSAZ, 1960l' ainsi quepar les données de l'Atlas des Oiseaux Nicheurs de Suisse (SCHIFFERLI etal., 1980).
C'est donc sans surprise que l'on note sur le tableau 10 et la figure 11 que lesGrives musiciennes arrivent massivement dans la région Rhône-Alpes dès le début d'oc-tobre et que le gros passage se poursuit jusqu'au 15 du mois qui lui succàCe, avec unrecord de 27 reprises dans les 15 premíers jours de novembre. Par la suite, leurs effec-tifs s'amoindrissent et les reprises hivernales n'atteignent jamais la dizaine par tranchede quinze jours. Les 9 recaptures de début janvier doivent correspondre à un petit ar-rivage tardif d'oiseaux découragés par le froid dans leurs tentatives d'hivernage plus auNord (rappelons à cet égard que chez le Merle noir, l'amorce d'une remontée des effec-tifs régionaux à l'issue d'un creux similaire se manifestait dès le début de décembre,soit un mois plus tôt, et concernait une majorité d'individus appartenant aux popula-tions du Nord de l'Europe). Chez la Grive musicienne, contrairement à la situationdécrite pour le Merle, il ne semble pas que les individus d'Europe du Centre migrentplus töt que les populations du Nord de I'Europe;en effet, les deux populations fré-quentent la région Rhône-Alpes en proportions quasiment équivalentes pendant lesdeux mois de pointe const¡tués par octobre et novembre. Au cours de la petite (pous-sée> de janvier cependant, il semblerait que les oiseaux bagués en Europe centrale(Suisse, Allemagne, Tchécoslovaquie) figurent davantage que les sujets igsus de contréesplus nordiques, ce qui tend à s'écarter de notre remarque sur le Merle noir. Toutefois,nos résultats restent en accord avec les conclusions plus générales tirées par d'autresauteurs sur le cas de la Grive musicienne. SIMMS (1978) parle de mouvements liés aufroid en cours d'hiver; ASHMOLE (1962) précise que ces déplacements sont rares àHéligoland, Fair lsle et au Danemark mais plus sensibles en d'autres lieux (lrlande parexemple). Le même auteur finit par écrire que globalement, les mouvements hivernauxde Musiciennes sont moins fr@uents que ceux d'autres espèces de Turdidés.Comme chez le.Merle noir, la fermeture de la chasæ intervient avant la désertion de larégion Rhône-Alpes par les migrateurs et les hivernants étrangers. ll est donc impossible,par reprises de bagues interposées, de préciser selon quelles modalités s'opèrent les mou-vements de fin d'hivernage. Pourtant, un individu bagué le 3 octobre 1968 à Héligoland,fut trouvé encore présent dans la Loire le 19 avril suivant.
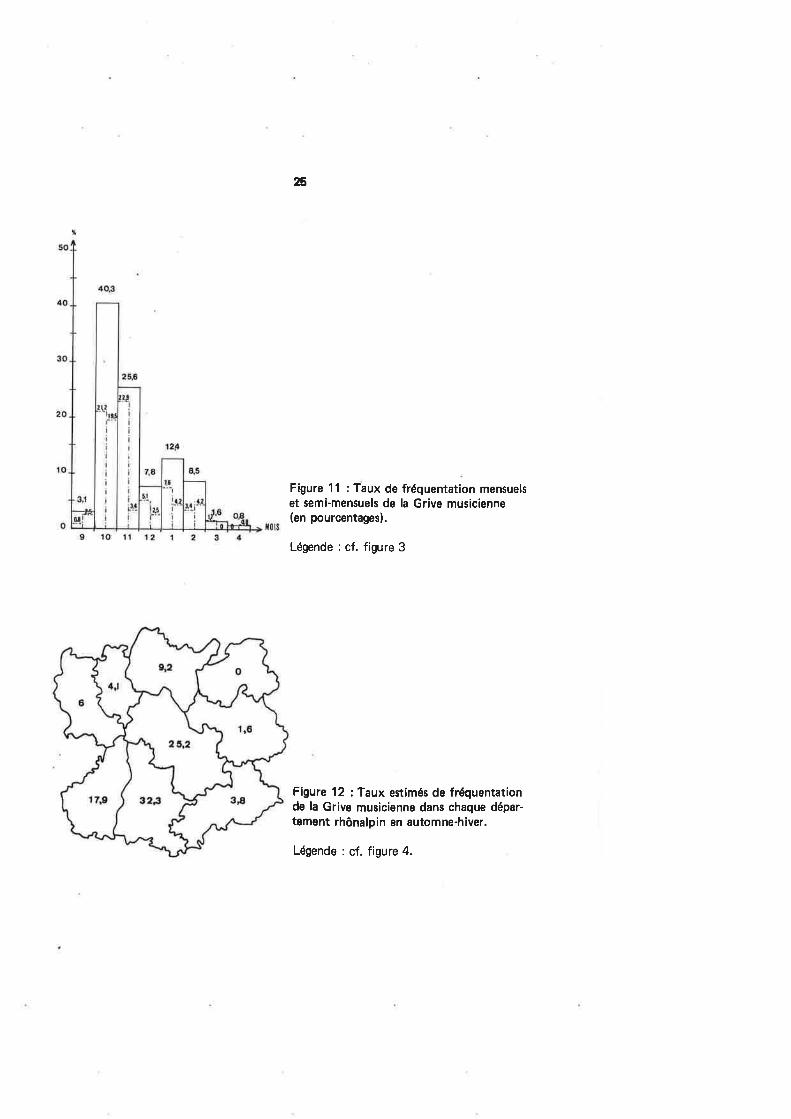
La figure 12 hisse, comme pour le Merle noir, l'Ardèche et surtout la Drômeen tête des départements largement fréquentés par la Musicienne. Les tests non par¿F
métriques utilisés confirment cette tendance. ll est donc clair que I'hivernage de l'es-pèce existe dans les deux départements les plus méridionaux de notre région, qui sem-blent constituer en cela la limite septentrionale de sa présence régulière au cours de lamauvaise saison.
Car de façon générale, les apparitions hivernales de cette Grive au Nord desdeux départements précédents sont rares ou du moins épisodiques, comme l'observa-tion le confirme. ll en est d'ailleurs de même dans les pays européens traversés à la

23
migration d'automne (tabl. 9) : novembre n'enregistre qu'une reprise en Allemagne del'Est, décembre aucune, alors que janvier constitue le mois de baguage d'un unique in-dividu en Allemagne de l'Ouæt. Ces deux reprises hivernales confirment les écrits deGEROUDET (1963) : <la tendance à demeurer en hiver sur les lieux de reproductionse manifeste déjà en Allemagne>. Mais il doit s'agir pour l'Europe centrale, de cas ex-ceptionnels, liés à des conditions hivernales (nourriture et climat) particulièrement fa-vorables certaines années. En Angleterre, Belgique et Hollande par contre, beaucoupd'oiseaux sont sédentaires à la faveur de températures hivernales modérées par I'in-fluence atlantique. c'est ainsi que LACK,n ASHMoLE (1962) prétend qu'en Grande-Bretagne, æuls 48 % des adultes el il o/o des juvéniles se livrent à des déplacementsqui les conduisent surtout en lrlande et dans l'Ouest français, mais aussi en Espagne eten Afrique du Nord. Cette double raison (proportion importante d'oiseaux sélentairesmais surtout quartiers d'hiver situés très à l'Ouest) explique la rareté des reprises d'ori-gine britannique dans notre région et leur absence totale en France méditerranéenne(JoNES, 1961). Les cartes dressées par cet auteur montrent que les populations d'Eu-rope centrale et de Finlande aboutissent dans le Midi de la France, ce qui reflète unesituation très proche de celle décrite pour la région Rhône-Alpes.
Les points jalonnant le bassin méditerranéen sur les figures 10 et 13 nous dési-gnent la France méditerranéenne, l'Espagne, l'ltalie et certaines îles (Baléares, Sardaigne)comme lieux d'hivernage des populations qui transitent par notre région.selon ASHMoLE (1962), les nicheurs germaniques (donc appartenant à la populationd'Europe centrale) hiverneraient principalement dans l'Ouest de la France et en Espagne,ce qui n'apparaît ni sur notre figure 13, ni dans les résultats de JoNES (1g61) dont iacarte montre que la présence des Musiciennes allemandes dans les départements méd¡-terranéens est bien fournie. Précisons cependant que JoNES (1961) présume que cer-taines Grives musiciennes migrent depuis le Midi de la France jusqu'à la côte orientaled'Espagne en traversant la Méditerranée via les Baléares. ll æ peut donc que des GrivesMusiciennes germaniques empruntent cette voie pour rejoindre leurs quartiers d'hiveribériques.L'autre population présente en automne dans la région Rhône-Alpes, celle du Nord-Estde l'Europe, baguée en début de saison sur les pourtours de la Baltique hiverneraitælon ASHMOLE (1962) en deux clans séparés : I'un sirué à l,Ouest du Rhône juqu,enEspagne ; l'autre à l'opposé, en ltalie. Bornons-nous à noter pour notre part que cesdeux répartitions existent sur nos figuræ 10 et 13.Les quartiers d'hiver des Musiciennes rhônalpines nicheuæs restent énigmatiques. Nousne disposons que d'une seule reprise hivernale pouvant correspondre à un reproducteurde notre région. ll s'agit d'un oiseau bagué dans l'Ain le 20 juillet igTo et tué cinqmois plus tard en ltalie (Salerne) (tabl. 1l), ce qui constitue malgré tout un indice digned'intérêt, bien qu'en désaccord avec l'hypothèæ selon laquelle les populations du Norddépassent en hiver celles des latitudes inférieures, selon le processus du <leap-frog> (lit-téralement, saut de grenouille), décrit par SALOMONSEN ,n ASHMOLE (1g62). Maisun seul cas ne peut évidemment pas donner lieu à des conclusions hâtives.

24
Ouelques longévités intéressantes :
- outre 3 cas de 6 ans, un individu capturé (adulte ? ) en Suisse le 20 juillet1967, fut tué en Ardeche le 8 novembre 1974 (donc à 7 ans 1/2 au moins).
- de même, un sujet bagué au nid en Tchécoslovaquie le 29 mai 1960, fut tuéà l'automne 1968 dans le département de l'Ain (soit âgé de 8 ans 1/2).
Figure 10 : Origine des Grives musiciennes -Turdus philomelos- reprises dans la régionRhône-Alpes.
Légende : cf. figure 2.
_ al cA! tout ufE LA
¡ur¡aa
I : a c^t o¡0 ue¡rt DE !r
25
Figure 11 : Taux de fréquentation mensuelset semi-mensuels de la Grive musicienne(en pourcentages).
Légende : cf. figure 3
Figure 12 : Taux estimés de fréquentationde la Grive musicienne dans chaque dépar-tement rhônalpin en automne-hiver.
Légende : cf. figure 4.

26
+ 1 REPRTSE SUR LE LIEU DE BAGUAGE
Figure 13 : Localités de reprise desGrives musiciennes baguées dans les
départements rhônalpins.
Légende dæ symboles : cf. figures2er5.
Tableau 9 : Périodes de baguage de la Grive musicienne par pays d'origine
Légende : cf. tableaux 1 et 3.
Le total des reprises utilisées est inférieur à l'effectif réel (cf. tableau 1), une fichen'ayant pu être utilisée en raison de æs coordonnées de baguage incomplètes.
\--¡OfS
'AYS \.2 3 4 5 6 7 I I 10 11 12 I0rlt
/rrv(
CH ¡ I I ¡ I t¡ 65 5o
/ sl I 0 0 a 2 5 4 a 6 5 ! t 5 il I 2t I 0 0 r39/l¡ I I 0 I ¡l aa 2 0

27
Tableau 10 : Périodes de reprise de la Grive musicienne par département rhônalpin
Légende: cf. tableaux 3 et 4.
I 'to 11 t2 1 2 3 4 T0ilL
38 I l3 I I 12
26 I t2 2 4 1 2 2 50
o7 I I 3 5 I 2 tl/ l5 ,l 3 25 23 27 a ô 3 I 5 I 5 2 0 0 I r29/ rntr 4 52 33 t0 t8 tl 2 I
Tableau 11 : Grives music¡ennes baguées dans la région Rhône'Alpes :
Départements de báguage et lieux de reprise
Légende : cf. tableaux 1 et 5.
R.A ;RANCE HORSN EfRAIOtn Iç (o
Âr
t¡¡.Dt(J)
¡t;!nt
{¡
t¡¡2oÉ
3
ct,,{Iu,
Ë
o.D
-o
EË
Þfl-a
o1 1 1 '| I 7
71 I I I I I I I t1
foTil, zuilt 0trrmtst
I 1 I 2 I 5 I2
2 E ll

28
- GRIVE DRAINE Turdus viæivorus
La distribution géographique de cette quatrième espèce recouvre la presque
totalité de l'Europe, à l'exclusion d'une surface importante de la Norvège, du Nord de
la Finlande et de la presqu'île de Kola. Elle se montre plus thermophile que la Musi'cienne puisque l'ensemble du bassin méditerranéen (Afrique du Nord y compris) est
concerné par sa présence estivale. Côté oriental en outre, deux fronts disjoints attei-gnent respectivement le Nord de l'lnde et l'Est de la Mongolie.Pourtant, en dépit d'une aire de reproduction aussi vaste, la Grive draine ne s'illustreque par 12 reprises (fig. 14), soit moins que le Merle à plastron à la répartition très
fragmentée. Cela suppose un effectif d'oiseaux marqués très inférieur aux autres espèces
de Turdidés. Plusieurs explications peuvent être invoquées à ce sujet : une première
réside dans la méfiance de cet oiseau qui fit écrire à GEROUDET (1963) que <la
Draine est la plus farouche de nos Grives; toujours sur æs gardes, prompte à la fuite>,donc de capture plus difficile. Mais il nous semble que ce seul argument ne suffit pas
à expliquer cette rareté des données, d'ailleurs générale à l'échelle de l'Europe (ASH-
MOLE, 1962). ll est probable aussi que la densité.de son peuplement n'est pas aussi
forte que celle d'autræ représentants du même genre; même si cette caractéristique
n'apparaît pas dans l'Atlas des Oiseaux Nicheurs de France (YEATMAN, 1976) qui ladonne <abondante> au même titre que la Grive musicienne. ll nous suffira de rappelerque cette clase d'abondance englobe des effectifs compris entre 100 000 et 1 000 000de couples pour montrer que notre supposition n'est pas forcément en désaccord avec
les chiffres de cet auteur; l'une (la Draine) pouvant se situer à la limite inférieure de
cette estimation, l'autre (la Musicienne) à l'extrémité inverse. Une telle hypothèæ se
trouve confortée par les décomptes plus affinés de l'Atlas britannique (SHARBOCK,
1975) qui avancent l'effectif de 3 500 OO0 couples de Grives Musiciennes, pour seule-
ment 300 à 600 000 couples de draines (il est vrai sous climat océanique)'
Malgré cette rareté des reprises, l'origine des Draines qui frequentent la région
Rhône-Alpes à la mauvaiæ saison semble relativement bien cernée par deux cas de ba'guages au nid (l'un en Suisse, l'autre en Allemagne orientale) et un troisième cas de
capture en période nuptiale dans le second de ces pays (fig. 1a). C'est en effet dans le
Centre de la France que les individus germaniques sont présumés hiverner (ASHMOLE,
1962). Ces trois cas paraissent assez significatifs à cet égard.
Le passage d'automne doit s'opérer en Suisse (col de Bretolet en particulier)en septembreoctobre.(tabl. 12) ; période également mentionnée pour la migration de
l'espèce dans l'Atlas des Oiseaux Nicheurs de ce pays (SCHIFFERLI et a|.,1980). Nospropres données ne permettent pas d'en ajouter plus sur le déroulement de la migrationen Europe.ASHMOLE (1962) s'inspirant de HAARTMAN & BERGMAN et de ULFSTRAND, pré-
cise que la Scandinavie est désertée dans la seconde moitié de æptembre et en octobre;alors que le Danemark est traversé fin octobre et début novembre (HANSEN /n ASH-MOLE, 19621 , ce qui fait apparaître un décalage par rapport aux dates données par

29
LIPPENS & wILLE ij9721 pour la Belgique. Selon ces deux auteurs, les passages se-raient en effet marqués en septembre et surtout octobre. Mais il peut s,agir deleuxpopulations différentes pour lesquelles læ périodes de pointe de leur migiation ne sontéventuellement pas les mêmes.
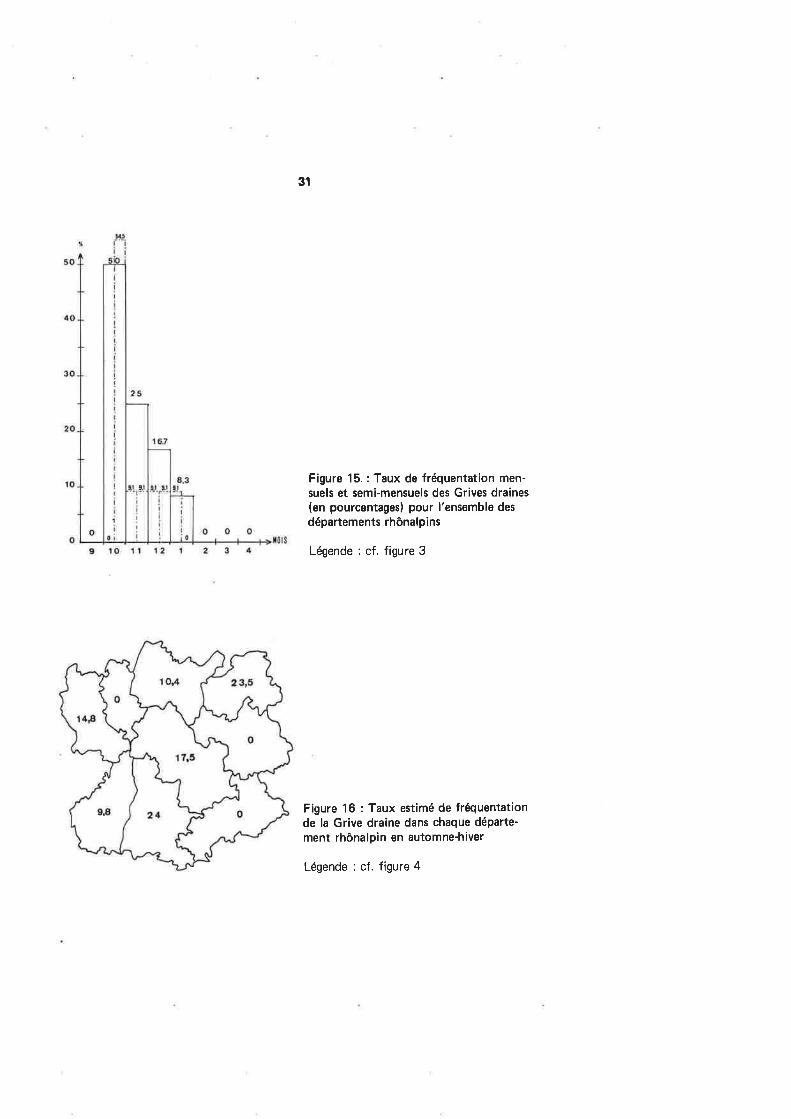
Les dates de reprises rhônalpines (tabl. 13 et fig. l5) s,échelonnent en nombredécroissant entre la mi-octobre et la mi-janvier. La seconde quinzaine d'octobre æmblecorræpondre au maximum d'affluence (50 % des reprises), remarque confirmant l.Atlasdes Oiseaux Nicheurs Rhônalpins. Cependant, des mouvements précoces s'effectuent dèsla dernière semaine d'août (obærvations personnelles dans les lr4onts du Forez (Loire)).Comme pour d'autres espèces de Turdidés (Merle noir en particulier), une période char-nière d'erratisme diffus d'oiseaux certainement locaux, doit avoir lieu en fin d'été avantle déclenchement de la migration proprement dite. Toutefois, des oiseaux belges auraientété repris dès le mois d'août en France (départements ? ) (LlppENS & WtLLE,1g72l.
Nul besoin de préciser que les douze reprises dont nous disposons ne permet-tent pas de montrer de différences de fréquentation des départements régionaux (fig.16).
Au cæur de la saison hivernale, la Grive draine est encore présente, car ellefait souvent preuve d'un tempérament sédentaire ou æ livre à des mouvements autom-naux de faible amplitude, un peu comparables à une transhumance. MAyAUD rnJONES (1961), cite des déplacements saisonniers aboutissant en Camargue à'partir desreliefs du Sud de la chaîne des Alpes,D'autres populations plus migratrices viennent se joindre aux hivernants locaux (voirplus haut le cas des nicheurs d'Allemagne). Ouoi qu,il en soit, la plupart du temps, laGrive draine s'en tient à des performances kilométriques plus modestes que les autresespèces du genre. cependant, des oiseaux tchèques, belges et norvégiens ont atte¡ntl'Espagne (JoNES, 1961). Mais ce même auteur déclare que l'hiverñage des populationsmigratrices est réduit au Sud des frontières françaises. ASHMOLE fi902) va plus loinpour sa part, en écrivant que les incursions en Espagne se limitent à un petit nombred'hivers, soulignant par là le caractère relativement exceptionnel de cette destination.Sans doute moins frileuse que les autres Grives et Merles, elle ne réagit généralementpas aux vagues de froid en cours d'hiver mais se maintient dans les sites qu'elle a choi-sis à I'automne (ASHMOLE, 19621. Cette particularité comportementale vis-à-vis desintempéries pourrait être en relation avec sa taille quí l'expoæ éventuellement moinsque d'autres especes de Turdidés à une déperdition calorifique (hypothèse suggérée parPh. LEBRETON (comm. pers.)). Toutefois, des déplacements en janvier et féviier sont,à titre d'exception, mentionnés par ASHMOLE (1962) pour l,lrlande.
Parmi les rares oiseaux bagués dans la région Rhône-Alpes, deux ont donnélieu à une reprise ultérieure (tabl. 14 et fig. 17) :
- une Draine capturée à Saint-Chamond (Loire) en décembre ,lg70, a été re-
trouvée à 15 km deux hivers plus tard :fidélité au lieu d'hivernage ou cas de sédenta-r¡té ?

30
- cette seconde alternative se vérifie chez un second sujet bagué en juin 1973dans le Pilat (Loire) à l'altitude de 1 100 m, et tiré sur la même commune en décembre1974. Ce cas particulier introduit donc quelques réserves dans le phénomène de trans-humance dont nous avons fait état un peu plus haut. ll prouve que certains oiseaux pas-sent l'hiver dans des localités montagneuses, où il paraît cependant bien difficile decroire qu'ils puissent rester en toutes circonstances quel que soit le degré d'enneigement.
Figure 14 des Grives draines -Turdus viscivorus- repriæs dans la région Rhône-
Légende : cf. ligure 2.
: OrigineAlpes
31
Figure 15.: Taux de fréquentation men-suels et semi-mensuels des Grives draines(en pourcentagesl pour l'ensemble des
départements rhônalpins
Légende : cf. figure 3
Figure 16 : Taux estimé de fréquentationde la Grive draine dans chaque départe-ment rhônalpin en automne-hiver
Légende : cf, figure 4

32
f z neenrsEs suR LE LIEU DE BAGUAGE
Figure 17 : Localités de reprise des Grives draines baguées dans les départementsrhônalpins
Légende des symboles : cf. f igures 2 er.5.
Tableau 12 : Périodes de baguage de la Grive draine par pays d,origine
Légende : cf. tableau 1.
\-- nOlSPIYS---\
1 2 3 4 5 6 7 I 9 10 t'l 12 TOIII/9 AYJ
s,/tsJ 0 o 00 0 0 0 0 1 0 I o 0 0 0 o 3 4 2 0 0 00 12/ lÍtl 0 0 0 0 t I I o 3 6 0 0

33
Tableau 13 : Périodes de reprise de la Grive draine par département rhônalpin
Légende : cf. tableaux 3 et 4.
9 10 11 12 2 3 4 IOTAI.
69 0
74 I t
o5 0
38 2 I 3
26 1 4
,nrrt / l5 J 0 0 0 I ll 0 0 0 0 0 0 0 12/ tnß. 6 2
Tableau 14 : Grives draines baguées dans la région Rhône-Alpes : départementsde baguage et lieux de reprise
Légende : cf. tableau 5.
Utr.rtlJI
OE
DE tPR tsr
sIOlJAOE
ñtç
CE<
êãé<É<€
42 2 2
l0T,.lt iltl,| 0E
RTPRISI
22
2

34
- GBIVE LITORNE Turdus pilaris
La <conquête de l'Ouest> à laquelle se livre cette Grive depuis le XlXe siècleest, avec le cas de la Tourterelle turque, un exemple classique de colonisation de mi-lieux inoccupés de la part d'espèces vraisemblablement au paroxysme de leur vitalitébiologique.La distribution actuelle de la Grive litorne couvre l'ensemble de l'Europe du Nord et duCentre, ainsi que la presque total¡té de la Russie occidentale. La limite sudiste euro-péenne fluctue grosso modo d'Est en Ouest entre le 48e et le 45e degrés de latltude N.Elle gagnerait d'ailleurs du terrain plus au Sud, à en croire MUNTEANU /n YEATMAN(1971l', au Nord de la Roumanie en particulier. Côté oriental, un large front atteintl'extrémité Est de la Mongolie. A l'ouest, les lles Britanniques ont été atteintes en1967 (SHARROCK, 1975\; de mêmeque la Belgique la même année (yEATMAN,1971). Paradoxalement, des contrées beaucoup plus lointaines ont été peuplées anté-rieurement: ainsi, l'lslande vers 1930 et le Groendland en 1g41 or) la population au-rait été décimée depuis, au cours d'un hiver rigoureux (SALOMONSEN /n YEATMAN,191 11.
A l'échelle de la France, le Massif central qui, toutes proportions gardées, est aussi detype <insulaire> (LEBRETON, 1975) [1 ], constitue à son tour l'un des tour récentspostes avancés de l'espèce (cocHET, 1977 er 1980), preuve de velléités expansionnis-tes croissantes. Cette colonisation à l'Ouest du Rhône a sans doute été facilitée parles nombreuses vallées orientées Est-Ouest qui constituent des couloirs naturels s'ou-vrant sur le Massif Central. On sait en effet que la Grive litorne utiliæ le tracé des coursd'eau pour appuyer son extension (YEATMAN, 1971).
L'origine des hivernants rhônalpins (fig. 18) est cernée de façon satisfaisantegrâce aux 60 reprises étrangères d'oiseaux bagués poussins (sur un total de 1b4 recap-tures). Avec 17 cas de naissance, la Finlande se hisse en tête de file des patries desGrives litornes passant par notre région. Les pays qui lui sont frontaliers sont égalementbien repréæntés : Norvège (3 cas), Suède (4), U.R.S.S. (3) ; ainsi que la pologne et laTchécoslovaquie (2 cas chacune), les deux Allemagne (15 cas en tout) et notre voisinla suisse (141. 18 autres captures estivales réparties dans les mêmes pays que les 60précédentes confortent ces données. Ainsi, notre région semble-t-elle fréquentée régu-lièrement [2]par deux populations distinctes : l'une nichant en Finlande (et dans unemoindre mesure en Scandinavie) ; l'autre, en Europe centrale au sens large du terme,c'est-à-dire de la Pologne à la Suisse. ll est cur¡eux de noter sur notre carte que lesrives de la Baltique (à l'exception de la partie qui borde les golfes de Finlande et deBotnie) sont très peu jalonnées par des points.Les Grives litornes reprises en France méditerranéenne proviennent des mêmes popula-tions que les nôtres (JONES, 1961) mais les oiseaux tchàques sont peu nombreux
['l ] <eien que réduit à vol d'oiseau, le fossé climatique du couloir rhôdanien fut et demeure suff isantcomme barrière biogéographique pour plusieurs oiseaux>,
[2]Voir plus loin les cas de reprise en provenance de l'Est.

35
semble-t-il au Sud de chez nous, de même que les nicheurs helvétiques ; alors que les oi-seaux originaires de Pologne ne sont apparemment pas du tout représentés.
La migration des Grives litornes, au vu du tableau 'l 5, est difficile à isoler de lapériode de reproduction dans les pays nordiques où celle-ci existe; les captures d'automnen'étant pas significativement différentes de celles d'été (d'autant que dans le Sud de laSuède et de la Norvège, des populations sont relativement sédentaires (ASHMOLE, 1962)).Ce même auteur, qui reprend HAARTMAN & BERGMAN et ULFSTRAND, signale desrassemblements prémigratoires dès le milieu de l'été ; mais c'est seulement vers la fin deseptembre que ces groupes nordiques s'animent. Au Danemark et en Belgique, nos donnéesmontrent que le passage est perceptible dès la seconde quinzaine de septembre, se poursui-vant jusqu'au 15 novembre, avec un maximum, dans le deuxième de ces pays, compris entrele i6 octobre et le 15 du mois suivant (9 captures sur un total de 12l.. Ces dates sont glo-balement en accord avec les travaux de WEIGOLD et HANSEN, /n ASHMOLE (1962).
Cependant, ces auteurssituent le premier jalon de la migration un mois plus tard, tandisque LIPPENS & WILLE (1972\ englobent le mois de décembre dans la période où s'ob-serve le maximum de passages en Belgique. Les lles britanniques seraient pour leur parttraversées massivement en octobre et dans les trois premières semaines de novembre(ASHMOLE, 1962). En Suisse, la période de pointe æ situe (sur notre tableau du moins)la seconde quinzaine de septembre, ce qui doit correspondre à des mouvements plus oumoins locaux d'erratisme (juvénile ? ).
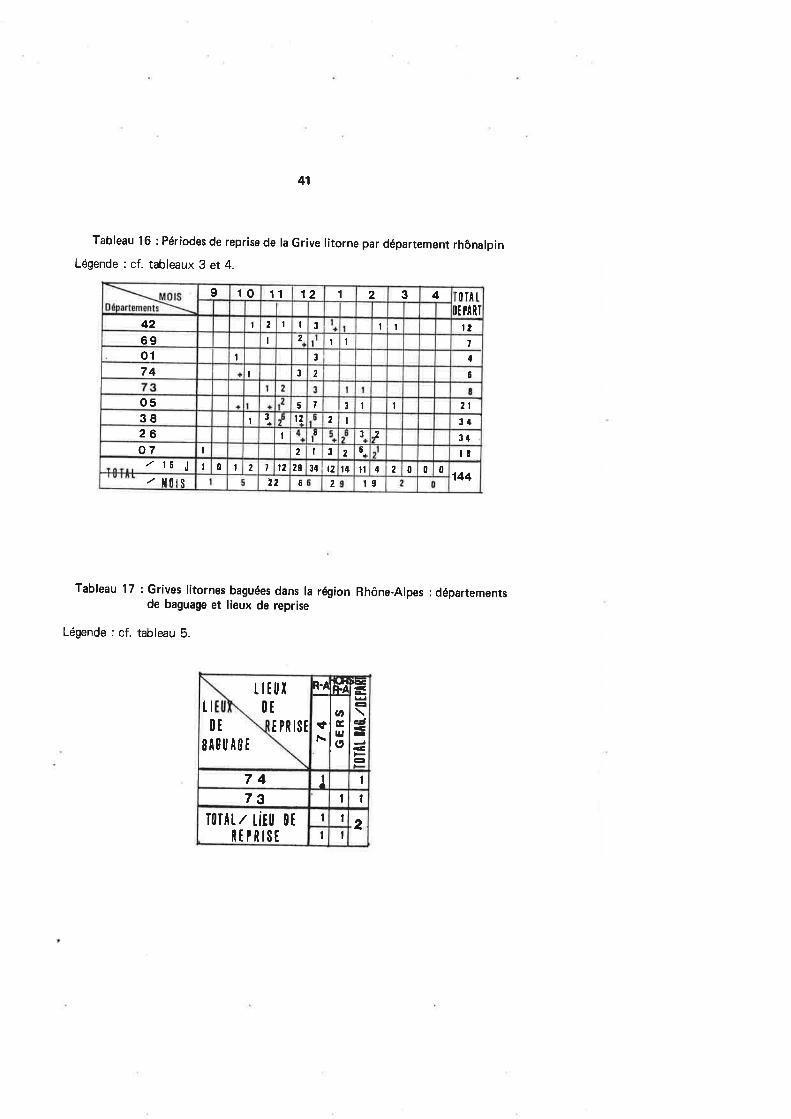
Les reprises régionales (tabl. 16 et fig. 19) deviennent importantes à partir denovembre (22cas pendant ce moisl, mais c'est en décembre que le maximum est atteint(66 cas), alors que janvier connaît une chute des effectifs, correspondant plus à une réa-
lité biologique (l'Atlas des Oiseaux Nicheurs Rhônalpins étant en parfaite entente à ce
sujet) qu'à une baisse de l'activité cynégétique (le tir des Grives étant, contrairement au
Merle, largement autor¡sé dans la plupart des départements au-delà de cette période).Cette chute de janvier est donc très certainement à relier à un tra¡t de comportementdéjà écrit pour d'autres espèces qui ont précédé, mais qui prend dans le cas précis de la
Grive litorne, une importance particulière. Celle-ci déserte toujours en effet les localitésencombrées par un enneigement persistant qui dérange sa prospection alimentaire, en
partie terrestre à ce moment de l'année. Or, janvier constitue souvent chez nous un moisdifficile au coursduquel la neige et le froid se combinent pour lui rendre l'existence plus
rude. Dès lors, sa seule alternative est de gagner un pallier inférieur qui n'en sera pas pourautant définitif. Cette fuite devant les intempéries est bien décrite par la plupart des au-
teurs. C'est ainsi qu'en Angleterre, un gros apport d'oiseaux en provenance de Norvèges'opère en février par temps défavorable (ASHMOLE, 1962). Cependant, SIMMS (1978)parle de mouvements lmportants à Fair lsle en décembre et janvier, apparemment nonprovoqués par une météorologie particulière. ASHMOLE (1962) pense que beaucoupd'oiæaux sont victimes du froid et que les populations peuvent subir de lourdes pertes
avant, pendant ou à la suite de leur exil forcé. Nous préciserons pour notre part que la
Grive litorne fa¡sait effectivement partie des espèces trouvées mortes lors d'une expédi-tion ornithologique au mois de février 1978 en Hollande par -20oC de température diurne.

36
L'Atlas des Oiseaux Nicheurs de notre région parle d'une remontée des effectifs en février,ce qui n'apparaît pas dans nos propres données de reprises, en dépit d'une certaine logiquequi voudraitque les oiseaux chassésdu Nord en janvier (ASHMoLE, 1g62) parviennentchez nous, compte tenu de la distance à couvrir, dans les semaines suivantes.
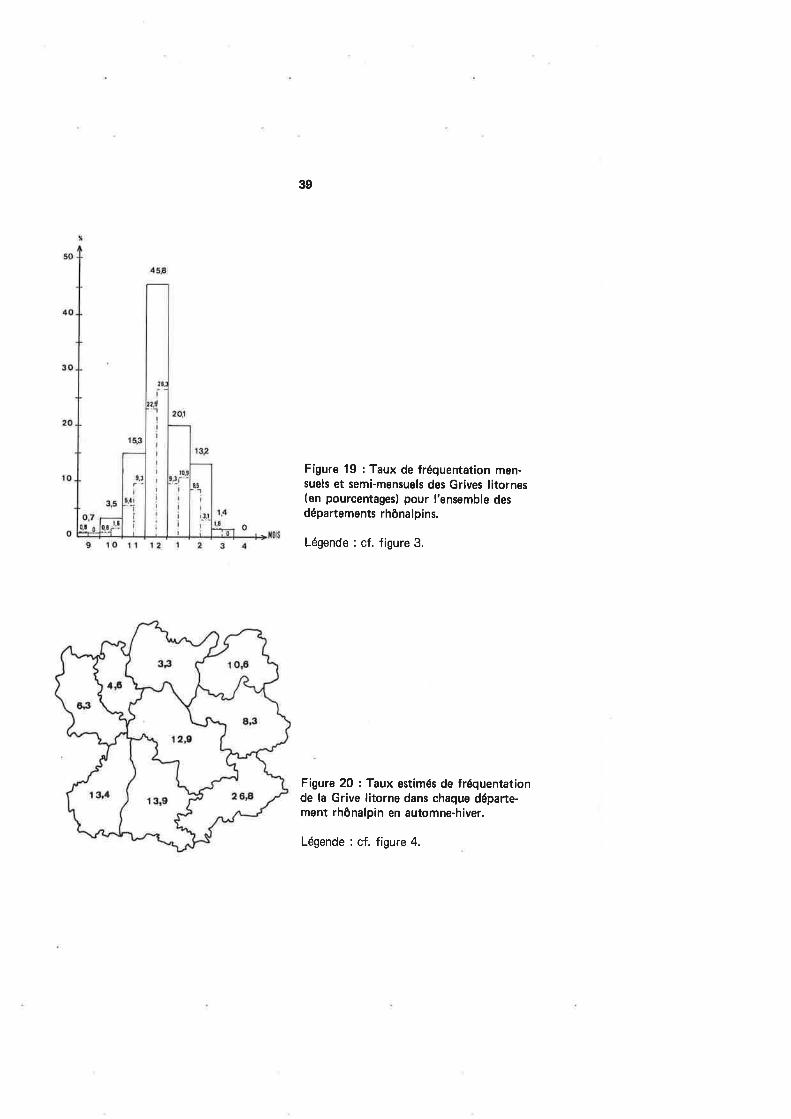
Les départements rhônalpins ne sont pas fréquentés de façon homogène par laGrive litorne : la figure 20 montre un foyer important de concentration de l'espèce dansles Hautes'Alpes. Contrairement aux espèces précédentes, il est difficile de trouver uneexplication à cette préférence.
Le tableau 15 montre que les données hivernales sont peu nombreuses dans lespays étrangers plus au Nord. Une présence assez continue (novembre, janvier et février)se manifeste dans le Sud de la Grande-Bretagne (ce qui est effectivement le cas d'aprèsASHMOLE, 1962). Notons que les 5 cas de captures britanniques ne donnèrent lieu àune reprise rhônalpine qu'au bout d'un ou plusieurs hivers. ll semble donc s'agir d,oi-seaux ayant radicalement modifié leurs emplacements successifs d'hivernage. Tel futsans doute le cas de cet ¡ndividu bagué le 12 février 1972 dans l'Huntingdon et tué leI février de l'année suivante dans les Hautes-Alpes. A ce propos, ASHMoLE (1g62)montre qu'une population nicheuse issue d'une même région peut essaimer à l'intérieurd'un angle de dispersion très ouvert au cours de I'hiver. Et l'auteur cite des cas æm-blables aux nôtres, d'individus s'étant considérablement écartés de leurs lieux de bagua-ge entre deux hivers différents, voire dans le cadre de la même saison. L'explicationdonnée est la suivante : la litorne serait la Grive la plus dépendante des baies en hiveret elle semblerait tout particulièrement capable d'adapter son comportement migrateurà la fructification des arbustes, variable dans l'espace et dans le temps. A cet égãrd,elle diffère considérablement d'une espèce comme le Merle à plastron, plus <rigìdementprogrammé> à hiverner dans un lieu immuable (commentaire personnel). ASHMOLE(1962) conclut qu'un tel comportement nomade est générateur d'une large zone dechevauchement entre les aires d'hivernage des différentes populations.Ailleurs en Europe du Nord et du Centre, les captures de décembre sont au nombre de3 (Danemark, Allemagne de l'Est, Autr¡che) ; celles de janvier se limitent à 2 (Belgiqueet Pays-Bas) ; tandis que février n'enregistre qu'un cas au Danemark. cette impress-ionde rareté hivernale qui se dégage à la lecture des données de baguage dont nous dispo-sons n'est en réalité qu'apparente. Les Grives litornes sont en effet visibles, au moinsen début d'hiver, un peu partout en Europe, alors qu'en février, à l'issue du <glisse-ment) dont nous avons parlé, les plus fortes concentrations peuvent s'observerdans leSud-Ouest de la France et dans les lles britanniques. Toutefois en février ig81 , d'impor-tantes troupes de ces Grives animaient localement la campagne hollandaise (observaiionpersonnelle).Au Sud, des sujets capturés en ltalie du Nord donnèrent lieu à des reprises dans lecadre rhônalpin :dans 4 cas sur 5, cela se passa dans les semaines ou les mois qui sui-virent le baguage, ce qui suppose une trajectoire abordant notre région par le Sud-Est.Nous verrons plus loin que certaines Grives mauvis scandinaves ontété capturées enItalie avant d'aboutir dans le Midi de la France. Aussi, les Grives litornes baguées enItalie sont-elles éventuellement de provenance nordique. Mais une origine orientale estégalement envisageable pour ces oiseaux; d'autant qu'un cas yougoslãve (le seul de

37
cette provenance pour l'ensemble des fiches des six espèces étudiées) peut étayer cettehypothèse ; de même que les deux cas soviétiques dont il est question un peu plus loinau chapitre des records.
, Nous ne possédons aucune indication sur les déplacements éventuels des Griveslitornes qui se reproduisent dans les départements du Nord-Est de notre région. Un oiæaumarqué adulte à Saint-Gervais (Haute-Savoie) le 4 septembre 1974, a été contrôlé surplace un an plus tard, preuve de sa fidélité à un site de reproduction antérieur ('fig.21et tabl. 17).
Ouelques longévités intéressantes :
- celle de'l ans 112 a été atteinte par un sujet bagué au nid en Finlande le 8juin 1960 et tué en Ardèche en janvier 1968.
- 8 ans 1/2 aura été la durée de vie d'un autre oiseau bagué poussin dans lemême pays le 26 mai 1963 et tué dans les Hautes-Alpes le 12 mars 1971.
- deux autres cas correspondent à des longévités de 6 ans environ.
Ouelques records de distance et de distance/temps intéressants :
- une Grive litorne a couvert la trajectoire Bergamo (ltal¡e) - Hautes-Alpes enune semaine.Bien plus intéressants encore sont les deux cas figurés par des symboles fléchés sur lafigure 18. Leurs lieux de baguage respectifs sont nettement excentrés (le second enparticulier) des contrées auxquelles nous ont habitués les autres reprises.
- la première de ces deux Grives litornes fut baguée au nid le 25 mai 1976 enU.B.S.S. par 57050'de latitude N et 56030'de longitude E;elle fut tuée le 25 dé-cembre 197'7 en Ardèche (à plus de 3 500 km de son lieu de marquage).
- le second cas est particulièrement remarquable :parti le 6 mai 1964 de larégion de Prokopievsk en Sibérie centrale (53o54'N 186"44'El , il échoua au terme d'unvoyage de I 000 km à Chalançon (Drôme) le 10 janvier 1966.ERARD (1967) cite 1'l cas français (dont celui qui nous intéresse) de reprises de litor-nes originaires du Plateau Central de Sibérie (1) et en arrive à la conclusion suivante :
des mouvements d'une telle ampleur sont indépendants des migrations régulières annuel-les. ll s'agit en réalité <de larges fluctuations de type invasionnel (qui) se produisentselon les disponibilités alimentaires présentées et le niveau de la population l'annéeconsidérée>. Et le même auteur de rattacher le phénomène aux invasions périodiquesde Becs-croisés et de Jaseurs, plus classiques. SVARDSON,n SIMMS (1978), rejointcette idée en affirmant qu'en SuàJe, des invasions hivernales inhabituelles se produisenttous les trois ou quatre ans sans retour apparent vers les lieux d'origine. ll pourraits'agir là de la clé régissant l'avancée spectaculaire de l'espèce dans la plupart des régionsrécemment colonisées. C'est d'ailleurs par cette tactique de non-réintégration des zonesde départ, que l'lslande et le Groenland ont été peuplés par des reproducteurs initiale-
(1) Ces oiseaux orientaux ne paraissent pas traverser la Mer du Nord, car aucune reprise semblablen'existe dans les lles br¡tanniques.

38
ment ¡nstallés en tant qu'hivernants (SALOMONSEN /n ASHMOLE, 1962 et YEAT-MAN, 1971). ASHMoLE (1962) pousse plus loin l'analyse en ajoutant que la nidifica-tion au Groenland a été rendue possible par l'arrivée tardive d'une troupe d'hivernantsà l'issue d'une vague de froid continentale en janvier. Deux mois plus tard, ces mêmesoiseaux ne semblent pas avoir été stimulés à faire demi-tour : ils ont donc estivé, puisniché et se sont ainsi sédentarisés, aidés en cela par un réchauffement climatique récent(de l'ordre de 5oC en période hivernale).
Figure 18 : Origine des Grives litornes -Turdus pilarís- reprises dans la région Rhône-Alpes.
Légende : cf . figure 2.Le gros symbole fléché met en relief deux oiseaux dont les lieux de baguage sontsitués en dehors des limites de la carte (cf. texte page précédente).
*.1
39
Figure 19 : Taux de fréquentation men-suels et serni-mensuels des Grives litornes(en pourcentagesl pour I'ensemble desdépartements rhônalpins.
Légende : cf. figure 3.
Figure 20 : Taux estimés de fréquentationde la Grive litorne dans chaque départe-ment rhônalpin en automne-hiver.
Légende : cf. figure 4.

Æ
I OISEAU REPRISSUR LE LIEU DEEAGUAGE
Figure 21 : Localités de reprisedes Grives litornes baguées dansles départements rhônalpins.
Légende des symboles : cf. figures2et5.
Tableau 15 : Périodes de baguage de la Grive litorne par pays d'origine
Légende : cf. tableau 1.
41
Tableau 16 : Périodes de reprise de la Grive litorne par département rhônalpin
Légende : cf. tableaux 3 et 4.
Tableau 17 : Grives litornes baguées dans la région Rhône-Alpes : départementsde baguage et lieux de reprise
Légende : cf. tableau 5.
r. ril,I0tU
0t t PR r$t
8A0llA0t
R-Á
çt\
IT¡IlF4
t,lc
lä
DÈ-t-tã-
lil.¡<É<ct
74 I I
73 I I
T0TAtz tittl 0tNEPRI$T
I I 2I I
9 10 11 12 1 2 3 4 r0t r
It rAnt42 I 2 t 3 I I t269 I 2
1
ol 3 4
74 3 2 I
0s 5 I 3 I 2t38 I 3¡ l2 2 I 3a26 ü
I 3 2 3{o7 2 I 3 2 tt tl/ 15 J 0 I 2 l2 29 34 t2 t4 ll 4 2 0 0 0
144/ tnts 22 6 2 l9

42
- GRIVE MAUVIS Turdus musicus (= T. iliacusl
Ce dernier représeñtant du genre Turdus (le plus nordique des six) est souventasocié à la Grive litorne dans les carnets de notes des ornithologues. Cela tient au faitque des bandes mixtes de ces deux espèces sont visibles l'hiver, lorsqu'elles se nourris-sent dans les prés et les labours ou s'abattent sur les haies en plaine et à moyenne al-
titude. D'ailleurs, cette coex¡stence se retrouve dans les vastes étendues boisées de latai'ga oir l'une et I'autre cohabitent pour nicher. Mais à l'encontre de la Grive litorne,la Grive mauvis n'a pas manifesté de tendance à essaimer à partir de son milieu origi-nel, restant l'hôte discret des forêts de conifères et de Bouleaux du Nord de l'Europe.L'aire de reproduction de la Mauvis couvre donc la Scandinavie (Sud de la SuMe ex-cepté), la Finlande, l'U.R.S.S. (avec groso modo une limite Sud oscillant d'Est enOuest entre les degrés 50 et 55 de latitude N), I'Ecosse (où elle n'est pas commune)et le pourtour de l'lslande. A l'Est, elle est le seul Turdidé européen à atteindre en
bordure de l'Océan Glacial Arctique la longitude de 160 degrés.
Pour différentes simil¡tudes écologiques réunissant les Grives litorne et mauvis, il nousa paru intéressant d'établir un parallèle entre les deux à chaque fois que l'occasionnous en était donnée. Nous nous sommes donc attachés à voir si leur statut rhônalpin,au travers des reprises de bagues dont nous disposions, se révélait ausi étroitementsemblable que nous le supposions a priori.
Le Centre Régional de Baguage Rhônalpin dispose de 57 reprises imputablesà la Grive mauvis. Ce nombre relativement restreint est-il en relation avec le caractèrenocturne de sa migration, la mettant plus que d'autres espèces de Turdidés, à l'abrides fusils rhônalpins (aux cols des Alpes et du Massif Central en particulier) ? ll estpermis de le croire, car SIMMS ('1978) qualifie la Grive mauvis de <typically night mi-grant). Ne perdons pas de vue cependant que l'aire de reproduction de cette espèce
est moins étendue que celle de la Grive litorne qui totalise 154 reprises étrangères (cf.
précédemment). Sur ces 57 fiches détenues par le centre (fig.22l,,15 correspondent à
des oiseaux bagués poussins : 9 en Finlande, 4 en Estonie, 1 en Norvège et 1 en Suède.Deux autres captures estivales figurent en Finlande. ll ressort donc que la populationfinnoise est très largement majoritaire parmi les migrateurs qui fréquentent notre ré-gion. Les oiseaux scandinaves, comme pour la Grive litorne, ne paraissent pas traverseren nombre les départements rhônalpins.
Les 40 reprises restantes sont trop insuffisantes pour établir un calendrier dela migration des Grives mauvis à travers l'Europe, Nous pouvons toutefois noter sur le
tableau 18 qu'en Norvège et Finlande, la date du 15 octobre epparaît comme une li-mite de leur préænce (on sait que cette Grive hiverne moins volontiers que la litorneà une latitude élevée). ASHMOLE (1962) mentionne pourtant quelques hivernants dansle Sud-Ouest de la Norvège ainsi qu'en lslande. Une capture dans la seconde quinzainede septembre en Allemagne de l'Ouest est à rapprocher de ce qu'écrit le même auteursur le déroulement de la migration d'automne : le passage débute fin æptembre, s'accen-tue en octobre et reste important jusqu'à la mi-novembre. Notons encore sur notre ta-

43
bleau que 2 individus étaient encore présents en Lithuanie dans la seconde quinzaine
d'octobre alors qu'à la même époque,5 oiseaux étaient déjà capturés en Suisse, or)
les premiers sont même notés fin septembre à Bretolet (DE CROUSAZ, 1960)'
Au niveau régional (tabl. 19 et fig.23), les reprises débutent à la mi-octobre.sàns accuser de hausse notable juqu'à la fin de novembre, Le début de décembre paraitpar contre, comme pour le Merle noir'et la Grive litorne, alimenté par une arrivée plus
massive de migrateurs, manifestement chassés plus au Nord par les intempéries. Ces
mouvements tardifs ont pu faire écrire à SIMMS (1978) : <Redwings are great hard-weather migrants). ASHMOLE 11562) décrit des passages importants à Fair lsle enjanvier ainsi qu'à Héligoland en décembre; mais ils seraient peu marqués au Danemarket en lrlande (HANSEN et BARRINGTON /n ASHMOLE, 1962l.Par contre, les repri-ses de bagues accusent une baisse importante en janvier dans le Nord de la France eten Belgique, signe d'une désertion de ces latitudes au profit de régions moins défavorisées.
Plus tard dans l'hiver, la situation rhônalpine des Grives mauvis et litorne diffère, car
contrairement à cette dernière, l'afflux de Grives mauvis qui avait marqué le début dumois ne se poursuit pas au-delà du 15 (comme si ces oiæaux ne faisaient que traversernotre région brièvement sans y séjourner vraiment). Très vite en effet, l'effectif des
reprises par quinzaine se stabilise à nouveau à ce qu'il était avant le pic; c'est-àdire,autour de la valeur 5, et ce jusqu'à la mi-février, avec toutefois une baisse à 1 à la findu mois de janvier. Ainsi donc, à l'exception d'une hausse confinée au début de décem-
bre et d'un creux (peut-être dû au hasard) deux mois plus tard, la présence de la Grivemauvis dans la région RhôneAlpes apparaît-elle comme stable d'un bout à l'autre de
la saison.
La figure 24 monTre, comme chez la Grive litorne, une attirance marquée de
la Grive mauvis pour le département des Hautes-Alpes. La Drôme et la Haute-Savoieparaissent également bien fréquentées sans qu'il soit possible d'en préciser la cause.
Cette Grive montre en général une forte tendance à la dispersion <en éventail>
à partir d'une même zone de reproduction ; aussi les populations sont-elles très mélan-
gées pendant l'hiver (ASHMOLE, 1962). Selon RENDAHL /n SIMMS (1978), ce sontles adultes qui se livrent à des mouvements extrêmes qui les poussent le plus à l'Est età l'Ouest. A l'échelle rhônalpine, ce caractère n'apparaÎt guère : l'origine des oiseaux
repris couvre en effet une zone assez bien délimitée dans l'espace. Les deux populationsqu¡ sont amenées à fréquenter, dans des proportions différentes, les départements qui
nous concernent (pour mémoire, les oiseaux finlandais et ceux, moins nombreux, de
Scandinavie) figurent aussi dans les cartes dressées par JONES (1961) pour lesdéparte-ments máliterranéens. Pourtant, un petit détail mérite notre attention : les oiseaux
issus de Norvège représentent I % de I'ensemble des repriæs enregistrées dans le Sud de
la France et seulement 2o/o chez nous. ll nous paraÎt intéressant de rattacher cette dif-férence à une hypothèse formulée par JONES (1961)qui présurne qu'une migrationvers le Midi via l'ltalie existe de la part de reproducteurs scandinaves. La comparaison
de nos données respectives cadre avec cette idée et nous incite à penser que des oiseaux

M
norvégiens doivent effectivement atteindre les départements de la MáJiterranée selonune trajectoire passant à I'Est de chez nous; tandis que d,autres, sans doute moinsnombreux, se déplacent plus à l'Ouest et aboutissent dans les départements rhônalpinspar le Nord' Toutefois, certains sujets dont l'axe de migration esi le plus oriental jarrapport à la chaîne des Alpes bifurquent ensuite à l'Ouest depuis I'ltalie et atteignentnotre région' A l'appui de cette affirmation, 4 repriæs rhônalpines enregistrées dãnsles semaines qui suivirent le baguage des oiseaux concernés en ltalie septentrionale.Quant aux trois reprises hivernales ayant pour origine les lles britanniques, deux re-marques peuvent être formulées à leur égard : tout d'abord, qu,il ne stagit probablementpas d'oiseaux locaux (la nidification en Ecosse restant clairæmée) mais ãe iujets scan_dinaves ou finlandais (1) ; ensuite, que dans les trois cas, les dates de baguage et dereprises sont séparées par une année. La conclusion logique qui s,impoæ est que deschangements de zones d'hivernage peuvent s'opérer d,une saison à liautre. Le fait estd'ailleurs bien connu et abondamment évoqué dans la littérature. GERouDET (1962)écrit par exemple : <... mais une certaine irrégularité se manifeste; des hivernants deGrande-Bretagne ayant été repris en Grèce et à chypre, lors de saisons ultérieures>.SIMMS (1978) pour sa part, attribue ces changements aux conditions météorologiquesannuelles ou à, selon ses.termes, <an unusually nomadic instinct>. ASHMoLE llgozlnote encore l'existence de concentrat¡ons temporaires massives de Grives mauvis en deslieux qui ne seront pas forcément peuplés les mois ou les hivers qui suivent, et proposeen explication, le succès localement plus ou moins grand de la fructification des arbus-tes en automne. Ces différents facteurs conjugués font de la Grive mauvis un visiteursoumis à d'importantes fluctuations annuelles dans le Midi de la France (JoNEs, 1961).ses incursions dans le sud de l'Espagne sont sans doute aussi très irrégulières, maisprobablement moins rares que ne l'écrivait GERouDET (1962) puiqLe pas moins de28 % des Grives mauvis baguées en Belgique ont donné lieu à une reprise dans la pénin-sule ibérique (LIPPENS & WtLLE, 1972).
Ouelques records de distance/temps intéressants ;
- un oiæau bagué le 26 octobre 1gl1 au col de Bretolet (valais), fut reprisdans la Drôme le lendemain.
- un individu marqué en u.R.s.s. (b5.11 N /20.49 E) te 1g octobre 1966, futtué en Hautesavoie dix jours plus tard, au terme d'un vol de 1 400 km environ.
{1} certainement pas islandais, car les Grives mauvis de c€tte provenance sont so¡t sédentaires, soitl jy:tl"ll en Ecosse, aux Hébrides.ou en certainei regions ae t;extremeöuãsiàlcàntinent (cuND-MUNDSSON in ASHMOLE, 1962).
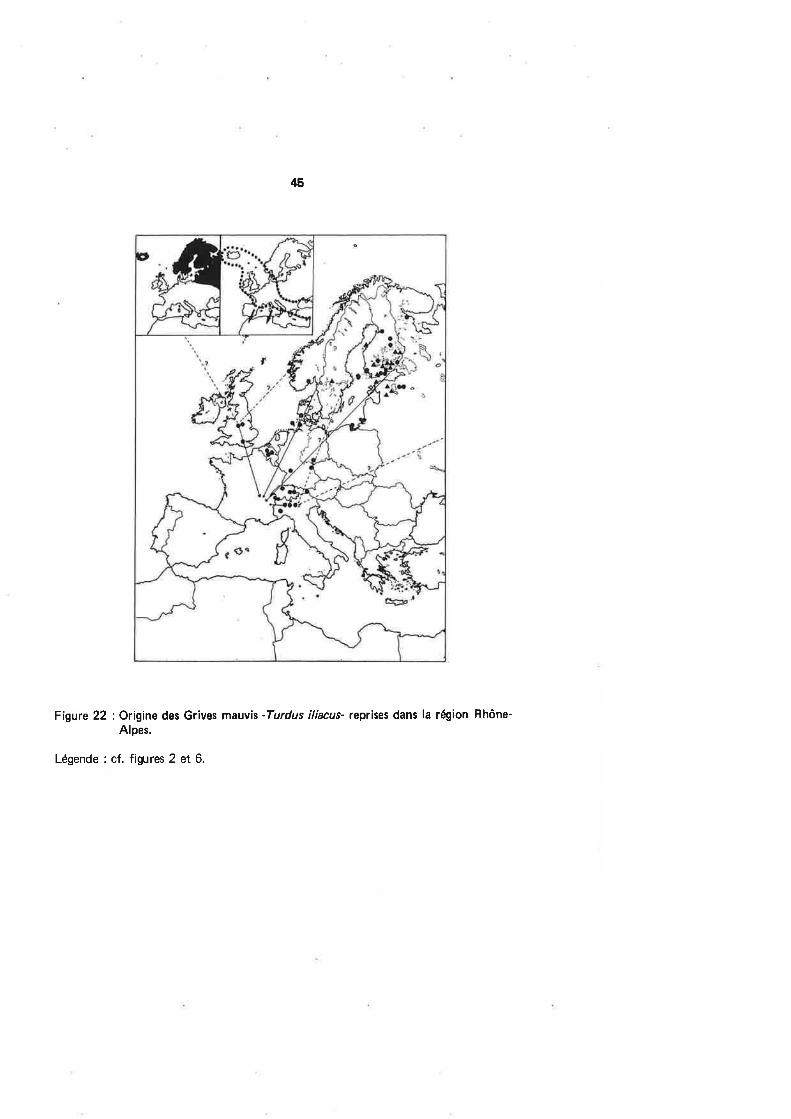
¡15
Figure 22 : Origine des Grives mauvis -Turdus iliacus'reprises dans la rfuion Bhône-Alpes,
Légende : cf. figures 2 et 6.
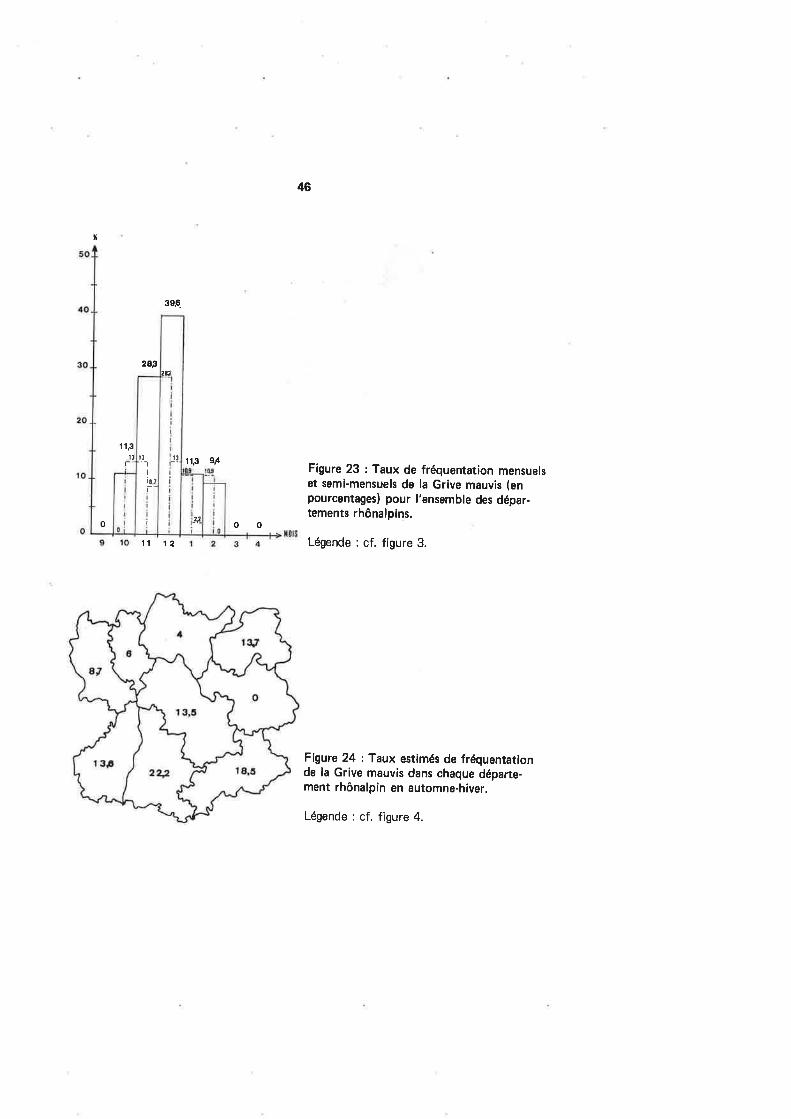
r
3e0
2432a;
t3
1 t,3lt
tt¡'-l
I
!-!¿I
1r3 ef
o ?i oo
46
1211
Figure 23 : Taux de fréquentation mensuelset semi-mensuels de la Grive mauvis (enpourcentages) pour l'ensemble des dépar-tements rhônalpins.
Légende : cf. figure 3.
Figure 24 : Taux estimés de fréquentationde la Grive mauvis dans chaque départe-ment rhônalpin en automne-hiver.
Légende : cf. figure 4.

47
Tableau 18 : Périodes de baguage de la Grive mauvis par pays d'origine
Légende : cf. tableau 1.
Tableau 19 : Périodes de reprise de ra Grive mauvis par département rhônalpin
Légende : cf. tableaux 3 et 4.
M0sPAYS
I 2 3 4 5 6 7 8 9 10 1f 12 IIITÀ
/ P^l
I 2 4t5 I 2 0 0 I 0 0 0 I 6 z 5 3 0 4 3 t0 ll I 0 '|
57/l¡ I 0.,
7 3 5 a 2 {
M0ts 9 10 't1 12 I 2 3 4 t0ï t
OTPAR
38 2 I 4 2 t026 t 5. 3 3 t9o7 I I + tjl I
/ t5 00 0 6 6 4 3 6 5t 5 0 0 0 00/ rl|ts 0 6 r5 2l 6 5 0 0

48
lV - Discussion
La fréquentation rhônalpine de 4 espèces de grands Turdidés (1) paraît soumiæà d'importantes fluctuations annuelles, comme le montre la figure 25 et le tableau 20.Les lignes qui précàJent ayant mis l'accent sur la réaction de fuite d'au moins 5 espècesde Grives et de Merles vis-à-vis d'aggravations météorologiques survenant en cours d'hi-ver, nous avons cherché à savoir si les indices de leur présence pouvaient être corrélésaux conditions climatiques qui ont caractérisé par le passé les saisons hivernales denotre région. Pour ce faire, nous nous sommes plus particulièrement penchés sur lescas de 5 saisons hivernales-test classées de <très doux> à <très froid> (avec trois inter-médiaires), Nous avons fait apparaître dans le tableau 21 qu'il n'existait sans doute pas
de lien étroit entre les rigueurs hivernales enregistrées à notre latitude et les taux an-nuels de fréquentation régionale des Merles et des Grives. Chez les Grives mauvis et leslitornes en particulier, les effectifs annuels semblent dans l'ensemble parfaitement dis-semblables. On remarque en effet sur la figure 25 que certains hivers particulièrementpeu fournis en Grives mauvis, étaient au contraire d'excellentes années à litornes et in-versement. Notons, par contre, que le très sévère hiver 1910-71 révèle une préæncemaximale rigoureusement identique pour les deux espèces.En conclusion de cet aspect particulier, nous supposerons au vu des résultats, que laprésence des Merles et des Grives dans la région Rhône-Alpes, n'est pas fondamentale-ment régie par les conditions climatiques locales. Nous avons de bonnes raisons decroire que la biomasæ alimentaire localement disponible (sous forme de baies en parti-culier) doit, au moins chez certaines espèces. jouer un rôle en tant que facteur annuel-lement limitant ou favorisant. Mais il est probable que des causes extérieures (c'est-à-dire sur les lieux de reproduction) à la fois complexes et variées, doivent égalementexercer leur influence. ll n'est qu'à se souvenir à ce sujet des cas de Grives litornesorientales épisodiquement retrouvées en France.
Tableau 20 : Répartition des reprises par espèce et par année
Le total des reprises utilísées est inférieur à l'effectif réel car certa¡nes données (moisde reprise, année de reprise...) sont absentes ou incomplètes, ce qui rend partiellementinutilisables les fiches correspondantes.
(1) Nous préférons laisser de côté le Merle à plastron et la Grive draine pour lesquels nos donnéessont insuffisantes.
I0Ttt/s il $0tt (%)
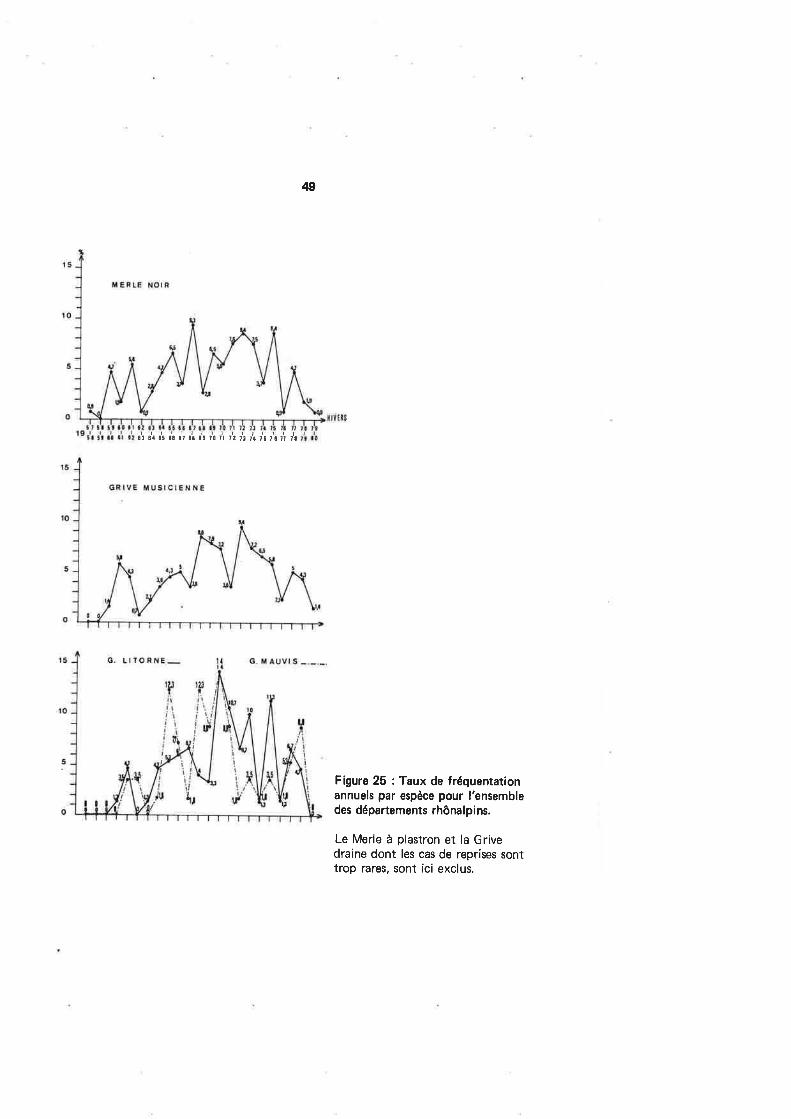
49
8l El 13 il l, 6t ll ,0 ll 12 t7 t. t5 tt 1, tt
Figure 25 : Taux de fréquentationannuels par espèce pour l'ensembledes départements rhônalpi ns.
Le Merle à plastron et la Grivedraine dont les cas de reprises sonttrop rares, sont ici exclus.
50
Tableau 21 : Taux de reprise par espèce au cours de cinq années-échantillon
Le Merle à plastron et la Grive draine dont les cas de reprises sont trop peu nombreuxne f igurent pas ici.
Codification utilisée : 6 points : très bon hivernage5 points : bon hivernage4 points : assez bon hivernage3 points : moyen hivernage2 points : faible hivernage1 point : très faible hivernage
Nous livrerons enfin quelques remarques sur l'âge-ratio des six espèces deMerles et de Grives figurant dans nos fichiers; non sans avoir auparavant précisé quele sujet a déjà fait couler beaucoup d'encre, les avis étant largement partagés. Uneopinion <classique> consiste à admettre (au moins pour le Merle noir, mieux étudié)que les individus juvéniles sont davantage enclins que leurs aînés à æ déplacer (GEROU-
DET,1S62, entre autres). LACK /n ASHMOLE (1962) prétend au contraire que 33 %
des Merles britanniques adultes se déplacent pour seulemenl2l o/o de juvéniles. SelonSNOW /n SIMMS (1978), le comportement migrateur du Merle noir n'est pas lié à
l'âge ; certains individus æ révélant sédentaires d'un bout à l'autre de leur existence,d'autres se dispersant en automne au-delà de un an. ASHMOLE (1962) pour sa part, se
situe dans un juste milieu en émettant l'hypothèse que certains sujets puisænt avoirun comportement hivernal non définitif ; tantôt migrant, tantôt restant sur place selonles années.Nos propres données sont elles-mêmes très contradictoires : sur l'ensemble des Merlesnoirs bagués chez nous (qu'ils soient reproducteurs ou de passage) et ayant donné lieuà une reprise plus ou moins lointaine dans les semaines ou les mois qui suivirent, l'âge-rat¡o est nettement déséquilibré en faveur des jeunes de l'année : 18 contre 8 adultes,soit près deTOo/o d'oiæaux de I'année. D'un autre côté, cette tendance se trouve tota-
-ì--,..- lllltRststtcr$---\
73-74TNES DOUX
75-76ASSEZ DOUX
68-69IOYENNÊMEXT FM
67-68FROID
7o-71rRES FROID
MEßLE NOIR,,J Ë
oaaatt,{Ë
oaaaa¿ toh
aY,JË
ooaoo5,OÀ
oaaGRIVE frluslclENl I,zh
aaaaJ,OÈaaa
o,oåaaaaa
J,OÈ
oaL¿h
aoaa
GRIì/E LITORNEaooaa aaaoaa .O
o,rõaao aooaoa
GnlvE ltauvts it,c'Ë
ooJ,5lO'
l z,J.É
aooaoa1 aua
14 t"aaaaao

51
lement inversée dans les reprises originaires de l'étranger pour lesquelles les adultes sont,dans la plupart des cas, majoritaires (fig. 26). Même si la détermination de l'âge des es-
pèces du genre Turdus n'est pas toujours aisée, nous ne pensons pas que ces résultats,sans doute surprenants, soient l'effet d'erreurs de la part des manipulateurs étrangers.Aussi, nous bornerons-nous à les considérer tels quels à défaut de pouvoir leur trouverune expl ication satisfaisante.
OISEAUX AAGUES AUETRANGER ET
R€PRrs gArE îtroNE
)ISEAUX REPRIS IOISEÀU REffi 15
- ,.rr lr,¡r¡cell PAR LÀsr.JtTE
Fõi]tïÉsfinEñltEsl
G. DRAINE12
,/ \,
23(60i JUV.)
MERLE NOIR
107
/\65 42
42f JW.
G LITORNE154
/\79 75
36 43
54r 'UV.
M. A F-ASIRON GRIVE MTJSICIENNE
16 139
" / \,
"o/ \un
( 25f JUV)
G. Mauvls57
/\42' f s
23 19
¡1 5f lUV.
38
47f JUV
38 27 62
Figure 26 : Répartition chronologique par espèce des reprises d'origine extérieure etâge-ratio des oiseaux repris l'année de baguaç.
Les taux obtenus pour le Merle à plastron et la Grive draine sont donnés sous touteréserue (entre parenthèses) compte tenu de l'effectif restreint de ces deux espèces.
V - Conclusion
Au-delà de l'aspect fondé sur une meilleure connaissance rhônalpine des espè-
ces appartenant au genre Turdus, nous espérons avoir montré que des données telles que
celles que nous avons utilisées pour les Grives et les Merles, sont disponibles et accessi-
bles à quiconque souhaitera à l'avenir s'intéresser par repriæs de bagues interposées, au
statut d'espèces fréquentant la région Rhône-Alpes. Les Anatidés, certains Laro-Limico-les. quelques espèces de Rapaces, ainsi que des groupes de Passereaux (Fringilles en par-
ticulier) et des espèces isolées (Rougegorge, Mésange bleue, Bruant des roseaux...)constituent autant de cas en attente pour lesquels les données sont d'ores et déjàtrès substantielles.

52
V - Bibliographie
ASHMoLE (M.J.) 1962 - rl.re. 1rp1atl91 of_european Thrushes : a comparative study based on rin-ging recoveries - lbis 1O4,914-946 et b22_bb9.AUSTIN (O.), SINGER (A.) t97g - Oiseaux - Ftammarion, Vérone, 317 p.COCHET (G.) 1977 - Nidification de la Grive litorne en Haute-Loire - Nos oiseaux,34 (96g), 134.cocHET (G.) 1980 -Trois nouveilesespèces nicheusesen Ardèche - r0z Nature, (pubrication dépar-tementale du C.O.R.A.), no 6, 14-16.DE cROUSAZ (G.l 1960 - Act¡vités de l'Observatoire ornithologique alpin du col de Bretolet en
1 959 - /Vos oisæux , ZE teæl , j 69-1 94.DURMAN (R.F.) 1976 - Ring Ouset migration _Bitd study,2g (31 ,1g7-2O5.ERARD (Ch.) 1967 - Sur la- présence hivernale en France de Grives litornes lurdus pilaris L. d,origine
sibérienne. Atauda,A5 l1l ,20_26.GEROUDET (P.) 1963 - lra
-u!e dgs^oigea^ux : les passereaux ll (des Mésanges aux Fauvertes) - Dela-chaux et Niestlé, Neuchátel,3Og p.
HE lNzE L _(H.), FtrrER (F.), pARs Low lJ.l 1972 - oisæux d,Europe, d'Afrique du No¡d et duMoyen.Orient - Delachaux et Niestlé, Neuchâtel, 320 p.JONES (P.H.) 1961 - Lieux d'origine des Grives de Ia France méditerranéenn e - Ois, Rev. Fr. Orn,,31 (3), 193-21 3.
LEBRETO.N (Ph.), CHABERT (8.) 1969 - compte rendu ornithologique annuel de l'automne 1967à r'été 1968 dans ra rfoion Rhône-Ar¡ies - But. mens. soc'. i.nn. lvãn,a,llo:zïsüii222-242.
LEBBETON (Ph.),CZAJK^Of/VS.K.| (MJ,CHABERT (8.) 1971 -Compterenduornithotogiqueannuelde |automne 196€.à rété r969 dans ra région nñone-Ãipäi-- eliL,ßn| soà.-iî;;.î'i;;;3,80-92 er4,i14-124.
LEBREToN {Ph.}, THEVENoT (M..}, LEBREToN (J.D.) et BRAEMER (H.) 197r - Etude ornitho-tos ique du Massif du. pi rat lLoirer - Ata uda, 39 (4), sr z-s?b ãi +'ó i I i, sl -sz.
LEBRETON 1Ph'), CZAJKOWSKI (M.) 1972 - Compte rendu ornithologique annuel de l'automne1969 å t'été 1970dans ra région Rhône-Arpes -Bulr. mens. sõìli,*. tvolr, 1,1g-1 Ber4,57-64.LEBRETON (Ph.),CZAJK9WSKl (M.) 1973-Compterenduornithologiqueannuel del'automne
1970 à l'été 197r dans ra rfoion Rhône-Aipes -|uil. mens. sõ")liir. tvon,5,i12-128et 6, 133-142.
LEBRETON (Ph.),RlCHOtlx.(M.),CZAJKOWSKI (M.) 1974-Compterenduornithotogiqueannuetde t'automne 1971-à r'été 1972 dans ra région nhone-Àrpei-buli.-miis. soc. iñ;: Li;;-,1 ,21-32 et 3,80-91.
LEBRETON (Ph'), RICHO-U-X. (M'), CZAJKOWSKI (M.) 1975 - Compte rendu ornithologique annuelde l'automne 1sZÈ !9t! 1973 dans ta rfoion Rhône-Atpei autt.-mãns. soci. L¡ìi. ùòi-,4 , 1 23-1 28 et 7 , 2B0-296 .
LEBRETON (Ph.) 1975-B-iogéographiedesoiseauxduMassif Central -Actesdug8eCongrèsNatio-nal des Sociétés Sayantes (æction des Sciences, tome l, extrait), 3ã-9-346.
LEBRETON (Ph.) etC.O.R.A. 1977 -Lesoiseaux nicheurs Rhônatpins (Atlasornithologique Rhône-Alpes), C.O.R.A., Lyon,3S3 p.
LEBRETON (Ph') 1980 -Attasorn¡thotogique Rhône-Alpes,complémenrs 1976-1979 -LeBièvre,2,supplément,80 p.
LIPPENS (L.) , W I LLE lH 'l 1972 - Atlas des oiæaux de Belgique et d'Europe occidentdte - Lannoo,liel't,847 p.
PERRINS rc) 1974 - Birds - Coilins, Londres, 173 p.

53
PETERSON (R.) , MOUNTFORT (c.),lloLLOM (P.), GEROUDET (P.) 1976 - Guidedes Oiseauxd'Europe - Delachaux et Niesilé, Neuchâte|,447 p.
RICHOUX (M.), PERSAT (H.1, COBNILLON (8.) 1977 - Compte rendu ornithologiqueannuel del'automne 1973 à l'été 1974 dans la rfoion Rhône-Alpes - Bull, mens, Soc. Linn. Lyon,'10, 342-360.
'RICHOUX (M.) 1979 - Compte rendu ornithologique annuel de l'automne 1974 à l'été 1975 dans larégion RhôneAlpes - La Bièvre, 1 (1), 53-73.
BICHOUX (M,), CORNILLON (8.) 1979 - Compte rendu orn¡thologique annuel de l'automne 1975 àl'été 1976 dans la région Rhône-Alpes - Le Biàvre, 1 (21 ,'17-35.
SCHIFFERLI (4.),GEROUDET (P.),WINKLER (R.) 1980-AttasdesOiseaux nicheursdesuisse,Station Ornithologique de Sem pach, 462 p.
SHABROCK (J.T.F,) 1975 - The Atlas of Breúing Bìrds in Britain and lratand - British Trust of Orni-thology, lrish Wildbird Conservancy,4T9 p,
SIMMS (8,) 1978 - British Thrushes - Collins, Londres,304 p.
SPENCER (8,) 1975 - Changes in the distr¡bution of recoveries of ringed Blackbirds - Bird Study,22(3),177-190.
YEATMAN (L.J.) 1971 - Histoirc des Oiseaux d'Europa - Bordas, 363 p.
YEATMAN (L.J,l 1976 - Atlas des Oiseaux Nicheurs de France, Société Orn¡thologique de France,Paris,282 p.
Vl - Remerciements
Diverses personnes et d¡vers organ¡smes ont, à t¡tres divers, apporté leur sou-tien à cette étude ou fourni des renseignements ayant servi à son élaborat¡on, Nous te-nons tout d'abord à remercier Hervé COOUILLART et Stéphane AULAGNIER pourleur contribution au traitement statistique de certaines données. Guy JARRY duC.R.B.P.O. nous a mis en contact avec les responsables de la convention EURING.MarieFrance LE PENNEC a pris part à la traduction de la littérature anglaise consultée.Pierre GUYOT nous a transm¡s la documentation cynégétique dont nous avions besoin.Nous ne saurions de même oublier les responsables des Fédérations Départementalesde Chasseurs pour leur aide s'étant à la fois manifestée dans leur réponse à notre en-quête et dans la centralisat¡on sans cesse répétée des bagues obtenues dans leurs dépar-tements respectifs. Enfin, nous exprimons notre grat¡tude à M. BOURNAUD,P. CORDONNIER et Ph. LEBRETON qui ont bien voulu critiquer une première ver-soin de ce travail.
54
MERLE NOIR (jeune mâle)

55
Hervé COOUILLART
L,AVIFAUNE DES GRAVIERES DE BAS.EN.BASSET (Haute-LoiIeI
Béférence.' COOUILLABT (H.) 1981 - L'avifaune des gravières de Bas-en-Basset (Haute-Loirel - Bièvre,3 (1), 55 - 78. - C.O.R.A., Université LYON 1,69622 VILLEURBANNECEDEX.
Résumé; L'avifaune de gravìères, part¡ellement exploitées, situées sur le cours supérieurdelaLoire,enHaute-Loire(France),aétéétudiée,de1973à l979.Desrelevésóevégé-tation ont mis en évidence l'hétérogénéité des milieux, favorable à une avifaune abon-dante et riche, tant en nidif ication qu'en migration ou en hivernage. Le problème duréaménagement et de la gest¡on de ce type de milieu est enfin aborde,Summary..The avifauna of a gravel pit, in the upper part of the Loire valley (France)was studied from 1973 to 1979. Our work has shown that the vegetat¡on is very heteroge-neous. This favours many different çecies of birds, as much during nest¡hg as duringmigratign and wintering periods. The problem of rehabilitation and managãment of th¡skind of landscape is, at last, considered.
| - lntroduction
La HauteLoire est un département essentiellement montagneux (Masifs duMeygal et Monts du Velay). Elle présente malgré tout un certain nombre de zones hu-mides : tourbières, marais, quelques lacs et rives de fleuves (Loire et Allier). Un aperçugénéral des milieux humides de HauteLoire a été donné par COCHET (1979).
De 1973 à 1979, nous avons recensé l'avifaune de l'un d'entre eux : la gravièrede Bas-en-Basset et ses annexes (les prairies humides situées plus au nord : les Gouttes,les Fontaines). 10 à 15 visites annuelles conduiront à un premier bilan. A part¡r de1976, un recensement d'oiseaux d'eau hivernants a été effectué lors de chaque dénom-brement national des Anatidés. Ces relevés quantitatifs réalisés par B. BRUN nous per-mettront une tentative d'interprétation du rôle de la gravière en dehors de la saison denidification. Nous avons effectué quelques relevés de végétation, afin de définir les principaux milieux naturels occupés par les oiseaux.
' La dépression de Bas-en-Basset, Monistrol-sur-Loire (HauteLoire) située à 44O md'altitude, sur le cours de la Loire est une plaine alluviale aux sédiments marno-sableuxdu tertiaire, isolée entre les gorges de Pont de Lignon en amont et d'Aurec en aval. plusau Nord la Loire débouche dans la plaine du Forez, vaste dépression connue pour æsnombreux étangs; Bas-en-Basset est à 28 km de Saint-Rambert, extrémité Sud de laplaine du Forez (fig. 1). Ces gravières semblent donc jouer un rôle de relais pour leso¡seaux traversant le Massif Central.

56
IN
I
Ptlilt 0ut0nrz
\
[r Puy
-Jt'Ji,t;,,
B¡¡-en-8¡¡¡¡t
il0lIst}U
YtytRils
EcHELLE .2o Km,
Figure 1 : Situation des gravières de Basen-Basset sur le cours supérieur de la Loire.
ll - lnventaire des milieux naturels et description générale (fig.2 et 3 - Annexe l)
Nous avons recensé huit types de milieux différents, pour chacun d'eux nousavons dresé la liste des principales espèces végétales (en général seules les plantes supé-rieures ont été retenues). Chaque groupement est décrit brièvement ci-dessous et lesæpèces d'oiseaux nicheurs les plus caractéristiques sont citées.
{a l.rl. a'¡¡.lrltrl¡.¡¡ : Ê¡hl¡, ¡arr¡l¡ ¡r l) !atal a'ararrr
3 ; ¡:li::ì:l:",iiiil ¡f *''|rr' r't.E - l.hl[¡..1t ar !tr¡|atr.F : l.rt.t a'al¡.t¡ ..allla.¡C : l.f|a¡ ¡'il¡¡|t ¡f, ra¡¡.laa¡H : ll.|lr a. t.l.lrI : ¡¡!rtrr,Crll!¡at
Figure 2 : Carte schématique des gravières de Basen-Basset présentant les milieux décrits.(Seule la rive gauche est prise en compte).

5l
| - ?rlrr¡rr
C - Prlrrrr¡ n¡!¡
[ - lrrt.¡ ar! alr¡ta r¡dlfia!r
larl¡r
Figure 3 : Structure de la végétation dans les huit formations définies dans les gra-
vières de Bas-en-Basset :
- l'axe des abscisses montre les surfaces (en pourcentage de la surface totale) occupéepar chaque strate de végétation.
- l'axe des ordonnées, la hauteur de chaque strate, en mètre, selon une échelle géomé-trique.de bas en haut : strates herbacées, buissonnantes, arbustives et arborées.
Les surfaces hachurées schématisent la partie de l'æpace occupée par des végétaux.
I - Salrt¡. Irrlrlr rr
0 - 0.rtr!t. ¡! l.rllh.
-S-
e.n
-a¡ - na¡¡larrtrt¡ aa !.rll¡ta¡
G - lrr¡ar ltt atr¡tt t¡¡ lrllriaat

58
A - Pelouses sèches et landes rases
Etablies sur une cóuche assez importante de sédiments très perméables (sables,graviers). ces formations ont un aspect xérique et paraissent stabil¡sées au stade de pe-louses. Les buissons d'Aubépine et de Prunellier ne semblent pas s'étendre. Les Peupliersdont les racines vont puiser l'eau très profondément, constituent les seules essences li-gneuses notoires. lls expliquent l'occupation de toutes les strates de végétation, avec unfaible pourcentage de recouvrement (fig. 3 A).
Ces pelouses regroupent :
- certains secteurs, bouleversés récemment par des travaux, qui présentent desespèces pionnières (Vergette du Canada, Bouillon blanc).
- d'autres infiuencés par le p¡ét¡nement ou des passages d'engins (on y rencon-tre le Serpolet, le Plantin majeur, le Paturin annuel).
- des creux où s'accumulent des matières organiques et des lieux où ont étédéposés des déchets (près du camping) avec des plantes nitrophiles (l'Armoise vulgaire,I'Ortie dioique, le Chénopode bon-henri).
Les oiseaux les plus caractéristiques sont : l'Alouette lulu, le Bruant zizi et laFauvette griætte qui niche dans les buisons d'aubépine.
B - Zone de galets presque dépourvue de végétation
ll s'agit de la zone située autour des postes d'exploitation y compris les tasde galets fréquemment remaniés par les passages d'engins, à l'exclusion des bancs degalets de la Loire, liés aux fluctuations de débit du fleuve.
Les galets forment un substrat particulièrement impropre à la végétation, quiprésente æulement quelques touffes éparses n'occupant pas plus de la moitié de l'es-pace (cf. fig. 3 B) Bromus tectorum eT. Berterola incana son| les espèces dominantes.
Ce æcteur est hab¡té par le petit Gravelot, dont le mimétisme est remarqua-ble, et le Traquet motteux, qui utiliæ læ tas de galets comme poste de chant. Notonsaussi la Pie grièche à tête rousse, espèce fráquente en milieu méditerranéen aride. Elleprofite ici des quelques bu¡ssons d'Aubépine et de pins rabougris pour nicher.
C - Pelouses rases
Ce milieu est encore plus aride que les pelouæs proches du camping (g A). lls'étend à l'ouest du Gaille (fig.2l près du hameau de Bouzai'. Le tapis végétal presquecontinu est composé d'espèces ayant soit une résistance particulière au desèchement(Lichens), soit des systèmes de rétention d'eau lsedum). ce milieu peut probablement

59
être considéré comme l'aboutissement naturel de la végétation sur une zone de galets
telle que celle citée au $ B.
ll y a quelques années (1973-19741,l'Oedionème nichait sur cette pelouæ, à
l,'époque plus vaste. Celle-ci régresse actuellement, morcelée par l'exploitation des gra-
vières.
D - Bosquets de feuillus \
ll s'agit, sans doute, de lambeaux de la ripisylve existant avant l'installation de
la gravière. C'est un milieu très hétérogène où toutes les strates de végétation sont re-
présentées (fig.2 D). La richesse spécifique importante témoigne d'un milieu biologi-quement riche.
On pourrait distinguer deux types principaux :
- l'un humide, à Aulne glutineux, Saule et Peuplier ;
- l'autre sec, à Chêne sessile, Prunellier et Cornouiller sanguin.
Les trous des troncs morts permettent la nidification de beaucoup de passe-
reaux (cavernicoles> (Mésanges, Grimpereaux). De nombreuses espèces forestières sontprésentes (Accenteur mouchet, Fauvette à tête noire, Pouillot véloce).
E - Reboisement artificiel de conifères
C'est un milieu très homogène avec un boisement dense; les arbres ont tousle même âge. Le sol est recouvert soit de litière, soit de Graminéæ aux endroits suffi-samment éclairés. Ce milieu est caractérisé par un nombre d'espèces d'oiseaux plus fai-ble que celui des bois de feuillus ; toutefois, il est le seul à abriter le Roitelet huppé etla Mésange noire, inféodée aux conifères.
F - Etangs lécents et leurs pourtours
ll s'agit de trous d'eau artificiels ou d'anciens étangs totalement remaniés pourl'exploitation des sédiments. Le niveau de l'eau a baissé dans ceux situés près de laLoire, depuis que le fleuve a été remanié (1). Les berges sont abruptes et la végétationmal implantée. Ceci interdit l'accès de ces étangs à certaines espèces d'Oiseaux que l'onobservera uniquement sur le Gaille non remanié. Cette morphologie des rives est aussi
défavorable aux populations de Poissons. En effet, beaucoup d'espèces ont besoin de
hauts fonds avec des herbiers pour se reproduire et se nourrir (2). Ces rives abruptessont donc la cause d'une faible productivité en Vertébrés (Oiseaux et Poissons).
(1 ) Le fond de la Loire a été surcreusé, donc le niveau de la nappe phréatique qu¡ alimente les étangsa baissé.
(2) Le Brochet, par exemple, ne peut se reproduire que sur les végétaux immergés. De plus, ces herbiersaquatiques sont favorables aux populations d'invertébrés (larves d'l nsectes, Crustacés,.,) dont se nourris-sent les Poissons.

60
G - Marais ou étang non remaniés (secteur du Gaille)
La végétation est beaucoup plus riche et diversifiée que sur les étangs récents.on peut distinguer différentes associations correspondant groisièrement aux ceinturesde végétation des étangs classiquement décrites :
- une étendue d'eau ribre avec ditnrs végétaux immergés ou frottants (Myrio-phylles, Lentilles d'eau...)
- une large ceinture de Typha dans la zone d'atterrisæment du marais, habitéepar la Rousserolle effarvatte et la poule d,eau.
- enfin la sulaie immergée (ce type de végétation très intéressant a malheu_reusement trop souvent été détruit). Ce milieu permet la nidification du Héron biho_reau, espèce qui devient rare à l'échelle de toute notre région et mérite un effort deprotection particulier. ll est donc absolument nécessaire q-u.
"" groupement végétal
soit conservé intact et il ærait souhaitable que l'eau soit mainteñue ious les Saules auniveau que l'on connaissait avant 1g77.
En outre, ce milieu fermé joue un rôle important lors de la migration et deI'hivernage (les Sarcelles d'hiver se posent souvent dans les Typhas, la Bóuscarle est ap-parue plusieurs fois).
H - Bords de Loire
Ce secteur est en perpétuel remaniement.
- la Loire, au courant assez fort, dépose des sédiments de grosse taille (galets),donc seules quelques touffes de Renoncules aquatiques s,installent.'c,est l,habitat duCincle, espèce assez commune.
- les plages de galets sont favorables au Chevalier guignette.
- l'érosion des rives concaves provoque l'apparition de falaises de sable où ni-chent souvent de petites colonies d,Hirondelies de rivage.
- les talus en bordure de la Loire sont prantés de grands peupliers occupés parla Huppe et le Loriot.
| - Prairie humide (bocage) au nord de la gravière
cette zone est citée, malgré r'absence de rerevés de végétation, car eile com-plète la mosaiþue des milieux rencontrés sur les gravières elles--mêmes et abrite cer-taines espèces d'oiseaux caractéristiques. c'est le-cas du vrnn"áu qri y niche. on yrencontre fréquemment des Bécassines des marais et des Busards Såint-Martin et cendré.Le Courlis corlieu a aussi été observé en ces lieux.

61
lll - Nombre d'espèces d'oiseaux recensées et cycle annuel de l'avifaune
lll-l - Oiseaux recensés sur la gravière
, La listedes oiseaux nicheurs et non nicheursestdonnée en annexe ll. La listedæ nicheurs (76 æpèces) n'est encore pas complète, malgré les observations régulières
depuis 6 ans. Nous poursuivons actuellement nos observations afin de prouver la nidi-fication de quelques espèces supplémentaires.
D'autres espèces non nicheuses doivent être recherchées :
Oies, Anser sp. (en hivernage, citées par un chasseur)- Faucon hobereau, Falco subbuteo
Perdrix rouge et Perdrix griæ, Alectoris rufa et Perdrix perdrixPluvier doré, Pluvialis apricariaCourlis cendré, Numerius arquataChevalier sylvain, Tringa glareolaGuifette moustac, Chlidonias hybridaChouette effraie, T¡rto albaEngoulevent d'Europe, Caprimulgus europaeusBec-croisé des sapins, Loxia curvirostrà.,.
Depuis cinq ans, deux espèces ont disparu des gravières de Bas-en-Basæt:
- le Faucon pélerin qui a totalement disparu des gorges de la Loire. ll venaitparfois chasser sur les gravières en 1973-11974.
- l'Oedicnème criard nicheur jusqu'en 1974.
Sur les 141 espèces enregistrées, 76 soit 54 % nichent dans la plaine de Bas'
en-Basset ; 71, soit 50 %, nichent dans la gravière au sens strict, parmi ces dernières,65 soit 46o/o se reproduisent chaque année depuis le début de cette étude. Les 65 es-
pèces non nicheusæ, soit 46 % du total, se répartisænt en 31 soit22 % d'accidentels(observés une fois ou deux sur la gravière) et 34, soit 24 % d'oiæaux observés régu-
lièrement.
lll-2 - Comparaison de l'avifaune de Bas-en-Basset à celle de régions proclres
La plus proche région présentant des milieux aquatiques ¡mportants est laplaine du Forez. Nous comparons donc les avifaunes de ces deux districts tout en res-
tant conscients de la différence de superficie (Forez :500 km2 - plaine de Bas-en-
Basæt : 15 km2 environ). Nous comparons aussi l'avifaune de Bas-en-Basset à celle duPilat, massif cristallin proche, où les milieux aquatiques sont très peu représentés (don-
NéCS dC LEBRETON Et FAURE 1969, LEBRETON Et AI. 1972, LEBRETON 'I977).

62
Nombre d'espèces nicheuses non nicheuses tota I
Bas-en-Basset
Pilat
Forez
76
104
122
65
40
85
141
144
207
Le nombre d'espèces nicheuses apparaît asez faible, ce qui était prévisiblecompte tenu de la faible superficie de la plaine de Bas-en-Basset. on remarque que lenombre d'espèces non nicheuses est important, supérieur à celui du pilat, mais infé-rieur à celui du Forez.
Toutes les espèces rencontrées à Bas-en-Basset l'ont été aussi en Forez, à uneseule exception près: le Courlis corlieu dont trois individus ont été obærvés le 11avril 1976. C'esr la seule citation dans la région. LHERITIER (19741a étudié un mi_lieu comparable en HauteLoire, sur I'Allier : la Limagne brivadoise. Son aire d'étudeest plus importante et inclut certains milieux collinéens; il note 108 espèces nicheusesdont 82 occupant les biotopes proches de l'Allier (grèves, berges, bras-morts, ripisylves).Ce résultat semble tout à fait comparable à celui obtenu à Bas-en-Basset (76 eipèäesnicheuses). Les différences sont liées:
- à I'absence dans le secteur que nous avons étudié, de trois espèces : leButor étoilé Botaurus stellaris qui a niché une année dans l,Allier, le phragmite desjoncsAcrocephalus schoenobaenus et le Gobe-mouche gris Muscicapa striata.
- au doute concernant la nidification de certaines espèces à Bas-en-Basset :
Pipit des arbres, Pie-Grièche grise, Torcol, Bruant jaune, Bruant des roseaux, Râled'eau (cf. époques de présence dans l'annexe ll).
. Ouelques espèces (Rouge-queue noir, Traquet motteux, Foulque) ne nichentqu'à Bas-en-Basset.
lll-3 - Nidif ication
Les 76 espèces nicheuses se répartissent en 55, soit72 % d'oiseaux migrateurset 21, so¡t 28 o/o de sédentaires et erratiques. Le pourcentage relativement fort d,oi-æaux migrateurs n'est pas surprenant dans un milieu pionnier offrant des ressourcesen nourriture concentrées, tant dans I'espace que dans le temps (1). Dans les milieuxtrès stables (forêt) le pourcentage d'oiseaux sédentaires est généralement supér¡eur.
(1) Des explosions de population de-plantes pionnières, d'insectes, provoquent l,apparition rapide deressources trophiques très v¡te épuisées.

63
Le régime alimentaire des oiseaux nichant à Bas-en-Basset montre une nette do-minance des espèces insectivores par rapport aux trois autres catégories (carnivores : ra-paces et oiseaux piscivores, polyphagæ et granivores)(fig. a).
- la forte production biologiquedes lisières milieu aquatique - milieu terrestre(marais, saulaies) profite à de nombreuses espèces :
. oiseaux aquatiques (canards, foulques) æ nourrissant dans l'eau
. espèces se nourrissant dans la végétation (Fauvettes, Mésanges)ì espèces se nourrissant au vol (Hirondelles, Martinets) qui consomment les insec-
tes éclos en milieu aquatique.
llir!rrrrlr !t I l¡r¡t!rr!
Figure 4 : Régime alimentaire des oiseaux ob-servés sur les gravières de Bas-en-Basset(Pourcentage de chaque groupe par rapportau nombre total d'espèces)
I : insectivoresC : carnivores (prédateurs de vertébrés)G : granivoresP : polyphages
Une mention particulière est réservée aux rapaces qui nichent sur les côteauxautour de Bas-en-Basset et viennent se nourrir sur la gravière :
. les Milans noir et royal, qui consomment surtout des cadavres et des déchets
. le Circaète Jean-le-Blanc, prédateur des Reptiles et des batraciens
. l'Epervier et l'Autour qui chassent des oiseaux
. la Bondrée apivore qui se nourrit d'insectes.
La moæiþue des milieux explique la présence d'espèces ayant des exigences écologiques très différentæ. On remarque la nidif ication des trois espèces de Traquet à quel-ques centaines de mètres de distance les uns des autres :
le Traquet tarier, habituellement préænt dans les prairies humides (d'altitudesurtout)
. le Traquet pâtre, caractéristique des bocages et des landes
. le Traquet motteu& nichant dans les régions arides où l'élément rocheux voisineavec des milieux herbacés ras (éboulis de montagne, rochers maritimes...).
lci, cæ espèces occupent respectivement les prairies humides autour de la gravière,les landes sèches près du camping et les monticules de galets du poste d'exploitation.

64
Dans la famille des Sylviidés, on observe un phénomène æmblable : la Fauvettegrisette, la Fauvette à tête noire, la Fauvette des jardins et la Rouserolle effarvatte quis'échelonnent habituellement des milieux ouverts et secs aux milieux fermés et humides,nichent ici à moins de cent mètres les unes des autres.
Dans ces gravières on peut multiplier les exemples de ce type. Remarquons aussile contraste entre les æpèces d'origine septentr¡onale évoquant pour l'ornithologue laforêt médioeuropéenne (Bouvreuil, Rouge-gorge, Sittelle, Verdier) et les espèces méridionales évoquant souvent des milieux plus ouverts (Huppe, Pie-grièche à tête rousse, Loriot).
Signalons, enfin, la nidification régulière d'espèces rares dans la région :
.. Hirondelle de rivage. Piegrièche à tête rousse. Martin-pêcheur, plus abondant que partout ailleurs dans les départements de la
Loire et de la Hauteloire. Héron bihoreau, æpèce qui a toujours été très localisée et subit actuellement
une régression alarmante.
Malgré le manque de données quantitatives concernant la nidification, nous allonstenter de situer les gravières de Bas-en-Basset par rapport aux 1 1 gravières et aux 35 étangsétud¡és par le C.O.R.A. (BOURNAUD 1978). La nidification de la Foulque, du Canardcolvert, ela fortíorí du Héron bihoreau, incite à situer le secteur étudié au sein du groupedes étangs plutôt que parmi les gravières. Les espèces régulièrement rencontrées sur lesgravières (Cincle, Chevalier guignette, Martin pêcheur, Poule d'eáu...) sont aussi présentesà Bas-en-Basset. En fait, il semble impossible de situer globalement ces gravières dans legradientdéfiniparBOURNAUD(1978),desgravières récentes,auxétangspermettantI'installation de colonies de Mouettes rieuses. ll serait plus réaliste de tenter de classerchaque petite surface d'eau élémentaire, depuis les bassins profondément remaniés, prèsdu poste d'exploitation, jusqu'aux étangs abritant la colonie de Hérons bihoreaux et denombreuses Rousserolles. Ceci n'est pas réalisable à cause du manque de précision desdonnées, récoltées au niveau de l'enæmble de la gravière et de l'aspect très morcelé de cesplans d'eau. Cette tentative de comparaison met encore une fois en évidence l'hétérogênéité qui confère un intérêt particulier à ces gravières.
ll14 - Migration et hivernage
Plusieurs migrateurs rares ont été observés:Balbuzard, Garrot à æil d'or,Bécasseau de Temmink, Chevalier arlequin, Guifette leucoptère. Liés au milieu aquati-que, ils font escale sur la gravière.
Contrairement à ce qui a été observé pour les nicheurs, on ne remarque pasla dominance d'un type de régime alimentaire par rapport aux autres (fig. 4). Les in-sect¡vores et les polyphages sont relativement nombreux (37 % et 35 % de l'ensemble),

65
alors que les granivores sont peu représentés (8 %). L'importance des carnivores est liée
aux nombreux oiseaux piscivores qui font halte sur la gravière (Hérons, Grand Cormoran,Guifettes et Sternes, Balbuzard...).
Si beaucoup d'espèces préæntent, en hiver, sur la gravière, des effectifs faibles,la masse des hivernants est toujours constituée par quelques troupes d'oiæaux grégaires(Alouette des champs, Grive litorne, Pinson des arbres, Pinson du Nord, Moineaux do-mestiques et fr¡quets, Etourneaux).
La figure 5 permet de visualiser les fluctions moyennes des effectifs de quel-ques espèces d'anatidés et de limicolæ, au cours de l'hiver. Le tableau 1 préænte ces
résultats pour les trois années (1976, 1977 , 1978) pendant lesquelles des comptagesont été effectuéS régulièrement à Basen-Basset.
Les effectifs de Canard colvert, assez abondants en hiver, semblent diminuerfortement au mois de mars, époque à lquelle la nidification est commencée pour cetteespèce. De 1976 à 1978, la population hivernante de cette espèce á doublé (27 indivi-dus en moyenne pendant l'hiver 1976 et 67 actuellement). Ce résultat a été obtenugrâce à la mise en réserve de chasæ de la gravière.
Les populations de Foulques préæntent des effectifs assez constants en hiver.
Les Sarcelles d'hiver sont présentes pendant toute la saison hivernale. Cetteespèce ne niche pas dans la région.
D'autres espèces d'Anatidés séjournent irrégulièrement pendant l'hiver (Milouin,Chipeau...). En conséquence, læ gravières de Bas-en-Basset jouent un rôle (il est vraimodeste) de reposoir hivernal, surtout depuis la mise en réærve de ctlasse.
La Bécassine des marais æmble surtout abondante au passage d'automne, trèséchelonné (de septembre à décembre) et au passage de printemps (février, mars). Onpeut favoriser le stationnement de cette espèce en maintenant certaines zones immer-gées dans quelques centimètres d'eau seulement (HARRISON 1974).
Le Vanneau huppé rare en hivernage, apparaît en février, mars. ll niche dansles prairies humides au nord de la gravière.
La Mouette rieuse est préænte uniquement pendant la migration (février, mars),la gravière jouant le rôle de relais sur son trajet (quelques individus non nicheurs fré-quentent les lieux tout l'été).
Ces quelques exemplæ montrent le rôle actuel de la plaine de Bas-en-Basset pourles oiseaux d'eau migrateurs ou hivernants, ils font encore bien plus apparaÎtre les po-
tent¡alités d'un tel milieu.
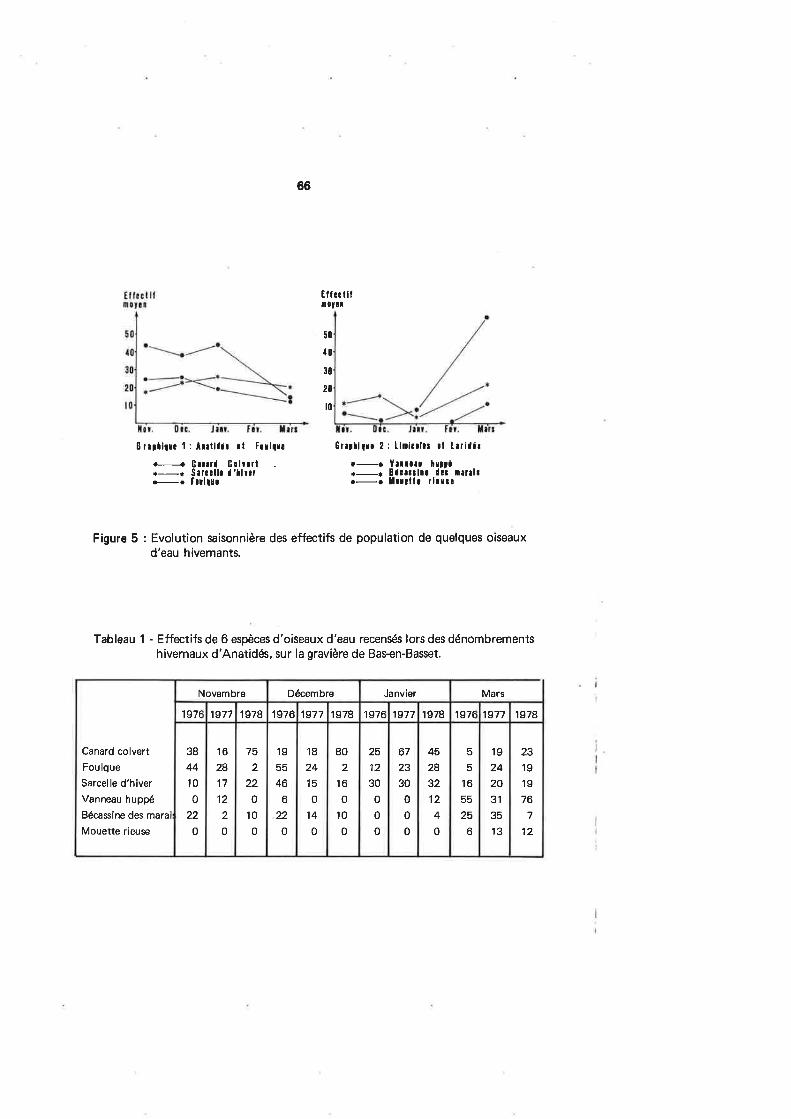
66
tll!c t lllr0trr
50
t0
30
20
t0
G r¡tlitrr I : Ar¡tilat rl frrl¡rrr--------¡ C¡r¡rl G¡lr¡rt*-* S¡rr!lh a'llrtr+o frrl¡lr
Êr¡tlilr¡ 2 : Ilnicrl¡¡ ¡t [¡rili¡o-. Y¡rrl¡I hlrrar_* Ba3trrlta d!¡ n¡t¡i¡¡-¡ l¡¡¡ll¡ ;l¡¡¡¡
Figure 5 : Evolution saisonnière des effectifs de population de quelques o¡seauxd'eau hivernants.
Tableau 1 - Effectifs de 6 espèces d'oiseaux d'eau recensés lors des dénombrementshivernaux d'Anatidés, sur la gravière de Basen-Basset.
Canard colvert
Foulque
Sarcelle d'hiver
Vanneau huppé
Bécasine des marai
Mouette rieuse
Novembre Décembre Janvier Mars
197( '1977 '1978 1976 1977 1 978 1976 1977 1978 197f 1977 1978
38
44
10
0
22
0
't6
28
17
12
2
0
75
2
22
0
10
0
19
55
46
6
22
o
18
24
15
0
14
0
80
2
16
0
10
0
25
12
30
o
0
o
67
23
30
0
0
0
45
28
32
12
4
0
5
5
16
55
25
6
19
24
20
31
35
13
23't9
19
76
7
12

67
lV - Réaménagement et protection.du milieu
Nous allons envisager successivement différents points quí méritent une atten-tion particulière lors d'un réaménagement de la zone d'extraction après la fin dæ tra-v,aux.
lV-l - Problèmes hydrologiques
Depuis que l'on extrait des graviers dans le lit mineur de la Loire, le niveaude la nappe phréatique baisse de plus en plus et on peut craindre que certains étangsne se trouvent aséchés. ll pourrait être nécesaire d'alimenter artificiellement la gra-
vière en eau, grâce à des prélèvements effectués dans la Loire, près du camping. ll esttrès important de réalimenter en eau la saulaie où se trouve la colonie de Hérons Biho-reaux ($ l-G).
lV-2 - Aménagement des étangs creusés
Plusieurs buts peuvent être poursuivis :
- maintenir et améliorer læ populations de poisons et les conditions depêche ;
- favoriser l'hivernage des Anatidés ;
- favoriser la nidification des Oiseaux.
Ces différents buts peuvent être conciliés, comme le prouve l'expérience bri-tannique dans la réserve de Sevenoaks (1). Le WAGBI (1) a réusi à aménager, dansune ancienne gravière, un milieu favorable au gibier d'eau, en toute saison, tout en
maintenant une activ¡té de pêche à la ligne importante (HARRISON 1974).
ll faut augmenter au maximum l'hétérogénéité des berges (obtenir une longueurde berge maximale pour une surface d'eau donnée). (ARRIGNON 1975 - F.R.A.P.N.A.1978).
ll peut être utile de créer des jetées de terre s'avançant sur les étangs et des
îles pour augmenter les populations d'Anatidés nicheurs, en particulier de Canards
colverts. En effet les mâles défendraient un terr¡toire défini par leur champ de visionsi celui-ci est lim¡té par une butte de terre ; une population plus importante peut ni-cher sur une même surface d'eau (d'après HARRISON 1974).
ll est important que certaines berges aient une pente douce pour que les végé-
taux supérieurs émergés et immergés puissent s'y installer (BUGNON et FELLRATH,1979).
(1) WAGBI : Wildfowler's Association of Great Britain and lreland. Association de chasseurs de gi-bier d'eau qui gère la réserve de Sevenoaks sur une gravière,

68
sí possible, íl est souhaitable qu'une telle pente douce soit créée avec de laterre arable ou de la vase pour faciliter la croissance des végétaux. ll peut être utile deplanter certaines æpèces végétales devant coloniser normalement les bordures de plansd'eau, pour fixer les bergæ. Les peupliers colonisant facilement les gravières et fréquem-ment plantés æmblent derúoir être évités. La décomposition des feuilles qui tombentdans l'eau dès l'été consomme beaucoup d'oxygène, ce qui asphyxie ra faune aquatiqueet provoque une eutrophisation accélérée du plan d'eau. (TREMOLIERES et GARBIE-NER 1978, DELBECOUE et at. 19791.
D'autres berges, sableuses, devront rester abruptes pour permettre la nidifica-tion du Martin-pêcheur et des Hirondelles de rivage.
ll æt important d'aménager des postes de pêche sur certaines rives tout enrendant d'autres zones (rives très favorables à la nidification) inaccessibles. Ce type deréaménagement des gravières favorise l'installation de l'avifaune comme en témoigneles résultats de FROCHOT et al. (1979) dans le val de Saône.
ll est utile qu'une grande surface d'eau libre demeure sur l'un des étangs lesplus vastes de la gravière. Ceci joue un rôle attractif sur les canards de passaç ou hi-vernants qui ont besoin d'aires de repos.
En un mot, il faut maintenir ou accroltre encore l'hétérogénéité de l'écosys-tème.
lV-3 - Protection du secteur du Gaille
. Le tracé sinueux des berges préconisé cidessus subsiste dans le secteur duGaille (fig. 2). Ces confluents et ces bras morts présentent une hétérogénéité remarquable, justifiant pour une grande part l'intérêt général de la gravière.
une mention particulière doit être réservée à la saulaie, jadis immergée, oùniche la colonie de Hérons bihoreaux. En effet, de telles formations sont assez raresdans la région. En Forez, il n'y a que deux ou trois saulaies assez vastes pour accueil-lir une telle colonie.
Ce mil ieu doit absolument être épargné par les extractions de granulats. ll jus-tifie des mesures de protection particulières, évitant tout dérangement. Ceci peut êtrefait en plantant des ceintures denæs de plantes peu franchissables (aubépines, ronces).
lV-4 - Aménagements divers
ll est indispensable d'éliminer la décharge d'ordures. De moins en moins con-trôlée, elle s'étend sur une grande partie de la gravière lors des crues; BRULAT (1976)fait remarquer :

69
- l'aspect inesthétique de tels dépôts,- la pollution de la nappe phréatique et probablement des cours d'eau pro
ches, du fait des infiltrations d'eau de pluie au niveau des décharges. Celleci est établiesur des sédiments sablonneux, très perméables, qui ne peuvent pas offrir toutes læ ga-
rant¡es nécessaires pour le maintien de la qualité de l'eau.
ll faudrait pouvoir circonscrire dans l'espace, si ce n'est dans le temps, certainesactivités nuisibles à l'environnement (trial, etc...) et peut-être canaliser les voitures danscertains secteurs (FROCHOT et al. 19791.
Certains aménaçments fins pourraient être envisagés :
- postes d'observation des oiseaux,- circuits d'initiation à la botan¡que...
lnsistons sur l'intérét de l'aménagement des plans d'eau artificiels (barrages etgravières) pour l'accueil du gibier d'eau, ceci afin de pallier, dans une certaine mesure,à la destruction continue des milieux palustres naturels (TAMISIER, 1979). Espéronsque telle sera la vocation de la gravière de Baven-Bæset où, malgré l'exploitation, unmilieu naturel particulièrement riche a pu subsister.
V - Conclusion : intérêt de la gravière
Les résultats présentés cidesus nous permettent de mettre en évidence et d'ex-pliquer l'intérêt biologique de ce milieu. La diversité spécifique de l'avifaune est remar-quable en toute saison. Un nombre important d'espèces rares frfiuentent la gravière,
tant en période de nidification que de migration ou d'hivernage. Cette zone est un re-lais sur le trajet migratoire de nombreux oiseaux. Elle joue, sans doute, aussi un rôletrophique pour ces espèces de passaç, ainsi que pour d'autres nichant aux alentours.
La richese avifaunistique s'explique par la juxtaposition des milieux très con-trastés :
- par leur hydrographie (étang, marais et pelouses sèches)- par leur topographie (landes plates, talus, tas de graviers)- par leur végétation qui intègre les 2 points cidessus.
Cette hétérogénéité, liée à un effet de mosaiþue (morcellement important dessurfaces occupéæ par chaque groupement végétal) crée l'intérêt de la gravière. Ce con-traste entre des biotopes variés révèle la position de carrefour entre les avifaunes médio-européenne et méditerranéenne qui est une caractéristique essentielle de l'enæmble dela région.

70
Vl - Remerciements
Nous tenons à remercier les membres du C.O.R.A. et du C.O.A. qui ont col-laboré à cette étude, en particulier B. BRUN et R. FAURE qui ont participé active-ment au travail de terrain, G. COCHET, A. DELPUECH, G. TERROT et Y. THON-NERIEUX qui ont fourni des observations, S. AULAGNIER, J. BROYER et J.D.LEBRETON qui ont apporté des compléments d'information et relu le manuscrit.
Vll - Bibliographie
ARRIGNON (J.) 1975-Aménagementp¡sc¡coledescarrièreshumides. Butt. lnfo. C.S.P. 101,3345,BOURNAUD(M.) 1978-L'eçaceétangdanssesrapportsavecl'avifauneenpériodedenidification.
Ed. C.O.R.A., Ministère de l'environnement, Tg p.
B R U LAT (F. F.) 1975 - Les balastières dans la vallée de la R iste. Butl. tnfo. C.S.p. 1O9,21-92.BUGNON (F.), FELLRATH (M.) 1979 - Rapport sur la végétation des gravières de la plaine de la
Tille, Etude écologique des sablières de la plaine de Saône bourguignonne ei perspectivesd'aménagements. Ed. Greminat lJniv. Dijon, 9-15.
CLAVEL (P.), CU INAT (R.), HAMON (Y.), ROMANEIX (C.) 1977 - Effet des extractions de maté-riaux alluvionaires sur l'environnement dans le cours supérieur de la Loire et de I'Allier,Bull. Fr. Pisci.50 e68l ,122-154.
COCHET (G.) 1979 - Les zones humides de la Haute-Loire. Nature 43.5,17-19,DELBECOUE (J.P.), FROCHOT (8.), PAGES (J.), ROLL atique des
gravières de la plaine de Tille. Etude écologique ône bour-guignonne et perspectives d'aménagements. €d.
FOUBN IE R (P.) f 961 - Les quatre flores de Fnnce. Ed. Le Chevalier, Paris, l 1 0b p.
F.R.A.P.N,A, 1978 - Les carrières humides, agressions potentielles, aménagements possibles aprèsexploitation. La Loíre et I'aménagement du Bassin ligérien. lJniy. Sa¡nt-Etienne,212-219.
FROCHOT (8.), LACROIX (P.), MESNIEB (D.ì 1979 - Les oiseaux des gravières de la plaine deSaône. Etude écologique des sablières de la plaine de Saône bourguignonne ei perspectivesd'a ménai¡emen Ts. Ed, G re m i na t tJ n i v. D ij o n', 4 1 -SZ,
HABBISON (J.l 1974-TheSevenoakgravel pitreserve.A WAGBtConservatíonpublication, 116p.LEBRETON (J.D.), FAURE (R.) 1973 - Les oiseaux des gorges de la Loire. Doc. polycopié c.o.R.A.,
4p.LEBBETON (Ph.) 1977 - Aths nhône-Alpes des oiseaux nicheurs. Ed, C.R.D,p. Lyon,3b3 p.
LEBRETON (P.),feUnp (J.M,) 1969 - Contribut¡on à la connaissance avifaunistique de la plaine duForez, Centre d'Etudes Foréziennes ; Mélanges:255-28g.
LEBRETON (Ph.),THEVENOT (M.), LEBRETON (J.D.), BRAEMER (H.) 1972 - Etude ornitholosi-quedu massif du Pilat lLoirel.Atauda,39 (4),255-289 er 40 (ll,97-52.
LH ER lTl ER (J.N.) 1974 - L'avifau ne nicheuse de la L imagne brivadoise (Haute-Loirel. A lauda, 42(1),79-110.
TAMISIER (4.) 1979 - Réalités biologiques et gestions des populations de gibier : avenir de la chasseau gibier d'eau, Courriar Naturc, b9, 1-10.
TREMOLIEB'.ES (M.), CARBINIER (R.) 1978 - Mesures de l'activité anti-oxygène de macérats defeuilles d'automne dans les eaux de zurface, essai d'évaluation de l'impact écologique. C.Fl.Aæd. Sci., 286, 17 19-1722.

71
ANNEXE I : Liste des plantes rencontrées sur les gravières de Bas-en'Basset
Elle n'est pas exhaustive, nous avons noté les espèces les plus remarqua-
bles lors d'un seul relevé effectué pendant l'été 1978.
La nomenclature est celle de FOURNIER (1961).
M ILIEUX
ESPECES
oEo=tsøoo,o
o
9@
CL
oo
oo
aÐó
ÐG
o
IoCL(.)
ooEoE
oCD
c¡
o,o=oo@
=too.2o.E¡oCEt¡J
Ëo0\o
oõ
t¡JlJ-
p6Eo
o
Ðet¡¡(j,
o':oJsoEooo64
ESPECES LIGNEUSES
GYMNOSPERMES
Pseudotsuga douglasii : DouglasPicea exceln : Epicéa
Pinus laricio : Pin noirPinus sylvestris : Pin sylvestre
DICOTYLEDONES
Rubus sp. : BonceRon sp. : EglantierPrunus spinoats : PrunellierCrataegus oxyacantha : AubépineSarothamnus scoparius : Genêt à balais
Cornus sanguinea : Cornouiller sanguin
Evonymus europaeus : FusainFraxinus excelsior : FrêneAlnus glutinosa : AulneOuercus robur sessiliflora : Chêne sessile
Corylus avellana : NoisetierSalix sp. : Saule
Populus nigra : Peuplier noirHumulus lupulus : Houblon
a
aaaa
aaaa
oaa
aoo
a
aoaa
aa
aa
oa
aa
aa
o
o
aa

72
Hedera helix : LierreAcer pæudoplatanus : Erpble sycomoreLigustrum vulgare : TroëneLonicera periclymenum : ChèvrefeuilleSambucus nigrum : Sureau noir
ESPECES HERBACEES
MONOCOTYLEDONES
Typha latifolia : RoseauPhalaris arundinacea : BaldingèreBromus tectorum : Brome des toitsLolium perenne : lvraieBrachipodium pinnatum : BrachipodeAgropyrum repens : ChiendentPoa annua : Paturin annuelGraminées non déterminéesLemna minor : Lentille d'eauJuncus effusus : Jonc épars
Juncus glaucus: Jonc glauqueAllium sp. : Ail
DICOTYLEDONES
Urtica dioica : OrtieRumex crispus : ParelleRumex acetoe : OseillePolygonum cuspídatum : RenouéePolygonum aviculare : Renouée des oiseauxPolygonum persicaria : Renouée persicairePolygonum amphibium : Renouée amphibieChenopodium bonus-henricus : Epinard sauvage
Chenopodíum albumAmarantus hybridusMelandryum rubrumSaponaria officinalisFicaria ranunculoídes
Epinard: AmaranteCompagnon rouge
Saponai re
FicaireBatrachium sp. : Renoncule aquatiqueAlliaria officinalis : AlliaireBerterola incana : Crucifère blanche
ooa
oO
oO
oo
O
o
oao
oo
oo
oo
oo
ooo
oo
a
oo
ooo o
ooa
O
O
ao
o
o a

73
ooaooaaa
o
o
LICHEN
Cladonia sp. a

74
Resda luteola : Réséda jaune
Sedum album '. Sédum blancSdum acre : Poivre de murailleGeum rurale : BenoiteAgrimonia eupatoria : AigremoineSpiraea ulmaria : Reine des prés
Trifolium pratense : Trèfle des prés
Trifolium repens : Trèfle rampantMelilotus albus '. Mélilot blancMedicago ætiva : LuzerneOnonis spinoæ : Arrête bæufLotus corniculatus : LoTier
Oenothera biennis : Onagre bisannuelleEpilobium hirsutum : EpilobeMyriophyllum sp. : MyriophyleGeranium robertianum ; Géranium herbe à RobertDaucus carotta : CarotteLysimachia vulgaris : grande LysimaqueConvolvulus æpium : Liseron des haiesPulmonaria officinalis : PulmonaireMyosotis palustris : MyosotisEchium vulgare : VipérineVerbascum sp. : MolèneSolanum dulcamara : DouceamèreEuphrasia stricta : EuphraiseLamium maculatum : LamierThymus serpyllum : SerpoletMentha rotundifolia : Menthe à feuille rondeSalvia officinalr's : Sauge
Plantago major : Plantain majeurPlantago lanceolata : Plantain lancéoléGalium verum i Gaillet jaune
Galium aparine : GratteronGalium mollugo : GailletDipsacus silvestris : Cardère
Scabios columbaria ; ScabieuseEryngium campestre : Panicaut champêtreBryonia dioi'ca : BryoneErigeron canadensis: Vergerette du Canada
Cirsium nutans : Chardon penché
o oao
O
oooooaaooo
o
ooo
oaaooO
oaaatao
o
o
a
O
a
o
o o
o
o
ooo
oO
ao
aao
o o
oo
o
oO
a O
oo
oa
oo

75
ANNEXE ll :Epoque de présence des oiseaux sur la gravière de Bascn-Basset
Ce tableau présente la liste complète des oiseaux observés à Bas-en-Basset
depuis 6 ans.
, Chaque point correspond à une observation effectuée par l'un des observa-teurs, ou à une série d'observations répétées d'année en année aux mêmes dates.
Les traits continus correspondent à une présence constante de l'espèce pen-
dant la période de temps concernée, les traits pointillés indiquent une présence irrégu-lière ou douteuse.
espèce nicheuseespèce présente en migrationespèce présente en hivernageespèce présente toute l'année
Grèbe à cou noir Podiceps nigricollisGrèbe castagneux Pod iceps ruf ico I I is
G rand cormoran Phalacrocorax carboAigrette garzette Egretta garzetta
Héron cendré Ardea cinerea
Héron pourpré Ardea purpurea
Héron bihoreau Nycticorax nycticoraxCanard colvert Anas platyrhynchosSarcelle d'hiver Anas crecca
Sarcelle d'été Anas querquedula
Canard siffleur Anas penelope
Canard chipeau Anas strepenCanard pileT Anas acutaCanard souchet Anas clypeataFuligule milouin Aythya ferinaFuligule morillon Aythya fuligulaGarrot à æil d'or Bucephala clangulaBuse variable Buteo buteo
Légende I !
H
HP
P
P
H
P
N
C
HP
P
HP
P
P
HP
P
P
P
C

76
Epervier d'Europe Accipiter nisusAutour des palombes Accipiter gentilisMilan royal Milvus milvusMilan noir Milvus migransBondré apivore Pernis apivorusBusard Saint-Martin Circus cyaneusBusard cendré Circus pygargusCircaète jean-leblanc Circaätus gal licusBalbuzard pêcheur Pandion haliaëtusFaucon pélerin Falco perqrinusFaucon crécerelle Falco tinnunculusCaille des blés Coturnix coturnixFaisan de colchide Phanius colchiusRâle d'eau Rallus aquaticusPoule d'eau Gallinula chloropusFoulque macroule Fulica atraVanneau huppé, Vanellus vanellusPetit gravelot Charadrius dubiusBécassine des marais Gallinago gallinagoCourlis corlieu Numarius phaeopusBarge à queue noire Limosa limoæCheval ier aboyeur Tringa nebu leriaCheval ier culblanc Tringa ochropusChevalier arlequin Tringa erythropusChevalier guignette Tringa hypoleucosBécasseau de Temminck Calidris temminckiiOed icnème criard Burh inus æd icnemusGoéland argenté Larus argentatusMouette rieuæ larøs ridibundusGuifette noire Clidonias nigerGuifette leucoptère Clidonias leucoptrusSterne pierregarin Sterna hirundoPigeon colombin Columba ænasPigeon ramier Columba palumbusToureterelle des bois Streptopelia turturCoucou gris Cuculus canorusHibou moyen-duc Asio otusChouette chevêche Athene noctuaChouette hulotte Stríx alucoMartinet noir Apus apus
HP
HP
PN
N
N
HP
PN
N
P
(P)
C
N
C
PH
cc
N
HP
P
P
P
P
P
N
P
N}
P
P
P
P
P
HP
C
N
N
N
N
N?
N

77
Martin pêcheur Alcedo atthisHuppe fasciée Upupa epops
Pic vert Picus viridisPic épeiche Dendrocopos majorPic épeichette Dendrocopos minorTorcol fourmilier Jynx torquillaAlouette lulu Lullula arboreaAlouette des champs Alauda arvensis
Hirondelle de rivage Riparia ripariaHirondelle de rocher Ptyonoprogne rupestrisHirondelle de cheminée Hirundo rusticaHirondelle de fenêtre Delichon urbicaPipit roussel ine Anthus campestrisPip¡t des arbres Anthus trivialisPipit farlouse Anthus pratensisPip it. spioncelle A nthus spi no lettaBergeronnette printanière Motacilla flavaBergeronnette des ruisseaux Motacilla cinereaBergeronnette grise Motaci lla a lba
Pie-grièche écorcheur Lanius collurioPiegrièche à tête rousse Lanius senatorPiegrièche grise Lanius excubitorCincle plongeur Cinclus cinclusTroglodyte Troglodytes troglodytesAccenteur mouchet Prunella modularisTraquet Tarier Saxicola rubetraTraquet pãf.re Saxicola torquataTraquet motteux Oenanthe ænantheR ougequeue noir Phoenicu rus ochrurosBougequeue à front blanc Phoenicurus phoenicurusRougegorge Erithacus rubeculaRossignol philomèle Luscinia megarhynchosGrive litorne Turdus pilarisMerle noir Turdus merulaGrive mauvis Turdus iliacusGrive musicienne Turdus philomelosGrive draine Turdus viæivorusBouscarle de cetti Cettia cettiRousserolle effarvate Acrocephalus scirpaceusHypolai's polyglotte Hippolais polyglotta
C
N
C
cC
N?
PN
C
PN
PN?
PN
PN
P
PN?
HP
HP
P
cC
N.N
C?
PH
cC
N
PN
N
N
N
cN
H
C
H
PH
PH
P
N
N?

78
Fauvette des jardins Sylvia borinFauvette à tête noire Sylvia atricapillaFauvette grisette Sylvia communisPou i I lot filis Phylloscopus trochi lusPouil lot véloce Phyl loscopus collybitaPouillot de Bonelli Phylloscopus bonelliRoitelet hvppé Rqulus rqulusRoitelet triplebandeau Rqulus ignicapillusGobemouche noir Ficdula hypoleucaGobemouche gris Musciapa striataMésange à longue queue Aegithalos caudatusMésange nonnette Parus palustrisMésange boréale Parus montanusMésange huppée Parus cristatusMésange noire Parus aterMésange bleue Parus caeruleusMésange charbonnière Parus majorSittelle torchepot Sitta europaeaGrimpereau des jardins Cenhia brachydactylaBruant proyer Emberiza calandraBruant jaune Emberiza citrinellaBruant zizi Emberiza cirlusBruant fou Emberiza ciaBruant des roseaux Emberiza schoeniculusPinson des arbres Fringilla coelebsPinson du Nord Fringilla montifringillaVerdier Carduelis chlorisCha rd onnerel Ca rd ue I is ca rduel isTarin des aulnes Carduelis spinusLinotte mélodieuse Carduelis cannabinaSerin cini Serinus serinusVenturon montagnard Serinus citrinel laBouvreuil pivoine Pythula pyrrhulaG ros-bec Coccothraustes coccothraustesMoineau domestique Passer domesticusMoineau friquet Paser montanusEtourneau sansonnet Sturnus vulgarisLoriot jaune Oriolus oriolusGeai des chênes Garrulus glandariusPie bavarde Pica picaChoucas des tours Corvus mondulaCorneille noire Corvus corone coroneGrand corbeau Corvus corax
N
PN
N
N?PN
P
PN
P
P
P
cC
H
c?cccC
CN
PH
cH
HPC
H
C
C
H
PN
PN
H
C
PH
cC
cN
C
C
PNcP

7S
François J. BAUD
CONTRIBUTION A LA CONNAISSANCE DU REGIME ALIMENTAIREHIVERNAL DU GENRE MARTES EN HAUTESAVOIE
Référence ; BAUD (F.J.) 1981 - Contribution à la connaissance du rfuime alimentairehivernal du genre Martes en Haute-Savoie - Bièvre,3 (1),79-84. Muséum d'Histoire natu-relle, case postale 284, cH-1211 GENEVE 6.
Résumé.' L'étude de 35 estomacs des deux espèces du genre Martes capturées en Haute-Savoie nous montre un éclectisme plus grand dans le choix des proies chez M. martes etconf irme le statut de commensal de l'homme de M. foina dans nos rfuions, déf ini parWAECHTER (1976). Elle relève un nombre élevé de Hérissons lErinaceus europaeuslconsommées en hiver par M. martes ainsi qu'un cannibalisme ou une nécrophagie existant,au moins à cette saison, chez les deux espèces. Elle signale enf in, pour la première fois,la présence du Muscardin lMuscardinus avellanariusl comme proie chez M, martes,
Summary .'A contribution to the knowledge of the winter diet of the genus Martes cag-tured in Haute-Savoie (France).The study of 35 stomachs of the two species of the genus Martes ?Âptured in Haute-Savoie reveals a greater eclectism in the choice of Wev in M. martes and conf irms theman commensal status of M. foina in our regions, as def ined by WACHTE R (1976), ltshows a high number of hedgehogs lErinaceus europaaus) consumd bV M. mattes inwinter, as wellas cannibalism or necrophagy in both species, at least in this season. ltf inally records, for the f irst time, the presence of the dormouse (Muscardinus avella-nariusl as prey of M. martes.
Le genre Martes, bien que commun en Europe, n'est étudié au point de vueécologique que depuis peu d'années. Des travaux généraux tels que ceux de BAUMANN(1949) et HAINARD (196i) donnaient quelques indications sur le rég¡me de ces ani-maux, basées sur l'observation ou sur des études plus précises, telle celle de SCHMIDT(1943) pour l'Allemagne.
C'est LOCKIE (1961)qui présente le premier travail systématique sur le ré-gime alimentaire de la Martre en Ecosse, et il faut attendre la monographie de WAECH-TER (1975) pour mieux connaître l'écologie de la Fouine en Alsace. Puis GOSZCZYN-SKI (1976) étudie ce problème en Pologne, alors que DELIBES (1978) et AMORES(1980) le reprennent pour le nord et l'ouest de l'Espagne.
La présente contribution nous permet d'approcher cette question dans notrerégion.

80
| - Matériel et méthode
En 1976 nous obtehions, grâce à la collaboration de M. RENAUD, lieutenantde louveterie à Annemasse, 4 fouinæ et 8 Martræ. Puis, intéressé par le problème desproies des Mustelidae, M. GRILLET de la DDA d'Annecy nous faisait parvenir 26 esto-macs de ces deux espèces en 1977 et 1978. Toutes les bêtes examinées ont été tuéesentre les mois de janvier et mars et proviennent de la <Vallée Verte> au sens large,c'est-àdire de la dépression située immáliatement au sud-est de la chaine const¡tuéepar les Voirons, la Tête du Char, le Mont Forchat et le Mont d'Hermone, ainsi que dutriangle Magland-St Gervais-Megève dans la vallée de l'Arue. L'altitude varie de 500 à
1 100 mètres.
Comme la majeure partie de notre matériel était constituée de contenus sto-macaux, nous nous sommes cantonnés à I'analyse de ces contenus, sans tenir comptedu tractus digestif, même s'il était présent dans certains cas. Nous avons ainsi un nom-bre restreint de proies, mais qui ne sont que très partiellement digérées, permettant dedéterminer plus finement leur origine.
La détermination des parties dures s'est faite ælon les procédés classiques (col-lections du muséum, etc) et les poils ont été examinés par KELLER dont les techni-ques d'empreintes et de coupes histologiques sont décrites dans ses travaux de 1978et 1980.
ll - Résultats des analyses
Sur 14 estomacs de Fouines et24 de Martres, une M. martes était inutilisableet un individu de chaque espèce avait la cavité stomacale pleine de mucus, état consácutif au stress du piège (patte avant arrachée).
Pour 13 fouines lM. foina\, nous avons :
lnæctivores : néant
Rongeurs : Apodemus flavicollis Melch. (Mulot à collier) 1
Apodemus sp. (Mulots indéterminés) 3Rattus sp. (Rat indéterminé) 1
Flongeurs indéterminés 4
Martes sp, main presque complète... (Fouine ou Martre) 1
Restes de Passereaux 2
Cynorrhodons, herbe, terre, bois, restes de baies non déter-minables, carottes et ragoCrt de bæuf...
Nombre de proies moyen : 0,9 par individu, pour un total de 12 proies d'ori-gine animale.
Autres :
Divers :

81
Pour 22 martres lM. martesl , nous avons :
lnsectivores : Erircceus europeus L. (Hérison)Talpa europaa L. {Taupe)
31
1
1
31
21
1
5
1
1
1
231
Ronçurs :
Autres :
Oiseaux :
furex araneus L. (Muæraigne carrelet)
Lepus europaeus Pall. (Lièvre)Sciurus vulgaris L. (Ecureuil)Muscardinus avella¡nrius (L.), main complète (muscardin)Apodemus sp. (Mulots indéterminés)C lethrionom ys glareo lus (Sch reb ) (campagnol roussâtre)Microtus agrestis L. (Campagnol agreste)Autres campagnols indéterminés
Felis catus L. juv. (Chat domestique)Martes sp., une phalange avec griffe (Martre ou Fouine)
Erithacus rubecula (L.) (Rougegorge)hrus sp (Mésanges indéterminées)Passereaux indéterminésGallus domestrbus (L.) (Poule)
Divers : Cynorrhodons, poires à cidre, noyau-x ¿e cei'ileJ'ltli herbes,avoine, mousse, aiguilles de sapin.Puce de I'Ecureuil lMonopsyllus æiuroruml
Nombre de proies moyen : 'l ,5 par individu, pour 33 proies d'origine animale.
ll est à remarquer que sur 13 fouines,2 n'avaient mangé que des végétaux etune présentait un régime mixte et que sur 22Martres,3 avaient un contenu stomacaluniquement végétal et 5 mixte.
En ce qui concerne les proies animales, elles æ répartissent ainsi :
Fouine '.75 o/o de rongeurs,8,3 % autres que rongeurs (dans notre cas uneMartes sp.) et 16,7 % d'oiseaux.
Martre : 15,2 o/o d'inæctivores,57,6 o/o de rongeurs et lagomorphes, 6 % autres(dans notre cas chat et Martes sp.) et 21 ,2 o/o d'oiseaux.
Nous insistons sur le fait que ces résultats ne sont pas significatifs, mais qu'ilsnous donnent une bonne idée, bien que certainement partielle, du régime hivernal dugenre Martes en HauteSavoie.
lll - Discussion
Relevons tout d'abord l'abænce d'lnsectivores dans les proies de nos Fouines.

82
Ce fait provient certainement du faible nombre d'échantillons autant que de la techni-que de chasse de cet animal consistant à effectuer un parcours stéréotypé autour des
zones habitées, capturant ainsi essentiellement des espècæ commensales de l'homme,surtout en hiver où l'on trouve dans nos régions desApodemus jusque dans les mai-
sons. Ce type de chasse, démontré par WAECHTER (1975), explique également la re-
lative <monotonie> des espèces capturées par M. foina dans la région étudiée oùt seule-
ment deux genres de rongeurS Sont représentés alors que M. martes a un éventail de
6 genres de ce groupe (Lagomorphes compris).
Un autre point qui est intéressant à signaler est la préænce de3 Erinaceus
europaeus sur 22 contenus stomacaux de M. martes, nombre identique à celui des
écureuils, proie considérée comme typique de I'espèce. Nous savons qu'en hiver lehérisson entre en léthargìe au-dessous de I à 1OoC et qu'à ce moment sa température
descend et que ses réactions aux excitations sont nulles. C'est à cette occasion qu'ildoit être victime de la Martre. Une proportion si importante de Hérissons n'a jamais
été observée dans le régime des Mar-tres européennes et la présence de cet inæctivoren'est signalée que par GOSZCZYNSKI (1976) en Pologne chez Martes sp'
Enfin, I'apport végétal dans le régime alimentaire de nos deux espèces, bien
que difficilement quantifiable, n'est pas négligeable, puisque sur 35 bêtes, 5 avaient
l'estomac plein de fruits et d'herbes et 6 s'étaient nourries d'une manière mixte. Pour
ces 11 individus nous retrouvons régulièrement des restes de cynorrhodons -Æosa sp.-
en assez grande quantité (recherche de vitamíne C ? ), mais également des poires et
des pommes. Chez deux Martres nous avons même trouvé des noyaux de cerises prove-
nant selon toute vraisemblance de fruits ayant séché sur l'arbre car aucune bête cap-
turée n'a été appâtée à la cerise durant ces trois hivers.
lV - Cannibalisme
La découverte de restes tout à fait identifiables (main presque complète,phalange avec griffe, le tout avec peau et poils) appartenant au genre Martes dans
l'estomac d'une Martre et dans celui d'une Fouine pose un problème important. Bien
qu'il nous soit impossible actuellement de différencier ces deux espèces par l'examen
des poils, il semble exclu que nous soyons en présence de prédation réciproque, leur
biologie, quoique relativement proche, ne justifiant guère une compétition agressive
(WAECHTER, 1975).
Ouoi qu'il en soit, le cannibalisme est connu et a été observé chez plusieurs
espèces, soit au moment de la parturition en ce qui concerne les femelles primipares,
soit au moment de I'accouplement comme chez les félidés lorsque le mâle mord par
hasard trop fortement la femelle à la nuque et que le goÛt du sang supprime ses inhi-
bitions. Dans le cas qui nous occupe, la Martre provient de la réserve des Voirons et a
été tuée r.n 3 mars et la Fouine de la région de St-Gervais au mois de février. Les res-

83
tes trouvés dans leur estomac sont suffisamment complets pour dire qu'ils appartiennentà des animaux adultes. Notre opinion est confirmée par le fait que la mise bas commenceen mars et plutôt à la fin de ce mois chez nous. Une explication serait donc une ba-taille entre adultæ de la même espèce durant la période de <faux rut> qui s'étend entre
lanvier et début mars, avec ingestion du perdant, l'inhibition étant moins forte que du-rant la période de rut effectif de juillet et aott.
Une autre interprétation pourrait s'appuyer sur les diverses citations de tracesde Martres ou de Fouines auprès de cadavres divers. Une nécrophagie extra ou intraspé-cifique serait nullement à exclure, surtout en période hivernale.
V - Conclusion
Bien que provenant d'un petit nombre d'individus notre analyse qualitative desproies du genre Martes complète parfaitement les données des auteurs qui ont eu à leurdisposition un plus grand nombre de matériel. Grâce aux techniques d'analyse fines despoils et au fait que dans l'estomac les proies ne sont pas totalement digérées, nous avonspu déterminer avec plus de précision les espèces ingérées. Pour mieux comparer les diversrésultats, nous avons établi le tableau suivant, donnant le nombre de genres de Mammi-fères et d'Oiseaux cités comme proies dans les divers travaux.
I nsectivores Rongeurs Lagomorphes Carnivores O iseau x
LOCKIE 1196'll M. martes(337 fèces examinées)
WAECHTEB (1975)M. foina (431 feces)
GOSZCZYNSKT (1976)M. sp (835 fèces)
DELIBES (978l, M. foina( 184 feces, 14 estomacs)
AMORES tt198Ol M. foina(539 fèces)
BAUD (1981) M. foina(35 estomacsl M. martes
2
3
3
1
1
3
5
6
7
6
25
2
2
2
1 1 Putorius
1
2
'15
6
'13
12
13
3
Cette concordance est d'autant plus étonnante que dans notre cas nous n'avonspas seulement un nombre restreint d'échantillons, mais également que le contenu stoma-cal correspond à une durée de chase très courte précáCant la mort du prédateur car lebol al imentaire ne séjourne guère plus de 2 heures dans l'estomac.

84
Vl - Bibliographie
AMORES (F.) 1980 - Feedings habits of the Stone Martens, Martes foina (Erxleben, 1777l, ,insouth western Spain. - Säugetierk. Mitt. - 28 Øl :816-822,
BAUMANN (F.) 1949 - Dic freilebenden Säugatiere der Sdtweiz. Huber Verlag, Bern,492 p.
DAY (M'G.) 1966 - ldentification of hair and feather remains in the gut and faces of stoars andweasels - J. Zool, - 148,201-217,
DELIBES (M.) 1978 - Feeding habits of the Stone Marten, Martes foina Erxleben, 17771 , in nor-thern Burgos, Spain. - Z. Säugetierk, 43 lïl, 2BZ-289.
GOSZCZYNSKI (J.) 1976 - Composition of the Food of Martens -Acta theriol.,21 (36) , 527-594.HAINARD (R,) 1961 - Mammifères sauvages d'Europe, l. - Delachaux Niestlé, Neuchâtel, 322 p,
KELLER (4.) 1978 - Détermination des mammifères de Suisse par leur pelage l. Talpidae erSoricidae, - Rev, suisse Zool., 85 l4l ,758-761.
1980 - Détermination des mammifères de Suisse par leur pelage : ll, Diagnose desfamilles - lll. Lagomorpha, Rodentia (partim.). - Bev. suisse 2oot.,gl (3),7ól-796.
LOCKIE (J'D.) 1961 - The food of the Marten Martes martes in west Ross.Shire, Scotland. - Proc.zool. Soc. London,136, 187-195,
SCHMIDT (F.) 1943 - Natürgeschichte dar Baum- und Steinmatders, Monographien der Wildsäuge-tiere, 10. Verlag P. Schöps, Leipzig,258 p.
WAECHTER (4.) 1975 - Ecologie de la Fouine en Alsace. -Terre Vie,29,399-457.

85
Jean-François DESMET
. DONNÉES BÉCENTES SUR LA MUSARAIGNE ALP¡NESorex alpinus (Schinz, 1837) DANS LES ALPES FRANçAISES
Référence; DESMET (J.F.) 1981 - Données récentes sur la Musaraigne alpineSorex alpinus(Schinz, '1837) dans les Alpesfrançaises- Bièvre,3 (1), 85-88. 74340 SAMOENS.
Résumé.'Cette note relate les observations de Musaraigne alpine effectuées en été 1972etle 20 septembre 1977 ainsi que la récolte d'un individu de cette espèce le 18 juillet 1978dans la région de Samoëns (Haute-Savoie). A notre connaissance, cette espèce avait été cap-turée dans les Alpes françaises uniquement à Chamonix (Haut+Savoie) par MILLER en1912et à Argentières (Haute-Savoie) par CATZEFLIS en ,l97S.
Summary.' Recent data about the Alpine Shrew (Sorax a lpinusl in the French Alps.
The species was observed in Summer 1972and on September 2Ùth 1977, and one individualwas captured on July 18th 1978 in the Samoens area (Haute-Savoie, France),
To our knowledge, the only previous captures in the French Alps were made by M ILLERat Chamonix (Hte Savoie) in 1912 and by CATZEFLIS at Argentières (Hte-savoie) in 1975.
Les observations, captures ou récoltes de restes de Musaraigne alpine Sorex'alpinus (Schinz, 1837) sont rares dans les Alpes françaises, ceci étant probablement dûen grande partie au faible nombre d'observateurs et donc à un défaut de prospectiondans les zones a priori favorables. On ne sa¡t par ailleurs que très peu de choses concer-nant la biologie de cet animal.
ll semble toutefois que cette espèce recherche des endroits assez frais et mêmehumides, forestiers ou non, jusqu'à 2500 m. d'altitude. Nous avions déjà eu l'occasionà deux reprises de noter la présence de la Musaraigne alpine dans la Haute vallée duGiffre (Haute-Savoie) aux alentours de Samoëns ma¡s, faute de preuves matérielles, lavaleur accordée à ces informations éta¡t l¡mitée.
La première fois, en été 1972, un individu était observé de très près, à environ1350 m. d'altitude, sur un chemin de montagne traversant des prairies d'alpages humi-des entourées par de la pessière subalpine.
D'autre part, le 20 septembre 1977, une Musaraigne alpine était observée àproximité d'un chaletd'alpages, à environ 1500 m. d'altitude, en même tempsqueplusieurs autres Soricidés (Sorex araneus Linnaeus, 1758 très probablement). Toutesces musaraignes para¡ssaient très actives aux abords des toilettes extérieures, intéressées,semblet-il, par les nombreux ¡nsectes attirés par ces lieux. De nombreuses séances depiégeage effectuées à cet endroit -à l'aide de boîtes de conserve enterrées afin d'es

86
sayer de récolter des animaux vivants- permirent de capturer uniquement des Musarai-gnes carrelets Sorex araneus (Linnaeus, 1758).
Outre ces deux obærvations, plus récemment, le 18 juillet 1978, nous trou-vions une femelle de Musaraigne alpine morte sur un chemin traversant une hêtraie-pessière à environ 1000m d'altitude, toujours dans la même région des environs deSamoeins.
Le cadavre paraissait très frais et permettait donc de déterminer très facile-ment l'animal simplement par sa coloration et sa morphologie. Assez grand, il possé-
dait en effet un pelage entièrement sombre aux reflets cendrés, avec les pattes et ledessous de la queue un peu plus clairs. La queue semblait être d'autre part aussi longueque la tête et le corps réunis. Des mensurations plus précises ainsi que les critères dela denture nous confirmaient ensuite effectivement l'identification.
Données morphologiqøes (fig. 1)
Longueur de tête + corps : 75 mmLongueur de la queue (sans le pinceau de poils) :74 mmLongueur du pied (sans les ongles) : 16 mmCondylobasale : 20,8 mmLargeur du rétrécissement interorbitaire : 5,2 mmLargeur du crâne : 9,4 mm
1
Longueur de la rangée dentaire supérieure sans l,,' :7,7 mmLongueur de la rangée dentaire inférieure sans l' : 5,8 MM
(Le crâne ainsi que la peau tannée ont été conservés par l'auteur).
Rappelons enfin que MILLER (1912) signale la présence de la Musaraigne al-pine à Chamonix (HauteSavoie) etque plus récemment, en 1975, CATZEFLIS a cap-turé cet animal à 1320 m d'altitude à Argentières (environs de Chamonix, Haute-Savoie) (comm. pers.).
Par ailleurs, signalons que MEYLAN (1961) mentionne l'espèce en Suisse dansle Valais, entre Champéry et le col de Bretolet, soit à une distance d'environ 70 km à<vol d'oiseau> de notre lieu de récolte et il est très probable que cette Musaraigne estbeaucoup plus répandue que la rareté des récoltes pourrait nous le laisser croire.
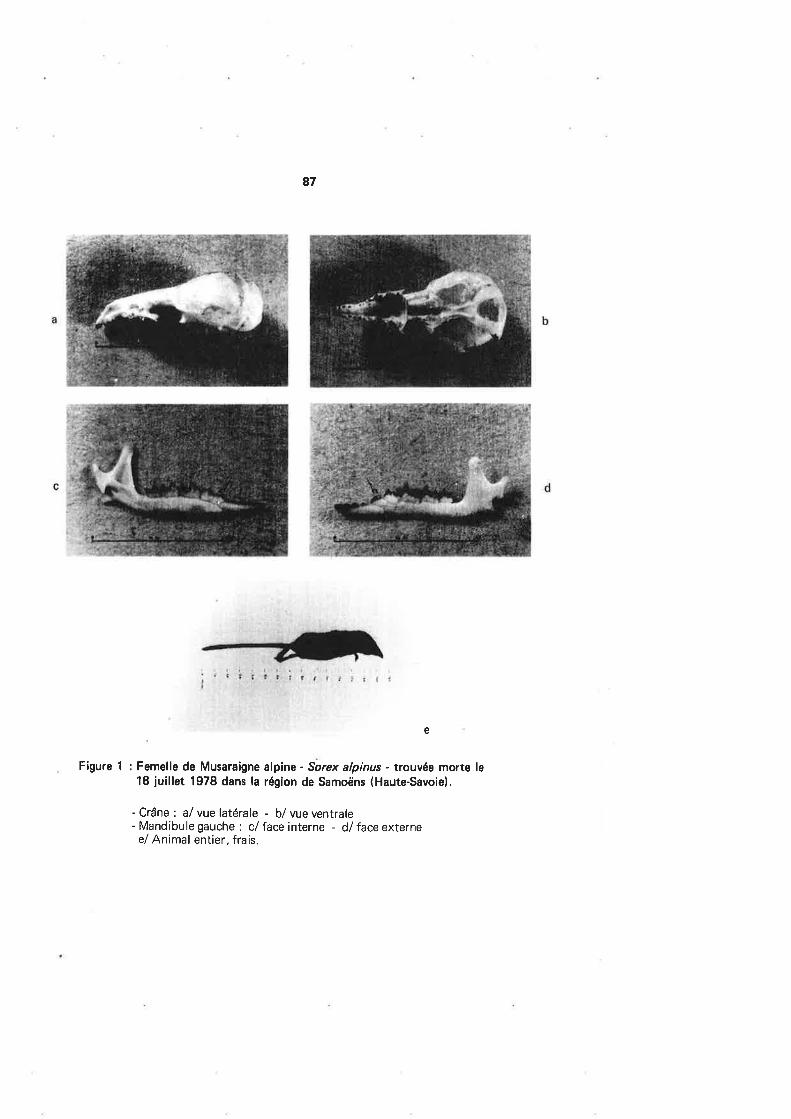
87
e
Figure 1 : Femelle de Musaraigne alpine - Sorex alpinus - trouvée morte le18 juillet 1978 dans la région de Samoëns (Haute-Savoiel.
- Cráne : a/ vue latérale - b/ vue ventrale- Mandibule gauche : c/ lace interne - dl face externe
e/ Animal entier, frais.

88
Bibliographie
AR IAGNO (D. ) 1976 - Essai de synthès€ sur les Mamm ifères sauvages de la région Bhône-Alpes -Mammalia, 40 (1 l, 1 2E-160.
DESMET (J.F.) 1975 - Essai d'inventaire des Vertébrés de la vallée du Haut-Giffre {Haute-Savoie) -Annales Centre univ. Savoie, numêro spécial, 179-200,
HAINARD (E.) 1 961 - Mammifères sauvagas d'Europe - I - lnsectivores, Cheiroptères, Carnivoras -Neûchatel, Delachaux Niestlé, 352 p.
MEYLAN (A.) 1961 - lnsectivores et Rongeursdans la région d€ Bretolet (résumé) - Rev, suissa Zoot.,68.
MILLER (G.S.) 1912 -øtalogueof theMammalsof Western Europø lEuropeaxclusiveof Russial -London, Brit, Mus,, 1019 f.
SAINTGIRONS (M.C.) 1973-¿es MammifèresdeFranceetduBenelux -Paris,Doin,488p.
Remerciements
Nous tenons à remercier ici M. CATZEFLIS de l'lnstitut de Zoologie et d'Ecolo-gie Animale de Lausanne pour les intéressantæ informations communiquées, MM. BAUDet WEBER du Muséum d'Histoire Naturelle de Genève pour leur accueil chaleureux, letemps qu'ils ont bien voulu nous consacrer et leurs très précieux conseils concernant laprésente note ainsi que M. FAYARD, Conservateur du Muséum d'Histoire Naturelle deGrenoble, et M. RAYNAUD, Chirurgien-dentiste à Samoëns, pour la réalisation de cer-uinæ photographies du crâne.

89
Stéphane AULAGNIER, PatricK BRUNET-LECOMTE et Hervé coouILLART
NOTE PRELIMINAIRE SUR LES CHIROPTERES
DU DEPARTEMENT DE LA LOIRE
R.érérence.'AULAGNtER (s.), BffUNET-LECOMTE (p.), ctoutLLABT (H.) 1981 -
\o^t9_nfli1!ry-iry sg¡ !e¡ c!¡gqtp¡gs du département de tá Loire - e¡¿we, à iil, eé_ss.C.O. R.A., 69622 VI LLEUR BANNE.
Résumé.' Les chiroptères du département de la Loire étaient jusqu,ici tout à fait incon-nus. 8 espèces de ces Mammifères viennent d'être signalées. cé travail présenìe cesdon-nées et les compare à une ancienne liste (CANTUEI tS¿S].
l!ry*rv " unt_iltoday, chiroptera of the department of the Loire were quite unknown.
Eight species of these mammars were recentry reported. Theseìata .rã órËén1ä inrhis nore and can be compared to an older liú (dANTUEL ig49).
Dans une synthèse sur les chiroptères de la région Rhône-Alpes, TUpINIER(1971) remarque en introduction : <la plupart des donñées proviennent de trois dépar-tements : l'Ain, l'lsère et l'Ardèche. pour le Rhöne, les monts du Beaujolais et la valléede l'Azergues ont été les mieux étudiés. Par contre, la Drôme et la Savoie sont assezpauvres. La Loire, enfin, n'est pas citée>. La Loire est également absente de la syn-thèse sur les Chiroptères rhônalpins de ARIAGNO (1976). Les seules données biblio-graphiques concerRant le département sont rapportées par cANTUEL (1g49). Ellessont relativement anc¡ennes et demandent à être conf irmées, surtout en raison de larégression consrante des effectifs de Chauve-souris (SAINT GIRONS , jg7gl.
Cette méconnaissance d'un département au demeurant fort riche en Mammi-fères, est la conséquence directe de l'absence de relief karstique ou de grottes naturel-les propices à la biospéléologie. De plus, les mines et souterrains, façonnés par l,hommeaux siècles derniers, mal localisés, sont peu prospectés. Néanmoins, à la faveur de nosrecherches sur les micromammifères du Forez, nous avons recueilli quelques Chirop-tères qui s'ajoutent aux rares données existantes. Læ quelques observations en notrepossession sont rasæmblées dans cette note afin de combler, partiellement, un vide etde susciter des recherches plus systématiques des chauves-souiis.
Rhinolophus hipposideros (BECHSTEIN, lBOO) : petit Rhinolophe fer à cheval.
un individu en hibernation dans une cave a été observé en main par l,un d,en-tre nous en novembre 1973 à Pouilly sous Charlieu (Roannais).

90
Myotis emarginatus (GEOFFROY, 1806) : Vespertilion à oreille échancrée.
Un crâne de cette espèce a été trouvé dans une pelote de réjection de Tytoalba, sur la commune de Saint-Just sur Loire en 1978 (tabl. 1).
Myotis sp. : Vespertilion à moustaches ?
Un Vespertilion de petite taille a été observé un mat¡n d'août 1972 (6 h) àPouilly sous Charlieu. L'aninial, à oreilles courtes non échancrées et tragus pointu de
longueur subégale à la moitié du lobe, était éveillé au pied d'un marronnier. Cette ob-
servation ne permet pas de conclure s'il s'agissailde M. mystacinus ou M' brandti ,
espèce très rarement observée en France (TUPINIER et AELLEN, 1978).
Myotis nattereri (KUHL, 1818) : Vespertilion de Natterer.
Un crâne de cette espèce a été trouvé par DESTRE dans une pelote d'Effraieà Sévelinges (Monts du Lyonnais) en 1978 (détermination : MEIN).
Myotis bechsteini (KUHL, 1818) : Vespertilion de Bechstein.
La seule colonie recensée à ce jour, dans le département de la Loire, appar-
tient à cette espèce. Une vingtaine d'individus ont été observés par BUSSY dans une
cave de Magneux Haute Rive, en fin d'été 1973-1974-1975. Un Vespertilion de Bech-
stein a également été trouvé dans un lot de pelotes provenant de Saint Marcellin en
Forez (tabl. 1).
Tableau 1 : Mesures (en mm) recueillies sur deux crânes de Vespertilions.largeur bizygomatique Longueur de la mandibulelargeur entre les M3 hauteur coronoideLongueur entre canine et troisième molaire (M3)
l. bizyg. . v3-rrl3 L. C-M3 L, mand h. coron t-, c-ua
Myotis emarginatus
Myotis bechsteini
9.10
t0.(n
6.52
7.00
5.90
6.99
I t.07
t2.52
3.06
3.96
6.33
7-30
Pipistrellus pipistrellus (SCHREBER , 17741 : Pipistrelle commune.
Cette petite Chauvesouris a été déterminée de manière précise à deux reprises,
à Pouilly sous Charlieu en 1972 et à Saint-Etienne en 1977.
Pipistrellus kuhli (KUHL, 1819) : Pipistrelle de KÜhl.
Deux individus morts ont été recueillis en 1979, l'un à Thélis la Combe (Pilat)
l'autre dans un quartier de Saint-Etienne (La Métare).

91
Plecotus auritus (L., 1758) : Oreillard septentrional.
ARIAGNO a déterminé un animal de cette espèce provenant de Saint-Etienne,trouvé dans le même site que la Pipistrelle commune. Un autre individu a été trouvé à
Sail sous Couzan (Montsdu Forez, BRUGIERE) ; un troisième Oreillard, enfin, a étéeitrait d'une pelote d'Effraie provenant de Poncins, mais les fragments n'ont pas permisde déterminer précisément l'espèce.
Notre connaisance des Chiroptères est ponctuelle en dépit de I'existence pro-bable de gîtes d'hivernage ou de reproduction. L'absence d'espèces arboricoles parmiles proies deTyto alba TanT dans la Loire qu'en lsère ou dans le Rhône (NOBLET,1980; ARIAGNO et al., à paraître) semble accréditer l'hypothèse selon laquelle cetteChouette capturerait les chauvesouris troglophiles dans leur lieu de séjour estival (clo-
cher, grenier...) (UTTENDORFER et al., 1952). La prédation qu'exerce l'Effraie surles populations de Chiroptères reste très faible dans notre région (4 individus sur14000 proies dans la Loire,3 sur'10000 dans le Rhône).
La liste établie par CANTUEL (19491 (tabl. 2) à partir d'animaux naturalisés,fait état de I espèces pour la Loire. Les observations récentes concernent I espèces
dont 4 au moins n'étaient pas encore signalées dans le département. Ce nombre est net-tement inférieur à celui des départements voisins, Rhône, Ardèche, Ain... (tabl.3). Cettefaible diversité peut s'expliquer par le manque de recherches systématiques des Chirop-tères du département, mais également par divers facteurs régionaux.
Tableau 2 : Espèces observées dans la Loire, par CANTUEL (1949) et plus récemment.(R : Rare ; AC : Assez Commun ; C : Commun)
R hi noloph us ferrumequi nu m
Rh inoloph us hipposideros
Myotis myotis (7 )Myotis emarginatus
Myotis mystacinus
Myotis nattereriMyotis bechsteiniEptesicus serotinus
Nyctalus noctulaPi p istrel I u s p i p istre I I u s
Pipistellus kuhliPlecotus auritusBarbastel la baúastel la
1 949 1980
F
ccR
c
AC
AC
c
AC
+
+?
++
+++

92
Tableau 3 : Comparaison du nombre d'espèces de Chiroptères observées dans la Loire etdans les départements voisins
Département Nombre d'espèces
LOIRERHONEAINARDECHEISERE
I15
13
15
23
(AR IAGNO et al. - à paraître)(FAYARD et at. 19791(TUPTNtER, 1971 - FAUGtER, 1979)(NOBLET, 1980}
RHONE-ALPESFRANCE
27 (NOBLET, 1980)30 (NoBLET, 1980)
A la rareté des gÎtes souterrains naturels s'ajoutent une forte préænce humaine(résidences secondaires), un développement industriei important dans certa¡ns secteurs(Vallées de l'Ondaine et du Gier) et une modernisation agricole poussée (pesticides,(entretien des haies>...) dans les plaines du Forez et du Roannais notamment.
L'isolement de la Loire au sein du Massif central, à l'écart de la vallée du I
Bhône, et le climat de type continental modéré, sont à même de freiner la pénétrationd'espèces mfuiterranéennes (Pipistrellus nvii, Myotis capaccini, Tadarida teniotisl. LesChauvesouris observées ont une aire de répartition large, médioeuropéenne à l'excep-tion de Pipistrellus kuhli, espèce méridionale qui a néanmoins été observée plus auNord (SAINT GIRONS, 1973).
En dépit de cæ facteurs défavorables (climatiques, géologiques et (économ¡-ques>) susceptibles de limiter la diversité spécifique ¿ei Cn¡ropt¿r"r, ¡l ,"rt. probablequ'une prospection plus systématique permettrait d'allonger la liste des Mammíffèresde la Loire' De plus, une.connaissance plus approfondie des Chauve-souris du départ+ment est nécessaire pour instaurer une protection efficace de ces espèces qui ont acquisrécemment le statut d'espèces protégées, et de sauvegarder les quelques mílieux qui '
leur sont favorables.
Bibliographie
ARIAGNO (D.) 1976 - Essai de synthèse sur les mammifères de la rfuion Rhône-Alpes. Mammatia,40 (1).125-160.

93
ARIAGNO (D.), AULAGNIER (S.), BROYER (J.), BRUNET LECOMTE (P.} 1981 (à paraître) -Les mammifères du département du Rhône.
CANTUEL (P.) 1949 - Faune des Vertébrés du Massif Centnl da la France -400 p.
Lecheval ier, Paris,
FAUGIER (C.) 1979-Evolutiondespopulationsdechauve-sourisenArdèche.07Nature,5,7-11.
FAYARD (4.), ROLANDEZ (J.L.l , RONCIN (P.) 1979 - Les mammifères du département de l'Ain.Bièvre, 1 Í1 ,1-26.
NOBLET (J'F') 1980 - Nouvellesdonnées sur la répartition des chlroptèresde larfoion Rhône-Alpes. Cas particulier de l'lsère. Ciconia,4 (1), S-1 1.
SAINT GIRONS (M.C.) 1973 - Les mammifères de Fnnce et du Benelux. Doin, paris,4B1 p.
TUPINIER (Y.l 1971 - Chiroptères de la région Rhône-Alpes. Actes lve Congrès suisse Spéléo\.,Neuchâtel, 205-212.
TUPINIER (Y.), AELLEN (V.) 1978 - Présence de Myotis brandti (EVERSMANN ,18451 lChirop-teral en France et en Suisse. Rev. suisse Zool., 85 l2l, 449456,
UTTENDORFER (O.), BODENSTEIN (G.), KUHK (R.) 1952 - Neue Ergebnisse über die Ernàhrung.der Greifuöçl und Eulen. Ulmer, Stuttgart, 149-164.
Remerciements
Nous remercions toutes les personnes citées dans le texte qui ont communiquéleurs données à P. GUYor et particulièremenr D. ARIAGNo et P. MEIN qul ont con-firmé certaines déterminations délicates.


95Le Bièvre, 1981, 3(1 )
Hervé COOUILLART (1)
XXe COLLOOUE INTERRÉGIONAL D'ORNITHOLOGIE
Saint-Etienne, les 15 et 16 novembre 1980
organisé à l'université de sAINT-ETIENNE, ce colloque a réuni plus de 400
participantJvenus de France et de Suisse (pays organisant alternativement ces colloques),
mais aussi de Belgique, d'Allemagne Fédérale et d'ltalie.
euelques allocutions ont permis de rendre hommage aux fondateurs de ces col-
loques interrégionaux Paul GEROUDET, Camille FERRY, Philippe LEBRETON, et de
mesurer le chèmin parcouru en 20 ans dans la connaissance et la protection des oiæaux.
La mémoire de Michel BROSSELIN, décédé en juillet 1980, et la contribution essen-
tielle de son æuvre à l'efficacité et à la crédibilité actuelles de la protection de la nature
ont été saluées avec émotion.
Nous présentons ci-dessous les résumés des communications et des posters.
Ont également eu lieu :
1- une excursion sur le terrain pendant laquelle ont été observés de nombreux
Rapaces (Buæs, Milans royaux, Eperviers, Faucons crécerelles) et des Sizerins flammés
(observations rares dans la Loire).
2- une séance de projections oùl furent visionnés, outre quelques diapositives
de Rapaces, un remarguable fonduenchaîné sur la LOIRE et un f¡lm récent de J.F' et
M. TERRASSE <Tunisie, refuge aux oiseaux>.
3- des tables rondes, qui ont débattu de cinq sujets de protection de la nature :
. l'impact des aménagements et de I'industrie sur l'avifaune ;
. les trafics d'animaux sauvages : les participants ont analysé les textes lé-
gislatifs, leurs applications et discuté du pouvoir réel des associations. lls ont déploré
L'inaction et souvent l'ignorance des administrations dans ce domaine ;
les grandi aménagements en projet ou en cours de réalisation dans la
plaine du Forez ont permis d'aborder à travers une sortie sur le terrain un sujet auquel
beaucoup de protecteurs de la nature sont confrontés. Une motion a été votée par l'en-
semble des participants du colloque ;
.la protection des rapaces, le bilan du travail réal isé par le FlR,les objec-
tifs et les moyens concrets de progresser dans ce domaine cher à beaucoup d'ornitholo-gues comme en témoignent les 120 personnes réunies autour de ce thème;
(1) C.O.R.A. - Section Loire

96
. les relations chasseurs-protecteurs de la nature, administratíons qui ontfait I'objet d'un large débat, prenant acte :
- de la complexité des répartitions de responsabilités entre les ad-ministrations et de l'abænce de coordination ;
- de la remise en cause actuelle, par certains responsables cynégé-tiques, des mesures légales nécessaires à la survie de la faune sauvage.
Les participants proposent une série de mesures regroupées autour de deuxthèmes :la prise en compte de la position de la F.F.S.p.N. et la plus grande utilisationdes médias,
Ce Colloque aura permis la présentation d'excellentes communications scienti-fiques, mais aussi des échanges, des discussions et des act¡ons à travers lesquels æ tisseI'avenir de la protection de la Nature en France et en Europe.
| - Résumé des communications
l-1 'Données quantitatives sur l'avifaune d'une futaie de pins sylvestres de 125 hadans les Vosges du Nord, par Yves MULLER (1)
Les résultats ont été obtenus par l'étude de :
- un quadrat de 12ha et 5 IPA répartis sur ce quadrat (1979-1980). Les '10
IPA réalisés au cours des deux années permettent ainsi d'obtenir des coeffícients deconversion I PA/densité absolue.
- un quadrat sur l'enæmble de la forêt de 125 ha (1980), pour les oiseaux àgrands territoires et les æpèces à faible distribution. Les onze espèces les plus comnlu-nes (densité de plus de deux couples aux 10 h en 1979) n'ont pas été recensées sur,cequadrat, mais elle I'ont été par la méthode des IPA (12lpA' sur les 12b ha). La densi-té de ces espèces sur l'ensemble de la forêt est alors obtenue en convertissant les lpAen densité absolue.
L'avifaune totale de cette futa¡e de Pins sylvestres avec un sous-étage de feuil-lus est donc constitué de 38 espèces totalisant 752 couples.
Cette technique a permis d'établir les caractéristiques du peuplement étudié,et d'envisager les comparaisons possibles avec d'autres travaux réalisés dans des forêtsde conifères.
(11 28, ¡ue Vauban - 57230 BTTCHE

97
l-2 - Variabilité du nombre d'espèces d'oiseaux contactées en un point d'écoute de20 minutes, par Michel BOURBON et Michel BOURNAUD (1).
Texte de la communication à paraître dans Bièvre 3 (2).
l-3 - Une nouvelle méthode pour l'étude du régime alimentaire des rapaces nocturnes
cavernicoles, par Michel JULLIARD (2).
Jusqu'à présent, l'étude du régime alimentaire des rapaces nocturnes était ba-
sée sur l'analyse des pelotes de réjection. Or, cette méthode n'est pas absolue pour plu-
sieurs espèces car les proies invertébrées (lombric, papillon, chenille, etc...) ne laissent
aucune trace macroscopique dans les pelotes et même si certaines en laissent, il est im-possible d'obtenir des données qualitatives valables sur la nourriture ingérée.
Dans le cadre d'un travail de thèæ effectué à l'lnstitut de zoologie de l'Univer-sité de Neuchâtel et à la station ornithologique suisse à Sempadr, nous avons mis au
point une caisse spéciale que nous substituons à un nichoir occupé et qui nous permet
de prendre des photographies. A l'aide de pellicule infra-rouge et d'un système de dé-
clenchement automatique des prises de vue, nous enregistrons systématiquement toutesles alléæ et venues des oiseaux, ce qui permet d'obtenir l'intégralité des proies appor'tées aux pousins. Une horloge, placée dans le nichoir, permet de voir à quelle heureles rapaces nourrissent leurs jeunes et, comme chaque oiseau est bagué, nous pouvons
aussi calculer les fréquences des nourrissages par æxe.
Cette méthode, inspirée du travail du japonais ROYAMA sur les Mésangês, a
été utilisée en Suisæ pour étudier la nourriture des jeunes Chouettes chevèches lAthenenoctual dans le Jura, dans la campagne genevoise et dans la plaine du Rhône en Valais.
Efle a aussi permis l'étude de deux couples de Chouettes hulottes (Strix alucol d'uncóuple de Chouettes effraies lTyto albal et d'un couple de Chouettes de Tengmalm
lA.egolius funereusl.
l-4 - Les espèces d'oiseaux méridionales le long de la côte du Rhône ardéchoise,par Jean-Dominique LEBRETON (3).
Texte de la communication : voir Bièvie 2 (l), pages 63 à 97.
(1) C.O.R.A., Université LYON 1 ,69622 VILLEURBANNE CEDEX(2) Tilleuls 4, CH 29000 PORRENTRUY(3) C.O.R.A. et Labo Biométrie - Université LYON 1 ,69622 VILLEURBANNE CEDEX

98
l-5 - L'avifaune des prés humides d'Alsace - Etude quantitative et écologique,par Christian KEMPF (1).
Au cours de l'année 1979, des comptages sur bandes ont été menés, sur 51hectares, dans des milieux prairiaux inondables d'Alsace.
Au total, 118 relevés permettent de dégager les densités (entre 4 et 10 cou-ples aux 10 ha) ainsi que la variété d'oiseaux.
ces résultats sont comparés à ceux des cultures de mai's, beaucoup plus pau-vres, ainsi qu'à divers travaux récemment menés en Europe, et permettent de dégagerles facteurs limitant esentiels de ces types de milieu :
- eau- couvert végétal.
Les cortèges d'oiseaux inféodés à ces milieux sont décrits, ils vont de l'association à Barge à queue noire (Limos timosal et à Räle des genêts (Crex crexl auxmilieux à alouette des champs (Alauda arvensisl.
La conclusion fait ressortir la rareté de ces biotopes de plaines inondables enFrance et l'urgence de leur conseruation : en Alsace, 80 % de leur surface a été dé-truite en 20 ans par le mai's et les gravières.
l-6' Les réserves cynégétiques sur les plans d'eau artificiels dans le bassin du Rhônefrançais, par Jean-Pierre CHOISY (2).
Les ordres de grandeur des effectifs hivernants sur l'ensemble des réserves cy-négétiques des plans d'eau artificiels (retenues.et anciennes carrières de granulats) sontdonnés par le tableau ci-dessous ; ils peuvent être temporairement beaucoup plus éle-vés : coup de froid, passage).
ANATIDES
Cygne tubercu lé (Cygnus olorlcolv€rr (Anas platyrhynchoslChlpeau lAnas streperalSarcelle d'hiver lAnas creccalMitouin lAythya ferinalMorillon (Aythya fuligulal
TOTAL
effectif tendance
30-401 500- I 600
50-70300 - 400
1000-1200100 - 150
€nviron3OOO-35OO
;
AUTRES FAMILLESGrèbe huppé ( tus)Grèbe castagn€ux ruficollislHáron cendré (Foutque (Fulica atrclPoule d'eau lGallinula chloropusl
TOTAL
80 - 100- de lOO50- 100
1 300- 1 500quelques centaines
2 O0O environ
+
Tendance: + = augmentation ;-=en dim¡nution
(1) A.C.l,N.E.R. - 36, rue de Sélestat - 67830 MUTTERSHOLTZ(2) G.D.E.R.V. - Vercheny - 263/:0 SAILLAN

99
La baisse des effectifs de Sarcelle d'hiver (Anas creccal s'explique par le boisement spontané des vasières de la réserve de l'Etournel. Certains de ces plans d'eau ontdéjà fait l'objet de publications. Deux articles sont en cours de rédaction concernantceux de la BaseVallée du Rhône et de la Basselsère (toute donnée complémentaireserait bienvenue...).
Nous proposons de travailler à augmenter le nombre et la superficie de ces ré-serves selon diverses modalités :
- dès maintenant par accord avec les sociétés de chasse, les fédérations et l'Offi-ce National de la chasse ;
- par accord avec l'Administration (D.D.4., O.N.C.) dans quelques années lorsde l'expiration des baux de location (presque tous ces plans d'eau sont du domaine del'Etat).
- par location du droit de chasæ sur ces mêmes plans d'eau lors des adjudica-tions : les prix dérisoires actuellement pratiqués (de l'ordre de trois cents francs par anpour plus de 15 km de cours d'eau) doivent faire envisager cette éventualité.
- par demande de mise en réserve cynégétique de la totalité des plans d'eau quiseront créés à l'avenir dans le cadre des (mesures compensatoires>.
UN POINT IMPORTANT:
Lors de la constítution de dossiers en vue de miæ en réserve, on ne peut abso-lument pas se baser sur l'absence actuelle d'hivernage pour préjuger du manque d'inté-rêt d'une telle mesure : pour chacune des réserves actuellement existante, l'hivernageétait quasi nul (de l'ordre de 1o/o des effectifs actuels) avant la mise en réserve. On peuttoujours préconiser deux ans de mise en réserve <à l'essai> et étudier l'évolution des ef-fectifs; d'autre part, dès maintenant, et avant les adjudications (dans 5 ou 6 ans) onpeut recueillir des informations très précieuæs en allant observer les stationnementslors de la migration de printemps dès la fermeture de la chasse, soit dès le premiermars,
l-7 - Suivi de la migration sur le marais de Thèse (Alpes de Haute Provence),par Roger GARCIN et Michel BOUVIER (1).
Texte de la communication à paraître dans Bièvre 3 (2).
l-8 - Exploitation des reprises de bagues de grives et merles enregistrées dans larégion Rhône-Alpes, par Yves THONNERIEUX (2).
Texte de la communication pages 1 à 54.
(1) Charance Nature - 05000 GAP(2) Centre régional de Baguage d'Oiseaux de Lyon, C.O.R.A, - Université Lyon 1
69622 Villeurbanne Cedex

100
l-9 - Etude d'une population d'Engoulevents d'Europe ' Les premiers résultats,par René AUCLAIR (1).
Synthèse de six années d'études de l'Engoulevent d'Europe (Caprimulgus
europaeusl dans l'Allier en forêt domaniale de plaine, sur trois parcelles en régénéra-
tion naturelle.
56 nids ont été découverts et 89 oiseaux bagués dont 36 adultes.
Chaque couple fait normalement deux pontes de deux æufs chacune, à même
le sol. La réussite semble affectée par les intempéries (pluies de 1977], et la prédation(carnassiers).
Les échecs sont beaucoup plus ngmbreux sur les nids situés en végétationhaute et serrée.
Le cycle de reproduction des deux nichées s'étale sur près de quatre mois.
25 contrôles effectués sur 14 oiæaux différents bagués adultes montrent :
- une grande fidélité au site de reproduction,- 3 cas de fidélité de couple,- 3 cas de quatre années de longévité,- certaines particularités dans le comportement.
Des mæures de biométrie, il ressort :
- une taille et un poids moyen supérieur chez les femelles,- des différences assez importantes d'un sujet à l'autre,- des longueurs d'ailes pliées qui ne sont pas toujours en rapport avec i
les poids correspondants.
Cette espèce est donnée comme étant en diminution. Son avenir semble liédans notre région au maintien des biotopes favorables.
;
f -10 - Un couple mixte Hippolais polyglotta X icterina a élevé deux jeunes en
Bourgogne,.par Camille FERRY (2).
Au prìntemps 1980, un couple mixle Hippolais polyglotta mâle x H. icterinafemelle s'est reproduit en Côte d'Or.
(1) Hyds, 03600 COMMENTRY(2) C.E.O.B. - Faculté des Sciences Mirande - 21000 DIJON

101
L'identité spécifique des deux adultes, reconnue à leurs émissions vocales, aété confirmée par les caractères biométriques apÈes capture au filet. Le nid, contenantcinq æufs, n'a donné que deux jeunes à l'envoi, nouriir par les deux parents. un pre-mier cas identique polyglotta mâle x icter¡na femelle aur¡t ¿t¿ soupçonné en 1975,sans qu'on ait pu avoir la confirmation par la capture des adultes qúi avaient disparuaprès destruction de leur nid contenant cinq æufs.
On discute les conditions et les consQuences éventuelles d'une telle hybrida-tion; elle reste exceptionnelle puisque dans les stations de sympatrie en Bourgognenous n'avons observé que.ces deux cas depuis 1962, alors que 82 couples étaiõni formésd'oiæaux de la même espèce.
l'11 - Deuxième étude sur le Grand-Duc en Auvergne, par Didier cHoussy (1),Gitbert COCHET et Alain DELpUECH (déOi¿e à ta mémoire de MichetBROSSELIN)
cette étude porte sur les quatre départements de la région Auvergne où lestrois observateurs se sont efforcé de dénombrer les couples de órands Ducs lBubo bubolactuellement existant. Elle se divise comme suit :
1- répartition de la population de Grands-Duæ d,Auvergne :
lls'agit ici d'envisager la répartition actuelle des différents couples en Auvergne,d'établir des densités tant globales que par sites.
2- évolution des populations :
Dans cette deuxième partie, nous mettons en parallèle trois répartitions diffé-rentes : la répartition optimale telle qu'elle était avant l'introduction de la myxomatoæ,la répartition en 1971 et la répartition actuelle.
3- biologie de l'espèce :
Nous étudions le cycre annuer du coupre, cantonnement, reproduction, érevage.Nous parlons également du régime alímentaire, de la susceptibilité ei du comportemeñtdes adultes (évoquant entre autres le Grand-Duc comme super prédateur).
4- conclusion sur l'avenir de l,espece, les dangers qui la menacent.
(1) V¡lle Haute - 63420 ARDES sur COUZE.

102
ll - Résumé des Posters
ll-l - Répartition alt¡tudinale de I'avifaune des monts du Forez,par Régis ANDRE
(1) et Hervé COOUILLART (2)
sur la bordure Est du Massif central, les Monts du Forez présentent une su-
perficie de 25 000 hectares. lls sont orientés Nord-Sud et culminent à 1 643 m'
La ré faune (tableau) a en évi-
dence la prése nes ou nordiques' ents
(ligne à très h enrésinement inte de tour-
bières, routes milieux naturels, andes
pseudo-alpines sommitales appelées (HAUTES CHAUMES)'
Landes à genêts - pinèdes
Hêtra ies-sap in ières
Lisières de forêts -
clai ri ères
Hautes Chaumes
Tou rbières
Rochers sommitaux
Bruant lou Emberiza ciaPouillot de Bonelli Phylloscopus bonelli
Pouillot siffleur Phylloscopus sibilatrixGrimpereau des bois Certhia familiarisBeccroisé des sapins Loxia curvirostraVenturon montagnard Serinus citrinella
Gelinotte des bois Tetrastes bonasia
Pipit des arbres Anthus trivialisMerle à plastron Turdus torquatusPouillot filis Phylloscopus trochilus
Pipit spioncelle Anthus spinolettaBruant ortolan Emberiza hortulana
Pipit farlouse Anthus PratensisTraquet larier Saxicola rubetra
Merle de roches Montícpla nxatilisTraquet motteux Oenanthe oenanthe
ll-2'D¡stribution des Chouettes chevêches lAthene noctual nicheuses en Suisse'
par Francis BENOIT (3)
É.n 1g7j,la liste rouge des espèces menacées de Suisse indique pour les Chouet-
tes chevêcfrþs.<Forte diminution dèpuis le début du sièilä, aócélérée depuis 1950; èffec-
tifs inconnus>.
(1) 33, rue Camélinat'42OOO SAINT-ETIENNE(2) C.O.R.A., Université Lyon 1 , 69622 VILLEURBANNE Cedex(3) Station ornithologique suisse CH 6204 SEMPACH

103
Après deux ans de prospection ornithologique, le tableau récapitulatif des cou-ples de Chouette chevêche nicheuse en Suisse se présente ainsi :
Ajou JU. 50 couplesCampagne genevoise 40 couplesGuerbetal DE. 25 couplesGrand Marais B.E.F.R. 25 couplesVallée du Rhône US. 10 couplesPlaine de Magadino Tl. 10 couplesReste de la Suisse 25 couples
TOTAL 185 couples
ll-3 - Migration du Grand Cormoran (Phalacrocorax carbol dans la région Rhône-Alpes (Hautes-Alpes inclues), par Jean-Pierre CHOISy (1)
L'estivage est exceptionnel.
L'hivernage n'est notable qu'au Léman, où il est en augmentation (quelquesdizaines dans les années cinquante, plus de cent actuellement). Ailleurs, les effectifsobservés sont de 1 à 3, exceptionnellement '10; essentiellement sur les retenues duRhône, de la Loire et à Serre-Ponçon.
Le pasuge d'automne est très étalé : amorcé en Septembre, en très faible quan-tité, il culmine au cours de la dernière semaine d'octobre et de la première de Novembre,décline ensuite progressivement au cours de ce mois.
Le passage de printemps est au contraire très concentré, la très grande majoritédes oiseaux passant au cours de la seconde semaine de mars. ce pic commence trèsbrusquement, presque sans transition avec les très faibles effectifs hivernants, mais ætermine progressivement courant avril.
La très grande majorité des observations au passage sont fournies par la Valléedu Rhône en aval de Lyon et la Dombes (bizarrement très peu par le Forez).
Six reprises d'oiseaux bagués pousins ont été faites en période de migrationde 1927 à 1978, toutes en Dombes ; quatre provenaient de Hollande (Lekkerkerk) etdeux du Danemark (Horsens Fjord).
ll-4' L'électrocution des rapaces sur les lignes électriques, par Jean-Michel FATON (1)
Le Fond d'lntervention pour les Rapaces (FlR) reçoit régulièrement des lettressignalant des cas d'électrocution de Rapaces dans toute la France. Dans le département
(1) G.D.E.R.V. Vercheny -26430 SATLLANS(2) Chargé de l'étude par le FIR - FRAPNA Drôme, ancienne mairie, quai St Nicolas
26500 BO U RG.LES-VALENCE

104
de la Drôme, en 1976 et 1980, nous avons trouvé plus de 20 rapaces électrocutés surles lignes moyenne tension. Pendant la même période, EDF a noté vingt coupures decourant provoquées par l'électrocution de rapaces. Ce rapide sondage nous laisse pen-ær que les électrocutions sont l'une des principales causes de mortalité artificielle desoiseaux de proie.
Nous avons proposé à EDF deux types de protection contre les pylônes meur-triers :
1- la pose de gaines isolantes en PVC sur la phase centrale;2- la pose de perchoirs en bois sur les equipements aériens spéciaux.
EDF est prête à accepter de modifier les pylônes (ayant déjà tué> après ex-périmentation.
Nous lançons un appel à tgus les naturalistes: les électrocutions que vousconstatez doivent être recensées et les pylônes dangereux identifiés.
FRAPNA 26 enverra des fiches spéciales sur simple demande.
ll-5 - L'Aigle de bonelli lHieraetus fasciatusl:des stades juvéniles a suU-a¿ulte,par Jacques FRIER (1)
Ces obærvations se rapportent à un sujet måle connu en plumage ¡uvenite:, al'âge de un an, au printemps 1979. RIVOIRE et HUE (O.R.F.O. 194g) signalent ceplumage jusqu'au deuxième automne (18 mois). Le26 novembre 1979 (à 21 mois),l'oiseau reste complètement sombre sur les part¡es pectorales, le dessous de l'aileconserve le plumage juvénile, mais une minuscule tache blanche est déjà visible entreles épaules. Le 2 mars 1980 (à 24 moisl,la poitrine est bariolée de blanc et la barresub-terminale des rectrices est visible, à 27 mois la poitrine est entièrement blanche etlæ couvertures secondaires se sont éclaircies. Le dessous de l'aile montre une large bandenoire, moins importante que chez l'adulte; le bord d'attaque n'est pas encore blanc.Le2fiévrier 1980 (à 32 mois), ce plumage subadulte n'a pas évolué : c,estcelui d,unadulte dont les faces ventrales et dorsales ne seraient pas contrastées. Les culottes res-tent encore rousses, comme celles d'un jeune.
ll-6' Les bassins amortisseurs de crue des autoroutes et leur intérêt pour quelquesoiseaux d'eau, par Jean-Pierre REITZ (1)
L'affiche présentée ne constitue en aucun cas un plaidoyer en faveur des auto-routes, mais illustre le résultat d'une intervention de la Section protection de la nature
(1) Les Mouettes - 38310 LES ROCHES DE CONDFTEU(2) Le Pâquis - CH - 1008 Jouxtens-Mézery

105
et de la Conærvation de la Faune du canton de Vaud, auprès du Bureau de construc-tion des autoroutes (BAR) et la réaction positive de ce dernier.
Au lieu de se contenter de construire pour les besoins techniques, une simplefosse de rétention, le BAR a mis au point au <Grand Roæy>, commune de penthazlr'D,un type de bassin amortisseur de crue alliant la nature à un ouvrage de génie civil (di-mension max. 120 x 60 m).
Les plantes palustres ont colonisé spontanément les lieux; les oiseaux d'eaus'y sont installés, comme nicheurs ou en escale.
4 autres bassins plus petits ont été réalisés dans la partie vaudoise de la Valléedu Rhône.
Caractéristþues générales des ouvrages :
Berges en pente douce; arborisation partielre ; clôtures, acèès, chasæ et pêchei nterd ites.
ll-7 - L'habitat du Faucon pélerin, par Dagmar VOGT (1)
J'étudie depuis 10 ans la structure de l'habitat du Faucon pélerin (Falco peregrinusl en Allemagne, en Suisse et en France. Je tiens à attirer l'attention des proiec-teurs de la nature, en particulier ceux qui s'intéressent au Faucon pélerin, sur le faitquer cet oiseau pourrait survivre en région boísée, plutôt mieux qubn région ouverte.D'autres résultats, en particulier la nécessité d'un versant ouvert et d'unã rivière prochede la falaise, ont été mis en évidence.
, Pour plus d'information, voir: VOGT (D.) 1g7g : Guide de l,habitat pour lele^lTsement, et.l'appréciarion de l'habitat du Faucon pélerin. Traduction ¿e : vocr (o.)l9lð : Eln Habitatschüssel zur Erfassung und Beurteilung von WanderfalkenhabitatenLusciana,
VOçT (D.) 1978 : Untersuchungen zur Habitatstruktur mittelenropai'scher, felænbrü-tendes Wanderfal ken lFa lco perqrinus). D ie Vogelvvett gg, 201 -222.
lll - Motion votée au XXe Colloque interrégional d,ornithologie
La plaine du Forez est l'une des régions d'étangs les plus riches de France aupoint de vue naturaliste :
(1) D-5400 KOBLENTZ, Zwickauesrr. b, R.F.A.

106
- d'importants stationnements hivernaux d'oiseaux d'eau l'ont fait clasær com-me zone d'intérêt national par le bureau MAR (bureau responsable de l'inventaire deszones humides européennes).
- elle a été retenue dès 1970 au préinventaire des richesses naturelles de laLoire.
- elle constitue un sanctuaire important à l'échelle internationale pour la nidi-fication du canard Chipeau, de la nette rousse, de la guifette moustac, du Grèbe à counoir.
Les ornithologues francophones réunis pour leur XXe Colloque interrégionalà Saint-Etienne les 15 et 16 novembre 1980 tiennent à souligner solennellement cetintérêt exceptionnel, et æ déclarent stupéfaits et indignés de la multiplication d'aména-gements lourds en cours de réalisation ou en projet à brève échéance :
- tracé retenu pour l'autoroute B 71 ;
- modification du cours de la Loire et importance des gravières;
- projet d'aérodrome de l'Hôpital-le-Grand ;
- modification accélérée des pratiques agricoles (ensilage, remembrement...) ;
- recalibrage des affluents de la Loire et modifications hydrologiques de toutenature, notamment par l'extension de l'irrigation par aspersion dans les milieux aussiremarquables que les chaninats ;
- sondages et projets d'extraction d'uranium ;
- <mitage> du bocage par les constructions résidentielles.
L'accumulation de ces diverses opérations menace de détruire à brève échéanceun équilibre entre l'espace naturel, les activités agricoles, piscicoles et cynégétiques,fouilibre qui est le fruit d'une longue adaptation de l'activité humaine au milieu foré-zien.
En conséquence, les ornithologues francophones réunis à Saint-Etienne esti-ment indispensables des actions visant à maintenir cet équilibre et à sauvegarder l'es-pace forézien (moyens de préservation de l'espace naturel, gestion concertée, réexa-men des projets d'impact écrasants) et ils se déclarent solidaires de toute action allantdans ce sens.
lV - Table ronde : relations protecteurs de la nature, chasseurs, administrations
Réunis en table ronde sur le thème <relations protecteurs, chaseurs, adminis-trat¡ons), les participants du XXe Colloque d'Ornithologie interrégional de Saint-Etienne

707
les 15 et 16 novembre 1980, ont fàit porter leurs réflexions dans trois directions :
lV-l - Après avoir essayé de définir des possibilités d'intervention etde participationdes naturalistes dans les structures officielles aux niveaux départemental, régional et na-tional, nous constatons :
- Au niveau des structures administrativæ concernées par la faune sauvage ladistribution des attributions est extrêmement confuse.
Exemple : au niveau du département læ compétences en matière de faune sau-vage (dont le gibier) sont attr¡buées quasi exclusivement au Directeur départemental del'Agriculture qui dépend du Ministère de l'Agriculture, alors qu'au niveau national cesmêmes attributions sont dévolues à la Direction de Protection de la Nature du Ministèrede l'Environnement et du Cadre de Vie.
- Sauf exception, les administrations départementales chargées des problèmesconcernant la faune sauvage et leur habitat n'ont pas les connaissances suffisantes dansces domaines.
- ll n'existe aucune inærtion officielle des naturalistes non chasæurs dans lesorganismes chargés de la gestion de la faune sauvage et de la chasse au niveau départe-mental.
Exemple :dans la commission départementale du plan de chasse, notre panici-pation n'est que consultative et ne dépend que du bon vouloir du Préfet.
- Le röle du conseiller biologiste ne dépend que du bon vouloir des fonction-naires et le Ministère de l'Environnement et du Cadre de Vie n'a pas encore reconnuoff iciellement leur participation.
lV-2 - Nous prenons acte de la campagne nationale menée par des responsables cyné-gétiques importants visant, tout en essayant de conserver aux yeux du grand public uneimage de marque de protecteur, à sauvegarder la pratique actuelle de la chasæ en Francesans tenir compte des connaissances acquises.
Exemple : ouverture anticipée de la chasæchasse de nuit, etc...
- Nous prenons acte des nombreuses déclarations publiques de ces mêmes res-ponsables qui prônent un retour à des pratiques officiellement condamnées :
Exemple : déclassement d'oiseaux protégésutilisation de poison rémanent (strychnine). Cf déclarations de
Monsieur HAMELIN, Président des présidents de Fédérations de Chasse, Prési-dent des chasseurs européens, Revue nationale de la Chasæ,397, oct. 1980.

108
lV'3' Nous réclamons la prise en compte des positíons officielles de la Fédérationfrançaise des Sociétés de Protection de la Nature sur la chasse :
- organiser des structures de gestion de la faune sauvage où scientifiques, chas-seurs, non chasseurs seraient représentés à égalité à tous les niveaux.
- subordonner la délivrance de tous permis à une vérification des connaissan-ces biologiques (examen obligatoire pour tous les chasseurs).
- asseoir le prélèvement sur des études et des inventaires permanents.
- limiter le prélèvement en fonction des impératifs écologiques et éthologiquesdes espèces et des biocoénoses.
- laisser aux propriétaires non chasseurs la possibilité d'interdire la chasse chezeux.
- donner la priorité à la conservation et à l'aménagement des milieux favora-blesàlafaunesauvagedanslesbudgetscynégétiques.
- arriver à supprimer tout commerce de gibier vivant, mort ou transformé,priorité étant accordée aux espècæ migratrices ou aux espèces rares. Au repeuplementartificiel doit se substituer une bonne organisation du territoire (réserves) et du prélè-vement (plan de chasse).
- moderniser et distinguer très nettement de la chasæ la limitation des espècesqui posent des problèmes aux activités humaines.
- application immédiate se traduisant dans les faits des connaissances déjàacq u ises.
- application immédiate et stricte de la législation existante et d'une réglemen-tation précise et adaptée en fonction des connaissances et orientations nouvelles.
- Nous engageons les associations de protection de la nature à poursuivre ledialogue avec les chasæurs responsables (à tous les niveaux) quand ceux-ci ont fait lapreuve de leur volonté d'appliquer les principes modernes de gestion de la faune sau-vage.
- Nous engageons par ailleurs les sociétés de protection de la nature à utiliserdavantage et plus efficacement les moyens d'information touchant le grand public.
Ouelques points précis sont suggérés dans ce sens :
- organisation de stages de formation de relations publiques ;- faire des communiqués de prese plus fréquents à tous les niveaux ;- inviter aux conférences de presse des naturalistes, les responsables officiels
de la protection de la nature.

109Le Bièvre, 1981,3(1)
Michel BOURBON (1)
DEUX NOUVEAUX NICHEURS PROBABLES POUR LA GORSE :
HIBOU GRAND-DUC (Bubo bubo)
HIBOU MOYEN-DUC (Asio otus)
Si le Hibou petit duc (Otus scos) apparaît à un quelconque observateur commeétonnamment commun en Corse (THIBAULT, 1978), par contre ies deux autres ne sem-blent pas connus comme nicheurs ou simplement soupçonnés (yEATMAN,1gi6).
Le premier juillet 1975 je remontais, à la nuit tombante, la petite route qui vade Lozari à Palasca en Balagne ; je tentais d'établir, en sifflant, un contact rapprochéavec les (petits Ducs> qui sont ici assez nombreux, lorsque j'entendis distinctement lescris plaintifs et aigus de mendicité de jeunes hiboux. lls provenaient d'une oliveraie quime paraissait abandonnée tant elle était envahie par les Cistes hauts de un à deux mè-treg; là, deux jeunes Hiboux d'une taille nettement supérieure au (petit duc>, perchésau sommet d'un olivier, me considéraient de leurs grands yeux oranges ébahis ; i,und'eux montra¡t nettement ses aigrettes et par deux fois un ou deux ãdultes me survolaen tentant l'intimidation. Comme la nuit s'épaississait et que je ne désirais pas provoquerunrdérangement prolongé, j'interrompis mon obærvation avec l'intention de la repren-dre plus tôt le lendemain. Or le lendemain les cris de mendicité s'étaient fortementéloignés et dispersés dans l'océan des Cistes; il me parut, en tout cas, difficile de rejoin_dre efficacement les animaux. Le surlendemain, plus rien.
Cette observation incomplète et acc¡dentelle témoignerait de la reproduction duHibou moyen duc en Corse, car la seule confusion qui mérite quelque attention, vue lataille et l'allure des oiseaux, est celle du Hibou brachyote, sans doute encore moinsvraisemblable ici. Par ailleurs, il ærait impossible de considérer que ces oiæaux a¡ent puparcourir une grande distance avant que je les observe ; pourtant, les résineux qui ontordinairement leur préférence pour l'installation de la couvée sont absents ici, sauf unetrès jeune plantation de Pins maritimes à Lozari; par contre, les corneilles (corvuscorone cornixl dont les vieux nids sont fréquemment utilisés, sont abondantes ici.
Au reste, le Hibou moyen duc est une espèce suffisamment discrète pour quesa reproduction puisæ paser inaperçue et ce sont, bien souvent, les cris persistants desjeunes qui att¡rent l'attention.
(1) C.O.R.A., Universiré Lyon t, 69622 VTLLEURBANNE CEDEX.

110
Le 7 juillet 1975, Michel Dupupet (également membre du C.O.R.A.) et moi'même descendions de la Bocca Stranciacone à Barghiana en suivant le ruisseau de Taita.affluent du Fango dans la forêt communale du Filosorma ; lorsqu'à la nuit tombantenous installâmes notre bivouac parmi les laissées de Mouflons, nous entendimes distinc-tement les chants de Hiboux grands ducs : le mâle d'abord, puis la réponse perlée etdescendante de la femelle. Ces chants semblaient provenir d'une gorge rocheuse domi-née par la Punta Lucsi.
A cette époque les chants de Hiboux grands ducs sont plutôt rares et, commeprécédemment, ce sont surtout les cris de mendicité des jeunes qui attirent l'attention ;
cependant, l'éloignement et les grondements du ruisseau de Taita ne nous permirentpas de percevoir pendant toute la nuit ces cris de mendicité.
Les 21 et 22 mai 1979 une nouvelle expédition nous fit prospecter le versant
nord de la Stranciacone, par le ruisseau de Maghine, mais aucune observation ne putêtre faite pendant les deux nuits passées à Broximité de l'abri de Maghine.
Encore plus fragmentaire, cette observation n'a certainement pas la prétentiond'établir la reproduction du Hibou grand duc en Corse mais d'attirer l'attention des ob-
servateurs éventuels sur une vallée particulièrement déserte et difficile d'accès où ce
rapace nocturne s'est probablement installé.
En conclusion, il apparait que I'avifaune de la Corse s'enrichit encore assez
régulièrement d'espèces nouvelles à mesure que le flot touristique déverse ses pâles
cont¡ngents de vacanciers dans des endroits insolites ou d'intérêt secondaire. La liste
des espèces nicheuæs est sans doute bientôt close ; quant aux espèces migratrices, des
régions entières, en raison de leurs difficultés d'accès dans un milieu trop hostile ou,pour leur banalité, réservent encore des surprises ; à preuve de récentes observationsdans le désert des Agriates du 24 au 27 mai 1979 :
- Pluvier argenté (Pluvialis squatarolal , un individu sur les dunes de l'Ostriconi.- Echasse blanche (Himantopus himantopusl , un individu.- Héron crabier (Ardeola ralloidesl , un adulte en plumage nuptial à l'étang de
Loto.
BibliographieBOURBON (M.) 1977 - Le désert des Agriates -Courrier Nature,50, 175-1 81 .
BRUN (8. & L.), CONRAD (M.), GAMISAN (J.) 1975 - CORSE - Horizon de France,222 p.
GEROUDET (P.) 1965 - Les Rapaces diurnes et nocturnes d'Europ - Delachaux & Niestlé,Neucháre|,426 p.
THIBAULT (J.C.) 1978 - Statut et effectifs des Rapaces en Corse - Courrier Parc Corse,30,6-31 .
YEATMAN (L.) 1971 - Histoire des oiseaux d'Europe - Bordas, Paris - 362 p.
YEATMAN (L.) 1976 - Atlas des oiseaux nicheurs de France - Ministère de la Oualité de la Vieet de l'Environnement, Paris,2B2 p.


















