
III-1 : LA RETINE : ORGANISATION...
49
Œil et Physiologie de la Vision – III-1 III-1 : LA RETINE : ORGANISATION SCHEMATIQUE Florence Rigaudière Pour citer ce document Florence Rigaudière, «III-1 : LA RETINE : ORGANISATION SCHEMATIQUE», Oeil et physiologie de la vision [En ligne], III-La physiologie rétinienne, mis à jour le 18/06/2013, URL : http://lodel.irevues.inist.fr/oeiletphysiologiedelavision/index.php?id=212, doi:10.4267/oeiletphysiologiedelavision.212 Plan I - Neurorétine Trois étages Premier étage : les cônes et les bâtonnets Les cônes Les bâtonnets Relation avec l’épithélium pigmentaire Couche plexiforme externe : les cellules horizontales Cellules HI Cellules HII Cellules HIII Jonctions gap entre cellules horizontales Deuxième étage rétinien : les cellules bipolaires Généralités Cellules bipolaires de cônes L ou M, naines Cellules bipolaires de cônes L et M, diffuses Deux maillages pour les cônes L et/ou M Cellules bipolaires de cônes S Cellules bipolaires de bâtonnets Cellules interplexiformes Couche plexiforme interne : les cellules amacrines Des cellules spécifiques Parmi vingt variétés, une essentielle : la cellule amacrine AII Troisième étage : les cellules ganglionnaires Nombre et densité Particularités Cellules ganglionnaires naines Cellules ganglionnaires parasols Cellules ganglionnaires bistratifiées Autres cellules ganglionnaires à destination extragéniculée Trois voies rétinocorticales : organisation et destinée Voie P Voie M Voie K Séparation des trois voies jusqu’au cortex Tests des voies P et K 1
Transcript of III-1 : LA RETINE : ORGANISATION...

Œil et Physiologie de la Vision – III-1
III-1 : LA RETINE : ORGANISATION SCHEMATIQUE Florence Rigaudière
Pour citer ce document
Florence Rigaudière, «III-1 : LA RETINE : ORGANISATION SCHEMATIQUE», Oeil et physiologie de la vision [En ligne], III-La physiologie rétinienne, mis à jour le 18/06/2013, URL : http://lodel.irevues.inist.fr/oeiletphysiologiedelavision/index.php?id=212, doi:10.4267/oeiletphysiologiedelavision.212
Plan
I - Neurorétine
Trois étages Premier étage : les cônes et les bâtonnets
Les cônes Les bâtonnets
Relation avec l’épithélium pigmentaire Couche plexiforme externe : les cellules horizontales
Cellules HI Cellules HII Cellules HIII Jonctions gap entre cellules horizontales
Deuxième étage rétinien : les cellules bipolaires Généralités Cellules bipolaires de cônes L ou M, naines Cellules bipolaires de cônes L et M, diffuses Deux maillages pour les cônes L et/ou M Cellules bipolaires de cônes S Cellules bipolaires de bâtonnets
Cellules interplexiformes Couche plexiforme interne : les cellules amacrines
Des cellules spécifiques Parmi vingt variétés, une essentielle : la cellule amacrine AII
Troisième étage : les cellules ganglionnaires Nombre et densité Particularités Cellules ganglionnaires naines Cellules ganglionnaires parasols Cellules ganglionnaires bistratifiées Autres cellules ganglionnaires à destination extragéniculée
Trois voies rétinocorticales : organisation et destinée Voie P Voie M Voie K Séparation des trois voies jusqu’au cortex Tests des voies P et K
1

Œil et Physiologie de la Vision – III-1
Deux systèmes rétiniens: photopique et scotopique Système photopique Système scotopique Explorations différentielles du système photopique et scotopique
Les cellules de Müller
II - L’épithélium pigmentaire Description Vascularisation
III - Application à la lecture des images rétiniennes en OCT
Conclusion
Texte intégral
Remerciements à C. Curcio pour ses illustrations.
La rétine est une structure complexe organisée en deux parties : la neurorétine et l’épithélium pigmentaire.
La neurorétine est essentiellement « photosensible ». Elle est capable de convertir les photons lumineux en influx visuels transmis jusqu’aux différentes aires visuelles pour aboutir à la vision. Elle peut être explorée tant en surface, que selon ses strates par des tests fonctionnels d’électrophysiologie visuelle.
Une partie de la neurorétine dont l’importance fonctionnelle se révèle grandissante, est « luminosensible ». Elle réagit à la lumière et à ses caractéristiques, ne permet pas de voir, mais de réguler l’humeur et le cycle nycthéméral. Il en sera dit un mot, bien que cette neurorétine ne soit pas actuellement explorable par des tests visuels cliniques.
En complément des explorations fonctionnelles rétiniennes par électrophysiologie, l’imagerie anatomique rétinienne par OCT (Optical Coherence Tomography) est de pratique courante.
Mettre en parallèle les images obtenues par OCT et les différentes structures rétiniennes -récepteurs, couches nucléaires, épithélium pigmentaire- permettront d’en mieux situer les anomalies.
I - Neurorétine
Seule l’architecture des principaux éléments utiles à la compréhension de l’exploration visuelle clinique est présentée ici ; ses aspects fonctionnels sont développés dans la partie suivante III-2, ses caractéristiques macroscopiques et vasculaires sont décrites au chapitre IV.
La structure de la neurotéine peut être regroupée selon trois étages, trois voies (P, M, et K) et deux systèmes photopique et scotopique. L’exposé ci-dessous des grands piliers de son architecture résume les travaux effectués chez l’homme et les grands primates dont le système visuel est proche de celui de l’humain [Kolb H, 2006], [Kolb H et al.].
2

Œil et Physiologie de la Vision – III-1
Trois étages
D’un point de vue fonctionnel, la neurorétine peut être stratifiée en trois étages (figure III-1-1), sauf au niveau de la fovéola correspondant à un seul étage, constitué essentiellement par les articles externes de cônes (figure III-1-2).
Remarque importante. Figure III-1-3. Convention d’orientation dans la description de la neurorétine. Elle correspond au sens de propagation du signal électrophysiologique qui est initié par les photorécepteurs ou 1er étage. La propagation se poursuit schématiquement vers les cellules bipolaires ou 2ième étage ; la sous couche-a de la couche plexiforme interne est rencontrée la première, elle est dite sous couche-a superficielle, la sous couche-b vient ensuite, elle est dite sous couche-b profonde. Le signal atteint enfin les cellules ganglionnaires ou 3ième étage ; la propagation se poursuit le long des nerfs optiques…
Les photorécepteurs forment le premier étage rétinien ou étage réceptoral. Ils font synapse à la couche plexiforme externe avec des cellules d’association, les horizontales et leurs cellules bipolaires qui constituent le deuxième étage rétinien jusqu’à la couche plexiforme interne. Cette dernière est essentiellement divisée en deux sous couches, la sous couche-a, superficielle et la sous couche-b, plus profonde. Le troisième étage correspond aux cellules ganglionnaires qui font synapse à la couche plexiforme interne avec les cellules bipolaires sus-jacentes et des cellules d’association, les amacrines (figure III-1-1).
Les axones des cellules ganglionnaires se myélinisent à partir de la lame criblée de la papille, pour former les fibres des nerfs optiques.
Premier étage : les cônes et les bâtonnets
Les cônes
Les cônes sont répartis sur toute la rétine et cohabitent avec les bâtonnets sauf au niveau de la fovéola où ils sont seuls (figure III-1-2, figure III-1-4)
Nombre Le comptage des cônes chez l’homme s’est effectué sur des prélèvements de fragments de tissu rétinien [Curcio CA et al., 1990] ou grâce à des méthodes de visualisation des photorécepteurs in vivo [Miller DT et al., 1996], [Delint PJ et al., 1997], [Marcos S et al., 1997], [Roorda A, 2000].
Chez l’adulte, les cônes sont en moyenne 4,6 millions (de 4 à 5,3 millions selon les sujets analysés), soit 20 fois moins environ que le nombre des bâtonnets (92 millions) [Curcio CA et al., 1990]. Ils sont répartis sur toute la rétine, de façon plus régulière en rétine temporale qu’en rétine nasale et ce, à tous les âges et pour les deux sexes [Kimble TD, Williams RW, 2000].
En dehors de la fovéola… …des bâtonnets s’intercalent régulièrement en mosaïque, entre les cônes. Les 10 degrés centraux contiennent 500 000 cônes environ, soit moins de 10% du total. Au-delà de six degrés d'excentricité, un cône peut être complètement entouré de bâtonnets voire jusqu'à 48 bâtonnets (figure III-1-5).
La fovéola C’est une exception rétinienne avec un seul étage rétinien. Sur cette surface très restreinte (< 0,1mm2), il n’y a que 10 000 cônes environ [Wassle H et al., 1989].
3

Œil et Physiologie de la Vision – III-1
Elle abrite essentiellement les articles externes des cônes et 300 à 500 noyaux de leurs articles internes [Ahnelt PK, 1998]. La majeure partie des noyaux des cônes fovéolaires sont déjetés en dehors de la dépression fovéale ; ils forment ses berges ou clivus fovéal. Les synapses de ces cônes avec les cellules sous-jacentes se font en dehors de la fovéola.
Densité La densité des cônes est maximale à la fovéola. Elle varie entre 300 000/mm2 et 80 000/mm² selon les sujets ; c’est la plus importante de toute la rétine (figure III-1-6). Elle décline très rapidement en dehors des quelques degrés centraux pour atteindre une valeur d’environ 5000/mm² à quelques degrés d’excentricité.
Article externe
Morphologie Pour une même région rétinienne et d'un point de vue histologique, les articles externes de cônes adjacents sont semblables et de forme conique (figure III-1-7, figure III-1-8) d'où leur nom. Ils ne peuvent être différenciés que par l’utilisation de coloration spécifique (figure III-1-9) [McCrane EP et al., 1983], [Curcio CA et al., 1991].
L’article externe est constitué d’une seule membrane qui sépare le milieu extracellulaire du milieu intracellulaire. Une partie de cette membrane présente des plis sur lesquels se trouvent inclus les photopigments des cônes qui sont ainsi en relation directe avec le milieu extracellulaire (figure III-1-7).
Le diamètre de l'article externe des cônes varie selon les individus (figure III-1-10) et en fonction de l’excentricité, de 1,5 µm environ à la fovéola [Curcio CA, Hendrickson AE, 1991] à 6 µm environ à la périphérie. Son diamètre est deux à trois fois supérieur à celui des bâtonnets et ce, pour toutes les excentricités rétiniennes (figure III-1-5).
La longueur des cônes varie entre 80 µm en zone fovéolaire et 22 µm à la périphérie [Willmer EN, 1987]. En un lieu donné, elle est constante grâce à un équilibre entre la phagocytose de son extrémité distale par l’épithélium pigmentaire et sa croissance qui se situe au niveau du cil connecteur à l’article interne (figure III-1-8) [Anderson DH et al., 1978], [Steinberg RH et al., 1980] [Boesze-Battaglia K, Goldberg AF, 2002], [Kevany BM, Palczewski K, 2010].
Trois photopigments différents pour trois types de cônes L, M, S Les photopigments portés par la membrane des articles externes sont de compositions différentes, à l’origine des propriétés caractéristiques de trois types de cônes dits L, M ou S selon leur probabilité d’absorption des photons, en fonction de la longueur d’onde de la stimulation.
Les cônes L présentent une probabilité maximale d’absorption des photons pour les grandes longueurs d’onde (Long wavelengths) située vers 560 nm, les cônes M, une probabilité maximale d’absorption des photons pour les moyennes longueurs d’onde (Middle wavelengths) située vers 530 nm et les cônes S, une probabilité maximale d’absorption des photons pour les courtes longueurs d’onde (Short wavelengths) située vers 420 nm (figure II-17).
Ces trois types de cônes se répartissent en deux groupes, les cônes L et M d’une part et les cônes S d’autre part. Ils sont de nombre, de répartition et d’origines différentes.
Cônes L et M Les cônes L et M sont les plus nombreux environ 90 % des 4 à 5,3 millions de cônes ; ils sont présents sur toute la rétine [Curcio CA et al., 1990], [Schiller PH, 1996].
4

Œil et Physiologie de la Vision – III-1
Leurs photopigments L et M sont codés par des gènes différents, situés à proximité l’un de l’autre sur le chromosome X [Sharpe LT et al., 1999].
La composition anormale ou la déficience de l’un ou l’autre des photopigments ou des deux, sont à l’origine de dyschromatopsies héréditaires liées à l’X [Rigaudiere F et al., 2006].
La proportion relative des cônes L et M chez l’homme est très variable d’un individu à l’autre. Elle peut osciller de 70% de cônes L avec 20% de cônes M [Cicerone CM, Nerger JL, 1989], [Kremers J et al., 1999], [Lennie P, 2000] à des valeurs plus extrêmes allant de 83% à 36% de cônes L, pour 7% à 54% de cônes M [Carroll J et al., 2000].
Les cônes L sont sensiblement deux fois plus nombreux que les cônes M dans la fovéola [Cicerone CM, Nerger JL, 1989], en parafovéa (c’est-à-dire jusqu’à quatre degrés d’excentricité [Nerger JL, Cicerone CM, 1992]) et probablement aussi pour des excentricités supérieures [Otake S et al., 2000]. Mais ils se répartissent au hasard l’un par rapport à l’autre [Roorda A, Williams DR, 1999], [Roorda A et al., 2001] plutôt que de façon homogène et isotrope [Gowdy PD, Cicerone CM, 1998] (figure II-16).
Cônes S Les cônes S correspondent à 10 % environ de tous les cônes [Ahnelt PK et al., 1987]. Le photopigment S porté par leur article externe est codé par le chromosome 7 [Fitzgibbon J et al., 1994].
Ils sont absents du cœur de la fovéola sur un diamètre d’environ 100 µm ou surface vue sous un angle de 20’ environ (0,35°) [Bumsted K, Hendrickson A, 1999], [Curcio CA et al., 1991] (figure III-1-11). Les cônes L et M y sont seuls présents, à l’origine d’un phénomène connu de longue date : la tritanopie fovéale [Williams DR et al., 1981].
La vision des couleurs normale se fait à l’aide de la synthèse des signaux issus des trois photopigments décrits : L (ou premier pigment), M (ou deuxième pigment) et S (ou troisième pigment). La tritanopie correspond à an-opie (en grec) : absence de vision, du troisième pigment : tri.
Ceci signifie que sur cette surface très restreinte, la vision des couleurs est dichromate, ne se faisant que grâce à la synthèse des signaux issus de deux types de cônes présents L et M.
En dehors de cette zone, un cône S peut être entouré par 5, 6 ou 7 cônes L et M [Ahnelt PK et al., 1987], ils se répartissent de façon régulière sur toute la rétine [Calkins DJ, 2001], sous forme d’une mosaïque de géométrie intermédiaire entre un triangle, un carré [De Monasterio FM et al., 1985], [Curcio CA, Hendrickson AE, 1991] ou un hexagone (figure III-1-12).
Les cônes S n’ont pas la même densité sur toute la rétine. La majorité des cônes S se trouvent dans les 12 degrés centraux avec une densité maximale allant de 1000 à 5000/mm2 sur un anneau situé entre 0,75 et 1,5° d’excentricité (berge fovéale) [Ahnelt PK, 1998], la densité des cônes S étant de l’ordre de 1000/mm2 sur le reste de la rétine [Curcio CA et al., 1991], [De Monasterio FM et al., 1985] (figure III-1-13).
Jonction article externe-article interne, article interne L’article externe est connecté à l’article interne par un cil (figure III-1-8), zone de croissance régulière de l’article externe [Roepman R, Wolfrum U, 2007], [Trojan P et al., 2008]. Cette croissance se fait par évagination de la membrane au niveau du cil connecteur article externe-article interne, avec des modalités différentes pour les cônes et les bâtonnets [Eckmiller M, 1997].
L’article interne contient le noyau et tous les organites nécessaires au fonctionnement métabolique du cône.
5

Œil et Physiologie de la Vision – III-1
Synapses chimiques à la couche plexiforme externe Si les articles externe et interne des trois types de cônes sont semblables, leurs zones synaptiques avec les cellules bipolaires et horizontales sont de morphologies différentes bien que difficilement visibles sur des coupes histologiques.
Les cônes L et M ont un pédicule large tandis que les cônes S ont un pédicule étroit [Kolb H et al., 1997] (figure III-1-14). Cette variation morphologique correspond à des modes de jonctions synaptiques différentes.
Pédicules larges Les pédicules des cônes L et M sont de forme évasée et pyramidale. Ils sont le lieu de deux types de synapses avec leurs cellules sous-jacentes : des synapses invaginantes et des synapses par contacts superficiels (figure III-1-15, figure III-1-16).
°Synapses invaginantes. Elles correspondent aux invaginations des dendrites des cellules bipolaires de cônes dites ON, entourées par plusieurs dendrites des cellules horizontales HI, HII et HIII, dans de petites logettes du pédicule.
Les synapses invaginantes sont des synapses chimiques avec °une zone présynaptique au niveau de la membrane des cônes et des vésicules de stockage du neurotransmetteur : le glutamate, °un espace intersynaptique et °des sites récepteurs spécifiques en zone postsynaptique au niveau des cellules bipolaires de cônes. La transmission de l’influx est unidirectionnelle.
Le nombre de synapses invaginantes par pédicule de cônes L ou M est de 22 à 25, en zone fovéale [Calkins DJ et al., 1996] et d’environ 40, à la périphérie vers 30° d'excentricité [Chun MH et al., 1996] La majorité des invaginations est dévolue aux cellules bipolaires naines ON (environ 80%), tandis que les autres sont utilisées par les cellules bipolaires diffuses ON (environ 20%) [Calkins DJ et al., 1996].
°Synapses par contacts superficiels. Les synapses se font à la surface du pédicule par contact des dendrites de cellules bipolaires de cônes dites OFF (soit juste de part et d'autre des synapses invaginantes pour les cellules bipolaires naines OFF, soit plus à distance des synapses invaginantes, pour les cellules bipolaires diffuses OFF) (figure III-1-16). Ce sont aussi des synapses chimiques, différentes des précédentes. On ne retrouve aucune vésicule dans la zone présynaptique des cônes alors que la zone postsynaptique des cellules bipolaires OFF de cônes présente bien les caractéristiques d'une membrane postsynaptique.
Les synapses par contacts superficiels sont beaucoup plus nombreuses que les synapses invaginantes, entre 180 à 270 [Chun MH et al., 1996].
Remarque. Le glutamate est le neurotransmetteur libéré au niveau de ces deux types de synapses invaginantes ou par contacts superficiels.
Pédicules étroits Ce sont les pédicules des cônes S. Ils sont de morphologie complexe, bilobée et font des synapses uniquement invaginantes avec les dendrites de cellules bipolaires de cônes S et les cellules horizontales HII [Haverkamp S et al., 2000]. Ce sont des synapses chimiques ayant pour neurotransmetteur le glutamate.
Corrélation anatomo-fonctionnelle Ce mode de connexion synaptique avec les cellules bipolaires sous-jacentes par invaginations d’une part et par contacts superficiels d’autre part, est corrélé à leur mode de fonctionnement.
6

Œil et Physiologie de la Vision – III-1
Cellules bipolaires ON Les cellules bipolaires de cônes qui s’invaginent dans les pédicules de cônes, se terminent toutes à la sous couche-b de la couche plexiforme interne.
Elles se dépolarisent lorsque le cône avec lequel elles sont directement liées, est stimulé ; leur réponse est dite de type ON. Elles sont dites cellules bipolaires ON.
Cellules bipolaires OFF Les cellules bipolaires de cônes qui font synapse par contacts superficiels avec les pédicules des cônes, se terminent toutes à la sous couche-a de la couche plexiforme interne.
Elles s’hyperpolarisent lorsque le cône avec lequel elles sont directement liées, est stimulé ; leur réponse est dite de type OFF. Elles sont dites cellules bipolaires OFF (figure III-1-17).
Remarque. Schématiquement, une cellule se dépolarise lorsque la différence de potentiel entre son milieu intérieur et extérieur diminue, facilitant ainsi son excitation. Une cellule s’hyperpolarise lorsque la différence de potentiel entre son milieu intérieur et extérieur augmente, rendant plus difficile son excitation.
Jonctions gap Un autre type de synapse interconnecte les cônes entre eux mais aussi les cônes et les bâtonnets : ce sont les jonctions gap. Ces jonctions correspondent à des zones latérales de contiguïté entre six à douze expansions du pédicule du cône avec le sphérule du bâtonnet.
Les jonctions gap sont formées par les protéines transmembranaires de deux membranes plasmiques accolées ou canaux ioniques. Elles diffèrent des synapses chimiques orientées où les zones présynaptiques, intersynaptiques et postsynpatiques sont bien individualisées.
Leur fonctionnement est différent de celui des synapses chimiques puisqu’un passage rapide d'ions et de petites molécules peut s’effectuer de part et d'autre du point de contact, dans les deux sens, assurant ainsi une transmission rapide de l'information.
Les bâtonnets
Nombre et densité Chez l'adulte, les bâtonnets sont en moyenne de 92 millions (78 - 108 millions) [Curcio CA et al., 1990]. Ils sont présents sur toute la rétine, intercalés régulièrement entre les cônes (figure III-1-5) sauf à la fovéola où ils en sont absents.
En dehors de la fovéola, leur densité croît régulièrement jusqu'à une zone comprise entre 10° et 20° d'excentricité où elle est maximale, d'une valeur proche de 176 000/mm2, puis la densité décroît régulièrement pour atteindre une valeur de l'ordre de 60 000/mm2 à la périphérie. La densité des bâtonnets est légèrement supérieure en rétine nasale qu'en rétine temporale, de même en rétine supérieure qu'inférieure.
Article externe L'article externe des bâtonnets est de forme cylindrique (figure III-1-8). Il est limité par une membrane externe qui sépare le milieu extracellulaire du milieu intracellulaire dans lequel flotte un empilement de 700 à 1 100 disques selon la localisation [Young RW, 1971] (figure III-1-7).
La membrane de chaque disque est complètement indépendante de la membrane externe. Elle sépare le milieu intracellulaire du milieu intradiscal. Les molécules de rhodopsine, pigments photosensibles des bâtonnets, sont incluses sur cette membrane discale (figure III-1-7).
7

Œil et Physiologie de la Vision – III-1
Le diamètre de l'article externe d’un bâtonnet varie entre 1 µm dans la région fovéale et 2,5 µm à la périphérie, sa longueur est d'environ 30 µm.
Jonction article externe-article interne L’article externe est connecté à l'article interne par un cil (figure III-1-8). Comme pour le cône, c’est à ce niveau que l’article externe croît régulièrement, assurant ainsi son maintien durant la phagocytose de son extrémité par l'épithélium pigmentaire.
Article interne Il contient le noyau ainsi que les organites nécessaires au fonctionnement métabolique des bâtonnets et à la régénération des disques. Les articles internes des bâtonnets sont plus effilés que ceux des cônes.
Synapses à la couche plexiforme externe Les synapses des bâtonnets ont la forme d’une petite sphère, dite sphérule. Elles font °des synapses chimiques avec les cellules sous-jacentes et °des jonctions gap, entre bâtonnets et cônes adjacents et entre bâtonnets.
Synapses chimiques entre bâtonnets et cellules bipolaires de bâtonnets Deux à sept dendrites de cellules bipolaires de bâtonnets s’invaginent à l’intérieur des sphérules des bâtonnets, entourées des terminaisons axoniques des cellules horizontales HI (figure III-1-18).
Les sphérules de bâtonnets présentent une zone membranaire présynaptique qui contient des vésicules de stockage du neurotransmetteur : le glutamate.
Les cellules bipolaires de bâtonnets se terminent toutes à la sous couche-b de la couche plexiforme interne ; elles se dépolarisent lorsque les bâtonnets sont stimulées. Ce sont des cellules bipolaires ON.
Jonctions gap entre bâtonnets et entre cônes et bâtonnets Elles sont nombreuses entre sphérules de bâtonnets mais aussi entre pédicules de cônes et sphérules de bâtonnets adjacents comme il a déjà été dit.
Ces jonctions gap sont particulièrement actives lorsque les niveaux lumineux sont mésopiques voire photopiques [Bloomfield SA, Dacheux RF, 2001].
Relation avec l’épithélium pigmentaire
La partie distale de l'article externe des photorécepteurs se situe entre les expansions apicales de l'épithélium pigmentaire (figure III-1-19). Ce couplage est important d’un point de vue fonctionnel.
Couche plexiforme externe : les cellules horizontales
Ce sont les cellules d’association de la couche plexiforme externe. Trois variétés de cellules horizontales ont été identifiées dans la rétine humaine : les cellules HI, HII et HIII [Kolb H et al., 1992], [Kolb H et al., 1994], [Wassle H et al., 2000] (figure III-1-20).
8

Œil et Physiologie de la Vision – III-1
Cellules HI
Leurs dendrites ont une arborisation exubérante, radiante, la largeur de leur champ dendritique est croissante du centre vers la périphérie.
Rappel de la définition du champ dentritique ou axonique : c’est la surface de neurorétine occupée par l’ensemble de l’arborisation dendritique ou des terminaisons axoniques (figure III-1-21).
Elles font des synapses invaginantes dans les pédicules de cônes voisins L et M, -sept puis neuf puis 15 à 20 plus à la périphérie- où elles entourent les dendrites de cellules bipolaires invaginantes [Dacey DM et al., 1996] (figure III-1-15, figure III-1-16).
Leur axone est épais (2 µm) de longueur variable allant de quelques micromètres à 100 µm ; il est généralement issu du corps cellulaire. Il se termine dans le sphérule de 350 à 500 bâtonnets.
Cellules HII
Elles sont moins nombreuses que les cellules HI avec une proportion variable selon l’excentricité [Wassle H et al., 2000]. Leurs dendrites ont une arborisation très fine, d'aspect laineux et entrelacé, ce qui les distingue des dendrites des cellules HI.
La taille de leurs champs dendritiques est soit plus grande, soit plus petite que celle des cellules HI, selon l'excentricité.
Leur mode de connexion synaptique avec les cônes est plus difficilement analysable à cause des convergences ou recouvrements dendritiques vers les cônes. Il s’agit vraisemblablement de synapses invaginantes avec des cônes L et M d’une part et des cônes S d’autre part et ce, pour toutes les excentricités [Dacey DM, 2000].
Leur axone est généralement issu de l'extrémité d'une dendrite ; il est plus fin (0,5 µm) que celui des cellules HI. Il sinue dans la couche plexiforme externe sur une grande distance pouvant aller jusqu'à 300 µm et se termine dans le pédicule de cônes S.
Cellules HIII
Elles ne se différencient franchement des cellules HI qu'à une excentricité d'environ 12°, par un corps cellulaire plus grand que celui des cellules HI, mais surtout par une arborisation dendritique asymétrique et de champ large.
Leurs synapses se feraient de façon irrégulière en manquant certains cônes adjacents, contrairement aux synapses faites par HI où tous les cônes voisins sont connectés. Cependant, le nombre de cônes interconnectés, probablement de type L et M, est de 20 à 30% supérieur à celui connecté par les cellules HI.
Le corps cellulaire des cellules HIII émet un axone dont la terminaison est difficile à identifier ; il émet aussi des prolongements qui peuvent atteindre la sous couche-a de la couche plexiforme interne.
Jonctions gap entre cellules horizontales
Les cellules horizontales communiquent entre elles par de nombreuses jonctions gap situées non seulement entre les corps cellulaires, mais aussi entre leurs dendrites et leurs terminaisons axoniques.
9

Œil et Physiologie de la Vision – III-1
Deuxième étage rétinien : les cellules bipolaires
Les cellules bipolaires forment le deuxième étage rétinien. Elles sont le lien entre les photorécepteurs et les cellules ganglionnaires et vont de la couche plexiforme externe à la couche plexiforme interne. Elles sont visibles sur une couche histologique essentiellement grâce à l’ensemble de leurs corps cellulaires (figure III-1-1).
Elles se répartissent en cellules bipolaires de cônes à l’origine de trois voies P, M et K et cellules bipolaires de bâtonnets.
Généralités
Les cellules bipolaires sont de disposition radiaire. Leurs dendrites sont situées au niveau de la couche plexiforme externe ; elles font synapse avec les photorécepteurs, soit par invaginations à l’intérieur des pédicules des cônes L, M, S et des sphérules des bâtonnets, soit par contacts superficiels uniquement avec les pédicules des cônes L et M.
Leurs axones se terminent à la couche plexiforme interne sur l’une ou l’autre des deux sous couches principales : la sous couche-a, la plus superficielle ou la sous couche-b, la plus profonde.
La sous couche-a est le lieu de terminaison des bipolaires de cônes qui font synapse par contacts superficiels avec les pédicules de cônes L et M. Elles s’hyperpolarisent lorsque les cônes L et/ou M sont stimulés. Ce sont les cellules bipolaires OFF de cônes.
La sous couche-b est celle de terminaison des bipolaires qui font synapses par invaginations avec les photorécepteurs. Elles se dépolarisent lorsque les photorécepteurs sont stimulés. Ce sont les bipolaires de cônes ON ou les bipolaires de bâtonnets ON.
Il existe des variations de terminaison en des sous niveaux différents (ou strates) des sous couches-a et –b selon les variétés de cellules bipolaires. Il est renvoyé à l'article de Kolb H. et coll. (1992) pour plus de détails. [Kolb H et al., 1992].
Sur 150 rétines humaines analysées, neuf variétés de cellules bipolaires ont été identifiées [Kolb H et al., 1992]. Elles se répartissent en quatre groupes : les cellules bipolaires de cônes soit L, soit M (ou bipolaires naines), les cellules bipolaires de cônes L et M (ou bipolaires diffuses), les cellules bipolaires de cônes S et les cellules bipolaires de bâtonnets.
Cellules bipolaires de cônes L ou M, naines
Les cellules bipolaires naines font synapse avec des cônes de même type, soit des cônes L, soit des cônes M.
Maillage serré Les cellules bipolaires naines se trouvent sur toute la rétine, en dehors de la fovéola. Elles forment un maillage serré. En rétine centrale, leurs champs dendritiques sont restreints à un seul cône de type L ou M [Gouras P, 1992], [Wassle H et al., 1994]. Plus en périphérie, une cellule bipolaire naine fait synapse avec seulement deux voire trois cônes de même type.
Un cône : une bipolaire naine ON et une OFF On distingue deux types de cellules bipolaires naines, selon que leurs synapses avec les cônes L ou M se font par invaginations ou par contacts superficiels (figure III-1-22).
10

Œil et Physiologie de la Vision – III-1
°par invaginations. Une cellule bipolaire naine peut faire synapse par environ 20 invaginations dendritiques par pédicule de cône [Calkins DJ, 2000] ; sa terminaison axonique se fait à la sous couche-b de la couche plexiforme interne : c’est une cellule bipolaire de cônes naine ON.
°par contacts superficiels. Une autre cellule bipolaire naine peut faire synapse avec un cône L ou M par contacts superficiels sur un pédicule de cône L ou M ; sa terminaison axonique se fait à la sous couche-a de la couche plexiforme interne : c’est une cellule bipolaire de cônes naine OFF.
Au centre de la rétine, chaque cône L ou M est donc en relation avec deux cellules bipolaires naines, l’une ON et l’autre OFF [Boycott B, Wassle H, 1999] (figure III-1-22).
Cette association fonctionnelle est à l’origine de l’analyse du contraste et de l’antagonisme spectral.
Plus en périphérie, chaque couple de cellules bipolaires naines ON et OFF est en relation avec deux ou trois cônes d’une même catégorie L ou M [Kolb H, Marshak D, 2003].
Les cellules bipolaires naines forment la 1ère partie de la voie P ou Parvo.
Cellules bipolaires de cônes L et M, diffuses
Les cellules bipolaires diffuses font synapse avec des cônes L et M sans en faire la distinction (figure III-1-23).
Maillage lâche Les cellules bipolaires diffuses se trouvent, elles aussi, sur toute la rétine, en dehors de la fovéola. Elles forment un maillage lâche. En rétine centrale, leurs connexions dendritiques se font avec au moins cinq cônes voisins et en périphérie, avec dix à 15 cônes voisins [Calkins DJ, 2000].
Un cône : plusieurs bipolaires diffuses ON et OFF On distingue également deux types de cellules bipolaires diffuses différentes, selon que leurs synapses avec les cônes L et M se font par invaginations ou par contacts superficiels (figure III-1-16).
°par invaginations. Une cellule bipolaire diffuse fait synapse par environ 2 à 5 invaginations dendritiques par pédicule de cône [Calkins DJ, 2000], en des sites [Calkins DJ et al., 1996] et des modes [Hopkins JM, Boycott BB, 1996] d’invagination différents de ceux des cellules bipolaires naines. Sa terminaison axonique se fait à la sous couche-b de la couche plexiforme interne : c’est une cellule bipolaire de cônes diffuse ON.
°par contacts superficiels. Une autre cellule bipolaire diffuse peut faire synapse par contacts superficiels sur plusieurs pédicules de cône L et M ; sa terminaison axonique se fait à la sous couche-a de la couche plexiforme interne : c’est une cellule bipolaire de cônes diffuse OFF.
Chaque cône L et M est donc en relation avec plusieurs couples de cellules bipolaires diffuses ON et OFF (jusqu’à 7 couples, [Boycott BB, Wassle H, 1991]). De même, un couple de cellules bipolaires diffuses ON et OFF fait synapse avec plusieurs cônes L et M sus-jacents [Calkins DJ et al., 1996].
Les cellules bipolaires de cônes diffuses forment la 1ère partie de la voie M ou Magno ; elles sont également en relation avec la 2ième partie de la voie K ou Konio (figure III-1-24).
11

Œil et Physiologie de la Vision – III-1
Deux maillages pour les cônes L et/ou M
Ainsi, les cônes L et/ou M font-ils synapse avec deux maillages : l’un serré formé par les cellules bipolaires naines ON et OFF et l’autre, plus lâche, par les cellules bipolaires diffuses ON et OFF. Chaque cône soit L, soit M est donc en relation avec au moins 4 cellules bipolaires : une naine ON et une naine OFF, une diffuse ON et une diffuse OFF…
Chacun de ces maillages est le point de départ de deux voies : respectivement la voie P (renvoi ci-dessous au § voie P) ou Parvo pour les cellules bipolaires naines et la voie M ou Magno pour les cellules bipolaires diffuses. Chacune traite de façon spécifique et différenciée les paramètres de la stimulation codés initialement par les cônes L et M.
Densité des cellules bipolaires de cônes L et/ou M Dans les 20° centraux, il y a un parallélisme étroit entre la densité des cônes et celle des cellules bipolaires de cônes L et/ou M, avec un rapport pratiquement constant de 2,5 cônes pour 4 bipolaires [Martin PR, Grunert U, 1992].
Bien qu’il y ait plus de cellules bipolaires de cônes, que de cônes, la densité relative des cellules bipolaires de cônes est pratiquement la même que celle des cônes avec l’excentricité.
Synapses des cellules bipolaires de cônes L et/ou M avec les cellules sous-jacentes Les axones des cellules bipolaires de cônes naines et diffuses font des synapses de type chimique :
°à la sous couche-a, pour les cellules bipolaires naines ou diffuses OFF, avec les dendrites des cellules ganglionnaires naines ou parasols OFF (figure III-1-22, figure III-1-23),
°ou à la sous couche-b pour les cellules bipolaires naines ou diffuses ON, avec les dendrites des cellules ganglionnaires naines ou parasols ON (figure III-1-22, figure III-1-23).
Cellules bipolaires de cônes S
Les dendrites des cellules bipolaires de cônes S font synapse uniquement par invaginations avec deux à trois cônes S espacés selon leur répartition spatiale rétinienne. Il y aurait environ 35 invaginations par cône [Calkins DJ, 2000] ; leurs champs dendritiques sont donc larges.
Leurs terminaisons axoniques s'effectuent principalement au niveau de la sous couche-b de la couche plexiforme interne avec une couverture large. 60% des terminaisons font des synapses chimiques avec des cellules ganglionnaires bistratifiées (figure III-1-24) et les 40% restant font des synapses avec des cellules amacrines [Calkins DJ, 2000].
Les bipolaires de cônes S fonctionnent sur un mode ON. Elles forment la 1ère partie de la voie K ou konio.
De rares cellules bipolaires de cônes S de type OFF ont été décrites chez le primate ; elles sont petites, issues de synapses par contacts avec des cônes S et font synapse avec des cellules ganglionnaires naines OFF [Klug K et al., 2003].
12

Œil et Physiologie de la Vision – III-1
Cellules bipolaires de bâtonnets
Caractéristiques Les cellules bipolaires de bâtonnets apparaissent dans la rétine à partir de trois à cinq degrés d'excentricité.
Leurs dendrites sont entourées des terminaisons axoniques des cellules horizontales HI ; elles font synapse par invaginations dans les sphérules de bâtonnets et uniquement avec des bâtonnets (figure III-1-22).
Les dendrites d'une même cellule bipolaire de bâtonnets se distribuent à 30-35 bâtonnets près du centre et à 40-45 bâtonnets en périphérie suivant une augmentation progressive de leurs champs d’arborisation du centre jusqu'à la lointaine périphérie.
Leurs terminaisons axoniques se font au niveau de la sous couche-b de la couche plexiforme interne, caractéristiques des cellules bipolaires fonctionnant sur un mode ON. Leurs terminaisons axoniques ont un champ de taille croissante du centre vers la périphérie.
Synapses à la couche plexiforme interne Les cellules bipolaires de bâtonnets ne font jamais de synapses directes avec des cellules ganglionnaires, les cellules amacrines AII et A17 étant leurs intermédiaires.
Leur arrangement est spécifique. Les cellules bipolaires de bâtonnets font des synapses chimiques à la sous couche-b avec des cellules amacrines de type A17 mais principalement de type AII [Wassle H et al., 1991]. Le neurotransmetteur libéré est le glutamate.
A la sous couche-b Les axones des cellules bipolaires de bâtonnets font une synapse chimique avec des prolongements des cellules amacrines AII dites expansions (figure III-1-22).
Par d’autres expansions, mais toujours à la sous couche-b, ces mêmes cellules amacrines AII font synapse par des jonctions gap, avec les cellules bipolaires de cônes naines ON, juste avant que ces cellules bipolaires de cônes ne fassent leur synapse chimique avec des cellules ganglionnaires naines ON (figure III-1-22).
Cette configuration permet à la voie des bâtonnets de rejoindre la 2ième partie de la voie P ON.
A la sous couche-a D’autres expansions des cellules amacrines AII font des synapses chimiques glycinergiques à la sous couche-a, à côté des terminaisons axonales de cellules bipolaires de cônes naines OFF, pour faire synapse avec une cellule ganglionnaire naine OFF.
Cette disposition permet à la voie des bâtonnets de rejoindre avec la 2ième partie de la voie P OFF.
Figure III-1-25. Superposition des cellules bipolaires et ganglionnaires décrites, loin de la réalité beaucoup plus complexe !
13

Œil et Physiologie de la Vision – III-1
Cellules interplexiformes Les cellules interplexiformes sont des cellules d’association qui font le lien entre les couches plexiformes externe et interne en formant une voie rétinienne centrifuge. Un seul type a été observé chez l'homme, à partir de la moyenne périphérie.
Leur corps cellulaire est de dimension moyenne (11 µm). Leurs expansions font synapse avec des cellules horizontales à la couche plexiforme externe sur plus de 300 µm et, de façon diffuse, avec des cellules bipolaires et des cellules amacrines à la couche plexiforme interne.
Leurs modes de connexions suggèrent qu'elles sont des analogues aux cellules interplexiformes GABAergiques observées chez le chat (GABA : Gamma-AminoButiric Acid). Cependant, chez l'homme, elles ne semblent pas être dopaminergiques comme c'est le cas dans d'autres espèces [Witkovsky P et al., 2008].
Couche plexiforme interne : les cellules amacrines
Les cellules amacrines sont des cellules d’association situées au niveau de la couche plexiforme interne, entre les cellules bipolaires et les cellules ganglionnaires [Kolb H et al., 1992], [Morgan IG, 1992], [Kolb H, 1997],[Bloomfield SA, Dacheux RF, 2001], [Kolb H, 2006].
Des cellules spécifiques
Les cellules amacrines sont spécifiques. Elles possèdent un corps cellulaire de taille variable duquel partent des expansions plus ou moins nombreuses et longues qui ont à la fois les caractéristiques de dendrites et d’axones.
En effet, les membranes des expansions possèdent des sites aussi bien présynaptiques que postsynaptiques, sur une courte distance et sans systématisation. Les cellules amacrines peuvent donc émettre et/ou recevoir des influx tout le long de leurs expansions.
Tailles du champ de leurs expansions Elles sont variables. On trouve des °cellules amacrines à petits champs ; leurs expansions sont nombreuses couvrant une surface de diamètre d’environ 100 µm. La densité de leurs corps cellulaires est importante sur toute la rétine. Ces cellules sont typiquement glycinergiques (dont les cellules AII).
On rencontre aussi des °cellules amacrines à champs larges ; leurs expansions sont souvent limitées à une seule strate et occupent une surface de diamètre supérieure à 500 µm ; elles se composent de différentes variétés de cellules mais essentiellement de cellules GABAergiques.
Terminaisons de leurs expansions Les cellules amacrines ont des expansions qui peuvent se terminer à la même strate, sur deux, trois ou toutes les strates de la couche plexiforme interne ; elles sont dites uni- bi-, tristratifiées ou diffuses.
Connexions synaptiques Les cellules amacrines peuvent faire des synapses chimiques ou par jonctions gap avec des cellules bipolaires, des cellules ganglionnaires, d'autres cellules amacrines et des cellules interplexiformes.
14

Œil et Physiologie de la Vision – III-1
Parmi vingt variétés, une essentielle : la cellule amacrine AII
Plus d’une vingtaine de cellules amacrines différentes ont été identifiées, sensiblement également réparties entre cellules glycinergiques [Wassle H et al., 2009] et cellules GABAergiques, tous ces neurotransmetteurs étant inhibiteurs. Les mieux connues sont respectivement les cellules amacrines A17 et les cellules amacrines AII qui assurent la connexion entre la voie des bâtonnets et les cellules ganglionnaires naines.
La plupart des cellules amacrines GABAergiques peuvent également libérer d’autres substances actives ou neuromodulateurs –substance P, somatostatine, cholécystokinine, sérotonine, dopamine, acétylcholine, adénosine… [Witkovsky P, Dearry A, 1992], [Kolb H, 2006] qui jouent un rôle dans différents couplages cellulaires.
Cellules amacrines AII glycinergiques Les cellules amacrines AII glycinergiques sont un exemple de cellules bistratifiées. Elles assurent le lien entre les cellules bipolaires de bâtonnets et les cellules ganglionnaires naines, aucune cellule bipolaire de bâtonnets ne faisant directement synapse avec des cellules ganglionnaires [Kolb H, 2006].
Elles font une synapse chimique avec les cellules bipolaires de bâtonnets à la sous couche-b de la couche plexiforme interne. Elles transmettent les influx reçus, à deux sous couches : d’une part, à la sous couche-a, par une synapse chimique, à côté de cellules bipolaires de cônes naines OFF puis vers des cellules ganglionnaires naines OFF et, d’autre part, à la sous couche-b, par une jonction gap sur le pédicule de cellules bipolaires de cônes naines ON, vers des cellules ganglionnaires naines ON comme il a déjà été dit (figure III-1-22).
Cellules amacrines A17 GABAergiques Elles ont été décrites chez le chat, le lapin et aussi chez le rat [Menger N, Wassle H, 2000]. Elles sont GABAergiques avec des champs d’expansion larges. Elles font synapse avec plus de 1000 cellules bipolaires de bâtonnets à la sous couche-b de la couche plexiforme interne par des jonctions gap ; elles permettent l’intégration d’informations dispersées sur une large surface [Nelson R, Kolb H, 1985].
Neuromodulateurs Ils sont libérés par la plupart des cellules amacrines GABAergiques, par des synapses non conventionnelles [Kolb H, 2006]. Ces neuromodulateurs diffusent à distance dans la rétine pour modifier l’organisation de la circuiterie au cours des changements l’éclairement ou à différents moments du cycle nycthéméral. A la lumière, la dopamine est libérée par des cellules spécialisées ; elle permet le découplage des jonctions gap en particulier entre cellules horizontales et probablement entre cellules amacrines AII [Bloomfield SA, Dacheux RF, 2001].
Troisième étage : les cellules ganglionnaires
Les cellules ganglionnaires forment le troisième étage fonctionnel de la neurorétine à partir de la couche plexiforme interne, en contiguïté avec les cellules bipolaires. Elles recouvrent plus d’une quinzaine [Field GD, Chichilnisky EJ, 2007], voire une vingtaine de variétés [Curcio CA, Allen KA, 1990].
Nombre et densité
Le nombre total moyen de cellules ganglionnaires chez l'homme est de l'ordre de 1 million, avec des variations allant de 700 000 à 1,5 million. La moitié des cellules ganglionnaires se trouve dans la zone rétinienne centrée sur la fovéola et vue sous un angle de 32° ; cette zone ne correspond qu'à 7% de la surface rétinienne totale.
15

Œil et Physiologie de la Vision – III-1
En dehors de la fovéola où il n'y a pas de cellules ganglionnaires, la densité des cellules ganglionnaires est maximale sur un anneau périfovéolaire compris entre 1,5° et 7°, entre 32 000 et 38 000 cellules/mm2 selon les sujets étudiés.
Elle diminue rapidement à une valeur d’environ 10 000 cellules/mm2 à une excentricité de 10° pour n'être plus que de quelques 300 cellules/mm2 à la périphérie.
Ces caractéristiques de densité expliquent en grande partie pourquoi l’onde N95 du P-ERG reflète le fonctionnement des corps des cellules ganglionnaires situées essentiellement dans 15 degrés centraux
Particularités
Asymétrie de répartition Le nombre de cellules ganglionnaires ainsi que leur densité sont plus grands en rétine nasale qu'en rétine temporale. De même, le nombre et la densité des cellules ganglionnaires sont plus importants en rétine supérieure qu'en rétine inférieure.
Différences interindividuelles Le nombre des cônes et celui des cellules ganglionnaires ne sont pas corrélés d'un sujet à l'autre. Le rapport : nombre de cellules ganglionnaires sur nombre de cônes, peut varier de 3 à 7, pour les 10 degrés centraux.
Le rapport n’est plus que de 1, à 7,5° d’excentricité et tombe à une valeur inférieure à 0,5, lorsque l’excentricité est supérieure à 19° et ce, en étroite relation avec la résolution spatiale de ces zones [Sjostrand J et al., 1999].
Plusieurs groupes de cellules ganglionnaires ont été individualisés. Les techniques de transport rétrograde permettent d’en retenir essentiellement trois : les cellules ganglionnaires naines (environ 80% du total) , les parasols (environ 10% du total) et les bistratifiées (quelques pourcents du total) [Calkins DJ, 2000].
Stratification à la sous couche-a ou à la sous couche-b Comme pour les cellules bipolaires, les cellules ganglionnaires ont leurs dendrites stratifiées pour les unes, °à la sous couche-a de la couche plexiforme interne, où elles reçoivent les influx provenant des cellules bipolaires de cônes OFF et pour d’autres, °à la sous couche-b où elles recueillent les influx provenant des cellules bipolaires de cônes ON (figure III-1-22, figure III-1-23, figure III-1-24).
Cellules ganglionnaires ON – cellules ganglionnaires OFF Les cellules ganglionnaires qui font synapse à la sous couche-a de la couche plexiforme interne, avec des cellules bipolaires sont dites cellules ganglionnaires OFF ; de même, les cellules ganglionnaires qui font synapse à la sous couche-b avec des cellules bipolaires sont dites cellules ganglionnaires ON.
Cette terminologie, reliée à la zone anatomique synaptique, prend tout son sens lorsque les aspects fonctionnels sont abordés ; un rappel succinct est présenté ci-dessous.
Réponse des cellules ganglionnaires à une stimulation : potentiels d’action En l’absence de stimulation, les cellules ganglionnaires émettent spontanément des potentiels d’action dont les fréquences temporelles sont aléatoires, correspondant à leur rythme de base.
16

Œil et Physiologie de la Vision – III-1
Lorsque les photorécepteurs qui leur sont directement sus-jacents sont stimulés et que les cellules ganglionnaires répondent par une augmentation de la fréquence temporelle de leurs potentiels d’action par rapport à leur rythme de base (ou réponse « ON »), les cellules ganglionnaires sont dites ON (figure III-1-26).
De même, lorsque les photorécepteurs qui leur sont directement sus-jacents sont stimulés et que les cellules ganglionnaires répondent par une diminution de la fréquence temporelle de leurs potentiels d’action par rapport à leur rythme de base (ou réponse « OFF »), les cellules ganglionnaires sont dites OFF (figure III-1-26).
Cellules ganglionnaires naines
Elles correspondent schématiquement à deux variétés : les cellules ganglionnaires naines P1 et P2. Les ganglionnaires naines P1 se trouvent essentiellement entre la fovéola et la moyenne périphérie ; les ganglionnaires naines P2 sont plus nombreuses à la moyenne périphérie que sur le reste de la rétine (figure III-1-22).
Champs dendritiques Les champs dendritiques des cellules ganglionnaires naines P1 augmentent de 5 µm à 20 µm entre la région fovéale et la moyenne périphérie ; ceux des cellules ganglionnaires naines P2 sont plus larges, variant de 10 µm à 100 µm entre la région fovéale et la périphérie.
Leurs dendrites sont stratifiées soit à la sous couche-a, pour les cellules ganglionnaires naines P1 ou P2 dites OFF, soit à la sous couche-b, pour les cellules ganglionnaires naines P1 ou P2 dites ON.
Corps cellulaires Le corps cellulaire des cellules ganglionnaires naines P1 est plus petit que celui des cellules ganglionnaires naines P2.
Relation avec photorécepteurs sus-jacents Au centre de la rétine, en zone extrafovéale ou périfovéale, chaque cellule ganglionnaire naine P1 ON ou OFF est reliée avec une cellule bipolaire de cônes naine ON ou OFF et donc un cône L ou M.
En périphérie, il y a convergence : une cellule ganglionnaire naine P2 ON ou OFF reçoit les influx de quelques cellules bipolaires de cônes naines ON ou OFF ; elle est donc en relation avec de petits groupes de cônes L ou M.
Les cellules ganglionnaires naines sont aussi en relation avec des bâtonnets par l’intermédiaire des cellules amacrines AII et des cellules bipolaires de bâtonnets.
Projections axoniques sur les corps géniculés latéraux Les axones des cellules ganglionnaires naines sont de petits calibres ; ils forment environ 80% des fibres du nerf optique. Ils se projettent sur les couches parvocellulaires des corps géniculés latéraux (figure IV-38). On trouve davantage d’axones de cellules ganglionnaires naines P1 que P2 se projetant sur les zones de représentation fovéale.
Relation avec le cortex visuel Après relais synaptique et par les radiations optiques, leurs projections se font au niveau de la couche 4-C-béta du cortex visuel primaire.
17

Œil et Physiologie de la Vision – III-1
Cellules ganglionnaires parasols
Elles sont présentes sur toute la rétine, depuis le centre jusqu'à la périphérie (figure III-1-23).
Champs dendritiques, corps cellulaires, axones Elles se distinguent des cellules ganglionnaires naines par la taille de leurs champs dendritiques qui sont larges allant de 20 µm à 330 µm, par leur corps cellulaire qui est beaucoup plus grand que celui des cellules ganglionnaires naines et par leur axone qui est de gros calibre (1,5 à 2 µm).
Relation avec photorécepteurs sus-jacents Les cellules ganglionnaires parasols font synapse avec plusieurs cellules bipolaires de cônes diffuses °à la sous couche-a pour les cellules ganglionnaires parasols OFF et °à la sous couche-b pour les cellules ganglionnaires parasols ON.
Les cellules ganglionnaires parasols reçoivent donc par l’intermédiaire de plusieurs cellules bipolaires de cônes diffuses, des messages issus uniquement et conjointement des cônes L et M et jamais de ceux provenant des cônes S.
Projections axoniques sur les corps géniculés latéraux Leurs axones forment environ 10% des fibres des nerfs optiques. Ils se projettent sur les couches magnocellulaires des corps géniculés latéraux.
Relation avec le cortex visuel Après relais synaptique et par les radiations optiques, leurs projections se font sur la couche 4-C-alpha du cortex visuel primaire.
Cellules ganglionnaires bistratifiées
Elles sont peu nombreuses et bistratifiées, c'est-à-dire qu’elles possèdent des dendrites localisées dans les deux sous couches de la couche plexiforme interne (figure III-1-24).
Champs dendritiques, corps cellulaires, axones Leurs champs dendritiques sont larges, stratifiés essentiellement à la sous couche-b de la couche plexiforme interne et, dans une moindre mesure, à la sous couche-a [Dacey DM, Lee BB, 1994].
Relation avec cellules sus-jacents A la sous couche-b de la couche plexiforme interne, les cellules ganglionnaires bistratifiées font synapse avec les axones de deux à trois cellules bipolaires de cônes S, (qui fonctionnent sur un mode ON) chacune d’elles étant en relation avec quelques cônes S [Calkins DJ, 2000], [Herr S et al., 2003].
A la sous couche-a, les cellules ganglionnaires bistratifiées font synapse avec les axones de plusieurs cellules bipolaires de cônes diffuses (qui fonctionnent sur un mode OFF) chacune d’elles faisant synapse avec une vingtaine de cônes L et M [Calkins DJ, 2000].
Les cellules ganglionnaires bistratifiées font donc synapse d’une part avec les cônes S –par les cellules bipolaires de cônes S ON- et d’autre part conjointement avec des cônes L et M par les cellules bipolaires de cônes diffuses OFF. Elles sont dites cellules ganglionnaires bistratifiées ON-OFF.
18

Œil et Physiologie de la Vision – III-1
Projections axoniques sur les corps géniculés latéraux Les axones de ces cellules ganglionnaires bistratifiées sont de très petits calibres ; ils se projettent essentiellement sur deux intercouches dites koniocellulaires [Hendry SH, Yoshioka T, 1994], [Roy S et al., 2009], probablement entre les couches 2-3 et 3-4 dénommées intercouches K3 et K4 [Hendry SH, Reid RC, 2000] (figure IV-38).
Relation avec le cortex visuel Après relais aux corps géniculés latéraux, leurs radiations optiques se projettent vraisemblablement directement sur les couches 2 et 3 du cortex visuel (figure IV-38).
Autres cellules ganglionnaires à destination extragéniculée
Parmi les 10% environ des cellules ganglionnaires restantes, une quinzaine de variétés ont été identifiées. Leurs champs dendritiques sont souvent étendus, leurs corps cellulaires de tailles variables, leurs axones de différents calibres. Elles participent par exemple à la formation des voies optiques secondaires (figure IV-29).
Cellules ganglionnaires luminosensibles Certaines de ces cellules ganglionnaires sont des cellules luminosensibles. Ces cellules ganglionnaires n’utilisent ni la voie des cônes, ni celle des bâtonnets et ne permettent pas la vision [Hattar S et al., 2002]. Elles sont sensibles à la lumière et en liaison avec la régulation circadienne. Elles se projettent sur le noyau suprachiasmatique de l’hypothalamus par la voie rétinohypothalamique.
La mélanopsine Ces cellules ganglionnaires luminosensibles expriment spécifiquement un pigment la mélanopsine [Hannibal J, Fahrenkrug J, 2002] [Newman LA et al., 2003].
Ce photopigment présente une probabilité maximale d’absorption des photons entre 420 et 440 nm (courtes longueurs d’onde ou lumière bleue) avec une cascade de transduction similaire à celle des photorécepteurs [Newman LA et al., 2003],[Berson DM, 2007], [Hankins MW et al., 2008].
Un dysfonctionnement du gène codant pour la mélanopsine entraîne des perturbations du rythme circadien et du réflexe pupillaire [Brown RL, Robinson PR, 2004], [Guler AD et al., 2007].
Des expériences ont montré que des mammifères ne possédant ni cônes et ni bâtonnets mais des cellules ganglionnaires à mélanopsine intactes, gardent une sensibilité à la lumière avec conservation de leur rythme circadien et réflexes pupillaires, probablement grâce à leurs cellules ganglionnaires à mélanopsine [Foster RG, Hankins MW, 2002], [Guler AD et al., 2007]. De même, réciproquement, des animaux privés de leurs cellules ganglionnaires à mélanopsine, gardent-ils la capacité de réagir à des structures, donc de voir par la voie classique cônes-bâtonnets [Guler AD et al., 2008].
Rétine, lumière et neuromodulations Sous l’influence de la lumière, la neurorétine est le siège de sécrétion de nombreux neuromodulateurs ; la dopamine a été évoquée ; la mélatonine a une sécrétion locale bien qu’elle soit plus largement sécrétée par l’épiphyse [Macchi MM, Bruce JN, 2004] ; elle participe à la régulation du rythme circadien d’éveil et d’endormissement.
Trois voies rétinocorticales : organisation et destinée
Les cellules des trois étages rétiniens s’organisent en trois voies rétinocorticales : la voie P ou Parvo, M ou Magno et K ou Konio. Ces trois voies véhiculent les informations issues des cônes lorsque les niveaux lumineux sont photopiques et aboutissent à différentes couches des aires visuelles primaires.
19

Œil et Physiologie de la Vision – III-1
La 2ième partie de la voie P recueille les influx issus des bâtonnets lorsque les niveaux lumineux sont faibles, par une voie collatérale intrarétinienne formée par les cellules bipolaires de bâtonnets. Les informations issues des bâtonnets sont alors transmises jusqu’aux aires visuelles primaires.
Voie P
Stimulations de niveaux lumineux photopiques La voie P reçoit les informations d’une part des cônes L et d’autre part des cônes M, sans mélange de connexions. Elle commence aux cellules bipolaires naines ON et OFF (1ère partie de la voie P), puis se poursuit par les cellules ganglionnaires naines ON et OFF (2ième partie de la voie P), en respectant la séparation de la voie ON et OFF. Elle compose 80% des fibres des nerfs optiques (figure III-1-22).
Elle fait relais aux couches parvocellulaires des corps géniculés latéraux pour se terminer à la couche 4-C-béta des aires visuelles primaires.
Elle véhicule les informations issues des cônes soit L, soit M lorsque les niveaux lumineux des stimulations sont photopiques. Elle permet le codage de l’antagonisme rouge-vert -premier canal à l’origine de la vision des couleurs-, du contraste chromatique, du fort contraste ainsi que la discrimination des hautes fréquences spatiales (détails fins).
Stimulations de niveaux lumineux scotopiques Les cônes ne sont pas fonctionnels. La voie P est rejointe à la couche plexiforme interne (en sa 2ième partie) par l’information issue de la voie collatérale intrarétinienne des bâtonnets (formée par les cellules bipolaires de bâtonnets) par l’intermédiaire des cellules amacrines AII. Elle transmet donc les informations issues des bâtonnets, vers les centres visuels.
Voie M
Cette voie n’est fonctionnelle que pour des stimulations de niveaux lumineux photopiques. Elle reçoit les informations des cônes L et M, sans en faire la distinction.
Elle commence aux cellules bipolaires de cônes diffuses d’une part ON et d’autre part OFF (1ère partie de la voie M) qui connectent conjointement des cônes L et M puis elle se poursuit par les cellules ganglionnaires parasols ON et OFF (2ième partie de la voie M). Elle compose 10% environ des fibres du nerf optique (figure III-1-23).
Elle fait relais aux couches magnocellulaires des corps géniculés latéraux et se termine aux couches 4-C-alpha des aires visuelles primaires.
Elle traite de paramètres différents issus des cônes L et M comme le faible contraste, les fréquences temporelles élevées, donc le mouvement.
Jusqu’ici, il n’a pas été possible de mettre en évidence une relation anatomique entre la voie des bâtonnets intrarétinienne et la voie M, bien que certaines expériences psychophysiques effectuées chez l’homme penchent pour le contraire [Lennie P, Fairchild MD, 1994].
Voie K
C’est une voie confirmée il y a une décennie environ [Hendry SH, Yoshioka T, 1994], [Dacey DM, Lee BB, 1994], [Martin PR et al., 1997]. Elle est numériquement très faible, moins d’un % des fibres du nerf optique. Elle reçoit les informations issues des cônes S.
20

Œil et Physiologie de la Vision – III-1
Elle commence aux cellules bipolaires de cônes S (bipolaires ON) (1ère partie de la voie K), puis se poursuit par les cellules ganglionnaires bistratifiées (2ième partie de la voie K). Les cellules bistratifiées font synapse, d’une part sur un mode ON, avec les cellules bipolaires de cônes S et, d’autre part sur un mode OFF, avec les cellules bipolaires de cônes L et M, diffuses OFF (figure III-1-24).
La voie K fait relais sur les intercouches des corps géniculés latéraux pour se terminer sur les blobs des couches 2 et 3 des aires visuelles primaires…
Elle est à l’origine de l’antagonisme bleu-jaune, deuxième canal permettant la vision des couleurs.
Séparation des trois voies jusqu’au cortex
Après relais synaptiques aux corps géniculés latéraux, les voies P, M et K restent séparées jusqu’au cortex [Callaway EM, 2005] en conservant leurs deux modes de réponse ON et OFF.
Tests des voies P et K
Le fonctionnement des voies P et K peut être apprécié cliniquement
°par des tests de la vision des couleurs en particulier le test 15 HUE désaturé de Lanthony et le 15 HUE standard [Lanthony P, 1978], [Lanthony P, 1986], [Lanthony P, 1987], [Lanthony P, 1994], [Lanthony P, 1995], [Leid J, 2008].
°mais également, plus spécifiquement celui de la voie P, grâce à des paramètres précis servant à évoquer les PEV damier.
Deux systèmes rétiniens: photopique et scotopique
Il est possible de présenter l’architecture fonctionnelle de la neurorétine en regroupant l’ensemble des cellules qui la compose en systèmes fonctionnant, d’une part sous la dépendance des cônes ou système photopique et, d’autre part, sous celle des bâtonnets ou système scotopique.
En effet ces deux systèmes visuels -photopique et scotopique- sont intriqués dans la neurorétine [Rigaudière F., Le Gargasson J-F., 2007], [Behar-Cohen F et al., 2009].
Le premier a pour origine les cônes, photorécepteurs qui codent les stimulations de niveaux lumineux élevés ou photopiques et, le second, les bâtonnets, photorécepteurs qui codent les stimulations de niveaux lumineux faibles ou scotopiques.
Cette partition en deux systèmes correspond aux deux aspects fonctionnels de la neurorétine photosensible et au mode d’exploration mis en œuvre au cours de l’exploration visuelle par électrophysiologie qui peut en tester séparément le fonctionnement.
En effet, dans les pathologies rétiniennes, il y a fréquemment une atteinte dissociée ou prépondérante de l’un ou l’autre des systèmes.
21

Œil et Physiologie de la Vision – III-1
Système photopique
Le système photopique est constitué par l’ensemble
°des cônes L, M et S, °de leurs cellules bipolaires de cônes : naines et diffuses, ON et OFF et de cônes S et °de leurs cellules ganglionnaires, naines et parasols, ON et OFF et bistratifiées ON/OFF avec lesquelles ils font successivement synapse (figure III-1-25). Il n’est fonctionnel que lorsque les niveaux lumineux sont photopiques.
Système scotopique
Le système scotopique est initié par les bâtonnets au premier étage rétinien ; au deuxième étage rétinien, il se compose, d’une voie propre aux bâtonnets : les cellules bipolaires de bâtonnets ; elles sont en quelque sorte une voie collatérale, parallèle au système photopique jusqu’à la couche plexiforme interne.
A ce niveau, grâce aux cellules amacrines AII, la voie des bâtonnets rejoint la 2ième partie de la voie P avec ses deux modes ON et OFF au 3ième étage rétinien, comme il a déjà été dit (figure III-1-22).
Niveaux lumineux scotopiques Lorsque les niveaux lumineux sont scotopiques, les cônes ne sont pas fonctionnels, les différentes voies qui en sont issues -voies P, M et K- sont libres de toute transmission.
Les influx issus des bâtonnets sont véhiculés par la 2ième partie de la voie P après médiation des cellules amacrines AII [Vardi N, Smith RG, 1996] (figure III-1-22). Cette configuration anatomo-fonctionnelle permet une économie de circuiterie rétinienne lorsque les niveaux lumineux sont faibles.
Niveaux lumineux mésopiques Lorsque les niveaux lumineux augmentent (niveau lumineux mésopique), la transmission des signaux issus des bâtonnets est plus complexe. L’information chemine, d’une part, le long des cellules bipolaires de bâtonnets et la 2ième partie de la voie P et, d’autre part, se répartit au niveau de la couche plexiforme externe, vers les cellules bipolaires de cônes par les jonctions gap entre cônes et bâtonnets [Sharpe LT, Stockman A, 1999].
Quelles sont alors les modalités de transmission de cette information issue spécifiquement des bâtonnets, emprunte-t-elle les cellules bipolaires de cônes ? La question se pose [Sharpe LT, Stockman A, 1999], [Bloomfield SA, Dacheux RF, 2001]. Il pourrait s’agir de la médiation des cellules bipolaires OFF de cônes, hypothèse encore à étayer [Li W et al., 2010].
Explorations différentielles du système photopique et scotopique
De nombreux tests électrophysiologiques sont à notre disposition, tous complémentaires, permettant de tester séparément, du moins dans une large mesure, le fonctionnement :
°du système photopique et scotopique et ses différents niveaux : réceptoral et post-réceptoral, grâce à l’ERG flash et, dans certaines conditions, le niveau réceptoral du système scotopique grâce à l’EOG.
°des voies ON et OFF du système photopique, grâce à certaines séquences de l’ERG flash photopique (potentiels oscillatoires photopique) et à l’ERG ON-OFF.
°des corps des cellules ganglionnaires en zone centrale grâce au P-ERG et indirectement par les PEV …
22

Œil et Physiologie de la Vision – III-1
Les cellules de Müller
Ce sont les cellules gliales les plus nombreuses de la neurorétine. Elles s’étendent sur toute son épaisseur et soutiennent l’ensemble des cellules.
Elles sont radiales, de grande taille, disposées en palissade. Leur forme varie en fonction de l'excentricité rétinienne; elles sont plus longues et plus minces au centre où leur densité peut atteindre jusqu'à 30 000 cellules/mm2 qu'à la périphérie, où elles sont plus épaisses, de densité moindre : de l'ordre de 6 000 cellules/mm2 [Distler C, Dreher Z, 1996]. Leurs pieds sont à la surface du vitré et les extensions apicales de leur cytoplasme sont nombreuses, digitiformes allant jusque dans l’espace sous rétinien (figure III-1-27).
Elles sont très résistantes à l’ischémie, l’anoxie et l’hypoglycémie [Bringmann A et al., 2006]. Leur rôle est présenté succinctement au chapitre III-2.
Figure III-1-27. Cellule de Müller isolée de rat, à gauche et son schéma fonctionnel, à droite, avec ses deux types de canaux potassium. Kir4.1 (en rouge) sont localisés ; ils permettent les échanges bidirectionnels des K+ au cours l’éclairement. Kir2.1 (en bleu) sont sur toute la membrane cellulaire ; ils permettent l’entrée des K+ en provenance des espaces périneuronaux (D’après à gauche : Fort PE et al. 2008. Glia 56 (6) :597-610, à droite : Kofuji et al. 2002. Glia 39 (3), 292-303).
II - L’épithélium pigmentaire
Description
C’est un épithélium. Il se compose d’une seule couche de cellules de formes hexagonales ou pentagonales, très nombreuses au centre et plus rares à la périphérie. Les cellules sont reliées entre elles par des jonctions serrées. Les échanges entre la choroïde et l’espace sous rétinien ne sont possibles qu’au travers des cellules épithéliales et de façon orientée (figure III-1-19).
L'espace sous rétinien est limité en zone distale par la partie apicale des cellules épithéliales et en zone proximale par les segments externes des photorécepteurs et les extrémités distales des cellules de Müller.
L’espace sous rétinien est un milieu extracellulaire vaste par rapport à n'importe quel autre espace intercellulaire rencontré dans la neurorétine. Son homéostasie est régulée par l'ensemble des cellules le limitant dont celles de l’épithélium pigmentaire.
La partie basale de l'épithélium pigmentaire repose sur la membrane de Bruch ; elle contient de nombreux pigments de mélanine et de lipofuscine.
L'adhérence entre l'épithélium pigmentaire et la choroïde est solide, les décollements de rétine l'épargneront faisant passer le clivage entre l'épithélium pigmentaire et les photorécepteurs.
Sa partie apicale est formée d’expansions qui contiennent de la mélanine ; elle est en rapport étroit avec une partie de l'article externe des photorécepteurs. Ces expansions apicales contribuent au maintien de l'alignement des photorécepteurs et à la phagocytose de leur article externe.
Vascularisation
L'épithélium pigmentaire est avasculaire, nourri par la choriocapillaire dont il en assure le transfert des éléments nutritifs vers les cellules sensorielles et en particulier vers la zone fovéolaire.
Le fonctionnement plus spécifiquement électrophysiologique de l'épithélium pigmentaire est développé au chapitre III-3.
23

Œil et Physiologie de la Vision – III-1
III - Application à la lecture des images rétiniennes en OCT
Jean-François Le Gargasson
L’Optical Coherence Tomography a pris une importance majeure dans l’exploration de la rétine. C’est une technique interférentielle optique non invasive qui permet de voir les différentes couches de la rétine [Costa RA et al., 2006], [Cohen S.Y., Haouchine B., 2006],[Gaudric A., Haouchine B., 2007].
Principe succinct La technique actuelle est basée sur l’utilisation du spectromètre. Il permet la transposition d’une image spatiale en une image fréquentielle, aboutissant à des contrastes jusqu’ici inégalés entre les différentes strates rétiniennes.
L’OCT « spectral domain » est basé sur le fait que la vitesse de propagation de la lumière dans un milieu dépend de la longueur d’onde considérée. C’est ainsi que durant un même lapse de temps, une lumière de grande longueur d’onde parcourt plus de distance qu’une lumière de plus courte longueur d’onde.
Si l’on envoie dans l’œil un ensemble de longueurs d’ondes et que l’on enregistre les interférences produites par chacune d’entre elles, on a alors une image simultanée des interférences générées par les différentes couches rétiniennes, chacune d’elle étant représentée par une longueur d’onde donnée.
Les grandes longueurs d’ondes correspondent aux couches les plus externes, car la distance parcourue par unité de temps est plus importante que celle parcourue par les lumières de courtes longueurs d’ondes qui correspondent alors aux couches les plus internes de la rétine.
Le spectromètre est l’appareil qui mesure ces interférences produites par ces différentes longueurs d’ondes et donc par les différentes couches de la rétine et permet de les afficher.
Conséquence importante Il ne faut cependant jamais oublier que ce type d’imagerie n’est pas une visualisation directe des différentes couches de la rétine, mais le résultat d’une opération physique complexe, basée sur les interférences.
La constitution d’interférences plus ou moins nombreuses et amples dépend de la structure du tissu rencontré. Elle peut donner des résultats optiques apparemment paradoxaux dans certaines circonstances.
Anisotropie En effet, la précision actuelle de l’OCT n’est pas isotrope, c'est-à-dire n’est pas identique dans toutes les directions. En profondeur, la précision est de l’ordre de 10 µm, voire inférieure -donc meilleure-, alors que celle correspondant au plan de la rétine est beaucoup moins bonne, de l’ordre de plusieurs dizaines de micromètres et dépendant largement de la largeur de coupe choisie (figure III-1-28).
L’OCT est donc essentiellement un appareil destiné à imager les différentes strates de la rétine – donc en profondeur- qui se distingueront selon la variation de leur texture (figure III-1-29). Il n’autorise pas, tel que, la visualisation à l’échelle cellulaire, sans adjonction d’une autre méthode telle que l’optique adaptative.
L’optique adaptative est une technique qui permet de compenser de façon active les aberrations oculaires. Cette technique utilise un aéromètre auquel est couplé un miroir déformable capable de compenser les aberrations. Voir http://research.opt.indiana.edu/Labs/AdaptiveOptics/default.html
Deux classes de structures rétiniennes Les structures des couches rétiniennes peuvent être divisées en deux classes, selon qu’elles comportent ou non une architecture essentiellement parallèle au plan rétinien.
24

Œil et Physiologie de la Vision – III-1
Première classe Elle correspond à la limitante interne, les fibres du nerf optique, la plexiforme interne, la plexiforme externe, la limitante externe, l’interface entre les articles externes et internes de photorécepteurs, l’épithélium pigmentaire et la membrane de Bruch.
Cette première classe comporte des éléments réfléchissants de structure fibrillaire dont l’architecture est majoritairement parallèle au plan rétinien c'est-à-dire perpendiculaire au faisceau lumineux exploratoire (figure III-1-30). Ces éléments correspondront à une image claire donnée par un signal interférométrique intense.
On note par ailleurs que les structures de type fibrillaire de diamètre inférieur à la longueur d’onde, constituent un élément particulièrement réfléchissant qui garde une cohérence temporelle plus importante que celle induite par les autres éléments rétiniens. Ce phénomène génère un signal optique de retour plus intense que pour les autres types de structures. C’est ainsi que les couches des fibres du nerf optique et les couches plexiformes sont particulièrement réfléchissantes et sont donc à l’origine d’un signal interférométrique ample. Leurs images OCT sont donc vues très claires (figure III-1-28).
Deuxième classe Elle comprend la couche nucléaire interne, la couche nucléaire externe, les articles des photorécepteurs.
Cette seconde classe correspond soit
° soit à des éléments de beaucoup plus grande taille que la longueur d’onde et dont l’organisation cellulaire n’est pas régulière ; cette irrégularité va favoriser la dispersion du faisceau lumineux et donc la perte de signal ; l’image est sombre ;
° soit à des structures ayant un contraste optique très faible telles que les articles externes et internes des photorécepteurs ; l’image est également sombre.
Catégorie intermédiaire La couche des cellules ganglionnaires représente une catégorie intermédiaire avec un mélange de fibres et de corps cellulaires. Le signal n’est pas uniformément déstructuré et son image sera vue moyennement contrastée avec un aspect de structure inhomogène. La présence d’une couche de vaisseaux internes renforce cette inhomogénéité de l’image.
On note par ailleurs que les structures de type fibrillaire de diamètre inférieur à la longueur d’onde, constituent un élément particulièrement réfléchissant qui garde une cohérence temporelle plus importante que celle induite par les autres éléments rétiniens. Ce phénomène génère un signal optique de retour plus intense que pour les autres types de structures. C’est ainsi que les couches des fibres du nerf optique et les couches plexiformes sont particulièrement réfléchissantes et sont donc à l’origine d’un signal interférométrique ample. Leurs images OCT sont donc vues très claires (figure III-1-28).
Réfléctivité des surfaces et visibilités des structures On comprend donc ainsi comment les différences de propriétés optiques des éléments rencontrés se traduisent par des visibilités différentes entre strates rétiniennes.
La chance de l’examen OCT est que les structures générant des fortes interférences alternent avec celles qui sont plus dispersives. En effet dans les cas où cette alternance n’existe pas ou n’existe plus à cause d’une pathologie, il est impossible de distinguer les structures les unes des autres, comme dans le cas de néovascularisation sous rétinienne.
25

Œil et Physiologie de la Vision – III-1
Cet effet de surface optiquement réfléchissante et peu dispersive sur le plan optique peut primer sur l’absorption optique dans les tissus, pour certaines structures. Ce paradoxe apparent est bien illustré au niveau de l’épithélium pigmentaire.
En effet, alors que l'épithélium pigmentaire est un tissu mélanique absorbant largement la lumière, le signal optique de retour est intense (et se traduit par une image claire). Ce phénomène est expliqué par l’effet miroir de la surface membranaire cellulaire, beaucoup plus important que l’absorption mélanique.
Conséquence de la méthode interférométrique
Disparition anatomique ou désorganisation structurelle : piège ! Cette méthode interférométrique a des conséquences importantes, en particulier lors des changements d’orientation ou des pertes d’organisation des structures. Dans ce cas, il y a une perte du signal qui se traduit par une image sombre.
Une perte de signal n’est donc pas nécessairement équivalente à une disparition des éléments anatomiques, mais peut simplement correspondre à une modification ou désorganisation de la structure…
Individualisation de certaines strates rétiniennes sur l’image OCT Certaines strates rétiniennes sont normalement accolées. Elles ne s’individualisent que si elles sont séparées par un élément pathologique qui permet alors d’en avoir une image OCT.
C’est ainsi que °le décollement du vitré permet d’individualiser la hyaloïde postérieure de la rétine, que °le décollement séreux permet de séparer la neurorétine de l’épithélium pigmentaire ou encore que °le dépôt sous rétinien permet le plus souvent de distinguer l’épithélium pigmentaire de la membrane de Bruch (figure III-1-29 et figure III-1-30).
Conclusion
Il a été insisté sur l’architecture de la neurorétine photosensible avec mise en perspective de son organisation en trois étages, trois voies et deux systèmes pour permettre de mieux comprendre les bases et les résultats des explorations fonctionnelles visuelles cliniques en particulier par électrophysiologie.
Son imagerie par OCT s’appuie étroitement sur son organisation en trois étages.
La mise en parallèle de la structure rétinienne et des images obtenues à l’OCT permet de mieux interpréter ces deux types de résultats, les lésions anatomiques visibles à l’OCT ne s’accompagnant pas toujours d’anomalie fonctionnelle enregistrable par électrophysiologie et réciproquement…
26

Œil et Physiologie de la Vision – III-1
Figures
Figure III-1-1. Mise en évidence des trois étages de la rétine humaine : coupe histologique (J-Ph Onolfo) et schéma de rétine humaine (D’après Dowling JE et Boycott BB. Organization of the primate retina. Electron microscopy. Proc Royal Soc London 1966, 166B 80-111).
Figure III-1-2. Le creux fovéolaire est bien visible sur une image OCT.
27
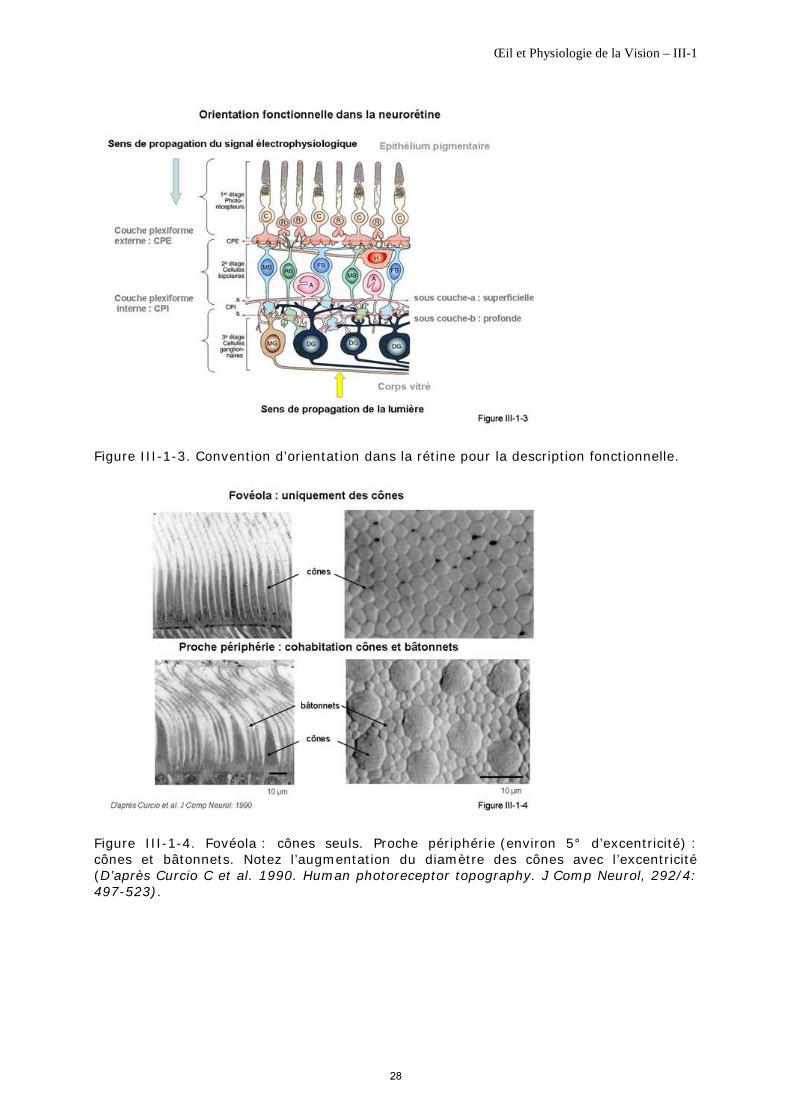
Œil et Physiologie de la Vision – III-1
Figure III-1-3. Convention d’orientation dans la rétine pour la description fonctionnelle.
Figure III-1-4. Fovéola : cônes seuls. Proche périphérie (environ 5° d’excentricité) : cônes et bâtonnets. Notez l’augmentation du diamètre des cônes avec l’excentricité (D’après Curcio C et al. 1990. Human photoreceptor topography. J Comp Neurol, 292/4: 497-523).
28
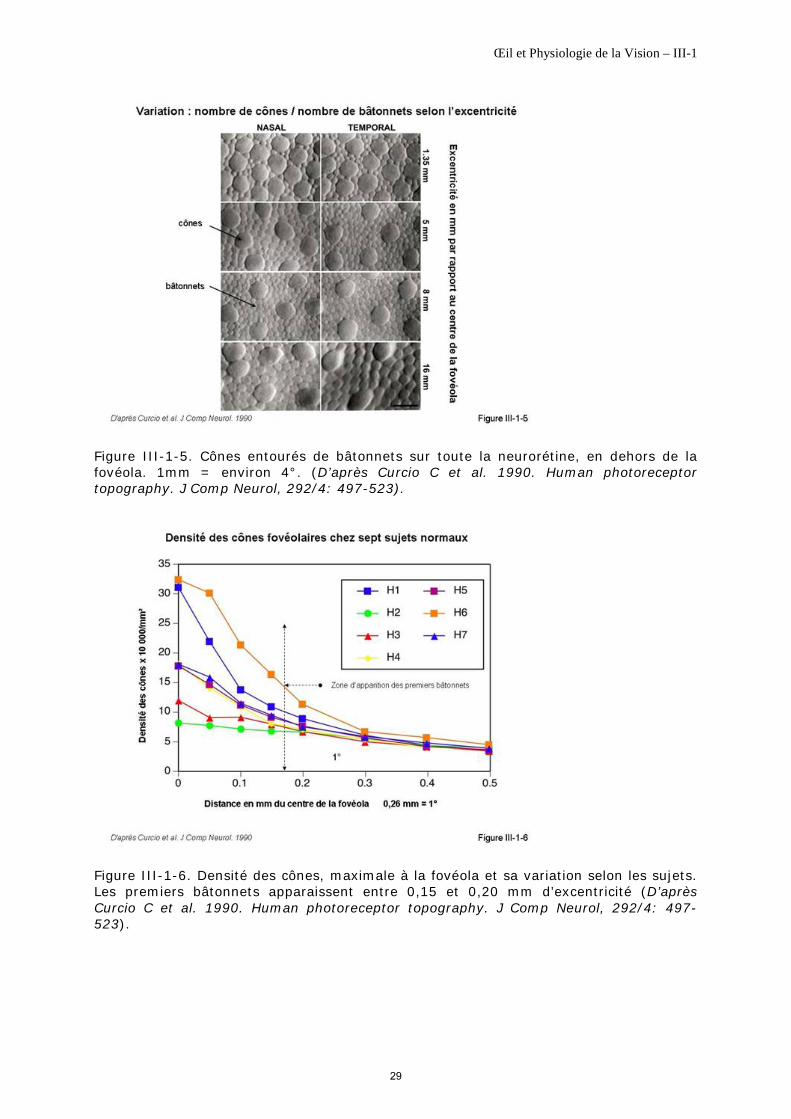
Œil et Physiologie de la Vision – III-1
Figure III-1-5. Cônes entourés de bâtonnets sur toute la neurorétine, en dehors de la fovéola. 1mm = environ 4°. (D’après Curcio C et al. 1990. Human photoreceptor topography. J Comp Neurol, 292/4: 497-523).
Figure III-1-6. Densité des cônes, maximale à la fovéola et sa variation selon les sujets. Les premiers bâtonnets apparaissent entre 0,15 et 0,20 mm d’excentricité (D’après Curcio C et al. 1990. Human photoreceptor topography. J Comp Neurol, 292/4: 497-523).
29
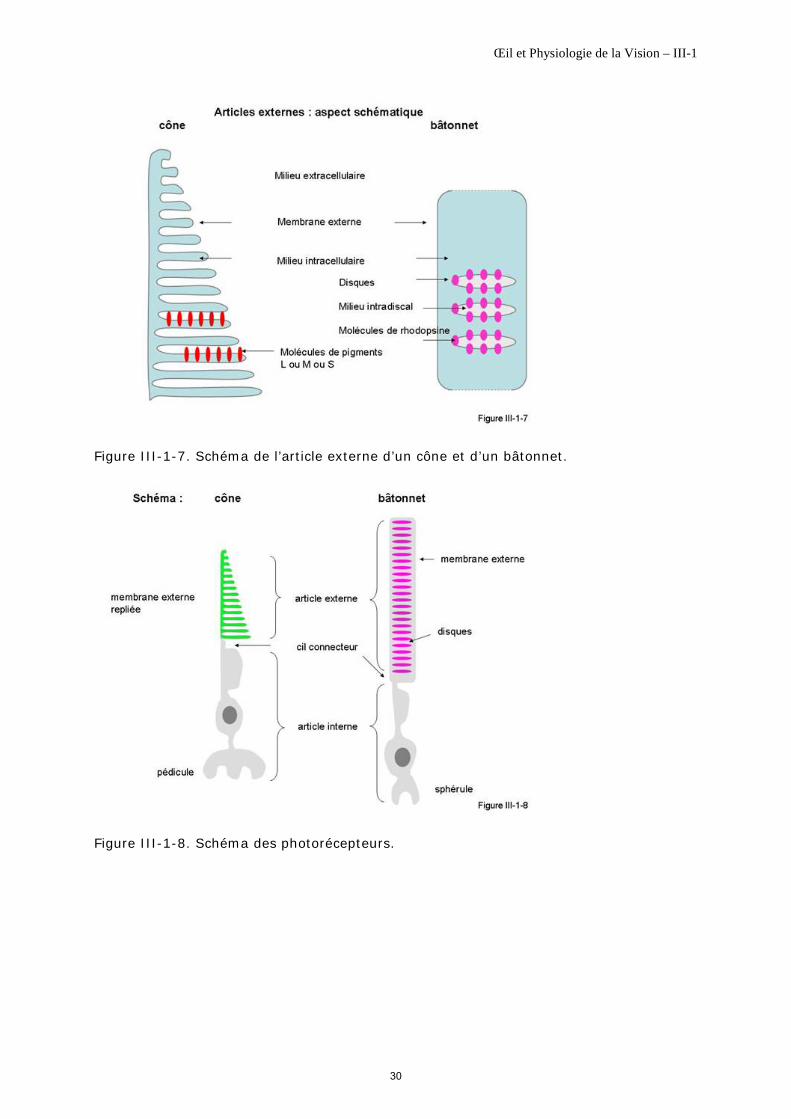
Œil et Physiologie de la Vision – III-1
Figure III-1-7. Schéma de l’article externe d’un cône et d’un bâtonnet.
Figure III-1-8. Schéma des photorécepteurs.
30

Œil et Physiologie de la Vision – III-1
Figure III-1-9. Mise en évidence de cônes S sur une rétine de macaque. Coloration par mélange de procion jaune et brun H3RS (microscopie par fluorescence) (D’après McCrane EP et al. 1983. Non-fluorescent dye staining of primate blue cones. Invest Ophthalmol Vis Sci, 24/11 : 449-1455).
Figure III-1-10. Cônes centraux. La flèche noire –en bas à droite- pointe l’intrusion d’un bâtonnet. (D’après Curcio CA, Hendrickson AE, 1991. Organisation and development of the primate photoreceptor mosaic. Progress in Retinal Research Vol 10: 89-120).
31
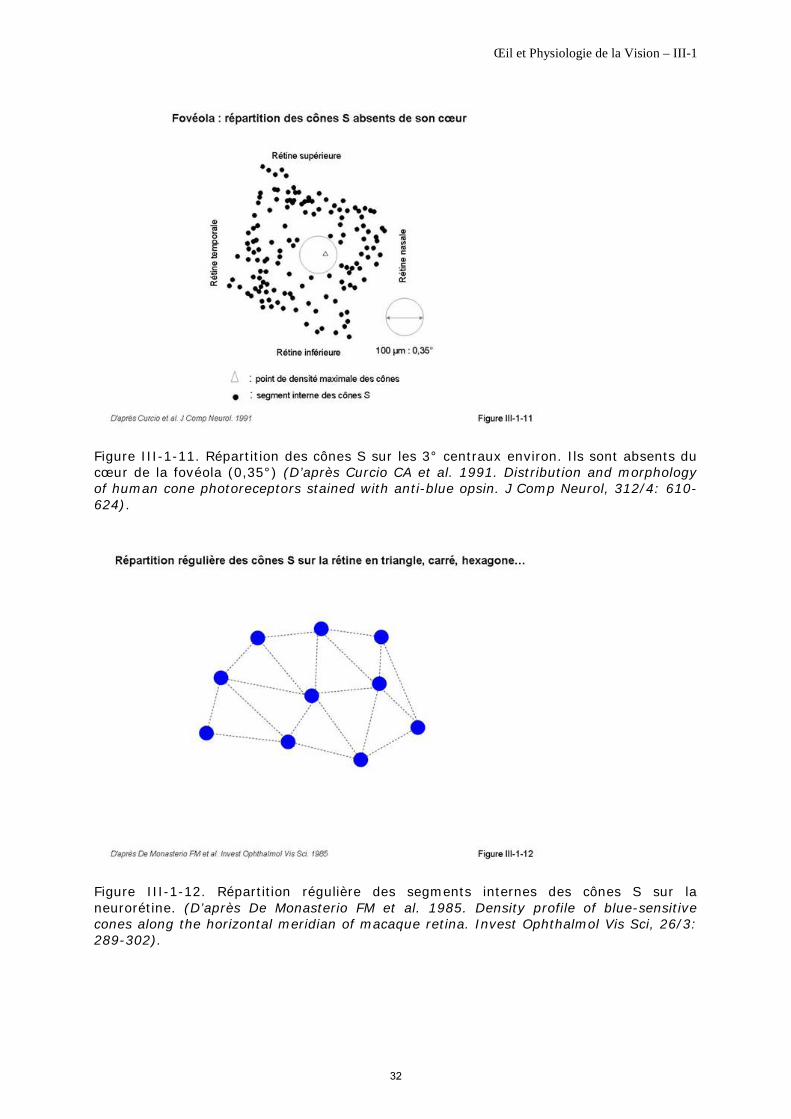
Œil et Physiologie de la Vision – III-1
Figure III-1-11. Répartition des cônes S sur les 3° centraux environ. Ils sont absents du cœur de la fovéola (0,35°) (D’après Curcio CA et al. 1991. Distribution and morphology of human cone photoreceptors stained with anti-blue opsin. J Comp Neurol, 312/4: 610-624).
Figure III-1-12. Répartition régulière des segments internes des cônes S sur la neurorétine. (D’après De Monasterio FM et al. 1985. Density profile of blue-sensitive cones along the horizontal meridian of macaque retina. Invest Ophthalmol Vis Sci, 26/3: 289-302).
32
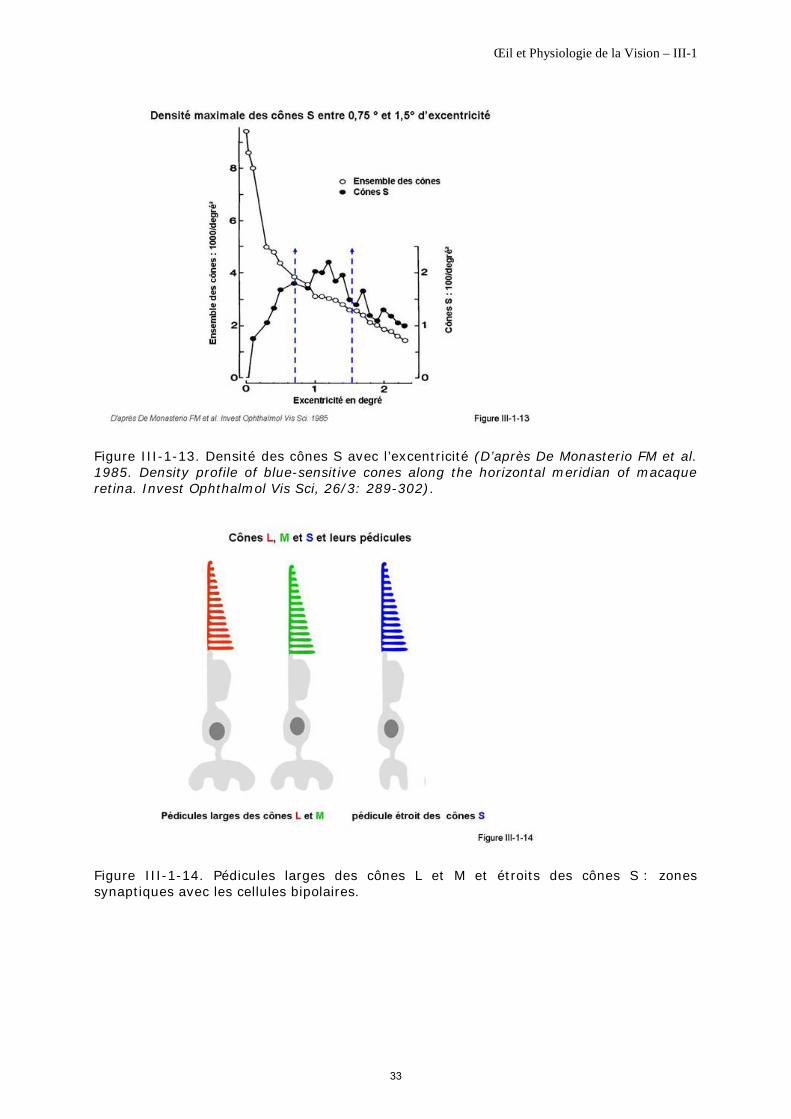
Œil et Physiologie de la Vision – III-1
Figure III-1-13. Densité des cônes S avec l’excentricité (D’après De Monasterio FM et al. 1985. Density profile of blue-sensitive cones along the horizontal meridian of macaque retina. Invest Ophthalmol Vis Sci, 26/3: 289-302).
Figure III-1-14. Pédicules larges des cônes L et M et étroits des cônes S : zones synaptiques avec les cellules bipolaires.
33
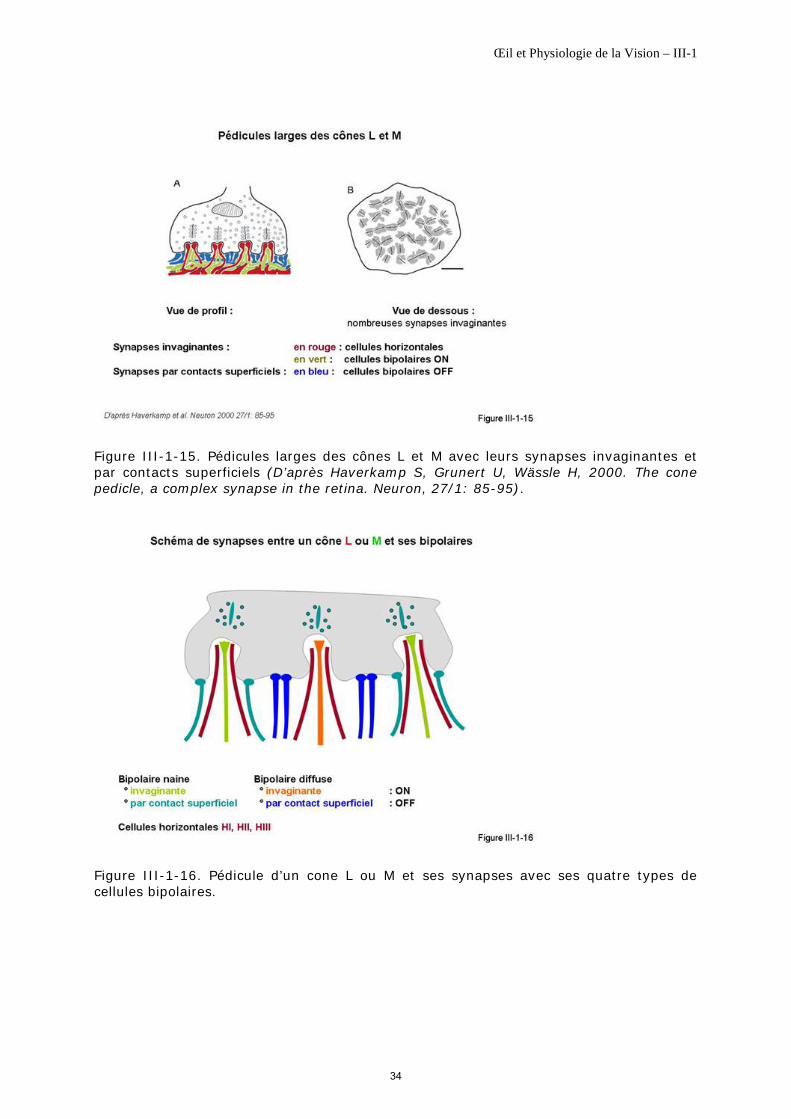
Œil et Physiologie de la Vision – III-1
Figure III-1-15. Pédicules larges des cônes L et M avec leurs synapses invaginantes et par contacts superficiels (D’après Haverkamp S, Grunert U, Wässle H, 2000. The cone pedicle, a complex synapse in the retina. Neuron, 27/1: 85-95).
Figure III-1-16. Pédicule d’un cone L ou M et ses synapses avec ses quatre types de cellules bipolaires.
34

Œil et Physiologie de la Vision – III-1
Figure III-1-17. Corrélation anatomo-fonctionnelle : bipolaires invaginantes ON : terminaison à la sous couche-b de la couche plexiforme interne – bipolaires par contacts superficielles OFF (cônes L – M) terminaison à la sous couche-a de la couche plexiforme interne.
Figure III-1-18. Cellules Horizontales HI : synapses invaginantes entre pédicules de cônes L et M et sphérules de bâtonnets.
35
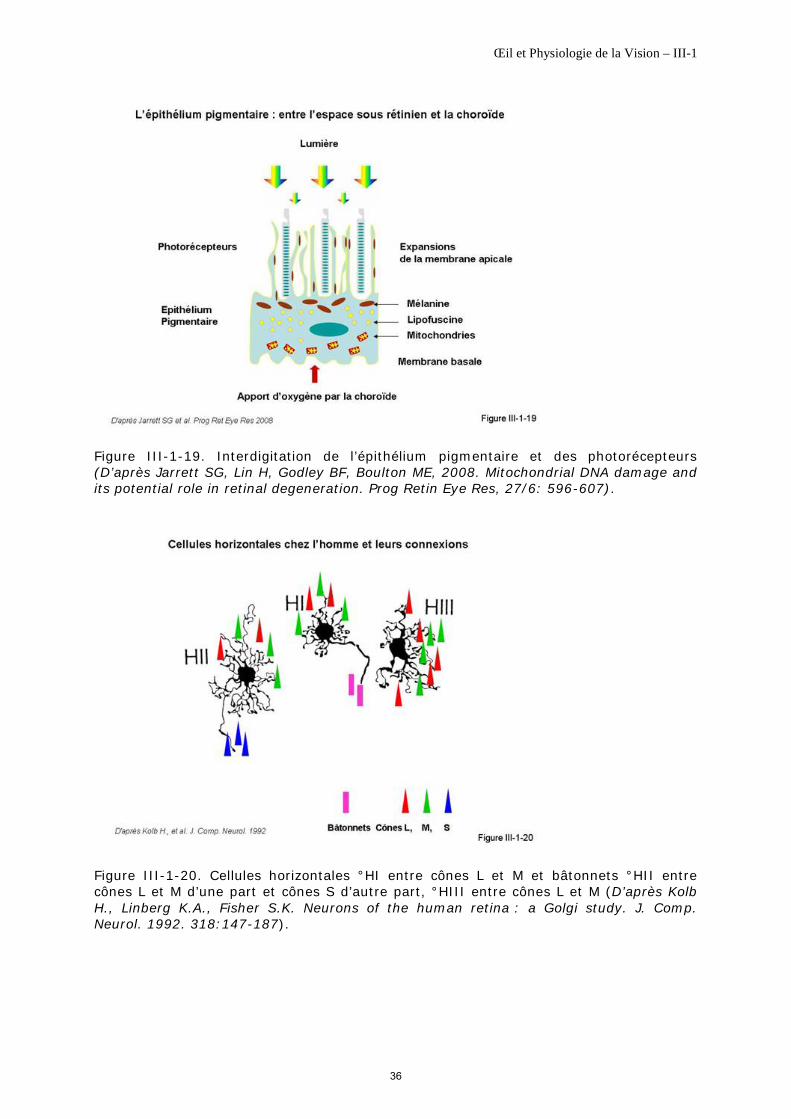
Œil et Physiologie de la Vision – III-1
Figure III-1-19. Interdigitation de l’épithélium pigmentaire et des photorécepteurs (D’après Jarrett SG, Lin H, Godley BF, Boulton ME, 2008. Mitochondrial DNA damage and its potential role in retinal degeneration. Prog Retin Eye Res, 27/6: 596-607).
Figure III-1-20. Cellules horizontales °HI entre cônes L et M et bâtonnets °HII entre cônes L et M d’une part et cônes S d’autre part, °HIII entre cônes L et M (D’après Kolb H., Linberg K.A., Fisher S.K. Neurons of the human retina : a Golgi study. J. Comp. Neurol. 1992. 318:147-187).
36
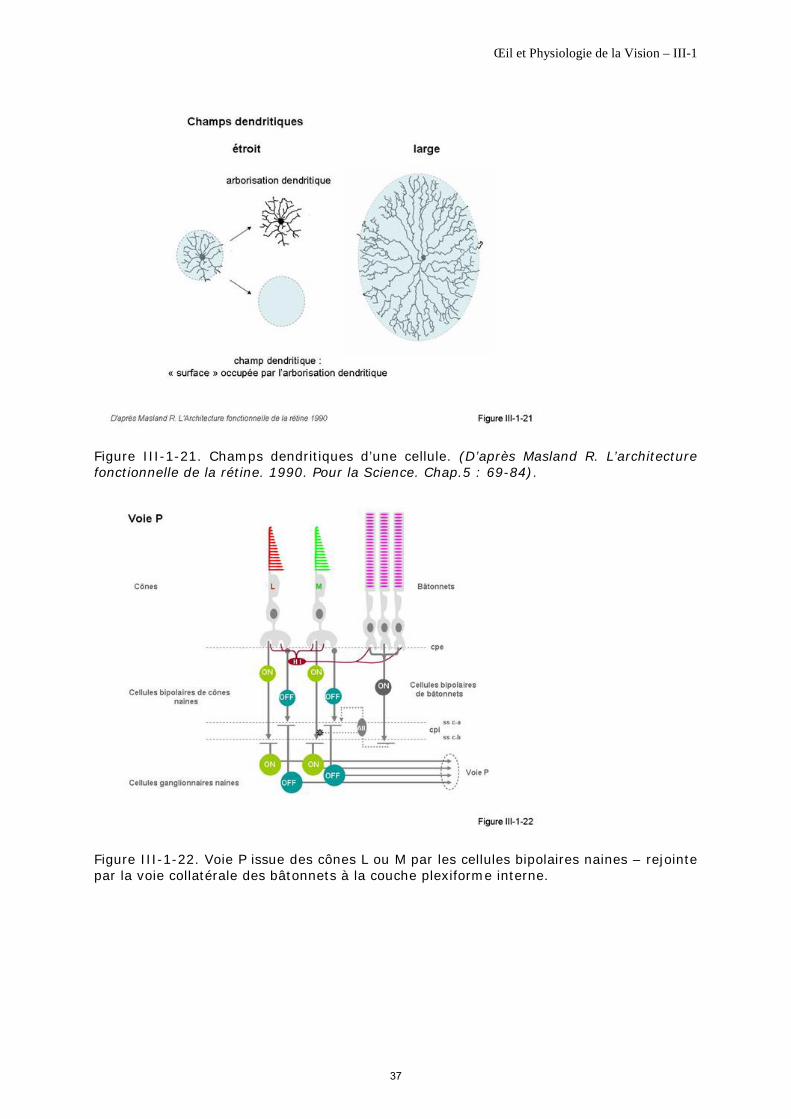
Œil et Physiologie de la Vision – III-1
Figure III-1-21. Champs dendritiques d’une cellule. (D’après Masland R. L’architecture fonctionnelle de la rétine. 1990. Pour la Science. Chap.5 : 69-84).
Figure III-1-22. Voie P issue des cônes L ou M par les cellules bipolaires naines – rejointe par la voie collatérale des bâtonnets à la couche plexiforme interne.
37
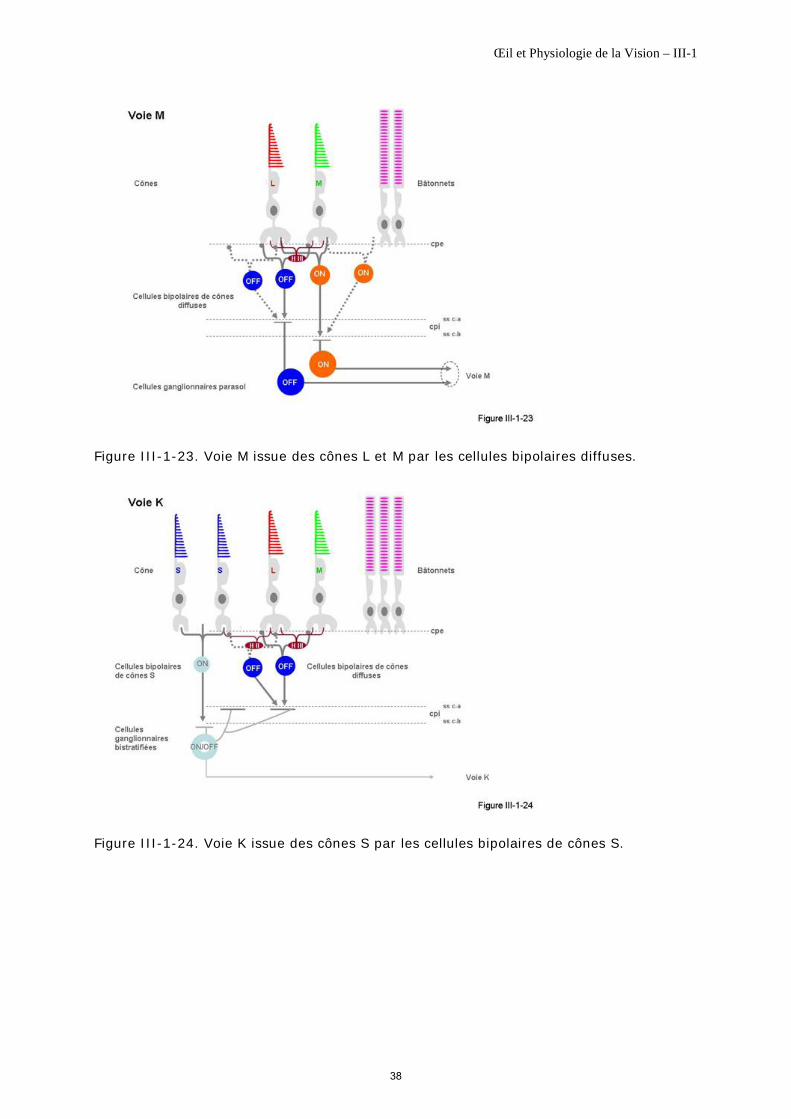
Œil et Physiologie de la Vision – III-1
Figure III-1-23. Voie M issue des cônes L et M par les cellules bipolaires diffuses.
Figure III-1-24. Voie K issue des cônes S par les cellules bipolaires de cônes S.
38
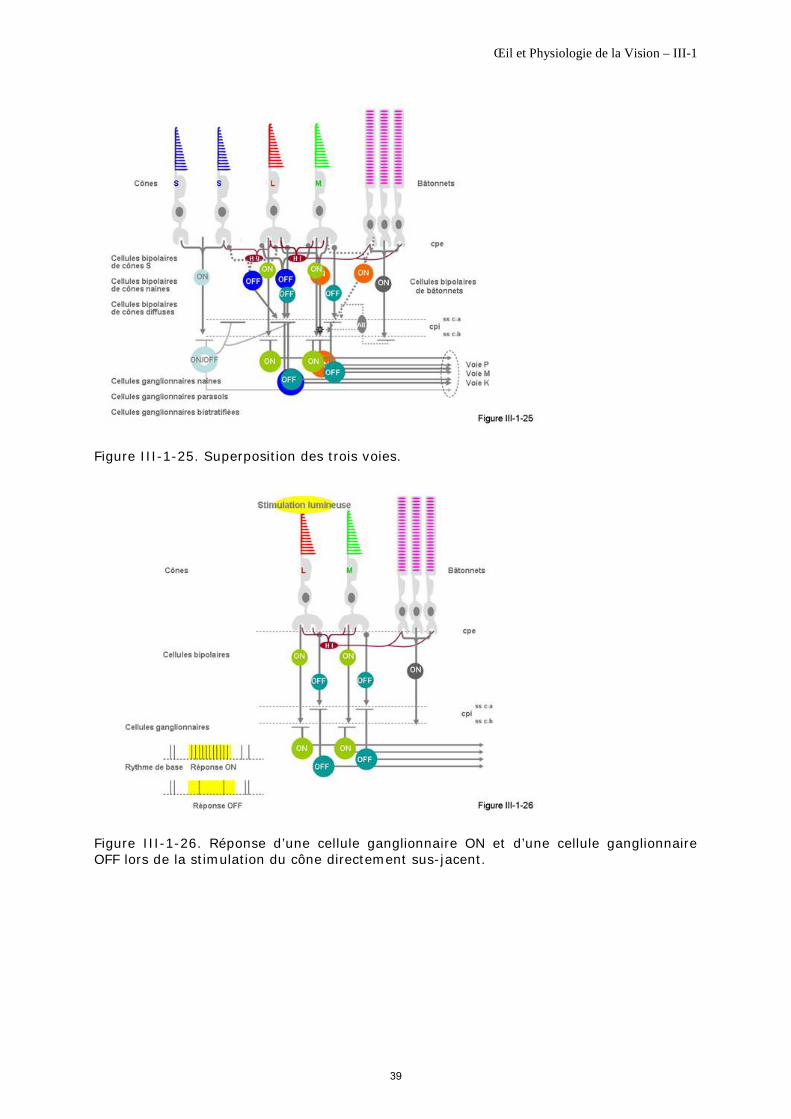
Œil et Physiologie de la Vision – III-1
Figure III-1-25. Superposition des trois voies.
Figure III-1-26. Réponse d’une cellule ganglionnaire ON et d’une cellule ganglionnaire OFF lors de la stimulation du cône directement sus-jacent.
39
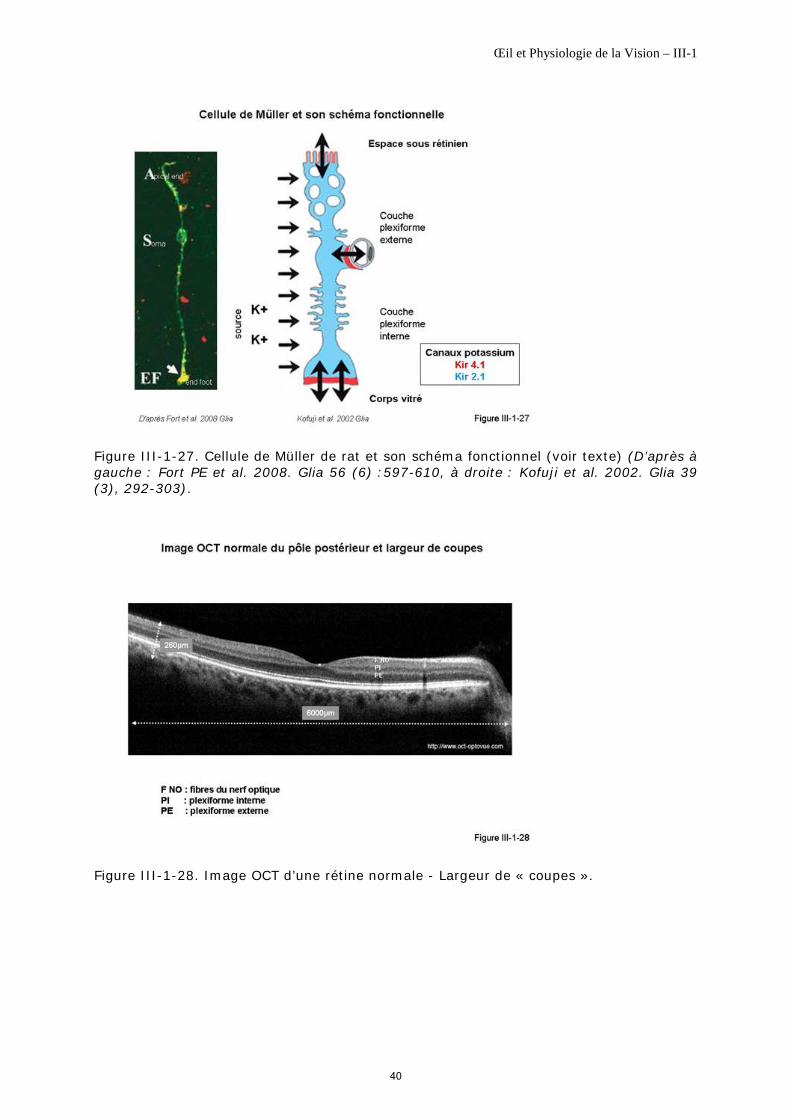
Œil et Physiologie de la Vision – III-1
Figure III-1-27. Cellule de Müller de rat et son schéma fonctionnel (voir texte) (D’après à gauche : Fort PE et al. 2008. Glia 56 (6) :597-610, à droite : Kofuji et al. 2002. Glia 39 (3), 292-303).
Figure III-1-28. Image OCT d’une rétine normale - Largeur de « coupes ».
40
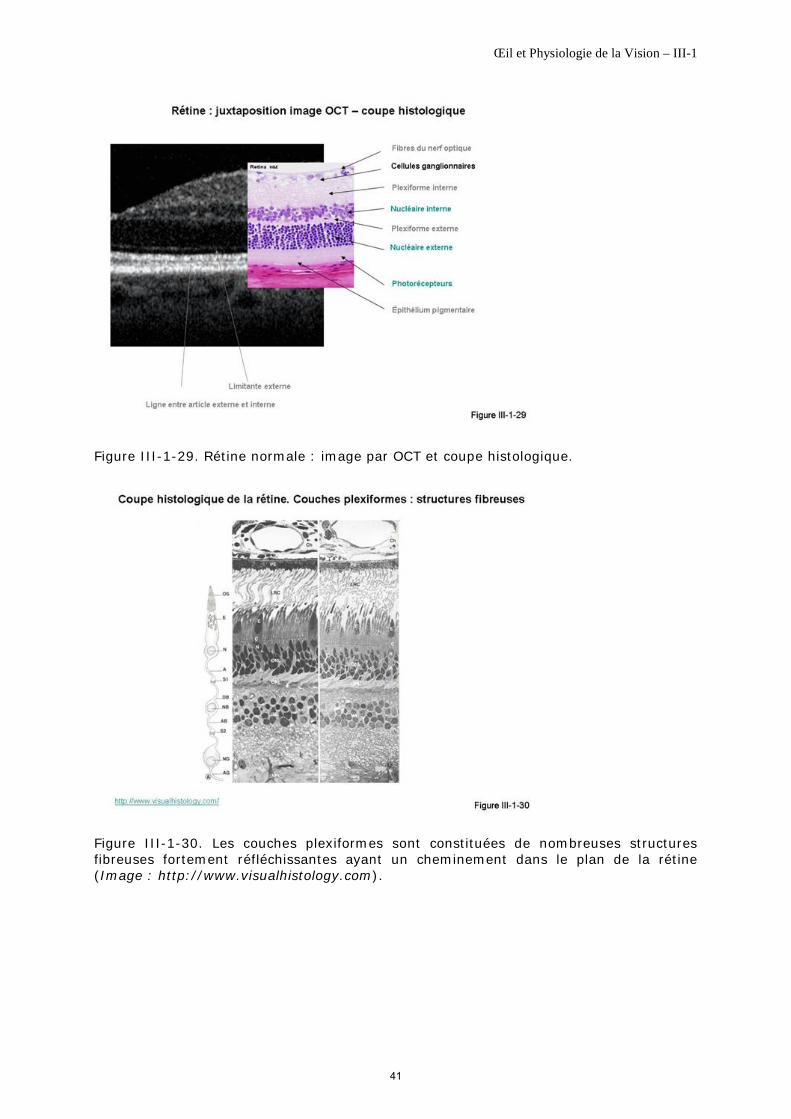
Œil et Physiologie de la Vision – III-1
Figure III-1-29. Rétine normale : image par OCT et coupe histologique.
Figure III-1-30. Les couches plexiformes sont constituées de nombreuses structures fibreuses fortement réfléchissantes ayant un cheminement dans le plan de la rétine (Image : http://www.visualhistology.com).
41
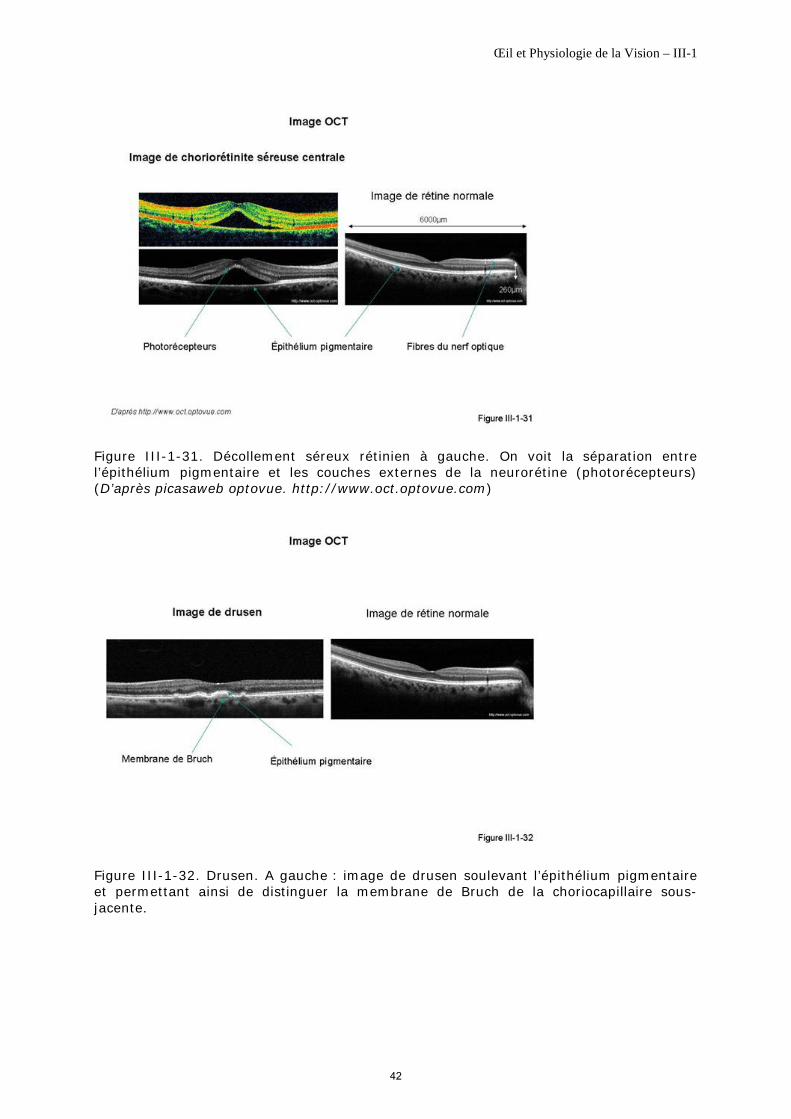
Œil et Physiologie de la Vision – III-1
Figure III-1-31. Décollement séreux rétinien à gauche. On voit la séparation entre l’épithélium pigmentaire et les couches externes de la neurorétine (photorécepteurs) (D’après picasaweb optovue. http://www.oct.optovue.com)
Figure III-1-32. Drusen. A gauche : image de drusen soulevant l’épithélium pigmentaire et permettant ainsi de distinguer la membrane de Bruch de la choriocapillaire sous-jacente.
42

Œil et Physiologie de la Vision – III-1
Bibliographie
Ahnelt, P.K. (1998). The photoreceptor mosaic. Eye, 12, 531-540. [Abstract]
Ahnelt, P.K., Kolb, H., & Pflug, R. (1987). Identification of a subtype of cone photoreceptor, likely to be blue sensitive, in the human retina. J Comp Neurol, 255 (1), 18-34. [Abstract]
Anderson, D.H., Fisher, S.K., & Steinberg, R.H. (1978). Mammalian cones: disc shedding, phagocytosis, and renewal. Invest Ophthalmol Vis Sci, 17 (2), 117-133. [Abstract]
Behar-Cohen F, Kowalczuk L, Keller N, Savoldelli M, Azan F, & Jeanny J-C (2009). Anatomie de la rétine. EMC Elsevier Masson, SAS, Paris, Ophtalmologie, 21-003-C-40, 1-14.
Berson, D.M. (2007). Phototransduction in ganglion-cell photoreceptors. Pflugers Arch, 454 (5), 849-855. [Abstract]
Bloomfield, S.A., & Dacheux, R.F. (2001). Rod vision: pathways and processing in the mammalian retina. Prog Retin Eye Res, 20 (3), 351-384. [Abstract]
Boesze-Battaglia, K., & Goldberg, A.F. (2002). Photoreceptor renewal: a role for peripherin/rds. Int Rev Cytol, 217, 183-225. [Abstract]
Boycott, B., & Wassle, H. (1999). Parallel processing in the mammalian retina: the Proctor Lecture. Invest Ophthalmol Vis Sci, 40 (7), 1313-1327. [Abstract]
Boycott, B.B., & Wassle, H. (1991). Morphological Classification of Bipolar Cells of the Primate Retina. Eur J Neurosci, 3 (11), 1069-1088. [Abstract]
Bringmann, A., Pannicke, T., Grosche, J., Francke, M., Wiedemann, P., Skatchkov, S.N., Osborne, N.N., & Reichenbach, A. (2006). Muller cells in the healthy and diseased retina. Prog Retin Eye Res, 25 (4), 397-424. [Abstract]
Brown, R.L., & Robinson, P.R. (2004). Melanopsin--shedding light on the elusive circadian photopigment. Chronobiol Int, 21 (2), 189-204. [Abstract]
Bumsted, K., & Hendrickson, A. (1999). Distribution and development of short-wavelength cones differ between Macaca monkey and human fovea. J Comp Neurol, 403 (4), 502-516. [Abstract]
Calkins, D.J. (2000). Representation of cone signals in the primate retina. J Opt Soc Am A Opt Image Sci Vis, 17 (3), 597-606. [Abstract]
Calkins, D.J. (2001). Seeing with S cones. Prog Retin Eye Res, 20 (3), 255-287. [Abstract]
Calkins, D.J., Tsukamoto, Y., & Sterling, P. (1996). Foveal cones form basal as well as invaginating junctions with diffuse ON bipolar cells. Vision Res, 36 (21), 3373-3381. [Abstract]
Callaway, E.M. (2005). Structure and function of parallel pathways in the primate early visual system. J Physiol, 566 (Pt 1), 13-19. [Abstract]
43

Œil et Physiologie de la Vision – III-1
Carroll, J., McMahon, C., Neitz, M., & Neitz, J. (2000). Flicker-photometric electroretinogram estimates of L:M cone photoreceptor ratio in men with photopigment spectra derived from genetics. J Opt Soc Am A Opt Image Sci Vis, 17 (3), 499-509. [Abstract]
Chun, M.H., Grunert, U., Martin, P.R., & Wassle, H. (1996). The synaptic complex of cones in the fovea and in the periphery of the macaque monkey retina. Vision Res, 36 (21), 3383-3395. [Abstract]
Cicerone, C.M., & Nerger, J.L. (1989). The relative numbers of long-wavelength-sensitive to middle-wavelength-sensitive cones in the human fovea centralis. Vision Res, 29 (1), 115-128. [Abstract]
Cohen S.Y., & Haouchine B. (2006). L'OCT en pratique: place et apport réel. PROPOS BIOPHARMA (pp. 1-62). Paris.
Costa, R.A., Skaf, M., Melo, L.A., Jr., Calucci, D., Cardillo, J.A., Castro, J.C., Huang, D., & Wojtkowski, M. (2006). Retinal assessment using optical coherence tomography. Prog Retin Eye Res, 25 (3), 325-353. [Abstract]
Curcio, C.A., & Allen, K.A. (1990). Topography of ganglion cells in human retina. J Comp Neurol, 300 (1), 5-25. [Abstract]
Curcio, C.A., Allen, K.A., Sloan, K.R., Lerea, C.L., Hurley, J.B., Klock, I.B., & Milam, A.H. (1991). Distribution and morphology of human cone photoreceptors stained with anti-blue opsin. J Comp Neurol, 312 (4), 610-624. [Abstract]
Curcio, C.A., & Hendrickson, A.E. (1991). Organisation and development of the primate photoreceptor mosaic. Progress in Retinal Research (pp. 89-120- Vol dix). Oxford: Pergamon Press.
Curcio, C.A., Sloan, K.R., Kalina, R.E., & Hendrickson, A.E. (1990). Human photoreceptor topography. J Comp Neurol, 292 (4), 497-523. [Abstract]
Dacey, D.M. (2000). Parallel pathways for spectral coding in primate retina. Annu Rev Neurosci, 23, 743-775. [Abstract]
Dacey, D.M., & Lee, B.B. (1994). The 'blue-on' opponent pathway in primate retina originates from a distinct bistratified ganglion cell type. Nature, 367 (6465), 731-735. [Abstract]
Dacey, D.M., Lee, B.B., Stafford, D.K., Pokorny, J., & Smith, V.C. (1996). Horizontal cells of the primate retina: cone specificity without spectral opponency. Science, 271 (5249), 656-659. [Abstract]
De Monasterio, F.M., McCrane, E.P., Newlander, J.K., & Schein, S.J. (1985). Density profile of blue-sensitive cones along the horizontal meridian of macaque retina. Invest Ophthalmol Vis Sci, 26 (3), 289-302. [Abstract]
Delint, P.J., Berendschot, T.T., & van Norren, D. (1997). Local photoreceptor alignment measured with a scanning laser ophthalmoscope. Vision Res, 37 (2), 243-248. [Abstract]
Distler, C., & Dreher, Z. (1996). Glia cells of the monkey retina--II. Muller cells. Vision Res, 36 (16), 2381-2394. [Abstract]
Eckmiller, M. (1997). Morphogenesis and renewal of cone outer segments. Progress in Retinal Research and Eye Research (pp. 401-441). Oxford: Pergamon.
44

Œil et Physiologie de la Vision – III-1
Field, G.D., & Chichilnisky, E.J. (2007). Information processing in the primate retina: circuitry and coding. Annu Rev Neurosci, 30, 1-30. [Abstract]
Fitzgibbon, J., Appukuttan, B., Gayther, S., Wells, D., Delhanty, J., & Hunt, D.M. (1994). Localisation of the human blue cone pigment gene to chromosome band 7q31.3-32. Hum Genet, 93 (1), 79-80. [Abstract]
Foster, R.G., & Hankins, M.W. (2002). Non-rod, non-cone photoreception in the vertebrates. Prog Retin Eye Res, 21 (6), 507-527. [Abstract]
Gaudric A., & Haouchine B. (2007). OCT de la macula. Collection Atlas en Ophtalmologie (pp. 1-349). Paris : Elsevier.
Gouras, P. (1992). Retinal circuitry and its relevance to diagnostic psychophysics and electrophysiology. Current Opinion in Ophthalmology, 3, 803-812.
Gowdy, P.D., & Cicerone, C.M. (1998). The spatial arrangement of the L and M cones in the central fovea of the living human eye. Vision Res, 38 (17), 2575-2589. [Abstract]
Guler, A.D., Altimus, C.M., Ecker, J.L., & Hattar, S. (2007). Multiple photoreceptors contribute to nonimage-forming visual functions predominantly through melanopsin-containing retinal ganglion cells. Cold Spring Harb Symp Quant Biol, 72, 509-515. [Abstract]
Guler, A.D., Ecker, J.L., Lall, G.S., Haq, S., Altimus, C.M., Liao, H.W., Barnard, A.R., Cahill, H., Badea, T.C., Zhao, H., Hankins, M.W., Berson, D.M., Lucas, R.J., Yau, K.W., & Hattar, S. (2008). Melanopsin cells are the principal conduits for rod-cone input to non-image-forming vision. Nature, 453 (7191), 102-105. [Abstract]
Hankins, M.W., Peirson, S.N., & Foster, R.G. (2008). Melanopsin: an exciting photopigment. Trends Neurosci, 31 (1), 27-36. [Abstract]
Hannibal, J., & Fahrenkrug, J. (2002). Melanopsin: a novel photopigment involved in the photoentrainment of the brain's biological clock? Ann Med, 34 (5), 401-407. [Abstract]
Hattar, S., Liao, H.W., Takao, M., Berson, D.M., & Yau, K.W. (2002). Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity. Science, 295 (5557), 1065-1070. [Abstract]
Haverkamp, S., Grunert, U., & Wassle, H. (2000). The cone pedicle, a complex synapse in the retina. Neuron, 27 (1), 85-95. [Abstract]
Hendry, S.H., & Reid, R.C. (2000). The koniocellular pathway in primate vision. Annu Rev Neurosci, 23, 127-153. [Abstract]
Hendry, S.H., & Yoshioka, T. (1994). A neurochemically distinct third channel in the macaque dorsal lateral geniculate nucleus. Science, 264 (5158), 575-577. [Abstract]
Herr, S., Klug, K., Sterling, P., & Schein, S. (2003). Inner S-cone bipolar cells provide all of the central elements for S cones in macaque retina. J Comp Neurol, 457 (2), 185-201. [Abstract]
Hopkins, J.M., & Boycott, B.B. (1996). The cone synapses of DB1 diffuse, DB6 diffuse and invaginating midget, bipolar cells of a primate retina. J Neurocytol, 25 (7), 381-390. [Abstract]
45

Œil et Physiologie de la Vision – III-1
Kevany, B.M., & Palczewski, K. (2010). Phagocytosis of retinal rod and cone photoreceptors. Physiology (Bethesda), 25 (1), 8-15. [Abstract]
Kimble, T.D., & Williams, R.W. (2000). Structure of the cone photoreceptor mosaic in the retinal periphery of adult humans: analysis as a function of age, sex, and hemifield. Anat Embryol (Berl), 201 (4), 305-316. [Abstract]
Klug, K., Herr, S., Ngo, I.T., Sterling, P., & Schein, S. (2003). Macaque retina contains an S-cone OFF midget pathway. J Neurosci, 23 (30), 9881-9887. [Abstract]
Kolb, H. (1997). Amacrine cells of the mammalian retina: neurocircuitry and functional roles. Eye (Lond), 11 ( Pt 6), 904-923. [Abstract]
Kolb, H. (2006). Functional organization fo the retina. Principles and practice of clinical electrophysiology of vision, Chap 6 (pp. 47-64). Cambridge MIT Press.
Kolb, H., Fernandez, E., Nelson, R., & . The organization of the retina and visual system. Webvision, www.webvision.med.utah.edu.
Kolb, H., Fernandez, E., Schouten, J., Ahnelt, P., Linberg, K.A., & Fisher, S.K. (1994). Are there three types of horizontal cell in the human retina? J Comp Neurol, 343 (3), 370-386. [Abstract]
Kolb, H., Goede, P., Roberts, S., McDermott, R., & Gouras, P. (1997). Uniqueness of the S-cone pedicle in the human retina and consequences for color processing. J Comp Neurol, 386 (3), 443-460. [Abstract]
Kolb, H., Linberg, K.A., & Fisher, S.K. (1992). Neurons of the human retina: a Golgi study. J Comp Neurol, 318 (2), 147-187. [Abstract]
Kolb, H., & Marshak, D. (2003). The midget pathways of the primate retina. Doc Ophthalmol, 106 (1), 67-81. [Abstract]
Kremers, J., Usui, T., Scholl, H.P., & Sharpe, L.T. (1999). Cone signal contributions to electroretinograms [correction of electrograms] in dichromats and trichromats. Invest Ophthalmol Vis Sci, 40 (5), 920-930. [Abstract]
Lanthony, P. (1978). The desaturated panel D-15. Doc Ophthalmol, 46 (1), 185-189. [Abstract] http://www.ncbi.nlm.nih.gov/pubmed/310382
Lanthony, P. (1986). [Evaluation of the desaturated Panel D-15. I. Method of quantification and normal scores]. J Fr Ophtalmol, 9 (12), 843-847. [Abstract]
Lanthony, P. (1987). [Color vision defects in optic neuropathies. Clinical forms]. Ophtalmologie, 1 (1), 27-30. [Abstract]
Lanthony, P. (1994). [Evaluation of desaturated Panel D-15. III. Evaluation of the validity of the desaturated Panel D-15]. J Fr Ophtalmol, 17 (1), 15-21. [Abstract]
Lanthony, P. (1995). [Evaluation of the desaturated Panel D-15. IV. Effect of the repetition of desaturated Panel D-15]. J Fr Ophtalmol, 18 (10), 578-583. [Abstract]
Leid, J. (2008). Spécial Vision des couleurs. Supplément Ophtalmologie n° 32 (pp. 1-86): Propos-Biopharma.
Lennie, P. (2000). Color vision: putting it together. Curr Biol, 10 (16), R589-591. [Abstract]
46

Œil et Physiologie de la Vision – III-1
Lennie, P., & Fairchild, M.D. (1994). Ganglion cell pathways for rod vision. Vision Res, 34 (4), 477-482. [Abstract]
Li, W., Chen, S., & DeVries, S.H. (2010). A fast rod photoreceptor signaling pathway in the mammalian retina. Nat Neurosci, 13 (4), 414-416. [Abstract]
Macchi, M.M., & Bruce, J.N. (2004). Human pineal physiology and functional significance of melatonin. Front Neuroendocrinol, 25 (3-4), 177-195. [Abstract]
Marcos, S., Tornow, R.P., Elsner, A.E., & Navarro, R. (1997). Foveal cone spacing and cone photopigment density difference: objective measurements in the same subjects. Vision Res, 37 (14), 1909-1915. [Abstract]
Martin, P.R., & Grunert, U. (1992). Spatial density and immunoreactivity of bipolar cells in the macaque monkey retina. J Comp Neurol, 323 (2), 269-287. [Abstract]
Martin, P.R., White, A.J., Goodchild, A.K., Wilder, H.D., & Sefton, A.E. (1997). Evidence that blue-on cells are part of the third geniculocortical pathway in primates. Eur J Neurosci, 9 (7), 1536-1541. [Abstract]
McCrane, E.P., de Monasterio, F.M., Schein, S.J., & Caruso, R.C. (1983). Non-fluorescent dye staining of primate blue cones. Invest Ophthalmol Vis Sci, 24 (11), 1449-1455. [Abstract]
Menger, N., & Wassle, H. (2000). Morphological and physiological properties of the A17 amacrine cell of the rat retina. Vis Neurosci, 17 (5), 769-780. [Abstract]
Miller, D.T., Williams, D.R., Morris, G.M., & Liang, J. (1996). Images of cone photoreceptors in the living human eye. Vision Res, 36 (8), 1067-1079. [Abstract]
Morgan, I.G. (1992). What do amacrine cells do? Progress in Retinal Research (pp. 193-214). Oxford: Pergamon.
Nelson, R., & Kolb, H. (1985). A17: a broad-field amacrine cell in the rod system of the cat retina. J Neurophysiol, 54 (3), 592-614. [Abstract]
Nerger, J.L., & Cicerone, C.M. (1992). The ratio of L cones to M cones in the human parafoveal retina. Vision Res, 32 (5), 879-888. [Abstract]
Newman, L.A., Walker, M.T., Brown, R.L., Cronin, T.W., & Robinson, P.R. (2003). Melanopsin forms a functional short-wavelength photopigment. Biochemistry, 42 (44), 12734-12738. [Abstract]
Otake, S., Gowdy, P.D., & Cicerone, C.M. (2000). The spatial arrangement of L and M cones in the peripheral human retina. Vision Res, 40 (6), 677-693. [Abstract]
Rigaudiere, F., Leid, J., Vienot, F., & Le Gargasson, J.F. (2006). [Neurophysiological basis and clinical tests for assessment of X-linked color vision deficiencies in school children]. J Fr Ophtalmol, 29 (1), 87-102. [Abstract]
Rigaudière F., & Le Gargasson J-F. (2007). Explorations électrophysiologiques sensorielles: électrorétinogramme, électro-oculogramme, potentiels évoqués visuels. EMC Elsevier Masson SAS, Paris, Ophtalmologie, (21-046-A-10), 1-23.
Roepman, R., & Wolfrum, U. (2007). Protein networks and complexes in photoreceptor cilia. Subcell Biochem, 43, 209-235. [Abstract]
47

Œil et Physiologie de la Vision – III-1
Roorda, A. (2000). Adaptive optics ophthalmoscopy. J Refract Surg, 16 (5), S602-607. [Abstract]
Roorda, A., Metha, A.B., Lennie, P., & Williams, D.R. (2001). Packing arrangement of the three cone classes in primate retina. Vision Res, 41 (10-11), 1291-1306. [Abstract]
Roorda, A., & Williams, D.R. (1999). The arrangement of the three cone classes in the living human eye. Nature, 397 (6719), 520-522. [Abstract]
Roy, S., Jayakumar, J., Martin, P.R., Dreher, B., Saalmann, Y.B., Hu, D., & Vidyasagar, T.R. (2009). Segregation of short-wavelength-sensitive (S) cone signals in the macaque dorsal lateral geniculate nucleus. Eur J Neurosci, 30 (8), 1517-1526. [Abstract]
Schiller, P.H. (1996). The ON and OFF channels of the mammalian visual system. Progress in Retinal and Eye Research, 15 (1), 173-195.
Sharpe, L.T., & Stockman, A. (1999). Rod pathways: the importance of seeing nothing. Trends Neurosci, 22 (11), 497-504. [Abstract]
Sharpe, L.T., Stockman, A., Jagle, H., Knau, H., & Nathans, J. (1999). L, M and L-M hybrid cone photopigments in man: deriving lambda max from flicker photometric spectral sensitivities. Vision Res, 39 (21), 3513-3525. [Abstract]
Sjostrand, J., Olsson, V., Popovic, Z., & Conradi, N. (1999). Quantitative estimations of foveal and extra-foveal retinal circuitry in humans. Vision Res, 39 (18), 2987-2998. [Abstract]
Steinberg, R.H., Fisher, S.K., & Anderson, D.H. (1980). Disc morphogenesis in vertebrate photoreceptors. J Comp Neurol, 190 (3), 501-508. [Abstract]
Trojan, P., Krauss, N., Choe, H.W., Giessl, A., Pulvermuller, A., & Wolfrum, U. (2008). Centrins in retinal photoreceptor cells: regulators in the connecting cilium. Prog Retin Eye Res, 27 (3), 237-259. [Abstract]
Vardi, N., & Smith, R.G. (1996). The AII amacrine network: coupling can increase correlated activity. Vision Res, 36 (23), 3743-3757. [Abstract]
Wassle, H., Dacey, D.M., Haun, T., Haverkamp, S., Grunert, U., & Boycott, B.B. (2000). The mosaic of horizontal cells in the macaque monkey retina: with a comment on biplexiform ganglion cells. Vis Neurosci, 17 (4), 591-608. [Abstract]
Wassle, H., Grunert, U., Martin, P.R., & Boycott, B.B. (1994). Immunocytochemical characterization and spatial distribution of midget bipolar cells in the macaque monkey retina. Vision Res, 34 (5), 561-579. [Abstract]
Wassle, H., Grunert, U., Rohrenbeck, J., & Boycott, B.B. (1989). Cortical magnification factor and the ganglion cell density of the primate retina. Nature, 341 (6243), 643-646. [Abstract]
Wassle, H., Heinze, L., Ivanova, E., Majumdar, S., Weiss, J., Harvey, R.J., & Haverkamp, S. (2009). Glycinergic transmission in the Mammalian retina. Front Mol Neurosci, 2, 6. [Abstract]
Wassle, H., Yamashita, M., Greferath, U., Grunert, U., & Muller, F. (1991). The rod bipolar cell of the mammalian retina. Vis Neurosci, 7 (1-2), 99-112. [Abstract]
48

Œil et Physiologie de la Vision – III-1
Williams, D.R., MacLeod, D.I., & Hayhoe, M.M. (1981). Foveal tritanopia. Vision Res, 21 (9), 1341-1356. [Abstract]
Willmer, E.N. (1987). Anatomy of the visual system. In The senses (pp. 35-45). Cambridge: University Press.
Witkovsky, P., & Dearry, A. (1992). Functionals role of dopamine in the vertebrate retina. Progress in Retinal Research (pp. 247-292). Oxford: Pergamon.
Witkovsky, P., Gabriel, R., & Krizaj, D. (2008). Anatomical and neurochemical characterization of dopaminergic interplexiform processes in mouse and rat retinas. J Comp Neurol, 510 (2), 158-174. [Abstract]
Young, R.W. (1971). The Renewal Of Rod And Cone Outer Segments In The Rhesus Monkey. J Cell Biol, 49 (2), 303-318. [Abstract].
49


















