I – Homéostasie énergétique - Thèses en ligne de l...
171
UNIVERSITE TOULOUSE III – PAUL SABATIER U.F.R. Sciences de la vie THESE en vue de l’obtention du DOCTORAT DE L’UNIVERSITE TOULOUSE III Ecole Doctorale Biologie, Santé et Biotechnologies de Toulouse Spécialité : Physiopathologie Moléculaire, Cellulaire et intégrée présentée et soutenue publiquement le 05 octobre 2007 par Elise GUILLOD-MAXIMIN SENSIBILITE AU GLUCOSE DU CERVEAU CHEZ LE RAT Implication de l’hypothalamus Détermination des acteurs cellulaires et moléculaires Membres du jury : Pr. Frances B. Pr. Hévor T. Pr. Moyse E. Dr. Lavialle M. Dr. Lorsignol A. Dr. Pénicaud L. Président Rapporteur Rapporteur Examinatrice Examinatrice Directeur de thèse
Transcript of I – Homéostasie énergétique - Thèses en ligne de l...

UNIVERSITE TOULOUSE III – PAUL SABATIER U.F.R. Sciences de la vie
THESE
en vue de l’obtention du
DOCTORAT DE L’UNIVERSITE TOULOUSE III
Ecole Doctorale Biologie, Santé et Biotechnologies de Toulouse
Spécialité : Physiopathologie Moléculaire, Cellulaire et intégrée
présentée et soutenue publiquement le 05 octobre 2007
par
Elise GUILLOD-MAXIMIN
SENSIBILITE AU GLUCOSE DU CERVEAU CHEZ LE RAT
Implication de l’hypothalamus
Détermination des acteurs cellulaires et moléculaires
Membres du jury :
Pr. Frances B. Pr. Hévor T. Pr. Moyse E. Dr. Lavialle M. Dr. Lorsignol A. Dr. Pénicaud L.
Président Rapporteur Rapporteur Examinatrice Examinatrice Directeur de thèse

- 2 -
Remerciements Voilà, mes études s’achèvent enfin. Je dois avouer qu’une étoile devait se trouver au-dessus de ma tête le jour de ma naissance ! En effet, un petit stage de « neuro » qui m’a permis d’appréhender le monde de la recherche au sein du laboratoire du Pr. Hévor à Orléans m’amena en DEA auprès du Dr. Pénicaud, directeur de l’UMR5018. Merci Luc de m’avoir donné cette chance. Je remercie les membres du jury qui ont accepté d’évaluer ce travail, notamment Messieurs les Pr. Hévor et Moyse qui m’ont fait l’honneur d’en être les rapporteurs. Le Dr. Lorsignol s’est chargée de ma formation. De tout cœur Anne, merci pour ton encadrement. J’ai adoré ta rigueur, que ce soit à la paillasse ou au bureau. Aujourd’hui, j’utilise encore cette façon de faire et je ne pense pas en changer. Je regrette seulement de n’avoir pas pu être témoin de ton ventre rond. Grâce à Anne et Luc, j’ai pu effectuer cette thèse, continuer mes travaux et ainsi répondre aux questions qui me turlupinaient (bizarre ce mannitol…). Votre franchise et soutien sont des qualités que j’apprécie beaucoup. Je n’oublie pas le reste de l’équipe toulousaine. Son ambiance me manque et voir l’écriture de mon cher petit Alaing sur un certain colis arrivé à Orsay, m’a serré le cœur. Par peur d’oublier quelqu’un, je vais choisir la facilité en faisant un tir groupé, mais sincère, de merci à toutes les personnes qui m’ont aidée de près ou de loin, niveau manips et niveau moral. Sachez que je pense souvent à vous, à nos discussions autour du café ou le midi dans la salle de réunion. Naturellement, mes pensées se tournent aussi vers ma famille et Yvan, devenu mon mari en cours de route. Il y a longtemps, alors que j’étais collégienne, je disais à mon père que je souhaitais faire de la recherche sur les cétacés (période du film Le Grand Bleu oblige). Il m’avait alors répondu : « Tu rêves ma petite fille ! On habite à Mareau-aux-prés ! ». Eh bien tu vois Papa, j’y suis quand même arrivée ! Enfin pour les dauphins c’est râté, et le poisson ne m’a pas attirée : indigne fille de pêcheur ! Ma mère a été très importante, si ce n’est essentielle, dans le choix de ma voie. En effet, elle m’a toujours poussée à faire un travail qui me plaisait, car comme elle me le répétait souvent, rien est pire que de passer ses journées au boulot à déprimer. Or, la recherche est un métier de passion qui promet de ne pas être de tout repos ! Bien sûr, mon parcours d’étudiante n’a pas été toujours rose. Merci Yvan de m’avoir suivie partout en France, supportée mes crises de nerfs et aidée à redresser la tête dans les périodes de doutes. Et dire que tu m’as attrappée à la sortie du lycée pour ne plus me lâcher ! Finalement, l’histoire s’est achevée sur un succès au concours d’ingénieur d’études de l’INRA, ce qui m’amène aux joyeux Guillod, Aline et Roland, que je remercie chaleureusement pour leur coaching de choc ! Et me voilà parisienne !

- 3 -
Les travaux présentés dans cette thèse ont fait l’objet de
Publications :
Pénicaud L, Leloup C, Lorsignol A, Alquier T, Guillod E Brain glucose sensing mechanism and glucose homeostasis Current Opinion in Clinical Nutrition and Metabolic Care, 2002, 5:539-543 Guillod-Maximin E, Lorsignol A, Alquier T, Pénicaud L Acute intracarotid glucose injection towards the brain induces specific c-fos activation in hypothalamic nuclei: involvement of astrocytes in cerebral glucose-sensing in rats Journal of Neuroendocrinology, 2004, 16:464-471 Pénicaud L, Bénani A, Fioramonti X, Alquier T, Guillod E, Lorsignol A, Leloup C Détecteurs de glucose et régulation de la prise alimentaire Cah. Nutr. Diét., 2007, 42(3) :134-138
Communications affichées :
Guillod-Maximin E, Lorsignol A, Pénicaud L (poster p369) Characterization of hypothalamic cells activated by a transient central hyperglycaemia using c-Fos 3rd Forum of European Neuroscience, Paris, Juillet 2002 Guillod-Maximin E, Lorsignol A, Pénicaud L (poster p389) Mechanism of acute cerebral hyperglycaemia-induced peripheral insulin secretion in the rat : glucokinase implication 4th Forum of European Neuroscience, Lisbonne, Portugal, Juillet 2004 Communication orale : Guillod-Maximin E Sensibilité au glucose du cerveau chez le rat IFR31 (CHU Rangueil), Toulouse, Mai 2004

- 4 -
Liste des abréviations
AGL : acides gras libres
AgRP : agouti-related peptide
α-MSH : alpha-melanocyte-stimulating hormone
AMPK : AMP-activated protein kinase
BDNF : brain-derived neurotrophic factor
BHE : barrière hémato-encéphalique
BSO : buthionine sulfoximine
CART : cocaine- and amphetamine-regulated transcript
CC : cortex cérébral
CCK : cholécystokinine
CNPase : 2’, 3’-cyclic nucleotide 3’-phosphodiesterase
CRH : corticotropin-releasing hormone
DAB : diaminobenzidine
Δ : différence de concentrations par rapport à la valeur basale au temps zéro
DMN : noyau hypothalamique dorsomédian
DMV : noyau dorsal moteur du vague
2-DG : 2-déoxyglucose
GABA : γ-aminobutyric acid
GE : gluco-excité
GI : gluco-inhibé
GFAP : glial fibrillary acidic protein
GFAT : glutamine fructose-6-phosphate aminotransférase
GK : glucokinase
GlcN : glucosamine
GlcNAc : N-acétylglucosamine
GLP-1 : glucagon-like peptide-1
[glucose]e : concentration extracellulaire en glucose
GLUT : transporteur de glucose
GS : glutamine synthase
H + L : heavy and light chains
IgG : immunoglobuline G
IRS : insulin receptor substrate
IVGTT : intravenous glucose-tolerance test

- 5 -
JAK : Janus kinase
Kir : inwardly rectifying K+ channel
Km : constante de Michaelis
LH : hypothalamus latéral
MCH : melanin-concentrating hormone
μU : micro-unité
MSO : L-méthionine sulfoximine
NA : noyau arqué
NaCl : Chlorure de sodium
NeuN : neuronal nuclei
NGS : Normal Goat Serum
NPY : neuropeptide Y
NTS : noyau du tractus solitaire
Ob-R : récepteur à la leptine
PBS : Phosphate-Buffered Saline
PI3K : phosphatidylinositol 3-kinase
PIP2 : phosphatidylinositol (4,5)-biphosphate
PIP3 : phosphatidylinositol (3,4,5)-triphosphate
POMC : pro-opiomélanocortine
PVN : noyau hypothalamique paraventriculaire
PYY : peptide YY
SNA : système nerveux autonome
SNC : système nerveux central
SUR : sulfonylurea receptor
TRH : thyrotropin-releasing hormone
VMN : noyau hypothalamique ventromédian

- 6 -
Liste des tableaux et figures
Tableau 1 : Neurotransmetteurs principaux impliqués dans la régulation de la prise alimentaire p.26 Tableau 2 : Effet d’un prétraitement intrapéritonéal de 18 heures de méthionine sulfoximine sur un test de tolérance au glucose p.105 Figure 1 : Illustration de l’homéostasie énergétique p.13 Figure 2 : Interactions établies entre les organes périphériques et le système nerveux central p.14 Figure 3 : Lieu de synthèse et action des principales hormones jouant sur l’homéostasie énergétique p.15 Figure 4 : Les différents effets de la leptine selon le tissu cible p.16 Figure 5 : Reflet de l’état énergétique d’un individu par sa leptinémie p.17 Figure 6 : Sécrétion de l’insuline en réponse au glucose par les cellules β du pancréas p.18 Figure 7 : Effets de l’insuline p.20 Figure 8 : Effets du système nerveux autonome sur l’homéostasie énergétique p.22 Figure 9 : Demi-coupes du cerveau d’un rat p.25 Figure 10 : Localisation des neurones hypothalamiques synthétisant les neuropeptides majeurs impliqués dans l’homéostasie énergétique p.32 Figure 11 : Intégration des signaux d’adiposité au niveau du noyau arqué p.33 Figure 12 : Modèle d’intégration GABA-ergique dans le noyau hypothalamique paraventriculaire des signaux neuropeptidergiques en provenance du noyau arqué p.35 Figure 13 : Connexions nerveuses intrahypothalamiques, impliquées dans l’homéostasie énergétique p.36 Figure 14 : Illustration de la boucle de contrôle existant entre les organes périphériques et le cerveau p.38 Figure 15 : Dialogue entre les voies de signalisation générées par la liaison de l’insuline et de la leptine à leur récepteur p.40 Figure 16 : Régulation de l’activité de l’AMPK du noyau arqué et du noyau hypothalamique paraventriculaire par les signaux hormonaux et le statut énergétique de l’organisme p.41 Figure 17 : Régulation de l’expression des neuropeptides par la leptine et l’insuline p.42

- 7 -
Figure 18 : Suggestion de mécanisme de régulation des canaux potassiques ATP-dépendants par la leptine, l’insuline et l’acide oléique p.46 Figure 19 : Effets des acides gras libres indépendants des hormones p.47 Figure 20 : Mécanismes de régulation mis en place suite à l’hyperglycémie post-prandiale p.49 Figure 21 : Voie des hexosamines p.51 Figure 22 : la voie des hexosamines au carrefour des métabolismes des différents types de nutriments, au travers de l’UDP-N-acétylglucosamine p.52 Figure 23 : Détection des variations glycémiques par le système nerveux central p.53 Figure 24 : Effet du glucose sur l’activité électrique de neurones du noyau arqué p.56 Figure 25 : Effets d’une hypoglycémie sur certains neurones hypothalamiques et influence de neuropeptides sur les neurones gluco-excités p.57 Figure 26 : Fonctions principales de l’astrocyte p.58 Figure 27 : Schéma du couplage astrocyte-neurone p.59 Figure 28 : Schéma illustrant la sensibilité au glucose dans les cellules nerveuses, comprenant les deux protéines-clé, GLUT2 et GK p.64 Figure 29 : Système de perfusion pour la fixation intracardiaque/intracarotidienne p.72 Figure 30 : Variation de la glycémie et de l’insulinémie suite à l’injection intracarotidienne vers le cerveau de sérum physiologique, de D-glucose ou de L-glucose p.81 Figure 31 : Variation de l’insulinémie suite à une deuxième injection de D-glucose (10 min après la première) p.82 Figure 32 : Variation de l’insulinémie après prétraitement à l’atropine méthylnitrate ou à l’oxymétazoline, en réponse à l’injection vers le cerveau de D-glucose p.83 Figure 33 : Validation de la technique utilisant l’immunohistochimie anti-c-Fos : effet d’une injection intrapéritonéale de 2-déoxyglucose versus une injection de sérum physiologique, sur l’expression de c-Fos p.84 Figure 34 : Validation de la technique utilisant l’immunohistochimie anti-c-Fos suite à l’injection de 2-déoxyglucose : spécificité du marquage p.85 Figure 35 : Effet de l’injection de D-glucose vers le cerveau sur l’expression de c-Fos dans différentes régions hypothalamiques p.86

- 8 -
Figure 36 : Caractérisation des cellules immunopositives pour c-Fos suite à l’injection de D-glucose vers le cerveau : double immunohistochimie c-Fos/marqueur cellulaire p.88 Figure 37 : Effet du glucose sur les astrocytes des noyaux arqué et hypothalamique paraventriculaire p.89 Figure 38 : Boucle de contrôle mise en place entre la détection cérébrale du glucose et la sécrétion d’insuline par le pancréas p.98 Figure 39 : Schéma hypothétique de l’action de la méthionine sulfoximine sur le couplage astrocyte-neurone p.99 Figure 40 : Variation de la quantité de glycogène dans le foie et le système nerveux central suite à un prétraitement à la méthionine sulfoximine p.101 Figure 41 : Effet d’un prétraitement intrapéritonéal à la méthionine sulfoximine sur l’expression de c-Fos induite par l’injection de D-glucose vers le cerveau p.102 Figure 42 : Effet d’un prétraitement intrapéritonéal à la méthionine sulfoximine sur la sécrétion d’insuline induite par l’injection de D-glucose vers le cerveau p.104 Figure 43 : Effet d’un prétraitement intrapéritonéal à la buthionine sulfoximine versus à la méthionine sulfoximine sur la sécrétion d’insuline induite par l’injection de D-glucose vers le cerveau, ainsi que sur la glycémie p.107 Figure 44 : Effet de l’injection intracarotidienne vers le cerveau de glucosamine sur la sécrétion d’insuline induite par celle simultanée de D-glucose p.113 Figure 45 : Effet de l’injection par voie intracarotidienne vers le cerveau de glucosamine sur un test de tolérance au glucose : effet sur la glycémie et sur l’insulinémie p.114 Figure 46 : Effet de l’injection intracarotidienne vers le cerveau de N-acétylglucosamine sur la sécrétion d’insuline induite par celle simultanée de D-glucose p.116 Figure 47 : Effet de l’injection intracarotidienne vers le cerveau d’alloxane au temps 0 sur la sécrétion d’insuline induite par celle simultanée de D-glucose p.117 Figure 48 : Effet d’une pré-injection par voie intracarotidienne vers le cerveau d’alloxane sur la sécrétion d’insuline induite par l’injection de D-glucose p.118 Figure 49 : Effets de la glucosamine sur la glucokinase et la voie des hexosamines, qui est aussi capable d’incorporer la N-acétylglucosamine p.121 Figure 50 : Détection d’une hyperglycémie cérébrale impliquant les astrocytes du noyau arqué et la glucokinase p.126

- 9 -
Table des matières
IINNTTRROODDUUCCTTIIOONN ____________________________________________________________ - 12 -
I – HOMEOSTASIE ENERGETIQUE _______________________________________________ - 13 - 1. GENERALITES ______________________________________________________________ - 13 - 2. ACTEURS HORMONAUX REGULANT L’HOMEOSTASIE ENERGETIQUE _________________ - 14 - 2.1. La leptine_______________________________________________________________ - 15 - 2.2. L’insuline _______________________________________________________________ - 18 - 3. REGULATION NERVEUSE DE L’HOMEOSTASIE ENERGETIQUE ______________________ - 21 - 3.1. Régulation par le système nerveux autonome ____________________________ - 21 - 3.2. Régulation par le système nerveux central _______________________________ - 23 - II – HYPOTHALAMUS ET METABOLISME ENERGETIQUE ____________________________ - 26 - 1. NEUROPEPTIDES PARTICULIERS_______________________________________________ - 26 - 1.1. Les neuropeptides orexigènes____________________________________________ - 27 - 1.1.1. Le neuropeptide Y (NPY) _______________________________________________ - 27 - 1.1.2. L’AgRP (Agouti-Related Peptide)________________________________________ - 28 - 1.1.3. La MCH (Melanin-Concentrating Hormone)_____________________________ - 29 - 1.1.4. Les orexines (hypocrétines) ____________________________________________ - 29 - 1.2. Les neuropeptides anorexigènes _________________________________________ - 30 - 1.2.1. L’α-MSH (alpha-melanocyte-stimulating hormone)______________________ - 30 - 1.2.2. Le CART (Cocaine- and Amphetamine-Regulated Transcript) ____________ - 30 - 2. ROLE MAJEUR DU NOYAU ARQUE______________________________________________ - 32 - 3. CONNEXIONS INTRAHYPOTHALAMIQUES ________________________________________ - 34 - 4. CONNEXIONS EXTRAHYPOTHALAMIQUES EN DIRECTION DU TRONC CEREBRAL _______ - 37 - III – SENSIBILITE HYPOTHALAMIQUE AUX INFORMATIONS METABOLIQUES ___________ - 39 - 1. SENSIBILITE AUX HORMONES _________________________________________________ - 39 - 1.1. Signalisation cérébrale de la leptine______________________________________ - 41 - 1.2. Signalisation cérébrale de l’insuline______________________________________ - 43 - 2. SENSIBILITE AUX METABOLITES _______________________________________________ - 44 - 2.1. Les acides gras__________________________________________________________ - 45 - 2.2. Le glucose______________________________________________________________ - 48 - 2.2.1. Généralités sur l’homéostasie glucidique _______________________________ - 48 - 2.2.2. Une voie annexe suivie par le glucose : la voie des hexosamines _________ - 50 - 2.2.3. L’hypothalamus, siège de la sensibilité cérébrale au glucose ____________ - 52 - 2.2.4. Acteurs cellulaires impliqués dans la détection du glucose ______________ - 55 - 2.2.5. Acteurs moléculaires __________________________________________________ - 60 - IV – NOS OBJECTIFS ET DEMARCHES SCIENTIFIQUES _____________________________ - 66 - 1. NOTRE MODELE D’HYPERGLYCEMIE CEREBRALE ________________________________ - 66 - 2. LE PROTO-ONCOGENE C-FOS _________________________________________________ - 67 - 3. OUTILS PHARMACOLOGIQUES_________________________________________________ - 67 -
MMAATTEERRIIEELLSS EETT MMEETTHHOODDEESS________________________________________________ - 69 -
V – ANIMAUX_________________________________________________________________ - 70 -

- 10 -
VI – ETUDE DE L’ACTIVATION CEREBRALE _______________________________________ - 70 - 1. VALIDATION DE LA TECHNIQUE FOS : INJECTION DE 2-DEOXYGLUCOSE ____________ - 70 - 2. MICROCHIRURGIE : POSE D’UN CATHETER « A DEMEURE » DANS LA CAROTIDE _______ - 70 - 3. PROTOCOLES D’INJECTION ___________________________________________________ - 71 - 4. FIXATION INTRACAROTIDIENNE ET INTRACARDIAQUE _____________________________ - 71 - 5. IMMUNOHISTOCHIMIE C-FOS _________________________________________________ - 72 - 6. DOUBLE IMMUNOHISTOCHIMIE C-FOS / MARQUEUR CELLULAIRE__________________ - 73 - 7. QUANTIFICATION DES NOYAUX IMMUNOPOSITIFS POUR C-FOS ET DES CELLULES DOUBLEMENT IMMUNOPOSITIVES C-FOS / GFAP __________________________________ - 74 - 8. EXPRESSION DES RESULTATS_________________________________________________ - 74 - VII – ETUDE DE LA REGULATION NERVEUSE DE LA SECRETION D’INSULINE __________ - 75 - 1. MICROCHIRURGIE : POSE D’UN CATHETER DANS LA CAROTIDE_____________________ - 75 - 2. PROTOCOLES D’INJECTION ___________________________________________________ - 75 - 3. TEST DE TOLERANCE AU GLUCOSE (IVGTT) ____________________________________ - 76 - 4. PRELEVEMENTS SANGUINS ___________________________________________________ - 76 - 5. DOSAGES__________________________________________________________________ - 77 - 6. EXPRESSION DES RESULTATS_________________________________________________ - 77 - VIII – METHIONINE SULFOXIMINE : DOSAGE DU GLYCOGENE CEREBRAL_____________ - 77 - IX – ANALYSES STATISTIQUES__________________________________________________ - 78 -
RREESSUULLTTAATTSS ________________________________________________________________ - 79 -
X – CARACTERISATION DE LA SENSIBILITE AU GLUCOSE ___________________________ - 80 - 1. RESULTATS I _______________________________________________________________ - 80 - 1.1. Mise en place et validation des modèles expérimentaux ___________________ - 80 - 1.1.1. Injection(s) aiguë(s) de glucose vers le système nerveux central : effets sur la glycémie et l’insulinémie _____________________________________________________ - 80 - 1.1.2. Mise en évidence des voies nerveuses impliquées _______________________ - 82 - 1.1.3. Validation de la technique immunohistochimique dirigée contre l’antigène c-Fos _______________________________________________________________________ - 84 - 1.2. Régions hypothalamiques activées par l’injection intracarotidienne de glucose _____________________________________________________________________________ - 86 - 1.3. Types cellulaires activés _________________________________________________ - 87 - 2. DISCUSSION I ______________________________________________________________ - 89 - 2.1. Réponse hormonale périphérique ________________________________________ - 89 - 2.2. Réponse hypothalamique________________________________________________ - 92 - 2.3. Réponses cellulaires ____________________________________________________ - 95 - XI – IMPLICATION DES ASTROCYTES DANS LA SENSIBILITE AU GLUCOSE_____________ - 99 - 1. RESULTATS II _____________________________________________________________ - 100 - 1.1. Validation de l’utilisation de la méthionine sulfoximine __________________ - 100 - 1.2. Conséquences de l’arrêt du métabolisme astrocytaire des carbohydrates_______________________________________________________________- 101 - 1.2.1. sur l’activation cérébrale induite par le bolus intracarotidien de glucose - 101 - 1.2.2. sur la sécrétion d’insuline ____________________________________________ - 103 - 1.3. Spécificité de la méthionine sulfoximine_________________________________ - 105 - 1.3.1. Effet uniquement cérébral ou aussi périphérique ? ____________________ - 105 - 1.3.2. Effet spécifique à l’inhibition de la glutamine synthase ? _______________ - 106 -

- 11 -
2. DISCUSSION II ____________________________________________________________ - 108 - XII – ACTEURS MOLECULAIRES IMPLIQUES DANS LA SENSIBILITE CEREBRALE AU GLUCOSE _____________________________________________________________________________- 112 - 1. RESULTATS III_____________________________________________________________ - 112 - 1.1. Etude de l’implication de la glucokinase_________________________________ - 112 - 1.1.1. Effet de la glucosamine sur la sécrétion d’insuline _____________________ - 112 - 1.1.2. Effet cérébral et non périphérique de la glucosamine __________________ - 113 - 1.1.3. Effet spécifique à l’inhibition de la glucokinase ? ______________________ - 115 - 1.2. Etude de l’implication du couple glucokinase/GLUT2____________________ - 116 - 1.2.1. Injection de l’alloxane au temps zéro __________________________________ - 116 - 1.2.2. Injection de l’alloxane au temps -10 minutes __________________________ - 117 - 2. DISCUSSION III ____________________________________________________________ - 119 -
CCOONNCCLLUUSSIIOONNSS EETT PPEERRSSPPEECCTTIIVVEESS ________________________________________ - 124 -
AARRTTIICCLLEE EETT RREEVVUUEESS PPUUBBLLIIEESS____________________________________________ - 131 -
RREEFFEERREENNCCEESS BBIIBBLLIIOOGGRRAAPPHHIIQQUUEESS _______________________________________ - 147 -

- 12 -
IInnttrroodduuccttiioonn

- 13 -
I – Homéostasie énergétique
1. Généralités
L’homéostasie énergétique est un état d’équilibre dynamique entre les
dépenses et les apports d’énergie, permettant le maintien de la stabilité du milieu
interne d’un organisme (Campbell NA, 1995). Les dépenses résultent
principalement de l’activité physique et du métabolisme basal, tandis que les
apports trouvent leur origine dans l’alimentation (Figure 1).
Par la tenue de cet équilibre, un individu conserve et défend un poids
corporel stable, compris dans un intervalle dont les limites inférieure et supérieure
seraient génétiquement définies. En cas d’éloignement au-dessus ou en-dessous de
cette fourchette, tout un système de régulations se met en place afin d’y revenir
(Woods SC et coll., 2004).
Figure 1 : Illustration de l’homéostasie énergétique
L’homéostasie énergétique est contrôlée et régulée à plusieurs niveaux.
Schématiquement, il existe un dialogue entre la périphérie et le système nerveux
central (SNC), faisant intervenir des signaux complémentaires et en perpétuelle
interaction, de natures hormonale, métabolique ou nerveuse (Broberger C, 2005).
Notons que cet équilibre est également sous l’influence de facteurs indépendants au
métabolisme de l’organisme comme par exemple les phénomènes sociaux, cognitifs
ou, d’une manière plus générale, l’environnement. Ils sont importants, notamment
chez l’Homme, mais difficilement contrôlables ou mesurables (Berthoud HR, 2004)
(Figure 2).

- 14 -
Figure 2 : Interactions établies entre les organes périphériques et le système nerveux central (SNC) ; CCK : cholécystokinine
A court terme, c'est-à-dire sur un seul repas, l’apport énergétique est
déterminé par la taille de ce dernier et sa valeur calorique. La boucle de régulation
entre la périphérie et le SNC aboutira à un signal de satiété, issu du tractus
digestif, normalement suivi d’un arrêt de la prise alimentaire chez un individu
normorégulé (Woods SC, 2005). Mais l’homéostasie énergétique ne se mesure pas
sur un unique repas, ni même sur une journée. Elle se réalise sur une plus longue
période comptant quelques jours. Ainsi, il est nécessaire de considérer une
régulation à plus long terme mettant en jeu des phénomènes de stockage et de
mobilisation des réserves (Schwartz MW et coll., 2000).
2. Acteurs hormonaux régulant l’homéostasie énergétique
De nombreuses hormones sont impliquées dans l’homéostasie énergétique,
jouant leurs rôles régulateurs en périphérie et/ou sur le cerveau comme la ghréline,

- 15 -
la cholécystokinine (CCK), le peptide YY (PYY), le glucagon-like peptide-1 (GLP-1), le
glucagon, la leptine et l’insuline (Ueno H et coll., 2005 ; Little TJ, 2005 ; McGowan
BMC et Bloom SR, 2004 ; Bojanowska E, 2005 ; Jiang G et Zhang BB, 2003 ;
Morton GJ et coll., 2006) (Figure 3).
Figure 3 : Lieu de synthèse et action des principales hormones jouant sur l’homéostasie énergétique ; AgRP : agouti related protein ; CCK : cholécystokinine ; GLP-1 : glucagon-like peptide-1 ; NPY : neuropeptide Y ; PYY : peptide YY
La leptine et l’insuline, considérées comme des signaux d’adiposité, font
l’objet des deux paragraphes suivants, consacrés à leurs effets généraux. Leurs
mécanismes d’action cérébraux sont développés dans le paragraphe III.1.
2.1. La leptine
La leptine est le produit du gène ob (« ob » pour obésité), communément
appelé « gène de l’obésité », localisé chez la souris en 1994 (Zhang Y et coll., 1994).
Il s’agit d’une cytokine de 16 kDa, secrétée principalement par le tissu adipeux et
de façon moindre par l’épithélium gastrique, le muscle squelettique ou le placenta
(Ahima RS et Osei SY, 2004).

- 16 -
Le récepteur à la leptine, Ob-R, se présente sous plusieurs isoformes divisées
en trois groupes, quelque soit l’espèce considérée : secrétée, courte et longue. La
première forme ne comporte que la partie extracellulaire du récepteur et se lie à la
leptine circulante. Les isoformes courte et longue comprennent les parties
transmembranaires, extra- et intracellulaires. C’est la longueur de la partie
intracellulaire qui, alors, les différencie. Les Ob-R n’ont pas d’activité enzymatique
intrinsèque, mais la partie intracellulaire s’associe avec les Janus Kinases (JAK),
membres de la famille des tyrosine kinases. Ce mécanisme est absent avec les Ob-R
courts. Ainsi, ces derniers auraient pour fonction d’assurer le transport de la
leptine à travers la barrière hémato-encéphalique (BHE) (Bates SH et Myers MG Jr,
2003).
Au niveau du SNC la signalisation induite par la leptine passe par la forme
longue du récepteur, Ob-Rb, dont l’expression la plus forte a été retrouvée dans
l’hypothalamus ventromédian, tandis que la signalisation périphérique emprunte
davantage les formes courtes qui ont une localisation ubiquitaire (Bates SH et
Myers MG Jr, 2003). Grâce à ses deux récepteurs, la leptine exerce une multitude
d’effets physiologiques (Moran O et Phillip M, 2003) (Figure 4).
Figure 4 : Les différents effets de la leptine selon le tissu cible

- 17 -
Son rôle dans la régulation de l’homéostasie énergétique est particulièrement
connu et étudié. En effet, les souris ob/ob, ainsi que les souris db/db (« db » pour
diabète) déficientes pour son récepteur Ob-R, sont obèses et diabétiques,
symptômes ressemblants au syndrome de l’obésité morbide rencontrée chez
l’humain, leur poids étant triple et leur masse adipeuse quintuple par rapport à des
souris sauvages soumises à un régime identique (Friedman JM et Halaas JL, 1998).
En effet, cette cytokine induit la réduction de la masse graisseuse en jouant sur le
métabolisme lipidique, diminue le contenu en triglycérides des différents tissus,
inhibe la synthèse d’insuline par les cellules β du pancréas, régule le métabolisme
du glucose et enfin, diminue la prise alimentaire (Moran O et Phillip M, 2003).
La leptinémie est proportionnelle à la masse du tissu adipeux, permettant
ainsi de refléter l’état énergétique global d’un organisme. Une augmentation de la
leptinémie induit une augmentation des dépenses et une diminution des apports
énergétiques (Friedman JM et Halaas JL, 1998) (Figure 5). Ces caractéristiques font
de la leptine un régulateur à long terme de l’homéostasie énergétique, d’autant plus
que sa concentration sanguine et l’adiposité n’augmentent pas de manière aiguë au
cours d’un repas.
Figure 5 : Reflet de l’état énergétique d’un individu par sa leptinémie

- 18 -
Toutefois, elle ne peut être utilisée dans les cas d’obésité induite par un
déséquilibre énergétique chronique, car chez ces patients est observée une
leptinorésistance. En revanche, elle s’avère efficace dans les cas d’obésité
congénitale, relativement peu fréquents, où le gène de la leptine ou celui de son
récepteur est muté.
2.2. L’insuline
Il s’agit d’une hormone quasi-exclusivement synthétisée par les cellules β du
pancréas. Une petite production cérébrale existe néanmoins même si elle prête
encore à controverse (Schwartz MW et coll., 1992 ; Banks WA, 2004).
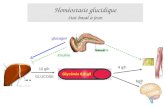
La fonction générale de l’insuline est le contrôle strict de la glycémie, ses
effets hypoglycémiants s’opposant à ceux du glucagon, qui lui est synthétisé par les
cellules α (Jiang G et Zhang BB, 2003). Dans le plasma, elle circule sous forme
libre, non liée à une « protéine de transport » (Baulieu EE et Kelly PA, 1990). Les
mécanismes impliqués dans la sécrétion d’insuline induite par le glucose sont bien
documentés (Doyle ME et Egan JM, 2003) (Figure 6).
Figure 6 : Sécrétion de l’insuline en réponse au glucose par les cellules β du pancréas

- 19 -
Les cellules β expriment un transporteur au glucose particulier, GLUT2 (cf. §
III-2.2.5), dont les caractéristiques biochimiques permettent un passage de cet
hexose dans la cellule de manière proportionnelle à sa concentration environnante.
Cette propriété aboutit à un équilibre de la concentration en glucose de part et
d’autre de la membrane cytoplasmique. En d’autres termes, la concentration
intracellulaire en glucose reflète la concentration extracellulaire. Cet hexose est
ensuite rapidement phosphorylé en glucose-6-phosphate par la glucokinase (GK ;
cf. § III-2.2.5) qui détermine le niveau de glycolyse et ainsi la quantité de pyruvate
qui entrera dans la mitochondrie pour emprunter le cycle de Krebs. Cette
succession de réactions enzymatiques conduit à une augmentation du rapport
ATP/ADP dans le cytoplasme, ce qui induit la fermeture de canaux potassiques
sensibles à l’ATP. Celle-ci engendre alors une dépolarisation membranaire et
l’ouverture de canaux calciques sensibles au voltage. L’augmentation de la
concentration intracellulaire en ions Ca2+ qui en résulte, provoque la fusion, à la
membrane plasmique, des vésicules contenant l’insuline et par-là même son
exocytose. De plus, l’ATP agit également directement sur ces vésicules en favorisant
leur migration vers la membrane cellulaire.
Il existe aussi une régulation nerveuse de cette sécrétion, mise en jeu avant
même l’ingestion des aliments, indépendante d’une variation de la glycémie,
nommée « phase céphalique ». En dehors de celle-ci, la sécrétion d’insuline en
réponse à une hyperglycémie est typiquement biphasique chez des individus sains
(Del Prato S, 2003). L’hormone est rapidement libérée et sa concentration maximale
quelques minutes seulement après l’absorption de glucose par voie orale ou son
injection par voie intraveineuse. Cette sécrétion initiale ne dure qu’une dizaine de
minutes et est suivie d’une autre plus soutenue, se prolongeant pendant plusieurs
heures, jusqu’à ce que la glycémie revienne à une valeur basale (5 mM, soit 1g/L).
Environ 3% du contenu en insuline des cellules β sont libérés lors de la phase
précoce de sécrétion, tandis que 20% le sont lors de la seconde phase.
Le récepteur à l’insuline est ubiquitaire, localisé dans la quasi-totalité des
tissus des mammifères. Ceux l’exprimant le plus sont le tissu adipeux, le foie et les
muscles, c’est-à-dire les tissus dits insulino-sensibles (Cheatham B et Kahn CR,
1995). Il est constitué de deux sous-unités α extracellulaires et de deux autres β
transmembranaires possédant une activité tyrosine kinase intrinsèque (Van
Obberghen et coll., 2001). Son activation, par liaison de l’hormone, induit la
phosphorylation de résidus tyrosines de divers substrats cellulaires, comme les

- 20 -
protéines de la famille des IRS (Insulin Receptor Substrate) ou des SHC (Src-
Homology-Collagen). Ce mécanisme est suivi par différentes cascades de
signalisation qui se traduiront par différents effets physiologiques.
Figure 7 : Effets de l’insuline
L’insuline assure de multiples fonctions (Figure 7), mais constitue surtout
un facteur anabolique primordial dans le maintien de l’homéostasie énergétique, au
point que des souris totalement déficientes en son récepteur meurent peu de temps
après leur naissance de cétoacidose et d’un dysfonctionnement métabolique (Van
Obberghen et coll., 2001). Elle assure le maintien des normoglycémie et
normolipidémie. Cette hormone intervient à plusieurs niveaux, d’où son
importance. D’une manière générale, l’insuline favorise l’utilisation du glucose par
les tissus dits insulino-sensibles. Elle augmente ainsi le transport du glucose dans
les cellules musculaires et adipocytaires en induisant la translocation du
transporteur au glucose, GLUT4, vers la membrane (Chang L et coll., 2004). Au

- 21 -
niveau du foie, l’insuline inhibe la glycogénolyse et la gluconéogenèse, tout en
stimulant la glycogenèse, cette activation s’observant également dans le muscle
strié. Dans le foie et le tissu adipeux, elle augmente la lipogenèse, en diminuant
dans le même temps la lipolyse (Baulieu EE et Kelly PA, 1990).
A ces effets périphériques s’ajoutent des effets centraux parmi lesquels une
inhibition de la prise alimentaire et du poids corporel (Stockhorst U et coll., 2004).
Les mécanismes mis en jeu font l’objet du paragraphe III-1.2.
3. Régulation nerveuse de l’homéostasie énergétique
Le système endocrinien n’est pas seul à réguler la balance énergétique. Un
autre système est primordial pour assurer cette tâche : le système nerveux,
autonome (SNA) et central (SNC). Leur implication sera traitée séparément bien
qu’ils soient indissociables puisque le SNA véhicule les informations provenant du
SNC vers la périphérie et vice versa.
3.1. Régulation par le système nerveux autonome
Le SNA contrôle les fonctions végétatives d’un organisme. Il est constitué des
systèmes nerveux para- et orthosympathique, distincts au niveau de l’anatomie, de
la physiologie et de la neurochimie. Généralement, leurs effets sont antagonistes et
complémentaires sur un organe-cible, impliquant comme neuromédiateurs
l’acétylcholine pour les fibres parasympathiques et la noradrénaline pour les fibres
orthosympathiques (appelées plus simplement sympathiques) (Campbell NA, 1995)
(Figure 8).
Par rapport à l’homéostasie énergétique, l’augmentation du tonus
parasympathique induit le stockage du glucose et des acides gras libres (AGL), alors
que celle de l’orthosympathique induit la mobilisation de ces composés à partir des
tissus de réserves (Scheurink AJW et Nolan LJ, 1996). D’un autre côté, des
variations dans les apports énergétiques provoquent des changements d’activité du
SNA. En effet, l’absence d’apports lipidiques provoque une activation du système
orthosympathique alors qu’un épisode hypoglycémique engendre celle du système
parasympathique.
Deux organes fondamentaux dans la régulation du métabolisme sont le
pancréas et le foie. Or, ils montrent une riche innervation par le SNA. Au niveau du
pancréas, à la fois les cellules α et β sont sous la dépendance de ce contrôle :
l’activation des nerfs splanchniques induit une augmentation de la sécrétion de

- 22 -
glucagon et celle du nerf vague stimule la libération d’insuline (Nonogaki K, 2000).
Ainsi, suite à une vagotomie ou à l’administration d’atropine (antagoniste des
récepteurs à acétylcholine), la première phase de sécrétion d’insuline n’apparaît
plus suite à l’injection de glucose ce qui illustre l’existence du contrôle
parasympathique sur les îlots de Langerhans et son implication lors de l’apparition
d’une hyperglycémie (Scheurink AJW et Nolan LJ, 1996). Le foie, quant à lui, se
trouve sous la régulation du SNA à la fois de manière directe et indirecte : directe
grâce à l’innervation des hépatocytes, indirecte par l’effet de l’insuline sécrétée sous
l’influence du système parasympathique (Nonogaki K, 2000). L’innervation
sympathique peut induire la libération de glucose dans la circulation sanguine,
mais ce mécanisme semble mineur en comparaison à l’action hormonale (via le
glucagon) stimulant la glycogénolyse ou la gluconéogenèse. De l’autre côté, l’action
directe du système parasympathique sur le foie est requise pour que l’insuline
conduise à une capture du glucose efficace (Püschel GP, 2004).
Figure 8 : Effets du système nerveux autonome sur l’homéostasie énergétique

- 23 -
Les autres organes sous l’influence du SNA sont les muscles squelettiques et
le tissu adipeux. Au niveau des adipocytes, les fibres orthosympathiques stimulent
la lipolyse et la libération des AGL, tandis que les parasympathiques jouent sur la
capture du glucose en augmentant la sensibilité à l’insuline (Fliers E et coll., 2003).
Au niveau des cellules musculaires, le système sympathique active la capture de
glucose et la glycogénolyse (Nonogaki K, 2000).
3.2. Régulation par le système nerveux central
L’implication du SNC, et plus particulièrement de certaines régions
cérébrales, dans le contrôle de l’homéostasie énergétique a été découverte par le
biais d’observations cliniques datant d’un siècle (Elmquist JK, 1999). Des patients
souffrant du syndrome de Fröhlich montrent, en plus des tumeurs hypophysaires,
des dépôts de graisses sous-cutanés importants. Or, Aschner démontra en 1912
que ce phénotype est dû à des dommages de l’hypothalamus et non à ceux de
l’hypophyse per se puisque des chiens privés de cette glande ne développent pas
d’obésité. C’est en 1940 que Hetherington et Ranson établirent que l’hypothalamus
régulait la prise alimentaire et le poids corporel, grâce à des expériences de lésions
électrolytiques bilatérales de sa partie ventromédiane : elles provoquent une
augmentation de l’adiposité aboutissant à un doublement du poids. Par ailleurs ces
chercheurs notèrent une diminution de la prise alimentaire lorsque la région de
l’hypothalamus latéral était lésée. Ces données aboutirent à la théorie d’un « centre
de la faim », désignant la région hypothalamique latérale (LH), et d’un « centre de la
satiété », correspondant au noyau hypothalamique ventromédian (VMN).
Depuis, des modèles expérimentaux d’obésité ont été mis au point par
traitement pharmacologique provoquant des lésions cérébrales plus ou moins
spécifiques et certains phénotypes observés furent reliés à des lésions cérébrales
données. Ainsi l’acide kaïnique induit une hypophagie liée à la lésion du LH
(Elmquist JK, 1999). Des rats rendus diabétiques par injection de streptozotocine
ou des souris rendues obèses par injection périnatale de monosodium de
glutamate, montrent des modifications du noyau arqué (NA) et de l’éminence
médiane sous-jacente, caractérisées par une dégénérescence des neurones, une
hypotrophie des tanycytes et des changements axonaux (Bestetti G et Rossi GL,
1980). Des souris recevant de l’aurothioglucose développent en plus des lésions
dans le VMN (Sanchis-Segura C et Aragon CMG, 2002). Enfin, le bipiperidyl
provoque des lésions dans le VMN et le noyau du tractus solitaire (NTS), également
associées à une obésité sévère (Laughton W et Powley TL, 1981).

- 24 -
L’hypothalamus contrôle l’activité des systèmes nerveux para- et
orthosympathiques par l’intermédiaire de projections sur le tronc cérébral, au
niveau du NTS, des noyaux moteurs du vague, et de la médulla ventrolatérale. Il en
découle que des lésions du diencéphale engendrent des modifications de l’activité
du SNA (Pénicaud L et coll., 1996). Ceci est illustré par le fait que la lésion du VMN
de souris ou de rats (entraînant le développement d’une obésité) se caractérise par
une augmentation du tonus parasympathique et une diminution de
l’orthosympathique. Il est intéressant de constater que ces altérations apparaissent
également chez des animaux dont l’obésité est de nature génétique. Elles
aboutissent en particulier à une sécrétion excessive d’insuline par le pancréas et
donc à une hyperinsulinémie.
Ces différentes données montrent que le SNC, et plus spécifiquement
certaines régions hypothalamiques que sont le VMN, le NA et le LH, régulent
l’homéostasie énergétique en contrôlant la prise alimentaire et le fonctionnement du
SNA, via d’importants relais au niveau du tronc cérébral (Figure 9). Ultérieurement,
nous verrons que les noyaux hypothalamiques paraventriculaire (PVN) et
dorsomédian (DMN) sont également importants dans ces contrôles.

- 25 -
Figure 9 : Demi-coupes du cerveau d’un rat, l’une située à 1,8 mm et l’autre à 3,3 mm en arrière du bregma (point repère utilisé en stéréotaxie, où se rejoignent les 4 plaques crâniennes). Sont indiquées les régions hypothalamiques impliquées dans le contrôle de l’homéostasie énergétique. 3V : troisième ventricule ; DMN : noyau hypothalamique dorsomédian ; LH : hypothalamus latéral ; NA : noyau arqué ; PVN : noyau hypothalamique paraventriculaire ; VMN : noyau hypothalamique ventromédian.

- 26 -
II – Hypothalamus et métabolisme énergétique
1. Neuropeptides particuliers
Les cellules hypothalamiques expriment et secrètent un certain nombre de
neurotransmetteurs permettant de « traduire » l’information de nature métabolique.
Les neurotransmetteurs non peptidiques tels que le glutamate, la dopamine, la
sérotonine ou le GABA, prennent part à ces mécanismes mais de manière non
spécifique, intervenant également dans d’autres grandes fonctions physiologiques.
Les prochains paragraphes ne traiteront que des neuropeptides dont le rôle majeur
concerne la régulation du métabolisme énergétique. Les uns sont qualifiés
d’orexigènes car mis en jeu dans des conditions de déficit énergétique. Leur
synthèse et libération entraînent ainsi une stimulation de la prise alimentaire ainsi
qu’une diminution des dépenses énergétiques. Les autres sont qualifiés
d’anorexigènes : leur libération est activée lors d’apports énergétiques importants et
s’accompagne d’une inhibition de la prise alimentaire et/ou une stimulation des
dépenses énergétiques (Dupin H et coll., 1992 ; Kalra SP et coll., 1999) (Tableau 1).
Tableau 1 : Neurotransmetteurs principaux impliqués dans la régulation de la prise alimentaire. Ceux notés en italique ne seront pas traités dans les paragraphes suivants. AgRP: agouti-related protein ; α-MSH: alpha-melanocyte-stimulating hormone ; CART: cocaine- and amphetamine-regulated transcript ; CRH: Corticotropin-Releasing Hormone ; GABA: γ-aminobutyric acid ; MCH: melanin-concentrating hormone ; NPY: neuropeptide Y ; TRH: thyrotropin-releasing hormone.

- 27 -
1.1. Les neuropeptides orexigènes
1.1.1. Le neuropeptide Y (NPY)
Le plus connu pour son rôle dans le contrôle de l’homéostasie énergétique est
le NPY. C’est un neurotransmetteur appartenant à la famille des polypeptides
pancréatiques, largement distribué dans le SNC. Au niveau hypothalamique, les
neurones à NPY proviennent essentiellement du NA, mais il en existe aussi
quelques uns éparpillés dans le LH, l’aire préoptique, les noyaux péri- et
paraventriculaires ainsi que dans le DMN (Fetissov SO et coll., 2004). Son action
passe par sa liaison à des récepteurs de type « 7 domaines transmembranaires ». A
ce jour, ils sont au nombre de six, nommés Y1 à Y6. Les récepteurs Y1, Y2 et Y5,
présentent une meilleure affinité pour le NPY et semblent être davantage impliqués
dans le contrôle de la prise alimentaire (Ramos EJB et coll., 2005).
Différents travaux de recherches ont montré que l’injection de NPY dans
l’hypothalamus stimule la prise alimentaire, diminue les dépenses énergétiques et
induit la lipogenèse dans le foie et le tissu adipeux. Ainsi l’apport chronique de NPY
aboutit à une prise de poids de part son action anabolique, parallèlement à la mise
en place d’une insulinorésistance (Schwartz MW et coll., 2000). De plus, injecté
spécifiquement dans le PVN, ce neuropeptide empêche la libération d’insuline et de
glucagon, diminue la thermogenèse et bloque l’activité du système nerveux
sympathique (Currie PJ, 2003).
Les concentrations hypothalamiques en NPY reflètent le statut énergétique de
l’organisme. En effet, la synthèse de ses ARNm et sa libération augmentent avec le
jeûne, tandis qu’elles diminuent avec la prise alimentaire (Wynne K et coll., 2005).
Donc, comme le montre l’augmentation des concentrations en NPY, ces voies
nerveuses sont activées lorsque le statut énergétique est négatif, par exemple en
période de jeûne, de lactation ou de déficience insulinémique chez les diabétiques
(Frankish HM et coll., 1995). Enfin, ces cellules sont stimulées par une baisse des
concentrations circulantes de la leptine et insuline, ces hormones inhibant aussi
directement l’expression du NPY (Williams G et coll., 2004).
Néanmoins, lorsqu’on analyse les modèles animaux d’obésité, on observe
certaines contradictions. Les souris obèses ob/ob montrent une augmentation de
l’activité des neurones à NPY, tandis que dans le cas d’une obésité induite par un
régime cette activité semble réduite (Williams G et coll., 2004). Par ailleurs, les
souris ayant une délétion du gène codant pour le NPY ne présente ni surpoids ni
augmentation de l’adiposité, ce qui suggère des phénomènes de redondance des

- 28 -
systèmes impliqués dans le contrôle de la balance énergétique (Wynne K et coll.,
2005).
1.1.2. L’AgRP (Agouti-Related Peptide)
Au niveau du SNC, ce neuropeptide est exprimé dans l’hypothalamus bien
qu’une petite production ait été mise en évidence dans le noyau subthalamique
(Stütz AM et coll., 2005). Lorsque l’on parle du système orexigène, il est
généralement mentionné le couple NPY/AgRP. En effet, 90% des neurones à NPY du
NA synthétisent et libèrent l’AgRP qui est aussi capable de stimuler la prise
alimentaire. Ce neuropeptide exerce son effet antagonisme direct de l’α-MSH (alpha-
Melanocyte-Stimulating Hormone ; cf. § II-1.2.1), neuropeptide anorexigène, sur les
récepteurs aux mélanocortines de type MC3-MC4. La stimulation des neurones
coexprimant l’AgRP et le NPY aboutit donc à une hyperphagie en stimulant d’une
part les récepteurs à NPY et en inhibant d’autre part les récepteurs aux
mélanocortines (Broberger C et coll., 1998).
Les mécanismes d’action impliqués dans l’effet orexigène de l’AgRP ne sont
pas totalement élucidés. Une seule injection intracérébroventriculaire est capable
d’induire une hyperphagie encore visible après 7 jours (Stütz AM et coll., 2005). Par
ailleurs, l’AgRP induit une diminution des dépenses énergétiques, notamment de la
thermogenèse, probablement en freinant l’influx nerveux sympathique.
Les souris transgéniques surexprimant l’AgRP sont hyperphagiques, obèses,
diabétiques et ont une hyperplasie du pancréas (Stütz AM et coll., 2005). En
revanche, l’invalidation de ce gène génère un phénotype normal suggérant la mise
en place de mécanismes compensateurs, comme pour le NPY. On observe la même
chose pour les rongeurs ayant une double délétion AgRP/NPY. Cependant, une
approche par ARN d’interférence au niveau hypothalamique, conduit à une
diminution du métabolisme et du poids corporel sans modification de la prise
alimentaire, suggérant une augmentation des dépenses énergétiques.
Chez l’Homme, une augmentation de la concentration en AgRP est observée
chez les individus obèses. De manière très intéressante, différentes mutations d’un
seul nucléotide du gène codant pour ce neuropeptide ont été décrites et sont
caractérisées par une résistance à l’obésité au sein d’un environnement favorable à
ce phénotype métabolique (Stütz AM et coll., 2005).

- 29 -
1.1.3. La MCH (Melanin-Concentrating Hormone)
La MCH est synthétisée par les neurones magnocellulaires du LH et de la
zona incerta.
L’injection de ce neurotransmetteur à des animaux induit une hyperphagie et
à long-terme une obésité, tandis que la délétion de son gène aboutit à un phénotype
inverse (hypophagie, diminution de la masse adipeuse et augmentation du
métabolisme). Il semblerait que la MCH agisse davantage sur le contrôle des
dépenses énergétiques. En effet, l’activation de ses voies neuronales stimule
l’adiposité, augmente la lipogenèse dans le foie et le tissu adipeux tout en
diminuant la thermogenèse (Angelopoulos N et coll., 2005 ; Williams G et coll.,
2004).
1.1.4. Les orexines (hypocrétines)
Les orexines sont au nombre de deux, l’orexine A et l’orexine B, dérivant d’un
précurseur commun, la prépro-orexine. Les neurones à orexine, peu nombreux,
sont entremêlés aux neurones à MCH dans le LH mais constituent bien une
population distincte, aux très nombreuses projections (bulbes olfactifs, cortex,
tronc cérébral, thalamus, hypothalamus…). D’autres neurones à orexines sont
présents au niveau de l’aire périfornicale et du DMN (Williams G et coll., 2004 ;
Kirchgessner AL, 2002).
L’injection intracérébroventriculaire ou spécifiquement dans le LH d’orexines
stimule la prise alimentaire. D’autre part, des souris dont le gène codant pour la
prépro-orexine est invalidé, sont narcoleptiques et hypophagiques (Kirchgessner AL,
2002). A l’heure actuelle, on pense donc que l’effet principal des orexines se situe
au niveau du contrôle des rythmes éveil/sommeil et que l’effet orexigène ne serait
que secondaire. Cette stimulation, à court-terme, de la prise alimentaire
nécessiterait l’activation des neurones à NPY du NA (Williams G et coll., 2004).
Néanmoins, les neurones à orexines répondent à différents signaux métaboliques.
Notamment, la concentration en ARNm de la prépro-orexine augmente après un
épisode de jeûne ou en réponse à une hypoglycémie induite par l’injection
d’insuline, ce qui mène à penser que ces neurones sont activés dans des conditions
de faim (Kirchgessner AL, 2002).

- 30 -
1.2. Les neuropeptides anorexigènes
1.2.1. L’α-MSH (alpha-melanocyte-stimulating hormone)
L’α-MSH est le principal neuropeptide anorexigène. Il est produit par clivage
d’un gros précurseur polypeptidique, la pro-opiomélanocortine (POMC). Deux
classes de peptides sont générées à partir de la POMC : les mélanocortines (α-MSH,
γ-MSH, β-MSH, ACTH) et les β-endorphines (Pritchard LE et coll., 2002). La délétion
du gène de la POMC entraîne le développement d’une obésité sévère accompagnée
d’insuffisance surrénalienne et d’anomalie de pigmentation (Yaswen L et coll.,
1999). La mutation avec perte des deux allèles est à l’origine du même phénotype
chez l’être humain (Krude H et coll., 1998 ; Pritchard LE et coll., 2002).
Les neurones à POMC du NA constituent la population la plus importante
du SNC. Néanmoins, certains sont aussi retrouvés au niveau de l’hippocampe et du
NTS (Zheng H et coll., 2005). Les neurones à POMC du NA, comme les neurones à
NPY/AgRP, projettent majoritairement vers le PVN et le LH.
L’implication de l’α-MSH, et plus généralement du système aux
mélanocortines, dans la régulation du métabolisme, a émergé après l’identification
de ses récepteurs : les récepteurs MC3-R et MC4-R. Des souris invalidées pour le
récepteur MC4-R sont hyperphagiques et développent une obésité sévère (Weide K
et coll., 2003). Différentes mutations du gène de ce récepteur sont désormais
associées au développement d’une obésité chez l’enfant (Tao YX et Segaloff DL,
2003). L’ensemble des études menées dans ce domaine suggère que le système à α-
MSH induit un tonus anorexigène permanent via le récepteur MC4-R. Ainsi une
stimulation de la prise alimentaire résulterait davantage de la suppression d’un
tonus satiétogène, notamment par l’intermédiaire du système à AgRP. Néanmoins,
l’injection cérébrale d’α-MSH est capable de diminuer la prise alimentaire et le poids
corporel (Ramos EJB et coll., 2005). De plus, elle augmente l’activité du SNA
sympathique et les dépenses énergétiques, plus particulièrement la thermogenèse
(Wynne K et coll., 2005).
1.2.2. Le CART (Cocaine- and Amphetamine-Regulated Transcript)
Dans le NA, ce peptide est retrouvé dans la majorité des neurones à POMC.
Ainsi, comme dans le système orexigène où l’on a les « neurones à NPY/AgRP », le
système anorexigène du NA est constitué de « neurones à POMC/CART »
(Angelopoulos N et coll., 2005). En revanche, dans le DMN et le LH, ce peptide
colocalise avec la MCH, neuropeptide orexigène. Ce peptide est également présent
dans les VMN, PVN et NTS. Au niveau du NTS, les neurones à CART expriment un

- 31 -
récepteur à la CCK, suggérant leur implication dans le mécanisme de satiété induit
par cette hormone. Les recherches concernant ce neuropeptide ne sont pas encore
bien développées et à ce jour, on ne connaît toujours pas son récepteur (Hunter RG
et coll., 2004).
L’injection intracérébroventriculaire chronique de CART induit une
diminution de la prise alimentaire parallèle à une perte de poids et à une
diminution des insulinémie et leptinémie. Toutefois, une injection réalisée au sein
de différentes régions de l’hypothalamus (PVN, DMN, VMN, NA, LH) montre des
résultats inverses sur l’ingestion de nourriture (Hunter RG et coll., 2004).
Enfin, des souris dont le gène codant pour CART est invalidé, auxquelles on
présente une alimentation hypercalorique riche en graisses, montrent une
hyperphagie, une augmentation du poids corporel et de la masse adipeuse par
rapport aux souris sauvages. Par ailleurs, quelques études réalisées chez des
humains révèlent un lien entre certains cas d’obésité et un polymorphisme du gène
CART (Hunter RG et coll., 2004).

- 32 -
La Figure 10 récapitule les zones hypthalamiques où se déroule la synthèse
majeure des différents neuropeptides mentionnés.
Figure 10 : Localisation des neurones hypothalamiques synthétisant les neuropeptides majeurs impliqués dans l’homéostasie énergétique. Les orexigènes sont en bleu et les anorexigènes en rouge. 3V: troisième ventricule ; AgRP: agouti-related protein ; CART: cocaine- and amphetamine-regulated transcript ; DMN: noyau hypothalamique dorsomédian ; em: éminence médiane ; fx: fornix ; LH: hypothalamus latéral ; MCH: melanin-concentrating hormone ; NA: noyau arqué ; NPY: neuropeptide Y ; POMC: pro-opiomélanocortine ; PVN: noyau hypothalamique paraventriculaire ; VMN: noyau hypothalamique ventromédian.
2. Rôle majeur du noyau arqué
Le NA est une région hypothalamique essentielle pour le contrôle de la
balance énergétique du fait 1) de la coexistence de deux populations neuronales
aux effets antagonistes et complémentaires, NPY/AgRP d’une part, POMC/CART
d’autre part, mais aussi 2) de la présence des récepteurs liant des hormones clés,
en particulier la leptine et l’insuline (cf. § III-1).

- 33 -
De plus, le NA possède des particularités anatomiques stratégiques pour un
tel rôle. En effet, il est situé à proximité du troisième ventricule et juste au-dessus
de l’éminence médiane. Il se trouve donc à l’interface entre le liquide céphalo-
rachidien et la circulation sanguine. De plus, l’endothélium vasculaire au niveau de
l’éminence médiane présente la particularité d’être fenestré. Il n’y aurait donc pas
de réelle barrière hémato-encéphalique (BHE) protégeant le NA et les échanges entre
le compartiment sanguin et le parenchyme nerveux y seraient facilités (Kalra SP et
coll., 1999).
Grâce à ces caractéristiques, les cellules nerveuses du NA peuvent
rapidement détecter des variations de concentrations plasmatiques hormonales ou
de paramètres métaboliques et ajuster, suite à des cascades de signalisation
intracellulaires, la libération des neurotransmetteurs. Ainsi, le NA joue un rôle
d’intégrateur métabolique (Niswender KD et coll., 2004) (Figure 11).
Figure 11 : Intégration des signaux d’adiposité au niveau du noyau arqué (NA). 3V : troisième ventricule ; AgRP: agouti-related protein ; em: éminence médiane ; insr: récepteur à l’insuline ; lepr: récepteur à la leptine ; NPY: neuropeptide Y ; POMC: pro-opiomélanocortine ; Schéma modifié à partir de Niswender KD et coll., 2004.

- 34 -
Lorsque l’adiposité augmente, l’insulinémie et la leptinémie s’élèvent, et les
signaux d’adiposité arrivant au niveau du NA s’intensifient. Il en résulte une
inhibition des neurones à NPY/AgRP, corrélée à une diminution de l’expression
et/ou de la libération de ces deux neurotransmetteurs, ainsi qu’une activation des
neurones à POMC, corrélée à une augmentation de la libération d’α-MSH par ces
neurones. Ces mécanismes coordonnés aboutissent à une réduction de la prise
alimentaire. La situation inverse se déroule dans le cas d’une diminution de la
masse adipeuse (Morton GJ et coll., 2006).
3. Connexions intrahypothalamiques
Au fur et à mesure des expériences de colocalisations, de traçages rétro-
et/ou antérogrades, d’activations nerveuses, la cartographie des circuits
intrahypothalamiques impliqués dans le contrôle de l’homéostasie énergétique se
dessine de plus en plus précise.
Comme mentionné dans le paragraphe précédent, de part ses
caractéristiques, le NA est une région cérébrale clé pour capter en premier les
signaux périphériques en relation avec le métabolisme. L’activité de ces neurones
influence celle des neurones dits « de second ordre », se situant dans d’autres aires
hypothalamiques (Wynne K et coll., 2005) (Figure 11). Les terminaisons nerveuses
orexigènes et anorexigènes du NA s’étendent vers la plupart des régions
hypothalamiques comme les noyaux VMN, DMN, PVN, la région périfornicale, le LH
et le noyau suprachiasmatique (Kalra SP et coll., 1999 ; Bouret SG et Simerly RB,
2004 ; Yi CX et coll., 2006). Quelques projections de neurones à NPY du NA sont
courtes et locales, se terminant sur les neurones à POMC voisins. Lorsque les
premiers sont activés par de la leptine, du NPY et du GABA sont libérés. Ces deux
neurotransmetteurs exercent un effet général inhibiteur, il en résulte donc une
diminution de l’activité des neurones adjacents à POMC (Cowley MA et coll., 2001).
A la fois les neurones à NPY et ceux à POMC projettent vers le PVN. Grâce à
ses travaux d’immunohistochimie et d’électrophysiologie, Michael A. Cowley et ses
collaborateurs ont pu établir un modèle d’intégration dans le PVN de signaux en
provenance du NA. Au niveau du PVN, les neurones à NPY et à POMC projetteraient
sur la même population de neurones cibles, à savoir des neurones GABA-ergiques.
Ces derniers contrôleraient l’activité de neurones anorexigènes à ocytocine, CRH
(corticotropin-releasing hormone) ou TRH (thyrotropin-releasing hormone)
(Cowley MA et coll., 1999 ; Schwartz MW et coll., 2000) (Figure 12).

- 35 -
Figure 12 : Modèle d’intégration GABA-ergique dans le noyau hypothalamique paraventriculaire (PVN) des signaux neuropeptidergiques en provenance du noyau arqué (NA). 3V: troisième ventricule ; AGRP: agouti-related protein ; α-MSH: alpha-melanocyte-stimulating hormone ; GABA: γ-aminobutyric acid ; GABAAR: récepteur au GABA ; MC4-R: récepteur à mélanocortine ; NPY-R: récepteur au NPY ; NPY: neuropeptide Y ; POMC: pro-opiomélanocortine ; Schéma modifié à partir de Cowley MA et coll., 1999.
Les neurones à NPY et à POMC du NA projettent également vers la région
périfornicale ainsi que le LH. Leur activité influence de manière opposée celles des
neurones à MCH et orexines (Schwartz MW et coll., 2000). De plus, il existe des
connexions réciproques, les neurones à orexines du LH innervant et activant, les
cellules à NPY du NA.
Le VMN, qui constitue une région cérébrale cruciale dans la régulation de
l’homéostasie énergétique, reçoit aussi des projections nerveuses à NPY/AgRP et
POMC du NA (Guan XM et coll., 1998). A ce niveau est synthétisé le BDNF (Brain-
Derived Neurotrophic Factor) dont l’expression est modulée par le statut
énergétique de l’organisme ou par des agonistes aux mélanocortines, suggérant un
relais de l’information en provenance des neurones à POMC (Xu B et coll., 2003).
Des projections nerveuses du VMN aboutissent ensuite dans le DMN (Wynne K et
coll., 2005).

- 36 -
Les connexions hypothalamiques entre le VMN et le DMN sont réciproques.
Ce dernier reçoit aussi des terminaisons nerveuses à NPY/AgRP trouvant leur
origine dans le NA et des efférences en partent en direction du PVN. Ce noyau
hypothalamique possède quelques neurones à NPY (Kalra SP et coll., 1999).
Ainsi, il existe un réseau riche de connexions intrahypothalamiques
relativement complexe, illustré par la figure 13.
Figure 13 : Connexions nerveuses intrahypothalamiques, impliquées dans l’homéostasie énergétique. Les neuropeptides orexigènes sont en bleu et les anorexigènes en rouge. 3V: troisième ventricule ; AgRP: agouti-related protein ; CART: cocaine- and amphetamine-regulated transcript ; CRH: Corticotropin-Releasing Hormone ; DMN: noyau hypothalamique dorsomédian ; em: éminence médiane ; fx: fornix ; LH: hypothalamus latéral ; MCH: melanin-concentrating hormone ; NA: noyau arqué ; NPY: neuropeptide Y ; NTS: noyau du tractus solitaire ; POMC: pro-opiomélanocortine ; PVN: noyau hypothalamique paraventriculaire ; TRH: thyrotropin-releasing hormone ; VMN: noyau hypothalamique ventromédian.

- 37 -
4. Connexions extrahypothalamiques en direction du tronc cérébral
L’hypothalamus est en relation avec d’autres grandes régions cérébrales,
indirectement liées à l’homéostasie énergétique, responsables des choix
alimentaires faisant référence à la mémoire ou à des phénomènes cognitifs,
gustatifs et/ou olfactifs. Il existe même des voies dites « de récompense »,
indépendantes du statut calorique, stimulant la prise alimentaire en dépit d’une
balance positive. Au contraire, d’autres aires y sont directement liées. Ce sont celles
du tronc cérébral qui permettent le relais de l’information nerveuse vers la
périphérie, refermant ainsi la boucle de rétrocontrôle, positif ou négatif, selon le
contexte énergétique de l’individu.
En ce qui concerne le tronc cérébral, il existe d’importantes connexions
réciproques entre l’hypothalamus et cette partie essentielle du SNC, en particulier
avec le NTS (Figure 13). Ce dernier reçoit les afférences du SNA parasympathique
provenant de la périphérie, dont le tractus gastro-intestinal, et se situe juste au
dessus du noyau dorsal moteur du vague (DMV), point de départ des neurones
préganglionnaires parasympathiques. De plus, tout comme le NA, le NTS est très
proche d’un organe circumventriculaire dépourvu de BHE, l’area postrema, où les
capillaires sont fenestrés. Cette caractéristique anatomique, le rend ainsi capable de
détecter lui-même des variations des concentrations plasmatiques des hormones
clés, comme l’insuline ou la leptine, d’autant plus que leurs récepteurs sont
présents. De manière intéressante, le NTS possède des neurones à POMC et
d’autres à NPY, ces derniers projetant vers le PVN. Enfin, il y a été révélée la
présence de récepteurs à mélanocortines (Wynne K et coll., 2005 ; Schwartz MW et
coll., 2000 ; Morton GJ et coll., 2006).
Par ailleurs, le VMN et le LH sont aussi reliés au NTS (Wynne K et coll.,
2005 ; Kirchgessner AL, 2002). La stimulation des neurones à orexines du LH
engendre l’activation de neurones du NTS et du DMV qui contiennent des
récepteurs pour ces neuropeptides. L’activité électrique des efférences vagales est
ainsi accrue, suggérant finalement la modulation des sécrétions et/ou de la
mobilité gastro-intestinales, en réponse à une activation des voies de transduction
NPY/AgRP et orexines/MCH. Le phénomène inverse est possible puisque des
neurones du NTS projettent vers le LH : des signaux de satiété en provenance de la
périphérie, par exemple de l’intestin ou du foie, moduleraient l’activité électrique
des neurones à orexines (Kirchgessner AL, 2002).

- 38 -
L’hypothalamus est donc le siège des contrôles de l’homéostasie énergique,
notamment grâce au NA capable d’intégrer les différents signaux hormonaux que
sont la leptinémie et l’insulinémie, et de relayer par différentes voies de
transduction les informations nerveuses qui aboutiront à un rétrocontrôle en
direction de la périphérie (Figure 14).
Figure 14 : Illustration de la boucle de contrôle existant entre les organes périphériques et le cerveau. (http://www-good.pasteur-lille.fr/accueil/presse/html/campagne-obe/Les_origines_ biologiques.htm)
Dans la prochaine partie, nous allons nous intéresser aux mécanismes de
sensibilité cérébrale aux hormones (leptine, insuline), mais aussi aux métabolites
(acides gras, glucose).

- 39 -
III – Sensibilité hypothalamique aux informations métaboliques
Ne sera traitée dans ce chapitre, que la sensibilité cérébrale directe, c'est-à-
dire par les cellules nerveuses, excluant ainsi la réception des informations
périphériques de nature métabolique relayées par le SNA. Ce mécanisme est
possible grâce à la circulation sanguine qui véhicule, de la périphérie vers le SNC,
des molécules liées au métabolisme telles que les hormones comme la leptine ou
l’insuline, et les métabolites comme les acides gras ou le glucose. Ils sont capables
d’accéder directement aux cellules cérébrales et, une fois détectés, sont à l’origine
de cascades de signalisation aboutissant à une information nerveuse. Ces composés
deviennent alors des signaux au-delà de leur rôle purement métabolique, nécessaire
au fonctionnement normal des cellules.
1. Sensibilité aux hormones
Comme nous l’avons vu précédemment la régulation endocrinienne est
primordiale pour le maintien de l’homéostasie énergétique. Or au niveau cérébral et
plus particulièrement de l’hypothalamus, sont exprimés des récepteurs spécifiques
aux hormones impliquées dans ce contrôle comme la leptine ou l’insuline. Ceux-ci
permettent la détection de leurs concentrations circulantes qui sont
proportionnelles à la masse adipeuse et donc liées à l’état énergétique de l’individu,
permettant un rétrocontrôle accordant la prise alimentaire à ces stockages d’énergie
(Figlewicz DP, 2003) (cf. § I-2.1, Figure 5).
De plus, il s’avère qu’il existe un dialogue entre ces deux signaux d’adiposité,
grâce à un croisement de leurs cascades de signalisation au niveau de la PI3K
(Phosphatidylinositol 3-Kinase) (Niswender KD et coll., 2004) (Figure 15). En se
liant à son hormone, le récepteur à l’insuline s’autophosphoryle au niveau de son
domaine intracellulaire grâce à son activité tyrosine kinase. Les protéines IRS
adhèrent alors au niveau des résidus phosphorylés, sont activées par
phosphorylation de leurs propres résidus tyrosines et de ce fait, deviennent
capables d’interagir avec la PI3K pour la stimuler. Cette enzyme catalyse la
phosphorylation du phosphatidylinositol (4,5)-biphosphate (PIP2) en
phosphatidylinositol (3,4,5)-triphosphate (PIP3), ce dernier permettant l’activation
d’enzymes telles que la PDK1 (3-Phosphoinositide-Dependent Kinase-1) et la PKB
(Protéine Kinase B). La leptine, quant à elle, en se liant à son récepteur, provoque
sa dimérisation et le recrutement de kinases JAK2, indispensables à la

- 40 -
phosphorylation de son domaine intracellulaire. Ses sites phosphorylés permettent
l’adhésion des molécules STAT3 (Signal Transducer and Activator of Transcription-
3), alors elles-mêmes phosphorylées et activées par les JAK2, aboutissant à leur
homodimérisation essentielle pour leur fonction de régulation de la transcription de
gènes cibles. Tout comme l’insuline, la leptine serait capable d’activer la PI3K par le
biais de JAK2 qui pourrait catalyser la phosphorylation des IRS. Ce mécanisme
existerait au sein de l’hypothalamus. En particulier, il a été montré que ces deux
hormones activaient les canaux potassiques ATP-dépendants via la stimulation de
la PI3K dans certains neurones. Ce phénomène conduirait ainsi à une
hyperpolarisation et donc à une inhibition de leur activité électrique (Obici S et coll.,
2002a) (Figure 18).
Figure 15 : Dialogue entre les voies de signalisation générées par la liaison de l’insuline et de la leptine à leur récepteur. Il se réalise au niveau de la phosphatidylinositol 3-kinase (PI3K) composée des 2 sous-unités p110 et p85. IRS: insulin receptor substrate ; JAK2: Janus-activated kinase-2 ; PDK1: 3-phosphoinositide-dependent kinase-1 ; PIP2: phosphatidylinositol (4,5)-bisphosphate ; PIP3: phosphatidylinositol (3,4,5)-triphosphate ; PKB: protéine kinase B ; PKC: protéine kinase C ; Socs3: suppressor of cytokine signaling -3 ; Stat3: signal transducer and activator of transcription-3 ; Schéma modifié à partir de Niswender KD et coll., 2004.
En plus de la PI3K, le dialogue leptine/insuline s’établirait également au
travers d’une autre enzyme, l’AMPK (AMP-activated protein Kinase) (Kahn BB et
coll., 2005). L’activité de cette enzyme est liée à la disponibilité cellulaire en
substrats métaboliques puisqu’elle est activée allostériquement par une
augmentation du rapport AMP/ATP. D’une manière générale, son activation conduit

- 41 -
à une inhibition des mécanismes anaboliques consommateurs d’ATP et à une
stimulation des voies cataboliques productrices d’ATP, par le biais de la régulation
de l’expression ou de l’activité d’enzymes clé du métabolisme des acides gras, du
cholestérol et du glucose (Xue B et Kahn BB, 2006). En plus de ce rôle exercé en
périphérie, l’AMPK contrôle la prise alimentaire et le poids corporel en étant
impliquée, de façon opposée, dans la transduction de signaux orexigènes (ghréline,
épisode de jeûne, hypoglycémie) et anorexigènes (leptine, insuline, hyperglycémie)
au niveau hypothalamique (Figure 16).
Figure 16 : Régulation de l’activité de l’AMPK (AMP-activated protein kinase) du NA (noyau arqué) et du PVN (noyau hypothalamique paraventriculaire) par les signaux hormonaux et le statut énergétique de l’organisme. Modifié à partir de Kahn BB et coll., 2005.
1.1. Signalisation cérébrale de la leptine
Dans le paragraphe I-2.1, nous avons vu que l’action majeure de la leptine se
déroulait dans le SNC via Ob-Rb, l’isoforme longue du récepteur. L’Ob-Rb est
présent un peu partout dans le SNC, mais de manière plus spécifique dans les
plexus choroïdiens, le cortex cérébral, l’hippocampe, le thalamus, l’hypothalamus et
le complexe dorsal vagal (Elmquist JK et coll., 1998 ; Funahashi et coll., 2003).
Dans ce dernier, les plus fortes concentrations sont localisées dans les noyaux
supraoptiques et périventriculaires, le PVN, le NA et le LH (Hakansson ML et coll.,
1998). Différentes études montrent que l’injection de leptine dans le SNC est plus
efficace que l’injection périphérique pour réduire la prise alimentaire et le poids
corporel. Au niveau de l’hypothalamus, ce sont les injections réalisées directement

- 42 -
dans le NA ou le VMN qui aboutissent aux effets optimaux. Enfin, si le gène codant
pour Ob-Rb est invalidé spécifiquement dans le SNC, les animaux deviennent
obèses (Sahu A, 2004).
Le contrôle de la leptine sur l’homéostasie énergétique passe
vraisemblablement par la modulation de l’expression des neuropeptides orexigènes
et anorexigènes décrits dans les chapitres précédents (Morton GJ et coll., 2006).
Cette régulation peut s’opérer directement puisque l’Ob-R est exprimé dans la
plupart de ces neurones : Ob-R colocalise avec la POMC et le NPY dans le NA, avec
la MCH et l’orexine dans le LH (Hakansson ML et coll., 1998 ; Kirchgessner AL,
2002 ; Figure 11).
Figure 17 : Régulation de l’expression des neuropeptides par la leptine et l’insuline. AgRP: agouti-related protein ; α-MSH: α-melanocyte-stimulating hormone ; MCR: récepteur aux mélanocortines. NPY: neuropeptide Y ; POMC: pro-opiomélanocortine. D’après Schwartz MW et coll., 2000.
Des travaux montrent qu’Ob-Rb, suite à la liaison de la leptine, stimule
l’expression du gène de la POMC et active, par dépolarisation, les neurones
exprimant cette prohormone (Cowley MA et coll., 2001). Ceci conduit à la libération
d’α-MSH inhibant la prise alimentaire par l’activation des récepteurs aux

- 43 -
mélanocortines. A l’inverse, la signalisation d’Ob-Rb inhibe l’expression du NPY et
de l’AgRP, neuropeptides orexigènes. Ces deux mécanismes simultanés et
coordonnés aboutissent finalement à une perte d’appétit et donc à une diminution
des apports énergétiques (Münzberg H et coll., 2005) (Figure 17). Enfin, l’injection
de leptine diminue la synthèse des ARNm de la MCH et de l’orexine A dans le LH
(Williams G et coll., 2004 ; Kirchgessner AL, 2002).
Les modèles animaux déficients en leptine (souris ob/ob) ou en récepteur
(souris db/db, rats Zucker fa/fa) confirment les données obtenues
expérimentalement. En effet, chez ces rongeurs, en condition basale, les neurones à
NPY du NA sont suractivés, et l’expression de l’AgRP est augmentée (Williams G et
coll., 2004 ; Stütz AM et coll., 2005). Ceci démontre que la leptine est un important
régulateur des neurones à NPY/AgRP et que l’inhibition de la prise alimentaire par
cette hormone se ferait via ces cellules. Cette hypothèse est renforcée par le fait que
la leptine est capable de les hyperpolariser et donc de diminuer leur activité
électrique. Par ailleurs, les rats fa/fa ou les souris ob/ob surexpriment la MCH par
rapport aux rats Zucker Fa/Fa ou aux souris à phénotype mince. Ainsi la leptine
inhiberait également cette population neuronale (Williams G et coll., 2004). Enfin, la
concentration en ARNm de la POMC est significativement plus basse chez ces
animaux obèses (Pritchard LE et coll., 2002).
1.2. Signalisation cérébrale de l’insuline
L’insuline est la première hormone dont l’implication dans le contrôle
nerveux du poids corporel a été démontrée (Schwartz MW et coll., 2000). Le
récepteur à l’insuline est exprimé un peu partout dans le SNC, avec de plus fortes
densités dans le tronc cérébral, le cervelet, le cortex, l’hippocampe et
l’hypothalamus. Dans ce dernier, il se concentre particulièrement dans le NA et de
manière plus faible dans les DMN, VMN et dans le LH (Schwartz MW et coll., 1992).
Par son action centrale, l’insuline a des effets similaires à ceux de la leptine,
impliquant probablement des mécanismes proches : elle inhibe la prise alimentaire
et augmente les dépenses énergétiques (Figure 17). Au niveau du PVN, elle diminue
la synthèse et la libération du NPY. Toutefois, il n’a pas été établi de manière
définitive que les neurones à NPY exprimaient le récepteur à l’insuline (Williams G
et coll., 2004 ; Stütz AM et coll., 2005). D’autre part, l’injection intrapéritonéale
d’insuline à des rats augmente l’expression des ARNm de la POMC dans le NA
(Pritchard LE et coll., 2002). Enfin, une diminution de 80 % de la quantité de
récepteurs à l’insuline spécifiquement dans le NA, par injection d’antisens, entraîne

- 44 -
une hyperphagie accompagnée d’une augmentation de la masse adipeuse et une
diminution de la production hépatique de glucose (Obici S et coll., 2002b).
Ainsi, l’hyperinsulinémie faisant suite à l’ingestion d’aliments induit un
rétrocontrôle négatif en partie de nature centrale afin d’empêcher un éventuel
apport d’énergie excédentaire (Figure 7).
2. Sensibilité aux métabolites
Les métabolites, dérivant de la prise alimentaire suite aux processus de
digestion et donc au métabolisme, sont des éléments essentiels à l’intégrité d’un
organisme. Ils sont classés en trois groupes : les protéines, les lipides et les
glucides. Au sein du SNC, leurs rôles consistent au maintien des structures et à
l’apport d’énergie. La respiration cellulaire est assurée essentiellement par les
lipides et glucides, les protéines ne devenant source d’énergie principale que
lorsque les autres molécules sont indisponibles.
Dans les années 50, on soupçonna l’implication des lipides et des glucides
dans d’autres processus physiologiques, donnant naissance aux théories
lipostatique et glucostatique. La première met en avant la fonction de stockage
d’énergie assurée par les adipocytes, tandis que la seconde concerne le rôle
primordial joué par le glucose dans l’ensemble du métabolisme énergétique des
cellules (Kennedy GC, 1953 ; Mayer J, 1953). Ainsi, le fait que ces métabolites
puissent eux-mêmes contrôler la balance énergétique commençait à faire son
chemin dans les esprits.
Ceci implique que ces composés soient capables de générer des cascades de
signalisation au sein de certaines cellules, en plus de leur action indirecte via la
sécrétion d’hormone. Précédemment nous avons vu qu’il existait un croisement des
voies de signalisation de la leptine et de l’insuline (Figures 15 et 16). Or, une
interaction a également été mise en évidence entre le glucose et les acides gras
grâce aux expériences de Philip Randle (Randle PJ et coll., 1963). Cette équipe de
chercheurs a montré que, sur le cœur de rat isolé, une augmentation de la
concentration des acides gras libres (AGL) dans le milieu de perfusion réduisait le
transport et l’utilisation du glucose par le myocarde. Ce concept d’interrelation
entre le métabolisme du glucose et celui des acides gras a été depuis étendu au foie
et aux cellules β du pancréas (voir § ci-dessous pour ces dernières) (Girard J, 2003).
Les molécules impliquées sont entre autres l’ATP et l’acétyl-CoA, cofacteurs générés

- 45 -
lors de l’oxydation des AGL et nécessaires à des étapes de la néoglucogenèse
hépatique.
2.1. Les acides gras
Parmi les lipides, ce sont les acides gras libres (AGL) qui tiennent le rôle de
molécule de signalisation, que ce soit en périphérie ou au niveau du SNC. Par
exemple, en plus d’être des substrats énergétiques majeurs, les AGL influencent la
sécrétion d’insuline par les cellules β du pancréas (Girard J, 2003). A court terme
(exposition aux AGL durant quelques heures), ils potentialisent cette libération
hormonale en réponse au glucose, alors qu’à long terme (plusieurs jours), ils
l’inhibent. Le premier mécanisme impliquerait l’augmentation de la concentration
intracellulaire d’acyl-CoA découlant de la dégradation normale des AGL. Or, cette
molécule a la capacité de stimuler directement l’exocytose, mais aussi
indirectement, via la formation de diacylglycérol, un activateur de la PKC. Quant à
l’effet inhibiteur, plusieurs hypothèses ont été formulées : inhibition de la fermeture
des canaux potassiques ATP-dépendants par les acyl-CoA, et/ou diminution de
l’expression de protéines comme le GLUT2 et la GK, et/ou effet découplant des AGL
sur la mitochondrie aboutissant à une diminution de la synthèse d’ATP (Figure 6).
En ce qui concerne la sensibilité centrale aux AGL, Loftus et ses
collaborateurs ont mis en évidence l’existence d’une relation entre lipides et
régulation de l’homéostasie énergétique (Loftus TM et coll., 2000). En effet,
l’inhibition pharmacologique de la synthèse d’acides gras à longues chaînes par le
C75 (inhibiteur de la Fatty Acid Synthase, FAS), au niveau cérébral, entraîne une
diminution de la prise alimentaire, accompagnée d’une perte de poids. De manière
intéressante, ce phénotype est corrélé à une diminution de l’expression du NPY
dans l’hypothalamus suggérant l’implication de ces neurones. Cet effet anorexigène
s’expliquerait par 1) l’accumulation de malonyl-CoA suite à l’inhibition de la FAS,
2) l’inhibition de l’entrée des acyl-CoA dans la mitochondrie due à l’excès de
malonyl-CoA et 3) l’accumulation cytosolique, secondaire, d’acyl-CoA. Cette
hypothèse selon laquelle l’accumulation d’AGL et plus particulièrement d’acyl-CoA
dans la cellule serait responsable de l’effet satiétogène des lipides a, depuis, été
étayée par différentes stratégies expérimentales (injection intracérébroventriculaire
d’AGL, injection d’un inhibiteur du transporteur aux acides gras mitochondrial).
Ces études révèlent, en plus de la diminution de la prise alimentaire, une
diminution de la production hépatique de glucose et suggèrent l’implication des

- 46 -
canaux potassiques ATP-dépendants (Obici S et coll., 2002a ; Cruciani-
Guglielmacci et coll., 2004). La figure 18 illustre cette hypothèse.
Figure 18 : Suggestion de mécanisme de régulation des canaux potassiques ATP-dépendants par la leptine, l’insuline et l’acide oléique. PI3K: phosphatidylinositol 3-kinase ; SU: sulfonylurée ; SUR: sous-unité du canal potassique (SU inhibe le canal par liaison à son récepteur SUR). D’après Obici S et coll., 2002a.
Par ailleurs, l’infusion pendant 2 jours d’une solution de triglycérides dans le
SNC, potentialise la sécrétion d’insuline en réponse à une hyperglycémie, indiquant
que l’action des AGL sur les cellules β mentionnée au début de ce paragraphe,
pourrait passer au moins en partie par une détection cérébrale (Clément L et coll.,
2002). De plus, dans ce cas aussi apparaît une désensibilisation hépatique à
l’insuline, limitant ainsi la libération de glucose dans la circulation sanguine.
Plus récemment, des travaux de recherche ont permis de mettre en évidence
de manière encore plus directe l’activation cérébrale induite par les lipides. En
particulier, l’étude de cette activation par l’utilisation de l’expression du proto-
oncogène c-Fos (marqueur de l’activation des neurones, cf. § IV-2) suggère que
l’infusion d’AGL par voie intracarotidienne vers le SNC inhibe les noyaux DMN,
VMN, PVN et NA, mais stimule le LH (Cruciani-Guglielmacci C et coll., 2004). Par
ailleurs, des données d’électrophysiologie indiquent clairement qu’il existe au sein
du NA une population de neurones sensibles aux AGL : ils augmentent ou
diminuent leur activité en réponse à l’application d’acide oléique (Wang R et coll.,
2004). Cette réponse est donc différente selon la cellule enregistrée, mais aussi
selon la concentration en glucose environnant. Cela renforce l’idée d’interrelation
entre les voies mises en jeu par le glucose et les AGL, déjà mentionnée dans le cas
des organes périphériques, mais cette fois-ci dans l’hypothalamus. Enfin, au sein
de notre laboratoire, il a été mis en évidence que l’injection intracérébroventriculaire
aiguë de triglycérides conduisait à la production rapide et transitoire d’espèces
libres de l’oxygène de nature mitochondriale et ce, au sein de l’hypothalamus

- 47 -
ventral (Benani A et coll., 2007). Or, cette synthèse est nécessaire au mécanisme
d’inhibition de la prise alimentaire normalement observée dans un contexte
d’hypertriglycéridémie, mettant en lumière un des éléments indispensables de la
cascade de signalisation.
Finalement, le lien entre la sensibilité hypothalamique aux AGL et la mise en
place des mécanismes régulateurs périphériques (désensibilisation hépatique à
l’insuline) s’établirait grâce au SNA. En l’occurrence, il a été démontré que le
système orthosympathique intervenait via les adrénorécepteurs α2A (Magnan C et
coll., 1999). Afin d’éclaircir ce point, il serait nécessaire d’entreprendre des études
plus approfondies, notamment sur le tronc cérébral.
Ainsi, les lipides circulants interviennent bien dans la régulation de
l’homéostasie énergétique et ce, grâce à une détection cérébrale de leur
concentration (Figure 19).
Figure 19 : Effets des acides gras libres (AGL) indépendants des hormones. NPY: neuropeptide Y ; SNA: système nerveux autonome

- 48 -
Les lipides ne sont pas les seuls à pouvoir intervenir dans le contrôle nerveux
de l’homéostasie énergétique. Le glucose, également étudié dans le cadre de la
théorie glucostatique, fera l’objet des prochains paragraphes.
2.2. Le glucose
2.2.1. Généralités sur l’homéostasie glucidique
Ce monosaccharide circule librement dans le sang à une concentration
d’environ 5 mM soit 1 g/L. Chez les individus sains, la glycémie est maintenue de
manière stricte autour de cette valeur, grâce à des mécanismes de régulation. En
particulier, la production de glucose endogène s’adapte aux besoins énergétiques
des tissus de l’organisme, mais aussi aux apports exogènes provenant de
l’alimentation. Lorsque ceux-ci sont en excès, se mettent en place des processus de
stockage sous forme de glycogène, au niveau hépatique ou musculaire notamment,
et de triglycérides, au niveau du tissu adipeux.
Les monosaccharides tels que le glucose ou le fructose sont directement
assimilables au niveau de la muqueuse digestive. Néanmoins, l’alimentation
quotidienne n’en apporte qu’une faible quantité puisqu’elle est composée
essentiellement de glucides plus ou moins complexes nécessitant une hydrolyse par
différentes enzymes du système digestif. Le glucose est absorbé par les entérocytes
de l’intestin grâce à un transport actif assuré par les SGLT-1 (sodium-glucose
cotransporteur). Du côté basal des cellules, l’hexose est libéré dans la circulation
sanguine grâce à un transport facilité assuré par GLUT2. Les capillaires intestinaux
convergeant en direction de la veine porte hépatique, une partie du glucose est
stockée à ce niveau sous forme de glycogène dans le foie et une autre redistribuée
vers les autres organes (Burcelin R et Pénicaud L, 2003).
Suite à un repas, il apparaît une hyperglycémie dont l’amplitude est
influencée entre autres par la quantité et la nature des glucides ingérés, par les
processus digestifs et par les facteurs hormonaux (Paquot N et coll., 2003). Le
déclenchement des mécanismes de régulation implique une détection préalable des
concentrations de glucose. Les systèmes de sensibilité à ce métabolite existent à
plusieurs endroits stratégiques de l’organisme (Figure 20).
Dans un premier temps, le passage des glucides dans le système gastro-
intestinal induit la libération d’hormones. Il s’agit entre autres de la CCK, du PYY et
du GLP-1, qui possèdent un effet anorexigène de nature cérébrale (Figure 3).

- 49 -
Plus en aval, il existe un système sensible à l’hyperglycémie au niveau de la
veine hépatoportale (Burcelin R et Pénicaud L, 2003). Ce détecteur de glucose, dont
l’organisation cellulaire est encore mal connue, relaye l’information
« hyperglycémie » aux fibres afférentes de la branche hépatique du nerf vague
(Marty N et coll., 2005). Cette information est ainsi acheminée jusqu’au tronc
cérébral au niveau du NTS. Son activation par l’hyperglycémie hépatoportale
entraîne la modulation adéquate de divers processus du métabolisme énergétique :
inhibition de la prise alimentaire, stimulation de l’utilisation du glucose par les
muscles et le tissu adipeux, stimulation du stockage de l’hexose au niveau
hépatique, inhibition de la production des hormones de contre-régulation
(catécholamines, glucagon) (Russek M, 1970 ; Burcelin R et coll., 2000 ; Donovan
CM et coll., 1994).
Figure 20 : Mécanismes de régulation mis en place suite à l’hyperglycémie post-prandiale. Les glucides sont détectés au niveau de l’intestin, et le glucose au niveau de la veine hépatoportale et des cellules β du pancréas. β: cellules β du pancréas ; SNA: système nerveux autonome
Les mécanismes cellulaires découlant de la sensibilité au glucose les mieux
décrits sont ceux déclenchés au niveau des cellules β (cf. § I-2.2, figure 6).
Schématiquement, cette sensibilité est générée par la glycolyse qui débute par la
phosphorylation de l’hexose, de manière proportionnelle à sa concentration, étape

- 50 -
assurée par la GK. La libération d’insuline est déclenchée par la détection directe
du glucose circulant et potentialisée par le GLP-1 libéré par la muqueuse
intestinale.
En plus des cellules β sécrétrices d’insuline, hormone anabolique, le
pancréas possède des cellules α sécrétrices de glucagon, hormone catabolique
entrant en jeu dans le cas d’une hypoglycémie. Son action permet de protéger
l’organisme contre un déficit énergétique pouvant être dangereux, en particulier
pour le SNC. Sa cible essentielle est le foie, au niveau duquel il stimule la
glycogénolyse et la gluconéogenèse afin de libérer du glucose dans la circulation
sanguine. La libération de glucagon est contrôlée par différents mécanismes (Marty
N et coll., 2005). Premièrement, l’insuline contenue dans les îlots de Langerhans la
régule, établissant ainsi un rétrocontrôle négatif levé lors d’une hypoglycémie.
Deuxièmement, la sécrétion de glucagon est sous l’influence du SNA sympathique
dont l’activité est modulée par l’hypoglycémie via une détection préalable du glucose
au niveau de la veine hépatoportale et du SNC (hypothalamus/tronc cérébral).
Enfin, il est probable que les cellules α soient directement sensibles au glucose.
Elles sont dotées, en effet, d’une GK et d’un canal potassique ATP-dépendant
(constitué des sous-unités Kir6.2 et SUR1) comme les cellules β. Néanmoins, ce
dernier mécanisme prête encore à controverse.
Avec les catécholamines, le glucagon fait partie des hormones dites de la
« contre-régulation ». De manière très simplifiée, ces hormones sont libérées lors
d’épisodes hypoglycémiques afin de contrer la diminution de la glycémie. Elles ont
donc des effets opposés à ceux de l’insuline.
2.2.2. Une voie annexe suivie par le glucose : la voie des hexosamines
Tout le glucose disponible dans une cellule n’emprunte pas la voie de la
glycolyse : une petite partie suit la voie des hexosamines. Elle a été découverte il y a
moins de dix ans et semble prometteuse en ce qui concerne la compréhension du
contrôle de l’homéostasie énergétique. Il semble qu’elle soit, au moins en partie,
responsable de la résistance à l’insuline observée chez les diabétiques, puisque
celle-ci est corrélée à une augmentation de l’activité de cette cascade enzymatique
(Buse MG, 2006). Toutefois, il n’a jamais été mis en évidence une relation directe
entre ces deux observations.
Physiologiquement, le glucose entre dans la cellule et est immédiatement
phosphorylé en glucose-6-phosphate par une hexokinase (la GK dans les cellules β),
puis transformé en fructose-6-phosphate (Figure 21). La majeure partie emprunte

- 51 -
alors la voie de la glycolyse, tandis qu’une infime partie (1 à 3%) est prise en charge
par la glutamine fructose-6-phosphate aminotransférase (GFAT) pour donner la
glucosamine-6-phosphate, ce qui constitue la première étape (limitante) de la voie
des hexosamines (Obici S et Rossetti L, 2003).
Figure 21 : Voie des hexosamines. AGL: acides gras libres ; Fruc-6-P: fructose-6-phosphate ; GFAT: glutamine fructose-6-phosphate aminotransférase ; GK: glucokinase ; Glc-6-P: glucose-6-phosphate ; GlcN-6-P: glucosamine-6-phosphate ; GlcNAc: N-acétylglucosamine ; GLUT: transporteur au glucose ; GS: glycogène synthase ; IR: récepteur à l’insuline ; IRS: insulin receptor substrate. D’après Wells L et coll., 2003.
Cette chaîne aboutit à la synthèse d’UDP-N-acétylglucosamine qui est son
produit final, quantifiable, reflétant son niveau d’activation. Il exerce un
rétrocontrôle négatif sur la GFAT, limitant par la même l’entrée de glucose, et
permet la O-glucosamine-N-acétylation de protéines cibles, notamment de facteurs
de transcription (c-myc) ou d’enzymes (glycogène synthase), et de manière
intéressante d’IRS-1 et probablement de GLUT4 (Buse MG, 2006). Ce processus
touchant à ce transporteur au glucose, serait responsable de la résistance à
l’insuline, en favorisant sa translocation à la membrane. Par ailleurs, l’UDP-N-
acétylglucosamine permet aussi la glycosylation de certaines protéines, comme le
transporteur au glucose astrocytaire, GLUT1. Différents niveaux de glycosylation de

- 52 -
ce transporteur pourraient alors être responsables du contrôle de ses mouvements
et des variations de son affinité (Medina RA et Owen GI, 2002). Ainsi la voie des
hexosamines engendrerait la régulation de protéines intervenant dans l’homéostasie
énergétique, en particulier des GLUTs, ce qui permettrait une influence directe sur
l’entrée du glucose dans la cellule et donc sur la sensibilité à cet hexose.
L’UDP-N-acétylglucosamine représente un « carrefour » de la sensibilité des
nutriments, puisque sa concentration intracellulaire est influencée par l’état
nutritionnel de l’organisme (Obici S et Rossetti L, 2003). Elle est nécessaire et/ou
sensible au métabolisme de l’ADP/ATP (« haute énergie »), du glucose, des acides
aminés, des acides gras et des nucléotides comme le montre sa composition
chimique (Wells L et coll., 2003) (Figure 22).
Figure 22 : la voie des hexosamines au carrefour des métabolismes des différents types de nutriments, au travers de l’UDP-N-acétylglucosamine (UDP-GlcNAc). D’après Wells L et coll., 2003.
Il est probable que la voie des hexosamines soit présente au sein du SNC,
pourtant, à ce jour, aucune donnée de la littérature ne le montre clairement.
2.2.3. L’hypothalamus, siège de la sensibilité cérébrale au glucose
Le glucose est la principale source d’énergie du SNC et sa concentration au
sein de cet organe est maintenue constante de façon drastique, sa pénurie pouvant
avoir de graves conséquences. Il traverse la BHE et pénètre dans les cellules grâce
aux transporteurs de glucose, GLUT1, localisé à la membrane des cellules
endothéliales et des astrocytes, et GLUT3, localisé à la membrane des neurones.

- 53 -
Au-delà du rôle énergétique basique, cet hexose joue un rôle dans la
sensibilité aux nutriments comme nous l’avons déjà abordé, c'est-à-dire que sa
concentration sanguine renseigne à tout moment le SNC sur le statut énergétique
d’un organisme. De manière logique, ce sont les régions cérébrales impliquées dans
l’homéostasie énergétique, l’hypothalamus et le tronc cérébral avec leur équipement
moléculaire particulier (cf. § II), qui sont capables de détecter et de répondre aux
variations de glucose. Une augmentation de sa concentration au sein de ces
structures cérébrales, est le reflet d’une balance énergétique positive, et provoquera
la mise en place de processus visant à freiner les apports et à favoriser les
dépenses. A l’opposé, une diminution de cette concentration fait suite à une
hypoglycémie générale, et il s’en suivra des mécanismes inverses, c'est-à-dire des
apports favorisés et des dépenses limitées. Cette détection va permettre le
déclenchement rapide et coordonné de mécanismes de régulation (sécrétion
pancréatique, contrôle du métabolisme glucidique hépatique, etc.) (Figure 23).
Figure 23 : Détection des variations glycémiques par le système nerveux central (SNC). β: cellules β du pancréas ; DMN: noyau hypothalamique dorsomédian ; LH: hypothalamus latéral ; NA: noyau arqué ; PVN: noyau hypothalamique paraventriculaire ; VMN: noyau hypothalamique ventromédian ; SNA: système nerveux autonome

- 54 -
Une nouvelle fois, le NA occupe une place clé dans ce processus de détection,
en raison de la BHE adjacente relâchée. De ce fait, le glucose peut la traverser et
atteindre plus librement l’hypothalamus. Il semble arriver directement au sein du
VMN puisque l’injection intrapéritonéale de glucose radioactif aboutit à des
quantités élevées de radioactivité dans ce noyau. De même, l’administration
d’aurothioglucose, une forme toxique de l’hexose, provoque une vaste lésion de cette
structure (King BM, 2006). De plus, il est vraisemblable qu’au moins au niveau du
NA, les concentrations extracellulaires en glucose avoisinent les valeurs de la
glycémie, ce qui n’est pas le cas dans des structures plus profondes telles que le
striatum ou l’hippocampe (McNay EC et Gold PE, 1999).
La sensibilité de l’hypothalamus au glucose peut être observée indirectement
grâce à l’activité du SNA. Par exemple, il a été démontré que l’injection de glucose
par la carotide induisait la stimulation des nerfs sympathiques innervant le tissu
adipeux brun chez le rat. Or, elle est diminuée de 60 % lorsque le VMN est
préalablement lésé (Sakaguchi T et Bray GA, 1990). De même, au sein de notre
laboratoire, a été mise au point une injection similaire de glucose vers le cerveau, à
une dose ne modifiant pas la glycémie périphérique (9 mg/kg). Ce bolus de glucose
provoque une augmentation très rapide et transitoire de l’insulinémie mettant en
jeu l’activation du SNA parasympathique (Atef N et coll., 1995 ; Alquier T et coll.,
2003). Ce protocole révèle donc la capacité du SNC à détecter une variation de la
glycémie. La rapidité de la réponse insulinémique (1 minute) ne peut guère que
laisser suggérer l’implication de l’hypothalamus et/ou du tronc cérébral dans cette
détection, en raison de leurs relations privilégiées avec le compartiment sanguin
(absence de BHE véritable).
D’autres travaux ont cherché à mettre en évidence, de manière plus directe,
la sensibilité globale au glucose de l’hypothalamus, grâce à l’utilisation du proto-
oncogène c-fos caractéristique des cellules nerveuses activées (principe de la
technique décrit au § IV-2). Ainsi, la perfusion de glucose par la carotide pendant 1
heure vers le SNC de rats stimule les VMN, DMN et PVN (Dunn-Meynell AA et coll.,
1997). L’injection intrapéritonéale simultanée de glucose et d’insuline provoque une
augmentation de l’expression de c-Fos dans les VMN, PVN, NA et le LH (Carrasco M
et coll., 2001).
A l’inverse, l’administration de 2-déoxy-D-glucose (2-DG), un analogue du
glucose non métabolisé induisant une glucopénie cellulaire, est suivie de l’activation

- 55 -
des PVN, NA, du noyau périventriculaire et de l’aire hypothalamique antérieure
(Briski KP, 1998).
2.2.4. Acteurs cellulaires impliqués dans la détection du glucose
Il existe deux grandes populations de cellules nerveuses : les neurones,
responsables de l’activité électrique du SNC, et les cellules de soutien appelées
cellules gliales, dix à cinquante fois plus nombreuses (Campbell NA, 1995). Parmi
ces dernières, ce sont les astrocytes qui nous intéressent, car il semblerait qu’ils
puissent être impliqués dans la sensibilité du glucose par le cerveau. Ils entourent,
entre autre, les vaisseaux sanguins, participant ainsi à la BHE, et ont un rôle dans
l’apport d’énergie aux neurones.
Les neurones
C’est en 1964 qu’a été établie la preuve de l’existence de neurones sensibles
au glucose dans l’hypothalamus (Anand BK et coll., 1964). Depuis, leur
caractérisation s’est développée simultanément aux techniques, notamment grâce
au patch-clamp qui consiste en l’enregistrement direct de l’activité électrique d’une
seule cellule. Selon leur réponse à une variation de la concentration extracellulaire
au glucose ([glucose]e), les neurones sont maintenant classés en trois grands
groupes : i) les neurones non sensibles au glucose, ii) les neurones gluco-excités
(GE) dont l’activité électrique est proportionnelle à la [glucose]e et iii) les neurones
gluco-inhibés (GI) dont l’activité électrique est inversement proportionnelle à la
[glucose]e. Actuellement, les travaux de recherches visent à définir de mieux en
mieux leur équipement moléculaire (Kang L et coll., 2004) (Figure 24).
Les neurones sensibles au glucose (GE et/ou GI) ont été localisés dans les
noyaux VMN, NA, PVN, le noyau suprachiasmatique et le LH, c’est-à-dire dans des
régions impliquées dans le contrôle de l’homéostasie énergétique (Levin BE et coll.,
2002). Dans le VMN, 14 % des neurones étudiés sont GE et 3 % sont GI (Song Z et
coll., 2001). Dans le NA, 20-25 % sont GE et 10-12 % GI, et chez le rat il existe une
régionalisation de ces 2 types, avec une majorité de GE dans le NA latéral et une
majorité de GI dans le NA médian (Fioramonti X et coll., 2004 ; Wang R et coll.,
2004).

- 56 -
Figure 24 : Effet du glucose sur l’activité électrique de neurones du noyau arqué. Neurone GE: gluco-excité ; Neurone GI: gluco-inhibé. D’après Fioramonti X et coll., 2004.
Il a aussi été démontré que les neurones à NPY/AgRP du NA et ceux à
orexines du LH sont activés par un état d’hypoglycémie : l’injection aiguë de 2-DG
(600 mg/kg) augmente, 2 heures après, la quantité d’ARNm NPY/AgRP, révélée par
hybridation in situ, et l’injection aiguë d’insuline (20 UI/kg) induit, 3 heures après,
l’expression de c-Fos dans des neurones immunopositifs pour l’orexine (Sergeyev V
et coll., 2000 ; Moriguchi T et coll., 1999) (Figure 25). En revanche, les neurones à
POMC sont inhibés dans ces conditions énergétiques : leur activité électrique,
mesurée par patch-clamp, diminue lorsque la [glucose]e passe de 10 à 5 mM
(Ibrahim N et coll., 2003). Enfin, l’injection de 2-DG directement dans le SNC induit
l’augmentation de la quantité des ARNm de l’AgRP accompagnée d’une stimulation
de la prise alimentaire (Stütz AM et coll., 2005). A l’inverse, l’application de ces
neuropeptides modulent l’activité des neurones de type GE qui se montrent
immunonégatifs pour le POMC : le NPY les inhibe, tandis que l’α-MSH les active
(Wang R et coll., 2004).

- 57 -
Figure 25 : Effets d’une hypoglycémie sur certains neurones hypothalamiques et influence de neuropeptides sur les neurones GE (gluco-excités). AgRP: agouti-related protein ; α-MSH: α-melanocyte-stimulating hormone ; CART: cocaine- and amphetamine-regulated transcript LH: hypothalamus latéral ; NA: noyau arqué ; NPY: neuropeptide Y ; POMC: pro-opiomélanocortine
Les astrocytes
Le neurone est considéré comme la cellule « noble » du cerveau, étant
responsable de son activité électrique et ainsi de la conduction des informations. Il
en résulte que les autres cellules nerveuses sont moins bien étudiées. Toutefois,
depuis plusieurs dizaines d’années, on observe un développement des travaux
concernant les astrocytes. En effet, il s’avère que ces cellules auraient une vocation
bien plus importante, ne s’arrêtant pas uniquement à un rôle de soutien.
Tout d’abord, ce type particulier de cellules gliales participent à la BHE,
protégeant ainsi le SNC des intrusions ou des influx aspécifiques (protéines, ions)
afin d’y maintenir au mieux l’homéostasie (Gloor SM et coll., 2001) (Figure 26).

- 58 -
Figure 26 : Fonctions principales de l’astrocyte. D’après http://learn.genetics.utah.edu/units/addiction/reward/cells.cfm
Les astrocytes sont aussi des fournisseurs d’énergie pour les neurones qui en sont
de gros consommateurs de part leur activité quasi permanente. Pour cela, ces
cellules gliales ont la capacité de stocker du glucose sous forme de glycogène,
stocks qui seront dégradés selon les besoins (Kong J et coll., 2002). La majeure
partie de l’énergie fournie aux neurones l’est sous forme de lactate provenant de la
glycolyse astrocytaire. En effet, les neurones sont capables de le capturer et de le
transformer en pyruvate dont le catabolisme aboutira à la synthèse d’ATP (Bouzier-
Sore AK et coll., 2002) (Figure 27).
D’autre part, les astrocytes ont un rôle important dans le maintien de la
fonctionnalité des synapses et donc des circuits neuronaux, notamment en régulant
les concentrations ioniques extracellulaires en potassium (Tsacopoulos M et
Magistretti PJ, 1996). De la même façon, les neurotransmetteurs libérés sont
recyclés par l’astrocyte afin d’éviter toute excitotoxicité par accumulation dans la
fente synaptique. Or 80 à 90 % des synapses sont de nature glutamatergique, c'est-
à-dire excitatrice (Paemeleire K, 2002). Le recyclage du glutamate s’imbrique dans

- 59 -
un couplage métabolique astrocytes-neurones (Tsacopoulos M et Magistretti PJ,
1996) (Figure 27).
Figure 27 : Schéma du couplage astrocyte-neurone. La flèche A indique la condition d’activation et la flèche B la condition basale. G: protéine G ; Gln: glutamine ; GLUT: transporteur au glucose ; GS: glutamine synthase ; Lac: lactate ; PGK: phosphoglycérate kinase ; Pyr: pyruvate. Modifié à partir de Tsacopoulos M et Magistretti PJ, 1996.
Le glutamate libéré par le neurone présynaptique est capturé par les
astrocytes adjacents, grâce à un cotransport fonctionnant avec le gradient sodique.
Ce mécanisme engendre une accumulation locale de sodium dans la cellule. Ceci
entraîne alors une augmentation de l’activité de la Na+/K+ ATPase et donc de la
consommation d’ATP. Il s’ensuit une stimulation de la glycolyse astrocytaire, elle-
même entraînant une augmentation du transport de glucose via GLUT1.
Simultanément à l’ATP, est produit du lactate qui sera utilisé par les neurones
adjacents (cf. plus haut). Le glutamate est transformé en glutamine via la glutamine
synthase (GS), qui pourra être redistribuée aux neurones afin de régénérer du
glutamate.
Ces différentes données montrent clairement qu’il existe un couplage entre
les neurones et les astrocytes, les seconds étant essentiels à la survie des premiers.

- 60 -
En ce qui concerne la sensibilité centrale au glucose, des travaux suggèrent
de plus en plus que les astrocytes interviendraient directement dans ce processus.
Une idée reçue est que les neuropeptides seraient uniquement dédiés aux
neurones. Or, ils sont aussi exprimés dans les cellules gliales et il existe une
régionalisation cérébrale de cette expression. Il semblerait en particulier que les
astrocytes pourraient synthétiser du NPY, du moins in vitro, et seraient
immunopositifs pour l’AgRP au niveau du NA et à proximité de l’éminence médiane
(Ubink R et coll., 2003 ; Dunbar J et coll., 2004). La présence de ces protéines
montre qu’ils disposent d’un équipement moléculaire typique du contrôle de
l’homéostasie énergétique en général. Cette hypothèse est renforcée par le fait que
quelques unes de ces cellules nerveuses possèdent aussi le récepteur à insuline et
d’autres, localisées dans le NA, sont anatomiquement proches de neurones
sensibles à la leptine (Schwartz MW et coll., 1992 ; Young JK, 2002).
Enfin, une possible implication directe des astrocytes dans la sensibilité du
glucose a été montrée au travers d’un article, dans lequel l’interruption du couplage
astrocytes/neurones par l’inhibition de la GS, bloque l’activation du NTS
normalement observée suite à l’injection de 2-DG (Young JK et coll., 2000).
Par ailleurs, une autre population de cellules gliales est sensible au glucose,
prouvant que cette sensibilité n’est pas une caractéristique exclusive aux neurones.
En effet, des travaux montrent que des épendymocytes isolés (cellules bordant les
cavités ventriculaires) sont sensibles à des changements de la concentration
extracellulaire en glucose, ceci étant étudié en suivant les fluctuations de leur
concentration intracellulaire en calcium (Moriyama R et coll., 2004). 30% de ces
cellules gliales répondraient soit à une diminution de 10 à 2 mM, soit à une
augmentation de 10 à 20 mM de la [glucose]e. De manière surprenante, dans les
deux cas, la réponse est une augmentation de la concentration intracellulaire en
calcium suite à une entrée de calcium par des canaux voltage-dépendants. Enfin, la
majorité de ces épendymocytes sensibles au glucose expriment la GK, enzyme-clé
de la sensibilité au glucose des cellules β du pancréas (cf. I-2.2 et figure 6).
2.2.5. Acteurs moléculaires
La sensibilité au glucose est particulièrement bien connue au niveau des
cellules β. Elle est indispensable à une sécrétion adéquate d’insuline en fonction
des variations de la glycémie. Or, on peut penser que ce type de mécanisme existe
également au niveau hypothalamique, suggérant que les cellules nerveuses

- 61 -
directement impliquées possèderaient des caractéristiques similaires, notamment la
présence du canal potassique ATP-dépendant composé des sous-unités Kir6.2 et
SUR1, du transporteur au glucose GLUT2 et de l’hexokinase particulière, la
glucokinase (GK) (Pénicaud L et coll., 2002). Notre projet s’intéresse aux deux
dernières protéines.
Le canal potassique ATP-dépendant Kir6.2-SUR1
Kir6.2 (inwardly rectifying K+ channel), formé de 4 sous-unités α, constitue le
pore et se trouve associé au complexe SUR1 (sulfonylurea receptor) composé de 4
sous-unités β. Grâce à lui, les ions potassiques sortent de la cellule et sa fermeture
provoque l’accumulation de charges positives aboutissant finalement à la
dépolarisation de la membrane cytoplasmique.
La mise en évidence de son implication dans la sensibilité au glucose s’est
d’abord faite dans la cellule β du pancréas (Aguilar-Bryan L et Bryan J, 1999)
(Figure 6). Kir6.2 intervient dans la détection pancréatique de l’hyperglycémie car,
contrairement à l’autre isoforme Kir6.1, il est très sensible à l’ATP (dont la
concentration augmente avec l’augmentation de [glucose]e) qui, en se liant au pore,
induit sa fermeture (Quast U, 2000). Dans ce type cellulaire, Kir6.2 est associé à
SUR1. Ce complexe est capable de lier l’ATP, mais il possède davantage d’affinité
pour l’ADP. Cette liaison est donc favorisée lorsque le rapport ATP/ADP diminue,
c’est-à-dire lorsque la [glucose]e diminue, et elle conduit à l’ouverture du canal.
Le canal Kir6.2-SUR1 a ensuite été recherché au niveau du SNC afin de
déterminer si là aussi, sa présence était indispensable à la détection du glucose. Or
les 2 isoformses ont bien été retrouvées, notamment dans les régions cérébrales
liées à l’homéostasie énergétique suggérant un rôle cérébral similaire (Dunn-
Meynell AA et coll., 1998). Cette hypothèse est renforcée par les résultats de RT-
PCR sur cellule unique appliquée à des neurones GE du VMN et du NA, indiquant
la synthèse de leurs ARNm (Ibrahim N et coll., 2003). Par ailleurs, l’application de
sulfonylurées, molécules qui vont se lier au complexe SUR et conduire ainsi à la
fermeture du canal, qui miment donc une augmentation de la [glucose]e, active les
neurones GE du VMN (Kang L et coll., 2004).
Toutefois, des travaux menés au laboratoire indiquent qu’il existe un
mécanisme de sensibilité cérébrale au glucose pour lequel les canaux potassiques
ATP-dépendant ne sont pas requis (Fioramonti X et coll., 2004). Ces deux processus
ne semblent pas exclusifs et doivent probablement se compléter en fonction de la
[glucose]e.

- 62 -
Le transporteur au glucose GLUT2
D’une manière générale, les transporteurs au glucose GLUT sont des
protéines à 12 domaines transmembranaires, assurant le transport facilité de cet
hexose et de ses analogues non métabolisables (entre autres, le 2-DG), ainsi que du
fructose ou de l’acide ascorbique (Medina RA et Owen GI, 2002).
Dans le SNC, les principaux GLUTs assurant le transport basal du glucose
nécessaire à la vie normale des cellules sont GLUT1, présent dans les cellules
endothéliales et les astrocytes, et GLUT3, spécifiquement neuronal. Tous les deux
présentent une constante de Michaelis (Km) faible, comprise entre 1 et 3 mM, c'est-
à-dire qu’ils présentent une forte affinité pour le glucose et sont donc saturés pour
une glycémie normale de 5 mM (Thorens B, 1992). A côté de ces transporteurs
ubiquitaires, d’autres GLUTs sont présents dans le SNC de manière plus localisée,
en plus faibles densités. C’est le cas de GLUT2, qui nous intéresse.
GLUT2 a été découvert, cloné et étudié pour la première fois par l’équipe de
B. Thorens en 1988. Il a d’abord été qualifié de « transporteur au glucose du foie »
bien qu’il soit aussi localisé dans l’intestin, le rein et les cellules β des îlots de
Langerhans et le SNC. Le transport est spécifique à l’énantiomère dextrogyre du
glucose (Thorens B et coll., 1988). Il se différencie des autres GLUT par son Km élevé
(environ 20 mM), ce qui révèle une faible affinité pour le D-glucose. Celui-ci est
donc transporté de manière proportionnelle à sa concentration environnante, sans
présenter de phénomène de saturation dans les gammes physiopathologiques de
glycémie (Thorens B, 1992).
La présence de GLUT2 (ARNm et protéine) dans le SNC a été démontrée au
sein de notre laboratoire en 1994, en particulier dans les NTS, DMV, PVN, NA et le
LH, renforçant l’hypothèse de son implication dans la régulation de l’homéostasie
énergétique (Leloup C et coll., 1994). Par la suite, des expériences de colocalisation
ont indiqué que ce transporteur était exprimé à la fois par des neurones, des
oligodendrocytes (cellules formant la gaine de myéline des neurones) et des
astrocytes (Arluison M et coll., 2004). Au sein du NA, GLUT2 est retrouvé le long du
troisième ventricule dans les épendymocytes, dans quelques astrocytes, mais
également dans les tanycytes, épendymocytes spécialisés dont le corps cellulaire
forme la paroi basale du troisième ventricule et qui possèdent de longs
prolongements atteignant l’éminence médiane, constituant ainsi un lien cellulaire

- 63 -
entre le liquide céphalo-rachidien et le sang (Ngarmukos et coll., 2001 ; Young JK et
McKenzie JC, 2004 ; Garcia MA et coll., 2003).
Par la suite, il a été démontré que GLUT2 occupait une place majeure dans la
sensibilité cérébrale au glucose. En effet, l’injection d’antisens GLUT2, directement
dans le NA, bloque l’augmentation de l’insulinémie normalement induite par
l’injection aiguë de glucose par la carotide vers le SNC de rats (cf. § III-2.2.3) (Leloup
C et coll., 1998). Simultanément, un autre groupe de chercheurs a suivi une
démarche similaire, où l’injection des antisens a été réalisée, cette fois, dans le
ventricule latéral. Les rats traités présentent une diminution du poids corporel et de
la prise alimentaire basale, qui n’est plus stimulée par du 2DG (Wan HZ et coll.,
1998).
Enfin, l’importance de GLUT2 dans la régulation de la balance énergétique
s’observe au travers du phénotype des souris qui en sont privées. L’invalidation
totale de son gène est létale. Néanmoins, un phénotype viable peut être obtenu en
compensant l’absence de GLUT2 par une expression de GLUT1 au niveau des
cellules β. Bien que l’insulinémie, la leptinémie et la glycémie de ces animaux soient
normales, ces souris présentent une augmentation de la prise alimentaire ainsi
qu’une absence de régulation des neuropeptides orexigènes/anorexigènes et des
concentrations circulantes de ghréline (Bady I et coll., 2006). De plus, l’injection
intrapéritonéale de glucose (400 mg/kg) n’inhibe plus, et celle de 2-DG (150 mg/kg)
ne stimule plus, la prise alimentaire. De même, l’expression des neuropeptides
hypothalamiques NPY, AgRP, POMC, CART, n’est plus modulée par l’injection
intracérébroventriculaire de glucose (5 μl, 20 mg/ml). Finalement, l’expression
exclusivement astrocytaire de GLUT2 chez ces souris, permet de restaurer une
sécrétion de glucagon après injection intrapéritonéale ou intracérébroventriculaire
de 2-DG, ainsi qu’une activation du NTS et du DMV révélée par l’expression de c-
Fos (Marty N et coll., 2005).
Ainsi, l’ensemble de ces données suggèrent fortement que GLUT2 fait partie
intégrante des mécanismes sous tendant la sensibilité au glucose dans le SNC.
La glucokinase (GK)
Il s’agit de l’hexokinase IV responsable de la sensibilité au glucose des
cellules β. Elle possède une faible affinité pour ce sucre et en assure sa
phosphorylation. Cette étape est limitante pour la glycolyse qui précède le cycle de
Krebs, fournisseur principal d’ATP. A la différence des autres hexokinases, la GK

- 64 -
n’est pas inhibée par son produit, le glucose-6-phosphate. Cette caractéristique
fondamentale permet un degré de phosphorylation du métabolite proportionnel à
son entrée dans la cellule et de ce fait, aboutit à un niveau de synthèse d’ATP en
relation directe avec les quantités intracellulaires de glucose. Ainsi, par analogie
avec la cellule β, on peut penser que dans les cellules cérébrales sensibles au
glucose, pourvues de GK, le ratio ATP/ADP augmenterait parallèlement à la
glycémie, provoquant la fermeture de canaux potassiques dépendants de l’ATP et à
une dépolarisation membranaire (Schuit FC et coll., 2001) (Figure 28).
Figure 28 : Schéma illustrant la sensibilité au glucose dans les cellules nerveuses, comprenant les deux protéines-clé, GLUT2 et GK. Ca2+: ions calciques ; CaVm: canal calcique voltage-dépendant ; GK: glucokinase ; GLUT2: transporteur au glucose 2 ; K+: ions potassiques ; KATP: canal potassique ATP-dépendant.
Des travaux d’immunohistochimie révèlent la présence de la GK le long des
parois du canal central et des différents ventricules (latéraux, troisième et
quatrième) (Maekawa et coll., 2000). Le marquage est cytoplasmique et la majorité
des cellules positives sont des épendymocytes. Aucun signal n’est observé dans
l’area postrema ni dans le NTS. Au niveau de l’hypothalamus, seuls les
épendymocytes et quelques cellules localisées à proximité du 3ème ventricule sont

- 65 -
marquées. Ces cellules gliales possédant la GK, sont aussi positives pour GLUT2 ;
le marquage GLUT2 est alors restreint à la partie ciliaire (Maekawa et coll., 2000). Il
est important de noter que les épendymocytes du canal central sont pour la plupart
sensibles aux variations de glucose (cf. § III-2.2.4). De manière intéressante, cette
sensibilité disparaît lorsque la GK est inhibée par l’application d’alloxane, composé
employé pour bloquer cette activité enzymatique (Moriyama R et coll., 2004).
Des résultats d’hybridation in situ confortent aussi l’hypothèse de
l’implication de la GK, couplée à GLUT2, dans la sensibilité cérébrale au glucose,
puisque leurs ARNm sont retrouvés colocalisant au niveau de la paroi du troisième
ventricule, comme dans le cas de la protéine. Néanmoins, un marquage au niveau
du noyau supraoptique, du NA, de l’éminence médiane, des VMN, DMN, PVN et le
LH est aussi révélé (Navarro M et coll., 1996 ; Lynch RM et coll., 2000). Au sein du
NA, des neurones à NPY, à POMC et à GABA expriment la GK (Dunn-Meynell AA et
coll., 2002). Enfin, des travaux ont permis de montrer que dans le tronc cérébral,
les neurones GE et GI du DMV l’exprimaient aussi, tandis que dans
l’hypothalamus, c’est le cas pour 64 % des GE et 43 % des GI du VMN (Balfour RH
et coll., 2006 ; Kang L et coll., 2004).
Ces diverses études, bien que parfois un peu contradictoires, laissent à
penser que, comme GLUT2, la GK est bien impliquée dans la sensibilité au glucose
dans le SNC au niveau des épendymocytes, et vraisemblablement des neurones et
des astrocytes.

- 66 -
IV – Nos objectifs et démarches scientifiques
Dans les modèles intégrés, la sensibilité cérébrale au glucose est le plus
souvent envisagée dans le contexte d’une glucoprivation par l’utilisation du 2-DG,
un analogue non métabolisable du glucose (Briski KP, 1998 et 2000 ; Young JK et
coll., 2000). Lorsque l’hyperglycémie est directement étudiée, elle se trouve
généralement dans un cadre à long terme, avec une injection chronique et/ou
périphérique du monosaccharide (Dunn-Meynell AA et coll., 1997 ; Carrasco M et
coll., 2001).
Contrairement à ces travaux, notre objectif est d’étudier les mécanismes de
la sensibilité cérébrale au glucose à très court terme, afin de mettre en évidence :
1. les régions cérébrales sensibles à une hyperglycémie et donc à une
augmentation de la concentration extracellulaire en glucose
2. les types cellulaires impliqués : neurones et/ou cellules gliales
3. les acteurs moléculaires mis en jeu : GLUT2 et/ou la GK.
1. Notre modèle d’hyperglycémie cérébrale
Nous avons opté pour une injection aiguë d’une solution de glucose via la
carotide et ce, chez le rat éveillé. Pour ce faire, nous avons repris et adapté un
modèle précédemment mis en place au laboratoire qui consiste en l’insertion d’un
cathéter dans la carotide en direction du cerveau, permettant l’injection d’un bolus
de glucose (9 mg/kg soit une solution à 115 mM) sans effet sur la glycémie
périphérique mais induisant très rapidement une stimulation transitoire de la
sécrétion d’insuline (Leloup C et coll., 1998). Ce protocole avait été jusqu’alors,
utilisé sur des animaux anesthésiés.
Une difficulté majeure dans l’étude de la sensibilité au glucose in vivo, est de
différencier son effet direct de celui de l’insuline, puisqu’une hyperglycémie induit
immédiatement la libération de l’hormone pancréatique. Comme le bolus injecté ne
modifie pas la concentration en glucose périphérique, mais vraisemblablement celle
dans le SNC au vu de la dose injectée, nous pouvons dire que les effets observés
mettent en jeu une détection cérébrale d’une hyperglycémie locale, et non de
l’insuline.
Il s’agit dès lors de déterminer si, dans ce protocole, l’hypothalamus, et plus
précisément certains noyaux, sont activés suite à cette injection aiguë,
responsables de la réponse nerveuse induisant la génération d’un « pic » d’insuline.
Pour cette mise en évidence, l’expression du proto-oncogène c-fos a été utilisée.

- 67 -
2. Le proto-oncogène c-fos
Cette technique est très largement utilisée dans le domaine des
neurosciences. Nous avons commencé par la valider en reprenant un modèle décrit
dans la littérature (Briski KP et Brandt JA, 2000).
c-Fos est une protéine qui se lie à d’autres produits de gènes précoces, le
plus souvent de la famille Jun, pour former des dimères s’accrochant au domaine
de liaison AP-1 de l’ADN. Ceci permet finalement un contrôle adéquat de la
transcription suite à un signal donné. A l’état basal, l’expression de c-Fos est faible
dans l’ensemble du SNC. Elle augmente fortement suite à un stimulus et ce, de
manière spécifique dans certaines régions cérébrales, en accord avec l’agent
inducteur. La protéine peut être détectée par immunohistochimie classique et le
marquage obtenu est nucléaire. Sa concentration est maximale environ 2 h après la
stimulation et diminue ensuite (Hoffman GE et coll., 1993).
L’utilisation de cette protéine a été validée en comparant les résultats
obtenus suite à un stimulus donné avec ceux d’autres techniques déjà connues
comme celle de l’autoradiographie via le 2-DG tritié. L’avantage crucial par rapport
à cette dernière, est que l’on obtient une cartographie d’activation cérébrale de
résolution cellulaire, permettant ainsi des doubles marquages dont nous avions
besoin pour déterminer ensuite le(s) type(s) cellulaire(s) entrant en jeu (Sagar SM et
coll., 1988).
3. Outils pharmacologiques
Nous avons entrepris la caractérisation du, ou des types cellulaires activés,
par double immunohistochimie dirigée contre c-Fos et un marqueur spécifique des
neurones ou des astrocytes ou des oligodendrocytes. Comme les données de la
littérature laissent à penser que les astrocytes seraient directement impliqués dans
la détection cérébrale du glucose, notre démarche a été d’altérer le métabolisme des
carbohydrates spécifiquement dans ces cellules, suggérant l’interruption de leur
couplage métabolique avec les neurones (Young JK et coll., 2000). La méthionine
sulfoximine (MSO) a été employée à cette fin.
Enfin, selon notre hypothèse, les cellules nerveuses sensibles au glucose
mettent en jeu des mécanismes similaires à ceux des cellules β du pancréas,
notamment le couple protéique GLUT2/GK (Pénicaud L et coll., 2002). Nous avons
donc opté pour une approche pharmacologique en utilisant des inhibiteurs de la
GK, l’alloxane et la glucosamine (GlcN). L’avantage du premier est qu’il serait

- 68 -
spécifiquement transporté par GLUT2 (Elsner M et coll., 2002) (cf. § III-2.2.5). Or il
n’existe pas d’inhibiteur sélectif de ce transporteur. La GlcN est transportée
indifféremment par les transporteurs au glucose.
Notons que l’utilisation d’agents pharmacologiques pose toujours le problème
de la spécificité de l’effet observé. C’est pourquoi, une série d’expériences
supplémentaires a été réalisée afin de limiter au plus cette éventualité.
Chaque agent pharmacologique sera développé dans la partie
«Résultats/Discussion ».

- 69 -
MMaattéérriieellss eett MMéétthhooddeess

- 70 -
V – Animaux
L’ensemble des travaux a été réalisé sur des rats mâles Wistar de 300 ± 30 g
(Charles River) maintenus en élevage au service de zootechnie de l’IFR 31 dans une
pièce éclairée 12 h par jour (de 7 h à 19 h), et dont la température est maintenue
constante à 22°C. Les animaux ont un accès libre à l’eau et à la nourriture (régime
standard de laboratoire). Les expériences se sont déroulées sur des rats nourris.
VI – Etude de l’activation cérébrale
1. Validation de la technique Fos : injection de 2-déoxyglucose
Les mises au point des techniques de fixation intracardiaque des rats et de
l’immunohistochimie Fos ont été faites à partir du modèle de Briski KP et Brandt
JA, 2000, dans lequel du 2-déoxyglucose (2-DG), analogue du glucose non
métabolisé par les cellules, est utilisé pour mimer une glucopénie cellulaire.
Les rats éveillés ont ainsi reçu une injection intrapéritonéale de 500 μl d’une
solution de 2-DG (160 mg/ml) dilué dans du NaCl 0,9 %, tandis que les rats
contrôles ont reçu 500 μl de la solution véhicule. La fixation intracardiaque a été
effectuée 2 h après.
2. Microchirurgie : pose d’un cathéter « à demeure » dans la carotide
L’expression de c-fos peut être induite par une large palette de stimuli
incluant notamment les stress de diverses origines comme les anesthésies ou
chirurgies. C’est pourquoi les rats ont été opérés plusieurs jours avant l’injection
intracarotidienne.
Les animaux sont anesthésiés par une injection intrapéritonéale de
pentobarbital sodique (60 mg/kg de poids, Centravet, Lapallise). Un cathéter (0,5 x
0,9 mm, silicone, Access Technologies, Netherlands) contenant du sérum
physiologique hépariné (100 U/ml, Sigma, St Quentin Fallavier) est inséré dans
l’artère carotidienne interne gauche en direction du cerveau et maintenu en place
par plusieurs ligatures à l’aide de fils non-résorbables. L’autre extrémité du cathéter
est alors passée sous la peau pour ressortir dorsalement au niveau du cou. Des
points de suture sont réalisés et permettent notamment le maintien en place du
cathéter. Ce dernier est ensuite rempli d’une solution héparinée de

- 71 -
polyvinylpyrrodilone à 20 % (Sigma) afin d’empêcher son obstruction par
coagulation sanguine, puis son ouverture bouchée.
Jusqu’au jour de l’expérimentation, les rats sont pesés quotidiennement pour
suivre la reprise de leur poids, bon indice de leur récupération. Le cathéter est rincé
avec du sérum physiologique hépariné et à nouveau rempli avec la solution
héparinée de polyvinylpyrrodilone. Cette manière de procéder, outre le fait qu’elle
évite tout risque d’obstruction du cathéter, permet d’habituer les animaux à être
manipulés par l’expérimentateur et à recevoir une « injection » à l’état éveillé.
3. Protocoles d’injection
Ils ont lieu 4 à 7 jours après la chirurgie. Toutes les injections carotidiennes
se déroulent le matin entre 9 et 11 h lorsque la concentration cérébrale en
glycogène est relativement stable (Helary-Bernard K et coll., 2000) et directement au
service de zootechnie, afin d’éviter toute variabilité liée au stress ou au statut
métabolique de chaque animal.
Les rats éveillés reçoivent une injection (130 μl en 35 s) de glucose (9 mg/kg,
300 mOsmoles, n=11) ou de sérum physiologique (n=7) via le cathéter
intracarotidien. Ce protocole est connu pour induire, chez le rat anesthésié, une
sécrétion rapide et transitoire d’insuline de nature nerveuse, sans modification de la
glycémie périphérique (Atef N et coll., 1995).
Deux autres groupes d’animaux ont été prétraités 18 h avant, avec une
injection par voie intrapéritonéale de L-méthionine sulfoximine (MSO, 100 mg/kg,
Sigma, MSO/glucose n=12, MSO/NaCl n=9) (cf. § III-2.2.4). La concentration et la
durée du prétraitement ont été choisies pour induire une accumulation maximale
du glycogène cérébral, reflet de l’altération du métabolisme des carbohydrates dans
les astrocytes, tout en limitant l’effet convulsif de la MSO (Young JK et coll., 2000).
4. Fixation intracarotidienne et intracardiaque
Les rats sont anesthésiés avec du pentobarbital sodique 2 h après le
protocole d’injection, au moment où la concentration en protéine c-Fos est
maximale suite à un stimulus (Hoffman GE et coll., 1993).
Le cathéter carotidien est relié à une seringue (installée sur un pousse-
seringue) contenant de la solution de fixation (formaldéhyde 4 % - acide picrique
0,2 %). Suite à une laparotomie, le cœur est mis à nu et la veine cave inférieure
sectionnée afin de permettre l’évacuation du sang, puis des liquides perfusés. Très
rapidement, une aiguille, reliée à un double système de perfusion (sérum

- 72 -
physiologique hépariné et solution fixatrice) est introduite dans le ventricule
gauche. Le débit est régulé par une pompe péristaltique (200 ml/h). Pendant les 3
premières minutes, l’injection de sérum physiologique hépariné permet l’élimination
du sang. La solution de fixation est alors perfusée pendant 1 h par les 2 voies
(carotidienne et cardiaque) (Figure 29).
Figure 29 : Système de perfusion pour la fixation intracardiaque/intracarotidienne. FA : formaldéhyde.
Après décapitation, le cerveau est prélevé, postfixé toute la nuit à 4°C dans la
même solution de fixation, puis laissé en imbibition dans du tampon sucrose 20 %
(rôle cryoprotecteur) pendant au moins 3 jours à 4°C. Une tranche épaisse
contenant la région hypothalamique est alors plongée dans un produit d’enrobage
pour congélation (Cryomount, Histolab) et le tout est congelé à -90°C dans de
l’isopentane refroidi à l’azote liquide. Le bloc ainsi obtenu est conservé à -80°C
jusqu’à utilisation.
5. Immunohistochimie c-Fos
Des coupes successives de 45 μm d’épaisseur situées dans la région
hypothalamique, de -0,80 mm à -3,8 mm par rapport au bregma en accord avec

- 73 -
l’atlas stéréotaxique (Paxinos G et Watson C, 1986), sont obtenues à l’aide d’un
cryostat (température de la chambre à -20°C, température du porte-objet à -15°C,
Leica, Rueil Malmaison). Elles sont immédiatement réparties dans des plaques de
24 puits contenant du PBS (Phosphate-Buffered Saline).
Une coupe sur deux est utilisée. Toutes les étapes se déroulent sous
agitation à température ambiante.
Les coupes flottantes sont tout d’abord lavées avec du PBS. L’activité des
peroxydases endogènes est ensuite neutralisée avec un traitement de 30 min dans
une solution de H2O2 0,3 % (Sigma) - PBS. Après une nouvelle série de lavages, les
sites antigéniques aspécifiques sont bloqués pendant au moins 2 h grâce à une
solution de triton 0,25 % (X100, Sigma) - NGS (Normal Goat Serum, Sigma) 3 % -
PBS. Les coupes sont mises en présence de l’anticorps primaire polyclonal anti-c-
Fos (antisérum Ab-5 de lapin dirigé contre les résidus 4 à 17 de la protéine c-Fos
humaine, Oncogene, USA) dilué à 1/10000 dans du triton 0,25 % - PBS, toute la
nuit à 4°C.
Après 4 lavages, les coupes sont plongées pendant 1 h dans une solution
d’anticorps secondaire biotinylé dirigé contre les IgG de lapin (antisérum de chèvre,
H + L, Jackson Laboratories, USA) dilué à 1/2000 dans du triton 0,25 % - PBS.
Après rinçages, elles sont alors mises à incuber pendant 1 h dans une solution de
streptavidine peroxydase (Jackson Laboratories) diluée à 1/2000 dans du triton
0,25 % - PBS. Les coupes sont une dernière fois lavées et l’activité peroxydase est
révélée grâce à une solution de diaminobenzidine/nickel ammonium sulfate (DAB-
Nickel) (Shu S et coll., 1988). La coloration est arrêtée avec de l’eau. Les coupes
sont montées sur lames silanisées, déshydratées et finalement incluses dans une
résine (Eukitt, CML).
Afin de contrôler la spécificité du marquage, l’anticorps primaire est omis à
chacune des premières coupes de cerveau (incubée alors dans du triton 0,25 % -
PBS). De plus, elle a également été vérifiée par compétition avec le peptide
immunogène (Oncogene) utilisé 100 fois plus concentré que l’anticorps.
6. Double immunohistochimie c-Fos / marqueur cellulaire
Dans le but de déterminer le ou les type(s) cellulaire(s) immunopositif(s) pour
c-Fos, des coupes ont fait l’objet d’une double immunohistochimie pour c-Fos et un
marqueur cellulaire : CNPase (2’, 3’-cyclic nucleotide 3’-phosphodiesterase) pour les

- 74 -
oligodendrocytes, GFAP (glial fibrillary acidic protein) pour les astrocytes et NeuN
(neuronal nuclei) pour les neurones.
La première étape consiste en l’immunohistochimie c-Fos comme décrite
précédemment. Suite à la révélation DAB-Nickel, les coupes flottantes sont rincées
avec du PBS puis mises à incuber, toute la nuit à 4°C, dans une solution
d’anticorps primaire dirigé contre le marqueur cellulaire concerné dilué dans du
triton 0,25 % - PBS : les anticorps monoclonaux anti-CNPase ou anti-GFAP sont
dilués à 1/500 (Sigma), l’anticorps monoclonal anti-NeuN est dilué à 1/100
(Valbiotech).
Après rinçages, dans le cas des marquages CNPase ou GFAP, les coupes sont
mises en présence, pendant 1 h, d’une solution diluée à 1/500 d’anticorps
secondaires dirigés contre les IgG de souris et conjugués à la peroxydase
(antisérum d’âne, H + L, Jackson Laboratories). Dans le cas du marquage neuronal,
les coupes subissent une étape d’amplification à l’aide d’un anticorps secondaire
biotinylé dilué à 1/1000 (antisérum d’âne anti-IgG de souris, H + L, Jackson
Laboratories), puis une troisième incubation d’1 h en présence d’une solution de
streptavidine peroxydase également diluée à 1/1000. Le marquage cellulaire est
révélé par une solution de DAB et arrêté avec de l’eau. Les coupes sont alors
montées sur lames silanisées, déshydratées et incluses dans une résine.
7. Quantification des noyaux immunopositifs pour c-Fos et des cellules
doublement immunopositives c-Fos / GFAP
Les cellules immunopositives pour c-Fos ont été quantifiées sans tenir
compte des variations d’intensité de coloration nucléaire. Cette quantification a été
réalisée au niveau de différentes régions hypothalamiques d’intérêt (Noyau arqué :
NA ; Noyau hypothalamique paraventriculaire : PVN ; Noyau hypothalamique
ventromédian : VMN) et dans une région constituant un contrôle négatif (cortex
cérébral : CC, non impliqué dans l’homéostasie énergétique), en prenant 3 coupes
distantes successivement de 80 μm, à l’aide d’un logiciel d’analyse d’image (Biocom
Visiolab), sous microscope à transmission (Leica).
Pour les cellules doublement positives c-Fos / GFAP, la quantification s’est
faite manuellement dans le NA et le PVN.
8. Expression des résultats
Les données individuelles de chaque rat correspondent à la somme des
noyaux immunopositifs pour c-Fos comptés bilatéralement sur 3 coupes

- 75 -
successives pour une région cérébrale. Le nombre de cellules immunopositives pour
c-Fos et de cellules doublement marquées par structure est exprimé en « moyenne ±
erreur standard à la moyenne ».
VII – Etude de la régulation nerveuse de la sécrétion d’insuline
1. Microchirurgie : pose d’un cathéter dans la carotide
Les rats sont anesthésiés par une injection intrapéritonéale de pentobarbital
sodique. Un cathéter (0,5 x 1 mm, polyéthylène, Portex, Scientific Laboratory, UK)
contenant du sérum physiologique hépariné (100 U/ml, Sigma) est inséré dans
l’artère carotidienne gauche en direction du cerveau et maintenu en place par
plusieurs ligatures. Une période de récupération d’au moins 15 min avant le début
du protocole d’injection est respectée permettant la stabilisation de la glycémie.
2. Protocoles d’injection
Les injections se déroulent uniquement le matin.
Les rats reçoivent une injection (100 μl en 30 s) de D-glucose (9 mg/kg, 300
mOsmoles) ou de sérum physiologique via le cathéter intracarotidien au temps zéro
(t0). Un autre groupe de rats contrôles a reçu une injection de L-glucose (9 mg/kg,
300 mOsmoles, énantiomère du glucose non actif métaboliquement) afin de valider
la spécificité de la réponse.
Pour déterminer les voies nerveuses ou les acteurs moléculaires cérébraux
impliqués dans la détection du glucose, différents agents pharmacologiques (Sigma)
sont utilisés. Les groupes de rats témoins reçoivent une injection de NaCl 0,9 % de
même volume.
Atropine méthylnitrate : antagoniste des récepteurs muscariniques à acétylcholine,
inhibe la voie parasympathique, ne traverse pas la barrière hémato-
encéphalique (Jansson L et Hellerstrüm C, 1986) ; 5 mg/kg, 400 μl injectés par la
veine fémorale, 20 minutes avant (t-20) l’injection de glucose ou de NaCl 0,9 % à t0.
Oxymétazoline : agoniste partiel des adrénorécepteurs α2A, active la voie
orthosympathique, agoniste des récepteurs à sérotonine 5-HT1A, 5-HT1B, 5-HT1D et
agoniste/antagoniste des 5-HT1C (Magnan C et coll., 1999) ; 1000 pmol/kg, 400 μl
injectés en intrapéritonéal à t-5.

- 76 -
L-Méthionine sulfoximine (MSO) (Figure 39) : inhibiteur de la glutamine synthase
localisée spécifiquement dans les astrocytes au niveau cérébral (Young JK et coll.,
2000), ainsi que de la γ-glutamylcystéine synthétase (Cooper AJL et coll., 1976) ; 100
mg/kg, 600 μl en intrapéritonéal 18 h avant t0.
Buthionine sulfoximine (BSO) : analogue de la MSO, inhibiteur uniquement de la γ-
glutamylcystéine synthétase et environ 100 fois plus efficace que la MSO (Griffith OW
et Meister A, 1979) ; 100 mg/kg, 600 μl en intrapéritonéal 18 h avant t0.
Glucosamine (GlcN) (Figure 49): inhibiteur de la glucokinase et activateur de la voie
des hexosamines (Yoshikawa H et coll., 2002) ; 80 mg/kg, 1800 mOsmol, 125 μl
injectés en 38 s par le cathéter intracarotidien 30 s avant t0.
N-acétylglucosamine (GlcNAc) (Figure 49): activateur de la voie des hexosamines,
inhibiteur de la glucokinase moins efficace que la GlcN. Le premier effet doit donc
prédominer (Wells L et coll., 2003 ; Agius L et Stubbs M, 2000) ; 80 mg/kg, 1800
mOsmol, 125 μl injectés en 38 s par le cathéter intracarotidien 30 s avant t0.
Alloxane : inhibiteur de la glucokinase, transporté spécifiquement par le transporteur
au glucose GLUT2 (Elsner M et coll., 2002) ; 1 mg/kg, 100 μl injectés en 30 s par le
cathéter intracarotidien soit à t-10, soit à t0, seul ou simultanément au glucose.
3. Test de tolérance au glucose (IVGTT)
Des IVGTT (intravenous glucose-tolerance test) ont été réalisés afin de vérifier
que les différentes injections d’agents pharmacologiques n’affectaient pas la
capacité du pancréas à secréter de l’insuline. Ainsi, ils permettent de mettre en
évidence que notre effet est spécifiquement cérébral.
L’agent pharmacologique est injecté de la même manière que décrite
précédemment (quantité, voie d’injection et temps identiques). Au temps 0 une
solution de glucose de très forte concentration (0,8 g/kg, 250 μl) est inoculée par la
veine fémorale, induisant une hyperglycémie périphérique massive. Si les fonctions
pancréatiques sont intactes la glycémie revient à une valeur normale sous l’action
de l’insuline.
4. Prélèvements sanguins
Des échantillons de sang (environ 200 μl) ont été prélevés au bout de la
queue, aux temps t-10 minutes (t-20 si l’agent pharmacologique est injecté à ce
moment-là), t0 (juste avant l’injection de glucose), t1, t3, t5, t10, t15 et t20. Dans le
cas des IVGTT les temps étudiés sont t-10, t0, t5, t10, t15 et t20 min.

- 77 -
Le sang, récupéré dans des tubes contenant un peu d’héparine, est
immédiatement centrifugé à raison de 6000 rotations par minute. Les plasmas ainsi
obtenus sont conservés dans de la glace avant d’être stockés à –80°C.
5. Dosages
La glycémie est mesurée extemporanément à partir d’une goutte de sang
déposée sur un analyseur de glucose (Glucotrend, Roche, Meylan).
L’insuline plasmatique est dosée à l’aide d’un kit radio-immunologique dont
le principe est basé sur la compétition entre l’insuline à mesurer et de l’insuline
marquée à l’iode 125 (quantité déterminée grâce à un compteur de rayons gamma)
vis à vis d’un anticorps (Diasorin, Antony).
6. Expression des résultats
La glycémie est exprimée en millimolaires (mM), l’insulinémie en micro-unités
par millilitre (μU/ml). Les valeurs présentées correspondent aux différences (Δ) de
concentrations mesurées aux différents temps par rapport à la concentration basale
mesurée au temps zéro. Ce sont les moyennes des Δ ± l’erreur standard à la
moyenne.
VIII – Méthionine sulfoximine : dosage du glycogène cérébral
Suite au prétraitement de 18 h, l’efficacité de la MSO a été vérifiée sur
plusieurs cerveaux. En effet, cet agent pharmacologique induit une forte
accumulation de glycogène dans les astrocytes (Hevor TK et coll., 1985). C’est
pourquoi la quantité de glycogène contenu dans les cerveaux de rats ayant reçu 18
h avant le prélèvement une injection de MSO ou de NaCl a été déterminée grâce à la
méthode colorimétrique de Trinder. Pour chaque animal, le dosage est effectué en
triplet (n=4 pour chaque groupe).
Les rats anesthésiés sont décapités et leur cerveau prélevé le plus
rapidement possible avant d’être plongé dans l’azote liquide et conservé à –80°C. La
rapidité est très importante, le glycogène se dégradant vite.
Pour chaque rat, un petit morceau (30 à 50 mg) du cortex frontal est mis en
présence de 50 μl de NaOH 1 N. Les tubes sont alors mis à 55°C pendant 1 h et
agités de temps en temps. 50 μl de HCl 1 N sont ensuite ajoutés et les tubes, une
fois vortexés, sont centrifugés 15 min à 13000 rotations par minute.

- 78 -
20 μl de surnageant sont alors mis en présence soit de 40 μl
d’amyloglucosidase (enzyme dégradant le glycogène en glucose) diluée à 1/500 dans
du tampon acétate de sodium 0,2 M, pH 4,8, soit de 40 μl de tampon acétate de
sodium. Ces derniers tubes servent à mesurer la quantité de glucose « libre »
présent dans le cerveau, non stocké sous-forme de glycogène. Tous les tubes sont
mis à incuber 30 min à 55°C sous agitation. Une gamme étalon de concentrations
connues en glucose est réalisée en doublet : 0 ; 0,5 ; 1 ; 2 g/l. 700 μl de solution de
Trinder sont finalement additionnés à tous les tubes. On laisse la coloration se
développer pendant 20 min avant de mesurer les densités optiques au
spectrophotomètre à la longueur d’onde de 505 nm.
Les résultats sont exprimés en μg de glucose par mg de tissu cérébral.
Chaque triplet est moyenné avant de calculer la moyenne ± l’erreur standard à la
moyenne de chaque groupe de rats.
IX – Analyses statistiques
Les moyennes de chaque groupe de données (quantification du nombre de
noyaux immunopositifs pour c-Fos par région cérébrale, glycémie, insulinémie,
quantité de glycogène) ont été comparées à l’aide du t-test non-apparié de Student.
La différence est considérée statistiquement significative lorsque p est inférieure à
0,05.

- 79 -
RRééssuullttaattss

- 80 -
Le but de l’ensemble de ces travaux a été l’étude de l’activation
hypothalamique par une hyperglycémie centrale aiguë. Or, le SNC peut être activé
par une grande variété de stimuli, comme les anesthésiques. Etant partis d’un
modèle d’injection de glucose par la carotide vers le cerveau réalisée chez le rat
anesthésié (Atef N et coll., 1995), des mises au point ont été nécessaires pour
l’appliquer à l’animal éveillé.
Par la suite, nous avons caractérisé les aires hypothalamiques sensibles
à une injection aiguë de glucose, puis étudié l’implication de cellules
particulières, les astrocytes, dans ce mécanisme et enfin entrepris de
déterminer si certains acteurs moléculaires, le transporteur au glucose GLUT2
et la glucokinase, y jouaient un rôle. Ces trois points constitueront ainsi les
trois grandes parties de mes résultats.
X – Caractérisation de la sensibilité au glucose
1. Résultats I
1.1. Mise en place et validation des modèles expérimentaux
Dans un premier temps, il a été indispensable de mettre en place et de
reproduire les protocoles expérimentaux de base, nécessaires pour toute la suite
des travaux entrepris.
1.1.1. Injection(s) aiguë(s) de glucose vers le système nerveux central :
effets sur la glycémie et l’insulinémie
Le glucose est retrouvé dans l’alimentation et l’organisme sous l’énantiomère
dextrogyre, soit le D-glucose. Il s’agit donc de la forme métabolisable qui est
injectée.
La concentration de D-glucose injectée de manière aiguë vers le cerveau via
la carotide, 9 mg par kg de poids de l’animal, n’induit pas d’augmentation de la
glycémie périphérique, tout comme chez les rats ayant reçu du sérum physiologique
ou l’énantiomère lévogyre (Figure 30A).
En revanche, seul le D-glucose induit une augmentation rapide et transitoire
de l’insulinémie (32,2 ± 6,1 μU/ml au temps 1 minute), significativement différente
comparée à celles des animaux « NaCl » (-3,1 ± 3,4 μU/ml) ou des animaux « L-

- 81 -
glucose » (10,56 ± 11,41 μU/ml) (Figure 30B). De 3 à 15 minutes, les insulinémies
des trois groupes sont de nouveau similaires.
Figure 30 : Variation de la glycémie (mM) (A) et de l’insulinémie (μU/ml) (B) suite à l’injection intracarotidienne vers le cerveau, au temps 0, de sérum physiologique, de D-glucose ou de L-glucose. Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. *** p < 0,001.
Cette augmentation de l’insulinémie induite par le bolus de D-glucose n’est
pas modifiée par une injection, 10 minutes auparavant, de D-glucose (9 mg/kg) ou
de NaCl 0,9% (Figure 31).
B
A

- 82 -
Figure 31 : Variation de l’insulinémie suite à une deuxième injection de D-glucose (10 minutes après la première). Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne.
Ce protocole d’injection du D-glucose chez le rat éveillé, induit donc une
hyperglycémie uniquement centrale et non périphérique, et il est suivi d’une
augmentation rapide et transitoire de l’insulinémie périphérique. Cet effet est
spécifique au D-glucose et reproductible à 10 minutes d’intervalle.
Remarque : dorénavant, la dénomination « glucose » désignera la forme dextrogyre,
D-glucose.
1.1.2. Mise en évidence des voies nerveuses impliquées
Si on procède à l’injection par voie intraveineuse (veine fémorale) d’atropine
méthylnitrate (antagoniste des récepteurs muscariniques à l’acétylcholine) à une
concentration de 5 mg par kg de poids de l’animal, 20 minutes avant d’injecter le
glucose (injection aux caractéristiques identiques à celles décrites dans le
paragraphe ci-dessus), cette dernière ne conduit plus à l’augmentation de
l’insulinémie périphérique observée chez le groupe de rats témoins recevant comme
substance de prétraitement du sérum physiologique (Figure 32).

- 83 -
Par ailleurs, lorsque l’on injecte par voie intrapéritonéale de l’oxymétazoline
(agoniste partiel des récepteurs 2A-adrénergiques à une concentration de 1000
pmol par kg de poids de l’animal), 5 minutes avant d’injecter le glucose au temps 0,
la réponse insulinémique est toujours présente à 1 minute, bien que d’amplitude
significativement plus faible que celle observée chez le groupe de rats témoins au
même temps (22,65 ± 4,62 versus 39,33 ± 5,88 μU/ml) (Figure 32).
Figure 32 : Variation de l’insulinémie après prétraitement à l’atropine méthylnitrate ou à l’oxymétazoline, en réponse à l’injection vers le cerveau de D-glucose au temps 0. Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. * p < 0,05 ; *** p < 0,001. ip : intrapéritonéal ; iv : intraveineux.
Un traitement à l’atropine (inhibition du SNA parasympathique) bloque
totalement l’hyperinsulinémie induite par le bolus de glucose, à l’inverse de celui à
l’oxymétazoline (activation du SNA orthosympathique) qui n’entraîne qu’une
diminution de l’insulinémie.

- 84 -
1.1.3. Validation de la technique immunohistochimique dirigée contre
l’antigène c-Fos
Cette validation s’est faite à partir d’un modèle déjà connu : l’injection
intrapéritonéale de 2-déoxyglucose (2-DG), analogue du glucose non métabolisé par
les cellules, mimant ainsi une glucopénie cellulaire (Briski KP et Brandt JA, 2000).
L’injection de 2-DG (500 μl d’une solution à 160 mg/ml par voie
intrapéritonéale à des rats éveillés) provoque une forte augmentation du marquage
c-Fos dans les noyaux hypothalamiques supraoptiques et paraventriculaires
(Figure 33A et 33B), tandis que chez les animaux contrôles très peu de cellules
sont immunoréactives (Figure 33C et 33D).
Figure 33 : Validation de la technique utilisant l’immunohistochimie anti-c-Fos : effet d’une injection intrapéritonéale de 2-déoxyglucose versus une injection de sérum physiologique, sur l’expression de c-Fos dans le noyau supraoptique (respectivement A et C) et le noyau hypothalamique paraventriculaire (respectivement B et D). Barres d’échelle = 100 μm ; 3ième V : troisième ventricule.
A
B
C
D

- 85 -
La spécificité de l’immunohistochimie est démontrée par l’absence de
marquage lorsque l’anticorps primaire anti-c-Fos est 1) omis (Figure 34A) ou 2) mis
en présence du peptide immunogène en excès, celui-ci entrant en compétition avec
la protéine endogène (Figure 34B).
Figure 34 : Validation de la technique utilisant l’immunohistochimie anti-c-Fos suite à l’injection intrapéritonéale de 2-déoxyglucose : spécificité du marquage. A : avec anticorps primaire anti-c-Fos (gauche) ou sans anticorps primaire (droite), dans le cortex piriforme. B : avec anticorps primaire anti-c-Fos (gauche) ou avec anticorps primaire en présence du peptide immunogène mis en excès (droite), dans le noyau supraoptique. Barres d’échelle = 100 μm
Ces données montrent que l’activation cérébrale observée par le biais de
l’immunohistochimie c-Fos est spécifique du stimulus créé en accord avec les données
de la littérature, que les anticorps secondaires ne se lient qu’à l’anticorps primaire et
qu’enfin, ce dernier est bien spécifique de son antigène, c-Fos.
A
B

- 86 -
1.2. Régions hypothalamiques activées par l’injection intracarotidienne
de glucose
L’injection aiguë de glucose (9 mg/kg de poids corporel) vers le cerveau via
un cathéter chronique placé dans la carotide, induit une augmentation significative
du nombre de noyaux immunopositifs pour c-Fos dans le noyau arqué (NA) et le
noyau hypothalamique paraventriculaire (PVN) comparé au groupe de rats recevant
du sérum physiologique par la même voie (Figure 35A). Cette augmentation est de
38 % dans le NA (421 ± 33 versus 305 ± 52 noyaux immunopositifs comptés
bilatéralement sur 3 coupes successives) et de 82 % dans le PVN (363 ± 57 versus
200 ± 27 noyaux immunopositifs).
Figure 35 : Effet de l’injection de D-glucose vers le cerveau sur l’expression de c-Fos dans différentes régions hypothalamiques. A : quantification du nombre de noyaux immunopositifs pour c-Fos dans le noyau arqué (NA), les noyaux hypothalamiques paraventriculaire (PVN), ventromédian (VMN) et le cortex cérébral (CC ; contrôle négatif). Les résultats sont exprimés en moyenne du nombre de noyaux positifs pour c-Fos comptés bilatéralement sur 3 coupes distantes les unes des autres de 90 μm ± l’écart type à la moyenne. * p < 0.05. B : illustration d’une immunohistochimie-type au niveau du noyau arqué. Les flèches indiquent des noyaux positifs pour c-Fos. Barre d’échelle = 190 μm ; 3ième V : troisième ventricule.
A
B

- 87 -
D’un point de vue qualitatif, on peut observer au niveau du PVN, un
marquage c-Fos particulièrement présent dans la partie parvocellulaire. En
revanche, il n’existe pas de régionalisation pour le NA, ni de différence
antéropostérieure pour ces deux noyaux hypothalamiques.
La Figure 35B illustre une immunohistochimie-type au niveau du NA après
l’injection intracarotidienne de glucose ou de NaCl 0,9 %.
Dans le noyau hypothalamique ventromédian (VMN), une augmentation de
39 % (même amplitude que pour le NA) du nombre de ces noyaux positifs est
observée suite à l’injection de glucose par rapport à celle du véhicule. Toutefois,
cette différence n’est pas statistiquement significative (165 ± 24 versus 119 ± 9
noyaux immunopositifs).
Enfin, dans le cortex cérébral (CC), région prise comme contrôle négatif
insensible au glucose, aucune différence n’est observée entre les deux groupes de
rats.
L’immunohistochimie c-Fos indique que l’injection aiguë de glucose vers le
cerveau induit spécifiquement l’activation du NA et du PVN. Il apparaît une tendance
à l’activation pour le VMN.
1.3. Types cellulaires activés
Après avoir déterminé les régions sensibles au glucose, nous avons entrepris
la caractérisation des types cellulaires impliqués et ce, grâce à un double marquage
immunohistochimique pour c-Fos et un marqueur cellulaire (CNPase, GFAP ou
NeuN). Les observations se sont tout naturellement focalisées sur le PVN et le NA,
au vu des résultats précédents. La CNPase (2’, 3’-cyclic nucleotide 3’-
phosphodiesterase) est une des enzymes de synthèse de la gaine de myéline. En
conséquence, la révélation de sa présence au niveau cytoplasmique, nous permet de
visualiser le cytoplasme et le début des prolongements des oligodendrocytes. Suite à
l’injection de glucose vers le cerveau, nous n’avons observé aucune cellule
doublement positive c-Fos/CNPase (Figure 36A).
Pour la mise en évidence des neurones, plusieurs anticorps primaires ont été
testés, mais seul celui dirigé contre NeuN (neuronal nuclei) a donné des résultats
exploitables. Il s’agit d’une protéine nucléaire tout comme c-Fos (Mullen RJ et coll.,
1992). Cependant, une amplification du signal NeuN par un système avidine-biotine
révèle une coloration cytoplasmique en plus du marquage nucléaire, créant une
auréole brune de DAB autour du noyau positif pour c-Fos coloré en noir par la

- 88 -
DAB-nickel (Figure 36B). Il apparaît que les cellules doublement immunopositives
c-Fos/NeuN sont en très grand nombre.
Enfin, la GFAP (glial fibrillary acidic protein) est une protéine du
cytosquelette, spécifique des astrocytes. L’immunohistochimie dirigée contre elle
permet d’observer le phénotype en forme étoilée caractéristique de ces cellules
gliales. Le double marquage c-Fos/GFAP révèle qu’il existe quelques cellules
doublement marquées au niveau hypothalamique (Figure 36C). Celles-ci restent
néanmoins minoritaires par rapport aux cellules doublement positives c-Fos/NeuN.
Notre attention s’est portée sur les astrocytes, car selon notre hypothèse de
départ, le glucose injecté en aiguë vers le cerveau serait détecté par GLUT2. Or ce
transporteur au glucose a été préférentiellement retrouvé dans ce type cellulaire en
particulier (Leloup C et coll., 1994).
Les astrocytes immunopositifs pour c-Fos sont le plus souvent observés dans
le NA (Figure 37A) et beaucoup plus rarement dans le PVN (Figure 37B). Nous
avons procédé à la quantification des cellules doublement marquées c-Fos/GFAP
Figure 36 : Caractérisation des cellules immunopositives pour c-Fos suite à l’injection de D-glucose vers le cerveau : double immunohistochimie c-Fos/marqueur cellulaire A c-Fos / CNPase (oligodendrocytes) B c-Fos / NeuN (neurones) C c-Fos / GFAP (astrocytes)
cellules immunonégatives pour c-Fos cellules immunopositives pour c-Fos
Barres d’échelle (A) et (B) = 50 μm, (C) = 25 μm
A B
C

- 89 -
dans ces deux régions cérébrales (Figure 37C) : l’injection aiguë de glucose vers le
SNC induit une augmentation significative du nombre d’astrocytes immunopositifs
pour c-Fos dans le NA, mais pas dans le PVN.
Figure 37 : Effet du glucose sur les astrocytes des noyaux arqué (NA : A) et hypothalamique paraventriculaire (PVN : B). cellules immunonégatives pour c-Fos ; cellules immunopositives pour c-Fos ; Barres d’échelle = 30 μm. C : quantification des astrocytes positifs pour c-Fos. Les résultats sont exprimés en moyenne du nombre de cellules positives pour c-Fos et GFAP comptées bilatéralement sur 8 coupes distantes les unes des autres de 90 �m ± l’écart type à la moyenne. * p < 0.05.
L’injection aiguë de glucose vers le cerveau induit essentiellement
l’activation de neurones hypothalamiques mais aussi de quelques astrocytes. Les
astrocytes immunopositifs pour c-Fos sont significativement plus nombreux dans le
NA par rapport au groupe témoin.
2. Discussion I
2.1. Réponse hormonale périphérique
Le protocole d’injection aiguë de glucose vers le SNC de rat que nous avons
choisi a été précédemment mis en place et plusieurs fois utilisé au sein de notre
A B
C

- 90 -
laboratoire (Atef N et coll., 1995 ; Leloup C et coll., 1998 ; Alquier T et coll., 2003).
La glycémie périphérique n’est pas modifiée, mais elle doit certainement augmenter
au niveau du système sanguin cérébral et notamment au niveau du système porte
hypothalamo-hypohysaire, c'est-à-dire de l’éminence médiane. En raison de
l’absence d’une barrière hémato-encéphalique (BHE) précisément à cet endroit
(l’endothélium y est fenestré), la concentration en glucose doit très
vraisemblablement augmenter dans le parenchyme nerveux du NA. Une
quantification de cette augmentation pourrait être réalisée par microdialyse, mais
cette technique est délicate et, à l’heure actuelle, non maîtrisée pour ce noyau
hypothalamique (Darbin O et coll., 2006). Par ailleurs, la technique des
« biosenseurs » commence à se développer même si elle reste à améliorer
notamment pour doser le glucose. Il s’agit d’une électrode miniaturisée dont la
surface est recouverte par une enzyme catalysant une réaction productrice d’un
gradient électrochimique mesurable et informant alors de la concentration du
composé d’intérêt. Des variations de cette concentration peuvent ainsi être suivies
au cours du temps et sur l’animal éveillé. Dans le cas du glucose, l’enzyme
employée est la glucose oxydase (Dixon BM et coll., 2002).
Quoiqu’il en soit, notre modèle d’injection vers le SNC permet de mettre en
évidence et ce, de manière relativement simple, la sensibilité cérébrale à une
variation de la glycémie grâce à l’augmentation de l’insulinémie périphérique qui est
rapide et transitoire. Cette réponse est spécifique au D-glucose, spécificité attendue
puisque les transporteurs au glucose (GLUTs) ne prennent pas en charge le L-
glucose (Medina RA et Owen GI, 2002). L’utilisation de la forme lévogyre (Figure
30), ne provoquant pas la réponse insulinémique, montre ainsi que le résultat
obtenu avec le D-glucose est bien sélectif, ne provient pas de la détection d’un
hexose quelconque ou d’un encombrement moléculaire. A noter qu’au cours de
l’année de DEA, le D-mannitol avait été utilisé pour le groupe témoin comme dans
les travaux de l’équipe de B. Levin (Dunn-Meynell AA et coll., 1997). Cet alcool de
sucre à 6 carbones possédant le même poids moléculaire, était supposé n’avoir
aucun effet biologique central et constituait donc un bon contrôle iso-osmotique.
Or, de manière inattendue, son injection aiguë vers le cerveau induit aussi une
augmentation de l’insulinémie (résultats non montrés). Finalement, devant cette
difficulté imprévue, nous avons opté pour une solution contrôle « classique », le
sérum physiologique, tout en contrôlant l’osmolarité de la solution de glucose
injectée par la carotide (solution elle-même préparée avec du NaCl).

- 91 -
Nous avons pu également constater lors de cette première expérience, que la
réponse insulinémique persistait même après une première injection aiguë de
glucose 10 min avant celle du temps 0, avec exactement les mêmes caractéristiques
temporelles et d’intensité (Figure 31). Ceci indique, que le SNC est capable de
détecter deux augmentations successives de concentration en glucose et montre à
cet égard une haute réceptivité. Il serait néanmoins intéressant de regarder si des
stimulations répétées au cours du temps n’entraînent pas l’apparition de
phénomènes de potentialisation ou de désensibilisation.
Les résultats obtenus suite à l’inhibition des voies nerveuses
parasympathiques et à l’activation des voies nerveuses orthosympathiques (Figure
32), indiquent clairement que le lien entre le SNC qui détecte l’augmentation de la
glycémie et le pancréas qui secrète en réponse de l’insuline, est le SNA. Si l’activité
du SNA est modifiée de manière pharmacologique, la réponse au glucose est altérée.
Comme montré par d’autres équipes, on observe que le système parasympathique
stimule la sécrétion des cellules β du pancréas, tandis que le système
orthosympathique exerce une influence inhibitrice (Rohner-Jeanrenaud F et coll.,
1983). De même, chez les rats obèses Zucker qui sont hyperinsulinémiques et
insulino-résistants, l’activité basale du parasympathique est augmentée,
simultanément à une baisse de celle de l’orthosympathique, et ceci est corrélé à une
perturbation de la réponse du pancréas à l’injection de glucose, indiquant une
possible cause de leur phénotype diabétique (Atef N et coll., 1995). De plus, ces
animaux révèlent une hypersensibilité cérébrale au glucose puisque son injection
intracarotidienne à une concentration de 3 mg/kg au lieu de 9 suffit à déclencher
l’augmentation de l’insulinémie (Alquier T et coll., 2003). Enfin, il s’avère que le
dysfonctionnement de l’activité du SNA apparaît très tôt au cours du
développement de l’obésité et pourrait être une des causes principales de cette
pathologie (Pénicaud L et coll., 1996).
Ainsi, l’ensemble de ces données indiquent que le SNC détecterait une
augmentation de la concentration en glucose au niveau de l’artère carotidienne, ce qui
induirait, après un certain nombre de relais nerveux qu’il reste à déterminer, une
stimulation du système nerveux parasympathique simultanée à l’inhibition de
l’orthosympathique aboutissant finalement à l’activation des cellules β du pancréas et
à la sécrétion d’insuline (Figure 38 ).

- 92 -
2.2. Réponse hypothalamique
Afin d’étudier l’activation cérébrale conséquente à l’injection aiguë de glucose
vers le cerveau, nous avons utilisé comme outil, l’expression du proto-oncogène c-
fos, caractéristique de la stimulation des neurones. La validité de cette technique a
été démontrée grâce à la reproduction de résultats publiés à propos de l’activation
cérébrale provoquée par l’injection intrapéritonéale de 2-DG comparée à celle du
NaCl (Briski KP et Brandt JA, 2000). Ce modèle a été choisi plutôt qu’un autre, car
ce composé a un lien relativement direct avec la sensibilité au glucose qui nous
intéresse. En effet, il s’agit d’un analogue non métabolisable du glucose, mimant
une glucopénie cellulaire et donc un état d’hypoglycémie. On pouvait donc
supposer, que notre propre protocole toucherait les mêmes régions cérébrales. Nos
résultats sont en parfait accord avec ceux publiés : le 2-DG induit bien l’activation,
dans l’hypothalamus, des noyaux supraoptique et paraventriculaire (Figures 33 et
34), structures impliquées entre autre dans l’homéostasie énergétique en particulier
par l’expression de l’ocytocine, neuropeptide anorexigène, et de la présence au
niveau du PVN des neurones cibles des projections NPY et POMC issues du NA
(Briski KP et Brandt JA, 2000) (Tableau 1 et figure 13).
L’expression de c-fos est induite par une très grande variété de stimuli ce qui
sous-entend quelques précautions à respecter. En particulier, l’anesthésie ou la
chirurgie doivent être proscrites et les phénomènes de stress les plus limités
possibles (Marota JJA et coll., 1992). C’est pourquoi, contrairement aux précédents
protocoles d’injection aiguë de glucose vers le cerveau réalisés par notre équipe, le
cathéter a été placé dans la carotide des rats une semaine avant le jour de
l’expérience per se. Le fait de devoir le rincer quotidiennement a permis d’habituer
les animaux à la manipulation et à une pseudo injection à l’état de vigilance. En
effet, nos résultats montrent, que chez le groupe contrôle il n’existe que quelques
noyaux immunopositifs pour c-Fos. Cette expression peut être due à la
manipulation malgré tout, à un changement dans l’environnement imperceptible à
l’humain, ou à l’expression basale du proto-oncogène. Celle-ci ne peut être
totalement absente puisqu’il s’agit d’un facteur de transcription intervenant dans la
vie normale d’une cellule. Par rapport aux neurones, cette protéine régulerait
notamment l’expression des neurotransmetteurs ou celle de leurs enzymes de
synthèse, or les cellules nerveuses sont souvent sollicitées (Hoffman GE et coll.,
1993). Toutefois, la majorité des cellules n’exprime pas c-Fos en conditions basales
comme nous l’observons.

- 93 -
Suite à l’injection aiguë de glucose vers le SNC, nous observons une
augmentation significative du nombre de noyaux immunopositifs pour c-Fos dans
le PVN et le NA (Figure 35). Cette spécificité est démontrée par l’absence
d’augmentation du nombre de cellules exprimant c-Fos dans le cortex cérébral, pris
comme région insensible au glucose, et chez les rats recevant une solution de NaCl.
La stimulation observée est en accord avec les données de la littérature, puisque le
PVN et le NA sont bien connus pour leur implication dans la régulation de
l’homéostasie énergétique (Morton GJ et coll., 2006). De plus, ils contiennent des
neurones sensibles au glucose (Levin BE et coll., 2002).
Avec la technique utilisant c-Fos, on ne peut savoir précisément si les
noyaux hypothalamiques sont directement activés par le glucose, ou indirectement
via l’activation d’un autre noyau situé en amont. Ainsi, nos expériences ne nous
permettent pas de conclure si l’activation du NA et celle du PVN sont indépendantes
l’une de l’autre ou si l’une entraîne l’autre. Néanmoins, comme nous l’avons vu
dans l’introduction (cf. § II-2), le NA a un rôle primordial en assurant l’intégration
des signaux périphériques liés au statut énergétique de l’organisme, cette structure
étant sensible aux concentrations hormonales circulantes (leptinémie, insulinémie)
et possédant deux populations de neurones aux effets opposés mais
complémentaires orexigènes/anorexigènes (Niswender KD et coll., 2004) (Figure
11). Ainsi, il permet la transformation des signaux endocriniens ou métaboliques en
signaux électriques. En raison de l’absence de BHE au niveau de l’éminence
médiane, il ne serait pas étonnant que le NA puisse intégrer très rapidement des
variations de la glycémie et être ainsi le premier noyau hypothalamique activé par
notre bolus intracarotidien de glucose. Grâce à ses très nombreuses projections,
l’information « glucose » initiée dans le NA, va donc pouvoir être acheminée vers
d’autres régions intra et/ou extra-hypothalamiques (Morton GJ et coll., 2006). Un
grand nombre des neurones du NA émettent des prolongements en direction du
PVN, comme les neurones orexigènes et anorexigènes à NPY et à POMC (Woods SC,
2005). Bien que cela reste à démontrer, on peut donc supposer que l’activation du
PVN par notre bolus de glucose est secondaire à celle du NA.
Une partie des neurones du PVN projette vers le tronc cérébral, permettant le
relais de l’information nerveuse vers la périphérie grâce au SNA (Swanson LW et
Sawchenko PE, 1980 ; Levin BE et coll., 2002). Le contrôle de l’activité du SNA
permet en particulier la modulation de l’activité des cellules β du pancréas (Rohner-
Jeanrenaud F et coll., 1983). Ceci suggère que la stimulation, directe ou indirecte,

- 94 -
du PVN par le glucose aboutirait à la sécrétion d’insuline en périphérie grâce au
SNA, comme observée au cours de notre première série de manipulations.
Par ailleurs, on aurait pu s’attendre à l’activation d’autres régions
hypothalamiques possédant aussi des connexions nerveuses avec le NA, connues
pour leur implication dans la régulation de la balance énergétique et pour la
présence de neurones sensibles au glucose, comme le LH, le DMN ou le VMN (Levin
BE et coll., 2002 ; Kalra SP et coll., 1999). Avec notre modèle d’injection, dans le LH
et le DMN, on n’observe pas de changement particulier dans le nombre de noyaux
marqués (résultats non montrés). Dans le VMN, on peut noter une tendance à
l’augmentation (Figure 35). En terme de pourcentage, elle est de 39 % c'est-à-dire
identique à celle obtenue pour le NA. Même si on ne peut vraiment conclure sur
l’implication de ce noyau dans la sensibilité au glucose, certains travaux de
recherches la suggèrent fortement. Notamment, une perfusion intracarotidienne de
glucose d’1 h induit une augmentation du nombre de noyaux positifs pour c-Fos
dans le PVN et le VMN (Dunn-Meynell AA et coll., 1997). Cette même équipe a
démontré que ce dernier renfermait des neurones qui pouvaient être excités ou
inhibés, de manière directe ou indirecte, par une augmentation de la concentration
extracellulaire de glucose (Song Z et coll., 2001). Ainsi, comme le PVN, le VMN
pourrait être activé par une hyperglycémie de manière directe et/ou indirecte.
Par ailleurs, il est curieux de constater que le protocole de perfusion
intracarotidienne de glucose de longue durée (1 heure) mentionné plus haut, s’il
induit une augmentation de l’expression de c-Fos dans les PVN et VMN, ne le fait
pas dans le NA (Dunn-Meynell AA et coll., 1997). Cette différence de résultats avec
les nôtres provient certainement du fait qu’il s’agit d’une injection chronique et non
aiguë de glucose. On peut aussi émettre l’hypothèse que la protéine c-Fos dans le
NA a eu le temps d’être dégradée avant le sacrifice des animaux. Au vu du rôle
central de cette structure cérébrale, la demi-vie des facteurs de transcription y est
peut-être plus courte pour permettre un traitement adéquat des signaux
périphériques.
Enfin, on ne peut exclure une possible activation de structures du tronc
cérébral, comme le NTS ou l’area postrema. Comme pour les PVN et VMN, elle
pourrait être indirecte, via les fortes connexions réciproques qui existent avec
l’hypothalamus, ou directe car l’endothélium y est aussi fenestré et les neurones
possèdent un équipement moléculaire adéquat pour une fonction régulatrice de la
balance énergétique (Wynne K et coll., 2005 ; Schwartz MW et coll., 2000). De plus,
des neurones sensibles au glucose sont présents dans cette région comme le

- 95 -
démontrent des enregistrements électrophysiologiques (Himmi T et coll., 2001).
Finalement, on ne peut exclure que l’activation cérébrale que nous observons soit
due à une détection de l’augmentation de la glycémie directement au niveau de la
carotide. En effet, une telle sensibilité a déjà été décrite (Alvarez-Buylla R et de
Alvarez-Buylla ER, 1988 ; Pardal R et Lopez-Barneo J, 2002). Cependant, la
probabilité qu’elle active exactement les mêmes régions hypothalamiques est faible
d’autant plus que pour le premier article, une augmentation de la concentration en
glucose induit une diminution des influx nerveux générés par les chimiorécepteurs
(ce qui irait plutôt dans le sens d’une inhibition cérébrale), et pour le second il s’agit
de la détection d’une hypoglycémie.
Nos résultats indiquent que l’injection aiguë de glucose vers le SNC, induit
l’activation du NA, ainsi que celle du PVN, de manière directe et/ou indirecte.
L’information nerveuse pourrait alors être transmise au tronc cérébral permettant la
modulation de l’activité du SNA, aboutissant à la sécrétion d’insuline par les cellules
β du pancréas (Figure 38 ).
2.3. Réponses cellulaires
Les résultats des doubles immunohistochimies montrent que les cellules
activées par le bolus de glucose, c'est-à-dire exprimant c-Fos, sont en très grande
majorité des neurones, et quelques unes des astrocytes (Figures 36 et 37).
Le fait que des neurones soient stimulés est tout à fait logique puisque ces
cellules permettent la « traduction » de la majeure partie des signaux intrin- ou
extrinsèques en information électrique. De plus, il est clair qu’il existe une
population de neurones sensibles au glucose, c'est-à-dire de neurones dont l’activité
électrique est modifiée par une augmentation ou une diminution de sa
concentration (Levin BE et coll., 2002 ; Fioramonti X et coll., 2004). Il est
vraisemblable que les neurones du NA soient les premiers à exprimer c-Fos en
réponse à notre protocole d’injection aiguë. Une fois activés, ils doivent stimuler à
leur tour d’autres neurones auxquels ils sont connectés, ce qui aboutit à la
propagation de l’activation par le glucose. Néanmoins, certaines précautions doivent
être prises. D’une part, puisque l’expression de c-Fos n’est pas systématiquement
induite par un stimulus, il ne peut être exclu que certains neurones sensibles au
glucose ne soient pas révélés par l’expression de c-Fos (Sagar SM et coll., 1988).
D’autre part, la présence de cette protéine dans un neurone donné, ne nous permet
pas de savoir si celui-ci est de type GE ou GI (excité ou inhibé par le glucose,

- 96 -
Figure 24). En effet, elle n’est pas systématiquement associée à une activation
électrique (dépolarisation) de cette cellule (Sabatier N et coll., 2003). Enfin, on sait
que c-Fos peut former différents hétérodimères avec d’autres produits de gènes
précoces comme ceux de la famille Jun (Hoffman GE et coll., 1993). Or, selon le
couple formé, il aura un effet stimulateur ou inhibiteur sur la transcription des
gènes cibles, en particulier ceux codant pour les neuropeptides (Hoffman GE et
coll., 1993). Ceci peut laisser suggérer que les neurones marqués du NA peuvent
tout aussi bien être à POMC/CART (probablement stimulés) et à NPY/AGRP
(probablement inhibés). Toutefois, des travaux récents menés dans notre
laboratoire montrent que les neurones à POMC, du moins chez la souris, sont
négatifs pour c-Fos suite à l’injection de glucose et ne sont pas GE d’un point de
vue électrophysiologique, alors que les neurones GI expriment le NPY (Fioramonti et
coll., 2007).
De manière intéressante, nous observons quelques astrocytes positifs pour c-
Fos. Ils sont localisés dans le PVN mais surtout dans le NA. Dans ce dernier,
l’injection aiguë de glucose induit leur augmentation de façon significative par
rapport aux rats recevant le sérum physiologique. Jusqu’à présent, aucun article ne
mentionnait la capacité des cellules gliales à exprimer cette protéine en réponse à
un stimulus.
Parmi les cellules gliales, une population particulière se démarque par
rapport à la sensibilité cérébrale au glucose : les épendymocytes. En effet, dans
l’introduction (cf. § III-2.2.4) nous avons vu que la plupart possède le couple
protéique GLUT2/GK (indispensable à la sensibilité au glucose pour les cellules β
du pancréas), mais aussi qu’ils pouvaient être sensibles à des changements de la
concentration extracellulaire en glucose (Leloup C et coll., 1994 ; Maekawa et coll.,
2000 ; Moriyama R et coll., 2004). Toutefois, sur nos coupes, nous n’observons pas
d’expression de c-Fos dans ce type cellulaire. Il est possible que notre protocole
d’injection soit trop rapide pour entraîner une modification de la concentration en
glucose dans le liquide céphalorachidien (LCR). En effet, les épendymocytes bordent
le troisième ventricule au niveau de l’hypothalamus et sont donc très
vraisemblablement sensibles aux variations de composition du LCR (Moriyama R et
coll., 2004).
Au niveau du NA, les épendymocytes présentent certaines particularités : on
parle alors de tanycytes. Leurs corps cellulaires bordent la base du troisième
ventricule, tandis que de longs prolongements atteignent l’éminence médiane et le

- 97 -
lit capillaire du système porte, ce qui forme un lien entre ces deux zones. Par ces
caractéristiques anatomiques, ils sont en contact permanent à la fois avec le LCR et
le sang, et sont donc à même de pouvoir détecter une modification de la
concentration en glucose dans ces deux compartiments. Cela rappelle curieusement
le mécanisme de sensibilité au glucose présente au niveau de la veine hépatoportale
dont le principe est basé sur une différence de concentration glucidique entre la
veine porte et l’artère hépatique (Burcelin R et Pénicaud L, 2003) (cf. § III-2.2.1). De
plus, ces cellules gliales contiennent des protéines, la sous-unité Kir6.1 du canal
potassique dépendant de l’ATP, GLUT2 et la GK, connues pour leur implication
dans la sensibilité au glucose des cellules β du pancréas (Garcia MA et coll., 2003).
Les neurones du NA sont pour la plupart au contact de ces cellules. Ces dernières
n’expriment pas la GFAP, protéine du cytosquelette servant à identifier les
astrocytes (Garcia MA et coll., 2003). De ce fait, ils n’ont pas été comptabilisés lors
du double marquage cFos/GFAP.
Ainsi, le bolus de glucose vers le cerveau induit majoritairement l’activation
de neurones, mais aussi de quelques astrocytes, ces derniers localisés dans le NA
(Figure 38 ).

- 98 -
Figure 38 : Boucle de contrôle mise en place entre la détection cérébrale du glucose et la sécrétion d’insuline par le pancréas. renvoient aux conclusions des trois paragraphes de discussion ; β : cellules β ; NA : noyau arqué ; PVN : noyau hypothalamique paraventriculaire ; SNA : système nerveux autonome ; SNC : système nerveux central.

- 99 -
XI – Implication des astrocytes dans la sensibilité au glucose
Après avoir montré qu’un bolus de glucose vers le SNC induisait l’activation
du PVN et du NA, et que le nombre d’astrocytes stimulés était significativement plus
grand dans ce dernier noyau par rapport aux rats « contrôle », nous avons cherché
à déterminer l’implication de ce type cellulaire.
La glutamine synthase (GS) est l’enzyme qui catalyse la synthèse de
glutamine à partir du glutamate (Figures 27 et 39). Cette réaction est spécifique
des astrocytes dans le SNC. La méthionine sulfoximine (MSO), un analogue du
glutamate, inhibe cette enzyme et a déjà été utilisée par d’autres équipes comme
inhibiteur astrocytaire (Young JK et coll., 2000).
Figure 39 : Schéma hypothétique de l’action de la méthionine sulfoximine sur le couplage astrocyte-neurone. GS : glutamine synthase ; MSO : méthionine sulfoximine.
En bloquant la GS, il est probable que la MSO empêche le recyclage du
glutamate libéré (comme neurotransmetteur) par une grande population de
neurones. En effet, une fois libéré au niveau synaptique, celui-ci est pris en charge

- 100 -
par l’astrocyte qui le transformera en glutamine, elle-même relibérée dans le milieu
extracellulaire et servant de précurseur à une nouvelle synthèse de glutamate par
les neurones glutamatergiques (Bouzier-Sore AK, et coll., 2002). Cette hypothèse
suggère donc une interruption du couplage astrocyte-neurone. Ainsi la MSO a été
utilisée dans le but de déterminer si les astrocytes étaient effectivement
primordiaux à la sensibilité neuronale au glucose.
1. Résultats II
1.1. Validation de l’utilisation de la méthionine sulfoximine
Les doses choisies ont été celles utilisées par Young et ses collaborateurs
(Young JK et coll., 2000). Un moyen de vérifier l’efficacité du protocole d’injection
(prétraitement de 18 h à 100 mg/kg de poids corporel, par voie intrapéritonéale) a
été de mesurer les variations de glycogène ainsi induites et précédemment décrites
chez la souris (Hévor TK et coll., 1985). En effet, si la MSO inhibe la GS, aucune
accumulation de glutamate n’a été observée. D’après ces auteurs, le glutamate
serait converti en glucose via la gluconéogenèse. Le glucose emprunterait alors la
voie de la glycolyse et celle de la glycogenèse conduisant à une accumulation de
glycogène (Hévor TK et coll., 1985) (Figure 39).
Suite à un prétraitement de 18 h à la MSO, on observe dans le foie une
diminution de 58% (soit un facteur de 2,4) de la concentration en glucose provenant
du glycogène par rapport au groupe de rats recevant du sérum physiologique (2,38
± 1,21 versus 5,71 ± 1,24 μg/mg de tissu) (Figure 40A).
En revanche, cette même injection est suivie dans le cortex cérébral d’une
augmentation de 414% (soit un facteur de 4) de la concentration en glucose
provenant du glycogène par rapport au groupe de rats témoins (0,41 ± 0,18 versus
0,10 ± 0,03 μg/mg de tissu) (Figure 40B).
Néanmoins, ces variations ne sont pas statistiquement significatives, mais
ces fortes tendances observées vont dans les mêmes sens que ceux décrits par
l’équipe du Pr. Hévor (Hévor TK et coll., 1985).

- 101 -
Figure 40 : Variation de la quantité de glycogène dans le foie (A) et le système nerveux central (B) suite à un prétraitement intrapéritonéal à la méthionine sulfoximine (MSO). Les résultats sont exprimés en moyennes ± l’écart type à la moyenne (n=4 pour les deux groupes).
La MSO, injectée selon notre protocole, donne bien les résultats escomptés
en accord avec la figure 39, à savoir une accumulation de glycogène cérébral.
1.2. Conséquences de l’arrêt du métabolisme astrocytaire des
carbohydrates :
1.2.1. sur l’activation cérébrale induite par le bolus intracarotidien de
glucose
Suite à un prétraitement de 18 h à la MSO par voie intrapéritonéale, nous
avons procédé à l’injection intracarotidienne de glucose selon notre modèle et à la
quantification des noyaux immunopositifs pour c-Fos dans les mêmes régions
cérébrales que précédemment (cf. § X-1.2, figure 35A) (Figure 41).
B
A

- 102 -
Figure 41 : Effet d’un prétraitement intrapéritonéal (ip) à la méthionine sulfoximine (MSO) sur l’expression de c-Fos induite par l’injection de D-glucose vers le cerveau : quantification du nombre de noyaux immunopositifs pour c-Fos dans le noyau arqué (NA), les noyaux hypothalamiques paraventriculaire (PVN), ventromédian (VMN) et le cortex cérébral (CC ; contrôle négatif). Les résultats sont exprimés en moyennes du nombre de noyaux positifs pour c-Fos comptés bilatéralement sur trois coupes ± l’écart type à la moyenne. * p < 0,05 ; ** p < 0,01 ; *** p < 0,005 ; NS : non significatif (p > 0,05).
Au niveau du NA, on peut observer que les rats recevant un prétraitement à
la MSO avant l’injection de glucose par la carotide ne montrent plus d’augmentation
du nombre de noyaux immunopositifs pour c-Fos contrairement à ce qui avait été
observé pour les rats non prétraités (263 ± 26 versus 421 ± 33 noyaux
immunopositifs) (Figure 41A barres rouge et jaune). Notons que le prétraitement à
la MSO en lui-même n’a pas d’effet sur l’expression de c-Fos dans cette région
hypothalamique : il n’existe pas de différence significative entre les deux groupes de
rats recevant du sérum physiologique vers le cerveau au temps 0, avec ou sans
prétraitement à la MSO (305 ± 52 versus 256 ± 34 noyaux immunopositifs) (Figure
41A barres grise et rose).
Dans le PVN, le prétraitement à la MSO ne modifie pas le nombre de noyaux
immunopositifs pour c-Fos quantifiés après l’injection de glucose (439 ± 55 versus
B A

- 103 -
363 ± 57 noyaux immunopositifs). En revanche, il l’augmente de manière
significative dans le VMN (239 ± 24 versus 165 ± 24 noyaux immunopositifs)
(Figure 41B barres rouge et jaune). Cependant, dans ces deux régions
hypothalamiques ainsi que dans le CC, l’injection intrapéritonéale de MSO,
précédant l’injection intracarotidienne de NaCl 0,9% induit l’expression de c-Fos de
manière significative par rapport au groupe ne recevant que le NaCl 0,9% au temps
0 (PVN : 453 ± 68 versus 200 ± 27 ; VMN : 308 ± 47 versus 119 ± 9 ; CC : 667 ± 132
versus 299 ± 56 noyaux immunopositifs) (Figure 41B barres rose et grise).
Un prétraitement à la MSO supprime l’activation du NA normalement induite
par l’injection aiguë de glucose vers le cerveau.
1.2.2. sur la sécrétion d’insuline
L’injection aiguë de glucose vers le cerveau induit non seulement une
augmentation du nombre de noyaux immunopositifs pour c-Fos mais aussi
l’élévation de l’insulinémie périphérique de manière rapide et transitoire en 1
minute (cf. § X-1.1.1, figure 30B). Puisque le prétraitement avec la MSO supprime
l’activation cérébrale du NA (cf. § XI-1.2.1), nous avons voulu regarder ce qu’il en
était au niveau de la réponse périphérique au bolus intracarotidien de glucose.
On observe que le prétraitement en intrapéritonéal à la MSO empêche
l’apparition de l’hyperinsulinémie périphérique au temps 1 minute, normalement
induite par l’injection seule de glucose vers le cerveau (6,5 ± 4,9 versus 32,2 ± 6,1
μU/ml) (Figure 42A).
Notons que les rats prétraités à la MSO montrent une faible, mais
significative, diminution du delta glycémique au temps 1 minute par rapport aux
deux autres groupes, « NaCl » et « glucose » seul (-0,4 ± 0,09 versus 0,1 ± 0,07 et 0,1
± 0,16 mM) (Figure 42B).

- 104 -
Figure 42 : Effet d’un prétraitement intrapéritonéal (ip) à la méthionine sulfoximine (MSO) sur la sécrétion d’insuline induite par l’injection de D-glucose vers le cerveau au temps 0 (A), ainsi que sur la glycémie (B). Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. ** p < 0,01.
Le prétraitement à la MSO empêche l’augmentation de l’insulinémie
normalement induite par l’injection aiguë de glucose vers le cerveau.
B
A

- 105 -
1.3. Spécificité de la méthionine sulfoximine
Un problème inhérent à l’utilisation de n’importe quel agent pharmacologique
est celui de sa spécificité. C’est pourquoi nous avons mené des expérimentations
supplémentaires, afin d’invalider au mieux ce doute.
1.3.1. Effet uniquement cérébral ou aussi périphérique ?
Comme la MSO est injectée par voie intrapéritonéale, il doit être envisagé la
possibilité que l’effet observé soit aussi dû à une action directe sur le pancréas,
c'est-à-dire que celui-ci serait, en fin de compte, incapable de secréter de l’insuline
correctement. Pour résoudre ce problème, nous avons réalisé un test de tolérance
au glucose injecté par voie intraveineuse (0,8 g/kg de poids corporel) sur des
animaux traités avec la MSO.
Tableau 2 : Effet d’un prétraitement intrapéritonéal de 18 heures de méthionine sulfoximine (MSO) sur un test de tolérance au glucose (0,8 g/kg de poids corporel) injecté par voie intraveineuse. Les ΔG et ΔI expriment les aires sous la courbe. Les valeurs correspondent aux moyennes ± l’écart type à la moyenne. * p < 0,05.
Au temps 0, les valeurs basales des glycémie et insulinémie ne sont pas
différentes entre les rats prétraités avec du sérum physiologique et ceux prétraités
avec de la MSO (Tableau 2).
L’injection intraveineuse aiguë de la solution riche en glucose entraîne une
augmentation de la glycémie périphérique qui retourne secondairement à son
niveau basal sous l’effet de l’insuline. L’hyperglycémie, mesurée sur 30 minutes, est
significativement plus importante pour le groupe de rats prétraités à la MSO par
rapport à celle obtenue pour les rats témoins (141,6 ± 21,8 versus 87,7 ± 8,6
mM/30 min). Néanmoins, il n’y a pas de différence significative lorsqu’on regarde la
quantité d’insuline secrétée sur cette même période (810,7 ± 161,1 pour les rats
prétraités au NaCl 0,9% et 868,9 ± 216,7 μU/ml/30 min pour les rats prétraités à
la MSO).

- 106 -
Ce résultat démontre que, même avec un prétraitement à la MSO, le
pancréas est toujours capable de secréter une quantité normale d’insuline en réponse
à une hyperglycémie.
Par ailleurs, quelques expériences d’injection de MSO par voie
intracérébroventriculaire (25 μg en μl, dans le troisième ventricule, au niveau de
l’hypothalamus, 2 heures avant le bolus de glucose vers le cerveau) confortent cette
conclusion. En effet, ce prétraitement cérébral et plus court dans le temps, aboutit
à une diminution significative de l’hyperinsulinémie périphérique apparaissant au
temps 1 minute (83,5 ± 5,2 μU/ml pour les 6 rats prétraités à la MSO versus
122,57 ± 13,2 μU/ml pour les 7 rats prétraités au sérum physiologique, p=0,0182,
résultat non montré). Ainsi, la seule inhibition de la GS cérébrale suffit à observer
une perte de la sensibilité au glucose injecté vers le SNC.
1.3.2. Effet spécifique à l’inhibition de la glutamine synthase ?
Il a été décrit dans la littérature, qu’en plus d’inhiber la glutamine synthase,
la MSO inhibait aussi la γ-glutamylcystéine synthase. Cette enzyme intervient dans
la synthèse du glutathion (Cooper AJL et coll., 1976). On peut alors légitimement
s’interroger sur le fait que l’effet observé provient bien uniquement de l’interruption
du métabolisme astrocytaire des carbohydrates et non de l’inhibition de la synthèse
de glutathion ou du cycle du γ-glutamyl.
Afin d’invalider cette hypothèse nous avons procédé à un prétraitement (aux
paramètres identiques que précédemment avec la MSO) des animaux avec de la
buthionine sulfoximine (BSO) analogue de la MSO. La BSO a été décrite comme
inhibant la γ-glutamylcystéine synthase environ cent fois plus que la MSO, mais
pas la GS de manière détectable (Griffith OW et Meister A, 1979).
Contrairement à ce qui avait été observé avec le prétraitement à la MSO,
celui avec la BSO n’empêche pas l’augmentation de l’insulinémie au temps 1 min
comme observée pour le groupe de rats témoins (63,65 ± 4,75 versus 50,96 ± 4,32
versus 6,5 ± 4,90 μU/ml) (Figure 43A).
En ce qui concerne la glycémie, l’injection par voie intrapéritonéale de BSO
18 h avant l’injection aiguë de glucose vers le cerveau n’a pas d’effet : les deltas ne
sont pas différents de ceux du groupe de rats témoins prétraités au sérum
physiologique (Figure 43B).

- 107 -
Figure 43 : Effet d’un prétraitement intrapéritonéal (ip) à la buthionine sulfoximine (BSO) versus à la méthionine sulfoximine (MSO) sur la sécrétion d’insuline induite par l’injection de D-glucose vers le cerveau au temps 0 (A), ainsi que sur la glycémie (B). Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. *** p < 0,005.
Ainsi, l’effet inhibiteur de la MSO sur l’augmentation de l’insulinémie induite
par l’injection du bolus de glucose vers le cerveau est bien spécifique à l’inhibition de
la GS.
B
A

- 108 -
2. Discussion II
L’implication des astrocytes dans la sensibilité au glucose est appuyée par le
fait que ces cellules sont les seules capables, dans le SNC, de stocker cet hexose
sous forme de glycogène (Kong J et coll., 2002). Ainsi, elles doivent être à même de
pouvoir détecter des modifications de sa concentration puisqu’elles gèrent ses
réserves énergétiques. Dans le but de clarifier le rôle des astrocytes dans ce
mécanisme, nous avons utilisé un inhibiteur de la glutamine synthase (GS), enzyme
permettant la synthèse de glutamine à partir du glutamate, ceci spécifiquement
dans ce type cellulaire au niveau central (Young JK et coll., 2000). Cet inhibiteur
est la méthionine sulfoximine (MSO), précédemment utilisée par une autre équipe
pour un objectif similaire mais dans un contexte hypoglycémique (Young JK et coll.,
2000). C’est pourquoi nous avons choisi un protocole de traitement identique.
D’après le schéma du couplage métabolique qui existerait entre les astrocytes et les
neurones, l’inhibition de la GS provoquerait son interruption puisque le glutamate
ne serait plus recyclé en glutamine (Tsacopoulos M et Magistretti PJ, 1996) (Figure
39).
Une des caractéristiques phénotypiques de l’administration de MSO, est
l’accumulation de glycogène astrocytaire. Son dosage dans le SNC montre que, suite
à notre protocole d’injection consistant en un prétraitement intrapéritonéal de 18 h
à 100 mg/kg, sa concentration quadruple (Figure 40). Cette variation n’est pas
statistiquement significative, ainsi que la diminution d’un facteur de 2,5 de la
quantité de glycogène hépatique. Toutefois, ces deux tendances sont similaires à ce
qui est décrit dans la littérature que ce soit dans le sens et l’amplitude (Hévor TK et
coll., 1985). La MSO inhibant la GS, le glutamate devrait s’accumuler dans le SNC.
Or, il n’en est rien d’où l’hypothèse selon laquelle, cet acide aminé serait converti
par néoglucogenèse en glucose, lui-même utilisé pour la glycogénogenèse (Hévor TK
et coll., 1985).
Utiliser la MSO n’est pas négligeable pour l’expression de c-Fos, puisque le
glutamate est un des neurotransmetteurs majeurs du cerveau. En effet, le nombre
de cellules immunopositives pour c-Fos est très augmenté dans les noyaux PVN et
VMN par cette drogue seule (Figure 41). Néanmoins, une augmentation similaire
dans le cortex cérébral suggère que ceci n’est pas spécifique à l’hypothalamus. Par
conséquent, il nous est impossible de conclure sur le rôle des astrocytes dans la
détection du glucose au niveau du PVN.

- 109 -
Dans le NA, nos résultats indiquent que le traitement à la MSO supprime
l’augmentation du nombre de noyaux positifs pour c-Fos en réponse au bolus de
glucose. Ainsi, dans cette structure hypothalamique, le couplage métabolique entre
les astrocytes et les neurones semble être important, sinon indispensable, à la
détection d’un changement de la concentration cérébrale en glucose. Les travaux
menés par l’équipe de Bernard Thorens vont aussi dans le sens d’une implication
des astrocytes (Marty N et coll., 2005). Des souris dépourvues de GLUT2, mais dont
le gène a été réinséré spécifiquement dans ce type cellulaire, sont de nouveau
sensibles à l’injection intrapéritonéale de 2-DG (150 mg/kg), comme le montrent la
sécrétion périphérique de glucagon et l’induction de l’expression de c-Fos au niveau
du tronc cérébral.
Selon notre hypothèse, certains astrocytes seraient les « détecteurs » du
glucose, en relation directe avec les neurones adjacents qui seraient les
« effecteurs ». En raison des nombreuses jonctions communicantes qui existent
entre les astrocytes, un petit nombre d’astrocytes « détecteurs » suffirait pour
étendre l’information à l’ensemble de la population astrocytaire du NA et de là, aux
neurones sensibles au glucose.
Le couplage astrocyte-neurone peut être de différentes natures. Il pourrait
mettre en jeu une synthèse astrocytaire de composés qui seraient ensuite libérés et
agiraient sur les neurones voisins. On peut naturellement penser au glutamate,
mais aussi à l’ATP ou au lactate (Fields RD et Stevens-Graham B, 2002). En ce qui
concerne le glutamate, on sait depuis peu, que les astrocytes sont capables de le
sécréter directement par des processus d’exocytose calcium-dépendante (Fields RD
et Stevens-Graham B, 2002). Des techniques d’imagerie ont aussi permis de
montrer qu’une augmentation de la concentration intracellulaire en calcium dans
ces cellules gliales pouvait être accompagnée d’une augmentation de la
concentration extracellulaire d’ATP (Fields RD et Stevens-Graham B, 2002). Pour le
lactate, il a été démontré par électrophysiologie que son application était capable de
stimuler les neurones répondants aux variations de glucose extracellulaire (Yang XJ
et coll., 1999). Et comme nous l’avons vu dans l’introduction (figure 27), ce
composé pourrait dériver de la glycolyse astrocytaire, sa concentration étant
proportionnelle à celle du glucose entrant. Une fois dans le neurone, il est pris en
charge par la lactate déshydrogénase qui le transforme en pyruvate directement
utilisable comme source d’énergie (Tsacopoulos M et Magistretti PJ, 1996). Ceci
sous-entend que le glucose agirait non seulement directement sur les neurones
sensibles au glucose via la glycolyse et le cycle de Krebs, mais aussi indirectement

- 110 -
via le métabolisme glial (Levin BE et coll., 2002). Enfin, les astrocytes sont capables
de modifier l’organisation des synapses. Cela passerait par la libération de
particules de cholestérol qui seraient ensuite intégrées par les neurones et
aboutirait à l’augmentation du nombre de synapses (Fields RD et Stevens-Graham
B, 2002). Ainsi, on peut envisager un couplage astrocyte-neurone via les lipides
permettant la modulation de l’activité synaptique et donc, de la transmission des
signaux. Le statut énergétique de l’organisme, communiqué notamment par le biais
de la concentration en glucose, pourrait influencer ces connexions interneuronales,
aboutissant probablement à un contrôle à plus long terme de l’homéostasie
énergétique. En faveur de cette hypothèse, des travaux très récents effectués sur le
foie, indiquent que le glucose est capable de stimuler un facteur de transcription,
LXR (Liver X Receptor), protéine assurant le contrôle du métabolisme du cholestérol
(Mitro N et coll., 2007). Ainsi, LXR intégrerait les métabolismes glucidique et
lipidique. Dès lors, il serait intéressant de déterminer s’il est présent au sein du
SNC, et en particulier dans l’hypothalamus.
L’absence d’activation du NA par le bolus de glucose suite au prétraitement à
la MSO est corrélée à l’absence de réponse insulinémique qui y fait normalement
suite (Figure 42). Ce résultat suggère fortement que la détection de la variation de
la concentration en glucose impliquerait les astrocytes du NA, et que l’activation de
ce noyau hypothalamique est indispensable à la stimulation des cellules β du
pancréas. Ceci est en accord avec le rôle central joué par cette structure cérébrale
dans la régulation de l’homéostasie énergétique, en particulier au niveau de
l’intégration des signaux périphériques qu’ils soient de nature hormonale ou
métabolique (cf. § III) (Niswender KD et coll., 2004 ; Wang R et coll., 2004 ; Briski
KP, 1998). Ceci renforce aussi l’idée selon laquelle le NA serait le premier à détecter
une modification de la glycémie, et stimulerait ensuite le PVN (Wynne K et coll.,
2005).
Afin de certifier au maximum la spécificité de l’effet observé avec l’utilisation
de la MSO, nous avons procédé à des expériences contrôles. Le test de tolérance au
glucose montre que le pancréas est tout à fait capable de sécréter de l’insuline en
quantités normales 18 h après l’injection de MSO par voie intrapéritonéale (Tableau
2). Ainsi, le traitement à la MSO a bien eu un effet cérébral (absence d’activation du
NA), et non un effet périphérique (altération des cellules β). En accord avec nos
résultats, d’autres auteurs ont montré que cette molécule n’affectait pas la
sécrétion d’insuline basale ou induite par le glucose (Schneid C et coll., 2003).

- 111 -
Notons toutefois que l’hyperglycémie créée par l’administration massive de glucose
par voie intraveineuse est significativement plus prononcée pour les rats ayant reçu
la MSO par rapport à ceux ayant reçu le sérum physiologique, suggérant une
possible résistance à l’insuline. Enfin, nos résultats obtenus suite à l’injection
intracérébroventriculaire de la MSO (résultats non montrés) excluent également un
hypothétique effet périphérique puisque ce protocole ne devait toucher que la GS
centrale.
Par ailleurs, il a été décrit que la MSO inhibait une autre enzyme, la γ-
glutamylcystéine synthase, qui intervient dans la voie de synthèse du glutathion
(Cooper AJL et coll., 1976). On pouvait donc se demander si l’effet que nous
observions était bien spécifique de l’inhibition de la GS. Un analogue de la MSO, la
buthionine sulfoximine (BSO), inhibe la γ-glutamylcystéine synthase environ cent
fois plus efficacement que la MSO (Griffith OW et Meister A, 1979). Un
prétraitement à la BSO à la même concentration que la MSO, ne montre aucun effet
sur la réponse insulinémique observée (Figure 43). On peut donc en conclure que
la disparition de l’hyperinsulinémie rapide et transitoire est spécifique à l’inhibition
de la GS cérébrale par la MSO et non à un effet sur la γ-glutamylcystéine synthase.
L’ensemble de ces données montre que des astrocytes sont effectivement
sensibles à l’augmentation de la concentration en glucose au niveau du NA, et
transmettraient l’information métabolique à certains neurones. L’information
transiterait alors par différentes structures pour conduire à la modulation du SNA
permettant finalement la sécrétion d’insuline par le pancréas afin de remédier à
l’hyperglycémie sensée exister (Figure 38 ).

- 112 -
XII – Acteurs moléculaires impliqués dans la sensibilité cérébrale au glucose
Un modèle de sensibilité au glucose bien connu est celui des cellules β du
pancréas (cf. § I-2.2 et figure 6). Dans celles-ci, intervient un couple de deux
protéines : le transporteur au glucose GLUT2 et la glucokinase (GK). Notre
hypothèse, renforcée par certaines données de la littérature, est que ces deux
acteurs moléculaires rempliraient des fonctions similaires au niveau du SNC. Pour
cette étude, nous avons choisi une approche pharmacologique appliquée à la
sécrétion d’insuline induite par l’injection aiguë de glucose vers le cerveau.
1. Résultats III
1.1. Etude de l’implication de la glucokinase
En ce qui concerne l’étude de l’implication de la GK dans la sensibilité
cérébrale au glucose, nous avons choisi la glucosamine (GlcN) comme inhibiteur de
son activité (Yoshikawa H et coll., 2002).
1.1.1. Effet de la glucosamine sur la sécrétion d’insuline
La GlcN (80 mg/kg) injectée simultanément au glucose par le cathéter
intracarotidien au temps 0 n’induit pas de variation de l’insulinémie contrairement
au groupe de rats témoins recevant à la place du NaCl de même osmolarité (1800
mOsmol) en plus du glucose (-9,77 ± 4,34 versus 54,52 ± 18,56 μU/ml) (Figure 44).

- 113 -
Figure 44 : Effet de l’injection intracarotidienne vers le cerveau de glucosamine (GlcN) au temps 0 sur la sécrétion d’insuline induite par celle simultanée de D-glucose. Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. ** p < 0,01 par rapport au groupe témoin.
La GlcN supprime l’hyperinsulinémie normalement observée en réponse au
bolus de glucose vers le SNC.
1.1.2. Effet cérébral et non périphérique de la glucosamine
Comme dans le cas de l’utilisation de la MSO (cf. § XI-1.3.1.), nous avons
vérifié que l’effet de la GlcN était bien dû à une action uniquement centrale et non à
une action globale, c'est-à-dire également périphérique avec inhibition de la GK
pancréatique. Nous avons ainsi réalisé un test de tolérance au glucose à l’aide d’une
charge intraveineuse de glucose (0,8 g/kg de poids corporel) sur des animaux
recevant la GlcN par la carotide.

- 114 -
Figure 45 : Effet de l’injection par voie intracarotidienne vers le cerveau de glucosamine (GlcN) au temps 0 sur un test de tolérance au glucose injecté par voie intraveineuse : effet sur la glycémie (A) et sur l’insulinémie (B). Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. * p < 0,05 par rapport au groupe témoin. iv : injection intraveineuse.
L’injection de glucose par voie intraveineuse induit une augmentation
massive de la glycémie périphérique que ce soit pour le groupe de rats témoins avec
injection vers le cerveau de NaCl ou le groupe de rats recevant la GlcN (Figure
45A). Notons qu’au temps 20 min, l’augmentation du delta glycémique est
significativement plus importante pour le groupe GlcN (3,82 ± 0,36 versus 1,80 ±
0,8 mM pour le NaCl).
B
A

- 115 -
Cette augmentation de la glycémie périphérique est accompagnée d’une
augmentation importante de l’insulinémie et ce, pour les deux groupes de rats
(Figure 45B). Toutefois, aux temps 15 et 20 min, cette élévation est
significativement plus faible chez les rats recevant la GlcN (23,16 ± 6,03 versus
65,77 ± 15,92 à t15 et 6,59 ± 9,23 versus 52,16 ± 13,61 μU/ml à t20).
Le test de tolérance au glucose montre, qu’au moins pendant les 10
premières minutes suivant l’injection de GlcN vers le cerveau, le pancréas est tout à
fait capable de secréter l’insuline normalement.
1.1.3. Effet spécifique à l’inhibition de la glucokinase ?
La GlcN peut néanmoins avoir un effet autre que celui d’inhiber la GK : elle
est en effet capable d’activer la voie des hexosamines (cf. § III-2.2.2, figure 21)
(Yoshikawa H et coll., 2002). Dans le but de déterminer de quel effet proviennent
nos observations, nous avons procédé à l’injection de N-acétylglucosamine (GlcNAc)
aux mêmes caractéristiques que celle de la GlcN. La GlcNAc a été décrite comme
activant la voie des hexosamines mais en inhibant la GK de manière beaucoup
moins efficace que la GlcN (Wells L et coll., 2003). Donc dans ce cas-là, c’est l’effet
« activation de la voie des hexosamines » qui prédomine.
Contrairement à la GlcN, l’injection de GlcNAc à la même dose (80 mg/kg)
vers le cerveau (simultanément au glucose) ne supprime pas la réponse
insulinémique induite par le glucose au temps 1 min (GlcN + glucose : -9,77 ±
4,34 ; GlcNAc + glucose : 46,90 ± 6,23 ; NaCl + glucose : 54.52 ± 18,56 μU/ml)
(Figure 46). Et même, de manière surprenante, si on injecte la GlcNAc
simultanément à du sérum physiologique au temps 0, on observe également au
temps 1 min une augmentation de l’insulinémie (29,94 ± 6,57 μU/ml) (Figure 46).

- 116 -
Figure 46 : Effet de l’injection intracarotidienne vers le cerveau de N-acétylglucosamine (GlcNAc) au temps 0 sur la sécrétion d’insuline induite par celle simultanée de D-glucose. Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. ** p < 0,01 ; *** p < 0.001
La GlcNAc ne bloque pas l’induction de l’hyperinsulinémie par le bolus de glucose.
Donc, l’effet inhibiteur de la GlcN sur cette réponse est bien spécifique à l’inhibition de
la GK et ne fait pas intervenir son effet activateur de la voie des hexosamines.
1.2. Etude de l’implication du couple glucokinase/GLUT2
L’alloxane est un inhibiteur de la GK ayant la particularité d’être transporté
spécifiquement par GLUT2 contrairement à d’autres inhibiteurs de cette hexokinase
(Elsner M et coll., 2002). C’est pourquoi notre choix s’est porté sur celui-ci plutôt
qu’un autre, sachant qu’au moment de ces expérimentations il n’existait pas
d’agent pharmacologique spécifique à GLUT2.
1.2.1. Injection de l’alloxane au temps zéro
L’injection simultanée vers le cerveau de glucose à 9 mg/kg et d’alloxane à 1
mg/kg (dose choisie après plusieurs essais) au temps 0 est suivie, une minute
après, d’une augmentation de l’insulinémie comme observée chez les rats ne

- 117 -
recevant que du glucose. Cette augmentation se prolonge jusqu’au temps 3 minutes
(delta de 64,20 ± 23,32 à 1 min et de 52,91 ± 21,75 μU/ml à 3 min) (Figure 47).
Figure 47 : Effet de l’injection intracarotidienne vers le cerveau d’alloxane au temps 0 sur la sécrétion d’insuline induite par celle simultanée de D-glucose. Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. NS : non significatif (p > 0,05). * p < 0,05 : significativement différent par rapport à t 0.
Puisque l’alloxane est un inhibiteur de la GK dont on envisage l’implication
dans la détection cérébrale de glucose, nous nous attendions à une disparition de
l’augmentation de l’insulinémie normalement induite par l’injection de glucose vers
le cerveau comme observé après l’administration de GlcN. Or nous avons obtenu
l’inverse. C’est pourquoi nous avons testé l’effet de l’alloxane seul : on observe une
augmentation du delta insulinémique au temps 3 minutes significativement
différente comparée au temps 0 (35,44 ± 8,68 μU/ml, p=0,0065).
1.2.2. Injection de l’alloxane au temps -10 minutes
L’injection d’alloxane vers le cerveau induit en elle-même une augmentation
de l’insulinémie au temps 3 minutes (cf. § précédent et figure 47). Cet effet peut

- 118 -
masquer une possible inhibition de la réponse au glucose. Nous avons donc
procédé à l’injection de l’alloxane 10 minutes avant celle de glucose, temps au bout
duquel l’hyperinsulinémie induite par l’alloxane seul disparaît.
Deux groupes de rats témoins ont été constitués. Un premier groupe a reçu
une solution de NaCl 0,9 % par voie intracarotidienne à la place de l’alloxane. Un
deuxième groupe de rats a reçu deux injections de glucose à 10 minutes d’intervalle
afin de mimer le premier pic d’insuline induit par l’alloxane chez le groupe
alloxane/glucose.
Figure 48 : Effet d’une pré-injection par voie intracarotidienne vers le cerveau d’alloxane sur la sécrétion d’insuline induite par l’injection de D-glucose au temps 0. Les résultats sont exprimés en delta par rapport aux valeurs basales mesurées à t 0, en moyennes ± l’écart type à la moyenne. NS : non significatif par rapport aux deux autres groupes à t 1 (p > 0,05).
Comme précédemment décrit (cf. § X-1.1.1, figure 31), une première
injection de glucose 10 minutes avant celle effectuée au temps 0, n’altère pas
l’augmentation de l’insulinémie observée au temps 1 min. Cette deuxième réponse a
la même amplitude que celle produite avec le groupe de rats prétraités au NaCl 0,9
% (Figure 48).
Lorsque les rats reçoivent une injection aiguë d’alloxane vers le cerveau 10
minutes avant le bolus de glucose, on observe toujours une augmentation de

- 119 -
l’insulinémie 1 minute après celle de glucose au temps 0. Toutefois, on peut
remarquer que son amplitude tend à être plus faible, mais cette différence reste
statistiquement non significative (23,70 ± 3,27 versus 37,43 ± 6,78 μU/ml) (Figure
48).
Ces résultats indiquent que l’alloxane per se induit la libération d’insuline en
périphérie et de ce fait, ne nous permettent pas d’émettre une quelconque conclusion
sur l’implication ou non du couple GK/GLUT2.
2. Discussion III
Selon notre hypothèse de départ, la sensibilité au glucose, très bien décrite
pour les cellules β, existerait également dans le SNC et impliquerait un mécanisme
similaire (Figures 6 et 28). D’ores et déjà, quelques uns de nos résultats apportent
des éléments en sa faveur, puisqu’ils mettent en lumière des similitudes. En effet,
une modification de la concentration intracellulaire en calcium constitue un
intermédiaire entre le stimulus et l’expression de c-Fos (Chaudhuri A, 1997). Or,
dans les cellules β, l’augmentation de la concentration en glucose intracellulaire,
proportionnelle à l’hyperglycémie, induit une entrée de calcium qui permet
l’exocytose de l’insuline (Doyle ME et Egan JM, 2003). De la même manière, on peut
imaginer que si cet influx calcique existe aussi dans les cellules cérébrales sensibles
au glucose, comme le suggère l’apparition de c-Fos après l’injection aiguë de
glucose par la carotide, il peut conduire cette fois-ci à l’exocytose de
neurotransmetteurs ou de molécules de signalisation indispensables au couplage
astrocyte-neurone (glutamate, ATP, lactate). Par ailleurs, le fait que la MSO
supprime l’activation du NA corrélée à l’absence de réponse insulinémique, suggère
que ce couplage impliquerait le glutamate puisque la GS, une fois inhibée, est
incapable de le transformer en glutamine. Or, depuis quelques années, des travaux
montrent que cet acide aminé interviendrait directement dans la sécrétion
d’insuline des îlots de Langerhans (Maechler P et coll., 2002). Il proviendrait
notamment du glucose par le biais de l’α-kétoglutarate, un intermédiaire du cycle
des acides tricarboxyliques. Cette transformation est assurée par la glutamate
déshydrogénase. Par analogie, les astrocytes pourraient également utiliser le
glutamate comme signal direct pour le neurone, s’il est lui-même libèré, et/ou
indirect, s’il contrôle l’exocytose d’autres molécules de signalisation.

- 120 -
Au-delà de ces premières similitudes, nous avons voulu déterminer quel était
l’équipement moléculaire présent dans le SNC et caractéristique des cellules
sensibles au glucose, en comparaison avec les cellules β. Depuis longtemps, au
niveau du pancréas, il a été établi qu’un couple protéique remplissait largement le
rôle de « détecteur » de la concentration extracellulaire en glucose. Il est constitué
du transporteur au glucose GLUT2 et de la glucokinase (GK), le premier permettant
l’entrée de l’hexose proportionnellement à sa concentration environnante et la
seconde, sa phosphorylation également de façon proportionnelle (Doyle ME et Egan
JM, 2003).
Dans un premier temps, nous avons procédé à l’inhibition de la GK grâce à la
glucosamine (GlcN) transportée par les différents GLUTs. Son injection aiguë par la
carotide vers le SNC simultanément au glucose, supprime la réponse insulinémique
périphérique (Figure 44). Ceci suggère fortement que l’activité normale de la GK est
indispensable à la sensibilité cérébrale à cet hexose. Par ailleurs, le test de
tolérance au glucose indique que cet effet est bien spécifique à la GK cérébrale
puisque le pancréas est toujours capable de secréter normalement de l’insuline, du
moins jusqu’au temps 10 minutes, c'est-à-dire pendant la durée de la réponse
insulinémique au bolus carotidien de glucose (Figure 45).
La GlcN est largement utilisée dans l’étude de la GK, notamment dans le but
de caractériser l’activité hépatique et pancréatique en réponse à une hyperglycémie,
ou bien dans celui de définir directement le fonctionnement de l’enzyme, c'est-à-dire
de déterminer ses sites de liaison (Balkan B et Dunning BE, 1994 ; Barzilai N et
coll., 1996 ; Agius L et Stubbs M, 2000). Dans le cadre de l’étude de la sensibilité
cérébrale au glucose, la GlcN a aussi été utilisée et supprime pour partie les
réponses des neurones sensibles au glucose venant ainsi conforter nos résultats
(Yang XJ et coll., 1999 ; Dunn-Meynell AA et coll., 2002).
Au-delà de son effet inhibiteur sur la GK pour lequel elle est le plus souvent
employée, la GlcN intervient dans une autre voie métabolique, la voie des
hexosamines. Or, il semblerait que cette cascade de réactions enzymatiques jouerait
un rôle comme indicateur du statut énergétique de l’organisme (Wells L et coll.,
2003). Dans ce contexte, la GlcN peut être utilisée comme activateur de la voie des
hexosamines (Figures 21 et 49).

- 121 -
Figure 49 : Effets de la glucosamine (GlcN) sur la glucokinase (GK) et la voie des hexosamines, qui est aussi capable d’incorporer la N-acétylglucosamine (GlcNAc).
Afin de s’assurer du lieu d’action de la GlcN (inhibition de la GK versus
activation de la voie des hexosamines), nous avons utilisé la N-acétylglucosamine
(GlcNAc) qui agit plus en aval dans la voie des hexosamines en servant de substrat
à la synthèse d’UDP-GlcNAc (Wells L et coll., 2003) (Figure 49). L’injection de
GlcNAc simultanément à celle de glucose vers le cerveau, n’altère pas la réponse
insulinémique rapide et transitoire, contrairement à la GlcN (Figure 46). L’effet
inhibiteur de la GlcN sur la sensibilité cérébrale au glucose dans notre modèle,
serait donc bien dû à l’inhibition de la GK et non à l’activation de la voie des
hexosamines. Par ailleurs, nos résultats montrent que l’activation de la voie des
hexosamines, seule, induit une réponse insulinémique même si à ce stade, on ne
peut pas exclure un effet périphérique de la GlcNAc. Néanmoins, ce résultat serait
en accord avec des données de la littérature suggérant une suractivation de cette
voie métabolique dans le diabète de type II (Wells L et coll., 2003). Il nous paraît
intéressant de noter que les résultats obtenus avec la GlcNAc sont toutefois en
contradiction avec ceux de Dunn-Meynell et coll. (2002) qui montrent que la GlcNAc
inhibe les réponses des neurones GE (stimulés par le glucose) au même titre que la
GlcN. Il est probable que cette discordance soit due aux conditions expérimentales
(in vivo versus in vitro).

- 122 -
Dans un second temps, nous avons décidé d’utiliser l’alloxane qui est un
inhibiteur de la GK sélectivement transporté par GLUT2, contrairement à d’autres
inhibiteurs de cette hexokinase (Elsner M et coll., 2002). Ainsi, son emploi
permettait de confirmer l’implication de la GK et de cibler le couple, et non une
seule des deux protéines mises en jeu dans les cellules β. A notre grande surprise,
non seulement l’injection aiguë d’alloxane vers le SNC n’empêche pas la réponse
insulinémique au glucose comme le fait la GlcN, mais en plus, elle induit per se une
hyperinsulinémie rapide et transitoire (Figure 47). Si l’on compare les effets de
l’injection de glucose seul, alloxane seul et glucose plus alloxane, il apparaît un effet
additif. Ceci suggère que ces deux composés empruntent des voies de signalisation
différentes ou aux cinétiques légèrement décalées.
Il a été démontré que cette toxine, injectée dans le troisième ventricule,
provoquait la mort des tanycytes qui le bordent (Sanders NM et coll., 2004). Ceci
indique qu’elle serait bien transportée spécifiquement par GLUT2 puisque ces
cellules spécialisées l’expriment (Garcia MA et coll., 2003). Par ailleurs, dans la
plupart des travaux, l’alloxane est utilisé comme agent diabétogène par une double
action sur les cellules β provoquant leur dysfonctionnement : l’inhibition de la GK
qui altère la sécrétion d’insuline, mais aussi la cytotoxicité induite par la génération
d’espèces actives de l’oxygène (EAOs) (Elsner M et coll., 2002). L’apparition de ces
radicaux libres est suivie d’une augmentation de la concentration intracellulaire en
calcium, ce qui aboutit à la sécrétion d’insuline (Kliber A et coll., 1996). Si les
cellules hypothalamiques sensibles au glucose fonctionnent vraiment comme les
cellules β du pancréas, on peut imaginer qu’il puisse se produire, dans les cellules
possédant GLUT2, une production d’EAOs entraînant un influx calcique suite à
l’injection d’alloxane. Il s’agirait d’un phénomène analogue à celui engendré par le
bolus de glucose (cf début de ce paragraphe de discussion), aboutissant à la
modulation de l’activité du SNA et à l’augmentation de l’insulinémie périphérique
effectivement observée. Enfin, au laboratoire, il a été démontré que les EAOs
d’origine mitochondriale, produites par le métabolisme du glucose, induisaient une
réponse insulinémique et seraient nécessaires à la sensibilité cérébrale à cet hexose
(Leloup C et coll., 2006). Ainsi, il est vraisemblable que l’alloxane injecté vers le SNC
induit une réponse insulinémique via ces mêmes composés.
Finalement, si on envisage que l’alloxane puisse exercer deux effets dans les
cellules exprimant GLUT2 (inhibition de la GK et production d’EAOs), il se peut que,
peut-être en raison de la dose d’alloxane que nous avons utilisée, l’effet inhibiteur

- 123 -
sur la GK soit masqué par une forte production d’EAOs. Il serait ainsi intéressant
de regarder si la co-injection, dans la carotide, de molécules antioxydantes et
d’alloxane induit une sécrétion rapide et transitoire d’insuline, comme observée
avec l’alloxane seul. Si cette réponse insulinémique disparaît, cela suggèrera
fortement l’implication de la production d’EAOs dans la réponse induite par
l’alloxane. L’addition de molécules antioxydantes à l’alloxane devrait donc permettre
de révéler uniquement l’effet inhibiteur de ce dernier sur la GK. Dans ce cas, notre
bolus de glucose, en présence d’alloxane et d’antioxydant ne devrait plus induire de
réponse insulinémique périphérique. En accord avec ce schéma, les travaux de
l’équipe de Dunn-Meynell montrent que les réponses au glucose des neurones GE
disparaissent bien en présence d’alloxane et de catalase (Dunn-Meynell AA et coll.,
2002). Néanmoins on ne peut pas exclure que, dans nos expériences, l’absence
d’inhibition visible de la GK par l’alloxane soit due à la durée de vie très courte de
cet agent pharmacologique puisqu’on l’injecte dans la carotide et non directement
sur les cellules sensibles au glucose (Lenzen S et Panten U, 1988).
La démarche pharmacologique que nous avons suivie (utilisation de la GlcN et de
l’alloxane), associée aux données quelques fois disparates de la littérature, nous permet ainsi
de conclure à l’implication de la GK dans la sensibilité cérébrale à une hyperglycémie
transitoire. Une étude systématique du contenu en ARNm des neurones sensibles au glucose
confirme effectivement la présence de la GK dans ces cellules (Kang L et coll.,2004).
Néanmoins, cette étude ne permet pas de savoir si les astrocytes en sont pourvus, ce qui dans
notre hypothèse de couplage entre astrocytes et neurones serait pertinent.D’autre part, cette
étude montre qu’il n’y a pas de corrélation entre la présence de GLUT2 et les réponses au
glucose d’un neurone, soulevant ainsi la question du transport de glucose à l’intérieur de ce
type cellulaire et de sa disponibilité pour la GK.

- 124 -
CCoonncclluussiioonnss
eett
PPeerrssppeeccttiivveess

- 125 -
CONCLUSIONS :
Les travaux menés au cours de cette étude aboutissent à plusieurs
conclusions pouvant se résumer ainsi :
L’injection aiguë de glucose par la carotide vers le cerveau de rat se
caractérise par :
le maintien d’une glycémie périphérique stable, suggérant qu’elle
n’augmente qu’au niveau central
l’apparition rapide d’une hyperinsulinémie périphérique transitoire
une spécificité pour le D-glucose.
Cette réponse au bolus de glucose implique :
la mise en jeu du SNA qui transmet l’information du SNC vers la
périphérie : le parasympathique active, tandis que l’orthosympathique
inhibe les cellules β du pancréas
l’hypothalamus avec l’activation spécifique du NA et du PVN
majoritairement l’activation de neurones, mais aussi de quelques
astrocytes
en particulier les astrocytes du NA : leur inhibition empêche
l’apparition de l’hyperinsulinémie induite par l’injection aiguë de
glucose vers le cerveau
une enzyme essentielle, la GK.
Ainsi, l’injection aiguë de D-glucose vers le cerveau induit l’activation
spécifique de l’hypothalamus, et en particulier du NA. La sensibilité de cet hexose
met en jeu la GK et les astrocytes, ces cellules gliales activant alors les neurones
auxquels elles sont couplées. Le signal métabolique est donc transformé en signal
électrique se propageant dans le SNC. Le NA doit activer d’autres régions
cérébrales, notamment le PVN, permettant la conduction de l’information vers le
tronc cérébral où l’activité du SNA est modulée, pour finalement aboutir à la
stimulation des cellules β du pancréas. Celles-ci libèrent de l’insuline, dans le but
de diminuer l’hyperglycémie générée par le bolus de glucose.

- 126 -
Figure 50 : Détection d’une hyperglycémie cérébrale impliquant les astrocytes du noyau arqué (NA) et la glucokinase (GK). β : cellule β du pancréas, PVN : noyau hypothalamique paraventriculaire, SNA : systène nerveux autonome.
Notre modèle d’hyperglycémie cérébrale permet d’étudier la sensibilité du
cerveau au glucose de manière indépendante de l’insuline, ce qui est impossible
dans les protocoles se rapprochant du cadre physiologique où ces deux paramètres
sont indissociables (absorption de sucres par voie orale, injection de glucose
intrapéritonéale, hypoglycémie induite par l’injection d’insuline ou le jeûne). De tels
travaux mettent l’accent sur le facteur « détection du glucose », or cette sensibilité
se trouve altérée dans des modèles animaux diabétiques et/ou obèses puisqu’elle
n’aboutit pas à une sécrétion d’insuline adéquate.

- 127 -
D’après nos résultats, les composés glutamine/glutamate semblent au
carrefour de plusieurs voies métaboliques en relation avec la détection du glucose.
Ils constituent un lien entre astrocytes et neurones (Figure 27), interviennent dans
l’étape limitante de la voie des hexosamines au niveau de la GFAT (Figure 21), et
finalement le glutamate peut aboutir à la formation de glucose par néoglucogenèse
(Figure 39). Ces molécules promettent d’être moins anodines qu’elles ne le
paraissent et approfondir cette piste pourrait mener à une meilleure compréhension
des dysfonctionnements métaboliques liés au glucose.
De même, dans le but de rétablir une détection normale du glucose, on peut
imaginer le développement de molécules thérapeutiques visant à augmenter
l’activité de la GK, ce qui devrait accroître la sensibilité à cet hexose. Par
conséquent, une hyperglycémie centrale de plus faible amplitude devrait aboutir à
une sécrétion d’insuline plus importante et donc permettre une normalisation plus
rapide de la glycémie.
PERSPECTIVES :
• Etude plus approfondie des régions cérébrales activées par une
hyperglycémie
Notre protocole d’injection aiguë de glucose à 9 mg/kg par l’artère
carotidienne vers le SNC ne modifie pas la glycémie périphérique. Nous supposons
qu’elle provoque une hyperglycémie locale, en particulier au niveau du système
porte hypothalamo-hypophysaire et du NA. Pour en être certains, il serait
nécessaire de mesurer la concentration en glucose, avant et après son
administration, au sein du NA. Or, des électrodes sont en cours de mise au point,
notamment pour leur miniaturisation. A l’heure actuelle, leur taille est trop
importante, lésant le noyau cérébral d’intérêt. La surface de cette électrode est
recouverte d’une enzyme, la glucose oxydase. La réaction dont elle est responsable,
va générer un gradient électrochimique qui pourra être mesuré et qui est
directement proportionnel à la quantité de substrat, le glucose (Dixon BM et coll.,
2002). Ainsi, il sera possible de doser les concentrations de ce sucre au cours du
temps, au sein d’un seul noyau hypothalamique et même sur animal éveillé. Par
ailleurs, on pourrait également déterminer, si cette concentration augmente dans
d’autres structures telles que les noyaux hypothalamiques PVN, VMN et le LH. En
ce qui concerne la première, cela permettrait de conforter ou d’invalider notre

- 128 -
hypothèse selon laquelle le NA serait à l’origine de son activation (la stimulation
observée par le biais de c-Fos ne serait donc pas directe).
Par rapport à cette dernière question, il est possible d’y répondre grâce aux
méthodes de traçage classiques utilisées en neuroanatomie. Elles consistent en
l’injection de molécules marquées dans un noyau cérébral donné, qui vont être
prises en charge par les synapses puis transportées le long du prolongement
nerveux pour aboutir dans le corps cellulaire. Dans le cas d’un transport
rétrograde, le marquage est retrouvé dans les neurones afférents, c'est-à-dire
excitateurs (Vercelli A et coll., 2000). Ainsi, administrés dans le PVN, ces traceurs
devraient être retrouvés dans des neurones du NA exprimant c-Fos en réponse au
bolus de glucose.
A la fin de ces travaux, nous avions envisagé de compléter nos résultats
d’activation cérébrale par le biais d’une technique innovante, l’imagerie par
résonance magnétique (IRM). En effet, depuis peu il existe des appareils étudiés
spécialement pour une application sur le petit animal (Tabuchi E et coll., 2002).
Cependant, cette technologie coûteuse, demande des connaissances pointues et
n’était pas encore totalement disponible sur le site. Cette approche, si elle est
réalisable, permettrait de suivre le décours de l’activation cérébrale, « en direct »,
suite au bolus de glucose chez le rat éveillé et il serait simple d’élargir notre champ
d’intérêt au cerveau entier. Enfin, comme le sacrifice de l’animal n’est pas
nécessaire, il peut être son propre témoin.
• Implication des astrocytes
A l’occasion de la discussion de nos résultats, nous avons suggéré que le
calcium semblait intervenir dans le mécanisme de sensibilité au glucose au niveau
cérébral, comme c’est le cas au niveau du pancréas. L’imagerie calcique, appliquée
aussi bien aux neurones qu’aux astrocytes sur des tranches fraîches de cerveau,
apportera une nouvelle compréhension du processus de détection du glucose. Il est
probable que l’administration de cet hexose induit des modifications de la
concentration intracellulaire en calcium dans certaines cellules du NA, notamment
des vagues calciques au sein de cellules gliales, mécanisme engendrant la
propagation rapide d’un signal à toute une population cellulaire, ce qui pourrait
expliquer que seuls, quelques astrocytes « détecteurs de glucose » soient nécessaires
pour engendrer une réponse. Finalement, en couplant la mesure de calcium
intracellulaire et l’enregistrement électrique, il serait possible de savoir si une telle

- 129 -
réponse calcique dans un astrocyte est capable d’induire une réponse électrique
donnée (de type GE par exemple) dans un neurone voisin.
En ce qui concerne les cellules gliales en général, nous sommes conscients
que l’antigène GFAP marqueur des astrocytes, ne permet pas de révéler les
tanycytes. Or il semblerait que ces cellules jouent un rôle important dans la
détection des variations de la concentration en glucose. Il nous paraît donc
important de déterminer s’ils expriment effectivement c-Fos en réponse à l’injection
aiguë de glucose, grâce à une double immunohistochimie c-Fos/vimentine,
marqueur de ces cellules, afin d’envisager une étude plus fine des mécanismes
moléculaires qui pourraient être mis en jeu dans une réponse éventuelle au glucose
de ces cellules.
• Etude de l’implication de la GK dans la réponse cérébrale à
l’hyperglycémie
Dans le cadre des traitements pharmacologiques GlcN et GlcNAc, il serait
intéressant de corréler les résultats que nous avons obtenus sur la réponse
insulinémique périphérique avec la présence (avec la GlcNAc) ou l’absence (avec la
GlcN) d’une activation de c-Fos au niveau de l’hypothalamus et plus
particulièrement du NA.
D’autre part, observer la réponse insulinémique suite à l’activation directe de
la GK par un agent pharmacologique spécifique, permettrait de renforcer
l’hypothèse de son rôle primordial dans la détection d’une hyperglycémie centrale.
Une telle injection devrait induire une augmentation de l’insulinémie au profil
identique, c'est-à-dire rapide et transitoire. Un article récent indique que deux
activateurs seraient disponibles, GKA1 et GKA2, caractérisés pour leur action sur le
métabolisme hépatique (Brocklehurst KJ et coll., 2004). Là aussi, une confirmation
des résultats pourrait être recherchée par immunohistochimie c-Fos.
Enfin, nous avons comme projet d’étudier le bagage moléculaire des
astrocytes d’intérêt. Pour ce faire, deux étapes sont nécessaires. Dans un premier
temps, il nous faut rechercher la présence, au niveau des astrocytes du NA, de la
GK. En effet, une colocalisation de l’enzyme et de la GFAP (recherchée par
immunohistochimie) renforcerait notre hypothèse de l’implication d’une GK
astrocytaire dans la sensibilité cérébrale au glucose. Ceci sera réalisé en couplant
hybridation in situ (pour la GK) et immunohistochimie (pour la GFAP).
Dans un second temps, il nous faudrait démontrer la présence de la GK
spécifiquement dans les astrocytes sensibles au glucose (c'est-à-dire activés par le

- 130 -
bolus de glucose). Pour cela, il conviendrait de réaliser notre protocole expérimental
et de procéder à une double immunohistochimie c-Fos/GFAP couplée à
l’hybridation in situ GK. Dans l’éventualité où l’hybridation in situ se révèlerait être
difficile à utiliser, nous pensons à utiliser la microdissection laser après double
immunohistochimie c-Fos/GFAP. Cette technique nous permet en effet d’avoir
accès au contenu en ARNm par RT-PCR (RetroTranscriptase Polymerase Chain
Reaction) pour une cellule donnée (dans notre cas, identifiée c-Fos +/GFAP +).
L’ensemble de ces travaux montre que l’hypothalamus, en particulier le
noyau arqué, les astrocytes et la glucokinase, tiennent un rôle important, si ce n’est
indispensable, dans la sensibilité au glucose du cerveau. Les futures investigations
proposées devraient permettre de mieux comprendre encore ces mécanismes, dans le
but d’éclairer la nature des dysfonctionnements observés dans le cas de
physiopathologies.

- 131 -
AArrttiiccllee eett rreevvuueess ppuubblliiééss

- 132 -

- 133 -

- 134 -

- 135 -

- 136 -

- 137 -

- 138 -

- 139 -

- 140 -

- 141 -

- 142 -

- 143 -

- 144 -

- 145 -

- 146 -

- 147 -

- 148 -

- 149 -

- 150 -
RRééfféérreenncceess
bbiibblliiooggrraapphhiiqquueess

- 151 -
Nombre de références bibliographiques : 170
A Agius L, Stubbs M Investigation of the mechanism by which glucose analogues cause translocation of glucokinase in hepatocytes: evidence for two glucose binding sites The Biochemical Journal, 2000, 346: 413-421 Aguilar-Bryan L, Bryan J Molecular biology of adenosine triphosphate-sensitive potassium channels Endocrine Reviews, 1999, 20(2):101-135 Ahima RS, Osei SY Leptin signaling Physiology & Behavior, 2004, 8: 223-241 Alquier T, Leloup C, Atef N, Fioramonti X, Lorsignol A, Pénicaud L Cerebral insulin increases brain response to glucose Journal of Neuroendocrinology, 2003, 15: 75-79 Alvarez-Buylla R, de Alvarez-Buylla ER Carotid sinus receptors participate in glucose homeostasis Respiration Physiology, 1988, 72: 347-360 Anand BK, Chhina GS, Sharma KN, Dua S, Singh B Activity of single neurons in the hypothalamic feeding centers: effect of glucose American Journal of Physiology, 1964, 207(5): 1146-1154 Angelopoulos N, Goula A, Tolis G Current knowledge in the neurophysiologic modulation of obesity Metabolism Clinical and Experimental, 2005, 54: 1202-1217 Arluison M, Quignon M, Nguyen P, Thorens B, Leloup C, Pénicaud L Distribution and anatomical localization of the glucose transporter 2 (GLUT2) in the adult rat brain – an immunohistochemical study Journal of Chemical Neuroanatomy, 2004, 28: 117-136 Atef N, Ktorza A, Pénicaud L CNS involvement in the glucose induced increase of islet blood flow in obese Zucker rats International Journal of Obesity and related metabolic disorders, 1995, 19(2):103-107

- 152 -
B Bady I, Marty N, Dallaporta M, Emery M, Gyger J, Tarussio D, Foretz M, Thorens B Evidence from Glut2-null mice that glucose is critical physiological regulator of feeding Diabetes, 2006, 55: 988-995 Balfour RH, Kruse Hansen AM, Trapp S Neuronal responses to transient hypoglycaemia in the dorsal vagal complex of the rat brainstem Journal of Physiology, 2006, 570.3: 469-484 Balkan B, Dunning BE Glucosamine inhibits glucokinase in vitro and produces a glucose-specific impairment of in vivo insulin secretion in rats Diabetes, 1994, 43: 1173-1179 Banks WA The source of cerebral insulin European Journal of Pharmacology, 2004, 490: 5-12 Barzilai N, Hawkins M, Angelov I, Meizhu H, Rossetti L Glucosamine-induced inhibition of liver glucokinase impairs the ability of hyperglycemia to suppress endogenous glucose production Diabetes, 1996, 45(10): 1329-1335 Bates SH, Myers MG Jr The role of leptin receptor signalling in feeding and neuroendocrine function Trends in Endocrinology and Metabolism, 2003, 14(10): 447-452 Baulieu EE, Kelly PA Hormones: from molecules to disease Hermann, publishers in arts and science, Paris, 1990 Benani A, Troy S, Carmona MC, Fioramonti X, Lorsignol A, Leloup C, Casteilla L, Pénicaud L Role for mitochondrial reactive oxygen species in brain lipid sensing Diabetes, 2007, 56:152-160 Berthoud H-R Mind versus metabolism in the control of food intake and energy balance Physiology & Behavior, 2004, 81: 781-793 Bestetti G, Rossi GL Hypothalamic lesions in rats with long-term streptozotocin-induced diabetes mellitus Acta Neuropathologica, 1980, 52: 119-127

- 153 -
Bojanowska E Physiology and pathophysiology of glucagon-like peptide-1 (GLP-1): The role of GLP-1 in the pathogenesis of diabetes mellitus, obesity, and stress Medical Science Monitor, 2005, 11 (8): RA271-278 Bouret SG, Simerly RB Minireview: leptin and development of hypothalamic feeding circuits Endocrinology, 2004, 145(6): 2621-2626 Bouzier-Sore AK, Merle M, Magistretti PJ, Pellerin L Feeding active neurons: (re)emergence of a nursing role for astrocytes Journal of Physiology - Paris, 2002, 96: 273-282 Briski KP Glucoprivic induction of Fos immunoreactivity in hypothalamic dopaminergic neurons NeuroReport, 1998, 9: 289-295 Briski KP, Brandt JA Oxytocin and vasopressin neurones in principal and accessory hypothalamic magnocellular structures express Fos-immunoreactivity in response to acute glucose deprivation Journal of Neuroendocrinology, 2000, 12: 409-414 Broberger C, Johansen J, Johansson C, Schalling M The neuropeptide Y/agouti gene-related protein (AGRP) brain circuitry in normal, anorectic, and monosodium glutamate-treated mice Proceedings of the National Academy of Sciences, 1998, 95: 15043-15048 Broberger C Brain regulation of food intake and appetite: molecules and networks Journal of Internal Medicine, 2005, 258: 301-327 Brocklehurst KJ, Payne VA, Davies RA, Carroll D, Vertigan HL, Whightman HJ, Aiston S, Waddell ID, Leighton B, Coghlan MP, Agius L Stimulation of hepatocyte glucose metabolism by novel small molecule glucokinase activators Diabetes, 2004, 53: 535-541 Burcelin R, Dolci W, Thorens B Portal glucose infusion in the mouse induces hypoglycaemia: evidence that the hepatoportal glucose sensor stimulates glucose utilization Diabetes, 2000, 49(10): 1635-1642 Burcelin R, Pénicaud L Rôle de l’axe intestin, foie, et système nerveux dans la régulation de la glycémie en période absorptive Médecine Clinique, endocrinologie & diabète, 2003, hors-série n°1, janvier-février Buse MG Hexosamines, insulin resistance, and the complications of diabetes: current status American Journal of Physiology – Endocrinology and Metabolism, 2006, 290: 1-8

- 154 -
C Campbell NA Biologie Editions De Boeck, Bruxelles, 1995, 3ième édition Carrasco M, Portillo F, Larsen PJ, Vallo JJ Insulin and glucose administration stimulates Fos expression in neurones of the paraventricular nucleus that project to autonomic preganglionic structures Journal of Neuroendocrinology, 2001, 13: 339-346 Chang L, Chiang SH, Saltiel AR Insulin signalling and the regulation of glucose transport Molecular Medicine, 2004, 10(7-12): 65-71 Chaudhuri A Neural activity mapping with inducible transcription factors NeuroReport, 1997, 8(16): v-ix Cheatham B, Kahn CR Insulin action and the insulin signalling network Endocrine Reviews, 1995, 16(2): 117-142 Clément L, Cruciani-Guglielmacci C, Magnan C, Vincent M, Douared L, Orosco M, Assimacopoulos-Jeannet F, Pénicaud L, Ktorza A Intracerebroventricular infusion of a triglyceride emulsion leads to both altered insulin secretion and hepatic glucose production in rats Endocrinology, 2002, 445: 375-380 Cooper AJL, Stephani RA, Meister A Enzymatic reactions of methionine sulfoximine The Journal of Biological Chemistry, 1976, 251 (21): 6674-6682 Cowley MA, Pronchuk N, Fan W, Dinulescu DM, Colmers WF, Cone RD Integration of NPY, AGRP, and melanocortin signals in the hypothalamic paraventricular nucleus: evidence of a cellular basis for the adipostat Neuron, 1999, 24: 155-163 Cowley MA, Smart JL, Rubinstein M, Cerdan MG, Diano S, Horvath TL, Cone RD, Low MJ Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus Nature, 2001, 411: 480-484 Cruciani-Guglielmacci C, Hervalet A, Douared L, Sanders NM, Levin BE, Ktorza A, Magnan C Beta oxidation in the brain is required for the effects of non-esterified fatty acids on glucose-induced insulin secretion in rats Diabetologia, 2004, 47: 2032-2038

- 155 -
Currie PJ Integration of hypothalamic feeding and metabolic signals: focus on neuropeptide Y Appetite, 2003, 41: 335-337
D Darbin O, Carre E, Naritoku D, Risso JJ, Lonjon M, Patrylo PR Glucose metabolites in the striatum of freely behaving rats following infusion of elevated potassium Brain Research, 2006, 1116: 127-131 Del Prato S Loss of early insulin secretion leads to postprandial hyperglycaemia Diabetologia, 2003, 46(suppl1): M2-M8 Dixon BM, Lowry JP, O'Neill RD Characterization in vitro and in vivo of the oxygen dependence of an enzyme/polymer biosensor for monitoring brain glucose Journal of Neuroscience Methods, 2002, 119(2):135-42 Donovan CM, Hamilton-Wessler M, Halter JB, Bergman RN Primacy of liver glucosensors in the sympathetic response to progressive hypoglycaemia Proceedings of the National Academy of Sciences USA, 1994, 91(7): 2863-2867 Doyle ME, Egan JM Pharmacological agents that directly modulate insulin secretion Pharmacological Reviews, 2003, 55: 105-131 Dunbar J, Lapanowski K, Barnes M, Rafols J Hypothalamic agouti-related protein immunoreactivity in food-restricted, obese, and insulin-treated animals: evidence for glia cell localization Experimental Neurology, 2005, 191: 184-192 Dunn-Meynell AA, Govek E, Levin BE Intracarotid glucose selectively increases Fos-like immunoreactivity in paraventricular, ventromedial and dorsomedial nuclei neurons Brain Research, 1997, 748: 100-106 Dunn-Meynell AA, Rawson NE, Levin BE Distribution and phenotype of neurons containing the ATP-sensitive K+ channel in rat brain Brain Research, 1998, 814(1-2):41-54 Dunn-Meynell AA, Routh VH, Kang L, Gaspers L, Levin BE Glucokinase is the likely mediator of glucosensing in both glucose-excited and glucose-inhibited central neurons Diabetes, 2002, 51: 2056-2065

- 156 -
Dupin H, Cuq JL, Malewiak MI, Leynaud-Rouaud C, Berthier AM Alimentation et nutrition humaines Editions ESF, Paris, 1992
E Elmquist JK, Bjorbaek C, Ahima RS, Flier JS, Saper CB Distributions of leptin receptor mRNA isoforms in the rat brain The Journal of comparative neurology, 1998, 395(4):535-547 Elmquist JK, Elias CF, Saper CB From lesions to leptin: hypothalamic control of food intake and body weight Neuron, 1999, 22: 221-232 Elsner M, Tiedge M, Guldbakke B, Munday R, Lenzen S Importance of the GLUT2 glucose transporter for pancreatic beta cell toxicity of alloxan Diabetologia, 2002, 45: 1542-1549
F Fetissov SO, Kopp J, Hökfelt T Distribution of NPY receptors in the hypothalamus Neuropeptides, 2004, 38: 175-188 Fields RD, Stevens-Graham B New insights into neuron-glia communication Science, 298:556-562 Figlewicz DP Adiposity signals and food reward: expanding the CNS roles of insulin and leptin American Jounal of Physiology – Regulatory, Integrative and Comparative Physiology, 2003, 284: R882-R892 Fioramonti X, Lorsignol A, Taupignon A, Pénicaud L A new ATP-sensitive K+ channel-independent mechanism is involved in glucose-excited neurons of mouse arcuate nucleus Diabetes, 2004, 53:2767-2775 Fioramonti X, Contié S, Song Z, Routh VH, Lorsignol A, Pénicaud L Characterization of glucosensing neuron subpopulations in the arcuate nucleus: integration in neuropeptide Y and pro-opio melanocortin networks? Diabetes, 2007, 56(5):1219-27 Fliers E, Kreier F, Voshol PJ, Havekes LM, Sauerwein HP, Kalsbeek A, Buijs RM, Romijn JA White adipose tissue: getting nervous Journal of Neuroendocrinology, 2003, 15:1005-1010

- 157 -
Frankish HM, Dryden S, Hopkins D, Wang Q, Williams G Neuropeptide Y, the Hypothalamus, and Diabetes: Insights Into the Central Control of Metabolism Peptides, 1995, 16(4): 757-771 Friedman JM, Halaas JL Leptin and the regulation of body weight in mammals Nature, 1998, 395:763-770 Funahashi H, Yada T, Suzuki R, Shioda S Distribution, function and properties of leptin receptors in the brain International Review of Cytology, 2003, 224:1-27
G Garcia MA, Millan C, Balmaceda-Aguilera C, Castro T, Pastor P, Montecinos H, Reinicke K, Zuniga F, Vera JC, Onate SA, Nualart F Hypothalamic ependymal-glial cells express the glucose transporter GLUT2, a protein involved in glucose sensing Journal of Neurochemistry, 2003, 86: 709-724 Girard J Rôle des acides gras libres dans la sécrétion et l’action de l’insuline : mécanismes de la lipotoxicité Médecine/Sciences, 2003, 19 : 827-833 Gloor SM, Wachtel M, Bolliger MF, Ishihara H, Landmann R, Frei K Molecular and cellular permeability control at the blood-brain barrier Brain Research Reviews, 2001, 36(2-3): 258-264 Griffith OW, Meister A Potent and specific inhibition of glutathione synthesis by buthionine sulfoximine (S-n-butyl homocysteine sulfoximine) The Journal of Biological Chemistry, 1979, 254 (16): 7558-7560 Guan XM, Yu H, Van der Ploeg LH Evidence of altered hypothalamic pro-opiomelanocortin/neuropeptide Y mRNA expression in tubby mice Brain Research Molecular Brain Research, 1998, 59: 273-279
H Hakansson ML, Brown H, Ghilardi N, Skoda RC, Meister B Leptin receptor immunoreactivity in chemically defined target neurons of the hypothalamus The Journal of Neuroscience, 1998, 18(1): 559-572

- 158 -
Helary-Bernard K, Ardourel MY, Cloix JF, Hevor TK The xenobiotic methionine sulfoximine modulates carbohydrate anabolism and related genes expression in rodent brain Toxicology, 2000, 153:179-187 Hevor TK, Delorme P, Gayet J Glycogen content and fructose-1,6-biphosphatase activity in methionine sulfoximine epileptogenic mouse brain and liver after protein synthesis inhibition Neuropathology and Applied Neurobiology, 1985, 11: 129-139 Himmi T, Perrin J, Dallaporta M, Orsini JC Effects of lactate on glucose-sensing neurons in the solitary tract nucleus Physiology & Behavior, 2001, 74: 391-397 Hoffman GE, Smith MS, Verbalis JG c-Fos and related immediate early gene products as markers of activity in neuroendocrine systems Frontiers in Neuroendocrinology, 1993, 14(3): 173-213 Hunter RG, Philpot K, Vicentic A, Dominguez G, Hubert GW, Kuhar MJ CART in feeding and obesity TRENDS in Endocrinology and Metabolism, 2004, 15(9): 454-459
I Ibrahim N, Bosch MA, Smart JL, Qiu J, Rubinstein M, Ronnekleiv OK, Low MJ, Kelly MJ Hypothalamic proopiomelanocortin neurons are glucose responsive and express KATP channels Endocrinology, 2003, 144(4): 1331-1340
J Jansson L, Hellerstrüm C Glucose-induced changes in pancreatic islet blood flow mediated by central nervous system American Journal of Physiology, 1986, 251: E644-E647 Jiang G, Zhang BB Glucagon and regulation of glucose metabolism American Journal of Physiology, Endocrinology and Metabolism, 2003, 284: E671-E678

- 159 -
K Kahn BB, Alquier T, Carling D, Hardie DG AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism Cell Metabolism, 2005, 1: 15-25 Kalra SP, Dube MG, Pu S, Xu B, Horvath TL, Kalra PS Interacting appetite-regulating pathways in the hypothalamic regulation of body weight Endocrine Reviews, 1999, 20(1): 68-100 Kang L, Routh VH, Kuzhikandathil EV, Gaspers LD, Levin BE Physiological and molecular characteristics of rat hypothalamic ventromedial nucleus glucosensing neurons Diabetes, 2004, 53: 549-559 Kennedy GC The role of depot fat in the hypothalamic control of food intake in the rat Proceedings of the Royal Society of London, 1953, 140(901): 578-596 King BM The rise, fall, and resurrection of the ventromedial hypothalamus in the regulation of feeding behaviour and body weight Physiology & Behavior, 2006, 87: 221-244 Kirchgessner AL Orexins in the brain-gut axis Endocrine Reviews, 2002, 23(1):1-15 Kliber A, Szkudelski T, Chichlowska J Alloxan stimulation and subsequent inhibition of insulin release from in situ perfused rat pancreas Journal of Physiology and Pharmacology, 1996, 47(2): 321-328 Krude H, Biebermann H, Luck W, Horn R, Brabant G, Grüters A Severe early-onset obesity, adrenal insufficiency and red hair pigmentation caused by POMC mutations in humans Nature Genetics, 1998, 19(2):155-157 Kong J, Shepel PN, Holden CP, Mackiewicz M, Pack AI, Geiger JD Brain glycogen decreases with increased periods of wakefulness: implications for homeostatic drive to sleep The Journal of Neuroscience, 2002, 22(13): 5581-5587
L Laughton W, Powley TL Bipiperidyl mustard produces brain lesions and obesity in the rat Brain Research, 1981, 221: 415-420

- 160 -
Leloup C, Arluison M, Lepetit N, Cartier N, Marfaing-Jallat P, Ferré P, Pénicaud L Glucose transporter 2 (GLUT2): expression in specific brain nuclei Brain Research, 1994, 638: 221-226 Leloup C, Orosco M, Serradas P, Nicolaïdis S, Pénicaud L Specific inhibition of GLUT2 in arcuate nucleus by antisense oligonucleotides suppresses nervous control of insulin secretion Molecular Brain Research, 1998, 57: 275-280 Leloup C, Magnan C, Benani A, Bonnet E, Alquier T, Offer G, Carrière A, Periquet A, Fernandez Y, Ktorza A, Casteilla L, Pénicaud L Mitochondrial reactive oxygen species are required for hypothalamic glucose sensing Diabetes, 2006, 55(7): 2084-2090 Lenzen S, Panten U Alloxan: history and mechanism of action Diabetologia, 1988, 31: 337-342 Levin BE, Dunn-Meynell AA, Routh VH CNS sensing and regulation of peripheral glucose levels International Review of Neurobiology, 2002, 51: 219-258 Little TJ, Horowitz M, Feinle-Bisset C Role of cholecystokinin in appetite control and body weight regulation Obesity reviews, 2005, 6: 297-306 Loftus TM, Jaworsky DE, Frehywot GL, Townsend CA, Ronnett GV, Lane MD, Kuhajda FP Reduced food intake and body weight in mice treated with fatty acid synthase inhibitors Science, 2000, 288: 2379-2381 Lynch RM, Tompkins LS, Brooks HL, Dunn-Meynell AA, Levin BE Localization of glucokinase gene expression in the rat brain Diabetes, 2000, 49(5):693-700
M Maechler P, Gjinovci A, Wollheim CB Implication of glutamate in the kinetics of insulin secretion in rat and mouse perfused pancreas Diabetes, 2002, 51(suppl. 1): S99-S102 Maekawa F, Toyoda Y, Torii N, Miwa I, Thompson RC, Foster DL, Tsukahara S, Tsukamura H, Maeda KI Localization of glucokinase-like immunoreactivity in the rat lower brain stem: for possible location of brain glucose-sensing mechanisms Endocrinology, 2000, 141: 375-384

- 161 -
Magnan C, Collins S, Berthault MF, Kassis N, Vincent M, Gilbert M, Pénicaud L, Ktorza A, Assimacopoulos-Jeannet F Lipid infusion lowers sympathetic nervous activity and leads to increased β-cell responsiveness to glucose The Journal of Clinical Investigation, 1999, 103(3): 413-419 Marota JJA, Crosby G, Uhl GR Selective effects of pentobarbital and halothane on c-fos and jun-B gene expression in rat brain Anesthesiology, 1992, 77: 365-371 Marty N, Dallaporta M, Foretz M, Emery M, Tarussio D, Bady I, Binnert C, Beermann F, Thorens B Regulation of glucagon secretion by glucose transporter type 2 (glut2) and astrocyte-dependent glucose sensors The Journal of Clinical Investigation, 2005, 115(12): 3545-3553 Mayer J Glucostatic mechanism of regulation of food intake The England journal of medicine, 1953, 249: 3-16 McGowan BMC, Bloom SR Peptide YY and appetite control Current Opinion in Pharmacology, 2004, 4: 583-588 McNay EC, Gold PE Extracellular glucose concentrations in the rat hippocampus measured by zero-net-flux: effects of microdialysis flow rate, strain, and age Journal of Neurochemistry, 1999, 72(2): 785-790 Medina RA, Owen GI Glucose transporters: expression, regulation and cancer Biological Research, 2002, 35: 9-26 Mitro N, Mak PA, Vargas L, Godio C, Hampton E, Molteni V, Kreusch A, Saez E The nuclear receptor LXR is a glucose sensor Nature, 2007, 445: 219-223 Moran O, Phillip M Leptin : obesity, diabetes and other peripheral effects – a review Pediatric Diabetes, 2003, 4: 101-109 Moriguchi T, Sakurai T, Nambu T, Yanagisawa M, Goto K Neurons containing orexin in the lateral hypothalamic area of the adult brain are activated by insulin-induced acute hypoglycemia Neuroscience Letters, 1999, 264: 101-104 Moriyama R, Tsukamura H, Kinoshita M, Okazaki H, Kato Y, Maeda KI In vitro increase in intracellular calcium concentrations induced by low or high extracellular glucose levels in ependymocytes and serotonergic neurons of the rat lower brainstem Endocrinology, 2004, 145(5): 2507-2515

- 162 -
Morton GJ, Cummings DE, Baskin DG, Barsh GS, Schwartz MW Central nervous system control of food intake and body weight Nature, 2006, 443:289-295 Mullen RJ, Buck CR, Smith AM NeuN, a neuronal specific nuclear protein in vertebrates Development, 1992, 116(1): 201-211 Münzberg H, Björnholm M, Bates SH, Myers Jr MG Leptin receptor action and mechanisms of leptin resistance Cellular and Molecular Life Sciences, 2005, 62: 642-652
N Navarro M, de Fonseca FR, Alvarez E, Chowen JA, Zueco JA, Gomez R, Eng J, Blazquez E Colocalization of glucagon-like peptide-1 (GLP-1) receptors, glucose transporter GLUT-2, and glucokinase mRNAs in rat hypothalamic cells: evidence for a role of GLP-1 receptor agonists as an inhibitory signal for food and water intake Journal of Neurochemistry, 1996, 67: 1982-1991 Ngarmuko C, Baur EL, Kumagai AK Co-localization of GLUT1 and GLUT4 in the blood-brain barrier of the rat ventromedial hypothalamus Brain Research, 2001, 900: 1-8 Niswender KD, Baskin DG, Schwartz MW Insulin and its evolving partnership with leptin in the hypothalamic control of energy homeostasis TRENDS in Endocrinology and Metabolism, 2004, 15(8): 362-369 Nonogaki K New insights into sympathetic regulation of glucose and fat metabolism Diabetologia, 2000, 43: 533-549
O Obici S, Feng Z, Morgan K, Stein D, Karkanias G, Rossetti L Central administration of oleic acid inhibits glucose production and food intake Diabetes, 2002a, 51: 271-275 Obici S, Feng Z, Karkanias G, Baskin DG, Rossetti L Decreasing hypothalamic insulin receptors causes hyperphagia and insulin resistance in rats Nature Neuroscience, 2002b, 5(6): 566-572

- 163 -
Obici S, Rossetti L Minireview: nutrient sensing and the regulation of insulin action and energy balance Endocrinology, 2003, 144(12): 5172-5178
P Paemeleire K The cellular basis of neurovascular metabolic coupling Acta Neurologica Belgica, 2002, 102: 153-157 Paquot N, Scheen AJ, Lefèbvre PJ Physiologie de l’hyperglycémie post-prandiale Médecine Clinique, endocrinologie & diabète, 2003, hors-série n°1, janvier-février Pardal R, Lopez-Barneo J Low glucose-sensing cells in the carotid body Nature Neuroscience, 2002, 5(3): 197-198 Paxinos G, Watson C The rat brain in stereotaxic coordinates Academic Press, Sydney, 1986, 2nd edition Pénicaud L, Cousin B, Leloup C, Atef N, Casteilla L, Ktorza A Changes in autonomic nervous system activity and consecutive hyperinsulinaemia: respective roles in the development of obesity in rodents Diabetes & Metabolism (Paris), 1996, 22: 15-24 Pénicaud L, Leloup C, Lorsignol A, Alquier T, Guillod E Brain glucose sensing mechanism and glucose homeostasis Current Opinion in Clinical Nutrition and Metabolic Care, 2002, 5: 539-543 Pritchard LE, Turnbull AV, White A Pro-opiomelanocortin processing in the hypothalamus: impact on melanocortin signalling and obesity Journal of Endocrinology, 2002, 172: 411-421 Püschel GP Control of hepatocyte metabolism by sympathetic and parasympathetic hepatic nerves The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology, 2004, 280A(1): 854-867
Q Quast U La structure des canaux potassiques ATP-dépendants : complexe moléculaire SUR/Kir6 Journées annuelles de diabétologie de l’Hôtel Dieu, 2000, 1-12

- 164 -
R Ramos EJB, Meguid MM, Campos ACL, Coelho JCU Neuropeptide Y, α-Melanocyte-stimulating hormone, and monoamines in food intake regulation Nutrition, 2005, 21: 269-279 Randle PJ, Garland PB, Hales CN, Newsholme EA The glucose-fatty acid cycle: its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus Lancet, 1963, 1: 785-789 Rohner-Jeanrenaud F, Bobbioni E, Ionescu E, Sauter JF, Jeanrenaud B Central nervous system regulation of insulin secretion Advances in Metabolic Disorders, 1983, 10: 193-219 Russek M Demonstration of the influence of an hepatic glucosensitive mechanism on food-intake Physiology & Behavior, 1970, 5(10): 1207-1209
S Sabatier N, Caquineau C, Dayanithi G, Bull P, Douglas AJ, Guan XMM, Jiang M, Van der Ploeg L, Leng G α-Melanocyte-stimulating hormone stimulates oxytocin release from the dendrites of hypothalamic neurons while inhibiting oxytocin release from their terminals in the neurohypophysis The Journal of Neuroscience, 2003, 23(32): 10351-10358 Sagar SM, Sharp FR, Curran T Expression of c-fos protein in brain: metabolic mapping at the cellular level Science, 1988, 240: 1328-1331 Sahu A Leptin signaling in the hypothalamus: emphasis on energy homeostasis and leptin resistance Frontiers in Neuroendocrinology, 2004, 24: 225-253 Sakaguchi T, Bray GA Ventromedial hypothalamic lesions attenuate responses of sympathetic nerves to carotid arterial infusions of glucose and insulin International Journal of Obesity, 1990, 14: 127-134 Sanchis-Segura C, Aragon CMG Consequences of monosodium glutamate or goldthioglucose arcuate nucleus lesions on ethanol-induced locomotion Drug and Alcohol Dependence, 2002, 68: 189-194

- 165 -
Sanders NM, Dunn-Meynell AA, Levin BE Third ventricular alloxan reversibly impairs glucose counterregulatory responses Diabetes, 2004, 53: 1230-1236 Scheurink AJW, Nolan LJ Food intake, Fuel homeostasis, and Autonomic Nervous system Appetite, 1996, 26: 304 Schneid C, De Bandt JP, Cynober L, Torres E, Reach G, Darquy S In vivo induction of insulin secretion by ornithine α-ketoglutarate: involvement of nitric oxide and glutamine Metabolism, 2003, 52(3): 344-350 Schuit FC, Huypens P, Heimberg H, Pipeleers DG Glucose sensing in pancreatic β-cells. A model for the study of other glucose-regulated cells in gut, pancreas, and hypothalamus Diabetes, 2001, 50:1-11 Schwartz MW, Figlewicz DP, Baskin DG, Woods SC, Porte D Jr Insulin in the brain: a hormonal regulator of energy balance Endocrine Reviews, 1992, 13(3): 387-414 Schwartz MW, Woods SC, Porte D Jr, Seeley RJ, Baskin DG Central nervous system control of food intake Nature, 2000, 404: 661-671 Sergeyev V, Broberger C, Gorbatyuk O, Hokfelt T Effect of 2-mercaptoacetate and 2-deoxy-D-glucose administration on the expression of NPY, AGRP, POMC, MCH and hypocretin/orexin in the rat hypothalamus Neuroreport, 2000, 11: 117-121 Shu S, Ju G, Fan L The glucose oxidase-DAB-nickel method in peroxidase histochemistry of the nervous system Neuroscience Letters, 1988, 85: 169-171 Song Z, Levin BE, McArdle JJ, Bakhos N, Routh VH Convergence of pre- and postsynaptic influences on glucosensing neurons in the ventromedial hypothalamic nucleus Diabetes, 2001, 50: 2673-2681 Stockhorst U, de Fries D, Steingrueber HJ, Scherbaum WA Insulin and the CNS: effects on food intake, memory, and endocrine parameters and the role of intranasal insulin administration in humans Physiology & Behavior, 2004, 83: 47-54 Stütz AM, Morrison CD, Argyropoulos G The Agouti-related protein and its role in energy homeostasis Peptides, 2005, 26: 1771-1781

- 166 -
Swanson LW, Sawchenko PE Paraventricular nucleus: a site for the integration of neuroendocrine and autonomic mechanisms Neuroendocrinology, 1980, 31(6):410-417
T Tabuchi E, Yokawa T, Mallick H, Inubushi T, Kondoh T, Ono T, Torii K Spatio-temporal dynamics of brain activated regions during drinking behavior in rats Brain Research, 2002, 951: 270-279 Tao YX, Segaloff DL Functional characterization of melanocortin-4 receptor mutations associated with childhood obesity Endocrinology, 2003, 144(10):4544-4551 Thorens B, Sarkar HK, Kaback HR, Lodish HF Cloning and functional expression in bacteria of a novel glucose transporter present in liver, intestine, kidney, and β-pancreatic islet cells Cell, 1988, 55: 281-290 Thorens B Molecular and cellular physiology of GLUT-2, a high-Km facilitated diffusion glucose transporter International Review of Cytology, 1992, 137A: 209-238 Tsacopoulos M, Magistretti PJ Metabolic coupling between glia and neurons The Journal of Neuroscience, 1996, 16(3): 877-885
U Ubink R, Calza L, Hökfelt T “Neuro”-peptides in glia: focus on NPY and galanin TRENDS in Neurosciences, 2003, 26(11): 604-609 Ueno H, Yamaguchi H, Kangawa K, Nakazato M Ghrelin: a gastric peptide that regulates food intake and energy homeostasis Regulatory Peptides, 2005, 126: 11-19

- 167 -
V Van Obberghen E, Baron V, Delahaye L, Emanuelli B, Filippa N, Giorgetti-Peraldi S, Lebrun P, Mothe-Satney I, Peraldi P, Rocchi S, Sawka-Verhelle D, Tartare-Deckert S, Giudicelli J Surfing the insulin signaling web European Journal of Clinical Investigation, 2001, 31: 966-977 Vercelli A, Repici M, Garbossa D, Grimaldi A Recent techniques for tracing pathways in the central nervous system of developing and adult mammals Brain Research Bulletin, 2000, 51(1): 11-28
W Wan HZ, Hulsey MG, Martin RJ Intracerebroventricular administration of antisense oligodeoxynucleotide against GLUT2 glucose transporter mRNA reduces food intake, body weight change and glucoprivic feeding response in rats The Journal of Nutrition, 1998, 128: 287-291 Wang R, Liu X, Hentges ST, Dunn-Meynell AA, Levin BE, Wang W, Routh VH The regulation of glucose-excited neurons in the hypothalamic arcuate nucleus by glucose and feeding-relevant peptides Diabetes, 2004, 53: 1959-1965 Weide K, Christ N, Moar KM, Arens J, Hinney A, Mercer JG, Eiden S, Schmidt I Hyperphagia, not hypometabolism, causes early onset obesity in melanocortin-4 receptor knockout mice Physiological Genomics, 2003, 13(1): 47-56 Wells S, Vosseller K, Hart GW A role for N-acetylglucosamine as a nutrient sensor and mediator of insulin resistance Cellular and Molecular Life Sciences, 2003, 60: 222-228 Williams G, Cai XJ, Elliott JC, Harrold JA Anabolic neuropeptides Physiology & Behavior, 2004, 81: 211-222 Woods SC, Benoit SC, Clegg DJ, Seeley RJ Regulation of energy homeostasis by peripheral signals Best Practice & Research, Clinical Endocrinology & Metabolism, 2004, 18 (4): 497-515 Woods SC Signals that influence food intake and body weight Physiology & Behavior, 2005, 86: 709-716

- 168 -
Wynne K, Stanley S, McGowan B, Bloom S Appetite Control Journal of Endocrinology, 2005, 184: 291-318
X Xu B, Goulding EH, Zang K, Cepoi D, Cone RD, Jones KR, Tecott LH, Reichardt LF Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor Nature Neuroscience, 2003, 6(7):736-742 Xue B, Kahn BB AMPK integrates nutrient and hormonal signals to regulate food intake and energy balance through effects in the hypothalamus and peripheral tissues The Journal of Physiology, 2006, 574(Pt 1):73-83
Y Yang XJ, Kow LM, Funabashi T, Mobbs CV Hypothalamic glucose sensor: similarities to and differences from pancreatic β-cell mechanisms Diabetes, 1999, 48: 1763-1772 Yaswen L, Diehl N, Brennan MB, Hochgeschwender U Obesity in the mouse model of pro-opiomelanocortin deficiency responds to peripheral melanocortin Nature Medicine, 1999, 5(9):1066-1070 Yi CX, van der Vliet J, Dai J, Yin G, Ru L, Buijs R Ventromedial arcuate nucleus communicates peripheral metabolic information to the suprachiasmatic nucleus Endocrinology, 2006, 147(1): 283-294 Yoshikawa H, Tajiri Y, Sako Y, Hashimato T, Umeda F, Nawata H Glucosamine-induced β-cell dysfunction: a possible involvement of glucokinase or glucose-transporter type 2 Pancreas, 2002, 24 (3): 228-234 Young JK, Baker JH, Montes MI The brain response to 2-deoxyglucose is blocked by a glial drug Pharmacology, Biochemistry and Behaviour, 2000, 67: 233-239 Young JK Anatomical relationship between specialized astrocytes and leptin-sensitive neurones Journal of Anatomy, 2002, 201: 85-90

- 169 -
Young JK, McKenzie JC GLUT2 immunoreactivity in Gomori-positive astrocytes of the hypothalamus Journal of Histochemistry & Cytochemistry, 2004, 52(11): 1519-1524
Z Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM Positional cloning of the mouse obese gene and its human homologue Nature, 1994, 372: 425-432 Zheng H, Patterson LM, Phifer CB, Berthoud H-R Brain stem melanocortinergic modulation of meal size and identification of hypothalamic POMC projections The American Journal of Physiology – Regulatory, Integrative and Comparative Physiology, 2005, 289: 247-258

- 170 -
Résumé :
Sensibilité au glucose du cerveau chez le rat : implication de l’hypothalamus,
détermination des acteurs cellulaires et moléculaires
Le contrôle de l’homéostasie énergétique met en jeu des interactions entre la
périphérie et le cerveau, de natures hormonale, métabolique et nerveuse. Notre
projet s’est porté sur la sensibilité cérébrale au glucose dans l’hypothalamus,
nécessaire au maintien de la normoglycémie. Tout d’abord, nous avons mis en
évidence l’activation spécifique de noyaux hypothalamiques grâce à l’expression de
c-Fos induite par l’injection aiguë de glucose par la carotide vers le cerveau. Nos
résultats indiquent que des neurones sont majoritairement impliqués et que la
population d’astrocytes du noyau arqué est essentielle à cette réponse cérébrale.
Enfin, la détection de l’hyperglycémie centrale fait intervenir la glucokinase, enzyme
assurant la première étape de la glycolyse par la phosphorylation du glucose.
Glucose-sensing in rat brain: involvement of the hypothalamus, determination
of cellular and molecular actors
Energy homeostasis is a well regulated mechanism that depends on
hormonal and nervous regulation. With this aim in view, central nervous system
continuously integrates hormonal and metabolic signals to elicit adaptive
responses. The present investigation aims to elucidate the mechanism of the
cerebral glucose-sensing in the hypothalamus, i.e. the ability of this cerebral area to
detect glycaemia changes. We have shown that the acute injection of glucose by the
carotid towards the brain activates specific hypothalamic nuclei as revealed by c-
Fos expression. Our results indicate that it mainly implies neurons and that
astrocytes in the arcuate nucleus are essential for this cerebral response. Finally,
glucokinase, which assumes the glucose phosphorylation in the first step of the
glycolyse, is crucial for the detection of central hyperglycaemia.

- 171 -
Thèse soutenue par Elise Guillod-Maximin
Intitulée Sensibilité au glucose du cerveau chez le rat : implication de
l’hypothalamus, détermination des acteurs cellulaires et moléculaires
Dirigée par le Dr. Luc Pénicaud
A l’Université Toulouse III – Paul Sabatier (UFR SVT)
Le 15 octobre 2007
Résumé : Le contrôle de l’homéostasie énergétique met en jeu des interactions
entre la périphérie et le cerveau, de natures hormonale, métabolique et
nerveuse. Notre projet s’est porté sur la sensibilité cérébrale au glucose dans
l’hypothalamus, nécessaire au maintien de la normoglycémie. Tout d’abord,
nous avons mis en évidence l’activation spécifique de noyaux
hypothalamiques grâce à l’expression de c-Fos induite par l’injection aiguë de
glucose par la carotide vers le cerveau. Nos résultats indiquent que des
neurones sont majoritairement impliqués et que la population d’astrocytes du
noyau arqué est essentielle à cette réponse cérébrale. Enfin, la détection de
l’hyperglycémie centrale fait intervenir la glucokinase, enzyme assurant la
première étape de la glycolyse par la phosphorylation du glucose.
Mots clés : glucose – cerveau – hypothalamus – noyau arqué – astrocyte –
glucokinase – insuline – c-Fos – méthionine sulfoximine – glucosamine – rat
Discipline : Physiopathologie moléculaire, cellulaire et intégrée
Laboratoire de Neurobiologie, Plasticité Tissulaire et Métabolisme Energétique
UMR5018-CNRS, IFR31, Institut Louis Bugnard, BP 84225
31432 TOULOUSE Cedex 4