
EXTRAIT DU RÈGLEMENT -...
38
Transcript of EXTRAIT DU RÈGLEMENT -...


EXTRAIT DU RÈGLEMENT
Le nombre des membres de la Société est illimité. Pour faire partie de la Société, il faut s'être fait présenter dans une des séances par deux membres et avoir été proclamé dans la séance suivante par le Président, après élection.
l a cotisation annuelle est fixée à trente francs. Un droit d'entrée de dix francs est dû par les nouveaux membres. Les cotisations doivent être versées dans le premier trimestre de chaque année (C. Ch. Postaux Nancy 45.24). En cas de non versement au premier avril, elles seront recouvrées par la poste contre un reçu de trente-cinq francs.
La Société tient ses séances mensuelles régulières à son siège social (Institut de Zoologie, 30, rue Sainte-Catherine, à Nancy), en principe le 2· vendredi de chaque mois à 17 heures, sauf pendant les mois d'août, de septembre et d'octobre. Elle tient en outre des séances volantes pour des conférences, visites et excursions, aux dates, lieux et heures indiqués aux sociétaires par des avis spéciaux ou par la voie de la presse locale.
Les ouvrages de la bibliothèque de la Société sont conservés à la bibliothèque de la Ville de Nancy, ce "qui, en vertu d'une entente avec la Municipalité, donne aux membres de la Société le droit de. recevoir un prêt temporaire, non seulement des ouvrages de la bibliothèque particulière de la Société, mais aussi de la bibliothèque générale de la Ville de Nancy.
La Société publie des Mémoires et un Bulletin Mensuel. Le Bulletin paraît régulièrement en neuf numéros mensuels par an. Il est envoyé gratuitement aux membres, numéro par numéro.
Les Mémoires, sous forme de numéros bis, sont publiés lorsque la situation financière de la Société le permet. Ils ne sont envoyés gratuitement qu'aux membres qui en font la demande, en joignant la somme nécessaire pour les frais de poste.
Publications dans le Bulletin et les Mémoires
Les notes destinées au Bulletin et aux Mémoires ne sont publiées qu'après leur examen par la Commission et avis du Conseil. Ce dernier, considération faite de l'état des finances de la Société, est toujours habilité à demander à l'auteur une contribution financière plus ou moins importante.
Les illustrations sont toujours à la charge de l'auteur; mais la commission du bulletin en règle l'exécution.
La Société n'offre pas de tirés à part; mais les auteurs peuvent en obtenir è leurs frais; dans ce cas, ils doivent le mentionner d'une façon très apparente sur la première page de leur manuscrit en indiquant le nombre de tirés à part, et s'ils désirent une couverture, avec ou sans titre.
Pour 1938, le prix des tirés à part sur papier du Bulletin est fixé ainsi qu'il suit. Le montant en est versé entre les mains du Trésorier sur présentation de facture.
PAR 25 EN EXEMPLAIRES 25 50 75 100 PLUS
Une feuille (16 pages) 45 00 75 00 20 3/4 feoille (12 pages) 35 50 65 75 16 1/2 feuille ( 8 pages) 25 35 45 50 10 1/4 feuille ( 4 pages) 15 20 25 30 6
La couverture tst comptée comme 1/4 de feuille. La planche tirée à part est comptée pour 1/4 de feuille. Les tirés à part peuvent être exécutés à la demande des auteurs sur un papier
différent Les prix sont alors fixés par l'imprimeur.
debeaupu
Crayon

SOCIÉTÉ DES SCIENCES DE NANCY
Convocation
La prochaine réunion mensuelle de la Société des Sciences de Nancy aura lieu le Vendredi n Février 1938, à i/ heures, à r ins t i tu t de Zoologie de la Faculté des Sciences, rue Sainte-Catherine.
Assemblée générale. Modification des statuts.
O R D R E DU JOUR
Communications: M. E R R A R D : Les cavernes de la vallée de l'Esse et leurs rapports avec la tectonique régionale.
Colonel Ch. G É R A R D : Les fossiles types d'espèces conservés dans les collections de Nancy.
Causerie de M. A. R O B A U X : Sondages et hydrologie.

Décembre 1937 Numéro 9
BULLETIN MENSUEL
SOCIÉTÉ DES SCIENCES DB
NANCY (Fondée en 1828)
SIÈGE SOCIAL: Institut de Zoologie, 30, Rue Sainte-Catherine - NANCY
SÉANCE DU 10 DÉCEMBRE 1937
Amphithéâtre de Zoologie de la Faculté des Sciences
Présidence de M· H. Joly
Un nombreux auditoire est présent lorsque M. JOLY ouvre la séance, à 17 heures. Après avoir fait adopter le procès-verbal de la réunion de novembre, M. le Président félicite M. ANZIANI de sa récente nomination au poste de Chef de travaux à la Faculté des Sciences, puis fait part aux membres d'une légère modification apportée à la parution du Bulletin. Pour aider à l'impression convenable de ce compte rendu mensuel de notre activité, il apparaît indispensable de retarder dorénavant d'un mois l'envoi de ce périodique; c'est ainsi que le numéro de décembre paraîtra seulement en Février 1938. Ce léger décalage permettra aux auteurs et à la Rédaction de corriger plus posément les épreuves d'imprimerie. Ce projet est adopté. On passe ensuite à l'élection de cinq membres nouveaux. Plusieurs candidatures sont présentées.
L'ordre du jour appelle une communication de M. P. FLORENTIN sur « Les réflexes photo-pigmentaires chez les Batraciens », puis une causerie de M. CUÉNOT sur « L'adaptation chez les animaux ». Cette très intéressante conférence, qui reprenait dans le domaine zoologique un thème analogue à celui que M. GUINIER avait traité au cours de la dernière réunion, obtint le plus vif succès et fut saluée de chaleureux applaudissements.
La séance est levée à 19 heures. Des convocations spéciales seront envoyées pour la réunion du 14 janvier 1938.

2ÓO SOCIETE DES SCIENCES DE NANCY
ELECTION DE MEMBRES NOUVEAUX
Aucune objection n'ayant été formulée au sujet des candidatures présentées en novembre, MM. W E I S , AUBRY, KLEIN, GROGNOT et
KOCHER ont été nommés, à l'unanimité, membres titulaires de la Société des Sciences de Nancy.
PRÉSENTATION D E MEMBRES NOUVEAUX
M. CERIGHELLI, Professeur à la Faculté des Sciences, Directeur de l'Institut Agricole de l'Université de Nancy, rue Sainte-Catherine, à Nancy, présenté par MM. BLACHE et JOLY.
M. CHALAUD, Maître de Conférences à la Faculté des Sciences, Institut Agricole de l'Université de Nancy, rue Sainte-Catherine, à Nancy, présenté par MM. JOLY et HAMANT.
M. JEAN-PROST (Pierre), Professeur à l'Ecole départementale d'agriculture de Château-Salins, présenté par MM. JOLY et DELA-FOSSE.
M1Ie BILLIONNET, Professeur à l'Ecole primaire supérieure, 5, rue de l'Ecole-Normale, à Nancy, présentée par M. JOLY et M l le TÉTRY.
M. M O I N E , Directeur d'Ecole primaire, à Millery. présenté par MM. JOLY et ROBAUX.
M. l'Abbé M O I N E , Professeur à l'Ecole Saint-Sigisbert, cours Leopold, Nancy, présenté par MM. JOLY et ROBAUX.
COTISATIONS
La Société des Sciences de Nancy exprime sa vive gratitude à ceux de ses membres dont les noms suivent, qui ont bien voulu témoigner du grand intérêt qu'ils lui portent en ajoutant à leur cotisation de 1937 un versement bénévole supplémentaire: MM. ANZIANI, EMÉRIQUE, GARDET, le colonel GÉRARD, JOLY, LEMAIRE, O U D I N , L. PELTIER,
SCHAEFFER.

SOCIÉTÉ DES SCIENCES DE NANCY 2ÓI
COMMUNICATIONS
Les réflexes photo-pigmentaires chez les Batraciens et la notion de physiologie spécifique
PAR
P. FLORENTIN
On sait depuis longtemps que les changements de couleur des téguments, chez certains Vertébrés, sont sous la dépendance de facteurs externes (chaleur, lumière, humidité, terrain) et qu'ils se réalisent spontanément dans la nature au cours des variations de l'éclairement ou de l'alternance des cycles saisonniers. Ils peuvent être reproduits expérimentalement. Les exemples abondent de ces modifications pigmen-taires fugaces ou durables, dont l'établissement est rapide ou progressif et qui, d'après L. CUÉNOT ( I ) , font partie du patrimoine héréditaire des individus tout en conservant pour certains une origine strictement individuelle.
Parmi les espèces animales, fort nombreuses, qui présentent à un degré très accusé ces aptitudes à ce qu'on a coutume d'appeler « l'homochromie », les Batraciens anoures apparaissent nettement privilégiés. Depuis quelques années déjà, l'étude systématique des réflexes photo-pigmentaires entreprise par les physiologistes et les histologîstes chez les Anoures, s'est révélée très féconde en résultats positifs et a conduit à une interprétation séduisante du mécanisme des variations pigmentaires réalisées expérimentalement chez ces animaux. Elle a souligné, en particulier, l'intervention indiscutable, dans le développement complet du processus, de principes endocriniens : le principe mélanophoro-dilatateur d'origine hypophysäre et l'adrénaline d'origine médullo-surrénale, son antagoniste, sans lesquels le phénomène d'homochromie ne peut se produire intégralement. Elle a montré au surplus la mise en

2Ô2 SOCIETE DES SCIENCES DE NANCY
jeu de phénomènes nerveux opposés: sympathique et parasympathique, qui semblent contrôler, sinon déterminer les facteurs endocriniens.
Toutefois, des variations individuelles existent et ne sont pas négligeables : il ne peut évidemment s'agir, dans les expériences entreprises et les conclusions qui en découlent que de phénomènes globaux, que seule l'étude d'un grand nombre d'individus permet de considérer comme présentant un réel intérêt. Il suffit de regarder, par exemple, un aquarium peuplé de Hyla viridis pour se convaincre que Thomochromie est souvent une réaction strictement individuelle, et que, placés dans les mêmes conditions de milieu, deux animaux voisins peuvent être de coloration entièrement différente. Seule, une statistique portant sur un grand nombre d'individus soumis aux mêmes excitants physiologiques définira les tendances générales de l'espèce.
Il peut se manifester par contre, des réactions totalement inversées chez deux espèces voisines et ce phénomène présente une portée biologique beaucoup plus grande. Cette constatation permet d'affirmer en effet qu'il existe chez les Batraciens à côté de ce qu'on peut appeler une « physiologie individuelle », une « physiologie spécifique », départageant d'une manière indiscutable les espèces les plus rapprochées dans la classification zoologique et traduisant ainsi l'adaptation différente de ces espèces vis-à-vis du monde extérieur.
Nous voudrions signaler, dans cette note, quelques faits positifs tendant à illustrer d'une manière frappante cette constatation.
L'expérience a montré qu'une Grenouille (Rana temporaria, Rana escalenta), placée dans l'obscurité complète, contracte ses mélanophores cutanés en un temps moyen de 2 à 3 jours ; elle devient très claire, puis fonce légèrement au 4e jour et se stabilise presque invariablement dans cet état intermédiaire correspondant à la position de repos des cellules pigmentaires (Fig. ι α). Remise à la lumière diffuse sur un fond de couleur neutre, elle reprend sa teinte initiale brun verdâtre, ou rougeâtre, teinte qui subit des variations diurnes et nocturnes légères. Placée sur fond blanc, la Grenouille pâlit fortement (Fig. ι 6), sur fond noir elle étale ses
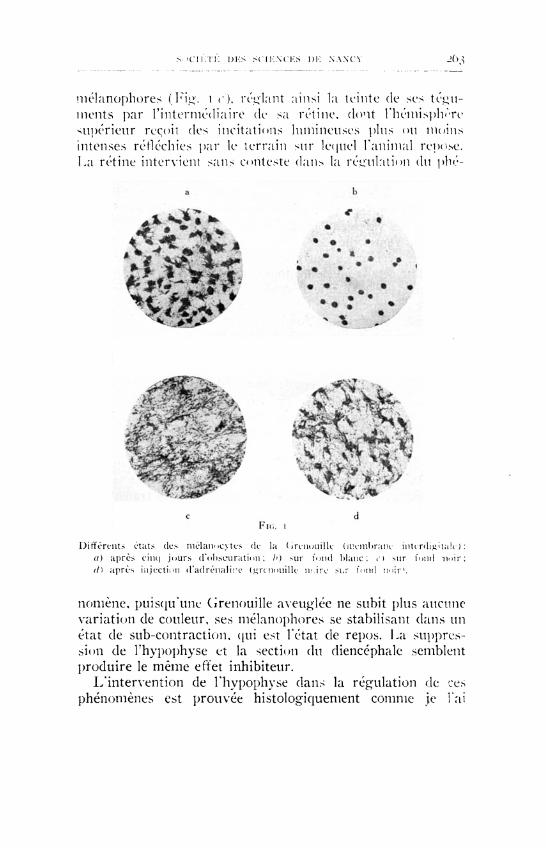
s icnVn: iJKS s c U : \ C K S DK NANCV J O ^
mélanophores (Kig. ι c), rel iant ainsi la teinte de ses téguments par l'intermédiaire de sa rétine, dont l'hémisphère supérieur reçoit des incitations lumineuses plus ou moins intenses réfléchies par le terrain sur lequel l'animal repose. La rétine intervient sans conteste dans la régulation du phé-
+ % H * * · *
•
Y*"'
"*. V " · '* «te *
-"?-•''• ' '·'·
> '.jagst; ^rfS» :,.- ν I'PMÎ^ ,-*."" >/;£#··£
. . ώ ^ ' - '
, "V*
*,>
; , , ; „ ·
; . ? _ . - . ; * ' ;
#
< '%.f + - ,vV"%-< "
' . ^ ' ^ '
-v. Ί ^
*f->
1? , c*
.. % .·
ν I
$ k
•C y
W %4
*\ %Λ*ί,
· * " * * * •
^ . '-*•• V
y is·'
/" i f Ma
Différents états des melanocytes de la Grenouille (membrane interdigitale): a) après cinq jours d'obseniration ; h) sur fond Mane; e ) sur fond noir : (i) après injection d 'adrénal i re (.urcnouille m i r e sur fond imir1.
noniène, puisqu'une Cirenouille aveuglée ne subit plus aucune variation de couleur, ses mélanophores se stabilisant dans un état de sub-contraction, qui est l'état de repos. La suppression de l'hypophyse et la section du diencéphale semblent produire le même effet inhibiteur.
L'intervention de l'hypophyse dans la régulation de ces phénomènes est prouvée histologiquement comme je l'ai
debeaupu
Crayon

2Ó4 SOCIÉTÉ DES SCIENCES DE NANCY
montré avec STUTINSKY (2). L'hypophyse d'une Grenouille (Rana temporaria L.) placée dans l'obscurité absolue prend des caractères cytologiques particuliers et, dans la plupart des cas étudiés, devient intensément eosinophile; on observe en outre une contraction très nette des cellules hypophysai-res et une suspension notable de l'élaboration et de l'excrétion de la substance colloïde, sécrétion visible de la pars intermedia reprise par le lobe nerveux et normalement déversée dans les vaisseaux sanguins ou le liquide céphalo-rachidien du 3e ventricule.
J'ai pu vérifier en outre qu'une Grenouille placée sur un fond blanc depuis plusieurs jours et dont la peau est devenue très claire présente une hypophyse à prédominance eosinophile (se rapprochant de la structure d'obscuration) et qu'une Grenouille placée sur fond noir et dont les téguments ont fortement noirci, possède une hypophyse en activité secrétoîre évidente: les cellules hypophysaires sont volumineuses, claires ou cyanophiles, et le pourcentage des cellules eosinophiles est considérablement réduit; d'autre part, le lobe intermédiaire semble en voie d'excrétion active et le lobe nerveux, dans ce dernier cas, est fortement imprégné de colloïde hydratée, principalement localisée au pourtour des vaisseaux élargis.
Il existe donc, chez la Grenouille, des réflexes photo-pigmentaires dont le point de départ doit être situé au niveau de la rétine; l'excitation de cette membrane se transmet à l'hypophyse par un relai diencéphalique probable, commandant lui-même l'excrétion par la glande d'un principe mélano-phoro-dilatateur actif. Le phénomène paradoxal réside toutefois dans ce fait que la lumière blanche provoque, comme l'obs-curation complète, une inhibition de l'hypophyse et une contraction des mélanophores. Il existe sans conteste un optimum de lumière pour lequel la sécrétion hypophysäre mélano-phorotrope s'établit, et cet optimum est réalisé pour les faibles intensités lumineuses. N'oublions pas aussi que le principe hypophysäre n'est pas le seul produit capable d'agir sur les mélanophores et que l'adrénaline intervient dans la contraction des cellules pigmentaires (fig. 1 d). Des dosages d'adrénaline dans la glande surrénale des Grenouilles, efifectués

SOCIÉTÉ DES SCIENCES DE NANCY 265
suivant une méthode physiologique par R. COLLIN, Th. FONTAINE et L. HENNEOUIN (3) ont montré que, chez les Grenouilles obscurées, l'excrétion de l'adrénaline est ralentie, et qu'au contraire, chez les Grenouilles blanches sur fond blanc, l'adrénaline a été excrétée dans une certaine mesure, et agit probablement en contractant les mélanophores. Cette interprétation n'est valable qu'en raison du phénomène que nous avons constaté plus haut : l'inhibition de l'hypophyse chez ces animaux, histologiquement confirmée.
La Grenouille a servi jusqu'à présent d'objet courant pour les recherches expérimentales sur les réflexes photo-pigmen-taires; toutefois l'étude d'une seule espèce ne permet pas de tirer de conclusions absolues sur le déterminisme de ces réflexes. Comme nous allons le voir maintenant, l'étude d'un Anoure qui semble très voisin, le Crapaud, va nous donner des résultats absolument inversés.
J'ai pu réaliser sur un lot assez important de Bufo vulgaris (une trentaine d'individus mâles et quelques femelles récoltés au moment du frai, en mars 1937), une série d'expériences surperposables à celles qui ont été décrites ci-dessus, et pratiquées sur la Grenouille.
Un lot de Crapauds a été placé dans l'obscurité absolue, deux autres groupes à la lumière diffuse sur fond blanc ou sur fond noir, quelques exemplaires enfin furent soumis à un éclairement artificiel continu, sur fond blanc, pendant une semaine. Les résultats enregistrés furent les suivants: six individus sur sept devinrent intensément noirs à l'obscurité complète, et ceci dès la seconde journée d'obscuration. Le même phénomène se produisit chez les Crapauds éclairés artificiellement. Les deux autres groupes enfin ne subirent pas de modifications tégumentaires notables; sur fond noir ou sur fond blanc, à la lumière du jour dififuse, les téguments demeurèrent assez pâles, d'une teinte jaune grisâtre uniforme. Quelques-uns de ces individus clairs furent placés ensuite dans l'obscurité totale: leurs melanocytes se dilatèrent au bout de 48 heures en moyenne. Une femelle claire, et demeurée telle sur fond noir et sur fond blanc, fut éclairée artificiellement pendant 8 jours consécutifs et présenta une intense dilatation des melanocytes au bout du troisième jour d'éclai-rement

266 SOCIÉTÉ DES SCIENCES DE NANCY
Il résulte de ces expériences préliminaires (que nous avons l'intention de poursuivre d'une manière systématique et avec plus de précision dès que nous pourrons nous procurer à nouveau une certaine quantité de Crapauds) que l'obscuration totale, comme l'irradiation continue, provoque chez Bufo vulgaris une dilatation extrême des mélanophores cutanés, tandis que la lumière diffuse et faible demeure sans action sur les téguments. Cest exactement l'inverse de ce que nous avons constaté chez la Grenouille.
Pour parfaire ces observations, j 'ai examiné histologîque-ment l'hypophyse et l'encéphale des animaux en expérience. Les individus obscures ou irradiés artificiellement et dont les mélanophores étaient dilatés ont présenté au bout de quelques jours des signes d'hyperactivité hypophysaire, avec imprégnation colloïde des centres diencéphaliques s'accom-pagnant de réactions neuronales accentuées, aboutissant même à la division mitotique de certains neurones du noyau préoptique. Les autres individus ne présentèrent aucune modification importante de leur complexe infundibulo-pituitaire. Les détails de ces modifications histologiques ont été donnés dans un note récente à la Société de Biologie de Nancy (4) à laquelle on pourra se reporter.
Ces expériences prouvent, en résumé, que la physiologie de chaque espèce est soumise à des lois définitives, préétablies, différentes pour chaque espèce, et que, dans l'espèce même, ces lois sont sujettes à des variations individuelles, de faible importance il est vrai, mais dont il est nécessaire de tenir compte lorsqu'on veut établir des statistiques et, partant, des principes fondamentaux. Nous ne pouvons conclure, en ce qui concerne les réflexes photo-pigmentai-res, que pour une espèce déterminée sans qu'il nous soit permis d'étendre nos conclusions à un groupe et a fortiori à une classe entière d'animaux, comme on est si souvent tenté de le faire en physiologie générale. Le fait qu'une Grenouille pâlit à l'obscurité et qu'au contraire un Crapaud obscure noircit vient nous prouver que ces deux espèces voisines sont différemment adaptées au milieu, et en particulier aux conditions d'éclairement. Nous le supposions déjà, en considérant le comportement de chaque espèce: le Crapaud

SOCIÉTÉ DES SCIENCES DE NANCY 267
étant plus franchement obscuricole que la Grenouille. Mais la notion qui se dégage des expériences précédentes est que l'adaptation spécifique comporte une série de réactions histo-physiologiques contrôlables au microscope: elle résulte, dans le cas concret qui nous occupe, soit de Tactivation, soit de l'inhibition de l'hypophyse, sous le contrôle de la rétine visuelle, réactions inverses déclenchées par un facteur identique : la suppression de la lumière.
Des recherches ultérieures me permettront peut être d'étendre à d'autres animaux cette notion si intéressante de la physiologie spécifique et individuelle, qui contribue à élargir le champ de nos investigations et à renforcer l'idée que l'individu, comme l'espèce, présente une incontestable autonomie.
(Laboratoire d'Histologie de la Faculté de Médecine)
1. CUÉNOT (L.). — La genèse des espèces animales, 2e édition, 1921,
P· 495· 2. C. R. Soc. Biol., t. 122, 1936, p. 674. 3. Congrès Ass. Physiol. Paris, juin 1937, pp. 248-250. 4. C. R. Soc. Biol, t. 126, 1937, pp. 33T-334, 1 H-

268 SOCIÉTÉ DES SCIENCES DE NANCY
L'adaptation chez les animaux
(Causerie avec présentat ion d'échantillons)
PAR
L . CUÉNOT
Le mot adaptation vient du latin adaptare, ajuster à; il signifie exactement un ajustement de l'organisme aux conditions extérieures de l'existence ; il renferme une idée de convenance, de rapport, bien plus que de nécessité ou même d'utilité; on peut dire aussi qu'une adaptation est une solution à un problème posé, le résultat devant être la persistance d'une formule de vie. Je citerai deux exemples : l'Insecte, avec son curieux appareil trachéen dont les ramifications vont porter l'air jusqu'aux cellules mêmes, est essentiellement terrestre; or, il y a un certain nombre de formes qui sont devenues aquatiques, et qui ont eu par suite à résoudre le problème de respirer dans l'eau avec un appareil fait pour prendre l'oxygène dans l'air ; comme on sait, les solutions sont variées : le Dytique vient à la surface capter une provision d'air sous ses élytres; la Nèpe l'aspire au moyen d'un long tube, de même que la larve d'Eristale dont le corps s'allonge en tuyau de lorgnette; d'autres, devenus vraiment animaux d'eau, ont trouvé une solution plus originale avec les trachéo-branchies qui prennent l'oxygène par osmose.
Autre exemple emprunté au règne végétal: M. GUINIER, dans sa charmante causerie de la séance précédente, vous a parlé de l'adaptation des plantes à l'hiver : les arbres perdent leurs feuilles et échappent ainsi aux dangers de la dessica-tion; d'autres plantes du type Laurier ont un feuillage persistant, mais leurs cellules sont organisées de telle façon que la transpiration est réduite au minimum; d'autres encore, les annuelles, font le sacrifice de l'individu, et laissent des graines protégées qui, l'hiver passé, donneront de nouveaux pieds.

SOCIÉTÉ DES SCIENCES DE NANCY 269
YJ adaptation spécifique est le phénomène global de la persistance d'une espèce; par définition, toute forme qui vit et qui dure est adaptée d'une façon nécessaire et suffisante au milieu qu'elle habite. Cette adaptation globale est une somme d'aptitudes physiologiques et de conformations organiques s'accordant aux conditions de l'habitat; certains dispositifs se répètent, chez les animaux ayant le même genre de vie, avec une telle similitude qu'ils deviennent quasi caractéristiques de celui-ci: ce sont les adaptations morphologiques: des membres conformés en nageoires comme ceux des Poissons, des Ichthyosaures, des Cétacés, des Phoques, des pattes palmées comme celles des Oiseaux, des Loutres, des rames de Dytiques, des queues aplaties transversalement comme celles des Serpents marins, des Crocodiliens, du Desman, du Potamogale, sont si habituels chez les animaux aquatiques qu'ils permettent au zoologiste de présumer du genre de vie d'individus, même lorsqu'il n'en voit que la dépouille ou l'empreinte. Sans doute, il pourra se tromper (très rarement), car la Nature a parfois une même solution pour des fins différentes: ainsi le Palmatogecko Rangez, Gecko des dunes côtières du sud de l'Afrique, a une palmure notable aux deux paires de pattes; or, il n'a jamais été aquatique, pas plus que les autres Geckos ; ce doit être une adaptation à la marche sur du sable fin et mobile; le Ptychozoon, Gecko de Malaisie, qui vit sur des troncs d'arbres, a aussi des palmures (et aussi des expansions sur les flancs et les bords de la queue aplatie de bas en haut) ; il est bien probable que cela répond à quelque condition de la vie de ce Lézard.
Dans un sens opposé, le zoologiste rencontrera deux authentiques Oiseaux d'eau, le Râle aquatique et la Poule d'eau, qui plongent et nagent avec facilité, bien que les doigts soient complètement libres. Nous verrons plus loin la signification de cette anomalie.
Mais à part ces quelques exceptions, du reste explicables, la conformation morphologique est bien révélatrice du genre de vie. Pour préciser ce rapport, qu'il est essentiel de mettre hors de doute, nous allons l'exprimer numériquement. Je prends un livre de Brasil sur Les Oiseaux d'eau, de rivage et de marais cfe France, et je relève le nombre des genres cités

2JQ SOCIÉTÉ DES SCIENCES DE NANCY
(113) en notant l'état de la palmure: il y en a 60 % qui ont des pattes palmées ou garnies de larges festons, et 40 % de non palmées; une autre statistique donnée par RABAUD dit 65 et 35 %. Parmi les Oiseaux d'eau, il y a donc une évidente majorité de palmés ; ce sont les Palmipèdes de l'ancienne classification, vraiment aquatiques, nageant fréquemment et facilement; ceux qui ne le sont que peu ou point (les Echas-siers de l'ancienne classification) sont des Oiseaux de rivage ou de plaines humides; la preuve de la corrélation est encore renforcée si l'on constate que la palmure complète n'existe chez aucun Oiseau franchement terrestre. Cependant la palmure n'est pas une nécessité, car les deux Rallides aquatiques cités plus haut ont les doigts entièrement libres. Cette convenance indiscutable entre la vie aquatique et la morphologie est ce que j 'ai appelé Xadaptation statistique ou étholo-giqne. On a trouvé assez bizarre ce mot de statistique ; il me paraît cependant significatif, en ce sens qu'il marque bien un rapport de fréquence, dans lequel n'intervient pas forcément la nécessité. Le parachute que l'on rencontre chez divers Mammifères arboricoles (Marsupiaux, Ecureuils, Galéopi-thèque) est aussi, bien que présenté par une petite minorité, une adaptation statistique, parce qu'on ne le trouve jamais chez les non-arboricoles.
Nous avons vu que l'adaptation statistique n'est pas nécessaire ; est-elle au moins utile à ceux qui, par fortune, la possèdent? Rien n'est plus confus que la notion d'utilité; toute chose a un bon et un mauvais côté; tel dispositif morphologique, très avantageux à certains moments, est gênant à d'autres, et il n'est pas facile de savoir de quel côté penche la balance. Toute adaptation morphologique est une spécialisation, et qui dit spécialisation dit restriction de la liberté d'action. Il est extrêmement probable qu'un Oiseau à pattes palmées nage avec plus d'aisance et de rapidité et moins de fatigue qu'un autre de même volume qui a les doigts séparés, mais le premier sera un mauvais marcheur (pensez à un Canard) et ne pourra courir sur un sol raboteux ; sa patoure, avantageuse quand il est dans l'eau, restreint donc ses possibilités; il ne pourra guère s'éloigner du rivage. Qu'un Ecureuil volant, comme le Sciuroptère qui a tfne membrane

SOCIÉTÉ DES SCIENCES DE NANCY 27I
étendue sur les flancs, puisse franchir en sautant un espace plus grand qu'un Ecureuil ordinaire, cela n'est pas douteux; maïs quand il grimpe ou marche, son patagium n'est-il pas encombrant ? Le Galéopithèque, type de grand planeur, grimpe très mal, comme une Chauve-Souris ; à terre il marche aussi péniblement qu'un Phoque; il paraît qu'il ne peut planer que lorsqu'il atteint une certaine taille; assurément il ne réalise pas un progrès sur l'agile Ecureuil, abondant en espèces et en individus et à large extension géographique En Australie, les Phalangers volants sont plus rares que les Phalangers sans patagium. Le Phoque, admirable nageur, est très empêtré quand il prend terre à l'époque de la reproduction, d'où la facilité du massacre par les chasseurs de fourrures. Une plante entomophile, dont les fleurs sont exclusivement fécondées par des Insectes, n'est-elle pas placée dans la dépendance de ceux-ci? Et si ces derniers font défaut au bon moment, par suite d'un hasard climatique (printemps froid et pluvieux de 1937), l'adaptation entomophile n'apparaîtra-t-elle pas comme des plus fâcheuses? Enfin il y a bien des caractéristiques statistiques négatives qui ne répondent à aucune utilité, par exemple la perte des yeux et du pigment chez la très grande majorité des cavernicoles; il y a convenance entre ces caractères et le milieu puisque ces animaux vivent dans une obscurité absolue; mais en quoi pigment et yeux pourraient-ils les gêner ? Toujours est-il que maintenant il leur est impossible de sortir du milieu très spécial qu'ils habitent.
Qu'advient-il lorsqu'une espèce présentant des adaptations statistiques en convenance avec un milieu A est transportée, par hasard ou de son gré, dans un autre milieu B, différent? Par exemple, les Pics sont des Oiseaux typiquement grimpeurs et arboricoles : leurs doigts arrangés en deux groupes, leur robuste bec capable de perforer le bois, leur longue langue, pointue et barbelée, bien faite pour piquer des Insectes lignicoles dans des fentes étroites, les pennes rigides de la queue fournissant un point d'appui solide lorsque l'animal grimpe sur un tronc, leur nidification dans un trou creusé en pleine matière ligneuse, tout cela convient bien à un Oiseau forestier. Mais un Pic de la Plata (Geocolaptes campestris)

2J2 SOCIÉTÉ DES SCIENCES DE NANCY
vit dans des plaines où ne pousse pas un arbre, où l'on ne voit tout au plus que de rares buissons; il se nourrit à terre de Termites, de Fourmis, et niche dans des terriers. De petits Moutons laponais, avec leur estomac et leur denture d'herbivores, mangent avec délices la chair en putréfaction des Baleines; le célèbre Kea de la Nouvelle Zelande {Nestor noia-bilis) qui est un Perroquet, jadis insectivore et granivore, a pris goût depuis 1868 à la chair fraîche des Moutons et se comporte comme un redoutable Rapace. La famille des Ecureuils, arboricoles typiques, a donné des formes tout à fait terrestres, les Xerns africains et les Tamias holarctiques, vivant dans des terriers creusés entre les racines des arbres ; les Kangourous, sauteurs, animaux de savanes, ont des genres qui vivent sur les rochers (Petrogale, grimpant comme un Singe sur des rochers à pic, d'où il saute), ou même sur les arbres (Dendrolagas aux ongles forts et recourbés). Les Rallides des prés humides comptent deux formes accommodées à une vie franchement aquatique, le Râle d'eau et la Poule d'eau.
D'une façon générale, ces emigrants gardent dans leur nouveau biotope les adaptations statistiques, plus ou moins atténuées, de l'ancien habitat, et s'en accommodent : les Ecureuils de terre ont toujours la longue queue touffue, le Kea le bec du Perroquet, le Pic de terre les pattes de grimpeur et le bec solide, les Râles d'eau les longues pattes et les doigts séparés. Parfois, cette persistance d'anciennes caractéristiques statistiques permet de reconstituer les étapes parcourues dans le temps ; on regarde l'Homme comme un Simien arboricole qui s'est spécialisé en animal marcheur dressé verticalement, parce qu'il a gardé de son ancien état la main prenante, qui est devenue son outil le plus précieux, et sa dentition d'omnivore. En même temps que la tendance aux nouveautés il y a souvent un conservatisme qui surprend: ne trouve-t-on pas, parmi les feuilles mortes des forêts, une petite Blatte (Ectobia) qui mène exactement la même vie que les grandes Blattes des forêts carbonifères? L'Orchidée Ôphrys apifera, qui s'autoféconde, a gardé une organisation nettement ento-mophile.

SOCIÉTÉ DES SCIENCES DE NANCY 2?3
Le fait de l'adaptation statistique entraîne une conséquence de haute importance: les animaux d'un même milieu ayant des mœurs analogues peuvent, en raison de leurs adaptations morphologiques communes, se ressembler d'une façon surprenante; pour reconnaître le groupe auquel ils appartiennent, il faut, laissant l'aspect de côté, s'adresser à des caractères plus permanents, tels que ceux de la dentition. Voici trois Mammifères non apparentés, tous trois fouisseurs, aveugles ou à petits yeux, à pattes antérieures munies de fortes griffes, à fourrure soyeuse et rase, à queue courte, qui ont ce qu'on appelle l'aspect talpoïde: c'est une Taupe qui est un Insectivore, un Myotalpa qui est un Rongeur, un Notoryctes d'Australie qui est un Marsupial. Cette ressemblance constitue le fait de la convergence.
Voici un exemple de convergence ichthyoïde : une nageoire dorsale triangulaire, sorte de quille supérieure, est apparue trois fois chez des animaux bons nageurs : le Requin (et bien d'autres Poissons), l'Ichthyosaure (Reptile jurassique), les Cétacés du groupe Dauphin, Orque, Marsouin. Cette adaptation morphologique, en rapport avec la vie marine, s'est réalisée d'une façon indépendante dans les trois formes, alors qu'elles étaient déjà aquatiques; elle fait défaut aux Plésiosaures, aux Phoques, aux Siréniens et aux Cétacés littoraux ou à grosse tête; elle paraît liée à la forme générale, qu'elle achève de rendre hydrodynamique, et par suite à la rapidité; on a l'impression que la dorsale fait partie d'un ensemble, comme si l'Ichthyosaure et le Dauphin avaient voulu se rapprocher le plus possible de la forme poisson, idéal nécessaire puisqu'ils sont l'un et l'autre chasseurs de proies agiles, Poissons et Céphalopodes; le galbe général du corps, la nageoire caudale, les pectorales et la nageoire dorsale ont dû aller de pair dans leur développement. Quant aux dorsales rudimentaires ou dégradées que l'on rencontre chez les Cétacés les plus évolués, on peut supposer qu'elles sont devenues telles parce qu'elles ont cessé de jouer un rôle utile, les animaux ayant changé de forme ou de comportement.
Il y a aussi une convergence ornithoïde chez les Vertébrés supérieurs capables de voler : Ptérosauriens, Oiseaux, Chau-

274 SOCIÉTÉ DES SCIENCES DE NANCY
ves-Souris, etc.. M. GUINIER VOUS a montré, le mois dernier, de magnifiques exemples de convergences végétales ou formes biologiques, qui vont presque jusqu'au mimétisme; je vous rappellerai en particulier les plantes « grasses » ou type eactoïde, qui sont des Cactacées, des Euphorbes, des Asclé-piadacées, des Composées, caractéristiques des climats chauds et secs.
La connaissance de la convergence a un intérêt pratique, au moins pour les étudiants ; il arrive que des examinateurs, voulant les habituer à regarder les choses de près et à ne pas se borner à l'impression de surface, leur donnent à reconnaître des espèces qui n'ont pas le faciès habituel du groupe dont elles font réellement partie; dans l'argot de laboratoire, ce sont des « colles », parfois redoutables. Il n'est pas commode de classer correctement un Typhlops qui est un Serpent fouisseur, un Orvet qui est un Lézard apode, une Cecilie qui est un Batracien apode; de séparer une Malacobdelle (Némerte) d'une Sangsue à trompe, un Armadillidhim (Crustacé Iso-pode) d'un Glomeris (Diplopode). Des Bryozoaires Ecto-proctes ressemblent à des Eponges, à des Hydraires, à des Madrépores, voire à des Algues ,et je vous présente un échantillon de ce groupe, une Flustre, que j 'ai retiré d'un herbier ancien. Les vieilles classifications, dites artificielles, étaient remplies d'erreurs dues à la convergence; l'ordre des Palmipèdes réunissait tous les Oiseaux à pattes palmées (sauf les Flamants), ce qui était fort commode, mais tout à fait artificiel, car il n'y a pas de parenté entre les Goélands d'une part, les Canards, Cormorans, Albatros d'autre part; dans les Rapaces, cohabitaient les Diurnes et les Hiboux; dans les Echassiers, on a rangé les Grues, les Outardes, les Hérons, les Flamants, les Autruches !
La convergence est l'ennemie particulière du biologiste qui recherche les affinités; si on arrive à peu près, sans trop de difficultés, à la dépister dans les ressemblances superficielles simples, nous sommes beaucoup moins sûrs de notre doctrine quand il s'agit d'organes internes, sur lesquels on se base justement pour établir les vrais rapports de parenté. L'Amphio-xus a des protonéphridies à solénocytes typiques, comme beaucoup de Vers et d'Annélides; faut-il donc le rapprocher de

SOCIÉTÉ DES SCIENCES DE NANC\ 275
ceux-ci, contre toute vraisemblance? Le Nematode et l'Arthropode ont en commun, seuls dans tout le règne animal, l'horreur du cil vibratile; doit-on pour cela en faire un groupe unique? Le Trilobite, qui converge avec les Phyllopodes Notostracés et les Xiphosures Hémiaspidés, est-il un ancêtre des Crustacés ou des Chélicérés? On est loin d'être fixé sur la signification du mimétisme: est-ce un phénomène qui a pour effet la protection des copieurs, comme on l'a pensé depuis Bates et Wallace, ou une convergence climatique ou mutative sans autre intérêt? Le démon convergence a encore bien des secrets à livrer.
* *
Le naturaliste se satisfait rarement des constatations de fait, comme celle de l'adaptation statistique ou de la caractéristique statistique; il cherche invinciblement une explication. Quelques-unes ont été proposées; il est tout indiqué, d'abord, de demander aux conditions extérieures l'agent même qui détermine l'acquisition organique; deux théories célèbres, le lamarckisme d'une part, le darwinisme et son fils le mutationnisme d'autre part, ont paru longtemps apporter des solutions acceptables.
Le lamarckisme peut être défini la causalité du désir ou du besoin, ou encore l'hérédité des effets de l'habitude : l'Oiseau que le besoin attire sur l'eau pour y trouver la proie qui le fait vivre, écarte les doigts de ses pieds lorsqu'il veut nager; la peau prend l'habitude de s'étendre, et ainsi se forme, par hérédité des effets de l'exercice, répété pendant de nombreuses générations, la palmure des animaux aquatiques. La gaine des griffes des Chats résulte de l'effort pour retirer en arrière les grandes griffes nécessaires pour déchirer leurs proies, mais gênantes dans la marche sur des sols pierreux. Pour Lamarck, « ce ne sont pas les oreanes d'un animal qui ont donné lieu à ses habitudes et à ses facultés particulières, mais ce sont au contraire ses habitudes, sa manière de vivre et les circonstances dans lesquelles se sont rencontrés les individus dont il provient, qui ont avec le temps constitué la forme de son corps, le nombre et l'état de ses

276 SOCIÉTÉ DES SCIENCES DE NANCY
organes ». Un changement de milieu ou d'habitudes doit donc toujours précéder les changements de la forme qui n'ont lieu que dans les générations suivantes.
L'explication lamarckienne n'est qu'un amas d'hypothèses ; elle suppose que l'organisme répond constamment au besoin, à l'effort, à l'action du milieu par une création ou une modification utile; c'est ainsi que la patte de l'Oiseau aquatique répondrait au besoin de mieux nager, à la friction de l'eau par la naissance d'une palmure. Comme l'a très justement dit BERGSON, c'est attribuer implicitement à la matière organisée une capacité sui generis, une mystérieuse puissance de monter des machines très compliquées, comme réaction aux excitations simples dont elle subit l'influence. L'exemple du Râle d'eau et de la Poule d'eau, nageurs sans palmure, montre bien que l'utilité ou le besoin d'un organe n'est pas une raison suffisante de son apparition. D'autre part, le lamarc-kiste raisonne comme si l'espèce était formée d'individus qui ne meurent jamais et qui, pendant des siècles, subissent l'action modelante (supposée!) du milieu; il fait abstraction de ce perpétuel recommencement et de la refonte de l'organisme par l'œuf, qui, lui, n'a pas d'organes, n'a pas fait d'effort ni éprouvé de besoin.
Le darwinisme peut être défini le hasard trié par la mort pour raison d'utilité : les divers individus d'une même espèce présentent de légères variations produites par l'action directe ou indirecte des conditions d'existence, de l'usage ou du défaut d'usage; étant de sens quelconque, elles peuvent être indifférentes, avantageuses ou désavantageuses. Il naît beaucoup plus d'individus qu'il n'en peut subsister à l'état adulte ; il y a donc nécessairement une mortalité considérable; or, il est logique que cette mortalité porte de préférence sur les individus qui présentent par hasard une variation désavantageuse et que survivent au contraire les individus à variations avantageuses; cette sélection naturelle du plus apte, conjuguée avec l'élimination du non apte, détermine un progrès automatique lent et continu dans les conformations utiles. Ainsi la palmure des Oiseaux aquatiques s'est développée par la survivance des meilleurs nageurs, exactement comme celle des Chiens-loutre d'Angleterre est due à la sélection par

SOCIETE DES SCIENCES DE NANCY 277
l'Homme des Chiens qui allaient le plus loin dans l'eau chercher le gibier blessé.
Au premier abord, le darwinisme, parfaitement logique, semble donner toute satisfaction; la sélection du plus apte a réponse à tout; elle intervient évidemment pour un organe utile; mais même s'il s'agit d'un organe indifférent, le darwiniste peut dire encore qu'il a une utilité qu'on n'a pas su découvrir ou qu'il est en corrélation avec un autre organe directement utile. On comprend l'enthousiasme des naturalistes, désormais libérés de l'obsession des causes finales; le darwinisme s'intégrait aisément dans la conception d'un Cosmos mécaniquement pur, sans but ni dessein, pâté de boue fait par deux enfants aveugles, la Matière et la Force, comme a dit HUXLEY; aussi son succès fut-il immédiat et complet. Presque tout le monde s'accorda pour reconnaître que le principe de sélection était la seule explication causale intelligible de toutes les formes d'adaptation.
Aujourd'hui le darwinisme ne nous paraît plus acceptable ; sa chaîne logique a été définitivement brisée lorsqu'on reconnut que la mort n'avait nullement cette fonction de triage automatique qui est la clé de voûte du système: il y a bien quelques tarés et anormaux qui sont éliminés dès le début, mais leur disparition a plutôt un effet conservateur du type moyen de l'espèce. Le reste est tué au hasard, comme les morts d'une bataille moderne ou d'un accident de chemin de fer; imaginons un pays où le nombre des Grenouilles vertes reste sensiblement le même d'une année à l'autre, ce qui est habituel ; qu'est-ce que cela signifie ? Etant donné qu'un couple de Grenouilles produit environ 5.000 œufs par an pendant η années, cela veut dire que sur 5.000 Χ η individus possibles, il survit seulement deux adultes, qui prendront la place du couple quand il disparaîtra. Il est évident que cette survie n'est pas due à quelque avantage physiologique ou anatomi-que, mais bien à la pure chance, d'autant plus que la destruction massive touche surtout les jeunes stades (têtards et jeunes métamorphosés).
Même si la sélection darwinienne existait, elle ne saurait expliquer la genèse graduelle d'une adaptation morphologique simple ou complexe; en effet, les étapes du début ne

278 SOCIETE DES SCIENCES DE NANCY
pourraient donner prise à la sélection, l'organe total fonctionnel n'étant pas encore formé; un organe électrique de Torpille en voie d'évolution qui ne donne pas encore de secousses n'est qu'une surcharge gênante, sans aucune utilité. DARWIN disait lui-même que le problème de la formation d'un organe complexe, tel que l'œil, quand il y songeait, lui donnait la fièvre.
Le mutationnisme, plus solide sur ses bases, a cependant des prétentions explicatives bien moindres que les théories qui l'ont précédé, puisqu'il n'accepte pas l'idée d'une liaison causale directe entre un caractère donné et les conditions du milieu auquel il paraît répondre, non plus que le triage automatique et utilitaire exercé au sein de l'espèce par la mort différentiatrice. Mais il rend bien compte de ce qui se passe lors d'un changement d'habitat ou du peuplement d'une place vide (voir plus haut les exemples du Geocolaptes% du Hibou de terrier, des Râles aquatiques) ; comme la Nature engendre éperduement,du pareil et du différent, elle offre à l'évolution d'innombrables combinaisons; parmi ces milliers d'essais, de temps à autre, il y en a un qui réussit ; un variant trouve une place vide ou mal défendue. Ce qui importe pour le succès, ce n'est pas tel ou tel détail morphologique, c'est la convenance physiologique globale d'une certaine constitution héréditaire (préadaptation = sélection écotypique de TURESSON) ; chaque être qui vit et qui dure a trouvé par hasard le milieu qui lui convient suffisamment; lorsque le milieu change, il s'accommode ou émigré si cela lui est possible, ou il meurt. D'où un renouvellement continu des flores et des faunes, des extinctions et des naissances d'espèces. Cette sélection globale des préadaptés a des conséquences d'apparence téléolo-gique; on pourrait croire que le nègre a été fait pour les régions tropicales, et le blanc pour les pays tempérés et nordiques, les plantes grasses pour les pays arides. Mais le point de départ, la variation mutative, est un fait de hasard; la trouvaille d'un milieu adéquat est un autre fait de hasard. Il paraît donc que le mutationnisme, hasard trié par la réussite globale, rend assez bien compte des adaptations statistiques, surtout dans le domaine végétal; on comprend qu'il y ait convergence de formes biologiques dans des stations à

SOCIÉTÉ DES SCIENCES DE NANCY 2 7 9
conditions tranchées: ainsi se développe une Laurisilva dans les climats humides et chauds, une Darisilva au pourtour de la Méditerranée, une Aestatisilva ou forêt feuillée en été dans les climats à hivers froids, une Hiemisilva ou forêt feuillée durant la saison des pluies dans les climats tropicaux. Et on comprend du même coup qu'il y ait des exceptions, des reliques d'époques antérieures qui se maintiennent par d'autres procédés que ceux de la masse, comme le Lierre et le Daphne laurèole dans Y Aestatisilva.
Je serais encore très disposé à croire que la sélection globale des préadaptés explique, au moins en partie, la ressemblance de couleur notée si fréquemment entre la teinte dominante de certains milieux et leurs habitants ; dans notre pays, il y a beaucoup de petits animaux verts dans le feuillage et les herbes; sur la terre nue, ils manquent presque absolument; dans les déserts par contre, il est bien connu qu'abondent les espèces de teinte grise ou jaunâtre comme le sol. Il a pu y avoir rassemblement fortuit d'espèces trouvant quelque avantage à vivre dans le milieu qui leur est horno-chrome.
Mais je ne crois pas qu'on puisse demander plus au muta-tionnisme; il ne saurait expliquer la genèse des adaptations morphologiques qui apparaissent après l'adaptation globale à un nouveau milieu, comme la nageoire dorsale des Ichthyo-saures et des Dauphins, la palmure des aquatiques, l'aspect talpoïde; et encore bien moins la formation de ces organes ressemblant à des outils, qui paraissent faits exprès pour une fin déterminée: le parachute du Sciuroptère, l'aile d'Insecte ou de Chauve-Souris, les instruments de musique, l'ombrelle plumeuse d'un fruit de Composée, la bascule staminale de la Sauge officinale, et mille autres. Le hasard heureux ne suffit pas.
Puisque le lamarckisme, le darwinisme et même le muta-tionnisme se montrent impuissants à donner une explication qui satisfasse totalement l'esprit, il y a encore une ressource, c'est de dire qu'il n'y a rien à expliquer, de nier ou de minimiser à l'extrême l'adaptation morphologique, ou tout au moins de la regarder, quand elle paraît exister, comme une coïncidence, un hasard qui joue la finalité. Cette attitude

28ο SOCIÉTÉ DES SCIENCES DE NANCY
vraiment hardie, j 'ai envie de dire paradoxale, est cede de la théorie « des causes actuelles » de DELAGE, reprise et développée par RABAUD; c'est une sorte de lamarckisme modernisé.
On sait que les caractères spécifiques sont renfermés en puissance dans l'œuf fécondé qui donne naissance à l'individu ; toute modification de ce dernier est la suite d'un changement dans une particule infiniment petite du patrimoine héréditaire. Ceci posé, imaginons que les conditions de vie d'une espèce se modifient; par exemple qu'un herbivore mange une plante arrivée nouvellement dans son district, que le climat devienne plus sec ou plus humide, ou qu'un carnivore soit contraint de chercher sa proie sur le rivage; si l'espèce persiste, il est probable que sa physiologie sera quelque peu changée; tous les tissus subiront le contre-coup de la modification du métabolisme; il se peut, alors, que le patrimoine héréditaire soit touché, et qu'en conséquence apparaissent des variations morphologiques de l'espèce. Ces changements de forme, évidemment, seront quelconques, sans rapport avec le bien-être ou l'utilité; généralement indifférents, il pourra se faire qu'ils aboutissent à des sortes d'organes, dont l'animal tirera parti tant bien que mal, et auxquels nous attribuerons à tort la signification d'adaptations morphologiques; ainsi pour la palmure; si celle-ci est tout de même plus fréquente chez les animaux vivant en milieux humides que chez ceux des endroits secs, on peut se demander si ce n'est pas une traduction quelconque, sans valeur fonctionnelle, des échanges métaboliques des êtres fréquentant les eaux ; ils réagissent de même façon, l'alimentation ayant une action morphogène eminente, et il en résulte un effet de convergence. Les exceptions marquent les différences de réaction en fonction des constitutions; elles prouvent nettement l'absence de toute valeur adaptative des formes.
A la base de cette audacieuse théorie, il y a une hypothèse : il n'est pas du tout certain, ni même probable, que de faibles modifications du métabolisme puissent influer sur les gênes du patrimoine héréditaire, abrités derrière la double cuirasse du cytoplasme et du noyau de la cellule sexuelle. Je trouve singulièrement prévenant ce hasard métabolique qui donne

SOCIÉTÉ DES SCIENCES DE NANCY 281
justement une palmure ou une queue aplatie à des animaux qui nagent (que cela leur soit très utile ou pas du tout), un parachute à quelques formes arboricoles, un aspect talpoïde à des animaux qui creusent, un aspect ichthyoïde avec nageoire dorsale à des Reptiles et Mammifères qui vivent dans la mer, etc., etc... DELAGE reconnaissait, du reste, qu'il serait fort embarrassé d'expliquer la formation d'un organe aussi compliqué et aussi adapté que l'oeil, le phénomène de la régénération et le fait qu'un développement est dès le début une préparation au futur.
* *
Résumons: l'adaptation morphologique et la convergence sont des faits, que la méthode statistique met parfaitement en évidence; il faut laisser de côté la question, à peu près dépourvue de sens, de savoir si l'adaptation morphologique augmente la facilité de vivre de l'espèce; elle est dans une mesure variable une spécialisation, c'est-à-dire un rétrécissement des possibilités; elle tend à confiner l'espèce dans un biotope déterminé où celle-ci peut se multiplier avec le minimum de concurrence.
En regardant les choses de plus haut, il apparaît que dans une région donnée, la spécialisation permet l'existence d'un maximum quantitatif de vie; toute association ou biocoenose d'animaux vivants ou fossiles, pour être en équilibre, doit comprendre des « fonctions » variées, herbivores, rongeurs, fouisseurs, carnivores, insectivores, nettoyeurs, arboricoles, terricoles, etc., comme le montre si clairement la célèbre faune marsupiale de l'Australie. Tout se passe comme si la Nature, conquérante d'espace et de matière, avait une fin particulière : maintenir la Vie sur la Terre, à travers les changements lents et continus des conditions cosmiques. La spécialisation, c'est-à-dire l'adaptation morphologique, concoure à cette fin : ainsi tout se peuple, la terre et les mers, l'eau douce, les arbres, les cavernes, l'air. Nous connaissons assez bien le mécanisme de l'adaptation nécessaire et suffisante, phénomène de hasard, rencontre heureuse du milieu et de l'espèce adéquate: nous n'en saurions dire autant du mécanisme de l'adaptation morphologique; une aile d'Oiseau, un parachute de Sciurontère,
debeaupu
Crayon

282 SOCIÉTÉ DES SCIENCES DE NANCY
une nageoire dorsale de Marsouin, semblent si manifestement répondre à un but et réaliser un dessein que nous hésitons à invoquer encore le hasard heureux des variations; mais on n'aperçoit clairement aucune cause naturelle qui puisse expliquer leur développement dirigé. Aussi terminerons-nous cette causerie en disant, comme Du Bois-REYMOND dans son célèbre discours de 1880 sur la finalité de la Nature: Ignoramus!

SOCIETE DES SCIENCES DE NANCV 283
LISTE DES MEMBRES COMPOSANT LA SOCIÉTÉ DES SCIENCES
(ARRÊTÉE LE 31 DÉCEMBRE 1937)
Membres titulaires
MM.
1936 ANZIANI (P.), docteur es Sciences, chef de travaux à l'Institut •Chimique, 9, rue de Villers, Laxou.
1924 ASSOCIATION DES ANCIENS ELÈVES DE LA FACULTÉ DE PHAR
MACIE DE NANCY.
1937 AUBRY, professeur d'agriculture à l'Ecole d'agriculture Ma-thieu-de-Dombasle, Tomblaine.
1936 BAUBILLIER (M l le), professeur à l'Ecole supérieure de jeunes filles, 13, rue de Serre, Nancy.
1936 BEAUDOUIN (D r), médecin-chef à l'Asile de Maréville, Pension Sainte-Anne, Laxou.
1936 BERTRAND, ingénieur commercial, 5, rue Migette, à Metz (Moselle).
1937 BESSON (M l le), pharmacien, 59, rue Maréchal-Oudinot, Nancy. 1930 BIZOT, pharmacien, 19, rue Condorcet, Dijon. 1937 BLACHE, professeur de Géographie à la Faculté des Lettres,
42, avenue Anatole-France, Nancy. 1936 BOQUET (Colonel), 0 # , 24, boulevard Jean-Jaurès, Nancy. 1937 BOUILLON, instituteur, 21, boulevard de la Pépinière, Nancy. 1937 BOUR (M1Ie Th.), licenciée es Sciences, 31 bis, rue de Lorraine,
Nancy. 1903 BRUNTZ (Dr), C # , recteur de l'Université de Nancy, 56, bou
levard Albert-Ier, Nancy. 1937 CALAFAT (P.), membre de la Société Linoéenne de Lyon, mem
bre de la Société d'Histoire naturelle de la Moselle, 29, rue de la Chèvre, Metz (Moselle).
1936 CAMO, professeur à l'Ecole normale de Nancy, 11, rue de Thionville, Nancy.
1936 CHAVAROT ( D T M.), pharmacien, licencié es Sciences, 98, avenue de Bouffiers, Nancy.
1936 CHEVALIER (P.), assistant à l ^ o l e de Brasserie, 6, rue Maurice-Barres, Nancy.

284 SOCIÉTÉ DES SCIENCES DE NANCY
1937 CHEVALLIER, professeur à la Faculté des Sciences, 86, rue Isabey, Nancy.
1936 CÉzARD, jardinier-chef du Jardin Botanique, 11 bis, rue Godron, Nancy.
1936 COLLIN (D r R.), * , professeur à la Faculté de Médecine, 33, boulevard Charles-V, Nancy.
1936 COLSON (D r P.), 16, rue Isabey, Nancy. 1930 COMBES, pharmacien, Sancey-le-Grand (Doubs). 1936 L E COMITÉ DES FORGES ET M I N E S DE FER DE MEURTHE-ET-
MOSELLE.
1937 COMPAGNIE D E SAINT-GOBAIN, CHAUNY ET CIREY, à Varan-
géville (M.-et-M.). 1912 COMPAGNIE LORRAINE D'ELECTRICITÉ, 62-64, rue du Faubourg-
Stanislas, Nancy. 1925 CONTAUT, ingénieur chimiste, 92, avenue Carnot, à Saint-Max
(M.-et-M.). 1937 CRESSOT, directeur de l'Ecole normale de Montigny-les-Metz
(Moselle). 1937 CROIX (M.-C) , principal honoraire du Collège de Commercy,
Avesnes (Nord). 1936 DELAFOSSE, professeur agrégé au Lycée, 44, place de Chambre,
Metz (Moselle). 1937 DESMONS (Colonel), 0 # , 18, rue Lyautey, à Nancy. l937 DEVILLE (M l le), licenciée es Sciences, 45, rue Saint-Gorges,
Nancy. 1936 DOMBRAY (D r P.), docteur es Sciences, chargé de cours à la
Faculté de Médecine, 1, rue de Serre, Nancy. 19^1 DONZELOT, professeur à la Faculté des Sciences, Nancv. 1890 DOREZ, pharmacien de ιΓΘ classe, 6yy rue de la Ravinelle, Nancy. 1936 DURUY, ingénieur en chef des Mines, place Carnot, Nancy.
1937 L ' E C O L E SUPÉRIEURE DE LA MÉTALLURGIE ET DES M I N E S , à
Nancy. 1923 EMERIQUE, 29, rue des Bégonias, Nancy.
1906 ENGEL, pharmacien de i r e classe, à Tucquegnieux (M.-et-M.).
1937 ERRARD, licencié es Lettres, instituteur à Villey-le-Sec (Meurthe-et-Moselle).
1936 FALLOT, ^ , professeur à la Faculté des Sciences, Institut de Géologie, rue de Strasbourg, Nancy.
1930 FLORENTIN (D r P.)., professeur abrégé à la Faculté de Médecine, 29, rue Henri-Déclin, Nancy.
1936 FLORENTIN (D r R.), * , Robert-Espagne (Meuse).

SOCIÉTÉ DES SCIENCES DE NANCY 2 8 5
1936 FONTAINE (Μ11Θ Th.), licenciée es Sciences, 83, rue Denfert-Rochereau, Paris (XIVe).
1934 FOURCHY, assistant à la Conservation du Musée Lorrain, 13, rue de Verdun, Nancy.
*937 FRANÇOIS (M l le), professeur à la Faculté de Pharmacie, Nancy. 1930 FRIBOURG (René), pharmacien, avenue Paul-Déroulède, Laxou. 1910 GAIN, # , professeur honoraire à la Faculté des Sciences de
Nancy, ancien directeur de l'Institut agricole et colonial, 9, rue de l'Oratoire, Nancy.
1922 GARDET, commis à l'Inspection Académique de Nancy, 3 bis, rue de La Salle, Nancy.
1920 GARNIER (Emile), θ # , sous-directeur de l'Ecole supérieure de la Métallurgie et de l'Industrie des Mines, avenue Carnot, à Saint-Max.
1929 GÉRARD (Colonel), 0 # , S, cours Leopold, Nancy. 1923 GÉRARDIN (André), 32, quai Claude-le-Lorrain, Nancy.
1901 GiRARDET, professeur agrégé libre à la Faculté de Pharmacie de Nancy, 6, rue de la Côte, Nancy.
1920 GODFRIN (Louis), # , pharmacien de i r e classe, 35, rue Saint-Dizier, Nancy.
1900 GOURY, chargé de cours d'archéologie préhistorique à la Faculté des Lettres, 5, rue des Tiercelins, Nancy.
1937 GROGNOT, Médecin-capitaine, 3, avenue de France, Nancy. 1936 GROUPE DES ETUDIANTS CATHOLIQUES, 35, cours Leopold,
Nancy. 1937 GUILLAUME (L.), chef de travaux à la Faculté des Sciences de
Strasbourg (Institut de Géologie). 1910 GUINIER, 0 # , directeur et professeur à l'Ecole nationale des
Eaux et Forêts, 16, rue Girardet, Nancy. 1936 GUYOT, inspecteur principal des Eaux et Forêts, Metz (Moselle).
1936 HAMANT, licencié es sciences, Institut de Botanique, 30 bis, rue Sainte-Catherine, Nancy.
1937 HATTON (M. l'abbé), 24, rue des Brice, Nancy.
1937 H E I M DE BALSAC (H.), docteur es Sciences, 34, rue Hamelin, Paris (16e).
1936 HENNEQUIN (D r L.), chef des travaux à la Faculté de Médecine, 30, rue Lionnoîs, Nancy.
1929 HERMANN (R.), ingénieur I.E.N., professeur à l'Institut Elec-trotechnique, 1 bis, rue Sadi-Carnot, à Malzéville (M.-et-M.).
1936 HERRY, ingénieur à la Société Générale Electrique, 5. rue Lafayette, Nancy.

286 SOCIÉTÉ DES SCIENCES DE NANCY
1937 HORNECKER, architecte diplômé du Gouvernement, 4, rue des Jardiniers, Nancy.
1922 HUSSON, professeur à la Faculté des Sciences de Nancy, 107 bis, rue Isabey, Nancy.
1936 HUSSON (R.), licencié es Sciences, 107 bis, rue Isabey, Nancy. 1892 IMBEAUX (Dr), 0 # , ingénieur en chef des Ponts et Chaussées,
professeur à l'Ecole nationale des Ponts et Chaussées, docteur en médecine, 18, rue Emile-Gallé, Nancy.
*937 JACOBI (M l le), professeur de Sciences naturelles au Collège Jules-Ferry, à Saint-Dié (Vosges).
1937 JÉRÉMiNE (Mme E.), assistante de Géologie à la Sorbonne, Laboratoire de Géologie de la Sorbonne, 2, rue V.-Cousin, Paris.
1936 JOLIBOIS, licencié es Sciences, 35, rue de Villers, Nancy. 1902 JOLY (H.), # , professeur à la Faculté des Sciences de Nancy,
11, boulevard Georges-Clemenceau, Nancy. 1937 JUPILLE (Mlle G.), licenciée es Sciences, 34, rue Sellier, Nancy. 1937 KAPLAN (Α.), licencié es Sciences, 55, rue Gambetta, Van-
dœuvre. Ι93Γ KARPINSKI, professeur de géologie, Teachar's College, Terres-
Hautes (Indiana). 1936 KISSEL (Dr P.), professeur agrégé à la Faculté de Médecine de
Nancy, 17, avenue Foch, Nancy. 1937 KLEIN, professeur d'agriculture à l'Ecole d'agriculture Mathieu- ·
de-Dombasle, Tomblaine. 1937 KOCHER, commandant à l'Etat-Major de la 20e Région. 1930 Mme LASSEUR-DUPAIX (Andrée), pharmacien, Faculté de Phar
macie, place Carnot, Nancy. 1937 LEBLANC (M.), herboriste, trésorier de la Société de Myco
logie, 16, rue Saint-Jean, Nancy. 1937 LEMAIRE (M.), professeur au Collège de Saint-Mihiel (Meuse). 1936 LEMASSON (P.), professeur à l'Ecole supérieure de garçons, 12,
place Aimé-Morot, Nancy. 1931 LENOIR, maître de Conférences de Botanique, Faculté des Scien
ces, Rennes. 1936 LEROY (le R.P.), Institut de Zoologie, 30, rue Sainte-Catherine,
Nancy. 1936 LÉVY-NEYMARCK ( D T ) , 2, rue des Chanoines, Nancy. 1926 LIENHART, # , chargé de Conférences à la Faculté des Sciences
de Nancy, 61, rue Isabey, Nancy. 1923 LONGCHAMBON, # , professeur à la Faculté des Sciences de
Nancy, 5, rue J.-J.-Rousseau, à Laxou (Meurthe-eet-Moselle).

SOCIÉTÉ DES SCIENCES DE NANCY 287
1937 MACQUART (Μ1 , Θ M.), pharmacien, licenciée es Sciences, 26, rue Héré, Nancy.
1899 MAIRE, # , professeur à la Faculté des Sciences d'Alger. 1936 MARTIN, directeur d'Ecole honoraire, 3, rue du Puits-de-Jacob,
Caen (Calvados). 1934 MAUDUIT, professeur à la Faculté des Sciences de Nancy, 16,
passage Sébastien-Bottin, Nancy. 1928 MERKLEN (Dr), professeur agrégé à la Faculté de Médecine,
1, rue de la Commanderie, Nancy. 1937 METRIS (M.-G.), professeur de Sciences naturelles au Collège
de Lunéville. 1930 MEUNIER, chargé de cours à la Faculté de Pharmacie, 11 bis,
rue du Lieutenant-Crépin, Nancy. 1936 MOREAUX (Dr R.), 20, rue Verlaine, Nancy. 1930 MOREL (Maurice), pharmacien, EpinaJ. 1937 MORELLE (Dr E.), # , docteur en pharmacie, Conseiller général
de la Meuse, Maire de Commercy, 1, place de l'Hôtel-de-Ville, Commercy (Meuse).
1936 MOISSON (R.), # , ingénieur E.C.A.M., 36 bis, rue Grandville, Nancy.
1931 MORISOT (Μ1 1 Θ) , pharmacien, 42, rue Emile-Gebhart, Nancy. 1937 NEUGNOT (M.), professeur de Sciences naturelles au Collège
de Neuhchâteau (Vosges). 1912 NICOLAS, # , greffier honoraire à la Cour d'Appel de Nancy,
avenue et château de Beauregard, Nancy. 1905 NOËL, ancien élève de l'Ecole normale supérieure, ingénieur,
106, rue du Faubourg-des-Trois-Maisons, Nancy. 1936 OUDIN, 0 # , conservateur, chargé de cours à l'Ecole des Eaux
et Forêts, 24, rue Saint-Michel, Nancy. 1937 PAILHOUX (M.-L.), ingénieur agronome, directeur de l'Ecole
d'agriculture Mathieu-de-Dombasle, à Tomblaine (M.-et-M.). 1937 PARISOT (Mm6), professeur de Sciences naturelles au Lycée de
Nancy, 12, place de la Croix-de-Bourgogne, Nancy. 1937 PARMENTELOT (Α.), licencié es Sciences, maître d'internat au
Lycée Henri-Poincaré, Nancy. 1923 PELTIER (Jean), ingénieur I.E.N., docteur es sciences de l'Uni
versité de Nancy, 8, rue de la Monnaie, Nancy. 1929 PELTIER (L.), professeur à l'Institut commercial et à l'Institut
colonial et agricole, 20, rue du Téméraire, Nancy. 1937 PERRIN (R.), professeur de Sciences au Collège de Commercy,
86, rue des Capucins, Commercy (Meuse). 1911 PÊTELOT, professeur au Lycée d'Hanoï, 4, rue Do-Hui-Vi.

288 SOCIÉTÉ DES SCIENCES DE NANCY
1936 PICARD (D.), interne des hôpitaux, 42, boulevard Albert-I r, Nancy.
1931 PIERRET, Institut de Physique, Faculté des Sciences, Nancy. 1936 EiTON (Dr L.), Jouet-sur-1'Aubois (Cher). 1930 PRÉVOST, pharmacien, place de l'Arsenal, Nancy. 1929 PRIVAT DE FORTUNIÉ (Dr), asile de Maréville, près de Nancy. 1936 RAILLARD, 24, rue de la Salle, Nancy.
1936 RAUX, directeur de l'Ecole de Brasserie, rue Grandville, Nancy. 1934 ROBAUX, assistant à l'Institut de Géologie, rue Monseigneur-
Turinaz, Nancy. 1922 ROBERT (Dr), # , professeur de Chimie à la Faculté de Méde
cine de Nancy, 30, rue Lionnois, Nancy. 1937 ROBLOT, instituteur à Xivray et Marvoisin (Meuse). 1936 ROESCH (Dr J.), 2, rue de la Botte, Belfort. 1937 RiOLS (M.), licencié es Sciences, professeur d'Horticulture,
Services agricoles, 26, rue Gustave-Simon, Nancy. !933 ROL, inspecteur des Eaux et Forêts, chef de section à la Station
de recherches de l'Ecole nationale des Eaux et Forêts, 33, rue Henri-Déglin, Nancy.
1925 SAINT-JUST PÉQUART, 3, avenue Paul-Déroulède, à Laxou (M.-et-M.).
1925 Mme SAINT-JUST PÉQUART, 3, avenue Paul-Déroulède, à Laxou (M.-et-M.).
1936 SANTENOISE (Dr D.), # , professeur de Physiologie à la Faculté de Médecine, 30, rue Lionnois, Nancy.
1936 SCHAEFFER, inspecteur des Eaux et Forêts, chef de section à la Station de Recherches des Eaux et Forêts, 21, avenue Foch, Nancy.
1930 SERRA (Jean), pharmacien, 22, boulevard Clemenceau, Oran 1936 SEXE (D1* J.), 2, rue Victor-Delavelle, Besançon (Doubs). 1921 SEYOT, * , doyen honoraire, professeur à la Faculté de Phar
macie de Nancy, 2, rue Montesquieu, Nancy. *937 SIMONIN, géomètre expert, à Saint-Nicolas. 1937 SIMONIN (Dr P.), professeur de Thérapeutique à la Faculté de
Médecine, 3, rue Saint-Lambert, Nancy. 1912 SOCIÉTÉ ANONYME DES MINES DE SEL GEMME ET SALINES DE
BOSSERVILLE, à Laneuveville-devant-Nancy. 1936 SOCIÉTÉ DES HAUTS-FOURNEAUX ET FONDERIES DE PONT-A-
MOUSSON (Meurthe-et-Moselle). 1927 SOCIÉTÉ DES SALINES DE DOMBASLE.
1912 SOCIÉTÉ SOLVAY et Cle, à Varangéville-Dombasle (M.-et-M.).

SOCIÉTÉ DES SCIENCES DE NANCY 289
1911 SPILLMANN (D r Louis), C # , doyen de la Faculté de Médecine de Nancy, 14, rue Saint-Léon, Nancy.
1936 STAUB (D r M.), # , médecin-commandant, 2, rue Saint-Sympho-rien, Metz.
1936 STEIMETZ, chef des travaux à la Faculté de Pharmacie, place Carnot, Nancy.
1933 TÉTRY (M l le Α.), assistante à l'Institut de Zoologie, avenue de Gentilly, Maxéville.
1935 THEOBALD, inspecteur d'Académie, Albi (Tarn). 1922 THIÉBAUT, &, chaigé de cours de géologie appliquée à l'Ins
titut de Géologie de Nancy, 17, rue des Goncourt, Nancy. 1937 TIIIÉBAULT (M.), ingénieur cartographe, 3, rue Sigisbert-
Adam, Nancy. 1908 T H I R I E T , # , docteur en pharmacie, droguiste, 26, rue des Ponts,
Nancy. 1937 · T H I R I O T , notaire honoraire, 21, r. Poincaré, Commercy (Meuse). 1936 TILLO Y (R.), licencié es Sciences, ingénieur géologue, Sainte-
Ménehould (Marne). 1934 TRAVERS, # , professeur à la Faculté des Sciences de Nancy,
directeur de l'Institut Chimique, 2, rue Messier, Nancy. 1937 U R I O N , professeur à la Faculté des Sciences, Institut Chimique,
Nancy. 1936 VÉRAIN (D r M.), 58 bis, rue de la Commanderie, Nancy. 1923 VERNIER (D r), chargé de cours à la Faculté de Pharmacie de
Nancy, 11, rue de Metz, Nancy. 1931 VIGNERON, pharmacien, 16, rue Léopold-Bourg, Epinal. 1936 VIGNERON (P.), architecte, 30, rue Carnot, Nancy. 1936 VIVIER, inspecteur adjoint des Eaux et Forêts, Station hydro
logique, Le Paraclet (Somme). 1937 W E I S (D r M.), ancien préparateur à la Faculté de Médecine
de Nancy, Montmédy. 1889 W Œ L F L I N , * , capitaine du génie démissionnaire, 2, rue
Hermite, Nancy.

2 9 0 SOCIÉTÉ DES SCIENCES DE NANCY
TABLE DES MATIÈRES POUR L'ANNÉE 1937
Comptes rendus des séances Pages
Séance du 15 janvier 1937 1 Séance du 12 février 1937 37 Séance du 12 mars 1937 69 Séance du 16 avril 1937 101 Séance du 14 mai 1937 125 Séance du 11 juin 1937 155 Séance du 9 juillet 1937 187 Séance du 12 novembre 1937 227 Séance du 10 décembre 1937 259
Conférences et Excursions
Conférence de Mn> CAILLIÈRE: Etude microscopique des minerais métalliques (10 février 1937) 70
Conférence de M. KAPLAN : Eclipse totale de soleil du 19 juin 1936 (14 février 1937) 74
Excursion du 11 avril 1937 : Forêt de Haye 102 Excursion du 25 avril 1937 : Glaciaire du Bassin Mosellan 111
Nécrologie
Louis MANGIN (1852-1937) et la Société des Sciences de Nancy, par E. NICOLAS 38
Notice nécrologique de M. Paul M E N T R E (1889-1936), par A. MAUDUIT 45
Présentations d'ouvrages
Extension et exploitation des forêts (par M. R O L ) 3 Carte des sols de Meurthe-et-Moselle (par M. O U D I N ) 25 Carte de la géographie zoologique de la France (par M. L.
CUÉNOT) 41
Les fossiles du Jurassique de la Belgique. Deuxième partie : Lias inférieur (par M. M. JOLY) 229

SOCIÉTÉ DES SCIENCES DE NANCY 29I
Communications Pages
BERTRAND (Th.). — Présence en Moselle du Chalcidien Poda* grion Pachymerum Walker 189
CÉzARD (N.). — Stations nouvelles de Vallisneria spiralis L. et Lepidinm latifolinm L 10
— Notes pour la flore de Lorraine Lycopersicnm esculentum Mill, sur les bords de la Meurthe 49
COLLIN (R.). — Le facteur svmpathique dans le mécanisme de la pigmentation cutanée 149
CONTAUT (H.). — Sur le régime de la Haute Meuse au Flandrien. 5 CUÉNOT (L.). — L'adaptation chez les animaux 268 FLORENTIN (P.). — Les réflexes photo-pigmentaires chez les
Batraciens et la notion de physiologie spécifique 261 GUINIER ,Ph.). — Science et Sylviculture. A propos des commu
nications de MM. O U D I N et R O L 30 — A propos d'un chêne fossile des Alluvions de la Meurthe.
Observations botaniques et forestières 90 — Les Broméliacées epiphytes (présentation d'échantillons). . . : 93 — L'adaptation chez les végétaux. (Causerie avec présentation
d'échantillons) 239 H E I M DE BALSAC (H.). — La Testacelle en Lorraine. Quelques
points de son écologie 146 HUSSON (R.). — Reproduction non saisonnière des Polydes-
midées (Dipl.) des galeries de mines 195 JOLY (M.). — Sur un chêne semi-fossile des Alluvions de la
Meurthe 84 — Le Kiméridgien de la région de Pierrefitte-sur-Aire et en
général, de la feuille de Bar-le-Duc 231 KAPLAN (Α.). — Organisation de la mission de la Société Astro
nomique de France et les observations faites par elle au cours de l'éclipsé totale de soleil du 19 juin 1936 à Beloretchens-kaya (U.R.S.S·) 127
MOREAUX (René).. — Le rôle de l'appareil de coaptation alaire che2 l'Abeille 60
— Contribution à l'étude d'une nouvelle maladie des Abeilles.. 179 ROBAUX (Α.). — Deux sondages de recherches d'eau en Meurthe-
et-Moselle (Rozières-en-Haye et Mor fontaine) 190 TÉTRY (Μ11β Α.). — Sur la synonymie d'un ver de terre commun
en Lorraine (Allolobophora terrestris Saviçny) 51 — Sur le Podagrion pachymerum parasite des Oothèques de
Mantes (deuxième note) Q8
debeaupu
Crayon

292 SOCIETE DES SCIENCES DE NANCY
— Description d'une nouvelle espèce de Lombricien (Allolobo-phora cupulifera) 119
THEOBALD (U.). — Découverte d'une défense de Mammouth (Elephas primigenius Blum.) dans les alluvions de la plaine rhénane à Walff (Bas-Rhin) 142
— Note complémentaire sur les insectes fossiles oligocènes des gypses d'Aix-en-Provence 157
TiLLOY (R.). — Contribution à l'étude de la Faune quaternaire des environs de Sainte-Ménehould (Marne) 12
— La fonction athrocytaire chez les Hirudinées (Essais d'explication de la sélection des colorants) 199
URION (E.). — Mécanisme de déshydratation du divinylglycol.. 66
debeaupu
Crayon



















