
Etudes embryologiques dans le genre Cynorkis...
14
ADANSONIA, ser. 2, 12 (3) : 384-402. 1972. ÉTUDES EMBRYOLOGIQUES DANS LE GENRE CYNQRKIS (ORCHIDACEAE) par Yvonne VEYRET RÉSUMB : Les C. ridleyi Dur. et Schinz., C. anzpullacea H. Perr. et l’hybride présumé C. lilacina X C. ridleyi H. Perr. sont apomictiques autonomes absolus. L’archéspore est la CMM; elle avorte ou dépasse rarement le stade dyade suivant les espèces. Ses produits ne sont pas fonctionnels. L’embryonie est soit nucellaire, soit omnisaccale diploïde; ce dernier mode est nouveau. SUMMARY : C. ridleyì Dur. et Schinz., C. anipidlacea H. Perr. and the presumed hybrid C. Zilaciria x C. ridleyi H. Perr. are obligate autonomous apomicts. Archespore is the MMC; generaly is aborts or give rise to dyad cells according to the species. Its products are not functionnal. Embryony is nucellar or omnisaccal diploid; this last modality is new. i Le genre Cynorkis groupe actuellement environ 140 espèces; l’une d’elles est asiatique, un petit nombre se rencontrent en Afrique tropicale et australe, les autres sont spéciales 8. la région malgache : Madagascar, Comores, Mascareignes, Seychelles ; mais la majorité sont endémiques de la Grande Ile. I Ces espèces sont réparties dans 6 sections suivant le nombre de viscidies et diverses caractéristiques du rostellum. Les espèces présentement étudiées appartiennent toutes à la première section du genre, qui rassemble plus de la moitié des Cynorkis, mais elles y sont classées dans des groupes différents : le C. ridleyi Dur. et Schinz. dans le premier, le C. anipzillacea H. Pen. dans le troisième, l’hybride présumé C. lilacilia s C. ridleyi a le premier de ses parents dans le deuxième groupe. Chez toutes ces espèces le mode de reproduction asexué était évident avant d‘en avoir la preuve microscopique; en effet le pollen peut être avorté ou les pollinies rester en place et le stigmate ne porter aucune trace de pollen; malgré cela elles développent des ovules pourvus d‘embryons. Dans tous les cas l’apomixie est autonome et absolue mais les modalités de la reproduction apomictique présentent des différences suivant ces espèces et en conséquence nous examinerons celles-ci successivement.
Transcript of Etudes embryologiques dans le genre Cynorkis...

ADANSONIA, ser. 2, 12 (3) : 384-402. 1972.
ÉTUDES EMBRYOLOGIQUES DANS LE GENRE CYNQRKIS (ORCHIDACEAE)
par Yvonne VEYRET
RÉSUMB : Les C. ridleyi Dur. et Schinz., C. anzpullacea H. Perr. et l’hybride présumé C. lilacina X C. ridleyi H. Perr. sont apomictiques autonomes absolus. L’archéspore est la CMM; elle avorte ou dépasse rarement le stade dyade suivant les espèces. Ses produits ne sont pas fonctionnels. L’embryonie est soit nucellaire, soit omnisaccale diploïde; ce dernier mode est nouveau.
SUMMARY : C. ridleyì Dur. et Schinz., C. anipidlacea H. Perr. and the presumed hybrid C. Zilaciria x C. ridleyi H. Perr. are obligate autonomous apomicts. Archespore is the MMC; generaly is aborts or give rise to dyad cells according to the species. Its products are not functionnal. Embryony is nucellar or omnisaccal diploid; this last modality is new.
i
Le genre Cynorkis groupe actuellement environ 140 espèces; l’une d’elles est asiatique, un petit nombre se rencontrent en Afrique tropicale et australe, les autres sont spéciales 8. la région malgache : Madagascar, Comores, Mascareignes, Seychelles ; mais la majorité sont endémiques de la Grande Ile. I
Ces espèces sont réparties dans 6 sections suivant le nombre de viscidies et diverses caractéristiques du rostellum. Les espèces présentement étudiées appartiennent toutes à la première section du genre, qui rassemble plus de la moitié des Cynorkis, mais elles y sont classées dans des groupes différents : le C. ridleyi Dur. et Schinz. dans le premier, le C. anipzillacea H. Pen. dans le troisième, l’hybride présumé C. lilacilia s C. ridleyi a le premier de ses parents dans le deuxième groupe.
Chez toutes ces espèces le mode de reproduction asexué était évident avant d‘en avoir la preuve microscopique; en effet le pollen peut être avorté ou les pollinies rester en place et le stigmate ne porter aucune trace de pollen; malgré cela elles développent des ovules pourvus d‘embryons. Dans tous les cas l’apomixie est autonome et absolue mais les modalités de la reproduction apomictique présentent des différences suivant ces espèces et en conséquence nous examinerons celles-ci successivement.

- 390 -
C. ridleyi Dur. et Schinz
Le C. ridZeyi est décrit par H. PERRIER DE LA BATHIE comme étant G très variable quant à la forme du lobe médian du labelle qui peut être orbiculaire, largement flabelliforme, obréniforme, largement ovale ou même étroitement oblong, et denté-crénelé ou échancré-apiculé au bord antérieur D.
Le mode de développement apomictique de l’embryon dans les 4 formes que nous avons observées, est particulièrement évident lorsque le pollen est avorté, ce qui est le cas dans 2 de celles-ci. Parmi ces 4 formes, 3 poussent côte à côte en sous-bois d’Eucalyptus dans la réserve de l’Angavokely, près de Tananarive; nous les numéroterons de 1 à 3; la 4e provient de la forêt primaire de la Montagne d’Ambre, ali nord de Yîle. Pour tolites la floraison a lieu en septembre.
Le no 1 a une feuille petite, courte, résillée de vert plus clair, à limbe horizontal. Le lobe médian du labelle est cordiforme; l’appareil pollinaire est apparemment normal.
Le no 2 présente 1 à 2 petites feuilles d‘un vert foncé en-dessus, pourpre en-dessous. Le lobe médian du labelle est plus large que long, tronqué-crénelé à son extrémité; les pollinies sont entières.
Le no 3 ne possède qu’une feuille, finement maculée de blanc, princi- palement de part et d’autre de la nervure médiane sur la face supérieure de la feuille, dressée. Le lobe médian du labelle est de forme générale triangulaire, à extrémité obtuse, 2,5 fois plus long que large en son milieu. Le pollen est avorté.
Le no 4. est une plante beaucoup plus grande que les précédentes, à 2 feuilles dressées, régulièrement et hement tachetées de blanc. Le lobe médian du labelle est arrondi-crénelé, pourvu lui-même d’un court lobe médian tronqué-crénelé. Le pollen est avorté.
La planche 1 représente la fleur et le labelle de ces différentes formes. Nos examens microscopiques ont plus spécialement porté sur le no 3
et la majorité des microphotographies se rapportent à cette forme (Pl. 2 et 3, fig. 1 à 5).
La cellule archésporiale atteint une certaine taille, mais elle ne se divise pas et présente très rapidement des signes de dégénérescence. Pendant ce temps 1 OLI 2 cellules de l’épiderme nucellaire s’accroissent considérable- ment de taille et l’on pourrait penser que ce sont les initiales d’embryons adventifs nucellaires comme on l’a déjà constaté chez diverses plantes depuis fort longtemps, parmi elles les Orchidacede : Nigritella nigra Reichb. f. (K. A~z~kIus) , Zeuxine szflcata Lindl. (K. N. SESHAGIRIAH, B. G. L. SWAMY) et Cynorkis lilacina Ridley (Y. VEYRET) ; mais les embryons développés aux dépens de ces initiales ne sont pas orientés comme des embryons nucellaires qui se fixent par leur suspenseur sur l’organe dont ils sont issus. ALI contraire c’est leur sommet qui se différencie sur le bord de cet organe; et la polarité de ces embryons est ainsi la même que celle des embryons sexués. En conséquence, nous pensons que les initiales de ces embryons doivent en réalité être considérées comme les initiales de sacs embryonnaires
jc

- 391 -
3 a
P1. 1. - C. ridleyi Dur. et Schinz : 1,2,3,4, se rapportent aux formes décrites sous les mêmes numéros (fleur 1; 2,4; labelle x 3).
somatiques virtuels. Le gamétophyte demeure unicellulaire et participe en totalité, à I’édification de l’embryon. L’évolution de cellules somatiques - généralement de la clialaze, parfois du nucelle - en initiales de sacs embryonnaires se produit fréquemment dans les cas d‘apogamogonie présentés par les Graminées, les Composées, les Rosacées. Un tel phéno- mène est très souvent précédé de la dégénérescence de l’archéspore ou de ses produits (dyade ou tétrade), et l’initiale somatique du sac embryon- naire engendre un gamétophyte à 4 ou 8 noyaux suivant les cas. On dit qu’il y a aposporie somatique. Chez le C. ridleyì on retrouve la même initiale somatique, mais elle évolue directement en embryon. Nous pro- posons pour désigner ce phénomène le terme d’embryonie omnisaccale diploïde.
Les microphotographies que nous avons sélectionnées représentent la formation de l’embryon aux dépens d’une cellule de l’épiderme nucel- laire, par l’intermédiaire d’un gamétophyte virtuel. A cet effet, après l’im- portant accroissement de taille déjà signalé, la première segmentation est transversale [fig. 4). La différenciation de l’embryon proprement dit semble
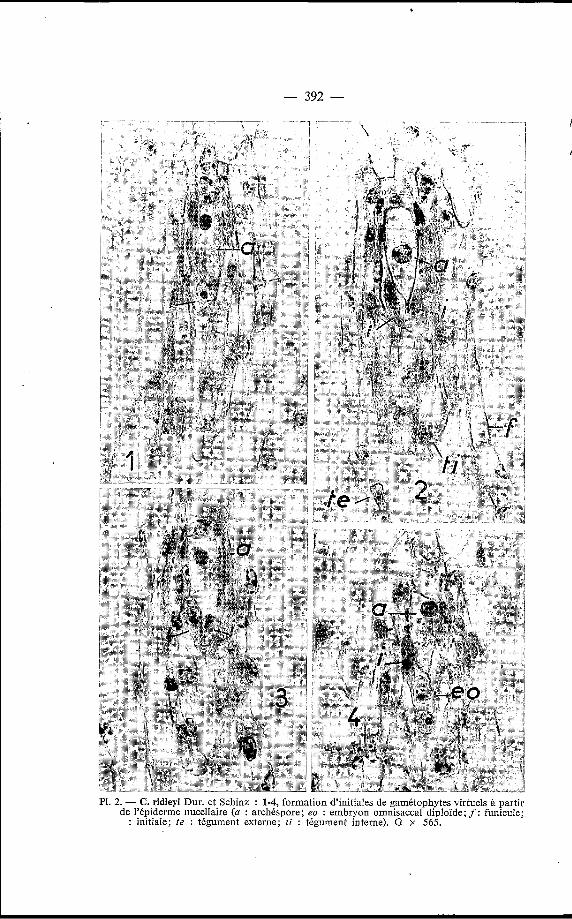
PI. 2. - C. ridleyi Dur. et Schinz : 1-4, formation d’initiales de gamétophytes virtuels à partir de l’épiderme nucellaire (a : archéspore; eo : embryon omnisaccal diploïde; f: funicule;.
: initiale; fe : tégument externe; f i : tégument interne). G x 565.

- 393 -
C. ampullacea H. Perr.
La forme du C. anzpiillacea examinée provient d‘un lambeau forestier du massif de 1’Itremo I. Elle diffère du type décrit par H. PERRIER DE LA BATHIE par une feuille relativement plus large et des fleurs environ 2 fois plus grandes, morphologiquement un peu différentes. C‘est ainsi que le labelle n’est pas courtement trisinué ou trilobulé, mais à lobes bien nets, le lobe central étant plus grand que les lobes latéraux; la présence de glandes pédiculées s’étend, de plus, sur la partie inférieure de la face supérieure du labelle, la base de l’épero11 et la face inférieure des sépales. Cette espèce varie dans sa morphologie dans les diverses stations de l’île où nous l’avons
.qui lui est propre. La planche 4 représente la fleur et les pièces florales de la forme étudiée.
l’anthèse, c’est-à-dire que l’archéspore est individualisée et que le tégument interne commence à se former. L’ovaire continu de grossir sans qu’il y ait pollinisation; si même on supprime les pollinies les ovules n’en pour- suivent pas moins leur développement. Dans le jeune ovule le nucelle est représenté par une rangée axiale comportant en général 4 cellules enve- loppées par l’épiderme nucellaire. La dernière cellule de cette rangée sera l’archéspore qui fonctionne directement coinme cellule-mère des macro- spores. I1 semblerait qu’au cours de la méiose de la CMM l’asyndèse soit totale et que la répartition des chromosomes se fasse un peu au hasard, d‘une manière assez semblable à ce qui se produit dans certains ovules d’ilntennaria, d‘après les observations de G. L. STEBBINS, et où tous, ou la majeure partie des gamétophytes issus des macrospores réduites, périssent. Chez le C. ainpullacea les chromosomes retardataires sont fréquents et ils peuvent parfois simuler un pont (Pl. 3, fig. 7-8, P1. 5, fig. 9 et 10); le premier cloisonnement de l’archkspore se produit souvent avant que les chromosomes aient achevé leur migration vers les pôles (Pl. 3, fig. 7-8).
1. Nous remercions nos collègues F. FRIEDMAN qui a découvert cette forme du C. ainpii//aceu au col d’Itremo et en a récolté des pieds B notre intention, et J. BOSSER qui a bien voulu se charger de sa détermination.
1 rencontrée; chaque variété y fleurit également à une époque de l’année
L’ovple de ce C. avlzpirllacea est légèrement ébauché au moment de 3
~\ .se faire après le stade tétracellulaire par un premier cloisonnement longi- tudinal de la cellule apicale voisine de I’archéspore (fig. 6). Bien que très fréquemment 2 cellules du nucelle semblent évoluer en initiales de gainéto- phytes on ne trouve généralement qu’un seul embryon dans la graine et même dans l’ovule; ce qui prouverait qu’une seule des initiales soit normale- ment fonctionnelle.
Ce phénomène est semblable dans les 4 foimes examinées; cependant j l peut arriver dans la forine no 4 que des embryons nucellaires se déve- loppent également, soit concurremment avec l’embryon omiiisaccal, dans u n même ovule, soit isolément.
1
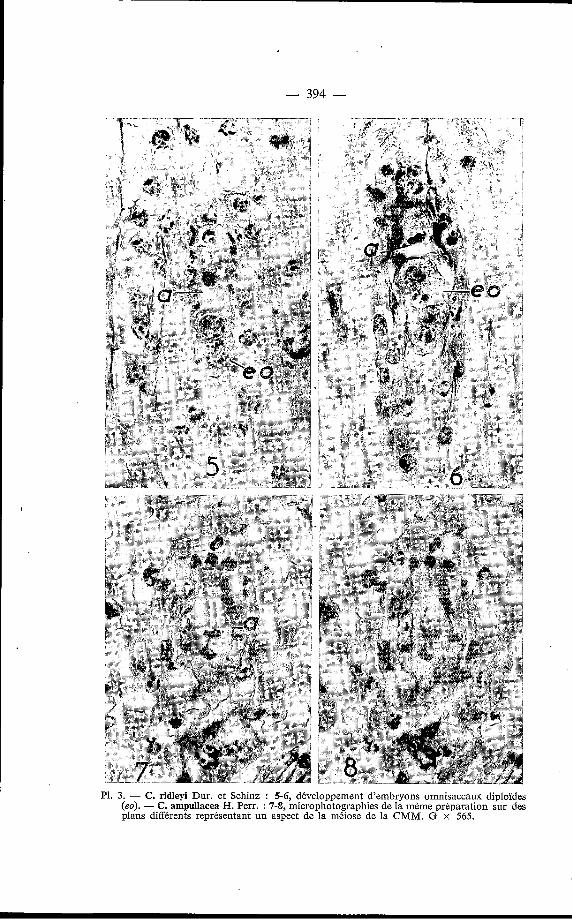
- 394 -
P1. 3. - C. ridleyi Dur. et Schinz : 5-6, développement d'embryons omnisaccaux diploïdes (eo). - C. ampullacea H. Perr. : 7-8, microphotographies de la même préparation sur des plans différents représentant un aspect de la méiose de la CMM. G x 565.

- 395 -
6
PI. 4. - C. ampdlacea H. Perr. : 1, fleur x 2; 2, sépale latéral; 3, sépale médian; 4, pétale latéral; 5, labelle X 2,s; 6, pollinaire; 7, gynostème x 5.
En fin de première division des chromosomes se trouvent isolés en dehors des noyaux télophasiques (fig. 7 à 10). Nous avons pu compter jusqu’à 60 chromosomes au cours de la première division de l’archéspore, mais nous préférons écrire 212 = ca60 au cas oÙ quelques-uns de ces chromo- somes petits et nombreux auraient échappé à notre observation. L’espèce serait donc tétraploïde par rapport à quelques espèces sexuées oÙ 2n = ca30.
Le plus souvent une cloison sépare les noyaux engendrés par l’arché- spore; ce sont alors de véritables dyades; parfois ces 2 noyaux se disposent chacun à une extrémité de la cellule simulant un sac embryonnaire binucléé (F‘l. 6, fig. 13). Par la suite, ces noyaux ne se divisent plus et dégénèrent assez rapidement (Pl. 6, fig. 14 à 16), sauf si l’ovule n’est pas embryonné. Dans ce cas, les produits de l’archéspore peuvent s’accroître d’une manière relativement importante et former une polyade ayant jusqu’à 6 noyaux, en partie cloisonnée (Pl. 5, fig. 12).
Tous les embryons développés dans les ovules de ce Cynorkis le sont à partir d‘une cellule nucellaire épidermique qui peut évoluer de deux manières différentes, soit qu’elle se transforme en initiale d’un proembryon adventif nucellaire, soit qu’elle devienne l’initiale d’un gamétophyte virtuel comme nous l’avons déjà décrit plus haut chez le C. ridleyi, initiale qui se transforme directement en embryon omnisaccal diploïde. Par la suite les embryons omnisaccaux seront munis d‘un suspeqseur filamenteux comme dans les embryons sexués des Ophrydoideae (Pl. 6, fig. 13 à 15), tandis que les embryons nucellaires auront un aspect plus globuleux avec un court suspenseur passant insensiblement à l’embryon proprement dit (Pl. 6, fig. 16). 11 n’est pas toujours aisé de prévoir le devenir d‘une initiale, sauf
i
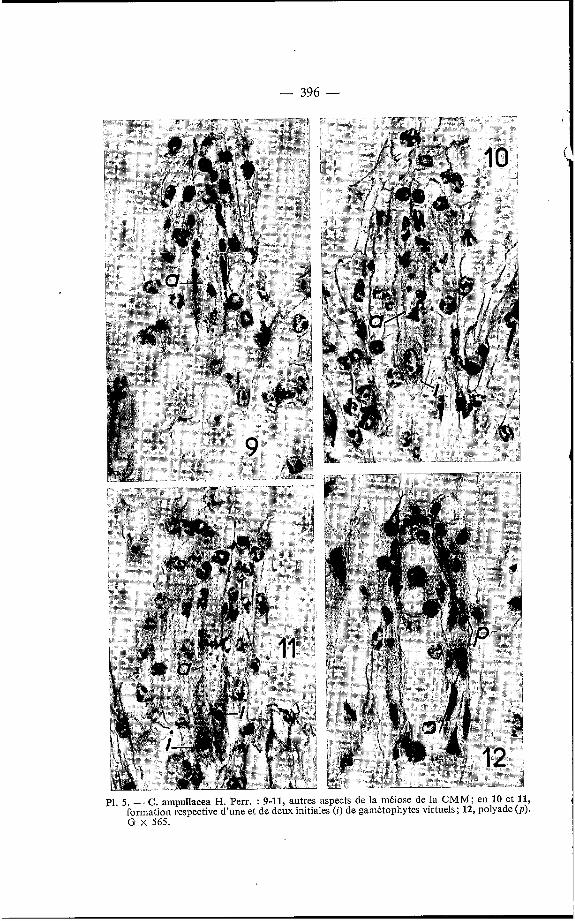
- 396 -
P1. 5 . - C. ampullacea H. Perr. : 9-11, autres aspects de la méiose de la CMM; en 10 et 11, formation respective d’une et de deux initiales (i) de gamétophytes virtuels; 12, polyade (p ) . G x 565.
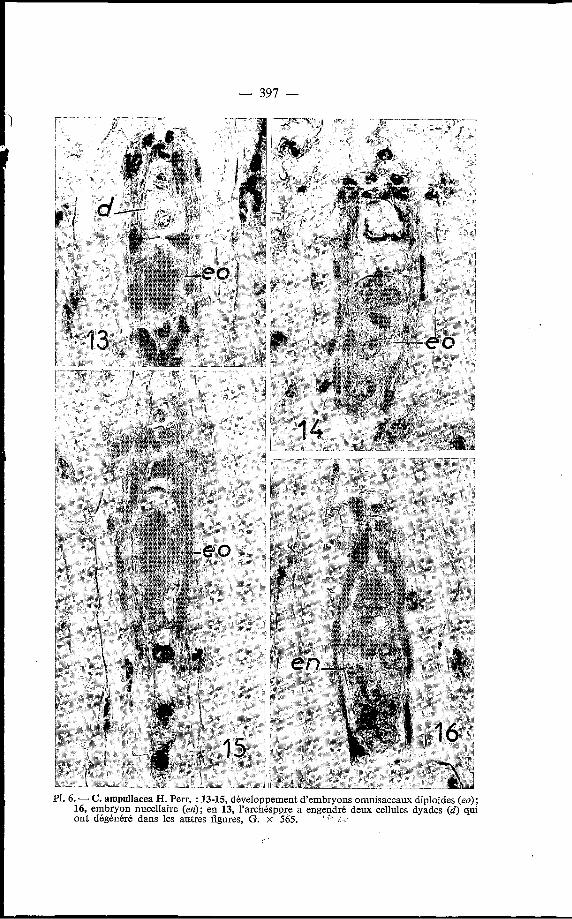
- 397 -
PI. 6. - C. ampullacea H. Perr. : 13-15, développement d'embryons omnisaccaux diploïdes (eo); 16, embryon nucellaire (en); en 13, I'archéspore a engendré deux cellules dyades (d) qui ont dégénéré dans les autres figures, G. x 565. . I

-- 398 -
lorsque sa forme préfigure l'orientation de l'embryon qui en tirera- [son origine soit directement, soit indirectement. Dans la figure 10 de la Planche 5, l'initiale est sans aucun doute celle d'un gamétophyte virtuel comme le laisse penser sa forme arrondie au sommet et en pointe à son extrémité. Par contre, dans la figure 11 on ne peut encore savoir quel sera le sort des initiales.
Du côté des microspores on peut noter la présence assez fréquente de tétrades aberrantes à plus de 5 noyaux, généralement de 5 à 6 , plus rarement 7, de taille irrégulière, certains à I'état de micronoyaux. Les pollinies semblent d'ailleurs peu fonctionnelles ; d'autre part, les sécrétions des stigmates sont faibles et lorsqu'on tente la pollinisation peu de pollen y adhère, dont la germination est particulièrement rare et peu vigoureuse.
Hybride présumé C. Macina x C. ridleyi H. Perr.
Cette plante est l'ancien Microtheca madagascarica R. Schlechter. H. PERRIER DE LA BATHIE lui a donné ce nouveau statut car il pensait que cet état hybride et cette parenté devaient être attribués à l'ancien Microthecn parce que ses fleurs sont (< toujours stériles, à anthères avortées D d'une part, et qu'il cohabite avec ses 2 parents présumés en quelques stations d'autre part. Cette plante est, en effet, remarquable par l'avortement de son appareil pollinaire; cependant, il peut arriver que quelques fleurs présentent une pollinie plus ou moins bien développée. Un si faible pour- centage de pollen serait insuffisant pour assurer une fertilisation régulière et pourtant le Microtheca fructifie régulièrement et abondamment.
Les plantes qui ont fait l'objet de nos observations embryologiques proviennent d'un sous-bois clair d'Eucalyptus situé en bordure du chemin qui relie la RN 2 à Ambatomena, à 30 km environ de Tananarive. Parmi les fleurs d'une dizaine de pieds examinés aucune ne présentait de pollinies et les examens microscopiques ont montré, ou mieux confirmé notre opinion que cette espèce était apomictique. Elle l'est, de plus, et forcément, d'une manière autonome et absolue.
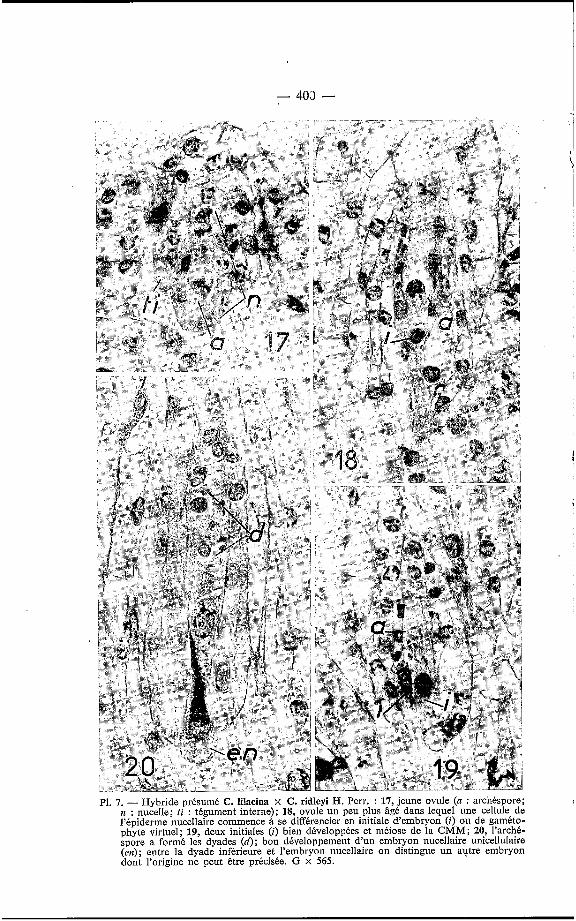
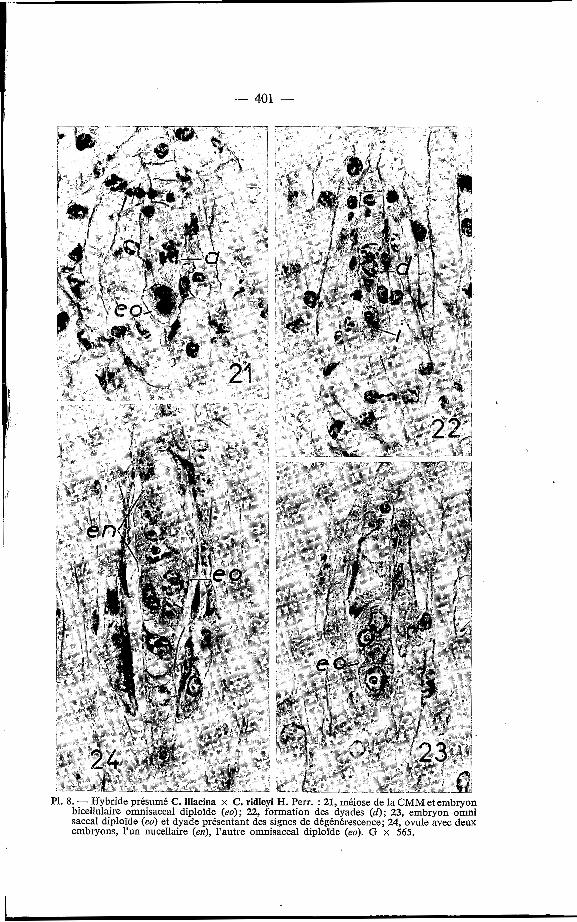
L'apomixie se présente soit sous forme d'embryonie adventive nncel- laire, soit sous forme d'embryonie omnisaccale diploïde, celle-ci paraissant plus fréquente que la première. Les embryons ainsi formés suppléent au manque de fonctionnement des spores. En effet, l'archéspore, provenant directement d'une cellule nucellaire sous-épidermique (PI. 7, fig. 17), n'engendre jamais plus de 2 cellules dyades (Pl. 7, fig. 20) dont aucune ne sera à l'origine d'un gamétophyte femelle, et qui dégénéreront avant la lin de l'embryogenèse. Ici aussi l'avenir des initiales est parfois difficile à prévoir, par exemple pour celles des figures 18 et 22 des planches res- pectives 7 et 8. Par contre, dans la figure 20 (PI. 7), l'embryon unicellulaire est indiscutablement nucellaire et dans la figure 19 (PI. 7) les initiales parais- sent être celles de gamétophytes virtuels avec leur extrémité distale en pointe. Dans un même ovule les deux sortes d'initiales peuvent se former puisque l'on peut y rencontrer les deux formes d'embryons; en effet dans la figure 26 (Pl. 8), on peut voir un embryon adventif nucellaire et un embryon omni- sacca1 diploïde opposés par leur sommet.
. .

- 399 - 1 Quant à l’origine de cette plante, si elle est de nature hybride, elle
ne peut l’être entre les espèces présumées par H. PERRIER DE LA BATHIE, puisque toutes les formes des C. ridleyi et C. lilacina que nous avons eu l’occasion d‘examiner se sont révélées être apomictiques autonomes ab- salues et que l’apomixie paraît donc caractéristique de ces espèces. Comme tous les autres Cyrtorkis apomictiques elle nous paraît devoir mériter un nom d’espèce qui lui soit propre; son ancien nom devrait lui être res titué ou plus probablement une combinaison faisant état de ses autres dénominations lui être attribuée.
Chez tous ces Cyrzorkis apomictiques : C. ridleyi, C. ainpullacea, l’hybride présumé C. lilacina >i C. ridleyi, la sexualité est très dégradée puisque nous avons constaté des anomalies polliniques ou l’avortement du pollen, et une gamétogenèse femelle anormale par la méiose de l’arché- spore, tout au moins chez le C. ainpullacea, et incomplète puisque le sac embryonnaire n’est jamais formé, la CMM ne dépassant généralement pas le stade dyade.
Chez le C. lilucirza nous avions vu, il y a quelques années, que la sexua- lité était légèrement moins dégradée puisqu’il peut se former 4 spores femelles ou un sac embryonnaire bien que celui-ci atteigne rarement le stade trinucléé. Cependant, nous ignorons la nature de la première division de l’archéspore. Chez tous les Cynorkis apomictiques, en plus des embryons adventifs nucellaires, on trouve soit des embryons sporiques, c’est-à-dire formés aux dépens d’éléments issus de l’archéspore, chez le C. Macina, soit omnisaccaux diploïdes chez les autres connus à ce jour. La présence des embryons sporiques pourrait, peut-être, être en rapport avec cet état moins dégradé de la sexualité du C. lilacina. .
Néanmoins il y a de fortes présomptions pour que le C. Macina comme les autres Cyrzo~kis apomictiques soit de nature hybride comme peuvent le suggérer les anomalies dans les gamétogenèses ou l’état du pollen et le devenir de l‘archéspore.
Si donc nous supposons une origine hybride pour ces Cynorkis la variation morphologique interspécifique qu’ils présentent - il ne s’agit pas de microformes écologiques - peut s’expliquer par ce phénomène d‘hybridation où plusieurs espèces ont pu intervenir et se recroiser. I1 n’est pas improbable aussi que les espèces aient varié avant de se croiser comme pourrait le laisser penser la présence de diverses formes chez des espèces sexuées comme les C. fastigiuta Thou. ou C. aizgtistipetala Ridley.
Hybridité et polyploïdie semblent donc dans le cas des Cynorkis être les facteurs favorables à l‘induction de leur état apomictique. Si l’hybri- dité a favorisé le rassemblement des gènes nécessaires à l’apparition de ce phénomène, la polyploïdie a pu modifier la physiologie de l’ovule si bien que l’action première du pollen qui, chez les Orcltidaceae, est celle d’un stimulus auxinique indispensable pour parfaire le développement de l’ovule, est ici inutile.
1. J. BOSSER pense que cette espèce existe aussi a la Réunion O ~ I elle est connue sous le nom d‘Act-ostyliaparadoxa,Frapp., lequel est l’ancien Hemiperis fissirostris Frapp. Mais ceci a besoin d‘être confirmé.
)
- 403 -
P1. 7. - Hybride présumé C. lilacina x C. ridleyi H. Perr. : 17, jeune ovule (a : archéspore; n : nucelle; ti : tégument interne); 18, ovule un peu plus âgé dans lequel une cellule de l'épiderme nucellaire commence à se différencier en initiale d'embryon (i) ou de gaméto- phyte virtuel; 19, deux initiales (i) bien développées et méiose de la CMM; 20, l'archk- spore a formé les dyades (d); bon développement d'un embryon nucellaire unicellulaire (en); entre la dyade inférieure et l'embryon nucellaire on distingue un autre embryon dont l'origine ne peut être précisbe. G x 565.
.- 401 -
PI. 8. - Hybride présumé C. Macina x C . ridleyi H. Perr. : 21, méiose de la CMM et embryon bicellulaire omnisaccal diploïde (eo); 22, formation des dyades (d) ; 23, embryon omni sacca1 diploïde (eo) et dyade présentant des signes de dégénérescence; 24, ovule avec deux embryons, l'un nucellaire (ed, l'autre omnisaccal diploïde (eo). G x 565.

- 402 -
BIBLIOGRAPHE
AFZELIUS, K. - Die Embryobildung bei NigritelIa nigra; Svensk Bot. Tidskr. 22, 1-2 :
GUSTAFSON, A. - Apomixis in higher plants Part I. The mecanism of apomixis; Lunds
PERRIER DE LA BÂTHIE, H. - in H. HUMBERT, Flore de Madagascar, 4ge famille, Orchi-
SCHLECHTER, R. - Systematischer Teil; Fedde Repert. 33 : 76-78 (1925). SESHAGIRMH, K. N. - Morphological studies in Orchidaceae; I. Zeiixine siilcata Lindley,
STEBBINS, G. L. - Cytology of Antennaria. II. Parthenogenetic species, Bot. Gaz. 94 :
. SWAMY, B. G. L. - The embryology of Zeuxine sulcata Lindl., New Phytol. 45, 1 :
VEYRET, Y . - L’apomixie chez’le Cynosorchis Macina Ridley (Orchidacées), C. R. Acad.
82-91 (1928).
Univ. Arsskrift. NF2, 42, 3 : 3-66 (1946).
dées, l (1939).
J. Indian bot Soc. 20 : 357-365 (1941).
322-344 (1932) (d’après GUSTAFSON, 1946).
132-136 (1946).
Sci., sér. D, 265 : 1713-1716 (1967).
Centre O.R.S.T.O.M. de Tananarive et Institut de Botanique, 91405-0RSAY.


















