
Etude de la résistance à la colistine de souches hospitalières
36
République Algérienne Démocratique et Populaire Ministère de l’Enseignement Supérieur et de la Recherche Scientifique Université Aboubekr Belkaïd Tlemcen Faculté des Sciences de la Nature et de la Vie, des Sciences de la Terre et de l’Univers Département de Biologie Laboratoire : Antibiotiques Antifongiques : physico-chimie, synthèse et activité biologique Mémoire En vue de l’obtention du diplôme de Master En sciences biologiques Option : Biochimie appliquée Présenté Par BENBOUZIANE Meriem Thème Soutenu le 11 Juillet 2017 Devant le jury Pr Boucherit-Otmani Zahia Université de Tlemcen Président Dr Sari-Belkherroubi Lamia Université de Tlemcen Examinatrice Dr Kazi-Tani Zahira Zakia Université de Tlemcen Promoteur Année universitaire 2016-2017 Etude de la résistance à la colistine de souches hospitalières d’Escherichia coli
Transcript of Etude de la résistance à la colistine de souches hospitalières

République Algérienne Démocratique et Populaire
Ministère de l’Enseignement Supérieur et de la Recherche Scientifique
Université Aboubekr Belkaïd Tlemcen
Faculté des Sciences de la Nature et de la Vie, des Sciences de la Terre et de l’Univers
Département de Biologie
Laboratoire :
Antibiotiques Antifongiques : physico-chimie, synthèse et activité biologique
Mémoire
En vue de l’obtention du diplôme de Master
En sciences biologiques
Option : Biochimie appliquée
Présenté
Par
BENBOUZIANE Meriem
Thème
Soutenu le 11 Juillet 2017
Devant le jury
Pr Boucherit-Otmani Zahia Université de Tlemcen Président
Dr Sari-Belkherroubi Lamia Université de Tlemcen Examinatrice
Dr Kazi-Tani Zahira Zakia Université de Tlemcen Promoteur
Année universitaire 2016-2017
Etude de la résistance à la colistine de souches hospitalières
d’Escherichia coli

Dédicace
Je dédie ce modeste travail
A
Mes chers parents que je remercie infiniment pour leurs conseils, encouragements et
leurs prières tout au long de mon cursus,
Papa, maman ; ce travail est le fruit de tous vos sacrifices, je vous aime,
Que Dieu vous garde pour nous
Mon mari BOUBAKEUR, qui m’a aidé et encouragé durant toute la période de ce
travail
Mes beaux-parents pour leurs encouragements
Mes sœurs FATIMA ZOHRA et KHADIJA
Mes frères MOHAMED et SALLAH EDDINE
Mes belles sœurs HADJER, FATIMA ZOHRA et AFAF
Mes beaux-frères FAYSAL et KHALED
Ma chère grand-mère
Mes cousines SABRINA, AMINA, FATIMA ZOHRA, MARWA et KAWTER.
La mémoire de ma tante ALIA et ma cousine FATIHA
Puisse Dieu tout puissant vous accorder sa clémence, sa miséricorde et vous
accueillir dans son saint paradis.

Remerciements
Je tiens tout d’abord à remercier Dieu le tout puissant et miséricordieux, qui nous a
donné la force et la patience d’accomplir ce modeste travail.
Je tiens à exprimer ma profonde gratitude à Mme Kazi Tani-Baba Ahmed Z.Z., Maître
de conférences classe B, au département de Biologie de la faculté des Sciences de la
Nature et de la Vie, Sciences de la Terre et de l’Univers de l’Université Aboubekr
Belkaïd Tlemcen, d’avoir accepté la charge de m’encadrer. Je la remercie vivement
pour l’aide scientifique précieuse et tous les conseils qu’elle a pu me fournir pendant
la durée de ce mémoire.
J’adresse mes vifs remerciements à Mme Boucherit-Otmani Z., Professeur au
département de Biologie de la faculté des Sciences de la Nature et de la Vie, des
Sciences de la Terre et de l’Univers de l’Université Aboubekr Belkaïd Tlemcen, pour
l’honneur qu’elle m’a fait en acceptant de présider ce jury. Qu’elle trouve ici l’assurance
de profond respect.
Mes remerciements s’adressent également à Mme Sari-Belkherroubi L., Maître de
conférences classe A au département de Biologie de la faculté des Sciences de la
Nature et de la Vie, des Sciences de la Terre et de l’Univers de l’Université Aboubekr
Belkaïd Tlemcen, pour l’honneur qu’elle m’a fait en acceptant de présider ce jury.
Qu’elle trouve ici l’assurance de ma respectueuse gratitude.
Je remercie également les doctorants du laboratoire "Antibiotiques Antifongiques
physico-chimie, synthèse et activité biologique", pour leur aide et conseils
Enfin, j'adresse mes plus sincères remerciements à toute personne qui a participé de
près ou de loin à l’accomplissement de ce modeste travail.

Sommaire
Première partie : Synthèse bibliographique ……………………………………………..... 1
Deuxième partie : Matériels et méthodes ………………………………………………….. 5
1. Matériel .………………………………………………………………………………… 6
1.1. Souches bactériennes ………………….………………………………………….. 6
1.2. Milieux de culture…………………………………………………………............... 6
1.3. Tests biochimiques et antibiotiques……………………………………................ 6
2. Méthodes…………………….………………………………………………………… 6
2.1. Identification ……………………..………………………………………………….. 6
2.2. Etude de la sensibilité aux antibiotiques …………………………………………. 7
2.2.1. Antibiogramme……………………………………………………...……………... 7
2.2.2. Détermination de la Concentration Minimale Inhibitrice ………………………. 7
Troisième partie : Résultats et discussion ………………………………………..….......... 8
1. Identification biochimique ……………………………….…………………………… 9
2. Sensibilité aux antibiotiques…………………………….……………………………. 10
Quatrième partie : Conclusion …………………………………………………………….... 15
Cinquième partie : Références bibliographiques …………………………………........... 17
Sixième partie : Annexes ………………………………………………………………........ 24

Liste des abréviations
L-Ara4N : 4-amino-4-désoxy- L arabinose
pEtN : phosphatidyléthanolamine
BLSE : β-lactamases à spectre élargi
LPS : Lipopolysaccharide
CTX-M: cefotaxime hydrolyzing capabilities
NDM: New Delhi métallo-beta-lactamase
OXA: oxacillin hydrolyzing capabilities
AAC : aminoglycosides acétyltransférases
aac : aminoglycosides acétyltransférases
sul: sulfamide
qnr : quinolone resistance gene
pmr : polymyxin resistance operon
Dab : acide L-α, γ-diaminobutyrique
mcr : mobilized colistin resistance gene
MCR : mobilized colistin resistance proteine
bla : β-lactamase gene
CMI : Concentration Minimale Inhibitrice
CLSI : Clinical and laboratory standard institute
UFC : Unité Formant Colonie

Liste des figures
Figure 1. Structure chimique de la colistine………………………………... 2
Figure 2. Mécanisme de résistance chromosomique acquise à la
colistine…………………………………………………………………………. 3
Figure 3. Identification d’une souche d’Escherichia coli (Ec2) par la
galerie API 20 E………………………………………………………………... 9
Figure 4. Résultat du E-test pour la colistine………………………………. 13
Figure 5. Phénotype de résistance d’une souche d’Escherichia coli
(Ec15) aux antibiotiques………………………………………………………. 14

Liste des tableaux
Tableau 1. Résultats d’antibiogramme des 15 souches d’Escherichia
coli vis-à-vis des molécules de β-lactamines ……………………………… 11
Tableau 2. Résultats d’antibiogramme des 15 souches d’Escherichia
coli vis-à-vis des antibiotiques testés ………………………………………. 12

Première partie
Synthèse bibliographique

Première partie Synthèse bibliographique
- 1 -
La résistance aux antimicrobiens continue d’être un problème de santé publique
grave tant en Algérie qu’à l’étranger. Elle soulève la possibilité que des infections
courantes et traitables redeviennent des infections mortelles, particulièrement
illustrées par la dissémination d’entérobactéries productrices de cabapénémases
(Dortet et coll., 2016).
Escherichia coli est une bactérie à Gram négatif, appartenant à la famille des
Enterobacteriaceae (Tenaillon et coll., 2010), anaérobie facultative, en forme de
bâtonnet, oxydase négative, nitrate positive, fermente le glucose et produit l’indole
en présence de trypthophane (Kaper et coll., 2004). Elle est considérée comme un
hôte normal, c'est-à-dire commensal, de la microflore digestive de l’homme et de la
plupart des animaux à sang chaud. La niche écologique de cette bactérie se trouve
dans la couche du mucus secrétée par l’épithélium du côlon (Russo et Johnson,
2000). Cependant, il existe des souches d’Escherichia coli pathogènes qui se
distinguent des souches commensales par l’acquisition de propriétés de virulence à
l’égard de l’hôte. Ces propriétés leur permettent de s’affranchir des mécanismes de
défense de l’hôte afin de s’établir dans de nouvelles niches écologiques et d’exprimer
leur pathogénicité (Sylvie, 2010). Selon les facteurs de virulence acquis, adhésines
ou toxines (Kaper et coll., 2004), les souches d’Escherichia coli pathogènes peuvent
être à l’origine d’infections du tractus digestif, de l’arbre respiratoire et du tractus
urinaire, mais également de méningites et de septicémies (Sylvie, 2010).
Escherichia coli est caractérisée par une aptitude particulière à acquérir les
résistances à des antibiotiques habituellement actifs (Oteo et coll., 2002).
La résistance aux β-lactamines est dominée par la production de β-lactamases à
spectre élargi (BLSE) et de carbapénémases (Chouchani et coll., 2011).
En Algérie, seules les BLSE de type CTX-M3 et CTX-M15 ont été identifiées chez
Escherichia coli (Baba Ahmed-Kazi Tani et Arlet, 2014). Ces enzymes confèrent
une résistance à la quasi-totalité des β-lactamines, excepté les céphamycines et les
carbapénèmes. Elles sont inhibées par les inhibiteurs de β-lactamases comme
l’acide clavulanique, le tazobactam et sulbactam (Bradford, 2001). Les gènes codant
pour ces enzymes sont principalement situés sur des éléments génétiques mobiles
expliquant la rapidité de leur diffusion (Partidge, 2011). Concernant la résistance aux
aminosides et aux quinolones, elle est marquée par la dissémination de nouveaux
déterminants de résistance tels les méthylases de l’ARN 16S(16S-RMTase), les
gènes qnr ou encore l’enzyme bi-fonctionnelle AAC(6’)-1b-cr (Baba Ahmed-Kazi

Première partie Synthèse bibliographique
- 2 -
Tani et Arlet, 2014). En Algérie, les gènes codant les aminoglycosides
acétyltransférases aac(3’)-II et aac(6’)-Ib ont été identifiés sur des plasmides
associés aux gènes de résistance aux β-lactamines (blaCTX-M) et aux quinolones
(qnrB) (Meradi et coll., 2011). La résistance aux sulfamides et au triméthoprime a
été liée à la présence d’intégrons de classe 1 portant les gènes sul1 et dfr-like et au
gène sul2 (Baba Ahmed-Kazi Tani et coll., 2013).
La multirésistance d’Escherichia coli aux antibiotiques a engendré le retour de la
colistine comme traitement de dernier recours, abandonné dans les années 80 en
raison de sa toxicité rénale et neurologique [(Abass et coll., 2012) ; (Biswas et coll.,
2012)]. Cet antibiotique appartient à la famille des polymyxines du groupe E.
Il est produit par Bacillus polymyxa subspecies colistinus (Frasca et coll., 2008).
Il est constitué d’un mélange complexe d’au moins 30 composés, dont les deux
majeurs sont la colistine A et la colistine B. Chaque composé de ce polypeptide est
un décapeptide cyclique lié à un acide gras (Figure 1) [(Turkoglu et coll., 2012) ;
(Li et coll., 2006)].
Figure 1. Structure chimique de la colistine (Li et coll., 2006)
La colistine exerce son activité antibactérienne en se liant au lipide A du
lipopolysaccharide (LPS), par interaction électrostatique entre les résidus d’acide
L-α-γ-diaminobutyrique (Dab) chargés positivement de la colistine et les groupes
phosphates chargés négativement du lipide A (Velkov et coll., 2009). Elle déplace

Première partie Synthèse bibliographique
- 3 -
par la suite de manière compétitive les cations divalents calcium (Ca 2+) et
magnésium (Mg 2+) qui stabilisent le LPS (Yahav et coll., 2012). La colistine forme
des pores dans la membrane externe, ce qui favorise son absorption dans la cellule
et permet le passage de molécules différentes (Fathy Mohamed et coll., 2016). Cela
conduit à la lyse de la cellule, la fuite du contenu intracellulaire et la mort cellulaire
(Rhouma et coll., 2016).
La résistance acquise d’Escherichia coli à la colistine est principalement liée à des
mutations chromosomiques conduisant à des modifications de charge du LPS
(Dortet et coll., 2016) par addition de phosphatidyléthanolamine (pEtN)
et/ou du 4-amino-4-désoxy- L-arabinose (L -Ara4N) (Bergen et coll., 2012).
Ces modifications impliquent l’activation des systèmes de régulation à deux
composants PhoP/PhoQ et PmrA/PmrB (Dortet et coll., 2016) en présence de
stimuli environnementaux ou par des mutations spécifiques (Figure 2) (Olaitan et
coll., 2014 ).
Figure 2. Mécanisme de résistance acquise chromosomique à la colistine
(Dortet et coll., 2014).
Le système PhoP/PhoQ est régulé par la protéine transmembranaire MgrB (Dortet
et coll., 2016). En condition de faible concentration en Mg2+, le récepteur
transmembranaire PhoQ phosphoryle PhoP. Ce dernier se lie au niveau de l’ADN et
induit la transcription de pmrD (Velkov et coll., 2014). Le gène mgrB exerce un
rétrocontrôle négatif sur le système PhoP/PhoQ en stimulant l’activité phosphatase

Première partie Synthèse bibliographique
- 4 -
de PhoQ, limitant ainsi l’activation de PhoP (Lippa et Goulian, 2009). Une délétion
au niveau du gène mgrB conduit à la régulation positive des gènes régulés par PhoP
(Cannatelli et coll., 2013). Le récepteur transmembranaire PmrB s’autophosphoryle
en présence de concentrations micromolaires de Fe3+, de Al3+ et d’un pH légèrement
acide (Prost et coll., 2007). Il transfère ses groupements phosphoryles à PmrA qui
se lie aux promoteurs des opérons arnBCADTEF (également nommé pmrHFIJKLM)
et pmrCAB (également nommé eptA), médiateurs de la synthèse du PEtN et de la L-
Ara4N ainsi que de leur transfert sur le lipide A (Olaitan et coll., 2014).
Récemment, des gènes de résistance plasmidique à la colistine, mcr-1 et mcr-2,
ont été décrits chez Escherichia coli [(Yi et coll., 2016) ; (Xavier et coll., 2016)].
Les protéines MCR font partie de la famille des phosphoéthanolamines transférases
dont l’expression aboutit à l’addition de pEtN sur le lipide A et conduit à une
diminution de sensibilité à la colistine (Liu et coll., 2016). Les modifications du LPS
par l’ajout de pEtN, confère un niveau de résistance à la colistine plus faible que ceux
conférés par l’ajout de L-Ara4N (Tamayo et coll., 2005). La protéine MCR-2 ne
possède que 81% d’identité en acides aminés avec MCR-1 suggérant une origine
différente des gènes mcr-1 et mcr-2 (Liu et coll., 2016). En Algérie, des études
récentes ont permis d’établir la présence du gène mcr-1 chez des souches
d’Escherichia coli d’origine clinique en 2011 (Berrazeg et coll., 2016) et d’origine
animale en 2015 (Yanat et coll., 2016).
La découverte du gène mcr-1 en Algérie nous a conduits à étudier le profil de
résistance à la colistine d’une collection de souches hospitalières d’Escherichia coli.

Deuxième partie
Matériels et méthodes

Deuxième partie Matériel et Méthodes
- 6 -
1. Matériel
1.1. Souches bactériennes
Une collection de quinze souches d’Escherichia coli du Laboratoire "Antibiotiques
Antifongiques : physico-chimie, synthèse et activité biologique" isolées au service de
réanimation du CHU de Tlemcen, a fait l’objet de notre étude. Ces souches ont été
entretenues par repiquage régulier et conservées à 4°C sur gélose nutritive.
La souche de référence Escherichia coli ATCC 25922 a été utilisée comme contrôle
interne.
1.2 Milieux de culture
• Bouillon nutritif (BN) (Institut Pasteur d’Algérie)
• Bouillon cœur cerveau (BHIB) (Fluka)
• Gélose Mac Conkey (Fluka)
• Gélose Mueller Hinton (Fluka)
1.3. Tests biochimiques et antibiotiques
• Galerie API 20 E (Bio Mérieux), disques d’oxydase (HIMEDIA)
• Ticarcilline (75µg), Ticarcilline / acide clavulanique (85µg), Céfamandol
(30µg), Céfoxitine (30µg), Céftazidime (30µg), Aztréonam (30µg), Imipenème
(10µg), Gentamicine (10µg), Tobramycine (10µg), Amikacine (30µg),
Ciprofloxacacine (5µg), Rifampicine (5µg), Colistine (10µg), Triméthoprime /
sulfamethoxasole (25μg).
2. Méthodes
2.1. Identification
L’identification des souches a été réalisée par galerie API 20E et test d’oxydase.
La galerie API 20E est une version miniaturisée et standardisée des techniques
biochimiques conventionnelles pour l’identification des entérobactéries.
L’ensemencement des 20 microtubes de la galerie a été réalisé par une suspension
bactérienne équivalente à 0,5 Mc Farland. La lecture des réactions produites pendant
la période d’incubation a été réalisée en se référant au tableau de lecture(Annexe 1)
et d’identification (Annexe 2).

Deuxième partie Matériel et Méthodes
- 7 -
Le test d’oxydase a été réalisé en ajoutant un disque d’oxalate N-diméthylphénylène-
diamine à une suspension bactérienne dense en eau physiologique. Les réactions
d’oxydation se sont traduites par une coloration violette.
2.2. Etude de la sensibilité aux antibiotiques
2.2.1. Antibiogramme
L’activité de 14 antibiotiques a été déterminée par la méthode de diffusion en gélose
Mueller Hinton. Un inoculum de 106 UFC/ml, obtenu à partir d’une dilution au 1/100
d’une culture de 18 heures en bouillon cœur cerveau (BHIB), a été ensemencé par
écouvillonnage à la surface du milieu gélosé. Les milieux ont été incubés à 37 C̊
pendant 18 à 24 heures. Les souches ont été catégorisées sensibles (S),
intermédiaires (I) ou résistantes (R) selon les diamètres critiques du Clinical and
Laboratory Standard Institut (CLSI, 2016)
2.2.2. Détermination de la Concentration Minimale Inhibitrice de la colistine
Les CMIs ont été déterminées par les méthodes de dilution et de diffusion (E-Test)
de la colistine en gélose Mueller Hinton selon les recommandations du CLSI (2016).
Les inoculums utilisés étaient de 104 UFC/spot et de 106 UFC/ml respectivement.
Pour la méthode de dilution, la colistine a été incorporée au milieu gélosé à des
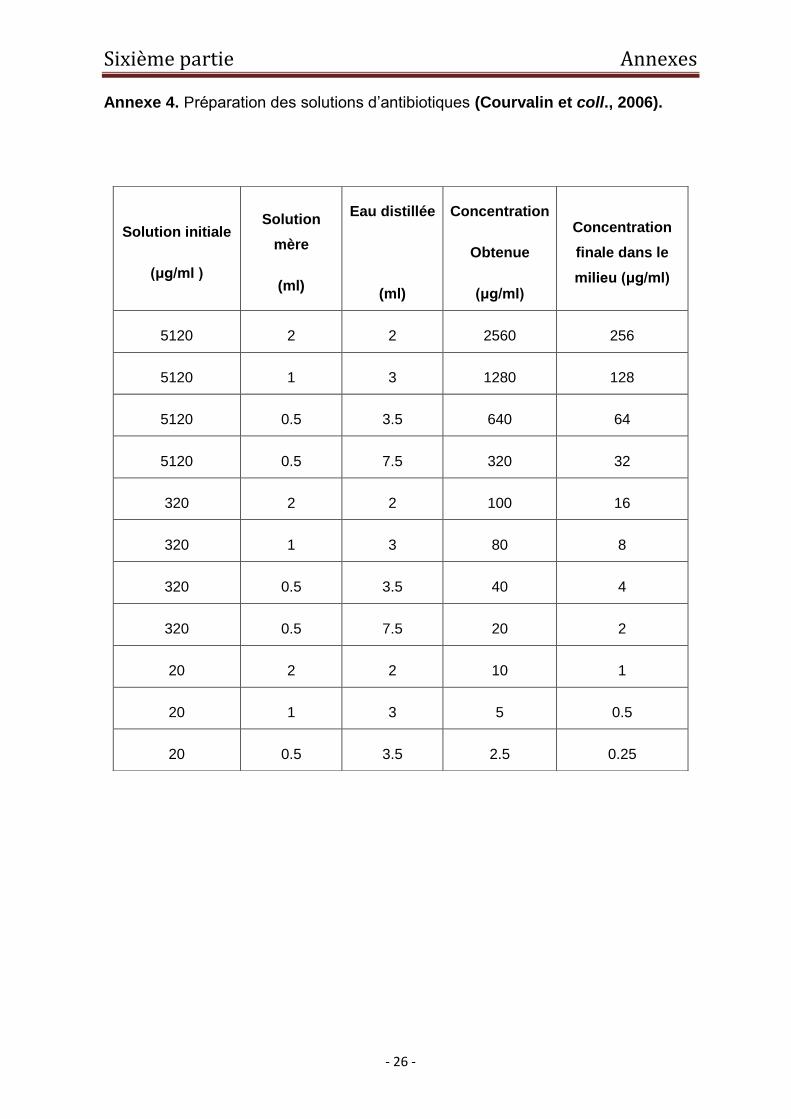
concentrations croissantes de raison 2 comprises entre 0,125 et 32 μg/ml
(Courvalin et coll., 2006) (Annexe 3). Après incubation des milieux à 37 ̊C pendant
18 à 24 heures, la CMI a été définie comme la plus petite concentration de colistine
inhibant toute croissance bactérienne visible. Pour la méthode de diffusion, la valeur
de la CMI correspondait au point d’intersection entre la limite de la zone d’inhibition
et la bande E-test.

Troisième partie
Résultats et discussion

Troisième partie Résultats et Discussion
- 9 -
1. Identification biochimique
Les résultats d’identification biochimique ont montré que les 15 souches étudiées
présentaient une béta-galactosidase, une lysine décarboxylase et une ornithine
décarboxylase ; elles fermentaient les sucres et donnaient une réponse négative aux
tests ADH, CIT, H2S, TDA, VP, GEL et uréase. A l’aide de ces résultats et en se
référant au catalogue d’identification API 20E (Annexe 1), les souches ont été
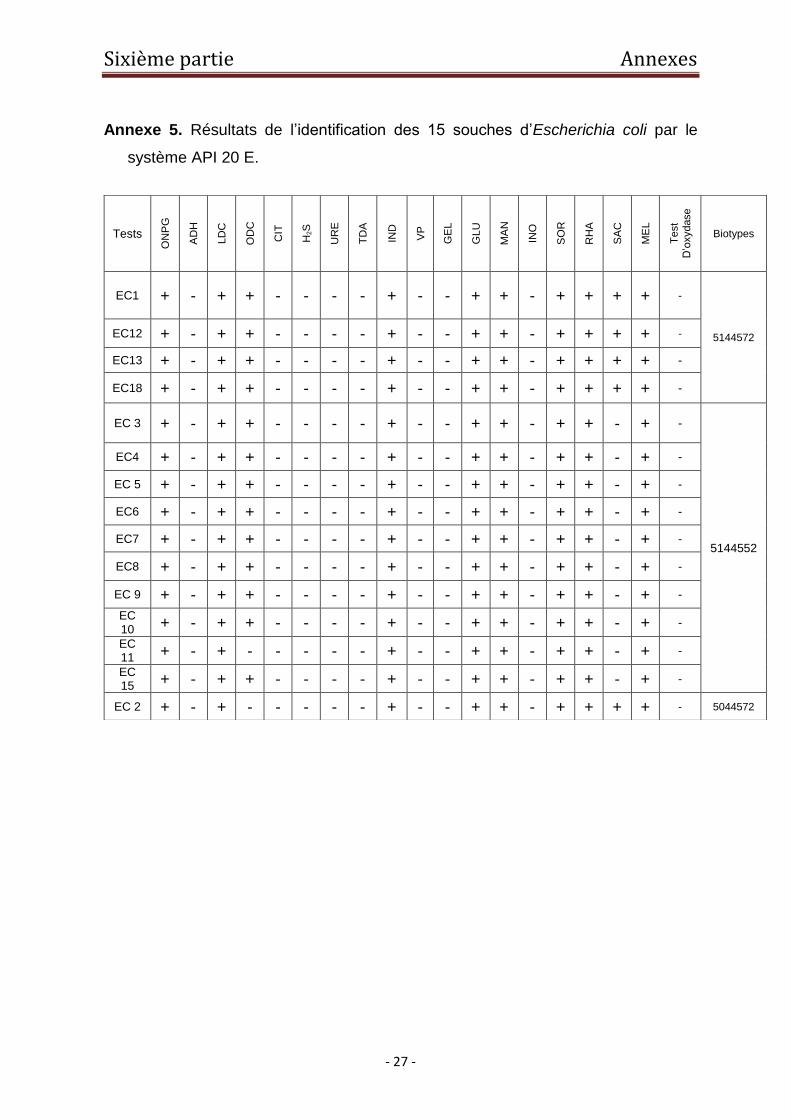
assignées au genre espèce Escherichia coli (Figure 3).
Figure 3. Identification d’une souche d’Escherichia coli (Ec2) par galerie API 20 E
La comparaison des biotypes, déterminés sur la base des réactions biochimiques
caractéristiques du genre Escherichia, nous a permis de classer les souches en 3
groupes. Le premier est représenté par le biotype 5144572 et regroupe 4 souches
(Ec1, Ec12, Ec13, Ec18). Le second correspond au biotype 5044572 et est
représenté par une seule souche (Ec2), alors que le troisième, représenté par le
biotype 5144552, comprend 10 souches (Annexe 5). L’appartenance des 10
souches au même biotype suggère un caractère clonal. L’utilisation des biotypes
comme marqueur phénotypique constitue un des premiers outils dont dispose le
laboratoire dans les investigations épidémiologiques. Cependant, démontrer ou
infirmer l’existence d’une souche clonale nécessite d’utiliser plusieurs approches afin
d’être certain de l’identité ou de la différence des souches (Bidet et Bingen, 2012).
Les souches d’Escherichia coli identifiées dans cette étude appartiennent au
laboratoire "Antibiotiques Antifongiques physico-chimie, synthèse et activité
biologique". Elles ont été isolées au service de réanimation du CHU de Tlemcen.
Dans ce service, le développement de mutants d’Escherichia coli responsables
d’infections nosocomiales s’est accéléré au cours des dix dernières années. Le
traitement de ces infections reste souvent difficile, de par la résistance naturelle et
acquise à de cette espèce à de nombreux antibiotiques (Oteo et coll., 2002).

Troisième partie Résultats et Discussion
- 10 -
2. Sensibilité aux antibiotiques
L’étude de la sensibilité des souches aux antibiotiques a été réalisée selon les
recommandations du CLSI (2016).
Toutes les souches d’Escherichia coli ont été testées par la méthode de diffusion en
milieu gélosé vis-à-vis de 14 molécules d’antibiotiques appartenant à différentes
familles, dont 7 β-lactamines ,3 aminosides, une fluoroquinolone, une rifamycine, une
polymixine et à l’association triméthoprime / sulfamethoxasole.
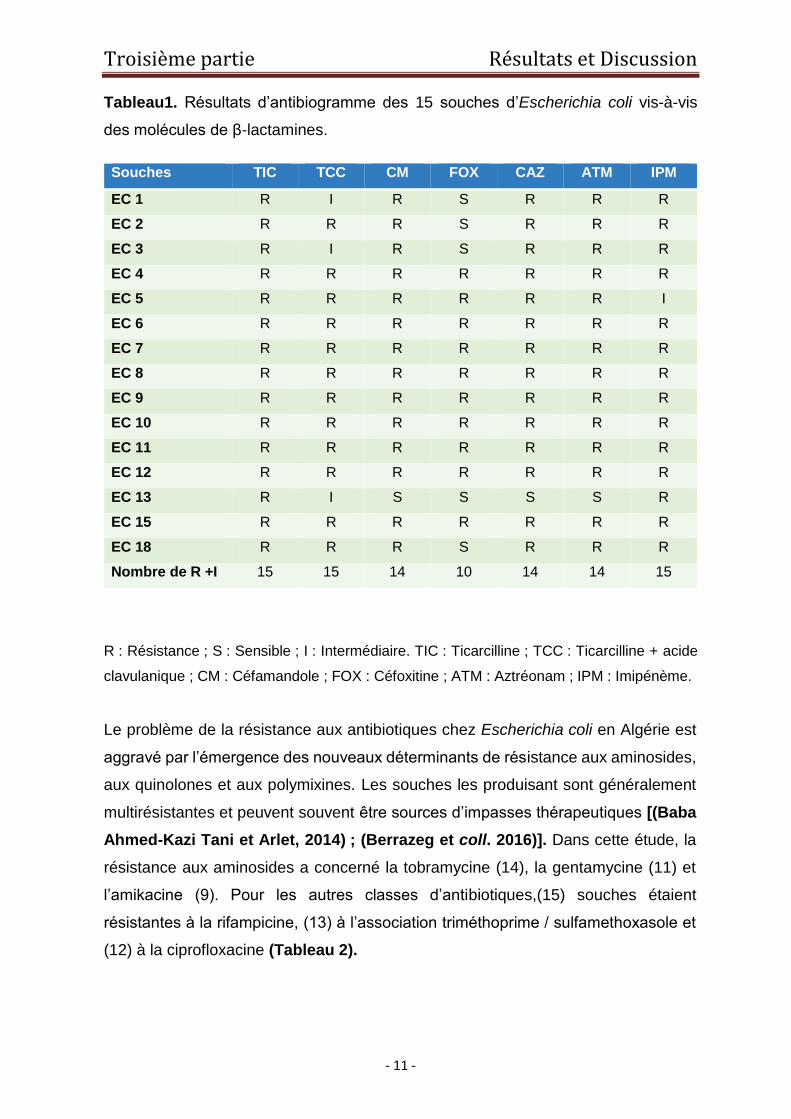
Les résultats d’antibiogramme vis-à-vis de la famille des β-lactamines ont montré que
les 15 souches étudiées étaient résistantes (R+I) à la ticarcilline et à l’association
ticarcilline / acide clavulanique. Quatorze (14) souches présentaient une résistance
au céfamondole, à la céftazidime et à l’aztréonam, et (10) souches à la céfoxitine
(Tableau 1).
Ces résultats sont en accord avec ceux des études précédentes réalisées sur les
souches d’Escherichia coli isolées au service de réanimation du CHU de Tlemcen
[(Baba Ahmed Kazi-Tani et coll., 2012) ; (Ayad et coll., 2016)] et ceux rapportés
par le 16ème rapport d’évaluation de la surveillance de la résistance des bactéries aux
antibiotiques publié par le ministère de la santé de la population et de la réforme
hospitalière d’Algérie (2015) (Rahal et coll., 2017).
L’utilisation croissante des antibiotiques au niveau du service de réanimation du CHU
de Tlemcen a contribué à l’émergence et à la diffusion de souches résistantes à la
quasi-totalité des β-lactamines. En effet, une résistance marquée à l’imipénème (15
souches) a été rapportée dans cette étude. Le mécanisme essentiel de cette
résistance peut être dû soit à une résistance enzymatique par production de
carbapénémases, soit à l’association de mécanismes tels une imperméabilité et une
céphalosporinase (Nordmann et Poirel, 2014).
Troisième partie Résultats et Discussion
- 11 -
Tableau1. Résultats d’antibiogramme des 15 souches d’Escherichia coli vis-à-vis
des molécules de β-lactamines.
R : Résistance ; S : Sensible ; I : Intermédiaire. TIC : Ticarcilline ; TCC : Ticarcilline + acide
clavulanique ; CM : Céfamandole ; FOX : Céfoxitine ; ATM : Aztréonam ; IPM : Imipénème.
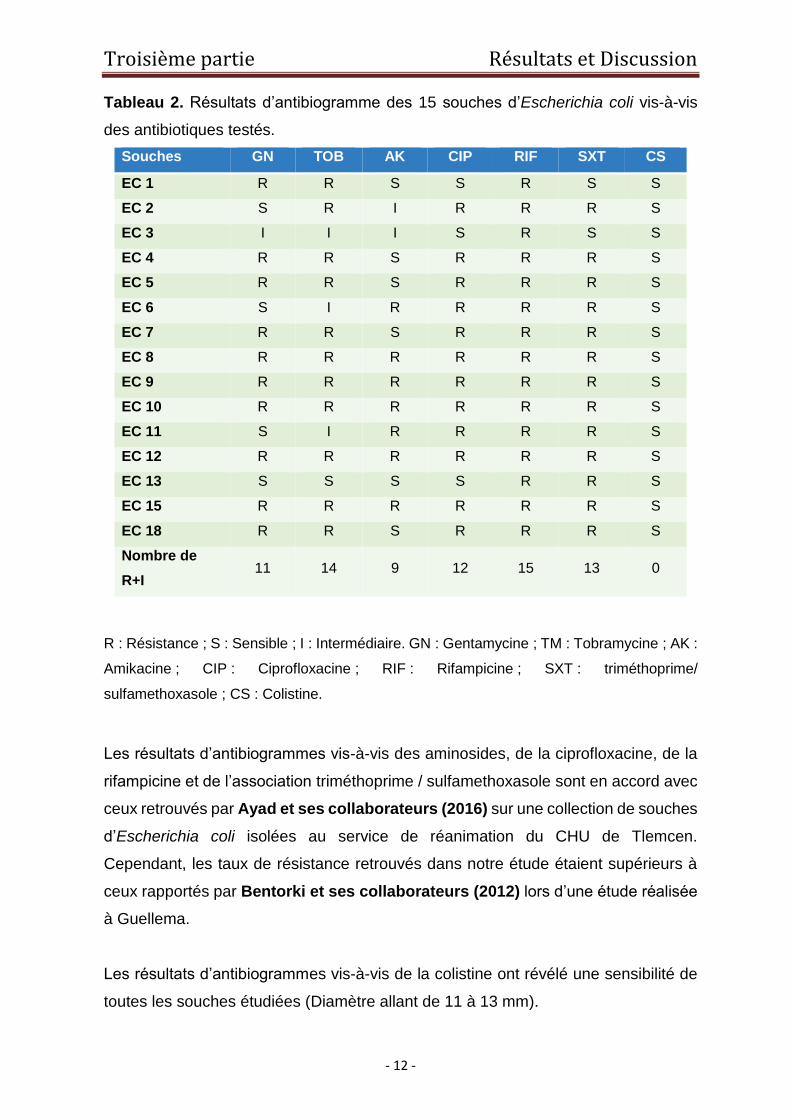
Le problème de la résistance aux antibiotiques chez Escherichia coli en Algérie est
aggravé par l’émergence des nouveaux déterminants de résistance aux aminosides,
aux quinolones et aux polymixines. Les souches les produisant sont généralement
multirésistantes et peuvent souvent être sources d’impasses thérapeutiques [(Baba
Ahmed-Kazi Tani et Arlet, 2014) ; (Berrazeg et coll. 2016)]. Dans cette étude, la
résistance aux aminosides a concerné la tobramycine (14), la gentamycine (11) et
l’amikacine (9). Pour les autres classes d’antibiotiques,(15) souches étaient
résistantes à la rifampicine, (13) à l’association triméthoprime / sulfamethoxasole et
(12) à la ciprofloxacine (Tableau 2).
Souches TIC TCC CM FOX CAZ ATM IPM
EC 1 R I R S R R R
EC 2 R R R S R R R
EC 3 R I R S R R R
EC 4 R R R R R R R
EC 5 R R R R R R I
EC 6 R R R R R R R
EC 7 R R R R R R R
EC 8 R R R R R R R
EC 9 R R R R R R R
EC 10 R R R R R R R
EC 11 R R R R R R R
EC 12 R R R R R R R
EC 13 R I S S S S R
EC 15 R R R R R R R
EC 18 R R R S R R R
Nombre de R +I 15 15 14 10 14 14 15
Troisième partie Résultats et Discussion
- 12 -
Tableau 2. Résultats d’antibiogramme des 15 souches d’Escherichia coli vis-à-vis
des antibiotiques testés.
R : Résistance ; S : Sensible ; I : Intermédiaire. GN : Gentamycine ; TM : Tobramycine ; AK :
Amikacine ; CIP : Ciprofloxacine ; RIF : Rifampicine ; SXT : triméthoprime/
sulfamethoxasole ; CS : Colistine.
Les résultats d’antibiogrammes vis-à-vis des aminosides, de la ciprofloxacine, de la
rifampicine et de l’association triméthoprime / sulfamethoxasole sont en accord avec
ceux retrouvés par Ayad et ses collaborateurs (2016) sur une collection de souches
d’Escherichia coli isolées au service de réanimation du CHU de Tlemcen.
Cependant, les taux de résistance retrouvés dans notre étude étaient supérieurs à
ceux rapportés par Bentorki et ses collaborateurs (2012) lors d’une étude réalisée
à Guellema.
Les résultats d’antibiogrammes vis-à-vis de la colistine ont révélé une sensibilité de
toutes les souches étudiées (Diamètre allant de 11 à 13 mm).
Souches GN TOB AK CIP RIF SXT CS
EC 1 R R S S R S S
EC 2 S R I R R R S
EC 3 I I I S R S S
EC 4 R R S R R R S
EC 5 R R S R R R S
EC 6 S I R R R R S
EC 7 R R S R R R S
EC 8 R R R R R R S
EC 9 R R R R R R S
EC 10 R R R R R R S
EC 11 S I R R R R S
EC 12 R R R R R R S
EC 13 S S S S R R S
EC 15 R R R R R R S
EC 18 R R S R R R S
Nombre de
R+I 11 14 9 12 15 13 0

Troisième partie Résultats et Discussion
- 13 -
La détection de la résistance à la colistine étant difficile par la méthode de diffusion
en milieu gélosé (Dortet et coll., 2016), la détermination des concentrations
minimales inhibitrices (CMIs) a été réalisée pour l’ensemble des souches étudiées
par la méthode de dilution en gélose Mueller Hinton. Les valeurs de CMI obtenus
(1 à 2µg/ml) nous ont permis d’assigner les souches à la catégorie sensible.
La détermination des CMIs vis-à-vis de la colistine a également été réalisée par la
méthode du E-test pour trois souches (Ec2, Ec3, Ec13). Les CMIs obtenus
(0.75 µg/ml) nous ont permis de catégoriser les souches sensibles (Figure 4).
Figure 4. Résultat du E-test pour la colistine
Les valeurs de CMIs obtenus ont révélé une bonne concordance avec les profils
d’antibiogrammes et nous ont permis de confirmer la sensibilité des souches étudiées
vis-à-vis de la colistine.
En Algérie, la prévalence de la résistance à la colistine reste basse avec seulement
deux études rétrospectives qui ont rapporté l’existence d’une résistance plasmidique
à la colistine parmi des souches d’Escherichia coli isolées en milieu hospitalier dont
deux de l’ouest algérien [(Berrazeg et coll. 2016); (Yanat et coll. 2016)].
La pression de sélection exercée au niveau des établissements de soins intensifs,
notamment lorsque seuls les antibiotiques les plus performants demeurent encore
actifs et utilisables, contribue probablement à faire évoluer les souches bactériennes
vers les pires phénotypes de résistance, ceux résistants “a tout” ou presque (Jeannot

Troisième partie Résultats et Discussion
- 14 -
et Plésiat, 2006). En effet, dans notre étude, les souches sensibles à la colistine,
étaient résistantes à au moins 5 antibiotiques. Quatre (4) étaient résistantes à 11
antibiotiques, (7) à 12 antibiotiques et une souche (Ec15) à 13 antibiotiques (Figure
5). Il en résulte que au niveau du service de réanimation du CHU de Tlemcen,
circulent des souches multirésistantes, clonales ou non, contre lesquelles les options
thérapeutiques s’avèrent très limitées.
Figure 5. Phénotype de résistance d’une souche d’Escherichia coli (Ec15)
aux antibiotiques.
TIC : Ticarcilline ; TCC : Ticarcilline + acide clavulanique ; CM : Céfamandole ; FOX :
Céfoxitine ; ATM : Aztréonam ; IPM : Imipénème ; GN : Gentamycine ; TM : Tobramycine ;
AK : Amikacine ; CIP : Ciprofloxacine ; RIF : Rifampicine ; SXT : triméthoprime/
sulfamethoxasole ; CS : Colistine.

Quatrième partie
Conclusion

Quatrième partie Conclusion
- 16 -
Escherichia coli est la première espèce d’entérobactérie impliquée en pathologie
infectieuse en milieu hospitalier. Cette bactérie est caractérisée par une aptitude
particulière à acquérir des mécanismes de résistances à des antibiotiques
habituellement actifs.
Les résultats des tests de sensibilité réalisés au cours de cette étude ont permis de
révéler d’importants taux de résistance aux β-lactamines y compris à l’imipénème.
Pour les autres classes d’antibiotiques, les résistances les plus élevées ont
concernées la rifampicine, l’association triméthoprime / sulfaméthoxazole suivies de
la gentamycine et de la ciprofloxacine. Seule la colistine était active sur toutes les
souches étudiées.
Cet antibiotique, initialement abandonné en réanimation, en raison d’une toxicité
supposée, revient en première ligne pour le traitement des infections dues à des
souches d’Escherichia coli multirésistantes. Une prescription abusive de cette
molécule pourrait contribuer à faire évoluer ces souches vers les phénotypes de
résistance à tous les antibiotiques.
Il serait donc important d’entreprendre une action de sensibilisation au bon usage
des antibiotiques et d’instaurer des réseaux de surveillance afin de limiter
l’émergence et la diffusion des souches multirésistantes et de préserver les
molécules les plus actives.

Cinquième partie
Références bibliographiques

Cinquième partie Références bibliographiques
- 18 -
1. Abbas M., Cherkaoui A., Fankhauser C., Harbarth S. (2012).
Carbapénémases implications cliniques et épidémiologiques pour la Suisse.
Rev. Med. Suisse, 8: 882-889.
2. Ayad A., Drissi M., Curraize C., Dupont C., Hartmann A., Solanas S et coll.
(2016). Occurrence of ArmA and RmtB Aminoglycoside resistance 16 S rRNA
methylases in extended-spectrum β-lactamases producing Escherichia coli in
Algerian hospitals, Front. Microbiol , 7 : 1409.
3. Baba Ahmed, Ayad A., Mesli E., Messai Y., Bakour R., Drissi M. (2012). CTX-
M-15 extended-spectrum β-lactamases in Enterobacteriaceae in the
intensive care unit of Tlemcen Hospital, Algeria. Pub Med., 18(4) : 382-385.
4. Baba Ahmed-Kazi Tani Z., Decre D., Genel N., Boucherit Otmani Z., Arlet G.,
Drissi. (2013). Molecular and Epidemiological Characterization of
enterobacterial Multidrug-Resistant Strains in Tlemcen Hospital (Algeria)(2008-
2010).Microbial Drug Resistance. 19 (3) : 185-190.
5. Baba Ahmed-Kazi Tani Z., Arlet G. (2014). News of antibiotic resistance among
Gram negative bacilli in Algeria. Pathol. Biol., 62: 169-78.
6. Bentroki A., Gouri A., Yakhlef A., Touaref A., Gueroudj A., Bensouilah T.
(2012). Résistance aux antibiotiques de souches isolées d’infections urinaires
communautaires entre 2007 et 2011 à Guelma (Algérie). Ann. Biol.Clin., 70
(6) : 666-8.
7. Bergen J.P., Landersdorfer C.B., Lee H.J., Li J., Nation R.L. (2012). ‘Old’
antibiotics for emerging multidrug-resistant bacteria. Current opinion in
infectious diseases, 25 (6): 626-633.
8. Berrazeg M., Hadjadj L., Ayad A., Drissi M., Rolain J. (2016). First Detected
Human Case in Algeria of mcr-1 Plasmid-Mediated Colistin Resistance in a
2011 Escherichia coli Isolate. Antimicrobial Agents and Chemotherapy,
60(11):6996-6997.
9. Bidet P et Bingen E. (2012). Apports des marqueurs moléculaires dans
l’analyse des mécanismes d’acquisition et dans le suivi des infections
nosocomiales. Mt. pédiatrie, 15 : 46-61.
10. Biswas S., Brunel J., Dubus J., Reynaud-Gaubert M., Rolain JM. (2012).
Colistin: an update on the antibiotic of the 21st century. Expert Rev. Anti.
Infect. Ther., 10: 917-34.

Cinquième partie Références bibliographiques
- 19 -
11. Bradford P A., (2001). Extended-spectrum beta-lactamases in the 21st century:
characterization,epidemiologie ,and detection of this important resistance
threat. Clin. Microbiol. Rev.,14:933-51.
12. Cannatelli A., D'andrea M.M., Giani T., Diplato V., Arna F., Amretti S et
coll.,(2013). In Vivo Emergence of Colistin Resistance in Klebsiella
pneumoniae Producing KPC-Type Carbapenemases Mediated by Insertional
Inactivation of the PhoQ/PhoP mgrB Regulator. Antimicrobial Agents and
Chemotherapy, 57 (11): 5521-5526.
13. Chouchani C., Marrakchi R., EL SA. (2011). Evolution of beta-lactams
resistance Gram-negative bacteria in Tunisia. Crit. Rev. Microbiol., 37: 167-
77.
14. Clinical and laboratory standard intitute (CLSI) (2016) Reference method for
broth dilution antimicrobial susceptibility testing of yeasts ; approved standard.
26th ed Document M11- A 8 .Wayne, PA : USA.
15. Courvalin P., Leclerck R., Bingen E. (2006). Antibiogramme. Paris.
ESKA :2éme édition. p :141-162.
16. Dortet L., Bonnin R., Jousset A., Gauthier L., Naas T. (2016). Émergence de
la résistance à la colistine chez les entérobactéries : une brèche dans le
dernier rempart contre la pan-résistance. Journal des Anti-infectieux,
18(4) :139-159.
17. Fathy Mohamed Y., Moustafa Abou-Shleib H., Mohamed Khalil A., Mohamed
El-Guink N., El-Nakeeb M.A.(2016). Membrane permeabilization of colistin
toward pan-drug resistant Gram-negative isolates. Brazilian Journal of
Microbiologie, 47: 381-388.
18. Frasca D., Dahyot-Fizelier C., Mimoz O. (2008). La colistine en réanimation.
Réanimation, 17: 251-258.
19. Jeannot K et Plésiat P. (2006). Implications thérapeutiques de la résistance
aux antibiotiques chez Pseudomonas aeruginosa. La lettre du pneumologue,
151-158.
20. Kaper, J. B., Nataro J. P., Mobley H. L. (2004). "Pathogenic Escherichia coli".
Nat Rev Microbiol, 2(2): 123-140.
21. Li J., Nation RL., Turnidge JD., Milne RW., Coulthard K, Rayner CR et coll.
(2006). Colistin: the re-emerging antibiotic for multidrugresistant Gram-
negative bacterial infections. Lancet. Infect. Dis, 6:589-601.

Cinquième partie Références bibliographiques
- 20 -
22. Lippa AM., Goulian M. (2009). Feedback inhibition in the PhoQ/PhoP signaling
system by a membrane peptide. PLOS Genet., 5: 1000788.
23. Liu YY., Wang Y., Walsh TR., Yi LX., Zhang R., Spencer J. et coll., (2016).
Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in
animals and human beings in China: a microbiological and molecular
biological study. Lancet. Infect. Dis, 16: 161-168.
24. Meradi L., Djahoudi A., Abdi A., Bouchakour M., Perrier Gros Claude JD.,
Timinouni M. (2011). Qnr and aac(6’)-Ib-cr types quinolone resistance among
Enterobacteriaceae isolated in Annaba, Algeria. Pathol. Biol., 59:73-8.
25. Nordman P et Poirel L. (2014). Résistances aux antibiotiques émergentes et
importantes chez les bactéries Gram négatif : épidémiologie,aspects
théoriques et détection. Revue Médicale Suisse, 10 : 902-7.
26. Olaitan A.O., Morand S., Rolain J.M. (2014). Mechanisms of polymyxin
resistance: acquired and intrinsic resistance in bacteria. Frontiers in
Microbiology, (5) 643: 18.
27. Oteo J., Campos J., Baquero F. (2002). Antibiotic resistance in 1962 invasive
isolates of Escherichia coli in 27 Spanish hospitals participating in the
European Antimicrobial Resistance Surveillance System (2001). Antimicrob.
Chemother., 50: 945-52.
28. Partridge SR., (2011). Analysis of antibiotic resistance regions in Gram-
negative bacteria. FEMS Microbiol. Rev., 35: 820-55.
29. Prost L.R., Daley M.E., Le Sage V., Bader M.W., Le Moual H., Rachel E et
coll. (2007). Activation of the Bacterial Sensor Kinase PhoQ by Acidic pH.
Molecular Cell, 26(2): 165-174.
30. Rahal K., Belouni R., Tali maamar H., Boudouane M., Missoum K., Benslimani
A., Aboun A. (2017). Surveillance de la résistance des bactéries aux
antibiotiques. 16ème Rapport d’évaluation.
31. Rhouma M., Beaudry F., Thériault W., Letellier A. (2016). Colistin in Pig
Production: Chemistry, Mechanism of Antibacterial Action, Microbial
Resistance Emergence, and One Health Perspectives. Front. Microbiol., 7:
1789.

Cinquième partie Références bibliographiques
- 21 -
32. Russo T. A., et Johnson J.R. (2000). Proposal for a new inclusive designation
for extraintestinal pathogenic isolates of Escherichia coli: ExPEC. J. Infect.
Dis. 181:1753‐4.
33. Sylvie M., (2010). Facteurs de virulence d’Escherichia coli adhérents et
invasifs associés à la maladie de Crohn : caractérisation et régulation de leur
expression. Thèse de doctorat. Microbiologie- Universite Blaise Pascal, p.8
34. Tamayo R., Prouty AM., Gunn JS. (2005). Identification and functional analysis
of Salmonella enterica serovar Typhimurium PmrA regulated genes. FEMS.
Immunol. Med. Microbiol., 43: 249-58.
35. Tenaillon O., Skurnik D., Picard B., Denamur E. (2010). The population
genetics of commensal Escherichia coli. Nat. Rev. Microbiol., 8: 201-217.
36. Turkoglu M., Dizbay M., Çiftçi F., Aygencel G. (2012). Colistin therapy in
critically ill patients with chronic renal failure and its effect on development of
renal dysfunction. Antimicrobial Agents, 39 (2) : 142-145.
37. Velkov T., Thompson P. E., Nation R. L., Li J. (2009). Structure-activity
relationships of polymyxin antibiotics. J. Med. Chem., 53: 1898-1916.
38. Velkov T., Deris ZZ., Huang JX., Azad MA., Butler M., Sivanesan S., et coll.
(2014). Surface changes and polymyxin interactions with a resistant strain of
Klebsiella pneumoniae. Innate. Immun. 20:350-63.
39. Xavier BB., Lammens C., Ruhal R., Kumar-Singh S., Butaye P., Goossens H.,
et coll .(2016). Identification of a novel plasmid-mediated colistin-resistance
gene, mcr-2, in Escherichia coli, Belgium, June 2016. Presse Med., 7(21)
:1560-7917.
40. Yahav H., Farbman L., Leibovici L., Paul M. (2012). Colistin: new lessons on
an old antibiotic. Clinical Microbiology and Infection, 18 (1): 18-29.
41. Yanat B., Machuca J., Dali Yahia R., Touati A., Pascual A., José-Manuel
Rodríguez-Martínez J.M. (2016). First report of the plasmid-mediated colistin
resistance gene mcr-1 in a clinical Escherichia coli isolate in Algeria.
International Journal of Antimicrobial Agents, 48(6): 760-761.
42. Yi L, Zhang R.,Spencer J., Doi Y., Tain G., Dong B et coll .(2016). Emergence
of plasmid-mediated colistin resistance mechanism MCR-1 in animals and
human beings in China: a microbiological and molecular biological
study.Infectious Deseases, 16(2): 161-168.

Sixième partie
Annexes
Sixième partie Annexes
- 23 -
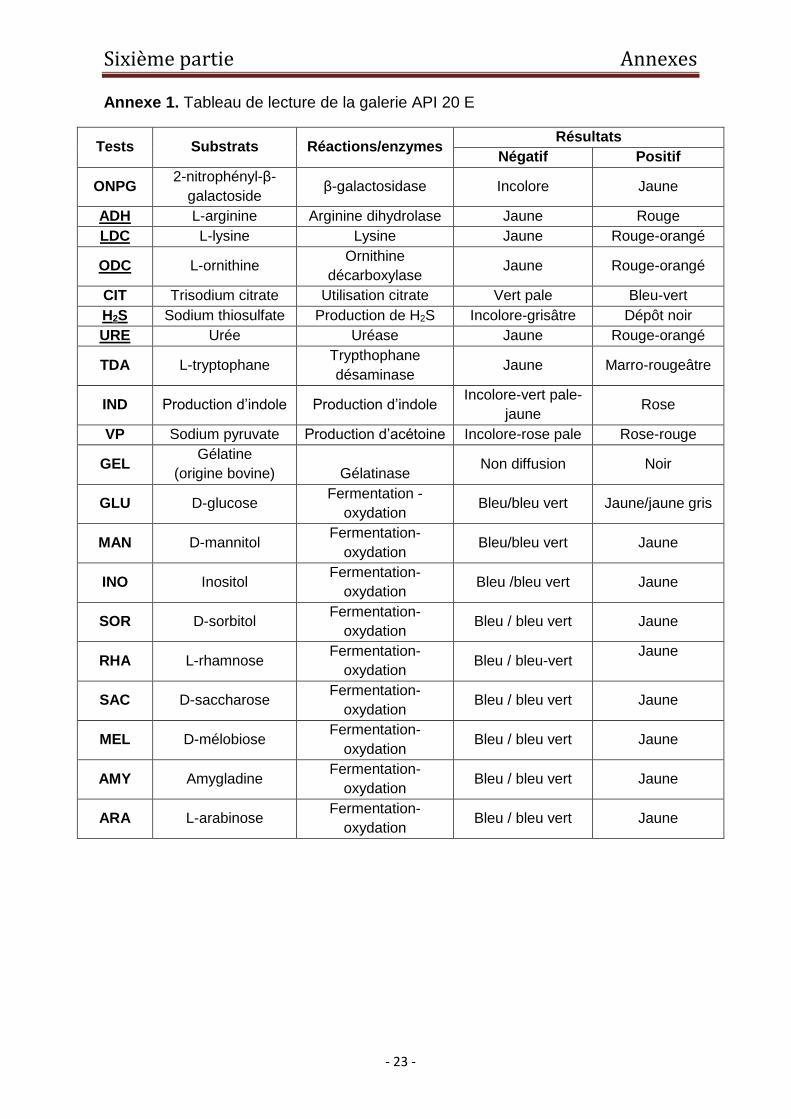
Annexe 1. Tableau de lecture de la galerie API 20 E
Tests Substrats Réactions/enzymes Résultats
Négatif Positif
ONPG 2-nitrophényl-β-
galactoside β-galactosidase Incolore Jaune
ADH L-arginine Arginine dihydrolase Jaune Rouge
LDC L-lysine Lysine Jaune Rouge-orangé
ODC L-ornithine Ornithine
décarboxylase Jaune Rouge-orangé
CIT Trisodium citrate Utilisation citrate Vert pale Bleu-vert
H2S Sodium thiosulfate Production de H2S Incolore-grisâtre Dépôt noir
URE Urée Uréase Jaune Rouge-orangé
TDA L-tryptophane Trypthophane
désaminase Jaune Marro-rougeâtre
IND Production d’indole Production d’indole Incolore-vert pale-
jaune Rose
VP Sodium pyruvate Production d’acétoine Incolore-rose pale Rose-rouge
GEL Gélatine
(origine bovine)
Gélatinase Non diffusion Noir
GLU D-glucose Fermentation -
oxydation Bleu/bleu vert Jaune/jaune gris
MAN D-mannitol Fermentation-
oxydation Bleu/bleu vert Jaune
INO Inositol Fermentation-
oxydation Bleu /bleu vert Jaune
SOR D-sorbitol Fermentation-
oxydation Bleu / bleu vert Jaune
RHA L-rhamnose Fermentation-
oxydation Bleu / bleu-vert
Jaune
SAC D-saccharose Fermentation-
oxydation Bleu / bleu vert Jaune
MEL D-mélobiose Fermentation-
oxydation Bleu / bleu vert Jaune
AMY Amygladine Fermentation-
oxydation Bleu / bleu vert Jaune
ARA L-arabinose Fermentation-
oxydation Bleu / bleu vert Jaune

Sixième partie Annexes
- 24 -
Annexe 2. Tableau d’identification du catalogue analytique API 20 E
Sixième partie Annexes
- 25 -
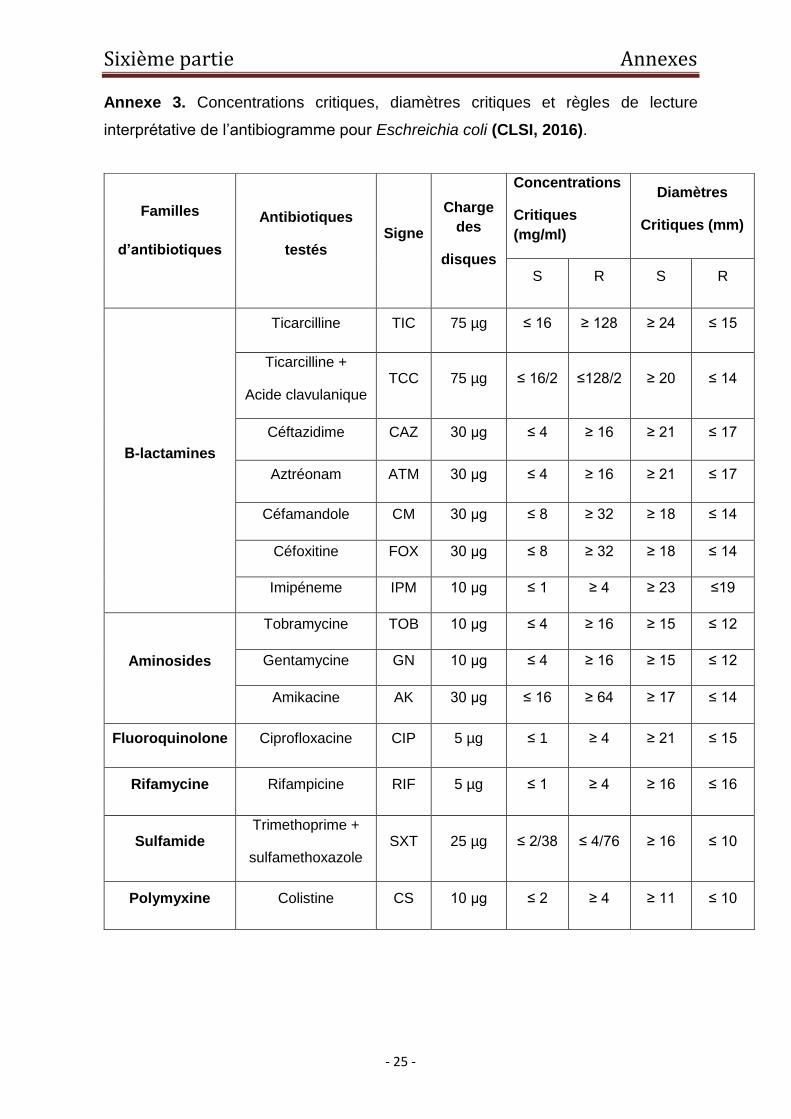
Annexe 3. Concentrations critiques, diamètres critiques et règles de lecture
interprétative de l’antibiogramme pour Eschreichia coli (CLSI, 2016).
Familles
d’antibiotiques
Antibiotiques
testés Signe
Charge
des
disques
Concentrations
Critiques
(mg/ml)
Diamètres
Critiques (mm)
S R S R
Β-lactamines
Ticarcilline TIC 75 µg ≤ 16 ≥ 128 ≥ 24 ≤ 15
Ticarcilline +
Acide clavulanique TCC 75 µg ≤ 16/2 ≤128/2 ≥ 20 ≤ 14
Céftazidime CAZ 30 μg ≤ 4 ≥ 16 ≥ 21 ≤ 17
Aztréonam ATM 30 μg ≤ 4 ≥ 16 ≥ 21 ≤ 17
Céfamandole CM 30 μg ≤ 8 ≥ 32 ≥ 18 ≤ 14
Céfoxitine FOX 30 μg ≤ 8 ≥ 32 ≥ 18 ≤ 14
Imipéneme IPM 10 μg ≤ 1 ≥ 4 ≥ 23 ≤19
Aminosides
Tobramycine TOB 10 μg ≤ 4 ≥ 16 ≥ 15 ≤ 12
Gentamycine GN 10 μg ≤ 4 ≥ 16 ≥ 15 ≤ 12
Amikacine AK 30 μg ≤ 16 ≥ 64 ≥ 17 ≤ 14
Fluoroquinolone Ciprofloxacine CIP 5 µg ≤ 1 ≥ 4 ≥ 21 ≤ 15
Rifamycine Rifampicine RIF 5 µg ≤ 1 ≥ 4 ≥ 16 ≤ 16
Sulfamide Trimethoprime +
sulfamethoxazole SXT 25 µg ≤ 2/38 ≤ 4/76 ≥ 16 ≤ 10
Polymyxine Colistine CS 10 μg ≤ 2 ≥ 4 ≥ 11 ≤ 10
Sixième partie Annexes
- 26 -
Annexe 4. Préparation des solutions d’antibiotiques (Courvalin et coll., 2006).
Solution initiale
(μg/ml )
Solution
mère
(ml)
Eau distillée
(ml)
Concentration
Obtenue
(μg/ml)
Concentration
finale dans le
milieu (μg/ml)
5120 2 2 2560 256
5120 1 3 1280 128
5120 0.5 3.5 640 64
5120 0.5 7.5 320 32
320 2 2 100 16
320 1 3 80 8
320 0.5 3.5 40 4
320 0.5 7.5 20 2
20 2 2 10 1
20 1 3 5 0.5
20 0.5 3.5 2.5 0.25
Sixième partie Annexes
- 27 -
Annexe 5. Résultats de l’identification des 15 souches d’Escherichia coli par le
système API 20 E.
Tests
ON
PG
AD
H
LD
C
OD
C
CIT
H2S
UR
E
TD
A
IND
VP
GE
L
GLU
MA
N
INO
SO
R
RH
A
SA
C
ME
L
Te
st
D’o
xydase
Biotypes
EC1 + - + + - - - - + - - + + - + + + + -
5144572 EC12 + - + + - - - - + - - + + - + + + + -
EC13 + - + + - - - - + - - + + - + + + + -
EC18 + - + + - - - - + - - + + - + + + + -
EC 3 + - + + - - - - + - - + + - + + - + -
5144552
EC4 + - + + - - - - + - - + + - + + - + -
EC 5 + - + + - - - - + - - + + - + + - + -
EC6 + - + + - - - - + - - + + - + + - + -
EC7 + - + + - - - - + - - + + - + + - + -
EC8 + - + + - - - - + - - + + - + + - + -
EC 9 + - + + - - - - + - - + + - + + - + -
EC 10 + - + + - - - - + - - + + - + + - + -
EC 11 + - + - - - - - + - - + + - + + - + -
EC 15 + - + + - - - - + - - + + - + + - + -
EC 2 + - + - - - - - + - - + + - + + + + - 5044572

ملخص
ينتمون لاشيريشيا كولي سلالة 15 من لمجموعة مضاد حيوي 14لـ للبكتيريا المضاد النشاط تقييم إلى الدراسة هذه هدفت
قابلية ) بطريقة التحليل "البيولوجي النشاط و الفيزيائية، الكيمياء: الفطريات ومضادات يويةالح المضادات" مختبر إلى
E-test-) وفقا للمعايير المقترحة من قبل و التخفيف في وسط صلبCLSI . من كبيرة معدلات النتائج أظهرت
نسب الحيوية، المضادات من الأخرى ا يخص الفئاتالإيميبينيمن. و فيم ذلك في بما اكتام الحيوية بيتا للمضادات المقاومة
نشطة كوليستين فقط. وسيبروفلوكساسين التوبراميسين سلفاميثوكسازول/ ميثوبريم ثم ريفامبين، شملت قد العليا المقاومة
.المدروسة السلالات جميع على
مقاومة. –تخفيف –مضادات حيوية –تحليل –ا نشاط مضاد للبكتيري –Escherichia coli :الكلمات المفتاحية
Résumé
Cette étude avait pour but d’évaluer l’activité antibactérienne de 14 antibiotiques pour
une collection de 15 souches d’Escherichia coli appartenant au laboratoire
"Antibiotiques Antifongiques : physico-chimie, synthèse et activité biologique" par les
méthodes de diffusion (antibiogramme, E-Test) et de dilution en milieu gélosé (CLSI
2016). Les résultats obtenus ont révélé d’importants taux de résistance aux β-
lactamines y compris à l’imipénème. Pour les autres classes d’antibiotiques, les
résistances les plus élevées ont concernées la rifampicine, l’association
triméthoprime / sulfaméthoxazole suivies de la tobramycine et de la ciprofloxacine.
Seule la colistine était active sur toutes les souches étudiées.
Mots clés : Escherichia coli- activité antibactérienne-diffusion-antibiotiques-dilution-
résistance.
Abstract
The aim of this study was to evaluate the antibacterial activity of 14 antibiotics for a
collection of 15 strains of Escherichia coli belonging to the laboratory "Antibiotics
Antifungals : physico-chemistry, synthesis and biological activity" by diffusion
methods (antibiogram, E -Test) and dilution in agar medium (CLSI 2016). The results
obtained revealed high levels of resistance to β-lactams including imipenem. For the
other classes of antibiotics, the highest resistances were rifampicin, trimethoprim /
sulfamethoxazole followed by tobramycin and ciprofloxacin. Only colistin was active
on all strains studied.
Keywords : Escherichia coli - antibacterial activity -diffusion-antibiotics-dilution-
resistance.


















