
ETUDE DE LA DIVERSITE TAXONOMIQUE DES POACEAE …
102
UNIVERSITE D’ANTANANARIVO FACULTE DES SCIENCES DOMAINE : SCIENCES ET TECHNOLOGIE MENTION : BIOLOGIE ET ECOLOGIE VEGETALES Mémoire pour l’obtention du diplôme de MASTER Parcours : SYSTEMATIQUE ET GESTION DURABLE DE LA DIVERSITE VEGETALE (SYGEDUR) ETUDE DE LA DIVERSITE TAXONOMIQUE DES POACEAE MALGACHES : Cas Du Parc National d’Isalo Présenté par RAKOTOMALALA Nantenaina Herizo Soutenu publiquement le 07 Août 2019 devant la commission d’examen composée de : Président : Prof. Vonjison RAKOTOARIMANANA Rapporteurs : Prof. Vololoniaina JEANNODA Dr Maria VORONTSOVA Examinateurs : Dr Hélène RALIMANANA Dr Mijoro RAKOTOARINIVO
Transcript of ETUDE DE LA DIVERSITE TAXONOMIQUE DES POACEAE …

UNIVERSITE D’ANTANANARIVO
FACULTE DES SCIENCES
DOMAINE : SCIENCES ET TECHNOLOGIE
MENTION : BIOLOGIE ET ECOLOGIE VEGETALES
Mémoire pour l’obtention du diplôme de MASTER
Parcours : SYSTEMATIQUE ET GESTION DURABLE DE LA DIVERSITE
VEGETALE (SYGEDUR)
ETUDE DE LA DIVERSITE TAXONOMIQUE DES
POACEAE MALGACHES : Cas Du Parc National
d’Isalo
Présenté par RAKOTOMALALA Nantenaina Herizo
Soutenu publiquement le 07 Août 2019 devant la commission d’examen composée de :
Président : Prof. Vonjison RAKOTOARIMANANA
Rapporteurs : Prof. Vololoniaina JEANNODA
Dr Maria VORONTSOVA
Examinateurs : Dr Hélène RALIMANANA
Dr Mijoro RAKOTOARINIVO

UNIVERSITE D’ANTANANARIVO
FACULTE DES SCIENCES
DOMAINE : SCIENCES ET TECHNOLOGIE
MENTION : BIOLOGIE ET ECOLOGIE VEGETALES
Mémoire pour l’obtention du diplôme de MASTER
Parcours : SYSTEMATIQUE ET GESTION DURABLE DE LA DIVERSITE
VEGETALE (SYGEDUR)
ETUDE DE LA DIVERSITE TAXONOMIQUE DES
POACEAE MALGACHES : Cas Du Parc National
d’Isalo
Présenté par RAKOTOMALALA Nantenaina Herizo
Soutenu publiquement le 07 Août 2019 devant la commission d’examen composée de :
Président : Prof. Vonjison RAKOTOARIMANANA
Rapporteurs : Prof. Vololoniaina JEANNODA
Dr Maria VORONTSOVA
Examinateurs : Dr Hélène RALIMANANA
Dr Mijoro RAKOTOARINIVO

Photo de couverture : groupe d’Hyparrhenia rufa, fréquent dans le circuit Malaso, Isalo.
Crédit photo: DAVID RABEHEVITRA © 2018.

Remerciement
Grâce soit rendue à Dieu primordialement car c’est par son amour miséricordieux et sa
bénédiction que je suis arrivée au terme de ce mémoire.
La présente étude est le fruit de la collaboration entre l’Université d’Antananarivo, la
Mention Biologie et Ecologie Végétales, le Royal Botanic Gardens (RBG) Kew et le Kew
Madagascar Conservation Centre (KMCC). Ainsi j’adresse mes sincères gratitudes au personnel
et aux enseignants de ces institutions, ainsi qu’à toutes les personnes qui ont contribué de loin et
de près leur aide dans la réalisation de cette étude. Ainsi je remercie :
Les membres du jury :
❖ Professeur Vonjison RAKOTOARIMANANA, Enseignant-Chercheur, Mention Biologie
et Ecologie Végétales, Faculté des Sciences, Université d’Antananarivo, qui m’a fait un grand
honneur de présider le jury et qui a apporté des critiques très instructives sur la présentation de
cette étude grâce à ses expériences sur la rédaction scientifique. Je vous témoigne ma sincère
gratitude.
❖ Professeur Vololoniaina JEANNODA, Enseignant-Chercheur, Mention Biologie et
Ecologie Végétales, Faculté des Sciences, Université d’Antananarivo, qui a montré la volonté de
diriger cette recherche et de m’encadrer durant la réalisation de cette étude, depuis le terrain
jusqu’à sa présentation, malgré ses lourdes responsabilités. Veuillez accepter mes sincères
remerciements, Madame.
❖ Docteur Maria VORONTSOVA, Botaniste et Chercheur, Royal Botanic Gardens Kew,
Londres, qui m’a accordée confiance et support durant la réalisation de cette recherche. Je la
remercie de m’avoir supervisée et d’avoir eu l’amabilité de me transmettre ses connaissances
sur les Poaceae de Madagascar. Je vous adresse mes remerciements les plus sincères.
❖ Docteur Hélène RALIMANANA, Chercheur et team leader, Kew Madagascar
Conservation Centre, Royal Botanic Gardens Kew ; qui a assuré l’organisation administrative
de cette recherche, et qui accepté sans hésitation à examiner cette étude toute en apportant les
améliorations possibles. Veuillez recevoir mes reconnaissances les plus vives, Madame.
❖ Docteur Mijoro RAKOTOARINIVO, Maître de conférences, Mention Biologie et
Ecologie Végétales, Facultés des Sciences, Université d’Antananarivo, qui a consacré son temps

à examiner ce mémoire malgré ses occupations. Je lui suis humblement reconnaissante pour ses
précieux critiques et conseils qui ont significativement permis d’améliorer ce mémoire. Je vous
adresse ma reconnaissance le plus profond, Monsieur.
Le Kew Madagascar Conservation Centre (KMCC), ainsi que son personnel, qui m’ont
accueillie, m’ont rendue service, et m’ont conseillée lors la réalisation de cette recherche,
particulièrement à :
❖ David RABEHEVITRA, Chercheur, botaniste et photographe, qui nous a accompagnés
durant les travaux sur terrain et a réalisé la prise et le traitement de quelques photos presentées
dans le présent manuscrit. Veuillez accepter mes vifs remerciements.
❖ Andotiana Mihaja ANDRIAMANOHERA, Chercheur et botaniste, qui nous a
également apportés son aide et son assistance lors des travaux sur terrain, et qui n’a pas hésité à
donner des conseils amélioratifs à ce travail. Qu’elle croit à ma sincère reconnaissance.
❖ Lova Cédrique SOLOFONDRANOHATRA, Etudiante chercheur de l’Université
d’Antananarivo qui fait sa thèse de Doctorat sur l’Ecologie des savanes malgaches, qui a offert
son assistance technique et instructive, et a été toujours partante pour discuter ou pour échanger
des idées sur la réalisation de cette recherche, malgré ses occupations. Qu’elle croit à ma
sincère gratitude.
❖ Tatamo RANAIVOMANANA, Chauffeur : sans lui, nous n’avons pas pu effectuer les
inventaires sur terrain. Il a fait preuve d’effort en nous conduisant à Isalo et dans les différents
sites de collectes, des plus faciles aux plus difficiles à franchir. Je vous suis vraiment
reconnaissante.
❖ L’équipe responsable de la base de données de KMCC : Linah RABARIVOLA,
Henintsoa RAZANAJATOVO, Velosoa RAZAFINIARY, qui m’a apprise à servir le logiciel
BRAHMS et m’a donnée la base de données nécessaire requise dans cette recherche. Je vous
remercie du fond du cœur.
Le Département Flore au sein du Parc Botanique et Zoologique de Tsimbazaza (PBZT) et son
personnel, qui ont contribué par leur assistance dans cette étude, particulièrement :

❖ Docteur Solohery Jean Victor RAPANARIVO, Chercheur et chef du département Flore,
qui m’a accueillie au sein de l’Herbarium et le laboratoire de TAN afin que je puisse effectuer
l’identification de mes spécimens. Veuillez accepter mes vifs remerciements.
❖ Madame Jacqueline RAZANATSOA, Botaniste et technicien qui a participé aux
collectes de données sur terrain et a transmis ses acquis sur le mode de collecte et le maniement
des spécimens d’herbiers. Je vous remercie sincèrement.
❖ Monsieur Rokiman LETSARA, Botaniste et Chercheur, qui m’a accueillie au sein du
laboratoire de CAS et m’a apprise les techniques d’usage du microscope utilisé lors de
l’identification des spécimens. Veuillez accepter mes vifs remerciements, Monsieur.
J’adresse aussi une particulière reconnaissance à Monsieur Roger Lala ANDRIAMIARISOA,
Chercheur et illustrateur botanique basé au sein de Missouri Botanical Gardens Madagascar
qui m’a volontairement donnée conseils et m’a apprise les techniques sur l’illustration
botanique. Qu’il croit à ma sincère gratitude.
J’adresse encore et également mes vifs remerciements au personnel du Madagascar National
Parks Ranohira, particulièrement :
❖ Monsieur Ranaivojaona RANDRIANTSITOHAINA, Responsable du site, qui nous a
accueillis et donnés l’autorisation de recherche dans le parc, et a supervisé notre investigation à
Isalo. Je vous remercie de votre collaboration, Monsieur.
❖ Messieurs Ivon TOVONDRAINY et Noel RANDRIANASOLO, Agents du MNP
Ranohira, qui ont prouvé leur sympathie et leur motivation de nous assister et aider durant notre
séjour de collecte à Isalo. Sans eux, je n’ai pas pu réussir à récolter tous ces spécimens. Je vous
suis de tout cœur reconnaissante.
Je tiens aussi à adresser mes profonds remerciements à tous les Enseignants-Chercheurs de la
Mention Biologie et Ecologie Végétales, Université d’Antananarivo, qui nous ont formés durant
les années d’études à devenir des chercheurs. Je vous suis sincèrement reconnaissante.
Je tiens également à remercier ma grand-mère (Bebe Henriette, paix à son âme), mes parents,
mes frères (Nambinina, Nasandratra et Naliniaina), ma tante et mon oncle, ainsi que toute la

famille pour leurs soutiens spirituels, moraux et matériels durant mes années d’étude. Sans vous
je n’ai pas pu réussir et achever mes études, je vous suis à jamais reconnaissante !
Et finalement, je remercie mes meilleurs amis MIHARIVOLA Hasina Norbert, et Radoniaina
RAKOTOMANANDRIANA, qui m’ont toujours soutenue et encouragée. Merci mes amis !
Et merci à tous

TABLE DE MATIERES
LISTE DES FIGURES .................................................................................................................... i
LISTE DES PHOTOS...................................................................................................................... i
LISTE DES TABLEAUX................................................................................................................ i
LISTE DES ANNEXES ..................................................................................................................ii
GLOSSAIRE....................................................................................................................................ii
I. INTRODUCTION ................................................................................................................... 1
I. MILIEU D’ÉTUDE ................................................................................................................. 4
II.1.1 - Localisation et rattachement administratif ................................................................................ 4
II.1.2 - Climat ........................................................................................................................................ 5
II.1.3 - Géomorphologie et topographie ................................................................................................ 6
II.2 - MILIEU BIOLOGIQUE ................................................................................................................... 7
II.2.1 - Flore et végétation ..................................................................................................................... 7
II.2.1.1 - Bois de Tapia ......................................................................................................................... 7
II.2.1.2 - Forêts galeries ........................................................................................................................ 7
II.2.1.3 - Végétation rupicole ................................................................................................................ 8
II.2.1.4 - Savanes et prairies .................................................................................................................. 8
II.2.1.5 - Forêts et fourrées secondaires ................................................................................................ 9
II.2.1.6 - Marais et marécage ................................................................................................................ 9
II.2.2 - Faune ......................................................................................................................................... 9
II.2.3 - L’Homme et ses activités .......................................................................................................... 9
II. MATERIELS ET METHODES ............................................................................................ 11
III.1.1 - Caractères morphologiques ................................................................................................... 11
III.1.1.1 - Tige ..................................................................................................................................... 11
III.1.1.2 - Feuilles ................................................................................................................................ 12
III.1.1.3 - Inflorescence ....................................................................................................................... 12
III.1.1.4 - Epillets ................................................................................................................................ 12

III.1.1. 5 - Fleuron et fleur .................................................................................................................. 13
III.1.1.6 - Caryopse ............................................................................................................................. 14
III.1.2 - Base de données sur les Poaceae ........................................................................................... 15
III.2 - METHODES ................................................................................................................................. 15
III.2.1 - Etude préliminaire ................................................................................................................. 15
III.2.1.1 - Recherche bibliographique ................................................................................................. 15
III.2.1.2 - Apprentissage de l’identification des Poaceae .................................................................... 16
III.2.2 - Collectes de données .............................................................................................................. 16
III.2.2.1 - Téléchargement et vérification des données existantes ...................................................... 16
III.2.2.2 - Collecte de données sur terrain, identification et transcription des données sur BRAHMS
............................................................................................................................................................ 16
III.2.3 - Compilation des données et listage des espèces .................................................................... 18
III.2.4 - Calcul du nombre cumulé de collectes et d’espèces .............................................................. 19
III.2.5 - Aire de distributions des espèces ........................................................................................... 19
III.2.6 - Etablissement d’une clé de détermination pour les Poaceae de l’Isalo ................................. 19
III.2.7 - Analyse et étude de la diversité ............................................................................................. 20
III.2.7.1 - Richesse spécifique ............................................................................................................. 20
III.2.7.2 - Richesse spécifique et degré d’endémisme au niveau des habitats .................................... 20
III. RESULTATS ET INTERPRETATIONS ............................................................................. 22
IV.1 - LISTE DES ESPECES DE POACEAE DE L’ISALO ...................................................... 22
IV.1.1 – Sites de collectes ................................................................................................................... 23
IV.1.2 - Collecteurs ............................................................................................................................. 24
IV.1.3 - Découvertes pour Isalo .......................................................................................................... 25
IV.1.4 - Découverte pour la science .................................................................................................... 26
IV.2- EVOLUTION DE LA CONNAISSANCE SUR LES POACEAE DE L’ISALO ......................... 26
IV.2.1 - Espèces natives ...................................................................................................................... 28
IV.2.2 - Espèces introduites ................................................................................................................ 29
IV.2.3 - Espèces endémiques .............................................................................................................. 29
IV.2.4 - Espèces endémiques locales de l’Isalo .................................................................................. 29

IV.3 - DIVERSITE TAXONOMIQUE DES POACEAE DE L’ISALO ................................................ 32
IV.3.1 - Diversité au niveau des sous familles .................................................................................... 32
IV.3.2 - Diversité au niveau des tribus ................................................................................................ 33
IV.3.3 - Diversité au niveau des genres .............................................................................................. 33
IV.4 - CLE DE DETERMINATION DES GENRES DE POACEAE DE L’ISALO ............................. 34
IV.5 - ZONE DE DIVERSITE SPECIFIQUE DANS LE PARC ........................................................... 34
IV.6 - DIVERSITE ET ENDEMISME PAR TYPE DE VEGETATION ............................................... 36
IV.6.1 - Tendance de distribution des espèces dans le parc ................................................................ 36
IV.6.2 - Richesse spécifique et endémisme au niveau des habitats .................................................... 37
V. DISCUSSION .......................................................................................................................... 41
V.1 - FLORE DE POACEAE DE L’ISALO ................................................................................ 41
V.2 - PLAUSIBLE ORIGINE ANCIENNE DE LA VEGETATION DE L’ISALO .............................. 43
IV. CONCLUSION ..................................................................................................................... 46
V. BIBLIOGRAPHIE................................................................................................................. 48
ANNEXES ....................................................................................................................................... I

i
LISTE DES FIGURES
Figure 1: Situation géographique de l'Isalo .................................................................................................. 4
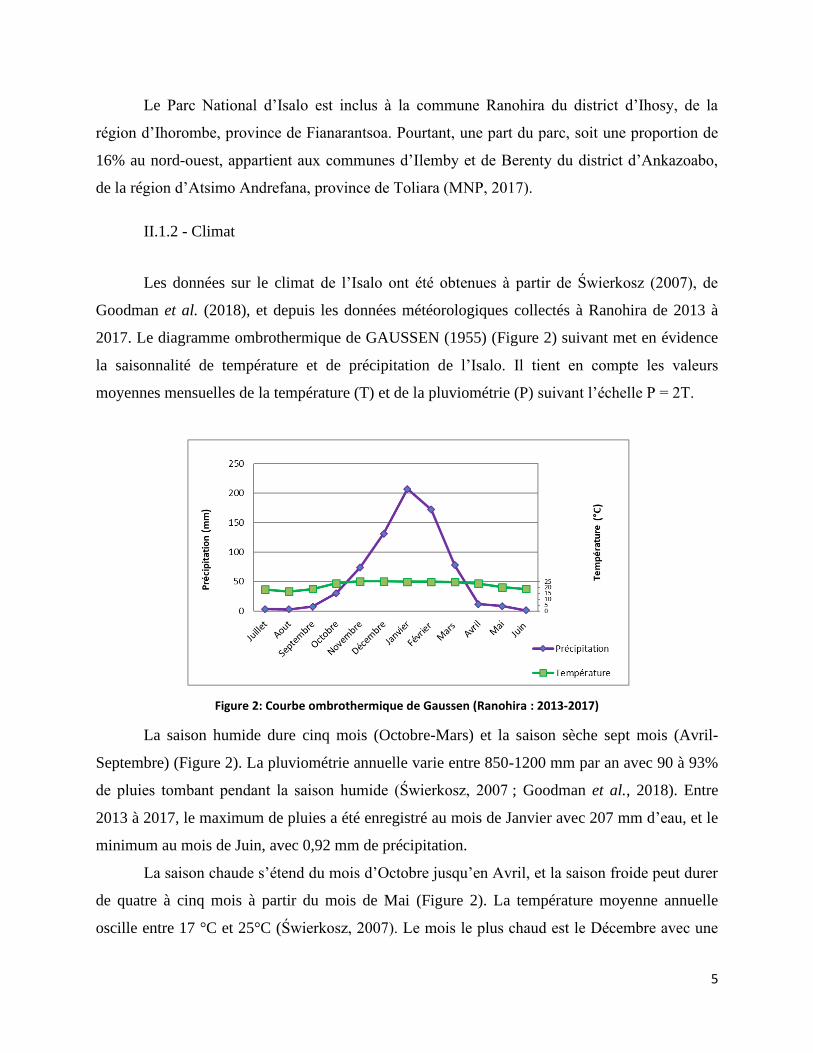
Figure 2: Courbe ombrothermique de Gaussen (Ranohira : 2013-2017) ..................................................... 5
Figure 3: Morphologie de l’inflorescence, de l’épillet, du fleuron et de la fleur chez les Poaceae ............ 13
Figure 4: Collectes à Sakamalio Isalo (A) et dissection des spécimens à TAN (B) ................................... 18
Figure 5: Extraits de photos des Poaceae de l’Isalo. ................................................................................... 22
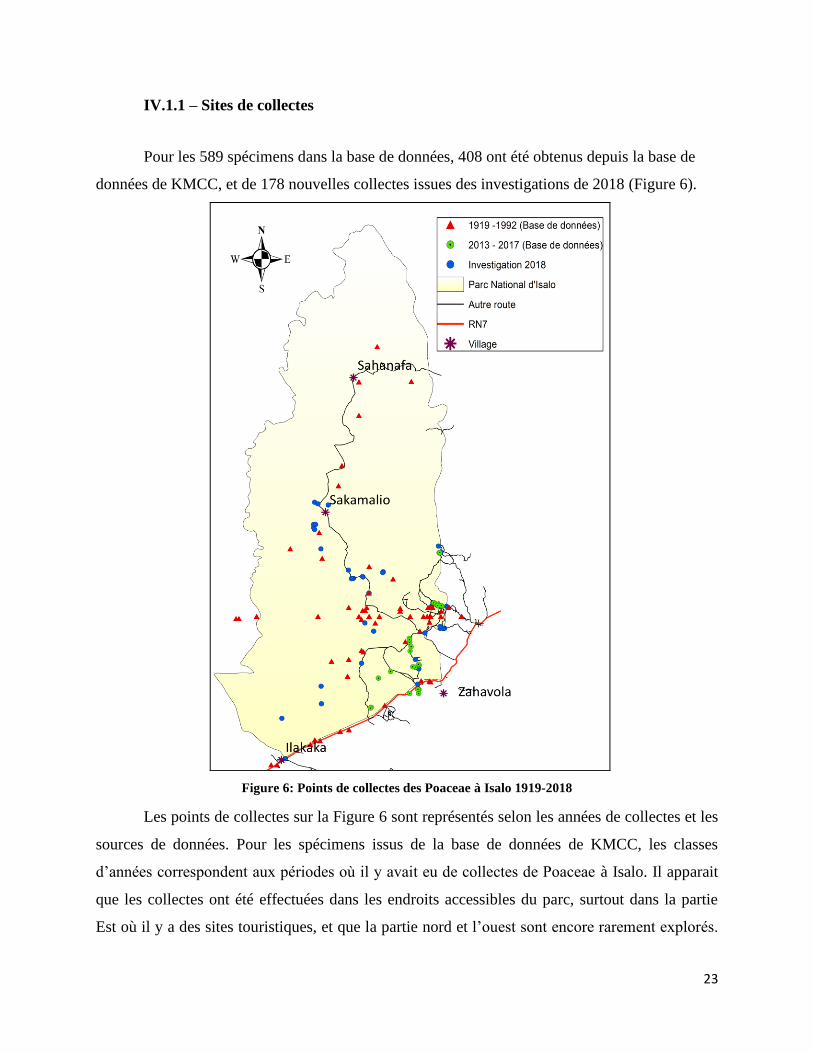
Figure 6: Points de collectes des Poaceae à Isalo 1919-2018 ..................................................................... 23
Figure 7: Herbiers de la nouvelle espèce de Bambou dans le Parc de l’Isalo ............................................ 26
Figure 8: Evolution sur la connaissance des Poaceae de l’Isalo ................................................................. 27
Figure 9: Proportion en espèces de Poaceae natives, endémiques et introduites présentes à Isalo ............. 28
Figure 10: Illustration de la triade d'épillets, caractéristiques du genre Tristachya .................................... 30
Figure 11: Points de collectes des Tristachya dans le parc de l’Isalo ......................................................... 30
Figure 12: Points de collectes de Styppeiochloa sp nov. ined dans le parc de l’Isalo ................................. 31
Figure 13: Points de collectes de Pseudolasiacis neoperrieri var. neoperrieri .......................................... 32
Figure 14: Zones potentielles de diversité en espèces de Poaceae dans le parc .......................................... 35
Figure 15: Tendance de distribution des espèces dans les habitats disponibles de l’Isalo .......................... 37
Figure 16: Richesse spécifique, espèces endémiques, introduites et natives par habitat ............................ 38
Figure 17: Espèces en C3 et en C4 par habitat ........................................................................................... 38
LISTE DES PHOTOS
Photo 1: Massif de l’Isalo vu depuis Korobe ................................................................................................ 6
Photo 2: Caractères de distinction des Poaceae .......................................................................................... 11
LISTE DES TABLEAUX
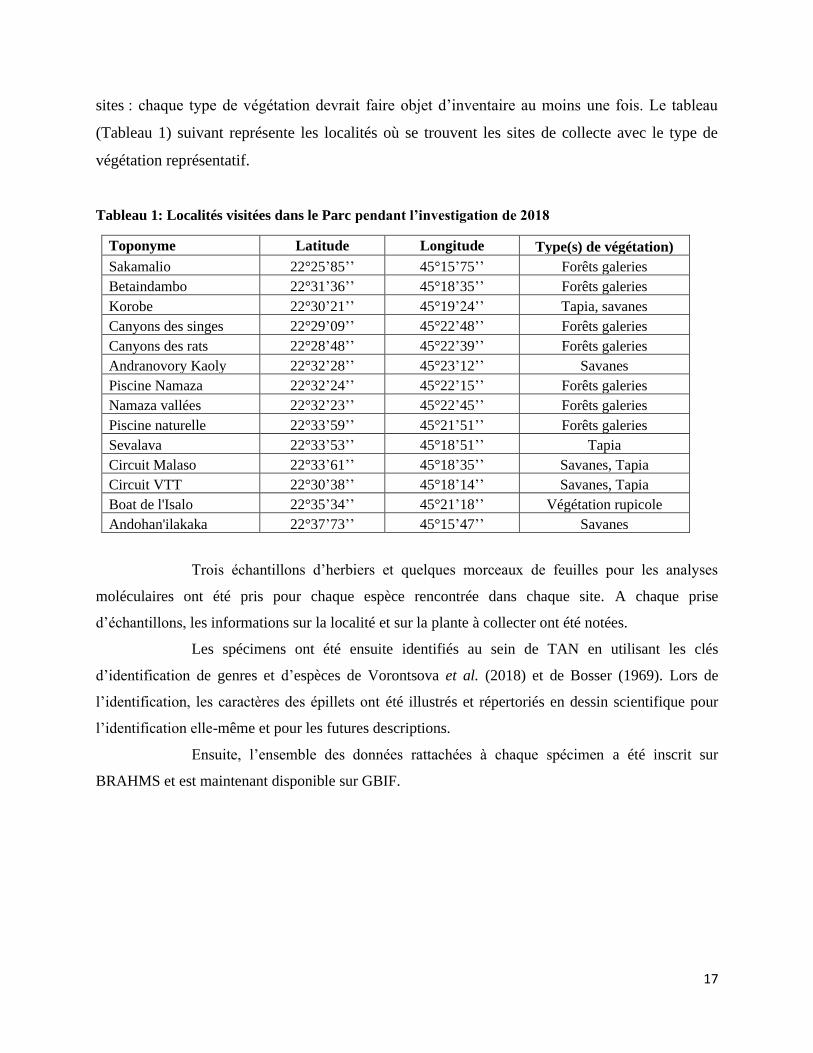
Tableau 1: Localités visitées dans le Parc pendant l’investigation de 2018 ............................................... 17
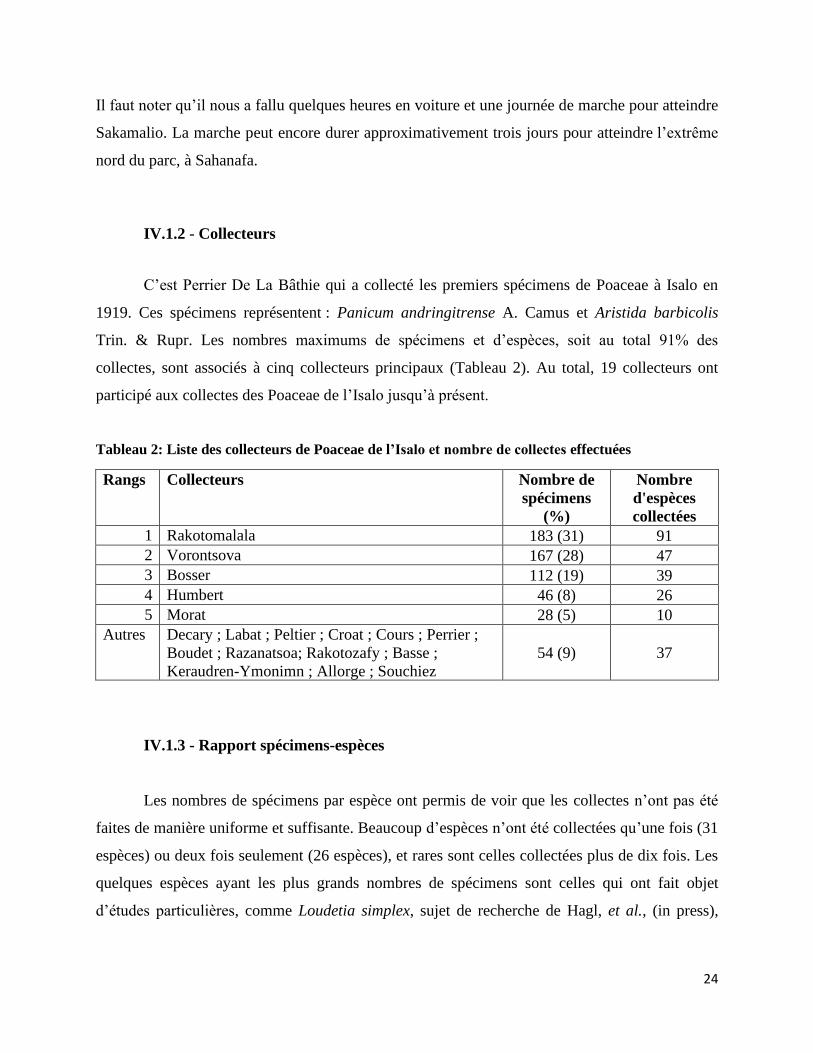
Tableau 2: Liste des collecteurs de Poaceae de l’Isalo et nombre de collectes effectuées ......................... 24

ii
Tableau 3: Affinité des Poaceae natifs rencontrés à Isalo à ceux des autres régions du monde ................. 28
Tableau 4: Clades et sous familles des Poaceae à Isalo par ordre de richesse en espèces, genres et tribus 33
Tableau 5: Liste et diversité des tribus des Poaceae à Isalo en fonction du nombre d’espèces et de genres
.................................................................................................................................................................... 33
Tableau 6: Liste et diversité spécifique des genres de Poaceae à Isalo en fonction du nombre d’espèce .. 34
LISTE DES ANNEXES
ANNEXE I: PHOTOS DES TYPES DE VEGETATIONS DE L’ISALO. ................................................... I
ANNEXE II: ETHNIES, ACTIVITES ECONOMIQUES ET RESSOURCES UTILISEES AUX
ENVIRONS DU PARC. ............................................................................................................................... II
ANNEXE III: LISTE DES ESPECES DE POACEAE RENCONTREES A ISALO ................................. III
ANNEXE IV: CLE DE DETERMINATION DES GENRES DE POACEAE DE L’ISALO ET
ILLUSTRATIONS .................................................................................................................................... XIII
GLOSSAIRE
Aile : partie aplatie et amincie prolongeant ou adhérant latéralement à certains organes. Les
carènes de certaines lemmas portent parfois des crêtes qui méritent cette appellation.
Arête : appendice filiforme, raide, droit, enroulé, ou coudé, naissant sur le dos ou à l'extrémité
d'un organe.
Aristé : désigne un organe muni d’arêtes.
Carène : désigne le plus souvent une nervure en relief sur un organe (feuilles, glumes,
glumelles), rappelant de façon plus ou moins prononcée la quille d'un bateau. Si elle est
nettement en relief, en lame aplatie, la carène est dite ailée.
Caryopse : désigne le fruit des graminées, le grain ; c'est un fruit sec, la graine étant soudée aux
enveloppes du fruit.
Cespiteux : en touffes denses et compactes, à chaumes dressés.

iii
Chartacé : ayant la consistance d'un papier fort.
Chaume : désigne les tiges florifères des graminées.
Contracté : se dit d'une inflorescence (panicule) dont les ramifications ont des pédoncules
relativement courts, ce qui donne à l'ensemble un aspect plus dense se rapprochant de l'épi.
Digité : qualifie une disposition des épis ou des racèmes groupés au sommet des chaumes et
divergeant, comme les doigts d'une main.
Dorsiventral : désigne des épis ou des racèmes dissymétriques, les épillets étant portés sur une
seule face de l'axe.
épi : inflorescence simple formée d'un axe portant des épillets sessiles.
Gaine : partie basale de la feuille naissant aux nœuds et entourant la tige.
Gazonnant : formant un gazon, c'est-à-dire se ramifiant dans toutes les directions au niveau du
sol et le couvrant.
Géminé : disposé par paire.
Genouillé : se dit d’une plante qui se fléchie (comme un genou) a la base avant de se redresser.
Gibbeux : présentant de profil une bosse nettement marquée.
Glumelles : bractées de la fleur. On distingue une glumelle inférieure ou lemma et une glumelle
supérieure ou paléa
Herbacé : parlant de glumes et de glumelles : de texture molle, ayant la consistance d'une
feuille, et de couleur verte.
Hyaline : de texture membraneuse, fine, transparente, non coloré en vert (les paléas sont souvent
hyalines).
Inféodé : qui est restreint à un endroit donné.
Lancéolé : en forme de fer de lance, la plus grande largeur étant au 1/3 inférieur, le rapport de la
largeur à longueur étant d'environ 1 à 3.
Ligule : membrane ou ligne de poils transversale marquant, sur la face supérieure, la séparation
entre le limbe et la gaine de la feuille chez les graminées.
Linéaire : long, étroit, à bords parallèles, qui est fréquent pour les feuilles de graminées.
Lodicules : petites pièces, peu visibles, au nombre de 2 ou 3 à la base de l'ovaire chez les
graminées, représentant des vestiges du périanthe.
Membraneux : de texture fine et mince mais non transparente par comparaison à hyaline.
Mutique : dépourvu d'arête.

iv
Nervures : faisceaux vasculaires souvent en relief sur les feuilles, les glumes, les glumelles.
Oblong : de forme allongée, à bords parallèles, à extrémités arrondies.
Ob- : préfixe signifiant le contraire, par exemple : obové, le contraire d'ové, organe de forme
ovée mais fixé par le pôle le plus étroit.
Ové : ayant la forme d'un œuf, donc à pôles arrondis l'un étant plus étroit.
Panicule : inflorescence ramifiée qui est une grappe composée irrégulière.
Plumeux : portants des poils disposés comme les barbes d'une plume.
Pseudo-pétiole : partie étroite de la base d'une feuille chez certaines graminées, simulant un
pétiole.
Racème (ou grappe) : inflorescence ou portion d'inflorescence formée d'un axe portant des
épillets pédicellés.
Rachéole : axe de l'épillet.
Rachis : axe d'une inflorescence ou d'une portion d'inflorescence.
Réfléchi : se dit d'un organe courbé et dirigé vers le bas faisant un angle plus ou moins aigu avec
l'axe principal.
Rhizome : tige souterraine portant des feuilles réduites à des écailles et s'enracinant aux noeuds.
Sessile : fixé directement sur l'axe sans l'intermédiaire d'un pédicelle ou d'un pétiole.
Soie : poil long et raide (poil sétacé),
Stérile : se dit d'une fleur dont l'ovaire absent ou avorté ne donne pas de fruit.
Stolon : tige couchée sur le sol, non souterraine, se ramifiant et s'enracinant aux noeuds.
Stolonifère : émettant des stolons
Stigmate : extrémité des styles sur lesquels se fixe le pollen ; chez les graminées, ils sont
plumeux.
Style : partie amincie du pistil, au sommet de l'ovaire.
Sub- : préfixe signifiant presque : (subégales : presque égales).
Triade : désigne des épillets groupés par trois.
Tuberculé : portant de petits mamelons arrondis.
Unisexué : fleur, épillet ou inflorescence où un seul sexe est représenté

I. INTRODUCTION

1
L’exploration de l’exceptionnelle biodiversité de Madagascar a débuté depuis le
XVIIème siècle et se trouve encore loin de son achèvement (Lowry et al., 2018 ; Goodman et
al., 2018). Suite aux efforts continuels de collectes et de révisions taxonomiques, plus de
nouvelles espèces sont décrites et le taux de la biodiversité augmente avec. En prenant le cas
des plantes malgaches, entre 60 et 90 espèces nouvelles à la science sont décrites chaque année
(Goodman et al., 2018). En 2017, Madagascar Catalogue (2017) (http1) reconnait 249 familles,
1698 genres et 11 649 espèces de plantes vasculaires natives présentes à Madagascar. Avec le
rythme de découverte au cours des trois dernières décennies, la flore malgache devrait compter
environ 14 000 espèces selon Phillipson et al. (2017). Cela est d’ailleurs fort possible car seule
72,4 % des collections de plantes à Madagascar sont identifiées jusqu’au niveau spécifique
(Goodman et al.,2018). De plus certains taxa, exceptionnellement riches en espèces, cas de
plusieurs familles de monocotylédones, sont encore mal inventoriés à Madagascar. C’en est le
cas de la famille des Poaceae, qui fait sujet de la présente étude.
La famille des Poaceae représente la cinquième famille d’Angiospermes la plus riche en
espèces dans le monde, comptant environ 12 000 espèces (Kellogg, 2015 ; Soreng et al., 2017).
A Madagascar, elle se trouve à la quatrième place après la famille des Orchidaceae (902 spp.),
des Rubiaceae (767 spp.), et celle des Fabaceae (592 spp.) (Goodman et al., 2018). Elle y compte
provisoirement 541 espèces avec un taux d’endémisme estimé à 40% (Vorontsova et al., 2016).
Les espèces de Poaceae constituent principalement les écosystèmes herbeux malgaches qui
couvrent environ 65% de la superficie totale de Madagascar (Moat et Smith, 2007). Elles sont
sources de nourritures primordiales pour les Malgaches en produisant les céréales, comme le riz
(Oryza sativa L.), le maïs (Zea mays L.), le blé (Triticum aestivum L.), l’orge (Hordeum vulgare
L.), l’avoine (Avena sativa L.), et le Sorgho (Sorghum sp.), et en fournissant des forages pour les
bétails.
Malgré la présence significative de la famille des Poaceae à Madagascar, il s’y trouve
qu’elle est encore mal étudiée. La difficulté de distinguer les espèces de Poaceae entre elles, à
cause de leurs complexes et minuscules pièces florales, ainsi que le manque de matériels
adéquats pour les identifier, auraient découragés beaucoup de chercheurs à les étudier
systématiquement. De même, en étant cosmopolites, les Poaceae sont souvent considérés comme
des mauvais herbes et d’aucun intérêt. Les recherches faites sur cette famille sont encore rares et
la plupart se portent sur des espèces d’importances économiques uniquement (par exemple :

2
Rakotonjanahary, 1993 ; Raharimanana, 2007 ; Rabealaina, 2010). L’ouvrage de Bosser (1969) :
‘‘Graminées des pâturages et de culture’’ a été longtemps la seule référence sur la diversité des
Poaceae malgaches, mais encore, il ne décrit que la moitié des espèces sensées se trouver à
Madagascar, et son application se limite dans le domaine des hauts plateaux malgaches. Ce
manque d’intérêt est pourtant en contraste absolu avec l’évolution des recherches sur ce groupe
dans les autres pays, tenant compte que c’est l’une des familles de plantes les plus étudiées
globalement. De nombreux pays ont déjà publié leur flore de Poaceae, tels que l’Afrique de l’est
(Clayton, 1970 ; Clayton & Renvoize 1982 ; Clayton et al., 1974), ou l’Amérique du centre
(Davidse et al., 1994) etc. La flore en ligne des Poaceae a été même créée récemment et est
maintenant disponible sur le site du Royal Botanic Gardens Kew (RBG, Kew) (http2). Quant à
Madagascar, la description des espèces de Poaceae, largement effectuée par Aimée Camus, ainsi
que les collectes de spécimens semblent s’arrêter en 1970 (Vorontsova et al., 2018).
En effet, la connaissance sur la diversité des Poaceae malgaches est encore pauvre et
archaïque. Cependant, elle est particulièrement indispensable dans la détermination de l’origine
des écosystèmes herbeux malgaches, qui fait pourtant sujet d’un grand débat depuis très
longtemps jusqu’à maintenant. Ces écosystèmes ont toujours été considérés comme des
formations secondaires, ayant substituées les végétations primaires dans les zones à saisonnalité
marquée, après que l’Homme ait débarqué sur l’Ile (Perrier de la Bâthie, 1921 ; Humbert, 1927 ;
Koechlin, Guillaumet & Morat, 1974 ; Lowry et al., 1997). Toutefois, selon les récentes
découvertes, il se peut qu’une part de ces écosystèmes existait à Madagascar bien avant l’arrivée
de l’Homme, et soit alors naturelle. Les pollens et les dépôts de charbons pré-anthropiques que
l’on a découverts récemment indiquent la présence du feu et d’habitats ouverts précédant
l’arrivée de l’Homme (Burney,1987b, 1997 ; Gasse & Van Campo, 1998). Cela a été appuyé par
la présence de plantes et d’animaux endémiques dans les écosystèmes herbeux malgaches, ainsi
que d’une biodiversité plus riche que l’on croyait auparavant (Bond et al.,2008 ; Vorontsova et
al.,2016).
Tenant compte de cette lacune de connaissance au niveau de la diversité des Poaceae
malgaches, il apparaît indispensable de mener d’avantages d’inventaires de Poaceae dans les
régions et stations malgaches, peu ou pas encore explorées. La présente recherche se réalise ainsi
dans le Parc National l’Isalo, une zone potentielle pour l’étude des Poaceae par son paysage

3
principalement constitué de savanes. Des recherches portant sur l’écologie des savanes et les
traits fonctionnels des espèces de Poaceae dans les différents biomes ont été antérieurement
effectuées aux environs du sud-ouest et des hauts plateaux de Madagascar incluant l’Isalo
(Morat, 1973 ; Solofondranohatra et al., 2018). Mais bien que ces études aient concerné les
Poaceae et l’Isalo, la diversité exacte de cette famille y reste encore inconnue. Dans le cadre du
projet « Grasses and Grasslands of Madagascar » mené par RBG Kew et Kew Madagascar
Conservation Centre (KMCC), cette recherche a pour principal objectif de documenter les
Poaceae dans le Parc National d’Isalo. Pour l’atteindre, les objectifs spécifiques sont de :
➢ Inventorier les espèces de Poaceae dans tous les habitats disponibles de l’Isalo ;
➢ Etablir la liste des Poaceae de l’Isalo ;
➢ Quantifier et analyser la diversité et l’endémicité des Poaceae rencontrés dans le parc ;
➢ Discuter de l’origine des savanes de l’Isalo à partir des données disponibles.
Pour cette étude, les hypothèses suivantes ont été émises :
➢ Des espèces de Poaceae nouvelles à la science ou pour Isalo, y seraient potentiellement
collectées ;
➢ Les savanes de l’Isalo auraient une plausible origine ancienne.
Le présent travail se divise en quatre parties. La première présente les caractéristiques du milieu
d’étude. La seconde décrit le matériel d’étude et la méthodologie adoptée. La troisième présente
les résultats et leurs interprétations, lesquelles seront discutées dans la quatrième partie.

I. MILIEU D’ÉTUDE

4
II.1 -MILIEU PHYSIQUE
II.1.1 - Localisation et rattachement administratif
Le présent travail a été réalisé dans l’une des plus anciennes aires protégées de
Madagascar, le Parc National d’Isalo, créé en 1962. Il représente la quatrième plus grande parcs
nationaux malgaches avec une superficie de 81 540 Ha (Madagascar National Parks, 2017).
Isalo se situe entre 22°13’ et 22°45’ de latitude Sud et entre 45°13’ et 45°29’ de longitude Est, et
se trouve à 700 km de la capitale Antananarivo, le long de la Route Nationale numéro 7 (RN7)
menant vers Toliara (Figure 1).
Figure 1: Situation géographique de l'Isalo
5
Le Parc National d’Isalo est inclus à la commune Ranohira du district d’Ihosy, de la
région d’Ihorombe, province de Fianarantsoa. Pourtant, une part du parc, soit une proportion de
16% au nord-ouest, appartient aux communes d’Ilemby et de Berenty du district d’Ankazoabo,
de la région d’Atsimo Andrefana, province de Toliara (MNP, 2017).
II.1.2 - Climat
Les données sur le climat de l’Isalo ont été obtenues à partir de Świerkosz (2007), de
Goodman et al. (2018), et depuis les données météorologiques collectés à Ranohira de 2013 à
2017. Le diagramme ombrothermique de GAUSSEN (1955) (Figure 2) suivant met en évidence
la saisonnalité de température et de précipitation de l’Isalo. Il tient en compte les valeurs
moyennes mensuelles de la température (T) et de la pluviométrie (P) suivant l’échelle P = 2T.
Figure 2: Courbe ombrothermique de Gaussen (Ranohira : 2013-2017)
La saison humide dure cinq mois (Octobre-Mars) et la saison sèche sept mois (Avril-
Septembre) (Figure 2). La pluviométrie annuelle varie entre 850-1200 mm par an avec 90 à 93%
de pluies tombant pendant la saison humide (Świerkosz, 2007 ; Goodman et al., 2018). Entre
2013 à 2017, le maximum de pluies a été enregistré au mois de Janvier avec 207 mm d’eau, et le
minimum au mois de Juin, avec 0,92 mm de précipitation.
La saison chaude s’étend du mois d’Octobre jusqu’en Avril, et la saison froide peut durer
de quatre à cinq mois à partir du mois de Mai (Figure 2). La température moyenne annuelle
oscille entre 17 °C et 25°C (Świerkosz, 2007). Le mois le plus chaud est le Décembre avec une

6
température moyenne de 30,6°C, et le plus froid le mois d’Aout avec une température moyenne
de 10,2 °C. La température peut encore descendre jusqu’à 7,3°C pendant ce mois le plus froid
(Goodman et al., 2018).
Avec une saison chaude plus longue et une moyenne de précipitation annuelle comprise
entre 950 à 1500 mm, le bioclimat que subit Isalo est de type subhumide tempéré (Koechlin et
al., 1974).
II.1.3 - Géomorphologie et topographie
L’Isalo est un grand massif de grés siliceux, une roche sédimentaire de faible dureté, de
coloration rouille et ocre. Ces grès ont résisté à l’érosion par leur porosité et par la présence
d’armature siliceuse et d’horizon ferrugineux, d’où l’aspect ruiniforme et déchiqueté des reliefs.
Ces hauts reliefs sont très découpés de 20 à 30 kilomètres de largeur et apparaissent dans
l’ensemble comme une alternance de vaste plaine, de côtes et de falaises. Par l’effet de la
tectonique, certaines falaises, que l’on appelle canyons, atteignent jusqu’à 200 m de profondeur
(Morat, 1973 ; Świerkosz, 2007). Les caractéristiques géomorphologiques du massif de l’Isalo
confèrent la particularité et l’unicité de son paysage (Photo 1). L’altitude varie de 541 – 1268 m.
Photo 1: Massif de l'Isalo vu depuis Korobe

7
II.2 - MILIEU BIOLOGIQUE
II.2.1 - Flore et végétation
Selon les divisions phytogéographiques d’Humbert (1955), Isalo appartient aux secteurs
des pentes occidentales, du domaine du Centre, de la région orientale malgache.
La flore et la végétation de l’Isalo est encore peu étudiée et les descriptions suivantes sont
obtenues de Morat (1973), de Świerkosz (2007), et de Goodman et al., 2018). Un total de 101
familles, dont trois endémiques de Madagascar, et 431 espèces de plantes se rencontrent à Isalo
(Goodman et al., 2018). Les espèces endémiques comptent 227 espèces (70,7%), dont 13 espèces
ligneuses sont restreintes au massif de l’Isalo. On y rencontre à la fois de la végétation primaire
(bois de Tapia, forêts galeries, végétations rupicoles) et de la végétation modifiée (savanes, forêts
et fourrées secondaires). Quelques photos de ces types d’habitats sont représentées dans
l’Annexe I.
II.2.1.1 - Bois de Tapia
Le bois de Tapia ou forêts sclérophylles de moyenne altitude se rencontrent dans
le tiers ouest et dans le Nord-Est de l’aire protégée. Cette formation occupe 30,87% du Parc. Sa
physiognomie est caractérisée par une strate arborée ouverte à 50% et un sous-bois plus sec de
type éricoïde. Les arbres sont en boule, à troncs tortueux et au feuillage terne. Les taxa
caractéristiques sont Uapaca bojeri (Phyllantaceae) et quelques espèces de la famille des
Sarcolaenaceae (Humbert, 1955).
II.2.1.2 - Forêts galeries
Les forets galeries regroupent les formations forestières tributaires de la présence
d’un cours d’eau à proximité plus ou moins immédiate (Morat, 1937). On y distingue :
- les forêts ripicoles des canyons occupent 2,18% du parc, et se rencontrent à
l’ouest, au centre, et surtout à l’est du parc (MNP, 2017). Cette formation se trouve dans un
environnement ombragé à l’abris du feu et bénéficie d’une certaine humidité édaphique. Elles

8
sont représentées par Ravenea glauca (Arecaceae), Breaunidia salcinia (Rubiaceae) et
Weinmania stenostachya (Cunoniaceae).
- les forêts ripicoles des vallons peu profonds et des ruisseaux temporaires sont
dominées par Pandanus namakiensis (Pandanaceae) et quelques espèces d’arbres. Un exemple
pertinent de cette formation se trouve auprès de la piscine naturelle de l’Isalo.
- les forêts humides semi-décidues occupent 1,95% du parc et colonisent les trois
vallées du centre au nord du parc, là où trois principales rivières circulent : Sahanafa,
Ankademoky et Sakamalio. La composition arbustive de cette formation est similaire à celle des
forêts ripicoles des canyons mais elle abrite, en plus, des espèces d’arbres introduites ou
spontanément naturalisées, comme : Melia azedarach L. (Meliaceae), Mangifera indica L.
(Anacardiaceae) et Eugenia sp. (Myrtaceae).
II.2.1.3 - Végétation rupicole
Une importante végétation rupicole pousse sur les affleurements rocheux des
falaises de l’Isalo, plus ou moins à l’abris du feu. Elle est caractérisée par des espèces
xérophytiques de la famille des Apocynanceae (Cynanchum spp., Pachypodium spp.), des
Euphorbiaceae (Euphorbia spp.), et des Asphodelaceae (Aloe spp.).
II.2.1.4 - Savanes et prairies
Les savanes couvrent plus de 40% de la superficie du Parc. Cette formation est
constituée par des espèces associées au régime du feu, surtout de la famille des Poaceae, qui
empêchent la prolifération des arbres. Les Poaceae qui y sont dominant sont : Loudetia simplex
(Nees) C.E.Hubb., Loudetia filifolia Schweick., Trachypogon spicatus Kuntze, Chrysopogon
serrulatus Trin.. Des ligneux y sont parfois présents, dont plusieurs sont endémiques, tels que
Menabea venenata Baill. et Secamone tenuifolia Decne. (Apocynaceae), Tephrosia cf.
isaloensis. Du Puy & Labat (Fabaceae), Polygala isaloensis H.Perrier (Polygalaceae), et
Monotes madagascariensis Humbert (Dipterocarpaceae).

9
II.2.1.5 - Forêts et fourrées secondaires
Ces formations se rencontrent sur les pentes externes du massif en contact avec
les cultures. Leur composition floristique est similaire à celle des formations secondaires du
centre de Madagascar. Elles sont constituées surtout par une communauté de Vangueria
madagascariensis J.F.Gmel. (Rubiaceae), Aphloia theiformis Benn. (Aphloiaceae) et de
Tamarindus indica L. (Fabaceae).
II.2.1.6 - Marais et marécage
Des marais et marécages se rencontrent sporadiquement dans le parc, surtout dans
la partie sud. Ils sont composés par des espèces de Poaceae, dont Phragmites mauritianus Kunth
est le plus représentatif, et de Cyperaceae.
II.2.2 - Faune
Le parc abrite plusieurs espèces de vertébrés incluant des espèces endémiques restreintes
à Isalo, dont quatre amphibiens (Gephyromantis corvus, Mantella expectata, Mantidactylus
narolattae, Scaphiophryne gottlebei), un reptile (Trachylepis nancycoutuae), et un rongeur
(Eliurus danieli). Au total, on y rencontre 94 espèces d’oiseaux, 22 espèces d’amphibiens, 46
espèces de reptiles, 04 espèces de tenrecidés, 01 espèce de musaraignes (Sorididae), 02 espèces
de carnivores, 15 espèces de chauvesouris, et 07 espèces de Lémuriens (Goodman et al., 2018).
II.2.3 - L’Homme et ses activités
Les Bara étaient, jadis, les seuls ayant habité l’Isalo. Depuis 1845 que d’autres ethnies
venaient s’y installer aussi, mais les Bara restent les plus important en effectif (Deschamps,
1965). Le royaume Bara fut établi aux canyons de Ranohira, à la limite du parc du côté est, et le
reste de l’Isalo a été presque pratiquement vide d’habitants : le massif servait plutôt de lieu
d’enterrement et de pâturage (Rajaofera, 2007 ; Zafisoalizy, 2011).

10
Actuellement, le parc de l’Isalo est entouré par 39 villages à l’ouest, à l’est et au nord,
supportant un total de 16 382 habitants (MNP, 2017). Les différentes ethnies avec leurs activités
économiques qui ont une implication directe ou indirecte au Parc National d’Isalo et ses
environs, sont représentées dans l’Annexe II.

II. MATERIELS ET
METHODES

11
III. 1 - MATERIELS D’ETUDES La famille des Poaceae ou Gramineae constitue le matériel végétal dans le cadre de cette
étude. Les caractères morphologiques et la taxonomie des Poaceae à Madagascar, ainsi que leur
base de données actuelle seront mis en évidence dans cette partie.
III.1.1 - Caractères morphologiques
Les Poaceae appartiennent au groupe des Monocotylédones, caractérisés par un embryon
à un seul cotylédon et par des pièces florales en trimère chacune. Ils se reconnaissent directement
par la morphologie uniforme de leurs racines, tiges et feuilles (Photo 2). Les Cyperaceae peuvent
être confondus avec les Poaceae, mais chez eux, les tiges sont anguleuses au touché et sont
solides, et les ligules font défauts au niveau de leurs feuilles. Il en est de même pour les
Juncaceae, mais leur port est dur et de couleur plus foncée que celle de leurs pièces forales, et
leur feuilles dépourvues de ligule.
Photo 2: Caractères de distinction des Poaceae
III.1.1.1 - Tige
Les Poaceae sont généralement des plantes herbacées de petite taille, à part les
bambous ligneux et larges. La tige est le plus souvent à section arrondie et, est constituée par une
succession de nœuds solides séparés par des segments, longs ou courts, appelés entrenœuds. Les
CHAMONT S. (INRA)

12
tiges aériennes, dressées (chaumes) ou couchées (stolons), portent des feuilles au niveau des
nœuds, alors que les tiges souterraines (rhizomes) en sont dépourvues.
III.1.1.2 - Feuilles
Les feuilles naissent successivement au niveau des nœuds en deux rangs alternes,
et sont composées de trois principales parties : une gaine tubulaire qui entoure le chaume ; un
limbe vert qui peut être étroit ou allongé, avec des veines ou nervures parallèles ; et une ligule
qui peut être une membrane fine ou une ligne de poils au niveau de la jonction interne (ou
rarement à l’externe) entre la gaine et le limbe.
III.1.1.3 - Inflorescence
Le port des Poaceae se terminent en une inflorescence, ou parfois appelée
synflorescence, lors du stade reproductif. Chaque inflorescence est constituée par des petits
éléments ressemblant à des petites fleurs, appelés épillets. Ils s’arrangent directement sur le
rachis, axe principal de l’inflorescence, ou sur les ramifications secondaires du rachis. Cet
arrangement peut prendre la forme d’un épi, de racèmes ou de panicules selon les groupes. La
Figure 3 illustre les types d’inflorescences et les détails de chaque partie à partir de l’épillet.
III.1.1.4 - Epillets
Un épillet est constitué par un ou plusieurs fleuron(s), surmonté(s) par deux
bractées vides appelés glume inferieure et glume supérieure. Ces éléments s’arrangent en
alternance le long de la rachéole, axe principal d’un épillet. Selon le nombre des fleurons, un
épillet peut être uniflore, biflore, ou pluriflore. Un épillet peut être soutenu par un pédicelle ou
être sessile.

13
Figure 3: Morphologie de l’inflorescence, de l‘épillet, du fleuron et de la fleur chez les Poaceae
III.1.1. 5 - Fleuron et fleur
La fleur des Poaceae est enveloppée par des bractéoles ou glumelles vides appelés
lemme ou lemma (l’inferieur) et paléa ou paléole (le supérieur). L’ensemble [lemma + paléa +
fleur] donne ce qu’on appelle : fleuron.
La fleur des Poaceae ont des structures similaires aux fleurs ordinaires mais
réduites et modifiées. Le périanthe est représenté par deux ou trois lodicules insérés sur le
pédoncule floral.

14
Ce sont des pièces charnues qui gonflent pour assurer l’ouverture des glumellules lors de la
période de floraison. Les étamines sont souvent au nombre de trois, mais quelquefois au nombre
de six. Chaque étamine est constituée d’un filament fragile et d’une large anthère. Quant au
pistil, il comprend un seul ovule surmonté par deux styles séparés se terminant chacun en un
stigmate plumeux.
III.1.1.6 - Caryopse
Le fruit des Poaceae est un fruit sec à une seule graine, appelée caryopse. Il est
caractérisé par un endosperme féculant, un embryon latéral et un péricarpe fusionnant avec la
graine. La structure, la texture, la taille et la position de l’embryon sont aussi utiles dans
l’identification de l’espèce.
III.1.2 - Position taxonomique des Poaceae et leur taxonomie à Madagascar
La famille des Poaceae appartiennent à l’Ordre des Poales (Grass Phylogeny Working
Group I, GPWG, 2001). La dernière classification de la famille des Poaceae validée par
Angiosperm Phylogeny Group III en 2009 est la suivante :
Règne : PLANTAE
Embranchement : MAGNOLIOPHYTA
Sous-embranchement : ANGIOSPERMES
Classe : LILIOPSIDA
Sous-classe : COMMELINIDAE
Ordre : POALES
Famille : POACEAE
La famille comprend 12 sous-familles, 52 tribus, 768 genres et environ 12.000 espèces
(GPWPII, 2012 ; Kellogg, 2015 ; Soreng et al., 2015 ; Soreng et al., 2017). On en distingue trois
groupes de sous-familles, tels qu’un groupe de trois sous-familles ancestrales (Anomochloideae,
Pharoideae, Puelioideae), un autre groupe de trois sous-familles rassemblées dans le clade BEP
(Bambusoideae, Ehrhartoideae, Pooideae), et un groupe de six sous-familles dans le clade
PACMAD (Panicoideae, Aristidoideae, Chloridoideae, Micrairoideae, Arundinoideae,
Danthonioideae).
A Madagascar, les trois groupes de sous-familles sont tous présents. Toutefois, les deux
sous-familles ancestrales : Anomochloideae et Puelioideae y font défaut, et la sous-famille :

15
Pharoideae est seulement représentée par l’espèce Leptapsis zeylanica. En tout, la famille des
Poaceae à Madagascar compte dix sous-familles, 24 tribus, 144 genres et 541 espèces
(Vorontsova et al., 2018).
Les connaissances sur les Poaceae de Madagascar ne cessent d’évoluer. On a récemment
publié : une mise à jour sur les genres de Poaceae endémiques de Madagascar (Vorontsova &
Rakotoarisoa, 2014), une checklist des Poaceae d’Itremo (Nanjarisoa et al.,2017), cinq nouvelles
espèces décrites (Vorontsova et al.,2013 ; Vorontsova 2014 ; Vorontsova et al.,2015 ;
Vorontsova 2017), un guide d’identification des genres de Poaceae à Madagascar (Vorontsova et
al.,2018), ainsi qu’un Atlas des Poaceae à Madagascar (Rabarivola et al., 2019d).
III.1.2 - Base de données sur les Poaceae
La base de données sur les Poaceae de Madagascar a été créé par KMCC depuis 2018.
Elle compile les données sur tous les spécimens de Poaceae collectés depuis le XVIIIe siècle
jusqu’à cette date. Elle englobe environ 12 000 collections de Poaceae à l’issue des collections
dans les Herbaria de Tsimbazaza Antananarivo Madagascar ou TAN, et de Museum National
d’Histoire Naturelle ou MNHN Paris France, et aussi des collectes récentes faites par Maria
Vorontsova (RBG, Kew) et KMCC.
Les informations sur chaque spécimen sont enregistrées et conservées dans un logiciel
Botanical Research And Herbarium Management System ou BRAHMS (http3). Ainsi, pour
chaque échantillon (herbier, tubercule, graine, etc.), les renseignements sur la localité, le(s)
collecteur(s), le numéro de collecte, la date de collecte, le déterminateur, la date de
détermination, l’altitude, l’habitat et d’autres notes concernant le spécimen et l’Herbarium y sont
transcrites. Les données inscrites par KMCC dans BRAHMS sont disponibles et téléchargeables
sur le site de Global Biodiversity Information Facility ou GBIF (http4).
III.2 - METHODES
III.2.1 - Etude préliminaire
III.2.1.1 - Recherche bibliographique
Pour toute étude, la revue bibliographique sur le milieu et le(s) matériel(s) passe
toujours au préalable. De ce fait, les connaissances actuelles sur les Poaceae de l’Isalo ont été

16
synthétisées depuis Bosser (1969), Morat (1973), Świerkosz (2007), Vorontsova et al., 2016), et
de Solofondranohatra et al., 2018).
III.2.1.2 - Apprentissage de l’identification des Poaceae
L’identification des Poaceae consiste d’abord à voir un à un les caractéristiques de
chaque partie de la plante, allant du port visible aux minutieux paléas, lemmas et fleurs. Il est
indispensable de disséquer ces minuscules caractères et de les observer sous une loupe
binoculaire ou microscope. Un ou plusieurs guides d’identification des taxa devrait être
ultérieurement suivi(s) suivant les caractères observés du spécimen pour aboutir au nom de
l’espèce. Quand le nom a été déterminé, il faut comparer le spécimen aux herbiers de référence
correspondant au nom de l’espèce indiqué. Afin d’acquérir l’habileté dans la manipulation et
dans la dissection des spécimens, ainsi que pour maitriser les terminologies de description
propres aux Poaceae, des exercices d’identification ont été faites à TAN au préalable.
III.2.2 - Collectes de données
III.2.2.1 - Téléchargement et vérification des données existantes
Les données sur les précédentes collections de Poaceae de l’Isalo ont été extraites
depuis la base de données des Poaceae de Madagascar mentionnée precedemment. Les noms de
la localité : ‘Isalo’ ou ‘Ranohira’ ont été utilisés comme filtre. Afin d’éviter la répétition d’un
même spécimen dans l’extrait de données pour l’Isalo, les échantillons ont été triés par le nom du
collecteur, le numéro de collecte outre la localité, pour pouvoir supprimer les doublons. Un
numéro ne doit représenter qu’un seul spécimen.
III.2.2.2 - Collecte de données sur terrain, identification et transcription des
données sur BRAHMS
Le présent travail relève le défi de mener des inventaires dans les endroits
inexplorés du Parc, mais aussi de faire des inventaires plus complets dans ceux visités par les
botanistes précédents. Une investigation de 15 jours a été réalisée dans le Parc National d’Isalo
entre Mars et Avril 2018, période incluse dans la saison de floraison des Poaceae. Des collectes
non sélectives ont été effectuées dans les différents sites accessibles du Parc, du plus près au plus
loin. Outre les conditions d’accessibilité, le type de végétation a aussi influencé le choix des
17
sites : chaque type de végétation devrait faire objet d’inventaire au moins une fois. Le tableau
(Tableau 1) suivant représente les localités où se trouvent les sites de collecte avec le type de
végétation représentatif.
Tableau 1: Localités visitées dans le Parc pendant l’investigation de 2018
Toponyme Latitude Longitude Type(s) de végétation)
Sakamalio 22°25’85’’ 45°15’75’’ Forêts galeries
Betaindambo 22°31’36’’ 45°18’35’’ Forêts galeries
Korobe 22°30’21’’ 45°19’24’’ Tapia, savanes
Canyons des singes 22°29’09’’ 45°22’48’’ Forêts galeries
Canyons des rats 22°28’48’’ 45°22’39’’ Forêts galeries
Andranovory Kaoly 22°32’28’’ 45°23’12’’ Savanes
Piscine Namaza 22°32’24’’ 45°22’15’’ Forêts galeries
Namaza vallées 22°32’23’’ 45°22’45’’ Forêts galeries
Piscine naturelle 22°33’59’’ 45°21’51’’ Forêts galeries
Sevalava 22°33’53’’ 45°18’51’’ Tapia
Circuit Malaso 22°33’61’’ 45°18’35’’ Savanes, Tapia
Circuit VTT 22°30’38’’ 45°18’14’’ Savanes, Tapia
Boat de l'Isalo 22°35’34’’ 45°21’18’’ Végétation rupicole
Andohan'ilakaka 22°37’73’’ 45°15’47’’ Savanes
Trois échantillons d’herbiers et quelques morceaux de feuilles pour les analyses
moléculaires ont été pris pour chaque espèce rencontrée dans chaque site. A chaque prise
d’échantillons, les informations sur la localité et sur la plante à collecter ont été notées.
Les spécimens ont été ensuite identifiés au sein de TAN en utilisant les clés
d’identification de genres et d’espèces de Vorontsova et al. (2018) et de Bosser (1969). Lors de
l’identification, les caractères des épillets ont été illustrés et répertoriés en dessin scientifique pour
l’identification elle-même et pour les futures descriptions.
Ensuite, l’ensemble des données rattachées à chaque spécimen a été inscrit sur
BRAHMS et est maintenant disponible sur GBIF.

18
Figure 4: Collectes à Sakamalio Isalo (A) et dissection des spécimens à TAN (B)
III.2.3 - Compilation des données et listage des espèces
Afin d’avoir la liste des espèces de l’Isalo, les données extraites de la base de données de
KMCC et les données issues de notre investigation ont été compilées en une seule base de
données sur BRAHMS.
La compilation des données a montré que certaines espèces sont citées par leur synonyme
mais non par leur nom accepté actuel. Afin d’éviter qu’une même espèce porte deux noms
différents dans la base de données, le site Plants of The World Online de Kew Science (http5) a
été consultée pour vérifier le nom correct et à jour de chaque espèce.
Cette étude concerne le Parc National d’Isalo, ainsi seules les collectes effectuées dans la
limite du parc ont été considérées. Pour ce faire, les coordonnées géographiques rattachées à
chaque spécimen ont été projetées sur une carte montrant la limite du parc, à l’aide du logiciel
ArcGis 10.5. Les collectes qui sont en dehors et très éloignées de la limite du parc ont été
supprimées.
Après ces vérifications, les données sur tous les spécimens ont été enregistrées sous
format Ms Excel afin d’établir la liste des espèces. Les affinités biogéographiques [de Bosser
(1969) et de Rabarivola et al., 2019d)], et le type de photosynthèse (de Soreng et al., 2017) de
chaque espèce ont été ajoutées à cette liste.
Rabehevitra, 2018 Miharivola, 2019

19
III.2.4 - Calcul du nombre cumulé de collectes et d’espèces
Cette partie a pour but de présenter l’évolution de la connaissance des Poaceae de l’Isalo
suivant le temps. Le nombre de collectes (spécimens) faites pour chaque espèce et tous les ans a
été compté. Ensuite, ces nombres ont été ajoutés entre eux de façon cumulative toutes les
décennies. Le nombre de collectes de la première décennie sera ajouté à celui de la décennie
suivante, et ainsi de suite jusqu’à la dernière date où l’on ait fait de la collecte de Poaceae à
Isalo. Il a été fait de même pour les espèces. Ensuite, à l’aide du logiciel R, les courbes
d’accumulation des collectes et des espèces suivant le temps ont été tracées sur un même plan.
Cela afin de voir dans l’ensemble le rythme de collecte et de découverte d’espèces, ainsi les
interprétations à prévoir sont les suivantes :
➢ La courbe pour les collectes et les espèces devraient augmenter ensemble vers le début et
pendant un certain temps ;
➢ La courbe de l’espèce n’augmentera plus avec celle des collectes quand il n’y a plus de
nouvelles espèces à collecter dans le site, et dans ce cas, la liste des espèces proposée serait
quasi-complète.
III.2.5 - Aire de distributions des espèces
Dans une étude taxonomique, il est pratique d’illustrer l’aire de distribution des espèces à
l’aide d’une carte de distribution sous forme de pointillés (Brown et Lomolino, 1998). Les points
expriment les références géographiques des localités où les spécimens ont été collectés.
Dans le présent travail, deux types de carte en pointillés ont été élaborés avec le logiciel ArcGis
10.5, tels que :
➢ Une carte exprimant les localités où toutes les espèces de Poaceae ont été collectées à
Isalo ;
➢ Une carte représentant les points de distribution de chaque espèce endémique restreinte
de l’Isalo à l’intérieur du parc.
III.2.6 - Etablissement d’une clé de détermination pour les Poaceae de l’Isalo
Une clé de détermination des genres de Poaceae de l’Isalo a été établie dans cette étude.
Elle a été basée sur la clé des genres de Madagascar de Vorontsova et al. (2018). Cette clé

20
préétablie a été modifiée et adaptée selon les caractéristiques des espèces représentant chaque
genre à Isalo. Ces caractéristiques ont été observées directement et vérifiées dans l’ouvrage de
Bosser (1969).
III.2.7 - Analyse et étude de la diversité
La diversité est ici exprimée par le nombre de taxon dans un endroit donné. Afin de
connaitre la diversité de chaque rang taxonomique des Poaceae présent à Isalo, les positions
taxonomiques de chaque espèce ont été consultées sur Vorontsova et al. (2018) et Soreng et al.
(2017).
III.2.7.1 - Richesse spécifique
La richesse floristique est exprimée ici par la diversité α (alpha) ou le nombre
d’espèces présents dans une unité de surface dans le parc. Afin de calculer et de présenter la
richesse spécifique des Poaceae à Isalo, une carte de profil montrant la variation spatiale de la
densité des espèces de Poaceae dans le parc a été élaborée en suivant les étapes suivantes :
➢ Subdiviser la surface du parc en un système de grille (carré) de 2.5×2.5 km, une surface
régulière pouvant représenter approximativement une localité (voir Tableau 1) ;
➢ Calculer le nombre d’espèces de Poaceae rencontrées dans chaque grille. Les espèces ont
été codées afin que le logiciel puisse calculer la diversité spécifique dans chaque grille ;
➢ Fournir des classes de densité en espèce en fonction de la différence entre les nombres
d’espèces dans chaque grille. En fonction de ces classes, les grilles auront différentes couleurs
selon le niveau de la densité en espèces de chaque grille.
Cette analyse mettra en évidence la variation de la richesse spécifique dans
l’espace tout en spécifiant les sites les plus riches en espèces de Poaceae dans le parc de l’Isalo.
III.2.7.2 - Richesse spécifique et degré d’endémisme au niveau des habitats
Les différentes notes sur les lieux de collectes dans la base de données ont été
synthétisées afin d’avoir à la fois :
➢ La liste des habitats qu’occupent chaque espèce ;
➢ La composition en espèces dans chaque type de végétation ou habitat dans le parc.

21
Le nombre d’espèces endémiques a été aussi compté pour chaque type d’habitat.
Ceci afin de mettre en évidence la présence d’endémisme dans les savanes.
La préférence écologique d’une espèce de Poaceae vis-à-vis des conditions a une
certaine relation avec la stratégie de photosynthèse utilisée par l’espèce. Globalement, une
espèce utilisant la stratégie en C4 peut coloniser une grande variété d’habitats à condition qu’il y
ait assez de lumière, tandis qu’une espèce en C3 est limité dans les endroits humides, froids, et
moins éclairés. Ainsi , la distribution des espèces en C3 et C4 dans chaque type d’habitat a été
aussi mis en évidence.

III. RESULTATS ET
INTERPRETATIONS

22
IV.1 - LISTE DES ESPECES DE POACEAE DE L’ISALO
La compilation des données a permis de rassembler 589 spécimens représentant 112
espèces incluant trois variétés de Poaceae qui sont listées dans l’Annexe III. Les informations sur
les collecteurs, le nombre de spécimens par espèce, ainsi que la date de la première collecte ont
été synthétisées et ajoutées à cette liste. Parmi ces 589 spécimens, une vingtaine restent non
identifiables. La figure suivante (Figure 5) représente quelques extraits de photos montrant les
espèces de Poaceae rencontrées à Isalo.
Figure 5: Extraits de photos des Poaceae de l’Isalo. A : Tristachya humbertii - B : port, C : épillet en floraison ;
D : épillet mature de Tristachya (Isalus isalensis) – E : port, F : racème de Humberthochloa bambusiuscula – G :
port, H : épillet de Panicum ibitensis – I : racème de Pseudechinolaena perrieri – J : Andropogon eucomus. Crédits
photos : Rakotomalala Nantenaina (A,B,D,J), Rabehevitra David (E,F,G,H,I), Andriamanohera Andotiana (C).
23
IV.1.1 – Sites de collectes
Pour les 589 spécimens dans la base de données, 408 ont été obtenus depuis la base de
données de KMCC, et de 178 nouvelles collectes issues des investigations de 2018 (Figure 6).
Figure 6: Points de collectes des Poaceae à Isalo 1919-2018
Les points de collectes sur la Figure 6 sont représentés selon les années de collectes et les
sources de données. Pour les spécimens issus de la base de données de KMCC, les classes
d’années correspondent aux périodes où il y avait eu de collectes de Poaceae à Isalo. Il apparait
que les collectes ont été effectuées dans les endroits accessibles du parc, surtout dans la partie
Est où il y a des sites touristiques, et que la partie nord et l’ouest sont encore rarement explorés.
24
Il faut noter qu’il nous a fallu quelques heures en voiture et une journée de marche pour atteindre
Sakamalio. La marche peut encore durer approximativement trois jours pour atteindre l’extrême
nord du parc, à Sahanafa.
IV.1.2 - Collecteurs
C’est Perrier De La Bâthie qui a collecté les premiers spécimens de Poaceae à Isalo en
1919. Ces spécimens représentent : Panicum andringitrense A. Camus et Aristida barbicolis
Trin. & Rupr. Les nombres maximums de spécimens et d’espèces, soit au total 91% des
collectes, sont associés à cinq collecteurs principaux (Tableau 2). Au total, 19 collecteurs ont
participé aux collectes des Poaceae de l’Isalo jusqu’à présent.
Tableau 2: Liste des collecteurs de Poaceae de l’Isalo et nombre de collectes effectuées
Rangs Collecteurs Nombre de
spécimens
(%)
Nombre
d'espèces
collectées
1 Rakotomalala 183 (31) 91
2 Vorontsova 167 (28) 47
3 Bosser 112 (19) 39
4 Humbert 46 (8) 26
5 Morat 28 (5) 10
Autres Decary ; Labat ; Peltier ; Croat ; Cours ; Perrier ;
Boudet ; Razanatsoa; Rakotozafy ; Basse ;
Keraudren-Ymonimn ; Allorge ; Souchiez
54 (9) 37
IV.1.3 - Rapport spécimens-espèces
Les nombres de spécimens par espèce ont permis de voir que les collectes n’ont pas été
faites de manière uniforme et suffisante. Beaucoup d’espèces n’ont été collectées qu’une fois (31
espèces) ou deux fois seulement (26 espèces), et rares sont celles collectées plus de dix fois. Les
quelques espèces ayant les plus grands nombres de spécimens sont celles qui ont fait objet
d’études particulières, comme Loudetia simplex, sujet de recherche de Hagl, et al., (in press),

25
collectées 50 fois. Elle est suivie par Tristachya (Isalus isalensis), Schizachyrium sanguineum et
Craspedorhachis africana, toutes recensées à Isalo plus de 30 fois.
IV.1.3 - Découvertes pour Isalo
Les 181 spécimens de Poaceae collectés en 2018 représentent 93 espèces. Parmi elles, 38
sont des espèces nouvelles pour Isalo dont quatre espèces endémiques de Madagascar dont l’une
du genre endémique malgache : Neostapfiella. La plupart de ces espèces nouvellement connues à
Isalo sont représentées en générale, soit par des :
➢ herbes communes comme : Chloris pycnothrix Trin., Eleusine indica Gaertn. Cynodon
dactylon (L.) Pers.) ;
➢ petites herbes difficiles à remarquer comme : Aristida cumingiana Trin. & Rupr.;
Schizachyrium brevifolium (Sw.) Nees ex Büse.
➢ Herbes de grande taille qui ne sont pas attrayantes et semblent difficiles à collecter
comme : Neyraudia arundinacea (L.) Henrard; Rottboellia cochinchinensis (Lour.) Clayton.
La plupart de ces nouvelles espèces pour Isalo ont été connues precedemment se
distribuer dans des zones très éloignées de l’Isalo (voir : Rabarivola et al., 2019d). Comme
Neostapfiella perrieri A.Camus, elle a été rencontrée uniquement au nord-ouest et de l’extrême
sud de Madagascar (Vorontsova & Rakotoarisoa, 2014 ; Rabarivola et al., 2019d). Il faut
toutefois noter que cette espèce a été représentée par un groupe d’environs cinq individus
rencontrés dans un seul endroit qui est très fréquenté. Il se pourrait qu’elle soit accidentellement
introduite dans le parc par les visiteurs, et que sa présence n’y est pas naturelle.
L’inventaire sur terrain n’a pas seulement permis la découverte de plus d’espèces mais a
aussi permis d’ajouter d’avantages de spécimens pour les espèces rares. Le tout troisième
spécimen de Pseudechinolaena perrieri (voir Figure 5), espèce endémique de Madagascar, a été
fourni par cette étude. Les deux premiers spécimens ont été collectés par Henri Perrier De La
Bâthie (I1224) à Ankarafantsika, et par Jean Bosser (19124) à Isalo en 1964.

26
IV.1.4 - Découverte pour la science
Une espèce de bambou pousse aux environs des rivières de Namaza et de Sakamalio. Elle
a été collectée pour la première fois par Maria Vorontsova en 2013, et n’a pas encore fleuri
depuis. Cette espèce mystérieuse de bambou forme une population archée et pachymorphe
indiquant une nette affinité morphologique avec la sous-famille Bambusoideae, tribu
Bambuseae, sous-tribus Hickeliinae (Vorontsova et al., 2018). Des analyses phylogénétiques de
ses marqueurs chloroplastiques ont révélés que l’espèce est plus proche du genre Perrierbambus
(Hackel et al, 2018), mais les caractères des tiges ne conviennent pas du tout avec ce taxon. Il se
peut alors que ce soit un nouveau genre pas encore décrit, probablement endémique de l’Isalo.
Figure 7: Herbiers de la nouvelle espèce de Bambou dans le Parc de l’Isalo (port à droite et feuille du chaume à
gauche)
IV.2- EVOLUTION DE LA CONNAISSANCE SUR LES POACEAE DE L’ISALO
La collecte de Poaceae à Isalo a débuté en 1919, mais n’a véritablement démarré qu’à
partir de 1950 (Figure 8). De 1919 à 1950, donc en trois décennies, seule 12% des espèces de
Poaceae de l’Isalo ont été recensées. Mais les efforts de documentation de ce groupe ont connu
un grand essor de 1950 à 1970, et les espèces connues ont augmenté de 52%. Cette progression
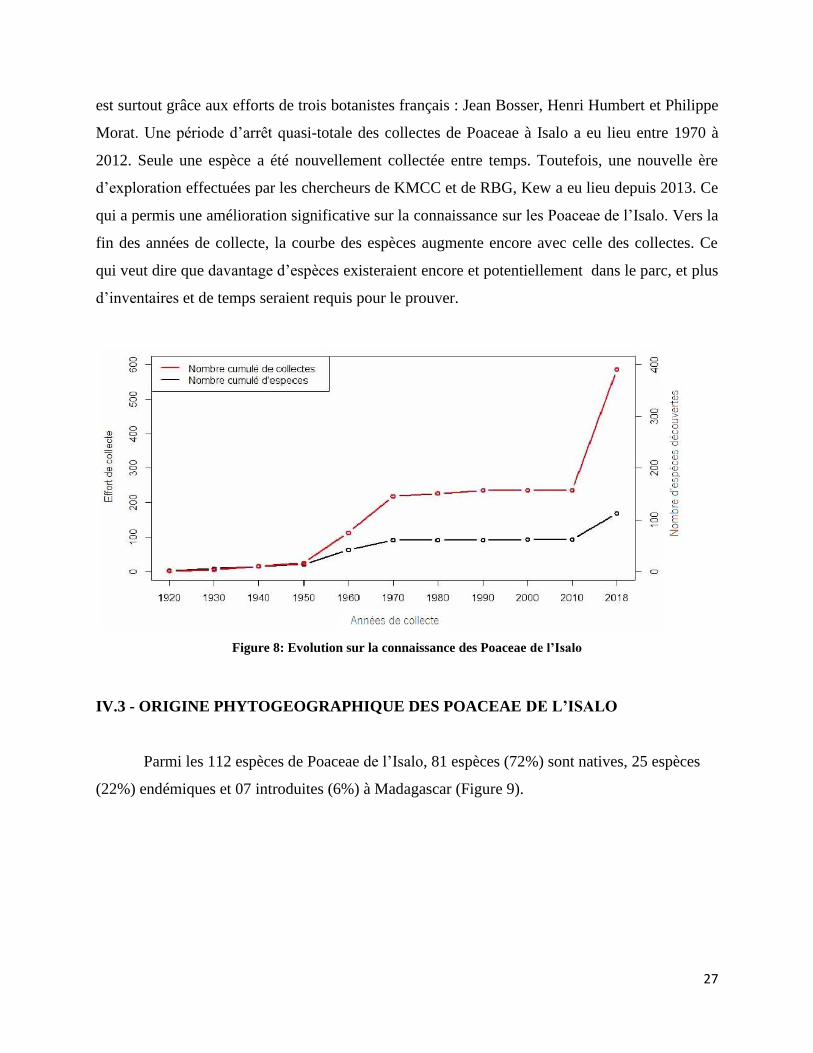
27
est surtout grâce aux efforts de trois botanistes français : Jean Bosser, Henri Humbert et Philippe
Morat. Une période d’arrêt quasi-totale des collectes de Poaceae à Isalo a eu lieu entre 1970 à
2012. Seule une espèce a été nouvellement collectée entre temps. Toutefois, une nouvelle ère
d’exploration effectuées par les chercheurs de KMCC et de RBG, Kew a eu lieu depuis 2013. Ce
qui a permis une amélioration significative sur la connaissance sur les Poaceae de l’Isalo. Vers la
fin des années de collecte, la courbe des espèces augmente encore avec celle des collectes. Ce
qui veut dire que davantage d’espèces existeraient encore et potentiellement dans le parc, et plus
d’inventaires et de temps seraient requis pour le prouver.
Figure 8: Evolution sur la connaissance des Poaceae de l’Isalo
IV.3 - ORIGINE PHYTOGEOGRAPHIQUE DES POACEAE DE L’ISALO
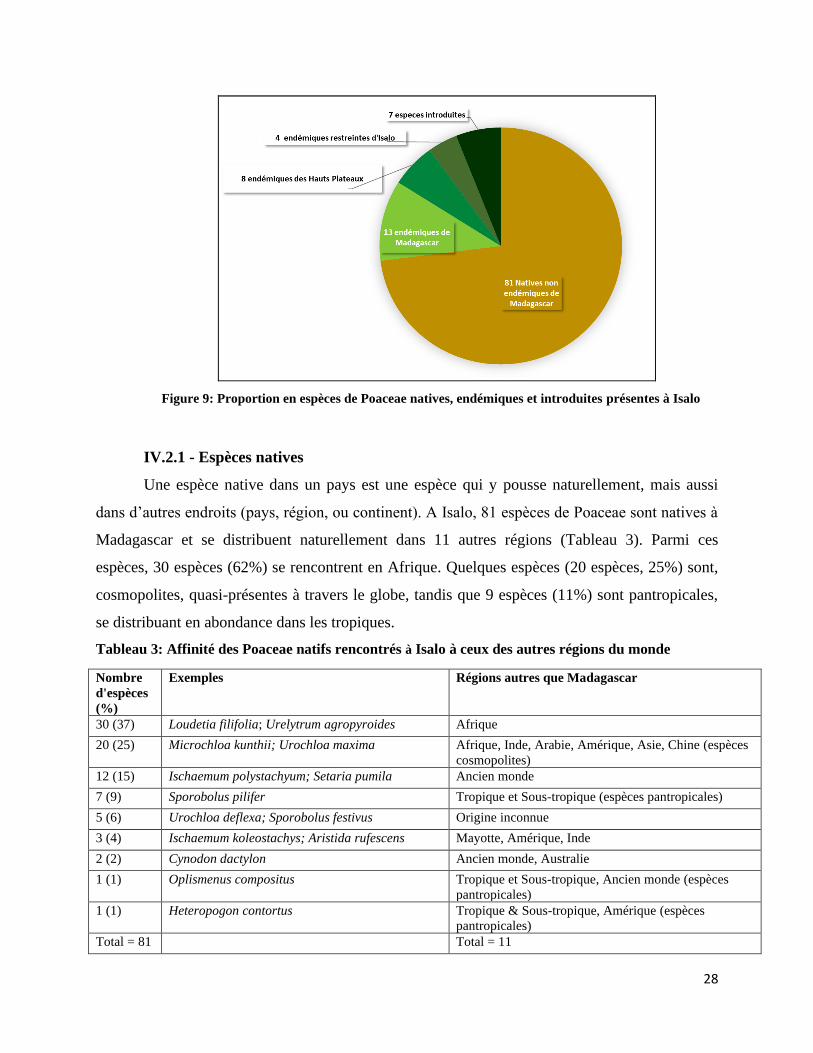
Parmi les 112 espèces de Poaceae de l’Isalo, 81 espèces (72%) sont natives, 25 espèces
(22%) endémiques et 07 introduites (6%) à Madagascar (Figure 9).
28
Figure 9: Proportion en espèces de Poaceae natives, endémiques et introduites présentes à Isalo
IV.2.1 - Espèces natives
Une espèce native dans un pays est une espèce qui y pousse naturellement, mais aussi
dans d’autres endroits (pays, région, ou continent). A Isalo, 81 espèces de Poaceae sont natives à
Madagascar et se distribuent naturellement dans 11 autres régions (Tableau 3). Parmi ces
espèces, 30 espèces (62%) se rencontrent en Afrique. Quelques espèces (20 espèces, 25%) sont,
cosmopolites, quasi-présentes à travers le globe, tandis que 9 espèces (11%) sont pantropicales,
se distribuant en abondance dans les tropiques.
Tableau 3: Affinité des Poaceae natifs rencontrés à Isalo à ceux des autres régions du monde
Nombre
d'espèces
(%)
Exemples Régions autres que Madagascar
30 (37) Loudetia filifolia; Urelytrum agropyroides Afrique
20 (25) Microchloa kunthii; Urochloa maxima Afrique, Inde, Arabie, Amérique, Asie, Chine (espèces
cosmopolites)
12 (15) Ischaemum polystachyum; Setaria pumila Ancien monde
7 (9) Sporobolus pilifer Tropique et Sous-tropique (espèces pantropicales)
5 (6) Urochloa deflexa; Sporobolus festivus Origine inconnue
3 (4) Ischaemum koleostachys; Aristida rufescens Mayotte, Amérique, Inde
2 (2) Cynodon dactylon Ancien monde, Australie
1 (1) Oplismenus compositus Tropique et Sous-tropique, Ancien monde (espèces
pantropicales)
1 (1) Heteropogon contortus Tropique & Sous-tropique, Amérique (espèces
pantropicales)
Total = 81
Total = 11

29
IV.2.2 - Espèces introduites
Une espèce introduite est une espèce non native d’un lieu donné. On rencontre 07 espèces
originaires d’Amérique, d’Afrique, Asie, Australie, du Nouveau monde et d’Inde à Isalo (voir
Annexe III). Ce sont : Cenchrus echinatus L., Cenchrus polystachios (L.) Morrone, Cenchrus
purpureus (Schumach.) Morrone, Chloris virgata P.Durand, Digitaria violascens Link,
Echinochloa colona (L.) Link, Enteropogon prieurii (Kunth) Clayton.
IV.2.3 - Espèces endémiques
Une espèce endémique est une espèce native dans un lieu donné et y est restreinte. A
Isalo, 25 espèces sont endémiques de Madagascar dont 02 espèces des genres endémiques
malgaches : Neostapfiella et Pseudolasiacis. Le taux d’endémisme des Poaceae pour Isalo est
donc 22%. Selon la distribution de ces espèces, 03 espèces et une variété (4%) sont restreintes à
Isalo, 08 espèces (7%) restreintes aux hauts plateaux, et 12 espèces (11%) sont à large répartition
à Madagascar (voir Figure 9).
IV.2.4 - Espèces endémiques locales de l’Isalo
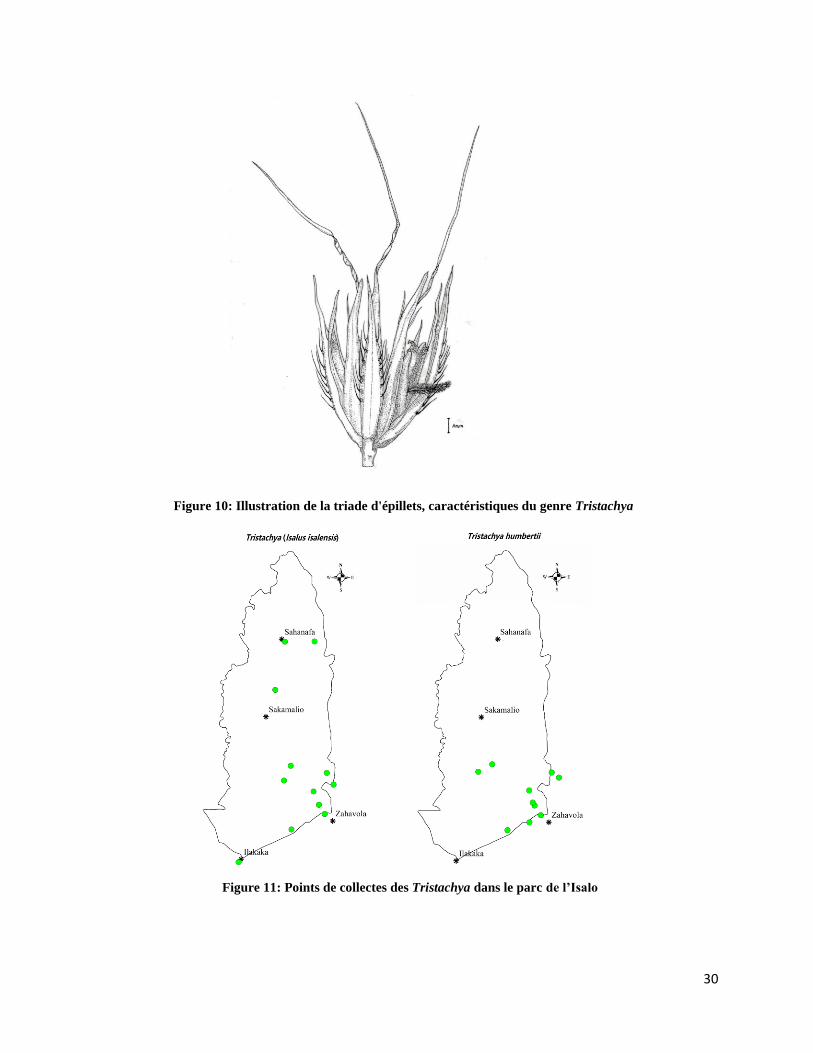
➢ Tristachya (Isalus isalensis) et Tristachya humbertii
Deux espèces de Tristachya (sur 03 dans tout Madagascar) se rencontrent à Isalo et y sont
restreintes. Le genre Tristachya se reconnait facilement par leurs rachéoles se terminant par un
lot de trois épillets munis de poils à base tuberculée (Figure 10). Les deux espèces de l’Isalo se
distinguent entre elles par la morphologie de leur port et celle de leurs feuilles, et sont
actuellement en cours de révision taxonomique (voir Figure 5). Tristachya (Isalus isalensis) a
été placé dans le genre Isalus J. B. Phipps et sera combiné au genre Tristachya. Dans le parc de
l’Isalo uniquement, ces espèces ont été collectées des localités (Figure 11) associées, en général,
à des habitats ouverts ou semi ouverts, et sur des substrats rocheux ou sableux. Par contre,
Tristachya (Isalus isalensis) a été rencontrée une fois dans le canyon de Ranohira, qui est un
environnement humide et moins claire.
30
Figure 10: Illustration de la triade d'épillets, caractéristiques du genre Tristachya
Figure 11: Points de collectes des Tristachya dans le parc de l’Isalo
31
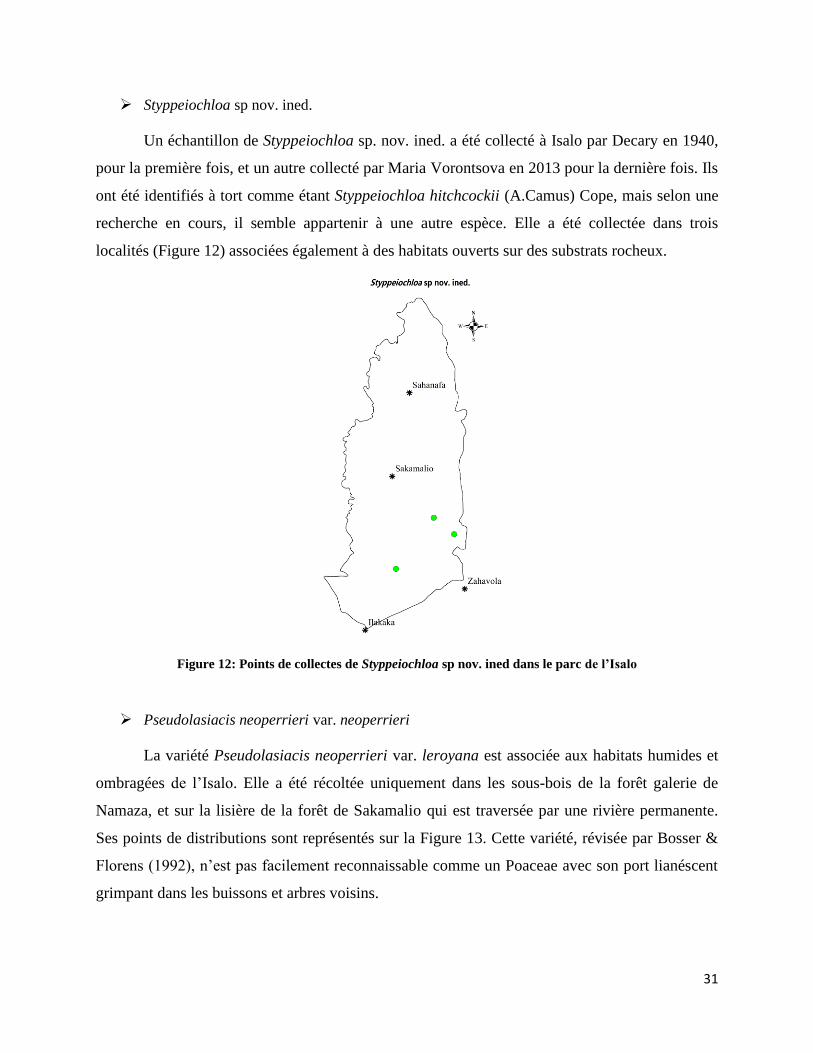
➢ Styppeiochloa sp nov. ined.
Un échantillon de Styppeiochloa sp. nov. ined. a été collecté à Isalo par Decary en 1940,
pour la première fois, et un autre collecté par Maria Vorontsova en 2013 pour la dernière fois. Ils
ont été identifiés à tort comme étant Styppeiochloa hitchcockii (A.Camus) Cope, mais selon une
recherche en cours, il semble appartenir à une autre espèce. Elle a été collectée dans trois
localités (Figure 12) associées également à des habitats ouverts sur des substrats rocheux.
Figure 12: Points de collectes de Styppeiochloa sp nov. ined dans le parc de l’Isalo
➢ Pseudolasiacis neoperrieri var. neoperrieri
La variété Pseudolasiacis neoperrieri var. leroyana est associée aux habitats humides et
ombragées de l’Isalo. Elle a été récoltée uniquement dans les sous-bois de la forêt galerie de
Namaza, et sur la lisière de la forêt de Sakamalio qui est traversée par une rivière permanente.
Ses points de distributions sont représentés sur la Figure 13. Cette variété, révisée par Bosser &
Florens (1992), n’est pas facilement reconnaissable comme un Poaceae avec son port lianéscent
grimpant dans les buissons et arbres voisins.

32
Figure 13: Points de collectes de Pseudolasiacis neoperrieri var. neoperrieri
IV.3 - DIVERSITE TAXONOMIQUE DES POACEAE DE L’ISALO
Les Poaceae de l’Isalo se répartissent en 07 sous familles, 16 tribus, 58 genres et 112 spp.
dont trois variétés. Ils appartiennent largement au clade PACMAD avec 97% des espèces, mais
aussi au clade BEP (3% des espèces). La sous famille ancestrale y fait défaut. Globalement les
taxa les plus diverses appartiennent toujours aux Panicoideae. La sous familles des Pooideae,
celle la plus riche parmi les sous familles de Poaceae (Kellogg, 2015), n’a pas été rencontrée
dans le parc. C’est parce qu’elle exige une certaine altitude et un climat plus humide.
IV.3.1 - Diversité au niveau des sous familles
Sept sur 10 sous familles de Poaceae à Madagascar sont présentes à Isalo. La plupart des
tribus (38%), des genres (36%) et des espèces (66%) appartiennent à la sous-famille des
Panicoideae (Tableau 4). La sous famille des Chloridoideae démontre aussi une certaine diversité
taxonomique comparée aux autres familles. La sous famille des Bambusoideae est la moins
représentée à Isalo, avec seulement une espèce.
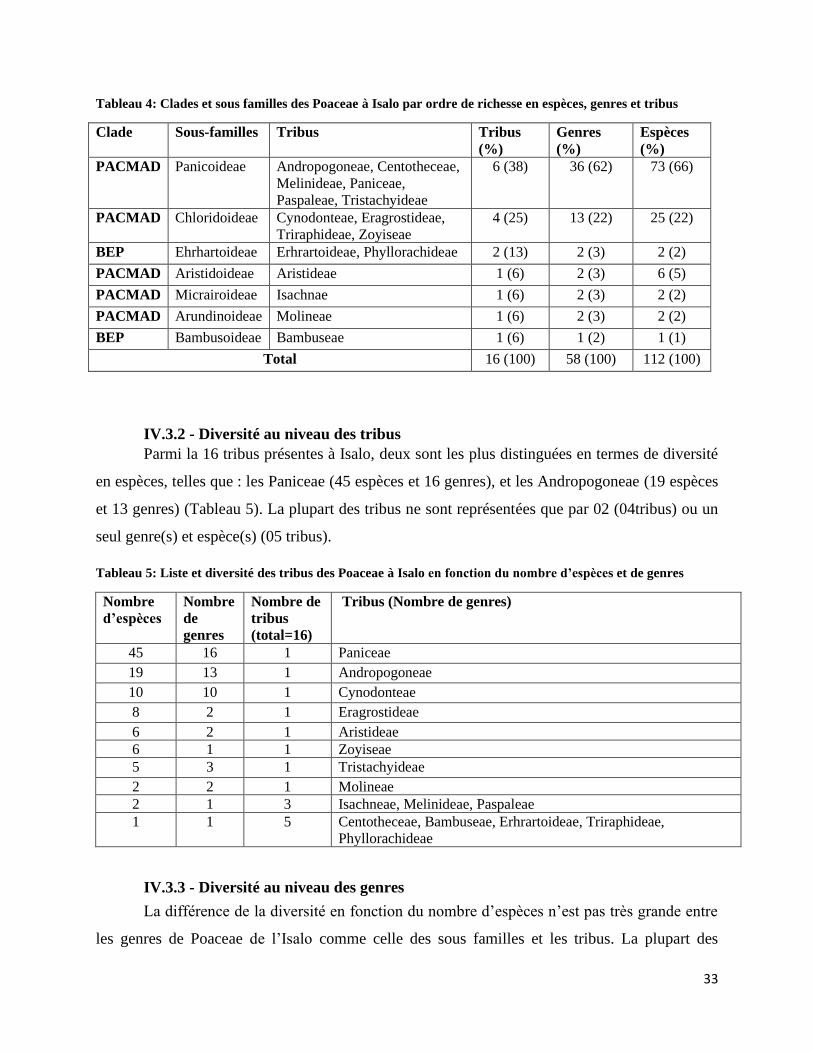
33
Tableau 4: Clades et sous familles des Poaceae à Isalo par ordre de richesse en espèces, genres et tribus
Clade Sous-familles Tribus Tribus
(%)
Genres
(%)
Espèces
(%)
PACMAD Panicoideae Andropogoneae, Centotheceae,
Melinideae, Paniceae,
Paspaleae, Tristachyideae
6 (38) 36 (62) 73 (66)
PACMAD Chloridoideae Cynodonteae, Eragrostideae,
Triraphideae, Zoyiseae
4 (25) 13 (22) 25 (22)
BEP Ehrhartoideae Erhrartoideae, Phyllorachideae 2 (13) 2 (3) 2 (2)
PACMAD Aristidoideae Aristideae 1 (6) 2 (3) 6 (5)
PACMAD Micrairoideae Isachnae 1 (6) 2 (3) 2 (2)
PACMAD Arundinoideae Molineae 1 (6) 2 (3) 2 (2)
BEP Bambusoideae Bambuseae 1 (6) 1 (2) 1 (1)
Total 16 (100) 58 (100) 112 (100)
IV.3.2 - Diversité au niveau des tribus
Parmi la 16 tribus présentes à Isalo, deux sont les plus distinguées en termes de diversité
en espèces, telles que : les Paniceae (45 espèces et 16 genres), et les Andropogoneae (19 espèces
et 13 genres) (Tableau 5). La plupart des tribus ne sont représentées que par 02 (04tribus) ou un
seul genre(s) et espèce(s) (05 tribus).
Tableau 5: Liste et diversité des tribus des Poaceae à Isalo en fonction du nombre d’espèces et de genres
Nombre
d’espèces
Nombre
de
genres
Nombre de
tribus
(total=16)
Tribus (Nombre de genres)
45 16 1 Paniceae
19 13 1 Andropogoneae
10 10 1 Cynodonteae
8 2 1 Eragrostideae
6 2 1 Aristideae
6 1 1 Zoyiseae
5 3 1 Tristachyideae
2 2 1 Molineae
2 1 3 Isachneae, Melinideae, Paspaleae
1 1 5 Centotheceae, Bambuseae, Erhrartoideae, Triraphideae,
Phyllorachideae
IV.3.3 - Diversité au niveau des genres
La différence de la diversité en fonction du nombre d’espèces n’est pas très grande entre
les genres de Poaceae de l’Isalo comme celle des sous familles et les tribus. La plupart des
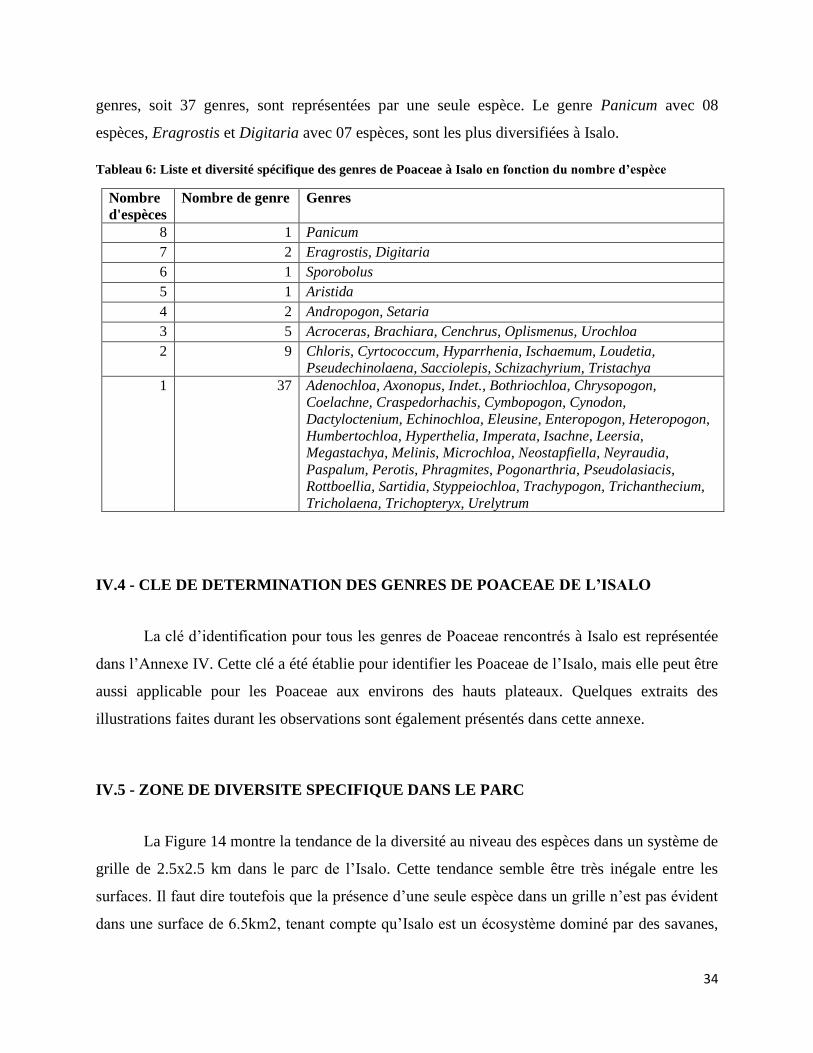
34
genres, soit 37 genres, sont représentées par une seule espèce. Le genre Panicum avec 08
espèces, Eragrostis et Digitaria avec 07 espèces, sont les plus diversifiées à Isalo.
Tableau 6: Liste et diversité spécifique des genres de Poaceae à Isalo en fonction du nombre d’espèce
Nombre
d'espèces
Nombre de genre Genres
8 1 Panicum
7 2 Eragrostis, Digitaria
6 1 Sporobolus
5 1 Aristida
4 2 Andropogon, Setaria
3 5 Acroceras, Brachiara, Cenchrus, Oplismenus, Urochloa
2 9 Chloris, Cyrtococcum, Hyparrhenia, Ischaemum, Loudetia,
Pseudechinolaena, Sacciolepis, Schizachyrium, Tristachya
1 37 Adenochloa, Axonopus, Indet., Bothriochloa, Chrysopogon,
Coelachne, Craspedorhachis, Cymbopogon, Cynodon,
Dactyloctenium, Echinochloa, Eleusine, Enteropogon, Heteropogon,
Humbertochloa, Hyperthelia, Imperata, Isachne, Leersia,
Megastachya, Melinis, Microchloa, Neostapfiella, Neyraudia,
Paspalum, Perotis, Phragmites, Pogonarthria, Pseudolasiacis,
Rottboellia, Sartidia, Styppeiochloa, Trachypogon, Trichanthecium,
Tricholaena, Trichopteryx, Urelytrum
IV.4 - CLE DE DETERMINATION DES GENRES DE POACEAE DE L’ISALO
La clé d’identification pour tous les genres de Poaceae rencontrés à Isalo est représentée
dans l’Annexe IV. Cette clé a été établie pour identifier les Poaceae de l’Isalo, mais elle peut être
aussi applicable pour les Poaceae aux environs des hauts plateaux. Quelques extraits des
illustrations faites durant les observations sont également présentés dans cette annexe.
IV.5 - ZONE DE DIVERSITE SPECIFIQUE DANS LE PARC
La Figure 14 montre la tendance de la diversité au niveau des espèces dans un système de
grille de 2.5x2.5 km dans le parc de l’Isalo. Cette tendance semble être très inégale entre les
surfaces. Il faut dire toutefois que la présence d’une seule espèce dans un grille n’est pas évident
dans une surface de 6.5km2, tenant compte qu’Isalo est un écosystème dominé par des savanes,

35
donc aussi par les Poaceae. Ce cas ne peut être expliquée que par l’insuffisance de collectes dans
ces sites. Toutefois, la richesse spécifique est bien exposée les sites presque suffisamment
investigués.
Le site Namaza détient le maximum de diversité en Poaceae dans le parc, abritant jusqu’à
64 espèces parmi 112 spp. Il est suivi par Ranohira qui abrite une vingtaine d’espèces. Cette
localité est caractérisée par la présence de canyons où une diversité de microhabitats sont
disponibles. Etant l’environnement humide et surtout moins éclairés de l’Isalo, les canyons
offrent des niches pour les espèces de Poaceae qui exigent ces conditions. Ce qui pourrait
expliquer la potentielle diversité spécifique dans ces sites, outre le nombre des collectes.
Figure 14: Zones potentielles de diversité en espèces de Poaceae dans le parc

36
IV.6 - DIVERSITE ET ENDEMISME PAR TYPE DE VEGETATION
IV.6.1 - Tendance de distribution des espèces dans le parc
La présence d’une espèce dans un habitat donné indique que les conditions physiques de
cet habitat sont adéquates pour l’espèce. Ici, les habitats ont été prédéterminés et sont représentés
en gros par les types de végétations de l’Isalo. Au total, six types ont été catégorisés selon les
informations sur la base de données, tels que : les savanes, les forêts galeries, les Tapia, les zones
humides, les rochers, et les bords de la RN7. Les zones humides regroupent les marais et les
habitats humides dans les canyons à l’exclus des forêts et leurs sous-bois même étant traversés
par des cours d’eau. Les bords de route (savanes) ont été spécifiés particulièrement comme un
type d’habitat très fréquenté. La liste des habitats occupées par chaque espèce est présentée dans
l’Annexe III.
La Figure 15 suivante montre la capacité des espèces de Poaceae à coloniser les habitats
de l’Isalo. Il apparaît que la plupart des espèces (63 espèces) ne se rencontre que dans un seul
habitat, et qu’aucune espèce ne colonise à la fois les six habitats. Tristachya (Isalus isalensis) a
été retrouvée dans les cinq habitats à part les sous-bois des forêts galeries. Il en est de même pour
Urochloa maxima (Jacq.) R.D.Webster qui a été rencontré dans tous les types d’habitats sauf sur
les rochers.
Dire que la famille des Poaceae est cosmopolite et les espèces sont capables de dominer
plusieurs endroits, une certaine exigence ou préférence se remarque au niveau des espèces vis-à-
vis des paramètres qui conditionnent le milieu.
37
Figure 15: Tendance de distribution des espèces dans les habitats disponibles de l’Isalo
IV.6.2 - Richesse spécifique et endémisme au niveau des habitats
La Figure 16 permet de synthétiser la richesse spécifique par habitat en fonction du
nombre des espèces dans chaque habitat. La Figure 17 montre la composition des espèces en
fonction du type de photosynthèse utilisé par chaque espèce. Les espèces de Poaceae de l’Isalo
en C4 comptent au total 89 espèces (79%) et les C3, 23 espèces (21%). La liste des habitats
occupés par chaque espèce avec leur type de photosynthèse est représentée en Annexe III.
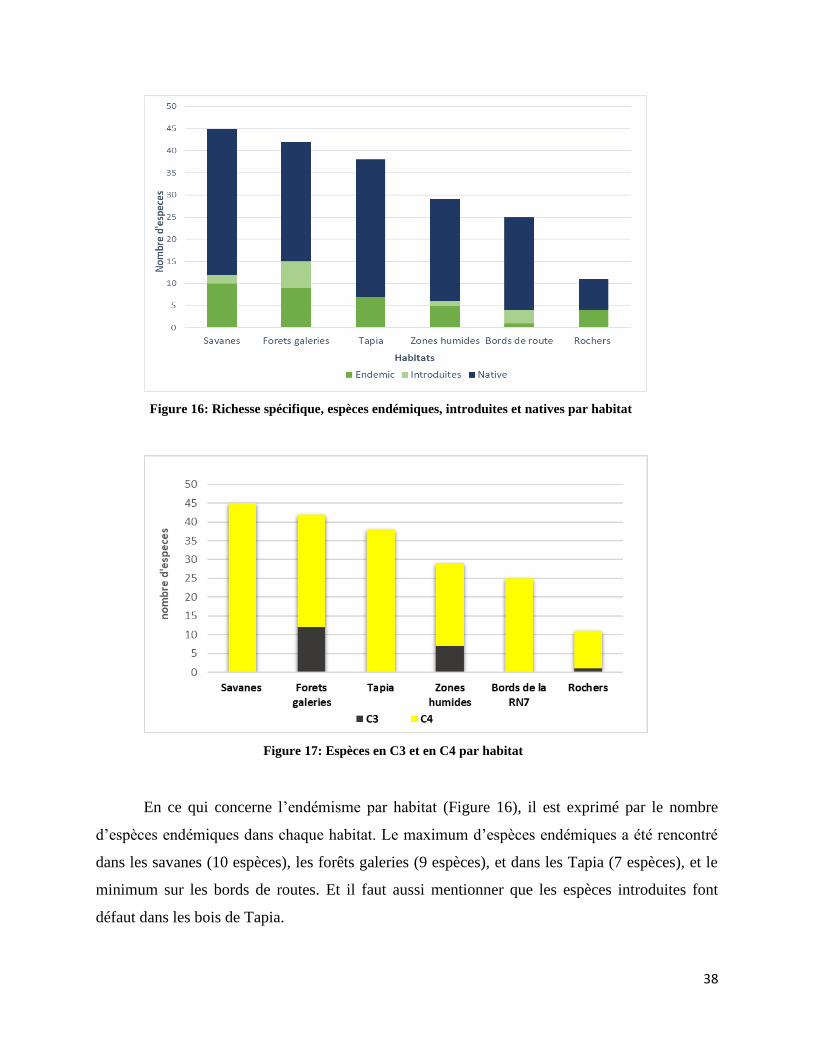
38
Figure 16: Richesse spécifique, espèces endémiques, introduites et natives par habitat
Figure 17: Espèces en C3 et en C4 par habitat
En ce qui concerne l’endémisme par habitat (Figure 16), il est exprimé par le nombre
d’espèces endémiques dans chaque habitat. Le maximum d’espèces endémiques a été rencontré
dans les savanes (10 espèces), les forêts galeries (9 espèces), et dans les Tapia (7 espèces), et le
minimum sur les bords de routes. Et il faut aussi mentionner que les espèces introduites font
défaut dans les bois de Tapia.

39
En ce qui concerne la richesse spécifique (Figure 16), les savanes sont les plus diverses,
suivies de près par les forêts galeries et par les Tapia, et les rochers sont les moindres diverses.
Cette analyse ne tient pas compte de la taille des habitats, mais si autrement, la diversité dans les
savanes serait la moindre car c’est, de loin, l’habitat le plus vaste. Les espèces de chaque habitat
démontrent une certaine affinité à chaque habitat selon une analyse descriptive de la composition
en espèce suivante et la Figure 17.
➢ Forêts galeries
Les forêts galeries comportent 42 espèces dont 28 y sont inféodées. Ces espèces sont
en générale caractérisées par un port rampant ou genouillé, avec des feuilles suffisamment larges
pour acquérir de la lumière, qui est limitante dans certains endroits. Cet habitat à l’abri du feu et
conditionnée par une certaine humidité et un éclairage limité, abrite la plupart des espèces en C3
(12 espèces) rencontrées à Isalo. Parmi les espèces, il y a également des espèces sont des espèces
C4 (30 espèces). La plupart des Poaceae de cet habitat est membre des Paniceae.
➢ Zones humides
Les zones humides abritent 29 espèces dont 17 y sont toutes inféodées. Cet habitat
caractérisé par la présence d’eau en permanence est habité par des espèces naturellement
aquatique ou opportunistes à ces conditions, comme les espèces de Phragmites, Leersia,
Sacciolepis, Andropogon et certaines Panicum, appartenant surtout aux tribus des Paniceae.
Parmi ces espèces, 22 sont en C4 et sept en C3.
➢ Savanes
Dans les savanes, 45 espèces y sont rencontrées parmi lesquelles huit espèces y sont
inféodées. Ces espèces sont en générale caractérisées par un port cespiteux formant une touffe
dense et portant des feuilles à bords enroulés et des bourgeons apicaux souterrains pour survivre
aux feux fréquents. Ces espèces sont tous des C4 (Figure 17) et la plupart est membre des
Andropogoneae.

40
➢ Bois de Tapia
Dans les sous-bois de Tapia, 38 espèces de Poaceae sont rencontrées avec six espèces
inféodées. Les compositions taxonomiques des Tapia et des savanes sont presque similaires, car
ils sont formés par un même tapis herbacé continu. Seulement le couvert herbacé dans les Tapia
sont moins éclairés que dans les savanes. Les espèces y sont également tous des C4 et membre
d’Andropogoneae pour la plupart.
➢ Bords de la RN7
Cet habitat est constitué par des savanes mais dans un milieu plus perturbé. Sa
composition floristique est similaire aux savanes. Il abrite 25 espèces au total Seulement une
espèce du sud-ouest (Cenchrus echinatus L.) le distingue des savanes et des autres habitats.
➢ Rochers
Les rochers abritent seulement onze espèces avec trois espèces inféodées. Cet habitat
est caractérisé par des espèces xérophytiques naines qui se répartissent au hasard, laissant des
sols nus a grande proportion. Les Tristachyideae (Loudetia simplex et Tristachya spp.) sont les
plus fréquents sur les rochers. Une espèce endémique Sartidia isaloensis pousse particulièrement
sur les escarpements des rochers de l’Isalo.

V. DISCUSSION

41
V.1 - FLORE DE POACEAE DE L’ISALO
Cette étude présente la première recherche visant à fournir des détails complets sur la
diversité taxonomique des Poaceae à Isalo. Les données présentées dans les publications
antérieures ont été basées sur des données insuffisantes et sont de nature plus écologique que
systématique. La MNP Ranohira cite 25 genres et de 38 espèces de graminées à Isalo en 2017
(MNP, 2017), tandis que la synthèse de Solofondranohatra et al. (2018) a avancé la présence de
71 espèces de Poaceae dans les hauts plateaux, incluant l’Isalo, en 2018. Après la synthèse des
collections historiques et celles issues des plus récentes investigations faites lors de cette étude,
les Poaceae connus à Isalo atteignent 112 spp., 58 genres, 16 tribus et sept sous-familles. Cette
étude présente la liste de ces taxa et propose une guide d’identification des genres de Poaceae
pour Isalo et ses environs.
L’évolution historique de la connaissance sur les Poaceae de l’Isalo correspond bien à
l’histoire de l’exploration botanique dans cette zone, qui a commencé en 1910, ainsi que dans
tout Madagascar (Goodman et al.,2018). Le boom de découverte de 1950 à 1970 était dans le
cadre d’une campagne d’inventaire des biotes natifs et de l’évaluation des ressources naturelles
de l’ile, et la suspension des recherches vers la fin du XXème siècle, étaient commune dans toute
l’ile et pour tous taxa. A ce jour, la famille des Poaceae constitue la première famille de plantes
ayant fait l’objet d’une checklist à Isalo. Les recherches sur les autres familles de plantes, à Isalo,
semblent être encore en suspension et encore peu nombreuses, même pour les familles
endémiques malgaches qui y sont présentes.
Grâce à des recherches plus dédiées à la documentation de la diversité des Poaceae
depuis 2013, le nombre d’espèces connues à Isalo a été doublé avec une nouvelle espèce de
Bambou découverte récemment. Cette énorme évolution en si peu de temps rappelle la réelle
insuffisance et l’ancienneté des données disponibles sur les Poaceae de Madagascar
actuellement, tenant compte que l’Isalo n’est que le deuxième site ayant une liste complète de ses
Poaceae après Itremo (Nanjarisoa et al., 2017). La présente étude témoigne la signification d’un
effort de collectes non sélectives, dans les zones déjà explorées ou pas, dans l’évolution de la
connaissance d’un taxon donné. Adoptant la même méthode, Nanjarisoa et al (2017) ont même

42
triplé le nombre d’espèces de Poaceae connues dans la Nouvelle Aire Protégée d’Itremo.
Naturellement, le manque d’effort délibéré de collectes ne peut pas beaucoup avancer les
recherches sur la diversité, bien qu’il s’agisse d’un taxon très diversifié et dans des zones
potentiellement riches en espèces. Cela reflète les faibles nombres d’espèces Poaceae connus
dans le Parc National d'Andringitra (18 espèces) dans l’étude de Lewis et al. (1996), et dans la
réserve de Manongarivo (42 espèces) dans celui de Gautier (1997). Toutefois, malgré notre
effort, cette liste a été démontrée ne pas encore être totalement exhaustive, du fait que certains
spécimens (dans la base de données) n’ont pas pu bien être identifiés et beaucoup d’endroits
potentiels dans le parc n’a pas encore été explorés.
Concernant la diversité, la famille des Poaceae représente la famille la plus riche en
espèce listée dans le parc. Les Fabaceae (42 espèces sur 592 à Madagascar) et les Asteraceae (21
sur 516 espèces), familles globalement très riches en espèces, y sont présents mais ont encore
peu de données (Du Puy, 2002 ; MNP, 2017). Le nombre de Poaceae à Isalo est similaire à celui
d’Itremo qui abrite 100 espèces (Nanjarisoa et al., 2017). Ces deux sites ont également les
mêmes compositions taxonomiques, surtout au niveau des sous familles, tribus et genres. De
plus, une cinquantaine d’espèces (56 espèces) se partagent entre eux. Ce résultat est conforme
avec la découverte de Vorontsova et al. (2016), qui aient démontré la forte affinité en
composition floristique de deux sites de la même écorégion.
Cette étude a permis de rajouter des données sur l’endémicité de la flore de l’Isalo. Soit
227 sur 401 espèces de plantes sont endémiques dans le parc, faisant un taux de 70,1%
(Goodman et al.,2018), qui est de loin supérieur à celle de la famille des Poaceae. Mais le taux
d’endémisme en Poaceae est généralement plus faible que celui des autres familles
d’Angiospermes à Madagascar (Callmander et al., 2011). Au niveau spatial, ce taux
d’endémisme de 22% de Poaceae à Isalo est faible par rapport à celui d’Itremo qui a un taux de
36% (Nanjarisoa et al. (2017), et l’est encore plus par rapport à celui d’Andringitra (97%) et de
Manongarivo (58%). Ces sites des hautes montagnes démontrent déjà un certain niveau
d’endémisme en Poaceae élevé même avec de faibles nombres préliminaires d’espèces connues
(Lewis et al.,1996 ; Gautier, 1997). Mais il faut rappeler que dans tout Madagascar, l’endémisme
en Poaceae est le plus faible dans les hauts plateaux (Bond et al., 2008). Suivant une échelle plus

43
grande, le niveau d’endémisme des Poaceae malgaches (40%) figure généralement parmi les plus
élevés dans le monde (Vorontsova el al., 2016), et le cas des Poaceae de l’Isalo se conforme
avec. Le taux d’endémisme en Poaceae à Isalo est élevé par rapport à celui du National Parc de
Mkomazi (3%) et à celui de la Réserve de Selous Game (12%) en Tanzanie Afrique (Vollesen et
al.,1999 ; Williams et al.,2016). Il faut quand même noter que dans le continent africain, une
extinction massive d’espèces est survenue autrefois, et cela pourrait être l’une des raisons de ce
faible taux d’endémisme.
Dans cette étude, des analyses sur la diversité des Poaceae dans le parc de l’Isalo ont été
avancées, et a présenté les zones potentielles de diversité. Par défaut de données écologiques, et
de temps pour les collecter, seules des données descriptives ont été avancées concernant ce sujet.
Les paramètres environnementaux qui conditionnent la distribution et la variation de la diversité
des espèces à Isalo peuvent faire un sujet de recherche ultérieure pour Isalo et ses Poaceae, en
utilisant des méthodes plus analytiques.
V.2 - PLAUSIBLE ORIGINE ANCIENNE DE LA VEGETATION DE L’ISALO
Les écosystèmes herbeux malgaches sont considérés comme la répercussion de la
déforestation surgissant à plusieurs reprises depuis l’arrivée de l’Homme dans l’ile vers 1350-
1100 années BP. (Anderson et al., 2018). Jugés par leur faible diversité et la quasi-absence
d’espèce endémique, ils sont considérés comme des habitats dégradés et secondaires (Morat,
1973). Pourtant, plusieurs lignes d’évidences peuvent nous éloigner de ce point de vue.
Outre les plantes endémiques malgaches comme Uapaca bojeri, Monotes
madagascariensis (restreintes à Isalo), qui constituent et se trouvent naturellement dans les
savanes de l’Isalo (Solofondranohatra et al., 2018), s’ajoutent neuf espèces endémiques de
Poaceae trouvées lors de cette étude. Trois parmi ces espèces sont des espèces endémiques
restreintes de l’Isalo. En outre, comme les Poaceae de l’Isalo sont des C4 et sont très similaires à
celui d’Afrique, ils peuvent faire partie de l’expansion des Poaceae en C4 vers Miocène (Bond et

44
al., 2008 ; Hackel et al., 2018). Ce qui indiquerait une présence antérieure des savanes à l’arrivée
de l’Homme.
Faramalala (1988) et Rakotoarimanana (2008) ont mentionné auparavant que la
physionomie et la composition spécifique se diffèrent entre les écosystèmes dans les différentes
écorégions malgaches. L’étude de cette différence en termes de composition floristique a été
démontrée pour les Poaceae à Madagascar, et cela servait d’interprétation sur leur origine. Si les
Poaceae et les savanes ne se sont répandues dans tout Madagascar qu’après l’arrivée de
l’Homme, la composition floristique entre les régions malgaches varierait peu. Autrement dit, de
similaires taxa devraient se rencontrer dans tout Madagascar. Toutefois, il a été découvert que les
Poaceae dans deux écorégions malgaches démontrent une dissimilarité et que la composition en
taxa varie beaucoup d’une écorégion à une autre (Vorontsova et al., 2016). C’est le
‘régionalisme’, et les Poaceae aux environs de l’Isalo ont été inclus dans cette considération.
Pour Isalo, la composition taxonomique en Poaceae démontre presque zéro affinité avec celle de
l’Andringitra (Lewis et al., 1996), par exemple, là où les C3 et les Pooideae sont abondants.
Ainsi, le cas de l’Isalo est en accord avec un fort régionalisme à Madagascar, donc à une origine
ancienne probablement.
La dynamique des savanes est maintenue par le feu, et cela représentait aussi un prétexte
pour juger une origine récente et non naturelle de ces biomes. Cependant, les feux ne sont pas
toujours d’origine anthropique mais peuvent être aussi générés par des phénomènes naturels
comme les tonnerres. Et il faut rappeler que des feux naturels ont survenu maintes fois avant
l’arrivée de l’Homme à Madagascar (Burney,1987b, 1997 ; Gasse & Van Campo, 1998). En
outre, le type de climat aride et la saison sèche prolongée favorisent naturellement l’expansion
des feux, et donc favorisent les savanes aux forêts. Durant cette saison, les biomasses cumulées
des savanes deviennent sèches et sensiblement inflammables. De tel scenario se passe surtout
dans le centre et centre-ouest de Madagascar, incluant Isalo, là où la saison sèche dure six mois
ou plus et où une proportion considérable de la végétation est constituée par des savanes. Il se
trouve ainsi que des paramètres naturels assurent la dynamique de ces écosystèmes herbeux sans
que l’Homme intervienne. Ainsi, il se peut que les savanes se répandent dans cette partie de l’ile
avant l’arrivée de l’Homme.

45
Historiquement, l’Isalo a été habité primordialement par des Bara. Ils étaient jadis semi-
nomadiques pour la quête de meilleur pâturage pour leurs troupeaux (Rajaofera, 2007 ;
Zafisoalizy, 2011). Ils s’installassent définitivement à Isalo puisqu’ils y trouvèrent satisfaction
dans leur quête. Cela indique que les savanes existaient déjà avant leur arrivée. Leur mode de vie
traditionnelle peut également refléter cela. Les Bara vivent en dépit des savanes plutôt qu’aux
arbres et aux forêts. Ils brulent régulièrement les savanes pour maintenir le pâturage, et leurs
maisons sont faits de l’argile mais pas de bois, et de graminées pour le toit (Rajaofera, 2007 ;
Zafisoalizy, 2011). Cette ethnie, connue comme bons éleveurs mais agriculteurs fainéants,
détruit moins les forêts pour l’agriculture. Une récente étude a même mentionné qu’elle est de
nature protecteur de la forêt (Saandi, 2012). Ces faits contrastent plus ou moins la présence de
forêts denses, dits naturelles, à Isalo avant l’arrivée de l’Homme (les Bara). De plus, Isalo est très
étendue et a été toujours moins peuplé, du coup il est difficile d’imaginer qu’une grande surface
de forêts ait pu être détruite pour aboutir à ces vastes terrains de savanes.
Il faut rappeler qu’une savane est dite naturelle quand sa présence et son développement
ont eu lieu avant l’arrivée de l’Homme, selon sa définition la plus simple (Veldman et al., 2015 ;
Veldman, 2016). A partir de ces lignes d’évidences citées ci-dessus, il est probable qu’il existait
déjà de savanes à Isalo avant que l’Homme y débarque, mais on ne peut savoir exactement la
proportion et ni leur localisation. Mais l’étude sur l’origine des savanes est un sujet très
complexe, doit être bien approfondie et donc considérant tous disciplines susceptibles.

IV. CONCLUSION

46
La présente étude a traité un sujet concernant l’une des familles globalement riches en
espèces, les Poaceae, mais qui est encore négligée par la science à Madagascar. Dans le but
d’enrichir les connaissances sur la diversité des Poaceae et de la flore malgache, la
documentation des Poaceae de l’Isalo a été effectuée. Comprendre les complexes et minuscules
pièces florales des Poaceae constituent l’un des barrières qui empêchent beaucoup de chercheurs
à les étudier, du coup une description morphologique simple et complète de la famille des
Poaceae a été avancée en premier lieu.
Cette recherche a été réalisée dans le parc d’Isalo, pour lequel une liste des taxa de
Poaceae devrait être établie. En se basant sur les données issues des bases de données
préexistantes sur les Poaceae et celles issues de nos inventaires sur terrain, nous avons listé 112
espèces de Poaceae à Isalo. Ces effectifs ont été obtenus à partir des collectes réalisées dans la
partie Est et Sud du parc seulement, et la partie Nord et Ouest du parc devraient être investiguées
dans le futur afin de compléter cette liste. Nous avons proposé une clé de détermination des
genres et des illustrations de quelques espèces afin d’encourager et de faciliter les recherches sur
ce groupe dorénavant.
Outre la liste des espèces, des analyses sur la diversité des espèces, de leur endémicité et
de leur distribution dans le parc ont été avancées. Isalo abrite 112 espèces de Poaceae
appartenant à 58 genres, 16 tribus et sept sous familles. Parmi ces espèces, 25 (22%) sont
endémiques de Madagascar dont trois espèces et une variété sont retreintes à Isalo. Les cartes de
distribution de ces espèces endémiques restreintes sont aussi présentées dans cette étude pour les
futures recherches sur ces espèces. A partir des points d’occurrence de chaque espèce de Poaceae
dans le parc, les zones où il y a des canyons sont présents, disposent la plus grande diversité
spécifique, dont Namaza et Ranohira. Ces études sont de natures descriptives et peuvent servir de
base pour les futures analyses écologiques plus approfondies.
Cette étude a également fourni des données pour appuyer les futures recherches sur
l’origine des savanes de l’Isalo ou encore celles des hauts plateaux. D’après les synthèses des
informations liées à la diversité des organismes dans ce type de végétation et les relations
historiques entre Isalo et ses premiers occupants, nous penchons vers une origine ancienne et

47
naturelle des savanes de l’Isalo. Toutefois, nous n’avons fait que de rassembler la moindre des
données nécessaires pour traiter ce sujet. Ainsi, de sérieuses analyses sont indispensablement
requises si on veut aboutir à une conclusion.
En bref cette étude a permis de documenter les Poaceae de l’Isalo et de fournir des
données de base pour les futures recherches relatant les Poaceae de l’Isalo et/ou les Poaceae
Madagascar.

V. BIBLIOGRAPHIE

48
ANDERSON, A., CLARK, G., HABERLE, S., HIGHAM, T., NOWAK-KEMP, M.,
PRENDERGAST, A., ET AL. (2018) New evidence of megafaunal bone damage
indicates late colonization of Madagascar. PLoS ONE 13: e0204368. doi:
10.1371/journal.pone.0204368
BOND, W. J., SILANDER, J. A., RANAIVONASY, J. & RATSIRARSON, J. 2008. The
antiquity of Madagascar’s grassland and the rise of C4 grassy biomes. Journal of
Biogeography (J. Biogeogr.) 35 : 1743-1758.
BOSSER, J. 1969. Graminées des pâturages et de culture. Mémoire O.R.S.T.O.M. Paris. 440p.
BROWN, J.H. & LOMOLINO, M.V. 1998. Biogeography. Sinauer Associates, INC. 2nd
Edition, 692 p.
BURNEY, D. A. 1987b. Late Quaternary stratigraphic charcoal records from Madagascar.
Quaternary Research 28: 274- 280.
BURNEY, D. A. 1997. Theories and facts regarding Holocene environmental change before and
after human colonization. In: Goodman S. M. & Patterson B. D. (Eds.), Natural change
and human impact in Madagascar. Washington, D.C.: Smithsonian Institution Press.
75–89.
CALLMANDER, M. W., PHILLIPSON, P. B., SCHATZ, G. E., ANDRIAMBOLOLONERA,
S., RABARIMANARIVO, M., RAKOTONIRINA, N., RAHARIMAMPIONONA, J.,
CHATELAIN, C., GAUTIER, L. & LOWRY, P. P. 2011. The endemic and non-
endemic vascular flora of Madagascar updated. Plant Ecology and Evolution 144: 121-
125.
CHRISTIN P.A. & OSBORNE C. P. 2014. Review, The evolutionary ecology of C4 plants. New
Physiology: 765-781.
CLAYTON, W. D. & RENVOIZE, S. A. 1982. Flora of tropical East Africa. Gramineae. Part 3.
AA Balkema, Rotterdam. 448p.
CLAYTON, W. D., PHILLIPS, S. M. & RENVOIZE, S. A. 1974. Flora of tropical East Africa:
Gramineae. Part 2. Crown Agents for Overseas Governments and Administrations,
London. 449p.

49
CLAYTON, W. D. 1970. Flora of tropical East Africa: Gramineae. Part 1. Crown Agents for
Overseas Governments and Administrations, London. 176p.
DAVIDSE, G., SOUSA, S. M. & CHATER, A. 1994. Flora Mesoamericana 6: Alismataceae to
Cyperaceae. UNAM, Mexico City, México. 430p.
FARAMALALA, M. H. 1988. Etude de la Végétation de Madagascar à l’aide des données
spatiales. Université Paul Sabatier, Toulouse, France.
PHILLIPSON, P. B., ANDRIAMBOLOLONERA, S., LOWRYII, P. P., MANJATO, N.,
RABARIMANARIVO, M., RAKOTONIRINA, N., RAVOLOLMANANA, N. &
SCHATZ, G. E, 2017. The Madagascar Catalogue, progress to date and prospects for the
future. Abstracts, XXI AETFAT Congress, Nairobi, Kenya, 241 p.
GAUSSEN, A. 1955. Détermination des climats par méthodes des courbes ombrothermiques.
Académie des Sciences et techniques 240: 642-643.
GASSE, F. AND VAN CAMPO, E. 1998. A 40,000-yr pollen and diatom record for Lake
Tritrivakely, Madagascar, in the southern tropics. Quaternary Research 49 : 299-311.
GPWG II – Grass Phylogeny Working Group II. 2012. New grass phylogeny resolves deep
evolutionary relationships and discovers C4 origins. New Phytologist 193: 304–312.
GOODMAN, S. M., RAHERILALAO, M. J. AND WOHLHAUSER, S. 2018. The terrestrial
protected areas of Madagascar: Their history, description and biota. Association
Vahatra, Antananarivo.
HACKEL, J., VORONTSOVA, M. S., NANJARISOA, O. P., HALL, R. C., RAZANATSOA,
J., MALAKASI, P. et BESNARD, G. 2018. Grass diversification in Madagascar: in situ
radiation of two large C3 shade clades and support for a Miocene to Pliocene origin of
C4 grassy biomes. Journal of Biogeography 45: 750-761.
http://dx.doi.org/10.1111/jbi.13147
HAGL, P. A., GARGIULO, R., FAY, M., SOLOFONDRANOHATRA, S. L.,
RAKOTOMALALA, N. H., LEHMANN , C. E. R., GESNARD, G, PAPADOPULOS,
S. T., & VORONTSOVA, M. S. Geographic structure of genetic diversity within the
Loudetia simplex complex in Africa and Madagascar. Botanical Journal of the Linnean
Society, in review.
HUMBERT, H. 1927. Destruction d’une flore insulaire par le feu : principaux aspects de la
végétation à Madagascar. Mem. Acad. Malgache 5: 1-80.

50
KELLOG, E. 2015. Flowering plants monocots. Poaceae. In: Kubitizki K. (ed), The families and
Genera of Vascular Plants, Volume XII. Berlin: Springer Verlog. 1-413.
KOECHLIN, J., GUILLAUMET J.L. et MORAT, P. 1974. Flore et végétation de Madagascar.
A.R.G. GantnerVerlog FL 9490 Vaduz. 687 p.
LOWRY, P. P, SCHATZ, G. E., AND PHILLIPSON, P. B. 1997. The Classification of Natural
and anthropic Vegetation in Madagascar. In: Goodman S. M. & Patterson B. D. (eds.).
Natural change and human impact in Madagascar. Smithsonian Institution Press,
Washington D. C. 113–114.
MADAGASCAR NATIONAL PARKS. 2017. Plan d’aménagement et de Gestion. Plan
quinquennal de mise en oeuvre 2017-2021 : Parc National d’Isalo. Régions Ihorombe et
Atsimo Andrefana. Madagascar. Antananarivo. 94 p.
MOAT, J. ET SMITH, P. 2007. Atlas de la végétation de Madagascar. Royal Botanic Gardens,
Kew. 124pp.
MORAT, P. 1973. Les savanes du Sud-Ouest de Madagascar. Mémoire O.R.S.T.O.M. Paris.
235pp.
NANJARISOA, O. P., BESNARD, G., RALIMANANA, H., JEANNODA, V. et
VORONTSOVA, M. S. 2017. Grass survey of the Itremo Massif records endemic central
highland grasses. Madag. Conserv. Dev. 12 : 34-40. doi:10.4314/mcd.v12i1.6
PERRIER DE LA BATHIE, H. 1921. La végétation malgache. Annales du Musée colonial de
Marseille, 3ème ser. 9 : 1-266.
RABARIVOLA, M. L., RAZANAJATOVO, H., RAZAFINIARY, V., RASOLOHERY, A.,
RALIMANANA, H. & VORONTSOVA, M., 2019d. Madagascar Grass Atlas. Kew
Madagascar Conservation Centre, Madagascar, 189 pp.
RABEALAINA, B. B. 2010. Etude comparative des lignées doubles haploïdes du riz (Oryza
sativa L.) avec la variété parent. DEA en Biologie et Ecologie Végétales – Option
Physiologie Végétale. Faculté des Sciences Antananarivo. 46p.
RAHARIMANANA, V. 2007. Contribution à l’étude de la conservation des ressources
génétiques pour leur utilisation durable : Cas de Zea mays L. à Madagascar. DEA des
sciences Agronomiques Département « Agriculture ». Université d’Antananarivo. 57p.
RAJAOFERA, C. 2007. Contribution à l’histoire des Zafimanely, l’exemple du royaume
Barabe, d1e sa création au début de la période coloniale. Formation Doctorale

51
Multidisciplinaire. Université de Toliara, Madagascar. < http://biblio.univ-
ntananarivo.mg/pdfs/rajaoferaChristophe_FR_M2_07.pdf>
RAKOTOARIMANANA, V. 2008. Feu, pâturage et dynamique des savanes à Madagascar.
Habilitation à Diriger des recherches en Biologie et Ecologie Végétale. Unpubl. Ph. D.
thesis, Université Antananarivo, Madagascar.
RAKOTONJANAHARY, X. 1993. Les populations de riz des Hautes terres Centrales et du lac
Alaotra : caractérisation, évaluation, utilisation (Oryza sativa L.). Thèse d’Ecologie et
Physiologie Végétale Appliquée. L’Etablissement d’Enseignement Supérieur des
Sciences. Université d’Antananarivo. 132p.
SAANDI, Y.M. 2012. Dynamique de gestion des ressources naturelles dans le sud-ouest
malgache : cas du Parc National Zombitse Vohibasia et ses zones périphériques. Diplôme
d’Etudes Approfondies. Université de Toliara, Madagascar. Http://biblio.univ-
antananarivo.mg/pdfs/saandiYousoufM_GEO_DNR_12.pdf
SOLOFONDRANOHATRA, C., VORONTSOVA, M. S., HACKEL, J., BESNARD, G.,
CABLE, S., JEANNODA, V., & LEHMANN, C. E. R. 2018. Grass functional traits
differentiate forest and savannah in the Madagascar central highlands. Frontiers of
Ecology and Evolution: 1-20.
SORENG, R. J., PETERSON, P. M., ROMASHENKO, K., DAVIDSE, G., TEISHER, J. K.,
CLARK, L. G., BERBERA, P. et al.,2017. A worldwide phylogenetic classification of
the Poaceae (Gramineae) II: An update and a comparison of two 2015 classifications.
Journal of Systematics and Evolution 55: 259-290.
SORENG, R. J., PETERSON, P. M., ROMASHENKO, K., DAVIDSE, G., ZULOAGA, F. O.,
JUDZIEWICZ, E. J. et al., 2015. A worldwide phylogenetic classification of the Poaceae
(Gramineae). Journal of Systematics and Evolution 53: 117-137.
SWIERKOSZ, K. 2007. Vegetation of the southern part of the Isalo Sandstone Massif (Central
Madagascar, Africa) – differentiation and threats. In: Migón P. & Kasprzak M. (eds)
Sandstone Landscapes. Poland: Museum of Natural History. 158 – 160.
VELDMAN, J. W. 2016. Clarifying the confusion: old-growth savannahs and tropical ecosystem
degradation. Phil. Trans. R. Soc. B 371 : 20150306.

52
VELDMAN, J. W., BUISON, E., DURIGAN, G., FERNANDES; W., LE STRADIC, S. ET AL.,
2015. Toward an old-growth concept for grasslands, savannas, and woodlands. Front.
Ecology and Evolution 13 (3): 154-162.
VOLLESEN, K., ABDALLAH, R., COE, M. AND MBOYA, E. 1999. Checklist: vascular plants
and pteridophytes of Mkomazi. In: Coe M. (ed.), Mkomazi: The Ecology, Biodiversity and
Conservation of a Tanzanian Savanna. London: Royal Geographical Society. 81–116.
VORONTSOVA, M. S., DRANSFIELD, S., RENVOIZE, S., BESNARD, G., MCROBB, A.,
RAZANATSOA, J. & RALIMANANA, H. 2018. Identification guide to grasses and
bamboos in Madagascar. 169p.
VORONTSOVA, M. S., BESNARD, G., FOREST, F., MALAKASI, P., MOAT, J., CLAYTON,
W. D., et al.,2016. Madagascar's grasses and grasslands: anthropogenic or natural? Proc.
R. Soc. B Biol. Sci. 283: 20152. http://dx.doi.org/ 10.1098/rspb.2015.2262.
VORONTSOVA, M. S., CLAYTON, D., BRYAN, K. S. 2015. Grassroots e-floras in the
Poaceae: growing GrassBase and Grass World. PhytoKeys 48: 73–84.
VORONTSOVA, M. S. & RAKOTOARISOA, S. E. 2014. Endemic non-bambusoid genera of
grasses (Poaceae) in Madagascar: Review of current knowledge. Malagasy Nature 8: 14-
34.
VORONTSOVA MS, RATOVONIRINA G, RANDRIAMBOAVONJY T. 2013. Revision of
Andropogon and Diectomis (Poaceae: Sacchareae) in Madagascar and the new
Andropogon itremoensis from the Itremo Massif. Kew Bulletin 68(2): 193-207.
WILLIAMS, E., NTANDU, J. E., FICINSKI, S. AND VORONTSOVA, M. S. 2016. Checklist
of Serengeti Ecosystem Grasses. Biodiversity Data Journal 4: e8286.
ZAFISOALIZY, M. 2011. La société Bara et les feux de brousses (L’exemple du district de
Sakaraha). Diplôme d’Etudes Approfondies de Sciences Sociales. Université de Toliara,
Madagascar. Http://biblio.univ-
antananarivo.mg/pdfs/zafisoalizyMonica_GEO_M2_11.pdf

53
WEBOGRAPHIE :
Http1 : Madagascar catalogue 2017 < http://legacy.tropicos.org/Project/Madagascar> [Site
consulté le 5 Janvier 2018]
Http2 : Grassbase, The Online World Grass Flora (2015), Royal Botanic Gardens Kew. URL :
http://www.kew.org/data/grasses-db/sppindex.htm/ [site visité le 4 Octobre 2018]
Http3: Botanical Research And Herbarium Management System (BRAHMS) (2014):
https://herbaria.plants.ox.ac.uk/bol/BRAHMS/ [Site visité le 26 Janvier 2019 ]
Http4 : Global Biodiversity Information Facility (2018). URL : https ://www.gbif.org/ [Site
visité le 6 Décembre 2018]
Http5: Plants Of The World, Kew Sciences (POWO) (2016). URL :
http://www.plantsoftheworldonline.org/ [site visité le 19 juin 2019]

ANNEXES

I
ANNEXE I: PHOTOS DES TYPES DE VEGETATIONS DE L’ISALO. A : FORETS GALERIES DE SAKAMALIO. B : MARAIS
DE PANDANUS AUX ENVIRONS DE LA PISCINE NATURELLE DE L’ISALO. C : SAVANES ET TAPIA LE LONG DU
CIRCUIT MALASO. D : VEGETATION RUPICOLE DE L’ISALO.
A B
C D
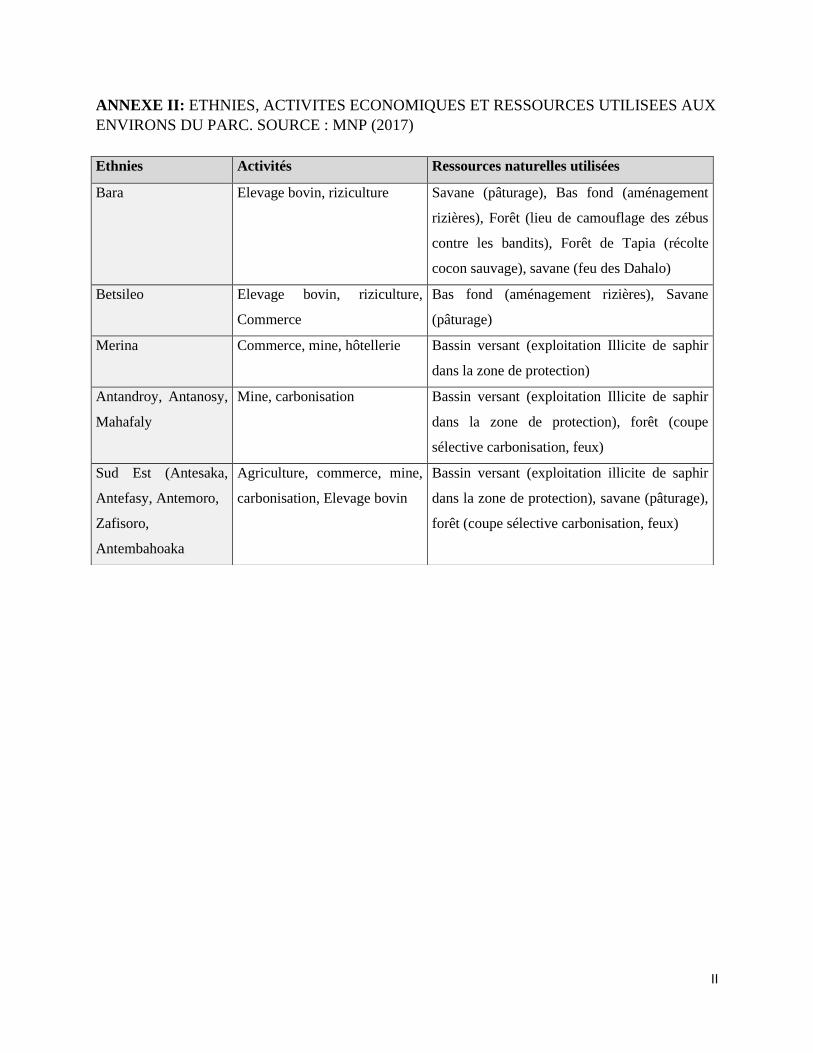
II
ANNEXE II: ETHNIES, ACTIVITES ECONOMIQUES ET RESSOURCES UTILISEES AUX
ENVIRONS DU PARC. SOURCE : MNP (2017)
Ethnies Activités Ressources naturelles utilisées
Bara Elevage bovin, riziculture Savane (pâturage), Bas fond (aménagement
rizières), Forêt (lieu de camouflage des zébus
contre les bandits), Forêt de Tapia (récolte
cocon sauvage), savane (feu des Dahalo)
Betsileo Elevage bovin, riziculture,
Commerce
Bas fond (aménagement rizières), Savane
(pâturage)
Merina Commerce, mine, hôtellerie Bassin versant (exploitation Illicite de saphir
dans la zone de protection)
Antandroy, Antanosy,
Mahafaly
Mine, carbonisation Bassin versant (exploitation Illicite de saphir
dans la zone de protection), forêt (coupe
sélective carbonisation, feux)
Sud Est (Antesaka,
Antefasy, Antemoro,
Zafisoro,
Antembahoaka
Agriculture, commerce, mine,
carbonisation, Elevage bovin
Bassin versant (exploitation illicite de saphir
dans la zone de protection), savane (pâturage),
forêt (coupe sélective carbonisation, feux)
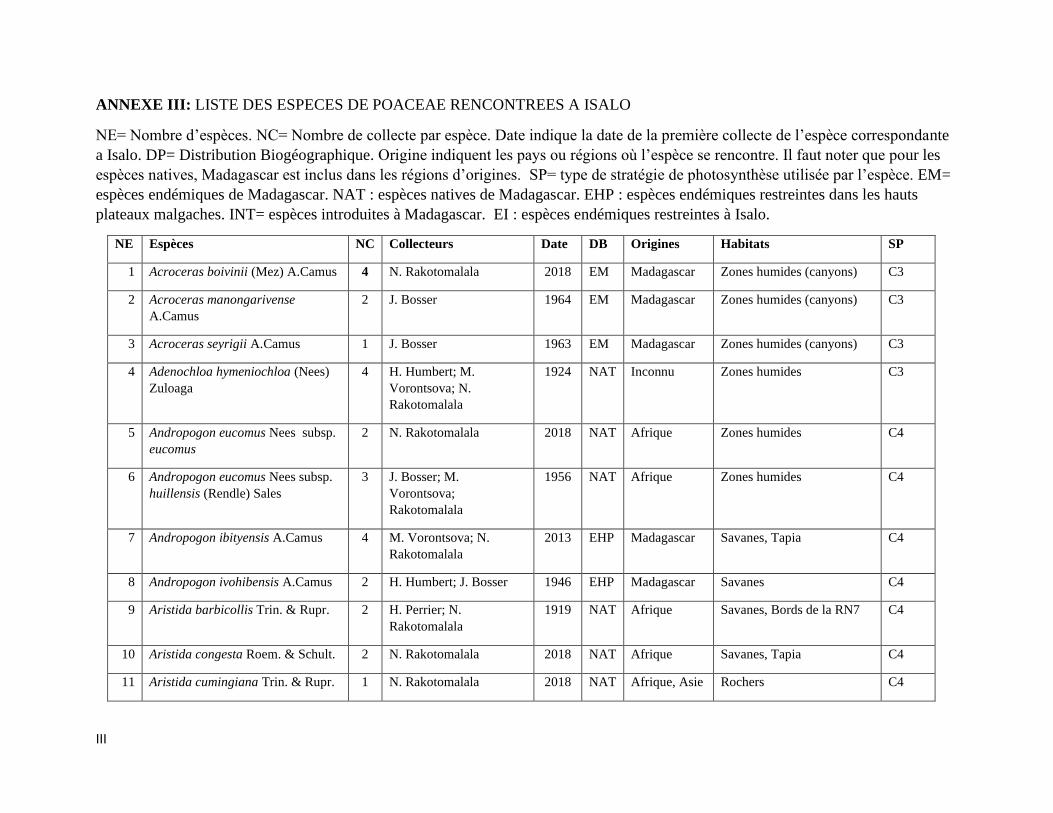
III
ANNEXE III: LISTE DES ESPECES DE POACEAE RENCONTREES A ISALO
NE= Nombre d’espèces. NC= Nombre de collecte par espèce. Date indique la date de la première collecte de l’espèce correspondante
a Isalo. DP= Distribution Biogéographique. Origine indiquent les pays ou régions où l’espèce se rencontre. Il faut noter que pour les
espèces natives, Madagascar est inclus dans les régions d’origines. SP= type de stratégie de photosynthèse utilisée par l’espèce. EM=
espèces endémiques de Madagascar. NAT : espèces natives de Madagascar. EHP : espèces endémiques restreintes dans les hauts
plateaux malgaches. INT= espèces introduites à Madagascar. EI : espèces endémiques restreintes à Isalo.
NE Espèces NC Collecteurs Date DB Origines Habitats SP
1 Acroceras boivinii (Mez) A.Camus 4 N. Rakotomalala 2018 EM Madagascar Zones humides (canyons) C3
2 Acroceras manongarivense
A.Camus
2 J. Bosser 1964 EM Madagascar Zones humides (canyons) C3
3 Acroceras seyrigii A.Camus 1 J. Bosser 1963 EM Madagascar Zones humides (canyons) C3
4 Adenochloa hymeniochloa (Nees)
Zuloaga
4 H. Humbert; M.
Vorontsova; N.
Rakotomalala
1924 NAT Inconnu Zones humides C3
5 Andropogon eucomus Nees subsp.
eucomus
2 N. Rakotomalala 2018 NAT Afrique Zones humides C4
6 Andropogon eucomus Nees subsp.
huillensis (Rendle) Sales
3 J. Bosser; M.
Vorontsova;
Rakotomalala
1956 NAT Afrique Zones humides C4
7 Andropogon ibityensis A.Camus 4 M. Vorontsova; N.
Rakotomalala
2013 EHP Madagascar Savanes, Tapia C4
8 Andropogon ivohibensis A.Camus 2 H. Humbert; J. Bosser 1946 EHP Madagascar Savanes C4
9 Aristida barbicollis Trin. & Rupr. 2 H. Perrier; N.
Rakotomalala
1919 NAT Afrique Savanes, Bords de la RN7 C4
10 Aristida congesta Roem. & Schult. 2 N. Rakotomalala 2018 NAT Afrique Savanes, Tapia C4
11 Aristida cumingiana Trin. & Rupr. 1 N. Rakotomalala 2018 NAT Afrique, Asie Rochers C4
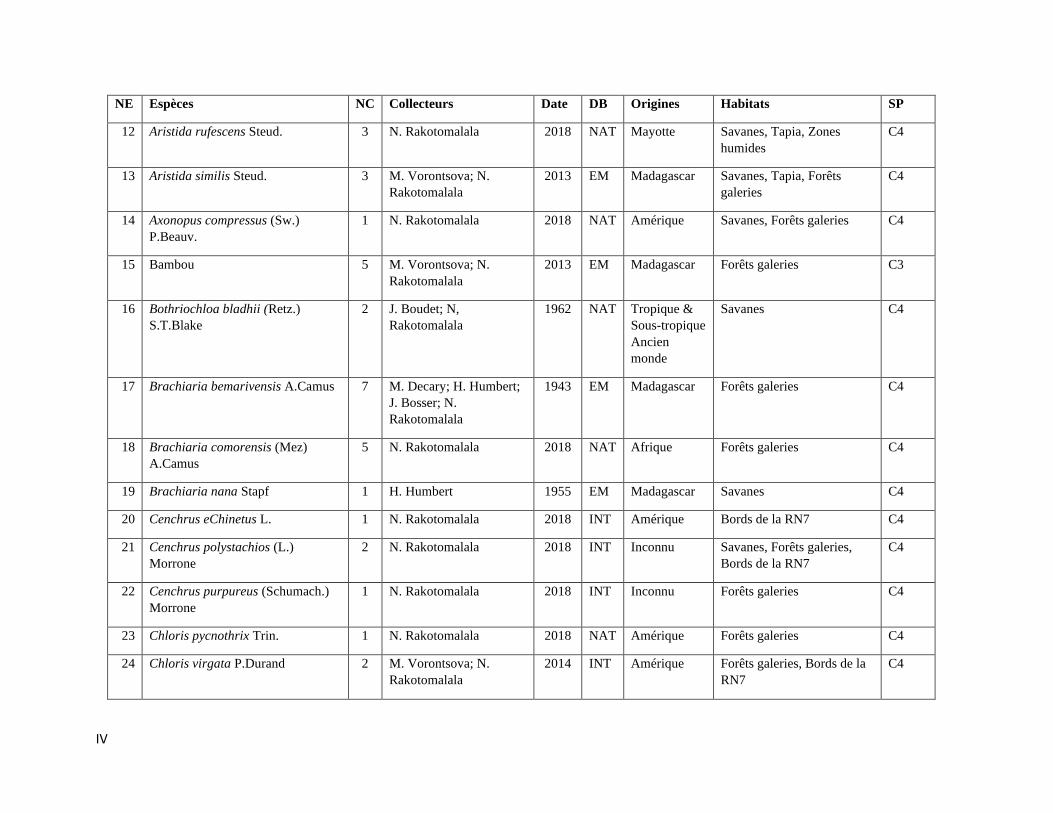
IV
NE Espèces NC Collecteurs Date DB Origines Habitats SP
12 Aristida rufescens Steud. 3 N. Rakotomalala 2018 NAT Mayotte Savanes, Tapia, Zones
humides
C4
13 Aristida similis Steud. 3 M. Vorontsova; N.
Rakotomalala
2013 EM Madagascar Savanes, Tapia, Forêts
galeries
C4
14 Axonopus compressus (Sw.)
P.Beauv.
1 N. Rakotomalala 2018 NAT Amérique Savanes, Forêts galeries C4
15 Bambou 5 M. Vorontsova; N.
Rakotomalala
2013 EM Madagascar Forêts galeries C3
16 Bothriochloa bladhii (Retz.)
S.T.Blake
2 J. Boudet; N,
Rakotomalala
1962 NAT Tropique &
Sous-tropique
Ancien
monde
Savanes C4
17 Brachiaria bemarivensis A.Camus 7 M. Decary; H. Humbert;
J. Bosser; N.
Rakotomalala
1943 EM Madagascar Forêts galeries C4
18 Brachiaria comorensis (Mez)
A.Camus
5 N. Rakotomalala 2018 NAT Afrique Forêts galeries C4
19 Brachiaria nana Stapf 1 H. Humbert 1955 EM Madagascar Savanes C4
20 Cenchrus eChinetus L. 1 N. Rakotomalala 2018 INT Amérique Bords de la RN7 C4
21 Cenchrus polystachios (L.)
Morrone
2 N. Rakotomalala 2018 INT Inconnu Savanes, Forêts galeries,
Bords de la RN7
C4
22 Cenchrus purpureus (Schumach.)
Morrone
1 N. Rakotomalala 2018 INT Inconnu Forêts galeries C4
23 Chloris pycnothrix Trin. 1 N. Rakotomalala 2018 NAT Amérique Forêts galeries C4
24 Chloris virgata P.Durand 2 M. Vorontsova; N.
Rakotomalala
2014 INT Amérique Forêts galeries, Bords de la
RN7
C4

V
NE Espèces NC Collecteurs Date DB Origines Habitats SP
25 Chrysopogon serrulatus Trin. 16 R. Decary; J. Bosser; P.
Morat; M. Vorontsova;
N. Rakotomalala
1940 NAT Afrique,
Asie,
Nouveau
monde
Savanes, Tapia C4
26 Coelachne afriana Pilg. 3 M. Decary; M.
Vorontsova; N.
Rakotomalala
1941 NAT Afrique, Inde Forêts galeries C3
27 Craspedorhachis africana Benth. 34 G. Cours; H. Humbert;
M. Peltier; J. Bosser; T.
Croat; N. Labat; M.
Vorontsova; N.
Rakotomalala
1955 NAT Afrique,
Europe
Savanes, rochers, Tapia,
Bords de la RN7
C4
28 Cymbopogon caesius (Hook. &
Arn.) Stapf
4 H. Humbert; J. Bosser;
N. Rakotomalala
1955 NAT Afrique, Inde
Arabie
Savanes, Forêts galeries,
forets sèches
C4
29 Cynodon dactylon (L.) Pers. 5 H. Humbert; J. Bosser;
N. Rakotomalala
1956 NAT Ancien
monde,
Australie
Forêts galeries, Bords de la
RN7
C4
30 Cyrtococcum deltoideum A.Camus 3 N. Rakotomalala 2018 EM Madagascar Forêts galeries C3
31 Cyrtococcum multinode (Lam.)
Clayton
1 N. Rakotomalala 2018 NAT Afrique Forêts galeries C3
32 Dactyloctenium aegyptium (L.)
Willd.
3 P. Morat; N.
Rakotomalala
1965 NAT Ancien
monde
Forêts galeries C4
33 Digitaria bicornis (Lam.) Roem. &
Schult.
2 G. Cours; M. Vorontsova 1955 NAT Inde, Asie,
Australie
Forêts galeries C4
34 Digitaria ciliaris (Retz.) Koeler 2 N. Rakotomalala 2018 NAT Ancien
monde
Tapia, Bords de la RN7 C4
35 Digitaria debilis (Desf.) Willd. 1 N. Rakotomalala 2018 NAT Afrique,
Europe
Tapia C4
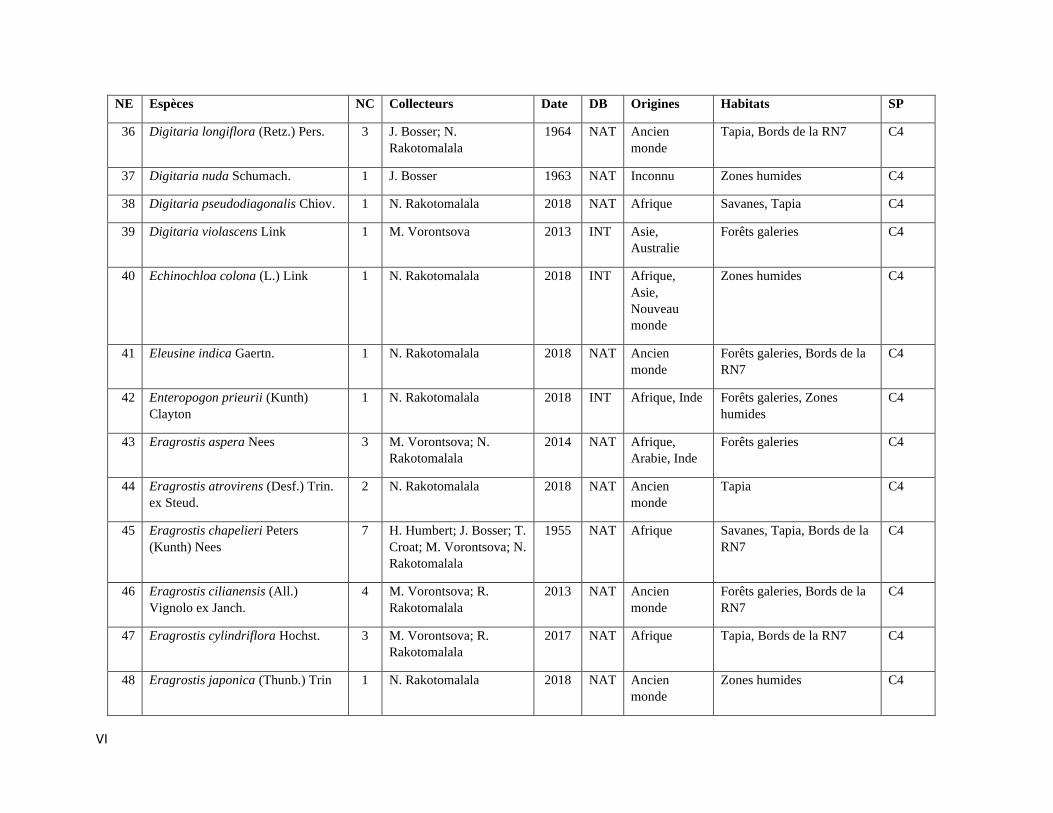
VI
NE Espèces NC Collecteurs Date DB Origines Habitats SP
36 Digitaria longiflora (Retz.) Pers. 3 J. Bosser; N.
Rakotomalala
1964 NAT Ancien
monde
Tapia, Bords de la RN7 C4
37 Digitaria nuda Schumach. 1 J. Bosser 1963 NAT Inconnu Zones humides C4
38 Digitaria pseudodiagonalis Chiov. 1 N. Rakotomalala 2018 NAT Afrique Savanes, Tapia C4
39 Digitaria violascens Link 1 M. Vorontsova 2013 INT Asie,
Australie
Forêts galeries C4
40 Echinochloa colona (L.) Link 1 N. Rakotomalala 2018 INT Afrique,
Asie,
Nouveau
monde
Zones humides C4
41 Eleusine indica Gaertn. 1 N. Rakotomalala 2018 NAT Ancien
monde
Forêts galeries, Bords de la
RN7
C4
42 Enteropogon prieurii (Kunth)
Clayton
1 N. Rakotomalala 2018 INT Afrique, Inde Forêts galeries, Zones
humides
C4
43 Eragrostis aspera Nees 3 M. Vorontsova; N.
Rakotomalala
2014 NAT Afrique,
Arabie, Inde
Forêts galeries C4
44 Eragrostis atrovirens (Desf.) Trin.
ex Steud.
2 N. Rakotomalala 2018 NAT Ancien
monde
Tapia C4
45 Eragrostis chapelieri Peters
(Kunth) Nees
7 H. Humbert; J. Bosser; T.
Croat; M. Vorontsova; N.
Rakotomalala
1955 NAT Afrique Savanes, Tapia, Bords de la
RN7
C4
46 Eragrostis cilianensis (All.)
Vignolo ex Janch.
4 M. Vorontsova; R.
Rakotomalala
2013 NAT Ancien
monde
Forêts galeries, Bords de la
RN7
C4
47 Eragrostis cylindriflora Hochst. 3 M. Vorontsova; R.
Rakotomalala
2017 NAT Afrique Tapia, Bords de la RN7 C4
48 Eragrostis japonica (Thunb.) Trin 1 N. Rakotomalala 2018 NAT Ancien
monde
Zones humides C4
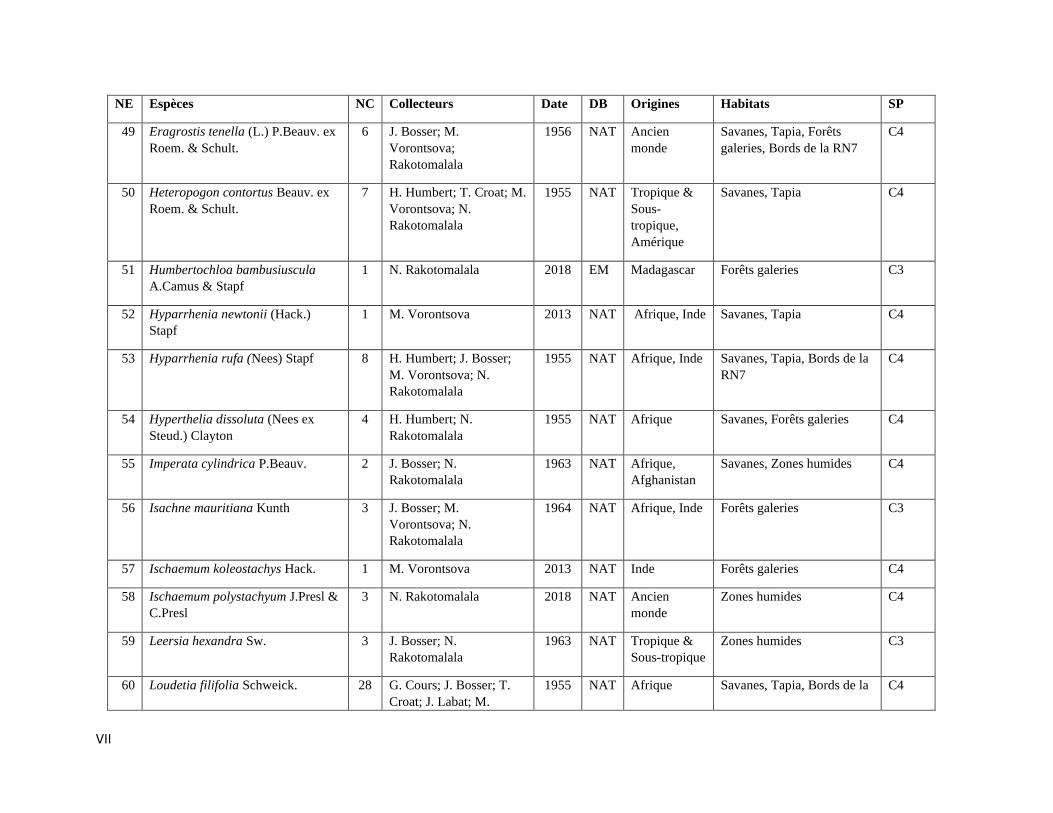
VII
NE Espèces NC Collecteurs Date DB Origines Habitats SP
49 Eragrostis tenella (L.) P.Beauv. ex
Roem. & Schult.
6 J. Bosser; M.
Vorontsova;
Rakotomalala
1956 NAT Ancien
monde
Savanes, Tapia, Forêts
galeries, Bords de la RN7
C4
50 Heteropogon contortus Beauv. ex
Roem. & Schult.
7 H. Humbert; T. Croat; M.
Vorontsova; N.
Rakotomalala
1955 NAT Tropique &
Sous-
tropique,
Amérique
Savanes, Tapia C4
51 Humbertochloa bambusiuscula
A.Camus & Stapf
1 N. Rakotomalala 2018 EM Madagascar Forêts galeries C3
52 Hyparrhenia newtonii (Hack.)
Stapf
1 M. Vorontsova 2013 NAT Afrique, Inde Savanes, Tapia C4
53 Hyparrhenia rufa (Nees) Stapf 8 H. Humbert; J. Bosser;
M. Vorontsova; N.
Rakotomalala
1955 NAT Afrique, Inde Savanes, Tapia, Bords de la
RN7
C4
54 Hyperthelia dissoluta (Nees ex
Steud.) Clayton
4 H. Humbert; N.
Rakotomalala
1955 NAT Afrique Savanes, Forêts galeries C4
55 Imperata cylindrica P.Beauv. 2 J. Bosser; N.
Rakotomalala
1963 NAT Afrique,
Afghanistan
Savanes, Zones humides C4
56 Isachne mauritiana Kunth 3 J. Bosser; M.
Vorontsova; N.
Rakotomalala
1964 NAT Afrique, Inde Forêts galeries C3
57 Ischaemum koleostachys Hack. 1 M. Vorontsova 2013 NAT Inde Forêts galeries C4
58 Ischaemum polystachyum J.Presl &
C.Presl
3 N. Rakotomalala 2018 NAT Ancien
monde
Zones humides C4
59 Leersia hexandra Sw. 3 J. Bosser; N.
Rakotomalala
1963 NAT Tropique &
Sous-tropique
Zones humides C3
60 Loudetia filifolia Schweick. 28 G. Cours; J. Bosser; T.
Croat; J. Labat; M.
1955 NAT Afrique Savanes, Tapia, Bords de la C4
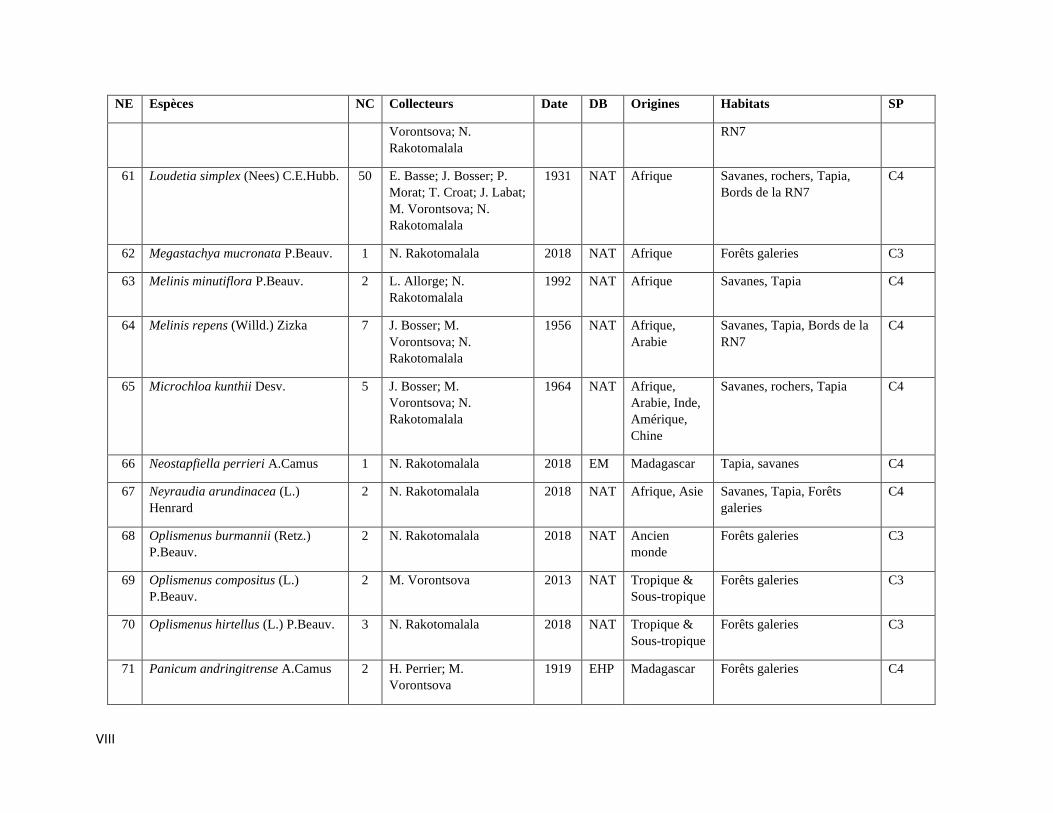
VIII
NE Espèces NC Collecteurs Date DB Origines Habitats SP
Vorontsova; N.
Rakotomalala
RN7
61 Loudetia simplex (Nees) C.E.Hubb. 50 E. Basse; J. Bosser; P.
Morat; T. Croat; J. Labat;
M. Vorontsova; N.
Rakotomalala
1931 NAT Afrique Savanes, rochers, Tapia,
Bords de la RN7
C4
62 Megastachya mucronata P.Beauv. 1 N. Rakotomalala 2018 NAT Afrique Forêts galeries C3
63 Melinis minutiflora P.Beauv. 2 L. Allorge; N.
Rakotomalala
1992 NAT Afrique Savanes, Tapia C4
64 Melinis repens (Willd.) Zizka 7 J. Bosser; M.
Vorontsova; N.
Rakotomalala
1956 NAT Afrique,
Arabie
Savanes, Tapia, Bords de la
RN7
C4
65 Microchloa kunthii Desv. 5 J. Bosser; M.
Vorontsova; N.
Rakotomalala
1964 NAT Afrique,
Arabie, Inde,
Amérique,
Chine
Savanes, rochers, Tapia C4
66 Neostapfiella perrieri A.Camus 1 N. Rakotomalala 2018 EM Madagascar Tapia, savanes C4
67 Neyraudia arundinacea (L.)
Henrard
2 N. Rakotomalala 2018 NAT Afrique, Asie Savanes, Tapia, Forêts
galeries
C4
68 Oplismenus burmannii (Retz.)
P.Beauv.
2 N. Rakotomalala 2018 NAT Ancien
monde
Forêts galeries C3
69 Oplismenus compositus (L.)
P.Beauv.
2 M. Vorontsova 2013 NAT Tropique &
Sous-tropique
Forêts galeries C3
70 Oplismenus hirtellus (L.) P.Beauv. 3 N. Rakotomalala 2018 NAT Tropique &
Sous-tropique
Forêts galeries C3
71 Panicum andringitrense A.Camus 2 H. Perrier; M.
Vorontsova
1919 EHP Madagascar Forêts galeries C4
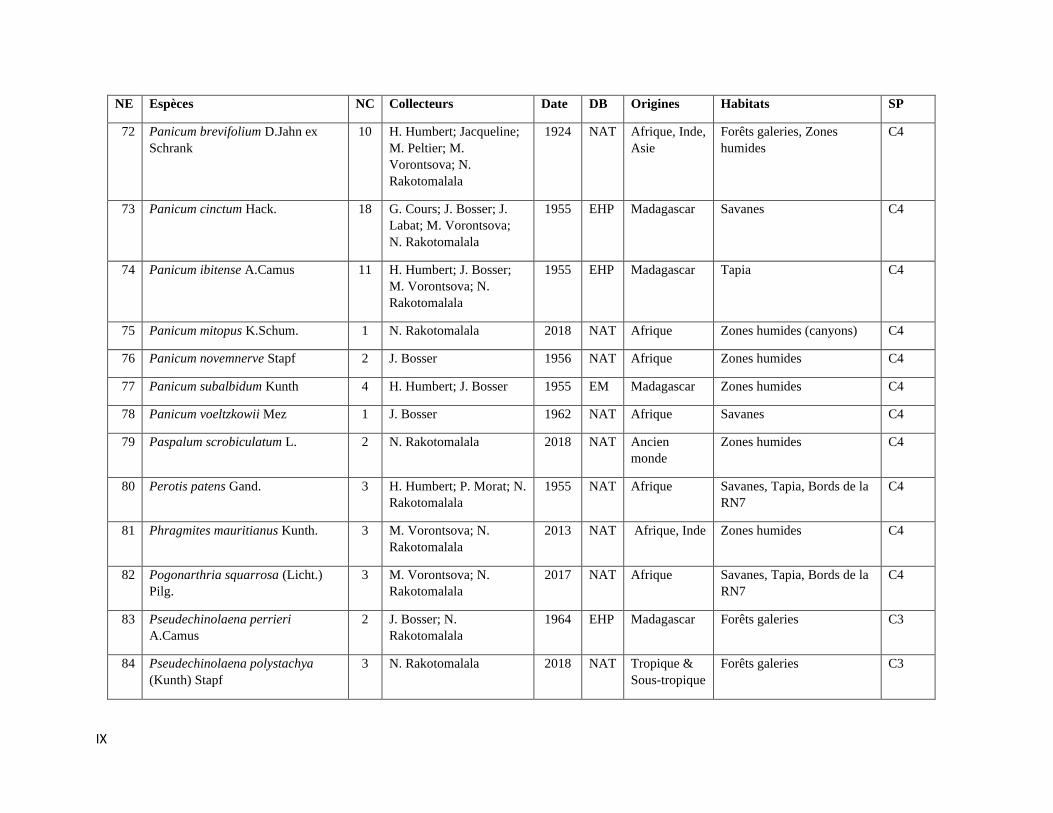
IX
NE Espèces NC Collecteurs Date DB Origines Habitats SP
72 Panicum brevifolium D.Jahn ex
Schrank
10 H. Humbert; Jacqueline;
M. Peltier; M.
Vorontsova; N.
Rakotomalala
1924 NAT Afrique, Inde,
Asie
Forêts galeries, Zones
humides
C4
73 Panicum cinctum Hack. 18 G. Cours; J. Bosser; J.
Labat; M. Vorontsova;
N. Rakotomalala
1955 EHP Madagascar Savanes C4
74 Panicum ibitense A.Camus 11 H. Humbert; J. Bosser;
M. Vorontsova; N.
Rakotomalala
1955 EHP Madagascar Tapia C4
75 Panicum mitopus K.Schum. 1 N. Rakotomalala 2018 NAT Afrique Zones humides (canyons) C4
76 Panicum novemnerve Stapf 2 J. Bosser 1956 NAT Afrique Zones humides C4
77 Panicum subalbidum Kunth 4 H. Humbert; J. Bosser 1955 EM Madagascar Zones humides C4
78 Panicum voeltzkowii Mez 1 J. Bosser 1962 NAT Afrique Savanes C4
79 Paspalum scrobiculatum L. 2 N. Rakotomalala 2018 NAT Ancien
monde
Zones humides C4
80 Perotis patens Gand. 3 H. Humbert; P. Morat; N.
Rakotomalala
1955 NAT Afrique Savanes, Tapia, Bords de la
RN7
C4
81 Phragmites mauritianus Kunth. 3 M. Vorontsova; N.
Rakotomalala
2013 NAT Afrique, Inde Zones humides C4
82 Pogonarthria squarrosa (Licht.)
Pilg.
3 M. Vorontsova; N.
Rakotomalala
2017 NAT Afrique Savanes, Tapia, Bords de la
RN7
C4
83 Pseudechinolaena perrieri
A.Camus
2 J. Bosser; N.
Rakotomalala
1964 EHP Madagascar Forêts galeries C3
84 Pseudechinolaena polystachya
(Kunth) Stapf
3 N. Rakotomalala 2018 NAT Tropique &
Sous-tropique
Forêts galeries C3
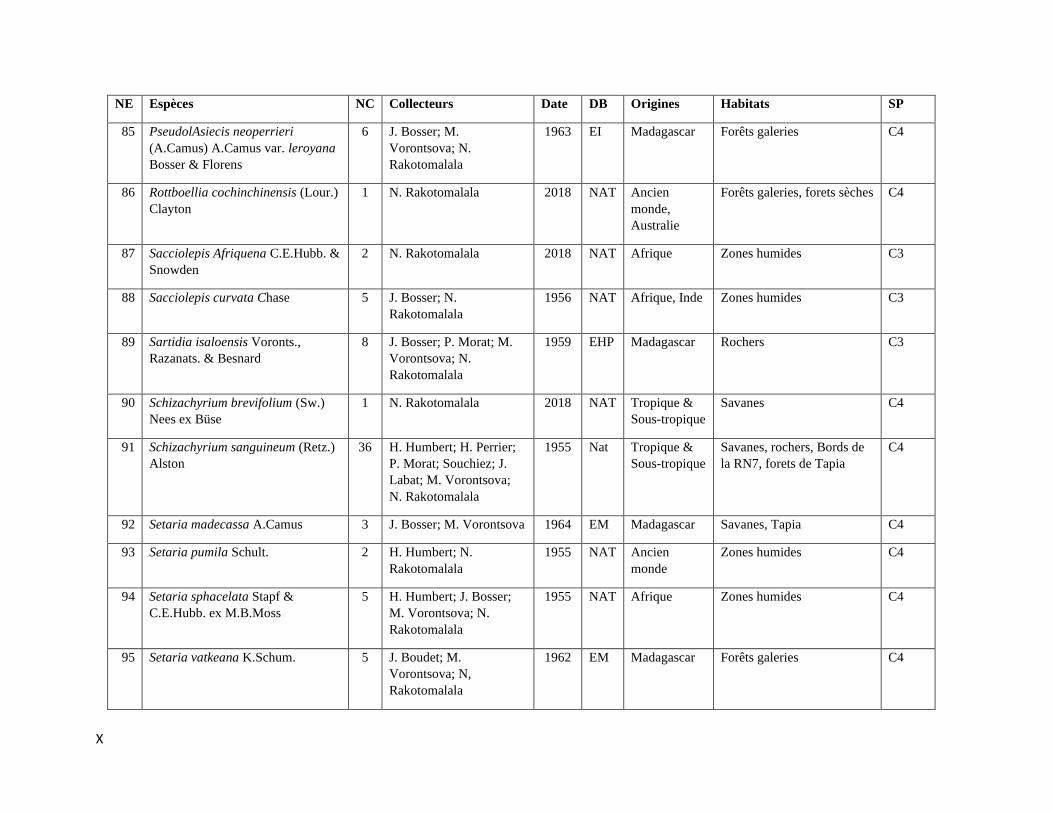
X
NE Espèces NC Collecteurs Date DB Origines Habitats SP
85 PseudolAsiecis neoperrieri
(A.Camus) A.Camus var. leroyana
Bosser & Florens
6 J. Bosser; M.
Vorontsova; N.
Rakotomalala
1963 EI Madagascar Forêts galeries C4
86 Rottboellia cochinchinensis (Lour.)
Clayton
1 N. Rakotomalala 2018 NAT Ancien
monde,
Australie
Forêts galeries, forets sèches C4
87 Sacciolepis Afriquena C.E.Hubb. &
Snowden
2 N. Rakotomalala 2018 NAT Afrique Zones humides C3
88 Sacciolepis curvata Chase 5 J. Bosser; N.
Rakotomalala
1956 NAT Afrique, Inde Zones humides C3
89 Sartidia isaloensis Voronts.,
Razanats. & Besnard
8 J. Bosser; P. Morat; M.
Vorontsova; N.
Rakotomalala
1959 EHP Madagascar Rochers C3
90 Schizachyrium brevifolium (Sw.)
Nees ex Büse
1 N. Rakotomalala 2018 NAT Tropique &
Sous-tropique
Savanes C4
91 Schizachyrium sanguineum (Retz.)
Alston
36 H. Humbert; H. Perrier;
P. Morat; Souchiez; J.
Labat; M. Vorontsova;
N. Rakotomalala
1955 Nat Tropique &
Sous-tropique
Savanes, rochers, Bords de
la RN7, forets de Tapia
C4
92 Setaria madecassa A.Camus 3 J. Bosser; M. Vorontsova 1964 EM Madagascar Savanes, Tapia C4
93 Setaria pumila Schult. 2 H. Humbert; N.
Rakotomalala
1955 NAT Ancien
monde
Zones humides C4
94 Setaria sphacelata Stapf &
C.E.Hubb. ex M.B.Moss
5 H. Humbert; J. Bosser;
M. Vorontsova; N.
Rakotomalala
1955 NAT Afrique Zones humides C4
95 Setaria vatkeana K.Schum. 5 J. Boudet; M.
Vorontsova; N,
Rakotomalala
1962 EM Madagascar Forêts galeries C4
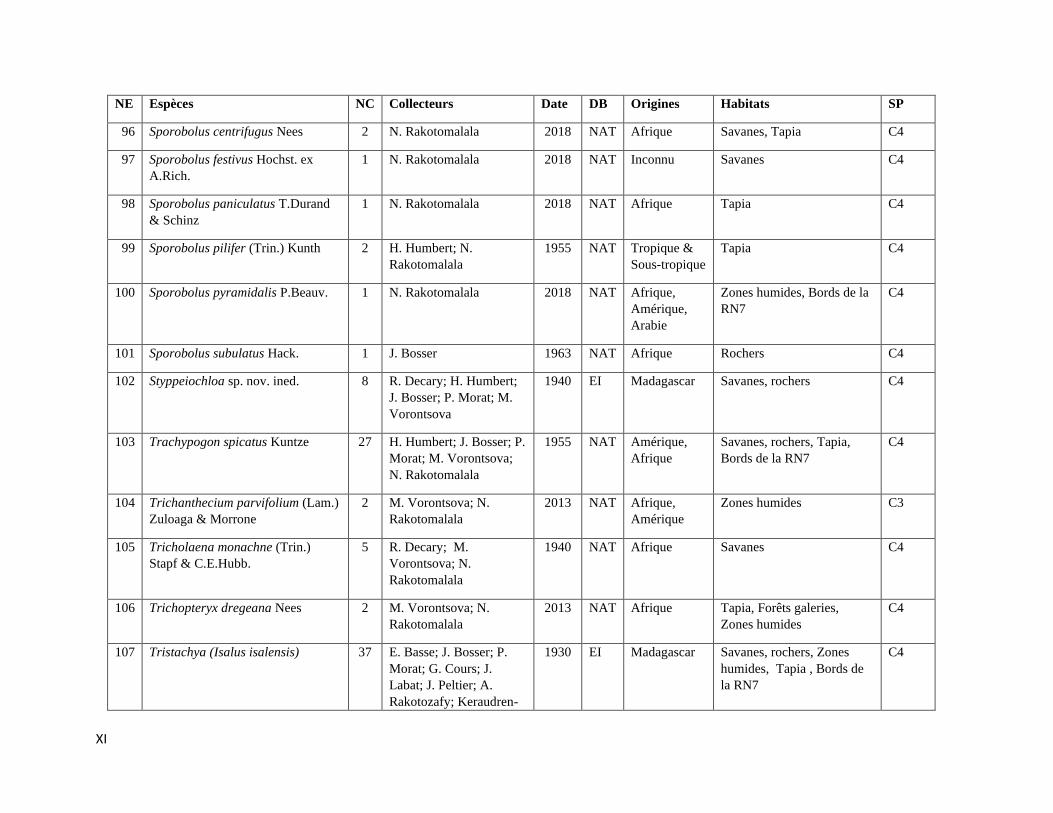
XI
NE Espèces NC Collecteurs Date DB Origines Habitats SP
96 Sporobolus centrifugus Nees 2 N. Rakotomalala 2018 NAT Afrique Savanes, Tapia C4
97 Sporobolus festivus Hochst. ex
A.Rich.
1 N. Rakotomalala 2018 NAT Inconnu Savanes C4
98 Sporobolus paniculatus T.Durand
& Schinz
1 N. Rakotomalala 2018 NAT Afrique Tapia C4
99 Sporobolus pilifer (Trin.) Kunth 2 H. Humbert; N.
Rakotomalala
1955 NAT Tropique &
Sous-tropique
Tapia C4
100 Sporobolus pyramidalis P.Beauv. 1 N. Rakotomalala 2018 NAT Afrique,
Amérique,
Arabie
Zones humides, Bords de la
RN7
C4
101 Sporobolus subulatus Hack. 1 J. Bosser 1963 NAT Afrique Rochers C4
102 Styppeiochloa sp. nov. ined. 8 R. Decary; H. Humbert;
J. Bosser; P. Morat; M.
Vorontsova
1940 EI Madagascar Savanes, rochers C4
103 Trachypogon spicatus Kuntze 27 H. Humbert; J. Bosser; P.
Morat; M. Vorontsova;
N. Rakotomalala
1955 NAT Amérique,
Afrique
Savanes, rochers, Tapia,
Bords de la RN7
C4
104 Trichanthecium parvifolium (Lam.)
Zuloaga & Morrone
2 M. Vorontsova; N.
Rakotomalala
2013 NAT Afrique,
Amérique
Zones humides C3
105 Tricholaena monachne (Trin.)
Stapf & C.E.Hubb.
5 R. Decary; M.
Vorontsova; N.
Rakotomalala
1940 NAT Afrique Savanes C4
106 Trichopteryx dregeana Nees 2 M. Vorontsova; N.
Rakotomalala
2013 NAT Afrique Tapia, Forêts galeries,
Zones humides
C4
107 Tristachya (Isalus isalensis) 37 E. Basse; J. Bosser; P.
Morat; G. Cours; J.
Labat; J. Peltier; A.
Rakotozafy; Keraudren-
1930 EI Madagascar Savanes, rochers, Zones
humides, Tapia , Bords de
la RN7
C4
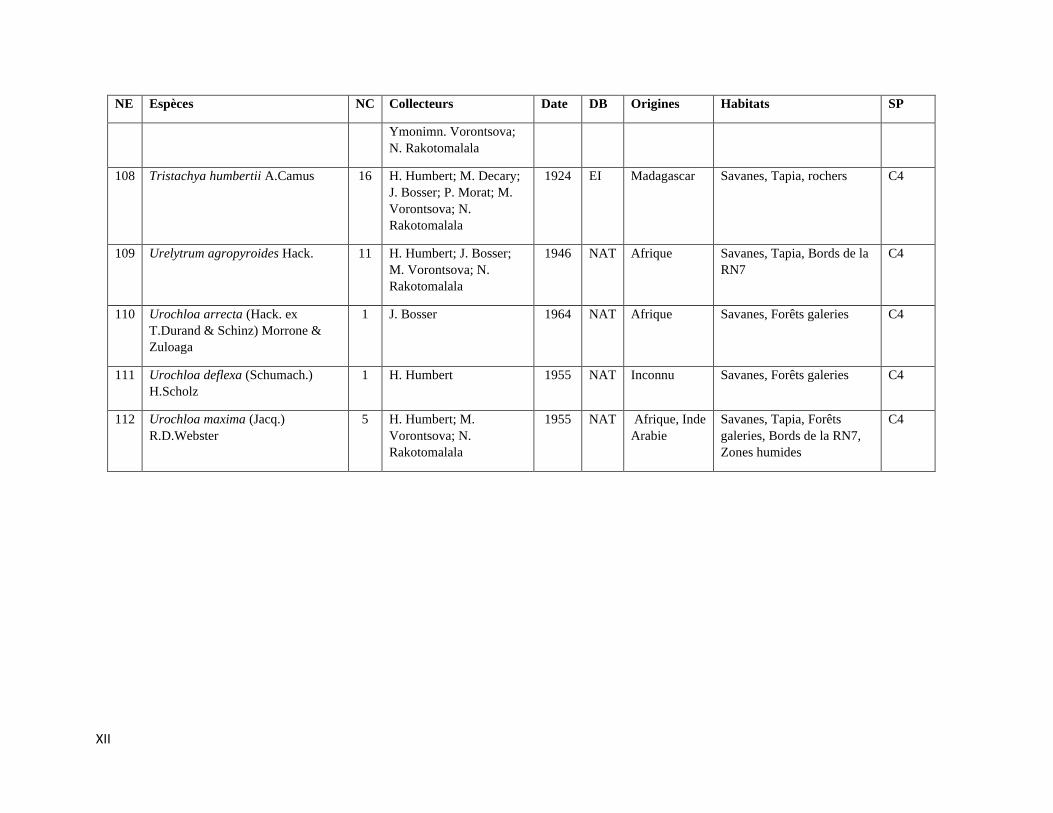
XII
NE Espèces NC Collecteurs Date DB Origines Habitats SP
Ymonimn. Vorontsova;
N. Rakotomalala
108 Tristachya humbertii A.Camus 16 H. Humbert; M. Decary;
J. Bosser; P. Morat; M.
Vorontsova; N.
Rakotomalala
1924 EI Madagascar Savanes, Tapia, rochers C4
109 Urelytrum agropyroides Hack. 11 H. Humbert; J. Bosser;
M. Vorontsova; N.
Rakotomalala
1946 NAT Afrique Savanes, Tapia, Bords de la
RN7
C4
110 Urochloa arrecta (Hack. ex
T.Durand & Schinz) Morrone &
Zuloaga
1 J. Bosser 1964 NAT Afrique Savanes, Forêts galeries C4
111 Urochloa deflexa (Schumach.)
H.Scholz
1 H. Humbert 1955 NAT Inconnu Savanes, Forêts galeries C4
112 Urochloa maxima (Jacq.)
R.D.Webster
5 H. Humbert; M.
Vorontsova; N.
Rakotomalala
1955 NAT Afrique, Inde
Arabie
Savanes, Tapia, Forêts
galeries, Bords de la RN7,
Zones humides
C4

XIII
ANNEXE IV: CLE DE DETERMINATION DES GENRES DE POACEAE DE L’ISALO ET
ILLUSTRATIONS
CLE DE DETERMINATION
1 Poaceae à chaume lignifié et à feuilles dimorphiques .….....……… Bambusoideae (une espèce).
1’ Poaceae à chaume non lignifiée, feuilles non dimorphiques ……………………………………... 2
2 Limbe munis de pseudo-pétioles, épillets unisexués (épillets mâles et femelles séparés) ……………
Humberthochloa bambusiuscula.
2’ Pseudo-pétioles absents, épillets bisexués ………………………………………………………… 3
3 Epillets un à plusieurs fleurons, si biflores : soit les deux fleurons sont bisexués, soit l’inferieur est
bisexué et le supérieur stérile ou mâle(si le fleuron inferieur est mâle ou hermaphrodite, et le fleuron
supérieur est femelle ou hermaphrodite, voir Isachne) ……….……..…………..……………………4
3’ Epillets toujours biflores avec le fleuron inferieur mâle ou stérile, et le supérieur bisexué …….... 23
4 Limbes lancéolés munis de veines transversales, épillets pluriflores latéralement compressés
………………………………………………………………………………. Megastachya mucronata.
4’ Limbes dépourvus de veines transversales…………………..…………………………………….. 5
5 Epillets uniflores bisexués…………………….……………………………………………………..6
5’ Epillets avec deux ou plusieurs fleurons bisexués ………………………………………………... 18
6 Lemma muni de trois arêtes fusionnées jusqu’au sommet ou se divisant et se terminant en trois
branches………………………………………………………………………………………….……. 7
6’ Lemma dépourvue d’arête ou à une seule arête .………………………………………………….... 8
7 Glumes à trois nervures, ne dépassent pas de 30 cm, plante poussant uniquement sur les escarpements
gréseux de l’Isalo ………………………………………………………..……………. Sartidia isalensis.
7’ Glume uninerve, plante plus grande ……………………………..……………. Aristida (5 espèces).
8 Paléa munie d’une carène, 3 à 7 nervures, épillet uniflore ……………..……… Leersia hexandra.
8’ Paléa à 2 carènes et 2 nervures ..………………………….………….……………………………. 9
9 Inflorescence en panicules …….………………………………………………………………….. 10
9’ Inflorescence en racèmes ...………………………………………………………………………. 12
10 Epillets biflores, taille ne dépassant pas de 25 cm, plante des zones humides strictes … Coelachne
africana.
10’ Epillets uniflores, taille dépassant 25 cm, poussent sur tout habitat .…… Sporobolus (6 espèces)
12 Racèmes solitaires …………….………………………………………………………………….. 13
12’ Deux ou plusieurs racèmes ……………………………………..……………………………….. 16
13 Epillets mutiques, glumes recouvrant les fleurons ………………………… Microchloa kunthii.
13’ Epillets aristés, glume supérieure mutique ou aristée au sommet …………………………….... 14

XIV
14 Epillets 2 à 5 fleurons, glumes membraneuses, courtes que l’épillet ………. Neostapfiella perrieri
14’ Epillets uniflore …………………………………………………………………………………. 15
15 Racème unilatéral, épillets à 2 à 3 fleurons, arêtes dressées ……………… Enteropogon prieurii.
15’ Racème bilatéral, racème violacé ou jaunâtre à maturité ……..…………….…..… Perotis patens.
16 Racèmes arrangés de long de l’axe de l’inflorescence, lemma recouverte par les glumes...………
Craspedorhachis africana.
16’ Racèmes digités, lemma non recouverte par les glumes …………………………………….…. 17
17 Epillets mutiques, plantes gazonnantes, à la fois stolonifère et rhizomateuse …. Cynodon dactylon.
17’ Epillets aristés, plantes grêles, ne possédant pas ce caractère ………… Chloris (C. pychnotrix ; C.
virgata).
18 Plantes à port rigide de grande taille, inflorescences en panicules larges et plumeuses ……….. 19
18’ Plantes herbacées de petite taille, inflorescences non plumeuses ………………………...…… 20
19 Lemma mutique, plante aquatique et de grande taille ressemblant aux bambous ….. Phragmites
mauritiana
19’ Lemma munie d’arêtes, plante de taille moins large, poussant sur des zones arides … Neyraudia
arundinacae
20 Inflorescence en panicules ………………………………………………………………………. 21
20’ Inflorescence racémeuse ………………………………………………………………………… 22
21 Lemma mutique, épillets pluriflores, plus de 5 fleurons …………………………...…… Eragrostis (7
espèces), Pogonarthria squarrosa (en combination avec Eragrostis)
21’ Lemma aristée, épillets 2 à 5 fleurons …………………………….... Styppeiochloa sp. nov. ined.
22 Racèmes digités, l'axe du racème est terminé par une pointe ……………….…….. Dactyloctenium
aegyptium
22’ Racèmes digités, l’axe du racème est terminé par un épillet ………….………….. Eleusine indica
23 Epillets se brisent à maturité et les glumes persistent ……………………………………………. 24
23’ Epillets tombant entière à maturité ………………………………………………..…………….. 26
24 Glumes dépourvues de poils à base tuberculée, épillets plus petits ……….. Trichopteryx dregeana
24’ Glumes munies de poils à base tuberculée plus de 6 mm …………..…………………………… 24
25 Chaque branche de l’inflorescence se terminant en une paire d’épillets ………...……….. Loudetia (L.
filifolia ; L. simplex)

XV
25’ Chaque branche de l’inflorescence se terminant en une triade d’épillets ……..………… Tristachya
(T. humbertii ; T. Isalus isalensis)
26 Epillets solitaires, avec ou sans arêtes ………………………………………………………….. 27
26’ Epillets géminés dont l’un sessile et aristé, et l’autre pédicellé et mutique…………………… 44
27 Epillets surmontés par des soies ou d'arêtes épineuses, tombant ensemble à maturité…. Cenchrus (3
espèces)
27’ Epillets non surmontés par des soies ou si soies présentes, elles ne sont pas épineuses ………. 28
28 Soies présentes et persistent sur les rachis après que les épillets tombent à maturité …… Setaria (4
espèces)
28’ Soies absentes ……………..…………………………………………………………………… 29
29 Lemma supérieure hyaline à coriacée à maturité, marges enroulées……………………………. 30
29’ Lemma supérieure cartilagineuse ou chartacée, marges aplaties ……….……………………… 42
30 Fleuron supérieur latéralement compressé ………..……………………….…………………... 31
30’ Fleuron supérieur dorsalement compressé………………….……..…………………………... 32
31 Inflorescence en panicules ouvertes ou semi-contractées, épillets très compressés latéralement, à profil
gibbeux sur le dos ………………………………. Cyrtococcum (C. deltoideum, C. multinode).
31’ Inflorescence racémeuse, épillets ne disposant pas ce caractère, glume supérieure surmontée par des
poils à base tuberculée ou par des ailes ………… Pseudechinolaena (P. perrieri, P. polystachya)
32 Inflorescence en panicules ouvertes ou contractées ……………………………………………. 33
32’ Inflorescence racémeuse …………………………….….……………………………………… 37
33 Epillets asymétriques, panicules ouvertes ou contractées prenant la forme d’un épi, glumes à nervures
saillantes .................................................................… Sacciolepis (S. curvata, S. africana).

XVI
33’ Epillets symétriques, panicules ouvertes ……………………………………………………….34
34 Glume supérieure et lemma inférieure compressées vers le sommet, lemma supérieure munies de
crêtes.…………………………………………………………………………. Acroceras (3 espèces).
34’ Glume supérieure et lemma inférieure non compressées, crêtes absentes au niveau de la lemma
supérieure ………………………………………………………………………………………….. 35
35 Chaume cylindrique, robuste, grimpant, glume inférieure de la taille des épillets ou encore plus longue
…………………………………………………… Pseudolasiacis neoperrieri var. leroyana.
35’ Chaume herbacé, glume inférieure nettement courte que la longueur de l’épillet……….…… 36
36 Plante de 5 à 30 cm de haut, feuilles à limbes vert glauques parfois violacées dessous, dressées puis
réfléchies à maturité …………………………………………… Trichanthecium parviflorum.
36’ Plante plus grande, de 0.1 à 2 m de hauteur, axes de l’inflorescence munis de plusieurs poils
glanduleux au sommet ……………………………………..………….. Adenochloa hymeniochila.
37 Glumes aristées ……………………………………………………………………………….. 38
37’ Glumes mutiques ………………………………………………………….…………………. 39
38 Epillets latéralement compressés ……………………………………….. Oplismenus (3 espèces).
38’ Epillets dorsalement compressés ……..…………………………………… Echinochloa colona.
39 Glume inférieure présente …………………………………………………..…………………. 40
39’ Glume inférieure absente ou vestigiale …………………………………………….………… 41
40 Lemma supérieure lisse …………………………………..……………… Brachiaria (3 espèces)
40’ Lemma supérieure rigoureuse …………………………………………….. Urochloa (3 espèces)
41 Epillets plan-convexes (ronds et aplatis)………………………………. Paspalum scrobiculatum.
41’ Epillets lancéolés à oblongs, feuilles à gaines comprimées .... …………. Axonopus compressus.

XVII
42 Epillets dorsalement compressés, racèmes digitées ……………………….. Digitaria (6 espèces).
42’ Epillets latéralement compressés ……………………………………..………………………. 43
43 Lemma supérieure latéralement compressée ………...……. Melinis (M. minutiflora, M. repens).
43’ Lemma supérieure dorsalement compressée …………………………… Tricholaena monachne.
44 Entre-noeud du rachis de l’inflorescence et le pédicelle de l’épillet pédicellé sont épais et solides,
racèmes solitaires et spiciformes ……………………………………………….………………….. 45
44 Rachéoles et pédicelles de l’épillet pédicellé minces et linéaires ………………………………. 46
45 Racèmes cylindriques dorsiventraux, épillets mutiques, rachis très fragiles ................. Rottboellia
cochinchinensis.
45’ Racèmes spiciformes solitaires, l’épillet sessile mutique et l’espèce pédicellé à glume inférieure
longuement aristée ………………………………………………………… Urelytrum agropyroides.
46 Epillets sessiles similaires aux épillets pédicellés, panicules en faux épis blancs et soyeux
………………………………………………………………………………….. Imperata cylindrica.
46’ Epillets sessiles similaires et épillets pédicellés différents, inflorescences non soyeux ……... 47
47 Epillet sessile mâleou stérile, épillet pédicellé bisexuée et aristé ………. Trachypogon spicatus.
47’ Epillet sessile bisexuée, épillet pédicellé mâleou stérile …………………………………..… 48
48 Inflorescences en panicules, épillets surmontés par une touffe de poils fauves à brun dorée qui persiste
à maturité ……………………………………………….………… Chrysopogon serrulatus.
48’ Inflorescence racémeuse ……………………………………………………………………… 49
49 Pédicelle et entre-noeuds du rachis présentant une ligne translucide au milieu …… Bothriochloa
bladhii.
49’ Pédicelle sans une ligne translucide ………………………………………….………………… 50
50 Racèmes solitaires ……………...……………………………………………….………………. 51
50’ Racèmes géminés ……………………………………………………………………………… 52
51 Racèmes asymétriques, se terminant par des arêtes fermes de couleur marron noirâtre
……………………………………………………………………………….. Heteropogon contortus.
51’ Racèmes symétriques, fragiles, souvent rougeâtres ……… Schizachyrium (S. sanguineum, S.
brevifolium).

XVIII
52 Le fleuron inférieur de l’épillet sessile est mâle et muni de paléa .……………….. Ischaemum (I.
koleostachys, I. polystachyum).
52’ Le fleuron inférieur de l’épillet sessile est stérile et sans paléa ………………..……..……..…. 53
53 Glume inférieure de l’épillet sessile bicarénée ………………………………….………………. 54
53’ Glume inférieure de l’épillet sessile arrondie sur le dos ………………………...…………..…. 55
54 Racèmes défléchis, feuilles aromatiques ……………… ……………..…… Cymbopogon caesius.
54’ Racèmes non défléchis, feuilles non aromatiques………………...……. Andropogon (4 espèces).
55 Glume inférieure de l’épillet creusée d’un sillon médian profond …………. Hyperthelia dissoluta.
55 Glume inférieure de l’épillet arrondie sur le dos et n’est pas creusée ……………. Hyparrhenia (H.
newtonii, H. rufa).
ILLUSTRATION
- Différents types d’épillets. A. Deux épillets géminés avec un pédicelle mutique et un sessile
aristé (Bothriochloa bladhii). B. Un épillet aristé, triflore, glumes mutiques, lemmas aristées
(Styppeiochloa sp. nov. ined.). C. Un épillet pluriflore, glume mutique, lemmas mucroné (Megastachya
mucronata). D. Un épillet biflore et mutique, ové (Pseudolasiacis neoperrieri var. neorperrieri). E. Un
épillet plurifore et mutique (Eragrostis atrivirens).
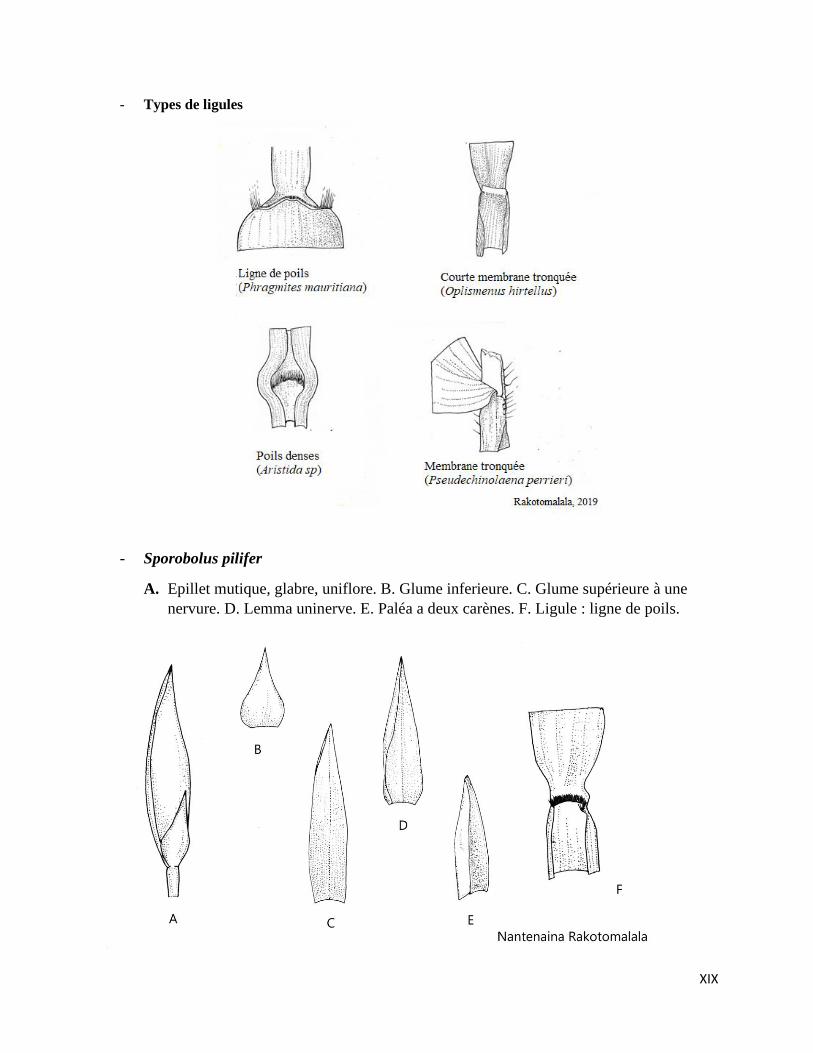
XIX
- Types de ligules
- Sporobolus pilifer
A. Epillet mutique, glabre, uniflore. B. Glume inferieure. C. Glume supérieure à une
nervure. D. Lemma uninerve. E. Paléa a deux carènes. F. Ligule : ligne de poils.
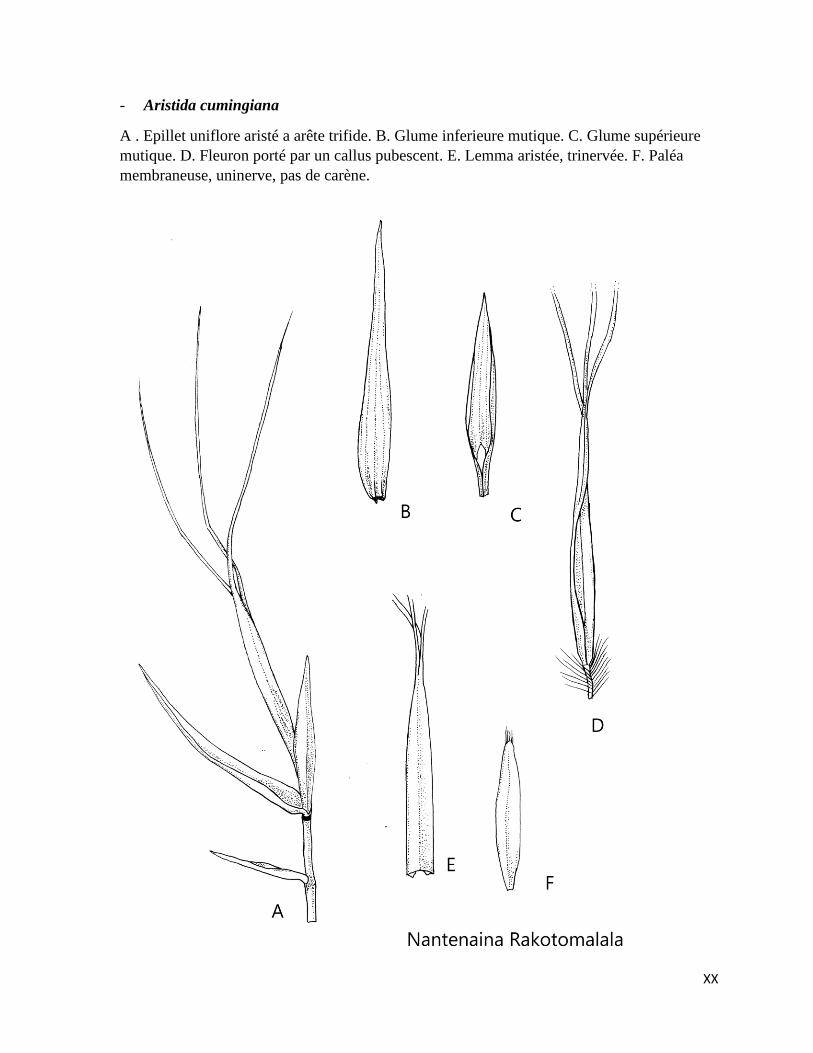
XX
- Aristida cumingiana
A . Epillet uniflore aristé a arête trifide. B. Glume inferieure mutique. C. Glume supérieure
mutique. D. Fleuron porté par un callus pubescent. E. Lemma aristée, trinervée. F. Paléa
membraneuse, uninerve, pas de carène.
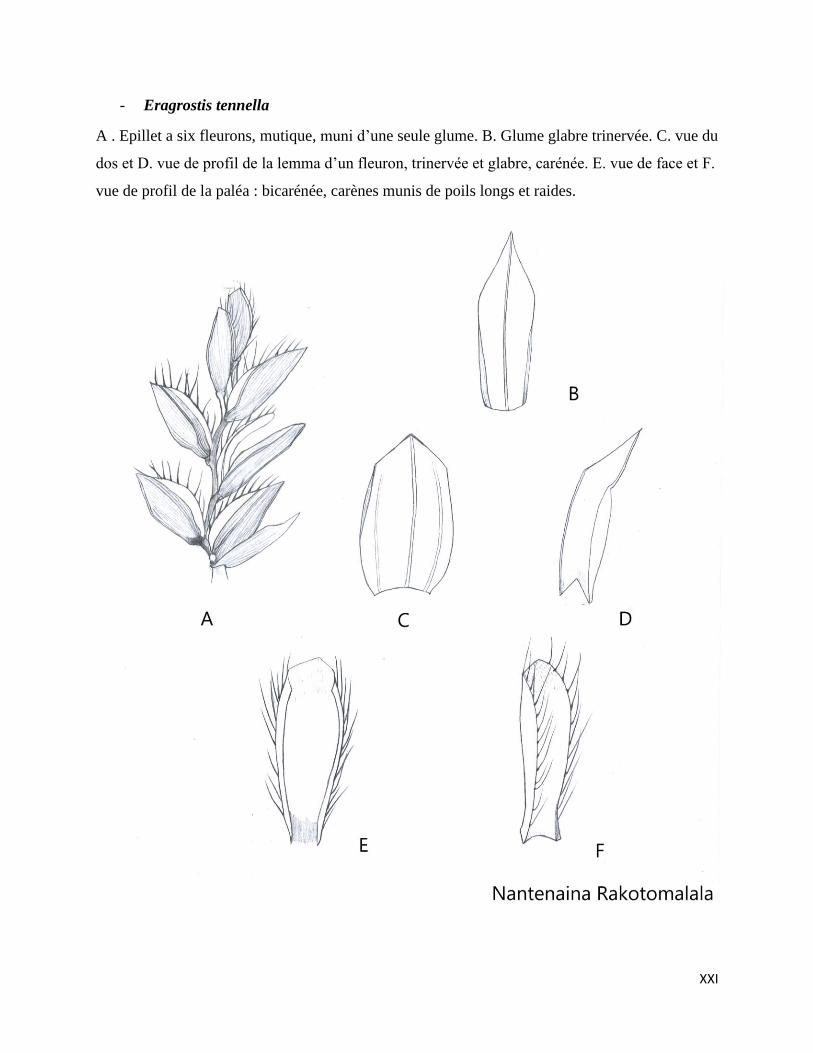
XXI
- Eragrostis tennella
A . Epillet a six fleurons, mutique, muni d’une seule glume. B. Glume glabre trinervée. C. vue du
dos et D. vue de profil de la lemma d’un fleuron, trinervée et glabre, carénée. E. vue de face et F.
vue de profil de la paléa : bicarénée, carènes munis de poils longs et raides.
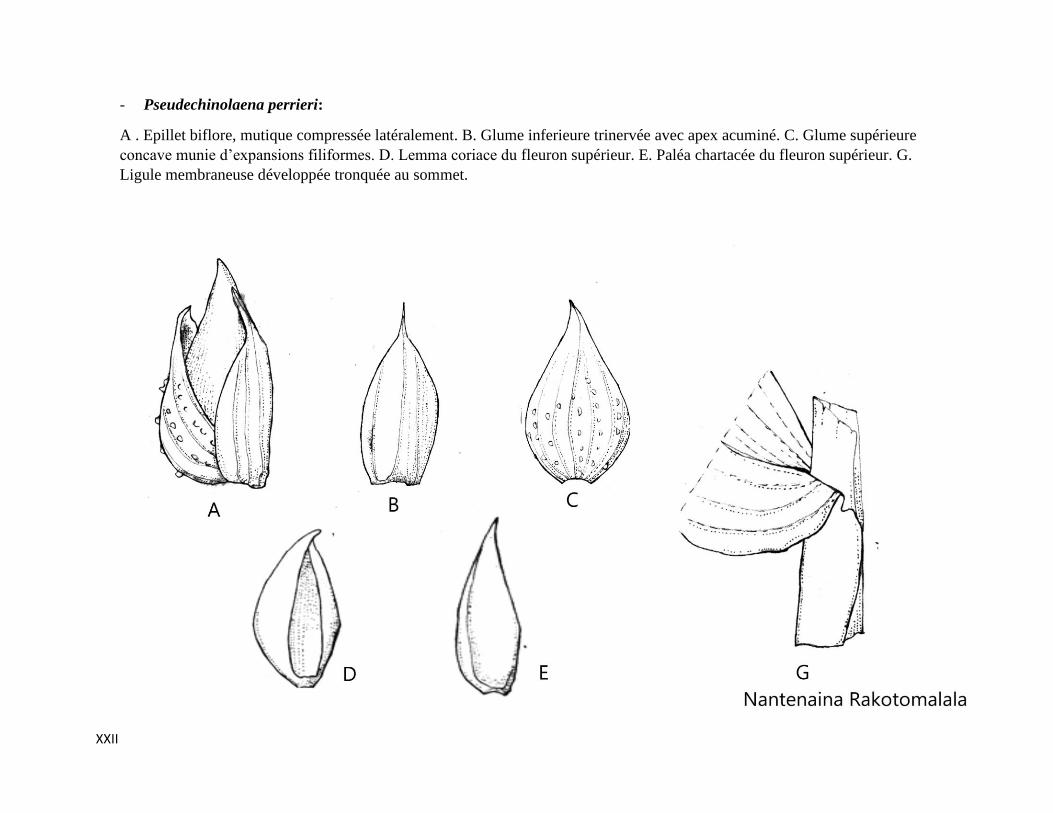
XXII
- Pseudechinolaena perrieri:
A . Epillet biflore, mutique compressée latéralement. B. Glume inferieure trinervée avec apex acuminé. C. Glume supérieure
concave munie d’expansions filiformes. D. Lemma coriace du fleuron supérieur. E. Paléa chartacée du fleuron supérieur. G.
Ligule membraneuse développée tronquée au sommet.
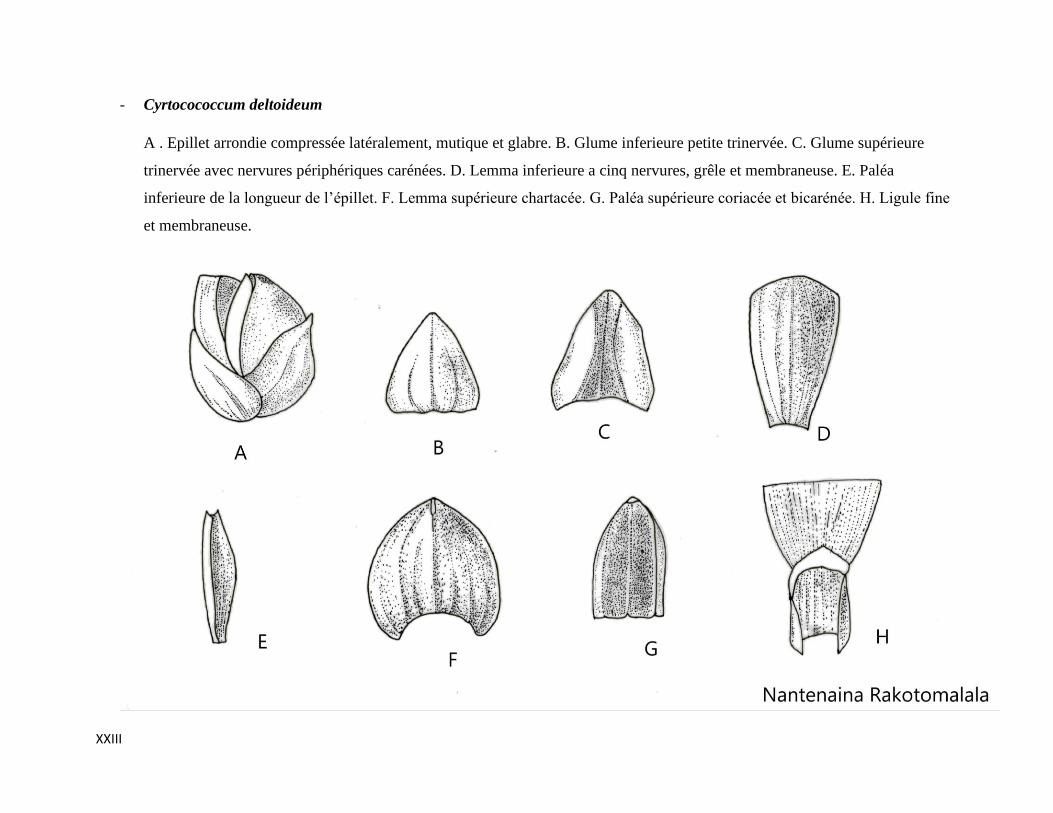
XXIII
- Cyrtocococcum deltoideum
A . Epillet arrondie compressée latéralement, mutique et glabre. B. Glume inferieure petite trinervée. C. Glume supérieure
trinervée avec nervures périphériques carénées. D. Lemma inferieure a cinq nervures, grêle et membraneuse. E. Paléa
inferieure de la longueur de l’épillet. F. Lemma supérieure chartacée. G. Paléa supérieure coriacée et bicarénée. H. Ligule fine
et membraneuse.
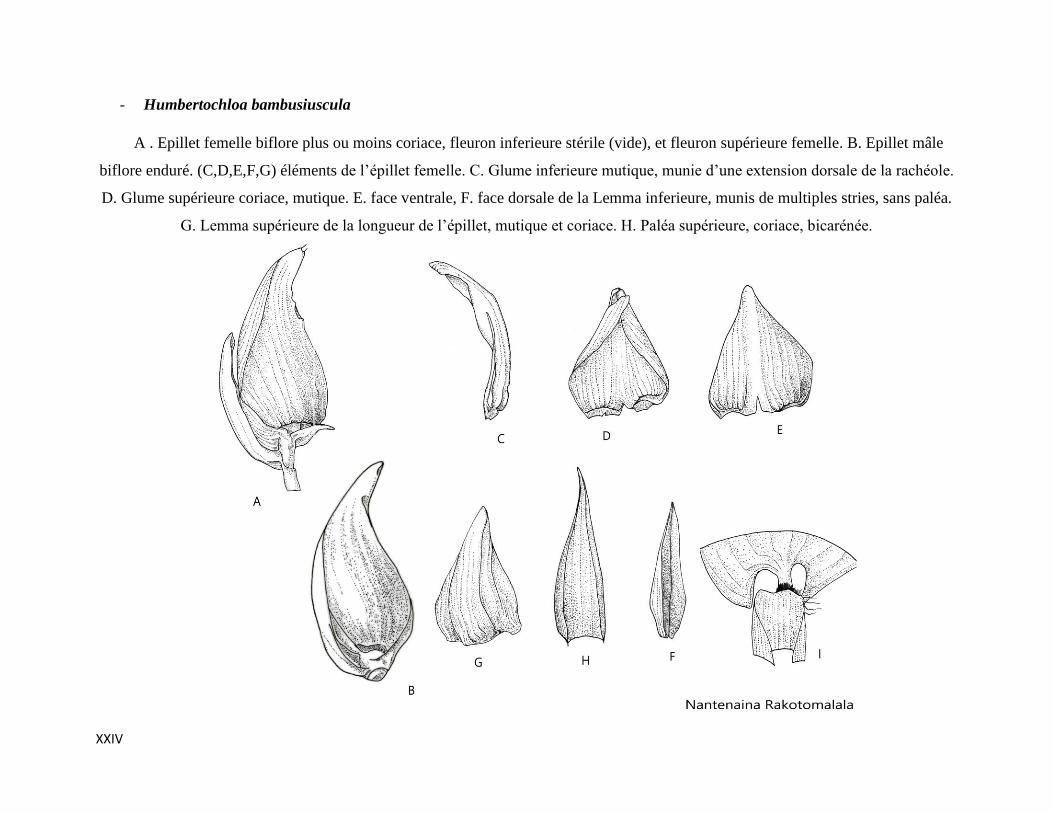
XXIV
- Humbertochloa bambusiuscula
A . Epillet femelle biflore plus ou moins coriace, fleuron inferieure stérile (vide), et fleuron supérieure femelle. B. Epillet mâle
biflore enduré. (C,D,E,F,G) éléments de l’épillet femelle. C. Glume inferieure mutique, munie d’une extension dorsale de la rachéole.
D. Glume supérieure coriace, mutique. E. face ventrale, F. face dorsale de la Lemma inferieure, munis de multiples stries, sans paléa.
G. Lemma supérieure de la longueur de l’épillet, mutique et coriace. H. Paléa supérieure, coriace, bicarénée.

XXV
- Tristachya humbertii
A Triades d'épillets. B : Glume inferieure avec poils à base tuberculée. C : Glume supérieure bilobée vers le sommet. D : Fleuron
inferieur mâle glabre munie de paléa et de lemma. E : Fleuron supérieur bisexué, avec lemma à trois lobes qui se finissent en arêtes
torsades. Lemma et callus pubescents.

RESUME
La famille des Poaceae (graminées) figure parmi les familles de plantes globalement mieux
documentées en termes de taxonomie, mais sont encore mal connues à Madagascar. Leur
diversité demeurait mal étudiée puisque les graminées et les savanes ont été toujours considérés
comme des mauvaises herbes et des terrains dégradés. Dans la présente étude, on vise alors à
établir une liste des graminées rencontreés dans le Parc National d’Isalo et à étudier leur
distribution, afin d’améliorer la connaissance et la compréhension sur leur diversité et
endémicité, ainsi que sur l’origine des écosystèmes herbeux de l’Isalo. Des revues de spécimens
d'herbiers de Poaceae existants et de nouveaux inventaires sur terrain ont été réalisés et ont
permis de générer 589 collections de graminées du site d’étude. Elles ont été attribuées à 112
espèces de Poaceae, dont 38 sont des premières collections pour l’Isalo. La flore mise à jour de
Poaceae de l’Isalo comprend sept sous-familles, parmi les 10 présentes à Madagascar, dont celle
des Panicoideae représente 65% du total des espèces. L’endémicité des Poaceae de l’Isalo est à
22%, incluant trois espèces et une variété restreintes à Isalo. Deux de ces espèces sont très
charismatiques et particularisent en quelques sortes les savanes de l’Isalo, à savoir : Tristachya
humbertii et Tristachya (Isalus isalensis). La diversité spécifique maximale a été rencontrée dans
les sites Namaza et Ranohira, là où de complexes habitats se rencontrent, entre et aux alentours
des canyons. L’habitat abritant la plus grande diversité correspond aux savanes, suivies par les
forêts humides. Les savanes se composent par des graminées d’origine natives, et abritent pas
mal d’espèces endémiques, et le déroulement du peuplement de l’Isalo rend moins évident la
présence de forêts (denses) et de massive déforestation à Isalo, autrefois. Ces faits, bien que très
simples, contrastent aux caractéristiques d’un écosystème secondaire. En bref, cette étude a
amélioré la connaissance sur les Poaceae et fournit des données de base pour les recherches
ultérieures sur ce groupe et sur les savanes de Madagascar.
Mots clés
Graminées, Madagascar, diversité, savanes, endémicité.

ABSTRACT
Poaceae family (grasses) is one of the globally best documented plant families in terms of
taxonomy, but still poorly known in Madagascar. Their diversity has always been rarely studied
since grasses and grasslands are considered by many to be weeds and degraded wastelands. In
this study, we then aimed to list the grasses in Isalo National Park and study their distribution, to
improve knowledge and understanding on their diversity and endemicity, as well as the origin of
Isalo’s grasslands. Reviews of herbarium specimens and new field inventories allowed us to
confirm 589 grass occurrence records from the study area. They were assigned to 112 species, of
which 38 are new records for Isalo. The updated grass flora of Isalo includes subfamilies, of the
10 present in Madagascar, of which the subfamily Panicoideae makes up 65% of the total
species. The grass endemism is 22% for Isalo, including three species and one variety restricted
to Isalo. Two of these species are very charismatic and specify somehow Isalo’s savanna, these
are: Tristachya humbertii and Tristachya (Isalus isalensis). The maximum grass diversity was
found in sites Namaza and Ranohira, where complex habitats occur, in and around canyons. The
habitat harbouring the highest grass species diversity goes to savanna, followed by the humid
forests. Isalo’s savanna is composed of native grasses and harbours endemic species, and the
human settlement process in Isalo does not evidence the presence of forests (dense) and massive
deforestation in Isalo, in the past. These facts, thought very simple, contrast to a secondary
grassland’s features. In summary, this study improved knowledge on Poaceae and fornicates data
baseline for future research on this group and on grasslands of Madagascar.
Key words
Grasses, Madagascar, diversity, savanna, endemism.


















