Direction de la recherche sur la faune1 Direction ...plaines inondables de la rivière Saint-Jean au...
59
Direction de la recherche sur la faune 1 et Direction régionale du Saguenay - Lac Saint-Jean 2 CARACTÉRISATION DE L’HABITAT DE REPRODUCTION DES CANARDS ARBORICOLES par Charles Maisonneuve 1 Raymond Mc Nicoll 1 Alain Desrosiers 1 et Gilles Lupien 2 Société de la faune et des parcs du Québec Septembre 2002
Transcript of Direction de la recherche sur la faune1 Direction ...plaines inondables de la rivière Saint-Jean au...

Direction de la recherche sur la faune1
et
Direction régionale du Saguenay - Lac Saint-Jean2
CARACTÉRISATION DE L’HABITAT DE REPRODUCTION
DES CANARDS ARBORICOLES
par
Charles Maisonneuve1
Raymond Mc Nicoll1
Alain Desrosiers1
et
Gilles Lupien2
Société de la faune et des parcs du Québec
Septembre 2002

ii
Référence à citer :
MAISONNEUVE, C., R. MC NICOLL, A. DESROSIERS et G.LUPIEN. 2002. Caractérisationde l’habitat de reproduction des canards arboricoles. Société de la faune et des parcs du Québec,Direction de la recherche sur la faune et Direction de l’aménagement de la faune du Saguenay –Lac Saint-Jean. 51 p.
Dépôt légal – Bibliothèque nationale du Québec, 2002ISBN : 2-550-39743-6

iii
RÉSUMÉ
Le garrot à œil d’or est l’une des plus grandes espèces d’oiseau à utiliser des cavités d’arbres pour
la nidification. La conservation d’une densité suffisante de gros arbres pour assurer le maintien
des populations de cette espèce est susceptible de servir aussi à plusieurs autres espèces qui
dépendent des cavités. Une étude a donc été initiée afin de mieux caractériser l’habitat de
nidification de cette espèce.
À la fin de la saison de reproduction, les canards arboricoles effectuent de la prospection afin de
localiser des cavités en prévision de l’année suivante. Ce comportement serait une adaptation
permettant à ces oiseaux d’avoir des sites de nidification alternatifs compte tenu de la précarité
des sites utilisés. On a donc profité de ce comportement en créant un réseau de nichoirs dans la
région de Portneuf et dans la réserve faunique Ashuapmushuan. Chacun des nichoirs était équipé
d’un mécanisme de déclenchement activé dès qu’un animal pénètre à l’intérieur et qui entraîne la
fermeture d’une porte bloquant l’issue du nichoir. Les femelles étaient munies d’émetteurs
permettant de suivre leurs déplacements et on pouvait ainsi suivre celles-ci vers les cavités
naturelles utilisées comme sites de nidification alternatifs.
Près de 50 garrots à œil d’or ont été capturés, de même que 44 harles couronnés. Tous les garrots
ont été munis d’émetteurs, de même que quelques harles, afin de suivre leurs déplacements et
localiser les nids. Les arbres utilisés pour la nidification avaient un diamètre à hauteur de poitrine
(DHP) de près de 50 cm et la distance moyenne du plan d’eau le plus proche était de 300 m. Ces
chicots ont une durée de vie limitée et des travaux devraient être réalisés pour évaluer si les
conditions actuelles permettent un renouvellement suffisant d’arbres de cette taille pour assurer le
maintien des populations d’espèces qui en dépendent. La faible proportion des cavités trouvées à
moins de 20 m des plans d’eau indique que la bande riveraine présentement laissée lors des
coupes est susceptible d’être insuffisante, d’autant plus qu’une partie des tiges exploitables est
aussi récoltée dans ces bandes.

iv

v
TABLE DES MATIÈRES
RÉSUMÉ .................................................................................................................................................................... III
TABLE DES MATIÈRES .......................................................................................................................................... V
LISTE DES TABLEAUX ..........................................................................................................................................VI
LISTE DES FIGURES..............................................................................................................................................VII
1. INTRODUCTION................................................................................................................................................... 1
2. AIRES D’ÉTUDE.................................................................................................................................................... 3
3. MÉTHODES ........................................................................................................................................................... 7
3.1 INSTALLATION ET VISITE DES NICHOIRS ............................................................................................................. 73.2 CAPTURE ET MARQUAGE ................................................................................................................................... 93.3 SUIVI TÉLÉMÉTRIQUE ...................................................................................................................................... 103.4 VISITE DE CAVITÉS UTILISÉES ANTÉRIEUREMENT PAR LE GARROT À ŒIL D'OR................................................. 113.5 TAUX D'OCCUPATION DU RÉSEAU DE NICHOIRS DANS LA RÉSERVE FAUNIQUE ASHUAPMUSHUAN ................... 11
4. RÉSULTATS ......................................................................................................................................................... 12
4.1 PÉRIODE DE CAPTURE...................................................................................................................................... 124.1.1 Effort et succès de capture............................................................................................................... 124.1.2 État reproducteur des femelles capturées ........................................................................................ 164.1.3 Masse des garrots à œil d’or............................................................................................................ 17
4.2 PÉRIODE DE NIDIFICATION ............................................................................................................................... 184.2.1 Proportion des femelles ayant niché................................................................................................ 184.2.2 Sites de nidification.......................................................................................................................... 194.2.2 Chronologie de nidification ............................................................................................................. 21
4.3 PÉRIODE D’ÉLEVAGE ....................................................................................................................................... 234.4 INSPECTION DES CAVITÉS UTILISÉES EN 1999 PAR DES GARROTS..................................................................... 254.5 TAUX D’OCCUPATION DU RÉSEAU DE NICHOIRS DE LA RÉSERVE FAUNIQUE ASHUAPMUSHUAN....................... 26
5. DISCUSSION ........................................................................................................................................................ 27
5.1 EFFICACITÉ DES MÉTHODES............................................................................................................................. 275.1.1 Capture et télémétrie........................................................................................................................ 275.1.2 Effets possibles des méthodes sur la nidification du garrot............................................................. 285.1.3 Examen des nids dans les cavités naturelles.................................................................................... 35
5.2 NIDIFICATION ................................................................................................................................................. 35
6. CONCLUSION ET RECOMMANDATIONS.................................................................................................... 41
6.1 MÉTHODES...................................................................................................................................................... 416.2 PRATIQUES FORESTIÈRES................................................................................................................................. 42
REMERCIEMENTS.................................................................................................................................................. 45
RÉFÉRENCES ........................................................................................................................................................... 46

vi
LISTE DES TABLEAUX
Tableau 1. Effort déployé et succès de capture obtenu avec les nichoirs de capture dansla réserve faunique Ashuapmushuan, 1999, et la région de Portneuf, 1999-2001 ......13
Tableau 2. Fréquence de capture de femelles de garrot à œil d’or et de harle couronné au coursd’une même saison dans les nichoirs de la région de Portneuf, 1999-2001................14
Tableau 3. Répartition des recaptures selon les années.................................................................14
Tableau 4. Efficacité des émetteurs utilisés en 1999.....................................................................15
Tableau 5. Proportions des femelles de garrot à œil d’or capturées dans la région de Portneufayant pondu un œuf lors de la capture.........................................................................17
Tableau 6. Proportions des femelles de garrot à œil d’or capturées dans la région de Portneufprésentant des signes de reproduction .........................................................................17
Tableau 7. Comparaisons (tests de Wilcoxon et de Kruskal-Wallis) de la masse des femellesde garrot à œil d’or capturées dans la région de Portneuf, 1999-2000........................18
Tableau 8. Proportions des femelles de garrot à œil d’or capturées dans la région de Portneufayant initié un nid ........................................................................................................19
Tableau 9. Proportions des femelles de harle couronné capturées dans la région de Portneufayant initié un nid ........................................................................................................19
Tableau 10. Caractéristiques des sites de nidification utilisés dans la région de Portneuf par les femelles marquées.................................................................................................21
Tableau 11. Occupation des nichoirs dans la réserve faunique Ashuapmushuan ..........................26
Tableau 12. Chronologie de la période de ponte du garrot à œil d’or mentionnée dans la littérature ....................................................................................................................30
Tableau 13. Masses de femelles de garrot à œil d'or mentionnées dans la littérature selon le stade de la période de reproduction et le statut reproducteur .....................................32

vii
LISTE DES FIGURES
Figure 1. Localisation de l’aire d’étude dans la réserve faunique de Portneuf et la ZEC –Bastican-Neilson…………………………………………………………………… 5
Figure 2. Localisation de l’aire d’étude dans la réserve faunique Ashuapmushuan…………... 6

11. INTRODUCTION
Plusieurs espèces de vertébrés dépendent de la présence de chicots pour s’abriter, s’alimenter et se
reproduire. Les oiseaux dominent généralement cette faune utilisatrice de bois mort et la situation
n’est pas différente au Québec où 36 espèces d’oiseaux sont associées aux arbres à cavité (Darveau
et Desrochers 2001).
Depuis longtemps, on soupçonne que la disponibilité de cavités dans les chicots représente un
facteur qui limite l’abondance des espèces qui nichent en cavité (von Haartman 1957). Plusieurs
études ont pu mettre en évidence certains éléments confirmant cette hypothèse; certaines ont indiqué
un accroissement notable des populations de certaines espèces nichant en cavité à la suite de
l’installation d’un réseau de nichoirs artificiels (Hamerstrom et al. 1973; Brush 1983; Newton 1994;
Holt and Martin 1997) alors que d’autres ont souligné une saturation des cavités naturelles
disponibles au cours de certaines années (van Balen et al. 1982) et même une diminution du nombre
d’oiseaux reproducteurs après l’obstruction de cavités sur un territoire où il y en avait en abondance
(Brush 1983).
L’exploitation forestière représente l’un des facteurs pouvant réduire l’abondance de chicots (Cline
et al. 1980) et plusieurs études ont démontré que les pratiques d’aménagement forestier peuvent
ainsi contribuer à réduire les populations d’espèces nichant en cavité (Haapanen 1965; Balda 1975;
Mannan et al. 1980, Morrison et Meslow 1983). Ces constatations ont mené les aménageurs
forestiers à tenter de développer des approches de bonnes pratiques forestières pour favoriser le
maintien de chicots (voir Darveau et Desrochers 2001). Par contre, au Québec, il n’existe
présentement aucune législation ou réglementation à cette fin.
Un aménagement forestier ciblant les besoins de la plus grande espèce de cavité présente dans une
région donnée fait partie des approches préconisées pour assurer la conservation de chicots de façon
à favoriser le plus grand nombre d’espèces fauniques possibles (Darveau et Desrochers 2001).
Quelle que soit cette espèce, celle-ci servirait ainsi de « parapluie » pour l’ensemble des espèces qui
dépendent des chicots. Cette approche permettrait d’assurer la conservation d’un nombre suffisant
des plus gros chicots qui demeurent généralement sur pied plus longtemps (Cline et al. 1980;
Schreber et Decalesta 1992).
Au Québec, les plus grandes espèces d’oiseau qui nichent dans des cavités d’arbre sont la chouette
lapone (Strix nebulosa), le grand harle (Mergus merganser), la chouette rayée (Strix varia), le garrot

2d’Islande (Bucephala islandica) et le garrot à œil d’or (Bucephala clangula). La chouette lapone et
le garrot d’Islande ont, au Québec, une répartition beaucoup trop limitée pour se prêter à un tel
exercice. Parmi les trois autres espèces, le garrot à œil d’or est celle qui a la plus vaste répartition
géographique, celui-ci étant présent jusqu’à la limite des arbres. De plus, des indices récents de
déclin de ses populations dans l’est de l’Amérique du Nord (Bordage 1995; Bordage et Plante 1997)
pourraient inciter à croire que des problèmes d’habitat affectent le garrot à œil d’or. De meilleures
connaissances des besoins de cette espèce en termes d’habitat de nidification peuvent donc s’avérer
utiles pour l’élaboration de normes de conservation de chicots visant à favoriser le plus grand
nombre possible d’espèces fauniques.
Mais, étonnamment, comme la majorité des études ont été réalisées sur des populations en nichoirs
(Eadie et al. 1995), il n’existait jusqu’à récemment que très peu d’informations publiées permettant
de bien caractériser les sites de nidification recherchés par le garrot. Les seules études publiées qui
donnent des détails concernant les arbres utilisés par des garrots à œil d’or ont été réalisées dans les
plaines inondables de la rivière Saint-Jean au Nouveau-Brunswick (Carter 1958; Prince 1968).
Cependant, cette région se trouve en périphérie de l’aire de reproduction du garrot à œil d’or qui est
surtout concentrée dans la forêt boréale où les habitats diffèrent considérablement. La présente étude
avait donc comme objectif principal la caractérisation des chicots utilisés pour la nidification en
forêt mixte et boréale par le garrot à œil d’or afin de fournir les données de base essentielles à
l’élaboration de normes d’intervention pouvant favoriser le maintien de l’espèce et des espèces qui
profitent de la présence de gros chicots. Pour tenter de répondre à cet objectif, nous avons tenté de
capturer des femelles adultes lors de leur arrivée sur les sites de reproduction et de les munir
d’émetteurs pour pouvoir suivre leurs déplacements et éventuellement découvrir des cavités
naturelles utilisées pour la nidification.

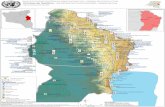
32. AIRES D’ÉTUDE
Deux principales aires d’étude ont été retenues dans des régions représentatives de deux
domaines bioclimatiques particuliers, soit la sapinière à bouleau jaune et la sapinière à bouleau
blanc. Dans le cas de la sapinière à bouleau jaune, on a délimité un territoire d’environ 365 km2
chevauchant la réserve faunique de Portneuf et la zone d’exploitation contrôlée (ZEC) Batiscan-
Neilson, situées au nord-ouest de la ville de Québec (figure 1). Ce territoire a été retenu en raison
du fait qu’un réseau de nichoir y avait été installé par le Service canadien de la faune (SCF) au
cours des années 1980 et qu’il existait donc une population de garrot à œil d’or susceptible de
fréquenter rapidement les nouveaux nichoirs (voir la section 3). La seconde aire d’étude,
représentative de la sapinière à bouleau blanc, est située dans la réserve faunique Ashuapmushuan
(figure 2). Deux secteurs ont été retenus de part et d’autre de la route 167. Le secteur sud, d’une
superficie de 141 km2, est essentiellement couvert de parterres de coupes récentes de grande
superficie où le milieu forestier est limité à des bandes riveraines et à quelques séparateurs de
coupe. Quelques parterres de coupe récente sont aussi présents en bordure du secteur nord (361
km2), mais celui-ci est mieux caractérisé par la présence de jeunes forêts issues de coupes
forestières pratiquées il y a quelques dizaines d’années.


Figure 1. Localisation de l’aire d’étude dans la réserve faunique Portneuf et la ZEC Batiscan-Neilson
5

Figure 2. Localisation de l’aire d’étude dans la réserve faunique Ashuapmushuan
6

7
3. MÉTHODES
3.1 Installation et visite des nichoirs
Au cours du mois de juin 1998, 50 nichoirs artificiels ont été installés dans chacune des aires
d’étude. L’installation des nichoirs à cette période de l’année devait normalement permettre aux
femelles qui font alors de la prospection (Eadie et Gauthier 1985; Zicus et Hennes 1989) de
localiser les nichoirs en prévision de l’année suivante où les travaux de capture étaient prévus.
Ces nichoirs sont fixés sur des arbres localisés en bordure de plans d’eau, à une hauteur variant de
4 à 12 m. Au cours de l'automne 1999, 25 nichoirs supplémentaires ont été installés dans chacune
des aires d'étude. Un seul nichoir était normalement installé par lac afin de réduire les probabilités
de recapture. Le lac Soixante, situé dans la ZEC Batiscan-Neilson et le lac du Coin, situé dans la
réserve faunique de Portneuf, représentent les seules exceptions; deux nichoirs y ont été installés
en raison de leur grande superficie et de l’historique d’utilisation des deux nichoirs installés
auparavant par le SCF sur le lac Soixante.
Chacun des nichoirs était équipé d’un mécanisme de déclenchement activé dès qu’un animal
pénètre à l’intérieur et qui entraîne la fermeture d’une porte bloquant l’issue du nichoir (Zicus
1989). Les mécanismes de déclenchement des systèmes de capture ont été améliorés au cours de
l’hiver 1999-2000 afin d'éliminer les problèmes rencontrés en 1999. Les portes, initialement en
contre-plaqué et placées à l'intérieur du nichoir, ont été remplacées par des portes de plastique
rigide et déplacées à l'extérieur avec le système de glissières afin de réduire les risques de blocage
lors du déclenchement du mécanisme. La corde servant de déclencheur utilisée en 1999 a aussi
été remplacée par un câble d'acier de façon à ce que les écureuils capturés ne puissent plus les
couper.
L’enclenchement des mécanismes a été prévu en fonction de la date d’arrivée des premiers
garrots dans les régions étudiées. Dans le comté de Portneuf, le dégel a été relativement hâtif en
1999, de sorte que les premiers nichoirs ont été activés dès le 12 avril. Au début, nous
soupçonnions que les nichoirs seraient visités par les garrots au fur et à mesure du dégel des lacs
en bordure desquels ils sont situés. Ainsi, seuls les nichoirs placés en bordure des plans d’eau

8
offrant une superficie d’eau libre étaient activés. La découverte, le 20 avril, d’un oeuf de garrot
dans un nichoir localisé en bordure d’un lac complètement gelé indiquait cependant que ces
oiseaux ne nichent pas nécessairement en bordure du plan d’eau qu’ils fréquentent à leur arrivée.
À partir de ce moment, l’ensemble des nichoirs ont été amorcés rapidement et les 50 nichoirs ont
été en opération à partir du 22 avril, et ce jusqu’au 6 mai. Dans la réserve faunique
Ashuapmushuan, les conditions hivernales persistantes du printemps 1999 ont retardé le début de
la saison jusqu’au 29 avril et l’effort de capture s’est prolongé jusqu’au 18 mai. En raison du
faible taux de capture obtenu sur ce territoire au cours de la saison 1999 (voir les résultats), les
travaux des années subséquentes ont été concentrés uniquement dans la région de Portneuf. En
2000, les premiers nichoirs ont été activés le 17 avril. L’ensemble des 75 nichoirs a été amorcé en
deux jours et désamorcé graduellement entre le 3 et le 9 mai. En 2001, cette période d’activité
s’est étendue du 24 avril au 11 mai.
Une fois qu’ils étaient amorcés, les nichoirs étaient visités quotidiennement pour limiter le temps
de captivité des femelles pouvant y être prises. Compte tenu de la grandeur du territoire à couvrir,
de son accessibilité restreinte et de la faible portance de la neige à cette période de l’année, les
visites devaient s'effectuer au moyen d’un hélicoptère.
Dans la région de Portneuf, compte tenu du nombre élevé de femelles capturées, les portes des
nichoirs ont été fermées à la fin de chacune des périodes de capture de façon à empêcher les
femelles marquées de sélectionner ceux-ci pour nicher. Ceci devait normalement inciter les
femelles à utiliser les cavités naturelles existantes comme sites alternatifs de nidification. Par
contre, l’accès aux nichoirs n’a pas été bloqué dans la réserve Ashuapmushuan en 1999. Le
nombre relativement faible de captures obtenu nous a plutôt incité à laisser libre accès aux
nichoirs de façon à augmenter la durée de la période pendant laquelle ceux-ci pouvaient être
visités par des femelles en prospection (Eadie et Gauthier 1985; Zicus et Hennes 1989) pour des
sites de nidification éventuels pour les années subséquentes.

9
3.2 Capture et marquage
Les femelles capturées étaient manipulées et relâchées sur place. Les ailes de chacune d’elles
étaient examinées (Carney 1983) afin de s’assurer que seules les femelles adultes seraient munies
d’émetteurs. Tous les oiseaux étaient munis d’une bague standard du U.S. Fish and Wildlife
Service et d’un émetteur fixé dorsalement au moyen d’un harnais (Dwyer 1972). Trois modèles
d’émetteurs ont été utilisés pendant la durée de l’étude. Le modèle d'émetteur 7pn TSA2000 de la
compagnie Advanced Telemetry Systems Inc. (ATS) a été utilisé au cours des trois années de
marquage. Le modèle RI-2CM, de la compagnie Holohil a été utilisé uniquement en 1999 (à
l’exception d’une seule femelle garrot marquée en 2000) en raison de différents problèmes
nuisant à sa fiablilité. Les premiers avaient une durée de vie prévue de six mois, tandis que les
seconds devaient fonctionner 18 mois. Les deux boucles du harnais étaient fixées autour des ailes,
de part et d’autre de l’émetteur, en croisant le harnais sur la poitrine, laissant ainsi le cou et la
partie abdominale dégagés. Finalement, en 2001, quelques essais ont aussi été effectués avec trois
harles couronnés (Lophodytes cucullatus) qui ont été munis d’émetteurs (Biotrack, modèle TW-5)
fixés sur la queue au moyen de fils et de colle (Giroux et al. 1990; Kenward 1978), tel que
préconisé par Pöysä et Virtanen (1994).
L’effort et le succès de capture ont été exprimés en termes de jours-pièges, cette unité faisant
référence à un potentiel de capture s’échelonnant sur une période de 24 heures pour chacun des
nichoirs. Le nombre de nichoirs déclenchés par des espèces autres que les canards ciblés, de
même que ceux déclenchés accidentellement, a été noté afin d’en tenir compte dans le calcul du
succès de capture (Nelson et Clark 1973).
Au moment de la capture, la masse de chacune des femelles était mesurée (� 5 g) au moyen d’une
balance à ressort de marque Pesola. Différents regroupements de femelles ont été faits selon
l’année et le statut reproducteur afin de procéder à la comparaison de la masse des individus ainsi
regroupés. Les femelles considérées comme nicheuses sont celles qui ont initié un nid, que ce soit
dans une cavité naturelle ou un nichoir. Les femelles dites reproductrices regroupent les femelles
nicheuses, celles qui ont pondu un œuf dans le nichoir lors de leur capture, de même que celles
qui présentaient un cloaque dilaté, indicateur d’une ponte récente. Ces comparaisons ont été faites

10
au moyen des tests de Wilcoxon et Kruskal-Wallis (PROC NPAR1WAY, SAS Institute Inc.
1989). La relation entre la masse des femelles capturées et la date de capture a été examinée au
moyen de la régression linéaire (PROC GLM; SAS Institute Inc. 1989) après vérification de
l'homogénéité de la variance et de la normalité des résidus. Pour toutes ces analyses, seule la
masse à la première capture a été considérée dans le cas des femelles capturées à plus d’une
reprise.
3.3 Suivi télémétrique
Les observations télémétriques étaient effectuées au sol au moyen de récepteurs à balayage et
d’antennes à trois éléments. Afin de pouvoir déterminer le statut des femelles (seules, en couple,
avec une couvée) et de bien caractériser l’habitat fréquenté, des localisations précises étaient
effectuées en se rendant le plus près possible. Cependant, comme le but principal du repérage
était de localiser les sites de nidification, compte tenu des contraintes d’accessibilité et du temps
requis pour se rendre à chacune des femelles marquées, seule une localisation grossière du secteur
fréquenté était faite lorsque le signal obtenu indiquait que les femelles étaient en alimentation.
Ceci était clairement perceptible par la baisse régulière de l’intensité du signal qui survient à
chaque fois qu’une femelle plonge. Quelques survols ont aussi été effectués afin de tenter de
localiser les femelles dont le signal radio n’était plus audible au sol.
Au début des travaux, un équipement de grimpeur (éperons, ceinture) était utilisé pour tenter
d’accéder aux cavités naturelles trouvées au moyen de la télémétrie et de vérifier le nombre
d’oeufs présents. Cette méthode a dû être mise de côté pour des raisons de sécurité. À partir de
l’année 2000, un système de caméra a été utilisé pour procéder à l'inspection des cavités
susceptibles d'être localisées. Une caméra infrarouge (Channel Vision, modèle 5007 CCTV)
permettant d'obtenir des images d'une très grande clarté même en absence de lumière était fixée à
l'extrémité d'une perche télescopique. Cette perche était manipulée de façon à pouvoir introduire
la caméra infrarouge dans la cavité. La taille compacte de cette caméra (50 mm x 50 mm x 60
mm) permet l'inspection de cavités de dimensions relativement réduites. Cette caméra était reliée
par des câbles à un caméscope qui permettait de visualiser les images au sol et de bien orienter
l'objectif dans la direction souhaitée pour bien voir l'intérieur de la cavité. Le caméscope

11
permettait aussi l'enregistrement des images sur une cassette pour un visionnement ultérieur sur
écran de téléviseur à l'aide d'un magnétoscope. Des échelles ont aussi dû être utilisées lorsque les
images fournies par la caméra semblaient indiquer la présence d'œufs qui ne pouvaient être
dénombrés du fait qu'ils étaient couverts de duvet.
3.4 Visite de cavités utilisées antérieurement par le garrot à œil d'or
Les trois chicots où des nids de garrot à œil d'or avaient été localisés en 1999 ont été visités en
2000 afin de vérifier s’ils étaient à nouveau utilisés. Le système de caméra a d'abord été utilisé
pour vérifier si la cavité était à nouveau fréquentée et, dans l'affirmative, une échelle a ensuite été
utilisée pour tenter de vérifier le nombre d'œufs et le statut du nid (abandon ou non). Aucune
visite des cavités trouvées en 1999 et 2000 n’a été effectuée en 2001.
3.5 Taux d'occupation du réseau de nichoirs dans la réserve faunique Ashuapmushuan
Les nichoirs de la réserve faunique Ashuapmushuan ont été inspectés entre le 19 et le 21 juin
2000, et entre le 28 et le 31 mai 2001. La présence d'œufs et/ou de coquilles et membranes était
notée de même que l'espèce ayant utilisé le nichoir. À la date tardive où les visites ont été
effectuées au printemps 2000, la présence d'œufs intacts et froids était considérée comme un
indice de l'abandon du nid. À chacune des visites, les nichoirs étaient nettoyés afin de favoriser la
réutilisation éventuelle du nichoir au cours de la prochaine saison.

12
4. RÉSULTATS
4.1 Période de capture
4.1.1 Effort et succès de capture
En raison de la date tardive d'installation des 25 nichoirs supplémentaires à l'automne 1999 et du
comportement de prospection des oiseaux qui se déroule normalement au cours de l’été, ceux-ci
ne pouvaient pas vraiment être considérés comme fonctionnels au cours de la saison 2000. Seules
deux femelles de harle couronné ont alors été capturées dans ces nichoirs. Ceux-ci ont donc été
exclus des calculs pour évaluer l'effort et le succès de capture pour l’année 2000, mais ont été
inclus au calcul pour la saison 2001. Dans la région de Portneuf, l’effort de capture s’est
graduellement accru d’une année à l’autre, passant de 693 jours-pièges en 1999 à 1207 jours-
pièges en 2001 (tableau 1). Au total, un maximum de 132 femelles ont été capturées. Cependant,
comme 17 cas de captures de harles couronnés n’ont pas fait l’objet de baguage en 2001, il est
possible qu’une partie des ces oiseaux ait été recapturée au cours de cette saison; un nombre
minimum de 115 captures auraient donc été effectuées. Près de la moitié de ces femelles étaient
des garrots à œil d'or, le harle couronné regroupant le reste des captures. Tous les individus
capturés présentaient le plumage caractéristique de femelles adultes. Pour l’ensemble des
femelles capturées en 1999, le succès de capture a été de 5,5 femelles/100 jours-pièges dans la
région de Portneuf, comparativement à 1,0 femelle/100 jours-pièges dans la réserve
Ashuapmushuan (tableau 1). Dans la région de Portneuf, le succès de capture a été relativement
semblable au cours des deux premières années, pour diminuer à près de la moitié en 2001
(tableau 1).
Au cours d’une même saison, la majorité des femelles ont été capturées à une seule reprise
(tableau 2). La proportion de femelles ayant fait l’objet de recaptures au cours d’une même saison
n’a pas varié significativement d’une année à l’autre, tant pour le garrot (test exact de Fisher,
P = 0,162) que pour le harle (1999 et 2000, P = 0,711). Plusieurs femelles ont été recapturées au
cours d’années subséquentes, deux femelles ayant même été capturées au cours de chacune des
trois saisons (tableau 3). Seulement trois individus (deux harles couronnés et un garrot à œil d’or)
ont été capturés dans le même nichoir d’une année à l’autre. La distance moyenne séparant les

13
deux nichoirs de capture s’élevait ainsi à 2,8 km. Un harle couronné a été capturé à quatre
reprises dans des nichoirs différents en 1999 et à huit reprises dans sept nichoirs différents en
2000 (tableau2). Deux des nichoirs où cette femelle a été prise en 1999 ont permis de la reprendre
en 2000. Le territoire englobant l’ensemble des nichoirs utilisés par cette femelle couvrait une
superficie de 34,3 km2. Soulignons que cette femelle, de même qu’une des deux autres harles
recapturées en 2000, avaient niché avec succès en 1999. L’un des garrots recapturés en 2000 avait
aussi niché avec succès en 1999. Aucun des oiseaux recapturés n’était encore muni des émetteurs
fixés auparavant.
Tableau 1. Effort déployé et succès de capture obtenu avec les nichoirs de capture dans la réservefaunique Ashuapmushuan, 1999, et la région de Portneuf, 1999-2001
Ashuapmushuan1999
Portneuf
1999 2000 2001
Nombre total de jours-pièges 846 693 944 1207
Nombre de garrots à œil d’or 3 18 23 14
Nombre de recaptures de garrot à œil d’or 0 1 7 4
Nombre de harles couronnés 5 17 18 n.d.a
Nombre de recaptures de harle couronné 0 7 12 16
Nombre de nichoirs déclenchésaccidentellement
15 17 42 62
Nombre de rongeurs capturés 3 15 60 75
Nombre d’individus d’autres espècescapturées
2 2 1 4
Succès de capture de garrots à œil d’or(femelles/100 jours-pièges)
0,4 2,8 2,9 1,4
Succès de capture de harles couronnés(femelles/100 jours-pièges)
0,6 2,7 2,3 n.d.
a Parmi les 51 captures de harles effectuées, 17 n’ont pas fait l’objet de baguage, empêchant de savoirs’il s’agissait de nouvelles captures ou de cas de recaptures

14
Tableau 2. Fréquence de capture de femelles de garrot à œil d’or et de harle couronné au cours d’unemême saison dans les nichoirs de la région de Portneuf, 1999-2001
Fréquencede capture
Garrot à œil d’or Harle couronné
1999 2000 2001 Global 1999 2000 2001b Global1 17 17 10 44 12 14 10 262 1 5 4 10 4 1 4 53 0 1 0 1 0 2 0 24 0 0 0 0 1 0 4 1� 5 0 0 0 0 0 1a 0 1
a Cette femelle a été capturée à 8 reprises.bLes valeurs pour 2001 représentent des minima du fait que 17 femelles n’ont pas été baguées, empêchant dedéterminer si celles-ci avaient été capturées auparavant.
Tableau 3. Répartition des recaptures effectuées dans les nichoirs de la région de Portneuf selonles années.
Année de capture Garrot à œil d’or Harle couronné Total1999 et 2000 4 2 61999 et 2001 2 2 42000 et 2001 2 2 41999, 2000 et 2001 1 1 2Nombre total de recaptures 9 7 16
Quelques autres espèces ont été prises dans les nichoirs de capture. L’écureuil roux
(Tamiasciurus hudsonicus) et le grand polatouche (Glaucomys sabrinus) ont été capturés à 150
occasions. Comme ces rongeurs réussissaient à s’échapper la plupart du temps en pratiquant des
trous dans les parois du nichoir ou dans la porte bloquant la sortie, il n’est pas possible de
connaître les proportions exactes des captures attribuables à chacune des deux espèces.
Finalement, deux crécerelles d’Amérique (Falco sparverius) et cinq pics flamboyants (Colaptes
auratus) ont aussi été capturés.
En 1999, à l’exception d’une femelle trouvée morte, tous les garrots (20) ont été munis
d’émetteurs, tandis que 12 harles couronnés ont été marqués, soit huit dans la région de Portneuf
et quatre dans la réserve Ashuapmushuan. En considérant qu’un émetteur a été changé sur une

15
femelle garrot qui a été recapturée, un total de 33 émetteurs ont été utilisés au cours de cette
saison. Parmi ceux-ci, 17 étaient fabriqués par la compagnie Holohil et 16 par la compagnie ATS
(tableau 4). Plusieurs problèmes sont survenus avec les émetteurs Holohil. Quatre des harnais se
sont brisés rapidement et les émetteurs sont tombés des oiseaux. Le signal émis par quatre autres
émetteurs s’est rapidement avéré anormal; les pulsations étaient souvent irrégulières et beaucoup
plus rapides que celles d’un signal normal ou même d’un signal produit lorsque l’option mortalité
est déclenchée. Dans deux de ces cas, l’émetteur produisait même un sifflement continu. L’un de
ces émetteurs a cessé d’émettre après seulement deux semaines et un autre après un mois. Une
femelle munie d’un de ces émetteurs a heureusement pu être recapturée et son émetteur a été
changé pour un émetteur ATS. Finalement, sans qu’on puisse déterminer si c’était à cause des
émetteurs ou au départ des femelles, le signal de six autres émetteurs Holohil a été rapidement
perdu après le marquage, tandis qu’un autre a cessé d’émettre pendant la période d’incubation de
la femelle marquée. Le seul problème connu avec les émetteurs ATS est la perte du signal de
deux d’entre eux en début de saison. La proportion d’émetteurs fonctionnels pour les besoins de
l’étude au cours de la saison 1999 s’est avérée significativement plus élevée chez les émetteurs
ATS (87,5 %) que chez les émetteurs Holohil (11,8 %) (�2 = 18,93, dl = 1, P = 0,001).
Tableau 4. Efficacité des émetteurs utilisés en 1999.
Marque
ATS Holohil
Signal perdu dès le départ 2 6
Signal perdu en cours d’incubation 0 1
Harnais brisé 0 4
Signal défectueux 0 4
Nombre total d’émetteurs posés 16 17
Pourcentage d’émetteurs fonctionnels 87,5 % 11,8 %

16
Au cours de la saison 2000, aucun harle couronné n'a été muni d'un émetteur. Parmi les émetteurs
posés sur les 23 garrots à œil d’or, deux seuls se sont détachés. L'un d'eux, le seul de la
compagnie Holohil à avoir été utilisé en 2000, est tombé à l'eau, fort probablement à cause du bris
du harnais, confirmant les problèmes vécus avec ces émetteurs au cours de la saison 1999. L'autre
émetteur de la compagnie ATS a été récupéré à l'eau. Le harnais était encore intact et il semble
que la femelle a pu s'en libérer sans défaire les fixations. Les autres émetteurs ont tous fonctionné
normalement par la suite. Finalement, au cours de la saison 2001, tous les émetteurs ATS posés
(14 garrots et 8 harles) ont été maintenus. Cependant les trois émetteurs Biotrack fixés sur la
queue de harles couronnés sont tombés rapidement après leur pose.
En 1999, deux femelles garrot ont produit un nid dans des nichoirs installés pour le projet dans la
ZEC Batiscan-Neilson. Le système de déclenchement de ces nichoirs a été bloqué. Les visites des
nichoirs étaient effectuées du haut des airs et, afin de minimiser les coûts associés au survol en
hélicoptère, l’équipe de terrain ne s’attardait pas aux nichoirs dont les portes étaient ouvertes. Les
mécanismes de déclenchement des nichoirs ayant été améliorés par la suite, aucun de ceux-ci ne
s'est bloqué au cours des saisons subséquentes. Aucun nid n'a donc été initié dans les nichoirs de
capture comme cela s'était produit en 1999.
4.1.2 État reproducteur des femelles capturées
La proportion des femelles de garrot ayant pondu un œuf dans le nichoir de capture (tableau 5)
semblait supérieure en 1999 (28 %) et en 2000 (26 %) comparativement à 2001 (7 %), mais cette
différence n’était pas significative (test exact de Fisher, P = 0,325). Seulement trois harles
couronnés ont pondu un œuf dans le nichoir de capture (1 en 1999 et 2 en 2001). La proportion de
femelles garrot en état reproducteur, c’est-à-dire incluant celles qui ont pondu un œuf et celles qui
présentaient une dilatation du cloaque lors de leur capture, n'était pas significativement différente
en 1999 (67 %) et en 2000 (61 %) (tableau 6) (�2 = 0,15, dl = 1, P = 0,702). Les observations sur
l’état du cloaque n’ont pas été notées en 2001, éliminant les possibilités de procéder à de telles
comparaisons.

17
Tableau 5. Nombre de femelles de garrot à œil d’or capturées dans la région de Portneuf ayantpondu ou non un œuf lors de la capture.
Catégorie 1999 2000 2001 Global
Ponte 5 6 1 12
Absence de ponte 13 17 13 43
Total 18 23 14 55
Tableau 6. État reproducteur des femelles de garrot à œil d’or capturées dans la région dePortneuf.
Catégorie 1999 2000 Global
Reproductrices 12 14 26
Non reproductrices 6 9 15
Total 18 23 41
4.1.3 Masse des garrots à œil d’or
En ne considérant que la masse des individus lors de leur première capture, la masse moyenne des
garrots à œil d’or était de 712 g (� 7 g) pour l’ensemble des trois années. Parmi toutes les
comparaisons de masses effectuées selon les différents regroupements (tableau 7), une seule
différence significative a été obtenue : la masse des femelles nicheuses était plus élevée que celle
des femelles non nicheuses au moment de la capture au cours de la saison 2000. Une différence
d’environ 85 g était ainsi obtenue entre les masses moyennes de ces deux groupes de femelles.
Au cours des années 1999 et 2001, aucune relation significative n’existait entre la masse des
garrots à œil d’or et la date de leur capture (1999 : F = 1,248, dl = 1, 16, P = 0,281; 2001 :
F = 0,980, dl = 1, 12, P = 0,341). Par contre, en 2000, une relation négative hautement
significative a été obtenue (F = 10,440, dl = 1, 20, P = 0,004), la masse des femelles capturées
diminuant à un taux d’environ 5,5 g/jour.

18
Tableau 7. Comparaisons (tests de Wilcoxon et de Kruskal-Wallis) de la masse des femelles degarrot à œil d’or capturées dans la région de Portneuf, 1999-2000.
Groupes comparés Masse (g) moyenne (� erreur type) Z ou X2 P
Toutes femelles 1999 712 � 10Toutes femelles 2000 702 � 14Toutes femelles 2001 726 � 10
1,703 0,427
Nicheuses 1999 738 � 39Non nicheuses 1999 709 � 9 1,081 0,280
Nicheuses 2000 775 � 28Non nicheuses 2000 690 � 14 2,156 0,031
Nicheuses 2001 725 � 0Non nicheuses 2001 727 � 12 0,278 0,781
Nicheuses 1999 738 � 39Nicheuses 2000 775 � 28Nicheuses 2001 725 � 0
0,802 0,406
Reproductrices 1999 714 � 12Non reproductrices 1999 706 � 17 0,428 0,630
Reproductrices 2000 705 � 19Non reproductrices 2000 697 � 21 0,335 0,738
4.2 Période de nidification
4.2.1 Proportion des femelles ayant niché
En ne retenant que les individus dont les émetteurs ont tenu assez longtemps pour permettre de
déterminer si celles-ci nichaient ou non, la proportion de femelles ayant niché a été presque deux
fois plus élevée en 1999 (30 %) qu’au cours des deux autres saisons (14 %) (tableau 8). Cette
différence n’est cependant pas significative (test exact de Fisher, P = 0,598). La proportion des

19
harles couronnés ayant niché était aussi similaire en 1999 et en 2001 (tableau 9, test exact de
Fisher, P = 0,613).
Tableau 8. Proportions des femelles de garrot à œil d’or capturées dans la région de Portneufayant initié un nid.
Catégorie 1999 2000 2001 Global
Nicheuses 3 3 2 8
Non nicheuses 7 18 12 37
Total 10 21 14 45
Tableau 9. Proportions des femelles de harle couronné capturées dans la région de Portneuf ayantinitié un nid.
Catégorie 1999 2001 Global
Nicheuses 2 2 4
Non nicheuses 15 8 23
Total 17 10 27
4.2.2 Sites de nidification
Malgré l’inefficacité des émetteurs Holohil utilisés en 1999, cinq nids ont pu être localisés dans
des cavités naturelles, soit trois nids de garrot à oeil d’or et deux nids de harle couronné. Tous ces
nids étaient localisés dans la région de Portneuf. Deux autres harles couronnés marqués ont aussi
niché, mais dans des nichoirs; l’une de ces femelles a utilisé un des nichoirs probablement
installés par les propriétaires de chalets situés en bordure du lac Tessier dans la ZEC Batiscan-
Neilson, tandis que l’autre a niché dans l’un de nos nichoirs de la réserve faunique
Ashuapmushuan dont l’accès n’avait pas été bloqué après la période de capture. Malgré
l’utilisation d’émetteurs plus fiables au cours des deux autres années, seulement sept nids ont pu
être localisés, dont cinq dans des cavités naturelles. Les deux autres nids ont été initiés dans des
nichoirs n'appartenant pas à notre réseau.

20
Les cavités naturelles utilisées par les garrots pour nicher étaient situées à une distance moyenne
de 3,0 km du nichoir où ces femelles avaient été capturées, tandis que la valeur obtenue dans le
cas du harle couronné était de 3,3 km.
Plus de la moitié des nids des femelles marquées étaient dans des cavités naturelles localisées
dans des bouleaux jaunes (Betula alleghaniensis), quatre autres espèces d’arbre abritant le reste
des cavités (tableau 10). Un seul des nids a été trouvé dans un chicot de conifère, soit un nid de
harle dans un sapin baumier (Abies balsamea). La moitié des cavités utilisées étaient des cavités
latérales fort probablement creusées par des grands pics (Dryocopus pileatus). Le diamètre à
hauteur de poitrine (D.H.P.) médian obtenu pour les arbres utilisés par le garrot à œil d'or était de
49 cm, cette valeur étant de 47 cm pour les arbres utilisés par le harle couronné (tableau 7). La
distance médiane entre la cavité de nidification et le plan d’eau le plus proche était de 300 m pour
le garrot à œil d’or et de 135 m pour le harle couronné.

21
Tableau 10. Caractéristiques des sites de nidification utilisés dans la région de Portneuf par lesfemelles marquées.
Espèce Espèce d'arbre Diamètreà hauteur
depoitrine
(cm)
Hauteurde l’arbre
(m)
Hauteur dela cavité
(m)
Type decavité
Distance duplan d’eau
le plusproche (m)
Garrot à œil d’or Bouleau jaune 51 8 7 latérale 630
Garrot à œil d’or Bouleau jaune 69 8 8 cheminée 82
Garrot à œil d’or Bouleau jaune 58 12 11 latérale 83
Garrot à œil d’or Hêtre à grandesfeuilles
49 9 8 cheminée(nœud)
150
Garrot à œil d’or Érable rouge 36 15 7 cheminée 600
Garrot à œil d’or Érable à sucre 37 7 6,5 latérale 1750
Harle couronné Sapin baumier 30 8 6 cheminée 15
Harle couronné Bouleau jaune 50 10 8 latérale 200
Harle couronné Bouleau jaune 44 8 7 latérale 418
Harle couronné Bouleau jaune 84 25 20 branchecreuse cassée
70
4.2.3 Chronologie de nidification
La date d’initiation des nids de garrot à œil d’or a été évaluée grossièrement pour les deux
espèces au moyen d’un rétrocalcul tenant compte d’une période d’incubation de 30 jours et d’un
taux de ponte d’un oeuf par deux jours (Dugger et al. 1994; Eadie et al. 1995), ainsi que de la
date à laquelle les couvées ont été observées pour la première fois et du stade de développement
des canetons dans le cas des femelles ayant mené leur nichée à terme. Comme les harles
couronnés fréquentaient souvent des habitats où il est pratiquement impossible de voir les
canetons (voir la section suivante), cette date n’a pu être déterminée pour tous les individus.
En 1999, l’un des deux nids de garrot ayant été menés à terme aurait été initié le 16 mai, tandis
que l’autre l’aurait été entre le 10 et le 15 mai. Parmi les femelles capturées au cours de cette
même saison, six avaient pondu au moins un oeuf dans le nichoir au moment de leur capture, soit
cinq garrots et un harle. Deux des nichoirs avaient même commencé à être utilisés avant que les

22
mécanismes ne soient amorcés pour la capture en début de saison, de sorte qu’une femelle garrot
a été capturée après avoir pondu deux oeufs, et le harle couronné avait pondu trois oeufs. La
période d’initiation de la ponte de ces oeufs se serait échelonnée du 19 avril au 4 mai. Lorsqu’un
oeuf était présent, l’accès au nichoir était bloqué pendant deux à trois jours afin d’inciter la
femelle à nicher ailleurs. Le suivi de ces femelles n’a mené à la découverte que d’un seul nid. Ce
garrot, qui avait pondu un oeuf dans notre nichoir le 4 mai, s’est déplacé de 7,3 km pour initier
son second nid. La date d’initiation de ce nid a été estimée au 16 mai (voir ci-dessus). Il y a donc
eu un décalage de 12 jours entre le moment de la capture et l’initiation du second nid.
Parmi les trois nids localisés en 2000, un seul a été incubé jusqu’à l’éclosion des œufs. Les
canetons ont été entendus dans le nichoir le 28 juin, indiquant l’éclosion récente puisque ceux-ci
quittent normalement le nid entre 24 et 36 heures après l’éclosion (Eadie et al.1995). En tenant
compte du nombre d’œufs pondus par cette femelle, soit sept, la date d’initiation de ce nid peut
être estimée au 15 mai. Cette femelle avait été capturée à trois reprises avant l’initiation de ce nid.
Ces trois captures avaient été effectuées le 22 avril, le 5 mai et le 8 mai. Une période de sept jours
se serait donc écoulée entre le moment de la dernière capture et l’initiation du nid. Il faut
souligner que, lors de la dernière capture de cette femelle, elle avait alors pondu un œuf dans le
nichoir de capture, indiquant qu’elle était alors prête pour la ponte. Parmi les autres garrots à œil
d’or capturés, cinq avaient aussi pondu un oeuf dans le nichoir au moment de leur capture. Ces
pontes ont été notées aux dates suivantes : 20 avril, 24 avril, 4 mai, 6 mai et 9 mai.
Les deux autres nids initiés au printemps 2000 par les femelles marquées ont été abandonnés
avant la fin de l’incubation. Celui qui a été initié dans une cavité naturelle a été découvert le 24
mai. Le nid n’a pas été inspecté à ce moment, de sorte qu’il n’est pas possible de savoir si la
femelle était alors en ponte ou en incubation. L’inspection ultérieure de ce nid (15 juin) a révélé
la présence de huit œufs abandonnés sous une couche de duvet. Le dernier nid, initié dans un
nichoir, a été découvert le 25 mai. Encore une fois, il n’est pas possible de savoir si la femelle
était alors en ponte ou en incubation. Une visite ultérieure (5 juin) a révélé la présence de 14 œufs
froids dans ce nichoir, soit huit œufs de garrot et six de harle couronné.

23
En 2001, seul un nid de harle, celui situé dans un sapin baumier de faible diamètre, a été
abandonné. Deux des trois autres femelles qui ont niché dans une cavité naturelle avaient
initialement pondu un œuf dans les nichoirs au moment de leur capture. L’une d’elles, un garrot à
œil d’or, a été capturée le 25 avril. Avec une date d’éclosion estimée au 21 juin et sept œufs
pondus, la date d’initiation du nid est estimée au 7 mai. Ceci laisse un délai de 12 jours entre le
moment de la capture et la date d’initiation du nid. L’autre femelle, un harle couronné, a été
capturée à deux reprises et avait pondu un œuf à chacune de ces occasions. L’éclosion des œufs
de la nichée de cinq œufs qu’elle a initiée par la suite est estimée au 25 juin, ce qui permet
d’estimer la date d’initiation de ce nid au 15 mai. Ceci laisse une période de huit jours entre le
moment de la capture et l’initiation du nid. Basé sur une éclosion datée du 3 juillet et d’une
nichée de quatre œufs, le dernier garrot à avoir niché a vraisemblablement initié ce nid autour du
26 mai.
4.3 Période d’élevage
En 1999, des cinq femelles marquées qui ont niché dans des cavités naturelles, seulement trois
ont produit des couvées qui ont pu être suivies un certain temps, soit deux garrots et un harle.
L’émetteur posé sur une femelle garrot a cessé d’émettre avant la fin de l’incubation, tandis
qu’une femelle harle a été tuée par un prédateur pendant cette période d’incubation. Néanmoins,
le harle qui a niché dans un nichoir dans la ZEC Batiscan-Neilson a aussi pu être suivi pendant la
période d’élevage.
L’une des deux femelles garrot a mené sa couvée sur le lac du Coin, dans la réserve faunique de
Portneuf, près duquel était le chicot utilisé pour nicher. La couvée a été suivie sur ce même lac
jusqu’à ce que le harnais se brise, soit pour une période de 11 jours. Le secteur fréquenté était
situé à 560 m du nid. L’autre femelle garrot a élevé sa couvée sur le lac Petit Delaney dans la
ZEC Batiscan-Neilson. Bien que le lac Saverne ait été à moins de 100 m du chicot utilisé pour
nicher par cette femelle, celle-ci a amené sa couvée jusqu’au Petit Delaney, à 950 m, en moins
d’une journée après la sortie du nid. Toutes les observations de ces couvées se sont effectuées en
eau libre, facilitant le dénombrement des jeunes. Les deux couvées comportaient initialement sept
canetons chacune. Un seul des canetons a disparu pendant la période de suivi, soit après environ

24
11 jours. La période de suivi a été de 14 jours dans le cas de la femelle ayant perdu son radio et de
15 jours pour l’autre femelle.
Après la sortie du nid, les harles couronnés se sont rapidement déplacés vers des étangs à castor
relativement récents. L’encombrement d’arbres morts sur les sites fréquentés empêchait toute
approche discrète, de sorte qu’il a été pratiquement impossible de dénombrer les canetons. L’une
des femelles a été tuée par un prédateur une vingtaine de jours après la sortie du nid, sans que les
jeunes n’aient été vus une seule fois. L’étang fréquenté par cette couvée était situé à 910 m du
nichoir où la femelle avait niché. La couvée de l’autre femelle a pu être observée à une occasion
sur le lac Morasse, dans la ZEC Batiscan-Neilson, environ 14 jours après la sortie du nid. La
couvée était alors vraisemblablement en déplacement entre le premier étang à castor où elle avait
été observée, à 810 m du nid, et un deuxième où elle a été observée le lendemain à 1,9 km de
l’étang précédent. C’est donc le seul moment où la couvée a été observée dans un habitat dégagé
permettant l’observation des jeunes. Cinq canetons étaient présents à ce moment. Il s’agissait
d’une couvée mixte, c’est-à-dire qu’elle était composée de trois harles et de deux garrots.
En 2000, comme cela a été mentionné ci-dessus, l’éclosion des œufs du seul nid mené à terme a
eu lieu le 28 juin, date à laquelle les canetons ont été entendus dans le nichoir. L’inspection
subséquente du nichoir a révélé que sept œufs sur neuf avaient éclos. L'un des œufs abandonnés
contenait un embryon, tandis que l'autre était stérile. Cinq jours après l’éclosion, cette couvée a
été localisée sur un élargissement de cours d’eau à environ 1,5 km du nichoir. La femelle n’était
alors accompagnée que de trois canetons. Ce cours d’eau n’étant pas du tout relié au lac où la
nidification a eu lieu, il semble que la seule façon que la couvée pouvait avoir d’y accéder était en
marchant. Finalement, le 5 juillet, soit sept jours après l’éclosion, la femelle a été repérée seule
sur le lac Saverne, en bordure duquel le nichoir où elle a niché était installé.
Parmi les quatre nids localisés en 2001, un nid de harle couronné a été abandonné avant la fin de
l’incubation. Le seul harle ayant mené sa nichée à terme a effectué un déplacement de un
kilomètre avec sa couvée pour se rendre au lac le plus rapproché, soit le lac Bourgeault. Elle y est
demeurée jusqu’au 19 juillet, date à laquelle le suivi a été arrêté, soit pendant une période d’au

25
moins 24 jours. À sa sortie du nid, l’une des couvées de garrot a été localisée sur un cours d’eau à
350 m du nid. La couvée semblait emprunter ce cours d’eau vers l’amont en direction du lac de
l’Étang qui était le lac le plus rapproché. La couvée semble ne pas être restée longtemps sur ce lac
puisque, cinq jours plus tard, elle était localisée sur le lac des Marais situé en amont du précédent.
Les déplacements effectués par cette couvée, en empruntant les cours d’eau reliant ces lacs,
totalisent près de 4,5 km. La couvée est par la suite demeurée sur ce lac jusqu’à la fin du suivi,
soit pendant une période minimum de 23 jours. Le dernier garrot à avoir initié un nid a effectué
un déplacement de 550 m pour se rendre avec sa couvée au plan d’eau le plus proche, soit le
Bassin 1. Elle y est aussi demeurée jusqu’à la fin du suivi et y a ainsi séjourné pendant au moins
14 jours. Il faut souligner que, pendant cette période, cette femelle a été repérée à plusieurs
kilomètres du site d’élevage pour ensuite retourner auprès de sa couvée.
4.4 Inspection des cavités utilisées en 1999 par des garrots
Parmi les trois cavités utilisées en 1999 par des femelles nicheuses de garrot à œil d’or, une seule
a révélé des signes d’utilisation à nouveau au printemps 2000. Une première inspection effectuée
le 19 mai au moyen du système de caméra a permis de déterminer la présence de deux œufs. Lors
d’une visite subséquente, effectuée au cours de la semaine du 5 juin, les images visionnées
indiquaient que les œufs avaient été couverts de duvet et il était impossible de dénombrer ceux-ci.
Lors de la dernière visite (16 juin) les œufs n’étaient toujours pas visibles et l’escalade du chicot
abritant le nid présentait un risque pour la sécurité du personnel en raison de son état avancé de
décomposition. Une échelle a été appuyée à la tête du chicot, sous la cavité, et une pression a été
exercée pour en faire casser la tête. Ceci a permis de constater que la ponte s’était arrêtée à deux
œufs et que l’incubation avait cessé ou n’avait tout simplement pas été initiée.
L’inspection d’une seconde cavité au moyen du système de caméra n’a révélé aucune trace
d’utilisation. La grande profondeur et l’incurvation de la dernière cavité ont empêché toute
inspection adéquate de celle-ci, même avec le système de caméra. Aucun indice extérieur (traces
de duvet ou autres) ne pouvait laisser croire à une réutilisation de cette cavité.

26
4.5 Taux d’occupation du réseau de nichoirs de la réserve faunique Ashuapmushuan
Aucun des 25 nichoirs installés à l’automne 1999 n’a été fréquenté au printemps 2000. Ceux-ci
ont commencé à être utilisés en 2001, année où trois nids y ont été initiés (deux garrots à œil d’or
et un harle couronné). De plus, deux des nichoirs installés en 1998 ont été retrouvés en 1999 sur
des arbres couchés par terre dans des chablis. Parmi les 48 nichoirs potentiels restants pour la
nidification, 19 % ont été utilisés par les canards arboricoles (tableau 11). Un nichoir abritait un
nid en incubation contenant un œuf de garrot et huit œufs de harle. Au moment où les visites ont
été effectuées, le taux d’abandon des nids de garrot a été de 20 % tandis qu'aucun nid de harle n'a
été abandonné. En 2001, le taux d’occupation des 70 nichoirs encore disponibles s’élevait à 24 %.
Au cours des deux années de ce suivi, des nids actifs de crécerelle d’Amérique et d’hirondelle
bicolore (Tachycineta bicolor) ont aussi été observés, et des plumes de pics flamboyants ont été
trouvées dans certains. Finalement, des amoncellements de matériel indiquant une utilisation par
les écureuils ont été notés dans neuf nichoirs.
Tableau 11. Occupation des nichoirs dans la réserve faunique Ashuapmushuan.
Espèce 2000 2001
Garrot à œil d’or 5 8
Harle couronné 4 9
Garrot + Harle 1
Hirondelle bicolore 2
Crécerelle d’Amérique 1 6

27
5. DISCUSSION
5.1 Efficacité des méthodes
5.1.1 Capture et télémétrie
Le nombre relativement élevé de captures de femelles adultes de canards arboricoles obtenu au
cours de cette étude indique clairement que la méthode des nichoirs de capture est efficace. Les
émetteurs utilisés en 2000-2001 ont aussi démontré une très grande fiabilité, éliminant les
problèmes rencontrés au cours de l’année 1999. La très grande majorité des femelles suivies ont
pu être localisées assez régulièrement pour permettre d’atteindre les objectifs de l’étude. Les
quelques cas où le signal ne pouvait être localisé au sol ont facilement été repérés lors des survols
télémétriques en avion.
Malgré que les méthodes utilisées puissent sembler au point, un nombre relativement faible de
cavités naturelles utilisées pour la nidification ont pu être localisées. En 1999, les déboires
avaient été attribués aux nombreux problèmes associés aux émetteurs utilisés. De nombreux bris
de harnais et une faible durée de vie des batteries avaient réduit l’efficacité de la majorité des
émetteurs de marque Holohil, compromettant ainsi les possibilités de localiser des femelles au
nid. Cependant en 2000, les émetteurs utilisés, de la compagnie ATS, ont été très efficaces. Sur
les deux qui sont tombés des femelles marquées, l’un était le seul de la marque Holohil posé en
2000 et son harnais a cédé comme cela s’était fréquemment produit en 1999. L’autre émetteur, de
marque ATS, a été retrouvé intact dans l’eau d’un lac. On soupçonne que la femelle munie de cet
émetteur aurait considérablement maigri au cours de la saison (voir plus loin), de sorte qu’elle a
pu se défaire du harnais qui serait devenu trop ample. En 2001, seuls les trois émetteurs de
marque Biotrack fixés à la queue de harles couronnés sont tombés, continuant à démontrer la
fiabilité des émetteurs de marque ATS.
En plus des problèmes précédents, certains oiseaux ont niché dans des nichoirs n’appartenant pas
à notre réseau. Si ces nichoirs n’avaient pas été accessibles, il est probable que ces femelles
auraient pu tenter de nicher dans des cavités naturelles. L’installation de nichoirs pour favoriser la
nidification de canards connaît une popularité grandissante et il demeure inévitable que certains

28
propriétaires de chalets situés dans notre aire d’étude continuent à en installer et que certaines des
femelles marquées y soient attirées.
5.1.2 Effets possibles des méthodes sur la nidification du garrot
Il serait possible de croire que les méthodes utilisées, tant pour la capture que pour le marquage,
dérangent les femelles ainsi manipulées et nuisent à la reproduction. L’efficacité des nichoirs
pour la capture de garrots a déjà été expérimentée sans que des effets négatifs n’aient été
mentionnés (Eadie et Gauthier 1985; Eadie 1989; Zicus 1989). Cependant, seule l’étude de Zicus
et Riggs (1996), réalisée au Minnesota, couvrait sensiblement la même période de capture qu’au
cours de notre étude, soit la période de ponte et les jours qui la précèdent. Au cours de leur étude,
les femelles capturées n’étaient pas munies d’émetteurs et leur nidification était confirmée
lorsqu’elles utilisaient par la suite des nichoirs de leur réseau. La majorité des femelles capturées
qui ont niché par la suite auraient utilisé un nichoir différent de celui où elles avaient été
capturées (Zicus, comm. pers.). La capture ne semble donc pas empêcher les femelles de nicher,
mais pourrait les inciter à utiliser un site alternatif. Ces auteurs ont pu établir que 36 % des
femelles capturées avant l’incubation auraient ainsi niché. Cette proportion n’est pas
significativement différente de celles obtenues en 1999 (30 %) et en 2001 (14 % ) dans la région
de Portneuf (test exact de Fisher, 1999 P = 1,0, 2001 P = 0,189), mais diffère de celle obtenue en
2000 (14 %) (�2 = 3,16, dl = 1, P = 0,076), bien qu’à un seuil de signification de 0,1. Cependant,
la proportion des femelles nicheuses obtenue par Zicus et Riggs (1996) doit être considérée
comme un minimum puisque, comme leurs femelles n’étaient pas suivies par télémétrie, il ne leur
était pas possible de vérifier si certaines des autres femelles ont pu nicher dans des cavités
naturelles pouvant exister dans leur aire d’étude. Il est donc fort probable que la différence dans
les proportions de femelles nicheuses obtenues au Minnesota et dans la région de Portneuf en
2000 soit bien réelle. Ainsi, compte tenu de l’emploi de méthodes de capture identiques, le fait
que la proportion de femelles nicheuses dans la région de Portneuf ait été similaire en 1999 et en
2001 à celle obtenue au Minnesota en 1999 indique qu’un facteur autre que la capture semble
avoir nui à la nidification au cours du printemps 2000.

29
Au cours des travaux de suivi télémétrique, la très grande majorité des localisations des femelles
marquées ont été faites sur des lacs où celles-ci étaient accompagnées d'un mâle. Ces couples
étaient fréquemment observés sur les mêmes lacs et ne franchissaient pas de grandes distances
lorsqu'ils se déplaçaient. Ce comportement laisse fortement croire que ces individus étaient des
oiseaux reproducteurs qui devaient pondre dans le secteur. De plus, plusieurs femelles capturées
dont le suivi n’a pas mené à la découverte d’un nid avaient pondu un œuf dans le nichoir au
moment de leur capture, et d’autres présentaient une dilatation du cloaque caractéristique de
femelles en cours de ponte. Il n’y avait ainsi aucune différence entre 1999 et 2000 dans la
proportion de femelles en état reproducteur au moment de leur capture. Nous croyons donc que
plusieurs des femelles capturées au printemps 2000 ont ainsi pu initier la ponte mais qu’en raison
des conditions climatiques rigoureuses, la plupart d'entre elles ont abandonné le nid, soit pendant
la ponte, soit en début d’incubation. Deux des nids initiés par les garrots à œil d’or ont été
abandonnés au cours de ce printemps, de même que le nid initié dans une cavité localisée l’année
précédente, alors qu’aucun des nids de garrot initiés en 1999 et en 2001 n’a été abandonné. Selon
Eadie et al. (1995), la principale cause d'insuccès chez le garrot à œil d’or serait effectivement
l'abandon du nid. Dans une étude s'échelonnant sur une période de 20 ans, Fredga et Dow (1984)
ont établi que le nombre de nids abandonnés par les garrots à œil d'or était supérieur au cours des
printemps tardifs. Le dégel tardif connu au printemps 2000 pourrait donc avoir influencé les
résultats.
La date d’initiation des nids trouvés dans les cavités naturelles pourrait sembler relativement
tardive. En effet, l’ensemble des nids localisés de 1999 à 2001 ont été initiés entre le 7 et le 26
mai. Il existe un décalage important entre le moment où les femelles ont été capturées et celui où
ces nids ont été initiés. Des décalages de 7 à 12 jours ont ainsi été observés au cours de nos
travaux. Les cas connus de renidification chez le garrot à œil d’or sont extrêmement rares; un seul
est mentionné dans la littérature avec un décalage de 19 jours entre l’abandon du premier nid et
l’initiation du second (Zicus 1990). Cela pourrait porter à attribuer la nidification tardive de nos
femelles marquées à l’influence combinée de la capture et du marquage. Néanmoins, la durée de
la période comprise entre le premier œuf pondu dans nos nichoirs (19 avril) et la date la plus
tardive d’initiation d’un nid par nos femelles marquées (26 mai) se compare favorablement à la

30
durée de périodes de ponte mentionnées pour le garrot à œil d’or dans des aires d’études situées à
des latitudes similaires (tableau 12).
Tableau 12. Chronologie de la période de ponte du garrot à œil d’or mentionnée dans lalittérature.
Date d’initiation du nid n Endroit Source
Médiane Étendue
4 mai 4 avril-1 juin 159 Colombie-Britannique Eadie et al. 1995
7 mai 16 avril-26 mai 96 Kirkland Lake, Ontario Eadie et al. 1995
11 mai 24 avril-27 mai 60 Sudbury, Ontario Eadie et al. 1995
11-20 mai 7 avril-31 mai Nouveau Brunswick Eadie et al. 1995
Le décalage noté précédemment entre la ponte d’un œuf dans un nichoir de capture et l’initiation
subséquente d’un nid pourrait plutôt être expliqué par un comportement noté chez les femelles
de garrots. Eadie (1989, 1991) mentionne que, au cours d’une même saison, une femelle peut
pondre des œufs dans le nid d’une autre même si elle initie elle-même un nid. Ainsi dans son
étude, sur 33 femelles considérées comme parasites, 10 avaient aussi initié leur propre nid
pendant ou après avoir pondu dans le nid d’une autre femelle. Zicus (comm. pers.) a aussi
observé que certaines femelles peuvent pondre des œufs dans l’eau au pied des nichoirs avant
d’initier leur nid, indiquant que, pour des raisons inconnues, la ponte peut être déclenchée avant
qu’une femelle ne sélectionne un nid. Il est donc possible que les femelles capturées en ponte
aient pu continuer à pondre leurs œufs dans le nid d’autres individus ou ailleurs avant d’initier
leur propre nid. Dans le cas de la femelle pour laquelle un écart de sept jours séparait la capture et
l’initiation du nid, cela impliquerait la ponte de trois œufs supplémentaires pendant cette période.
En incluant les sept œufs de son nid et l’œuf pondu dans le nichoir de capture, cette femelle aurait
pondu un total de 11 œufs, ce qui se rapproche sensiblement de la valeur de 10 œufs rapportée par
Eadie (1989) pour le nombre d'œufs pouvant être pondus en une saison par une femelle qui
parasite. Par contre, dans le cas d’une autre femelle, le même calcul mène à un total de 15 œufs,
ce qui dépasse largement le nombre d'œufs pouvant être pondus par une seule femelle. Il est donc
plus logique de croire que ceci représente un cas de renidification.

31
L’abondance de nourriture a été mentionnée à plusieurs reprises comme un facteur pouvant
influencer la reproduction chez différentes espèces de canards. Bengtson et Ulfstrand (1971) ont
observé que la proportion non reproductrice (adultes n’ayant pas niché) de femelles d'arlequins
plongeurs (Histrionicus histrionicus) peut atteindre près de 90 % lors de saisons caractérisées par
une faible abondance de larves de simuliidés qui constituent la principale source de nourriture
chez cette espèce. La disponibilité de nourriture pendant l'incubation semble aussi influencer la
constance d'incubation des femelles de canards souchets, espèce qui présente un comportement
d’incubation semblable à celui du garrot à œil d’or (MacCluskie et Sedinger 1999). La diminution
de masse de femelles de canards branchus (Aix sponsa) pendant l'incubation peut être accentuée
lors de saisons où la disponibilité de nourriture est réduite (Harvey et al. 1989). L'examen
préliminaire des échantillons d'invertébrés prélevés dans des lacs de la réserve faunique des
Laurentides dans le cadre d'une étude sur les effets des coupes forestières semblait indiquer que
les conditions météorologiques particulières qui ont prévalu au printemps 2000 ont effectivement
eu un effet sur l'abondance des invertébrés dans les lacs étudiés (P. Bérubé, FAPAQ, comm.
pers.). Le printemps 2000 aurait ainsi été caractérisé par un décalage de quelques semaines dans
l'émergence des invertébrés. Bien que l'aire d'étude où cet échantillonnage s'est déroulé ne
présente pas nécessairement les conditions rencontrées dans la région de Portneuf, les résultats
n'en indiquent pas moins que les conditions rigoureuses du printemps 2000 ont eu des
répercussions à plusieurs niveaux et qu'il est fort probable qu'elles aient aussi agi négativement
sur l'effort de nidification des garrots à œil d'or.
L’examen de la masse des femelles capturées peut permettre d’appuyer cette théorie d’un effet du
manque de nourriture au cours du printemps 2000. La période pendant laquelle les captures ont
été effectuées correspond vraisemblablement à la période de ponte chez le garrot à œil d'or. Ceci
est confirmé par le fait que des œufs ont été pondus dans les nichoirs de capture, et ce jusqu'à la
dernière journée de capture au cours de chacune des deux années. La masse moyenne des
femelles capturées dans la région de Portneuf (712 g) est effectivement comparable aux valeurs
mentionnées dans la littérature pour des femelles en ponte ou en début d'incubation, masse qui est
généralement supérieure à ce qui peut être observé plus tard au cours de la saison (tableau 13).

32
Les garrots à œil d'or pondent généralement de six à neuf œufs pesant près de 60 g chacun (Eadie
et al. 1995). La masse totale de la ponte peut ainsi représenter jusqu'à 85 % de la masse de la
femelle. Or il a été démontré que, durant la ponte, la masse des femelles de garrot à œil d'or ne
varie pas significativement (Zicus et Riggs 1996), indiquant que les femelles dépendent alors
grandement de sources exogènes de nourriture pour répondre à leurs besoins énergétiques.
Conformément aux résultats de ces auteurs, la masse des femelles capturées dans la région de
Portneuf n'a pas varié significativement pendant les saisons 1999 et 2001. Cependant, un déclin
significatif a été observé au cours du printemps 2000, la masse des femelles capturées diminuant
de 5,5 g/jour. Ainsi, en 2000, la masse des femelles nicheuses était significativement plus élevée
que celle des non nicheuses alors qu’aucune différence n’a été obtenue entre les masses de ces
deux groupes en 1999 et en 2001. Ceci est un indice que seules les femelles possédant les
meilleures réserves énergétiques à leur arrivée ont pu initier un nid au cours de la saison 2000.
Les conditions météorologiques particulièrement rigoureuses du printemps 2000 peuvent ainsi
avoir eu un effet négatif sur la disponibilité des invertébrés qui constituent la base de
l'alimentation chez cette espèce. Il a été démontré, chez le canard branchu, que la survie des
femelles est reliée à leur masse à la fin de l’incubation (Hepp et al. 1990).
Comme la masse des femelles de garrot à œil d'or diminue graduellement pendant l'incubation
(Mallory et Weatherhead 1993; Zicus et Riggs 1996), il est possible que le succès de leur
nidification soit grandement influencé par leur masse à la fin de la ponte. Une diminution trop
considérable de la masse des femelles de garrots à œil d’or pourrait ainsi inciter celles-ci à cesser
l'incubation afin d'éviter de risquer leur propre survie (Mallory et Weatherhead 1993). Donc, les
éléments suivants tendent à faire ressortir les conditions météorologiques du printemps 2000 pour
expliquer, du moins en partie, le faible taux de nidification obtenu au cours de ce printemps :
1) une proportion de femelles nicheuses (14 %) en 2000 significativement inférieure à celle
obtenue dans le cadre d’une étude similaire réalisée au Minnesota (36 %);
Tableau 13 . Masses de femelles de garrot à œil d'or mentionnées dans la littérature selon le stadede la période de reproduction et le statut reproducteur.

33
Stade de la périodede reproduction
Statut des femelles Masse n Source
Nicheuses 768 ± 9 16 Zicus et Riggs 1996Pré-ponte et ponteInconnu (peut incluredes femelles nicheuseset/ou non nicheuseset/ou qui parasitentd'autres nids)
733 ± 8 29 Zicus et Riggs 1996
Nicheuses couvantensuite
693 ± 16 10 Eadie 1989Ponte
Parasites (ne couvantpas)
659 ± 19 6 Eadie 1989
Incubation (début) Nicheuses 698-715 ? Zicus et Riggs 1996
Incubation (global) Nicheusesexpérimentées
717 ± 18 5 Mallory et al. 1994
Premières nidifications 688 ± 9 19 Mallory et al. 1994Nicheuses 635 ± 10 14 Eadie et Gauthier 1985Nicheuses 660 ± 7 19 Zicus et Hennes 1989Nicheuses sans succès 577 ± 3 44 Zicus et Hennes 1989Juvéniles non nicheuses 581 ± 6 18 Zicus et Hennes 1989
Incubation (fin) Nicheuses 616 20 Zicus et Riggs 1996Non nicheuses (juvéniles+ nidification ratée)
575 ± 7 17 Eadie et Gauthier 1985
Élevage En prospection dans desnichoirs
613 ± 13 6 Zicus et Hennes 1989
Prises au filet sur leslacs
613 ± 11 12 Zicus et Hennes 1989
2) un déclin significatif de la masse des femelles capturées en 2000, contrairement à
l’absence de tendance obtenue aux printemps 1999 et 2001 ainsi que dans une autre étude
réalisée au Minnesota ;
3) l’abandon fréquent de nids observés en 2000 alors qu’aucun nid de garrot n’a été
abandonné au cours des autres saisons;

34
4) le décalage observé au printemps 2000 dans l’abondance des invertébrés dans les
échantillons prélevés dans les lacs de la réserve faunique des Laurentides;
5) l'absence notée de couvées de garrot sur le territoire dans les semaines qui ont suivi
l’éclosion.
Néanmoins, malgré de meilleures conditions au cours du printemps suivant, un nombre encore
très faible de femelles nous ont mené à la découverte de cavités de nidification. Il semble donc
que d’autres facteurs viennent influencer nos résultats et, parmi ceux-ci, il faut considérer le
comportement de parasitisme des canards arboricoles.
Le taux de parasitisme serait vraisemblablement relié directement à la disponibilité de cavités
propices à la nidification (Eadie 1989, 1991) et certains indices laissent croire que cette
disponibilité serait relativement limitée dans la région de Portneuf. Premièrement, les données
d’utilisation des nichoirs installés antérieurement par le SCF démontraient une fréquence
relativement élevée de nichoirs contenant plus de 10 œufs (J. Rodrigue, SCF, données non
publiées), ce qui est un indice de parasitisme intraspécifique (Eriksson et Andersson 1982). Ces
mêmes nichoirs renfermaient aussi régulièrement des œufs de garrot à œil d’or et de harle
couronné. Un de ces nichoirs a même déjà contenu un total de 26 œufs, soit 13 de chacune des
deux espèces. La découverte en 1999 de deux canetons de garrot dans l’une des couvées de harle
couronné suivies confirme la persistance du parasitisme interspécifique dans la région.
Deuxièmement, les distances obtenues entre le nichoir de capture et la cavité utilisée pour nicher
(médiane de 2,5 km, maximum de 7,3 km) par nos femelles marquées étaient relativement plus
élevées que celles mentionnées ailleurs. Dans les populations établies dans des réseaux de
nichoirs, les femelles utilisent souvent le même nichoir d’une année à l’autre (Johnson 1967;
Dow et Fredga 1985; Savard et Eadie 1989). Dans leur étude réalisée en Suède, Dow et Fredga
(1983) mentionnent que les femelles qui changeaient de nichoirs nichaient en moyenne à 0,75 km
du nichoir utilisé l’année précédente. Les femelles nichant pour la première fois le font aussi
généralement à moins d’un km du nichoir où elles sont nées (Dow et Fredga 1983 ; Pöysä et al.
1997). Les distances plus considérables que nous avons obtenues pourraient représenter un indice
d’une faible disponibilité de cavités propices à proximité des nichoirs dans notre aire d’étude.

35
Mais les faibles distances mentionnées ci-dessus proviennent toutes d’études où une grande
disponibilité de cavités était maintenue artificiellement au moyen de nichoirs et ne sont pas
nécessairement représentatives de ce qui prévaut dans des conditions naturelles. Une étude
récente sur le canard branchu et le harle couronné indique d'ailleurs que les femelles de ces
espèces capturées dans des nichoirs peuvent ensuite parcourir jusqu'à 6,7 km pour nicher (Blums
et al. 2000).
Sans pouvoir totalement exclure un effet possible des harnais sur le comportement reproducteur
des femelles marquées, nous croyons que, comme l'avaient conclu Pietz et al. (1993), si ceux-ci
ont vraiment un effet, celui-ci se fait surtout sentir au cours de saisons où les oiseaux subissent
déjà un stress causé par des conditions environnementales plus rigoureuses.
5.1.3 Examen des nids dans les cavités naturelles
Le dénombrement des œufs est essentiel à la détermination du succès de nidification. L’obtention
du succès de nidification de femelles nichant dans des paysages forestiers différents permettrait
des comparaisons entre ceux-ci et aiderait à identifier les effets possibles des pratiques forestières
particulières à chacune des régions étudiées. Or, la seule cavité accessible trouvée en 1999 était
trop profonde pour que les œufs puissent être visibles, et l’escalade des autres chicots était trop
risquée. Ces complications avaient mené à la recommandation de la mise au point d’un système
de caméra pour procéder à l’inspection des cavités.
Le système utilisé en 2000 et 2001 s’est avéré fort utile pour déterminer si une cavité était
utilisée. Comme les femelles recouvrent leurs œufs de duvet avant de quitter le nid pour aller
s’alimenter, il faut que les visites coïncident avec la présence de la femelle au nid. Celle-ci étant
alors dérangée, elle ne prend pas le temps de recouvrir les œufs de duvet. Afin de réduire les
risques d’abandon du nid en raison du dérangement, il faut que ces visites soient planifiées au
cours des deux dernières semaines d’incubation.
5.2 Nidification

36
Dans une monographie récente faisant la synthèse des connaissances sur le garrot à œil d’or, il a
été souligné que la très grande majorité des études récentes sur l’espèce ont été réalisées sur des
populations se reproduisant dans des nichoirs (Eadie et al. 1995). Le même phénomène semble
aussi s’appliquer au harle couronné (Dugger et al. 1994). Ceci fait en sorte que les connaissances
sur les sites de nidification utilisés habituellement par ces espèces en conditions naturelles sont
relativement limitées. En Amérique du Nord, les seules études publiées qui donnent des détails
concernant les arbres utilisés par des garrots à œil d’or ont été réalisées dans les plaines
inondables de la rivière Saint-Jean au Nouveau-Brunswick (Carter 1958; Prince 1968) où les
habitats diffèrent considérablement de ceux rencontrés dans les forêts plus nordiques. Néanmoins,
certaines caractéristiques méritent de faire l’objet de comparaisons avec nos résultats.
Au Nouveau-Brunswick, Prince (1968) précise que 56 % des nids étaient localisés dans des
érables argentés (Acer saccharinum), 38 % dans des ormes (Ulmus sp.) et 6 % dans des noyers
(Juglans cinerea). Le diamètre moyen des arbres utilisés par le garrot était de 66 cm. L’étude de
Carter (1958) indiquait que 89 % des nids trouvés dans des arbres étaient dans des érables
argentés et des érables rouges (Acer rubrum). Étonnamment, cet auteur rapporte un diamètre
moyen de huit pouces (20 cm) pour ces arbres, ce qui laisse croire à une erreur car cette valeur
correspond sensiblement au diamètre de l'intérieur des cavités mentionné par Prince (1965) pour
la même région, soit 6 à 10 pouces (15 à 25 cm). Il semble que les garrots n’exercent pas
nécessairement une sélection pour les espèces d'arbres mentionnées ci-dessus, mais que les
cavités disponibles se retrouvent essentiellement dans celles-ci. Dans la région de Portneuf, les
cavités utilisées par les canards arboricoles étaient localisées dans une bonne diversité d’espèces
d’arbres et les diamètres obtenus étaient similaires à ceux mentionnés au Nouveau-Brunswick.
Les chicots de cette taille fournissent vraisemblablement des cavités dont les parois plus épaisses
assurent une meilleure isolation du nid et réduisent les risques de bris de l’arbre au niveau de la
cavité (Miller et Miller 1980; Raphael et White 1984).
Les essences sélectionnées pour la nidification sont donc susceptibles de varier d’une région à
l’autre selon leur disponibilité. Le seul nid de garrot à œil d’or que nous ayons pu localiser dans la
sapinière à bouleau blanc a été trouvé à l’est de la réserve faunique Ashuapmushuan dans le cadre

37
de travaux exploratoires (Maisonneuve et al. 1998). Ce nid était situé dans un peuplier faux-
tremble (Populus tremuloides) de 38 cm de diamètre. Cette espèce d’arbre semble être
particulièrement utilisée en forêt boréale. En effet, dans le cadre d’une étude réalisée en Alberta
sur l’habitat utilisé par le grand pic (Dryocopus pileatus) (Bonar 2000), 93 % des cavités utilisées
par le garrot à œil d’or et le garrot d’Islande étaient dans des peupliers faux-trembles, la seule
autre essence utilisée par les garrots étant le peuplier baumier (Populus balsamoides) (R.L.
Bonar, comm. pers.). De nombreuses études réalisées en Amérique du Nord ont d’ailleurs
démontré l’importance relative du tremble pour la nidification de plusieurs espèces d’oiseau de
cavité (Scott et al. 1980; Winternitz et Cahn 1983; Peterson et Gauthier 1985; Runde et Capen
1987; Li et Martin 1991; Dobkin et al. 1995; Martin et Eadie 1999), indiquant que, dans les
régions où le peuplier faux-tremble est présent, il peut être particulièrement important.
En Alberta, le diamètre moyen des arbres dans lesquels ont été localisés des garrots des deux
espèces était de 45,8 cm, cette valeur étant de 43,6 cm pour les arbres utilisés seulement par le
garrot à œil d’or (Bonar, comm. pers.). Le plus petit diamètre d’un arbre utilisé par le garrot à œil
d’or était de 36,8 cm, valeur relativement rapprochée de ce que nous avons trouvé près de la
réserve faunique Ashuapmushuan. L’atlas des oiseaux nicheurs de l’Ontario (Peck et James 1987)
fait aussi mention de neuf cas de nidification; huit nids ont été localisés dans des peupliers et un
autre dans un bouleau. Le seul diamètre indiqué par ces auteurs était de 35,5 cm. Les seules
données disponibles concernant les arbres utilisés par le harle couronné proviennent aussi de cet
atlas. On y précise que cette espèce niche tant dans les feuillus que les conifères et on mentionne
des diamètres de 20,5 cm et de 45,5 cm pour les arbres utilisés. Ces observations confirment nos
résultats en ce sens que cette plus petite espèce semble pouvoir utiliser des cavités de plus petite
taille, ce qui lui permettrait de nicher à l’occasion dans des conifères.
Le grand pic, qui creuse au moins une nouvelle cavité de nidification par année (Gauthier et
Aubry 1995) est pour cette raison souvent considéré comme une espèce-clé des communautés
d’oiseau puisque plusieurs autres espèces dépendent directement de ces cavités (Daily et al. 1993;
Newton 1994). Nos résultats démontrent que les canards arboricoles profitent aussi de la présence
du grand pic, la moitié des nids de garrot à œil d’or et de harle couronné ayant été localisés dans
des cavités creusées par cette espèce. Les travaux de Bonar (2000) en Alberta soulignent aussi

38
l’importance des cavités creusées par le grand pic pour les autres espèces, dont les garrots. Deux
des sept nids de garrot à œil d’or et quatre des 11 nids de harle couronné localisés dans le cadre
de l’atlas des oiseaux nicheurs d’Ontario étaient dans des cavités creusées par des grands pics
(Peck et James 1987). Le pic noir (Dryocopus martius), qui consitue probablement un équivalent
écologique du grand pic en Europe, est aussi considéré comme une espèce-clé sur ce continent et
le garrot à œil d’or utilise aussi les cavités creusées par cette espèce (Johnsson 1993).
Il est souvent mentionné que le garrot à œil d’or et le harle couronné nichent normalement à
proximité de l’eau. Cependant, cette croyance découle probablement du fait que la majorité des
observations proviennent d’études effectuées sur des populations utilisant des réseaux de nichoirs
établis en bordure de plans d’eau. Des études réalisées tant sur le garrot à œil d’or (Pöysä et al.
1999) que sur le harle couronné (Morse et al. 1969 ) ont en effet démontré que, lorsqu’elles ont le
choix entre des nichoirs établis en bordure de plans d’eau et d’autres situés un peu plus loin dans
la forêt, les femelles ont une nette préférence pour les nichoirs les plus rapprochés de l’eau. La
réduction des distances que les couvées ont à franchir sur la terre ferme, diminuant ainsi les
risques de prédation, expliquerait ce comportement. Néanmoins, les conditions offertes par les
réseaux de nichoirs ne sont pas nécessairement représentatives des conditions qui prévalent en
milieu naturel. En sapinière boréale, les gros arbres susceptibles de fournir éventuellement des
cavités de nidification propices au garrot à œil d’or sont beaucoup plus abondants en milieu
mésique et sont pratiquement absents en milieu riverain (Courteau et al. 1997). Dans la région de
Portneuf, les cavités naturelles utilisées par les femelles marquées étaient ainsi situées, en
moyenne, à près de 400 m des plans d’eau les plus rapprochés.
Une étude récente réalisée dans les marécages de la rivière Saint-Jean au Nouveau-Brunswick
indique qu’à la suite des effets de la maladie hollandaise de l'orme et de coupes de bois,
l'abondance de gros arbres a diminué et la densité moyenne de cavités adéquates pour la
nidification de la sauvagine est passée de 5,5 cavités/ha en 1963-1964 à 1,0 cavité/ha en 1991
(Johnsen et al. 1994). Cette diminution a coïncidé avec une réduction de 33 % dans le nombre de
couvées de garrot à œil d'or sur ce même territoire (Johnsen 1992). Compte tenu des résultats
préliminaires récoltés jusqu’à maintenant dans le cadre de nos travaux, il est possible de croire

39
que les pratiques forestières courantes sont susceptibles d’influencer les populations de garrot à
œil d’or en forêt mixte et boréale en réduisant la disponibilité de gros arbres. Parmi les essences
utilisées jusqu’à maintenant par les garrots marqués pour nicher, le bouleau jaune et le hêtre sont
exploités comme bois d’œuvre et comme bois de chauffage, et le peuplier faux-tremble, bien que
non récolté au cours des années 1980, fait maintenant l’objet d’une récolte (Courteau et al. 1997).
Cette récolte de peupliers est maintenant pratiquée de façon intensive, ces arbres étant réduits en
copeaux pour fournir les fabricants de panneaux de particules.
Deux raisons semblent expliquer la présence des chicots utilisés par les garrots dans la région de
Portneuf : 1) les sites étaient trop escarpés pour en permettre l’accès à l’exploitation forestière et
2) ces arbres étaient déjà morts ou trop malades pour être récoltés au moment où les coupes
forestières ont eu lieu dans le secteur. Ces chicots ont cependant une espérance de vie limitée et il
faudrait prévoir des prescriptions de récolte qui permettront d’assurer le maintien d’un nombre
suffisant de gros arbres susceptibles de fournir éventuellement des cavités propices à la
nidification.
Une pratique sylvicole remarquée dans les parterres de coupes effectuées dans la région de
Portneuf est aussi susceptible de nuire à la disponibilité de chicots pour la nidification par les
espèces nichant en cavité. Les plus gros arbres, majoritairement des bouleaux jaunes, d’un
diamètre souvent près de 60 cm, considérés comme « moribonds » et peu intéressants pour la
récolte sont laissés sur place au moment de la coupe et servent temporairement de semenciers.
Mais afin d’accélérer le dépérissement et la chute de ces gros arbres, les exploitants pratiquent la
méthode de l’annelage qui consiste à faire un trait de scie mécanique sur le pourtour du tronc à la
base de l’arbre. Cette pratique fait en sorte que ces arbres sont attaqués par des champignons de
surface au lieu d’être attaqués par la carie et ils pourrissent de l’extérieur au lieu de pourrir par
l’intérieur, ce qui aurait pour effet de fournir éventuellement des cavités (M. Darveau, comm.
pers.). L’arrêt d’une telle pratique pourrait être profitable au garrot à œil d’or et aux autres
espèces qui utilisent des cavités pour nicher et s’abriter. Des travaux réalisés en Suède ont
démontré que de tels gros arbres isolés sur des parterres de coupe peuvent effectivement être
utilisés pour la nidification par différentes espèces, dont le garrot à œil d’or qui a obtenu un

40
succès de nidification de plus de 30 % dans un tel environnement (Johnsson 1993). Cette valeur
est cependant beaucoup plus faible que celles généralement mentionnées pour des territoires
vraisemblablement moins perturbés. Ainsi on mentionne des valeurs de succès de nidification de
64 % (Eadie et al. 1995) et de 81 % (Allen et al. 1990; Ludwichowski 1998) pour des populations
de garrot utilisant surtout des nichoirs. Mais les valeurs obtenues chez de telles populations
peuvent ne pas être représentatives de celles qui prévalent lorsque les oiseaux nichent dans des
cavités naturelles. La valeur de 50 % obtenue dans les plaines inondables du Nouveau-Brunswick
(Prince 1965) est ainsi plus faible que les précédentes, mais demeure néanmoins supérieure à
celle mentionnée pour les paysages de coupe forestière de la Suède. Il est donc probable que,
même si les canards arboricoles sont susceptibles de nicher dans des chicots isolés laissés sur des
parterres de coupe, ces sites ne présentent pas nécessairement les conditions idéales pour assurer
un succès. Il est cependant probable que la présence de tels gros arbres pourrait devenir
particulièrement importante dans des secteurs de coupes sélectives ou de jardinage laissant un
meilleur couvert.

41
6. CONCLUSION ET RECOMMANDATIONS
6.1 Méthodes
Les méthodes utilisées antérieurement pour tenter de capturer des femelles adultes de garrot à œil
d’or s’étaient avérées relativement inefficaces (Maisonneuve et al. 1998). Le faible nombre
d’individus capturés, malgré les efforts considérables qui ont été déployés, et le fait que les
femelles capturées étaient souvent des sous-adultes faisaient en sorte qu’il était difficile de songer
à une étude télémétrique pour caractériser l’habitat de nidification et d’élevage de cette espèce en
forêt boréale. Les résultats de nos travaux démontrent clairement que l’utilisation de nichoirs
munis de dispositifs de capture représente une excellente solution aux problèmes rencontrés
jusqu’à maintenant. Non seulement cette méthode permet-elle de capturer un nombre intéressant
d’individus, du moins sur des territoires où les garrots nichent dans de telles structures depuis un
certain nombre d’années, mais elle permet aussi de cibler spécifiquement les femelles adultes.
L'expérimentation de nichoirs à captures multiples récemment utilisés au Missouri pour la capture
de canards branchus et de harles couronnés (Blums et al. 2000) permettrait de vérifier l'utilité de
tels dispositifs pour accroître le nombre de captures. Finalement, l’utilisation d’émetteurs fiables
a démontré qu’il était possible de suivre avec une relative facilité les déplacements de garrots
femelles en période de ponte et d’incubation.
Malheureusement, certains facteurs incontrôlables peuvent agir sur le comportement des oiseaux
marqués et nuire à la nidification de ceux-ci, limitant par le fait même le nombre de nids pouvant
être localisés au moyen de ces méthodes. Les conditions météorologiques variables d’une année à
l’autre peuvent parfois retarder le dégel printanier et entraîner l’abandon de la nidification du
garrot à œil d’or (Fredga et Dow 1984), ce qui s’est vraisemblablement produit au cours du
printemps 2000. La méthodologie utilisée dans notre étude était basée sur le comportement de
prospection des femelles (Eadie et Gauthier 1985; Zicus et Hennes 1989) et sur l’hypothèse selon
laquelle ce comportement est une adaptation leur permettant de prévoir des sites de nidification
alternatifs pour l’année suivante dans l’éventualité où le nid utilisé au cours d’une saison était
détruit. Nous espérions donc que la plupart des femelles, ayant d’abord sélectionné un nichoir
comme site de nidification, se seraient ensuite tournées vers de tels sites de nidification alternatifs
après leur capture dans ces nichoirs. Il a été démontré que le taux de parasitisme chez les canards

42
arboricoles serait inversement relié à la disponibilité des cavités (Eadie 1989, 1991). Le faible
nombre de femelles nous ayant mené à la découverte de sites alternatifs est fort probablement un
indice que les cavités naturelles susceptibles de servir à cette fin étaient pour la plupart déjà
occupées et, que les femelles en état de ponte n’ont eu d’autres choix que de pondre leurs œufs
dans les nids d’autres femelles. Le court laps de temps alors passé dans une cavité par chacune de
ces femelles était alors insuffisant pour permettre la localisation de ces nids.
Initialement, nous avions prévu poursuivre ces travaux sur une seconde aire d’étude localisée
dans la réserve faunique Ashuapmushuan. Des nichoirs y ont été installés depuis le début du
projet et un suivi annuel de leur utilisation était effectué afin de déterminer à quel moment le taux
d’occupation serait suffisant pour prévoir un nombre suffisant de captures. Mais les coupes
forestières couvrent des superficies beaucoup plus importantes sur ce territoire et il est fort
probable que la disponibilité de cavités naturelles dans des arbres de diamètre adéquat se font
encore plus rares que dans la région de Portneuf. Compte tenu des résultats obtenus jusqu’à
maintenant, il est logique de soupçonner que les possibilités de découvrir des cavités naturelles au
moyen de la même méthodologie sont encore plus faibles sur cette aire d’étude. Il n’est donc pas
recommandé de poursuivre les travaux plus longtemps.
Néanmoins, malgré tous ces problèmes, les résultats obtenus demeurent intéressants compte tenu
du fait qu’on ne disposait jusqu’à maintenant d’aucune information publiée quant aux sites de
nidification utilisés par le garrot à œil d’or en forêt mixte et boréale. Ces résultats permettent ainsi
d’élaborer des recommandations quant à des approches potentielles de bonnes pratiques
forestières qui assureraient la protection des chicots pour la faune.
6.2 Pratiques forestières
Le plus petit diamètre répertorié parmi les arbres utilisés pour la nidification par le garrot à œil
d’or (36 cm) pourrait être considéré pour établir le diamètre minimal d’arbre qu’il faudrait
maintenir pour assurer la nidification de l’espèce (Thomas et al. 1979). Cette approche
impliquerait cependant que plusieurs chicots seraient à peine plus gros que la cavité elle-même,

43
celle-ci devenant ainsi compressée, fragile et mal isolée (Conner 1979), pouvant entraîner des
effets négatifs sur le succès reproducteur et, à plus long terme, sur les populations.
L’utilisation du diamètre moyen des chicots utilisés par une espèce peut aussi être considérée
pour établir le diamètre minimal des chicots à conserver (Conner 1979; Hunter 1990). Cette
approche peut aussi être critiquée, comme le souligne l’exemple du grand pic qui niche dans des
arbres de 50 cm de diamètre dans l’ouest de l’Amérique du Nord, mais qui peut s’accommoder
d’arbres de 30 cm dans l’est (Lafleur et Blanchette 1993). Mais l’utilisation du diamètre moyen
des chicots utilisés par l’une des plus grandes espèces nichant en cavité dans une région donnée
pourrait permettre d’assurer la conservation de chicots de taille suffisante pour l’ensemble des
espèces. Les chicots de grands diamètres demeurent sur pied significativement plus longtemps
que les plus petits et ils peuvent fournir ainsi des sites de nidification sur une plus longue période
de temps (Cline et al. 1980; Raphael et White 1984; Schreiber et deCalesta 1992). Une valeur de
50 cm a été obtenue au cours de notre étude comme diamètre moyen pour les cavités utilisées par
le garrot dans la sapinière à bouleau jaune. Bien que basée sur un faible nombre de nids (6), cette
valeur se situe entre celle de 66 cm obtenue dans les plaines inondables du Nouveau-Brunswick
(Prince 1968) et les valeurs fournies par Richard Bonar pour l’Alberta (45 cm ). Il serait donc
raisonnable de considérer un diamètre de 50 cm comme diamètre minimal d’arbres à conserver,
d’autant plus que d’autres espèces (chouettes rayée et lapone, grand harle) plus grandes que le
garrot à œil d’or auront aussi besoin de chicots de fort diamètre pour assurer leur nidification. Il
faudrait cependant tenir compte du fait que des chicots de plus faible diamètre peuvent être requis
pour répondre aux besoins d’espèces de plus faible taille (Haggard et Gaines 2001). Un nombre
minimal de chicots ou de gros arbres à conserver sur un territoire donné devrait être conservé
pour assurer le maintien des espèces ciblées et ce nombre est susceptible de varier selon les
domaines bioclimatiques. Les recommandations émises en ce sens par Darveau et Desrochers
(2001) pourraient servir de base à l’élaboration de normes adéquates. Même s’il semble que
l’espèce d’arbre ne semble pas influencer l’utilisation d’une cavité par les canards arboricoles, il
faudrait éviter de considérer des espèces se détériorant rapidement, tel le bouleau à papier, dans
les arbres qui seraient maintenus sur un territoire donné.

44
Dans une revue récente des études ponctuelles effectuées au Québec sur les chicots, Darveau et
Desrochers (2001) en arrivaient à la conclusion qu’il existe un manque flagrant de standardisation
des méthodes d’inventaire visant à déterminer la densité de chicots sur un territoire, plus
particulièrement en ce qui a trait aux classes de diamètre utilisées. Dans plusieurs cas, la plus
grande classe utilisée englobe l’ensemble des chicots de 30 cm et plus. Il devient ainsi difficile de
connaître la contribution des chicots de plus grande taille dans les densités qui sont présentées. Il
serait donc important de développer une technique standard de relevés de chicots qui permettrait
d’évaluer la disponibilité des chicots de 50 cm et plus. Cette évaluation est un préalable dans
l’élaboration de recommandations visant à intégrer aux normes d’intervention en milieu forestier
les besoins des espèces fauniques qui dépendent des chicots de cette taille pour se reproduire.
L’annelage des bouleaux jaunes moribonds, qui semble pratiqué couramment dans la région de
Portneuf, est une pratique qui nuit à la conservation des plus gros arbres susceptibles de fournir
des cavités de nidification adéquates. Le but de cet annelage n’est que d’accélérer la chute de ces
arbres qui ne sont pas récoltés. Une campagne de sensibilisation à l’importance de ces gros arbres
et aux effets négatifs de la pratique de l’annelage devrait être initiée auprès des entreprises et
travailleurs forestiers de la région. Cette approche mènerait vraisemblablement à la conservation
d’un plus grand nombre de gros arbres à cavité dans la sapinière à bouleau jaune. Cependant,
même s’il a été démontré que de tels arbres isolés sur des parterres de coupe peuvent
effectivement être utilisés par les espèces qui nichent en cavité (Johnsson 1993), il serait
important de pouvoir éventuellement comparer le succès reproducteur obtenu sur de tels sites au
succès obtenu dans des chicots entourés de forêt et où les conditions pourraient être plus
favorables.
Finalement, les cavités creusées annuellement par le grand pic semblent particulièrement
importantes pour les canards arboricoles et vraisemblablement pour plusieurs autres espèces qui
nichent en cavité. Le grand pic doit donc être considéré comme une espèce-clé de nos forêts et il
sera particulièrement important d’intégrer les besoins de cette espèce dans l’aménagement
forestier.

45
REMERCIEMENTS
Nous tenons à remercier Philippe Beaupré, Dominique Côté, Raphaël Demers, Rolland Lemieux,
Sylvain St-Onge et Nicolas Wampach pour leur collaboration bien appréciée aux travaux de
terrain, de même que Geneviève Munger, qui a aussi participé aux travaux de terrain, à la
compilation et à la saisie des données. Nous tenons aussi à remercier Louis Villemure de la
Direction de l’aménagement de la faune région du Saguenay-Lac Saint-Jean pour avoir autorisé
un membre de son équipe à poursuivre sa collaboration aux travaux malgré que les activités aient
été concentrées dans la région de Portneuf au cours du printemps 2000 et 2001. Nous remercions
l’unité de gestion de Saint-Félicien du ministère des Ressources naturelles pour l’hébergement
offert dans la réserve faunique Ashuapmushuan pendant la période de capture au printemps 1999.
Nous remercions aussi Jean Rodrigue, du Service canadien de la faune, pour les données relatives
à l’occupation antérieure du réseau de nichoir dans la région de Portneuf, de même que Richard
L. Bonar pour les informations inédites concernant les cavités utilisées en Alberta par le garrot à
œil d’or. Nous remercions finalement Pierre Blanchette pour les commentaires ayant aidé à
améliorer ce document.

46
RÉFÉRENCES
ALLEN, R.B., P.O. CORR and J.A. DORSO. 1990. Nesting success and efficiency of waterfowlusing nest boxes in central Maine : a management perspective. Pages 291-296 In L.H.Fredrickson, G.V. Burger, S.P. Havera, D.A. Graber, R.E. Kirby and T.S. Taylor (Eds.).Proc. 1988 North American Wood Duck Symposium, St. Louis, Missouri.
ANONYME. 1994. Mise à jour du Plan nord-américain de gestion de la sauvagine 1994: Unengagement à la mesure des nouveaux défis. Service canadien de la faune, United StatesFish and Wildlife Service et Mexico Secretaria de Desarrollo Social. 30 p. + annexes.
BALDA, R. P. 1975. The relationship of secondary cavity nesters to snag densities in westernconiferous forests . US For. Serv., Southwestern Region, Wildl. Habitat Tech. Bull. 1 .
BENGTSON, S. and S. ULFSTRAND. 1971. Food resources and breeding frequency of theharlequin duck Histrionicus histrionicus in Iceland. Oikos 22:235-239.
BLUMS, P., C.W. SHAIFFER and L.H. FREDRICKSON. 2000. Automatic multi-capture nestbox trap for cavity-nesting ducks. Wildl. Soc. Bull. 28:592-596.
BONAR, R.L. 2000. Availability of pileated woodpecker cavities and use by other species.Journal of Wildlife Management 64:52-59.
BORDAGE, D. 1987. Suivi des couples nicheurs de canard noir en forêt boréale - 1985.Environnement Canada, Service canadien de la faune, région du Québec. Série de rapportstechniques no 18.
BORDAGE, D. 1988a. Suivi des couples nicheurs de canard noir en forêt boréale - 1986.Environnement Canada, Service canadien de la faune, région du Québec. Série de rapportstechniques no 36.
BORDAGE, D. 1988b. Suivi des couples nicheurs de canard noir en forêt boréale - 1987.Environnement Canada, Service canadien de la faune, région du Québec. Série de rapportstechniques no 35.
BORDAGE, D. 1995. Garrot à oeil d’or, p. 328-331. In Gauthier J. et Y. Aubry (sous la directionde). Les Oiseaux nicheurs du Québec: Atlas des oiseaux nicheurs du Québec méridional.Association québécoise des groupes d’ornithologues, Société québécoise de protection desoiseaux, Service canadien de la faune, Montréal. 1295 p.
BORDAGE, D. et N. PLANTE. 1997. Tendance des effectifs nicheurs de canard noir et de canardcolvert au Québec méridional 1985-1995. Service canadien de la faune, Région du Québec,Série de rapports techniques no 300. 46 p.
BRUSH, T. 1983. Cavity use by secondary cavity-nesting birds and response to manipulations .

47
Condor 85:461-466 .
CARNEY, S. M. 1983. Species, age, and sex identification of Nearctic goldeneyes from wings. J.Wildl. Manage. 47: 754-761.
CARTER, B. C. 1958. The American goldeneye in central New Brunswick . Can, Wildl. Serv.,Wildl. Manage. Bull. Series 2 No 9 .
CLINE, S. P., A. B. BERG and H. M. WIGHT. 1980. Snag characteristics and dynamics inDouglas-fir forests, western Oregon . J. Wildl. Manage. 44:773-786 .
COMITÉ ÉVALUATION ET RECHERCHE DU PCHE-QUÉBEC. 1996. Identification desbesoins en évaluation et recherche au Québec dans le cadre du Plan conjoint des habitats del’Est. Canards Illimités Canada, Service canadien de la faune et Ministère del’Environnement et de la Faune. 27 p.
COURTEAU, M., M. DARVEAU et J.-P. SAVARD. 1997. Effets des coupes forestières sur ladisponibilité de sites de nidification pour le garrot à œil d’or (Bucephala clangula) ensapinière boréale. Série de rapports techniques No 270. Service canadien de la faune,Région du Québec, Environnement Canada, Sainte-Foy. 22 pages.
CONNER, R.N. 1979. Minimum standards in forest wildlife management. Wildl. Soc. Bull.7:293-296.
DAILY, G.C., P.R. ERLICH and N.M. HADDAD. 1993. Double keystone bird in a keystonespecies complex. Proc. Natl. Acad. Sci. USA 90:592-594.
DARVEAU, M. and A. DESROCHERS. 2001. Le bois mort et la faune: état des connaissancesau Québec.
DOBKIN, D. S., A. C. RICH, J. A. PRETARE and W. H. PYLE. 1995. Nest-site relationshipsamong cavity-nesting birds of riparian and snow-pocket aspen woodlands in thenorthwestern Great Basin . Condor 97:694-707 .
DOW, H. and S. FREDGA. 1983. Breeding and natal dispersal of the goldeneye, Bucephalaclangula. J. Anim. Ecol. 52: 681-695.
DOW, H. and S. FREDGA. 1984. Factors affecting reproductive output of the goldeneye duckBucephala clangula. J. Anim. Ecol. 53: 679-692.
DOW, H. and S. FREDGA. 1985. Selection of nest sites by a hole-nesting duck, the GoldeneyeBucephala clangula. Ibis 127: 16-30.
DUGGER, B. D., K. M. DUGGER and L. H. FREDRICKSON. 1994. Hooded merganser(Lophodytes cucullatus) . The Birds of North America, No. 98 (A. Poole and F. Gill, Eds).

48
The Acad. Nat. Sci., Philadelphia, and Am. Ornithol. Union, Washington DC .
DUPUIS, P., A. BOURGET et H. LÉVESQUE. 1995. Estimation des prises d’oiseauxmigrateurs considérés comme gibier par la chasse sportive au Québec pour les années 1973à 1992, p. 1236-1237. In Gauthier J. et Y. Aubry (sous la direction de). Les Oiseauxnicheurs du Québec: Atlas des oiseaux nicheurs du Québec méridional. Associationquébécoise des groupes d’ornithologues, Société québécoise de protection des oiseaux,Service canadien de la faune, Montréal. 1295 p.
DWYER, T. J. 1972. An adjustable radio-package for ducks . Bird-Banding 43: 282-284 .
EADIE, J.M. 1989. Alternative reproductive tactics in a precocial bird: the ecology and evolutionof brood parasitism in goldeneyes. Ph. D. Thesis, Univ. British Columbia, Vancouver. 202p.
EADIE, J. M. 1991. Constraint and opportunity in the evolution of brood parasitism in waterfowl.Acta XX Congr. Int. Ornithol. 2: 1031-1040.
EADIE, J. M. and G. GAUTHIER. 1985. Prospecting for nest sites in cavity-nesting ducks of thegenus Bucephala. Condor 87: 528-534.
EADIE, J. M., M. L. MALLORY and H. G. LUMSDEN. 1995. Common goldeneye, Bucephalaclangula. The Birds of North America 170: 1-31.
ERIKSSON, M. O. G. and M. ANDERSSON. 1982. Nest parasitism and hatching success in apopulation of Goldeneyes Bucephala clangula. Bird Study 29: 49-54.
FREDGA, S. and H. DOW. 1984. Factors affecting the size of a local population of goldeneye,Bucephala clangula (L.), breeding in Sweden. Viltrevy 13:225-255.
GAUTHIER J. et Y. AUBRY (sous la direction de). 1995. Les Oiseaux nicheurs du Québec:Atlas des oiseaux nicheurs du Québec méridional. Association québécoise des groupesd’ornithologues, Société québécoise de protection des oiseaux, Service canadien de lafaune, Montréal. 1295 p.
GIROUX, J-F, D. BELL, S. PERCIVAL and R.W. SUMMERS. 1990. Tail-mounted radio-transmitters for waterfowl. Journal of Field Orntihology 61(3): 303-309.
HAAPANEN, A. 1965. Bird fauna of the Finnish forests in relation to forest succession . Ann.Zool. Fenn. 2:153-196
HAGGARD, M. and W.L. GAINES. 2001. Effects of stand-replacement fire and salvage loggingon a cavity-nesting bird community in Eastern Cascades, Wasington. Northwest Sci.75:387-396.

49
HAMERSTROM, F. F. N. HAMERSTROM and J. HART. 1973. Nest boxes: an effectivemanagement tool for kestrels. J. Wildl. Manage. 37:400-403.
HARVEY, W.F., G.R. HEPP and R.A. KENNAMER. 1989. Body mass dynamics of wood ducksduring incubation: individual variation. Can. J. Zool. 67:570-574.
HEPP, G. R., R. A. KENNAMER and W. F. HARVEY. 1990. Incubation as a reproductive costin female wood ducks. Auk 107:756-764.
HOLT, R. F. and K. MARTIN. 1997. Landscape modification and patch selection: thedemography of two secondary cavity nesters colonizing clearcuts. Auk 114:443-455.
HUNTER, M.L. 1990. Wildlife, forest, and forestry: Principles of managing forests for biologicaldiversity. Prentice Hall, Englewood Cliffs, New Jersey, USA.
JOHNSEN, E.P. 1992. The effects of Dutch elm disease and timber harvesting on cavity-nestingwaterfowl in New Brunswick. M. Sc. Thesis, Univ. New Brunswick, Fredericton.
JOHNSEN, E.P., T.G. DILWORTH and P. KEHOE. 1994. Change in waterfowl nest cavitydensity in a New Brunswick floodplain forest. Écoscience 1:271-274.
JOHNSON, L. L. 1967. The common goldeneye duck and the role of nesting boxes in itsmanagement in north-central Minnesota . J. Minn. Acad. Sci. 34: 110-113 .
JOHNSSON, K. 1993. The Black Woodpecker Dryocopus martius as a keystone species inforest. Swedish University of Agricultural Sciences, Dep. Wildl. Ecol. Institutionen förViltekology, Rapport 24.
KENWARD, R.E. 1978. Radio transmitters tail-mounted on hawks. Ornis Scandinavica 9: 220-223.
LAFLEUR, P.-E. and P. BLANCHETTE. 1993. Développement d'un indice de qualité de l'habitatpour le grand pic (Dryocopus pileatus L.) au Québec . Projet de développement de lagestion intégrée des ressources, Doc. Tech. 93/3. MLCP, 36 pages.
LI, P. and T. E. MARTIN. 1991. Nest-site selection and nesting success of cavity-nesting birds inhigh elevation forest drainages. Auk 108:405-418.
LUDWICHOWSKI, I. 1998. The influence of female age on breeding parameters in commongoldeneyes Bucephala clangula in northern Germany. Der Ornithologische Beobachter95:1-10.
MACCLUSKIE, M. and J. S. SEDINGER. 1999. Incubation behavior of northern shovelers in thesubarctic: a contrast to the prairies. Condor 101:417-421.
MAISONNEUVE, C., R. MCNICOLL, A. DESROSIERS et G. LUPIEN. 1998. Évaluation del’efficacité de méthodes expérimentées pour la capture de canards noirs et de garrots à œil

50
d’or. Ministère de l'Environnement et de la Faune, Direction de la faune et des habitats etDirection régionale du Saguenay - Lac Saint-Jean. 38 p.
MAISONNEUVE, C., R. MCNICOLL, A. DESROSIERS et G. LUPIEN. 1999. Caractérisation del'habitat de reproduction du garrot à œil d'or en forêt boréale. Rapport d’étape interne -activités 1999. Société de la faune et des parcs, Direction de la faune et des habitats etDirection régionale du Saguenay-Lac Saint-Jean. 32 p.
MALLORY, M. L. and P. J. WEATHERHEAD. 1993. Incubation rythms and mass loss of commongoldeneyes. Condor 95:849-859.
MALLORY, M. L., D. K. MCNICOL and P. J. WEATHERHEAD. 1994. Habitat quality andreproductive effort of common goldeneyes nesting near Sudbury, Canada. J. Wildl.Manage. 58:552-560 .
MANNAN, R. W., E. C. MESLOW and H. M. WIGHT. 1980. Use of snags by birds in Douglas-fir forests, western Oregon . J. Wildl. Manage. 44:787-797 .
MARTIN, K. and J. M. EADIE. 1999. Nest webs: A community-wide approach to themanagement and conservation of cavity-nesting forest birds. For. Ecol. Manage. 115:243-257.
MILLER, E. and D. R. MILLER. 1980. Snag use by birds . Pages 337-356 in Management ofwestern forests and grasslands for nongame birds, workshop proceedings. USDA For.Serv. Gen. Tech. Rep. INT-86 .
MORSE, T. E., J. L. JAKABOSKY and V. P. MCCROW. 1969. Some aspects of the biology of thehooded merganser. J. Wildl. Manage. 33: 596-604.
NELSON, L. and F. W. CLARK. 1973. Correction for sprung traps in catch/effort calculations oftrapping results. J. Mamm. 54: 295-298.
NEWTON, I. 1994. The role of nest sites in limiting the numbers of hole-nesting birds: a review .Biol. Conserv. 70:265-276 .
PETERSON, B. and G. GAUTHIER. 1985. Nest site use by cavity-nesting birds of the CaribooParkland, British Columbia . Wilson Bull. 97:319-331 .
PIETZ, P.J., G. L. KRAPU, R.J. GREENWOOD and J.T. LOKEMOEN. 1993. Effects of harnesstransmitters on behavior and reproduction of wild mallards. J. Wildl. Manage. 57:696-703.
PÖYSÄ, H. and J. VIRTANEN. 1994. Habitat selection and survival of common goldeneye(Bucephala clangula) broods - preliminary results . Hydrobiologia 279/280:289-296 .
PÖYSÄ, H., M. MILONOFF V. RUUSILA and J. VIRTANEN. 1999. Nest-site selection inrelation to habitat edge: experiments in the common goldeneye. J. Avian Biol. 30:79-84.

51
PÖYSÄ, H., M. MILONOFF and J. VIRTANEN. 1999. Nest-site selection in relation to habitatedge : experiments in the Common Goldeneye. J. Avian Biol. 30: 79-84.
PÖYSÄ, H., P. RUNKO and V. RUUSILA. 1997. Natal philopatry and the local resourcecompetition hypothesis: data from the Common Goldeneye. J. Avian Biol. 28: 63-67.
PRINCE, H. H. 1965. The breeding ecology of wood duck (Aix sponsa L.) and commongoldeneye (Bucephala clangula L.) in central New Brunswick. Ms Thesis, Univ. NewBrunswick, Fredericton. 109 p.
PRINCE, H. H. 1968. Nest sites used by wood ducks and common goldeneyes in NewBrunswick. J. Wildl. Manage. 32: 489-500.
RAPHAEL, M. G. and M. WHITE. 1984. Use of snags by cavity-nesting birds in the SierraNevada . Wildl. Monogr. 86:1-66 .
ROGERS, J. P., and J. H. PATTERSON. 1984. The black duck population and its management.Trans. North Am. Wildl. Nat. Resour. Conf. 49: 527-534.
RUNDE, D. E. and D. E. CAPEN. 1987. Characteristics of northern hardwood trees used bycavity-nesting birds.
SAVARD, J.-P. and J. M. EADIE. 1989. Survival and breeding philopatry in Barrow's andCommon Goldeneyes. Condor 91: 198-203.
SCHREIBER, B. and D. S. DECALESTA. 1992. The relationship between cavity-nesting birdsand snags on clearcuts in western Oregon . For. Ecol. Manage. 50:299-316.
SCOTT, V. E., J. A. WHELAN and P. L. SVOBODA. 1980. Cavity-nesting birds and forestmanagement . Pages 311-324 in Management of western forests and grasslands fornongame birds, workshop proceedings. USDA For. Serv. Gen. Tech. Rep. INT-86.
THOMAS, J. W., R. G. ANDERSON, C. MASER and E. L. BULL. 1979. Snags Pages 60-77 inThomas, J.W. (Tech. Ed.), Wildlife Habitats in Managed Forests - the Blue Mountains ofOregon and Washington. USDA For. Serv., Agric. Handbook No. 553.
VAN BALEN, J. H. C. J. H. BOOY, J. A. VAN FRANEKER and E. R. OSIECK. 1982. Studieson hole-nesting birds in natural nest sites: availability and occupation of natural nest site.Ardea 70:1-24 .
VON HAARTMAN, L. 1957. Adaptation in hole-nesting birds. Evolution 11:339-347.
WINTERNITZ, B. L. and H. CAHN. 1983. Nestholes in live and dead aspen. Pages 102-106 InSnag Habitat Management: Proceedings of the Symposium. J.W. Davis, G.A. Goodwinand R.A. Ockenfels (tech. coords.). U.S. Dept. Agric., For. Serv., Rocky Mountain For.Range Exp. Stn. Gen. Tech. Rep. RM-99.

52
ZICUS, M. C. 1989. Automatic trap for waterfowl using nest boxes. J. Field Ornithol. 60: 109-
111.
ZICUS, M. C. 1990. Renesting by a common goldeneye. J. Field Ornithol. 61: 245-248.
ZICUS, M. C. and M. R. RIGGS. 1996. Change in body mass of female common goldeneyesduring nesting and brood rearing . Wilson Bull. 108:61-71.
ZICUS, M. C. and S. K. HENNES. 1989. Nest prospecting by common goldeneyes. Condor 91:807-812.