Constitutive Activation ofStat3inFibroblasts Transformed by...
10
Vol. 8, 1267-1276, December 1997 Cell Growth & Differentiation 1267 Constitutive Activation of Stat3 in Fibroblasts Transformed by Diverse Oncoproteins and in Breast Carcinoma Cells’ Roy Garcia, Chao-Lan Yu, Anne Hudnall, Robyn Catlett, Knstie L Nelson, Thomas Smithgall, Donald J. Fujita, Stephen P. Ethier, and Richard Jove2 Molecular Oncology Program, H. Lee Moffitt Cancer Center and Research Institute, and Department of Biochemistry and Molecular Biology, University of South Florida College of Medicine, Tampa, Florida 33612 [A. G., C-L V., A. H., R. C., A. J.]; Eppley Institute for Cancer Research, University of Nebraska Medical Center, Omaha, Nebraska 68198 [K. L N., T. S.]; Department of Medical Biochemistry, University of Calgary Medical Center, Calgary, Alberta, T2N4N1 Canada [D. J. F.]; and Department of Radiation Oncology, University of Michigan Medical School, Ann Arbor, Michigan 48109 [S. P. E.] Abstract Signal transducers and activators of transcription (STATs) were originally identified as key components of signaling pathways involved in mediating responses to IFNs. Previous studies showed that the Src oncoprotein constitutively activates one STAT family member, Stat3. In this study, we investigated STAT activation in a panel of rodent fibroblast cell lines stably transformed by diverse viral oncoproteins. Using a temperature-sensitive mutant of v-Src, we determined that Stat3 is activated within 15 mm of shift from nonpermissive to permissive temperature for cell transformation. This finding indicates that v-Src tyrosine kinase activity is required for Stat3 activation and suggests that Stat3 is proximal to signaling initiated by Src. In addition, Stat3 activation is induced by another nonreceptor tyrosine kinase, v-Fps; by polyoma virus middle T antigen, which activates Src family kinases; and by v-Sis, which acts as a ligand for the platelet-derived growth factor receptor. In contrast, SV4O large T antigen, which transforms cells through different mechanisms, and the v-Ras and v-Ref oncoproteins, which lie in signaling pathways downstream of tyrosine kinases, do not activate Stat3. We did not detect significant activation of Stati , Stat5, or Stat6 in fibroblasts transformed by the viral oncoproteins investigated. Moreover, Stat3 is activated in response to epidermal growth factor (EGF) but not heregulins in immortalized normal human breast Received 5/7/97; revised 9/10/97; accepted 10/2/97. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to mdi- cate this fact. I This work was supported by NIH Grants CA55652 (to A. J.) and CA58667 (to T. S.), DOD Grant DAMD1 7-94-J4382 (to S. E.), and funds from the National Cancer Institute of Canada/CBCRI (to D. F.). 2 To whom requests for reprints should be addressed, at Molecular On- cology Program, H. Lee Moffitt Cancer Center and Research Institute, i2902 Magnolia Drive, Tampa, FL 33612. Phone: (813) 979-6725; Fax: (813) 979-6700; E-mail: [email protected]. epithelial cells. Because constitutive activation of c-Src and EGF receptor kinases is associated with the progression of breast cancer, we examined activation of STATs in human cell lines derived from breast carcinomas. We detected constitutive activation of Stat3 in five of nine breast carcinoma cell lines but not in normal breast epithelial cells. Furthermore, experiments with an EGF receptor-specific inhibitor indicated that the constitutive activation of Stat3 in these breast carcinoma cell lines is not necessarily dependent on signaling through the EGF receptor, although EGF stimulation further increases Stat3 activation. Taken together, our results demonstrate that selective activation of Stat3 is a common event during oncogenic transformation that directly or indirectly involves activation of specific tyrosine kinase signaling pathways. Introduction STATs3 comprise a family of structurally related cytoplasmic proteins that serve as intermediaries in cytokine-dependent signal transduction from the cell surface to the nucleus (i). STATs are latent transcription factors that, following tyrosine phosphorylation and dimerization induced by cytokmne re- ceptor-associated JAKs, translocate to the nucleus, where they directly bind to elements in the promoters of specific cellular genes and regulate their transcription (2). Activation of STATs has also been demonstrated in response to ligands that activate growth factor receptors with tyrosmne kinase activity, such as EGF and PDGF (3-7). These findings sug- gest that activation of STATs participates in mediating growth regulatory signaling induced by specific growth fac- tors. We and others reported previously that constitutive activation of Stat3 occurs in fibroblasts stably transformed by v-Src, the transforming protein of Rous sarcoma virus (8, 9). In addition, activation of specific JAKs and STATs has been demonstrated in cells transformed by human T-cell leukemia virus-I and AbI oncoproteins (iO-i2) as well as various human blood malignancies, including lymphomas and leukemias (i 3-i 5). These results raise the possibility that constitutive signaling by certain STAT proteins may partici- pate in the process of cellular transformation. Aberrantly elevated expression and/or activity of the EGFR family tyrosine kinases, HER1/EGFR and HEA2/neu, and aberrant expression of EGF-like ligands have been associ- ated with the progression of breast and colon carcinomas (16). In addition, activation of c-Src, the normal cellular ho- 3 The abbreviations used are: STAT, signal transducers and activators of transcription; JAK, Janus kinase; EGF, epidermal growth factor; EGFR, EGF receptor; EMSA, electrophoretic mobility shift assay; FBS, fetal bo- vine serum; hSlE, high-affinity sis-inducible element.
Transcript of Constitutive Activation ofStat3inFibroblasts Transformed by...

Vol. 8, 1267-1276, December 1997 Cell Growth & Differentiation 1267
Constitutive Activation of Stat3 in Fibroblasts Transformed byDiverse Oncoproteins and in Breast Carcinoma Cells’
Roy Garcia, Chao-Lan Yu, Anne Hudnall,Robyn Catlett, Knstie L Nelson, Thomas Smithgall,Donald J. Fujita, Stephen P. Ethier, andRichard Jove2
Molecular Oncology Program, H. Lee Moffitt Cancer Center andResearch Institute, and Department of Biochemistry and MolecularBiology, University of South Florida College of Medicine, Tampa,Florida 33612 [A. G., C-L V., A. H., R. C., A. J.]; Eppley Institute forCancer Research, University of Nebraska Medical Center, Omaha,Nebraska 68198 [K. L N., T. S.]; Department of Medical Biochemistry,University of Calgary Medical Center, Calgary, Alberta, T2N4N1 Canada[D. J. F.]; and Department of Radiation Oncology, University ofMichigan Medical School, Ann Arbor, Michigan 48109 [S. P. E.]
AbstractSignal transducers and activators of transcription(STATs) were originally identified as key components ofsignaling pathways involved in mediating responses toIFNs. Previous studies showed that the Srconcoprotein constitutively activates one STAT familymember, Stat3. In this study, we investigated STATactivation in a panel of rodent fibroblast cell linesstably transformed by diverse viral oncoproteins. Usinga temperature-sensitive mutant of v-Src, wedetermined that Stat3 is activated within 15 mm of shiftfrom nonpermissive to permissive temperature for celltransformation. This finding indicates that v-Src
tyrosine kinase activity is required for Stat3 activationand suggests that Stat3 is proximal to signalinginitiated by Src. In addition, Stat3 activation is inducedby another nonreceptor tyrosine kinase, v-Fps; bypolyoma virus middle T antigen, which activates Srcfamily kinases; and by v-Sis, which acts as a ligand forthe platelet-derived growth factor receptor. In contrast,SV4O large T antigen, which transforms cells throughdifferent mechanisms, and the v-Ras and v-Refoncoproteins, which lie in signaling pathwaysdownstream of tyrosine kinases, do not activate Stat3.We did not detect significant activation of Stati , Stat5,or Stat6 in fibroblasts transformed by the viraloncoproteins investigated. Moreover, Stat3 is activatedin response to epidermal growth factor (EGF) but notheregulins in immortalized normal human breast
Received 5/7/97; revised 9/10/97; accepted 10/2/97.The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to mdi-cate this fact.I This work was supported by NIH Grants CA55652 (to A. J.) andCA58667 (to T. S.), DOD Grant DAMD1 7-94-J4382 (to S. E.), and fundsfrom the National Cancer Institute of Canada/CBCRI (to D. F.).2 To whom requests for reprints should be addressed, at Molecular On-cology Program, H. Lee Moffitt Cancer Center and Research Institute,i2902 Magnolia Drive, Tampa, FL 33612. Phone: (813) 979-6725; Fax:(813) 979-6700; E-mail: [email protected].
epithelial cells. Because constitutive activation of c-Src
and EGF receptor kinases is associated with theprogression of breast cancer, we examined activation
of STATs in human cell lines derived from breastcarcinomas. We detected constitutive activation ofStat3 in five of nine breast carcinoma cell lines but notin normal breast epithelial cells. Furthermore,experiments with an EGF receptor-specific inhibitor
indicated that the constitutive activation of Stat3 inthese breast carcinoma cell lines is not necessarilydependent on signaling through the EGF receptor,although EGF stimulation further increases Stat3activation. Taken together, our results demonstratethat selective activation of Stat3 is a common eventduring oncogenic transformation that directly orindirectly involves activation of specific tyrosine kinase
signaling pathways.
IntroductionSTATs3 comprise a family of structurally related cytoplasmicproteins that serve as intermediaries in cytokine-dependent
signal transduction from the cell surface to the nucleus (i).STATs are latent transcription factors that, following tyrosinephosphorylation and dimerization induced by cytokmne re-ceptor-associated JAKs, translocate to the nucleus, wherethey directly bind to elements in the promoters of specificcellular genes and regulate their transcription (2). Activationof STATs has also been demonstrated in response to ligandsthat activate growth factor receptors with tyrosmne kinaseactivity, such as EGF and PDGF (3-7). These findings sug-gest that activation of STATs participates in mediatinggrowth regulatory signaling induced by specific growth fac-tors. We and others reported previously that constitutiveactivation of Stat3 occurs in fibroblasts stably transformedby v-Src, the transforming protein of Rous sarcoma virus (8,9). In addition, activation of specific JAKs and STATs hasbeen demonstrated in cells transformed by human T-cellleukemia virus-I and AbI oncoproteins (iO-i2) as well asvarious human blood malignancies, including lymphomasand leukemias (i 3-i 5). These results raise the possibility thatconstitutive signaling by certain STAT proteins may partici-pate in the process of cellular transformation.
Aberrantly elevated expression and/or activity of the EGFRfamily tyrosine kinases, HER1/EGFR and HEA2/neu, andaberrant expression of EGF-like ligands have been associ-
ated with the progression of breast and colon carcinomas(16). In addition, activation of c-Src, the normal cellular ho-
3 The abbreviations used are: STAT, signal transducers and activators oftranscription; JAK, Janus kinase; EGF, epidermal growth factor; EGFR,EGF receptor; EMSA, electrophoretic mobility shift assay; FBS, fetal bo-vine serum; hSlE, high-affinity sis-inducible element.

A B Cts v-src 3Y1 + IFN’y SRI
I,-
1
�
1 I
��WWL1.I-I--I-#{149}‘-c’’o��o++++++
F 1
2WW�I�I.-
��o++++++
�. WWWWIIIST3
w. #{149} ��STi b� �ST3
-1234567
I Ii234567
1268 Constitutive Activation of Stat3 in Oncogenesis
1 234 5678
mologue of the Rous sarcoma virus-transforming protein,
also occurs at high frequencies in these tumors (i7-2i).
EGFR family and c-Src kinases have been shown to coop-erate in mitogenesis and transformation of model cell lines
(22). In some breast carcinoma cell lines, the constitutiveactivation of c-Src kinase has been associated with activa-
tion of EGFR and/or neu proteins (i9-2i). These findings
suggest a function for activation of c-Src in normal growth
regulation and malignant transformation of breast epithelial
cells induced by signaling through the EGF pathway. Be-
cause STAT activation has been demonstrated to occur in
response to both ligand-dependent activation of EGFR and
constitutive activation of c-Src (3-6, 8, 9), these results fur-
ther suggest the possibility that STAT signaling may be an
important event in malignant progression of human tumors,
especially those such as breast carcinomas, which possess
frequent activation of these kinases.
To explore further the potential role of STATs in oncogen-esis, we have determined the time course of STAT activationby v-Src and examined STAT activity in fibroblasts trans-
formed by other viral oncoproteins. In addition, we have
examined STAT DNA-binding activity in human breast can-
cer cell lines possessing different levels of activation of theEGFR/neu/c-Src proteins. We report that Stat3 activation
occurs rapidly after activation of a temperature-sensitive v-
Src protein, suggesting it is an early event in cellular trans-
formation induced by the oncoprotein. We also show that
constitutive activation of Stat3 occurs in fibroblasts stably
transformed by the v-Fps, v-Sis, and polyoma virus middle T
oncoproteins, all of which directly or indirectly activate tyro-
sine kinase signaling pathways. In addition, we show that
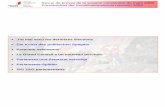
Fig. 1 . Stat3 activation in v-Src-transformed fibroblasts. hSIE-binding activity of nuclear extractsfrom 3Yi fibroblasts transformedwith a temperature-sensitive (ts) v-Src mutant (PAiO4) or wild-typev-Src (SRi) was examined byEMSA at various times after shiftto permissive temperature fortransformation. A, EMSA patternof ts-v-Src-transformed 3Yi nu-clear extracts following tempera-ture shift for various times in mm-utes. B, competition/supershiftcontrols for IFN-’y-stimulated 3Yifibroblasts. C, competition/super-shift controls for SRi cells. ST1,Stati; 5Th, Stat3; *, supershifted
band.
EGF induces rapid activation of STATs in normal breastepithelial cells, whereas heregulins that activate different
EGFR family kinases, including HER2/neu, do not. Further-more, we demonstrate that constitutive activation of Stat3 is
frequent in human breast carcinoma cell lines harboring ac-
tivated EGFR and c-Src kinases. Although the observed
constitutive Stat3 activation is not necessarily dependent on
EGFR signaling, EGF induces further activation of Stat3 inthe breast carcinoma cell lines.
ResultsStat3 Activation Occurs with Rapid Kinetics after Activa-tion of Temperature-sensitive v-Src. To characterize
the kinetics of Stat3 activation by v-Src, the time course of
Stat3 activation in response to activation of a temperature-
sensitive v-Src mutant (23) was examined in 3Y1 rat fibro-
blasts. For these experiments, the cells were incubated atnonpermissive temperature for v-Src kinase and transform-
ing activities, and nuclear extracts were prepared at various
times after shift to permissive temperature. DNA-binding
activity of Stat3 in the nuclear extracts was detected by
EMSA using 32P-labeled hSIE probe, which binds both Stati
and Stat3 with high affinity (8, 24). As shown in Fig. iA,
nuclear extracts prepared from temperature-sensitive v-Src-
transformed cells (Lanes 2-6) exhibited increasing Stat3
DNA-binding activity within iS mm of shift to permissive
temperature (Lane 4), with maximal stimulation occurring
after 60 mm (Lane 6). For comparison, high levels of Stat3
activity were detected in the same cells continuously grown
at permissive temperature (Fig. iA, Lane 7). These results

A B C A B C
v.sis
-
oujw i-.l..
0I*..C �u++ ++
v-fps
+ +++
ST3I ST3
NIH
�
#{149}b-- .‘�ST3
I 2345
3Yi
I-2 E �if a”;. ‘�
z>Q_ >>
#{149}w� ‘4ST3
12345
PymT
�
QC.c �
�
� 4 ST3
�Wwwt.
BALB
00
z> Cl)
4
12312345
12345 1 2 34 5
Cell Growth & Differentiation 1269
Fig. 2. Stat3 activation in NIH 3T3 and BALB/c 3T3 fibroblasts trans-formed by viral oncoproteins. DNA-binding activity of nuclear extractsfrom NIH 3T3 and BALB/c 3T3 fibroblasts stably transformed with viraloncoproteins was examined by EMSA using radiolabeled hSIE probe. A,EMSA pattern of nuclear extracts from normal (N) and transformed NIH3T3 cells. B, competition/supershift controls for v-Sis-transformed NIH3T3 cells. C, EMSA pattern of nuclear extracts from normal (N) andtransformed BALB/c 3T3 cells.
establish that v-Src tyrosine kinase activity is required for
Stat3 activation, and that Stat3 activation occurs rapidly
following activation of the v-Src kinase.
Fig. iB shows results of competition and supershift con-
trols using nuclear extracts from IFN-y-stimulated 3Y1 cells,
which exhibit Stati activation. As shown in Fig. iB, Lanes
2-4, binding to radiolabeled hSlE probe was competed by
excess unlabeled hSIE and to a lesser extent by unlabeled
MGFe probe, which binds to Stat5 (25) and other STATs, but
not by the unlabeled FIRE probe, which does not bind STAT
proteins (8). Fig. iB, Lanes 5-7, demonstrate that anti-Statiantibody specifically blocked the IFN-y-induced binding to
the probe, which is characteristic of this antibody. Fig. i Cshows results of competition/supershift controls for nuclear
extracts from 3Yi cells stably transformed with wild-type
v-Src (SRi cells). The v-Src-induced complex was specifi-
cally supershifted by anti-Stat3 antibody (Fig. iC, Lane 6)
with no effect by anti-Stati or the anti-StatS antibody, which
can supershift Stat5 containing complexes (see below).
These results demonstrate the presence of activated Stati in
IFN-y-treated cells as expected and the presence of acti-
vated Stat3 but not Stati in v-Src-transformed 3Yi fibro-
blasts, consistent with our previous findings (8).
Fig. 3. Stat3 activation in 3Vi fibroblasts transformed by viral oncopro-teins. DNA-binding activity of nuclear extracts from 3Yi fibroblasts stablytransformed with viral oncoproteins was examined by EMSA using radio-labeled hSIE probe. A, EMSA pattern of nuclear extracts from normal (t�and transformed 3Vi cells. B, competition/supershift controls for v-Fps-transformed 3Vi cells. C, competition/supershift controls for polyomamiddle T antigen (Py mT)-transformed 3Vi cells.
Specificity of Stat3 Activation by Viral Oncoproteins.To determine if activation of Stat3 is a universal event duringtransformation of fibroblast cell lines by viral oncoproteins,
DNA-binding activity was measured in a panel of stablytransformed NIH 3T3, BALB/c 3T3, and 3Yi cells. As shownin Fig. 2A, increased Stat3 DNA-binding activity was de-
tected in nuclear extracts of NIH 3T3 cells transformed by thev-Src and v-Sis oncoproteins (Lanes 2 and 3) but not in
extracts of two independent clones of cells transformed bythe v-Ras protein (Lanes 4 and 5). Fig. 2B shows that theDNA-binding activity from v-Sis-transformed NIH 3T3 cells
was specifically competed by unlabeled hSlE probe (Lane 3)
and supershifted by anti-Stat3 antibody (Lane 4). DNA-bind-ing activity was also examined in nuclear extracts obtainedfrom BALB/c 3T3 cells transformed by v-Src and SV4O large
T antigen. As shown in Fig. 2C, elevated Stat3 DNA-binding
activity also occurs in v-Src-transformed BALB/c 3T3 cells
but not in cells transformed by SV4O. These results indicatethat activation of Stat3 is mediated by specific oncoproteins
and is not a universal event in cellular transformation.
These experiments were extended using a panel of 3Y1cells transformed by additional viral oncoproteins. As shown
in Fig. 3A, elevated Stat3 activity was detected in 3Y1 cellstransformed by polyoma virus middle T antigen and v-Fps(Lanes 3 and 4) but not in cells transformed by v-Raf (Lane 5).
Fig. 3, B and C, shows results of competition/supershiftcontrols for nuclear extracts of 3Y1 cells transformed by
v-Fps and polyoma middle T antigen, respectively. Bothcomplexes were specifically competed by unlabeled hSIEprobe (Fig. 3, Lanes 3) and supershifted by anti-Stat3 anti-body (Fig. 3, Lanes 4), confirming the specificity of Stat3
activation by the viral oncoproteins.

A B
NIH 3T3 MCF1OA
C D
MCF-1OA M07e
+EGF +EGF +EPO
F�!W�’
P!�e?, �s#{248}�c�++++
‘-i�:-----’
o�111W#{149}+
#{149}wW��1
�o++++++
Ii 1
�0 �U) U)
I #{149}��8T1:3
.#{149}
�W�STi:3
WW’hI ‘�ST5
1�’ �
123456
I I 234567
I 2345
1270 Constitutive Activation of Stat3 in Oncogenesis
Fig. 4. EGF-induced STAT activ-ity in NIH 3T3/hEGFR fibroblastsand MCF-iOA normal human
C breast epithelial cells. DNA-bind-ing activity of nuclear extractsfrom NIH 3T3/hEGFR and MCF-iOA cells was examined by EMSAusing the hSlE probe after treat-ment with EGF for 1 0 mm at 37CC.A, competition/supershift controlsfor NIH 3T3/hEGFR fibroblaststreated with i6.5 ni,i EGF. B, in-
duction of DNA-binding activity inMCF-iOA cells stimulated with 10nM EGF. C, competition/supershiftcontrols for EGF-treated MCF-i OAcells. D, competition/supershiftcontrols for M07e cells treatedwith erythropoietin using the ra-diolabeled MGFe probe.
The results presented above indicate that those oncopro-teins that constitutively activate Stat3 induce very little or nodetectable Stati activation. To determine if additional STATscould be activated during transformation by any of these viral
oncoproteins, the DNA-binding activities of nuclear extracts
were examined by EMSA using the MGFe probe, whichbinds Stat5 proteins (25), and the 6spA probe, which bindsStat6 (26). Although MGFe-binding complexes were de-tected only in cells transformed by v-Src and v-Fps, thelevels of this DNA-binding activity were very low, and it wasdifficult to confirm the presence of StatS proteins in these
complexes by supershift analysis (data not shown). Further-more, EMSA analysis using radiolabeled 6spA probe did notreveal any specific complexes in v-Src or v-Fps-transformed3Yi cells (data not shown). These results suggest that Stati,StatS, and Stat6 are not significantly activated in fibroblaststransformed by the oncoproteins investigated.
Stat3 Is Activated in Response to EGF Stimulation inNIH 3T3 Fibroblasts and MCF-iOA Normal Human BreastEpithelial Cells. For comparison with human breast epithe-
hal cells, the DNA-binding activity of nuclear extracts pre-
pared from NIH 3T3 cells overexpressing the human EGFRprotein (NIH 3T3/hEGFR cells) was examined following treat-ment with EGF. As expected from earlier studies (3-6), EGFstimulation of NIH 3T3/hEGFR cells induced formation of
three distinct complexes detectable by EMSA (Fig. 4A). Anti-
Stat3 antibody resulted in complete supershifting of theslower migrating complex and partial shifting of the complex
possessing intermediate mobility, which contains a het-erodimer of Stati and Stat3 (Lane 5). The faster migrating
complex was blocked by anti-Stati antibody (data notshown), whereas incubation with anti-StatS antibody had noeffect on migration of the EGF-induced complexes (Fig. 4A,
Lane 6).
To determine if this response is shared with immortalized,normal human breast epithelial cells, DNA-binding activitywas assayed in MOF-i OA cells stimulated with EGF. Asshown in Fig. 4B, three distinct complexes were also de-tected by EMSA using nuclear extracts prepared from EGF-treated MCF-i OA cells (compare Lanes 1 and 2). Thesecomplexes were fully competed by unlabeled hSIE (Fig. 4C,Lane 3), and partial competition was observed using excessunlabeled MGFe probe (Lane 4). Anti-Stati antibody blocked
formation of the two faster migrating complexes (Fig. 4C,Lane 5), whereas anti-Stat3 antibody specifically super-shifted the slowest migrating complex (Lane 6). As a positivecontrol for the anti-StatS antibodies, Fig. 4D shows thattreatment of M07e megakaryoblastic cells with erythropoi-etin resulted in formation of a single complex containing
StatS (25) that was specifically supershifted by anti-StatSantibody (Lane 5). These results demonstrate that EGF-induced activation of Stat3, and to a lesser extent Stati , is aresponse shared by the fibroblast and epithelial cell typesexamined.
To investigate the potential role of other EGFR family ki-nases in STAT signaling in breast epithelial cells, MCF-iOA

A B C
MCF.1OA MCF-1OA MCF-1OA + EGF
�u_�zo�.ow=xo+++
�I).r?z(,..ouJ=IC.)+++
�1O,�Cl)U�
�w0�ee
U+++
�IIIST3
- � AAA� �
I I1234 1234
hSlEProbe MGFoProb12345MGFe Probe
Cell Growth & Differentiation 1271
I-l-� � � I- I.- _ -�
‘�, � ,�, I-. I.- ‘��- ‘�‘ I- -�‘� Z �� _l.__�(o� �<WC,�Q.0.O)o�ocs1�‘�i’;#{149}’’2’t’;- I U � I
�� �;;�i- ��,-u)cocncI) Cfli-U)�I�
Fig. 5. Heregulins do not induce STAT activity in MCF-iOA cells. DNA-binding activity in nuclear extracts from MCF-iOA cells was examined byEMSA using the hSIE and MGFe probes after treatment with 10 n� EGF,i 0 nM heregulin xi ,or 5 n� heregulin j3i for i 0 mm at 37CC. A, EMSApattern using the hSIE probe with nuclear extracts from treated MCF-iOAcells. B, EMSA pattern using the MGFe probe with nuclear extracts fromtreated MCF-iOA cells. C, supershift controls for EGF-stimulated MCF-iOA cells using the radiolabeled MGFe probe.
cells were treated with heregulins ai and pi . As shown inFig. 5, A and B, heregulins did not induce activation of STAT
proteins (Lanes 3 and 4) in these cells, although Stati , Stat3,
and StatS were induced by EGF (Lanes 2), as expected.Supershift analysis with anti-StatS antibody (Fig. SC) con-
firmed the specific induction of StatS by EGF, consistent with
previous reports using other cell types (27). Earlier studies
have shown that MCF-iOA cells respond to heregulin stim-
ulation (28), indicating that these cells express functional
EGFR family receptors, including HER2/neu, which bind
heregulins. These results suggest that STATs represent a
divergence in signaling pathways activated by related Ii-
gands of the EGF family.
Stat3 Is Constitutively Activated in Human BreastCarcinoma Cells. Activation of c-Src and EGFR kinases is
associated with malignant progression of human breast can-
cer. Because these kinases can activate STAT proteins in
other cell types, the DNA-binding activity of nuclear extracts
prepared from model human breast carcinoma cell lines
possessing different levels of EGFR/neu/c-Src activationwas examined by EMSA using radiolabeled hSIE probe. Fig.6 shows that elevated Stat3 activity was detected in five of
nine breast carcinoma cell lines examined (Lanes 3, 7, 8, 9,
and 12) but not in any of three cell lines derived from normal
breast epithelial tissue (Lanes 1, 2, and 1 1).
To determine the identity of proteins present in the com-
plexes detected by EMSA, competition/supershift controls
were performed using nuclear extracts of i 31 S-M02, SK-
1 2 3 4 5 6 7 8 9 10 ii i2
Fig. 6. Breast carcinoma cell lines possess constitutive Stat3 DNA-bind-ing activity. DNA-binding activity of nuclear extracts from cell lines estab-lished from normal breast epithelium and breast carcinoma cell lines wasexamined by EMSA using radiolabeled hSIE probe. N, cell lines obtainedfrom normal breast epitheliurn; T, breast carcinoma cell lines.
BR-3, and MDA-MB-468 cells. As shown in Fig. 7, the com-plexes were specifically competed by unlabeled hSIE probe(Lanes 3) and supershifted by anti-Stat3 antibody (Lanes 5)but not by anti-StatS antibody (Lanes 6), demonstrating ac-tivation of Stat3 in these cells. A second, abundant complex,
which migrated faster in EMSA, was also detected in these
experiments using nuclear extracts from all three cell lines;however, the identity of its components remains unclear. To
assess if additional activated STAT proteins were present inthe breast cancer cell lines, EMSAs were performed using
radiolabeled MGFe probe. Although elevated MGFe-bindingactivity was detected in the same five cell lines possessing
constitutive Stat3 activation, the levels of this DNA-bindingactivity were very low and difficult to confirm by supershift
analysis using anti-StatS antibodies (data not shown). Fur-themiore, EMSAs performed using nuclear extracts from
SK-BR-3 and MDA-MB-468 cells with radiolabeled 6spA didnot reveal formation of Stat6-containing complexes (data notshown). These results are consistent with the profile of acti-

A B
SK-BR-3- #{149}1�100 � � �_ ,_
C� 15 Cl) (I)
gizc� � is0 + + + + +
131 5-M02
- #{149}c’)I�)0 �j�j � �-. �_
- C� C� Cl) Cl)
gc�.c� aC.) + + + + +
S
�w � a 15T3
�. �
a
123456
A
C
MDA-MB-468
� u1uJ if �
ou..cS a a0 + + + + +
0 ��:sT3
.. ..
12 34 S61234�6
F- I- (0a. a. c�i ‘�‘
4 (“1 0) 0 th c’0 0 11) � � � ci:� �� Cl) U) ‘- � � �
U � � � I- �
� o
�
�
1 2 3 4 5 6 7 8a-Phosphotyrosine
Fig. 7. Identification of Stat3 DNA-binding activity in breast carcinomacell lines. Competition/supershift assays were performed with nuclearextracts from breast carcinoma cell lines possessing elevated hSIE-bind-ing activity to determine the identity of STAT proteins present in corn-plexes detected by EMSA.
I 2 3 4 5 6 7
a-Stat3
C �-. � .,. ‘ -P-Stat3
I 2 3 4 5 6 7
a-Phospho-Stat3
Fig. 8. Tyrosine phosphorylation of cellular proteins and Stat3 in breastcarcinoma cell lines. Western blot analyses were performed on whole-celllysates of the indicated cell lines lysed in boiling SOS sample buffer. Equalamounts of total protein were loaded on each lane and, following transferto nitrocellulose, the blots were probed with the indicated antibodies. A,phosphotyrosine content of total cellular proteins. B, steady-state levelsof Stat3 proteins. C, levels of tyrosine-phosphorylated Stat3 protein.
1272 Constitutive Activation of Stat3 in Oncogenesis
B � � - �- � � -Stat3
vated STATs observed above in fibroblasts transformed byviral oncoproteins, suggesting that similar signaling path-
ways may participate in activation of STAT proteins duringbreast cancer progression.
It was of interest to examine whether the activation of
Stat3 detected in extracts from breast carcinoma cells cor-related with an overall increase in tyrosine phosphorylation of
total cellular proteins. Western blot analysis revealed that, in
contrast to v-Src-transformed fibroblasts (SRi), the breastcarcinoma cell lines with elevated Stat3 DNA-binding activ-ities shown in Fig. 6 did not exhibit large increases in overall
levels of phosphotyrosine in cellular proteins compared tonormal breast epithelial cells (Fig. 8A). This finding suggests
that constitutive activation of Stat3 in breast carcinoma cells
is the result of selective tyrosine phosphorylation of Stat3 by
specific tyrosine kinases. Moreover, whereas most of the cell
lines contained detectable levels of Stat3 protein, only breast
carcinoma cells harboring elevated Stat3 DNA-binding ac-tivities also displayed increased tyrosine phosphorylation of
Stat3 protein (Fig. 8, B and C). Taken together, these resultsestablish that constitutive, ligand-independent tyrosinephosphorylation and activation of Stat3 signaling pathways
is a frequent event in human breast carcinoma cell lines.Constitutive Activation of Stat3 in Breast Carcinoma
Cell Lines Is Not Necessarily Dependent on Signaling bythe EGFR Tyrosine Kinase. To more directly examine the
role of the EGFR kinase in constitutive activation of Stat3 in
breast cancer cells, cultures of the SK-BA-3 and MDA-MB-468 cells were pretreated with the pharmacological inhibitor,PD1 58780, which is selective for the EGFR kinase (29). Fol-lowing pretreatment with inhibitor, cells were incubated with
or without EGF, and then STAT activation was analyzed by
EMSA using the hSIE probe. Fig. 9 shows that pretreatmentof both cell lines with PDiS8780 in the absence of exoge-
nous EGF resulted in slightly elevated levels of constitutiveStat3 DNA-binding activity (compare A and B, Lanes 1 and
2). As observed in other cell lines, treatment of SK-BR-3 andMDA-MB-468 cells with EGF induces formation of three spe-cific complexes containing Stati and Stat3 (Fig. 9, Lanes 3).
The formation of EGF-induced STAT complexes, but not theconstitutive Stat3 complex, was abrogated in both cell linesfollowing treatment with PD1 58780 prior to addition of ligand(Fig. 9, Lanes 4). These results indicate that the constitutive
activation of Stat3 observed in the SK-BR-3 and MDA-MB-
468 cell lines is not dependent on signaling by the EGFRkinase.
DiscussionA unique feature of STAT proteins is their capacity to directlylink receptor-induced signals generated at the plasma mem-
brane to transcriptional events in the nucleus. This charac-teristic, and the activation of these proteins in response to
various stimuli involved in mitogenic signaling (3-7, 27), raise
the possibility that STAT proteins may have important roles in
normal cellular growth control as well as malignant transfor-mation. In this report, we demonstrate that increased Stat3DNA-binding activity occurs rapidly after activation of a tem-

A
SK-BR.3
5OnMEGF: - - + +
PD158780: - + - +
- #{149}i�.. =�Jl:3-ST3
MDA-MB-468
5OnMEGF: - - + +
PD158780: - + - +
12 34
hSIE Probe12 34
hSIE Probe
Fig. 9. Inhibition of EGFR kinase activity does not block constitutiveStat3 activation in model human breast epithelial cell lines. DNA-bindingactivity was examined by EMSA using the hSIE probe with nuclear ex-tracts from SK-BR-3 and MDA-MB-468 cells treated with the EGFR se-lective inhibitor, PD1 58780, prior to the addition of EGF. Shown are resultsusing extracts from SK-BR-3 (A) and MDA-MB-468 (B) cells pretreatedwith drug for 15 h prior to stimulation with 50 ns�i EGF at 37CC for 10 mm.
perature-sensitive mutant of v-Src upon shift to permissive
temperature for transformation. Because transformation re-
quires activation of v-Src kinase activity, these results mdi-
cate that v-Src kinase activity is also required for activation
of Stat3. Moreover, these findings suggest that Stat3 acti-
vation may be an early event in signaling leading to transfor-
mation by the Src oncoprotein. Oao et a!. (9) have confirmed
Stat3 activation by v-Src and also demonstrated direct in-
teraction between the v-Src and Stat3 proteins, suggesting
the potential for direct activation of Stat3 by v-Src. Alterna-
tively, the finding that JAK family kinases are activated in
v-Src-transformed cells (30) raises the possibility that v-Src
indirectly activates STATs through activation of JAKs. These
two possibilities are not mutually exclusive, and both are
consistent with the rapid kinetics of Stat3 activation by v-Src.
We also demonstrate in this report that transformation by
polyoma virus middle T antigen and v-Sis, which indirectly
result in activation of c-Src (3i), induces elevated Stat3
DNA-binding activity. These findings support a role for Stat3
activation in the transmission of transforming signals gener-
ated upstream of c-Src. In addition, Stat3 activation was
observed in fibroblasts transformed by the v-Fps tyrosine
kinase but not in cells transformed by v-Ras or v-Raf, which
lie in signaling pathways downstream of tyrosine kinases.
These results suggest that Stat3 activation is not merely a
secondary effect, or consequence, of cell transformation. On
the other hand, Stati , StatS, and Stat6 are not significantly
activated by any of the oncoproteins investigated here, dem-
onstrating specificity of these oncoproteins for activation of
Stat3 in fibroblasts. Taken together, our findings suggest that
activation of Stat3 is not a universal event during cellular
transformation but, rather, may be important to the mecha-
Cell Growth & Differentiation 1273
4 R. Garcia, D. Fujita, S. Ethier, and R. Jove, unpublished results.
B nisms of transformation of specific oncoproteins that directly
or indirectly activate tyrosine kinases.
____________ Constitutive activation of EGFR and neu receptor proteins
as well as c-Src has been detected in a large percentage of
breast tumors (16). Because the EGFR mitogenic pathway
has been previously linked to activation of STATs by othersw W � � E�:3 (3-6) and previous studies demonstrated that Stat3 is asso-
ciated with cellular transformation induced by activated c-
es � Src (8, 9), it was of interest to examine DNA-binding activity
, V I in nuclear extracts obtained from normal human breast ep-ithelial and breast carcinoma cell lines. Initially, the DNA-binding activity of MCF-i OA immortalized normal breast ep-
ithelial cells was examined following treatment with EGF to
determine if ligand-dependent activation of STATs could also
occur in this cell type. Consistent with previous studies using
other cell types (3-6), we found that Stati and Stat3 were
activated rapidly in MCF-iOA cells following stimulation with
EGF, indicating that this pathway is functionally intact inthese cells. On the other hand, heregulins ai and f3i ,which
activate different EGFR family members (32-34), did not
induce STAT activation in MCF-1 OA cells. These results sug-
gest that, in contrast to EGFR, other EGFR family members
like HER2/neu do not signal through STAT pathways, mdi-
cating a divergence in the signaling pathways used by EGFR
family members.
We detected constitutive activation of Stat3 by EMSA
using nuclear extracts prepared from five of nine breast
carcinoma cell lines examined. Interestingly, although EGF
stimulation of MCF-iOA normal breast epithelial cells in-
duces activation of Stati and Stat3, only Stat3 was consti-
tutively activated in the carcinoma cell lines. The observation
that activation of Stat3 in breast carcinoma cells can occur
without a large increase in overall phosphotyrosine content
of total cellular proteins further suggests that Stat3 is selec-
tively phosphorylated by specific kinases in these cells. All
five Stat3-positive breast carcinoma cell lines possess over-
expressed or activated HERi/EGF-R, whereas only SK-BR-3
cells possess activated HER2/neu; SK-BR-3 and MDA-MB-
468 cells also possess elevated c-Src activity.4 Because
activation of Stat3 without Stati is characteristic of cells
harboring activated Src, this raises the possibility that con-
stitutive STAT activation is associated with signaling through
c-Src in at least some of the carcinoma cell lines. Consistent
with this model, treatment of SK-BR-3 and MDA-MB-468
cells with the EGFR selective inhibitor, PDiS878O, did not
diminish the constitutive activation of Stat3 in these cells.
This finding demonstrates that, in these two breast carci-
noma cell lines, EGFR kinase activity is not required for
constitutive Stat3 activation. In contrast, other studies have
suggested that in another breast carcinoma cell line, SUM-
i O2PT, constitutive activation of Stat3 is associated with
signaling through the EGFR receptor (35). Thus, it is possible
that both EGFR-dependent and -independent pathways may
be involved in constitutive activation of Stat3 in different
breast carcinoma cells. Furthermore, the finding that EGF
stimulation can further increase Stat3 activation above the

1274 Constitutive Activation of Stat3 in Oncogenesis
constitutive levels in these breast carcinoma cell lines sup-ports a model in which EGFR and c-Src cooperate in onco-genesis (22) through additive or synergistic activation ofStat3 signaling. Studies of primary tumors indicate that ele-vated Stat3 activation also occurs in human breast tumorscompared to adjacent normal tissue (36).� Taken together,these results establish that constitutive activation of Stat3 isa frequent event in breast carcinoma cells, consistent with a
role for Stat3 signaling in breast cancer progression.Although activation of Stat3 is correlated with transforma-
tion by Src (8, 9) and the oncoproteins reported here, trans-formation by other oncoproteins correlates with activation ofdifferent combinations of STAT family members. For exam-pIe, transformation by v-AbI is associated with activation ofStatS and Stat6 (1 i), BCA-Abl transformed cells possessactivated Stati and StatS (1 2), transformation by Eyk corre-lates with activation of Stati (37), and transformation byhuman T-cell leukemia virus-I is accompanied by activationof Stat3 and StatS (1 0). In addition, Stati , Stat3, and StatSare activated to differing extents in various human leukemiasand lymphomas (i 3-i 5). Furthermore, recent findings mdi-cate that Stati and Stat3 are activated in T cells transformed
by the oncogenic herpesvirus saimiri (38) and myeloid cellstransformed by v-Src (39). In the context of oncogenesis,therefore, the most frequently activated STAT family mem-bers are Stati , Stat3, and Stat5. It is possible that both thespecific cell type and the particular oncoprotein determinewhich STAT family members are activated. This diversity inthe profile of activated STAT family members, in combinationwith simultaneous activation of other signaling pathways,may reflect the diversity of transformed properties elicited bydifferent oncoproteins.
A critical question concerning the role of STATs in cellular
growth control and transformation is: Does activation ofSTATs direct normal cellular proliferation and/or malignanttransformation? Several studies involving genetic manipula-tions of STATs demonstrate a function for these proteins intransduction of growth regulatory signals. For example, nullmutation of the Stat4 gene abrogates interleukin i 2-inducedproliferation of Thi helper T cells (40, 4i). Similarly, nullmutation of the Stat6 gene diminishes interleukin-4-inducedproliferation of Th2 helper T cells (42). Genetic studies havealso directly implicated STAT proteins in tumorigenesis. Infruit flies, reduced expression of the product of the Marellegene, a Drosophila STAT homologue, inhibits hematopoietic
neoplasia associated with activation of the Hopscotch gene,which encodes a JAK kinase (43). Moreover, we have shownthat disrupting Stat3 signaling in NIH 3T3 fibroblasts blockscell transformation by v-Src, providing evidence that Stat3 isone of the signaling pathways that contributes to Src onco-genesis.6 Our results reported here further support the notionthat activated STAT proteins participate in oncogenic trans-formation of mammalian cells, including human breast car-cinoma cells.
5 J. Bromberg and J. E. Darnell, personal communication.
6 J� Turkson, T. Bowman, R. Garcia, E. Caldenhoven, A. P. de Groot, and
A. Jove. Stat3 activation by Src induces specific gene regulation and isrequired for cell transformation, submitted for publication.
Materials and MethodsCell Unes and Culture Conditions. NIH 3T3, 3Vi , and BALB/c 3T3fibroblasts transformed by viral oncogenes and the temperature-sensitive
v-Src mutant PA1O4 have been described previously (8, 23, 44). NIH
3T3/hEGFA cells were obtained from Dr. S. Decker (Parke-Davis Phar-maceutical Research Division, Ann Arbor, Ml). The human breast carci-
noma cell lines, SUM-44PE (45), SUM-52PE (46), SUM-1O2PT, SUM-149PT, and SUM-159PT, were grown in DMEM:Fi2 (i:i) mediasupplemented with 5% FBS, 5 �g/ml insulin, and 1 pg/mI hydrocortisone.
i315-M02 cells were grown as above with iO ng/mI EGF substituted forhydrocortisone in the growth media. 1 2-4 HME normal human breast
epithelial cells were grown in DMEM:Fi2 supplemented with insulin, hy-
drocortisone, EGF, and cholera toxin. MCF-iOA immortalized normalhuman breast epithelial cells (47) were grown in DMEM:Fi2 media sup-
plemented with 5% FBS, 10 �ig/ml insulin, 500 ng/rnl hydrocortisone, 20ng/ml EGF, and 100 ng/rnI cholera toxin. SK-BR-3, MDA-MB-468, andBT483 breast carcinoma cell lines and Hs578Bst cells, established fromnormal breast epithelium, were grown in DMEM supplemented with 10%FBS. Human recombinant EGF was purchased from Ufe Technologies,Inc. (Grand Island, NV). Heregulmns ai and �31 were obtained from Dr. T.Veatman (H. Lee Moffitt Cancer Center, Tampa, FL). For stimulation ofMCF-iOA cells with EGF and heregulins, the cells were preincubatedovernight in media lacking serum, EGF, and insulin, after which ligands
were added directly to the media at the indicated concentrations, followedby incubation at 37#{176}Cfor 10 mm prior to preparation of nuclear extracts.Fortreatment with the EGFR selective inhibitor, PD158780, the compound
was added to a concentration of 500 np�i, and cells were incubated at 37#{176}Cfor 3 or i 5 h prior to stimulation with 50 n� EGF for i 0 mm and preparationof nuclear extracts. PD158780 (29) was a kind gift of Drs. A. Kraker and D.Fry (Parke-Davis Pharmaceutical Aesearch Division).
Nuclear Extract Preparation and EMSA. The procedures were per-
formed as described previously (8). Briefly, nuclear extracts were pre-pared in hypertonic buffer [20 m�i HEPES (pH 7.9), 420 rns�i NaCI, i m�
EDTA, i mM EGTA, 20% glycerol, 20 m� NaF, 1 m�i Na3VO4, 1 mp�i
Na�P2O7, 1 mM DTT, 0.5 mM phenylmethylsulfonyl fluoride, 0.1 p.s�i apro-tinin, 1 �1.M leupeptin, and 1 � antipain]. Preparation of nuclear extracts
to detect STAT activities in transformed cells is critical because loss ofStat3 activation occurs rapidly in cytoplasmic extracts due to dephos-phorylation by phosphatases. Normalized extracts containing 3-8 �.tg oftotal protein were incubated with 32P-radiolabeled oligonucleotide probescontaining the following sequences: (a) hSIE derived from the c-fos gene
promoter (sense strand, 5’-AGCTTCATTTCCCGTAAATCCCTA) thatbinds Stati and Stat3 (24); (b) mammary gland factor element (MGFe)from the bovine (3-casein gene promoter (sense strand, 5’-AGATTTCTAG-GAATTCAA) that has been shown to bind Stat5 (25); and (c) 6spA probe(sense strand, 5’-GATCTGCUCCTrGGAACGT), which selectively binds
Stat6 (26). Protein-DNA complexes were resolved by nondenatunngPAGE and detected by autoradiography as described previously (8).
Competftion and Supershift Controls for EMSA. For competition
controls, nuclear extracts were incubated with radiolabeled probes in the
presence of 100-fold molar excess unlabeled competitor probe prior toPAGE. The FIRE probe (sense strand, 5’-AGCGCCTCCCCGGCCGGGG)was used as nonspecific competitor in our assays. This probe does not
contain conserved sequence elements involved in recognition/binding of
STAT proteins (8). Anti-Stati , -Stat3, and -Stat5 polyclonal antibodieswere purchased from Santa Cruz Biotechnology. For use in supershiftassays, 1-pi volumes of the STAT antibodies were incubated with nuclearextracts for 20 mm at room temperature prior to the addition of radiola-beled probe and PAGE.
Western Blot Analyses. Cell lysates were prepared in boiling SDSsample buffer to preserve the in vivo phosphorylation state of proteins,
and normalized amounts of total cellular proteins were resolved by SDS-PAGE. Following transfer to nitrocellulose, blots were probed with anti-phosphotyrosine antibodies (4GiO; Upstate Biotechnology), anti-Stat3
(Santa Cruz Biotechnology), or anti-phospho-Stat3 antibodies (New
England Biolabs) according to the suppliers’ recommendations. Primaryantibodies bound to blotted proteins were detected by ECL (Arnersharn)
using appropriate horseradish peroxidase-conjugated secondary anti-
bodies as described (8).

Cell Growth & Differentiation 1275
i6. Khazaie, K., Schirrmacher, V., and Uchtner, R. B. EGF receptor inneoplasia and metastasis. Cancer Metastasis Rev., 12: 255-274, i 993.
AcknowledgmentsWe thank members of the laboratory for stimulating discussions, J. Brom-berg and J. E. Darnell for communicating unpublished results, A. Krakerand D. Fry for PDi58780, T. Veatman for heregulmns ai and f3i ,M. Sudoland S. Decker for providing cell lines, A. Pang for technical assistance,
and the Moffitt Cancer Center Molecular Imaging Facility.
References1 . Darnell, J. E., Kerr, I. M., and Stark, G. A. Jak-STAT pathways and
transcriptional activation in response to IFNs and other extracellular sig-
naling proteins. Science (Washington DC), 264: 1415-1421, 1994.
2. Schindler, C., and Darnell, J. E. Transcriptional responses to polypep-tide ligands: the JAK-STAT pathway. Annu. Aev. Biochem., 64: 62i-651,i995.
3. Ruff-Jarnison, S., Chen, K., and Cohen, S. Induction by EGF andinterferon-y of tyrosine phosphorylated DNA binding proteins in mouseliver nuclei. Science (Washington DC), 261: 1733-1736, 1993.
4. Fu, X. V., and Zhang, J. J. Transcription factor p91 interacts with the
epidermal growth factor receptor and mediates activation of the c-fosgene promoter. Cell, 74: 1 135-i i 45, 1993.
5. Zhong, Z., Wen, Z., and Darnell, J. E. Stat3: a STAT family memberactivated by tyrosmne phosphorylation in response to epidermal growth
factor and interleukin-6. Science (Washington DC), 264: 95-98, 1994.
6. Leaman, D. W., Pisharody, S., Flickinger, T. W., Commane, M. A.,Schlessinger, J., Kerr, I. M., Levy, D. E., and Stark, G. A. Aoles of JAKs inactivation of STATs and stimulation of c-fos gene expression by epidermalgrowth factor. Mol. Cell. Biol., 16: 369-375, 1996.
7. vignams, M-L, Sadowski, H. B., Watling, D., Rogers, N. C., and Gilman,
M. Platelet-derived growth factor induces phosphorylation of multiple JAKfamily kinases and STAT proteins. Mol. Cell. Biol., 16: i759-i769, 1996.
8. Vu, C-L, Meyer, D. J., Campbell, G. S., Lamer, A. C., Carter-Su, C.,Schwartz, J., and Jove, A. Enhanced DNA-binding activity of a Stat3-related protein in cells transformed by the Src oncoprotein. Science(Washington DC), 269: 81-83, 1995.
9. Cao, X., Tay, A., Guy, G. A., and Tan, V. H. Activation and associationof Stat3 with Src in v-Src-transformed cell lines. Mol. Cell. Biol., 16:
i595-i603, 1996.
io. Migone, T. S., Un, J. X., Cereseto, A., Mulloy, J. C., O’Shea, J. J.,Franchini, G., and Leonard, W. J. Constitutively activated Jak-STAT path-way in T cells transformed with HTLV-l. Science (Washington DC), 269:
79-81, 1995.
1 i . Danial, N. N., Pernis, A., and Aothman, P. B. Jak-STAT signalinginduced by the v-abl oncogene. Science (Washington DC), 269: 1875-
1877, i995.
i2. Carlesso, N., Frank, D. A., and Griffin, J. D. Tyrosyl phosphorylation
and DNA binding activity of signal transducers and activators of transcrip-
tion (STAT) proteins in hematopoietic cell lines transformed by Bcr/AbI. J.Exp. Med., 183: 8ii-820, 1996.
i3. Weber-Nordt, A. M., Egen, C., Wehinger, J., Ludwig, W., Gouilleux-
Gruart, V., Mertelsrnann, A., and Finke, J. Constitutive activation of STATproteins in primary lymphoid and rnyeloid leukemia cells and in Epstein-Barr virus (EB’I)-related lymphoma cell lines. Blood, 88: 809-8i6, 1996.
i4. Gouilleux-Gruart, V., Gouilleux, F., Desaint, C., Claisse, J. F., Capiod,J. C., Delobel, J., Weber-Nordt, A., Dusanter-Fourt, I., Dreyfus, F., Groner,
B., and Prin, L STAT-related transcdption factors are constitutively acti-vated in peripheral blood cells from acute leukemia patients. Blood, 87:i692-i697, 1996.
15. Zhang, Q., Nowak, I., vonderheid, E. C., Rook, A. H., Kadin, M. E.,Nowell, P. C., Shaw, L M., and Wasik, M. A. Activation of Jak/STAT
proteins involved in signal transduction pathway mediated by receptor forInterleukin 2 in malignant T lymphocytes derived from cutaneous anaplas-
tic large T-ceIl lymphoma and Sezary syndrome. Proc. NatI. Acad. Sd.USA, 93: 9148-9153, 1996.
17. Bolen, J. B., Veillette, A., Schwartz, A. M., DeSeau, V., and Rosen, N.
Activation of pp60c-src protein kinase activity in human colon carcinoma.
Proc. NatI. Acad. Sci. USA, 84: 2251-2255, 1987.
18. Garcia, A., Pankh, N. U., Saya, H., and Gallick, G. E. Effect of herbi-mycin A on growth and pp6oc-src activity in human colon tumor cell lines.
Oncogene, 6: 1983-1989, 1991.
19. Ottenhoff-Kalif, A. E., Rijksen, G., van Beurden, E. A. C. M.,
Hennipman, A., Michels, A. A., and Steal, G. E. J. Characterization ofprotein tyrosmne kinases from human breast cancer: involvement of the
c-src oncogene product. Cancer Res., 52: 4773-4778, 1992.
20. Luttrell, D. K., Lee, A., Lansing, T. J., Crosby, A. M., Jung, K. D.,Willard, D., Luther, M., Aodriguez, M., Berman, J., and Gilmer, T. M.
Involvement of pp90cS�c with two major signaling pathways in human
breast cancer. Proc. NatI. Acad. Sci. USA, 91: 83-87, i994.
21 . Muthuswamy, S. K., and Muller, W. J. Direct and specific interactionof c-Src with Neu is involved in signaling by the epidermal growth factorreceptor. Oncogene, 1 1: 271-279, 1995.
22. Maa, M-C., Leu, T-H., McCarley, D. J., Schatzman, A. C., andParsons, S. J. Potentiation of epidermal growth factor receptor-mediatedoncogenesis by c-Src: implications for the etiology of multiple humancancers. Proc. NatI. Acad. Sci. USA, 92: 6981-6985, 1995.
23. Jove, R., Mayer, B. J., Iba, H., Laugier, D., Poiner, F., Calothy, G.,
Hanafusa, T., and Hanafusa, H. Genetic analysis of p6Ov-src domains
involved in the induction of different cell transformation parameters.J. Virol., 60: 840-848, 1986.
24. Wagner, B. J., Hayes, T. E., Hoban, C. J., and Cochran, B. H. The SIFbinding element confers sis/PDGF inducibility onto the c-fos promoter.EMBO J., 9: 4477-4484, 1990.
25. Pallard, C., Gouilleux, F., Charon, M., Groner, B., Gisselbrecht, S., andDusanter-Fourt, I. lnterleukin-3, erythropoietin, and prolactin activate aSTATS-like factor in lymphoid cells. J. Biol. Chem., 270: 15942-15945,1995.
26. Seidel, H. M., Milocco, L H., Lamb, P., DarneIl, J. E., Jr., Stein, A. B.,
and Rosen, J. Spacing of palindromic half sites as a determinant ofselective STAT (signal transducers and activators of transcription) DNAbinding and transcriptional activity. Proc. NatI. Acad. Sci. USA, 92: 3041-
3045, 1995.
27. Auff-Jarnison, S., Chen, K., and Cohen, S. Epidermal growth factor
induces the tyrosine phosphorylation and nuclear translocation of Stat5 inmouse liver. Proc. NatI. Acad. Sci. USA, 92: 4215-4218, 1995.
28. Ram, T. G., Kokeny, K. E., Dilts, C. A., and Ethier, S. P. Mitogenic
activity of neu differentiation factor/heregulin mimics that of epidermalgrowth factor and insulin-like growth factor-I in human mammary epithe-
hal cells. J. Cell. Physiol., 163: 589-596, i995.
29. Aewcastle, G. W., Palmer, B. D., Thompson, A. M., Bridges, A. J.,Cody, D. A., Zhou, H., Fry, D. W., McMichael, A., and Denny, W. A.Tyrosine kinase inhibitors. 10. Isomenc 4-[(bromophenyl)amino]pyndo[d]-
pyrimidmnes are potent Al? binding site inhibitors of the tyrosine kinase
function of the epidermal growth factor receptor. J. Med. Chem., 39:1823-1835, 1996.
30. Campbell, G. S., Vu, C-L, Jove, A., and Carter-Su, C. Constitutiveactivation of JAK1 in Src-transformed cells. J. Biol. Chem., 272: 2591-2594, 1997.
31 . Courtneidge, S. A. Protein tyrosine kinases, with emphasis on the Srcfamily. Sernin. Cancer Blol., 5: 239-246, i994.
32. Carraway, K. L, Ill, and Cantley, L C. A neu acquaintance for ErbB3
and ErbB4: a role for receptor heterodimerization in growth signaling. Cell,78: 5-8, 1994.
33. Chang, H., Aiese, D. J., II, Gilbert, W., Stern, D. F., and McMahan,U. J. Ligands for ErbB-family receptors encoded by a neuregulin-likegene. Nature (Lond.), 387: 509-512, 1997.
34. Carraway, K. L, Ill, Weber, J. L, Unger, M. J., Ledesma, J., Vu, N.,
Gassrnann, M., and Lai, C. Neuregulmn-2, a new ligand of ErbB3/ErbB4-
receptor tyrosine kinases. Nature (Lond.), 387: 5i2-5i6, 1997.
35. Sartor, C. I., Dziubinski, M. L, Vu, C. L, Jove, A., and Ethier, S. P.Role of epidermal growth factor receptor and STAT-3 activation in auton-
omous proliferation of SUM-i O2PT human breast cancer cells. CancerRes., 57: 978-987, 1997.

1276 Constitutive Activation of Stat3 in Oncogenesis
36. Watson, C. J., and Miller, W. A. Elevated levels of members of theSTAT family of transcription factors in breast carcinoma nuclear extracts.Br. J. Cancer, 71: 840-844, i995.
37. Zong, C., Van, A., August, A., Darnell, J. E., and Hanafusa, H. Uniquesignal transduction of Eyk: constitutive stimulation of the JAK-STAT path-way by an oncogenic receptor-type tyrosine kinase. EMBO J., 15: 4�15-
4525, i996.
38. Lund, T. C., Garcia, A., Medveczky, M. M., Jove, A., and Medveczky,P. G. Activation of STAT transcription factors by herpesvirus saimin Tip-484 requires p56Ick. J. virol., 71: 6677-6682, i997.
39. Chaturvedi, P., Sharma, S., and Reddy, E. P. Abrogation of interleu-kin-3 dependence of myeloid cells by the v-sm oncogene requires SH2
and SH3 domains which specify activation of STATs. Mol. Cell. Biol., 17:3295-3304, i997.
40. Thierfelder, W. E., van Deursen, J. M., Vamamoto, K., Tripp, A. A.,Sarawar, S. A., Carson, R. T., Sangster, M. V., Vignali, D. A. A., Doherty,P. C., Grosveld, G. C., and IhIe, J. N. Requirement for Stat4 in interleukin-12-mediated responses of natural killer and T cells. Nature (Lond.), 382:i7i-174, i996.
41 . Kaplan, M. H., Sun, V-L, Hoey, T., and Grusby, M. J. Impaired IL-i2
responses and enhanced development of Th2 cells in Stat4-deficientmice. Nature (Lond.), 382: 174-177, 1996.
42. Takeda, K., Tanaka, T., Shi, W., Matsumoto, M., Minarni, M.,Kashiwamura, S-i, Nakanishi, K., Voshida, N., Kishirnoto, T., and
Akira, S. Essential role of Stat6 in IL-4 signaling. Nature (Lond.), 380:627-630, 1996.
43. Hou, X. S., Melnick, M. B., and Perrirnon, N. Marelle acts downstreamof the Drosophila HOP/JAK kinase and encodes a protein similar to the
mammalian STATs. Cell, 84: 41 1-419, 1996.
44. Vu, C-L, Prochownik, E. v., Imperiale, M. J., and Jove, A. Attenuationof serum inducibility of immediate early genes by oncoproteins in tyrosinekinase signaling pathways. Mol. Cell. Biol., 13: 2011-2019, 1993.
45. Ethier, S. P., Mahacek, M. L, Gullick, W. J., Frank, T. J., and Weber,
B. L Differential isolation of normal lurninal mammary epithelial cells andbreast cancer cells from primary and rnetastatic sites using selectivemedia. Cancer Aes., 53: 627-635, 1993.
46. Ethier, S. P., Kokeny, K. E., Ridings, J. E., and Dilts, C. A. ErbB familyreceptor expression and growth regulation in a newly isolated human
breast cancer cell line. Cancer Res., 56: 899-907, 1996.
47. Soule, H. D., Maloney, T. M., Wolman, S. A., Peterson, W. D., Brenz,A., McGrath, C. M., Ausso, J., Pauley, A. J., Jones, A. F., and Brooks,S. C. Isolation and characterization of a spontaneously immortalizedhuman breast epithelial cell line, MCF-10. Cancer Res., 50: 6075-6086,1990.