
Comparaison des cinétiques de coagulation enzymatique et mixte ...
13
Comparaison des cin´ etiques de coagulation enzymatique et mixte du lait. Influence du calcium Y. No¨ el, N. Lehembre, A. Dulac, M.C. Clavaud To cite this version: Y. No¨ el, N. Lehembre, A. Dulac, M.C. Clavaud. Comparaison des cin´ etiques de coagulation enzymatique et mixte du lait. Influence du calcium. Le Lait, INRA Editions, 1989, 69 (6), pp.479-490. <hal-00929178> HAL Id: hal-00929178 https://hal.archives-ouvertes.fr/hal-00929178 Submitted on 1 Jan 1989 HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci- entific research documents, whether they are pub- lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L’archive ouverte pluridisciplinaire HAL, est destin´ ee au d´ epˆ ot et ` a la diffusion de documents scientifiques de niveau recherche, publi´ es ou non, ´ emanant des ´ etablissements d’enseignement et de recherche fran¸cais ou ´ etrangers, des laboratoires publics ou priv´ es.
Transcript of Comparaison des cinétiques de coagulation enzymatique et mixte ...

Comparaison des cinetiques de coagulation enzymatique
et mixte du lait. Influence du calcium
Y. Noel, N. Lehembre, A. Dulac, M.C. Clavaud
To cite this version:
Y. Noel, N. Lehembre, A. Dulac, M.C. Clavaud. Comparaison des cinetiques de coagulationenzymatique et mixte du lait. Influence du calcium. Le Lait, INRA Editions, 1989, 69 (6),pp.479-490. <hal-00929178>
HAL Id: hal-00929178
https://hal.archives-ouvertes.fr/hal-00929178
Submitted on 1 Jan 1989
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinee au depot et a la diffusion de documentsscientifiques de niveau recherche, publies ou non,emanant des etablissements d’enseignement et derecherche francais ou etrangers, des laboratoirespublics ou prives.

Lait (1989) 69, 479-490© Elsevier/INRA
479
Article original
Comparaison des cinétiques de coagulation enzymatiqueet mixte du lait. Influence du calcium
Y. Noël 1
avec la collaboration technique de N. Lehembre 2, A. Dulac 2 et M.C. Clavaud 2
1 INRA, station de recherches en technologie et analyses laitières, BP 89, 39800 Poligny;2 INRA, laboratoire de génie des procédés alimentaires, 25 avenue de la République, 91300 Massy,France
(reçu le 10 novembre 1988, accepté le 27 juin 1989)
Résumé - Les coagulations enzymatique et mixte du lait sont étudiées avec une méthode viscoé-lasticimétrique. Alors que les cinétiques de coagulation enzymatique comportent 2 étapes, les cinéti-ques de coagulation mixte en font apparaître 4 : 1) une phase de latence qui correspond à l'hydro-lyse enzymatique et à l'agrégation des micelles, suivie par 2) la formation et le raffermissement ducoagulum, puis 3) une phase de diminution de la fermeté associée à la déminéralisation du réseauprotéique, enfin 4) une nouvelle phase d'augmentation de la fermeté, associée à une réorganisationde la structure du coagulum. L'influence du calcium est examinée par comparaison entre les 2 typesde coagulation, mixte et enzymatique, sur les différents paramètres descriptifs : temps de latence,fermeté, vitesse de coagulation et indice de viscoélasticité.
coagulation enzymatique - coagulation mixte - propriétés viscoélasticimétriques - cal-cium
Summary - Comparison between enzymatic and mixed enzymatlc and lactic acid coagula-tion of milk. Influence of calcium addition. Milk enzymatic coagulation and its combination withlactic acidification were studied using a viscoelasticimetric method. While two phases characterizeenzymatic coagulation, four steps were distinguished for the combined enzymatic and lactic coagula-tion : 1. a lag phase during which proteolysis and micelle aggregation occurred, 2. gel formation andfirming, 3. adecrease in firmness due to the demineralization of the protein structure, 4. a second in-crease in firmness associated with a reorganization of the structure of the coagulum. Calcium influ-ence was examined by comparing the two coagulation types, using descriptive parameters : lagphase, firmness, coagulation rate, viscoelasticity index.
enzymatic coagulation - combined enzymatic and lactic coagulation - viscoélasticimetricproperties - calcium

La coagulation enzymatique du lait a faitl'objet de nombreuses recherches sur les-quelles il existe plusieurs revues bibliogra- Le viscoélasticimètre GVT-INRA, utilisé pour
suivre les cinétiques de coagulation, fonctionnephiques : en particulier, Thomasow & Voss sur le principe du cisaillement harmonique en(1977), Prokopek (1978) sur les méthodes entrefer large. Les essais ont été conduits avecd'étude, Green (1977) sur les agents coa- une amplitude du mouvement du cylindre degulants, Walstra & Van Vliet (1986) sur la mesure de 4 degrés et une fréquence de 10-1
Hz. Le mouvement du cylindre de mesure estphysicochimie de la coagulation. En re- connu, à chaque instant, grâce à un capteur po-vanche, la coagulation mixte résultant de tentiométrique de déplacement angulaire Bo-l'action conjointe d'enzymes coagulantes nohm (Le Prototype Mécanique Industrie, F-et de l'acidification par les bactéries lacti- 78260 L'Etang-La-Ville). La résistance du coa-ques a été moins étudiée (Roefs, 1986; gulum en formation est mesurée par un capteur
de couple à jauges de contrainte (Société Euro-Van Hooydonk et al., 1986b). Trois types péenne d'Extensométrie Appliquée, F-28000de difficultés peuvent expliquer cette situa- Chartres). La saisie des données est assuréetion : d'une part, la maîtrise des dévelop- par un calculateur Hewlett Packard HP9816 etpements bactériens n'est pas acquise, un oscilloscope numérique à mémoire Nicolet
Explorer 2090 iliA (tiroir 206), accompagnésd'autre part la coagulation mixte met en des périphériques suivants : un disque durjeu des phénomènes physicochimiques HP9134, une imprimante HP 2225AB et uneplus complexes que la coagulation enzy- table traçante Tektronix 4662.matique, enfin les instruments de mesure Le comportement rhéologique du coagulumappropriés sont rares. Or, la technologie de lait est observé, avec le viscoélasticimètrefromagère est souvent basée sur des coa- GVT-INRA, dans le domaine non linéaire
(Gervais et al., 1982b), ce qui se traduit pargulations mixtes. Il importe donc de pro- l'existence d'harmoniques dans le signal de ré-gresser dans la connaissance des phéno- ponse. L'analyse de Fourier permet alors de sé-mènes qui leurs sont associés, parer la partie linéaire (signal fondamental) de laconnaissance particulièrement nécessaire partie non linéaire (signaux harmoniques). Onà l'optimisation de capteurs pour le ne s'intéressera par la suite qu'à la partie li-
néaire du signal (Kobayashi et al., 1982).contrôle automatique en fabrication froma-
La contrainte est calculée à partir de la me-gère. sure du moment de couple, en tenant compte
Nous présentons ici les premiers résul- des caractéristiques géométriques du cylindre ettats obtenus en coagulation mixte avec le après correction de l'inertie de l'équipage mobile
constitué par le cylindre et le couplemètreviscoélasticimètre GVT-INRA (Gervais et (Lefèvre, 1988).al., 1982a), et nous les comparons avec On calcule enfin le déphasage entre le signalceux obtenus en coagulation enzymatique. sinusoïdal du mouvement du cylindre et le si-De plus, les effets de l'addition du calcium, gnal fondamental du couple mesuré, ou angledont le mécanisme d'action au cours des de perte.différentes étapes de la coagulation n'est / /pas encore complètement élucidé (Van!/. .Hooydonk et al., 1986c; Bringe & Kinsella, Le lait1986) ont été examinés et sur les 2 typesde coagulation est discutée leur comparai-son.
480
INTRODUCTION
Y. Noël et al.
MATÉRIELS ET MÉTHODES
Mesures rhéologiques
Le lait utilisé pour les essais (5 kg par essai) estreconstitué dans de l'eau bidistillée à 45 oC,sous agitation, à partir d'une poudre de lait écré-
Coagulation enzymatique et mixte 481
mé Nilac (Nizo, Hollande) du type basse tempé-rature, à raison de 100 g de poudre par kg delait reconstitué. Le mélange est agité sanschauffage (1 250 t/min) pendant 1 h.
La teneur moyenne en calcium total du laitreconstitué est de 1,30 g/kg. Les solutions dechlorure de calcium ajoutées au lait ont été pré-parées à partir de CaCI2, 2H20 cristallisé (Prola-bo),
Coagulation enzymatique
Le lait destiné aux essais en coagulation enzy-matique est additionné d'un antibactérien (1 mld'une solution de thiomersal à 100 gl1 pour 1 1de lait). Après reconstitution, le lait est versédans la cuve du viscoélasticimètre GVT-INRA,où il s'équilibre au repos pendant 16-18 h à30 -c (Famelart, 1982; Roefs, 1986).
La solution enzymatique est préparée à par-tir d'une présure en poudre Hansen solubiliséedans un tampon acétate de sodium 0,05 molli àpH 5,5. La quantité ajoutée, soit 0,5 ml, repré-sente 222 J.lgde chymosine active par kg de laitreconstitué. L'activité chymosine de la présureutilisée représentait 99,3% de l'activité coagu-lante totale, déterminée selon la norme FIL 110A (FIL, 1987).
Les apports en chlorure de calcium corres-pondent à 9 niveaux compris entre 0 et 400 mglkg de lait. Lors de la reconstitution, le pH finaldu lait est ajusté à 6,60 par addition d'une quan-tité appropriée d'une solution normale d'acidechlorhydrique ou de soude.
Coagulation mixte
Le lait destiné aux essais en coagulation mixteest reconstitué sans antibactérien, puis versédans la cuve du viscoélasticimètre thermostatéeà 30 oC; il est maintenu pendant 1 h avant l'addi-tion des ferments lactiques.
Les 5 kg de lait sont ensemencés avec 50 mlde ferments préparés à partir d'une souche deStreptococcus lactis ssp diacetylactis (soucheINRA n° 125). Le pH du lait ensemencé est suivien continu au moyen d'un pHmètre Orion SA720 équipé d'une électrode de pH Ross 81.62(ET = 0,02). Lorsque le pH est à 6,00 (tempst'o), la solution enzymatique, préparée commeprécédemment à partir de présure en poudreHansen, est ajoutée au lait à la dose de 88,8 J.lgde chymosine active par kg de lait.
Les apports de chlorure de calcium corres-pondent à 6 niveaux entre 0 et 400 mg/kg delait.
RÉSULTATS
Coagumtionenzymatique
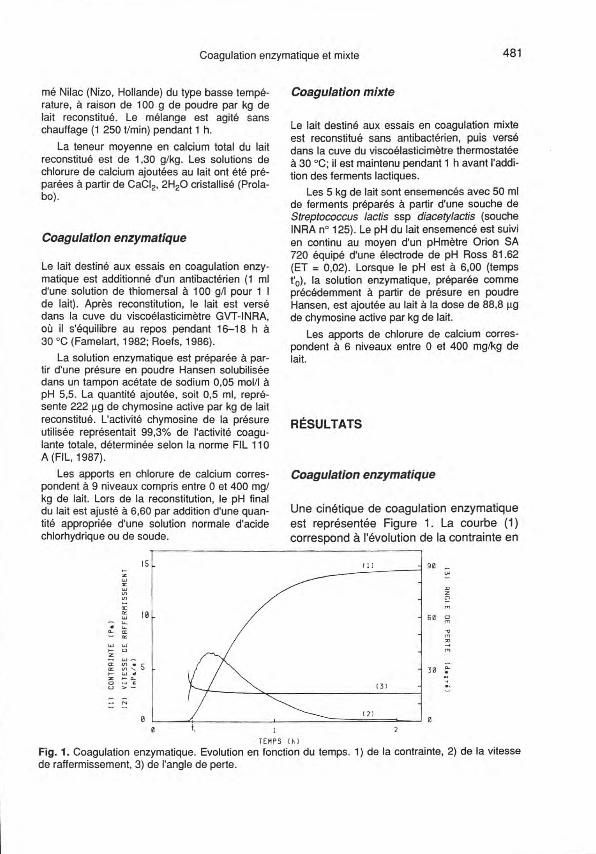
Une cinétique de coagulation enzymatiqueest représentée Figure 1. La courbe (1)correspond à l'évolution de la contrainte en
15~z
~<n
r
'" rawu,
:. ...~
w w~ caz
w -co <n • 5'" <n <,~ w •
~ ::~> -
- N::B
(Il
60 ~
30 p..
( 31
(21
1TEMPS (hl
Fig. 1. Coagulation enzymatique. Evolution en fonction du temps. 1) de la contrainte, 2) de la vitessede raffermissement, 3) de l'angle de perte.
482
fonction du temps. Le temps de latence, t1,
souvent appelé «temps de gel», dont ladétermination dépend du seuil de mobilitéde l'appareil, est associé arbitrairement àune contrainte de 0,5 Pa. La contraintemaximum relevée sur la cinétique corres-pond au plateau. La dérivée de lacontrainte en fonction du temps, qui n'estautre qu'une vitesse, est aussi représen-tée Figure 1 (courbe 2) : le maximum de ladérivée est pris comme vitesse maximalede raffermissement. Enfin, l'évolution del'angle de perte (courbe 3) est indiquée surla même figure : on observe une diminu-tion rapide et importante de l'angle deperte dès le début de la formation du gel,puis celui-ci tend vers une valeur limite, del'ordre de 15 degrés d'angle.
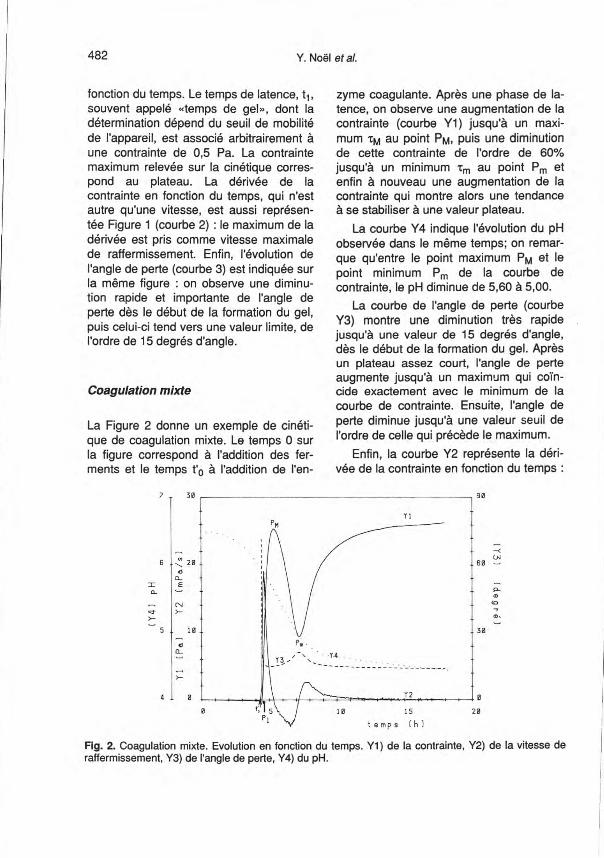
Coagulation mixte
La Figure 2 donne un exemple de cinéti-que de coagulation mixte. Le temps 0 surla figure correspond à l'addition des fer-ments et le temps t'a à l'addition de l'en-
Y. Noël et al.
zyme coagulante. Après une phase de la-tence, on observe une augmentation de lacontrainte (courbe Y1) jusqu'à un maxi-mum 'tM au point PM, puis une diminutionde cette contrainte de l'ordre de 60%jusqu'à un minimum 'tm au point Pm etenfin à nouveau une augmentation de lacontrainte qui montre alors une tendanceà se stabiliser à une valeur plateau.
La courbe Y4 indique l'évolution du pHobservée dans le même temps; on remar-que qu'entre le point maximum PM et lepoint minimum Pm de la courbe decontrainte, le pH diminue de 5,60 à 5,00.
La courbe de l'angle de perte (courbeY3) montre une diminution très rapidejusqu'à une valeur de 15 degrés d'angle,dès le début de la formation du gel. Aprèsun plateau assez court, l'angle de perteaugmente jusqu'à un maximum qui coïn-cide exactement avec le minimum de lacourbe de contrainte. Ensuite, l'angle deperte diminue jusqu'à une valeur seuil del'ordre de celle qui précède le maximum.
Enfin, la courbe Y2 représente la déri-vée de la contrainte en fonction du temps :
3e ,----- ---, ge
Fig. 2. Coagulation mixte. Evolution en fonction du temps. Y1) de la contrainte, Y2) de la vitesse deraffermissement, Y3) de l'angle de perte, Y4) du pH.
6 ~ 20e
CL:r: Ec,
Nq >->-- le~
"CL
~>-
Tl
-{
'"6e
-P-(1)
CD
-<(1),
3e
T2le 15
temps (hl2e
483
15~1'I . _
Coagulation enzymatique et mixte
Q) 1251'1oc;ID
CoC12 (mg/kg)
25
; Ht· --: 15 tCE:~_B_~ _~h'" h"' __"?CC:-::-:~--~
CI Hl l -~_..__+- __ . ~J1'1 81'1 161'1 241'1 320 41'11'1
CoCl2 (mg/kg)
31'1 • •• _._._
1 /0,_
~ 25 ~/
;::1' "I/=~~'~ ~ ~I~ ~u Hl:. <& e 1,
15 l ---1
1'1 80 151'1 ]Hl 320 H10
[0.[17 [mg/kg)
25 r-_~--~"r -~::\,t·~'~~__8>1 161'1 241'1 -3C:-:"---''''
CoCl2 (mg/kgl
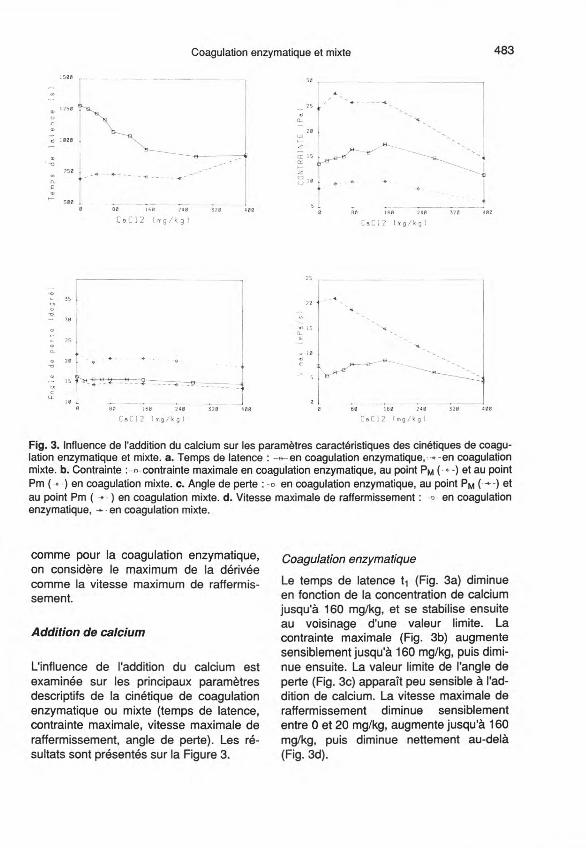
Fig. 3. Influence de l'addition du calcium sur les paramètres caractéristiques des cinétiques de coagu-lation enzymatique et mixte. a. Temps de latence: -a-en coagulation enzymatique,- ...-en coagulationmixte. b. Contrainte :-A-- contrainte maximale en coagulation enzymatique, au point PM (- e- -) et au pointPm (.) en coagulation mixte. c. Angle de perte: -e- en coagulation enzymatique, au point PM ( ...-) etau point Pm (. - ) en coagulation mixte. d. Vitesse maximale de raffermissement: -B-- en coagulationenzymatique, ~ -en coagulation mixte.
comme pour la coagulation enzymatique,on considère le maximum de la dérivéecomme la vitesse maximum de raffermis-sement.
Addition de calcium
L'influence de l'addition du calcium estexaminée sur les principaux paramètresdescriptifs de la cinétique de coagulationenzymatique ou mixte (temps de latence,contrainte maximale, vitesse maximale deraffermissement, angle de perte). Les ré-sultats sont présentés sur la Figure 3.
Coagulation enzymatique
Le temps de latence t1 (Fig. 3a) diminueen fonction de la concentration de calciumjusqu'à 160 mg/kg, et se stabilise ensuiteau voisinage d'une valeur limite. Lacontrainte maximale (Fig. 3b) augmentesensiblement jusqu'à 160 mg/kg, puis dimi-nue ensuite. La valeur limite de l'angle deperte (Fig. 3c) apparaît peu sensible à l'ad-dition de calcium. La vitesse maximale deraffermissement diminue sensiblemententre 0 et 20 mg/kg, augmente jusqu'à 160mg/kg, puis diminue nettement au-delà(Fig. 3d).

484
La reproductibilité des mesures avaitété par ailleurs évaluée sur 3 laits, addi-tionnés de 160 mg de calcium par kg delait : les coefficients de variation observésétaient respectivement de 1% sur le tempsde latence et la contrainte maximale, de2% sur la valeur limite de l'angle de perteet de 3% sur la vitesse maximale de raffer-missement.
Coagulation mixte
Dans le cas de la coagulation mixte, l'in-fluence de l'addition du calcium est exami-née sur les caractéristiques des points re-marquables de la cinétique (Fig. 2, courbeY1), c'est-à-dire:
-le point P1 qui a pour abscisse le tempsde latence t1;
- le point maximum PM;
-le point minimum Pm de la cinétique.Chaque point est caractérisé, d'une
part par ses coordonnées (contrainte ettemps) sur la cinétique de coagulation,d'autre part, par le pH et l'angle de pertecorrespondant. Les valeurs des temps prisen compte pour l'analyse des résultatssont données en considérant comme ori-
Tableau 1. Temps tM et tm associés respective-ment aux points PM et Pm de la cinétique de co-agulation mixte en fonction de la quantité deCaCI2 ajouté.
CaCI2 tM tm(g/kg) (5) (5)
0 3460 9650
40 3900 10850
80 3880 10540
120 4650 11250
240 5200 12250
400 7200 14800
Y. Noël et al.
gine le temps de l'addition de la solutionenzymatique.
Le temps de latence (t1) n'est pas sen-sible à l'addition du calcium, sauf à 400mg/kg où l'on note une augmentation trèsnette (Fig. 3a). Au contraire, les temps as-sociés aux points PM et Pm augmententavec la quantité de calcium ajouté, avecun plateau entre 40 et 80 mg/kg de Ca(Tableau 1).
La Figure 3b présente les valeurs de lacontrainte aux points PM et Pm en fonctionde la quantité de calcium ajouté. Lacontrainte en PM augmente légèrementjusqu'à 40 mg/kg de CaCl2 puis diminuefaiblement jusqu'à 160 mg/kg et très nette-ment ensuite. L'évolution de la contrainteen Pm n'est pas si différente de celle enPM, sauf pour 80 mg/kg de CaCI2·
L'angle de perte en PM et Pm manifesteune légère tendance à la baisse lorsque laquantité de calcium ajouté augmente (Fig.3c), avec un minimum plus prononcé vers40 mg/kg. La vitesse maximum de raffer-missement, calculée sur la partie de lacourbe de contrainte comprise entre P1 etPM, augmente un peu entre 0 et 40 mg/kgde calcium ajouté, mais diminue ensuitenettement (Fig. 3d).
La reproductibilité a été estimée dansle cadre d'un plan d'expérience dont les ré-sultats seront présentés et discutés dansun prochain article. Les coefficients de va-riation des paramètres étudiés ici étaient< 5%, sauf celui de la contrainte au pointPm pour lequel il atteignait 16%.
1
1
1
1
1
1
1
DISCUSSION
Cinétiques de coagulation
La cinétique de coagulation enzymatique,représentée par l'évolution de la contrainte

Coagulation enzymatique et mixte 485
mesurée en fonction du temps, est classi-que. Pendant la phase de latence où lacontrainte mesurée n'est pas significative-ment différente de zéro, se déroulent l'hy-drolyse enzymatique de la caséine kappaet l'agrégation des micelles de caséines.Puis, la formation du coagulum se traduitpar l'apparition d'une contrainte significati-vement différente de zéro, à laquelle estassocié le temps de latence ou temps degel.
L'organisation du réseau et le raffermis-sement du gel se traduisent ensuite parune augmentation de la fermeté, d'abordassez rapide, puis plus lente. La contraintetend alors vers un plateau qui matérialiseun comportement rhéologique stable ducoagulum puisque l'angle de perte aussiest constant. L'évolution de l'angle deperte traduit ici l'apparition et le développe-ment d'une composante élastique impor-tante dès le début de la formation du gel.L'existence d'un plateau atteint très rapide-ment suggère que l'acquisition du compor-tement rhéologique caractéristique du coa-gulum se produit très tôt après la transitionliquide-gel. De plus, si la part de la compo-sante élastique par rapport à la compo-sante visqueuse est constante dès quel'angle de perte atteint ce plateau, l'aug-mentation de la contrainte, c'est-à-dire dela fermeté, observée dans le même temps,indique que les amplitudes de ces 2 com-posantes augmentent mais dans lesmêmes proportions. Des résultats compa-rables, obtenus dans le domaine linéaire,sont présentés par Walstra & Van Vliet(1986), ainsi que par Dejmek (198?), cequi pourrait suggérer que l'évolution obser-vée de l'angle de perte n'est pas affectéepar le caractère linéaire ou non de l'essaiharmonique. Toutefois, Lee (1986) ob-serve, dans le domaine linéaire, une dimi-nution régulière de l'angle de perte depuisle point de gel jusqu'à l'état d'équilibre, at-teint après des temps particulièrementlongs. Ces différents auteurs ne fournis-
sant pas de données quantitatives sur laprécision des mesures, la discussion deces résultats contradictoires est limitée.Néanmoins l'hypothèse selon laquelle lanature des liaisons déterminant le compor-tement rhéologique du coagulum enzyma-tique n'évoluerait pas (Dejmek, 198?; Wal- //stra & Van Vliet, 1986) doit être considéréeavec prudence.
Le recours à un essai en cisaillementharmonique dans des conditions d'ampli-tude, de fréquence et de température nondestructives pour le gel, a permis aussi desuivre des cinétiques de coagulation mixtependant plusieurs heures sans que l'onobserve de fissures ou de synérèse.Quatre étapes à partir de l'addition de l'en-zyme coagulante ont ainsi été mises enévidence. Van Hooydonk et al. (1986b) ontprésenté un résultat qui suggère égaIe-ment l'existence de plusieurs étapes, maiscelles-ci n'apparaissaient pas nèttementdifférenciées.
Les 2 premières parties de la cinétiquede coagulation mixte correspondent aux 2premières étapes de l'action de l'enzymecoagulante : hydrolyse enzymatique de lacaséine kappa et agrégation des micellespendant le temps de latence, puis coagula-tion proprement dite avec formation du ré-seau protéique et augmentation de la fer-meté de celui-ci. La diminution de lacontrainte que l'on observe ensuite, qui tra-duit une diminution de fermeté, peut êtreimputée à la déminéralisation du réseauprotéique liée à l'évolution du pH. En effet,on sait qu'à pH = 5,10, la déminéralisationdes caséines est presque complète (Even- ~huis & De Vries, 1959; Heertje et al., 1985;Van Hooydonk et et., 1986a).
Après le point minimum Pm, la remon-tée de la contrainte et par suite de la fer-meté pourrait être expliquée par la mise enplace de liaisons nouvelles du type decelles d'un coagulum lactique.
L'évolution de l'angle de perte pendantles 2 premières parties est similaire à celle

486
que l'on observe en coagulation enzymati-que. En revanche, au voisinage du pointPm, la variation de l'angle de perte signifieque la composante visqueuse a sensible-ment augmenté au détriment de la compo-sante élastique, révélant ainsi une réorga-nisation de la structure du réseau. Lavaleur de "angle de perte atteinte au ni-veau des zones «plateau» (14-15 degrés)est légèrement inférieure à celle qui estobservée pour le coagulum obtenu parvoie enzymatique (15-16 degrés), maiselle est nettement différente de la valeurde 25 degrés mesurée pour un coagulumlactique (Lehembre, 1986). Walstra & VanVliet (1986) montrent une évolution simi-laire de l'angle de perte en fonction du pH.Si l'on retient l'hypothèse émise par cer-tains auteurs (Walstra & Van Vliet, 1986;Roefs, 1986; Bohlin et al., 1984) selon la-quelle l'angle de perte rendrait compte dutype de liaisons spécifiques d'un coagu-lum, nos résultats suggèreraient que lastructure d'un coagulum mixte est assezproche de celle d'un coagulum enzymati-que. Cependant, il convient aussi d'envisa-ger l'hypothèse selon laquelle ce para-mètre ne discriminerait pas suffisammentles différences de structure.
Addition de calcium
La discussion des résultats relatifs à l'in-fluence comparée du calcium en coagula-tion enzymatique et en coagulation mixtedoit être précédée de 2 remarques. Les 2expérimentations avaient été réaliséesdans des conditions différentes, notam-ment avec des concentrations en présureet un mode de préparation du lait diffé-rents. Toutefois, l'intérêt de faire une ana-lyse comparée des résultats a conduit àrapprocher les 2 séries d'expérimentation.On peut raisonnablement supposer que
. l'incidence de la différence de concentra-
Y. Noël et al.
tion en agent coagulant porte surtout surles valeurs absolues des paramètres detemps, mais que les hypothèses formuléesà partir des observations relatives à l'ac-tion du calcium restent valables. La se-conde remarque porte sur les différencesde préparation des échantillons. La coagu-lation par voie enzymatique autorise l'utili-sation d'un antibactérien et par suite untemps d'équilibrage long à la températurede l'essai (16 hl. En coagulation mixte,cette procédure étant exclue, le tempsd'équilibrage a été limité à 1 h à la tempé-rature de l'essai avant d'ajouter les fer-ments lactiques. Des mesures de pH ef-fectuées par ailleurs en laboratoiremontrant que le pH à 1 h est peu différentde celui mesuré à 24 h : en conséquence,on supposera que les équilibres minéraux,notamment calcique, sont pratiquementréalisés. Enfin, le pH du lait a été corrigéaprès l'addition du Ca dans le cas de lacoagulation enzymatique; cette correctionn'a pas été faite en coagulation mixte maisl'emprésurage a toujours été effectué aumême pH de 6,00.
On sait que l'addition de calcium à pHconstant n'a pas d'influence sur la réactiond'hydrolyse enzymatique, mais qu'elle ac-célère la réaction d'agrégation des mi-celles hydrolysées (Van Hooydonk et al.,1986c), probablement en provoquant spé-cifiquement une diminution du potentiel çen valeur absolue (Dalgleish, 1984), ce quise traduit par une diminution du temps delatence (Van Hooydonk et sl., 1986c).Toutefois, cet effet n'est observable quepour les quantités limitées de CaCI2 ajouté: au-delà de 200 mg/kg, l'addition de cal-cium ne modifie plus le temps de latence(Van Hooydonk, 1987).
En coagulation enzymatique, le tempsde latence diminue avec la quantité de cal-cium ajouté jusqu'à 160 mg/kg : la diminu-tion est lente entre 0 et 40 mg/kg, ten-dance observée par Kowalchyk & Oison

Coagulation enzymatique et mixte 487
(1979), et plus rapide entre 40 et 160mg/kg. Le pH du lait ayant été ajusté, lesvariations du temps de latence jusqu'à 160mg/kg traduiraient bien une accélérationde la phase d'agrégation. Au-delà de 160mg/kg, ce paramètre n'évolue plus. Des ré-sultats comparables ont été obtenus, no-tamment par Van Hooydonk (1987) et Lee(1986).
En coagulation mixte, il y a peu d'effetde l'apport de calcium sur le temps de la-tence. Pour expliquer ce phénomène, il estutile d'examiner la répartition probable ducalcium entre les phases micellaire et so-luble. La quantité de Ca fixée par les mi-celles, au pH du lait (pH 6,60), a été don-née par Brulé (1981) pour des apports encalcium de 0 à 4 g/kg. En particulier, dansla gamme des faibles apports en calcium(250 mg/kg de calcium fixé pour 386,4 mg/kg de calcium ajouté), on détermine sim-plement un coefficient d'interpolation li-néaire (Cafixé= 0,647 x CaajOUlé),qui per-met d'évaluer la répartition entre lesphases micellaire et soluble pour 40, 200et 400 mg/kg (Tableau Il). Connaissant parailleurs l'effet du pH sur la solubilisation ducalcium du lait en l'absence d'apport decalcium (Brulé, 1981; Rajput et al., 1983;
Van Hooydonk, 1986c), on a estimé la ré-partition Ca micellaire/Ca soluble à pH6,00 dans la lait (Tableau Il). On constatealors que la quantité de Ca soluble dansun lait à pH 6,00 est supérieure à celled'un lait additionné de 400 mg/kg de Ca àpH 6,60 : par conséquent, tout ajout deCaCI2 au lait emprésuré à pH 6,00 aug-mente cette fraction soluble. Dans le Ta-bleau Il, on a estimé enfin les quantités decalcium soluble à pH 6,00 et pour diffé-rents niveaux d'apports en calcium, ensupposant que les variations de cette frac-tion dépendaient seulement de la quantitéde CaCI2 ajouté et non du pH, hypothèseque les résultats de Van Hooydonk et al.(1986a) confirmeraient. En analysant lesrésultats obtenus en coagulatiop enzymati-que par rapport au calcium soluble, onconstate que lorsque cette fraction est> 450 mg/kg environ, le temps de latencen'est plus sensible à l'ajout du calcium. Or,les résultats observés en coagulationmixte correspondent précisément à cettesituation avant même tout ajout de CaCI2,
ce qui expliquerait l'absence d'effet duCaCI2 ajouté sur le temps de latence.
En coagulation enzymatique, jusqu'à 40mg/kg, la contrainte maximale augmente
Tableau Il. Répartition du calcium entre les phases micellaire et soluble (calculée d'après les résultatsde Brulé, 1981, indiqués en caractères gras). CaT : calcium total (g/I); Cam: calcium micellaire (g/I);Cas: calcium soluble (g/I).
1
Il pH= 6,60/1
pH= 6,00
~ICar
7 / 7 (Cam Cas Cam Cas
Lait 1,25 0,850 0,400 0,696 0,554+ 0,040 g(Ca)/kg 1,29 0,876 0,414 0,717 0,573+ 0,200 g(Ca)/kg 1,45 0,979 0,471 0,799 0,651+ 0,400 g(Ca)/kg 1,65 1,109 0,541 0,901 0,749

488
modérément. En revanche, la vitessemaximale de raffermissement diminuesensiblement, confirmant les observationsde Kowalchyk & Oison (1979). Un ajout decalcium limité à 40 mg/kg se manifesteraitessentiellement par une diminution de lavitesse d'organisation du gel.
Entre 40 et 160 mg/kg de Ca ajouté, lacontrainte maximale et la vitesse maxi-male de raffermissement augmentent, indi-quant à la fois un effet positif sur la vitessed'organisation du réseau ainsi qu'une aug-mentation de la fermeté. L'ajout de cal-cium se traduirait par une fixation intramo-léculaire, entraînant une diminution de lacharge nette des micelles (Dalgleish,1984) et une diminution de l'hydratation(Baumy & Brulé, 1986), pour finalemententraîner une formation plus rapide d'ungel plus ferme. On peut en effet penserque le calcium favorise le pontage entreles agrégats (Baumy & Brulé, 1986), doncl'organisation du gel. Au-delà de 160 mg/kg, la contrainte maximale et la vitessemaximale de raffermissement baissent for-tement. Tandis que la phase d'agrégationn'est plus affectée par les ajouts de cal-cium, l'organisation du réseau est forte-ment ralentie et conduit à l'obtention d'ungel plus mou. On peut penser que le cal-cium bloque certains sites réactifs par fixa-tion intermoléculaire (Baumy & Brulé,1986), en particulier sur les caséines as et~ (Mac Mahon et al., 1984), sites qui de-viennent indisponibles pour la réticulationet gênent la coagulation en conduisant àdes effets retard (Mac Mahon et el., 1984;Patel & Reuter, 1986). De plus, l'ajoutconcomitant des ions chlorure entraîneune augmentation de la force ionique, di-minuant les liaisons électrostatiquescontribuant à limiter le degré d'organisa-tion du gel.
En coagulation mixte, on remarque,d'une part, qu'une addition de calcium> 300 mg/kg environ conduit à des teneursen calcium micellaire comparables à celles
Y. Noël et al.
qui peuvent être observées sur des laits àpH 6,60, additionnés ou non de calcium(Tableau Il).
D'autre part, la vitesse de raffermisse-ment et la contrainte au point PM présen-tent des valeurs du même ordre de gran-deur que les paramètres équivalents encoagulation enzymatique. Ces résultatspourraient suggérer que l'ajout de calcium,en limitant la déminéralisation des micellesdue à l'acidification, empêcherait à pH6,00 la libération de certains sites actifspour la réticulation du coagulum. Ce phé-nomène expliquerait alors la diminution dela fermeté et de la vitesse de raffermisse-ment pour des quantités importantes decalcium ajouté (Fig. 3d). Des résultatsanalogues ont été observés par Marshallet al. (1982).
L'évolution de l'angle de perte, dans lecas de la coagulation enzymatique, n'estpas sensible à l'ajout de calcium. Ce résul-tat a aussi été observé dans le domaine li-néaire par Bohlin et al. (1984), ainsi quepar Zoon et al. (1988). On peut donc rai-sonnablement penser que ce résultat n'estpas affecté par les conditions de l'essaiharmonique, domaine linéaire ou non. Encoagulation mixte, l'évolution de l'angle deperte en PM et en Pm en fonction de laconcentration en calcium indiquerait que lastructure du coagulum est légèrement plusélastique à 40 ou 80 mg/kg de CaCI2 ajou-té qu'en l'absence d'addition, mais quecette caractéristique est ensuite peu modi-fiée par la quantité de calcium ajouté.
L'étude de l'influence de "addition decalcium suggère que le meilleur compro-mis entre la fermeté du coagulum, sonélasticité et la vitesse de raffermissementest obtenu au voisinage de :- 160 mg/kg de Ca pour un coagulationenzymatique à pH 6,60;- et de 40 mg/kg de Ca en coagulationmixte avec emprésurage à pH 6,00.

Coagulation enzymatique et mixte 489
Le raffermissement est un peu moinsrapide sans adjonction de calcium, mais lecoagulum est moins ferme et moins élasti-que. Jusqu'à 160 mg/kg, la fermeté etl'élasticité sont peu modifiées mais le ra-lentissement de la cinétique est sensible.Au-delà de 160 mg/kg, le ralentissementest de plus en plus marqué, la fermeté ducoagulum diminue alors que son élasticitévarie peu. Jager (1986) a aussi montré leseffets négatifs en fromagerie d'une addi-tion trop importante de calcium. Il est inté-ressant de noter que la pratique fromagèrecourante recommande l'ajout de 40 mg/kgde CaCI2, exceptionnellement 80 mg/kg, li-mite fixée par la législation française.
CONCLUSION
Ces premiers résultats montrent l'intérêt del'analyse rhéologique, et en particulieravec un viscoélasticimètre du type GVT-INRA, pour l'étude des phénomènes liés àla coagulation du lait. La possibilité de ca-ractériser, d'une part, les cinétiques de co-agulation à la fois sur le plan de la fermetéet de la viscoélasticité, et d'évaluer d'autrepart la vitesse de raffermissement, apporteun ensemble d'informations qui, associéesà l'examen des effets des paramètres tech-nologiques, devrait conduire à unemeilleure compréhension des mécanismesmis en jeu. Dans le cas de la coagulationmixte, même si en fromagerie le temps dedécaillage intervient avant la réorganisa-tion du réseau protéique, ce type 'd'étude,associé à la connaissance de l'évolutiondu pH et de la répartition du calcium entreles formes micellaire et soluble (ionique ounon), pourrait notamment contribuer àmieux cerner le potentiel d'évolution dugrain de caillé lors de l'égouttage.
Enfin, nos résultats confirment la néces-sité de raisonner sur la fraction de calcium
soluble et non sur la quantité de calciumajouté pour comprendre les mécanismesde la coagulation.
REMERCIEMENTS
Nous sommes particulièrement reconnaissantsà R. Grappin, directeur de la station de re-cherches en technologie et analyses laitières(INRA-Poligny) ainsi qu'au Pr G. Brulé (ENSA-Rennes) pour leurs conseils et les discussionscritiques lors de la rédaction de cet article.
Nous remercions spécialement S. Aligé et J.Rault (laboratoire de génie de l'hygiène et desprocédés alimentaires, INRA-Massy) pour leurassistance technique.
RÉFÉRENCES
Baumy J.J. & Brulé G. (1986) Etude comparéede la solubilité et de la viscosité des solutionsde caséinate et paracaséinate de sodium enprésence de calcium. Lait66, 65-77Bohlin L., Hegg P.O. & Ljusberg-Wahren H.L.(1984) Viscoelastic properties of coagulatingmilk. J. Dairy Sei. 67, 729-734Bringe NA & Kinsella J.E. (1986) Influence ofcalcium chloride on the chymosin-initiated coa-gulation of casein micelles. J. Dairy Res. 53,371-379Brulé G. (1981) Les minéraux du lait. Rev. Lait.Fr. 400, 61-65Dalgleish D.G. (1984) Measurement of electro-phoretic mobllities and zeta-potentials of par-ticles from milk using laser Doppler electropho-resis. J. Dairy Res. 51, 425-438Dejmek P. (1987) Dynamic rheology of rennetcurd. J. Dairy Sei. 70, 1325-1330Evenhuis N. & De Vries T.M.R. (1959) Thecondition of calcium phosphate in milk. Neth.Milk DairyJ. 13, 1-15Famelart M.H.(1982) Facteurs pnysico-chimiques de la gélification du lait par la pré-

490 Y. Noël et al.
sure. DEA sei. alimentaires, Paris VII Paris XI-ENSIAFIL(1987) Determination of chymosin and bo-vine pepsin contents. Norme FIL 11OAGervais A, Vermeire D., Cerf O. & Toux J.(1982a) Procédé et appareil de mesure des pro-priétés rhéologiques de corps semi-solides parcisaillement harmonique de rotation. Demandede brevet français n° 82.17835, des Etats-Unisn° 7130-US-2712, européen (Allemagne,Grande-Bretagne, Pays-Bas, Italie) n°83.402065.3 (déposé par l'INRA)Gervais A., Vermeire D., Cerf O. & Toux J.(1982b) Caractère viscoélastique non linéairedu caillé présure. xxJ8 Congr. Int. Lait. Moscou)\180-181Green M.L. (1977) Review of the progress ofdairy science : milk coagulants. J. Dairy Res.44,159-188Heertje 1., Visser J. & Smits P. (1985) Structureformation in acid milk gels. Food Microstruct. 4,267-277Jager H. (1986) Das Calcium und sein Einflussin der Kâserei. Dtsch. Milchwirtsch. 37, 496-498Kobayashi M., Ishikawa S. & Samejima M.(1982) Application of non Iinear viscoelasticanalysis by the oscillation method to sorne phar-maceutical ointments in the Japanese pharma-copeia. Chem. Pharm. Bull. 30, 4468-4478Kowalchyk AW. & Oison N.F.(1979) Milk clot-ting and curd firmness as affected by type ofmilk clottinq enzyme, calcium chloride concen-tration and season of year. J. Dairy Sci. 62,1233-1237Lee K.T.(1986) Contribution à l'étude cinétiquede la coagulation du lait par la présure: compa-raison de deux méthodes rhéologiques. Thèsede docteur-ingénieur, Paris VII-Paris XI-ENSIALefèvre V. (1988) Etude rhéologique de la for-mation des gels de caséines obtenus par traite-ment enzymatique. Thèse phys., Paris VII
Lehembre N. (1986) Contribution à l'étude de lacinétique de coagulation mixte d'un lait reconsti-tué suivie par une méthode rhéologique : effetdu calcium et étude multifactorielle de J'in-fluence de la température, du pH et de la dosede présure. ISAA science et technologie alimen-taire. INA Paris-Grignon. DAA. ENSA Toulouse.
McMahon D.J., Brown R.J., Richardson G.H. &Ernstrom CA (1984) Effects of calcium, phos-
phate and bulk culture media on milk coagula-tion properties. J. Dairy Sei. 67, 930-938Marshall R.J., Hatfield D.S. & Green M.L.(1982) Assessment of IWoinstruments for conti-nuous measurement of the curd-firming of ren-neted milk. J. Dairy Res. 49,127-135Patel R.S. & Reuter H. (1986) Effect of sodium,calcium and phosphate on properties of rennetcoagulated milk. Lebensm.-Wiss. Technol. 19,288-291Prokopek D. (1978) Bestimmung der Festigkeitder Gallerte beider Kascherstellung. Dtsch.Milchwirtsch.29,534-543
Rajput Y.S., Bhavasasan M.K. & Ganguli N.C.(1983) Changes in the chemical status of cal-cium in casein micelles with the pH of milk.Milchwissenschaft 38, 211-213
Roefs S.P.F.M. (1986) Structure of acid caseingels. A study of gels formed after acidification inthe cold. Thesis, Agricultural University, Wage-ningen, NetherlandsThomasow J. & Voss E. (1977) Methods for thedetermination of the firmness of milk coagulum.Bull. Fed. Int. Lait. 99, 1-9Van Hooydonk A.C.M. (1987) The renneting ofmilk. A kinetic study of the enzymic and aggre-gation reactions. Thesis Ph. D., Université deWageningen, Pays-BasVan Hooydonk AC.M., Hagedoorn H.G. & Boer-rigter I.J. (1986a) pH-induced physico-che~icalchanges of casein micelles in milk and thelr ef-fect on renneting. 1. Effect of acidification onphysico-chemical properties. Neth. Milk Dairy J.40,281-296Van Hooydonk AC.M., Boerrigter I.J. & Hage-doorn H. G. (1986b) pH-induced physico-chemical changes of casain micelles in milk andtheir effect on renneting. 2. Effect of pH on ren-neting of milk. Neth. Milk Dairy J. 40, 297-313Van Hooydonk AC.M., Boerrigter I.J. & Hage-doorn H.G. (1986c) The effect of various cationson the renneting of milk. Neth. Milk Dairy J. 40,369-390Walstra P. & Van Vliet T. (1986) The physicalchemistry of curd making. Neth. Milk Dairy J.40,241-259Zoon P., Van Vliet & Walstra P. (1988) Rheolo-gical properties of rennet-induced skim milkgels. 3. The effect of calcium and phosphate.Neth. Milk Dairy J. 42, 295-312


















