Cartographie génétique des génomes eucaryotes I ...pdv.univ-lille1.fr/labo/figB4part2.pdf ·...
65
Cartographie génétique des génomes eucaryotes I. Construction de cartes génétique 1. Marqueurs génétiques 2. Types de descendances 3. Méthodes d'établissement des cartes génétiques 4. Caractéristiques des cartes génétiques II. Utilisation des cartes génétiques 1. Localisation de gènes à effets majeurs 2. Localisation de locus contrôlant la variation de caractères quantitatifs 3. Clonage positionnel
Transcript of Cartographie génétique des génomes eucaryotes I ...pdv.univ-lille1.fr/labo/figB4part2.pdf ·...

Cartographie génétique des génomes eucaryotes
I. Construction de cartes génétique1. Marqueurs génétiques2. Types de descendances3. Méthodes d'établissement des cartes génétiques4. Caractéristiques des cartes génétiques
II. Utilisation des cartes génétiques 1. Localisation de gènes à effets majeurs2. Localisation de locus contrôlant la variation de
caractères quantitatifs3. Clonage positionnel

I. Construction de cartés génétique
2. Types de descendances
1. Rappels: méiose et génétique formelle
2. Descendances donnant accès à la phase haploïde
3. Descendances F2 et dérivées de F2
4. Descendances issues de parents hétérozygotes

1. Rappels: méiose et génétique formelle
Phase I carte génétiqueEtablissement des groupes de liaison
Tests liaison/indépendance de tous les couples de marqueurs possibles
Marqueurs assignés à des
groupes de liaison
différents
Marqueurs assignés au
même groupe de liaison
Estimation de la fréquence de recombinaison
Indépendance génétique entre 2 locus
Liaison génétique entre 2 locus
Phase IIEtablissement de l'ordre des marqueurs au sein des
groupes de liaison
Association des allèles portés par
les gamètes à l'issue de la méiose

M1
L2
L1
M2
Cellule avant la méïose2n = 4, 2C
Génotype: L1L2 M1M2
M1
L2
L1
M2
M1
L2
M2
L1Phase S: réplication de l'ADN; 2n = 4, 4C2 chromatides sœurs/ chromosomes
Prophase I méiose:appariement des
chromosomes homologues2n = 4, 2C
Echanges de segments d'ADN entre chromatides non sœurs = crossing-over =
Brassages génétiques intra-chromosomiques(pas de conséquences sur la ségrégation des
allèles dans ce cas)
M1
M2
M1
M2L2
L1
L2
L1
Plaque équatoriale
Métaphase I méiose:ségrégation aléatoire des
chromosomesBrassages génétiques inter-chromosomiques
P = 1/2
ou
M1
M2
M1
M2
L2
L1
L2
L1
P = 1/2
2 marqueurs , L et M, présents sur des chromosomes différents

M1
M2L2
L1
M1
M2L2
L1 M1
M2
L2
L1 M1
M2
L2
L1
Division I: séparation des chromosomes homologues
(réductionnelle)
n, 2C n, 2C n, 2C n, 2C
n, C
n, C
n, C n, C
L1M1 freq. = 1/4L2M2 freq. = 1/4
L1M2 freq. = 1/4L2M1 freq. = 1/4
M2L2
M1L1
M1L2
M2L1
M1L1
M2L2 M1L2
M2L1
Division II: séparation des centromères (équationnelle)
n, C
n, C n, C n, C
M1
M2
M1
M2L2
L1
L2
L1
M1
M2
M1
M2
L2
L1
L2
L1
Métaphase Iméiose (2n, 4C)

2 marqueurs, L et M présents sur des chromosomes différents
Gamètes parentaux (γP) Gamètes recombinants (γR)
L1M1 freq. = 1/4L2M2 freq. = 1/4
L1M2 freq. = 1/4L2M1 freq. = 1/4
r = freq (γR) = 0,5
freq (γR) = freq (γP) = 0,5
Indépendance génétique entre ces 2 locus
Définition: r est la fréquence d'apparition des γR = fréquence de recombinaison (%)
(1-r) = freq (γP) = 0,5d'où

2 marqueurs, L et M, présents sur le même chromosome
Pas de C-Oentre chromatides
non sœurs
1 C-O impliquant 2 chromatides
1
3
Autres possibilités
Génotype des gamètes
Association des allèles
Prophase I méiose
L1M1L1M1L2M2L2M2
PPPP
Aucune
1-42-32-4
L1M1L1M2L2M2L2M1
1-31-42-4
L1M1L1M2L2M1L2M2
PRPR
PRRP
2 C-O impliquant 2 chromatides
23
Génotype: L1L2 M1M22n =2, 2CL1 M1
L2 M2
L2
L1
L1
M1
M1
L2
M2
M2
12
34
Phase S
2n = 2, 4C2 chromatides
sœurs/ chromosomes

2 marqueurs, L et M, présents sur le même chromosome
2 C-O impliquant les 4
chromatides
Génotype: L1L2 M1M22n =2, 2CL1 M1
L2 M2
L2
L1
L1
M1
M1
L2
M2
M2
12
34
Phase S
2n = 2, 4C2 chromatides
sœurs/ chromosomes
Autres possibilités
Génotype des gamètes
Association des allèles
Prophase I méiose
2 C-O impliquant 3 chromatides
123
1-2-42-3-41-3-4
Aucune
L1M1L1M2L2M1L2M2
PRRP
L1M2L1M2L2M1L2M1
RRRR

Cas général: plusieurs locus présents sur le même chromosome
A B DC Liaison physique entre A, B, C et D→ locus synténiques
Indépendance génétique → A-C, A-D, B-D
Liaison génétique → A-B, B-C, C-D

• Indépendance génétique entre ces 2 locus
! 4 types de gamètes attendus (sauf cas de liaison absolue entre L et M)
2 marqueurs, L et M présents sur le même chromosome
Génotype: L1L2 M1M2
L1 M1
L2 M2
L1M1: freq = (1-r)/2L2M2 : freq = (1-r)/2
L1M2 : freq = r/2L2M1 : freq = r/2
γP: freq = (1-r)
γR: freq = r
freq (γR) = freq (γP) = 0,5 = r
et, freq (L1M1) = freq (L2M2) = freq (L1M2) = freq (L2M1) = 1/4
• Cas général

• Liaison génétique entre ces 2 locus
freq (γR) < freq (γP)freq (γR) < 0,5 ⇒ 0 ≤ r < 0,5
freq (γP) > 0,5
! freq (γR) et freq (γP) exprimées en fonction de rfreq (L1M1) = freq (L2M2) = (1-r)
et, freq (L1M2) = freq (L2M1) = r/2
Cas extrême: liaison absolue entre L et MAucun γR produit ⇔ freq (γR) = 0 ⇔ r=0
⇒ freq (γP) = 1

Notions de couplage et de répulsion
• 2 modes d'association possibles des allèles dominants/récessifs chez un individu hétérozygote
• 2 locus (liés) L et M, allèles dominants/récessifs
! Les 2 allèles dominants/récessifs ont été transmis par le même parent
Écriture du génotype: Ll Mm ou L Ml m
γP → L M et l mγR → L m et l M
Gamètes produits par l'individu hétérozygote:
L M
l m
1. Allèles dominants/récessifs en phase de couplage(conformation en cis)

Notions de couplage et de répulsion
• 2 modes d'association possibles des allèles dominants/récessifs chez un individu hétérozygote
• 2 locus (liés) L et M, allèles dominants/récessifs
! Les 2 allèles dominants/récessifs ont été transmis par des parents différents
Écriture du génotype: Ll mM ou L ml M
γP → L m et l MγR → L M et l m
Gamètes produits par l'individu hétérozygote:
L m
l M
2. Allèles dominants/récessifs en phase de répulsion (conformation en trans)

I. Construction de cartés génétique
2. Types de descendances
1. Rappels: méiose et génétique formelle
2. Descendances donnant accès à la phase haploïde
3. Descendances F2 et dérivées de F2
4. Descendances issues de parents hétérozygotes

2. Descendances donnant accès à la phase haploïde2.1. Analyse directe des produits de méiose
Analyse génétique (extraction d'ADN, génotypage…)
Disposer en abondance de tissus ou d'organismes entiers haploïdes
Difficile dans le cas des organismes à cycle de vie diplobiontique (animaux, plantes supérieures)
La phase haploïde du cycle est limitée aux gamètes plus ou moins étroitement associés à des tissus diploïdes
Certaines possibilités: animaux, plantes→ Particularités de certains organismes→ Avancée des techniques de bioliogiemoléculaire, biotechnologies

• Organismes à cycle de vie haplodiplobiontique
→ Alternance de génération entre individus haploïdes (gamétophytes) et diploïdes (sporophytes)
- Levures (ascomycètes )- Nombreuses algues unicellulaires (Chlamydomonas) ou pluricellulaires (certaines algues rouges)-Plantes (mousses, fougères)
• Organismes à cycle de vie haplobiontique
→ Phase diploïde limitée au zygote qui se divise immédiatement par méiose après la fusion des noyaux reproducteurs
→ Phase haploïde du cycle prépondérante (individus haploïdes)
- Champignons filamenteux (Neurospora crassa)- Certaines algues

Exemple: cycle de vie de la levure (Saccharomyces cerevisiae)
Spores n MATα Spores n MATaReproduction
végétativeMITOSES
Plasmogamie CaryogamieConjugaison
Zygote 2nMITOSES
Tétrade de 4 spores n
MEIOSE
½ MATα ½ MATa
Matériel de départ pour des analyses génétiques
Depuis les années 1960:plus de 1200 marqueurs
localisés sur le génome de
la levure
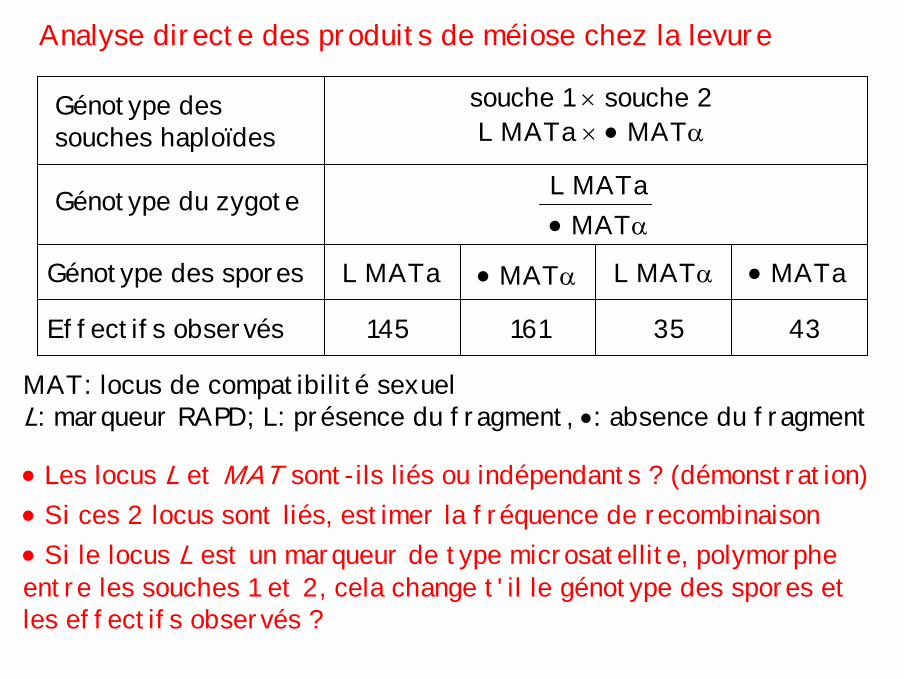
Analyse directe des produits de méiose chez la levure
MAT: locus de compatibilité sexuelL: marqueur RAPD; L: présence du fragment, •: absence du fragment
Génotype dessouches haploïdes L MATa × • MATα
souche 1 × souche 2
Génotype du zygote L MATa• MATα
Génotype des spores L MATa • MATα L MATα • MATa
Effectifs observés 145 161 35 43
• Les locus L et MAT sont-ils liés ou indépendants ? (démonstration)• Si ces 2 locus sont liés, estimer la fréquence de recombinaison• Si le locus L est un marqueur de type microsatellite, polymorphe entre les souches 1 et 2, cela change t'il le génotype des spores et les effectifs observés ?

Homme: essais de génotypage à partir de spermatozoïdes individuels (marqueurs issus de PCR) → A suivre …
• Certaines possibilités pour les organismes à cycle de vie diplobiontique ou assimilés
Plantes: 2 exemples
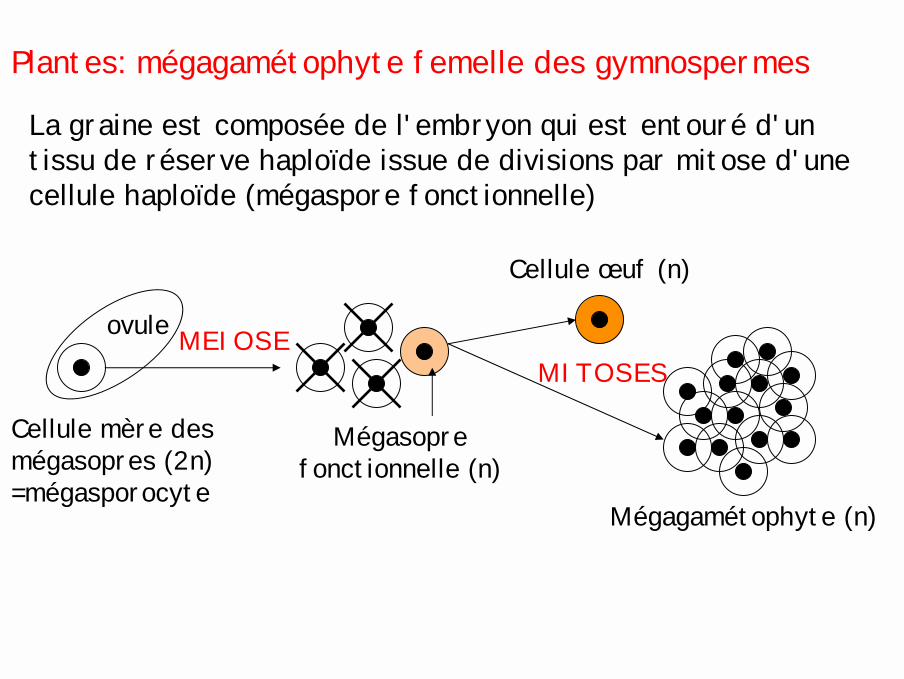
Plantes: mégagamétophyte femelle des gymnospermes
La graine est composée de l'embryon qui est entouré d'un tissu de réserve haploïde issue de divisions par mitose d'une cellule haploïde (mégaspore fonctionnelle)
Cellule mère des mégasopres (2n) =mégasporocyte
ovule
Mégasoprefonctionnelle (n)
MEIOSE
Cellule œuf (n)
Mégagamétophyte (n)
MITOSES
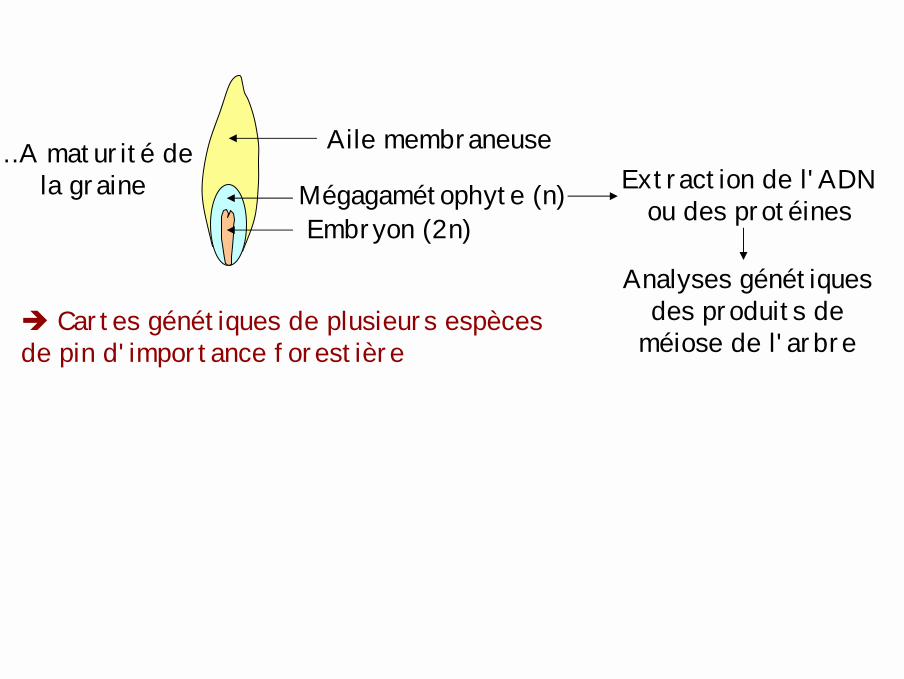
… A maturité de la graine
Aile membraneuse
Embryon (2n)Mégagamétophyte (n) Extraction de l'ADN
ou des protéines
Analyses génétiques des produits de
méiose de l'arbre! Cartes génétiques de plusieurs espèces de pin d'importance forestière

Plantes: lignées haploïdes doublées (Angiospermes)Biotechnologies végétales → culture in vitro
Culture d'ovairesimmatures = Gynogenèse
(betterave, tabac …)
Culture d'anthèresimmatures = Androgenèse
(maïs, orge …)
Mégaspores (n) Microspores (n)
Sacs embryonnaires
Grains de pollen
Embryogenèse haploïde
Milieu d'induction in vitro
Régénération d'individus haploïdes fonctionnels
Doublement chromosomique spontané ou provoqué (colchicine)
Individus "homozygotes parfaits" = lignées haploïdes doublées (HD)
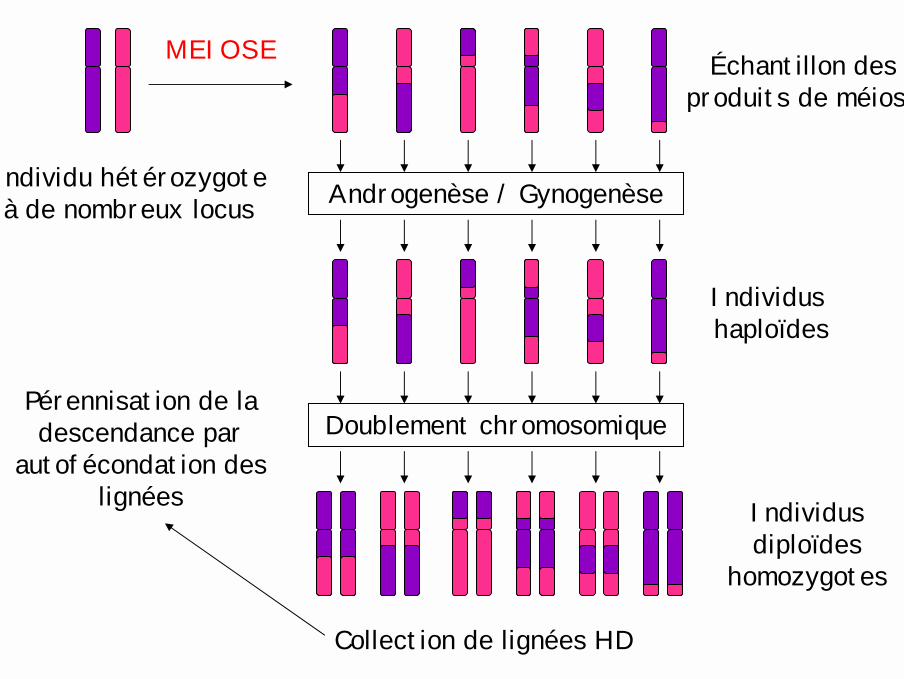
Individu hétérozygoteà de nombreux locus
Échantillon des produits de méiose
MEIOSE
Androgenèse / Gynogenèse
Individushaploïdes
Doublement chromosomique
Individus diploïdes
homozygotes
Collection de lignées HD
Pérennisation de la descendance par
autofécondation des lignées

2.2. Descendances donnant accès à la phase haploïde
→ Back-cross (croisement en retour)
P1 × P2 P1 et P2 = lignées
× P2 P2 = lignée récurrenteF1Hétérozygote à de nombreux locus
BC1 Étude indirecte des produits de méiose de l'individu F1
→ test-cross (croisement test): croisement d'individus F1 par une lignée homozygote récessive (≠ P1 et P2) pour 1 ou quelques locus donnés

Types d'organismes pour lesquels on peut étudier des descendances BC1 ou F2
→ Disposer de lignées et réaliser des croisements contrôlés
• Notion de lignée: croisements frères-sœurs pendant plusieurs générations → diminution du taux moyen d'hétérozygotie des individus à chaque génération
• Espèces modèles de laboratoire: drosophile, souris, rat, nématode
• Petits animaux d'élevage: poule, lapin, chien …
Animaux
→ Espèces prolifiques à cycle de vie court

• Plantes supérieures: majorité d'espèces hermaphrodites
• La plupart des espèces utilisées par l'homme sont capables de s'autoféconder (lignées)
→ Espèces préférentiellement autogames naturellement (blé, tomate)
→ Espèces préférentiellement allogames sélectionnée afin d'augmenter l'auto-compatibilité (tournesol, betterave)
Plantes
• De nombreuses espèces d'intérêt économique ont un cycle de vie annuel ou bisannuel • Production de graines / individu souvent élevée
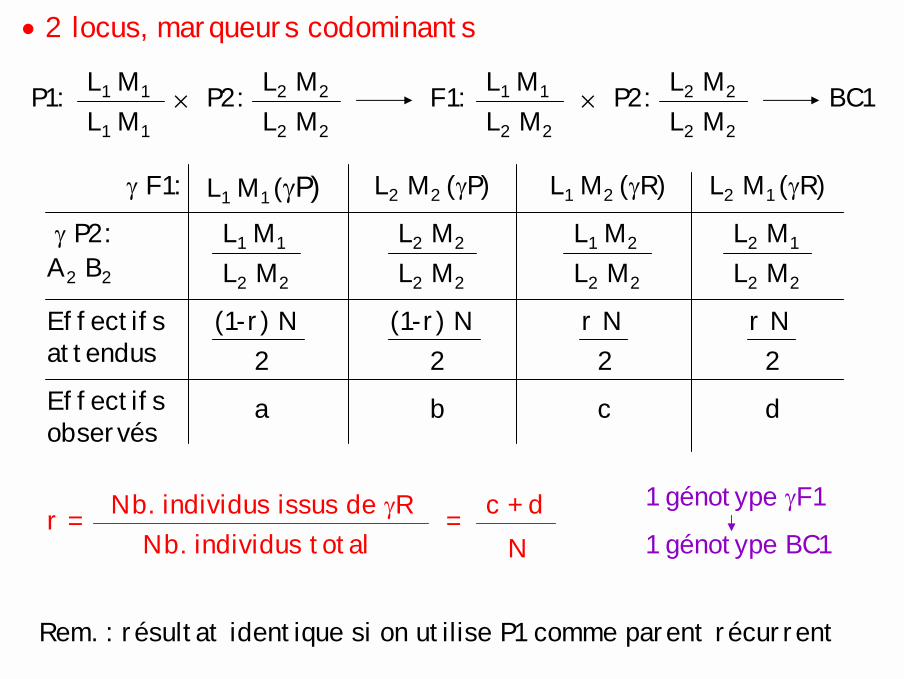
• 2 locus, marqueurs codominants
L1 M1
L2 M2
L2 M2
L2 M2
L1 M2
L2 M2
L2 M1
L2 M2
L1 M1
L1 M1
L2 M2
L2 M2× P2:P1: L1 M1
L2 M2F1: L2 M2
L2 M2× P2: BC1
(1-r) N2
(1-r) N2
r N2
r N2
a b c d
A2 B2
Effectifs attendus
L1 M1 (γP) L2 M2 (γP) L1 M2 (γR) L2 M1 (γR)γ F1:
γ P2:
Effectifs observés
r = Nb. individus issus de γRNb. individus total
Rem. : résultat identique si on utilise P1 comme parent récurrent
= c + dN
1 génotype γF1
1 génotype BC1

• 2 locus, marqueurs dominants, phase de couplage (1)
(1-r) N2
(1-r) N2
r N2
r N2
a b c d
Effectifs attendus
l m
L M (γP) l m (γP) L m (γR) l M (γR)γ F1:
γ P2:
Effectifs observés
r = Nb. individus issus de γRNb. individus total
= c + dN
1 génotype γF1
1 génotype BC1
× P2:P1: L ML M
l ml m
L Ml m
F1: × P2: l ml m
Homozygote récessif ☺
L ml m
L Ml m
l ml m
l Ml m

• 2 locus, marqueurs dominants, phase de couplage (2)
× P2:P1: L ML M
l ml m
L Ml m
F1:
Phénotypes
L M
L M (γP) l m (γP) L m (γR) l M (γR)γ F1:
γ P2: L mL M
L ML M
l mL M
l ML M
P1: Homozygote dominant #×
L ML M
100% [L M]
⇒ Impossible d'estimer r

• 2 locus, marqueurs dominants, phase de répulsion
Phénotypes
L m
L m (γP) l M (γP) L M (γR) l m (γR)γ F1:
γ P2:
× P2:P1: L mL m
l Ml M
L ml M
F1:
L ML m
L mL m
l ML m
l mL m
× P1: L mL m
⇒ Impossible d'estimer r
[L m] [L M] [L M] [L m]
Rem. : résultat identique si on utilise P2 comme parent récurrent

• 2 locus, 1 marqueur codominant, 1 marqueur dominant
× P2:P1: L1 ML1 M
L2 mL2 m
L1 ML2 m
F1: × P1: L1 ML1 M
# → cf. 2 marqueurs dominants, phase de répulsion
× P2:P1: L1 ML2 m
F1:L1 ML1 M
L2 mL2 m
× P2: L2 mL2 m
☺ → cf. 2 marqueurs dominants, phase de couplage (1)→ cf. 2 marqueurs codominants

Conclusions sur l'information obtenue à partir des différents types de descendances donnant accès à la phase haploïde
→ Accès direct (organismes ou tissus haploïdes, HD)
• Il suffit que les individus dont on étudie les produits de méiose soient hétérozygotes pour un maximum de locus
→ Accès indirect (back-cross)
• Pas d'effet du type de marqueurs, ni du mode d'association des allèles dans le cas de marqueurs dominants
• Individus F1 hétérozygotes pour de nombreux locus
• Marqueurs codominants
• Marqueurs dominants, phase de couplage, parent récurrent homozygote récessif pour un maximum de locus

I. Construction de cartés génétique
2. Types de descendances
1. Rappels: méiose et génétique formelle
2. Descendances donnant accès à la phase haploïde
3. Descendances F2 et dérivées de F2
4. Descendances issues de parents hétérozygotes

3. Descendances F2 et dérivées de F2
3.1. Descendances F2
P1 × P2 P1 et P2 = lignées
F1 × F1 → croisements frères-sœurs (animaux, plantes) → autofécondation (plantes)
× F1F1
Étude indirecte des produits de méiose et F2
MéioseMéiose

• 2 marqueurs codominants
Etude de la ségrégation de 2 locus
• 2 marqueurs dominants, phase de couplage• 2 marqueurs dominants, phase de répulsion• 1 marqueur codominant, 1 marqueur dominant
→ Il est toujours possible d'estimer r (même si le calcul n'est pas toujours évident !)
cf. TD
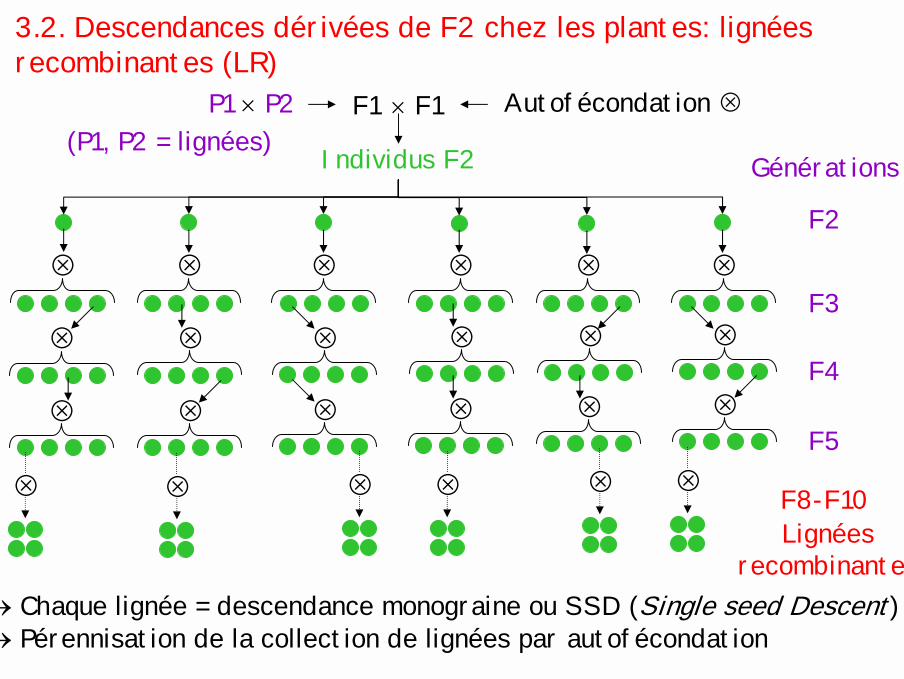
3.2. Descendances dérivées de F2 chez les plantes: lignées recombinantes (LR)
P1 × P2(P1, P2 = lignées) Individus F2 Générations
F2
! ! ! ! ! !F3
!! ! ! ! !F4
! ! ! ! ! !F5
! ! ! ! ! ! F8-F10Lignées
recombinantes
× F1F1 Autofécondation !
→ Chaque lignée = descendance monograine ou SSD (Single seed Descent)→ Pérennisation de la collection de lignées par autofécondation

Probabilité qu'un individu soit hétérozygote à un locus donné après n générations d'autofécondation: p = 1/2n
AA × aa F1: 100% Aa
F2: 25% AA 50% Aa 25% aa!
F3: 37,5% AA 25% Aa 37,5% aa!
F4: 43,75% AA 12,5% Aa 43,75% aa!
F5: 46,88% AA 6,25% Aa 46,88% aa!
F6: 48,44% AA 3,12% Aa 48,44% aa!
F7: 49,22% AA 1,56% Aa 49,22% aa!
F8: 49,61% AA 0,78% Aa 49,61% aa!p = 1/27

Individu F1 hétérozygoteà de nombreux
locus
Structure chromosomique des lignées recombinantes
MEIOSE
γ γ
n !n méioses n méioses
F2
F8-F10
→ Probabilité de détecter une liaison entre 2 locus plus élevée que dans le cas des autres types de descendances

Estimation de la fréquence de recombinaison entre 2 locus (LR)→ Estimation indépendante du type de marqueur
× P2:P1: L mL m
l Ml M
L ml M
F1: × F1: L ml M
F2 ! F8, F10 …
P P R R
L mL m
l Ml M
L ML M
l ml m
[L m] [l M] [L M] [l m]
(1-R) N2
(1-R) N2
R N2
R N2
type:
Génotype des lignéesEffectifs attendus
Eff. obs. a b c d
R = Nb. lignées recombinantesNb. lignées total
= c + dN

R: proportion de lignées recombinantes qui n'est pas une estimation de r→ fonction de r→ R > r car le nombre de recombinaisons occasionné par les autofécondations successives est supérieur à celui observé à l'issue de la méiose des individus F1
On peut montrer que:2r
(1 + 2r)R = , d'où r =
R2 (1 –R)
0,1 0,2 0,3 0,4 0,50
0,1
0,2
0,3
0,4
0,5
2r(1 + 2r)
R =
r = fréquence de recombinaison
R =
fréq
uenc
e de
lign
ées
reco
mbi
nant
es
Rem. Il est possible d'obtenir des LR chez les animaux (souris, drosophile, nématode) → croisements frères-soeurs

I. Construction de cartés génétique
2. Types de descendances
1. Rappels: méiose et génétique formelle
2. Descendances donnant accès à la phase haploïde
3. Descendances F2 et dérivées de F2
4. Descendances issues de parents hétérozygotes

4. Descendances issues de parents hétérozygotes
→ Organismes pour lesquels on ne dispose pas de lignées
F1'
Étude indirecte des produits de méiose et de P1 et P2
MéioseMéiose
P1 × P2 P1 et P2 ≠ lignées
Hétérozygotes ou homozygotes pour chacun
des locus considérés

• Gros animaux (d'élevage)
→ Temps de génération qui peut être important→ Espace d'élevage élevé→ Peu de descendants par génération
• Plantes→ Espèces auto-incompatibles ou qui subissent une très forte dépression de consanguinité (pomme de terre, chicorée)
→ Espèces pérennes à cycle de vie long: arbres (peuplier, eucalyptus)
→ solution à ce problème pour certains gymnospermes: analyse des mégagamétophytes femelles

• Homme: espèce très hétérozygote
→ Temps de génération élevé→ Peu de descendants par génération
→ Pas de croisements contrôlés (?) mais homogamie
! Homme et gros animaux d'élevage: cumul de l'information obtenue à partir de plusieurs fratries

Schémas de croisements possibles dans le cas de descendances issues de parents hétérozygotes
→ Pour un locus donné, il peut y avoir jusqu'à 4 allèles en ségrégation en F1' (marqueur codominant)
× P2:P1: L1
L2
L3
L4F1'
→ Si on considère un couple de locus et différents cas de figures:
- 2, 3 ou 4 allèles / locus- Marqueurs codominants ou dominants (couplage et répulsion
! 27 configurations parentales possibles

Cartographie génétique des génomes eucaryotes
I. Construction de cartes génétique1. Marqueurs génétiques2. Types de descendances3. Méthodes d'établissement des cartes génétiques4. Caractéristiques des cartes génétiques
II. Utilisation des cartes génétiques 1. Localisation de gènes à effets majeurs2. Localisation de locus contrôlant la variation de
caractères quantitatifs3. Clonage positionnel

3. Méthodes d'établissement des cartes génétiques
• Déterminer le génotypes des individus d'une ou plusieurs descendances pour un maximum de locus marqueurs
• Tests des ségrégations monolocus (χ2) → élimination des locus qui présentent de fortes distorsions de ségrégation
• Disposer de descendances en ségrégation pour de nombreux marqueurs

• Tests 2 points: tests de liaison génétiques en considérant tous les couples de locus possibles
Ex: 100 marqueurs → 4900 combinaisons
! Assigner chacun des marqueurs à un groupe de liaison
Groupe 1Groupe 2
Méthode statistique: χ2 d'indépendanceLimite: inadapté dans le cas d'effectifs réduits → Homme
Méthode probabiliste: LODscoreAdapté dans le cas d'effectifs réduits ; additif

LODscore (logarithm of the odd ratio) → logarithme décimal du rapport de vraisemblance de 2 hypothèses (Morton, 1955):
Hypothèse 1: il y a une liaison génétique entre 2 locus (r = θ < 0.5); la vraisemblance de cette hypothèse est notée eL(r = θ)
Hypothèse 2: il n'y a pas de liaison génétique entre ces 2 locus (r = 0.5); la vraisemblance de cette hypothèse est notée eL(r = 0.5).
LODscore = log10eL(r = θ)
eL(r = 0.5)
LODscore > 0 et à une valeur seuil → Liaison génétique
ex: LODscore = 3 → l'hypothèse de liaison est 1000 fois plus vraisemblable que l'hypothèse d'indépendance entre 2 locus
ex: LODscore = -1 → l'hypothèse de liaison est 10 fois moins vraisemblable que l'hypothèse d'indépendance entre 2 locus

• Tests 3 points et multipoints: détermination de l'ordre relatif des marqueurs au sein des groupes de liaison
Tests 3 points: ordre relatif de 3 locus appartenant au même groupe de liaison
Ordres possibles: A – B - C ?, A – C - B ?, B – A - C ?
Vraisemblance de chacun des ordres possibles: eL(ABC), eL(ACB) et eL(BAC)
Calcul et comparaison des valeurs de LODscore

LODscore (ABC/ACB) = log10eL(ABC)
eL(ACB)
LODscore (ABC/BAC) = log10eL(ABC)
eL(BAC)
LODscore (ACB/BAC) = log10eL(ABC)
eL(BAC)
Ex: LODscore (ABC/ACB) = 0.5
LODscore (ABC/BAC) = - 3
LODscore (ACB/BAC) = -2
Ordre le plus vraisemblable ?
BAC

Tests multipoint: ordres relatifs de 4, 5, 6, … locus
Complexité croissante des calculs de vraisemblance lorsqu'on augmente le nombre de locus, même avec des ordinateurs très puissants
Les logiciels de cartographie utilisent le principe suivant afin de réduire le nombre de calculs à réaliser:
Test 3 points? ? ?
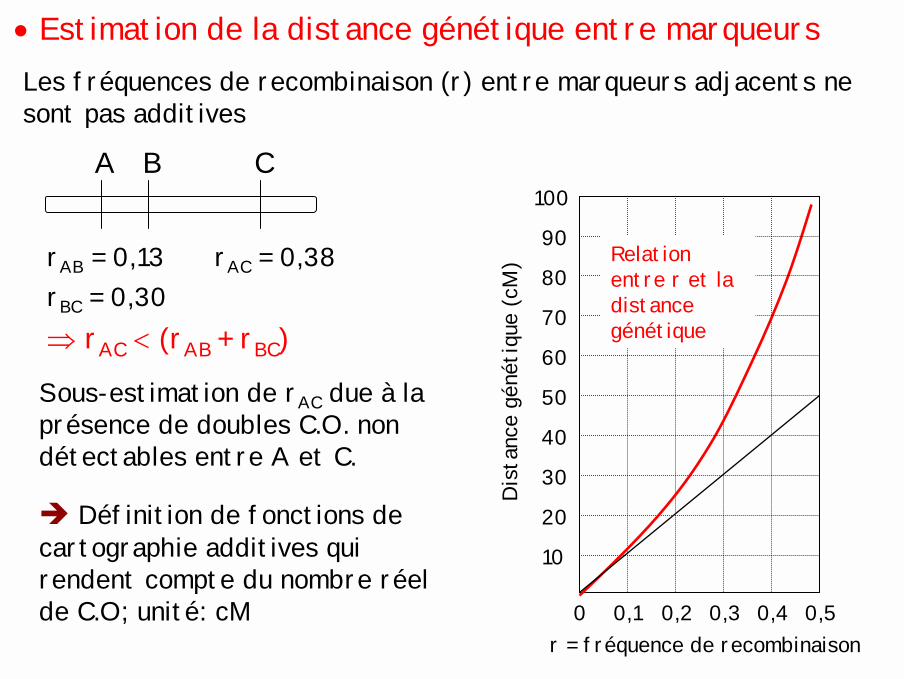
• Estimation de la distance génétique entre marqueursLes fréquences de recombinaison (r) entre marqueurs adjacents nesont pas additives
A B C
rAB = 0,13rBC = 0,30
rAC = 0,38
⇒ rAC < (rAB + rBC)
Sous-estimation de rAC due à la présence de doubles C.O. non détectables entre A et C.
0,1 0,2 0,3 0,4 0,50
10
20
30
40
50
r = fréquence de recombinaison
Dis
tanc
e gé
néti
que
(cM
)
60
70
80
90
100
Relation entre r et la distance génétique
! Définition de fonctions de cartographie additives qui rendent compte du nombre réel de C.O; unité: cM

Fonctions de cartographie
• Distance de Haldane
→ Prend en compte les C.O. multiples
dH = -50 ln (1-2r) unité: cM
• Distance de Kosambi
→ Prend en compte les C.O. multiples et l'existence d'interférence
Interférence: restriction à ce que 2 C.O. se produisent proches l'un de l'autre. Ce phénomène est d'autant plus marqué que la distance entre 2 marqueurs est faible.Origine probable: contraintes mécaniques lors de la méïose
dK = 25 ln [ (1+2r) / (1-2r) ] unité: cM

0,1 0,2 0,3 0,4 0,50
10
20
30
40
50
r = fréquence de recombinaison
Dis
tanc
e gé
néti
que
(cM
)
60
70
80
90
100Fonction de HaldaneFonction de Kosambi
r ≤ 0,1 ⇒ r×100 = dH = dK
r > 0,2 ⇒ dK > dH

Cartographie génétique des génomes eucaryotes
I. Construction de cartes génétique1. Marqueurs génétiques2. Types de descendances3. Méthodes d'établissement des cartes génétiques4. Caractéristiques des cartes génétiques
II. Utilisation des cartes génétiques 1. Localisation de gènes à effets majeurs2. Localisation de locus contrôlant la variation de
caractères quantitatifs3. Clonage positionnel

4. Quelques caractéristiques des cartes génétiques
Carte génétique saturée:
$ Liaison génétique de chaque point du génome à au moins un marqueurs
$ Nombre de groupes de liaison identique au nombre de chromosomes du génome haploïde; chaque marqueur de la carte est assigné à un groupe de liaison
$ Pas d'augmentation de la distance génétique couverte par la carte lors de l'ajout de nouveaux marqueurs
4.1. Saturation des cartes génétiques

Méthodes de saturation des cartes génétiques
$ Cartographie d'un maximum de marqueurs (× 100 chez la drosophile ou A. thaliana; × 1000 chez l'homme)
$ Localisation des régions télomériques à l'aide de marqueurs RFLP ou PCR spécifiques de ces régions (maïs)

4.2. Variations de longueurs des cartes génétiques
→ Distances génétique (i.e. fréquence de recombinaison) influencées par certains facteurs
Carte physique: distances absolues entre A, B et C (bp)
A B C
1
2
Cartes génétiques: ordre A, B, C conservé; distances variables entre A, B et C (cM)

Plantes: fréquence de recombinaison plus élevée lors des méioses femelles (tomate, orge) ou lors des méioses mâles (maïs, Arabidopsis thaliana)
Homme: fréquence de crossing-over plus importante chez les femmes que chez les hommes
Sexe
Eloignement génétique entre les parents de la decendanceRéduction de la distance des cartes génétiques construites àpartir de descendances issues de croisements inter-spécifiques
Facteurs génétiques et environnementaux
Drosophile: pas de C.O. chez les mâles

Carte génétique du chromosome 12 humain
Femme: 168 cMHomme: 78 cM
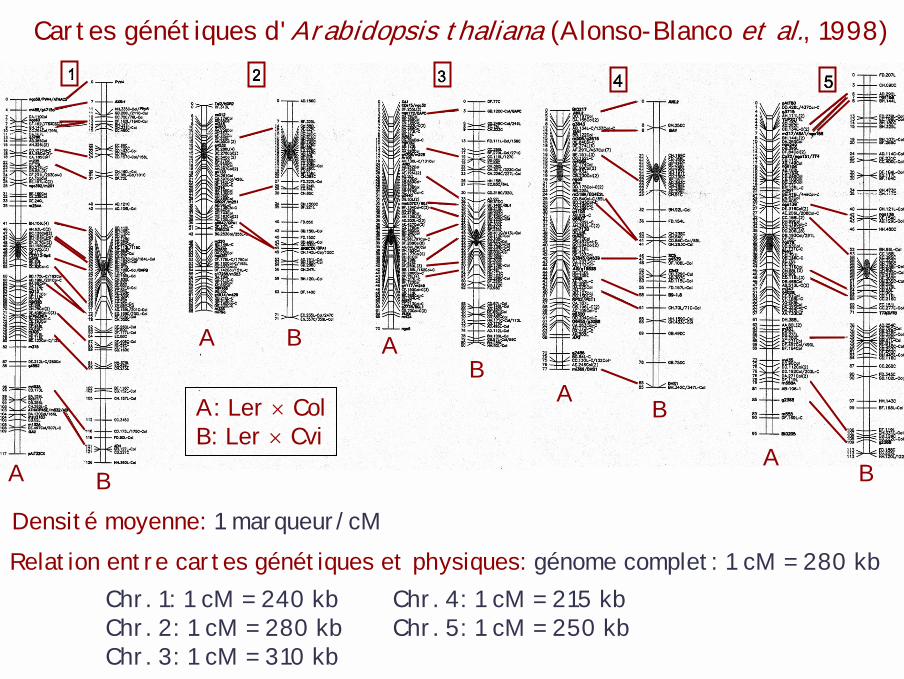
Cartes génétiques d'Arabidopsis thaliana (Alonso-Blanco et al., 1998)
A
A A
A
AB
BB
B
B
A: Ler × ColB: Ler × Cvi
Densité moyenne: 1 marqueur/cMRelation entre cartes génétiques et physiques: génome complet: 1 cM = 280 kb
Chr. 1: 1 cM = 240 kb Chr. 4: 1 cM = 215 kbChr. 2: 1 cM = 280 kb Chr. 5: 1 cM = 250 kbChr. 3: 1 cM = 310 kb

Le paradoxe de la valeur C: la quantité d'ADN par génome haploïde n'est pas toujours corrélée au degré d'évolution des espèces
4.3. Relation entre distance physique et distance génétique

4570170350016 000Blé690(H)-1180(F)120(H)-210(F)2800(H)–4780(F)3300Homme *
44013527001200Soja134019018602500Maïs 176010017003000Souris *
78075830650haricot7501051270950Tomate 11805510201200Colza
0.2-8000.16Phage T4 *2.6-17504.6E. Coli *2.8 260420012Levure *310320100Nématode *
200-260100-130480-630125A. Thaliana *60070280165Drosophile *2701301575430Riz *
(kbp)/cMChromosomeGénome(Mbp)
1800
Distance de carte(cM)
13 00015024 000Pin maritime
Longueur ADNLongueur du
génomeEspèce