
CAPES 2006-2007 cours: Christine Laclef Organogenèse des...
48
1 Organogenèse des membres I. Formation du bourgeon de membre de vertébré A-Organisation des membres chez les vertébrés B- Morphogenèse du membre de vertébré C- Spécification du territoire du membre D- Induction du bourgeon de membre II. Mise en place des axes de polarité du membre A- Régionalisation proximo-distale A.1. Le bourrelet apical ectodermique (AER: « Apical ectodermal ridge ») A.2. La zone de progression (« progress zone ») A.3. Expression régionalisée des gènes Hox B- Régionalisation antéro-postérieure B.1. La zone d’activité polarisante (ZPA) B.2. Shh est la molécule de signalisation produite par la ZPA C- Régionalisation dorso-ventrale III. Spécification de l’identité des membres, antérieur versus postérieur A- Spécification par les gènes Hox B- Spécification par les facteurs de transcription Tbx IV. Différenciation tissulaire dans le membre A- Le squelette B- Les muscles C- L’innervation D- la vascularisation E- L’individualisation des doigts nécessite une apoptose morphogène CAPES 2006-2007 cours: Christine Laclef
Transcript of CAPES 2006-2007 cours: Christine Laclef Organogenèse des...

1
Organogenèse des membres
I. Formation du bourgeon de membre de vertébréA-Organisation des membres chez les vertébrésB- Morphogenèse du membre de vertébréC- Spécification du territoire du membreD- Induction du bourgeon de membre
II. Mise en place des axes de polarité du membreA- Régionalisation proximo-distale
A.1. Le bourrelet apical ectodermique (AER: « Apical ectodermal ridge »)A.2. La zone de progression (« progress zone »)A.3. Expression régionalisée des gènes Hox
B- Régionalisation antéro-postérieureB.1. La zone d’activité polarisante (ZPA) B.2. Shh est la molécule de signalisation produite par la ZPA
C- Régionalisation dorso-ventrale
III. Spécification de l’identité des membres, antérieur versus postérieurA- Spécification par les gènes HoxB- Spécification par les facteurs de transcription Tbx
IV. Différenciation tissulaire dans le membreA- Le squeletteB- Les musclesC- L’innervationD- la vascularisationE- L’individualisation des doigts nécessite une apoptose morphogène
CAPES 2006-2007 cours: Christine Laclef

2
A- Organisation des membres de vertébrés
I – Formation du bourgeon de membre de vertébré
Différents axes de polarité bien visible au niveau de la main

3
Différents axes de polarité visiblesdans l’aile de poulet
A- Organisation des membres de vertébrés

4
Diversité de forme des éléments squelettiques,mais conservation des subdivisions:
- Stylopode: élément proximal- Zeugopode: élément médian- Autopode: élément distal
A- Organisation des membres de vertébrés

5
Morphogenèse du membre
embryons humains
29 jours 32 jours
M: mésenchymeE: ectoderme
AER

6
B- Morphogenèse du membre
aile de poulet patte antérieure de souris

7
C- Spécification du territoire du membre le long de l’axe antéro-postérieur

8
C- Spécification du territoire du membre le long de l’axe antéro-postérieur
Rôle des gènes Hox
vertèbre
somites
7 vertèbrescervicales
13 vertèbresthoraciques
6 vertèbreslombaires
4 vertèbressacrées
souris
Formation du membre antérieur à la limite ant.d’expression du gène Hoxc6

9
C- Spécification du territoire du membre le long de l’axe antéro-postérieur
Rôle des gènes Hox
souris
poulet
oie
python

10
1- Fgf10 sécrété par le mésenchyme (mésoderme latéral) induit l’expression de Fgf8 dans l’ectodermecompétent sus-jascent, à la limite entre les régions dorsales et ventrales (future AER).
2- Fgf8 sécrété par l’ectoderme induit la prolifération des cellules mésenchymateuses sous-jascentes(future zone de progression).
3- La sécrétion de Fgf8 par l’ectoderme induit l’expression de Shh à la limite postérieure du bourgeon demembre.
4- La sécrétion de Shh induit l’expression de Fgf4 dans l’ectoderme postérieur du bourgeon de membre.
D- Induction du bourgeon de membre

11
établissement des axes ? proximo-distalantéro-postérieurdorso-ventral
Spécification de l’identité antérieure vs postérieure ?
Processus morphogénétiques mis en jeu lors de la morphogenèse du membre ?

12
A- Régionalisation proximo-distale du membre
II – Mise en place des axes de polarité du membre
Le bourrelet apicale ectodermique ou AER (apical ectodermal ridge)
M: mésenchymeE: ectoderme
AER

13
Exp: ablation de l’AER à différents moments
du développement du membre
Les structures proximo-distales sont spécifiées
progressivement au cours du temps:
- spécification précoce des structures proximales
- spécification tardive des structures distales
A- Régionalisation proximo-distale du membre
L’AER est nécessaire à la croissancedu bourgeon de membre

14
Fgf8 est la molécule de signalisation exprimée dans l’AER
Hybridation in situ révélantl’expression de Fgf8
chez un embryon de pouletà 3 jours de développement
A- Régionalisation proximo-distale du membre L’AER est nécessaire à la croissance du bourgeon de membre
Voie de signalisation Fgf:

15
A- Régionalisation proximo-distale du membre
L’AER est nécessaire à la croissance du bourgeon de membre La zone de progression joue également un rôle important dans la croissance du membre
La zone de progression (progress zone): mésenchyme d’origine mésodermique adjacent à l’AER

16
Exp. de transplantation d’AERet de « progress zone » :
ectoderme d’âge quelconque+ mésoderme distal d’embryon âgé-> membre incomplet:(seuls les éléments squelettiques distaux se forment)
ectoderme d’âge quelconque+ mésoderme d’embryon jeune-> membre complet

17
Résumé des expériences réalisées sur l’AERet la zone de progression:

18
Expression des gènes Hox desgroupes de paralogue a et d dans lebourgeon d’aile de poulet
Expression différentielle le longde l’axe proximo-distal
Expression régionalisée desgènes Hox dans le membre

19
Le même profil d’expression proximo-distale se retrouve dans lesbourgeons de membre antérieur et postérieur (poulet)
Expression régionalisée des gènes Hox dans le membre

20
Corrélation entre les combinaisons de gène Hox exprimés dans le bourgeon de membreet la différentiation des différentes région du membre
Expression régionalisée des gènes Hox dans le membre

21
Délétion d’éléments squelettiques des membres suite à la délétion de gènes Hox:
Chez la souris: le double mutant Hoxa-11,Hoxd-11 n’a pas d’ulna et de radius
Chez l’homme: syndrome de synpolydactylierésultant de mutations homozygote de HOXD-13
Rôle des gènes Hox dans la morphogenèse du membre

22
Exp. de greffe d’une ZPAadditionnelle dans la régionantérieure du bourgeon de membre:
CCL: La ZPA a une activitépostériorisante
-> Duplication en miroir de la régiondistale du membre
La ZPA est localisée dans la région postérieuredes bourgeons de membre
B- Régionalisation antéro-postérieure du membre
II – Mise en place des axes de polarité du membre
La zone d’activité polarisante

23
Exp. de greffe d’une ZPAadditionnelle dans la régionantérieure du bourgeon demembre:
B- Régionalisation antéro-postérieure du membre
La zone d’activité polarisante
CCL: La ZPA a une activité postériorisante
-> Duplication en miroir de la région distale du membre

24
Shh est la molécule de signalisation produite par la ZPA
B- Régionalisation antéro-postérieure du membre
Hybridation in situ révélant l’expression de Shh dans larégion postérieure des bourgeons de membre
Profil d’expression de Shh:

25
Expérience réalisée pour tester le rôle polarisant de Shh:
1- Shh est transfecté dans des fibroblastes de poulet en culture
2- les fibroblastes transfectés (exprimant Shh) sont implantés dans larégion antérieure d’un bourgeon de membre
-> le résultat observé est identique à la transplantation de ZPA
CCL: - Shh a une activité polarisante identique à celle de la ZPA- Shh est la molécule de signalisation responsable de l’activité polarisante de la ZPA
Shh est la molécule de signalisation produite par la ZPA
B- Régionalisation antéro-postérieure du membre
Mise en évidence du rôle de Shh:

26
Activité inductrice de Shh est proportionnelle à sa concentration: Shh constitue un gradient morphogène

27
-> duplication en miroir de la région distale du membre-> action postériorisante similaire à celle de Shh
Exp. de greffe de billes imprégnées d’acide rétinoïque dans la région antérieure du bourgeon de membre
L’acide rétinoïque mime l’activité polarisante de la ZPA
B- Régionalisation antéro-postérieure du membre
Signification physiologique ???

28
Des gènes différents sont exprimés dans les régions dorsale et ventrale du bourgeon de membre
C- Régionalisation dorso-ventrale du membre
II – Mise en place des axes de polarité du membre

29
Position d’une cellule définie selon les 3 axes - proximo-distal (Fgf8, Fgf4)- antéro-postérieur (Shh)- dorso-ventral (Wnt7a/En1)
Intégration des différentes informations de position

30
1- établissement de la ZPA par l’AER
2- Induction de Fgf4 dans l’AER par Shh
3- Maintenance mutuelle de Fgf4 et Shh
4- Maintenance de Shh par Wnt7a
« Cross-talk » entre les différentes voies de signalisation

31

32
III – Spécification de l’identité du membre, antérieur vs postérieur
Exp. de transplantation de mésoderme:
mésenchyme prélevé au niveau dumembre antérieur
greffé au niveau du membre postérieur
-> formation d’un membre chimère
CCL: il existe une identité de membre antérieur
conservé dans l’explant

33
A- Rôle des gènes Hox dans la spécification de l’identité des membres
Le développement des membres est différentpour les membres antérieur et postérieur.
Des gènes sélecteurs différents sontexprimés dans le mésenchyme au niveauxoù sont induits les membres antérieurs etpostérieurs.
L’expression de ces gènes sélecteursdifférents est sous contrôle des gènes Hox:combinaisons de gènes hox différentesantérieurement et postérieurement

34
III – Spécification de l’identité du membre, antérieur vs postérieur
B- Rôle des facteurs de transcription à homéodomaine Tbx
Bourgeon d’aile
Bourgeon de patte
Expression de Tbx4dans le bourgeon de membre postérieur
Expression de Tbx5dans le bourgeon de membre antérieur

35
Si greffe à proximité du membre antérieur:-> induction de Tbx5 -> membre antérieur additionnel
Si greffe à proximité du membre postérieur:-> induction de Tbx4 -> membre postérieur additionnel
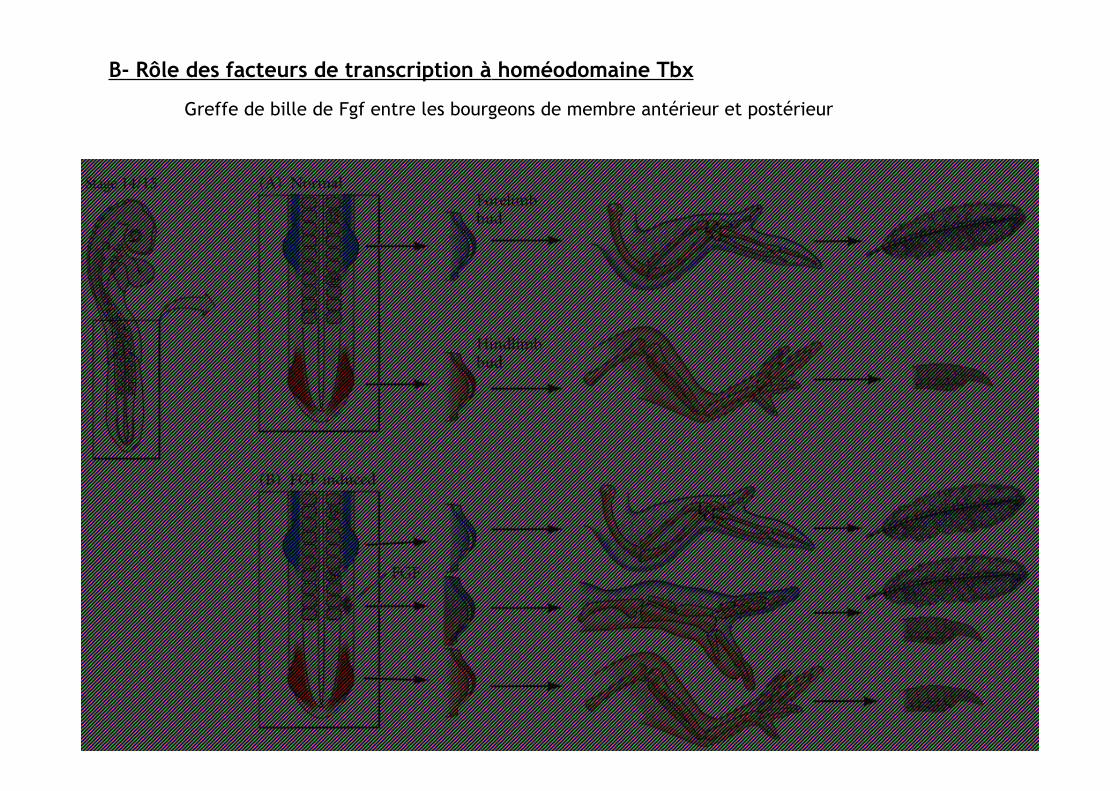
Greffe de bille de Fgf entre lesbourgeons de membre antérieuret postérieur
B- Rôle des facteurs de transcription à homéodomaine Tbx

36
Greffe de bille de Fgf entre les bourgeonsde membre antérieur et postérieur
Si greffe à équidistance des membres ant et post:
-> induction de Tbx5 ant
-> induction de Tbx4 post
-> membre additionnel chimérique
B- Rôle des facteurs de transcription à homéodomaine Tbx
37
Greffe de bille de Fgf entre les bourgeons de membre antérieur et postérieur
B- Rôle des facteurs de transcription à homéodomaine Tbx

38
IV – Différenciation tissulaire dans le membre
A- Le squelette, les tendons et le tissu conjonctif proviennent du mésoderme latéral
Ossification endochondrale progressive des éléments squelettiques mis en place
Poulet à 19 j
Souris 17 j

39
Exp. de greffe caille-poulet
B- Les muscles des membres sont formés à partir de précurseurs somitiques

40
DIFFERENCIATIONMyogénine, Desmine, MHC
DETERMINATION
MyoD, Myf5
Précurseursmyogéniques MyotubesMyoblastes
Fibresmusculaires
matures
MATURATION
Migration
Pax3, cMet, Lbx1
Morphogenèse
Différentes étapes de la myogenèse à l’échelle cellulaire
FUSION

41
Différentes étapes de la myogenèse à l’échelle del’embryon
1- Myogenèse somitique
E8.5 ………… E12.5

42
Différentes étapes de la myogenèse à l’échelle del’embryon
2- Myogenèse primaire
étape de morphogenèse musculaire
E12.5 ………………………..……… E15.5
3- Myogenèse secondaire
étape de croissance musculaire
par augmentation du nombre de fibres
…………………………….…… E18.5

43
Différents compartiments myogéniques
épaxial
hypaxial
44
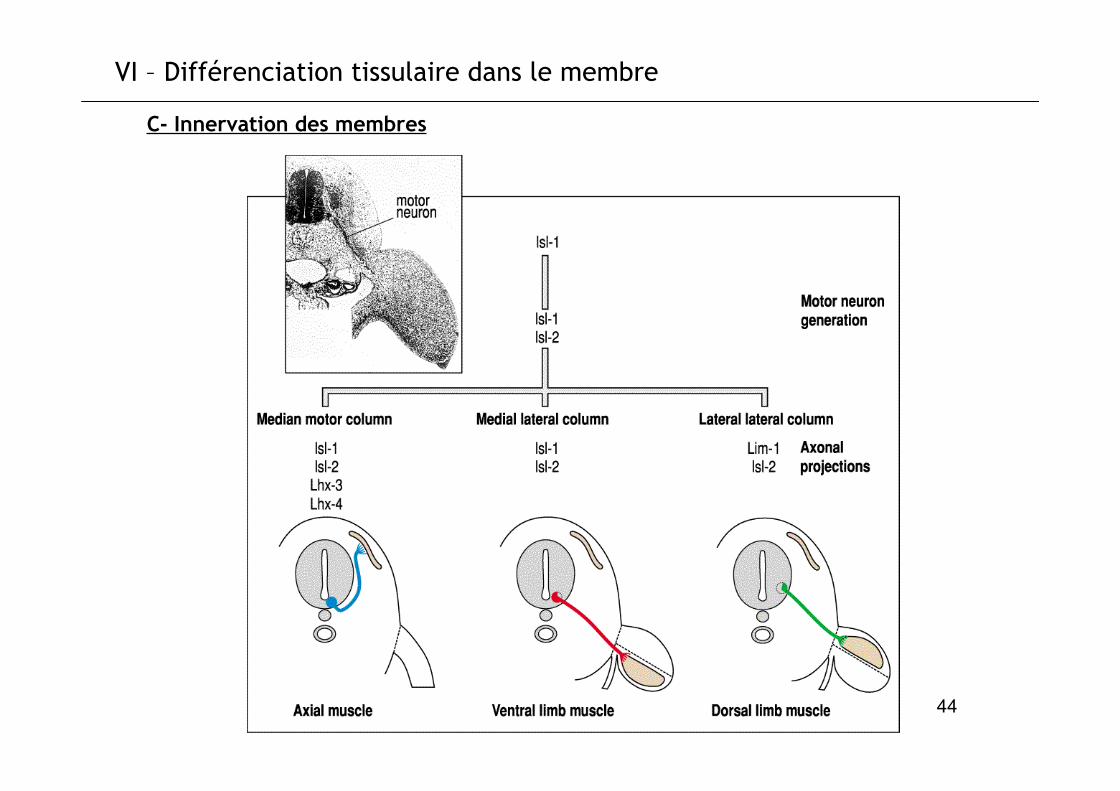
VI – Différenciation tissulaire dans le membre
C- Innervation des membres

45
D- La vascularisation

46
E- L’individualisation des doigts nécessite une apoptose morphogène
importance relative de l’apoptose morphogène variable en fonction des espèces
Exp. transplantation d’ectoderme etde mésoderme:

47
CCL: Grande conservation au cours de l’évolution des mécanismesmoléculaires impliqués dans la morphogenèse des membres

48
membre antérieur membre postérieur
Anatomie des élémentssquelettiques proximaux desmembres de vertébrés












