
BULLETIN - Université de Neuchâtel précédentes, l’auteur relève le lien entre la diversité...
200
BULLETIN DE LA SOCIÉTÉ NEUCHÂTELOISE DES SCIENCES NATURELLES FONDÉE EN 1832 Tome 121 1998 PUBLIÉ AvEC LE SOUTIEN DE L’ACADÉmIE SUISSE DES SCIENCES NATURELLES
-
Upload
hoangduong -
Category
Documents
-
view
224 -
download
0
Transcript of BULLETIN - Université de Neuchâtel précédentes, l’auteur relève le lien entre la diversité...

BULLETINDE LA
SOCIÉTÉ NEUCHÂTELOISEDES SCIENCES NATURELLES
FONDÉE EN 1832
Tome 121
1998
PUBLIÉ AvEC LE SOUTIEN DE L’ACADÉmIE SUISSE DES SCIENCES NATURELLES


avant-propos
Après les deux tomes du Bulletin et le mémoire parus l’année dernière, 1998 retrouve en quelque sorte sa vitesse de croisière. Le numéro 121 présente toutefois une particula-rité: l’abondance inusitée de planches en couleur, abondance qui n’est malheureusement pas signe de l’opulence de la S.N.S.N. En effet, ces illustrations sont à la charge des auteurs ... ou de leurs sponsors ! Il a toutefois paru élégant aux rédacteurs d’habiller cette livraison d’une couverture un brin plus soyeuse afin de souligner le fait.
Les différents articles couvrent une large partie des sciences naturelles, paléontologie, phanérogamie, algologie, écologie végétale et pédologie, entomologie, conservation de la nature. Répondant au vœu exprimé par des membres de la Société, les rédacteurs ont rédigé une brève introduction au contenu scientifique de ce numéro, sans pour autant la transformer en un glossaire des mots difficiles !
On remarquera d’emblée que la paléontologie se taille la part de ... l’allosaure ! L’imagerie a largement mis à la mode les dinosaures qu’avec plus ou moins de réussite
livres et films tentent de faire revivre. mais ce que l’on sait moins, c’est comment les paléontologues acquièrent les connaissances nécessaires à la reconstitution non seule-ment des formes, mais aussi du comportement et de l’écologie de ces animaux. J. Ayer a travaillé sur un site de fouilles aux USA. Il raconte comment les ossements fossiles sont dégagés du sol, préparés, et comment un environnement vieux de quelque 150 millions d’années peut être reconstitué. Cet article d’information générale frappera le lecteur par la qualité de son illustration. Il annonce une exposition “Dinosaures” qui sera présentée par le muséum d’histoire naturelle de Neuchâtel en 1999.
Les listriodons sont des mammifères fossiles à allure de sangliers qui ont vécu au miocène inférieur et moyen (il y a donc 15 à 20 millions d’années) en Europe et dans le sous-continent indien. Les ossements d’une espèce-type, Listiodon splendens, avaient été trouvés en 1839 par Célestin Nicolet à la Chaux-de-Fonds. Ils viennent d’être réexaminés par J. van Der MaDe spécialiste de ce groupe de mammifères. Son article est précédé par celui de m. S. Jacquat qui présente l’aspect historique et évoque la forte personnalité de C. Nicolet.
Troisième volet paléontologique, l’article de P. Morel examine des ossements d’élans trouvés il y a plus d’un siècle dans un gouffre situé près des verrières. La datation par le radiocarbone permet de situer dans le temps la présence de ces animaux dans les forêts jurassiennes, alors que leur aire de répartition européenne ne couvre plus aujourd’hui que la Scandinavie et l’est de notre continent.
La Phanérogamie est représentée par deux publications. Les Pandanacées constituent une importante famille de plantes monocotylédones dont on connaît actuellement plus de trois cents espèces dans l’Ancien monde intertropical; K. L. HuynH poursuit ses études sur le genre Pandanus en décrivant deux nouvelles espèces de madagascar.
Puis S. m. GHaffari et S. B. DJavaDi, deux botanistes iraniens, présentent l’étude chromosomique de neuf espèces de Cousinia, pour la plupart endémiques en Iran. Ce sont des Astéracées, famille que les plus âgés des lecteurs connaissent encore souvent sous le nom de Composées.
3

Après une absence qui suivit la disparition de marguerite Wüthrich, on retrouve dans le Bulletin, grâce à F. Straub, des articles d’algologie. Dans ce numéro, O. reyMonD, F. straub et H. R. preisiG signalent pour la première fois Chrysonebula holmesi en Suisse et discutent de sa position systématique. Il s’agit d’une algue qui forme des amas mucila-gineux sur les fonds caillouteux des cours d’eau propres. Les auteurs étudient, sous l’angle de la bioindication, les diatomées qui se développent dans le mucilage de Chryso-nebula.
L’écologie végétale est représentée par trois travaux. Dans le premier, G. vaDi et J.m. Gobat traitent du problème de la podzolisation dans le Jura à la fois sous l’angle de la pédologie et de la phytosociologie. Les poDzosols sont des sols de climats froids et humides. Ils sont caractérisés par des processus de chéluviation, c’est-à-dire d’entraîne-ment de matière organique et de métaux, de fer par exemple, de la surface vers la profon-deur. Les auteurs discutent de la présence de ces sols dans le Jura, en dépit de conditions pédologiques défavorables et établissent un lien entre la présence des poDzosols et celle de la pessière à blechnum (une fougère caractéristique).
Dans un milieu différent, P. cornali présente le second volet de son étude des pinèdes de la rive sud du lac de Neuchâtel. La biomasse végétale, la quantité de litière et la vitesse de sa décomposition, la productivité de cette forêt sont quantifiées. La dynamique du groupement végétal, examinée à la lumière de ces données, indique une évolution vers une forêt de feuillus.
Enfin, W. MattHey a procédé en 1998 à l’inventaire des gouilles dans la tourbière du Cachot (vallée de la Brévine). Ces points d’eau si caractéristiques des zones ouvertes à sphaignes avaient été recensés une première fois en 1968. La comparaison montre une diminution considérable de leur surface en une trentaine d’année.
L’entomologie est également présente. S. barbalat continue son étude des coléop-tères vivant dans les forêts des Gorges de l’Areuse (Bulletins 118, 119, 120). Son article est consacré aux lucanidés (famille du lucane cerf-volant) et aux scarabées mangeurs de feuilles et de fleurs (chacun connaît au moins la cétoine dorée). Comme dans ses publi-cations précédentes, l’auteur relève le lien entre la diversité structurale de la forêts (vieux arbres, lisières naturelles, clairières) et la diversité entomologique.
Puis, dans une note, E. Della santa mentionne la présence d’une espèce de fourmi tropicale trouvée sur un frangipanier au Papiliorama de marin.
Enfin, comme de coutume, le Bulletin contient quatre rapports scientifiques concer-nant la météorologie régionale, le diagnostic parasitaire, l’aéropalynologie et, pour la troisième fois, Nature neuchâteloise, qui renseigne sur l’activité des services cantonaux concernés, de celui de la Protection de la Nature en particulier, en 1997. Un article concernant le plan de gestion du Loclat près de Saint-Blaise complète le rapport.
De quoi occuper quelques longues soirées d’hiver !
Les rédacteurs Willy matthey et Jacques Ayer
4

bulletin De la société neucHâteloise Des sciences naturelles 121 : 5-36. 1998
5
à LA RECHERCHE DES DINOSAURESrécit D’une caMpaGne De fouilles paléontoloGiques Menée aux états-
unis(Howe rancH, sHell, wyoMinG, 4 au 30 août 1997)
JACqUES AyER
Géologue, responsable du département Géologie-Paléontologie au muséum d’histoire natu-relle de Neuchâtel. Rue des Terreaux 14, 2000 Neuchâtel, Suisse.
Participer une fois dans sa vie à une fouille de fossiles de dinosauriens est certainement le rêve du paléontologue, pour qui la fascination à l’égard de ces animaux n’a d’égale que le mystère qui les entoure. Un rêve devenu réalité pour Bernard Claude, collaborateur au muséum d’histoire naturelle de Neuchâtel, et moi-même, lorsque nous nous sommes rendus au début août 1997 dans une région retirée des Etats-Unis, située dans l’Etat du Wyoming.
Ce projet a trouvé son origine une année auparavant lors d’une discussion avec Hans-Jakob Siber, directeur du Saurier museum à Aathal, près de Zurich. Ce dernier dirige un musée consacré aux dinosaures et mène régulièrement depuis 1989 des campagnes de fouilles sur un site paléontologique célèbre situé dans le Nord du Wyoming. L’idée d’or-ganiser un jour une exposition sur le thème des dinosaures nous avait déjà effleuré l’es-prit mais il nous avait paru jusqu’alors délicat de traiter ce sujet déjà largement exploité. Or, la possibilité d’une participation à des recherches sur le terrain devenait soudain l’oc-casion idéale pour réaliser notre projet.
Cet article se compose de trois parties: la première présente brièvement le cadre géo-graphique, géologique et historique du site; la deuxième relate le déroulement des fouilles du mois d’août 1997 et décrit les techniques développées sur le terrain et en laboratoire, alors que dans la troisième partie, en guise de conclusion, l’origine géologique du site est discutée.
1ère partie: présentation Du site
contexte GéoGrapHique
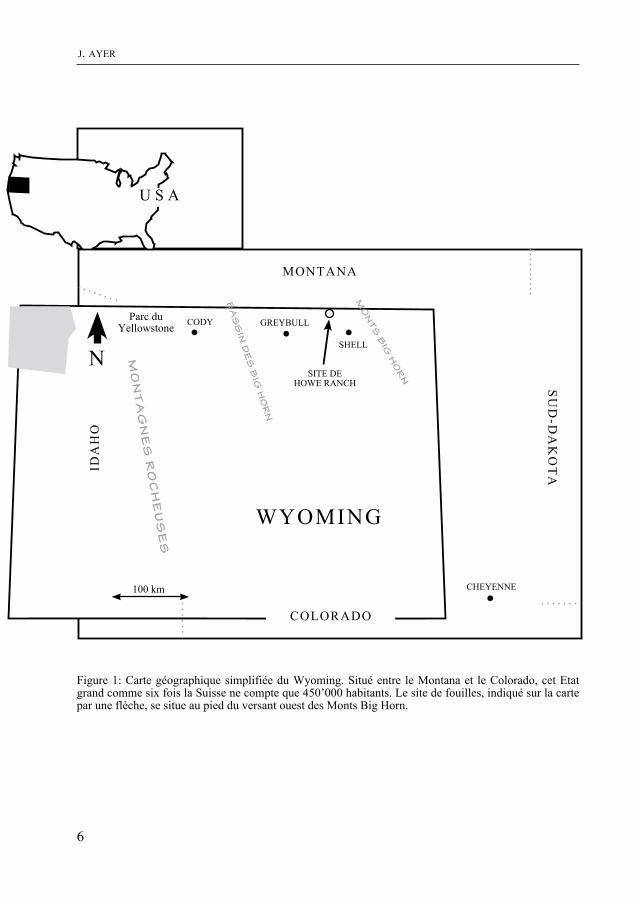
D’une superficie égale à six fois celle de la Suisse et avec une population de 450’000 habitants seulement, le Wyoming représente le deuxième état le moins peuplé des Etats-Unis après l’Alaska. Sa capitale, Cheyenne, est située à l’extrémité sud-est de l’Etat et ne compte que 50’000 âmes. Le nom Wyoming est à l’origine un mot indien qui signifie “Dans les Grandes plaines”. Le pays est d’ailleurs le berceau des tribus mythiques que
6
J. ayer
Figure 1: Carte géographique simplifiée du Wyoming. Situé entre le montana et le Colorado, cet Etat grand comme six fois la Suisse ne compte que 450’000 habitants. Le site de fouilles, indiqué sur la carte par une flèche, se situe au pied du versant ouest des monts Big Horn.
Mo
nt
s B
ig H
or
n
Ba
ss
in d
es
Big
Ho
rn
Mo
nt
ag
ne
s r
oc
He
us
es
GREyBULL
SHELL
CODy
CHEyENNE
Parc du yellowstone
SITE DE HOWE RANCH
WyOmING
COLORADO
mONTANA
IDA
HO
SUD
-DA
KO
TA
N
U S A
100 km

sont les Sioux, les Crow, les Shoshones ou encore les Cheyennes. L’Etat a connu au 19e siècle l’arrivée de centaines de milliers d’Européens venus conquérir de nouvelles terres cultivables ou prospecter les rivières aurifères. La célèbre épopée de William Cody, plus connu sous le nom Buffalo Bill, a notamment marqué cette époque. Il a donné son nom à la petite ville de Cody, située au Nord-Ouest du Wyoming, où un musée retrace sa vie.
Aujourd’hui, le Wyoming s’appuie économiquement sur l’extraction des richesses de son sous-sol, pétrole, gaz et charbon entre autres mais aussi sur l’éle-vage de bovins et de moutons. Le tou-risme, enfin, constitue une ressource non-négligeable, avec notamment la venue annuelle de plusieurs millions de visiteurs dans son célèbre Parc National du yel-lowstone (pitcHer, 1993).
Le site de fouilles se situe au centre-nord du Wyoming et porte le nom de Howe Ranch (fig. 1). La région marque la transition entre une plaine semi-déser-tique à l’ouest, le Bassin des Big Horn, et les premiers reliefs de la chaîne de mon-tagnes du même nom, à l’est. Perchés à 1600 mètres d’altitude, les lieux offrent un paysage aride jalonné de petites col-lines aux teintes variées. La végétation steppique est dominée par des buissons d’armoise. C’est le milieu de prédilection du serpent à sonnette - ou crotale - présent en grand nombre. Les coyotes fréquen-tent également régulièrement l’endroit. De plus, on assiste certaines années à de véritables invasions de lapins. Le climat de cette région du Wyoming est de type continental. Les étés y sont normalement très chauds et secs alors que la température peut descendre jusqu’à 40 degrés en des-sous de zéro durant l’hiver.
Le site de Howe Ranch comprend en fait deux carrières principales, Howe quarry et Howe Stephens quarry, et quelques points
de fouilles isolés (fig. 2). La région est loin de toute habitation, seuls les vestiges d’une bergerie attestent de la présence passée d’une ferme d’élevage. On y accède en une heure environ de voiture depuis le vil-lage le plus proche, Greybull. Les derniers kilomètres s’effectuent sur une piste diffi-cile, impraticable par mauvais temps.
caDre GéoloGique
Contexte stratigraphiqueLa série stratigraphique de la région
prospectée est bien développée. Une cou-verture sédimentaire allant du Cambrien au Crétacé repose sur un socle cristallin pré-cambrien affleurant au niveau d’un grand chevauchement au sein des monts Big Horn.
Dans les formations paléozoïques, de grandes falaises blanches de calcaire marin sont visibles depuis le site de fouilles. Cette formation, d’âge carbonifère (mis-sissipien), contient des niveaux d’algues marines fossiles que l’on retrouve très bien conservées dans des blocs erratiques proches de la carrière.
La série mésozoïque est caractérisée par une alternance de sédiments marins et continentaux. Le Jurassique est représenté dans la région par trois formations. La plus jeune d’entre elles, la Formation de mor-rison, est composée exclusivement de sédi-ments continentaux. Les fossiles de dino-saures du site de Howe Ranch proviennent de ces couches géologiques (fig. 2).
La Formation de morrison
La Formation de morrison est célèbre pour les nombreux fossiles de dinosaures qu’elle renferme. Au siècle passé déjà, elle fut à l’origine des premières grandes découvertes historiques de dinosaures géants sur le continent nord-américain. Les sites de Como Bluff au Wyoming et
7
à la recHercHe Des Dinosaures

8
J. ayer
Figure 3: vue panoramique en direction du sud sur la région de Howe Ranch et la Formation de mor-rison. On peut distinguer, au milieu de la photo, la carrière de Howe Stephens quarry.
Figure 2: Plan du site de Howe Ranch avec la localisation des deux principales carrières et des autres points de fouilles. En filigrane, l’extension schématique de la Formation de morrison, situé stratigraphi-quement entre une formation marine jurassique (Formation de Sundance) et une formation continentale crétacée (Formation de Cloverly).

du Dinosaur National monument en Utah sont les principaux gisements de ce type.
La Formation de morrison s’étend du Nouveau-mexique au sud du montana et de l’Utah à l’ouest jusqu’à l’Oklahoma à l’est, sur une superficie totale d’environ 1,5 millions de km2. Datée approximati-vement entre 155 et 145 millions d’an-nées, soit du Jurassique supérieur, elle a pour origine une grande plaine alluviale qui s’installa à l’époque après le retrait de la mer (mer de Sundance). De nombreux fleuves y drainaient de grandes quan-tités de sédiments, notamment depuis des reliefs situés au niveau des montagnes Rocheuses actuelles. Outre une grande quantité d’ossements de dinosaures, la Formation de morrison renferment éga-lement des fossiles de poissons, de batra-ciens, de crocodiliens ou encore de petits mammifères (benton, 1998; currie & paDian, 1997).
Dans la région de Howe Ranch, la Formation de morrison a été datée, par méthodes radiométriques, à 147 millions d’années. Constituée d’argiles, de grès et d’une faible proportion de calcaires, cette formation atteint une épaisseur d’environ 60 mètres (fig. 3).
Les argiles sont prédominantes. Elles se présentent sous deux formes: d’une part des bentonites gris-clair ou viola-cées riches en smectite, et d’autre part, des argiles bigarrées composées essen-tiellement d’illite et présentant des teintes tantôt rouges, tantôt verdâtres. On observe parfois dans ces argiles des fentes de dessication (mudcracks) et des nodules riche en calcaire appelés caliche nodules.
Les sédiments gréseux, de couleur beige-clair, forment des niveaux centimé-triques à décimétriques présentant par-fois des stratifications entrecroisées. On rencontre fréquemment dans ces roches des niveaux riches en matière organique, composée essentiellement de végétaux et
de bois sous forme de lignite dont certains fragments dépassent le mètre. Il n’est pas rare également d’observer des horizons à galets mous (fig. 4).
Enfin, de rares niveaux calcaires de cou-leur rouille se rencontre occasionnellement dans les argiles bigarrées.
Une interprétation de ces observations sédimentologiques est présentée à la fin de cet article sous le chapitre conclusions.
Contexte structuralLa géométrie des structures dans la
région étudiée est relativement complexe. Un chevauchement de grande envergure au Nord-Est a amené le socle cristallin sur les couches paléozoïques et mésozoïques au sein des monts Big Horn. Ces niveaux sédimentaires forment de grands plis cof-frés sur le versant ouest de la chaîne. Le site de Howe Ranch se situe sur le flanc oriental d’un grand synclinal kilométrique. La structure est chevauchante à l’ouest.
La géométrie des couches aux alen-tours du site de fouilles ne présente pas de grandes perturbations. Cependant, dans la carrière de Howe Stephens quarry, les niveaux gréseux forment une flexure avec un fort pendage local indépendant de l’in-clinaison général des couches. L’origine de cette structure reste pour le moment inconnue.
Historique Du site De Howe rancH
Les premières prospections en 1934Le site paléontologique prospecté a
fait l’objet de recherches il y a plus de soixante ans déjà. A cette époque, un chercheur de fossiles du nom de Barnum Brown fut envoyé par l’American museum of Natural History de New-york pour récolter des échantillons dans le Nord du Wyoming (fig. 5). Alors qu’il menait ses recherches sur le terrain, il fut informé de l’existence dans la région
9
à la recHercHe Des Dinosaures

10
J. ayer
Figure 4: Observations sédimentologiques dans la Formation de morrison. A: niveaux à galets mous centimétriques; B: stratifications entrecroisées dans les niveaux gréseux; C: horizons riches en débris végétaux fossiles; D: grand bois lignifié de plus d’un mètre de long.
A B
C D

11
de très grands ossements fossiles proche d’un ranch appartenant à un certain Barker Howe. Deux ans plus tard, soit en 1934, Brown se rendit sur les lieux pour diriger des fouilles. Un livre passionnant relate les aventures paléontologiques de Barnum Brown (birD, 1985) et consacre tout un chapitre à ce site du Wyoming. On y découvre dans un passage l’exaltation de Brown lorsqu’il contempla pour la pre-mière fois le gisement: «C’est un véri-table trésor de dinosaures - s’écria-t-il - un extraordinaire trésor de dinosaures !». On remarqua en effet très vite le caractère exceptionnel du site. Après six mois de fouilles, on avait dégagé dans la carrière prospectée, mesurant 14 x 20 m et baptisée Howe quarry, plus de 4000 ossements fos-
siles de dinosaures. Avec une telle concen-tration, ce gisement représente un des sites jurassiques de ce type parmi les plus riches au monde! Tous les ossements de ces fouilles furent transportés par train et entreposées dans les collections de l’Ame-rican museum of Natural History de New-york. Le matériel, d’une grande valeur scientifique, n’a pourtant fait l’objet d’au-cune étude détaillée au cours des années qui suivirent. A l’époque, Barnum Brown était apparemment très occupé par d’autres recherches. Ceci allait s’avérer d’autant plus regrettable que, quelques années plus tard, un terrible incendie ravagea les col-lections du muséum faisant ainsi dispa-raître le 90% du matériel trouvé à Howe quarry. Une riche et précieuse documen-
à la recHercHe Des Dinosaures
Figure 5: Lilian et Barnum Brown dans la carrière de Howe quarry en 1934, contemplant quelques osse-ments spectaculaires d’un dinosaure sauropode. (Neg. N° 2A6933 Photo. Barnum Brown. Courtesy Department Library Services, American museum of Natural History, New-york).

12
tation paléontologique venait de s’envoler en fumée.
De nouvelles fouilles à partir de 1989Suite à cette fin dramatique, le site allait
peu à peu tomber dans l’oubli. Aucune nouvelle prospection ne sera menée jusqu’à la fin des années 80, époque où Hans-Jakob Siber commença à s’inté-resser de très près à ce gisement fabuleux. En fait, c’est la lecture de l’ouvrage de birD (1985) qui le persuada de reprendre les recherches dans cette région. Après de longues et fastidieuses démarches auprès des propriétaires du terrain, il par-vint à obtenir le droit d’ouvrir à nouveau la carrière de Howe quarry et put ainsi démarrer son projet de fouilles paléon-tologiques qu’il nomma “The Big Horn mountain Dinosaur project ”. En 1992, une nouvelle carrière était ouverte par Siber et son équipe à environ 450 mètres au sud-ouest de Howe quarry. Le nou-veau gisement, baptisé Howe Stephens quarry, allait se montrer très vite tout aussi prometteur que le premier.
La législation américaine en matière de fouilles paléontologiques
Les lois américaines concernant la récolte et l’exportation de fossiles trouvés sur territoire national sont d’une grande sévérité (fig. 6). Cependant, lorsqu’il s’agit d’une parcelle privée, le gouvernement donne de larges droits aux propriétaires du terrain. Le site de Howe Ranch se situe précisément sur un domaine privé. Par conséquent, en payant à ses propriétaires une location annuelle, Hans-Jakob Siber s’octroie la concession paléontologique qui lui permet de dégager puis de ramener en Suisse tous les fossiles mis au jour sur son site de fouilles.
Il prit conscience, bien malgré lui, de la rigidité de cette législation lorsqu’en 1991, après avoir découvert et dégagé avec son
équipe un squelette d’allosaure presque complet, il se le fit confisquer par les auto-rités américaines. Le fossile, nommé Big Al One, une pièce exceptionnelle (fig. 7), se trouvait en fait à quelques centaines de mètres du gisement de Howe quarry, à la limite du terrain de l’Etat. Selon les habitants de la région, le lieu de la décou-verte appartenait au domaine privé. mal-heureusement, la venue de responsables de l’aménagement du territoire allait démontrer que les anciennes délimitations cadastrales étaient inexactes. Leurs appa-
J. ayer
Figure 6: La limite cadastrale actuelle entre le terrain privé et le terrain de l’Etat est matérialisée à Howe Ranch par un panneau de mise en garde. On peut y lire notamment: «Les restes fossiles animaux et végétaux ont une valeur scientifique et pédagogique. Ce terrain et les fossiles qui s’y trouvent sont la propriété du Gouvernement des Etats-Unis. Toute récolte non-autorisée sera pas-sible de poursuites judiciaires ».

13
reils de mesure indiquaient que l’allo-saure se trouvait sur le terrain de l’Etat, à moins de soixante mètres du terrain privé. En conséquence, les ossements, devenus propriété du Gouvernement, furent confis-qués et puis déposés dans les collections du museum of the Rockies à Bozeman, au montana.
Bilan des fouilles entre 1990 et 1996Les années suivantes allaient faire vite
oublier la malheureuse aventure du dino-saure Big Al One. Entre 1990 et 1996, Siber et ses collaborateurs ont dégagé sur le site de Howe Ranch près de quinze sque-lettes dont cinq sont complets. Ceux-ci appartiennent à au moins huit genres de dinosaures différents. Cette grande diver-
sité constitue, aussi bien sur le plan quan-titatif que qualitatif, une des collections les plus complètes de dinosaures du Jurassique supérieur sur le continent nord-américain.
Les fossiles extraits s’apparentent aux genres de dinosaures suivants: • Diplodocus, Camarasaurus, Apatosaurus et Barosaurus, dinosaures herbivores géants (sauropodes)• Stegosaurus, dinosaure ornithischien caractérisé par de grandes plaques dorsales• Allosaurus, dinosaure prédateur de grande taille (théropode), caractéristique du Jurassique supérieur• Othnielia et Dryosaurus, petits dino-saures ornithopodes.
à la recHercHe Des Dinosaures
Figure 7: Crâne de l’allosaure Big Al One en cours de dégagement. Le fossile, dans un état de conser-vation remarquable, a été découvert par Siber et son équipe en 1991, à 300 mètres au nord de Howe quarry. Sur la base de nouvelles mesures cadastrales, le squelette devenu propriété de l’Etat, leur sera confisqué quelques temps après par les Autorités américaines et déposé dans un musée au montana.(Photo H.-J. Siber)

14
J. ayer
Position systématique
Taille et poids (max.)
Régime alimentaire
Ornithischiens,Ornithopodes,Hypsilophontidae
1,4 m30 kg
herbivore
herbivore
herbivore
carnivore, parfois charognard
herbivore
herbivore
herbivore
herbivore
3,5 m75 kg
8 m4000 kg
7-12 m3000 kg
27 m10’000 kg
21 m25’000 kg
27 m?
25 m15’000 kg
Ornithischiens,Ornithopodes,Hypsilophontidae
Ornithischiens,Stégosaures,Stegosauridae
Saurischiens,Théropodes,Allosauridae
Saurischiens,Sauropodes,Diplodocidae
Saurischiens,Sauropodes,Diplodocidae
Saurischiens,Sauropodes,Diplodocidae
Saurischiens,Sauropodes,Camarasauridae
Othnielia
Dryosaurus
Stegosaurus
Allosaurus
Diplodocus
Apatosaurus
Barosaurus
Camarasaurus
Tableau 1: Silhouettes des huit genres de dinosaures trouvés à Howe Ranch. Les proportions entre les animaux sont approximativement respectées.

15
à la recHercHe Des Dinosaures
Une image de ces différents dinosaures est présentée dans le tableau 1.
Entre 1990 et 1991, la carrière de Howe quarry a produit entre autre près de 500 nouveaux ossements dont la plupart appartiennent au genre Diplodocus. Elle a permis également la mise au jour de plu-sieurs empreintes tridactyles d’Allosaurus, partielles ou complètes. Cette découverte est exceptionnelle car la présence à la fois d’ossements et d’empreintes de dinosaures sur un même gisement reste à ce jour un phénomène extrêmement rare.
Durant cette même période, la mise au jour d’empreintes de peau de dinosaures sauropodes, fossiles jusque là rarement découverts dans des couches jurassiques, a apporté de précieux renseignements sur la morphologie externe de ces animaux. La venue sur le site de Howe Ranch de Stephen Czerkas, spécialiste en la matière, a permis d’étudier dans le détail une cen-taine d’échantillons, dont certains pré-sentaient une forme triangulaire. Ces empreintes de peau insolites, dégagées à proximité des vertèbres caudales d’un diplodocus, ont amené le chercheur à attri-buer ces fossiles à une crête cornée partiel-lement conservée. La publication de ses observations l’ont amené ensuite à pro-poser une nouvelle image des sauropodes. Aujourd’hui, les reconstitutions de diplo-docus font apparaître une crête, ressem-blant à celle des iguanes, tout au long du dos et de la queue de ce dinosaure (tab. 1).
Ouverte en 1992, la carrière de Howe Stephens quarry allait réserver des sur-prises encore plus belles. De 1992 à 1995, ce ne sont pas moins de 4 squelettes par-tiels de diplodocus qui ont été dégagés. En 1992, la mise au jour d’un doigt de camarasaure dressé vers le ciel signalait en fait la présence d’un squelette complet que l’on surnomma pour la circonstance E.T.
Le dernier jour de la campagne de fouilles de 1995 a été le début d’un véri-table conte de fée. Ce jour-là, Hans-Jakob Siber a remarqué la présence d’un os res-semblant à un ischion (pièce du bassin) alors qu’il rangeait les outils pour la pause hivernale. Sa curiosité l’amena à dégager un peu plus le fossile qui semblait se pour-suivre dans le terrain. Des vertèbres sont apparues, puis un fémur et un tibia. Le mauvais temps et le calendrier allaient malheureusement contraindre le paléon-tologue à quitter les lieux sans connaître la nature de sa nouvelle découverte. De retour en Suisse, Hans-Jakob Siber eut tout l’hiver pour rêver à son mystérieux fos-sile. Parmi les interprétations avancées, on envisagea la présence d’un nouveau sque-lette de camarasaure. Les premiers jours de la campagne de fouilles 1996 allaient très vite apporter la réponse à cette énigme: il s’agissait en fait d’un squelette de sté-gosaure dont le dégagement allait de plus mettre au jour un deuxième trésor paléon-tologique: au niveau de la jambe du stégo-saure, on remarqua la présence de la queue d’un dinosaure carnivore. Après quelques semaines de fouilles, le constat dépassait toutes les espérances. Hans-Jakob Siber avait sous les yeux deux squelettes arti-culés et complets, ceux d’un stégosaure et d’un allosaure, surnommés respectivement victoria et Big Al Two.
A cette liste exceptionnelle s’ajoute la mise au jour d’un squelette complet de dryosaure et de deux squelettes partiel du genre Othnielia. Au total, plus de 2000 ossements ont été extraits parmi lesquels seul une vingtaine n’ont pu être attribués à un squelette identifié.
L’ouverture d’une petite fouille à quelques dizaines de mètres de Howe Ste-phens quarry (m-pit) a permis en 1995 la découverte d’un squelette d’apatosaure (max) dont il ne manque que les vertèbres caudales, et d’un squelette partiel de stégo-saure (moritz).

16
J. ayer
Tableau 2: Synthèse des découvertes faites sur le site de Howe Ranch lors des fouilles de Barnum Brown en 1934 et par l’équipe du Saurier museum d’Aathal de 1990 à 1996. La localisation exacte des carrières est présentée à la figure 2.

17
à la recHercHe Des Dinosaures
Le résumé de ces résultats est présenté dans le tableau 2.
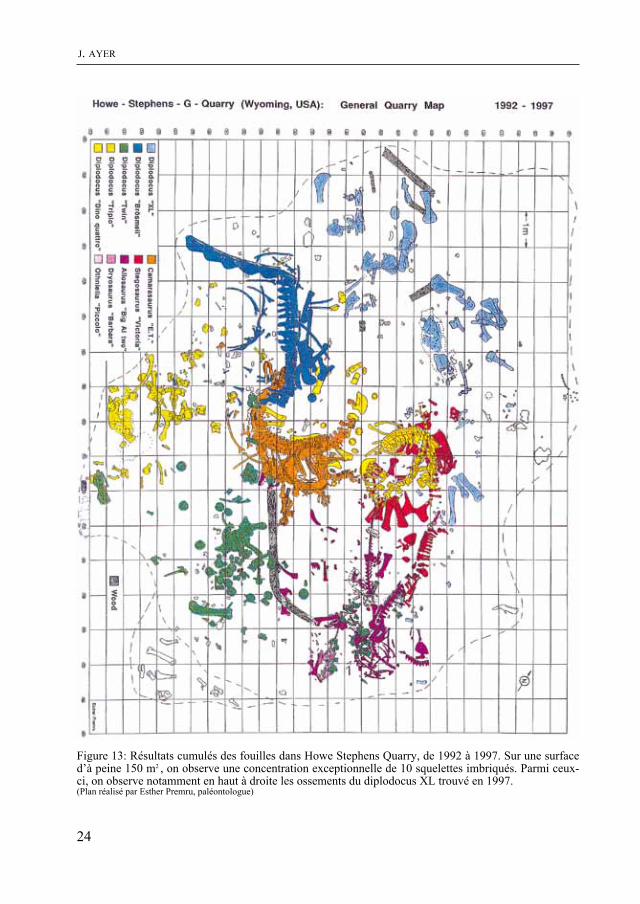
La campagne de fouilles 1997 s’est déroulée du 4 au 30 août. quelques semaines avant les recherches sur le ter-rain, un programme de fouilles a été mis sur pied afin de définir les objectifs et de tirer le meilleur profit du temps à disposi-tion sur le site. On a pris en compte notam-ment les résultats des fouilles précédentes, résumés sur un plan illustrant la répartition des fossiles dans le gisement de Howe Ste-phens quarry (fig. 14). Par extrapolation, on a tenté de repérer les niveaux géolo-giques et les secteurs de la carrière suscep-tibles de recéler de nouveaux squelettes. Le programme 1997 comportait deux objectifs principaux: le premier consistait à faire progresser le front de fouilles en direction du Nord afin de retrouver en pro-fondeur un niveau riche en fossiles alors que le second visait à dégager définitive-ment, sur le même gisement, les squelettes complets de l’allosaure Big Al Two et du stégosaure victoria, trouvés en 1996.
en route pour le site...Notre premier contact avec l’Ouest
américain ne s’est pas fait sans une cer-taine émotion. C’est à Billings, une des principales villes du montana, qu’a pris fin notre voyage en avion, après plus de quinze heures de vol. La location d’une voiture allait ensuite nous permettre de gagner par nos propres moyens le site de fouilles. La ville de Billings, disparue de notre champ de vision, a très vite laissé place à des paysages de plus en plus sau-vages, aux horizons infinis. Un décor gran-diose, aux formes géologiques parfaites, se déroulait sous nos yeux. La présence d’un couple d’aigles dorés perchés sur un rocher, quelques chiens de prairies à l’affût ou un troupeau de pronghorns, une espèce d’antilope, apportaient parfois une touche
de vie à cette immensité minérale. La route, fidèle à la théorie qui veut que la plus petite distance reliant deux points est une ligne droite, nous paraissait sans fin. Après deux heures et demi de voyage, la petite bour-gade de Greybull était atteinte (fig. 8). Cette ville est la dernière localité avant le site de fouilles. Peuplé d’environ 1800 habi-tants, Greybull doit son nom, raconte-t-on, à la présence passée d’un très grand bison albinos, vénéré à l’époque par les Indiens de cette région. De cet endroit, il fallait encore compter une heure de route: le regard fixé sur notre plan d’accès, nous nous sommes enfonçés progressivement dans un paysage envoûtant aux formes et aux teintes sur-naturelles (fig. 9). Après quelques hésita-tions de notre part, tout à fait justifiées au demeurant, nous sommes arrivés finalement au but par une route transformée sur les derniers kilomètres en une piste périlleuse. Le campement, regroupé autour de quelques arbres, était vide. Après avoir emprunté un chemin sur quelques centaines de mètres, nous avons rencontré nos compagnons de fouilles dans la carrière déjà mythique de Howe Stephens quarry, ce mardi 15 août 1997 en fin d’après-midi.
une Journée sur la fouille
L’équipe de base comptait six per-sonnes au total: Hans-Jakob Siber et sa fille maya; deux bénévoles: Christoph Keilmann et Esther Wolfensberger; et enfin Bernard Claude et moi-même du muséum de Neuchâtel. Elle fut complétée plus tard par la venue de Ben Pabst, préparateur au Saurier museum d’Aa-thal, Ioannis michelis, paléontologue à Bonn et Denis maurer, reporter vidéo à Colombier, mandaté par le muséum de Neuchâtel pour filmer le travail sur les fouilles (fig. 23).
vu l’isolement du site, notre campement devait pouvoir jouir d’une certaine auto-
2e partie: LA CAmPAGNE DE FOUILLES DE 1997

18
J. ayer
Figure 8: A l’entrée du village de Greybull, on est tout de suite plongé dans l’univers des dinosaures. Une artiste a décoré les murs en contre-bas de la route principale en s’inspirant de ces géants du passé.
Figure 9: Paysage de Western dans les environs du site de Howe Ranch. Au premier plan, on distingue quelques buissons d’armoise.

nomie. Les deux premiers jours avant notre arrivée avaient été réservés à la mise en place de toute l’infrastructure qui nous per-mettrait de séjourner un mois durant dans ces conditions particulières. L’eau potable était puisée dans une source voisine. Les tentes, installées les unes à côté des autres, accompagnaient une ancienne caravane faisant office à la fois de cuisinière à gaz et d’abri en cas de mauvais temps. Tous les trois jours, une équipe se rendait à Grey-bull pour le ravitaillement en nourriture et l’achat de matériel au besoin.
Les journées suivantes allaient être consacrées au travail de prospection propre-ment dit. Après un petit déjeuner copieux, toute l’équipe se rendait dans la carrière de Howe Stephens quarry en prenant soin d’emporter à chaque fois les litres d’eau indispensables à notre hydratation. La tem-pérature dans la carrière pouvait en effet s’élever à plus de 40 degrés, ceci couplé à un taux d’humidité très faible. Pour pallier quelque peu à ces conditions parfois diffi-ciles, quelques tentures étaient suspendues au-dessus de la carrière pour nous permettre de bénéficier d’un peu d’ombrage aux heures les plus chaudes. De plus, une petite cabane en bois fut construite aux abord du gisement afin d’entreposer les outils et de protéger le matériel photographique.
Le travail de fouilles faisait appel à des compétences très diverses. En véritables terrassiers, il nous fallait creuser à la pioche ou à la pelle et évacuer des centaines de kilos de gravats au moyen de brouettes. Puis, à chaque trouvaille, c’est avec la patience et la minutie d’un horloger que les fossiles étaient peu à peu dégagés. Notre polyvalence ne s’arrêtait pas là: il fallait encore pratiquer le travail du géomètre pour cartographier les fossiles ou celui du plâtrier pour consolider les ossements. Toutes ces activités étaient bien sûr complétées par des mesures géologiques sur le site et par des séances de discussion portant avant tout sur l’origine du gisement. Cette pluridisciplina-
rité rendait le travail de fouilles encore plus passionnant.
Le repas de midi était pris au campe-ment où l’ombre de quelques grands arbres nous apportait une délicieuse sensation de fraîcheur. La journée sur les fouilles se prolongeait jusqu’au crépuscule et se terminait souvent par le merveilleux spec-tacle du coucher de soleil embrasant les monts Big Horn (fig. 10). Le soir, nous prenions un bon repas autour du feu. Après quoi, la nuit illuminée de myriades d’étoiles et bercée parfois par le chant des coyotes nous plongeait dans un sommeil profond et réparateur.
Un imprévu est venu s’ajouter à notre programme quotidien. Le campement était relié à la carrière par une piste à peine carrossable qui devait permettre, à la fin des de fouilles, le transport par véhicules des fossiles. malheureusement, de fortes pluies printanières avaient grossi le petit ruisseau que le chemin enjambait, empor-tant du même coup le pont de fortune permettant l’accès au gisement. Nous n’avions alors pas d’autre alternative que de rétablir à tout prix le passage. Ainsi, pendant près d’une semaine et aux heures les moins chaudes, nous avons comblé le fossé au moyen de plusieurs dizaines de brouettes remplies de gravats.
Cette construction allait d’ailleurs nous causer quelque frayeurs lorsque le jeudi 28 août, à la fin des fouilles, les fossiles quittèrent la carrière pour leur trans-port jusqu’à l’aéroport. Les véhicules, en l’occurence une camionnette et deux bétaillères mises généreusement à dis-position par un propriétaire de ranch de la région, s’avançaient prudemment sur notre pont de fortune, à l’entrée du cam-pement, lorsqu’un pan de terre commença à glisser sous le poids du chargement. Un mois entier de travail de fouilles était soudain menacé. Heureusement, la dex-térité du chauffeur et la mise en place en catastrophe de poutrelles sous les roues allaient enfin permettre aux véhicules de
19
à la recHercHe Des Dinosaures

20
poursuivre leur chemin, indemnes.
une caMpaGne fructueuse
Les 4 semaines de fouilles ont abouti à de très bons résultats. Au total, plus de 5 tonnes de matériel ont été extraits, os et gangue de sédiments compris. Dès les premiers jours, nous mettions la main sur de grands ossements appartenant proba-blement au genre Diplodocus. On dégagea notamment de grandes vertèbres cervicales et dorsales, une omoplate de 1,5 m de long, un humérus et des pièces du bassin - pubis et ischion (fig. 11, en haut). Au total, près de 35 ossements ont été mis au jour. La grande dimension des fossiles nous amena à surnommer ce diplodocus XL.
quelques vertèbres et une griffe (fig. 11, en bas) d’un autre spécimen de diplodocus, surnommé Twin, sont venues compléter
les ossements trouvés durant les fouilles précédentes. Ce dinosaure intéresse parti-culièrement le muséum de Neuchâtel qui entend acquérir définitivement une partie du squelette, notamment une patte posté-rieure et le bassin complet, le tout d’une hauteur de 3,5 mètres.
La campagne 1997 a permis également d’extraire définitivement les squelettes de l’allosaure Big Al Two (fig. 12, en haut) et du stégosaure victoria, découverts tous deux en 1996. Leur dégagement a de plus abouti à d’étonnantes découvertes: le qua-trième pic caudal du stégosaure victoria qui manquait jusqu’ici fut notamment mis au jour. En outre, une dent de poisson, proba-blement du genre Ceratodus, a été localisée à proximité de la cage thoracique de l’al-losaure. Cette trouvaille pourrait apporter
J. ayer
Figure 10: A chaque fin de journée, le soleil couchant offrait un spectacle surnaturel. A l’arrière-plan: la chaîne des monts Big Horn.

21
à la recHercHe Des Dinosaures
Figure 11: En haut: vue d’ensemble des ossements du diplodocus XL. On distingue, en haut de la photo, de gauche à droite, un humérus et une omoplate de grande taille. En bas: Griffe du diplodocus Twin.

22
J. ayer
Figure 12: En haut: Ossements de la patte avant de l’allosaure Big Al Two. En bas: Ossements de la patte arrière d’un petit dinosaure, probablement du genre Othnielia.

23
à la recHercHe Des Dinosaures
de précieux renseignements sur le régime alimentaire de ce dinosaure prédateur.
Big Al Two et victoria sont deux spéci-mens parfaitement conservés et complets à près de 100%. Sur le plan mondial, ils représentent deux trouvailles d’importance majeure. Le squelette de l’allosaure, dont la taille est estimé à 8 mètres de long et à 3 mètres de haut, constitue du reste la trouvaille la plus complète jamais réalisée sur le continent américain. Le fossile, dont la préparation en laboratoire est estimée à plus d’une année, sera d’ailleurs l’attrac-tion principale de l’exposition à Neuchâtel, prévue en 1999.
Nos recherches ont abouti également à la découverte d’ossements appartenant à un dinosaure de très petite taille, probable-ment du genre Othnielia. Il s’agit en parti-culier d’un fémur et d’un tibia de quelques centimètres seulement (fig. 12, en bas).
La campagne de fouilles 1997 a donc été pleinement réussie. Elle constitue d’ailleurs une des plus riches de ces neuf dernières années. En ajoutant ces résultats aux découvertes précédentes, on obtient dans Howe Stephens quarry une concen-tration de squelettes exceptionnelle que l’on peut illustrer sur un plan (fig. 13).
les tecHniques De fouilles
Les méthodes pratiquées sur le gisement ressemblent aux techniques traditionnelles de prospection paléontologique. voici en résumé les différentes étapes qui sépa-rent la découverte d’un os fossilisé de son transport définitif:
La découverte d’un fossileet son dégagement
Le travail de fouilles consiste dans un premier temps à prospecter grossière-ment le terrain au moyen de pioches, de marteaux et de burins. A chaque coup, le contenu des gravats doit être vérifié
afin de repérer à temps la présence éven-tuelle d’un fragment d’os. Dans ce cas, on met immédiatement de côté les gros outils et on tente de localiser au plus vite d’où le débris d’os vient d’être arraché. Parfois, plusieurs heures d’excavation sont nécessaires avant de toucher l’objet tant recherché. La mise au jour du fossile récompense alors les durs efforts consentis à sa découverte.
Au moyen d’un pinceau ou d’un cou-teau, le contour et la surface du fossile sont dégagés minutieusement pour permettre plus tard son identification. Il est impor-tant que les limites de l’os ou du fragment soient parfaitement définies dans l’espace afin d’éviter toute perte au moment de son extraction. De plus, au cours du déga-gement, une goutte de colle forte insta-tanée est systématiquement appliquée sur chaque petite fissure. Le climat aride de cette région provoque une dessication qui fragilise considérablement le fossile. Pour le stabiliser, on imprégne l’os, à plusieurs reprises, d’une résine appelée fond d’ac-crochage acrylique.
Le dégagement doit tenir compte de deux aspects contradictoires. D’une part, il faut retirer le maximum de sédiments entourant les ossements afin de ne pas surcharger inutilement le lot de fossiles destiné au transport. D’autre part, un net-toyage excessif des ossements avec des outils inappropriés risque d’endommager le fossile ou de le fragiliser, compromet-tant son exportation vers la Suisse. La solution doit être adaptée à chaque cas. Par exemple, lorsqu’une portion de terrain ren-ferme une multitude de fragments osseux indéterminables ou, dans le cas d’un sque-lette articulé ne permettant pas d’isoler un à un les différents ossements, il est souhai-table de conserver et de circonscrire cette accumulation dans son ensemble en vue d’un dégagement plus complet en labora-toire. Dans ce cas, la gangue de sédiment, qui doit jouer le rôle de cocon protecteur pendant le transport, est imprégnée d’une
24
J. ayer
Figure 13: Résultats cumulés des fouilles dans Howe Stephens quarry, de 1992 à 1997. Sur une surface d’à peine 150 m2 , on observe une concentration exceptionnelle de 10 squelettes imbriqués. Parmi ceux-ci, on observe notamment en haut à droite les ossements du diplodocus XL trouvé en 1997. (Plan réalisé par Esther Premru, paléontologue)
25
solution de colle blanche plus ou moins diluée, destinée à renforcer l’ensemble.
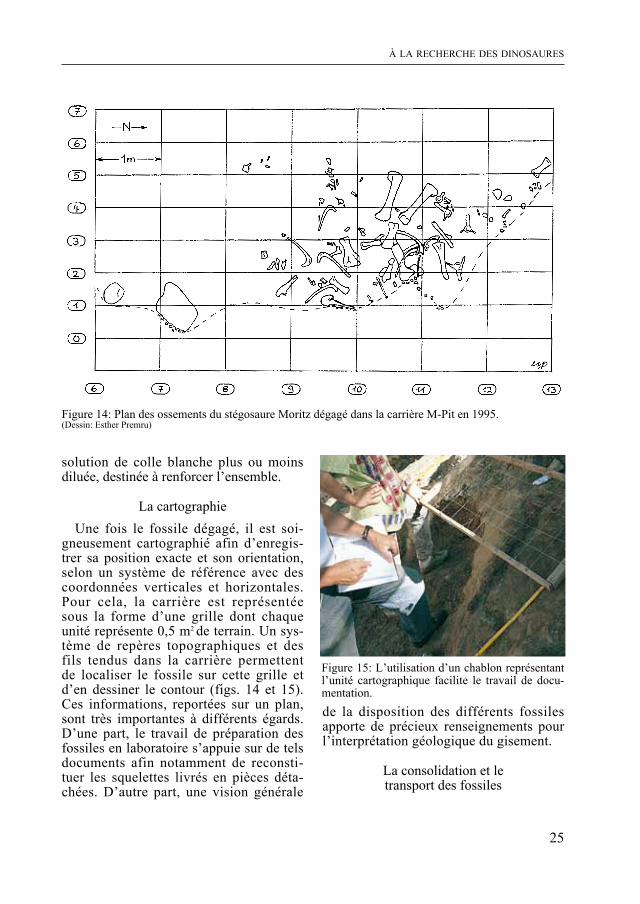
La cartographieUne fois le fossile dégagé, il est soi-
gneusement cartographié afin d’enregis-trer sa position exacte et son orientation, selon un système de référence avec des coordonnées verticales et horizontales. Pour cela, la carrière est représentée sous la forme d’une grille dont chaque unité représente 0,5 m2 de terrain. Un sys-tème de repères topographiques et des fils tendus dans la carrière permettent de localiser le fossile sur cette grille et d’en dessiner le contour (figs. 14 et 15). Ces informations, reportées sur un plan, sont très importantes à différents égards. D’une part, le travail de préparation des fossiles en laboratoire s’appuie sur de tels documents afin notamment de reconsti-tuer les squelettes livrés en pièces déta-chées. D’autre part, une vision générale
de la disposition des différents fossiles apporte de précieux renseignements pour l’interprétation géologique du gisement.
La consolidation et le transport des fossiles
à la recHercHe Des Dinosaures
Figure 15: L’utilisation d’un chablon représentant l’unité cartographique facilite le travail de docu-mentation.
Figure 14: Plan des ossements du stégosaure moritz dégagé dans la carrière m-Pit en 1995. (Dessin: Esther Premru)

Avant leur extraction et plus tard en vue de leur transport, les fossiles sont conso-lidés selon une méthode adaptée à la taille des ossements. Pour les petits fragments de quelques centimètres, aucune préparation particulière n’est nécessaire et le fossile est simplement extrait par sa base au moyen d’une lame de couteau par exemple. Pour les ossements complets isolés et les par-ties de squelettes, l’utilisation de plâtre est absolument indispensable. Pour ce faire, on découpe des bandes de tissu dans de la toile de jute, alors que du plâtre liquide est préparé simultanément. Ces bandes trempées dans le plâtre sont soigneusement appliquées sur toute la surface de l’os pré-alablement recouvert de feuilles d’alumi-nium (fig. 16). L’exercice doit s’effectuer rapidement car, au bout de deux à trois minutes déjà, le plâtre liquide durcit et devient inutilisable. Au bout de quelques heures, une fois séchés par le soleil, les fossiles sont prêts à être enfin libérés de leur linceul de pierre. La vision de ces momies immaculées posées sur le sol est tout à fait surprenante (fig. 17).
L’extraction définitive des ossements est un moment toujours délicat. Dans un premier temps, on sectionne le “cordon ombilical” qui lie encore le fossile plâtré à
son substrat. Puis, d’un seul mouvement, on retourne le tout afin de vérifier si aucun fragment d’os ne se trouve encore dans le sol. Le résultat de la manœuvre vient ainsi sanctionner en quelques secondes les heures de travail préliminaire passées à préparer le fossile.
La préparation pour le transport est adaptée à la dimension des échantillons. Les fossiles de petite et moyenne taille sont emballés minutieusement dans des feuilles d’aluminium et du papier journal avant d’être rangés dans des car-tons prêts au transport. Par contre, pour les grands fossiles, dont certains peu-vent peser jusqu’à 1000 kg, la démarche est plus délicate. Les ossements plâ-trés sont déposés tout d’abord sur une palette de transport. Puis, à la force des bras ou au moyen d’un treuil manuel, le tout est glissé, centimètre par centi-mètre, jusqu’aux véhicules. Pour faci-liter la manœuvre, des rondins de bois sont placées à l’avant et sous les palettes pour créer un effet de tapis roulant, à la manière des bâtisseurs de pyramides.
A la fin de la campagne de fouilles 1997, le transport s’est effectué dans un premier temps jusqu’à Greybull où le chargement fut déposé provisoirement dans un abri. quelques jours après, le tout était rangé dans un grand conteneur et conduit par camion jusqu’à l’aéroport de Denver, au Colorado. Il ne restait plus à nos dinosaures qu’à survoler l’At-lantique par avion jusqu’en Suisse pour être livrés par camion, quelques jours après et dans un parfait état de conserva-tion, devant la porte du Saurier museum d’Aathal.
le nettoyaGe Des osseMentsen laboratoire
Le travail en laboratoire représente une étape capitale, mais ô combien labo-
26
J. ayer
Figure 16: Application de bandes de jute trempées dans du plâtre sur la surface d’une vertèbre.

27
à la recHercHe Des Dinosaures
Figure 17: Une fois plâtrés, tels des sarcophages, les grands ossements offrent une vision insolite.

28
rieuse. En effet, on évalue que la prépara-tion et le montage définitif d’un squelette requièrent vingt fois plus de temps que son dégagement sur le terrain. La prépa-ration de l’allosaure Big Al Two dégagé durant les campagnes de fouilles 1996 et 1997 a été entreprise dès son arrivée au Saurier museum d’Aathal, soit en octobre 1997. Ben Pabst, le préparateur, a estimé la durée du travail à près de 2000 heures. Sans trop insister sur les techniques utili-sées, on peut résumer ainsi les principales étapes de la préparation des ossements en laboratoire:• Se référant aux données cartographiques de terrain, le préparateur tente d’abord d’identifier le fossile à dégager et estime sa position dans la gangue sédimentaire. Il peut s’aider également de représentations anatomiques tirées de la littérature.
• Le travail de dégagement des ossements s’effectue prudemment au moyen de petits outils ou de différentes fraises mécaniques. Le fossile est peu à peu libéré de sa coque de plâtre. Comme sur le terrain, les sur-faces osseuses nouvellement dégagées sont stabilisées au moyen de différentes colles et résines.• Une fois libérés de leur gangue, les fossiles sont nettoyés au moyen d’une sableuse. Cet appareil propulse sous pres-sion de la poudre minérale abrasive qui retire les derniers millimètres de sédiment collés aux ossements.• Un travail de reconstitution est ensuite nécessaire pour les pièces à recoller ou pour celles présentant des fissures. Un mastic mélangé à une pâte durcissante est alors appliqué aux endroits concernés.
J. ayer
Figure 18: Ossements de l’allosaure Big Al Two en cours de préparation. On distingue clairement une mâchoire supérieure et sa rangée de dents très bien conservées.

29
• L’os est finalement trempé quelques minutes dans une résine acrylique pour une dernière consolidation.• Une fois toutes les pièces du squelette nettoyées, le préparateur passe à la phase de montage qui est adaptée à chaque fos-sile. Les ossements trop fragiles ou d’une très grande importance scientifique seront
par exemple conservés dans les collections et seront remplacés par des moulages pour la présentation publique.Le squelette de l’allosaure Big Al Two sera tout de même présenté sous sa forme authentique durant l’exposition du muséum de Neuchâtel, qui s’ouvrira au printemps 1999. La fin du montage est
à la recHercHe Des Dinosaures
3e partie: interprétation GéoloGique Du site De Howe rancH
prévue pour novembre 1998. Les résultats intermédiaires sont d’ores et déjà spectacu-laires et laissent présager d’une présenta-tion finale impressionnante (fig. 18).
L’étude taphonomiqueLa taphonomie est une discipline de la
paléontologie qui tente de retracer sur un gisement les événements qui ont suivi la mort des organismes, notamment le trans-port, l’enfouissement et les processus de fossilisation. Couplée à des études sédi-mentologiques, elle permet d’une part de proposer une image reconstituée du site étudié et, d’autre part, d’envisager un scé-nario pour expliquer l’origine du gisement. Ces démarches s’appuient bien sûr sur des observations de terrain, mais également sur des analyses plus fines en laboratoire.
La taphonomie se concentre sur l’étude des ossements, de leur préservation et de la disposition des squelettes dans la car-rière prospectée. L’étude sédimentolo-gique analyse la nature des roches et la géométrie des strates sédimentaires. Elle s’appuie notamment sur des données car-tographiques détaillées. Elle a pour but de déterminer entre autre l’origine des sédiments et leurs conditions de dépôt. La composition minéralogique et le contenu fossilifère des roches apportent des infor-mations paléoécologiques.
Cette recherche est une véritable enquête policière où l’interprétation de tous les indices doit se faire pas à pas, sans céder à des conclusions trop hâtives.
Interprétations sédimentologiques du site de Howe Ranch
Ioannis michelis, paléontologue alle-mand, mène depuis quelques années des recherches dans la région de Howe Ranch, se consacrant à l’étude taphonomique du gisement de Howe quarry. Ce travail abou-tira en 1999 à une thèse de doctorat. Sa présence lors des fouilles en 1997 a permis de discuter du contexte sédimentologique du site. voici quelques-unes de ses obser-vations:
L’analyse des argiles a permis de distin-guer deux conditions de dépôt. L’origine des bentonites est rattachée généralement au dépôt éolien de particules d’origine volca-nique. A l’inverse, les argiles bigarrées ont été déposées par l’eau. Dans la Formation de morrison, ce dépôt a fait suite à des inondations. Des phénomènes d’altération ont modifié ensuite ces argiles: les teintes rouges attestent d’un drainage important responsable de l’oxydation du sédiment alors que les argiles verdâtres, mal drai-nées, ont été plutôt altérés par des circula-tions d’eau souterraine; enfin, la formation

30
de nodules calcaires, les caliche nodules, est un produit caractéristique d’altération sous des conditions climatiques arides, tout au moins saisonnières.
Dans la région de Howe Ranch, les grès ont pour origine le dépôt d’alluvions au sein d’un système de chenaux formé par l’écoulement d’une rivière de type braided river (“rivière à chenaux tressés”) (walker, 1984). Contrairement à la rivière à méandres, la braided river dépose ses sédiments sous la forme de dunes longitu-dinales entre lesquelles circulent le courant. Dans la région étudiée, la largeur de la rivière est estimée à au moins 50 mètres, avec des chenaux d’une profondeur de 3 à 6 mètres environ. De telles dimensions sont envisageables au vu de la taille des cadavres de dinosaures que la rivière a pu transporter à l’époque. Des indices de direction du courant, présents dans les niveaux gréseux, montrent un écoulement général du sud-ouest vers le nord-est.
Enfin, les rares niveaux calcaires ont pour origine l’évaporation à l’époque de points d’eau en période sèche.
Ces différents indices sédimento-logiques représentent des éléments pré-cieux pour l’interprétation des gisements de Howe quarry et de Howe Stephens quarry.
Howe quarry, il y a 147 millions d’annéesLa carrière de Howe quarry a déjà fait
l’objet d’une interprétation après les fouilles de 1934. L’image reconstituée du gisement au Jurassique supérieur montre un allosaure attaquant un troupeau de diplodocus (birD, 1985). Les observations faites par Hans-Jakob Siber et son équipe durant les fouilles de 1990 et de 1991 ont permis de proposer une nouvelle représentation. Ainsi, avec la participation de mark Hallett, célèbre illustrateur américain, une image de Howe quarry il y a 147 millions d’années a été réalisée (fig. 20). On y voit notamment un
troupeau de dinosaures sauropodes com-posé d’adultes et d’individus juvéniles venus se regrouper autour d’un ancien point d’eau, d’une largeur de quelques dizaines de mètres, dernier vestige d’une rivière asséchée par une longue période sans pluie. Les individus assoiffés constituent des proies faciles pour les allosaures, les grands prédateurs de l’époque. D’autres dinosaures affaiblis s’envasent. La végétation est repré-sentée par quelques fougères et de grands conifères, les araucarias.
Ce tableau est fortement inspiré des scènes animalières actuelles que l’on peut observer notamment dans la savane afri-caine. Il est en effet tentant de tirer des parallèles entre cette représentation et les nombreux films documentaires mettant en scène les stratégies d’attaque d’un groupe de lions, par exemple, sur un troupeau d’herbivores rassemblée autour d’un point d’eau. Cependant, même si l’image pro-posée comporte la touche personnelle de l’illustrateur, elle n’en repose pas moins sur une démarche scientifique rigoureuse dont les arguments sont les suivants:• Dans Howe quarry, la nature argileuse du niveau sédimentaire qui contient les ossements traduit des conditions hydro-dynamiques de très faible énergie qui ont accompagné le dépôt des fossiles. Un transport fluviatile est donc exclu. Cette conclusion est de plus renforcée par le fait que les sédiments de la carrière ne ren-ferment que peu ou pas du tout de débris végétaux.• La géométrie des couches dans la carrière laisse apparaître une sorte de cuvette. Cette paléotopographie serait le témoin du point d’eau décrit ci-dessus.• Bien qu’aucun squelette d’allosaure n’ait été trouvé sur ce gisement, la présence de ce grand carnivore est attestée d’une part par des empreintes trouvées aux côtés des ossements et d’autre part par la mise au jour de quelques dizaines de dents.
J. ayer

31
à la recHercHe Des Dinosaures
Figure 20: Représentation du paysage de Howe quarry, il y a 147 millions d’années. Réalisation artistique de mark Hallett.
Figure 19: Détail du plan de répartition des ossements trouvés dans Howe quarry, entre 1990 et 1991.

• La cartographie de tous les ossements montre une répartition des fossiles très hétérogène (fig. 19). De plus, aucun sque-lette complet n’a été trouvé. Il semble que l’activité prédatrice et occasionnellement charognarde des allosaures soit à l’origine de cette répartition désordonnée des osse-ments.• la préservation d’empreintes de peau implique des conditions de fossilisation particulières. D’une part, l’enfouissement des cadavres doit se faire rapidement et d’autre part, le sédiment doit être suffi-samment fin pour protéger les parties orga-niques de l’oxydation par l’air. De ce fait, l’envasement de certains individus peut être envisagé.
Parmi les questions non résolues, la dimension temporelle de la scène recons-tituée est encore très discutée. S’agit-il d’un événement dramatique ponctuel dans le temps, comme le laisse paraître la représentation de mark Hallett ? Ou faut-il plutôt imaginer une situation analogue qui s’est répétée dans le temps sur plu-sieurs décennies? Le travail de doctorat de Ioannis michelis amènera peut-être une réponse à cette question et permettra sans doute d’affiner l’interprétation du gise-ment de Howe quarry.
Première interprétation géologique de Howe Stephens quarry
Bien que distante de 450 mètres seule-ment de Howe quarry, la carrière de Howe Stephens quarry présente un contexte sédi-mentaire très différent. Les couches géo-logiques de la carrière sont de plus situées stratigraphiquement quelques mètres au-dessus des niveaux présents dans Howe quarry. L’interprétation géologique des deux gisements ne peut donc être en aucun cas similaire.
La carrière n’a pas fait encore l’objet d’une étude taphonomique détaillée. Cependant, des observations sédimentolo-giques faites durant les fouilles, une ana-
lyse de la disposition des squelettes dans la carrière et des relevés en laboratoire per-mettent de proposer les indices suivants:• Contrairement à Howe quarry, les sédi-ments sont clairement de nature gréseuse et les restes de bois et autres débris végé-taux y sont nombreux (fig. 4 c,D). Le dépôt des restes animaux et des sédiments s’est sans doute organisé dans le lit d’une rivière active.• Une analyse en laboratoire de la compo-sition des ossements a montré une relative-ment forte teneur en composés phosphatés, éléments originels de l’os. De plus, la pré-servation exceptionnelle de la plupart des squelettes, dont certains sont complets, implique un recouvrement très rapide des cadavres par les sédiments, les protégeant des agressions extérieures. • mis à part le diplodocus XL qui semble avoir une autre origine géologique, les neufs autres squelettes sont regroupés sur une surface de 10 m x 12 m seulement et sont contenus dans une couche d’un mètre d’épaisseur. Une telle imbrication semble indiquer un dépôt contemporain.• La présence de niveaux à galets mous (fig. 4 a) atteste d’une resédimentation consécutive à une inondation.• La disposition sur le terrain du squelette de l’allosaure Big Al Two, dont les ver-tèbres caudales étaient détachées du reste du squelette et reposaient sur l’avant de l’animal, tend à montrer que le cadavre a été transporté par le courant puis stoppé par un obstacle naturel. • Lors d’une campagne de fouilles menée sur le même gisement en août 1998, à laquelle Bernard Claude et moi-même avons participé partiellement, une décou-verte majeure a apporté un élément sup-plémentaire à cette enquête. L’obstacle naturel envisagé s’est soudain concrétisé sous la forme d’un gigantesque tronc silicifié (fig. 22). L’arbre, dont les bases des racines sont préservées, mesure plus d’un mètre de diamètre et sa longueur est
32
J. ayer

33
à la recHercHe Des Dinosaures
Figure 22: Tronc fossile silicifié mis au jour dans la carrière de Howe Stephens quarry en 1998. Cette découverte est d’une grande importance pour l’interprétation géologique du gisement.
Figure 21: Position et orientation du tronc silicifié reportées sur le plan des squelettes trouvées dans Howe Stephens quarry (fig. 13). E n n o i r : p a r t i e dégagée (7 m)En tirets: prolonga-tion estimée

34
J. ayer
estimée à plus de dix voire vingt mètres. La position et l’orientation de ce fossile specta-culaire semblent assez bien concorder avec la direction du courant envisagée du sud-ouest vers le nord-est (fig. 21).
Toutes ces observations permettent de proposer une première interprétation du gise-ment: La formidable concentration de squelettes articulés dans la carrière de Howe Ste-phens quarry semble être le résultat d’une crue dévastatrice. L’inondation aurait emporté les cadavres de dinosaures sur une certaine distance avant de les imbriquer un à un contre un gigantesque tronc faisant barrage au milieu de la rivière. L’énergie hydraulique devait être très importante pour transporter les très lourds cadavres de dinosaures. L’énorme quantité d’alluvions charriées par la rivière aurait ensuite recouvert rapidement les dépouilles.
A l’avenir, ce scénario devra être testé sur le terrain par des études sédimentologiques détaillées et des relevés cartographiques précis. même si cette première hypothèse est séduisante, de nombreux problèmes sont encore à résoudre, notamment la géométrie des couches au niveau du diplodocus XL qui présente un fort pendage indépendant de l’incli-naison générale des strates.
conclusion et reMercieMents
Les répercussions scientifiques de ces recherches au Wyoming sont multiples. Les fossiles, d’une préservation exceptionnelle, constituent sur le plan mondial un matériel paléontologique de grande valeur. De plus, l’étude paléoécologique de ce gisement ouvre une fenêtre nouvelle sur un monde disparu qui suscite depuis des décennies une fascina-tion sans pareille. A l’heure où la Dinomania fait encore rage avec son lot de spéculations et d’images erronées sur les dinosaures, la découverte et l’étude d’un site comme celui de Howe Ranch apportent aux chercheurs des éléments précieux pour leur permettre de pro-poser au public une vision de ces animaux qui soit plus proche de la réalité.
Outre l’intérêt scientifique, cette aventure merveilleuse vécue dans un groupe soli-daire restera gravée à tout jamais dans ma mémoire. A ce titre, toute ma reconnaissance s’adresse d’une part aux personnes qui ont permis la réalisation de ce projet et d’autre part, à toute l’équipe de fouilles, en particulier Bernard Claude et Hans-Jakob Siber, qui ont largement contribué à la réussite de cette aventure. Je tiens également à remercier Ioannis michelis pour la communication de ses observations et Jean-Paul Haenni, Chris-tophe Dufour et Johanne Blanchet, pour la relecture attentive du document.
biblioGrapHie
Références dans le textebenton, M. J. 1998. Atlas historique des dinosaures. Autrement, Paris. Trad. de l’anglais.
birD, r. t. 1985. Bones for Barnum Brown. Texas Christian University Press.
currie pH. J. & paDian, k. 1997. Encyclopedia of Dinosaurs. Academic Press, New-york.
pitcHer, D. 1993. wyoming Handbook. moon Publications, Inc. Chico, California, USA.

35
à la recHercHe Des Dinosaures
Figure 23: L’équipe de fouilles photographiée par H.-J. Siber dans Howe Stephens quarry: Du premier plan à l’arrière plan: Christoph Keilmann, maya Siber, Esther Wolfensberger, Bernard Claude, Jacques Ayer, Ioannis michelis, Denis maurer (à la caméra) et Ben Pabst.

walker, r. G. (éd.) 1984. Facies models. Geological Association of Canada Publications, Toronto.
lectures conseilléesbakker, r. t. 1986. The Dinosaur Heresies. Kensington Publishing Corp., New-york.
buffeteau, e. 1997. Dinosaures: A la recherche d’un monde perdu. L’archipel, Paris.
eHrlicH, G. 1996. La consolation des grands espaces. Albin michel, Paris. (Trad. de l’anglais)
Horner r. J. & GorMan J. 1988. Digging Dinosaurs. Harper & Row, Publishers, New-york.
norMan, D. 1991. La Grande Encyclopédie des dinosaures. Gallimard. (Trad. de l’anglais)
taquet, pH. 1997. L’empreinte des dinosaures. Ed. Odile Jacob. (2e éd.)
J. ayer
36 bull. soc. neucHâtel. sci. nat. 121 : 5-36. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 37-39. 1998
37
UNE PAGE RÉGIONALE D’HISTOIRE DES SCIENCES RE-LUE RÉCEmmENT...
I. CÉLESTIN NICOLET ET SES DÉCOUvERTES DE FAUNE mIOCÈNE à LA CHAUX-DE-FONDS
mARCEL S. JACqUAT
musée d’histoire naturelle, Av. Léopold-Robert 63, 2300 La Chaux-de-Fonds, Suisse.
mots-clés: La Chaux-de-Fonds, histoire des sciences, miocène, Lophiodon sp., Listriodon splendens
Key words: La Chaux-de-Fonds, history of sciences, miocen, Lophiodon sp., Listriodon splendens
Résumé:
Un regard moderne posé sur du matériel fossile recueilli en 1838 à La Chaux-de-Fonds permet de relire une page d’histoire des sciences du Jura neuchâtelois et de mettre en évidence l’intérêt d’une conservation à long terme.
introDuction
En octobre 1993, le musée d’histoire naturelle de La Chaux-de-Fonds accueillait le Dr Jan van der made, de l’Instituut voor Aardwetenschappen, Rijksuniversiteit, Utrecht, spé-cialiste de la paléontologie des suidés. Il venait étudier nos types de la collection Nicolet, trouvés en 1838 dans la molasse d’eau douce sous la Place Neuve (actuelle Place du marché) de La Chaux-de-Fonds, nous donnant ainsi l’occasion d’apprécier l’importance scientifique d’une toute petite partie des collections entreposées dans notre institution. A l’occasion de la 177e assemblée annuelle de l’Académie suisse des sciences naturelles en octobre 1997 à La Chaux-de-Fonds, nous avons sollicité du Dr van der made un témoi-gnage actuel de l’importance des découvertes de Célestin Nicolet. Il nous répondit par une contribution dont l’importance dépassait le cadre de la publication projetée1 et était mieux adaptée au Bulletin de la SNSN.
qui est Célestin Nicolet (1803-1871)? Pharmacien chaux-de-fonnier formé à Bâle, Besançon, Lausanne et Paris, il fut un des hommes les plus remarquables du XIXe dans le canton de Neuchâtel. Instigateur de la création de la Section des montagnes de la Société des sciences naturelles de Neuchâtel, fondateur du musée scolaire qui allait donner nais-sance au musée d’histoire naturelle et au musée d’histoire, créateur de la bibliothèque
* Les notes sont regroupées en page 7

38
de La Chaux-de-Fonds, protecteur du Dr Coullery, Célestin Nicolet prit aussi une part active à la Société d’histoire, dont il fut le premier président. Géologue, paléontologue, météorologue, historien, il fut aussi le botaniste des expéditions d’Agassiz, Desor, vogt et consorts au gla-cier de l’Aar, lors du fameux épisode de l’Hôtel des Neuchâtelois, dès 1840. On lui doit la première description et la pre-mière carte géologiques (en couleur!) de la vallée de La Chaux-de-Fonds, parue en 1839 2.
Nicolet avait découvert «en 1838, dans le terrain d’eau douce de La Chaux-de-Fonds, une incisive d’un grand mammi-fère. Elle ressemble à l’incisive externe de la girafe par un sillon et une échan-crure, qui paraissent la diviser en deux, et par la présence d’une saillie interne. Le bord interne est fortement usé; le grand lobe est externe, le petit lobe est interne; c’est par conséquent le contraire de ce qui s’observe dans l’incisive externe de la girafe. En comparant récemment cette dent avec les fossiles du Jardin des Plantes de Paris, m. Nicolet s’est assuré qu’elle cor-respond à l’incisive médiane supérieure du Lophiodon de Lartet, découvert en 1838, par Lartet, à Simorre, dans le département du Gers »3.
Lors d’une séance ultérieure, il fit à nouveau allusion à ses découvertes4: « m. Nicolet met sous les yeux de la Société des ossemens fossiles, provenant des marnes nymphéennes de la Chaux-de-Fonds, ainsi que les dessins qui les représentent et qui sont dus au beau crayon de m. Favre5. Ces fossiles appartiennent à deux espèces de mammifères communs à notre terrain lacustre et aux terrains tertiaires sub-pyré-néens du département du Gers; l’une est le dicrocère trapu de m. Lartet, l’autre est un Lophiodon découvert en 1838, par m. Lartet, à Simorre (Gers). Les osse-mens fossiles de notre bassin tertiaire, sont
dispersés çà et là dans les couches tour-beuses et marneuses, ou dans les galets tertiaires de l’étage supérieur du calcaire lacustre, qui occupe le centre de la vallée. Ces ossemens auront été probablement entraînés au fond de la vallée par les eaux alluviales. Les os longs et ceux du crâne sont plus ou moins altérés, broyés ou fracturés; les maxillaires manquent, les os courts, ceux du carpe et du tarse de plusieurs mammifères sont intacts. Les os du dépôt tourbeux sont teints d’une couleur noirâtre, ceux du dépôt marneux sont blanchâtres. Les dents sont peu ou pas altérées. Ces ossemens offrent de l’in-térêt sous le double rapport de la zoologie géographique, et de l’âge géologique des terrains tertiaires...
... Les molaires de notre Lophiodon vont en diminuant de longueur depuis la der-nière jusqu’à la première. Ce caractère appartient à tous les Lophiodons décrits par Cuvier. Des trois collines de l’arrière molaire inférieure, les deux premières sont bien séparées, transverses, tranchantes; une arête part de chacun des côtés externes des collines, et descend obliquement en dedans. Ces caractères appartiennent à l’espèce moyenne et à la grande espèce d’Issel, à l’espèce moyenne et à la très-petite d’Argenton, à la grande espèce de Buchsweiler. Les incisives ont une grande analogie avec celles du sanglier, les deux incisives médianes et supérieures sont très-larges, obliques, crénelées, divisées en deux lobes inégaux par un sillon. Le bord libre des incisives de la mâchoire inférieure est usé. La couronne des quatre incisives moyennes est quadrilatère, com-primée d’avant en arrière, large vers son bord libre, elle se rétrécit vers la racine et présente en avant de petits sillons longi-tudinaux, en arrière une arête saillante et des sillons longitudinaux. Les incisives médianes, sont plus petites et plus étroites que les suivantes qui sont larges; les côtés de celles-ci sont légèrement crénelés. Les deux incisives externes sont plus petites,
M. s. Jacquat

39bull. soc. neucHâtel. sci. nat. 121 : 37-39. 1998
Notes de renvoi dans le texte1 Célestin Nicolet (1803-1881), pharmacien, géologue, botaniste in: m. S. Jacquat, 1997: Des sciences dans les mont Jura, Editions de la Girafe, musée d’histoire naturelle, La Chaux-de-Fonds, pp. 82-832 Nicolet, A.C. 1839. Essai sur la constitution géologique de la vallée de La Chaux-de-Fonds, p. 1-26, avec une carte et des coupes en couleur. mémoires de la Société des sciences naturelles de Neuchâtel, tome 2. 3 cf. Bulletin de la Société des sciences naturelles de Neuchâtel, 1843-1844, tome 1: Section de La Chaux-de-Fonds, Séance du 2 décembre 1843, p. 34. 4 cf. Bulletin de la Société des sciences naturelles de Neuchâtel, 1843-1844, tome 1: Section de La Chaux-de-Fonds, Séance du 25 avril 1844, p. 124-126. 5 Il s’agit de Louis Favre (1822-1904), cofondateur du Club jurassien, animateur puis président d’honneur de la SNSN, romancier, naturaliste, préhistorien et mycologue de talent.6 voir à ce sujet: Stébler E., Note sur le Listriodon in Le Rameau de Sapin, novembre 1872, p. 41-44, et décembre 1872, p. 45-46
mais plus larges que les deux médianes; elles présentent en avant un sillon longitu-dinal, en arrière une arête saillante ».
On notera avec intérêt la référence aux travaux de Cuvier (1769-1832), naturaliste natif de montbéliard. Nicolet s’était par ailleurs adressé en 1846 au célèbre paléon-tologue allemand Hermann von meyer pour la détermination et la description de son matériel6 qui donna trois espèces nou-velles, dont le Listriodon splendens. Ainsi
les restes présumés de Lophiodon, sorte de tapir, proche des rhinocérotidés allaient-ils être attribués à un suidé d’une espèce nouvelle, dont le type est à La Chaux-de-Fonds !
Près de 160 ans après leur découverte, les reliques fossiles de ces suidés du mio-cène font à nouveau parler d’elles: d’abord par l’importante révision qu’en a faite Jan van der made en 1996, puis grâce à son article publié ci-après.
célestin nicolet et ses Découvertes De faune Miocène à la cHaux-De-fonDs
__________

40
Résumé de l’article de Jan van Der MaDe: Les ossements du premier Listriodon décrit.
Les listriodons sont des Suidae fossiles très caractéristiques du miocène inférieur et moyen. Ils sont apparus en Afrique et ont colonisé l’Europe, le nord de l’Asie et la Chine, il y a quelque 16,5 millions d’années. Une révision récente (van Der MaDe, 1996) reconnaît 5 genres et 25 espèces de listriodons. On peut estimer que leur poids allait de 8 kg (Lopholistriodon moruoroti) à plus de 3000 kg (Kubanochoerus mancharensis). Leur dentition suggère qu’ils se nourrissaient de feuilles plutôt que d’herbages. Plusieurs lignées de listriodons se sont éteintes rapidement suite à l’exten-sion des prairies.Les fossiles découverts à La Chaux-de-Fonds par nicolet constituent le matériel-type de Lis-triodon splendens, espèce-type du genre Listriodon, lui-même genre-type de la sous-famille des Listriodontinae. malgré leur importance, ces ossements ont été peu étudiés jusqu’ici.Les ossements du paratype sont illustrés ici pour la première fois (planches 1 et 2). Leurs dimen-sions sont données par van Der MaDe (1996, tab. 6-19).

bulletin De la société neucHâteloise Des sciences naturelles 121 : 40-44. 1998
41
II. THE FIRST DESCRIBED LISTRIODONT REmAINS
JAN vAN DER mADE
CSIC, museo Nacional de Ciencias Naturales, c. José Gutiérrez Abascal 2, 28006 madrid, España.
Listriodonts (Listriodontinae, Suidae) are a very characteristic element in Early and middle miocene faunas of the Old World. Between roughly 20 and 10 million years ago (ma), they evolved in Africa and the Indian subcontinent and spread some 16.5 ma to Europe, northern Asia and China (van Der MaDe, 1996).
nicolet (1844a) was the first to publish on a listriodont. He described a first upper incisor (Pl. 1, fig. 2), that he found in 1838 in La Chaux-de-Fonds. He mentioned the tooth in a discussion on fossil giraffes and noted that the grove that divides the tooth in two lobes resembles the “incisive externe” (lower canine) of giraffes, which has a similar grove. Later, vacek (1900) tentatively assigned a listriodont incisor to the giraffe Hel-ladotherium. However, Nicolet believed the incisor to belong to “Lophiodon” found by Lartet in Simorre (southern France), according to Nicolet, also in the year 1838. A skull from Simorre (figured by blainville, 1847) shows the association of “giraffe-like” inci-sors and tapir-like molars.
Nicolet (1844b) gave a further description of the material. He considered the material as Lophiodon, a Palaeogene tapir, apparently because of the lophodont structure of the molars. Lartet and also blainville (1847) considered the animal from Simorre a tapir and lyDekker (1868) described the first listriodont from the Indian subcontinent as Tapirus Pentapotamiae. nicolet (1844b) noted the resemblance of the incisors to those of a wild boar, which is surprising, since their resemblance to those of tapirs lead lein-Ders (1977) to interpret the Listriodon dietary adaptation as similar to those of tapirs. In particular the lower incisors (Pl. 1, figs. 4-5) are morphologically much closer to those of the tapirs than those of the living suids.
von Meyer (1846) became convinced that the animal from La Chaux-de-Fonds (and from Simorre) was indeed a pig and no tapir and that it had nothing to do with Lophiodon; he introduced the name Listriodon splendens for these fossils. In the same paper, he also introduced the names Calydonius trux and Calydonius tener (Pl. 1, fig. 1) for the suid canines from La Chaux-de-Fonds.
A recent revision of the listriodonts (van Der MaDe, 1996) recognizes 5 listriodont genera and some 25 species. Wide incisors with low crowns are typical for all of them and in nearly all lineages, still wider incisors evolved parallel. Lophodont teeth, as in tapirs, evolved at least twice. Incisor morphology, enamel thickness, lophodonty and tooth wear suggest that listriodonts were folivores. They may have ingested large quanti-ties of leaves and may have had a “grazing” type of feeding. However, their low crowned teeth suggest that they did not eat grass, which is very abrasive and which usually is eaten by species with high-crowned cheek teeth. Different listriodont lineages in Africa,
42
J. van Der MaDe
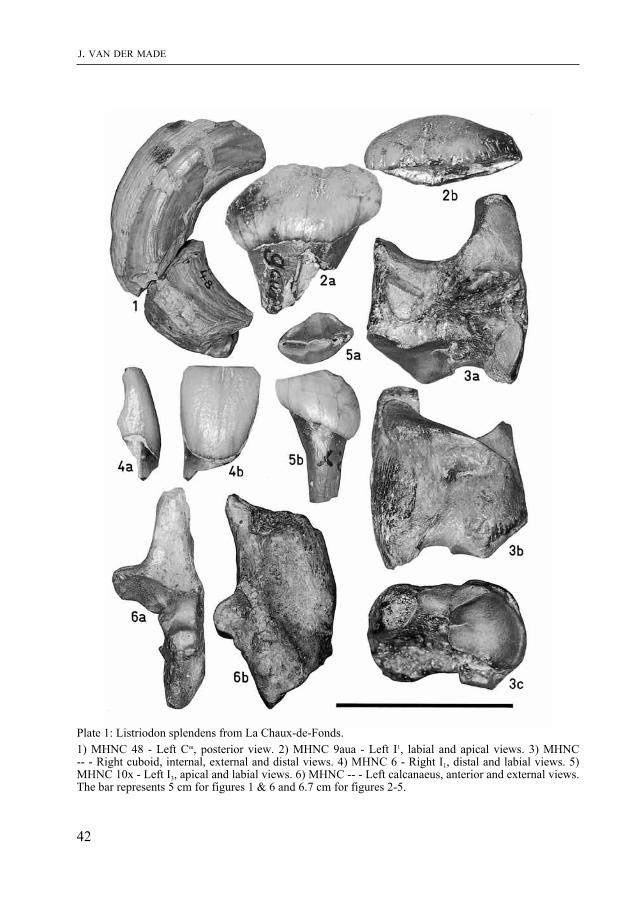
Plate 1: Listriodon splendens from La Chaux-de-Fonds.1) mHNC 48 - Left Cm, posterior view. 2) mHNC 9aua - Left I1, labial and apical views. 3) mHNC -- - Right cuboid, internal, external and distal views. 4) mHNC 6 - Right I1, distal and labial views. 5) mHNC 10x - Left I3, apical and labial views. 6) mHNC -- - Left calcanaeus, anterior and external views. The bar represents 5 cm for figures 1 & 6 and 6.7 cm for figures 2-5.
43
tHe first DescribeD listrioDont reMains
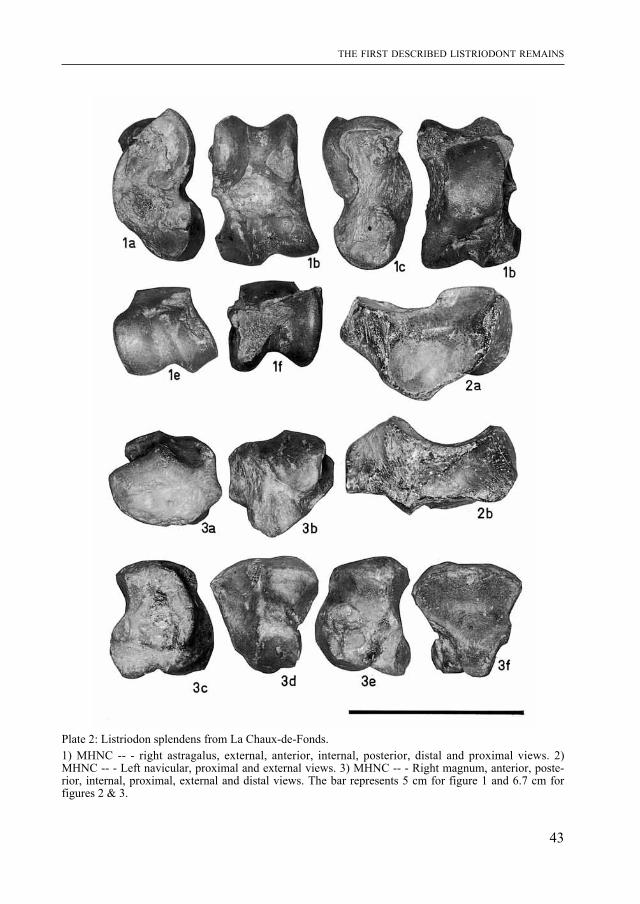
Plate 2: Listriodon splendens from La Chaux-de-Fonds.1) mHNC -- - right astragalus, external, anterior, internal, posterior, distal and proximal views. 2) mHNC -- - Left navicular, proximal and external views. 3) mHNC -- - Right magnum, anterior, poste-rior, internal, proximal, external and distal views. The bar represents 5 cm for figure 1 and 6.7 cm for figures 2 & 3.

44
Europe, China and the Indian Subcontinent disappeared in a short period following the spread of open grasslands.
Estimated body weights of the listrio-donts vary between 8 kg for Lopholis-triodon moruoroti and over 3000 kg for Kubanochoerus mancharensis. Kubano-choerus is a peculiar listriodont, in which the females has small protuberances above the orbits and the males, in addition to these, a large “horn” in the middle of the frontals. The “horn” was directed foreward and was probably not covered by keratin.
The increasing width of the incisors and increasing size of the canines in the diffe-rent lineages have stratigraphic value. The incisors of L. splendens suggest a position for La Chaux-de-Fonds between the older sample from La Grive and younger samples from Sant quirze and Castell de Barberà.
Though the fossils collected by Nicolet were the first described listriodont remains and constitute the type material of Lis-triodon splendens, type species of Lis-triodon, type genus of the Listriodontinae, the material was not often object of study. There are descriptions by bayle (1856) and steHlin (1899/1990). Stehlin and I figured some of the dental material (ste-
Hlin, 1899/1900; van Der MaDe, 1996, Pl. 41, figs. 1-14). None of the paratype bones have ever been figured, even though the listriodont ankle joint morphology has been interpreted as representing a particular cursorial specialisation (leinDers, 1976). Figures of the paratype astragalus (Pl. 2, fig. 1), cuboid (Pl. 1, fig. 3), navicular (Pl. 2, fig. 2) and calcanaeus (Pl. 1, fig. 6) are provided here. Also the magnum may turn out to have a phylogenetically impor-tant character. The anterior end of that bone may touch the unciform (as in the more derived suids) or not (as in a hyotheriine like Chleuastochoerus Pearson, 1928). In Listriodon, the magnum has a facet for the unciform that reaches the anterior end of the bone (Pl. 2, fig. 3). measurements of the specimens are given elsewhere (van Der MaDe, 1996, tables 6-19).
acknowleDGeMents
I thank marcel S. Jacquat for access to the material and for his hospitality during my stay in La Chaux-de-Fonds.
references
J. van Der MaDe
MaDe, J. van Der 1996. Listriodontinae (Suidae, mammalia), their evolution, systematics and distri-bution in time and space. Contributions to Tertiary and quaternary Geology, 33(1-4): 3-254 & 14 tables on a microfiche.
Nicolet, C. 1844 a. Sur une dent fossile de Lophiodon. Bull. Soc. neuchâtel. Sci. nat. 1843-1844: 34.
nicolet, C. 1844 b. Sur les ossements fossiles des marnes nymphéennes de la Chaux-de-Fonds. Bull. Soc. neuchâtel. Sci. nat. 1843-1844: 124-126.
more references in van Der MaDe, 1996.
__________
bull. soc. neucHâtel. sci. nat. 121 : 40-44. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 45-52. 1998
45
LES OS D’ÉLANS ALCES ALCES (L. 1758) HOLOCÈNES DU GOUFFRE 1 DES GRANDS BOIS
(JURA NEUCHÂTELOIS, LES vERRIÈRES NE)NOUvELLE DATATION RADIOCARBONE ET BRÈvE mISE AU POINT
DES CONNAISSANCES ACTUELLES.
PHILIPPE mOREL
Chemin de maujobia 8, CH-2000 Neuchâtel, Suisse.
Résumé
Les crâne ou fragments crâniens de trois élans holocènes mâles munis de leurs bois ont été décou-verts dans les années 1860 dans un gouffre du Jura neuchâtelois. Ces pièces, dispersées, ont été revues récemment et l’une d’entre elles a été datée à environ 2500 ans BP par le radiocarbone AmS. Deux ramures, à empaumure bien développée, ont appartenu à des adultes de forte taille et la troisième à un jeune, qui devait porter ses premiers bois. La présence d’élans en montagne, à des altitudes dépassant souvent 1000 m, est discutée et expliquée par l’écologie de cette espèce, qui se nourrit surtout de feuillage, et qui pouvait y trouver en été une nourriture de haute valeur nutritive.
Zusammenfassung
Die holozänen Elchknochen, Alces alces (L. 1758) aus dem Gouffre 1 des Grands Bois (neuenburger Jura, Les verrières NE): neue Radiokarbon-Datierung und Stand der heutigen Kenntnisse.
Ein Schädel und zwei Schädelteile von drei holozänen Elchen, alle mit Geweih, wurden in den Jahren 1860 in einer Schachthöhle im neuenburger Jura entdeckt. Diese Funde, die in verschie-denen museen verteilt sind, wurden neulich neu untersucht und einer davon mittels AmS-Radio-karbon auf 2500 Jahre BP datiert. Zwei der Geweihe, mit gut entwickelter Schaufel, gehörten stattlichen Individuen. Das dritte stammt von einem jungen Tier, möglicherweise einem Spiesser. Die Anwesenheit vom Elch in Gebirgslandschaften über 1000 m ü.m., wird dadurch erklärt, dass dieser Laubfresser dort bis spät im Sommer hochwertige Nahrung finden konnte.
Summary
The holocene remains of elk Alces alces (L. 1758) of the Gouffre 1 des Grands Bois (Swiss Jura, Les verrières, canton Neuchâtel): new radiocarbon dating and state of knowledge.
Skulls or skull fragments, with antlers, of three Holocene elk were discovered in the 1860s in a cave in the Swiss Jura (canton Neuchâtel). These finds, housed in various museums, were reana-lyzed recently. One has been AmS-radiocarbon dated to ca. 2500 BP. Two of the antlers with well developed palms belonged to robust individuals. The third comes from a younger animal that died with its first set of antlers. The presence of elk, a browser, in mountainous landscapes, often at

elevations of about 1000 m, can be explained by the presence of highly nutritional vegetation until late in the summer.
introDuction
Depuis les premières recherches archéo-logiques effectuées entre autres dans les sites littoraux des lacs jurassiens, on sait que depuis le retrait des derniers glaciers pléistocènes, l’élan a peuplé une bonne partie du territoire de la Suisse. La pré-sence de ce cervidé dans les montagnes du Jura était également établie à la suite de diverses découvertes, dont la plupart ont été faites dans des grottes ou des gouffres. Du point de vue de l’historique des recherches, les vestiges présentés ici font partie des découvertes d’ossements d’élan les plus anciennes du Jura, et sans doute de Suisse. Depuis plusieurs décen-nies, ces pièces, mentionnées par coulon en 1865 et DelacHaux en 1868, étaient considérées par l’auteur comme perdues, car aucune mention autre que celles faisant référence à ces premières publications n’a jamais été publiée à leur propos. En fait, ces ramures faisaient partie des collections du musée de Fleurier, petit musée d’his-toire naturelle aujourd’hui supprimé, et se trouvent aujourd’hui dans les collections du muséum d’histoire naturelle de Neu-châtel et de la maison des mascarons, à môtiers. Cette “redécouverte” a donné lieu à une datation radiocarbone de l’un de ces splendides trophées, publiée ici avec une petite mise au point sur ce qu’on connaît actuellement de ces trouvailles et de ce site.
Matériel
Dans son article, DelacHaux (1868) mentionne la découverte de trois “têtes” d’élans. Il s’agit en fait bien de trois pièces, mais leur état de conservation est variable. La première pièce (n° 1) est un fragment de crâne, dont les deux os frontaux sont
conservés, de même que la partie sagittale des pariétaux et l’amorce de l’occipital. Ses deux bois sont présents, comportant chacun sept andouillers, dont cinq sont brisés à plus ou moins grande distance de leur pointe (fig. 1). La deuxième pièce est un crâne presque entier, dont seuls les pré-maxillaires sont brisés. Aux deux rangées dentaires manquent la deuxième et troi-sième prémolaire. Toutes les jugales sont fortement usées, et la couronne de la pre-mière molaire est presque complètement abrasée, indiquant un âge avancé, certaine-ment de plus de dix ans (Heptner & nasi-MowitscH, 1974). Les deux bois sont pré-sents, mais celui de droite est partiellement détruit au niveau de l’empaumure. Le bois gauche possède sept andouillers, deux seu-lement subsistent sur le droit. La troisième pièce est un fragment de frontal gauche d’un individu jeune, muni d’un petit bois comportant deux andouillers, vraisembla-blement un daguet de première tête.
Les trois pièces sont bien conservées, de couleur beige clair. Les quelques cassures observables sont nettes, et peuvent avoir été produites lors de la chute des animaux ou lors du séjour des os dans les blocs des éboulis du fond du gouffre. Une seule du crâne n° 2, émoussée, semble avoir eu lieu du vivant de l’animal. L’empaumure du bois droit de la pièce n° 2 a été détruite par altération physico-chimique, vraisem-blablement dissoute par l’eau de percola-tion. Plusieurs fissures de dessiccation (qui peuvent cependant être anciennes) sont visibles sur les bois des adultes. quelques réparations ont été effectuées, en parti-culier sur la pièce n° 1, dont le bois droit a été recollé à son pédicule et une cas-sure, qui parcourt le frontal gauche, loca-lement consolidée. Des restes de plâtre sont visibles sur la cassure du bois droit du crâne n° 2, témoignant d’une tentative de reconstitution. Ces trois objets n’ont certainement jamais été traités avec un produit imprégnant pour les consolider (les pièces n° 2 et 3 n’ont même jamais été net-
46
pH. Morel
47
toyées et portent encore des restes de sédi-ments de la grotte). En tout cas, leur sur-face correspond bien à celle d’ossements issus de contextes comparables; les bois sont relativement poreux et aucune trace de vernis ou de paraffine n’est visible.
Les ramures des deux adultes sont de forte taille, et leurs empaumures sont bien formées (type palmata de nyGrén, 1986). La première comporte sept andouillers de chaque côté, et son envergure recons-tituée devait atteindre au moins 1,20 m (fig. 1). La deuxième comporte égale-ment sept andouillers sur le bois gauche – le droit, endommagé, ne permet pas de comptage. Son envergure peut être estimée à au moins 1,30 m. On peut noter que le contour des os nasaux, intacts, appar-tiennent au plus fréquent des quatre types
observés en Europe (type A de nyGrén, 1986, p. 185).
La partie basilaire des bois des pièces 1 et 3 est compacte et ne montre aucun signe de dissolution. Ces deux animaux ont donc trouvé la mort entre septembre et novembre pour l’adulte et entre septembre et février ou même mars pour le jeune (nyGrén, 1986). Par contre, un début de séparation des bois au sommet des pédi-cules est bien visible du côté ventral sur le crâne n° 2, indiquant un décès de l’animal peu avant la mue, donc entre octobre et février, mais plutôt en octobre puisque ses dents fortement usées indiquent un âge avancé (la période de mue se décale en effet avec le vieillissement de l’animal, nyGrén, 1986).
les os D’élan Holocènes Du Gouffre 1 Des GranDs bois: nouvelle Datation
Figure 1: Ramure d’élan du Gouffre 1 des Grands Bois, Les verrières NE, découverte en 1862 (pièce n°1). Echelle: 10 cm. Photo Ph. morel.

Le dessin publié par DelacHaux (1868), qui figure un crâne entier muni de ses deux bois complets, malgré une ressem-blance toute relative, correspond sans doute à la pièce n° 2. La manière dont le crâne est représenté, avec des prémaxil-laires surmontés d’une masse difforme, des os nasaux plutôt vagues, et sa mandi-bule entière munie de toutes ses incisives, et ses deux bois complets, montre que la pièce a visiblement été complétée, peut-être en s’inspirant de la pièce n°1 et de la mandibule censée avoir été trouvée dans ce gouffre. Il est en tout cas certain que la partie antérieure du crâne a été détruite et réduite à son état actuel bien avant sa découverte (et non après la réalisation du dessin). Les bois, quant à eux, sont bien ceux d’un animal de taille exceptionnelle, munis de sept andouillers à gauche, et cor-respondent sans aucun doute à ceux du crâne n° 2.
Le frontal de la pièce n° 1 porte une étiquette collée, portant la mention manus-crite “n°831/musée de Fleurier”. La pièce n°3 porte une étiquette identique. La pièce n°1 appartient au muséum d’histoire natu-relle de Neuchâtel, où elle est inventoriée sous le numéro 94.2604. Les deux autres pièces font partie de la collection de la maison des mascarons de môtiers. Une carte d’inventaire du musée de Fleurier (conservée à la maison des mascarons) a été retrouvée. Elle désigne une “Tête d’élan”, de dimension d’ “env. 1 m.”, et la rubrique provenance porte la mention: “Trouvée en 1862 dans une baume près du mt des verrières”. A la rubrique remarque, on lit: “C’est l’emblème de la société du musée”. Cette carte correspond sans doute à la pièce n° 2, puisque l’emblème cité comporte un crâne. Diverses recherches effectuées par l’auteur dans les musées de la région comportant des collections de sciences naturelles (musée de la Bande-rette à Travers, muséum d’histoire natu-relle de Neuchâtel, musée de l’Areuse de Boudry, musée d’histoire naturelle
de La Chaux-de-Fonds) laissent à penser qu’aucun autre os d’élan n’a été récolté à la même époque que les pièces mention-nées, et que la mandibule que DelacHaux (1868) dit avoir découverte a définitive-ment disparu.
lieu et circonstances De Découverte
Dans son article, DelacHaux (1868) dit que «La baume des Elans est formée d’abord d’une longue cheminée qui a une cinquantaine de pieds de profondeur au fond de laquelle on descend à l’aide d’une corde. On arrive sur un tas de neige qui ne fond jamais complètement durant toute l’année. » Cet auteur situe cette baume (qu’il appelle aussi Baume du mont des verrières) sur le mont des verrières, un haut plateau situé entre Les verrières et La Côte-aux-Fées. Dans son inventaire spéléologique du canton de Neuchâtel, GiGon (1976) considère, sans doute à juste titre, que cette baume est identique avec le Gouffre 1 des Grands Bois, appelé aussi Baume des élans (n°62.12, p. 202, à ne pas confondre avec la Baume de l’élan, n°53.2, p. 158). Cette cavité est formée de deux puits communiquants, dont le plus profond mesure 28 m. L’autre, comblé par de gros éboulis, n’atteint actuellement que quelques mètres. Cette profondeur ne correspond guère à la “cinquantaine de pieds” de DelacHaux, mais on peut envi-sager l’existence d’un névé important au fond du gouffre, ou simplement que l’esti-mation était fausse. En tout cas, une iden-tité avec la Baume à Jean du Lac (GiGon, 1976, 62.15, p. 203), et les gouffres 2 et 3 des Grands Bois (GiGon, 1976, 62.13 et 62.14, p. 202 et 203), petits puits voisins de quelques mètres de profondeur, dont la description ne correspond pas du tout et où aucun reste d’élan n’a été découvert, est sans doute à exclure.
Le mont des verrières est actuellement couvert d’une forêt de sapins entrecoupée
48
pH. Morel

de pâturages clairsemés d’arbres. Il s’agit d’une région fortement karstifiée, com-portant de grandes dolines humides, et plusieurs gouffres verticaux. On peut voir çà et là des zones de lapiaz partiellement dénudés.
Pendant des décennies, sans doute des siècles, le gouffre 1 des Grands Bois, comme tous les autres gouffres du mont des verrières, a servi de charnier, dans lequel des générations de paysans des environs se sont débarrassés de carcasses d’animaux non consommables. Pour cette raison, le bas du puits est jonché d’un énorme amas d’ossements et d’ordures diverses. C’est à cet endroit que trois Fleu-risans, mm. Petitpierre, Pillet et Lavaur ont découvert trois “têtes” d’élans, appa-remment en 1862 (selon la fiche du musée de Fleurier). GiGon (1976), mentionne deux dates contradictoires, août 1863, puis 1865. coulon (1865) ne parle que de “deux têtes d’élans pourvues de leurs bois”. L. Delachaux visita également le gouffre en 1867, et y découvrit une man-dibule qu’il attribua à l’un de ces crânes (DelacHaux, 1868; GiGon, 1976). J.-P. Jéquier y fit quelques recherches dans les années 1960 et y découvrit également quelques os de cette espèce (GiGon, 1976). Il est possible que les pièces sans indica-tion de provenance de la collection de ce chercheur, libellées n°52 dans l’inventaire de cHaix & Desse (1981) proviennent de cette cavité, mais elles ne doivent pas être confondues avec celle de la Baume de l’élan, synonyme du gouffre II de la Char-bonnière (Boveresse, NE), où deux sque-lettes d’élans furent découverts (Jéquier, 1963). L’auteur fit trois visites au gouffre 1 des Grands Bois en 1980, qui permirent la découverte d’os de cerf élaphe (Cervus elaphus), mais pas d’élan.
Datation
Une datation par le radiocarbone AmS a été effectuée en juillet 1998 par G. Bonani,
à l’Institut de physique des particules de l’Ecole polytechnique fédérale de Zurich, à partir d’un fragment scié par l’auteur dans le pariétal gauche de la pièce n°1. La datation a donné le résultat suivant:
ETH-18618: 2515±65 années BPAge calibré: 798-474 av. J.-C. (92,8%),
calculé avec le logiciel CalibETH (niklaus et al., 1992) avec un intervalle de confiance de deux écarts-types. L’état général de cette pièce et le bon état de conservation de l’os prélevé (qui était compact et très dense) permettent de considérer cette datation comme fiable.
L’âge calibré (corrigé) permet donc de dire que cet animal a probablement vécu pendant le premier âge du Fer, l’époque de Hallstatt. Cette date n’a en soit rien d’éton-nant, puisqu’on sait que l’élan était encore chassé en Suisse au moins à l’époque romaine (voir ci-dessous).
Discussion
Les trouvailles d’élans dans des gouffres naturels sont relativement fréquentes; on en connaît de nombreuses dans le Jura, et quelques-unes dans l’Arc alpin, celles-ci en grande partie inédites (revillioD, 1953; Jéquier, 1963; koby, 1967; weiD-Mann, 1974; cHaix, 1975; cHaix & Desse, 1981; Morel, 1986; 1992; 1993). Si on peut envisager que les élans se coincent facilement dans des fissures en paysage de lapiaz, dans lesquelles on les retrouve souvent, il est difficile de com-prendre pourquoi ils se font piéger dans des gouffres à orifice de grandes dimen-sions, comme celui des Grands-Bois 1. Dans ce contexte, on peut envisager que les animaux piégés sont tombés alors qu’ils étaient poursuivis, par exemple par des loups.
La présence de l’élan sur des hauts plateaux, au relief parfois fort accidenté, pourrait à première vue surprendre, car les longues pattes de cet animal en font un habitant des marécages et des paysages
49
les os D’élan Holocènes Du Gouffre 1 Des GranDs bois: nouvelle Datation

plats, où il se sent d’ailleurs probablement le plus à l’aise. On sait cependant que les élans actuels d’Eurasie peuvent vivre, en été, jusqu’à très haute altitude dans les montagnes, par exemple jusqu’à 2500 m dans l’Altaï (Heptner & nasiMowitscH, 1974; nyGrén, 1986; voir aussi Morel, 1992). Leur seule exigence semble être une couverture d’arbres et d’arbustes suffi-sante pour assurer leur subsistance, consti-tuée essentiellement de feuilles. Leur lèvre supérieure et l’ensemble de leur système digestif, de même que leur cou très court, font en effet des élans des herbivores spé-cialisés dans la nourriture foliée (nyGrén, 1986). vu leur préférence pour les par-ties les plus nourrissantes des arbres et arbustes, il est possible qu’ils effectuaient des migrations saisonnières vers les hau-teurs, suivant en été le développement des jeunes pousses, décalé progressivement avec l’altitude. On peut noter de plus que les hauts plateaux sur lesquels ces restes d’élans ont été retrouvés, s’ils ne sont pas toujours d’accès aisé, sont souvent relativement humides, et parfois même proches de zones marécageuses situées sur des couches géologiques imperméables. Il est difficile de dire si les élans pouvaient subsister à haute altitude pendant toute l’année. Si cela paraît envisageable dans le Jura, à des altitudes avoisinant habituelle-ment 1000 m, ce n’est probablement pas le cas pour les sites des Préalpes, situés entre 1500 et plus de 1800 m.
La présence de l’élan est attestée en Suisse ou en France voisine à partir de la fin du Pléistocène (Desbrosses & prat, 1974), mais il a commencé à recoloniser le Jura et le Plateau suisse après la der-nière glaciation, dès que la végétation a permis son retour, c’est-à-dire lorsque les arbres ont été assez nombreux pour lui permettre de trouver sa subsistance. Il est attesté, mais apparemment jamais fréquent, pendant tout l’Holocène, en par-ticulier dans les stations littorales du Néo-lithique et de l’âge du Bronze, mais aussi
dans les grandes vallées alpines et dans le Jura (bäcHler, 1911; cHaix & Desse, 1981; speck, 1987). On relève aussi sa présence dans de nombreux sites d’Al-lemagne et d’Autriche (willMs, 1987). Les exemplaires datés les plus récents de Suisse remontent au Bas-Empire ou au haut moyen Age et ont été trouvés à Hüttenböschen (mollis GL, gallo-romain; scHMiD, 1963; 1966) et dans le lapiaz des Innerbergli, Habkern BE (date radiocar-bone AmS inédite, ETH-17209: 1535 ± 55 années BP, CAL 424-633 AD). Il est vraisemblable que sa disparition en Suisse, consécutive à un retrait progressif vers le nord-est, remonte à la fin de l’époque romaine ou au début du moyen Age, soit entre les ve et vIIe siècle de notre ère. Comme le suppose willMs (1987), ce recul progressif vers le nord-est pourrait être dû à la présence de plus en plus mar-quée des établissements humains et des cultures, et à une réduction progressive de son habitat. On peut aussi supposer que la chasse a pu jouer un rôle dans cette dispa-rition, comme elle fut au moins en partie responsable de celle du chevreuil dans le Jura à la fin du moyen Age, et peut-être aussi du chamois à une époque indéter-minée au cours de l’Holocène (salzMann, 1975).
Si les découvertes de vestiges d’élans sont relativement fréquentes dans les gouffres du Jura, les pièces du gouffre 1 des Grands Bois, sans doute parmi les premières découvertes de cette espèce en Suisse, sont de taille exceptionnelle et ne méritaient pas l’oubli dans lequel elles étaient tombées depuis plus d’un siècle. Souhaitons que ce petit bilan donnera lieu à une publication plus détaillée, et en par-ticulier à une datation des autres ossements d’élans découverts dans le canton de Neu-châtel, et que les futures recherches des spéléologues permettront d’augmenter encore nos connaissances sur les modalités
50
pH. Morel

51
de la présence et sur le comportement de l’élan holocène dans les montagnes du Jura et des Alpes.
reMercieMents
les os D’élan Holocènes Du Gouffre 1 Des GranDs bois: nouvelle Datation
Je tiens à remercier C. Dufour, directeur du muséum d’histoire naturelle de Neuchâtel d’avoir bien voulu mettre la pièce présentée à ma disposition. m. Egloff, archéologue cantonal de Neuchâtel, a assuré le financement de la datation, qu’il en soit remercié chaleureusement. L’aide de L. vaucher, maison des mascarons de môtiers, J.-D. Blant, musée d’histoire naturelle de la Chaux-de-Fonds, et de m. Diana, musée de la Banderette du Club Jurassien, Travers, a été des plus précieuse dans les diverses recherches effec-tuées autour de ce matériel; que ces personnes soient assurées de toute ma reconnais-sance.
biblioGrapHie
bäcHler, E. 1911. Der Elch und fossile Elchfunde aus der Ostschweiz. Jahrbuch der St.Gallischen naturwissenschaftlichen Gesellschaft 1910: 72-181.
cHaix, L. 1975. A propos de l’Elan (Alces alces L.) en Suisse. Bolletino del Centro Camuno di studi preistorici 12: 12-13.
cHaix , L. & Desse, J. 1981. Contribution à la connaissance de l’Elan (Alces alces L.) postglaciaire du Jura et du Plateau suisse. Corpus de mesures. quartär 31/32: 139-190.
coulon, L. 1865. Têtes d’Elan de la grotte des verrières. Bull. Soc. neuchâtel. Sci. nat. 6: 263-264.
DelacHaux, L. 1868. La Baume des Elans. Le Rameau de sapin 2: 42-43.
Desbrosse, R. & prat, F. 1974. L’Elan magdalénien de Pierre-Châtel (Ain). quartär 25: 143-157.
GiGon, R. 1976. Inventaire spéléologique de la Suisse I. Canton de Neuchâtel. Comm. Spéléo. Soc. helvét. Sc. nat. Neuchâtel.
Heptner & nasiMowitscH, A. A. 1974. Der Elch. Neue Brehm-Bücherei, Wittenberg Lutherstadt.
Jéquier, J.-P. 1963. Les deux squelettes d’Elan du gouffre II de la Charbonnière (Boveresse - Jura neu-châtelois). Cavernes 8: 70-71.
koby, F.-E. 1967. vestiges préhistoriques d’élan et de castor dans le nord du Jura. Actes Soc. juras. émul. 78: 169-179.

52
Morel, P. 1986. Knochenfunde im P55 des “Réseau des Lagopèdes”, Schrattenfluh, Flühli, LU. Reflektor 7: 21-29.
--- 1992. Les Elans (Alces alces L.) holocènes en Suisse: quelques remarques à la lumière de décou-vertes spéléologiques récentes. Stalactite Supplément N° 13: 25-28.
--- 1993. Les ossements holocènes du gouffre des Bois de Châtelat, commune de Châtelat (BE). Le Jura souterrain 1993: 16-23.
nyGrén, K. 1986. Alces alces (Linnaeus, 1758) - Elch. In: nietHaMMer, J. & krapp, f. (eds.): Han-dbuch der Säugetiere Europas. Band 2/II: Paarhufer - Artiodactyla (Suidae, Cervidae, Bovidae), 173-197. Aula, Wiesbaden.
revilloD, P. 1953. Les élans et les ours dans le Jura des environs de Genève. Découvertes de nos spé-léologues. musées de Genève 1953.
salzMann, H. C. 1975. Die Geschichte der Gemsen im schweizerischen Jura. mitteilungen der natur-forschenden Gesellschaft in Bern N.F. 32: 15-35.
scHMiD, E. 1963. Die Tierknochen aus Hüttenböschen. Jahrbuch des Historischen vereins des Kan-tons Glarus 60: 17-19.
--- 1966. Ergebnisse der Ausgrabung 1964 beim gallorömischen Tempel auf Hüttenböschen (mollis). Jahrbuch des Historischen vereins des Kantons Glarus 61: 35-41.
speck, J. 1987. Spätglaziale und frühpostglaziale Überreste von Grosssäugetieren im Bereich des Reussgletschers. mitteilungen der naturforschenden Gesellschaft Luzern 29: 291-314.
weiDMann, m. 1974. Sur quelques gisements de vertébrés dans le quaternaire du canton de vaud. Bulletin de la Société vaudoise des sciences naturelles 72: 9-18.
willMs, C. 1987. Der Elch im nacheiszeitlichen Europa. Archeologia Polski 32 (2): 249-291.
__________
pH. Morel
bull. soc. neucHâtel. sci. nat. 121 : 45-52. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 53-60. 1998
53
THE GENUS PANDANUS (PANDANACEAE)IN mADAGASCAR (PART 2)
KIm-LANG HUyNH
Phanerogamy Division, Botanical Institute, University of Neuchâtel. P.O. Box 2, CH-2007 Neuchâtel, Switzerland.
mots-clés: Afrique, madagascar, Pandanus sect. Foullioya, Pandanacées, Taxonomie
Key-Words: Africa, madagascar, Pandanus sect. Foullioya, Pandanaceae, Taxonomy
Résumé
Deux espèces nouvelles de Pandanus de madagascar appartenant à la sect. Foullioya (P. barbel-latus Huynh, P. sylvicola Huynh), et une sous-section nouvelle de cette section (subsect. Barbellati Huynh), sont décrites. Une clé des espèces actuellement connues de la section est proposée.
Summary
Two new species of Pandanus from madagascar of sect. Foullioya (P. barbellatus Huynh, P. syl-vicola Huynh), and a new subsection of this section (subsect. Barbellati Huynh), are described. A key to the species of the section known at present is tentatively proposed.
Zusammenfassung
Zwei neue Pandanus-Arten aus madagaskar von Sektion Foullioya (P. barbellatus Huynh, P. sylvicola Huynh), und eine neue Subsektion dieser Sektion (subsect. Barbellati Huynh), werden beschrieben. Ein Schlüssel zur Bestimmung der gegenwärtig bekannten Arten der Sektion wird vorgeschlagen.
introDuction
Pandanus sect. Foullioya is endemic to madagascar. It is characterized by unilocular drupes with a stigma deeply divided into two lobes. The type species, P. pygmaeus Thouars, has been known since 1808. It is very characteristic with its low shrubby habit (about 1 m high), its short and narrow leaves (30-40 x 0.6-0.9 cm), and its infructescence with 4-5 small syncarps. It is widespread in the South-eastern part of madagascar where it forms dense thickets along streams, and was therefore the most collected of the section. However, for a century and a half after it was described, no other species were found in the section.
In 1951 four other species were added to the section: P. bilamellatus martelli, P. longecuspidatus Pic. Serm., P. malgassicus Pic. Serm., and P. pseudocollinus Pic. Serm., described in Martelli & picHi-serMolli (1951). P. bilamellatus showed that the

infructescence of the section may have up to 8 syncarps, while P. longecuspidatus and P. pseudocollinus showed that its dis-tribution also includes the North-eastern part of madagascar, thus extending almost from the North to the South of the island. Nevertheless, no species has been found in the Western part. P. boivinii Solms, from Nosy Be, for which only staminate plant was known, was later tentatively placed in the section by stone (1970: 592). Although efforts have been and will be made to hunt for its pistillate plant, it is doubtful that this species still exists.
In the course of the present study, two other species were found, also in the Eas-tern part, increasing the number of species of sect. Foullioya to seven. One of the new species is so peculiar as to form a new subsection. The new species and the new subsection are described below.
observations
1. Tentative key to the species of sect. Foullioya
1. Terminal syncarp of infructescence oblong ovoid, 8 x 5.5 cm, 3-angled, divided laterally into costal and intercostal faces (costal faces composed of broad and divergent drupes; intercostal faces, of broad and divergent drupes in the peri-pheral part but of narrow and convergent drupes in the central part). Stigma lobes strongly recurved so as to be face to face with one another at the drupe backside. Leaves 130-140 cm long
Subsect. BarbellatiP. barbellatus
- Terminal syncarp of infructescence glo-bose/subglobose, at most 2-3 cm in dia-meter, not divided laterally into costal and intercostal faces. Stigma lobes not recurved so as to be face to face with one another at the drupe backside. Leaves 30-60 cm long
Subsect. Foullioya 2
2. Leaf apex abruptly attenuate, cuspidate/subcuspidate 3
- Leaf apex gradually attenuate, not cuspi-date/subcuspidate 5
3. Pileus 2/3 of the total length of the drupe. Leaf margins with remote prickles in the upper part
P. pseudocollinus
- Pileus 1/4-1/3 of the total length of the drupe. Leaf margins with close prickles in the upper part 4
4. Leaves chartaceous, 30-40 cm long. Drupes 25-30 per syncarp; endocarp glo-bose; lower mesocarp occupying the lower third of the drupe
P. longecuspidatus
- Leaves coriaceous, 55-60 cm long. Drupes 70 per syncarp; endocarp clavate; lower mesocarp nil
P. sylvicola
5. Syncarps 8 per infructescenceP. bilamellatus
- Syncarps 4-5 per infructescence 6 6. Leaves 50 x 1.2 cm; lower marginal prickles close, 2 (- 3) mm apart
P. malgassicus
- Leaves 30-40 x 0.6-0.9 cm; lower mar-ginal prickles remote, 5-9 mm apart
P. pygmaeus
2. Pandanus sylvicola Huynh, sp. nov. (sect. Foullioya)
Folia 55-60 cm longa 2.3-2.5 cm lata in parte supera, 1.6 cm lata prope basim, in 2/10 superis abrupte attenuata, subcuspi-data, 3-4 cm caudata, tertia infera deorsum versus sensim attenuata, basi amplexi-caulia; in sicco coriacea, brunnea/brun-
54
k.-l. HuynH

neola, e basi laminae ad basim caudae fortiter revoluta; plicis subdistinctis, inermibus; venis longitudinalibus dis-tinctis in ambabus paginis sed transver-salibus invisibilibus, interdum obscuris in pagina abaxiali; denticulis margina-libus e circa 1 cm supra basim ad apicem praesentibus, omnibus antrorsis, albidis interdum brunneis in apice, in tertia infera minutis densiusculis deinde ad 1.5-2 mm longis 7-8 mm inter se separatis sursum versus ad apicem omnibus minutis, in media ad 4-5 mm inter se separatis, in supera inferne ad 3-4 mm inter se separatis superne densis; denticulis costalibus prae-sentibus in tertia supera solum, omnibus minutis plerumque magis distantibus quam marginalibus proximis; vagina circa 1 cm longa 2.5 cm lata, basim versus leviter expansa, pagina abaxiali dense longitu-dinaliter venata, adaxiali inferne dense longitudinaliter venata superne laevi niti-daque. Infructescentia 6-syncarpica; syn-carpiis racemosis, sessilibus, globosis vel subglobosis, mediis grandissimis 2 cm longis 1.8 cm latis circa 70 drupis com-positis; pedunculo 8.5 cm longo 6 mm lato, acute triquetro, dense longitudinaliter nervato. Basi infructescentiae ad apicem: bractea 14-15 cm longa 1.4 cm lata, syn-carpium infimum sustinens, naviculare, 3-4 cm caudata; deinde 8 cm longa 1.6 cm lata, naviculare, 2-3 cm caudata; 6 cm longa 2.4 cm lata, naviculare, vix caudata; 4.5 cm longa 2 cm lata, naviculare, non caudata; 3 cm longa 1 cm lata, navicu-lare, non caudata; postremo paulo bre-vior angustiorque, naviculare, non cau-data; margines omnes tote armati, costae mediae ± similiter armatae. Drupae ad 9 mm longae 4-5 mm latae 3-4 mm crassae, in 1/3-1/4 supera liberae; pileo late pyra-midali, 2-3 mm alto, partem liberam tote obtegenti; stigmate unico, bilobato, lobis divergentibus, verticalibus vel obliquis, vel horizontalibus cum superficiebus papillatis expositis; endocarpio tenuissimo praecipue in partibus lateralibus, usque ad basim
extenso, apice plano circiter 1/5 supera locato; mesocarpio supero semi-lunato, circa 0.8 mm alto, medulloso, intra fibris destituto; loculo seminali clavato, 6.5 mm longo 2.5 mm lato; mesocarpio infero nullo. (fig. 1-6).Type: Cremers 2121 (holo mO!); mada-gascar, in forest between Foulpointe and Andondabe, 12 may 1972.
Foulpointe, now mahavelona, is at about 17°41’S 49°31’E; Andondabe, 17°46’S 49°23’E.
P. sylvicola appears closest to P. longe-cuspidatus, from which it differs in the following characters. Its syncarps may have up to about 70 drupes; the pileus of its drupes is large pyramidal (fig. 4); its endocarp is clavate and its lower meso-carp nil (fig. 5); its leaves are coriaceous and 55-60 cm long. In P. longecuspidatus: the drupe number per syncarp does not exceed 25-30; the pileus is hemispheric; the endocarp is globose; the lower meso-carp occupies the lower third of the drupe; the leaves are chartaceous and 30-40 cm long (Martelli & picHi-serMolli, 1951: 107-108, fig. 18h). The leaves, also, are more abruptly attenuate in the upper part than those of P. sylvicola (compare fig. 18f in Martelli & picHi-serMolli, 1951, with fig. 2 in the present paper). Further-more, P. longecuspidatus was found in the Antalaha region (14°54’S 50°17’E), while P. sylvicola was found much further south (between 17°41’S 49°31’E and 17°46’S 49°23’E). Given the endemism of Pan-danus species in madagascar, this remote distribution also suggests that P. sylvicola is distinct from P. longecuspidatus.
The infructescence of P. sylvicola, com-posed of small and globose/subglobose syncarps (fig. 6), is typical of sect. Foul-lioya, as seen in other species in the sec-tion (Martelli & picHi-serMolli, 1951: fig. 18). Its drupe with lower mesocarp nil (fig. 5) recalls that of P. pseudocollinus where the lower mesocarp is almost nil
55
tHe Genus panDanus (panDanaceae) in MaDaGascar (part 2)
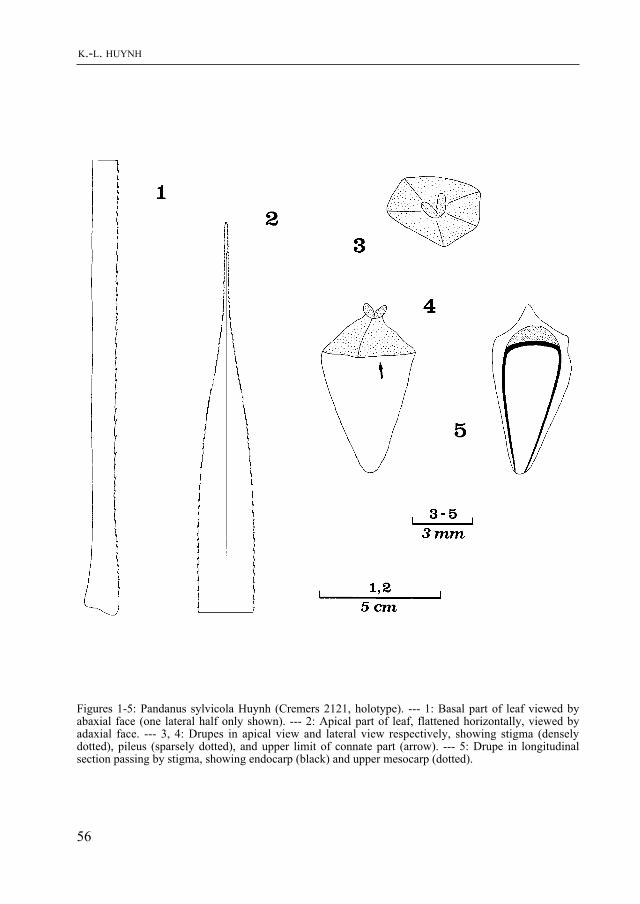
56
k.-l. HuynH
Figures 1-5: Pandanus sylvicola Huynh (Cremers 2121, holotype). --- 1: Basal part of leaf viewed by abaxial face (one lateral half only shown). --- 2: Apical part of leaf, flattened horizontally, viewed by adaxial face. --- 3, 4: Drupes in apical view and lateral view respectively, showing stigma (densely dotted), pileus (sparsely dotted), and upper limit of connate part (arrow). --- 5: Drupe in longitudinal section passing by stigma, showing endocarp (black) and upper mesocarp (dotted).

57
(Martelli & picHi-serMolli, 1951: 111, fig. 18r). In the drupe, the upper meso-carp is bordered by a layer of longitudinal fibres which extend from apex to base, thus also bordering the endocarp; however it does do not have fibres within.
3. Pandanus barbellatus Huynh, sp. nov. (sect. Foullioya)
Folia 135-140 cm longa 3.5 cm lata in medio 4 cm lata prope basim, in dimidio supero sensim attenuata, in sicco coriacea; plicis inermibus; venis longitudinalibus distinctis in pagina abaxiali, minus in adaxiali, transversalibus visibilibus in parte infera paginae adaxialis solum; den-ticulis marginalibus e circa 12 cm supra basim ad apicem praesentibus, stramineis sed apice brunneis, omnibus antrorsis, in tertia infera usque ad 4 mm longis 15 mm inter se separatis, in media 4 mm longis 10 mm inter se separatis, in supera 2-3 mm longis inferne 5 mm inter se separatis superne 2 mm vel minus; denticulis cos-talibus praesentibus in 2/3 superis solum, 1/3-1/2 brevioribus sed ± tam distantibus quam marginalibus proximis; vagina circa 9 cm lata in medio, ut videtur 5 cm longa, venis longitudinalibus partim visibilibus in ambabus paginis, transversalibus invi-sibilibus in pagina abaxiali partim visi-bilibus in adaxiali. Infructescentia 4-syn-carpica; pedunculo ex parte conservato 1 cm crasso, perspicue triquetro, multi-canaliculato inter angulos, sparsim verru-cato. Syncarpium terminale grandissimum, oblongo ovoideum, 8 cm longum 5.5 cm latum, circa 600 drupis praeditum, trique-trum; superficiebus costalibus drupis latis divergentibus compositis, intercostalibus drupis latis divergentibus in parte periphe-rica sed angustis convergentibus in cen-trali; bractea circa 6 cm longa 2.5 cm lata, naviculari, leviter acuminate, marginibus fere e basi ad apicem armatis, costa media apice in brevissimo spatio solum armata. Syncarpia infera leviter ovoidea, 5.5-6 cm longa 4-4.5 cm lata, sessilia. Drupae syn-carpii terminalis longissimae, circa 22 mm longae 5 mm latae 4 mm crassae, in tertia supera liberae; pileo pyramidali, dimidium superum partis liberae obte-genti; stigmatibus unicis, raro 2, bilobatis, lobis fortiter recurvatis; endocarpio circa 15 mm alto, 2 mm ab apice drupae distanti
tHe Genus panDanus (panDanaceae) in MaDaGascar (part 2)
Figure 6: Pandanus sylvicola Huynh (Cremers 2121, holotype). Infructescence (from the top downwards: the terminal syncarp, shown by the upper arrow, partially covered by bract; to the left, the third syncarp, almost entirely covered by bract; to the right, the fourth syncarp, the largest one visible; to the left, the fifth syncarp; to the right, the sixth syncarp, the lowermost; peduncle shown by the lower arrow; the second syncarp is invisible, superimposed by the terminal syncarp and the fourth syncarp). Scale bar = 2 cm.
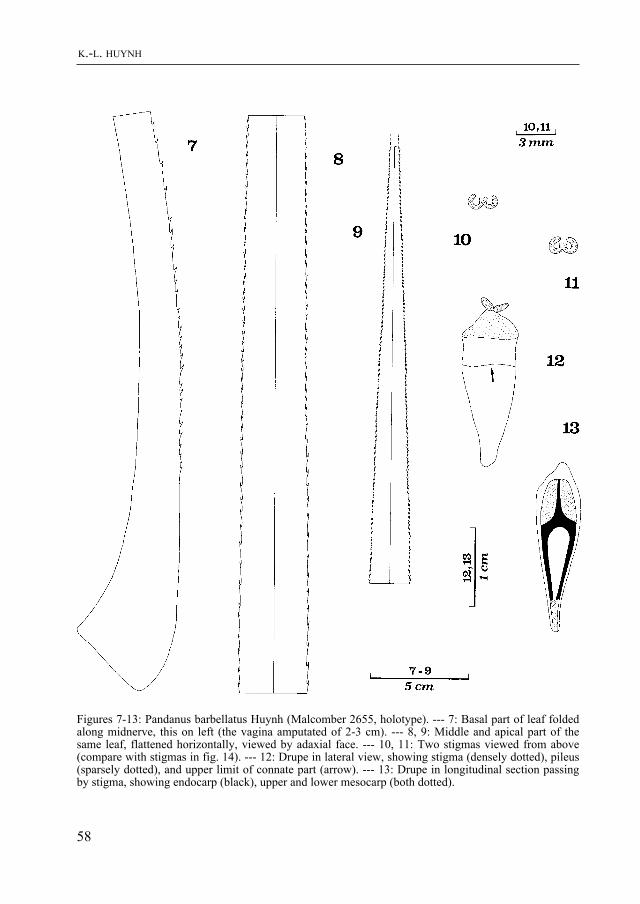
58
k.-l. HuynH
Figures 7-13: Pandanus barbellatus Huynh (malcomber 2655, holotype). --- 7: Basal part of leaf folded along midnerve, this on left (the vagina amputated of 2-3 cm). --- 8, 9: middle and apical part of the same leaf, flattened horizontally, viewed by adaxial face. --- 10, 11: Two stigmas viewed from above (compare with stigmas in fig. 14). --- 12: Drupe in lateral view, showing stigma (densely dotted), pileus (sparsely dotted), and upper limit of connate part (arrow). --- 13: Drupe in longitudinal section passing by stigma, showing endocarp (black), upper and lower mesocarp (both dotted).

59
tHe Genus panDanus (panDanaceae) in MaDaGascar (part 2)
4 mm a basi, ex apice loculi seminalis sursum versus sensim attenuato; loculo seminali 8-9 mm longo 3 mm lato, clavato, centro leviter inframediano; mesocarpio supero circa 6 mm alto, medulloso, intra fibris destituto, basi prope apicem loculi seminalis locato; mesocarpio infero 4 mm longo, non medulloso, intra fibris destituto. (fig. 7-14).Type: malcomber 2655 (holo P!; iso mO!); madagascar, NW of Tolanaro, Andohahela Réserve Intégrale, parcelle 2, 24°57’S 46°39’E, alt. 30 m, 24 December 1993; streamside tree 6 m tall, fruit green.
In the infructescence of P. barbellatus, the terminal syncarp has three costal faces and three intercostal faces; the latter faces are quite different in drupe width from the former faces, as described above; the narrowest drupes are located in the central part of the intercostal faces (fig. 14). The lower syncarps have smaller and fewer drupes; the difference in drupe width between the costal and intercostal faces is hardly visible. The second syncarp is 5.5 cm long and 4.5 cm wide, located at about 3.5 cm from the base of the terminal syncarp, separated from this by two inter-nodes, the upper internode about 1 cm long, the lower internode 2.5 cm. The third syncarp, 6 cm long and 4 cm wide, is at about 1.5 cm from the base of the second syncarp, and is separated from it by one internode. The fourth syncarp, 6 cm long and 3.8 cm wide, is at about 2.5 cm from the base of the third syncarp, and is sepa-rated from it by two internodes, the upper internode about 2 cm long, the lower inter-node 0.5 cm.
In the drupe of P. barbellatus, the two mesocarps are bordered by a layer of lon-gitudinal fibres which extend from apex to base. However they do not have fibres within. The layer also borders the endo-carp.
P. barbellatus is named in reference to its stigma whose lobes are recurved (fig. 10 and 11) like a moustache.
With its infructescence, stigma, and leaves, P. barbellatus appears isolated in sect. Foullioya, and can therefore be easily recognized. The terminal syncarp of the infructescence is oblong ovoid, tri-angled, up to 8 cm long and 5.5 cm wide, and com-prises a very large number of drupes (up to about 600). In lateral view, it shows six faces: three costal faces, composed of broad and divergent drupes; three intercostal faces, of broad and divergent drupes in the peripheral part, but of narrow and conver-gent drupes in the central part (fig. 14).
Figure 14: Pandanus barbellatus Huynh (mal-comber 2655, holotype). Terminal syncarp of a 4-syncarpic infructescence in lateral view, showing an intercostal face (compare stigmas in central part of the face with those in fig. 10 and 11). Scale bar = 2 cm.

60
This type of syncarp evokes that of sect. Heterostigma, another section of Pandanus in madagascar: in this respect, P. barbel-latus may constitute a phylogenetic link between these two sections. The stigma lobes are long and strongly recurved so as to be face to face with one another at the drupe backside (fig. 10 and 11), as can be seen on several drupe apices in the central part of fig. 14 (these are «white» points). The leaves are up to 140 cm. In the other species of the section, by contrast: the ter-minal syncarp is globose or subglobose, not tri-angled, at most 2-3 cm in diameter, not divided laterally into costal and inter-costal faces, and comprises at most about 70 drupes; the stigma lobes are divergent (fig. 3) and may be slightly curved, but are not recurved like those of P. barbel-latus (fig. 10 and 11); the leaves do not exceed 50-60 cm in length. With its unu-sual characters mentioned above, P. bar-bellatus strongly suggests a section of its own which is closest to sect. Foullioya. However, given its stigma deeply divided into two lobes, a characteristic to date considered proper to sect. Foullioya, P.
barbellatus is maintained in this section as the type of a subsection, which is defined below:
4. Pandanus sect. Foullioya Warb. subsect. Barbellati Huynh, subsect. nov.
Stigma 2 lobis fortiter recurvatis pro-funde divisum. Syncarpium terminale oblongo ovoideum, triquetrum, magnum (circa 8 x 5.5 cm); superficiebus costa-libus drupis latis divergentibus compositis, intercostalibus drupis latis divergentibus in parte peripherica sed angustis conver-gentibus in centrali; drupis numerosissimis (circa 600). Folium longum (circa 140 cm).
Type: P. barbellatus Huynh.
acknowleDGeMents
The author is deeply indebted to the missouri Botanical Garden, St. Louis, and the Laboratoire de Phanérogamie, muséum National d’Histoire Naturelle, Paris, for the loan of materials. Part of the Grant No. 31-45707.95 of the Swiss National Science Foundation was used.
k.-l. HuynH
references
Martelli, u. & picHi-serMolli, r. 1951. Les Pandanacées récoltées par Henri Perrier de la Bâthie à madagascar. mém. Inst. Sci. madagascar, Sér. B, Biol. vég. 3 (1): 1-174.
stone, b. c. 1970. New and critical species of «Pandanus» from madagascar. Webbia 24: 579-618.
__________
bull. soc. neucHâtel. sci. nat. 121 : 53-60. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 61-68. 1998
61
CHROmOSOmE STUDIES AND DISTRIBUTION OF NINE SPECIES OF COUSINIA SECTION STENOCEPHALAE (AS-
TERACEAE) IN IRAN
SEyyED mAHmOOD GHAFFARI 1 * and S. B. DJAvADI 2
1Institute of Biochemistry and Biophysics, University of Tehran, P.O. Box 13145-1384, Tehran, Iran. 2Plant pests and Diseases Research Institute, Section Botany, ministry of Agriculture, Tehran, Evin, Iran.
Résumé
quelque 9 espèces iraniennes de Cousinia sect. Stenocephalae (Asteraceae) font l’objet d’une étude caryologique. Les nombres chromosomiques de base, x=13, et la valence diploïde paraissent homogènes. Les régions du nord-est de l’Iran constituent le centre de diversité et d’origine de la section Stenocephalae.
Summary
Some nine Iranian species of Cousinia sect. Stenocephalae (Asteraceae) are the subject of a caryo-logical study. The base chromosome number is x=13, and the diploid valence appears homoge-nous. Northeastern Iran appears to be the center of diversity and origin of Cousinia section Steno-cephalae.
introDuction
Cousinia section Stenocephalae Bunge, with thirty-four species, is the third largest of the genus after sections Cynaroideae and Alpinae respectively (recHinGer, 1972). Of these, twenty-three are endemic to Iran. To date, chromosome studies on the section have been limited to six species: cHykasanova (see feDorov, 1974), afzal-rafii (1980), GHaffari & cHariat-panaHi (1985), and tscHerneva (1985). In this paper, we report on nine species that are endemic (except one) to Iran. We also present chromosome counts and notes on their meiotic behavior for the first time.
observations
The results of this study are summarized in Table 2. However, each species is dealt with in detail here.
Cousinia aggregata DC.* Author for correspondence

This species is endemic to Iran, and is found in the central and eastern parts of the country. meiosis was observed in three collections (Table 1), with 13 bivalents, three of which were associated with the nucleolus in diakinesis (fig. 1, 2). Occasio-nally, in some cells, one quadrivalent was observed. Chiasma frequency was 1.86, 1.80 and 1.68 per bivalent. This is the first chromosome count for this species.
Cousinia assyriaca Jaub. & Spach.This species is endemic to Iran. meiosis
in this species is regular with 13 bivalents, each of which usually had two terminal chiasmata per arm (fig. 3, 4).
The mean number of chiasmata was estimated at 1.56 for each bivalent at metaphase I. Occasionally, in some cells, one quadrivalent was observed. This is the first chromosome count for this species.
Cousinia calolepis Boiss.This species is endemic to Iran, and is
found in a restricted area of the country. meiosis in this species was regular and showed thirteen bivalents at metaphase I, most of which appeared in a ring confor-mation (fig. 5, 6). The mean number of chiasmata was estimated 1.82 for each bivalent at metaphase I. This is the first chromosome count for this species.
Cousinia commutata BungeThe previous report of this species is
2n=26 (afzal-rafii, 1980). In our study, meiosis was seen at diakinesis, metaphase I, and Anaphase one. Thirteen bivalents were observed at metaphase I, but also in some cells, one, two or three quadriva-lents were observed. Chiasma frequency was 1.84 per bivalent. In diakinesis, three bivalents of chromosomes were associated with nucleolus (fig. 7, 8).
Cousinia cylindracea Boiss.
This species is endemic to Iran, and is found in many parts of the country. meiosis in this species showed 13 biva-lents at metaphase I, but in some cells, one or two quadrivalents were observed (fig. 9, 10). The chiasma frequency determined from six cells, were 1.78 per bivalent. This is the first chromosome count for this spe-cies.
Cousinia gaubae Bornm.This species is endemic to Iran, and is
found in a restricted area of the country (Tehran, Karaj, and qazvin). The previous chromosome count of this species is n=13 (GHaffari & cHariat - panaHi, 1985). We found 13 bivalents of 11 bivalents and one quadrivalent at metaphase I. Three bivalents were associated with nucleolus at diakinesis stage. The mean number of chiasmata per each bivalent was 1.86 at metaphase I. Chromosome segregation at Anaphase I was also (13-13) (fig. 11,12).
Cousinia glaucopsis Bornm. & Rech.f.This species is endemic to Iran, and
is distributed in the areas of Damghan, Semnan, Firouz-kuh, and Gorgan. meiosis in this species was regular and showed 13 bivalents at metaphase I which most of them were in a ring conformation.
There are usually two terminal chiasma per arm. The mean number of chiasmata per each bivalent was 1.81 at metaphase I. In diakinesis stage 3 bivalents of chromo-somes were associated with nucleolus (fig. 13, 14). This is the first chromosome count for this species.
Cousinia nekarmanica Rech.f.This species is endemic to Iran, and is
found in a restricted area of the country (Semnan, Damghan, and Firouz-kuh). meiosis was regular and shows 13 biva-lents at metaphase I (fig. 15, 16). Occasio-nally, in some cells, one quadrivalent was
62
s. M. GHaffari et s. b. DJavaDi
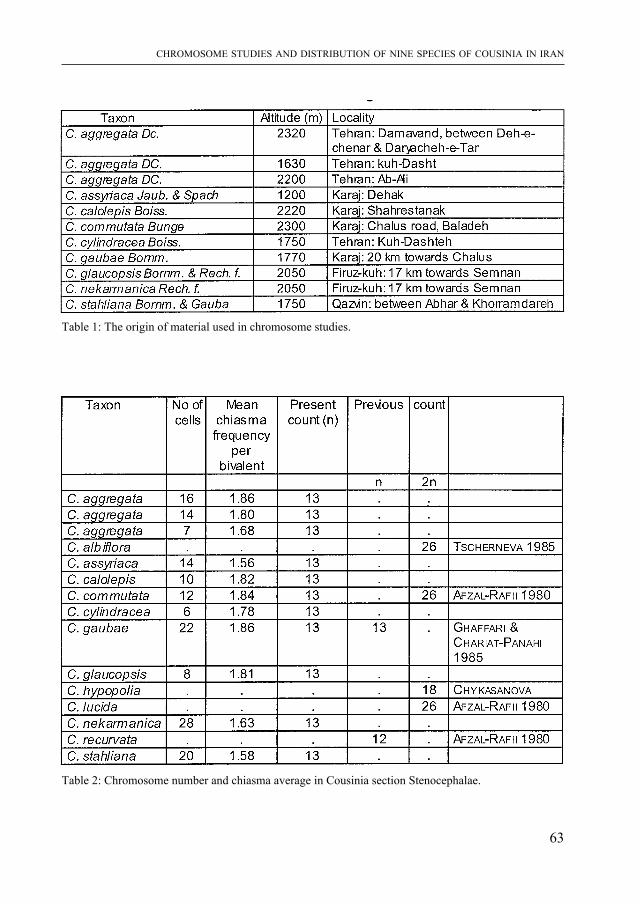
63
cHroMosoMe stuDies anD Distribution of nine species of cousinia in iran
Table 1: The origin of material used in chromosome studies.
Table 2: Chromosome number and chiasma average in Cousinia section Stenocephalae.
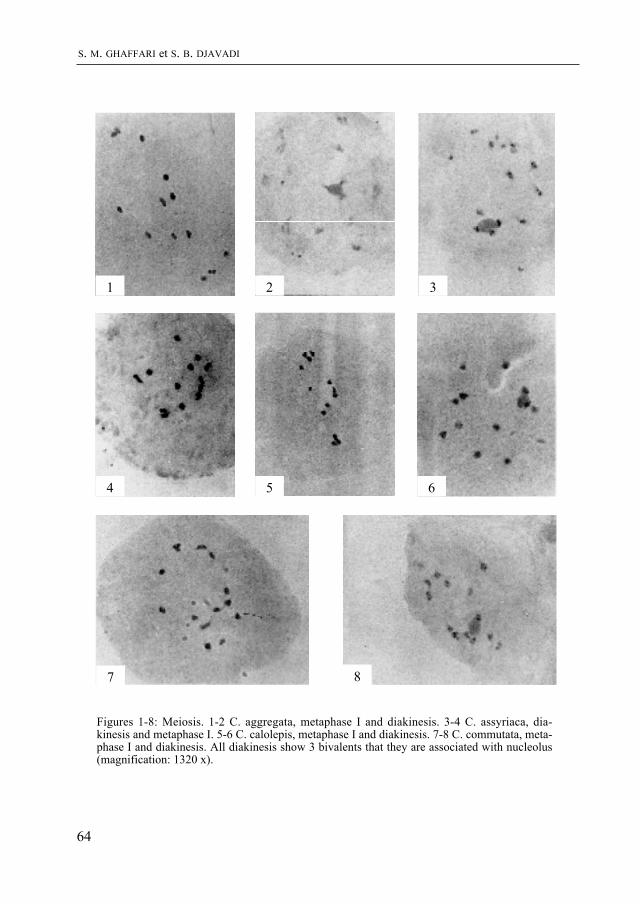
64
s. M. GHaffari et s. b. DJavaDi
Figures 1-8: meiosis. 1-2 C. aggregata, metaphase I and diakinesis. 3-4 C. assyriaca, dia-kinesis and metaphase I. 5-6 C. calolepis, metaphase I and diakinesis. 7-8 C. commutata, meta-phase I and diakinesis. All diakinesis show 3 bivalents that they are associated with nucleolus (magnification: 1320 x).
1 2 3
4 5 6
7 8
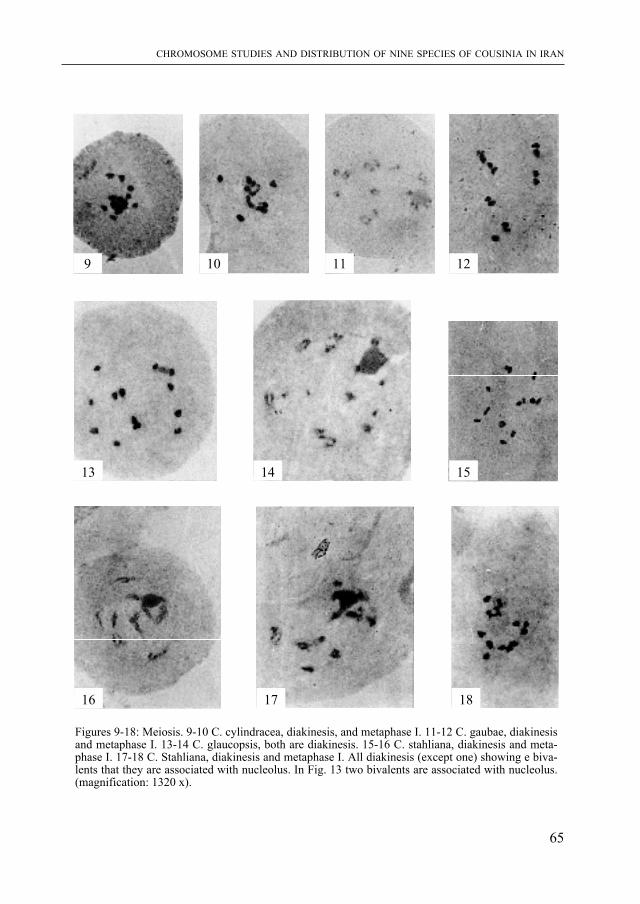
65
cHroMosoMe stuDies anD Distribution of nine species of cousinia in iran
Figures 9-18: meiosis. 9-10 C. cylindracea, diakinesis, and metaphase I. 11-12 C. gaubae, diakinesis and metaphase I. 13-14 C. glaucopsis, both are diakinesis. 15-16 C. stahliana, diakinesis and meta-phase I. 17-18 C. Stahliana, diakinesis and metaphase I. All diakinesis (except one) showing e biva-lents that they are associated with nucleolus. In Fig. 13 two bivalents are associated with nucleolus. (magnification: 1320 x).
9 10 11 12
13 14 15
16 17 18

66
observed. Chiasma frequency in 14 cells gave a mean of 1.69 per bivalent. This is the first chromosome count for this species.
Cousinia stahliana Bornm. & GaubaThis species is endemic to Iran and
Turkmenistan. Eleven bivalents and one quadrivalent were observed at metaphase I. Three pairs of chromosomes were asso-ciated with the nucleolus at diakinesis (fig. 17, 18). Chiasma frequency was 1.58 per bivalent. This is the first chromosome count for this species.
Discussion
recHinGer (1986) reported that Cou-sinia with 662 species is the third largest genus in the Asteraceae, and is also the largest in the tribe Cynareae. The distribu-
tion of Cousinia overlaps the Irano-Turk-menistanian region, but most of the species are concentrated in the mountains of Iran and Turkmenistan (knapp, 1987).
Cousinia section Stenocephalae is one of the largest sections of the genus, and is distributed in southwestern Asia (fig. 19). It appears that northeastern Iran is the center of diversity and origin of this section because of the 34 species in the section, 22 of the 25 species that occur in Iran are endemic. Iran shares 2 species with Turkmenistan (Cousinia stahliana and C. hypopolia Bornm. & Sint.) and one each with Iraq, Syria, and Turkey (C. stenocephala). Turkmenistan (Kopetdagh region) has 8 species, of which 6 are endemic (C. oreoxerophila, C. mucida, C. leptocephala, C. albiflora, C. chejra-badensis and C. chaetocephala). Cousinia
s. M. GHaffari et s. b. DJavaDi
Figure 19: Distribution of different species of the section Stenocephalae. Overlapping areas show the common species.

67
stenocephala Boiss. is the only species native to Iraq that ranges to Turkey and Syria. Turkey has 4 species, of which one (C. davisiana Hub.-mor.) is endemic (Huber-MoratH, 1975). Syria has 3 spe-cies, namely, C. foliosa Boiss. & Bal., C. stenocephala, and C. ramosissima Boiss., the former of which ranges to Lebanon (post, 1933). As is shown in fig. 19, the density and variation of the section decreases from east to west where only a single species occurs in Lebanon.
The conclusions obtained from the chro-mosome studies on pollen mother cells, show the same base chromosome number (x=13) in all species. From a cytological point of view, the presence of three chro-mosomes associated with the nucleolus in diakinesis stage, quadrivalent combi-nations, and eventually the similarity of chiasma frequency average for each biva-lent, verify the affinity of these species to another (table 2). Therefore, it appears that the placement of these species in sec-tion Stenocephalae based on Rechinger classification (recHinGer, 1972) is in accordance with their cytological affi-nities. There is only one base chromo-some number in section Stenocephalae, i.e. x=13. The presence of x=9 and x=12 seems to be contradictory (table 2).
In a cytological investigation of C. recur-vata DC., afzal-raffii (1980) reported its chromosome number as n=12. Since quadrivalent combination in meiosis of the species under this section were observed in plenty, the error in the chromosome counts is related to one tetravalent instead
of one bivalent (l.c.). cHykasanova (see feDorov, 1974) gave the chromosome count of C. hypopolia as 2n=18. In this species, it seems that the error in the chro-mosome count is either due to the misi-dentification of the species or simply a wrong chromosome count. Besides, in the other cases on chromosome reports, the writer (l.c.) disagrees with other investiga-tor’s opinions. For example, the chromo-some complement in C. karatavia Regel & Schmalh. was reported by him as 2n=26, was also counted as 2n=36 (tscHerneva, 1985). Also, 2n=18 for C. bekeri Trautv. which belongs to the section Cynaroideae, seems to be erroneous, because according to the other workers (GHaffari, 1984, 1986 and 1987; aryavanD, 1975; afzal-raffii, 1980 and tscHerneva, 1985), the base chromosome number in this section was reported as x=12. Consequently, the base chromosome number in section Ste-nocephalae, follows a general trend of sta-bility. It also appears that an important role was played by the heterozygotic transloca-tion in the evolution of this section.
acknowleDGeMents
Research was supported by the Plant Pests and Diseases Research Institute (Sec-tion Botany), ministry of Agriculture, and Institute of Biochemistry and Biophysics, University of Tehran. We thank Prof. m. Iranshahr for his help and identification of the plants.
references
cHroMosoMe stuDies anD Distribution of nine species of cousinia in iran
afzal-rafii, Z. 1980. Contribution à l’étude cytotaxonomique de quelques Cousinia d’Iran. Rev. Biol. Ecol. medit. 7: 6-14.
aryavanD, A. 1975. Contribution à l’étude cytotaxonomique de quelques Angiospermes de l’Iran. Bot. Notiser 128: 299-311.
feDorov, A.A. (ed.) 1974. Chromosome numbers of flowering plants. Königstein.
GHaffari, S.m. 1984. Chromosome counts LXXXIII. Taxon 33: 353.

68
GHaffari , S.m. 1986. Chromosome counts XCIII. Taxon 35 (4): 907.
GHaffari , S.m. 1987. Chromosome studies in some flowering plants of Iran. Rev. Cytol. Biol. veget-Bot. 10: 3-8.
GHaffari , s. & cHariat-panaHi, s. 1985. Chromosome count of some Angiosperms from Iran. Iran. J. Bot. 3 (1): 67-73.
Huber-MoratH, A. 1975. Cousinia Cass. In: Davis, P.H. et al. Flora of Turkey and the East Aegean Islands 5: 329-353. Edinburgh.
knapp, H.D. 1987. On the distribution of the genus Cousinia (Compositae). Pl. Syst. Evol. 155: 15-25.
post, G.E. - (1933). Flora of Syria, Palestine and Sinai, II: 84-87. American University of Beirut.
recHinGer, K. H. 1986. Cousinia - morphology, taxonomy and phytogeographic implications. Proc. Royal Soc. Edinburgh, 89 B: 45-58.
tscHerneva, O.v. 1985. Chromosome numbers in the species of the genus Cousinia (Asteraceae) in the flora of the USSR. Bot. Zurn. SSSR. 70 (7): 856-857. (in Russian).
wilson, G.B., 1945. The venetian turpentine mounting medium. Stain Technology 20: 133-135.
__________
s. M. GHaffari et s. b. DJavaDi
bull. soc. neucHâtel. sci. nat. 121 : 61-68. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 69-77. 1998
69
CHRySONEBULA HOLmESII LUND (ALGUE: CHRySOPHyCEAE),
PREmIÈRE mENTION POUR LA SUISSE
OLIvIER REymOND 1, FRANÇOIS STRAUB 2, HANS RUDOLF PREISIG 3
1Conservation de la faune, ch. du marquisat 1, 1025 St-Sulpice, Suisse.
2Laboratoire d’algologie, Gymnase cantonal, rue du Succès 45, 2300 La Chaux-de-Fonds, Suisse.
3Institut für Systematische Botanik, Universität Zürich, Zollikerstrasse 107, 8008 Zürich, Suisse.
mots-clés: Algues, Chrysophyceae, Hydrurales, Chrysonebula holmesii, Hydrurus, Cello-niella, macroinvertébrés, diatomées, mucilage, rivière, bioindicateurs, Suisse.
Key-words: Algae, Chrysophyceae, Hydrurales, Chrysonebula holmesii, Hydrurus, Cello-niella, macroinvertebrates, diatoms, mucilage, river, bioindicators, Switzerland.
Résumé
L’algue Chrysophycée Chrysonebula holmesii Lund se caractérise, macroscopiquement par une masse mucilagineuse blanchâtre attachée aux cailloux du fond de la rivière, et microscopiquement par des cellules portant un long filament de forme irrégulière. Cette espèce est signalée pour la première fois en Suisse, dans le Nozon, une rivière du Jura. A l’aide des indices biotiques utilisant les macroinvertébrés benthiques, ainsi que les diatomées prises dans le mucilage, on constate que cette algue se développe dans une eau d’excellente qualité. Sur le plan de la systématique, il est raisonnable de maintenir Chrysonebula et Hydrurus dans deux genres séparés, malgré beaucoup de similarités. Il en est de même entre Chrysonebula et Celloniella.
Summary: Chrysonebula holmesii Lund (Chrysophyceae), a species new to Switzerland.
The chrysophycean alga Chrysonebula holmesii Lund is macroscopically characterized by a creamy white mass of mucilage attached to the stones on the bottom of a stream. microscopically some cells possess a long and irregularly shaped thread arising from the posterior end. This spe-cies is recorded for the first time in Switzerland from the stream “Nozon” in the Jura mountains. Biological indices based on the biodiversity of the macroinvertebrates of the stream, as well as the diatoms of the mucilage, show that the water is of a very good quality. Concerning the systematics it appears to be justified to maintain Chrysonebula as a genus separate from Hydrurus and Cello-niella, though there are many similarities.

70
Dans le cadre de la surveillance biolo-gique des rivières du canton de vaud par la Conservation de la faune, des prélè-vements d’invertébrés benthiques (larves d’insectes) sont régulièrement effectués dans les principaux cours d’eau afin d’en connaître leurs qualités (lanG & rey-MonD, 1995; lanG, 1998).
Lors d’un prélèvement dans la partie haute du Nozon (lanG, 1998), nous avons remarqué que les cailloux qui forment le lit de la rivière étaient recouverts d’une géla-tine blanchâtre peu appétissante. L’ana-lyse au microscope optique nous a montré qu’il ne s’agissait pas de microorganismes typiques des rejets d’eaux usées, mais au contraire de Chrysonebula holmesii Lund, une algue Chrysophycée des cours d’eau apparemment non pollués. A notre connaissance cette algue n’avait jamais été signalée pour la Suisse.
malgré le très petit nombre de publica-tions qui la concerne, sa morphologie a été déjà bien décrite au microscope optique par lunD (1953) et coute (1976), et au microscope électronique par HibberD (1977). Ils nous a donc paru intéressant de discuter de cet organisme en mettant l’accent sur son environnement au moyen des indicateurs biotiques: les invertébrés benthiques et les diatomées.
Nous abordons aussi sa morphologie, sa systématique et ses affinités avec Hydrurus (coute, 1976, HibberD, 1977) et Cello-
niella (preisiG, 1995). Bien que très suc-cincts, quelques éléments géographiques et physiques concernant la rivière nous ont également parus dignes d’être mentionnés.
station et MétHoDes
Chrysonebula a été observée et prélevée le 8 mars 1996 au point 5202/1715 de la carte Nationale (Coordonnées suisses) à 920 m d’altitude, à environ 1,5 km de la source du Nozon, dans la chaîne calcaire du Jura. La masse gélatineuse contenant les algues a été délicatement décollée des cailloux du lit et mise dans des bocaux pour le transport jusqu’au laboratoire.
Afin de connaître au mieux la qualité environnementale des eaux du Nozon, les invertébrés benthiques ont été récoltés au cours de deux campagnes, le 31 janvier et le 8 mars 1996. En piétinant le fond de la rivière, les animaux ont été délogés et emportés par le courant dans un filet de maille 0,2 mm. Le matériel, récolté et fixé sur place, a été déterminé au laboratoire (lanG, 1998).
Pour étudier la composition du peuple-ment de diatomées, un peu de mucilage a été prélevé au coeur de la masse coloniale et traité au peroxyde d’hydrogène (H202 30%) pendant 2 semaines à température ambiante. Les frustules ainsi nettoyés ont été récupérés sur membrane de téflon (Sar-torius®, 5 µm de porosité, straub, 1981),
o. reyMonD et al.
Zusammenfassung: Chrysonebula holmesii Lund (Chrysophyceae), ein Neufund für die Schweiz.
Die Goldalge Chrysonebula holmesii Lund (Chrysophyceae) bildet auffällige weissliche Gallert-lager auf Steinen am Flussgrund. mikroskopisch ist die Alge durch ihre charakteristischen Zellen kennbar, die an ihrem Hinterende teilweise einen unregelmässigen, langfädigen Auswuchs tragen. Diese wenig bekannte Alge ist in der Schweiz erstmals im Flüsschen Nozon im Juragebiet (Kanton Waadt) gefunden worden. Im gleichen Gewässer vorkommende makroinvertebraten, die als Bioin-dikatoren dienen, wie auch in der Algengallerte vorhandene Diatomeen, zeigen an, dass diese Alge in Wasser von sehr guter qualität wächst. In systematischer Hinsicht scheint es gerechtfertigt zu sein, Chrysonebula neben Hydrurus und Celloniella als selbständige Gattung zu führen, obwohl diese Algen viele Ähnlichkeiten aufweisen.
introDuction

lavés à l’eau déionisée et montés entre lame et lamelle dans du Naphrax pour l’observation à fort grossissement (Leitz® Dialux au contraste de phase, Leica® DLmB au contraste interférentiel).
résultats
Systématique et morphologie
Le genre Chrysonebula (Hydrurales, Chrysophyceae) comprend trois espèces: C. holmesii lunD (1953) décrite pour l’Angleterre, C. flava starMacH (1966) ainsi que C. fonticola starMacH (1986), ces deux dernières étant décrites pour la Pologne (voir aussi starMacH, 1985). Ce genre a également fait l’objet de deux autres publications, l’une concernant la microscopie optique (coute, 1976), et l’autre concernant principalement l’ultras-tructure (HibberD, 1977).
Les cellules de notre matériel (Fig. 1) sont légèrement ovoïdes (longueur 10 - 12 µm, largeur 9 - 10 µm). Beaucoup de cellules possèdent un filament caracté-ristique, dont la longueur, très variable d’une cellule à l’autre peut atteindre 250 µm. Ce filament, souvent très sinueux et parsemé de boursouflures peut se diviser en plusieurs sous-unités. Les cellules pos-sèdent un ou deux plastes ainsi qu’un ou deux pyrénoïdes. Aucun kyste n’a été observé dans notre matériel. Les cellules forment des colonies globuleuses enrobées dans une masse gélatineuse formant des thalles de couleur crème de quelques cen-timètres d’épaisseur, tapissant les cailloux. La masse gélatineuse n’a pas une struc-ture très compacte, mais au contraire s’ef-filoche dans le courant en bras sinueux de plusieurs centimètres. De très nom-breux cristaux de carbonate de calcium (coute, 1976) ainsi que des diatomées (voir “Indice biotiques” ci-dessous), sont dispersés dans la gelée.
Paramètres physiques
En amont et à l’endroit de la récolte, la rivière, d’une largeur moyenne de 2 m, coule de façon rectiligne sur 500 m, du sud-ouest vers le nord-est sans qu’aucun rideau d’arbres, si ce n’est un encaisse-ment de 1 m environ, ne puisse faire de l’ombre sur le fond. Ce dernier est relati-vement plat, formé de cailloux de 1 à 10 cm, ovoïdes ou anguleux. La dénivellation de ce tronçon est de 1 % environ. Lors du prélèvement, la rivière était dans une période d’étiage: la profondeur de l’eau était de 20 cm, la vitesse du courant de 0,3 m/s, et la température de 3,2 °C.
Indices biotiquesLes macroinvertébrés
L’indice RIvAUD (lanG & reyMonD, 1995), calculés à partir des deux prélè-vements, nous donne une note de 18/20. L’indice IBGN (afnor, 1992), utilisé généralement en France, nous donne une note de 16/20. Ces deux indices sont liés à la diversité générale de la faune benthique ainsi qu’au degré de sensibilité de cer-taines espèces. Les taxons identifiés sont indiqués dans le tableau 1. Les résultats concernant le Nozon ainsi que d’autres rivières du Jura sont donnés par LanG (1998).
Les diatoméesL’observation de Chrysonebula au
microscope à 10 x 40 de grossissement permet de se rendre compte que la masse mucilagineuse est colonisée par d’autres microorganismes. Les plus évidents sont des diatomées (Bacillariophyceae), en particulier les colonies typiques en éven-tails de meridion circulare (Grev.) Ag. et les longues cellules de Fragilaria ulna (Nitzsch) Lange-B. On remarque égale-ment des cyanobactéries (Cyanophyceae) du genre Oscillatoria ainsi que des bacté-ries filamenteuses et des bacilles sporu-lants ou non.
71
cHrysonebula HolMesii (alGue cHrysopHyceae) preMière Mention pour la suisse
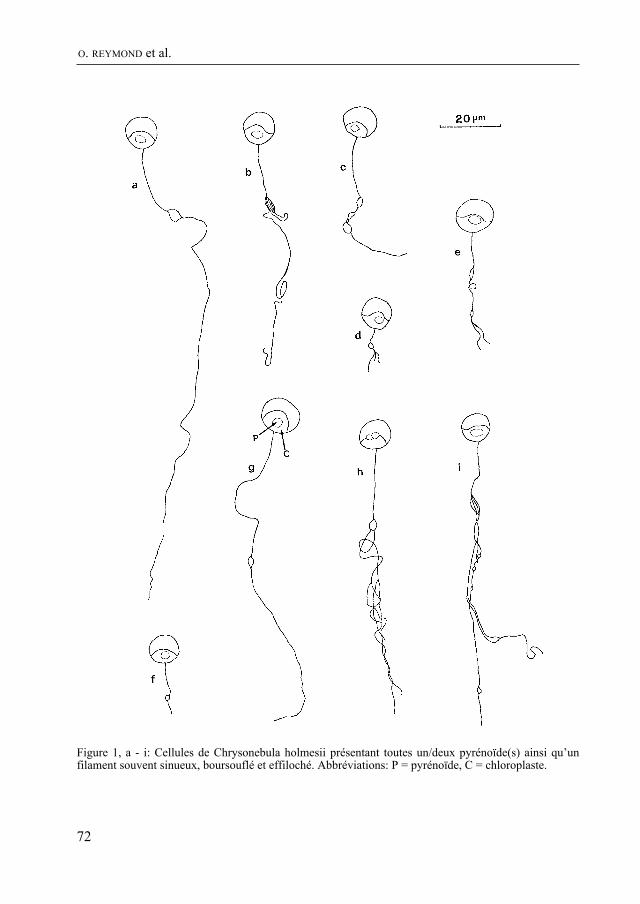
72
o. reyMonD et al.
Figure 1, a - i: Cellules de Chrysonebula holmesii présentant toutes un/deux pyrénoïde(s) ainsi qu’un filament souvent sinueux, boursouflé et effiloché. Abbréviations: P = pyrénoïde, C = chloroplaste.

73
L’identification des diatomées a suivi les propositions taxonomiques de kraMMer & lanGe-bertalot (1986-1991), reicHarDt & lanGe-bertalot (1991), sala et al. (1993), kraMMer (1997), kobayasi (1997) partiellement. L’examen détaillé a fourni une flore de 33 taxons (tableau 2) dont 15 forment la flore dominante (flore obtenue au cours d’un dénombrement de 500 valves de dia-tomées). Par ordre d’importance les cinq taxons suivants forment 93,8 % des indi-vidus de diatomées du mucilage: meridion circulare (espèce pionnière), Achnanthes minutissima var. minutissima, Encyonema
ventricosum, Achnanthes pyrenaica et Nitzschia fonticola. Ce peuplement com-prend 52,8 % de taxons rhéophiles (préfé-rant les rivières). D’autres sont cependant bien représentés par exemple Achnanthes minutissima var. minutissima (31,4%) ou Achnanthes pyrenaica (6,2%) qui vivent aussi bien en rivière qu’en eau stagnante, préférentiellement comme épiphytes sur les algues filamenteuses ou sur les plantes aquatiques, en particulier dans la zone de battement des vagues.
Discussion
cHrysonebula HolMesii (alGue cHrysopHyceae) preMière Mention pour la suisse
Tableau 1: Liste des taxons de macroinvertébrés répertoriés au cours des deux campagnes de prélève-ments. L’astérisque signale les taxons considérés comme sensibles aux pollutions lors du calcul de l’in-dice RIvAUD (lanG & reyMonD, 1995). Dans le calcul de l’indice l’IBGN (afnor, 1992), les Chloro-perlidae représentent le groupe faunistique indicateur.
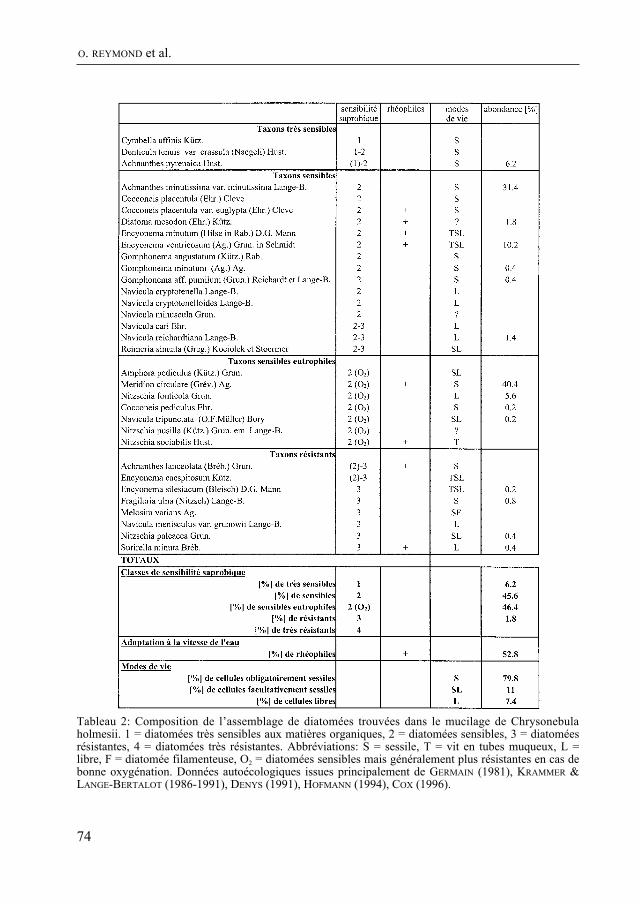
74
o. reyMonD et al.
Tableau 2: Composition de l’assemblage de diatomées trouvées dans le mucilage de Chrysonebula holmesii. 1 = diatomées très sensibles aux matières organiques, 2 = diatomées sensibles, 3 = diatomées résistantes, 4 = diatomées très résistantes. Abbréviations: S = sessile, T = vit en tubes muqueux, L = libre, F = diatomée filamenteuse, O2 = diatomées sensibles mais généralement plus résistantes en cas de bonne oxygénation. Données autoécologiques issues principalement de GerMain (1981), kraMMer & lanGe-bertalot (1986-1991), Denys (1991), HofMann (1994), cox (1996).

Environnement Les macroinvertébrés
Les notes très élevées des deux indices biotiques (rIvAUD: 18/20 et IBGN: 16/20), montrent qu’au lieu de récolte, l’état de la rivière est d’excellente qualité (lanG, 1998). malheureusement il n’existe aucune indication comparable pour les sta-tions mentionnées par lunD (1953), coute (1976), ou HibberD (1977).
Les diatoméesLa composition de la végétation diato-
mique par classes de sensibilité envers les matières organiques (tableau 2) permet de diagnostiquer l’état saprobique des eaux (kraMMer & lanGe-bertalot, 1986-1991). L’abondance relative de plus de 90% de taxons sensibles et la très faible abondance de taxons résistants indiquent des eaux ß-mésosaprobes typiques des cours d’eau jurassiens en bonne santé, en bonne corrélation avec l’image de l’état de santé de la station donnée par les macroinvertébrés. Cependant la forte abondance relative de taxons eutrophiles (taxons qui résistent mieux aux matières organiques en présence d’une bonne oxy-génation selon scHiefele, 1987 et rei-cHarDt, 1991), ainsi que la très faible représentation des taxons très sensibles indiquent la présence de matières orga-niques inhabituelles dans de tels cours d’eau. On peut penser que cette composi-tion peut fréquente est liée à la présence des matières organiques du mucilage de l’algue colonisée. Récemment czarnecki (1995) a fait une courte revue des popu-lations de diatomées qui colonisent les mucilages d’organismes aquatiques. Cet auteur présente les diatomées qui colo-nisent les mucilages du cilié péritriche Ophrydium versatile (O.F. müll.) Ehr. et cite celles qui ont été observées chez la cyanobactérie microcystis aeruginosa (Kütz.) Lemmermann ou dans les tubes muqueux d’autres diatomées. En parti-
culier, il relève la présence de Cymbella cesatii (Rab.) Grun. et Nitzschia flexoides Geit., diatomées semblerait-il obligatoire-ment associées à de tels mucilages. Dans la population relevée ici, Encyonema spp. et Nitzschia sociabilis peuvent sécréter leurs propres tubes muqueux, mais aucune des diatomées trouvées n’est obligatoire-ment associée à de telles formations extra-cellulaires d’autres organismes microsco-piques. On peut considérer que les diato-mées présentes dans ce mucilage sont soit fortuites, englobées de manière passive, soit qu’elles y vivent pour un intérêt parti-culier, par exemple sur le plan nutritionnel en profitant de la minéralisation active due à l’activité bactérienne ou en profi-tant de certaines molécules organiques excrétées par les divers microorganismes associés (tucHMan, 1996). En particulier ici, Achnanthes minutissima, Encyonema minutum, probablement aussi E. ventri-cosum, Navicula tripunctata, melosira varians et Nitzschia paleacea sont connues pour leur hétérotrophie facultative.
Affinités entre Chrysonebula et HydruruslunD (1953), mentionne que les colonies
des deux algues ont des aspects souvent très ressemblants. coute (1976) trouve les deux taxons dans la même station, et observe pour la première fois des kystes chez Chrysonebula. Le kyste d’Hydrurus étant déjà connu, cet auteur écrit: “L’ana-logie entre les kystes de ces deux algues est si frappante que l’on peut envisager l’hypo-thèse que ces deux organismes ne sont que des stades différents d’une seule et même algue”. Pour HibberD (1977), les kystes observés par coute (1976) pourraient en fait être ceux d’Hydrurus et non ceux de Chrysonebula; et bien que les deux taxons soient très similaires à plusieurs point de vue, il propose de maintenir les deux genres séparés. En ce qui concerne notre matériel, rappelons que nous n’avons pas observé de kystes. De plus nous n’avons
75
cHrysonebula HolMesii (alGue cHrysopHyceae) preMière Mention pour la suisse

76
o. reyMonD et al.
pas trouvé d’Hydrurus, algue que nous ren-controns pourtant fréquemment lors de la surveillance biologique des rivières. Sans nouveaux résultats probants, il ne nous paraît pas judicieux de placer les deux taxons sous la même dénomination spéci-fique ou générique.
Affinités entre Chrysonebula et CelloniellaDes thalles coloniaux et gélatineux simi-
laires à ceux de Chrysonebula et Hydrurus peuvent aussi être produits par la seule espèce connue de Celloniella, C. palensis (pascHer, 1929). Cette espèce, rarement mentionnée (preisiG, 1995), est extrême-ment polymorphique, mais généralement les cellules sont concentrées en une fine couche à la périphérie du thalle gélatineux.
Un long filament postérieur tel que celui de Chrysonebula n’a jamais été observé. Les cellules de C. palensis diffèrent appa-remment aussi par l’absence de pyrénoïde.
reMercieMents
ClauDe lanG a dirigé la campagne de surveillance des rivières et nous a auto-risé à publier les résultats concernant les macroinvertébrés.
biblioGrapHie
AFNOR. 1992. Essais des eaux. - Détermina-tion de l’indice biologique global normalisé (IBGN). Association française de normalisa-tion, Paris.
Coute, a. 1976. Sur la présence en France du Chrysonebula holmesii Lund (Chrysophyceae) et sur la structure de ses kystes. C.R. Acad. Sc. Paris 282: Sér.D, 2159-2161.
cox, E. 1996. Identification of freshwater diatoms from living material. Chapman & Hall, London.
czarnecki, D.B. 1995. Additions and confirmations to the algal flora of Lake Itasca (mN) State Park. III. The intramucilaginous diatom flora of the colonial peritrich ciliate, Ophrydium versatile (Ophry-diidae). In: Kociolek, J.P. & Sullivan, m.J. (eds). A century of diatom research in North America. A tribute to the distinguished careers of Charles W. Reimer and Ruth Patrick. Koeltz Sc. Books USA: 183-194.
Denys, L. 1991. A check-list of the diatoms in the holocene deposits of the western Belgian coastal plain with a survey of their apparent ecological requirements. I. Introduction, ecological code and complete list. Service géologique de Belgique, Professional paper 246: 1-41.
GerMain, H. 1981. Flore des diatomées. Boubée, Paris.
HibberD, D.J. 1977. The cytology and ultrastructure of Chrysonebula holmesii Lund (Chrysophyceae), with special reference to the flagellar apparatus. Br. phycol. J. 12: 369-383.
HofMann, G. 1994. Aufwuchs-Diatomeen in Seen und ihre Eignung als Indikatoren der Trophie. Bibliotheca Diatomologica 30: 1-241.
kobayasi, H. 1997. Comparative studies among four linear-lanceolate Achnanthidium species (Bacil-lariophyceae) with curved terminal raphe endings. Nova Hedwigia 65 (1-4): 147-163.
kraMMer, K. 1997. Die cymbelloiden Diatomeen. Eine monographie der weltweit bekannten Arten. Teil I: Allgemeines und Encyonema. Bibliotheca Diatomologica 36: 1-390.

77bull. soc. neucHâtel. sci. nat. 121 : 69-77. 1998
kraMMer, k. & lanGe-bertalot, H. 1986 - 1991. Bacillariophyceae 1-4. In: Ettl, H., Gerloff, J., Heynig, H. & mollenhauer, D. (eds): Süsswasserflora von mitteleuropa (begr. von A.Pascher), G. Fischer, Stuttgart, Band 2/1-4.
lanG, C. 1998. qualité biologique de 37 rivières vaudoises en 1996 indiquée par la diversité du zoo-benthos. Bull. Soc. vaud. Sc. nat. 84: 323-332.
LanG, c. & reyMonD, O. 1995. An improved index of environmental quality for Swiss rivers based on benthic invertebrates. Aquatic Sciences 57: 172-180.
lunD, J.W. 1953. New or rare British Chrysophyceae II. Hyalobryon polymorphum n. sp. and Chryso-nebula holmesii n. gen., n. sp. New Phytologist 52: 114-123.
pascHer, A. 1929. Über die Beziehungen zwischen Lagerform und Standortsverhältnissen bei einer Gallertalge (Chrysocapsale). Arch. Protistenk. 68: 637-668.
preisiG, H.R. 1995. A modern concept of Chrysophyte classification. In: Sandgren, C.D., Smol, J.P. & Kristiansen, J. (eds), Chrysophyte algae: Ecology, Phylogeny and Development. Cambridge Uni-versity Press, Cambridge: 46-74.
reicHarDt, E. 1991. Beiträge zur Diatomeenflora der Altmühl III: Wasserqualität und Diatomeenbe-satz. Arch. Hydrobiol. Suppl. 89, Alg. Studies 62: 107-132.
reicHarDt, e. & lanGe-bertalot, H. 1991. Taxonomische Revision des Artenkomplexes um Gom-phonema angustum - G. dichotomum - G. intricatum - G. vibrio und ähnliche Taxa (Bacillariophy-ceae). Nova Hedwigia 53: 519-544.
sala, S.E., Guerrero, J.m. & ferrario, m.E. 1993. Redefinition of Reimeria sinuata (Greg.) Kociolek & Stoermer and recognition of Reimeria uniseriata nov. spec. Diatom Research 8: 439-446.
scHiefele, S. 1987. Indikationswert benthischer Diatomeen in der Isar zwischen mittenwald und Landshut. Diplomarbeit am Lehrstuhl für systematische Botanik an der Ludwig-maximilians-Uni-versität münchen.
starMacH, K. 1966. Über neue und seltene Chrysophyceae in der Algenflora Polens. Acta Hydrobiol. 8: 5-14
starMacH, K. 1985. Chrysophyceae und Haptophyceae. In: Ettl, H., Gerloff, J., Heynig, H. & mol-lenhauer, D. (eds). Süsswasserflora von mitteleuropa (begr. von A. Pascher), G. Fischer, Stuttgart, Band 1.
starMacH, K. 1986. Chrysonebula fonticola n. sp. Bull. Pol. Acad. Sci., Biol. Sci. 34: 131-133.
straub, F. 1981. Utilisation des membranes filtrantes en téflon dans la préparation des diatomées épi-lithiques. Cryptogamie: Algologie 2: 153.
tucHMan, N.C. 1996. The role of heterotrophy in algae. In: Stevenson, R.J., Bothwell, m.L. & Lowe, R.L. (eds). Algal ecology of freshwater benthic ecosystems. Aquatic Ecology Series, Acad. Press, San Diego: 299-319.
__________
cHrysonebula HolMesii (alGue cHrysopHyceae) preMière Mention pour la suisse


bulletin De la société neucHâteloise Des sciences naturelles 121 : 79-91. 1998
79
LE PARADOXE DE LA PODZOLISATION EN DOmAINE JURASSIEN
ASPECTS PÉDOLOGIqUES ET PHyTOSOCIOLOGIqUES 1
GAËLLE vADI ET JEAN-mICHEL GOBAT
Laboratoire d’Ecologie végétale, Université de Neuchâtel, rue Emile Argand 11, CH-2007 Neuchâtel 7, Suisse.
mots-clés: Jura, poDzosols, podzolisation, pessière, Sphagno-Piceetum, relations sol-végéta-tion.
Key-words: Jura mountains, podzosoils (Spodosols), podzolisation, spruce forest, Sphagno-Piceetum, soil-vegetation relationships.
Résumé
Dans le Jura, un lien très étroit a été observé entre les sols podzolisés et la pessière à blechnum. Ce phénomène concerne des surfaces s’étendant entre quelques mètres carrés et un hectare. Les auteurs dressent le bilan des connaissances acquises sur cette relation et complètent les données sur la podzolisation jurassienne. La végétation, étudiée au moyen de la phytosociologie synusiale intégrée, correspond au Sphagno-Piceetum blechnetosum. Cette phytocénose montre deux formes et contient généralement sept synusies. Une description morphologique des sols et une analyse du comportement de trois formes du fer (extractions tétraborate, pyrophosphate et oxalate) ont été effectuées. La convergence de ces critères confirme la présence de poDzosols Meubles parfaite-ment différenciés dans le Jura. Un indice synthétique d’entraînement du fer a été défini; il permet de distinguer trois catégories de sols à intensité d’éluviation croissante, reflétant des étapes succes-sives de la pédogenèse: podzolisation ménagée, franche ou extrême.
Summary
In the Jura mountains a very close relationship has been observed between podzolised soils and forests characterised by Spruce (Picea abies) and the Hard Fern Blechnum spicant. This pheno-menon concerns surfaces ranging from a few square meters to a hectare. The authors review the current knowledge on this relationship and add new data on podzolisation processes in the Jura. The vegetation studied by the synusial phytosociological method belongs to the Sphagno-Piceetum blechnetosum. This phytocoenosis occurs in two forms and usually contains seven synusiae. A morphological description of the soils and an analysis of the behaviour of three forms of iron (tetraborate, pyrophosphate and oxalate extractions) were done. The convergence of these two cri-teria confirms the presence, in the Jura, of perfectly differentiated spoDosols (AFES classification: poDzosols Meubles). A synthetic index of iron leaching was defined; it allows the distinction of three soils categories characterised by an increasing eluviation intensity and reflecting the succes-sive stages of the pedogenesis: weak, clear and extreme podzolisation.
1 Cet article est issu du travail de diplôme du premier auteur

introDuction
La podzolisation ...mode d’évolution pédologique excep-
tionnel dans le Jura, la podzolisation est active dans 16 % de la couverture pédo-logique mondiale (peDro in bonneau & soucHier, 1994), principalement dans les zones boréales froides (taïga et étages subalpin et montagnard). Elle consiste en phénomènes de soustraction-redistribution de composés organo-métalliques (chélates) au sein de la colonne de sol, sous l’in-fluence de molécules organiques acides issues de la dégradation lente d’une litière acidifiante (résineux, éricacées) (DucHau-four, 1983). Impropre à une bonne acti-vité de la faune des décomposeurs et de la microflore bactérienne, la litière s’accu-mule pour former un épisolum humifère épais et acide de type moder ou mor.
Sous l’effet des précipitations, ces couches organiques libèrent des composés agressifs pour les minéraux des horizons sous-jacents. Ces substances complexantes arrachent des ions aluminium, puis fer (phénomène d’acido-complexolyse), qu’ils entraînent en profondeur par chéluviation. Au fur et à mesure de cette migration, les conditions physico-chimiques du sol se modifient et les composés chéluviés précipitent, s’accumulent et se conden-sent sous forme d’hydroxydes dans des niveaux préférentiels (soucHier, 1984). A cette différenciation chimique correspond une morphologie caractéristique influencée principalement par la coloration du fer dans ses diverses formes. Les horizons appauvris blanchissent (zone d’éluviation, horizons E) tandis que les horizons d’accu-mulation (zone d’illuviation, horizons BP) se colorent en ocre ou en orangé (DucHau-four, 1983; soucHier, 1984). Dans les poDzosols les plus évolués, l’illuviation procède en deux phases: la matière orga-nique se fige dans un horizon noir BPh qui surplombe un niveau rouge-orangé où se condensent les minéraux amorphes,
hydroxydes de fer et d’aluminium (horizon BPs) (riGHi in: AFES, 1995).
... impensable dans le Jura!La couverture pédologique jurassienne
est souvent peu épaisse en raison de sa relative jeunesse (âge tardi- à postwür-mien, décapage glaciaire) et d’une éro-sion importante (forte pluviosité, soutirage karstique) (bouyer et al., 1978). On peut la subdiviser en quatre catégories prin-cipales selon l’importance de la matière organique et des carbonates, ou selon les conditions locales d’altération et d’hydro-morphie:- sols humifères peu évolués (orGanosols, litHosols, réGosols),- sols calci-magnésiques (renDosols, ren-Disols),- sols brunifiés, dominants dans le Jura (calcosols, calcisols, brunisols, néo-luvisols),- sols hydromorphes (réDuctisols, Histo-sols).
Les conditions climatiques régnant à l’étage subalpin du Jura sont favorables à la podzolisation; toutefois celle-ci y est bloquée pour deux raisons (MicHalet & bruckert, 1986):a) La nature du soubassement rocheux, dominé par des sédiments jurassiques ou crétacés (ricHarD, 1961).
Ceux-ci présentent majoritairement des séquences calcaires et marneuses, recou-vertes parfois par des paquets de molasse tertiaire (aubert et al., 1979; bouyer et al., 1978).b) La trop faible épaisseur des sols.
Le flux des ions calcium en provenance de la roche-mère est maintenu par la pré-sence, au sein du sol, de fragments de roche (sols calcaires ou calciques) ou par le jeu du cycle biogéochimique (sols bru-nifiés). Ce n’est qu’au delà d’une couche de 35 à 40 cm de matériau silicaté (apport
80
G. vaDi et J.-M. Gobat

allochtone limoneux, résidu insoluble important de calcaires marneux, etc.) que le calcium perd son influence, laissant le sol s’orienter vers une pédogenèse acide. C’est le domaine des sols bruns ocreux (alocrisols) qui présentent un début de podzolisation (bruckert & Gaiffe, 1980; MicHalet & bruckert, 1986; Havlicek & Gobat, 1996). Ces sols ont longtemps été considérés comme les seuls représen-tants de la podzolisation dans le Jura.
Et pourtant...En 1961, ricHarD découvre un véri-
table poDzosol dans la Forêt des Ecorces (Département du Doubs, France), dont la présence résulte d’une combinaison de conditions stationnelles précises. La décou-verte d’autres sols de ce type dans la vallée de La Brévine, à Chasseral et aux Saignolis (chaîne de Pouillerel) permet à ricHarD de définir les composants de cette combi-naison: climat humide et froid, morpho-logie plane permettant un écoulement d’eau vertical, roche-mère riche en éléments sili-catés et végétation particulièrement aci-dophile, identique dans chaque cas réper-torié. Cette dernière est une sous-associa-tion de la pessière à sphaignes (Sphagno-Piceetum), à cortège floristique inédit. Caractérisée par une fougère acidophile, le blechnum en épi (Blechnum spicant), cette forme du Sphagno-Piceetum se distingue de l’autre sous-association (à Betula pubes-cens) par la nature du sol. S’il est fréquent de trouver les deux formes côte à côte, la sous-association à bouleau croît toujours sur la tourbe (Histosol) tandis que celle à blechnum le fait sur un sol podzolisé. Depuis ricHarD (1961), divers travaux ont mis au jour d’autres cas de présence conjointe du Sphagno-Piceetum blechne-tosum et de son étrange substrat (tab. 1).
L’existence paradoxale de poDzosols dans le Jura est ainsi aujourd’hui claire-ment établie et bien documentée, même si aucun travail global de description pédo-
logique n’a encore été effectué jusqu’ici. C’est pour combler cette lacune qu’a été réalisée la présente recherche dont les quatre buts sont de:- prouver la podzolisation jurassienne de
manière irréfutable,- la caractériser,- estimer son intensité,- préciser la nature du lien étroit existant
entre la végétation et le sol.
Matériel et MétHoDes
En vertu de la remarquable fidélité, soulignée par ricHarD (1961), entre le Sphagno-Piceetum blechnetosum et les sols podzolisés, notre recherche s’est orientée dans une double perspective: le sol et la végétation. Nous avons ainsi visité huit sites, dont sept choisis sur une base bibliographique (tab. 1).
Des fosses ont été creusées dans cha-cune des stations; les profils ont été décrits et analysés, puis les sols nommés selon le Référentiel pédologique (afes, 1995; Gobat & Havlicek, 1996). Des relevés phytosociologiques effectués selon la méthode synusiale intégrée (Gillet et al., 1991) complètent la caractérisation des stations. Les analyses usuelles en pédo-logie (pH, taux de saturation, perte au feu) ont été suivies d’extractions destinées à l’étude de la dynamique des métaux ché-latés puis précipités sous forme de ses-quioxydes. Les formes du fer ont été extraites de la manière suivante:- formes complexées mobiles: tétraborate-Na à pH 9,7 (rouiller, 1981),- formes complexées immobiles: pyro-phosphate-Na 0,1 m ( rouiller, 1981),- formes amorphes: tampon oxalique (Tamm) (bonneau & soucHier,1994),- formes cristallisées: tampon CBD (mehra-Jackson) (bonneau & soucHier, 1994).
81
le paraDoxe De la poDzolisation en DoMaine Jurassien

82
résultats
Concernant la végétationLes relevés effectués sur les huit sites
présentent un grande homogénéité floris-tique et structurelle (tab. 2). Ils s’inscrivent tout à fait dans la définition du Sphagno-Piceetum blechnetosum et sont compa-rables à ceux des tableaux de végétation disponibles dans la littérature (ricHarD, 1961; buttler et al., 1983; Gillet et al., 1984; cHristen & frutscHi, 1987).
Les espèces différentielles de sous-association (Blechnum spicant, Pre-nanthes purpurea, Athyrium filix-femina) et d’association (Equisetum sylvaticum) sont fidèles. L’épicéa (Picea abies) est constant et domine la strate arborescente, Abies alba et Fagus sylvatica restant sous forme arbustive. Les essences buisson-nantes principales sont Sorbus aucuparia, F. sylvatica et Lonicera nigra. L’essentiel du couvert herbacé est formé de vacci-nium myrtillus, fréquemment associé à Oxalis acetosella, Dryopteris dilatata et maianthemum bifolium.
A l’exception de deux cas, la phyto-cénose comprend sept synusies: arbo-
rescentes haute et basse, buissonnantes haute et basse, herbacées haute et basse et muscinale. Au Bois Raiguel, la synusie buissonnante haute est manquante, consé-quence de la violente tempête survenue une dizaine d’années auparavant; le vent a également couché tous les arbres, à l’ex-ception des grands épicéas, d’où le carac-tère monospécifique des synusies arbo-rescentes. A Raimeux, ce sont les hautes herbes qui manquent à cause de la faible luminosité au niveau du sol (fort taux de boisement et feuillages denses), du piéti-nement et de l’abroutissement; la pression du bétail se marque également au niveau du recrû: les plantules ne dépassent pas dix centimètres et seul l’épicéa participe aux strates buissonnantes et arborescentes.
Concernant les solsA titre d’exemple, le poDzosol de La
vattay est décrit dans la figure 1.
Des indices d’entraînement (DucHau-four, 1983) ont été estimés (tab. 3). Ils reflètent la redistribution des composés ferriques de l’horizon éluvial à l’ho-rizon principal de précipitation des ses-quioxydes. Ils ont été établis à partir des extractions au tétraborate (comportement
G. vaDi et J.-M. Gobat
Tableau 1: Identification des sites étudiés (CN No: numéro de la carte nationale).

83
le paraDoxe De la poDzolisation en DoMaine Jurassien
Tableau 2: Relevés phytosociologiques (nomenclature des espèces selon aescHiMann & burDet, 1994).

84
G. vaDi et J.-M. Gobat
Figure 1: Description du poDzosol Meuble de La vattay et analyses physico-chimiques.

85
des composés labiles), au pyrophosphate (composés de précipitation) et à l’oxa-late (composés amorphes). Les teneurs en formes libres (CBD) ont été abandonnées dans ces calculs, car nous les avons jugées trop influencées par la quantité de com-posés ferriques dans la roche-mère.
Séparément, ces indices reflètent des comportements différents d’un solum à l’autre mais ne concernent qu’un aspect particulier de l’évolution du fer au sein du profil. Nous nous sommes par conséquent servis d’un indice moyen (moyenne arith-métique des trois indices susmentionnés) pour placer nos sols sur une échelle évolu-tive générale selon l’intensité de l’éluvia-tion subie (fig. 2). Le tableau 4 et la figure 3 intègrent des données recueillies dans la littérature.
Discussion
Des plantes…Dans les stations étudiées, la phytocé-
nose se distribue en deux sous-ensembles:
a) En ceinture externe de tourbièreDans ce cas, les surfaces occupées par le
Sphagno-Piceetum blechnetosum forment de longs rubans qui s’intercalent entre la sous-association à bouleau de la pessière à sphaignes (vers l’intérieur de la tourbière) et une hêtraie. Aux Saignolis, les bandes de pessière à blechnum atteignent une cen-taine de mètres de longueur. Aux Pontins, le ruban se découpe en fragments d’une quinzaine de mètres tandis qu’au Creux de l’Epral il est fractionné au point d’être limité à quelques bosquets d’épicéas.b) En mosaïque avec des zones humides envahies par la mégaphorbiaie
Il est ici également possible de dégager plusieurs niveaux spatiaux d’organisation (Gobat et al., 1998). A La vattay, les surfaces de pessière à blechnum dépassent la centaine de mètres et sont séparées par
de véritables “bras” de mégaphorbiaie, situés sur des réDuctisols. Les sols de ces deux phytocénoses distinctes sont indé-pendants du point de vue évolutif mais, à long terme, le mouvement général de soli-fluxion affectant cette zone peut aboutir à leur superposition (fig. 1). Dans la Forêt des Ecorces, la pessière à blechnum occupe des parcelles de quelques dizaines de mètres carrés alors qu’au Bois Raiguel elle se limite à la couronne des épicéas, soit une circonférence de quelques mètres. Plus spectaculaire encore, à Raimeux, la phytocénose est strictement limitée à la base du tronc des épicéas, où l’on observe la formation de véritables “podzols en entonnoir” (DucHaufour, 1983).
…et un solLes neuf sols étudiés présentent un ou
plusieurs horizons BP, une caractéris-tique morphologique indispensable pour les attribuer à la référence poDzosol du Référentiel pédologique (afes, 1995). Le sol de La Grosse Prise est un intergrade entre le poDzosol ocrique (pas d’horizon éluvial) et le poDzosol Meuble (à horizon éluvial). Les autres sols sont des poDzo-sols Meubles. La plupart des formes d’humus observées sont des moders plus ou moins évolués, voire même des mors à La vattay et à la Forêt des Ecorces.
Les analyses confirment la podzolisa-tion: pH bas, acidité d’échange élevée, faible taux de saturation, répartition conforme des formes du fer et de l’alu-minim. L’échelle d’éluviation fondée sur l’indice global d’entraînement (fig. 2) déli-mite trois catégories principales parmi nos profils:a) Un sol à podzolisation ménagée, forte-ment isolé des autres (La Grosse Prise)
La redistribution des complexes fer-riques y est à peine amorcée puisque les horizons éluviaux Eh et AE, par ailleurs mal différenciés, sont plus riches que l’ho-rizon principal d’accumulation BPs. Ce
le paraDoxe De la poDzolisation en DoMaine Jurassien

86
G. vaDi et J.-M. Gobat
F i g u r e 2 : G r a d i e n t de podzol i sa t ion (en ordonnée) établi en fonc-tion de l’indice d’entraî-nement moyen du fer (sols étudiés dans le présent tra-vail). Les flèches rappor-tent au nom des sites.
Tableau 3: Indices d’entraînement du fer. L’indice moyen représente la moyenne arithmétique des indices calculés pour les extractions dans le tétraborate (tétra), le pyrophosphate (pyro) et l’oxalate (ox).

87
diagnostic confirme les observations réa-lisées sur le terrain, ce solum nous ayant tout de suite semblé morphologiquement moins différencié que les autres. La valeur très élevée de l’indice dans le tétraborate (tab. 3) indique cependant une formation importante de complexes mobiles du fer dans la zone éluviale. Ces composés étant les marqueurs de l’acidocomplexolyse, le sol se trouve dans une étape précoce de la podzolisation, où la redistribution des complexes labiles est encore modeste.
b) Une majorité de sols à podzolisation plus ou moins marquée mais franche (Les Saignolis, Creux de l’Epral, Les Pontins, Bois Raiguel, Raimeux)
Ce second ensemble contient les sols dont les indices d’entraînement globaux sont situés entre 0,1 et 0,5. Dans chaque cas, les phénomènes de soustraction-redis-tribution du fer sont avancés. L’examen détaillé des indices spécifiques (tab. 3)
le paraDoxe De la poDzolisation en DoMaine Jurassien
Figure 3:Gradient de podzolisa-tion (en ordonnée) établi en fonction de l’indice d’entraînement du fer amorphe (extrait par l’oxalate) (sols étudiés dans le présent travail et sols de la littérature). Les flèches rapportent à l’identité des sols.

88
met en évidence les particularités fonction-nelles propres à chaque situation.
A la tourbière des Saignolis, la migra-tion du fer dans le sol n° 2 n’est pas très marquée pour les complexes mobiles et immobiles (éléments typiques de la podzo-lisation) alors que le sol n° 1 est “actif” pour les trois extractants. La topographie est la principale responsable de cette diffé-rence. Le sol n° 2 est en effet situé en bor-dure d’une doline, d’où un fort drainage de l’horizon d’accumulation et un indice plus élevé que dans le sol n° 1, sis sur un replat.
Le sol de la tourbière du Creux de l’Epral est surtout actif par le fer com-plexé mobile (indice tétraborate), ce qui confirme l’actualité de la podzolisation. A la tourbière des Pontins, les valeurs relati-vement homogènes dénotent une podzoli-sation effective mais qui n’est plus active depuis l’enfouissement assez récent du sol sous une couche limoneuse dont l’évolu-tion s’oriente selon la même voie.
Le sol du Bois Raiguel présente une podzolisation active, particulièrement dans la redistribution des complexes du fer (indices pyrophosphate et tétraborate). A Raimeux, les petits indices pyrophos-phate et oxalate attestent une importante redistribution, tandis que la faible valeur
de l’indice tétraborate est la conséquence d’un changement de pédogenèse, intervenu probablement suite à l’accroissement de la charge minérale en surface (forte présence bovine).
c) Deux sols à podzolisation extrême (La vattay, Forêt des Ecorces)
Les indices sont ici infimes, voire nuls. A la Forêt des Ecorces, l’horizon élu-vial est très appauvri: les indices faibles (tab. 3) indiquent une podzolisation en phase finale. A La vattay, l’évolution est achevée et le sol est stable puisqu’il ne présente plus aucun mouvement de matière chélatée.
Comme il n’a pas été possible d’ob-tenir toutes les données bibliographiques nécessaires au calcul de l’indice global d’entraînement, c’est l’indice relatif au fer amorphe (extraction à l’oxalate) qui a servi de base de comparaison avec la littérature. Le tableau 4 montre que ces sols se placent dans une suite évolutive logique (fig. 3): plus la podzolisation est marquée, plus la redistribution de complexes ferriques est importante. A l’exception de celui de la Grosse Prise, les sols du Jura se répartis-sent dans les solums où la podzolisation est effective et non parmi les intergrades.
Des plantes sur un sol
G. vaDi et J.-M. Gobat
Tableau 4: Indice d’entraînement du fer des sols de la littérature, calculé pour l’extraction dans l’oxalate (indice ox) (nomenclature des horizons selon DucHaufour, 1983).

Les phytocénoses étudiées correspon-dent au Sphagno-Piceetum blechnetosum de ricHarD (1961) et sont parfaitement fidèles, dans le Jura, aux poDzosols. La pessière à blechnum existe sous deux formes: continue en bordure de tourbière ou en mosaïque et alternant avec la mégaphor-biaie. Dans les deux cas, on peut distinguer des niveaux d’organisation variant du mètre à la centaine de mètres, le sol subissant des contraintes identiques. A cet égard, les exemples extrêmes de La vattay et de Rai-meux mettent en évidence la variabilité de l’extension spatiale du phénomène de podzolisation ainsi que les relations étroites qui unissent le sol et la végétation.
Les couples pessière à blechnum–poDzosol, mégaphorbiaie-réDuctisol et pessière à bouleau-Histosol consti-tuent autant de duos étroitement liés dans leur fonctionnement et leur évolution. A l’échelle de temps de l’évolution pédo-logique habituelle, chacun est totalement indépendant de l’autre. En terrain plat, dans des conditions écologiques équili-brées ou sur un substrat “solide”, il n’est pas envisageable de voir la myrtillaie et son poDzosol gagner peu à peu sur la mégaphorbiaie et son réDuctisol. Ce sont des systèmes indépendants.
En revanche, un type d’évolution pédo-logique particulier, la solifluxion, a été observé à La vattay. Elle provoque le rem-placement progressif, par de lents mouve-ments de terrain concernant plusieurs ares, du couple mégaphorbiaie-réDuctisol par le couple pessière à blechnum-poDzosol, ce dernier venant “glisser” par-dessus l’autre. Plusieurs sondages pédologiques (Gobat & GallanDat, inédit) ont prouvé ce phénomène de lent rééquilibrage de l’ensemble de la forêt de La vattay, dû probablement à la qualité particulière du substrat géologique (molasse rouge de La vattay: favre et al., 1971; aubert, 1975; Gillet et al., 1984). Le processus inverse
est d’ailleurs aussi possible, dans lequel la mégaphorbiaie vient remplacer la pessière.
conclusion
La pessière à blechnum constitue un milieu équilibré, mais d’une grande fragi-lité. Peu productive, elle résiste mal à la compétition (ricHarD, 1961). L’augmen-tation de la luminosité au sol, qu’elle soit d’origine anthropique (gestion sylvicole) ou non (catastrophe naturelle), engendre la prolifération des espèces herbacées de la mégaphorbiaie ou de la hêtraie en bor-dure de la phytocénose. Peu à peu, ces espèces très concurrentielles envahis-sent la pessière, induisant un ensemble végétal constitué de plantes acido-tolé-rantes à vitalité réduite et d’acidophiles. D’autre part, les caractéristiques extrêmes du milieu ne lui permettent guère de tam-ponner les atteintes qui lui sont portées. Le cas de Raimeux en témoigne: cantonnée à des surfaces très réduites, la pessière à blechnum y est en outre gênée dans son développement par la forte pression du bétail, qui s’exprime par l’abroutissement (destruction des herbacées et du recrû), le tassement du sol (hydromorphie induite dans les horizons de surface) et l’augmen-tation de la charge minérale (croissance d’espèces nitrophiles et modification des conditions de pédogenèse).
Parfait exemple des interactions sol-végétation (Gobat et al., 1998), les destins de la pessière à blechnum et du poDzosol sont intimement liés dans le Jura. Par leur rareté et le paradoxe qu’ils illustrent, ces fragments paysagers consti-tuent des éléments précieux du patrimoine naturel de notre région. Suite aux dégra-dations que nous avons notées sur le ter-rain, nous pensons que des mesures de protection immédiates sont indispensables à leur maintien à long terme.
89
le paraDoxe De la poDzolisation en DoMaine Jurassien

90
reMercieMents
Les auteurs remercient Jean-Daniel Gallandat et François Gillet de leur appui en phytoso-ciologie, Claire Arnold pour la détermination des mousses, Edward mitchell pour le résumé anglais, ainsi que Carlos Beck et Lidia Paganuzzi, du laboratoire d’analyses de l’Institut de géologie de Neuchâtel, de leur aide technique lors des analyses de sols.
biblioGrapHie
G. vaDi et J.-M. Gobat
aescHiMann, D. & burDet, H.m. 1994. Flore de la Suisse et des territoires limitrophes. Ed. du Griffon, Neuchâtel.
AFES. 1995. Référentiel pédologique. INRA, Paris.
aubert, D. 1975. L’évolution du relief jurassien. Eclog. Geol. Helv. 68 (1): 1-64.
aubert , D., Gratier, m. & pocHon, m. 1979. Livret-guide de quelques sols types du Haut-Jura et du pied du Jura. Laboratoire de Pédologie, PED 4, EPF-Lausanne.
bartoloMe, B. 1990. Etude pédologique et phytosociologique de la tourbière des Saignolis (NE). Trav. dipl., Labo. Ecol. vég., Univ. Neuchâtel.
bonneau, m. & soucHier, B. 1994. Pédologie 2: constituants et propriétés du sol. masson, Paris.
bouDot, J.-P. 1982. Relations entre l’altération minérale et le cycle de l’azote sur matériel chloriteux du massif schisto-grauwackeux vosgien. Thèse Univ. Nancy I.
bouyer, y., Miserez, J.-J. & pocHon, m. 1978. Inventaire géochimique et bilan du fer dans le sol et les eaux du karst jurassien: état, importance, déplacements. Laboratoire de Pédologie, PED 1, EPF-Lausanne.
bruckert, S. 1979. Classification des sols bruns ocreux et cryptopodzoliques par analyse des extraits tétraborates tamponnés à pH 9,5. Geoderma 22: 205-212.
bruckert, S. & Gaiffe, m. 1980. Analyse des facteurs de formation et de distribution des sols en pays calcaire glaciaire et karstique. Plaine de Frasne – Bonnevaux et montagne du Laveron – Jura central. Ann. scientifiques Univ. Besançon, Biol. vég., 4ème série (1): 19-67.
buttler, a., cornali, p. & ricHarD, J.-L. 1983. La Tourbière des Pontins sur Saint-Imier. Phytoso-ciologie, éléments d’écologie et perspectives d’aménagement. mat. Levé géobot. Suisse Fasc. 59.
cHristen, a.-M. & frutscHi, C. 1987. Creux de l’Epral, étude phytosociologique et écologique d’une tourbière des Franches-montagnes (JU). Trav. dipl., Labo. Ecol. vég., Univ. Neuchâtel.
DucHaufour, P. 1983. Pédologie 1: pédogenèse et classification. masson, Paris.

91
favre, GoMes-silva, Hossin, MicHel & siwertz. 1971. Saint-Claude, feuille géologique n° XXXIII-28 de la Carte géologique de la France au 1:50 000. Bureau de recherches géologiques et minières, Service géologique national, Orléans.
Gillet, F., lHote, P. & trivauDet, m.-J. 1984. Etude cartographique de la végétation et des milieux naturels du Jura gessien. Document interne, Fac. Sc. et techn. Univ. de Franche-Comté, Besançon.
Gillet, f., De foucault, b. & Julve, P. 1991. La phytosociologie synusiale intégrée, objets et concepts. Candollea 46: 315-340.
Gobat, J.-m. 1984. Ecologie des contacts entre tourbières acides et marais alcalins dans le Haut-Jura suisse. Thèse Univ. Neuchâtel.
Gobat, J.-m., araGno, M. & MattHey, w. 1998. Le sol vivant. Presses polytechniques et universi-taires romandes, Lausanne.
Gobat, J.-M. & Havlicek, e. 1996. Clé de sol. Principaux sols de Suisse. Document interne, Labo. Ecol. vég., Univ. Neuchâtel.
Havlicek, E. & Gobat, J.-m. 1996. Les apports éoliens dans les sols du Jura. Etat des connaissances et nouvelles données en pâturages boisés. Etude et Gestion des sols 3 (3): 167-178.
kraeHenbueHl, C. 1968. Associations végétales du Jura bernois. Actes Soc. jurass. Emul. 71: 201-322.
MicHalet, R. & bruckert, S. 1986. La podzolisation sur calcaire du subalpin du Jura. Science du sol 24 (4): 363-375.
paternoster, m. 1984. Dynamique de la colonisation des moraines latérales historiques du grand glacier d’Aletsch par des groupements végétaux pionniers. Pedogenèse initiale. Bull. murithienne 100: 65-78.
ricHarD, J.-L. 1961. Les forêts acidophiles du Jura, étude phytosociologique et écologique. mat. Levé géobot. Suisse Fasc. 38.
rouiller, J. 1981. Analyses des sols, techniques de laboratoire. Centre de pédologie biologique, CNRS, Nancy. Note technique n° 32.
soucHier, B. 1984. Les podzols et la podzolisation en climats tempérés et montagnards. Livre jubi-laire du cinquantenaire de l’AFES, p. 77-96.
__________
le paraDoxe De la poDzolisation en DoMaine Jurassien


bulletin De la société neucHâteloise Des sciences naturelles 121 : 93-110. 1998
93
ÉCOLOGIE DES PINÈDES (PINUS SyLvESTRIS)DE LA RIvE SUD DU LAC DE NEUCHÂTEL (SUISSE)
II. PHyTOmASSE ET PRODUCTIvITÉ AÉRIENNES, DyNAmIqUE DU GROUPEmENT 1
PHILIPPE CORNALI
Laboratoire d’écologie végétale et de phytosociologie de l’Université de Neuchâtel. Ch. de Chantemerle 18, 2000 Neuchâtel, Suisse.
mots-clés: écosystème riverain, Pinus sylvestris, phytomasse, productivité primaire, dyna-mique.
Key-words: lakeshore ecosystem, Pinus sylvestris, biomass of plant community, primary pro-duction, dynamic.
Résumé
La sociologie et l’écologie générale des forêts de pin de la rive sud du lac de Neuchâtel font l’objet d’une investigation approfondie, dans le cadre d’un projet d’étude de l’ensemble de l’écosystème riverain exondé il y a environ 100 ans.
Ce deuxième article consacré à la pinède à Pinus sylvestris expose les principaux résultats obtenus en matière de phytomasse, productivité et dynamique de renouvellement de la matière organique. La dynamique de croissance de la pinède indique aujourd’hui une stabilisation de la productivité. A moyen terme, plusieurs facteurs, parmi lesquels un décalage fonctionnel dans la dynamique productive du groupement, permettent d’envisager l’hypothèse d’un remplacement progressif de la pinède par les espèces ligneuses actuellement compagnes.
Summary
A detailed study of the general phytosociology and ecology of the pine forests of the south shores of Lake Neuchâtel was undertaken in the framework of a research project concerning the whole of the shore ecosystem, which emerged about 100 years ago.
This second paper give the principal results concerning the biomass, primary production and dynamic of organic matter of the Pinus silvestris forest. At present the growth dynamics of the pine forest indicate a stabilizing of productive capacity. At middle term several factors, among which a functional displacement in production dynamics, lead to the hypothesis of a gradual replacement of the pine forests by the present neighbouring deciduous species.
1 Cet article constitue un condensé de chapitres choisis de la thèse de l’auteur, effectuée au Laboratoire d’écologie végétale et de phy-tosociologie de l’Université de Neuchâtel. Les caractéristiques générales du milieu et la phytosociologie du groupement ont été abor-dées dans un précédent article (cornali, 1997, chapitres 1-6). La minéralomasse et les cycles biogéochimiques des éléments majeurs du groupement seront abordés dans un troisième article (chapitres 11-12).

71. pHytoMasse et proDuctivité aérienne
7.1 IntroductionLes conditions générales du milieu et
la phytosociologie de la pinède à Pinus sylvestris de la rive sud du lac de Neu-châtel, unité fonctionnellement intégrée à l’écosystème riverain exondé il y a environ 110 ans, ont été décrites dans un premier article (cornali, 1997). Après avoir établi cette typologie détaillée, nous pouvons maintenant aborder les descripteurs phy-tomasse, productivité et cycles biogéochi-miques, véritables caractéristiques de base des écosystèmes (DuviGneauD, 1971), pour tenter de préciser la dynamique fonc-tionnelle de ce groupement végétal.
Par rapport à l’étude originale (cornali, 1992), seuls les résultats synthétiques sont présentés et discutés ici, à l’exclusion du nombre important d’analyses comparatives consacrées aux trois placettes d’étude, voire à la dynamique saisonnière de cer-tains phénomènes.
Dans un domaine où la littérature est abondante, il importe de préciser le sens de quelques termes utilisés (présentés en détail dans DuviGneauD, 1980):
- biomasse: masse de tous les organismes vivants au moment de l’observation, exprimée en masse de matière sèche par unité de surface, par ex. t/ha;- phytomasse: biomasse végétale dans laquelle on inclut ici la minéralomasse, fraction minérale évaluable par l’analyse des cendres;- litière: matériel végétal mort retournant au sol. Abstraction faite de l’humus, la totalité de la matière organique morte, particulièrement difficile à déterminer en forêt, comporte encore la nécromasse (bois mort sur pied et feuilles mortes attachées à la plante);
- productivité primaire nette: vitesse de pro-duction de la phytomasse additionnée de l’ensemble des pertes annuelles, exprimée en masse de matière sèche par unité de sur-face et de temps, par ex. t/ha.an. En forêt, elle correspond donc pour l’essentiel à l’incrément des tissus ligneux auquel il faut ajouter la totalité de la matière orga-nique morte dans le courant de l’année. Cette dernière comprend la chute de litière ainsi que la fraction de bois mort pendant l’année, fraction indéterminable dans la mesure où la chute de cette nécromasse est différée de plusieurs années. Une apprécia-tion peut toutefois en être faite en consi-dérant que la mortalité annuelle du bois correspond à la fine litière de rameaux capturée dans les cages à litière. Enfin, de manière simplifiée, la productivité annuelle de la strate herbacée peut être assimilée à sa biomasse (mesurée à bio-masse maximale).
Les pertes dues à la consommation (phy-tophages) et à l’exportation sont négligées.
Ces définitions sont également valables pour les parties souterraines, non invento-riées dans notre étude.
La mesure de la productivité primaire en milieu forestier présente d’importantes dif-ficultés (voir par ex. rieDacker, 1968a et 1968b; leMee, 1974; parDe, 1979; Duvi-GneauD, 1984). Il s’ensuit que nos résul-tats doivent être considérés comme des estimations (une amélioration notable de la précision implique en effet l’établissement de tarifs de cubage locaux, donc l’abat-tage exhaustif des surfaces échantillons, tâche qui dépasse de loin les disponibilités d’un modeste laboratoire, et qui n’est pas envisageable dans un milieu naturel aussi exceptionnel que celui de la rive sud du lac de Neuchâtel...!)
Sans entrer dans le détail des méthodes, il convient néanmoins de préciser les prin-cipales étapes de la démarche effectuée sur les trois placettes d’étude couvrant au total 1100 m2:
94
pH. cornali
1 La numérotation des chapitres, tableaux et figures continue celle de cornali (1997), les deux publications formant une suite.

a) Phytomasse aérienne:- arborescente (diamètre > 8 cm): mesure intégrale des DBH (diamètre à hauteur de poitrine), échantillonnage des hauteurs H par classes de diamètres de 2 cm, qui sont ensuite complétées par régression sur DBH, puis extraction des volumes frais en utilisant un tarif à double entrée (tables de cubage pour l’Allemagne du sud: Hilfstafeln für Die forsteinricHtunG, 1966). Pour les petits diamètres com-pris entre 5 et 8 cm, non inclus dans les tables, les volumes ont également été cal-culés par régression sur DBH. Le passage des volumes frais aux biomasses sèches est effectué par le biais de l’infradensité (masse sèche/volume frais), préalable-ment déterminée sur quelques échantillons par espèce. Les volumes des houppiers sont également fournis par les tarifs de cubage, alors que les aiguilles sont esti-mées directement en biomasse sèche sur la base des proportions moyennes par rapport aux ramilles totales (houppier) indiquées par burGer (1948). La biomasse foliaire des espèces caducifoliées est mesurée par les chutes annuelles recueillies dans des paniers à litière (cf. 7.3.1);- arbustive: même démarche, mais pesées directes pour les échantillons dont le dia-mètre ne dépasse pas 4 cm;- la fraction de la nécromasse constituée par les arbres manifestement morts sur pied est mesurée en diamètre et hauteur. Le passage aux masses sèches s’effectue en utilisant les équations de régression et les infradensités des espèces vivantes.- herbacée: échantillonnage au plus fort développement;b) Rhizomasse: à titre indicatif, mention des valeurs de la littérature;c) Productivité primaire aérienne: évalua-tion de l’accroissement courant par mesure des cernes de croissance (carottes préle-vées à la tarière presssler) d’un échan-tillon par classe de diamètres.
7.2 Phytomasse aérienneLes caractéristiques dendrométriques
moyennes, converties en unités à l’hec-tare, sont présentées sur le tableau 3. L’in-ventaire intégral du nombre de tiges par diamètre et par espèce a été regroupé en classes dont les intervalles ont été réduits pour la catégorie arbustes. En distinguant l’espèce dominante Pinus sylvestris, cette présentation est très révélatrice de la struc-ture actuelle du peuplement. Cette struc-ture confirme à elle seule une caractéris-tique dynamique importante, déjà mani-feste à l’examen du tableau de végétation (cornali, 1997, tab. 1), à savoir qu’en dehors des lisières la régénération de Pinus sylvestris est quasi nulle: l’absence de semis est totale, et la présence d’arbustes tout à fait accidentelle. En revanche les autres espèces de faible diamètre, feuillus et Juniperus communis surtout, sont parti-culièrement abondantes par endroits, et ont créé depuis longtemps déjà des conditions d’ombrage défavorables au développement initial du pin.
Au niveau de la phytomasse aérienne totale indiquée sur la tableau 4, les pinèdes de la rive sud du lac de Neuchâtel peu-vent être créditées d’un ordre de gran-deur de 191 t/ha, dont l’espèce dominante Pinus sylvestris représente actuellement les 82%. Parmi les principales autres espèces il y a lieu de mentionner Picea abies (5,4%), Populus nigra (4,3%), Alnus glutinosa (1,9%), Prunus avium (1,3%), Alnus incana (1%). quant au bois mort sur pied, les espèces autres que Pinus sylves-tris (66%) sont principalement Juniperus communis (15%), Fraxinus excelsior (8%), Alnus incana (7%).
7.3 Chute, accumulation etdécomposition de la litière
7.3.1 Chute de litièreLes retombées de litière ont été récol-
tées d’octobre 1982 à novembre 1987 à l’aide d’un dispositif de paniers collec-
95
écoloGie Des pinèDes (pinus sylvestris) De la rive suD Du lac De neucHâtel (ii)
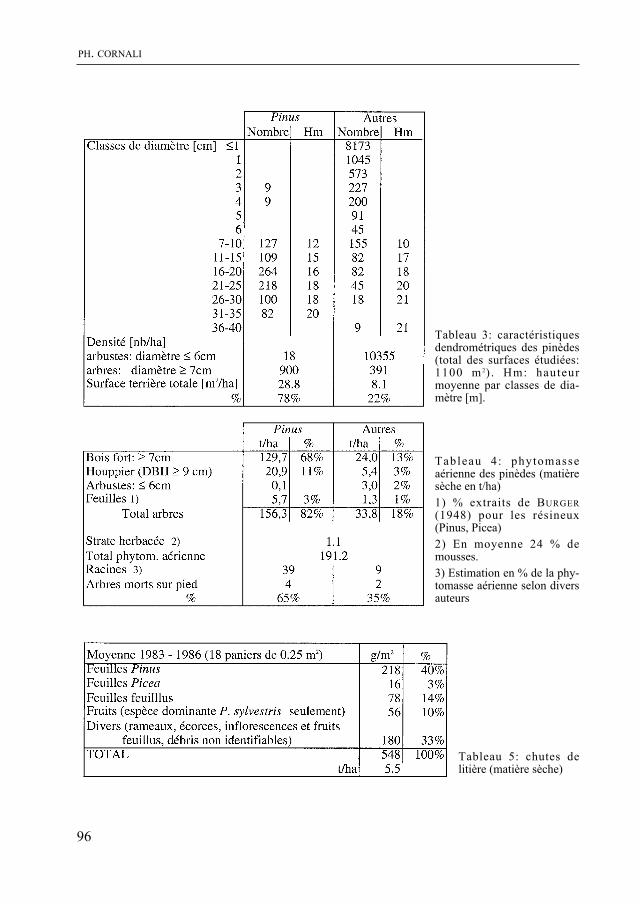
96
pH. cornali
Tableau 3: caractéristiques dendrométriques des pinèdes (total des surfaces étudiées: 1100 m 2) . Hm: hauteur moyenne par classes de dia-mètre [m].
Tableau 4 : phytomasse aérienne des pinèdes (matière sèche en t/ha)1) % extraits de burGer (1948) pour les résineux (Pinus, Picea)2) En moyenne 24 % de mousses.3) Estimation en % de la phy-tomasse aérienne selon divers auteurs
Tableau 5: chutes de litière (matière sèche)

97
teurs. Les récoltes ont été effectuées en relation avec le rythme saisonnier de la production végétale, tous les 2 à 3 mois pendant l’hiver et au printemps, tous les mois lors de la chute maximale, dès la fin de l’été jusqu’à l’arrière automne.
Le détail et la moyenne générale des chutes de litière pour les années 1983-1986 sont reportés dans le tableau 5. Calculées sur 4 ans, en considérant les stations comme homogènes et par consé-quent représentant un seul échantillon-nage, ces chutes présentent une moyenne de 549 g/m2 ou 5,5 t/ha.an. Les divers, à savoir 33%, se partagent en trois princi-pales sous-catégories qui ne figurent pas au tableau: rameaux et écorces 25%, inflo-rescences et fruits des espèces feuillues 4%, et enfin débris non identifiables 4%.
Sans qu’il y ait toujours matière à comparaison rigoureuse, nous pouvons mentionner chez quelques auteurs des totaux annuels très semblables aux 5,5 t/ha mesurés dans nos stations: 5,5 t/ha pour ovinGton (1959), 3-4 t/ha en pinède pure et 5-6 t/ha en peuplements mélangés (1/3 de hêtre en biomasse) pour Grünert (1964), 6 t/ha pour aussenac (1969).
7.3.2 Stock de matière organique au solAu début du mois d’octobre 1984, nous
avons récolté 12 échantillons de litière au sol. Seules deux placettes ont été échan-tillonnées, et les valeurs absolues ainsi que les proportions des différentes catégories
distinguées sont réunies dans le tableau 6. En raison des faibles masses considé-rées, mais aussi des difficultés à établir la différence entre mort et vivant, particuliè-rement en ce qui concerne les mousses, la strate herbacée a été entièrement assimilée à la matière organique morte. Pour cha-cune de ces catégories, la teneur en terre fine, impossible à séparer lors du tri, a été déterminée à l’aide de tests de lavage. Le tableau 6 présente des valeurs nettes, c’est-à-dire des valeurs brutes desquelles ont été soustraits les pourcentages moyens de terre fine suivants, assez constants: mousses 24%, herbes 8%, aiguilles 2%, litière non identifiée 35%, lignifié et fruits 3% (moyenne pondérée 8,5%).
Pour ce qui est de la masse totale de matière organique au sol, l’ordre de gran-deur de 12,3 t/ha peut être comparé aux 12,7 t/ha proposés par ovinGton (1954) ou aux 14,1 t/ha de GloaGuen & touffet (1980). En revanche, la littérature révèle aussi, s’agissant des pinèdes, une disper-sion considérable et assez surprenante qui peut varier de 25 à 85,5 t/ha chez eHwalD (1957), de 22 à 70 t/ha chez Grünert (1964), en passant par 45 t/ha pour ovinGton (1959) ou DuviGneauD (1980), et allant jusqu’à 110,6 t/ha pour ovinGton & MaGDwick (1959). Si cette comparaison sommaire n’apporte guère d’explications en dehors des indications classiques telles que contexte stationnel ou type d’exploi-tation, elle permet néanmoins de situer la rive sud du lac de Neuchâtel au bas de
écoloGie Des pinèDes (pinus sylvestris) De la rive suD Du lac De neucHâtel (ii)
Tableau 6: stock de matière organique au sol (matière sèche)

l’échelle pour ce qui est de l’accumula-tion au sein des horizons holorganiques en pinède. A chute de litière plus ou moins équivalente, il s’ensuit, comparativement, une vitesse de décomposition nettement plus rapide (voir 7.3.3). Sur le plan fonc-tionnel, ces conditions se rapprochent de celles des forêts caducifoliées (7-10 t/ha pour la rive sud du lac de Neuchâtel, cf. buecHe, à paraître, et chap. 9) et sont à mettre en relation avec la composition du groupement enrichie en espèces feuillues améliorantes. En termes d’évolution, ce constat renforce l’hypothèse d’un rem-placement possible, à moyen terme, de la pinède par la forêt feuillue (cf. cornali, 1997, chap. 4.4).
7.3.3 vitesse de décomposition et d’accumulation de la matière organiqueDeux méthodes complémentaires ont été
utilisées pour évaluer la vitesse de décom-position de la matière organique.
La première, proposée par Jenny et al. (1949), fait intervenir une constante annuelle de perte de poids (coefficient de Jenny) selon k’ = L / (L+ Xmin), où L repré-sente la chute annuelle de litière et Xmin le poids des horizons holorganiques avant la chute, pour la même unité de surface.
La deuxième consiste en la mise en décomposition d’échantillons disposés à l’intérieur de sachets en filet nylon eux-mêmes placés sous la couche de litière OL. Les sachets ont été remplis soit avec un mélange feuilles-aiguilles (pin sylvestre) récolté dans les paniers en station, et en proportions comparables à la moyenne des mois à forte chute, soit avec les mêmes essences, mais feuillus et résineux séparés.
L’intérêt d’une mise en oeuvre simul-tanée de ces deux méthodes est double. D’une part chacune permet d’évaluer de manière indépendante le taux de décom-position annuel de la matière organique. D’autre part, la méthode expérimentale de décomposition en filets permet de déter-miner un taux de décomposition instantané
dont la comparaison au taux annuel fournit une indication quant à l’état d’équilibre du système.
Pour les milieux forestiers, ces méthodes ont été appliquées par divers auteurs dont rapp (1971), leMee & bicHaut (1973), GloaGuen & touffet (1980), toujours en référence aux dévelop-pements théoriques proposés par deux pré-curseurs en la matière, Jenny et al. (1949) et olson (1963).
Ce dernier décrit un modèle général de type exponentiel pour l’évolution de la matière organique au sol, lequel intègre aussi bien l’accumulation avec chute annuelle de litière en fin de saison de croissance, que la décomposition en l’ab-sence de production (cas des filets). Les deux cas de figure obéissent aux équations suivantes:décomposition en filet: X/X0 = e-kt
accumulation au sol: X = (L/k’)(1-e-kt)avec:X: quantité de matière organique accu-mulée au temps t (années);X0: quantité de matière organique initiale;X/X0: fraction restante au temps t;L: chute annuelle de litière, théoriquement automnale;t: tempsk: taux de décomposition instantané, considéré constant, correspondant à la fraction du stock perdue par unité de temps (année ou autre);k’: taux de décomposition annuel avec chute automnale, initialement défini par Jenny:
k’ = 1-X/X0 = 1-e-kt
ce qui implique: k = -ln(1-k’)
A l’équilibre, l’apport compense la perte par décomposition:
L = k’(L+ Xmin)
98
pH. cornali
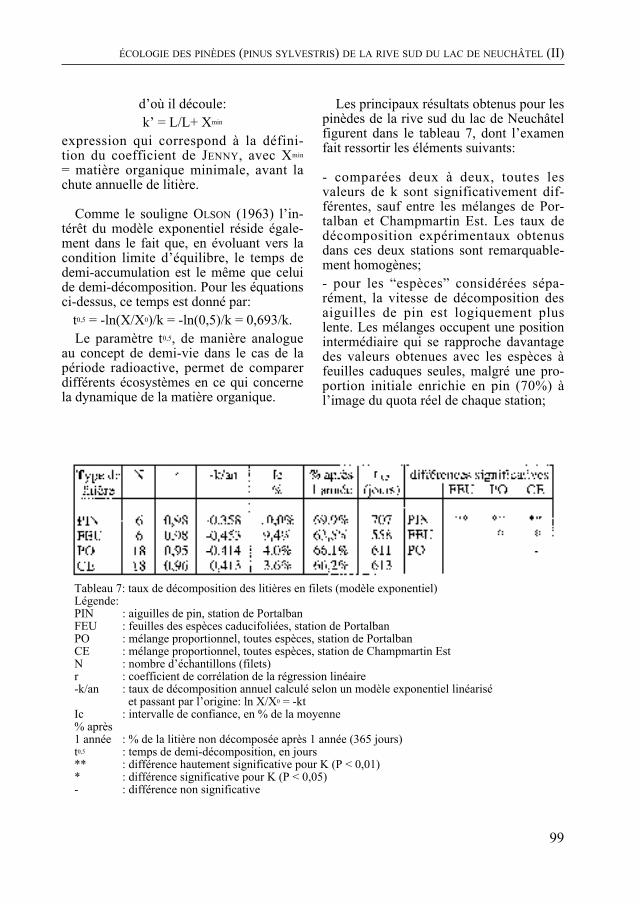
99
d’où il découle: k’ = L/L+ Xmin
expression qui correspond à la défini-tion du coefficient de Jenny, avec Xmin = matière organique minimale, avant la chute annuelle de litière.
Comme le souligne olson (1963) l’in-térêt du modèle exponentiel réside égale-ment dans le fait que, en évoluant vers la condition limite d’équilibre, le temps de demi-accumulation est le même que celui de demi-décomposition. Pour les équations ci-dessus, ce temps est donné par:
t0,5 = -ln(X/X0)/k = -ln(0,5)/k = 0,693/k.Le paramètre t0,5, de manière analogue
au concept de demi-vie dans le cas de la période radioactive, permet de comparer différents écosystèmes en ce qui concerne la dynamique de la matière organique.
Les principaux résultats obtenus pour les pinèdes de la rive sud du lac de Neuchâtel figurent dans le tableau 7, dont l’examen fait ressortir les éléments suivants:
- comparées deux à deux, toutes les valeurs de k sont significativement dif-férentes, sauf entre les mélanges de Por-talban et Champmartin Est. Les taux de décomposition expérimentaux obtenus dans ces deux stations sont remarquable-ment homogènes;- pour les “espèces” considérées sépa-rément, la vitesse de décomposition des aiguilles de pin est logiquement plus lente. Les mélanges occupent une position intermédiaire qui se rapproche davantage des valeurs obtenues avec les espèces à feuilles caduques seules, malgré une pro-portion initiale enrichie en pin (70%) à l’image du quota réel de chaque station;
écoloGie Des pinèDes (pinus sylvestris) De la rive suD Du lac De neucHâtel (ii)
Tableau 7: taux de décomposition des litières en filets (modèle exponentiel)Légende:PIN : aiguilles de pin, station de PortalbanFEU : feuilles des espèces caducifoliées, station de PortalbanPO : mélange proportionnel, toutes espèces, station de PortalbanCE : mélange proportionnel, toutes espèces, station de Champmartin EstN : nombre d’échantillons (filets)r : coefficient de corrélation de la régression linéaire-k/an : taux de décomposition annuel calculé selon un modèle exponentiel linéarisé et passant par l’origine: ln X/X0 = -ktIc : intervalle de confiance, en % de la moyenne% après 1 année : % de la litière non décomposée après 1 année (365 jours)t0,5 : temps de demi-décomposition, en jours** : différence hautement significative pour K (P < 0,01)* : différence significative pour K (P < 0,05)- : différence non significative

- le taux de décomposition instantané k est lié au coefficient de Jenny k’ par la rela-tion (voir plus haut):
k’ = 1-e-kt
dont l’application aux mélanges des 2 sta-tions (k = 0,414) fournit, pour t = 1 année:
k’ = 1-e-0,414 = 0,339Le coefficient de Jenny, nous l’avons
déjà mentionné, s’obtient également d’une tout autre manière: k’ = L / (L+ Xmin), où L représente la chute annuelle de litière et Xmin le poids des horizons holorganiques avant la chute. Pour l’année 1984, avec un stock au sol (récolté en octobre) diminué des chutes de litière du mois précédent, nous obtenons, sans tenir compte de la strate herbacée, une valeur moyenne de k’ = 0,352, proche de celle calculée à l’aide du taux de décomposition en filet. Il ne s’agit toutefois que d’un ordre de gran-deur dont l’évaluation peut être entachée d’imprécisions d’origines diverses (condi-tions de décomposition en filet éloignées des modalités naturelles, chute annuelle de litière théoriquement unique, ici étalée sur 2 à 3 mois). malgré ces réserves, le modèle exponentiel utilisé reste manifeste-ment bien adapté.
En termes d’accumulation, nous pou-vons également l’utiliser dans le but d’éva-luer la durée nécessaire pour atteindre l’équilibre annuel accumulation = décom-position. En utilisant la forme:
X = (L/k’)(1-e-kt) on tire: t = ln(1-X/(L/k’))/k
avec L/k’ = L+Xmin = valeur maximale annuelle.En fixant cette dernière à 99% de la valeur à l’équilibre, il vient:
t99% = -ln(1-0,99)/k = 4,6/ket finalement
t99% = 4,6/0,414 = 11 ans.Il s’agit là d’une durée très brève, com-
parée par exemple à la limite d’un siècle au minimum indiquée par Jenny et al.
(1949) pour la stabilisation des horizons holorganiques en forêt de pin. Plus près de nous pourtant, GloaGuen & touffet (1980) mentionnent un coefficient de Jenny de k’ = 0,318 pour les aiguilles de pin sylvestre seules (d’où k = 0,383 et t 99% = 12 ans) très semblable au nôtre. De même, en première estimation, Guittet (1967) évalue à 10 ans le temps néces-saire pour la transformation en humus d’aiguilles de Pinus sylvestris à partir de leur chute, en milieu ouvert sur substrat calcaire.
Il se confirme donc que le renouvelle-ment de la matière organique des pinèdes de la rive sud du lac de Neuchâtel est rela-tivement rapide. En dehors de conditions climatiques et édaphiques favorables, cette rapidité est notamment influencée par la composition hétérogène des pinèdes: même en quantités réduites (en moyenne 20% de la biomasse), les espèces feuillues améliorent notablement les processus de décomposition (tab. 7).
Par ailleurs, le fait que le k’ obtenu à partir de la mesure de k (décomposition instantanée en filet) soit sensiblement égal au k’ annuel de Jenny permet de consi-dérer l’état d’équilibre entre accumulation et décomposition comme pratiquement atteint.
7.4 Croissance et productivité aériennesAu niveau aérien, la productivité pri-
maire nette correspond théoriquement à l’addition de plusieurs termes (Duvi-GneauD, 1980):
PN1 = T1 + m1n + C2n + Ex1nsoit T1: incrément ligneuxm1n: tissus morts non consommésC2n, Ex1n: consommation respectivement exportation de tissus nouvellement formés durant l’unité de temps.
En négligeant les deux derniers termes, l’évaluation de PN1 peut être simplifiée en:
100
pH. cornali

101
PN1 = T1 + m1nL’évaluation de m1n présente encore
de grosses difficultés puisqu’elle inclut le bois mort annuellement produit mais pas forcément tombé. Là encore nous avons dû simplifier en assimilant les tissus morts non consommés à la chute de litière. De plus, en considérant cette dernière comme la somme du matériel récolté dans les paniers additionnée de la strate herbacée en fin d’année, nous avons négligé la chute des grosses branches. Cette sous-estima-tion a été très partiellement compensée par la prise en compte de la production moyenne du bois mort sur pied dès l’ori-gine du peuplement. Nous avons finale-ment retenu pour le calcul de PN1:PN1 = incrément ligneux + chute de litière (y.c. herbacée) + bois mort sur pied
quelle que soit la manière d’envi-sager la productivité primaire en milieu forestier, il est donc nécessaire d’avoir recours à l’incrément radial par unité de temps, c’est-à-dire à l’examen des cernes de croissance. Passage obligé, l’approche dendrochronologique permet en outre de multiples applications, clairement expo-sées par Munaut (1978).
Nos propres mesures concernent une soixantaine d’échantillons représentant au total 32% des pins et 45% des individus feuillus de diamètre ≥ 5 cm rencontrés dans les trois stations. Riche d’informa-
tions, la lecture des résultats peut être exploitée de trois manières visant chacune à compléter l’évaluation de la dynamique du système.
A) Age des pinèdesLes valeurs mesurées, réunies sur
le tableau 8, permettent d’évaluer l’âge moyen des pinèdes à environ 48 ans (± 3, à P = 0,01) pour l’ensemble échantillonné. Il s’agit de populations sensiblement équiennes comme l’indiquent les inter-valles de confiance.
Alors que le calcul de la moyenne permet surtout de préciser la dispersion des données, l’examen du tableau fait ressortir le caractère pionnier du pin syl-vestre, dont l’implantation progressive s’amorce entre 42 et 55 ans avant 1987, année des mesures, soit entre 1932 et 1945. Cette époque se situe environ 60 ans (entre 54 et 67 ans) après l’abaissement des eaux dû à la première correction des eaux du Jura, effectif dès 1878 (Müller, 1973).
Toujours sur le plan dynamique, l’ab-sence actuelle de jeunes pins est large-ment confirmée (cf. 7.2), quel que soit le diamètre considéré. En majorité plus récentes, mais aussi d’âges plus variés, les autres espèces ont peu à peu complété la colonisation de ces milieux ouverts sur sols très superficiels. A quelques excep-
écoloGie Des pinèDes (pinus sylvestris) De la rive suD Du lac De neucHâtel (ii)
Tableau 8: âge des pinèdes échantillonné à la tarière pressler

tions près, la compétition initiale s’est donc révélée favorable au pin sylvestre, du moins jusqu’à ce que le développement progressif du couvert en élimine les semis, très exigeants en lumière.
B) Dynamique par espèceL’examen des cernes de croissance
fournit de plus amples informations en ce qui concerne l’autoécologie des espèces. En effet les variations climatiques, principa-lement la température et les précipitations durant la période de croissance annuelle, influencent l’épaisseur et la densité des cernes, lesquels constituent en outre un bon indice pour juger de la vitalité d’un arbre (lenz. et al., 1988). Cette réponse est spéci-fique, et l’absorption de la variabilité indivi-duelle et microstationnelle nécessite la prise en compte d’un minimum empirique d’une dizaine d’individus par espèce (Munaut, 1978). Pour faciliter la comparaison de valeurs absolues parfois très différentes, les dendrochronologues ont introduit une repré-sentation graphique sur laquelle les années figurent en abscisse, alors que l’épaisseur des cernes est portée en ordonnée suivant une échelle logarithmique.
Nous avons reproduit de la sorte, sur la figure 3, l’évolution moyenne de l’épais-seur des cernes mesurée sur un échan-tillon de pins dans chaque station (32 mesures au total). A titre indicatif, nous y avons adjoint deux exemples de com-portement extrême, Picea et Juniperus, valeurs moyennes de respectivement 6 et 5 mesures. Bien qu’en toute rigueur ces deux nombres de mesures soient trop faibles, les valeurs individuelles pour chaque espèce sont suffisamment groupées pour que la tendance générale puisse être mentionnée. La pluviosité à Witzwil complète la figure, qui ne concerne ici que les 28 dernières années, de manière à éviter de prendre en compte les échantillons au début de leur période de croissance. Les mois retenus, avril à juillet, correspondent à l’essentiel
de la période de croissance annuelle. Ce graphe permet de constater que:- la reproductibilité des rythmes de crois-sance du pin sylvestre est très satisfaisante d’une station à l’autre;
- pour cette essence, l’épaisseur moyenne des cernes est en diminution, marquant par là un tassement progressif de la producti-vité (voir ci-après);
- l’épicéa en revanche présente une évolu-tion plutôt stable, alors que les genévriers échantillonnés, conformément à ce qui est observable en station, paraissent manifes-tement condamnés;
- la mise en parallèle de la pluviomé-trie permet de faire ressortir l’influence prépondérante de ce facteur. Les années sèches caractéristiques telles que 1962, 1964, 1976 ou 1979 sont parfaitement marquées au niveau des cernes de crois-sance. Ces années sont également repérées en tant qu’exceptionnelles dans l’étude sur le dépérissement du sapin blanc dans la région de Berne réalisée par lenz et al. (1988). mais, comme le notent ces auteurs, la causalité est souvent plus complexe et fait bien sûr intervenir la température, voire la saison, de telle sorte que l’ac-tion conjuguée de plusieurs facteurs est souvent nécessaire pour caractériser une année exceptionnelle. Ainsi par exemple la relative sécheresse de 1967 ne s’est guère traduite par une diminution de l’épaisseur des cernes.
Dans le but de confirmer l’existence de conditions originelles impropres à l’instal-lation de la pinède, nous avons également analysé des cernes de quelques échan-tillons de Pinus sylvestris le long d’un transect incluant une portion de pinède en microtopographie surélevée et les marais adjacents (site de Champmartin). Les mesures moyennes divergent de manière éloquente entre marais et pinède (y com-pris lisières), soit respectivement 8 cm
102
pH. cornali
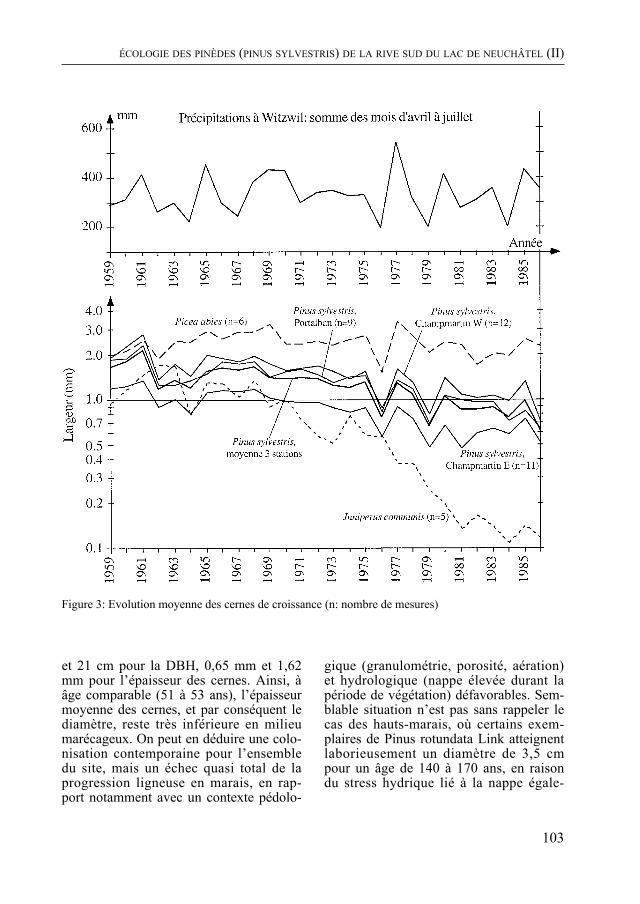
103
écoloGie Des pinèDes (pinus sylvestris) De la rive suD Du lac De neucHâtel (ii)
Figure 3: Evolution moyenne des cernes de croissance (n: nombre de mesures)
et 21 cm pour la DBH, 0,65 mm et 1,62 mm pour l’épaisseur des cernes. Ainsi, à âge comparable (51 à 53 ans), l’épaisseur moyenne des cernes, et par conséquent le diamètre, reste très inférieure en milieu marécageux. On peut en déduire une colo-nisation contemporaine pour l’ensemble du site, mais un échec quasi total de la progression ligneuse en marais, en rap-port notamment avec un contexte pédolo-
gique (granulométrie, porosité, aération) et hydrologique (nappe élevée durant la période de végétation) défavorables. Sem-blable situation n’est pas sans rappeler le cas des hauts-marais, où certains exem-plaires de Pinus rotundata Link atteignent laborieusement un diamètre de 3,5 cm pour un âge de 140 à 170 ans, en raison du stress hydrique lié à la nappe égale-
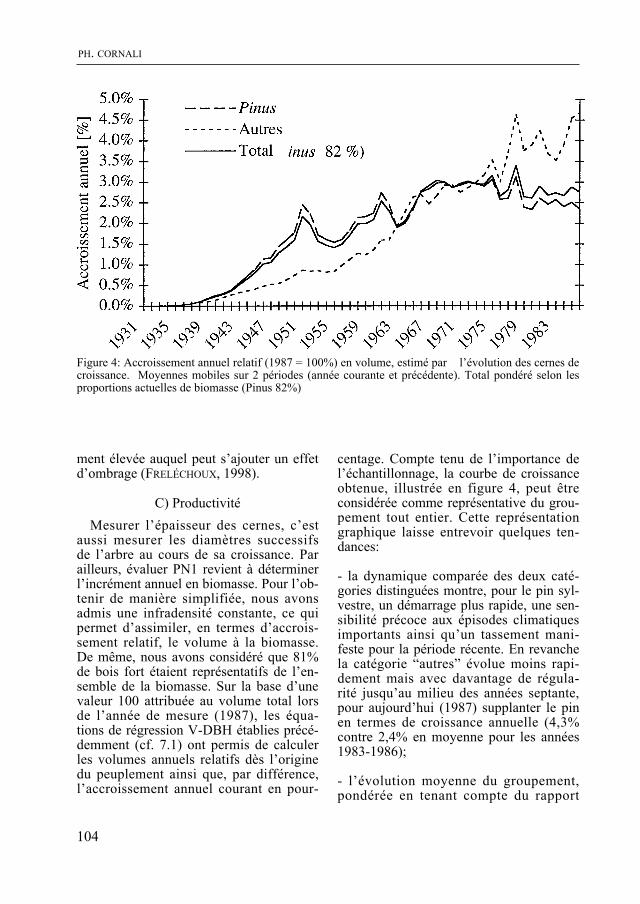
104
ment élevée auquel peut s’ajouter un effet d’ombrage (frelécHoux, 1998).
C) Productivitémesurer l’épaisseur des cernes, c’est
aussi mesurer les diamètres successifs de l’arbre au cours de sa croissance. Par ailleurs, évaluer PN1 revient à déterminer l’incrément annuel en biomasse. Pour l’ob-tenir de manière simplifiée, nous avons admis une infradensité constante, ce qui permet d’assimiler, en termes d’accrois-sement relatif, le volume à la biomasse. De même, nous avons considéré que 81% de bois fort étaient représentatifs de l’en-semble de la biomasse. Sur la base d’une valeur 100 attribuée au volume total lors de l’année de mesure (1987), les équa-tions de régression v-DBH établies précé-demment (cf. 7.1) ont permis de calculer les volumes annuels relatifs dès l’origine du peuplement ainsi que, par différence, l’accroissement annuel courant en pour-
centage. Compte tenu de l’importance de l’échantillonnage, la courbe de croissance obtenue, illustrée en figure 4, peut être considérée comme représentative du grou-pement tout entier. Cette représentation graphique laisse entrevoir quelques ten-dances:
- la dynamique comparée des deux caté-gories distinguées montre, pour le pin syl-vestre, un démarrage plus rapide, une sen-sibilité précoce aux épisodes climatiques importants ainsi qu’un tassement mani-feste pour la période récente. En revanche la catégorie “autres” évolue moins rapi-dement mais avec davantage de régula-rité jusqu’au milieu des années septante, pour aujourd’hui (1987) supplanter le pin en termes de croissance annuelle (4,3% contre 2,4% en moyenne pour les années 1983-1986);
- l’évolution moyenne du groupement, pondérée en tenant compte du rapport
pH. cornali
Figure 4: Accroissement annuel relatif (1987 = 100%) en volume, estimé par l’évolution des cernes de croissance. moyennes mobiles sur 2 périodes (année courante et précédente). Total pondéré selon les proportions actuelles de biomasse (Pinus 82%)

actuel des biomasses (82% de pin) indique une relative stabilisation de la producti-vité. Il s’agit probablement ici du seuil précédant la diminution classique de PN1 lorsque le système s’achemine vers le stade climax (cf. DuviGneauD, 1980, fig. 4.36). Les peuplements de pin sylvestre sont en effet susceptibles d’atteindre rapi-dement ce maximum, avant 30 ans même dans les plantations étudiées par ovinGton (1957).
Les réponses climatiques déjà visibles avec l’évolution de l’épaisseur des cernes (cf. fig. 3) sont naturellement maintenues.
Nous pouvons maintenant déterminer l’incrément ligneux nécessaire au calcul de PN1 en appliquant à la biomasse sur pied, soit 183 t/ha sans les feuilles (cf. tab. 4), le taux d’accroissement courant des années 1983-1986, en moyenne 2,72%:
183 t/ha x 2,72%/an = 5,0 t/ha.anA titre indicatif, en considérant un âge
moyen de 48 ans, l’accroissement moyen correspondant est de 3,8 t/ha.an, auquel on peut ajouter 0,13 t/ha.an de bois mort sur pied (3,3% de cet accroissement moyen), soit environ 4 t/ha.an.Nous obtenons finalement pour PN1:
PN1 = incrément ligneux + chute de litière + lit. herbacée + bois mort sur pied
= 5,0 + 5,5 + 1,1 + 0,2 = 11,8 t/ha.anCette évaluation de PN1 à environ 12 t/
ha.an est minimale dans la mesure où la chute des grosses branches est pratique-ment négligée.
8.oriGine et évolution Du GroupeMent
Déjà évoquée en considérant le contexte phytosociologique, pédologique et hydro-dynamique (cornali, 1997), la dynamique évolutive du groupement peut être reconsi-dérée à la lumière des précisions apportées par l’étude de la phytomasse et producti-vité aériennes.
Il résulte de nos observations que le pin s’est installé en pionnier, parfois dans des taillis comme le suggère berset (1949/50), en un laps de temps assez res-treint d’une quinzaine d’années. C’était il y a 50 ans, c’est-à-dire environ 60 ans après l’exondation consécutive à la pre-mière correction des eaux du Jura. Il a colonisé avec succès les zones légère-ment surélevées (placages morainiques) du substrat molassique, mais a échoué dans les dépressions marécageuses beau-coup plus inondées. D’autres espèces, en particulier feuillues, ont progressivement complété le couvert forestier, empêchant bientôt la régénération des semis de pin.
La phytomasse aérienne totale actuelle atteint environ 190 t/ha, dont 82% de pins âgés de 48 (± 3) ans. La reconstitution de l’évolution du peuplement grâce à l’examen des cernes de croissance révèle aujourd’hui une productivité primaire nette PN1 stabilisée à 12 t/ha.an, dont approxi-mativement 1 tonne de litière herbacée, le solde se partageant pour moitié entre incrément ligneux et chute de litière. Sur le plan fonctionnel, cette stabilisation de PN1 précède le stade climax, dont il y a tout lieu de penser qu’il est sur le point d’être atteint en raison d’un quasi équilibre entre accumulation et décomposition de la litière.
Plus difficile reste la prévision concer-nant l’évolution future du peuplement. Les âges respectifs aussi bien que la densité en individus à l’hectare (rapport de 1:500 en faveur des autres espèces que le pin pour les diamètres ≤ 6 cm), ou encore les tendances moyennes manifestées par les cernes de croissance, tout semble indiquer une dynamique individuelle divergente des espèces. Alors que les pins sont en légère régression, la majorité des autres espèces est plutôt stable ou en progression. En tout état de cause, rien n’indique l’in-dispensable diminution du couvert néces-saire à la régénération du pin, même si quelques espèces semblent aujourd’hui
105
écoloGie Des pinèDes (pinus sylvestris) De la rive suD Du lac De neucHâtel (ii)

mal supporter leur situation (par exemple Juniperus). Dans cette perspective on peut envisager, hypothèse à moyen terme, le remplacement progressif de la pinède par la forêt feuillue en raison non seule-ment des difficultés de régénération évo-quées, mais aussi grâce aux modifications des conditions pédologiques permettant de plus en plus aux concurrents de cette essence d’être compétitifs, ce qui n’était pas le cas à l’origine du peuplement. De ce fait, le terme climax qui vient d’être évoqué doit être compris davantage dans un sens écologique (climax “fonctionnel”) que dans une acception purement phytoso-ciologique.
9. potentialités fonctionnelles et conclusion
Afin de situer l’écosystème pinède de la rive sud du lac de Neuchâtel dans le cadre plus large de l’hémisphère nord, nous avons confronté ses principales caractéristiques quantitatives à la classification de roDin & bazilevitcH (1967) (cf. annexe 1) men-tionnée en annexe par rapp (1971). Ces auteurs proposent une quantification des principaux paramètres des cycles biogéo-chimiques selon une échelle à 10 classes (classe 1: valeurs faibles, classe 10: valeurs très élevées). Par rapport à cette classification, l’écosystème pinède de la rive sud du lac de Neuchâtel prend place dans les classes suivantes:
- tous les paramètres concernant la matière organique coïncident avec les classes 5-7 caractérisant des groupements moyenne-ment productifs. Dans cette catégorie, 190 t/ha de biomasse, 12 t/ha.an pour PN1 et 5,5 t/ha.an d’accroissement annuel sont plutôt élevés (classe 7). Le facteur de décomposition de 34-41% est quant à lui moyen (classe 6), alors que 5,5 t/ha.an de retombées de litière, mais surtout 12 t/ha pour l’horizon holorganique peuvent être considérés comme faibles (classe 5);
- pour les éléments minéraux, dont l’étude détaillée fera l’objet d’un troisième article, les paramètres sont davantage dispersés. La minéralomasse de 1314 kg/ha est qua-lifiée d’importante (classe 6) alors qu’une absorption de 192 kg/ha et une fixation annuelle de 30 kg/ha sont moyennes (classe 4). Enfin 188 kg/ha bloqués dans l’horizon holorganique et 122 kg/ha dans la litière (sans la strate herbacée), ainsi qu’une teneur minérale moyenne de 2,21% pour cette même litière, sont faibles (classe 3).
Pour une chute de litière assez impor-tante (6,6 t/ha y compris la strate her-bacée), la faible accumulation au sein des horizons holorganiques (11,3 t/ha) traduit une vitesse de décomposition élevée de la matière organique, du moins pour une pinède. En effet dans la classification de roDin & bazilevicH (op. cit.) les valeurs ci-dessus correspondent effectivement au type pinède pour biomasse et retombées de litière, mais traduisent nettement les forêts caducifoliées pour ce qui est de la teneur moyenne en éléments minéraux de la litière et surtout la vitesse de décompo-sition de la matière organique. Il s’ensuit une sorte de décalage fonctionnel dans la dynamique productive du groupement (analogue au décalage morpho-fonctionnel de certains sols de la rive sud du lac de Neuchâtel, buttler & Gobat, 1991), favorisé notamment par la présence d’es-pèces feuillues améliorantes. En d’autres termes, ce décalage montre que le fonc-tionnement actuel, déjà orienté du côté de la forêt feuillue, anticipe sur les structures physionomique et phytosociologique, qui restent encore celles d’une pinède. Cette situation renforce l’hypothèse d’un rem-placement progressif de la pinède par les espèces ligneuses actuellement com-pagnes. A titre d’exemple, wooDwell & wHittaker (1968) rendent compte d’un tel scénario (succession champ - pinède - forêt climacique feuillue) de manière parfaitement concordante à la dynamique
106
pH. cornali

107
observée sur la rive sud du lac de Neu-châtel pour les 60 premières années.
10. résuMé De la DeuxièMe partie
La phytomasse aérienne totale des pinèdes de la rive sud du lac de Neuchâtel avoisine 191 t/ha, dont l’espèce dominante Pinus sylvestris représente actuellement les 82%. La mesure des cernes de crois-sance a permis d’évaluer une producti-vité primaire nette PN1 stabilisée à 12 t/ha.an, compte tenu de l’apport de 1,1 t/ha par la strate herbacée, ainsi qu’un incré-ment ligneux courant de 5 t/ha.an. Sans la strate herbacée, les chutes de litière cor-respondent en moyenne à 5,5 t/ha.an. Bien qu’ils présentent d’importantes variations spatiales, les horizons holorganiques en place contiennent 11,3 t/ha de matière organique. Evaluée par deux méthodes, la vitesse de décomposition de la litière implique une libération annuelle d’environ 35% de la quantité initiale, c’est-à-dire 6,3 t/ha, ou encore 0,3 t/ha de moins que les chutes de litière, strate herbacée comprise. Le faible écart entre chute et décomposi-tion signifie une accumulation annuelle
théoriquement proche d’un équilibre dynamique entre les deux processus. Le renouvellement très rapide de la matière organique, du moins pour une pinède, est dû aux espèces feuillues dont la présence améliore notablement les processus de décomposition.
La dynamique de croissance de la pinède indique aujourd’hui une stabilisa-tion de la productivité. A moyen terme, plusieurs facteurs, parmi lesquels un déca-lage fonctionnel dans la dynamique pro-ductive du groupement, permettent d’en-visager l’hypothèse d’un remplacement progressif de la pinède par les espèces ligneuses actuellement compagnes.
reMercieMents
Je tiens à exprimer mes remerciements et toute ma gratitude aux personnes sans lesquelles le travail de thèse qui est à l’ori-gine de cette présentation n’aurait jamais pu être mené à bien. Il s’agit en particu-lier de J.-L. Richard, J.-m. Gobat, J.-D. Gallandat, J.-C. védy, m. Gratier, J.-P. Dubois, A. Buttler et m. Bueche.
écoloGie Des pinèDes (pinus sylvestris) De la rive suD Du lac De neucHâtel (ii)
biblioGrapHie
aussenac, G. 1969. Production de litière dans divers peuplements forestiers de l’est de la France. Oecol. Plant. 4: 225-236.
berset, J. 1949/50. La végétation de la réserve de Cheyres et des rives avoisinantes du lac de Neu-châtel. Bull. Soc. Frib. Sc. Nat. 40: 65-94.
buecHe, m. (A paraître). Ecologie des forêts feuillues RSLN. Thèse de doctorat. Institut de Botanique, Université de Neuchâtel.
burGer, H. 1948. Holz, Blattmenge und Zuwachs. 9. mitteilung: die Föhre. mitteilungen der schwei-zerrischen Anstalt für das forstliche versuchswesen 25: 435-493.
buttler, a. & Gobat, J.-M. 1991. Les sols hydromorphes des prairies humides de la rive sud du lac de Neuchâtel (Suisse). Bull. Ecol. 22 (3-4): 405-418.
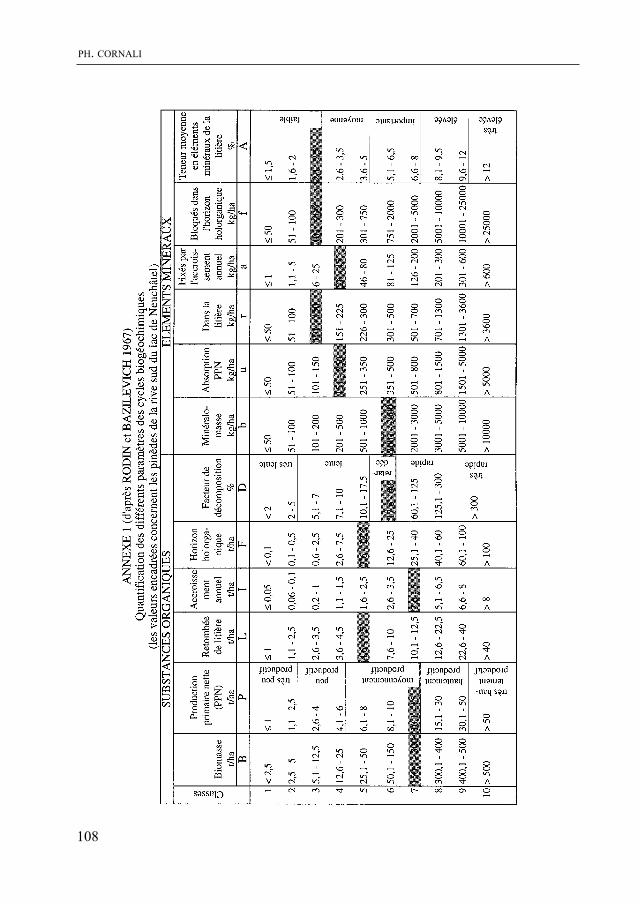
pH. cornali
108

109
cornali, p. 1992. Ecologie des pinèdes (Pinus sylvestris) de la rive sud du lac de Neuchâtel (Suisse): phytosociologie, pédologie, hydrodynamique. Thèse de doctorat. Institut de Botanique, Université de Neuchâtel.
cornali, P. 1997. Ecologie des pinèdes (Pinus sylvestris) de la rive sud du lac de Neuchâtel (Suisse). I Phytosociologie, pédologie, hydrodynamique. Bull. Soc. neuchâtel. Sci. nat. 120: 55-72.
DuviGneauD, P. 1971. Concepts sur la productivité primaire des écosystèmes forestiers. In “Producti-vité des écosystèmes forestiers”. Paris, UNESCO, Ecol. et Cons. 4: 110-140.
DuviGneauD, P. 1980. La synthèse écologique.. Doin, Paris.
DuviGneauD, P. 1984. L’écosystème forêt. Ecole nationale du génie rural, des eaux et forêts, Nancy.
eHwalD, E. 1957. Über den Nährstoffkreislauf des Waldes. S.B. dtsch. Akad.Wiss., Berlin, vol. 6 (1): 3-56.
frelécHoux, F. 1998. Étude du boisement des tourbières hautes de la chaîne jurassienne: typologie et dynamique de la végétation - approche dendroécologique et dendrodynamique des peuplements arborescents. Thèse de doctorat. Université de Neuchâtel.
GloaGuen, J.-C. & touffet, J. 1980. vitesse de décomposition et évolution minérale des litières sous climat atlantique. I. Le Hêtre et quelques Conifères. Acta Oecologica, Oecol. Plant., 1 (15), N° 1: 3-26.
Grünert, F. 1964. Der biologische Stoffkreislauf in Kiefern-Buchen-mischbeständen und Kiefernbes-tänden. Albrecht-Thaer-Archiv, archiv 8 (6/7): 435-452.
Guittet, J. 1967. Composition et évolution de la litière de pins sylvestres en peuplements ouverts sur pelouse xérophile. Oecol. Plant. 2: 43-62.
Hilfstafeln für Die forsteinricHtunG 1966. Bayerischen Staatsministerium für Ernährung, Land-wirtschaft und Forsten ministerialforstabteilung.
Jenny, H., Gessel, s.p. & binGHaM, f.t. 1949. Comparative study of decomposition rates of organic matter in temperate and tropical regions. Soil. Sc. 2: 419-432.
leMee, G. 1974. La productivité primaire de la forêt. In “Ecologie forestière” pesson, P., Gauthier-vil-lars, Paris: 135-153.
leMee, G. & bicHaut, N. 1973. Recherches sur les écosystèmes des réserves biologiques de la forêt de Fontainebleau. II. Décomposition de la litière de feuilles des arbres et libération des bioéléments. Oecol. Plant. 8 (2): 153-174.
lenz, o., noGler, p. & bräker, o. U. 1988. L’évolution du temps et le dépérissement du Sapin blanc dans la région de Berne. Institut fédéral de recherches forestières, Birmensdorf, rapport 303.
Müller, R. 1973. Les niveaux des lacs du Jura. In: scHwab, H., (1973): Die vergangenheit des See-landes in neuem Licht. Ed. Universitaires, Fribourg: 155-176.
Munaut, A.v. 1978. La dendrochronologie. Une synthèse de ses méthodes et applications. Lejeunia No 91, Liège.
olson, J.S. 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 44: 322-331.
écoloGie Des pinèDes (pinus sylvestris) De la rive suD Du lac De neucHâtel (ii)

110
ovinGton, J.D. 1954. Studies of the developement of woodland conditions under different trees. II. The forest floor. J. Ecol. 42: 71-80.
ovinGton, J.D. 1957. Dry matter production of Pinus sylvestris L. Annals of Botany 21: 287-314.
ovinGton, J.D. 1959. The Circulation of minerals in Plantations of Pinus sylvestris L. Annals of Botany, N.S. 23, No 90: 229-239.
ovinGton, J.D. & MaDGwick, H.A.I. 1959. Distribution of Organic matter and Plant Nutrients in a Plantation of Scots Pine. Forest Science 5 (4): 344-355.
parDe, J. 1979. Les biomasses forestières. I.N.R.A., Station de Sylviculture et de Production du C.N.R.F. Champenoux, 54280 Seichamps.
rapp, m. 1971. Cycle de la matière organique et des éléments minéraux dans quelques écosystèmes méditerranéens. Ed. Centre National de la Rech. Scient., Paris.
rieDacker, A. 1968 a. méthodes d’estimation de la biomasse d’un arbre. D.E.A. Biol. vég., Faculté d’Orsay.
rieDacker, A. 1968 b. méthodes indirectes d’estimation de la biomasse des arbres et des peuplements forestiers. Doc. int. Stn. sylvic. prod. C.N.R.F.
roDin, L.E. & bazilevicH N.I. 1967. Production and mineral cycling in terrestrial vegetation. Olivier and Boyd, Edinburgh.
wooDwell, G.M. & wHittaker, R.H. 1968. Primary production in terrestrial communities. Amer. Zool. 8: 19-30.
__________
pH. cornali
bull. soc. neucHâtel. sci. nat. 121 : 93-110. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 111-125. 1998
111
ÉvOLUTION DES POINTS D’EAU DANS LA TOURBIÈRE DU CACHOT
I. mORPHOLOGIE ET vÉGÉTATION DES GOUILLES
WILLy mATTHEy
Institut de Botanique de l’Université, rue Emile-Argand 11, CH-2000 Neuchâtel, Suisse.
mots-clés: Tourbière, gouille, butte, évolution naturelle, piétinement.
Key-words: Peat bog - hollow - hummock - natural evolution - trampling.
Résumé
43 gouilles de la tourbière du Cachot (canton de Neuchâtel, Suisse, coordonnées 541200 206350), ont été cartographiées en 1969. Un nouveau recensement effectué en 1998 met en évidence une diminution de leur surface de 176 m2 sur un total de 309 m2 , soit d’environ 57%. Le développe-ment des buttes du Sphagnetum magellanici a entraîné le comblement de 19 gouilles et le rétré-cissement de la plupart des 24 survivantes. Les effets à long terme (30 ans) du piétinement sur les sphaignes sont examinés. Cinq exemples sont présentés plus en détail.
Summary
In 1969, 43 hollows were recorded in the peat bog of le Cachot (Jura mountains, Switzerland, coordinates 541200 206350). A new survey has been made in 1998, showing that the total area covered by these hollows had grown 176 m2 smaller, that is 57%. Growing hummocks of Sphagnum mosses actually have filled 19 hollows and reduced the size of most of the remaining 24.
In this article, we study also the long term consequences of trampling on the hollow banks, and describe five exemples in detail.
introDuction
De nombreuses études ont été menées dans les tourbières du Haut-Jura. En zoologie, plusieurs d’entre elles le furent dans la tourbière du Cachot ou du Bas-du-Cerneux (tab. 1) au cours des 35 dernières années. Elles ont porté essentiellement sur l’écologie des Arthropodes, la définition de leurs niches écologiques et la structure de leurs peuple-ments. Une carte phytosociologique a servi de support à ces recherches. Etablie par Mat-tHey (1964 et 1971), elle a été complétée et améliorée à plusieurs reprises (auroi, 1975; GeiGer, 1978; neet, 1984; borcarD, 1988; von ballMoos, 1989 et vaucHer von ballMoos, 1997). Dans ce cadre, toutes les gouilles du haut-marais ont été répertoriées, mesurées et cartographiées. Il nous a semblé intéressant, trois décennies après le début

112
des travaux, de reprendre cet inventaire afin de mesurer l’évolution naturelle ou non de ces points d’eau si particuliers. De telles observations ont peu été publiées jusqu’ici sur les tourbières; elles consti-tuent pourtant des documents précieux pour définir les stratégies de conserva-tion, et aussi dans l’optique de recherches que cet aspect appliqué de l’écologie peut engendrer (MattHey, y., 1993; Grosver-nier, 1996). Dans le cas de la tourbière du Cachot, elles fourniront quelques points de repère aux botanistes qui reprendront peut-être cet inventaire vers 2030 !
2. MoDifications subies par la tourbière Du cacHot Dans son
enseMble
L’observation continue de plusieurs stations sur près de trois décennies (ce qui est toutefois bien court par rapport aux quelque 10000 ans de vie de la tour-bière) permet de constater qu’un certain nombre de transformations en ont passa-blement altéré le caractère (MattHey & borcarD 1996). Peu visibles en deux ou
trois ans, elles deviennent évidentes sur une trentaine d’années. Actuellement, les atteintes anthropiques directes ont pra-tiquement cessé, mais elles influencent encore l’hydrologie du massif tourbeux et la dynamique de la végétation. En particu-lier, le drainage et la mise en culture des zones avoisinantes affectent profondément le marais abaissé par l’exploitation de la tourbe, parfois abusivement qualifié de “bas-marais”.
En ce qui concerne le haut-marais, le centre humide subit un emboisement naturel de plus en plus perceptible, tandis que les sphaignes et les pins de montagne ont reconquis une partie des landes mises à nu par une exploitation qui a cessé vers la fin des années 50. La recolonisation de ces landes par la végétation du haut-marais a été accélérée par la fermeture, en 1978, des rigoles de drainage.
3. observation Des Gouilles
Peut-être est-il utile de rappeler d’abord le sens du mot. Selon le Petit Larousse, ce terme, propre à la Suisse romande,
w. MattHey
Tableau 1: Caractéristiques de la tourbière du Cachot
• Coordonnées (au centre): 541200 206350• Localisation géographique: vallée de la Brévine, Jura neuchâtelois, Suisse.• Altitude 1050 m, moyenne des précipitations atmosphériques proche de 1,50 m/an.• Surface: 7 ha, épaisseur maximale de la tourbe: 6,3 m dans les régions 5, 9 et 10 (fig. 2)• Principales formations végétales du haut-marais (fig. 1): Caricetum limosae = Scheuchzerietum, Sphagnetum magellanici et ses variantes, Pino mugo Sphagnetum, Sphagno-Piceetum betuletosum, landes d’exploitation à divers stades de recolonisation, associations du marais abaissé par l’exploitation de la tourbe.• Partie centrale (2,1 ha) sous protection depuis 1945. Extension de la réserve en 1961 (7 ha). Dernières modifications, concernant les zones-tampon, en cours de discussion suite à l’arrêt du Tribunal fédéral du 20 octobre 1997 (Jacot-DescoMbes et al., 1998).
_____________________________

113
signifie flaque ou mare. mais on peut le définir de manière moins succinte et mieux adaptée aux tourbières: les gouilles (Schlenken, hollows) sont des points d’eau en principe d’origine naturelle. Ce sont des microstructures topographiques qui se forment dans les tourbières ombrogènes, en milieu ouvert. Alimentées par les eaux qui s’écoulent dans les couches superfi-cielles, soumises aux fluctuations de la nappe phréatique, elles sont inondées une bonne partie de l’année, sauf en période de sec prolongé (profondeur maximale 20 à 25 cm) (voir aussi Gobat et al., 1998). Le fond est parcouru par les stolons de laiches et de scheuchzeries ou colonisé par des touradons de trichophore gazonnant. Il est partiellement recouvert par les feuilles mortes de ces espèces. Par temps sec, cette nécromasse est engluée dans une boue tourbeuse fluide qui peut se dessécher en une croûte résistante et protectrice. même en période de sec prolongé, le niveau de la nappe ne s’abaisse que de quelques centi-mètres au-dessous du fond, et l’empreinte du pied se remplit d’eau. Les gouilles sont des milieux aquatiques biologiquement très intéressants tant du point de vue bota-nique que zoologique. Leur écologie a été étudiée dans le Haut-Jura par MattHey (1971).
Les gouilles de la tourbière du Cachot sont occupées par le Caricetum limosae Br.-Bl.21 = Scheuchzerietum (Tx 37). Cette association végétale est carac-térisée par un petit nombre d’espèces: Carex limosa, Scheuchzeria palustris et Sphagnun cuspidatum flottant ou en tapis denses saturés d’eau qui pourrissent en période de sec pour se reconstituer ensuite. Cette composition de base est enrichie par un cortège d’espèces qui comprend en particulier Carex rostrata, C. lasiocarpa, C. chordorrhiza et Sphagnum subsecundum. oberDorfer (1977), royer et al. (1978) et Grosvernier et al. (1992) ont fait l’in-ventaire et l’analyse des groupements végétaux des gouilles de hauts-marais.
Ajoutons que les fossés d’exploitation ou canaux, également fréquents dans les tourbières jurassiennes, n’entrent pas dans la même catégorie de points d’eau que les gouilles, et ne sont pas traités ici.
3.1 coMparaison De l’état Des Gouilles en 1969 et en 1998
En 1969, la tourbière du Cachot était, dans le Jura neuchâtelois, celle qui possé-dait le plus beau Sphagnetum magellanici, bien ouvert et troué par une cinquantaine de gouilles regroupées en une dizaine de familles selon leur situation dans le terrain et leur composition végétale (fig. 1 et 2). Les 43 plus grandes représentaient glo-balement une étendue d’eau libre de 309 m2. Jusqu’en 1998, 19 d’entre elles ont disparu, diminuant cette surface de 69 m2. De plus, les 24 gouilles survivantes ont subi un rétrécissement de 107 m2 environ, si bien que la réduction totale atteint 176 m2, soit environ 57% de la surface mesurée en 1969.
Les gouilles sont réparties en deux ensembles séparés par une diagonale d’îlots de pins et de buttes:a) La partie NO comprend les groupes 11 à 15. C’est la zone où le comblement des gouilles est le plus avancé, puisque 15 d’entre elles, sur 26, ont disparu. La plupart des survivantes ne dépassent plus 1 m2, si bien que la surface d’eau ouverte a passé de quelque 110 m2 à 6 m2, ce qui signifie une diminution de 94,5%.b) La situation est un peu différente dans la partie SE du Sphagnetum magellanici, située dans la partie centrale de la tour-bière, où la tourbe est la plus épaisse (> 6 m). Les gouilles (groupes 5 à 10) s’y sont mieux maintenues, 3 d’entre elles seule-ment sur 16 ayant disparu. La diminution de surface de l’eau libre atteint néanmoins 66 m2 ou 33,5%, même si la surface et le périmètre de trois grandes stations (5, 9 et 10) n’ont guère changé en une trentaine d’années.
évolution Des Gouilles Dans la tourbière Du cacHot: MorpHoloGie et véGétation

114
La figure 2 met en évidence ces consta-tations.
3.2 quelques exeMples
Groupe 14.Surface en 1969: environ 37,5 m2; en 1998: environ 2, 5 m2. Profondeur de l’eau dans la station 14: de 0 à 15 cm en 1969, de 0 à 12 cm en 1998.
En 1969 déjà, ces gouilles étaient rela-tivement peu profondes. von ballMoos (1989) a considéré qu’elles pouvaient appartenir au Caricetum chordorrhizae. Leur végétation était caractérisée par Carex chordorrhiza, C. limosa, menyanthes trifoliata, de grosses touffes de Tricho-phorum caespitosum étayant leurs bords. Sphagnum magellanicum et surtout S. sub-
secundum en garnissaient le fond, indices d’un comblement en cours. En 30 ans, le développement centrifuge des buttes du Sphagnetum magellanici a fortement empiété sur les bassins. Dans la station 14, par exemple, la surface d’eau libre a passé de 17 m2 à 2,30 m2 (fig. 3).
En 1998, les emplacements des gouilles entièrement (14a, b, d) ou partiellement (14 et 14c) comblées sont marqués par de nombreuses feuilles de menyanthes trifo-liata noyées dans les sphaignes (Sphagnum rubellum, S. magellanicum et S. capilli-folium principalement). La diminution du volume des stations s’est accompagnée d’une forte régression de Carex chor-dorrhiza et Sphagnum subsecundum au profit de Carex limosa, C. rostrata et de menyanthes trifoliata. Les touradons de Trichophorum caespitosum ont été partiel-lement submergés par les sphaignes.
w. MattHey
Figure 1: Zones physionomiques dans la tour-bière du Cachot (D’après vernier, 1992). Les gouilles sont situés dans la zone centrale, en blanc sur le plan.
Forêt de pins de montagne (Pino mugo Sphagnetum)
Boisement mixte de pins et de bouleaux
Landes de dégradation à vacciniées et callune
Forêt de lisière (Sphagno-Piceetum betuletosum)
Zone centrale à tapis continu de sphaignes (Sphagnetum magellanici) contenant les gouilles (Caricetum limosae ou Scheuchzerietum)marais abaissé par l’exploitation de la tourbe au niveau des zones cultivées environnantes
Zones de transition tourbière - milieux cultivés
mur de tourbe effondré
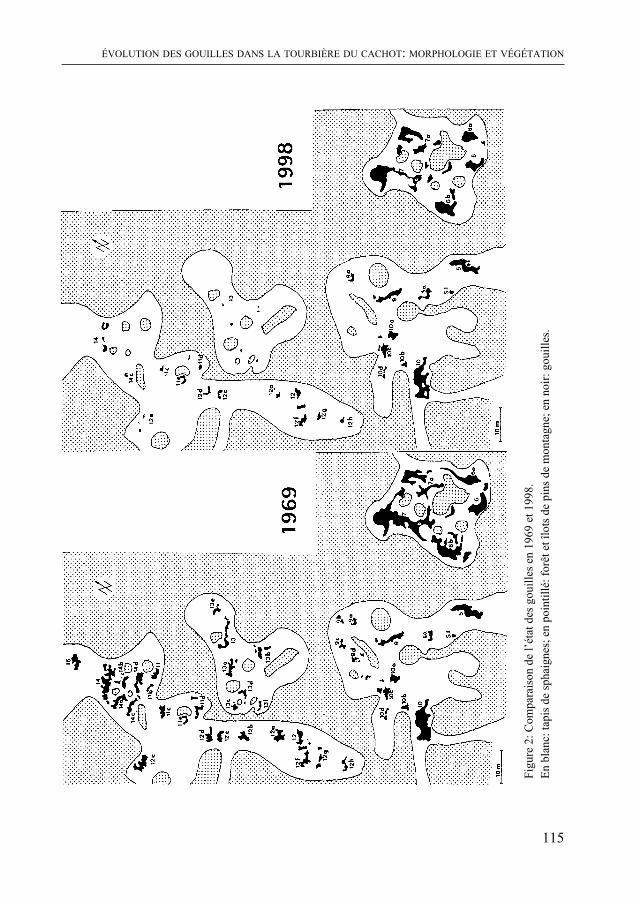
115
évolution Des Gouilles Dans la tourbière Du cacHot: MorpHoloGie et véGétation
Figu
re 2
: Com
para
ison
de
l’éta
t des
gou
illes
en
1969
et 1
998.
En b
lanc
: tap
is d
e sp
haig
nes;
en
poin
tillé
: for
êt e
t îlo
ts d
e pi
ns d
e m
onta
gne;
en
noir:
gou
illes
.

116
Groupe 13.Surface en 1969: environ 21 m2, en 1998 < 1m2.Profondeur de l’eau en 1969: de 0 à 17 cm.
Aucune des stations cartographiées en 1969, puis mentionnées encore en 1989 par von ballMoos ne mérite actuelle-ment le nom de gouilles, mais tout au plus celui de trou ou de fente. La croissance du Sphagnetum magellanici a été rapide dans cette zone colonisée par de nombreux îlots de pins autour desquels on observe un fort développement des buttes.
Groupe 10. Surface en 1969: 46 m2; en 1998: 41 m2 (diminution d’environ 10%).Profondeur de l’eau en 1969 et en 1998: de 0 à 10 cm.
La forme, la surface et la végétation de la station 10 (34 m2) sont restées iden-tiques durant trois décennies, à quelques détails près. Par ordre d’abondance, on
notait en 1969 Carex lasiocarpa, C. limosa, C. rostrata et quelques pieds de Scheuch-zeria palustris. Sur le fond de tourbe nue, à sec durant plus d’un mois par année, Oxycoccus quadripetalus était abon-dant. Sphagnum cuspidatum n’a pas été observé dans cette station trop sèche. mais depuis quelques années, un changement se dessine. Si la strate herbacée reste la même, trois massifs de sphaignes se déve-loppent sur le fond: un îlot de Sphagnum subsecundum (0,30 x 0,35 m), un autre de Sphagnum magellanicum + S. subse-cundum (0,80 x 0,40 m) et une “coulée” de Sphagnum magellanicum (0,80 x 0,70 m) issue d’une butte bordière haute de 0,50 cm. L’installation de ces sphaignes indique une augmentation d’humidité sur le fond de la station, elle représente aussi le pre-mier signe de comblement.
Station 7.Surface en 1969 (sans les îlots de sphaignes): 48 m2 , en 1998: 30 m2.
w. MattHey
Figure 3: Plans de la station 14.En haut: 1969, en bas: 1998.
Bord des buttes de sphaignes
Pins de montagne

117
Profondeur de l’eau en 1969, partie NO: 0 à 9 cm, partie SE: 0 à 10 cm.
En trente ans, la physionomie de cette grande gouille a passablement changé: les bras de ses diverticules ont été comblés par le Sphagnetum magellanici, ce qui a individualisé quatre petites gouilles secon-daires (fig. 4). Sur le fond, deux faciès végétaux, correspondant à la partie la plus sèche et la plus humide de la station, étaient bien marqués en 1969. Ils le sont encore plus nettement aujours’hui: a) Dans la moitié NO, le fond nu est occupé, comme il l’était déjà en 1969, par Carex rostrata (80 à 120 tiges/m2 ). mais d’épais touradons de Trichophorum caes-pitosum y sont maintenant installés. Leurs feuilles mortes abritent des sphaignes qui marquent le début du processus de coloni-sation décrit par y. MattHey (1993) (voir aussi MattHey y. & Jacot-DescoMbes,
1996). On peut remarquer que la moitié NO présente aujourd’hui une physionomie qui était celle de la moitié SE il y a trente ans.b) La partie SE a changé davantage. Les trois îlots centraux ont fusionné en une seule grande butte qui entre en contact avec le bord. Le fond humide, dégagé en 1969, est occupé maintenant par une dense couche de Sphagnum cuspidatum, épaisse d’une dizaine de centimètres.
La station 6b, dans le prolongement de 7, a subi la même évolution, marquée par une forte diminution de la surface d’eau libre (1969: 28 m2 ; 1998: 8 m2)
Station 5. Surface d’eau ouverte:- en 1962: 17 m2, profondeur de 0 à 25 cm. Cette station était la dernière gouille à s’assécher dans la tourbière.
évolution Des Gouilles Dans la tourbière Du cacHot: MorpHoloGie et véGétation
Figure 4: En 1969, les touradons de Trichophorum caespitosum (o) ont une disposition qui préfigure les étapes du comblement. En 1998, T. caespitosum colonise la partie NO, où il était rare en 1969.

118
- en 1998: 13 m2 (fig. 5 1998, niveau a), 21 m2 avec > 25 cm d’eau (fig. 5 1998, niveau b).
Dès 1962, cette gouille typique a été l’objet d’observations suivies sur les plans botanique d’abord, puis zoologique (Mat-tHey, 1964 et 1971, ). Il s’agissait d’un Caricetum limosae caractéristique, com-posé uniquement de Scheuchzeria palus-tris, Carex limosa et de Sphagnum cuspi-datum, forme flottante (fig.6a).
En 1988, Carex lasiocarpa a pris pied pour la première fois dans la station (von ballMoos, 1989), mais il ne s’est guère multiplié jusqu’à aujourd’hui (< 5 indi-vidus).
Les bords et arrière-bords de la station ont subi un piétinement plus intense que ceux des autres gouilles, le fond étant épargné. A la cadence de deux passages par semaine, de mai à octobre, cette pression anthropique s’est prolongée de 1964 à 1969. Après une année environ, les sphaignes sont mortes. Elles se sont décomposées après deux ans, transfor-mant les cheminements en surfaces de tourbe fluide profondément remuée (fig. 6b). Conséquence inattendue, des espèces rares, telles que Scheuchzeria palustris et Drosera rotundifolia ont proliféré sur ces surfaces bouleversées, étant sans doute favorisées par l’absence d’ espèces concur-rentes.
Actuellement, la station 5 a trouvé un nouvel équilibre (pl. 1, fig. 1). En 1998, sa forme est peu différente de celle de 1962 (fig. 5 et 6c). La partie NO est un bassin de 1,8 m x 0,70 cm, où la profondeur de l’eau atteint encore 7 cm quand le reste de la gouille est à sec; c’est là seulement qu’on peut encore reconnaître la physionomie originelle de la station (pl. 1, fig. 2). Les bords et le tapis de sphaignes des arrière-bords se sont reconstitués avec les mêmes espèces que précédemment (Sphagnum magellanicum et S. rubellum principa-lement). Le Sphagnetum magellanici a comblé la pointe ouest de la station sur 40 cm, et la progression des buttes latérales a modifié la forme de la rive ouest. Les pins situés en bordure de gouille ont une crois-sance très lente (tableau 2) et influencent vraisemblablement peu l’écologie de la station.
En 1998, la strate herbacée ancrée sur le fond comporte les mêmes espèces qu’en 1962, mais, dans les parties centrale et sud de la gouille, les proportions de Scheu-chzeria palustris et de Carex limosa ne sont plus identiques (77,4% / 22,6% en 1969 contre 55,6% / 44,4% en 1998). Sphagnum cuspidatum forme maintenant un tapis continu, épais d’une quinzaine de cm (>1000 capitulums /m2 en 1998 contre 400 en 1969) (pl. 2, fig. 3 et 4). Ce tapis reste encore très fragile, il se dissout et s’incorpore à la tourbe du fond en cas de sécheresse prolongée pour se reconstituer ensuite. Enfin, à l’extrémité SE, Drepano-
w. MattHey
Figure 5: Coupes transversales de la station 5.
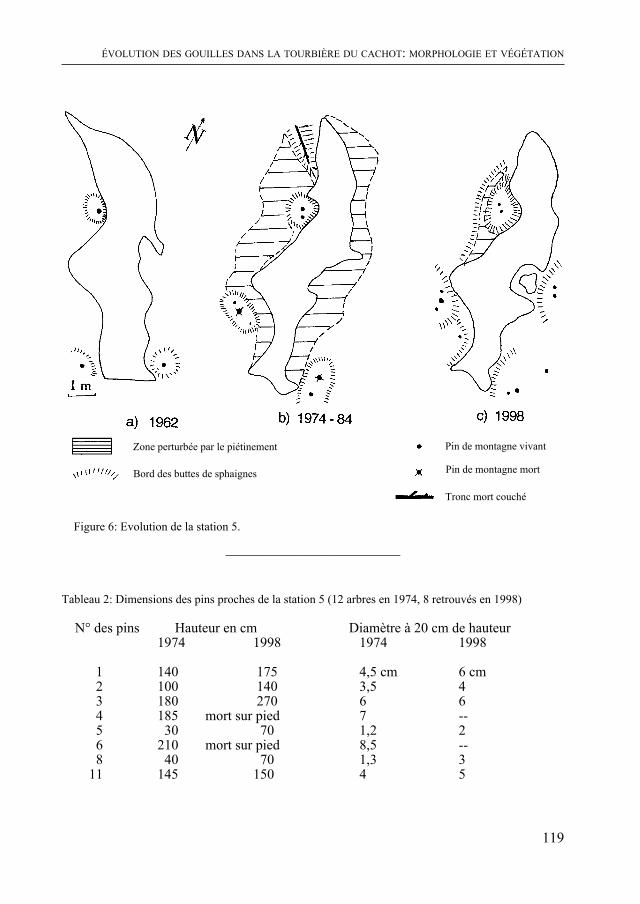
119
évolution Des Gouilles Dans la tourbière Du cacHot: MorpHoloGie et véGétation
Figure 6: Evolution de la station 5.
Tableau 2: Dimensions des pins proches de la station 5 (12 arbres en 1974, 8 retrouvés en 1998)
N° des pins Hauteur en cm Diamètre à 20 cm de hauteur 1974 1998 1974 1998
1 140 175 4,5 cm 6 cm 2 100 140 3,5 4 3 180 270 6 6 4 185 mort sur pied 7 -- 5 30 70 1,2 2 6 210 mort sur pied 8,5 -- 8 40 70 1,3 3 11 145 150 4 5
_________________________
Pin de montagne vivant
Pin de montagne mort
Tronc mort couché
Zone perturbée par le piétinement
Bord des buttes de sphaignes

120
cladus fluitans, d’abord confiné sous les bords, déborde actuellement sur le fond de la gouille. Dans l’ensemble, la station 5 évolue vers son comblement.
On peut aussi constater qu’en définitive, le piétinement n’a pas nui à la station. Sa forme et son profil en ont été modi-fiés, mais l’impact a sans aucun doute freiné l’avancée des buttes du Sphagnetum magellanici, contribuant ainsi à prolonger la survivance d’une association végétale rare et des espèces animales tyrphophiles et tyrphobiontes qui lui sont associées.
Le service d’une station météorologique, en fonction de 1969 à 1980, a entraîné un fort piétinement aux abords de la station 5a. Profondément destructuré, le tapis de Sphagnum magellanicum et de S. recurvum s’est effondré, provoquant un agrandissement de 2 à 3 m2 de cette sta-tion. Aujourd’hui, la zone perturbée s’est reconstituée, et seul un diverticule de 1,5 m2, visible sur la fig. 7, rappelle cet épi-sode. L’ensemble de la gouille abrite un dense peuplement de Scheuchzeria palus-tris et de Carex limosa.
4. Discussion
Le piétinement des sols fragiles de tour-bières a des conséquences très visibles dans certaines réserves naturelles (Mul-Hauser, 1996), ce qui a entraîné la mise en place de mesures de protection (sentiers balisés, mise à ban de zones sensibles, informations au public). Cependant, cet impact n’a que rarement fait l’objet de mesures précises (slater & aGnew, 1977). Parmi les observations et expé-riences effectuées au Cachot sur ce pro-blème, une seule, en rapport avec l’éco-logie des Oribates, a été publiée jusqu’ici (borcarD & MattHey, 1996).
Dans les conditions des tourbières juras-siennes, un piétinement répété déclenche
un processus de destruction rapide des tapis de sphaignes suivi, si le substrat le permet (Gobat et al., 1986), par une lente régénération qui ramène souvent, mais pas toujours, aux conditions initiales. Dans le cas du Sphagnetum magellanici de la tourbière du Cachot, on peut distinguer les phases suivantes 1):1. Piétinements répétés.2. mort des sphaignes.3. Désagrégation de la couche de sphaignes, qui se transforme en une tourbe fluide après deux ans environ.4. Formation d’une dépression de 5 à 20 cm de profondeur.5. Lente recolonisation du fond par, dans l’ordre, Oxycoccus quadripetalus, Andro-meda polifolia, Trichophorum caespi-tosum, Eriophorum vaginatum, E. angus-tifolium, Carex rostrata et C. lasiocarpa. Souvent, les premières mousses à réap-paraître dans la zone perturbée ne sont pas des sphaignes, mais appartiennent aux genres Aulacomnium et Dicranum. 6. La lente avancée des bords de sphaignes rétrécit la dépression tandis que Sphagnum recurvum réapparaît sur le fond sous les touffes de Trichophorum caespitosum ou d’Eriophorum vaginatum, selon le pro-cessus décrit par y. MattHey (1993).7. La durée de la cicatrisation est variable et dépend de la grandeur de la dépression et de la nature du tapis de sphaignes envi-ronnant. menyanthes trifoliata peut encore marquer l’emplacement de la gouille alors que le tapis de sphaignes l’a déjà effacé. 8. Plus rarement, ces dépressions se muent en agrandissements de gouilles existantes, ou vont jusqu’à en former de nouvelles, qui sont colonisées par le Caricetum limosae, à l’exemple du diverticule de la station 5a (fig. 7).
w. MattHey
1) Nous avons suivi la chronologie d’une telle régénération dans une petite zone circulaire piétinée par des chevreuils (diamètre 1,50 m). Le Sphagnetum magellanici (faciès à Sphagnum recurvum) s’est reconstitué en une vingtaine d’années, (observations non publiées).

121
5. conclusions
Les gouilles sont des milieux précieux par l’originalité de leur flore et de leur faune. Elles représentent une séquence dans l’évolution des haut-marais, étant destinées à être peu à peu nivelées par les replats et les buttes de sphaignes selon dif-férentes modalités.
En 1969, le centre intact de la tour-bière du Cachot était riche d’une cinquan-taine de gouilles de tailles diverses (de < 1m2 à 50 m2) réparties sur 1,5 ha environ. Elles ont évolué de manières différentes au cours des trois dernières décennies. a) Dans la moitié NO (figures 2 et 3), les gouilles caractérisées par Carex chordor-rhiza et Sphagnum subsecundum ont été presque toutes oblitérées ou réduites à des trous ou des fentes de petites dimensions par la coalescence des buttes du Spha-gnetum magellanici. Les surface d’eau
libre restantes sont insuffisantes pour per-mettre le maintien d’une flore originale et abriter des peuplements diversifiés d’in-vertébrés aquatiques. b) Dans la moitié SE, les gouilles sont perchées sur une plus grande épaisseur de tourbe, en gros dans le centre géogra-phique de la tourbière originelle selon l’axe transversal de la vallée. Plusieurs d’entre elles ont peu changé en trente ans, restant les dernières à abriter le grou-pement à Scheuchzeria palustris, Carex limosa et Sphagnum cuspidatum.
Cependant, on y observe aussi une ten-dance au comblement, particulièrement net dans les stations 6b et 7 (fig. 2 et 4).
Le processus de comblement est marqué par l’installation d’îlots ou de tapis perma-nents de sphaignes sur le fond des gouilles: Sphagnum cuspidatum (St. 5, 7), S. sub-secundum et S. magellanicum (St. 10), S. recurvum (St. 10a, b, c, c’, d). Seule
évolution Des Gouilles Dans la tourbière Du cacHot: MorpHoloGie et véGétation
Figure 7: Station 5a.En 1980, année de l’enlèvement de la cabane météo.En 1998, un diverticule s’est créé dans la zone piétinée et la butte centrale s’est étendue.
Emplacement de la cabane météo
Bord de la butte et pin de montagne
Partie en voie de comblement
Zone perturbée par le piétinement

122
la station 9 reste indemne. Le processus de comblement semble très lent... mais il faut faire montre de prudence dans ce genre d’affirmation. En effet, ni MattHey, W. (in Gobat et al. 1986), ni von ball-Moos (1989) n’avaient été particulièrement frappés par des changements de grande amplitude dans la partie NO du Spha-gnetum magellanici. Il semble donc que l’évolution se soit accélérée au cours des dix dernières années, quelles qu’en soient les causes (emboisement, modification de la circulation de l’eau dans l’acrotelm, fer-meture des drains superficiels sur la lande Pochon, etc).
Le piétinement a aidé la station 5 à maintenir ses limites, et a entraîné de grandes modifications à l’échelle de la stations 5a. mais l’homme n’est pas tou-jours à l’origine de ce genre d’impact. Nous avons observé à plusieurs reprises au Cachot que le tapis de sphaignes était gratté et bouleversé par des chevreuils
(reposées) ou des blaireaux, sur des sur-faces excédant rarement 2 m2. De même, l’emplacement d’un pin renversé peut évo-luer en une gouille permanente (S. 1, non décrite ici).
Naturelles ou anthropiques, ces atteintes aux tapis de sphaignes contribuent à entretenir le dynamisme du haut-marais humide. Elles freinent le comblement de gouilles, en créent parfois de nouvelles et maintiennent du même coup l’habitat de plantes rares et d’invertébrés tyrphophiles ou tyrphobiontes. Ainsi, un piétinement soigneusement programmé, pratiqué aux bons endroits et de durée limitée, pourrait entrer dans la panoplie des méthodes dont l’écologie appliquée fait déjà usage pour la conservation des tourbières (Grosvernier, 1997; MattHey, y., 1993; MattHey, y. & Jacot-DescoMbes, 1996).
reMercieMents
w. MattHey
A mon épouse, pour son assistance sur le terrain.
biblioGrapHie
auroi, C. 1979. Recherche sur l’écologie des Tabanidae, et de Hybomitra bimaculata (macquart) en particulier, dans une tourbière du Haut Jura neuchâtelois. Thèse de doctorat. Institut de Zoologie. Université de Neuchâtel. Suisse.
ballMoos, C., von. 1989. Contribution à l’étude des Odonates des tourbières ombrogènes. Peuplement d’un haut-marais du Jura neuchâtelois (vallée de la Brévine). mémoire de licence. Institut de Zoo-logie. Université de Neuchâtel. Suisse.
borcarD, D. 1988. Les Acariens Oribates des sphaignes de quelques tourbières du Haut-Jura suisse. Thèse de doctorat. Institut de Zoologie. Université de Neuchâtel. Suisse.
borcarD , D. & MattHey, W. 1995. Effect of a controlled trampling of Sphagnum mosses on their Oribatid mites assemblages (Acari, Oribatei). Pedobiologia 39: 219-230.
GeiGer, W. 1980. Phytosociologie des landes de dégradation dans la tourbière du Cachot (Jura neuhâ-telois, Suisse). Documents phytosociologiques N.S. 5: 5-10.
Gobat, J. m., Grosvernier, P. & MattHey, y (avec coll. de MattHey, W). 1986. Les tourbières du Jura suisse. Actes Soc. juras. d’émul. 213-315.

123
Gobat, J. m., araGno, m. & MattHey, W. 1998. Le sol vivant. Presses polytechniques et universi-taires romandes. Lausanne.
Grosvernier, P. R. 1996. Stratégies et génie écologique des sphaignes (Sphagnum sp) dans la restaura-tion spontanée des marais jurassiens suisses. Thèse de doctorat. Institut de Botanique. Université de Neuchâtel. Suisse.
Grosvernier, P., MattHey, y & MulHauser, G. 1992. Typologie des milieux tourbeux. Actes Soc. juras. d’émulation: 145-186.
Grosvernier, P., buttler, A., Gobat, J.-m. & MattHey, y. 1997. Régénération ou réimplantation des sphaignes dans les hauts-marais jurassiens suisses. Actes Soc. juras. d’émulation: 155-179.
Jacot-DescoMbes, p., fiecHter, a. & farron, L. 1998. Nature neuchâteloise 1997. Bull. Soc. neu-châtel Sci. nat. 121: 175-184.
MattHey, W. 1964. Observations écologiques dans la tourbière du Cachot. Bull. Soc. neuchâtel. Sci. nat. 87: 103-135.
MattHey, W. 1971. Ecologie des insectes aquatiques d’une tourbière du Haut-Jura. Rev. suisse Zool. 78 (367-536).
MattHey, W. & borcarD, D. 1996. La vie animale dans les tourbières jurassiennes. Bull. Soc. neu-châtel. Sci. nat. 119: 3-18.
MattHey, y. 1993. Typologie de la régénération spontanée des hauts-marais jurassiens non boisés et approche écologique de trois séries végétales caractéristiques de la dynamique secondaire. Thèse de doctorat. Institut de Botanique. Université de Neuchâtel. Suisse.
MattHey, y. & Jacot-DescoMbes, P. 1996. Les mesures de régénération et de cicatrisation des hauts-marais neuchâtelois. Premiers résultats et perspectives. Bull. Soc. neuchâtel. Sci. nat. 119: 154-162
MulHauser, B. 1996. La Gruère. Fondation Les Cerlatez. Saignelégier.
neet, C. 1984. Etude de l’écologie et de la biologie de Tetragnatha extensa (L.) (Araneae, Tetragna-thidae) dans une tourbière du Haut-Jura. mémoire de licence. Institut de Zoologie. Université de Neuchâtel. Suisse.
oberDorfer, E. Süddeutsche Pflanzengesellschaften I. Gustav Fischer verlag. Stuttgart.
royer, J.M., vaDaM, J. c., Gillet, J. p. & auMonier, M. f. 1978. Etude phytosociologique des tour-bières acides et alcalines du Haut-Doubs. Réflexions sur leur régénération et leur genèse. Publica-tions du Centre universitaire d’études régionales de l’Université de Franche-Comté 2: 109-185
slater, f. M. & aGnew, a. D. q. 1977. Observations on a peat bog’s ability to withstand increasing public pressure. Biol. Conserv. 11: 21-27
vaucHer-von ballMoos, c. 1997. Etude de six zones de transition entre tourbières acides et zones agricoles dans le Haut-Jura suisse. Thèse de doctorat. Institut de Zoologie. Université de Neuchâtel. Suisse.
vernier, R. 1992. Recherche écofaunistique sur les fourmis du genre Formica de la tourbière du Cachot (Jura neuchâtelois) et hauts-marais voisins (Hymenoptera, Formicidae) 1. Liste des espèces et leurs biotopes préférentiels. Bull. Soc. neuchâtel. Sci. nat.115: 61-82.
évolution Des Gouilles Dans la tourbière Du cacHot: MorpHoloGie et véGétation

124
w. MattHey

125
évolution Des Gouilles Dans la tourbière Du cacHot: MorpHoloGie et véGétation
bull. soc. neucHâtel. sci. nat. 121 : 111-125. 1998
Planche 1 (page 124):En haut: vue générale de la station 5. Sa forme est marquée par les feuilles jaune-brun des laiches des bourbiers (Carex limosa) et des scheuchzéries (Scheuchzeria palustris). (Photo F. matthey, le 30 septembre 1996)En bas: Extrémité nord-ouest de la station 5. Elle conserve, sur une surface réduite, la physionomie offerte par l’ensemble de la gouille en 1962 (voir texte). Sur le bord, au second plan, feuilles sèches d’une touffe de linaigrettes. (Photo F. matthey, le 30 mai 1998)
Planche 2 (page 125):
En haut: Centre de la station 5. Une couche dense de Sphagnum cuspidatum recouvre le fond, percée par les laiches des bourbiers et les scheuchzeries. (Photo F. matthey, le 30 mai 1998)En bas: Centre de la station 5. Détail de la couche de Sphagnum cuspidatum. (Photo F. matthey, le 30 mai 1998)


bulletin De la société neucHâteloise Des sciences naturelles 121 : 127-135. 1998
127
FAUNISTIqUE DE 9 SCARABAEIDAE PHyTOPHAGES ET DE 3 LUCANIDAE (COL., SCARABAEIDAE PLEUROS-
TICTI, LUCANIDAE) CAPTURÉS DANS LES GORGES DE L’AREUSE (NEUCHATEL, SUISSE)1
SyLvIE BARBALAT
Laboratoire d’écologie animale et d’entomologie, Université de Neuchâtel, Rue Emile-Argand 11, 2000 Neuchâtel, Suisse.
mots-clés: Scarabaeidae Pleurosticti, Lucanidae, faunistique, écologie, Jura suisse.
Key-words: Scarabaeidae Pleurosticti, Lucanidae, faunistics, ecology, Swiss Jura.
Résumé
Au moyen de pièges-fenêtres et d’assiettes colorées, nous avons recensé 9 espèces de Scarabaeidae phytophages et 3 de Lucanidae (Col., Scarabaeidae Pleurosticti, Lucanidae) dans les Gorges de l’Areuse (NE). Nous présentons le résultat de nos captures, ainsi qu’un aperçu de l’écologie et de la distribution générale, suisse et neuchâteloise de ces espèces.
Summary
By mean of window and water traps, we captured 9 phytophagous Scarabaeidae and 3 Lucanidae species (Col., Scarabaeidae Pleurosticti, Lucanidae) in the Areuse Gorges (NE). The results of our captures are presented, as well as an overview of the species’ ecology and distribution in the world, in Switzerland and in the canton of Neuchâtel.
Zusammenfassung
Durch Fensterfallen und Gelbschalen haben wir 9 phytophage Scarabaeidae- und 3 Lucanidaearten (Col., Scarabaeidae Pleurosticti, Lucanidae) in der Areuseschlucht (NE) gefangen. Die Ergebnisse der Fänge werden vorgestellt sowie einen Überblick der Ökologie und verbreitung der Arten in der Welt, in der Schweiz und im Kanton Neuenburg.
introDuction
Réunies dans la super-famille des Scarabaeoidea, les Scarabaeidae et les Lucanidae sont deux familles voisines, dont les représentants se caractérisent par des antennes ter-minées en lamelles, ce qui leur a jadis valu le nom de Lamellicornia. Les Scarabaeoidea comptent plus de 20’000 espèces dans le monde et près de 200 en Suisse. Les Sca-
1 Cet article fait partie de la thèse de l’auteur.

rabaeidae sont divisés en deux groupes dont l’écologie est très différente. Le pre-mier, celui des Laparosticti ou bousiers, au régime alimentaire essentiellement copro-phage, ne sera pas traité ici. Le second, celui des Pleurosticti, auquel cet article est consacré, comprend plutôt des espèces phytophages. Ces insectes ont une éco-logie assez différente selon qu’il s’agisse des larves ou des adultes. Les larves se nourrissent de racines vivantes, de bois ou de végétaux en décomposition. Les adultes peuvent se nourrir de feuilles, de bour-geons, de graines, de fleurs ou ne pas se nourrir du tout. La durée du développe-ment larvaire dépend fortement du climat. balacHowsky (1962) cite l’exemple du hanneton commun (melolontha melo-lontha L.) dont le développement dure 15 mois en laboratoire, 3 ans dans les régions méridionales et 4 ans dans les zones plus septentrionales. En Suisse, le cycle du han-neton commun est de 3 ans sur le Plateau et de 4 ans dans certaines vallées des Gri-sons et du valais (allenspacH, 1970).
La famille des Lucanidae est beau-coup moins nombreuse. En effet, elle n’est représentée en Suisse que par 7 espèces, dont la plus connue, le cerf-volant (Lucanus cervus L.), est l’un de nos plus spectaculaires Coléoptères. Les larves de Lucanidae se nourrissent de bois pour-rissant et les adultes de fruits ou de sève fermentés.
De nombreux représentants de ces deux familles se sont fortement raréfiés ces dernières décennies. Les espèces dont les larves se nourrissent de racines de plantes herbacées, dont les plantes cultivées, ont souffert de l’intensification de l’agriculture et de l’utilisation généralisée de pesticides. D’autres espèces vivant dans le bois pour-rissant ont décliné avec les enrésinements et la régression des vieux arbres. D’autres enfin, comme la Cétoine dorée (Cetonia aurata F.) ont su s’adapter aux activités humaines et ont trouvé un habitat de subs-titution dans les composts de jardin.
Description Des Milieux
Nous avons étudié la faune de trois groupements forestiers: la chênaie, la forêt mixte (feuillus et conifères) et la hêtraie pure. Nous avons choisi douze stations, entre Corcelles et Brot-Dessous, dont l’al-titude varie de 550 à 890 m (fig. 1). Les stations, exposées au sud ou au sud-est, se trouvaient en milieu semi-ouvert (tab. 1): cinq en lisière et sept dans des coupes d’abri. Ces dernières sont des trouées arti-ficielles ouvertes par les forestiers pour favoriser le recrû naturel d’essences hélio-philes comme le pin sylvestre (Pinus syl-vestris) ou les chênes (quercus sp.). Tous les arbres y sont abattus sauf quelques uns, conservés comme semanciers. De même, le sous-bois a été enlevé. Les souches sont laissées sur place, et les branches inutili-sables entassées aux abords de la coupe. Les coupes d’abri constituent des trouées aux limites franches, tandis que quatre des cinq lisières choisies sont étagées.
Matériel et MétHoDes
Les adultes des insectes étudiés ont été échantillonnés au moyen de pièges-fenêtres (2 par station) et d’assiettes colo-rées (une blanche et une jaune par station) (barbalat, 1995). Relevés tous les 10 jours, les pièges ont été fonctionnels de fin avril à début septembre en 1994 et 1995.
Les indications concernant l’écologie et la distribution générale des espèces pro-viennent des ouvrages de balacHowsky (1962), Horion (1958) et allenspacH (1970). Le régime alimentaire des larves, déterminant pour la présence ou l’absence d’une espèce dans un endroit donné, est également indiqué.
La distribution en Suisse et dans le canton de Neuchâtel a été établie à partir de cartes basées sur les données du Centre Suisse de Cartographie de la Faune (CSCF) à Neuchâtel, plus actuelles que le catalogue d’allenspacH (1970). La distribution neu-châteloise est présentée de façon générale et
s. barbalat
128
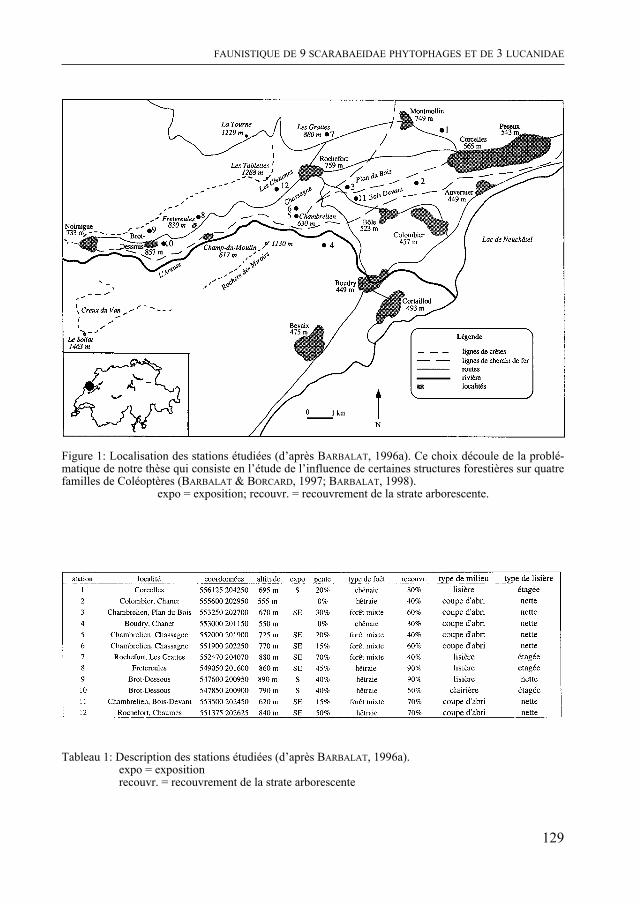
129
faunistique De 9 scarabaeiDae pHytopHaGes et De 3 lucaniDae
Figure 1: Localisation des stations étudiées (d’après barbalat, 1996a). Ce choix découle de la problé-matique de notre thèse qui consiste en l’étude de l’influence de certaines structures forestières sur quatre familles de Coléoptères (barbalat & borcarD, 1997; barbalat, 1998). expo = exposition; recouvr. = recouvrement de la strate arborescente.
Tableau 1: Description des stations étudiées (d’après barbalat, 1996a).expo = expositionrecouvr. = recouvrement de la strate arborescente

s. barbalat
130
la liste des localités citées n’est pas exhaus-tive. Le haut du canton comprend la zone, à environ 1000 m d’altitude (si l’on excepte les bords du Doubs à 650 m), au nord-ouest du val-de-Ruz et du val-de-Travers. Le fond de ces deux vallées est en moyenne à 750 m. Le bas se limite aux bords du Lac de Neuchâtel de 430 à 700 m et à la partie du canton située entre les lacs de Neuchâtel et de Bienne. La distribution dans le canton ne prend en compte que les données antérieures à notre recherche, qui a débuté en 1993 par une étude préliminaire, dont les résultats sont exposés dans deux publications (bar-balat, 1995; barbalat, 1996b). Le présent article inclut aussi une espèce capturée uni-quement en 1993, Protaetia cuprea.
présentation Des résultats
En 1994 et 1995, nous avons recensé 8 espèces de Scarabéides phytophages et 3 de Lucanides, représentant un total de 501 individus. La répartition de ces 2 familles dans les différentes stations est présentée dans le tableau 2. La nomenclature utilisée est celle de loHse & lucHt (1992). La systématique jusqu’à la sous-famille est celle de lawrence & newton (1995). A ces 11 espèces, il faut en ajouter une, Protaetia cuprea, recensée en 1993 lors de l’étude préliminaire et qui n’a pas été retrouvée en 1994 et 1995.
faunistique et écoloGie
lucaniDae
LucaninaePlatycerus caprea (Deg., 1774)Distribution générale: régions monta-gneuses d’Europe centrale.Distribution suisse: tout le pays, surtout en montagne.Distribution neuchâteloise: quelques don-nées éparses, plutôt à basse altitude (vau-seyon, Chambrelien, Colombier, bords du Doubs) mais également sur les reliefs (Les Bayards, Pouillerel).Régime alimentaire des larves: saproxylo-phage.Apparition des adultes: mai et juin.
Nous avons capturé cette espèce presque exclusivement dans les stations situées à plus de 750 m (stations 7, 8, 9, 10 et 12). Un seul individu sur 60 a été trouvé plus bas, à 550 m (station 4). Cela confirme les données de la littérature qui font état d’une vicariance altitudinale entre cette espèce et la suivante. Selon allenspacH (1970), les larves de Platycerus caprea se déve-loppent dans le bois pourrissant de divers arbres, aussi bien feuillus que résineux et les adultes volent volontiers dans les lisières en plein soleil.
Tableau 2: Liste des espèces capturées dans chaque station.

Platycerus caraboides (L., 1758)Distribution générale: Europe centrale et occidentale, Balkans (zaHraDnik & cHvala, 1990).Distribution suisse: Bassin lémanique, Jura, Plateau.Distribution neuchâteloise: une seule donnée au Landeron.Régime alimentaire des larves: saproxylo-phage.Apparition des adultes: avril et mai.
Contrairement au précédent, nous avons surtout trouvé ce Lucanide dans les sta-tions de basse altitude. La seule exception est l’individu capturé dans la station 12 à 840 m. allenspacH (1970) signale cette espèce sur le chêne, alors que zaHraDnik & cHvala (1990) la mentionnent sur le hêtre. Nos résultats nous feraient pencher pour la seconde information. Nous avons en effet trouvé 11 des 18 individus de Pla-tycerus caraboides à Colombier (station 2), une hêtraie située à basse altitude, contre 1 dans chacune des chênaies de Corcelles et Boudry (stations 1 et 4).
Syndesinae
Sinodendron cylindricum (L., 1758)Distribution générale: Europe et Sibérie occidentale (zaHraDnik & cHvala, 1990).Distribution suisse: surtout en valais. quelques données éparses dans le reste du pays.Distribution neuchâteloise: inconnue. Une seule donnée très ancienne et imprécise (Canton de Neuchâtel) atteste de la pré-sence de cette espèce dans le canton.Régime alimentaire des larves: saproxylo-phage.Apparition des adultes: de mai à août.Nous n’avons trouvé que deux individus de ce Lucanide, l’un en 1993 sur le versant nord de la montagne de Boudry et l’autre aux Grattes (station 7). Selon kocH (1992) et bily (1990), cette espèce est indicatrice
des vieilles forêts de hêtres. Nous l’avons effectivement trouvée dans une hêtraie et une forêt mixte, peu ou plus exploitées, présentant quelques vieux hêtres. Cepen-dant, zaHraDnik & cHvala (1990) la signalent sur d’autres essences comme le chêne, le tilleul (Tilia sp.), l’érable (Acer sp.), le frêne (Fraxinus sp.), le bouleau (Betula sp.) et les arbres fruitiers. allens-pacH (1970) indique également sa pré-sence sur de nombreuses essences. Nous avons observé Sinodendron cylindricum sous l’écorce d’un chêne dans l’est de la Pologne, région dont le hêtre est absent.
scarabaeiDae pleurosticti
melolonthinaeAmphimallon atrum (Hbst., 1790)Distribution générale: Europe occidentale.Distribution suisse: Jura et pied du Jura. quelques très rares anciennes données ailleurs.Distribution neuchâteloise: données éparses dans tout le canton (Couvet, Champ-du-moulin, Rochefort, Chaumont, St-Blaise, Neuchâtel-Chanet).Régime alimentaire des larves: probable-ment rhizophage.Apparition des adultes: juin et juillet.
Amphimallon atrum n’a été trouvé que dans les stations 7, 8, 9 et 10 et constitue dans le cas présent une espèce caractéris-tique des lisières d’altitude. En effet, il n’a été trouvé ni dans la station 12, une station d’altitude mais en coupe d’abri, ni dans la station 1, une lisière en situation thermophile. Nos constatations confirment celles de Horion (1958) et allenspacH (1970) qui signalent cette espèce surtout sur les pentes ensoleillées dans les régions de collines et de montagne, ainsi que celles de Horion (1958) et Janssens (1960) qui la mentionnent dans les lisières.
Hoplia argentea (Poda, 1761)
131
faunistique De 9 scarabaeiDae pHytopHaGes et De 3 lucaniDae

Distribution générale: Europe centrale et méridionale.Distribution suisse: Surtout en montagne. Population (ou prospection!) importante dans le nord du canton de Zurich.Distribution neuchâteloise: surtout sur les reliefs (Côte du Cerf, Ferme Robert, La Tourne, valanvron). Egalement à plus basse altitude (Chambrelien, Rochefort, Hauterive, bords du Doubs).Régime alimentaire des larves: probable-ment rhizophage.Apparition des adultes: de mai à juillet.
Cette espèce a été trouvée surtout dans la station 8, ce que nous expliquons par la présence conjointe d’une lisière étagée et d’une prairie maigre. En revanche, nos résultats ne nous permettent pas de considérer Hoplia argentea comme une espèce d’altitude, étant donné qu’elle n’est que très peu représentée dans les stations 7, 9 et 10, situées au dessus de 800 m. Cependant, sa distribution en Suisse, ainsi que nos observations personnelles, nous conduisent à penser que sa plus grande abondance en altitude est due à une agriculture plus extensive. Nous avons constaté une grande variation d’abon-dance entre 1994 et 1995, mais n’avons pas trouvé d’informations concernant le rythme de développement de cette espèce.
Omaloplia ruricola (F., 1775)Distribution générale: Europe centrale et méridionale.Distribution suisse: Très rare en dehors du valais central.Distribution neuchâteloise: une seule donnée très ancienne aux Bayards.Régime alimentaire des larves: peut-être myr-mécophile (kaufMann in paulian, 1959).Apparition des adultes: juin et juillet.
Selon allenspacH (1970), les adultes d’Omaloplia ruricola se tiennent volontiers sur les tiges sèches des graminées, ainsi que sur les fleurs et les buissons. Le seul individu de cette espèce a été capturé dans
la station 10, qui réunit prairie fleurie et buissons.
Rhizotrogus aestivus (Ol., 1789)Distribution générale: Europe centrale et méridionale. Asie mineure.Distribution suisse: Bassin lémanique et Plateau.Distribution neuchâteloise: Une seule donnée à Colombier-Planeyse.Régime alimentaire des larves: rhizophage.Apparition des adultes: d’avril à juin.
Nous n’avons trouvé cette espèce que dans la station 1, une lisière étagée ther-mophile. Dans le cas présent, Rhizotrogus aestivus privilégie nettement ce type de milieu, puisqu’il n’a été trouvé ni en alti-tude, ni en coupe d’abri. Nos observations confirment les données de la littérature en ce qui concerne sa prédilection pour les lisières mais aucune des sources consultées ne fait état du caractère thermophile de cette espèce.
Serica brunnea (L., 1758)Distribution générale: Sibérie, Europe sauf région méditerranéenne.Distribution suisse: tout le pays. Semble plus fréquente en montagne.Distribution neuchâteloise: surtout en alti-tude (Les Bayards, St-Sulpice, Pouillerel, La Chaux-du-milieu), mais également dans le bas du canton (Neuchâtel-Ermitage).Régime alimentaire des larves: rhizophage.Apparition des adultes: de juin à août.
Serica brunnea été capturée dans toutes les stations, sauf la 3 et la 6, avec une nette prédominance dans les stations 8 et 10, deux lisières étagées en hêtraie. Cette espèce se nourrit à l’état larvaire des racines de diverses plantes, notamment herbacées. Les adultes se nourrissent de feuilles et de bourgeons. Ils se rencontrent volontiers sur les arbres et buissons. (allenspacH, 1970; balacHowsky, 1962). C’est donc un insecte de milieux semi-ouverts par excel-lence. Sa présence plus forte dans les deux
s. barbalat
132

stations précitées confirme, d’une part, la plus grande fréquence de cette espèce en altitude constatée par allenspacH (1970). D’autre part, elle semble indiquer que l’es-pèce préfère les lisières aux coupes d’abri. Cela peut s’expliquer par la présence de la prairie jouxtant la lisière, dont la densité de racines est sans doute supérieure à celle d’une coupe d’abri. En effet, la densité de la strate herbacée en sous-bois est généra-lement assez faible et elle n’est guère plus fournie dans une coupe d’abri aussitôt après une coupe. La rareté de Serica brunnea aux Grattes (station 7) est un autre indice qui nous conduit à penser qu’une strate her-bacée fournie lui est favorable. Cette station (situation en altitude avec lisière étagée jouxtant une prairie maigre) présente pour-tant des caractéristiques assez comparables à celles des stations 8 et 10. Cependant, la prairie maigre des Grattes, poussant sur des dalles affleurantes est beaucoup moins fournie que celle des stations 8 et 10 crois-sant sur un sol plus profond.
Cet insecte est signalé comme forestier par balacHowsky (1962) et Hill (1987). Toutefois, nous ne l’avons trouvé qu’à un seul exemplaire en sous-bois en 1993, alors que nous l’avons capturé en bien plus grand nombre en milieu semi-ouvert.
Rutelinae
Phyllopertha horticola (L., 1758)Distribution générale: Europe, Sibérie.Distribution suisse: tout le pays. Semble plus fréquent en montagne.Distribution neuchâteloise: tout le canton.Régime alimentaire des larves: rhizophage.Apparition des adultes: de mai à juillet.
Cette espèce a surtout été capturée dans les lisières étagées d’altitude, avec une nette prédominance (105 individus) à Fre-tereules (station 8), où la lisière étagée comporte de nombreux buissons à fleurs, très favorables aux adultes phyllophages et floricoles. Elle borde une prairie de type mesobromion, propice aux larves qui se
nourrissent volontiers de racines de gra-minées. Phyllopertha horticola est moins abondante dans les autres stations d’alti-tude. Les structures à notre avis peu favo-rables à cette espèce, rencontrées dans les autres stations sont les suivantes: pour la station 7, une strate herbacée très clair-semée; pour la station 9, une lisière nette et pour la station 10, une absence de buis-sons à fleurs dans la lisière pourtant étagée (Corylus avellana étant le buisson domi-nant). D’après la répartition suisse et nos propres observations, Phyllopertha horti-cola se rencontre nettement plus fréquem-ment en montagne qu’en plaine. La litté-rature consultée ne mentionne pas cette préférence, même si elle indique que l’es-pèce peut se rencontrer jusqu’à 2500 m (allenspacH, 1970). A notre avis, cette plus grande abondance en altitude est due à une agriculture plus extensive qu’en plaine. Nous avons observé une très forte variation de l’abondance de cette espèce entre 1994 et 1995.
CetoninaeCetonia aurata (L., 1761)Distribution générale: Paléarctique.Distribution suisse: Bassin lémanique, Jura, valais, Tessin. Plus rare ailleurs.Distribution neuchâteloise: tout le canton, avec toutefois une plus grande abondance à basse altitude.Régime alimentaire des larves: saprophage.Apparition des adultes: de mai à juillet.
Cette espèce a surtout été capturée à Boudry (station 4), une coupe d’abri en chênaie. Etant donné que les adultes de Cetonia aurata sont floricoles, on aurait pu s’attendre à une plus grande abondance dans les lisières étagées comportant des buissons à fleurs, ce qui n’a pas été le cas. Les larves de cette espèce se dévelop-pent dans les vieilles souches pourries de feuillus, plus fréquentes dans une coupe d’abri que dans une lisière.
133
faunistique De 9 scarabaeiDae pHytopHaGes et De 3 lucaniDae

s. barbalat
134
Protaetia cuprea (Hbst., 1774)Distribution générale: Toute l’Europe (Janssens, 1960).Distribution suisse: Bassin lémanique, Jura, valais, Tessin, Grisons. Plus rare ailleurs.Distribution neuchâteloise: surtout dans le haut; quelques données éparses dans le reste du canton. Régime alimentaire des larves: myrméco-phile, saprophage.Apparition des adultes: de mai à août.
D’après allenspacH (1970), Pro-taetia cuprea préfère les régions chaudes et sèches. Les adultes sont floricoles sur divers arbres et buissons ou se nourrissent de la sève s’écoulant d’arbres blessés. Les larves vivent dans les nids de Formica rufa et se nourrissent de végétaux en décom-position. Nous n’avons trouvé qu’un seul individu de cette espèce en 1993 dans la station 5, effectivement assez thermophile.
Trichius fasciatus (L., 1758)Distribution générale: Europe, Caucase, Sibérie.Distribution suisse: tout le pays.Distribution neuchâteloise: tout le canton.Régime alimentaire des larves: saproxylo-phage.Apparition des adultes: de juin à août.
Nous avons trouvé Trichius fasciatus dans toutes les stations, sauf à Brot-Des-sous (station 9). Cette espèce saproxylo-phage profite des souches pourrissantes dans les coupes d’abri. En effet, elle est plus abondante dans ces dernières que dans les lisières.
conclusion
Avec 422 individus capturés, l’apport quantitatif des Scarabéides est relative-ment modeste. Leur intérêt dans le cadre de cette étude est surtout d’ordre éco-
logique car ils sont nettement favorisés par les lisières. On compte en effet une moyenne de 61,4 individus et 5 espèces dans les lisières contre respectivement 16,4 et 3 dans les coupes d’abri. Cette constatation s’explique facilement par la différence de régime alimentaire des larves et des adultes. Les larves ont besoin d’une strate herbacée fournie, dont elles mangent, pour la plupart, les racines. Les adultes se nourrissent volontiers de fleurs ou de feuilles d’arbustes. Les lisières leur sont donc tout à fait favorables. Parmi les espèces capturées, seul Trichius fasciatus paraît préférer les coupes d’abri.
Les Lucanides ne comptent que 79 individus mais sont représentés par trois espèces sténotopes, très caractéristiques des milieux où nous les avons rencontrées.
Le cas de Sinodendron cylindricum mérite tout particulièrement notre intérêt en raison des informations divergentes livrées par les différents auteurs consultés. Ce type de constatations, à première vue contradictoires, doit rendre attentif à la plasticité écologique des espèces. Elles sont nettement plus eurytopes lorsqu’on les considère sur l’ensemble de leur aire de répartition que lorsqu’on ne les étudie que dans une zone restreinte. Dans le cas pré-sent, il ne nous est pas possible de déter-miner si Sinodendron cylindricum est glo-balement eurytope et sténotope seulement par endroits en raison de facteurs précis (par exemple l’âge des forêts) ou si on se trouve en présence d’écotypes différents.
reMercieMents

135
Je tiens à exprimer ma vive reconnaissance au Professeur Willy matthey et au Dr Daniel Borcard pour le suivi de ce travail et la relecture de ce manuscrit, ainsi qu’au Dr yves Gonseth pour son aide dans l’analyse de la distribution. J’aimerais aussi remer-cier mm. Pascal Junod et milan Plachta, responsables des arrondissements forestiers concernés par cette étude, pour leur collaboration.
biblioGrapHie
allenspacH, v. 1970. Coleoptera: Scarabaeidae, Lucanidae. Insecta Helvetica. Lausanne.
balacHowsky, A. S. 1962. Entomologie appliquée à l’agriculture. Tome 1. vol. 1. masson. Paris.
barbalat, S. 1995. Efficacité comparée de quelques méthodes de piégeage sur certains Coléoptères saprophages ou xylophages et influence de l’anthophilie sur le résultat des captures. Bull. Soc. neu-châtel. Sci. nat. 118: 39-52.
barbalat, S. 1996a. Faunistique de quelques Coléoptères Buprestides capturés dans les Gorges de l’Areuse (Neuchâtel, Suisse). Bull. Soc. neuchâtel. Sci. nat. 119: 37-45.
barbalat, S. 1996b. Influence de l’exploitation forestière sur trois familles de Coléoptères liés au bois dans les Gorges de l’Areuse (Canton de Neuchâtel, Suisse). Rev. Suisse Zool. 103 (2): 1-12.
barbalat, S. & borcarD, D. 1997. Distribution of four beetle families in different forest ecotones in the Areuse Gorges (Neuchâtel, Switzerland). Ecologie 28 (3): 199-208.
barbalat, S. 1998. Effect of forest structures on four beetle families (Col., Buprestidae, Cerambycidae, phytophagous Scarabaeidae and Lucanidae) in the Areuse Gorges (Neuchâtel, Switzerland). Rev. Suisse Zool. 105: 569-580.
bily, S. 1990. Coléoptères. Gründ. Paris.
Janssens, A. 1960. Insectes: Coléoptères Lamellicornes. Faune de Belgique. Institut royal des sciences naturelles de Belgique. Bruxelles.
loHse, G. a. & lucHt, W. 1992. Die Käfer mitteleuropas Band 13 Goecke & Evers. Krefeld.
Hill, D. 1987. Agricultural insect pests of temperate regions and their control. Cambridge University Press. Cambridge.
Horion, A. 1958. Faunistik der mitteleuropäischen Käfer Band 6 Lamellicornia. Selbstverlag. Über-lingen.
kocH, K. 1992. Die Käfer mitteleuropas, Ökologie Band 3. Goecke & Evers. Krefeld.
paulian, R. 1959. Coléoptères Scarabéides. Faune de France 63. Librairie de la Faculté des Sciences. Paris.
zaHraDnik, J. & cHvala, m. 1990. La grande encyclopédie des insectes. Gründ. Paris.
__________
faunistique De 9 scarabaeiDae pHytopHaGes et De 3 lucaniDae
bull. soc. neucHâtel. sci. nat. 121 : 127-135. 1998


bulletin De la société neucHâteloise Des sciences naturelles 121 : 137-139. 1998
137
CARDIOCONDyLA EmERyI FOREL 1881 (HymENOPTERA FORmICIDAE) AU PAPILIORAmA
DE mARIN (NEUCHÂTEL, SUISSE)
EDOUARD DELLA SANTA
muséum d’Histoire naturelle, case postale 6434, 1211 Genève 6, Suisse.
mots-clés: Papiliorama - frangipanier -Cardiocondyla emeryi - Antilles.
Key-words: Papiliorama - temple tree - Cardiocondyla emeryi - Antilles.
Résumé
L’article mentionne la capture d’une fourmi exotique, Cardiocondyla emeryi, sur un frangipanier dans une serre tropicale du Papiliorama de marin, Neuchâtel, Suisse.
Summary
The article mentioned the capture of an exotic ant, Cardiocondyla emeryi, on a temple tree in a tro-pical garden, the Papiliorama of marin, Neuchâtel, Switzerland.
introDuction
En date du 18 mars 1997, lors d’une visite au Papiliorama de marin (Neuchâtel, Suisse), nous avons récolté quelques exemplaires d’une très petite espèce de fourmis, à allure de Leptothorax, sur les feuilles d’un frangipanier (Plumeria alba, famille des Apo-cynacées). Plumeria est un genre comprenant divers arbres et arbustes originaires d’Amé-rique centrale.
notes MyrMécoloGiques
Les fourmis récoltées, très actives à la surface des longues feuilles oblongues du fran-gipanier, différaient de toutes les espèces indigènes. Pour leur détermination, nous avons eu recours au Dr Janine Casewitz-Weulersse, myrmécologue au muséun national d’His-toire naturelle de Paris, qui a reconnu l’espèce Cardiocondyla emeryi Forel 1881.
Le genre Cardiocondyla, créé par Emery en 1869, est cosmopolite et comprend aujourd’hui une cinquantaine d’espèces (bolton, 1995). L’espèce-type, C. elegans Em. 1869, est l’une des deux espèces françaises. Elle est probablement très fréquente dans le midi, mais difficile à trouver (bernarD, 1968).
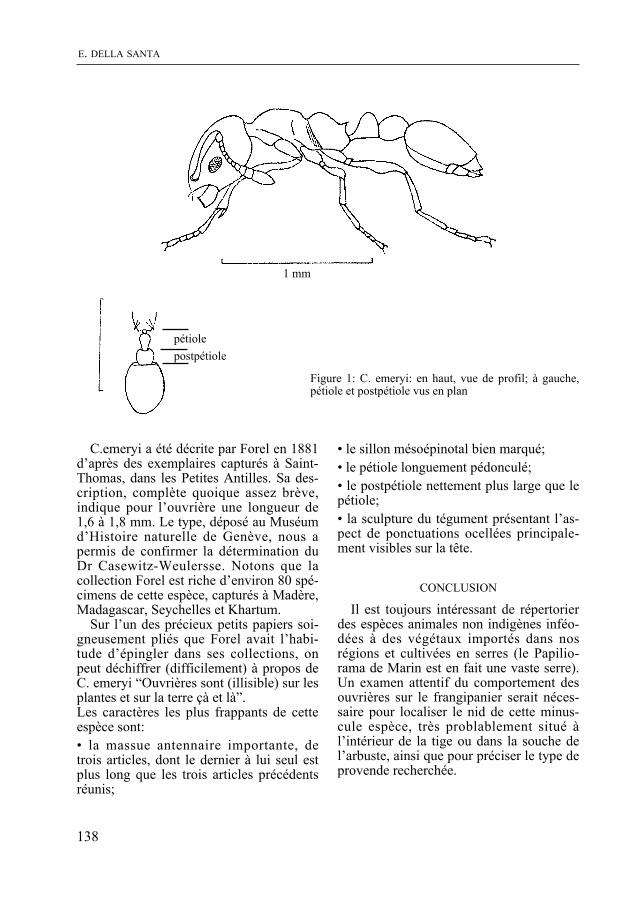
138
C.emeryi a été décrite par Forel en 1881 d’après des exemplaires capturés à Saint-Thomas, dans les Petites Antilles. Sa des-cription, complète quoique assez brève, indique pour l’ouvrière une longueur de 1,6 à 1,8 mm. Le type, déposé au muséum d’Histoire naturelle de Genève, nous a permis de confirmer la détermination du Dr Casewitz-Weulersse. Notons que la collection Forel est riche d’environ 80 spé-cimens de cette espèce, capturés à madère, madagascar, Seychelles et Khartum.
Sur l’un des précieux petits papiers soi-gneusement pliés que Forel avait l’habi-tude d’épingler dans ses collections, on peut déchiffrer (difficilement) à propos de C. emeryi “Ouvrières sont (illisible) sur les plantes et sur la terre çà et là”.Les caractères les plus frappants de cette espèce sont:• la massue antennaire importante, de trois articles, dont le dernier à lui seul est plus long que les trois articles précédents réunis;
• le sillon mésoépinotal bien marqué;• le pétiole longuement pédonculé;• le postpétiole nettement plus large que le pétiole;• la sculpture du tégument présentant l’as-pect de ponctuations ocellées principale-ment visibles sur la tête.
conclusion
Il est toujours intéressant de répertorier des espèces animales non indigènes inféo-dées à des végétaux importés dans nos régions et cultivées en serres (le Papilio-rama de marin est en fait une vaste serre). Un examen attentif du comportement des ouvrières sur le frangipanier serait néces-saire pour localiser le nid de cette minus-cule espèce, très problablement situé à l’intérieur de la tige ou dans la souche de l’arbuste, ainsi que pour préciser le type de provende recherchée.
e. Della santa
1 mm
pétiolepostpétiole
Figure 1: C. emeryi: en haut, vue de profil; à gauche, pétiole et postpétiole vus en plan

139
reMercieMents
Nous tenons à remercier très vivement le Dr J. Casewitz-Weulersse pour la détermi-nation de notre espèce, ainsi que notre collègue et ami Carlo Franscella, ancien directeur du Jardin botanique des Iles Brissago (TI) pour la documentation qu’il nous a fournie à propos de la plante-hôte.
biblioGrapHie
bernarD, F. 1968. Les Fourmis (Hymenoptera Formicidae) d’Europe occidentale. Faune de l’Europe et du Bassin méditerranéen. masson. Paris.
bolton, B. 1995. A New General Catalogue of the Ants of the World. Harvard University Press. Cambridge, massachussetts; London, England.
forel, A. 1881. Die Ameisen der Antille St Thomas. mitt. münchen. Ent. ver. 5: 1 -16.
__________
carDioconDyla eMeryi forel 1881 au papilioraMa De Marin (neucHâtel, suisse)
bull. soc. neucHâtel. sci. nat. 121 : 137-139. 1998


bulletin De la société neucHâteloise Des sciences naturelles 121 : 141-162. 1998
141
NATURE NEUCHÂTELOISE 1997
PHILIPPE JACOT-DESCOmBES1, ARTHUR FIECHTER2, LÉONARD FARRON3
1Office de la conservation de la nature, Parc 119, Case postale 1134, 2301 La Chaux-de-Fonds, Suisse.
2Service de la faune, Rue du Château 16, Case postale, 2001 Neuchâtel 1, Suisse.
3Service des forêts, Parc 119, Case postale 1367, 2301 La Chaux-de-Fonds, Suisse.
1. introDuction (pH. Jacot-DescoMbes)La protection de la nature s’appuie sur deux outils principaux, la protection par voie
d’autorité (arrêté, plan d’affectation cantonal, zones de protection communales) et la pro-tection par voie conventionnelle (contrat entre l’état et un partenaire). Ce dernier paraît au cours des ans prendre de plus en plus d’importance. Rien ne sert en effet de mettre en place des règles qui ne sont ni comprises ni acceptées. Le contrôle de l’application des mesures montre rapidement ses limites tant au niveau financier qu’au niveau de son acceptation par la population, les propriétaires et les exploitants directement touchés. L’arrêt du Tribunal fédéral concernant la protection des marais (voir point 2.1) constitue l’expression du premier principe mais ne dispensera pas notre canton de prendre en compte le second. Le plan de gestion du Loclat (voir point 7) constitue une autre voie qui cherche préalablement à mettre sur le papier les différents intérêts en présence, à en faire la synthèse puis à présenter un catalogue de mesures visant à assurer la pérénité du site et sa revitalisation. Ce “concept directeur” devient alors l’outil de travail privilégié du ges-tionnaire, indispensable à la mise sur pied d’une collaboration efficace entre les différents partenaires.
Deux outils complémentaires, deux visions de la protection de la nature dont le présent article vous donne un aperçu.
2. la léGislation (pH. Jacot-DescoMbes)2.1. L’arrêt du tribunal fédéral du 20 octobre 1997 concernant la protection des maraisSuite à la mise à l’enquête du plan de protection des marais d’importance nationale
par le canton de Neuchâtel en 1995, plusieurs opposants se sont manifestés. Parmi ceux-ci, les associations de protection de la nature reprochaient au canton de ne pas avoir rempli son mandat constitutionnel que lui avait confié le peuple en ne mettant pas en place autour de ces marais des zones-tampon (zones assurant une protection trophique,

142
pH. Jacot-DescoMbes et al.
hydrique et biologique du marais). Le Conseil d’Etat neuchâtelois considérait quant à lui que la législation fédérale allait plus loin que le mandat constitutionnel et souhaitait se donner le temps nécessaire pour négocier avec les propriétaires et les exploitants. Le Tri-bunal fédéral a tranché entre ces deux avis et a donné raison aux opposants en précisant quelques points importants:
Le Tribunal fédéral précise les fonctions que doivent assumer les zones-tampon:«Les zones-tampon sont des surfaces destinées à protéger les biotopes marécageux
ainsi que leur faune et leur flore spécifiques contre les menaces et les atteintes nuisibles en provenance des surfaces exploitées environnantes ... Les spécialistes distinguent trois catégories de zones-tampon selon les fonctions assignées à chacune d’entre elles. La zone-tampon hydrique comprend les surfaces adjacentes aux biotopes marécageux, dans lesquelles aucune modification du régime hydrique susceptible de compromettre l’approvisionnement en eau nécessaire à la conservation des marais n’est tolérée. La zone-tampon trophique inclut les terres agricoles cultivées, situées en dehors du biotope marécageux à protéger et soumises à des restrictions d’exploitation. Elle doit réduire ou prévenir l’engraissement indirect des marais pauvres en substances nutritives .... Une zone-tampon suffisante du point de vue écologique, au sens des art. 3 al. 1 OBm et OHm, doit en principe comprendre les surfaces nécessaires pour assurer les diverses fonctions énumérées ci-dessus...»
Le Tribunal fédéral ajoute:«L’art. 24sexies al. 5 Cst., adopté en votation populaire du 6 décembre 1987, place
sous protection les marais et les sites marécageux d’une beauté particulière et présentant un intérêt national. Selon les art. 18a al. 1, 23a, 23b al. 3 et 23c al. 1 LPN, il appartient au Conseil fédéral de désigner les biotopes d’importance nationale, après avoir pris l’avis des cantons, de délimiter leur situation et d’établir les buts de protection .... En vertu des art. 14 al. 2 let. D OPN, 3 al. 1 OHm et 3 al. 1 OBm, ces derniers (les cantons) sont notamment tenus de délimiter des zones-tampon suffisantes du point de vue écolo-gique..... La création de zones-tampon destinées à protéger les biotopes marécageux des menaces et des atteintes en provenance des terres agricoles environnantes fait partie des mesures de protection nécessaires à la conservation des objets protégés, de sorte que l’on ne saurait admettre que le Conseil fédéral aurait mis à la charge des cantons une obliga-tion nouvelle en leur imposant de délimiter des zones-tampon suffisantes du point de vue écologique.....»
Le Département de la gestion du territoire a donc repris le dossier, définit une planifi-cation sur trois ans des démarches à entreprendre et mandaté un bureau privé en écologie ainsi que le Service neuchâtelois de vulgarisation agricole pour appuyer l’Office cantonal de la conservation de la nature dans cette démarche.
Tous les partenaires concernés reconnaissent cependant aujourd’hui, que malgré l’arrêt du Tribunal fédéral, la discussion et la concertation avec les propriétaires et les exploi-tants agricoles sont aujourd’hui encore indispensables pour que les mesures qui seront prises soient effectives.
3. la faune sauvaGe (a. fiecHter)

143
nature neucHâteloise 1997
3.1. Faune terrestre et avifaune
a) LégislationLa loi sur la faune sauvage englobant la totalité des espèces, du 7 février 1995 et son
règlement d’exécution ainsi que le règlement de la chasse, du 27 novembre 1996 sont entrés en vigueur le 1er janvier 1997.
Si le règlement sur la faune permet légalement au Service de s’investir pour toutes les espèces, il ne change rien sur le terrain où ces tâches ont depuis toujours été accomplies.
Le règlement sur la chasse modifie profondément l’organisation de la chasse avec un permis englobant dès lors, le tir du chevreuil et du sanglier et une chasse à la carte pour d’autres animaux. Cette méthode permet au service de connaître les chasseurs et leur gibier préféré, de les réunir et de les instruire dans le but d’affiner la gestion au plus juste en ce qui concerne les objectifs choisis.
b) FauneLa grande faune qui habite nos forêts est de plus en plus dérangée par des activités et
manifestations sportives de toutes sortes mais aussi par les chiens accompagnés ou non qui se baladent dans les forêts. Entre différents services de l’Etat et les milieux de la pro-tection de la nature, nous travaillons dans le but de localiser voire canaliser ces activités afin de garantir des espaces calmes pour la grande faune.
Le KARCH, organisation suisse qui s’occupe des batraciens et reptiles a enfin opté pour la décentralisation de ces activités en la personne de m. F. Claude du CSCF à Neuchâtel. Nous avons trouvé là un responsable cantonal compétent et dynamique. Des relevés des sites de reproduction des batraciens effectués en 1997 par les gardes-faune lui ont été remis. En englobant les observations des spécialistes une première carte des sites est en préparation. L’importance de chaque site ainsi que la détermination des dif-férentes espèces reste à définir, ce qui va permettre une protection adaptée. Plus d’un km de barrière de protection a été acquis par le service au printemps 1998 pour la protection des batraciens et installées sur les voies de migration par gardes-faune et protecteurs de la nature.
L’activité du responsable cantonal des chauves-souris m. J.-D. Blant du musée d’his-toire naturelle de La Chaux-de-Fonds augmente d’année en année. mise à part l’informa-tion du public, c’est l’inventaire et la protection des sites ainsi que le sauvetage qui sont les préoccupations de ce spécialiste passionné ainsi que de ses aides.
La publicité qui a été faite pour le castor et son habitat dans notre canton a amené beaucoup de visiteurs sur le site. C’est probablement suite à ce dérangement que 2 de ces animaux au moins, sont partis vers d’autres rives.
Le grand tétras, extrêmement sensible au dérangement, occupe fréquemment notre service et notamment le garde-faune responsable de la réserve des Jordan. mis à part les photographes, ce sont les chiens errants qui nous préoccupent. L’influence négative du sanglier sur le grand tétras est également discutée.

144
pH. Jacot-DescoMbes et al.
3.2. Faune aquatique
a) LégislationLe règlement d’exécution de la loi sur la faune aquatique a été adopté par le Conseil
d’Etat le 5 novembre 1997 simultanément avec la loi sur la faune aquatique, du 28 août 1996. Cette nouvelle législation est entrée en vigueur le 1er janvier 1998.
b) FauneAprès avoir vidé le merdasson de tout poisson en 1996, une pêche électrique en 1997 a
réalisé le résultat formidable de 3300 poissons pêchés dont un grand nombre d’alevins de l’année. Ce résultat a démontré d’une part le bon fonctionnement de l’échelle à poissons permettant aux reproducteurs de monter dans l’Areuse et d’autre part, que la reproduction naturelle réussit parfaitement dans le merdasson.
Toujours dans le but de relier les différents tronçons d’un réseau aquatique pour la faune, un franchissement en forme d’échelle à poissons classique a été construit sur la chute du Pont de la Roche à Saint-Sulpice sur l’Areuse.
L’étude sur la truite du lac, mandatée par la Commission intercantonale sur la pêche dans le lac de Neuchâtel se poursuit. Pour la première fois, 21 truites ont pu être équipées d’un émetteur de 2,8 g, ceci à la pêcherie de Cortaillod. 150 truites supplémentaires ont pu être marquées à cette pêcherie, ce qui nous amène à un total de 507 truites qui se bala-dent dans notre lac équipées d’un numéro.
Les travaux à la nouvelle pisciculture de môtiers vont bon train. Cette dernière rempla-cera en partie la pisciculture de Boudry. Cette installation moderne permettra de produire différentes espèces de poissons dans de bonnes conditions. Des modifications de la pis-ciculture de Colombier sont prévues, afin de reprendre l’élevage de truites du lac et des ombles qui se réalisent actuellement à Boudry.
4. les forêts (l. farron)
4.1. Sylviculture et réserves forestièresL’activité sylviculturale, d’une activité purement éducative et extractive qu’elle fut
jadis est devenue, au fil des décennies, une véritable activité d’entretien mise au service des fonctions multiples de la forêt. marteau forestier et griffe des décideurs de terrain sont aujourd’hui, plus que jamais, des outils d’utilité publique.
C’est selon trois axes que le sylviculteur d’aujourd’hui entend agir en faveur de la bio-diversité, celui de la sylviculture proche de la nature proprement dite et toutes les précau-tions que cela suppose, celui de l’instauration de réserves forestières où la forêt est laissée à sa libre évolution naturelle et celui de surfaces de compensation écologique.
La sylviculture proche de la nature fait bien la part des choses entre les besoins de l’économie et les exigences de l’écologie (comprise dans le sens scientifique d’un terme parfois galvaudé par les temps qui courent). Le geste du sylviculteur vise à l’instauration de peuplements mélangés et diversifiés. Par la cohabitation intime d’arbres de toutes

145
nature neucHâteloise 1997
positions sociales et d’essences variées, il réalise cela de manière très appuyée dans les vallées et les montagnes par le jardinage en modelant des forêts où dominent, mélangés les uns avec les autres, l’épicéa, le sapin et le hêtre. Il le réalise aussi par la coupe pro-gressive en mosaïque dans les forêts du Littoral où le hêtre, le chêne et le pin sylvestre se partagent le terrain sans vraiment se mélanger. Dans les deux cas, l’objectif visé est l’obtention de peuplements forestiers productifs et conformes aux conditions de station. Il est vrai que les objectifs ne sont pas encore atteints partout, il reste un immense travail à poursuivre. L’œuvre du sylviculteur n’est jamais achevée, elle se place dans la continuité du temps qui passe; le sylviculteur n’a pas les facultés d’un magicien. Une belle forêt stable, équilibrée, productive, diversifiée ne se crée pas en deux coups de cuiller à pot. D’ailleurs un beau peuplement, une fois instauré n’a rien de définitif car il y a lieu d’en assurer la pérennité par une régénération bien orchestrée. Sans compter que les actes de nature sylviculturale se combinent avec les imprévus. La conduite des opérations sylvi-culturales c’est parfois la résolution de la quadrature du cercle; c’est toujours l’exercice d’un art où se mêlent l’intuition et le décryptage de situations infiniment complexes. Cela reste toutefois, quelque part, un art qui s’apprend aussi par les pieds, un art qui se nourrit de patience et du don de l’observation. La sylviculture, au sens neuchâtelois du terme, est un acte d’amour de l’humanité exploitante à l’égard du milieu forestier.
Les réserves forestières intégrales, deuxième pilier de la stratégie sylvestre, servent la cause du maintien de la biodiversité. Dans notre canton, elles sont actuellement au nombre de dix. Le résumé est présenté au tableau 1.
Nous avons donc au total 215 ha de réserves (24,5 ha appartiennent au Club Jurassien, 110 ha sont propriétés de l’Etat de Neuchâtel et 80,5 ha sont propriétés communales).
Il existe aujourd’hui plusieurs autres massifs qui sont déjà des réserves forestières inté-grales de fait, par exemple la plus grande partie de toute la bande boisée croissant dans les éboulis des Rochers des miroirs. Ce sont des parties de forêts à fonction économique peu affirmée, voire nulle. Il appartient aux propriétaires concernés de prendre à ce propos les décisions qui s’imposent lors de la révision des plans de gestion. Nous disposerons alors d’un réseau officiel de réserves qui devrait englober, à des degrés divers, tous les types d’associations forestières naturelles que compte notre canton.
quant aux surfaces de compensation écologique, troisième pilier des mesures à prendre en faveur du maintien de la biodiversité... de quoi s’agit il ?
Il s’agit de niches écologiques délibérément créées à l’intérieur même des surfaces qui affichent une vocation productrice. Cela sous forme d’arbres isolés ou de petits bou-quets d’arbres laissés à l’accomplissement du cycle complet de la vie et de la mort. Il peut s’agir aussi de façonner des lisières diversifiées et étagées car c’est dans ces zones de transition entre terrains agricoles et forestiers que la biodiversité est la plus affirmée, en particulier en ce qui concerne l’avifaune. Il peut s’agir également d’ouvrir dans les pessières et sapinières du Haut-Jura des clairières qui contribueront à créer les conditions favorables à la difficile survie du coq de bruyère et de la gélinotte. Il peut s’agir en outre de bien d’autres démarches très ciblées en faveur d’un petit étang, d’une colonie de four-milières, d’un réseau de terriers... Nous avons aujourd’hui de beaux exemples de lisières
146
pH. Jacot-DescoMbes et al.
Tableau 1: Résumé des réserves forestières intégrales

147
nature neucHâteloise 1997
étagées dans les hauts de Corcelles et des clairières sont aménagées dans la profondeur de nos joux noires.
4.2. LégislationLa loi cantonale sur les forêts, du 6 février 1996 est entrée en vigueur le 1er janvier
1997. Le Conseil d’Etat, le Département de la gestion du territoire et le service des forêts furent appelés à édicter plusieurs textes d’application. De plus un contact à été pris en septembre 1997 avec les principales sociétés et institutions neuchâteloises dont les acti-vités et les préoccupations touchent, d’une manière ou d’une autre, l’aire boisée. Les avis exprimés par ces groupements serviront à l’élaboration du Plan d’aménagement forestier.
5. les biotopes (pH. Jacot-DescoMbes)
5.1. Les maraisLes travaux de gestion dans les tourbières de la vallée des Ponts-de-martel se sont
poursuivis cette année. Ils se sont concentrés dans la région du Bois des Lattes où nous souhaitons aussi rapidement que possible terminer nos interventions afin de laisser la tourbière retrouver sa quiètude.
5.2. Les prairies et les pâturages maigresLe Service neuchâtelois de vulgarisation agricole a poursuivi cette année ses contacts
avec les agriculteurs qui exploitent des terrains agricoles maigres. Il a confirmé au cours de ce travail que de nombreuses parcelles inventoriées en 1986 avaient perdu une bonne partie de leur richesse floristique, suite à une modification de leur exploitation (sur ou sous-exploitation), conduisant à une banalisation de la flore ou à un embuissonnement excessif.
5.3. Le biotope cantonal des RoussottesCe biotope cantonal situé sur le territoire de la commune du Cerneux-Péquignot est une
ancienne retenue d’eau pour une scierie située en aval. Il a été mis sous protection canto-nale en 1969 et abrite batraciens et flore aquatique. La digue située en aval montrait ces derniers temps des signes de faiblesse, laissant par moment passer des eaux qui en ruisse-lant endommageaient un chemin agricole. Les fluctuations de la nappe étaient également peu favorables à la faune qui risquait de se retrouver par endroit à sec. C’est pourquoi nous avons entrepris des travaux de réaménagement de cette digue en marnant le secteur concerné. Cette réalisation a été complétée par la pose d’une barrière délimitant, mieux que précédemment, le biotope et sera poursuivie par quelques plantations qui compléte-ront le site. Restera alors à envisager d’entretenir régulièrement le plan d’eau qui a ten-dance, ces dernières années, à se combler.
6. Divers (pH. Jacot-DescoMbes)

148
pH. Jacot-DescoMbes et al.
6.1. Programme nature neuchâteloise 1997Cette année encore, la nature neuchâteloise a pu bénéficier du concours très apprécié
de nombreux collaborateurs au bénéfice de contrat de durée limitée dans le cadre du chô-mage. Depuis le début du mois de septembre, ce groupe s’est mis au travail pour soutenir les efforts de la Fondation de la Tourbière des Ponts, fondation dont font partie la com-mune des Ponts-de-martel, l’Etat de Neuchâtel et Pro Natura Neuchâtel. Cette fondation veut mettre en lumière les relations qui ont existé et existent encore entre l’homme et la tourbière. Ainsi, pourra se maintenir le souvenir d’un patrimoine culturel qui a marqué la nature, le paysage et l’histoire de cette vallée.
6.2. Le plan de gestion du LoclatSis sur le territoire communal de Saint-Blaise, ce petit lac présente un intérêt tout
particulier pour les batraciens. Il fait partie des sites de reproduction des batraciens d’im-portance nationale. Des passages à batraciens ont pu être aménagés en 1993 sous la route cantonale allant de Saint-Blaise à Cornaux facilitant ainsi la migration de ces espèces. mais cette seule mesure ne permet pas de résoudre à long terme tous les problèmes. La gestion de la végétation riveraine devient indispensable, l’abattage d’une populiculture doit être envisagé au cours de ces prochaines années, mais pour y mettre quoi ?... Toutes ces questions ont amené les parties intéressées à mandater un bureau privé d’écologie pour mettre sur pied un “concept directeur” ( voir point 7 du présent article).
6.3. Les agents chargés de la protection de la natureLes agents chargés de la protection de la nature (garde-faune, garde-forestier, ingénieur
forestier, etc...) continuent leur formation. Ils ont eu l’occasion cette année de découvrir le monde des prairies maigres sur les côteaux secs du pied du Jura dans la région du Lan-deron et de Lignières. Ainsi progressivement ils s’éloignent des domaines qui leurs sont plus familiers. mais ils ont pu se rendre compte que là aussi ils pouvaient faire valoir leurs connaissances puisqu’ils ont pu constater que depuis l’inventaire cantonal de 1996 de nombreuses prairies avaient tendance à être colonisées par des buissons puis par la forêt. La gestion de ce processus a pu être discutée à cette occasion.
6.4. Le Râle des genêtsLe râle des genêts (Crex crex) est la seule espèce migratrice recensée dans les listes
rouges au niveau mondial qui pourrait potentiellement nicher sur le territoire suisse. Cette année, il a été observé en plusieurs endroits dans la vallée des Ponts-de-martel et de La Sagne. Afin d’offrir à cet oiseau la possibilité de se reproduire, et à la demande de l’ASPO, l’Office de la conservation de la nature a passé avec le concours du Service de la faune et du Service neuchâtelois de vulgarisation agricole deux contrats d’exploi-tations avec les agriculteurs concernés afin que ces derniers ne fauchent leurs champs qu’au début du mois de septembre. De cette manière, nous avons donné à cet oiseau la possibilité de nidifier et aux oisillons de se développer suffisamment pour survivre. L’expérience a été suivie par télémétrie. De nombreuses observations ont été faites mais, malheureusement, aucune nidification n’a pu être constatée. Les informations recueillies à cette occasion sont cependant précieuses et permettent de mieux comprendre la bio-logie de cette espèce.

149
nature neucHâteloise 1997
6.5. Les talus routiersL’expérience mise en place en 1996 d’entretien extensif de certains talus routiers le
long des routes cantonales a été étendue cette année à l’ensemble des routes cantonales. Rappelons que dans les secteurs concernés les talus sont fauchés plus tardivement qu’à l’accoutumée. Les banquettes sont fauchées au printemps et l’ensemble du talus en automne. Le travail s’est bien déroulé cette année. Dans certains secteurs, la fauche en fin de saison a nécessité plusieurs passages, la végétation ayant eu le temps de prendre de la vigueur.
Ce travail d’extensification est possible avec un bilan financier positif ou neutre et un bénéfice pour la nature.
Fort de ce constat, le Département de la gestion du territoire a mis sur pied une séance d’information à l’attention des communes du canton pour leur faire part des essais qui ont été effectués, pour échanger des expériences et pour les inciter à se pencher sur leurs manières d’entretenir les talus routiers et les espaces verts.
6.6. Le projet de revitalisation du val-de-RuzA l’instigation de m. y. Gonseth du Centre Suisse de cartographie de la faune, un
projet soutenu financièrement par le Département de l’économie publique a été déve-loppé au courant de cette année.
Le val-de-Ruz, même s’il n’a pas globalement changé de vocation agricole au cours de ce siècle, a cependant été profondément modifié. Les remaniements parcellaires, l’industrialisation, l’exploitation de gravières et le développement des voies de commu-nication ont conduit à un isolement marqué des populations animales et végétales. Leur survie peut cependant être envisagée avec le concours de la nouvelle politique agricole qui permet de soutenir financièrement les exploitations agricoles qui acceptent les règles de la production intégrée et mettent une partie de leurs surfaces agricoles en surfaces de compensation écologique.
L’objectif premier de ce projet est de préparer un concept de réseau biologique puis de l’appliquer avec le concours des agriculteurs et des autres gestionnaires de l’espace extra urbain. Cette problématique semble avoir interpellé de nombreuses personnes puisque plusieurs services de l’Etat, des représentants d’associations de protection de la nature, de chasseurs et du milieu agricole y sont impliqués.
7. un tHèMe particulier
PLAN DE GESTION DU LOCLAT (ST-BLAISE - NE)

150
b. De MontMollin et al.
BERTRAND DE mONTmOLLIN1, GILLES mISEREZ2 et CATHERINE STREHLER1
1 Biol conseils S.A., rue de la Serre 5, 2000 Neuchâtel, Suisse.
2 Ecole d’Ingénieurs Agronomes HES, filière Gestion de la Nature, Centre de Lullier, 1254 Jussy, Suisse.
mots-clés: Loclat. Plan de gestion. Conservation
Résumé
Le Loclat est un lac naturel, bordé d’une mince ceinture végétale riveraine humide entourée de cultures et de jardins. Le site présente une grande diversité floristique et faunistique, mais l’effectif des populations est souvent faible.
Situé à proximité des zones d’habitations, le site subit une importante pression du public. De multiples activités telles que la pêche, le modélisme et le jardinage sont en effet pratiquées sur le lac et ses berges. Pour assurer le maintien et la revitalisation des biocénoses existantes, tout en conservant, dans la mesure du possible, les différents utilisateurs du site, un plan de gestion a été établi. Il repose sur des mesures de protection, d’aménagement, de revitalisation, d’entretien et de réglementation. Une conciliation des intérêts de la nature et des utilisateurs a été recherchée lors de l’élaboration des mesures. Une partie d’entre elles consistent en des interventions ponctuelles, sans interférences directes avec les utilisateurs du site et susceptibles par conséquent d’être réalisées rapidement. D’autres tendent à une revitalisation plus globale du Loclat. Elles sont plus difficiles à mettre en oeuvre car elles touchent de multiples utilisateurs et impliquent de leur part des conces-sions. Elles sont toutefois aussi les seules qui permettront de garantir à long terme la pérennité des milieux naturels existants.
introDuction
Le Loclat est un lac naturel d’origine glaciaire figurant à l’Inventaire des sites de reproduction de batraciens d’importance nationale de 1994.
Situé à quelques centaines de mètres des zones d’habitations de la commune de St.-Blaise, le site du Loclat est prisé du public qui y pratique diverses activités de loisirs telles que la pêche, le modélisme et le jardinage. La pression des différents utilisateurs sur les milieux naturels est importante. Elle se traduit notamment par le réempoissonne-ment du lac ainsi que par la régression de la surface, de la diversité et de la vitalité de la ceinture végétale du lac, qui ne comprend actuellement plus que des lambeaux de rose-lières et de prairies à laîches. La dégradation et la diminution des milieux naturels ayant pour corollaire la disparition de certaines espèces, l’Office cantonal de conservation de la nature a fait établir un plan de gestion destiné à:• protéger les reliques de végétation naturelle; • éviter la disparition des espèces les plus vulnérables; • préserver les populations de batraciens existantes et permettre leur reproduction au Loclat.
La réalisation d’un plan de gestion était également motivée par l’exploitation prévue de la populiculture existante.

151
plan De Gestion Du loclat (st-blaise - ne)
Les mesures proposées ont été accompagnées d’une évaluation de leur faisabilité tech-nique et économique, d’un calendrier de réalisation ainsi que d’un protocole de suivi.
bases MétHoDoloGiques
Le cadre géographique de l’étude correspond au périmètre de l’objet 3.1 de l’Inventaire Nature et Paysage du Plan directeur cantonal. Les données de base ont été recueillies auprès des services cantonaux et communaux concernés, d’associations de protection de la nature, des Universités de Neuchâtel et de Lausanne, du Centre Suisse de Cartographie de la Faune, du musée d’histoire naturelle de La Chaux-de-Fonds, du Centre de Coordi-nation pour la Protection des Amphibiens et des Reptiles de Suisse, du bureau Aquarius, des Chasseurs sans Fusil, de l’Association des Amis du Loclat et de naturalistes ama-teurs.
Ces données ont été en partie vérifiées et complétées par des inventaires de terrain effectués en 1996.
La rareté des espèces animales et végétales a été évaluée sur la base des:• Listes rouges des espèces animales menacées de Suisse (Duelli, 1994) • Plantes vasculaires menacées en Suisse. Listes rouges nationales et régionales (lan-Dolt, 1991)
La nomenclature utilisée pour les plantes est celle de l’Index synonymique de la flore suisse (aescHiMann & Heitz, 1996)
etat actuel
Biotope: Le Loclat est à la limite de deux zones biogéographiques: la zone du Plateau (2.1) et celle du Jura occidental (1.1) (lanDolt, 1991). Situé à une altitude de 432 m dans une plaine alluviale constituée de dépôts sédimentaires, sa profondeur maximale est de 9.2 m. Ses plus grandes dimensions sont de 480 m sur 100 m. Il n’a actuellement plus d’affluent ou d’effluent à l’air libre et est alimenté par des collecteurs de drainage. Son trop-plein se déverse dans le lac de Neuchâtel par l’intermédiaire d’une conduite qui rem-place l’ancien ruisseau du mouson.
Le caractère originel du site a progressivement été modifié au cours de ce siècle. Les terrains tourbeux attenant au lac ont été drainés pour permettre une intensification des cultures. Les prés à litière ont fait place à des terres ouvertes. Les rares couloirs d’échanges biologiques qui reliaient le lac aux autres écosystèmes ont progressivement disparu, ce qui a entraîné l’isolement du site. Une populiculture a été plantée, contribuant à assécher les milieux humides. Des jardins familiaux ont été aménagés, favorisant l’ar-rivée d’espèces non indigènes (carte 1).
Flore et végétation: La ceinture de végétation du Loclat est discontinue, parfois large de seulement 50 cm. Elle comprend des reliques:• de roselières• de mégaphorbiaies eutrophes en partie atterries et envahies par des aulnes et des saules• de prairies à grandes laîches
b. De MontMollin et al.
152
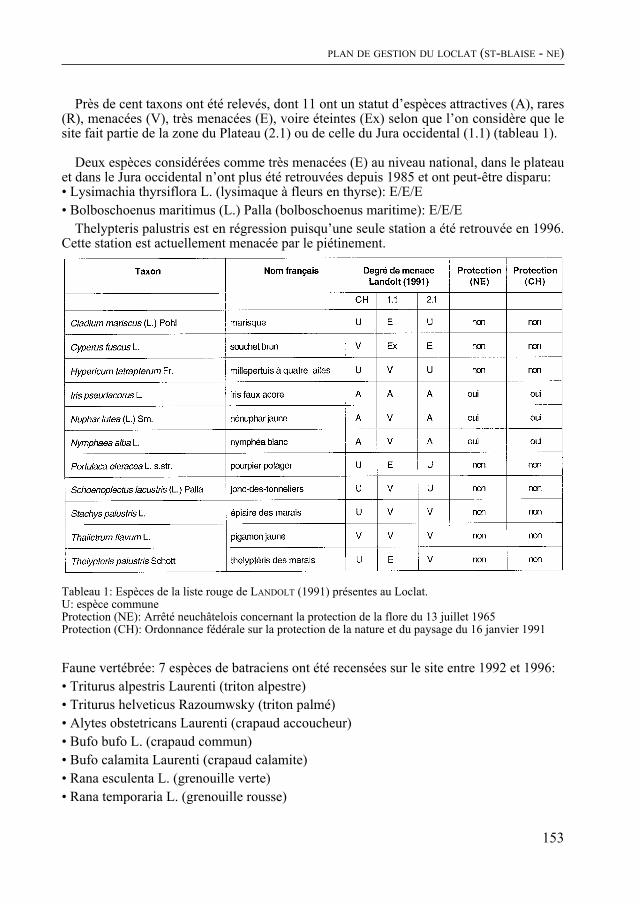
153
plan De Gestion Du loclat (st-blaise - ne)
Près de cent taxons ont été relevés, dont 11 ont un statut d’espèces attractives (A), rares (R), menacées (v), très menacées (E), voire éteintes (Ex) selon que l’on considère que le site fait partie de la zone du Plateau (2.1) ou de celle du Jura occidental (1.1) (tableau 1).
Deux espèces considérées comme très menacées (E) au niveau national, dans le plateau et dans le Jura occidental n’ont plus été retrouvées depuis 1985 et ont peut-être disparu: • Lysimachia thyrsiflora L. (lysimaque à fleurs en thyrse): E/E/E• Bolboschoenus maritimus (L.) Palla (bolboschoenus maritime): E/E/E
Thelypteris palustris est en régression puisqu’une seule station a été retrouvée en 1996. Cette station est actuellement menacée par le piétinement.
Faune vertébrée: 7 espèces de batraciens ont été recensées sur le site entre 1992 et 1996:• Triturus alpestris Laurenti (triton alpestre)• Triturus helveticus Razoumwsky (triton palmé)• Alytes obstetricans Laurenti (crapaud accoucheur)• Bufo bufo L. (crapaud commun)• Bufo calamita Laurenti (crapaud calamite)• Rana esculenta L. (grenouille verte)• Rana temporaria L. (grenouille rousse)
Tableau 1: Espèces de la liste rouge de lanDolt (1991) présentes au Loclat.U: espèce communeProtection (NE): Arrêté neuchâtelois concernant la protection de la flore du 13 juillet 1965Protection (CH): Ordonnance fédérale sur la protection de la nature et du paysage du 16 janvier 1991

154
b. De MontMollin et al.
A l’exception du triton alpestre et de la grenouille rousse, toutes ces espèces sont qualifiées de menacées dans le nord de la Suisse (GrossenbacHer, 1994). Le crapaud commun est actuellement le batracien le plus abondant au Loclat. Des passages sous la route cantonale ont été aménagés en 1992 pour faciliter sa migration vers l’adret du massif des Roches de Châtoillon où il hiverne. quelques améliorations ponctuelles per-mettraient de renforcer l’efficacité de ces passages.
Le lac abrite une quinzaine d’espèces de poissons. Plusieurs d’entre elles sont issues de réempoissonnements. Une espèce est considérée comme fortement menacée: Rhodeus sericeus amarus Bloch (bouvière).
En raison de la diversité de ses milieux et de la proximité de forêts, le Loclat permet à un grand nombre d’oiseaux nichant aux alentours de venir se nourrir. Situé sur un axe important de migration du plateau, il sert de relais pour les oiseaux migrateurs (repos, nourrissage). Sur la base des inventaires de MenDoça (1995) et de ceux effectués en 1996, 67 espèces, dont 42 nicheuses sont présentes sur le site. Deux d’entre elles sont menacées selon la liste rouge de zbinDen et al. (1994):• vanellus vanellus (vanneau huppé) • Picus canus (pic cendré)
Plusieurs espèces, autrefois nicheuses au Loclat, ont aujourd’hui disparu, comme par exemple la caille, le courlis cendré, la huppe, le torcol, la fauvette grisette ou le traquet tarier.
Hormis quelques loirs dans les chalets du Loclat, les mammifères du site sont com-muns.
Faune invertébrée: Le Loclat abrite au moins deux espèces d’odonates menacées au Nord des Alpes:• Erythromma najas Hansemann (la naïade à yeux rouges)• Gomphus pulchellus Selys (le gomphe joli)
Utilisation actuelle du sitePêcheurs: quelque 200 pêcheurs fréquentent annuellement Le Loclat, réempoissonné régulièrement avec des brochets, des brèmes et des gardons.
Aucun aménagement particulier n’a été réalisé jusqu’à présent pour les pêcheurs qui utilisent la totalité des rives. Ils se concentrent toutefois de préférence dans les secteurs où le plan d’eau est le plus accessible et le plus dégagé. Leur passage occasionne par endroits des dégradations importantes, en particulier dans les lambeaux de roselières et de cariçaies.Agriculteurs et locataires de jardins: Aux abords immédiats du lac, une majorité des sols voués à l’agriculture sont exploités de manière intensive sous forme de terres ouvertes (maïs, céréales). Ces terres sont cultivées jusqu’aux berges.
Les jardins familiaux occupent une grande partie des surfaces communales attenantes au lac. Ces jardins comportent pour la plupart une maisonnette et sont aménagés avec des arbres et arbustes exotiques: thuyas, sapins bleus, saules pleureurs, .... Devant les jar-dins, la végétation des rives est régulièrement tondue jusqu’au lac et les déchets de tonte laissés sur les berges.

155
plan De Gestion Du loclat (st-blaise - ne)
Autres utilisateurs: Le Loclat est également un lieu prisé des promeneurs et d’autres types d’utilisateurs tels que des modélistes. La pression des modélistes sur les rives est relati-vement faible et ponctuelle dans la mesure où elle s’exerce sur la rive sud-ouest unique-ment, déjà marquée par les activités humaines et qu’elle est limitée dans le temps.
Les promeneurs qui souhaitent faire le tour du Loclat sont contraints à certains endroits d’en suivre la berge en raison des clôtures des jardins familiaux. Ils contribuent ainsi à piétiner ce qui subsiste de végétation riveraine naturelle sur le site.
Le lac est peu fréquenté par les baigneurs, notamment en raison de ses berges abruptes.
conflits actuels et Menaces
Conflits utilisateurs - végétation (carte 2)
Pêcheurs: Les pêcheurs investissent la totalité des rives, mais se concentrent sur les points les plus attractifs, provoquant ainsi une forte pression sur la flore (piétinement). La station de Thelypteris palustris située sur le promontoire au nord du lac est régulièrement piétinée.Promeneurs: La principale pression qu’exercent les promeneurs sur la végétation est le piétinement. Bien qu’aucun chemin ne soit aménagé, les lieux de passage et d’arrêt (places de pique-nique) des promeneurs se marquent très clairement, d’autant plus dans les endroits les plus vulnérables (roselières, prairies à laîches). modélistes: Le conflit se limite à un seul secteur de la rive sud (promontoire). L’atteinte à la végétation est comparable à celle exercée par les pêcheurs et les promeneurs (piétine-ment). Jardiniers: Les locataires ou propriétaires de parcelles jardinées sur les rives du Loclat exercent une pression importante sur la végétation par le biais: • des engrais qui accentuent l’eutrophisation du lac et le développement d’espèces nitro-philes;• de l’entretien intensif qu’ils pratiquent sur les berges. Les tontes fréquentes empêchent la flore d’arriver à maturité et limitent les possibilités de semis naturel. La végétation aquatique subit la concurrence d’espèces plus aptes à supporter les coupes; • des espèces non indigènes qu’ils introduisent et qui modifient la composition originelle de la végétation;• des plantations qui modifient les conditions stationnelles (assèchement des terrains, diminution de l’ensoleillement au sol) et compromettent la pérennité de certaines espèces (Cyperus fuscus par exemple);• des clôtures en bordure du lac qui canalisent les promeneurs et les pêcheurs sur la frange de végétation la plus sensible (prairies à laîches, roselière).
Conflits utilisateurs-faunePêcheurs: Le réempoissonnement du Loclat avec des brochets nuit à la reproduction des batraciens. Ce phénomène est encore accentué par le déséquilibre actuel entre les pois-sons proies (poissons blancs) et les poissons prédateurs. De plus, les refuges pour batra-ciens sont très limités sur les rives en raison du manque de végétation.
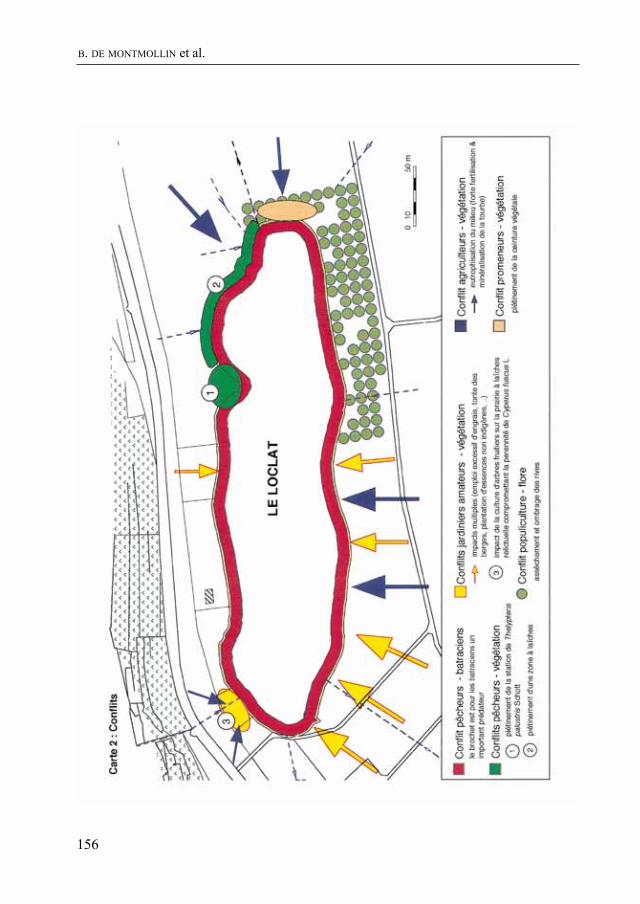
156
b. De MontMollin et al.

157
plan De Gestion Du loclat (st-blaise - ne)
Promeneurs: L’activité de reproduction des batraciens étant en majorité nocturne, les per-turbations occasionnées par les promeneurs sont minimes. En revanche, les chiens, s’ils ne sont pas en laisse, peuvent dégrader les sites de ponte et perturber la faune.
Conflits utilisateurs - biotope L’agriculture pratiquée autour du lac est intensive, exigeant une forte fertilisation. Les
cultures couvrent peu le sol, ce qui favorise son érosion et l’entraînement d’éléments nutritifs. De plus, la minéralisation de la tourbe, accélérée par les drainages, charge constamment les eaux de ruissellement et de percolation en substances nutritives, contri-buant à l’eutrophisation du lac et de sa flore.
Conflits entre utilisateurs (locataires de jardins - pêcheurs/promeneurs)Ce type de conflit provient essentiellement de la tendance de certains locataires de jar-
dins familiaux à accaparer ou à contrôler les rives du lac, ce qui entraîne régulièrement des frictions entre locataires et promeneurs ou pêcheurs.
Les pressions des différents utilisateurs s’ajoutant les unes aux autres, une conciliation devra permettre de ménager au mieux les intérêts de chacun.
Définition Des obJectifs
Pour permettre de concilier la valeur naturelle et paysagère du site avec les intérêts des utilisateurs, le plan de gestion doit intégrer non seulement des objectifs de nature écolo-gique, mais également les besoins et les attentes des utilisateurs qui sont nombreux dans cette région située à proximité immédiate de zones d’habitations. Il serait en effet illu-soire de vouloir restreindre la fréquentation du Loclat.
Les objectifs prioritaires retenus pour l’établissement du plan de gestion sont:Paysage• redonner au site un aspect proche de son caractère originel.
Flore et végétation• protéger la végétation naturelle subsistant afin d’assurer la survie à court et à long terme des espèces rares et menacées qu’elles abritent; • éviter la disparition des espèces les plus vulnérables présentes au Loclat de manière relictuelle; • renforcer la ceinture végétale aquatique naturelle; • recréer à moyen terme une végétation oligotrophe sur les berges du lac; • extensifier les cultures agricoles autour du Loclat; • supprimer progressivement les espèces non indigènes sur le site.
FauneBatraciens• préserver les populations existantes et permettre leur reproduction au Loclat; • limiter les obstacles sur le trajet de leur migration; • diversifier les sites de reproduction existants.

158
b. De MontMollin et al.
Poissons• recréer un équilibre naturel entre les espèces prédatrices et leurs proies. Cet aspect sera traité par le Service de la faune.
mammifères et avifaune• diversifier les niches écologiques sur le site; • relier le Loclat aux autres milieux naturels (corridors biologiques).
Invertébrés• intégrer la conservation des invertébrés dans le cadre des mesures prises pour conserver les habitats naturels du site.
UtilisateursLe but principal est de maintenir les différents utilisateurs actuels du site en conciliant
au mieux les objectifs écologiques cités ci-dessus avec leurs attentes qui sont:• pour le club de modélisme, le déplacement du ponton existant plus à l’ouest ainsi que l’amélioration des conditions de stationnement; • pour les promeneurs, l’aménagement d’un sentier, de places de repos, d’aires de pique-nique et de jeux dans la partie ouest du lac; • pour les jardiniers-amateurs, l’amélioration de l’émissaire du lac de manière à éviter les inondations hivernales de leurs jardins ainsi que la mise en place de poubelles pour les promeneurs et les pêcheurs; • pour les pêcheurs, un accès libre à la totalité des rives (débroussaillement), une régle-mentation en matière de pêche plus stricte et un rempoissonnement plus équilibré; • pour les agriculteurs, le maintien, voire l’amélioration des conditions de cultures exis-tantes.
plan De Gestion
Le plan de gestion du Loclat est prévu pour une durée de 15 ans. Il repose sur cinq types de mesures (tableau 2 et carte 3):
- des mesures de protection; - des mesures d’aménagement; - des mesures de revitalisation; - des mesures d’entretien; - des mesures de réglementation.Chaque mesure a fait l’objet d’une fiche qui contient les éléments nécessaires à sa mise
en oeuvre. La coordination et le contrôle des mesures seront assurés par le responsable du plan de gestion qui veillera au respect et à l’adaptation annuelle, si nécessaire, du calen-drier des travaux.
Les mesures proposées font l’objet d’une évaluation de leur priorité et de leur faisabi-lité selon une échelle allant de 1 à 3 définie comme suit:
Priorité 1: mesure indispensable pour éviter la disparition d’une espèce ou d’un milieu menacé et dont la mise en œuvre est urgente.
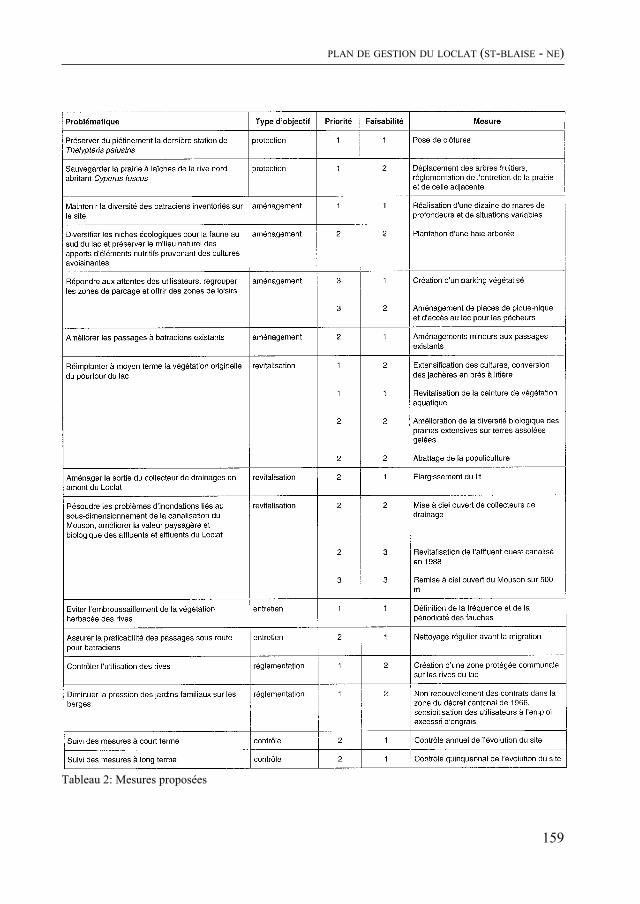
159
plan De Gestion Du loclat (st-blaise - ne)
Tableau 2: mesures proposées
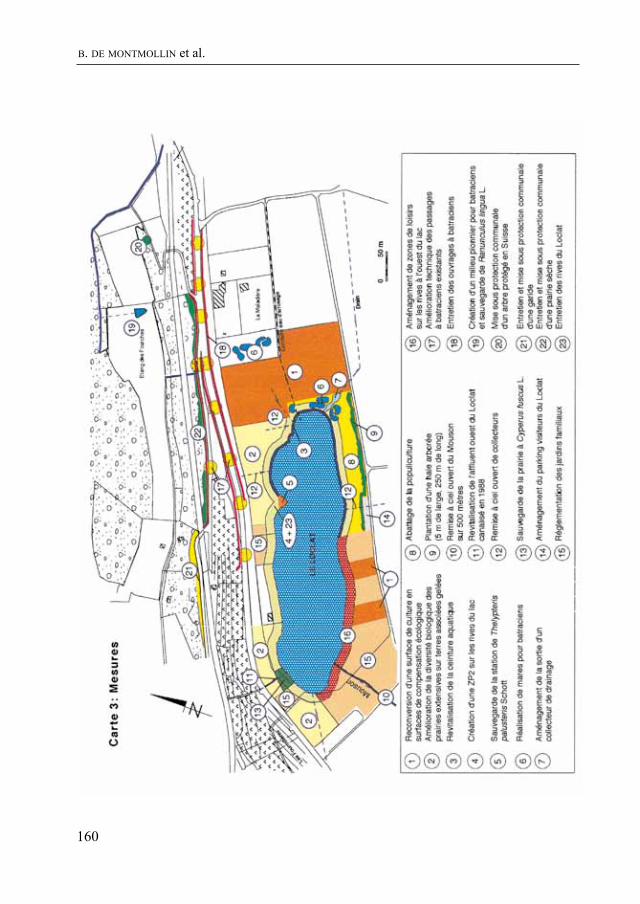
160
b. De MontMollin et al.

161
plan De Gestion Du loclat (st-blaise - ne)
Priorité 2: mesure nécessaire pour conserver et revitaliser les milieux existants et dont la mise en oeuvre devrait être assurée dans les 2 années à venir.Priorité 3: mesure souhaitable pour accroître la valeur paysagère et écologique du site ou destinée aux utilisateurs et dont la mise en oeuvre n’est pas urgente.Faisabilité 1: mesure peu coûteuse ou dont la réalisation peut être assurée par des chantiers de chômeurs et ne comportant pas ou peu d’obstacles administratifs.Faisabilité 2: mesure souvent relativement coûteuse, comportant des obstacles administratifs ou nécessitant des négociations (traitement des oppositions des propriétaires).Faisabilité 3: mesure onéreuse nécessitant des autorisations particulières.
La mise en oeuvre du plan de gestion ainsi que la réalisation des mesures proposées nécessiteront, en plus de moyens financiers, la participation active des autorités commu-nales et cantonales et des différents intervenants concernés.
reMercieMents
Nos plus vifs remerciements vont à mme m.-m. Duckert-Henriod et m. A. Scher-tenleib pour leur disponibilité et leur engagement ainsi qu’à m. Ph. Jacot-Descombes, Conservateur de la Nature pour son soutien.
Cette étude a été financée par l’Office cantonal de la conservation de la nature, avec le soutien du Service cantonal de la Faune et une subvention de l’Office fédéral de l’envi-ronnement, des forêts et du paysage. Elle a été en partie réalisée dans le cadre du travail de diplôme de Gilles miserez (cours ECOFOC de l’Université de Neuchâtel).
biblioGrapHie
aescHiMann, D. & Heitz, C. 1996. Index synonymique de la Flore de Suisse et territoires limitrophes (ISFS). Documenta Floristicae Helvetiae No 1. Genève.
Duelli, P. 1994. Listes rouges des espèces animales menacées de Suisse. Office fédéral de l’environ-nement, des forêts et du paysage, Berne.
GrossenbacHer, K. 1994. Liste rouge des amphibiens menacés de Suisse. In: Listes rouges des espèces menacées de Suisse. Office fédéral de l’environnement, des forêts et du paysage, Berne.
lanDolt, E. 1991. Plantes vasculaires menacées en Suisse. Listes rouges nationales et régionales. Office fédéral de l’environnement, des forêts et du paysage, Berne.
MenDoça, P. 1995. Suivi de la migration printanière des amphibiens de la région du Loclat à St-Blaise. KARCH, Berne.

162
zbinDen, n., Glutz von blotzHeiM, u.n., scHniD, H. & scHifferli, l. 1994. Listes des oiseaux nicheurs de Suisse avec le degré de menace dans les différentes régions. In: Listes rouges des espèces menacées de Suisse. Office fédéral de l’environnement, des forêts et du paysage, Berne.
__________
b. De MontMollin et al.
bull. soc. neucHâtel. sci. nat. 121 : 141-162. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 163-166. 1998
163
ANALySE AÉROPALyNOLOGIqUE à NEUCHÂTEL ET à LA CHAUX-DE-FONDS EN 1997
FRANÇOISE vUILLEmIN, mAy UDRIET et BERNARD CLOT
Institut Suisse de météorologie et Laboratoire de Phanérogamie, Université de Neuchâtel, Rue Emile-Argand 11, 2007 Neuchâtel, Suisse.
introDuction
Ce rapport présente les résultats des deux stations neuchâteloises du réseau national de mesure du pollen. La méthode de travail et l’emplacement des capteurs sont identiques à ceux des années précédentes (peeters et al., 1998).
Les analyses ont commencé à Neuchâtel le 6 janvier et à La Chaux-de-Fonds le 5 février; dans les deux stations elles ont pris fin le 30 septembre. Aucune panne n’a eu lieu durant la saison 1997.
résultats et Discussion
L’année 1997, avec des températures excédentaires de 1,5 à 2 degrés, compte parmi les plus chaudes du siècle; février et mars, ainsi que les mois d’août à décembre, trop chauds, ont largement contribué à ce fait. La végétation présentait ainsi une avance de 3 semaines au mois de mars. Bien que froid, avril a été le mois le plus ensoleillé de l’année avec par endroit 195 % de l’ensoleillement normal.
Les deux diagrammes de la page 166 résument la saison pollinique 1997. La rubrique “arbres divers” regroupe principalement Ulmus, diverses oléacées (sans Fraxinus), Pla-tanus, Acer et Tilia; “conifères divers” Picea, Cedrus et Abies; “Poaceae” les graminées sauvages et cultivées; “herbacées diverses” Plantago, Chenopodium, Cyperaceae et Aste-raceae (sans Artemisia ni Ambrosia). Le tableau I présente les valeurs importantes pour les pollens qui jouent les rôles les plus significatifs dans les cas d’allergies.
Les aunes ont abondamment fleuri en février. Au cœur de l’hiver, de nombreuses personnes ont ainsi souffert de pollinose. Celle-ci est alors souvent confondue avec un simple rhume. Un deuxième pic de pollen d’aune est apparu à Neuchâtel du 4 au 11 juin, dû à Alnus viridis. Ce pic est nettement moins important à La Chaux-de-Fonds cette année, alors que l’an passé la situation inverse était observée. Ce pollen est transporté par le vent depuis les Alpes, où l’aune vert est largement répandu, alors qu’il est presque inexistant dans le canton.
La quantité totale de pollen de noisetier observée cette année a été nettement plus faible que la moyenne: 650 pollens/m3 à Neuchâtel comparés à 1442 pollens/m3 en 1994,

164
année d’abondance. La floraison a duré un mois cette année contre deux et demi trois ans plus tôt. Rappelons que 1994 avait été particulièrement doux et humide par rapport à 1997.
A la mi-mars, avec plus de trois semaines d’avance grâce à février et mars plus chauds que normal, bouleaux et frênes ont commencé de fleurir. Ces arbres sont la principale source de pollinoses dans notre région après les graminées.
La pleine floraison des bouleaux a duré 18 jours à Neuchâtel, du 26 mars au 12 avril, avec une petite accalmie les 29 et 30 mars en raison de la pluie. A La Chaux-de-Fonds, le graphe montre l’existence de deux séries de pics à un mois d’intervalle, début avril et début mai (fig. 1). La première série concerne principalement du pollen transporté par le vent, la seconde correspond à la floraison des bouleaux locaux, dont le pollen est libéré plus tard dans la saison en raison de l’altitude de la ville du Haut.
La floraison du frêne a duré un mois et demi à Neuchâtel et plus de deux mois à La Chaux-de-Fonds. Elle a pris du retard en avril à cause du froid. Fin avril et début mai, chênes et platanes ont fait perdurer les allergies dues aux arbres.
Comme pour le bouleau, le décalage de floraison entre le bas et le haut du canton est marqué pour le frêne et plusieurs autres arbres à floraison printanière.
Depuis le début de mai, les pollens de graminées ont dominé dans l’air. La fin du mois de mai et les deux premières décades de juin ont ainsi été particulièrement pénibles pour les personnes allergiques, qui ont souffert des classiques symptômes du rhume des foins.
f. vuilleMin et al.
Figure 1: pollen de bouleau dans l’air en 1997
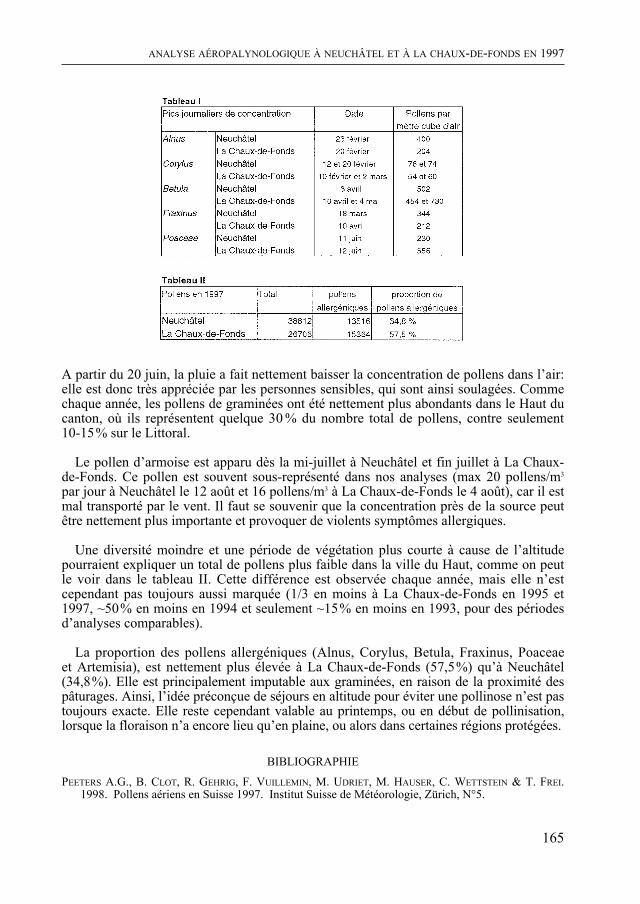
165
A partir du 20 juin, la pluie a fait nettement baisser la concentration de pollens dans l’air: elle est donc très appréciée par les personnes sensibles, qui sont ainsi soulagées. Comme chaque année, les pollens de graminées ont été nettement plus abondants dans le Haut du canton, où ils représentent quelque 30 % du nombre total de pollens, contre seulement 10-15% sur le Littoral.
Le pollen d’armoise est apparu dès la mi-juillet à Neuchâtel et fin juillet à La Chaux-de-Fonds. Ce pollen est souvent sous-représenté dans nos analyses (max 20 pollens/m3 par jour à Neuchâtel le 12 août et 16 pollens/m3 à La Chaux-de-Fonds le 4 août), car il est mal transporté par le vent. Il faut se souvenir que la concentration près de la source peut être nettement plus importante et provoquer de violents symptômes allergiques.
Une diversité moindre et une période de végétation plus courte à cause de l’altitude pourraient expliquer un total de pollens plus faible dans la ville du Haut, comme on peut le voir dans le tableau II. Cette différence est observée chaque année, mais elle n’est cependant pas toujours aussi marquée (1/3 en moins à La Chaux-de-Fonds en 1995 et 1997, ~50% en moins en 1994 et seulement ~15% en moins en 1993, pour des périodes d’analyses comparables).
La proportion des pollens allergéniques (Alnus, Corylus, Betula, Fraxinus, Poaceae et Artemisia), est nettement plus élevée à La Chaux-de-Fonds (57,5%) qu’à Neuchâtel (34,8%). Elle est principalement imputable aux graminées, en raison de la proximité des pâturages. Ainsi, l’idée préconçue de séjours en altitude pour éviter une pollinose n’est pas toujours exacte. Elle reste cependant valable au printemps, ou en début de pollinisation, lorsque la floraison n’a encore lieu qu’en plaine, ou alors dans certaines régions protégées.
biblioGrapHie
peeters a.G., b. clot, r. GeHriG, f. vuilleMin, M. uDriet, M. Hauser, c. wettstein & t. frei. 1998. Pollens aériens en Suisse 1997. Institut Suisse de météorologie, Zürich, N°5.
analyse aéropalynoloGique à neucHâtel et à la cHaux-De-fonDs en 1997

166
f. vuilleMin et al.
Neuchâtel
Pollens par mètre cube d'air par décade 1 à 9 10 à 99 100 à 9999 1000 et plus
janvier février mars avril mai juin juillet août septAlnusCorylusBetulaCarpinusSalixPopulusFraxinusQuercusFagusCastaneaArbres diversTaxus + CupressaceaePinusConifères diversPoaceaeRumexUrticalesArtemisiaAmbrosiaHerbacées diverses
4x3.3 mm
La Chaux-de-Fonds
Pollens par mètre cube d'air par décade 1 à 9 10 à 99 100 à 9999 1000 et plus
janvier février mars avril mai juin juillet août septAlnusCorylusBetulaCarpinusSalixPopulusFraxinusQuercusFagusCastaneaArbres diversTaxus + CupressaceaePinusConifères diversPoaceaeRumexUrticalesArtemisiaAmbrosiaHerbacées diverses
bull. soc. neucHâtel. sci. nat. 121 : 163-166. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 167-171. 1998
167
DIAGNOSTIC PARASITAIRE ET DE mALADIES TRANS-mISES PAR LES TIqUES DANS LE CANTON DE NEUCHA-
TEL / RAPPORT D’ACTIvITE 1997
yvES mOOSmANN1, BERNARD RUTTI1, HANS H. SIEGRIST2 et mICHEL BROSSARD1
1 Institut de Zoologie. Rue Emile-Argand 9, 2007 Neuchâtel, Suisse.2 Institut Neuchâtelois de microbiologie. Rue Sophie mairet 17, 2300 La Chaux-de-Fonds, Suisse.
introDuction
Les résultats des analyses effectuées au cours de l’année 1997 au Laboratoire de Dia-gnostic Parasitaire de l’Institut de Zoologie de l’Université de Neuchâtel sont présentés. Afin de donner un aperçu plus complet de l’activité diagnostique dans le canton de Neu-châtel, les résultats des examens parasitologiques de l’Institut Neuchâtelois de microbio-logie de La Chaux-de-Fonds sont inclus dans ce rapport. Des recherches de parasites par examen direct ont été effectuées chez 700 patients à Neuchâtel et 532 à La Chaux-de-Fonds. De plus, 2568 personnes ont subi un examen sérologique (sérologie parasitaire ou de maladies transmises par les tiques) dans notre laboratoire. La technique de mise en évidence par PCR (polymerase chain reaction) d’ADN de Borrelia burgdorferi, agent de la borréliose de Lyme, a été utilisée sur des échantillons provenant de 152 patients.
résultats et coMMentaires
maladies parasitairesParasites sanguins: examens directs
Une recherche de parasites sanguins sur frottis et gouttes épaisses a été effectuée pour 72 patients. Des plasmodies ont été retrouvées chez 12 d’entre eux (tab. 1), avec une pré-dominance de l’espèce Plasmodium falciparum (8 cas).
Parasites intestinaux: examens directsUn ou plusieurs échantillons pour 1160 personnes ont été analysés. Des formes évolu-
tives de protozoaires et d’helminthes ont été découvertes chez 257 d’entre elles (tab.1). Un quart environ (69 sur 257) des patients hébergeaient au moins deux espèces.
20 espèces de parasites ont été diagnostiquées, souvent chez des personnes ayant séjourné sur d’autres continents. Les voyages à l’étranger ne sont pas toujours signalés au laboratoire, ce qui explique le grand nombre de parasites de provenance inconnue. Parmi les protozoaires intestinaux, relevons ceux provoquant des signes cliniques: Enta-
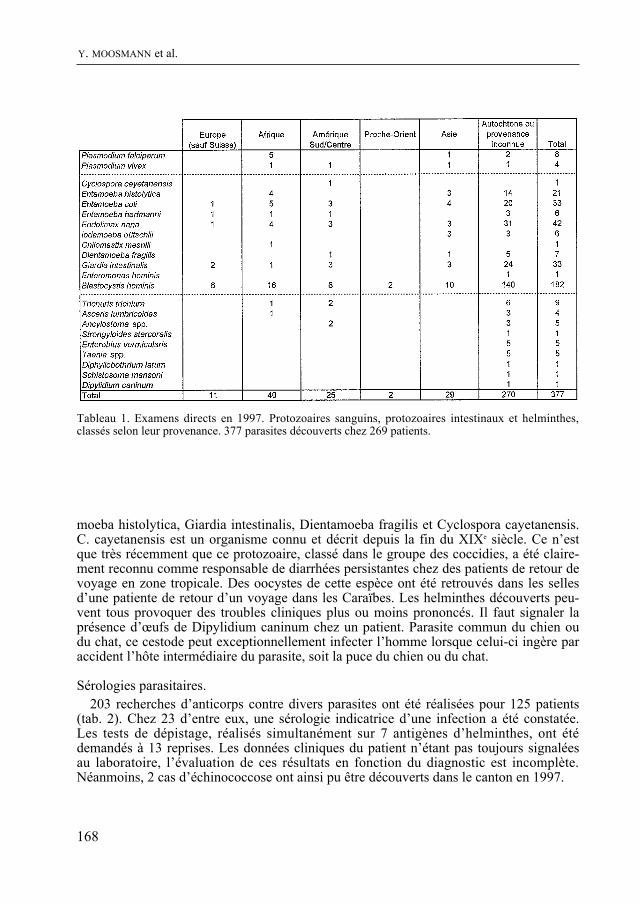
168
moeba histolytica, Giardia intestinalis, Dientamoeba fragilis et Cyclospora cayetanensis. C. cayetanensis est un organisme connu et décrit depuis la fin du XIXe siècle. Ce n’est que très récemment que ce protozoaire, classé dans le groupe des coccidies, a été claire-ment reconnu comme responsable de diarrhées persistantes chez des patients de retour de voyage en zone tropicale. Des oocystes de cette espèce ont été retrouvés dans les selles d’une patiente de retour d’un voyage dans les Caraïbes. Les helminthes découverts peu-vent tous provoquer des troubles cliniques plus ou moins prononcés. Il faut signaler la présence d’œufs de Dipylidium caninum chez un patient. Parasite commun du chien ou du chat, ce cestode peut exceptionnellement infecter l’homme lorsque celui-ci ingère par accident l’hôte intermédiaire du parasite, soit la puce du chien ou du chat.
Sérologies parasitaires.203 recherches d’anticorps contre divers parasites ont été réalisées pour 125 patients
(tab. 2). Chez 23 d’entre eux, une sérologie indicatrice d’une infection a été constatée. Les tests de dépistage, réalisés simultanément sur 7 antigènes d’helminthes, ont été demandés à 13 reprises. Les données cliniques du patient n’étant pas toujours signalées au laboratoire, l’évaluation de ces résultats en fonction du diagnostic est incomplète. Néanmoins, 2 cas d’échinococcose ont ainsi pu être découverts dans le canton en 1997.
y. MoosMann et al.
Tableau 1. Examens directs en 1997. Protozoaires sanguins, protozoaires intestinaux et helminthes, classés selon leur provenance. 377 parasites découverts chez 269 patients.
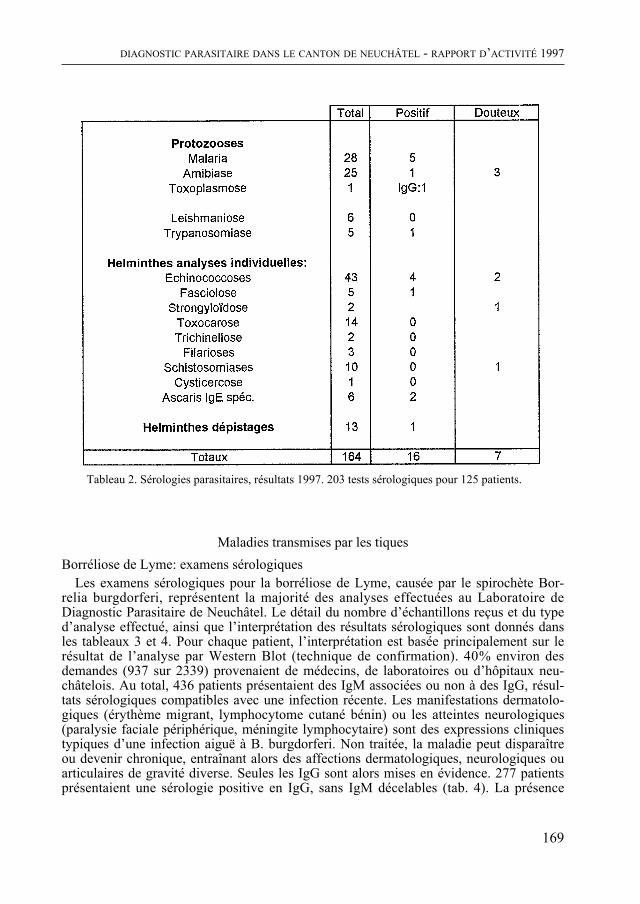
169
maladies transmises par les tiquesBorréliose de Lyme: examens sérologiques
Les examens sérologiques pour la borréliose de Lyme, causée par le spirochète Bor-relia burgdorferi, représentent la majorité des analyses effectuées au Laboratoire de Diagnostic Parasitaire de Neuchâtel. Le détail du nombre d’échantillons reçus et du type d’analyse effectué, ainsi que l’interprétation des résultats sérologiques sont donnés dans les tableaux 3 et 4. Pour chaque patient, l’interprétation est basée principalement sur le résultat de l’analyse par Western Blot (technique de confirmation). 40% environ des demandes (937 sur 2339) provenaient de médecins, de laboratoires ou d’hôpitaux neu-châtelois. Au total, 436 patients présentaient des Igm associées ou non à des IgG, résul-tats sérologiques compatibles avec une infection récente. Les manifestations dermatolo-giques (érythème migrant, lymphocytome cutané bénin) ou les atteintes neurologiques (paralysie faciale périphérique, méningite lymphocytaire) sont des expressions cliniques typiques d’une infection aiguë à B. burgdorferi. Non traitée, la maladie peut disparaître ou devenir chronique, entraînant alors des affections dermatologiques, neurologiques ou articulaires de gravité diverse. Seules les IgG sont alors mises en évidence. 277 patients présentaient une sérologie positive en IgG, sans Igm décelables (tab. 4). La présence
DiaGnostic parasitaire Dans le canton De neucHâtel - rapport D’activité 1997
Tableau 2. Sérologies parasitaires, résultats 1997. 203 tests sérologiques pour 125 patients.

170
d’IgG seules est parfois délicate à interpréter: simple reflet d’un contact ancien asympto-matique et sans relation avec la symptomatologie actuelle du patient, elle n’est pas forcé-ment le signe d’une borréliose de Lyme active ou passée. Ainsi, tout résultat sérologique doit être confronté systématiquement avec une clinique évocatrice.
Pour les 167 personnes testées au niveau du LCR (tab. 3), seules 26 (15%) possédaient des anticorps anti-B.burgdorferi (IgG et/ou Igm). Au moyen d’un test de ELISA-capture permettant la détermination d’un indice, nous avons comparé la sérologie dans le LCR à celle du sérum de ces patients. A l’aide de cet indice, une production intrathécale d’anti-corps spécifiques, caractéristique de la neuroborréliose, a été confirmée chez 6 patients (2 en IgG, 4 en IgG et Igm).
Borréliose de Lyme: examens par PCRLa mise en évidence directe de B. burgdorferi par culture dans des prélèvements de
peau, de liquide céphalo-rachidien (LCR) ou de ponction est possible mais difficile. La sensibilité de cette technique est toutefois médiocre, la bactérie étant présente en très faible quantité dans les tissus, particulièrement au stade chronique de la maladie. La technique de PCR permet l’amplification spécifique de matériel génétique cible (ADN bactérien par exemple). Cette nouvelle analyse, plus sensible, permet de détecter la pré-sence de 5 à 10 bactéries par millilitre d’échantillon. Des indications décisives pour le diagnostic sont ainsi apportées lors de suspicion d’arthrite de Lyme (positivité du liquide articulaire) ou de neuroborréliose (positivité du LCR et/ou de l’urine). L’élimination de borrélies dans les urines lors d’un traitement antibiotique peut également être suivie par PCR. 190 échantillons pour 152 patients ont été analysés cette année (tab. 3). Chez 15 patients, de l’ADN de B.burgdorferi a été amplifié dans du liquide articulaire, du LCR ou dans une biopsie cutanée.
Encéphalite à tique et rickettsioses: examens sérologiquesLes anticorps contre le virus de l’encéphalite à tique ont été dosés chez 59 personnes
(tab. 4). Douze montraient des taux faibles à élevés d’IgG, signe d’une infection ancienne ou résultant d’une vaccination. Chez une personne, la présence concomitante d’Igm signalait une infection ou une vaccination récente. Précisons toutefois que le canton de Neuchâtel, et plus généralement la Suisse romande, reste en bordure de la zone d’en-démie pour cette virose, et qu’aucun patient à examen sérologique positif n’a été infecté dans notre région.
Finalement, 45 sérologies pour la fièvre q ou pour d’autres rickettsioses (typhus ou fièvres boutonneuses) ont été demandées (tab. 4). Toutes se sont révélées négatives.
conclusions
En 1997, 3952 personnes ont subi une recherche par sérologie ou par PCR de maladie transmise par les tiques, un examen parasitologique ou une sérologie parasitaire. Le volume des analyses parasitologiques et d’agents transmis par les tiques est resté stable par rapport à l’année 1996 (moosmann et al., 1997). Par examen direct, des parasites ont été détectés chez 269 patients en 1997. Par sérologie, 23 personnes présentaient des anticorps spécifiques d’une parasitose, et 725 personnes des anticorps contre une maladie transmise par les tiques (maladie active ou cicatrice sérologique).
y. MoosMann et al.
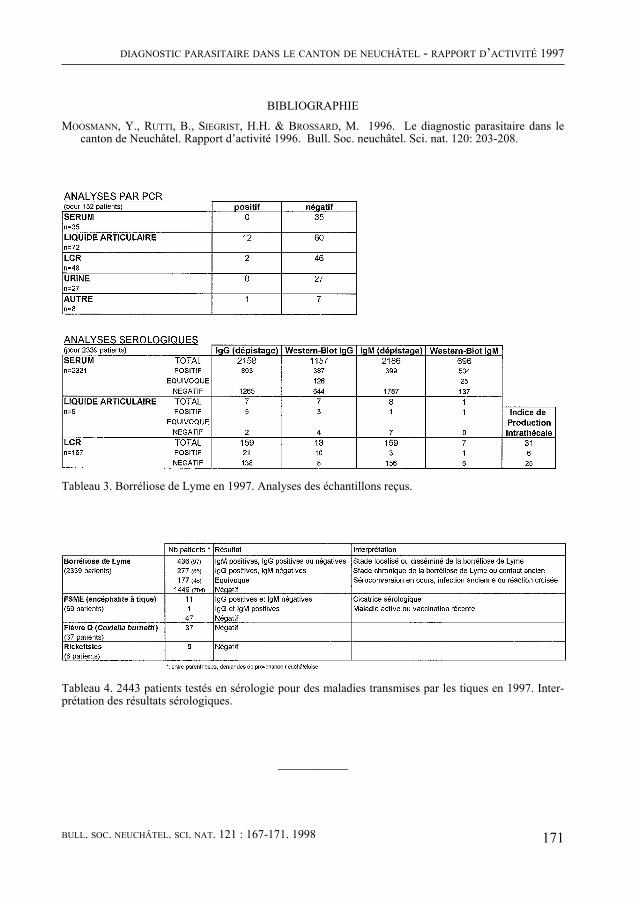
171
biblioGrapHie
MoosMann, y., rutti, b., sieGrist, H.H. & brossarD, M. 1996. Le diagnostic parasitaire dans le canton de Neuchâtel. Rapport d’activité 1996. Bull. Soc. neuchâtel. Sci. nat. 120: 203-208.
DiaGnostic parasitaire Dans le canton De neucHâtel - rapport D’activité 1997
bull. soc. neucHâtel. sci. nat. 121 : 167-171. 1998
Tableau 3. Borréliose de Lyme en 1997. Analyses des échantillons reçus.
Tableau 4. 2443 patients testés en sérologie pour des maladies transmises par les tiques en 1997. Inter-prétation des résultats sérologiques.
__________


bulletin De la société neucHâteloise Des sciences naturelles 121 : 173-180. 1998
173
OBSERvATIONS mÉTÉOROLOGIqUES FAITES EN 1997 àL’OBSERvATOIRE CANTONAL DE NEUCHTEL
GILBERT JORNOD
L’année 1997 a été dans son ensemble chaude, bien ensoleillée et pratiquement nor-male en précipitations. Au chapitre de ses particularités on se souviendra que janvier n’a reçu que 2 h de soleil (-95%), que juin et juillet ont été très pluvieux mais que par contre mars est déficitaire de 78 % en eau et qu’avril n’avait jamais été aussi bien enso-leillé depuis 1901 (+93 h, +54%). D’autre part on notera également que depuis 1981 la moyenne annuelle de la température de l’air a systématiquement dépassé ou égalé la norme, la moyenne la plus élevée étant de 11.3° en 1994.
teMpérature
La température moyenne de l’air est de 10.3° (1996: 9.3°) pour une valeur normale de 9.2° ; les moyennes saisonnières sont les suivantes: hiver: 1.7° (normale: 0.8°), prin-temps: 10.4° (9.0°), été: 18.1° (17.7°) et automne: 10.4° (9.5°) ; les moyennes mensuelles sont comprises entre –0.6° en janvier et 20.3° en août et les moyennes journalières entre 23.7° le 24 août et –6.2° le 21 janvier. Seuls janvier et juillet sont thermiquement défici-taires, avec un écart négatif maximal de -1.2° pour juillet tandis que février et mars ont été très chauds avec respectivement 3.5° et 3.3° de plus que leur valeur normale. Les lec-tures extrêmes du thermomètre, 30.8° le 11 août et –8.0° le 1er janvier, déterminent l’am-plitude annuelle absolue de la température: 38.8° (normale: 42.9°). L’année compte 42 jours d’été, 34 de gel et 15 d’hiver ; le dernier gel de printemps s’est produit le 22 avril et le premier gel de fin d’année date du 28 octobre.
insolation
L’insolation générale est excellente: 1812 heures (1996: 1622 h), soit 139 h (8 %) de plus que la valeur normale de ce critère ; les insolations saisonnières sont les suivantes: hiver: 138 h (normale: 147 h), printemps: 686 h (502 h), été: 580 h (704 h) et automne: 402h (304 h) et les insolations mensuelles s’échelonnent entre 2 h en janvier et 266 h en avril ; les écarts relatifs extrêmes appartiennent également à ces 2 mois, soit respective-ment –95 % (-36 h) et +54% (+93 h). Janvier, juin et juillet sont les seuls mois défici-taires de l’année. L’ensoleillement journalier maximal est de 14.3 h le 31 mai, 66 jours n’ont pas été ensoleillés et 27 autres l’ont été par moins de 1 h. L’année compte 147 jours très nuageux ou couverts et 64 jours clairs.
précipitations

174
Les précipitations reçues qui sont de 961 mm (1996: 887 mm) sont pratiquement normales (-15 mm, -2%) ; les valeurs saisonnières sont les suivantes: hiver: 181 mm (normale: 229 mm), printemps: 126 mm (211 mm), été: 389 mm (290 mm) et automne: 220 mm (246 mm). Juin, juillet et septembre sont les mois excédentaires, l’écart maximal étant de +58 mm (+60%), en juin, mars possédant l’écart négatif le plus important: -53 mm (-78%). Les précipitations journalières maximales au cours de l’année sont de 55 mm le 2 septembre au cours d’un orage, ces derniers étant au nombre de 19 en 1997. Il a plu au cours de 145 jours, neigé à 9 reprises, le sol a été recouvert par la neige pendant 23 jours, la couche maximale atteignant 10 cm le 2 janvier ainsi que les 3 et 4 décembre. La neige est tombée pour la dernière fois le 20 avril au printemps, pour faire sa réapparition le 2 décembre en fin d’année.
pression atMospHérique
La moyenne de la pression atmosphérique est de 960.8 hPa (normale: 959.6 hPa) ; les moyennes mensuelles sont comprises entre 967.0 hPa en mars et 953.6 hPa en novembre et les moyennes journalières entre 976.9 hPa le 27 février et 937.0 hPa le 6 novembre. L’amplitude absolue de la pression atmosphérique, 45.3 hPa (normale: 46.9 hPa), est donnée par les extrêmes de 979.6 hPa le 27 février et 934.3 hPa le 6 novembre.
HuMiDité De l’air
La moyenne de l’humidité relative de l’air est normale: 76 % ; les moyennes men-suelles vont de 91% en janvier à 59 % en avril et les moyennes journalières de 97% le 24 janvier à 43 % les 7, 15 et 23 avril. La lecture minimale de l’hygromètre date du 1er avril, 26%, et le brouillard au sol a sévi au cours de 21 jours, soit 6 en janvier, 1 en mars, 2 en mai, 4 en septembre, 1 en octobre et 7 en novembre.
vent
Les vents ont parcouru 64320 km à la vitesse moyenne normale de 2.0 m/seconde ; leur répartition selon les huit directions principales est assez classique: nord-est, est et ouest, 20% chacune du parcours total, suivies par sud-ouest, 17%, sud, 7%, nord, 6 %, nord-ouest, 6 % et sud-est, 4%. Le parcours mensuel maximal appartient à avril avec 8350 km (3.2 m/seconde), janvier avec 3070 km (1.1 m/seconde) étant le mois le plus calme ; les extrêmes journaliers sont compris entre 620 km le 13 février (26 km/h de vitesse moyenne) et 26 km le 5 novembre. La vitesse de pointe maximale atteinte par le vent à Neuchâtel-Observatoire en 1997 est de 105 km/h de l’ouest le 13 février, suivie par 80 km/h le 18 du même mois et le 31 mai, et 75 km/h les 12 octobre et 15 décembre ; les vents n’ont pas dépassé 65 km/h au cours des autres mois.
Note: Les données journalières peuvent être obtenues à l’Observatoire cantonal de Neuchâtel, rue de l’Observatoire 58, CH 2000 Neuchâtel
G. JornoD
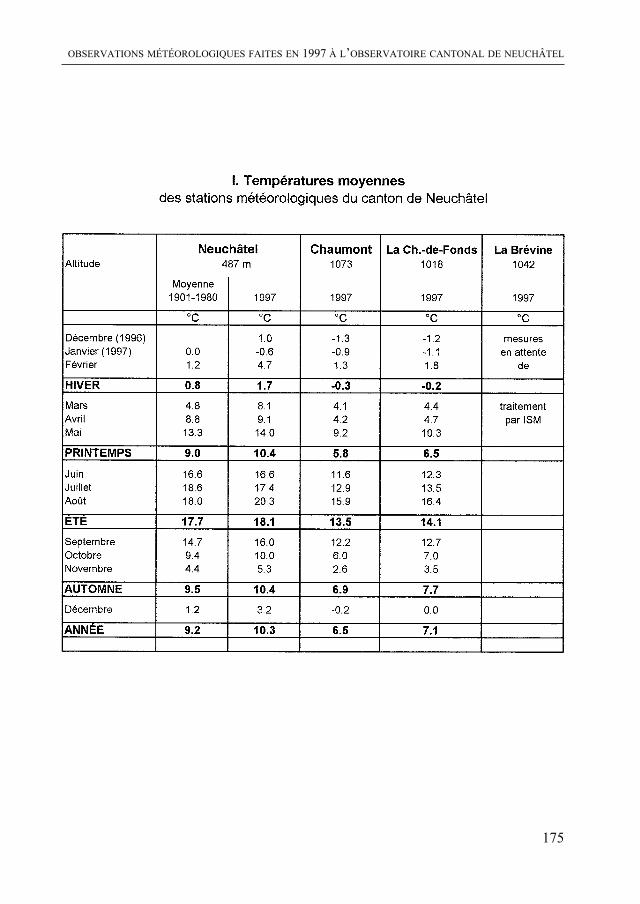
175
observations MétéoroloGiques faites en 1997 à l’observatoire cantonal De neucHâtel
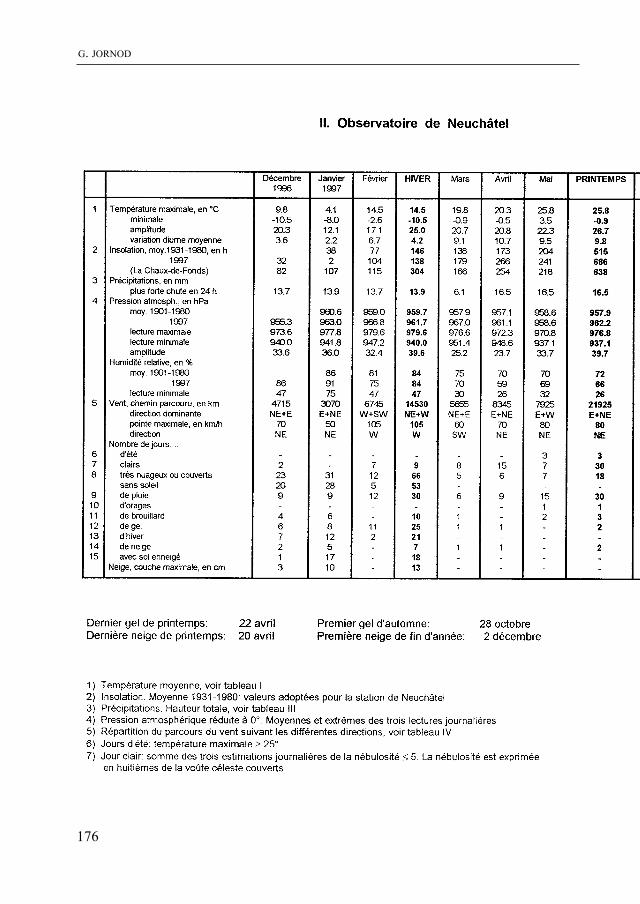
176
G. JornoD
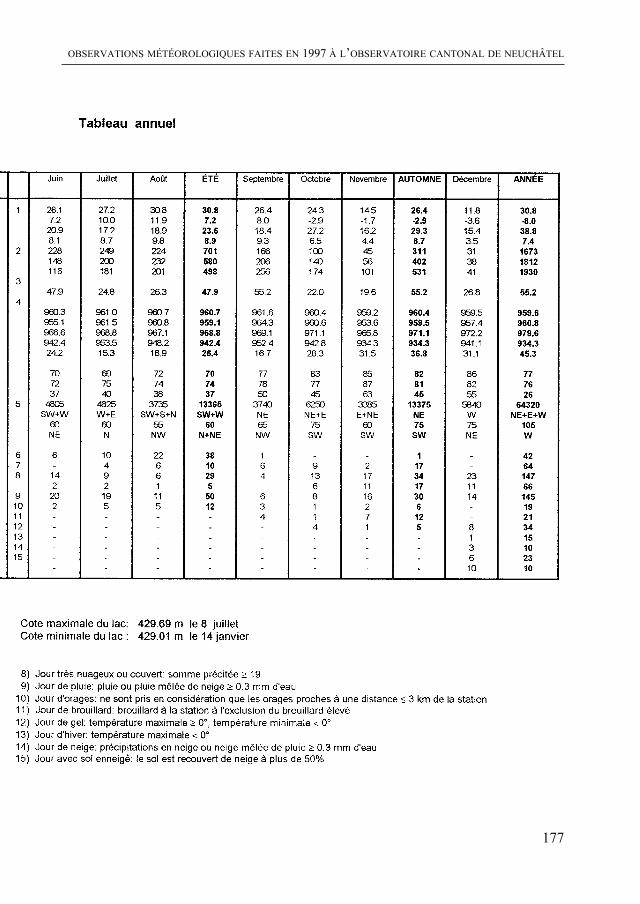
177
observations MétéoroloGiques faites en 1997 à l’observatoire cantonal De neucHâtel
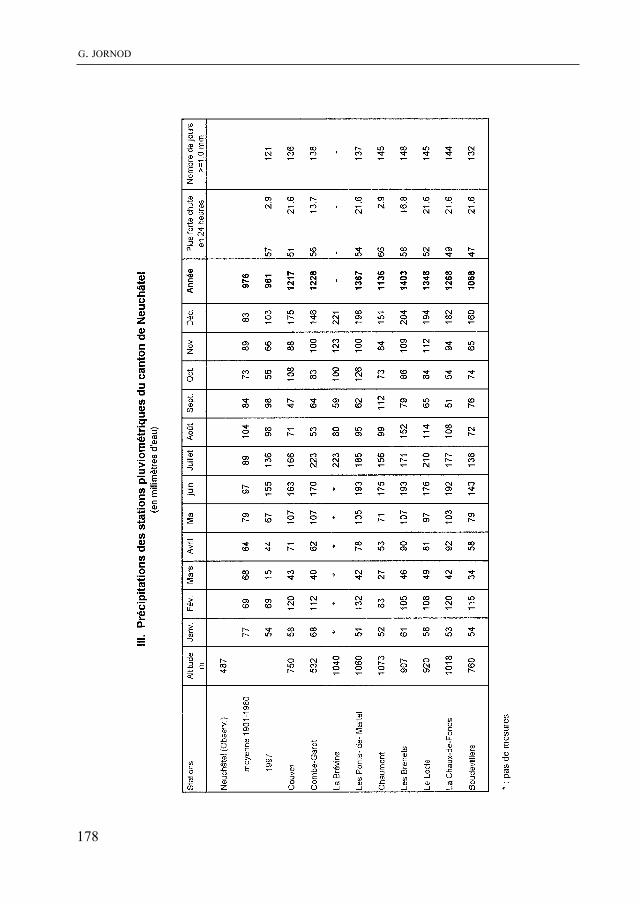
178
G. JornoD
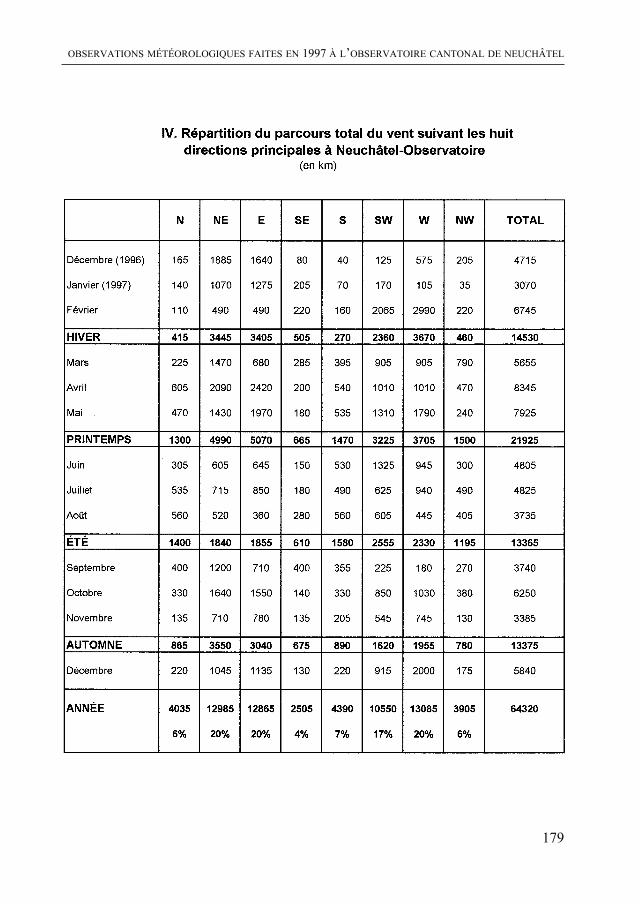
179
observations MétéoroloGiques faites en 1997 à l’observatoire cantonal De neucHâtel
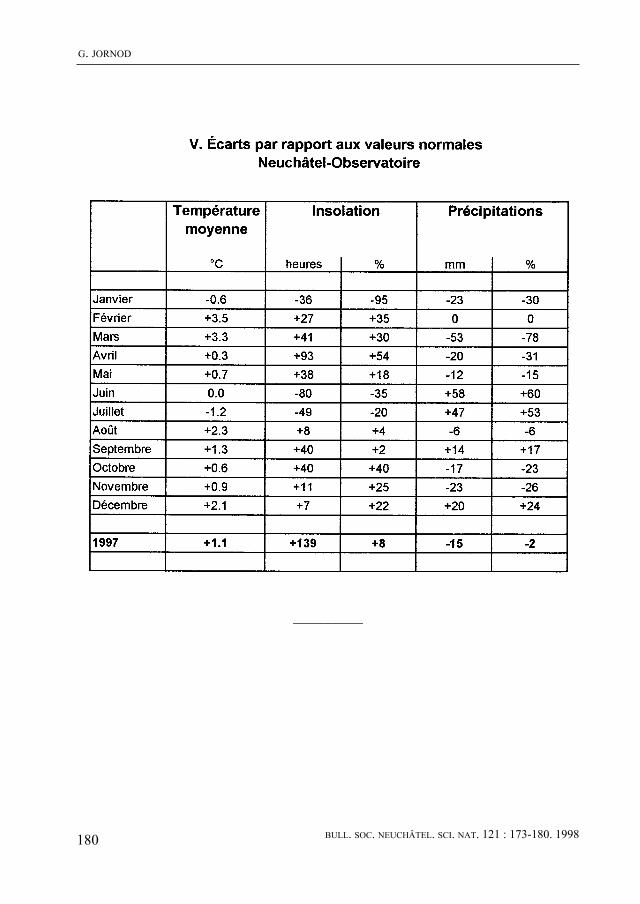
180
G. JornoD
__________
bull. soc. neucHâtel. sci. nat. 121 : 173-180. 1998

bulletin De la société neucHâteloise Des sciences naturelles 121 : 181-184. 1998
181
177E ASSEmBLÉE ANNUELLE DE L’ACADÉmIE SUISSE DES SCIENCES NATURELLES
à LA CHAUX-DE-FONDS8 - 12 octobre 1997
En date du 6 mai 1995, le Sénat de l’ASSN nommait le soussigné à la présidence annuelle 1997 et lui confiait ainsi, par le biais de la Société neuchâteloise des sciences naturelles, le soin de préparer le grand rassemblement annuel des scientifiques suisses à La Chaux-de-Fonds.
Le Comité annuel était bientôt constitué grâce à la collaboration de monsieur le professeur Willy matthey (vice-président), de madame Lise Gern (présidente SNSN jusqu’au printemps 97), de mm. Jean-Daniel Blant, yves Ehret (finances), Claude-Eric Hippenmeyer, marc Schlüssel (hébergement), François Straub (locaux de réunion), Roland Stettler (président SNSN dès le printemps 97) et Hugues Wülser. Pour faciliter la tâche du président, la Commune de La Chaux-de-Fonds lui accordait un congé d’un quart de poste durant une année. Au cours de deux réunions préparatoires, les différentes tâches étaient attribuées aux membres, alors qu’un gros travail préparatoire avait déjà été accompli par le vice-président et le président lors d’une dizaine de réunions.
C’est le thème général “Paysage calcaire de l’Arc jurassien: du minéral au vivant” qui avait été choisi pour l’assemblée. vingt-quatre sociétés spécialisées et commissions y ont participé, dont certaines siégaient déjà dès le matin de la journée inaugurale mercredi 8 octobre dans les pâturages du Jura neuchâtelois.
En fin d’après-midi, monsieur Jean-martin monsch, conseiller communal, directeur des affaires culturelles, accueillait les participants dans l’aula du Gymnase cantonal, en rap-pelant quelques grands traits de l’histoire de la ville et de sa vocation scientifique. Il était suivi par monsieur le professeur Bernard Hauck, président central de l’Académie, pour le discours d’ouverture officiel de la 177e assemblée annuelle. Il y fit part des préoccupations de l’Académie en cette fin de 20e siècle: maintien des ponts entre disciplines, dangers de l’hyperspécialisation, communications à l’intérieur même de la société et ouverture vers le public profane, rôle de l’ASSN en tant que réseau d’experts et enfin, pour l’immédiat, l’initiative pour la protection génétique, dont l’acceptation serait lourde de conséquences pour les projets de recherche des hautes écoles suisses. Le président achevait son propos en insistant sur la nécessité d’une attitude critique et responsable des scientifiques vis-à-vis de leur recherche, condition nécessaire pour avoir la confiance de nos concitoyens.
Après avoir remercié les autorités communales, cantonales et académiques pour leur appui à l’organisation de l’assemblée, le président annuel faisait ensuite une incursion dans l’histoire des sciences régionales, en rappelant l’apport de trois naturalistes des 18e, 19e et 20e siècles, Abraham Gagnebin, Célestin Nicolet et Albert monard, au développe-ment du mouvement scientifique des montagnes neuchâteloises. Si Abraham Gagnebin (1707-1800) doit en être considéré comme l’initiateur, ses apports à la botanique, à la paléontologie, à la météorologie, à l’utilisation de la nomenclature linnéenne ont été quelque peu oubliés. En contact avec les plus grandes célébrités de son temps, en France, en Suisse, en Italie, Abraham Gagnebin fut, avec son frère Daniel, le créateur du premier cabinet de curiosités privé connu loin à la ronde.

M. s. Jacqat
182
Célestin Nicolet (1803-1871), pharmacien, géologue, botaniste, a marqué le 19e régional par ses idées originales et son immense activité au service de la science et de l’éducation (fondateur de la bibliothèque de la ville, du musée polyvalent dont est issu le musée d’histoire naturelle, de la section des montagnes de la Société neuchâteloise des sciences naturelles, etc.). Collaborateur d’Agassiz, dont il fut le botaniste lors de l’ex-ploration du glacier de l’Aar, Nicolet fut aussi l’auteur de découvertes sensationnelles en matière de faune tertiaire à La Chaux-de-Fonds et présida la 38e session annuelle de la Société helvétique des sciences naturelles à La Chaux-de-Fonds en 1855.
Scientifique marquant des montagnes au 20e siècle, Albert monard (1886-1952), auteur du fameux Petit botaniste romand, fut un pionnier de l’hydrobiologie et un spécialiste renommé des harpacticides en même temps qu’un vulgarisateur de talent. Conservateur du musée d’histoire naturelle de La Chaux-de-Fonds dès 1920, il entreprit, entre 1928 et 1947, quatre expéditions en Angola, Guinée Bissau et Cameroun qui contribuèrent de manière remarquable à l’enrichissement des collections de l’institution et à la connais-sance des faunes africaines. Auteur de quelque 65 publications, il a laissé son nom à une foule de taxons et aux Principes de monard, bien connus en biocénotique, sans compter le fait qu’il fut à l’origine de très nombreuses vocations de naturalistes.
Le président annuel achevait son intervention en relevant la large ouverture d’esprit, la connaissance transversale et multidisciplinaire de nos prédécesseurs, dont la passion, l’en-têtement, la grande capacité de travail nous laissent pantois et nous enseignent l’humilité.
La conférence inaugurale de monsieur le professeur André Giordan, directeur du Laboratoire de didactique et d’épistémologie des sciences de l’Université de Genève por-tait sur le thème “Le généraliste aura-t-il encore sa place dans la recherche du troisième millénaire ? quel enseignement pour quelle recherche au-delà de l’an 2000 ?” .
Pour répondre à ces questions, le conférencier, biologiste de formation, sut, en alter-nant anecdotes, éléments provocateurs et constats personnels, mettre en évidence les problèmes de communication que peuvent rencontrer les spécialistes de deux laboratoires contigus. Selon lui, la recherche est le faite de deux groupes: les spécialistes ou micro-spécialistes, les généralistes ou naturalistes. Ces derniers deviennent de plus en plus rares. Cette situation est malsaine, car le développement de la recherche scientifique influence de plus en plus le quotidien de notre société. On peut se demander si les spécialistes sont encore capables de répondre aux questions générales. Giordan voit trois causes essen-tielles à cette situation: 1. les conditions de la reconnaissance d’un chercheur tiennent souvent au nombre de publications qu’il a à son actif, nombre forcément supérieur pour un spécialiste membre d’un groupe de recherche comparé à un naturaliste de terrain qui doit commencer par récolter des données.2. les programmes de recherche sont définis par des fonctionnaires fédéraux ou inter-nationaux qui évaluent la qualité d’un projet au nombre de publications, voire à la renommée de la revue dans laquelle elles paraissent. Les revues des généralistes ont à cet égard moins bonne presse. Les bailleurs de fonds ont aussi tendance à mieux consi-dérer un projet lorsqu’il s’accompagne de grosses dépenses d’équipement, alors que la recherche du généraliste nécessite peu de matériel, mais un engagement personnel très important, de longue durée. 3. les étudiants sont aujourd’hui confrontés à un flux d’informations, dont il est exigé la régurgitation sans contrôle de la compréhension. Les méthodes de travail, les analyses sys-tématiques, les modèles et la communication font défaut, ce qui développe une conception linéaire des problèmes, éloignée des réalités et des problèmes globaux du monde actuel.

177e asseMblée annuelle De l’assn à la cHaux-De-fonDs
183
En résumé, André Giordan voit une place pour les généralistes dans la recherche de l’avenir. Il pense que ce sont eux qui seront capables d’intégrer et de coordonner le savoir des spécialistes. Des lacunes dans ce domaine ne peuvent que contrecarrer le développe-ment future de la recherche. La question qui reste posée est celle de savoir comment on peut préparer un terrain favorable à ces nouveaux généralistes !
La remise du Prix Schläfli par le Président central à m. Andrew Hemphill, précédée de sa laudatio par monsieur le professeur Urs Rüegg, puis l’inauguration de l’exposition de la Fondation “La science appelle les jeunes” et un vin d’honneur offert par la ville de La Chaux-de-Fonds achevaient cette cérémonie d’ouverture enrichie des remarquables pres-tations musicales de mm. Olivier Theurillat, trompette et John-Edwin Graf, piano.
quarante-huit invités, dont le Professeur Jean-Paul Poirier, représentant de l’Académie des sciences de Paris et le Professeur Lothar Eissmann, délégué par l’Académie des sciences de Saxe, participaient au repas des invités du mercredi soir excellemment servi au Club 44. La soirée fut marquée notamment par la présence des présidents des quatre Académies de Suisse. L’ancien président de la Confédération Pierre Aubert, le conseiller national Rémy Scheurer, madame Josiane Nicolet, présidente de la ville du Locle, mon-sieur Jean-martin monsch, conseiller communal, directeur des affaires culturelles nous faisaient aussi l’honneur de leur compagnie.
Deux événements importants étaient organisés par le Comité central jeudi 9 octobre. Un lunch-débat au sujet de la place de la recherche dans l’Expo 2001, devenue Expo 01, pre-nait place aux heures du midi. madame Jacqueline Fendt, directrice, traita le sujet avec brio et passion. “Il faut créer un monde extraordinaire” sur des bases écologiquement correctes. Les projets les plus audacieux sont permis et c’est l’occasion de lancer des produits nova-teurs. Selon madame Fendt, l’exposition représentera pour les chercheurs jeunes d’esprit, qui y amèneront “leurs cerveaux et leurs cœurs, la possibilité de participer à l’élaboration de la politique de la science en Suisse... Ce sera aussi l’occasion de faire sortir la science de sa tour d’ivoire, partant du principe que haute qualité n’est pas incompatible avec popularité”.
Dès 17 h suivait un forum consacré au Génie génétique. Une présentation générale par monsieur le professeur Jean-marc Neuhaus, directeur du laboratoire de biochimie de l’Université de Neuchâtel, permettait de rappeler que le génie génétique est un formi-dable champ de recherches dont on ne vit que la préhistoire; c’est un pas vers l’avenir, mais un pas que le grand public ne franchit pas volontiers. Il appartenait à monsieur Gil Baillod, directeur des rédactions de l’Impartial et de l’Express de jouer le rôle de l’avocat du diable au milieu d’un collège d’experts comprenant madame le professeur Heidi Dig-gelmann, présidente du Fonds national suisse de la recherche scientifique, ancienne direc-trice de l’Institut suisse de recherche expérimentale sur le cancer (ISREC) à Epalinges, monsieur le professeur Klaus Amann, directeur du Jardin botanique de l’Université de Berne, monsieur le professeur michel Aguet, directeur actuel de l’ISREC et le Dr Walter Lesch, maître assistant à l’Institut interdisciplinaire d’éthique et des droits de l’Homme de l’Université de Fribourg. Une communication qui fonctionne mal entretient d’autant plus la crainte. Si la majorité de la population ne semble pas hostile à la recherche des scientifiques, elle ne croit plus à une science toujours au service de la bonne cause, car les convictions individuelles, parfois irrationnelles, interfèrent fréquemment avec elle.
Un apéritif clôturait cette riche journée organisée au Club 44, ce qui leur assurait une plus
grande audience parmi le grand public habitué aux conférences données dans ce cadre.

M. s. Jacqat
184
Le symposium principal occupait l’après-midi du vendredi 10 octobre. Durant près de 3 heures et devant un public extrêmement attentif, les trois conférenciers surent tenir en haleine leurs auditeurs sur le thème général de la 177e assemblée annuelle de l’ASSN. Alors que le Professeur michel monbaron traitait de “Jurassic Adventure” ou géo-histoire de la perception du relief calcaire jurassien par les hommes, le Professeur Jean-michel Gobat présentait “Sols et végétation du Jura: entre méditerranée et toundra scandinave”. Il appartenait au Professeur Jean-michel Le Tensorer de présenter “Les conquêtes d’Homo jurassicus”, parachevant ainsi ce panorama géologique, botanique et faunistique de l’Arc jurassien qui fait l’objet d’une publication spécifique.
Le Cabaret Louis Pasteur, dû à Daniel Raichvarg, de Paris, ravissait un public de connaisseurs le vendredi soir au Club 44. Dernière acte officiel de l’assemblée de La Chaux-de-Fonds, cette soirée où le professeur parisien d’histoire des sciences et de didactique devenait homme de théâtre permit de présenter de manière très vivante la vie et l’oeuvre de Louis Pasteur, au travers d’une foule d’objets et de diapositives accompa-gnant une mise en scène dynamique et très attractive.
Plusieurs excursions organisées par les sociétés spécialisées et le Comité annuel avaient lieu le samedi matin 11 octobre. Alors que l’on pouvait craindre le pire de Dame météo, le ciel s’éclaircissait bientôt et permettait à Roland Stettler et à son groupe d’aller à la découverte de quelques paysages typiques du Jura neuchâtelois dans la vallée de la Sagne et de la Brévine, au lac des Taillères, à Saint-Sulpice et à môtiers, alors que les mathématiciens et les ornithologues étaient encore en session scientifique. Ce n’est que le dimanche après-midi que les derniers naturalistes quittaient la région après une excursion au bord d’un Doubs brumeux, moins accueillant qu’à l’accoutumée.
Ainsi s’achevait la 177e assemblée de l’ASSN, tenue essentiellement dans les locaux du Gymnase cantonal. quelque cent-soixante symposiums, conférences et présentations, nombre auquel il faut ajouter la vingtaine d’interventions des mathématiciens et la cin-quantaine des physiciens faisant l’objet de programmes séparés ont permis à plus de 700 participants d’assister à tout ou partie de notre assemblée, la très large majorité d’entre eux s’étant inscrits lors de leur arrivée.
Chacun a reçu une petite serviette marquée du logo de l’assemblée, ainsi qu’une docu-mentation touristique régionale et une plaquette éditée spécialement à cette occasion par les Editions de la Girafe du musée d’histoire naturelle de La Chaux-de-Fonds. Il s’agis-sait du portrait de 41 scientifiques provenant des hauteurs de l’Arc jurassien.
Notons encore que les comptes ont bouclé avec un montant de quelque CHF 35’000.- inférieur au budget prévu et qu’une publication des textes des intervenants du symposium principal restera le souvenir tangible de cette troisième réunion (après 1855 et 1931) des scientifiques suisses à La Chaux-de-Fonds.
marcel S. JacquatConservateur du musée d’histoire naturelle de La Chaux-de-Fonds
Président annuel ASSN 1997
bull. soc. neucHâtel. sci. nat. 121 : 181-184. 1998

185
PROCÈS-vERBAUX DES SÉANCESANNÉE 1997
22 janvier 1997Auditoire du muséum d’histoire naturelle de Neuchâtel
BULGARIE, CARREFOUR DE LA DIvERSITÉ BIOLOGIqUEDr. Pierre Galland (Pro Natura, Bâle)
La Bulgarie, de par sa position centrale entre l’Europe centrale, l’Asie mineure et le bassin méditerranéen, héberge des richesses naturelles remarquables. Bien qu’encore intactes, elles sont menacées par les changements que traverse le pays. Dans le cadre général du suivi de Rio, la Confédération a attribué un crédit important à la conser-vation de la biodiversité dans la région balkanique. Un programme bulgaro-suisse a débuté en 1994 sous la responsabilité de la Ligue suisse pour la Protection de la Nature (aujourd’hui Pro Natura); il comprend huit projets de conservation de régions de mon-tagnes et de milieux humides le long de la côte de la mer Noire. Ours, loup, putois marbré, vautour moine, cormoran pygmée, tortue sont quelques-unes des espèces dont la protection est intégrée à un développement durable.
5 février 1997Auditoire du muséum d’histoire naturelle de Neuchâtel
LA DÉCOUvERTE DES PREmIÈRES PLANÈTES EXTRA-SOLAIRES, UN PREmIER PAS vERS LA DÉCOUvERTE D’AUTRES mONDES.
Dr. Didier queloz (Observatoire de Genève)
Depuis l’annonce, en octobre 1995, de la découverte par michel mayor et Didier queloz, deux astronomes suisses, de la première planète en orbite autour d’un astre sem-blable au soleil, plusieurs compagnons planétaires autour d’une étoile ont été détectés.
L’existence de certaines de ces planètes pose de nombreuses questions aux astrophysi-ciens, car aucun scénario de formation planétaire n’avait prévu leur existence.
Le conférencier a présenté les résultats récents et a montré les problèmes qu’ils sou-lèvent au vu des scénarios classiques de formation des planètes. Les méthodes actuelles de détection ont également été présentées, ainsi que les moyens d’investigation du futur. Enfin, la recherche de la vie sur d’autres mondes a été discutée en regard des découvertes actuelles et des expériences prévues pour le début du prochain millénaire.
19 février 1997

186
Auditoire du muséum d’histoire naturelle de NeuchâtelCARTOGRAPHIE DE LA vULNÉRABILITÉ DES BASSINS
D’ALImENTATION DES SOURCES KARSTIqUES PAR UNE mÉTHODE mULTICRITÈRE, LA mÉTHODE EPIK
Dr. Nathalie Doerfliger (Centre d’Hydrogéologie de l’Université de Neuchâtel)
Cette méthode permet d’établir des cartes de vulnérabilité des aquifères, des sources ou des captages en milieu karstique. Basée sur l’organisation spécifique des écoulements dans les aquifères, elle prend en compte quatre critères:
1.- le développement de l’épikarst,2.- l’importance de la couverture protectrice,3.- les conditions d’infiltration,4.- le développement du réseau karstique.Chacun de ces critères est subdivisé en classes, et ces dernières sont cartographiées sur
l’ensemble du bassin considéré. A l’aide d’un système d’information géographique, ces cartes sont ensuite pondérées et combinées dans le but d’obtenir une carte finale de vul-nérabilité.
Cette méthode a fait l’objet de tests sur plusieurs sites en Suisse, là où des problèmes de contamination des sources par l’agriculture se posent régulièrement. Ces tests ont abouti à l’établissement de nouvelles zones de protection. Un cas a été détaillé dans le cadre de l’exposé.
5 mars 1997Auditoire du muséum d’histoire naturelle de Neuchâtel
A) ASSEmBLÉE GÉNÉRALEB) NOTRE ATmOSPHÈRE, NOTRE AvENIR.
DIAGNOSTIC ET mESURESm. René Longet (Société suisse pour la protection de l’Environnement, Genève)
Le conférencier a traité, sous une forme très accessible et sous un angle très actuel, des points suivants:a) - Rappel des fonctions de l’atmosphère.b) - Le diagnostic, les moyens de savoir, le débat, une déontologie; - Les effets des modifications anthropogènes de l’atmosphère du point de vue du climat, de la santé, des relations nord-sud, de la politique de l’énergie.c) - Les mesures prises et leur efficacité; - Les mesures en discussion et à prendre, les perspectives.
5 novembre 1997Auditoire du muséum d’histoire naturelle de Neuchâtel

187
L’ORIGINE DE LA vIEDr. Danielle Decrouez (muséum d’Histoire naturelle de Genève)
Du fait que la nature n’est pas plus respectueuse que l’homme des vestiges et des témoins de son histoire, plus on s’avance dans le passé, plus il est difficile de découvrir des terrains géologiques intacts qui révèlent une époque précise. Toutefois, les connais-sances scientifiques actuelles attestent que la vie est apparue relativement tôt après la for-mation de la Terre. Durant pratiquement 4 milliards d’années, l’évolution fut semble-t-il très lente, et voici 540 millions d’années survint le “big bang” de l’évolution. Force est de constater que depuis cet événement, les mécanismes évolutifs paraissent s’être opposés à toute modification importante de l’organisation structurale des êtres vivants.
L’exposé a abordé les problèmes et les questions que posent la faune d’Ediacara, la faune tommotiennne à petites coquilles, la faune des schistes du Burgess et l’invention du squelette, vraisemblablement le changement le plus important que l’on puisse trouver dans les archives fossiles.
19 novembre 1997Auditoire du muséum d’histoire naturelle de NeuchâtelHISTOIRES DE GÈNES, HISTOIRE DE LANGUES
Professeur André Langaney (Laboratoire de génétique et de biométrie de l’Université de Genève et Laboratoire
d’Anthropologie biologique du musée de l’Homme à Paris).
Au cours des dernières années, les recherches de génétique moléculaire sur les popula-tions humaines ont montré que les six milliards d’humains actuels descendent d’une seule petite population de la préhistoire récente. C’est l’argument qui est développé dans l’ex-position “Tous parents, tous différents” pour affirmer les liens généalogiques étroits entre tous nos contemporains, malgré la très grande diversité génétique individuelle et les dif-férences d’aspect physique externe, souvent spectaculaires, entre populations lointaines.
Du coup, les théories minoritaires de certains linguistes sur l’origine unique de toutes les familles de langues ont reçu une confirmation indirecte inattendue, puisqu’il n’était pas imaginable que quelques dizaines de milliers de chasseurs préhistoriques aient parlé des dizaines de “protolangues” appartenant à des familles linguistiques indépendantes.
3 décembre 1997Auditoire du muséum d’histoire naturelle de Neuchâtel
APPLICATIONS DU GÉNIE GÉNÉTIqUE AU DOmAINE DES PLANTES

188
Professeur Jean-marc Neuhaus(Institut de Botanique, Laboratoire de biochimie, Université de Neuchâtel)
Le génie génétique est un ensemble de méthodes applicables à tous les domaines de la biologie. Chez les plantes, il permet de comprendre le fonctionnement de la cellule et de l’organisme à un niveau inimaginable jusqu’ici. L’exposé a présenté des exemples de travaux de recherche fondamentale, ainsi qu’un projet de biotechnologie de la vigne en cours dans le Laboratoire du Professeur Neuhaus.
Le Président: Roland StettlerRAPPORT SUR L’ACTIvITÉ DE LA SOCIÉTÉ
NEUCHÂTELOISE DES SCIENCES NATURELLES EN 1997

189
présenté à l’Assemblée générale de la SNSN du 18 février 1998
ComitéA fin 1997, la composition du comité est la suivante:• R. Stettler (président) qui a succédé dès le 14 juin à mme Lise Gern (présidente sortante)• F. Felber (vice-président)• y. Delamadeleine (trésorier)• m. Burkhard (secrétaire aux verbaux)• W. matthey (rédacteur du bulletin)• J. Ayer (rédacteur technique)• m. Aragno (président du comité de lecture)• P. Küpfer (délégué ASSN)• C. Dufour, B. de montmollin, P. Schürmann (assesseurs)
Le secrétariat est assuré pour 2 ans consécutifs par mme C. Bettinelli (Laboratoire de physiologie végétale), depuis septembre 1997.
Effectifs Au 31 décembre 1997, la SNSN comptait 444 membres, dont 333 actifs, 6 membres à vie, 1 membre d’honneur, 65 membres de plus de 40 ans de sociétariat et 40 membres collectifs (presse, bacheliers lauréats du prix SNSN, sociétés soeurs, divers). En 1997, nous avons enregistré 2 décès (le Dr. Olivier Clottu et m. max Steiner), 3 démissions, 5 nouveaux membres et aucune radiation.
Nouveaux membres: mm. François Calame, Edward mitchell, Blaise mülhauser, Jean-marc Neuhaus, Jean-Pierre Perrenoud.
manifestationsPremière série de conférences:22 janvier Dr. Pierre Galland (Ligue Suisse pour la protection de la nature): Bulgarie, carrefour de la diversité biologique.
5 février Dr. Didier queloz (Observatoire de Genève): la découverte des premières planètes extra-solaires, un premier pas vers la découverte d’autres mondes.
19 février Dr. Nathalie Doerfliger (Centre d’hydrogéologie de Neuchâtel). Cartographie de la vulnérabilité des bassins d’alimentation des sources karstiques par une méthode multicritère, la méthode EPIK.5 mars m. René Longet (Directeur de la Société suisse pour la protection de l’environnement, Genève): notre atmosphère, notre avenir - diagnostic et mesures. (L’assemblée générale a également eu lieu le 5 mars, présidée par mme Lise Gern).

190
14 juin Sortie annuelle: région de l’Orbe souterraine (grottes et musée du fer), Dent-de-vaulion et cascade de la Tine de Conflens.
manifestations spéciales (participation de la SNSN):• 12ème congrès international de spéléologie du 9 au 16 août 1997 à La Chaux-de-Fonds.
• 177ème assemblée annuelle de l’Académie suisse des sciences naturelles (ASSN) à La Chaux-de-Fonds, du 8 au 12 octobre 1997, regroupant environ 700 personnes de 22 sociétés différentes.
Cette manifestation organisée de main de maître par le Président du Comité annuel, marcel S. Jacquat, conservateur du musée d’histoire naturelle de La Chaux-de-Fonds avait pour thème: “Paysage calcaire de l’Arc Jurassien: du minéral au vivant”. Cer-tains membres de la SNSN ont apporté leur aide au bon déroulement de ces journées, notamment pour les excursions et pour la création de panneaux consacrés à l’histoire des sciences neuchâteloises (mm. y. Delamadeleine, W. matthey, J.-P. Schaer).
Un forum “Génie génétique” grand public a été organisé le jeudi 9 octobre au Club 44.
Deuxième série de conférences:5 novembre Dr Danielle Decrouez (muséum d’histoire naturelle de Genève): l’origine de la vie.
19 novembre Prof. André Langaney (Laboratoire de génétique de l’Université de Genève): histoire de gènes, histoires de langues.
3 décembre Prof. Jean-marc Neuhaus (Laboratoire de biochimie de l’Université de Neuchâtel): application du génie génétique au domaine des plantes.
Bulletins et mémoire (par Willy matthey, rédacteur, et Jacques Ayer, rédacteur technique)
L’année 1997 a été caractérisée par une intense activité de publication. De manière inha-bituelle, le Bulletin N° 120 a été édité en deux fascicules:
• Un premier tome de 232 pages comprenant un article d’information générale, 11 articles scientifiques, les Actes du deuxième congrès international sur les Tipulomorpha (Diptera) tenu à Aletsch en juillet 1996 et rassemblés par le Dr. C. Dufour, et enfin quatre rapports scientifiques (Nature neuchâteloise, Aéropalynologie, Immunologie, météorologie) et deux rapports sur la marche de la Société.
• Les 146 pages du second tome, rédigé par le Dr J-S. meia, contiennent les Actes du XXe Colloque francophone de mammologie, tenu à Neuchâtel en octobre 1996.

191
Les rédacteurs ont également travaillé avec mme Anna Sommaruga et l’Institut de Géo-logie de l’Université (Prof. m. Burkhard) à l’édition du 12ème mémoire de la Société neuchâteloise des Sciences naturelles. Rédigé en anglais, intitulé GEOLOGy OF THE CENTRAL JURA AND THE mOLASSE BASIN, ce mémoire de 176 pages est com-plété par une vingtaines de profils sismiques en dépliants hors texte. L’ouvrage est pourvu d’une préface du Prof. m. Burkhard et d’un large résumé en français, toutes les légendes étant de plus rédigées dans les deux langues, afin que les membres uniquement francophones de la Société y trouvent aussi leur intérêt.
Toutes ces publications ont pu paraître à temps pour être présentées lors de la 177ème Assemblée de l’Académie suisse des Sciences naturelles, tenue du 8 au 12 octobre 1997 à La Chaux-de-Fonds.
Les rédacteurs remercient le musée d’Histoire naturelle de Neuchâtel, par son directeur le Dr. C. Dufour, de les accueillir sur son logiciel quark X-Press, ainsi que m. Olivier Attinger qui s’occupe avec compétence de l’édition des publications SNSN. Puis leurs remerciements vont à l’ASSN d’abord, mais aussi aux autres donateurs, pour leur soutien financier qui a permis la sortie de ces différentes publications.
EchangesLe tome No 120 (fascicules 1 et 2) du Bulletin de la Société des Sciences naturelles a été échangé avec 412 périodiques et revues nationales et internationales, dont l’inventaire est disponible auprès de la Bibliothèque publique et universitaire de Neuchâtel.
Prix aux lycéensSections A et B: Philippe Robert Section C: Tatiana mantuanoSection D: Christelle GertschSection G: Janique Plancherel
Subventions et donsComme de coutume, nous remercions sincèrement l’Etat et la ville de Neuchâtel, ainsi que l’Académie Suisse des Sciences naturelles dont les subventions ont permis la publi-cation de notre double bulletin et du mémoire de madame Somaruga.
Le Président: R. Stettler

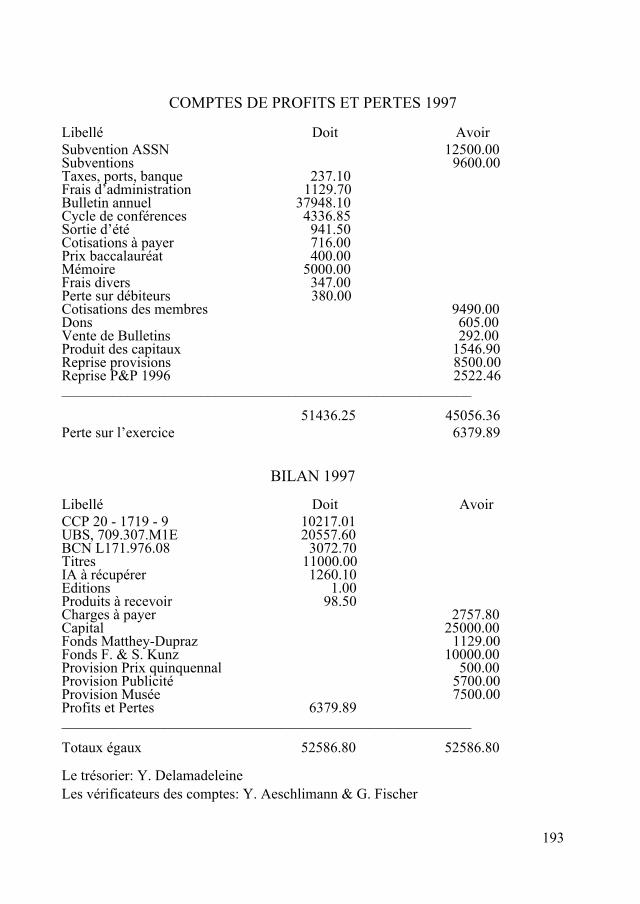
193
COmPTES DE PROFITS ET PERTES 1997
Libellé Doit AvoirSubvention ASSN 12500.00Subventions 9600.00Taxes, ports, banque 237.10 Frais d’administration 1129.70 Bulletin annuel 37948.10 Cycle de conférences 4336.85 Sortie d’été 941.50 Cotisations à payer 716.00 Prix baccalauréat 400.00 mémoire 5000.00 Frais divers 347.00 Perte sur débiteurs 380.00 Cotisations des membres 9490.00Dons 605.00vente de Bulletins 292.00Produit des capitaux 1546.90Reprise provisions 8500.00Reprise P&P 1996 2522.46 ________________________________________________________ 51436.25 45056.36Perte sur l’exercice 6379.89
BILAN 1997 Libellé Doit AvoirCCP 20 - 1719 - 9 10217.01 UBS, 709.307.m1E 20557.60 BCN L171.976.08 3072.70 Titres 11000.00 IA à récupérer 1260.10 Editions 1.00 Produits à recevoir 98.50 Charges à payer 2757.80Capital 25000.00Fonds matthey-Dupraz 1129.00Fonds F. & S. Kunz 10000.00Provision Prix quinquennal 500.00Provision Publicité 5700.00Provision musée 7500.00Profits et Pertes 6379.89 ________________________________________________________ Totaux égaux 52586.80 52586.80 Le trésorier: y. Delamadeleine Les vérificateurs des comptes: y. Aeschlimann & G. Fischer


195
T A B L E D E S m A T I È R E SDU TOmE 121 (1998)
. Pages
Willy matthey & Jacques Ayer - Avant-propos ......................................................................................... 3
Article d’information généraleJacques Ayer - A la recherche des dinosaures. Récit d’une campagne de fouilles paléontologiques
menée aux Etats-unis (Howe Ranch, Shell, Wyoming, 4 au 30 août 1997). ....................................... 5
Communications scientifiquesmarcel S. Jacquat - Une page régionale d’histoire des sciences relue récemment...
I. Célestin Nicolet et ses découvertes de faune miocène à La Chaux-de-Fonds. .............................. 37
Jan van der made - Une page régionale d’histoire des sciences relue récemment... II. The first described listriodont remains. ........................................................................................ 40
Philippe morel - Les os d’élans Alces alces (L. 1758) holocènes du gouffre 1 des Grands Bois(Jura neuchâtelois, Les verrières NE). Nouvelle datation radiocarbone et brève mise au point des connaissances actuelles. ..................................................................................................... 45
Kim-Lang Huynh - The genus Pandanus (Pandanaceae) in madagascar (Part 2). .................................. 53
Seyyed mahmood Ghaffari & S.B. Djavadi - Chromosome studies and distribution of nine species of Cousinia section Stenocephalae (Asteraceae) in Iran. ..................................................... 61
Olivier Reymond, François Straub & Hans Rudolf Preisig - Chrysonebula holmesii Lund(Algue: Chrysophyceae). Première mention pour la Suisse. ............................................................. 69
Gaëlle vadi & Jean-michel Gobat - Le paradoxe de la podzolisation en domaine jurassien.Aspects pédologiques et phytosociologiques. ................................................................................... 79
Philippe Cornali - Ecologie des pinèdes (Pinus sylvestris) de la rive sud du lac de Neuchâtel (Suisse).II. Phytomasse et productivité aériennes, dynamique du groupement. ............................................. 93
Willy matthey - Evolution des points d’eau dans la tourbière du Cachot. I. morphologie et végétation des gouilles. ...................................................................................... 111
Sylvie Barbalat - Faunistique de 9 Scarabaeidae phytophages et de 3 Lucanidae (Col., Scarabaeidae Pleurosticti, Lucanidae) capturés dans les Gorges de l’Areuse (Neuchâtel, Suisse). 127
Edouard Della Santa - Cardiocondyla emeryi Forel 1881 (Hymenoptera Formicidae) au Papiliorama de marin (Neuchâtel, Suisse). ..................................................................................... 137

196
Rapports scientifiques
Philippe Jacot-Descombes, Arthur Fiechter & Léonard Farron - Nature neuchâteloise 1997. .......... 141
Bertrand De montmollin, Gilles miserez & Catherine Strehler - Plan de gestion du Loclat (St-Blaise - NE). ............................................................................................................................. 150
Françoise vuillemin, may Udriet & Bernard Clot - Analyse aéropalynologique à Neuchâtel et à La Chaux-de Fonds en 1997. ....................................................................................................... 163
yves moosmann, Bernard Rutti, Hans H. Siegrist & michel Brossard - Diagnostic parasitaire et demaladies transmises par les tiques dans le canton de Neuchâtel. Rapport d’activité 1997. ........... 167
Gilbert Jornod - Observations météorologiques faites en 1997 à l’Observatoire cantonal de Neuchâtel. ................................................................................................................................. 173
vie de la Société
m. S. Jacquat - 177e Assemblée annuelle de l’Académie suisse des Sciences naturelles à La Chaux-de-Fonds (8-12 octobre 1997). ...................................................................................... 181
Procès-verbaux des séances et rapports sur l’activité de la S.N.S.N. en 1997. ................................... 185

197
instructions à l’intention Des auteurs
I. Remise des documents
Les manuscrits inédits, rédigés en français ou en anglais, doivent être remis jusqu’au 15 février pour paraître dans le Bulletin de l’année. Les textes sont présentés dans leur rédaction définitive, prêts à l’im-pression. Les auteurs utilisant un traitement de texte remettent, avec leur manuscrit, une disquette de 3,5 ou 5,25 pouces en précisant le type de logiciel.
Les figures, tableaux et autres planches sont remis sous la forme de tirages ou de dessins définitifs, de qualité optimale, prêts à l’impression. Ils peuvent être également fournis sur disquette s’ils ont été réa-lisés au moyen de logiciels d’utilisation courante (Excel ou Word par ex.)
Les auteurs reçoivent deux épreuves, la deuxième signée faisant office de bon-à-tirer. (Pour la partici-pation éventuelle aux frais de publication, voir deuxième page de couverture du Bulletin).
II. Remarques pour la rédaction
Titres: En cas de nombreux titres ou sous-titres d’importances diverses, indiquer leur hiérarchie par des soulignages de couleurs différentes.
mots-latins: Ils doivent être en italique dans le texte imprimé (p. ex. Scilla bifolia) et seront soulignés une fois dans le manuscrit dactylographié. Les noms français d’espèces sont écrits sans majuscules.
Noms d’auteurs: Ils doivent être composés en petites capitales s’il s’agit d’une citation, p. ex. (Linné 1758), linné (1758) ou (Goto & ozaki, 1930), et sont soulignés deux fois dans le manuscrit dactylo-graphié. quand plus de deux auteurs signent un travail, le nom du premier est suivi de la mention «et al.» (p. ex. Cort, W. et al.). Cette remarque n’est pas valable pour la bibliographie. Si, au contraire, le nom d’auteur est associé à celui d’un taxon, il s’écrit en romain et ne doit pas être souligné (p. ex. Scilla bifolia Linné); il est recommandé dans ce cas de l’abréger, en se conformant aux abrévations usuelles (p. ex. Scilla bifolia L.). Dans le texte, les titres d’ouvrages ou d’articles sont placés entre guillemets.
Numérotation des figures et des planches dans le texte: Entre parenthèses et en chiffres arabes pour les figures et tableaux (fig. 12, tab.1), en chiffres romains pour les planches (pl. III, fig.5). Dans le texte courant et dans les légendes, les mots «figure» et «planche» ne sont pas abrégés.
Abréviations et symboles usuels: a) Abréviations (suivies d’un point)vol. = volume fig., tab. = figure, tableaut. = tome pl. = planchep. = page (pp.= pages) chap. = chapitre
b) Symboles (non suivis d’un point)m = mètre g = grammecm = centimètre kg = kilogrammemm = millimètre mg = milligramme

198
µm = micromètre, ou micron µg = microgramme (et non µ) 1µm = 10 -6 m h = heurenm = nanomètre (et non mµ) mn = minute 1 nm = 10 -9 m s = secondekm = kilomètre o C ou o K = degré Celsius ou Kelvinl = litre 1er, 2e = premier, deuxièmeml = millilitre N S E W = points cardinauxµl = microlitre NE SSW = (ni trait d’union, ni espace)
Pour les autres unités: faire référence à une liste officielle (par ex: Ecological Society of America).
c) Coordonnées géographiques: les valeurs seront indiquées en nombres décimaux, séparées par un espace (ni point, ni virgule).
Bibliographie: A établir par ordre alphabétique des noms d’auteurs et selon les modèles suivants:Livres:bernarD, f. 1968. Les Fourmis (Hymenoptera Formicidae) d’Europe occidentale. Faune de L’Europe
et du Bassin méditerranéen. masson. Paris.
Citations:reznikova, l. 1983. Interspécific communications between ants. Behaviour 80: 84-85.sHiMazu, s.; Hirano, t. & ueMatsu, T. 1987. Shape-Selective Hydrogenation by Ruthenium-Hecto-
rite Catalysts with various Interlayer Distances. Appl. Catal. 34: 255.
Dans une série d’articles du même auteur, le nom de celui-ci est répété à chaque fois. Si deux articles consécutifs ont été publiés dans la même revue, le nom de celle-ci ne sera pas répété, mais remplacé par Ibid.
Référence au Bulletin de la Société neuchâteloise des Sciences naturelles: Bull. Soc. neuchâtel. Sci. nat.
Légendes: Elles seront réunies à la fin du texte dactylographié.
Résumés: Il est recommandé de rédiger trois résumés (en français, en allemand et en anglais), donnant l’essentiel des résultats avec, si nécessaire, la traduction du titre de la publication. Les articles en anglais comprennent un résumé étendu en français. Des mots-clés en français et anglais sont également indiqués.




















