BC Master CSQ1
78
Module :« Energie et fonctionnement cellulaire » Master S.V.T Le Cytosquelette
description
Â
Transcript of BC Master CSQ1
Module :« Energie et fonctionnement cellulaire »
Master S.V.T
Le Cytosquelette
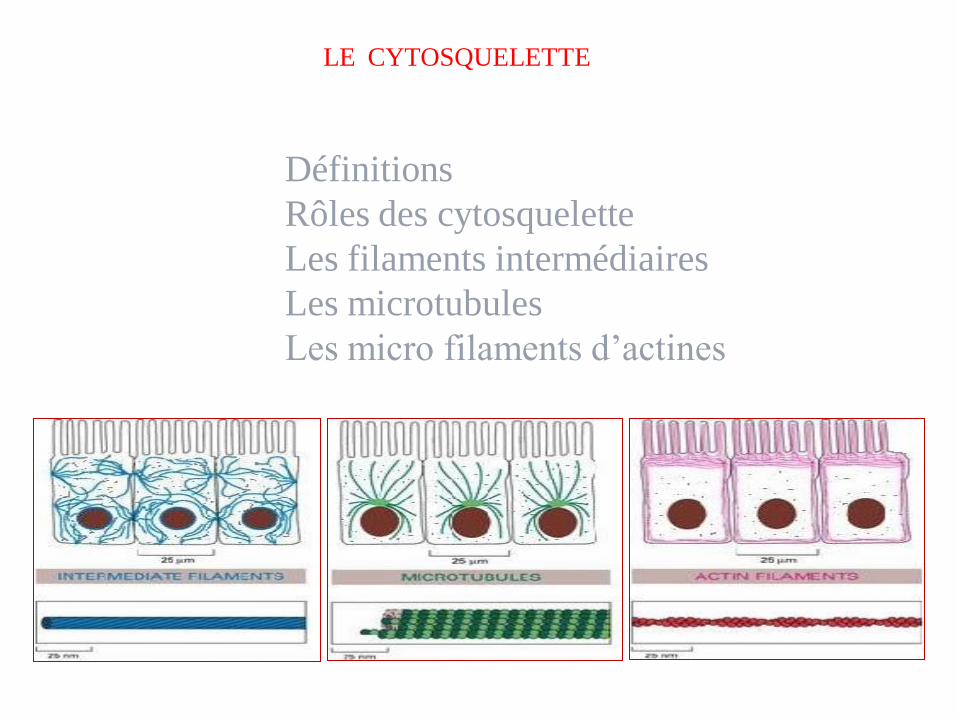
LE CYTOSQUELETTE
Définitions
Rôles des cytosquelette
Les filaments intermédiaires
Les microtubules
Les micro filaments d’actines

Définitions
Le CSQ est l’ensemble organisé des polymères biologiques qui lui
confèrent l'essentiel de ses propriétés mécaniques :
la structure
la déformation
la mobilité cellulaire
Localisation
Il est localisé en périphérie cellulaire dans le cytoplasme et
dans le nucléoplasme
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
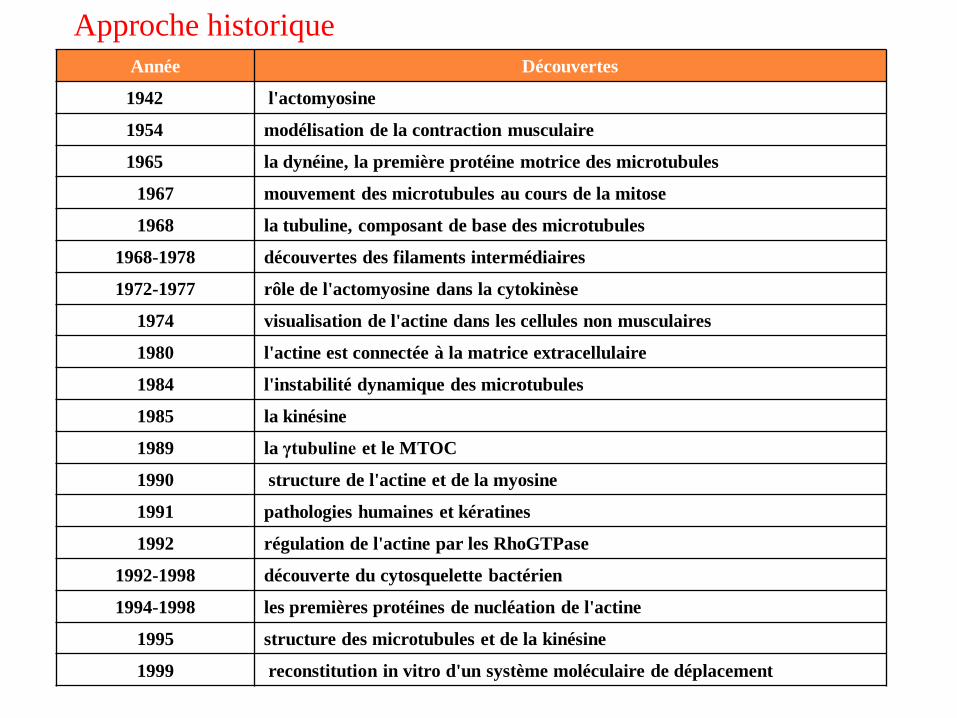
Année Découvertes
1942 l'actomyosine
1954 modélisation de la contraction musculaire
1965 la dynéine, la première protéine motrice des microtubules
1967 mouvement des microtubules au cours de la mitose
1968 la tubuline, composant de base des microtubules
1968-1978 découvertes des filaments intermédiaires
1972-1977 rôle de l'actomyosine dans la cytokinèse
1974 visualisation de l'actine dans les cellules non musculaires
1980 l'actine est connectée à la matrice extracellulaire
1984 l'instabilité dynamique des microtubules
1985 la kinésine
1989 la γtubuline et le MTOC
1990 structure de l'actine et de la myosine
1991 pathologies humaines et kératines
1992 régulation de l'actine par les RhoGTPase
1992-1998 découverte du cytosquelette bactérien
1994-1998 les premières protéines de nucléation de l'actine
1995 structure des microtubules et de la kinésine
1999 reconstitution in vitro d'un système moléculaire de déplacement
Approche historique
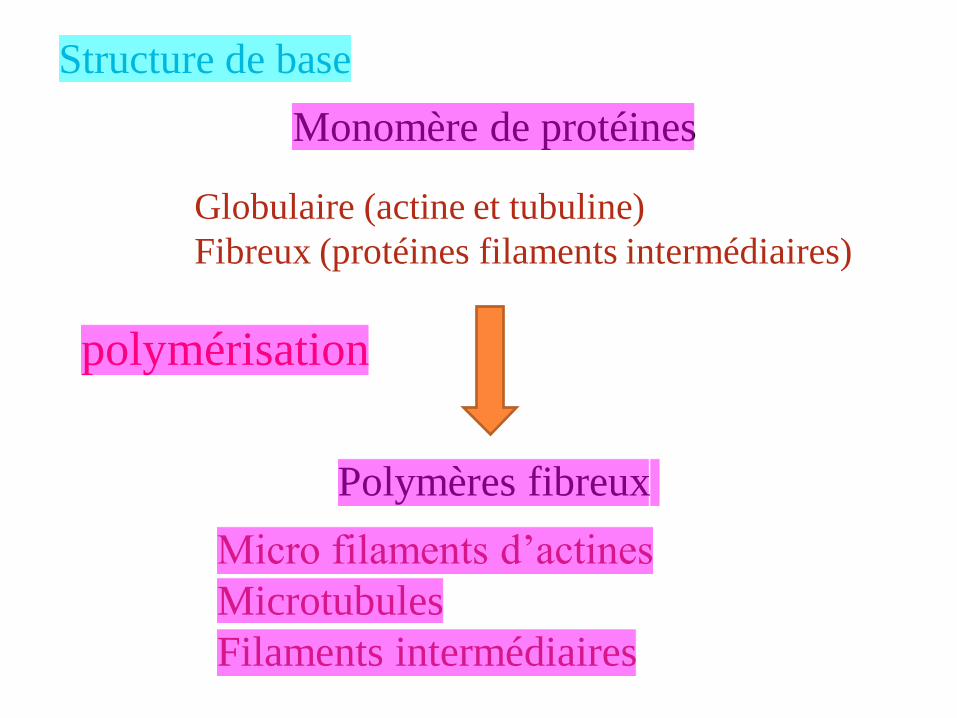
Structure de base
Monomère de protéines
Polymères fibreux
polymérisation
Globulaire (actine et tubuline)
Fibreux (protéines filaments intermédiaires)
Micro filaments d’actines
Microtubules
Filaments intermédiaires
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Association des monomères
Protéines des FI
Phosphorylation
O.Glycolysation
Déphosphorylation
Filaments intermédiaires
Actine ATP Micro filaments d’actines
ATP
GDP
Tubuline GT¨P
GTP
GDP
Microtubules
Monomères polymères
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Rôles des cytosquelette
Les polymères assurent:
La mobilité cellulaire:
Migration cellulaire
Contractilité cellulaire
Actine Migrations des organites et vésicules
Mouvement de la membrane plasmique
La structure de la cellule (microvillosités et
flagelles) et des organites (Microtubules pour le
Golgi, lamine pour le noyau)
tokyo
Highlight
tokyo
Highlight
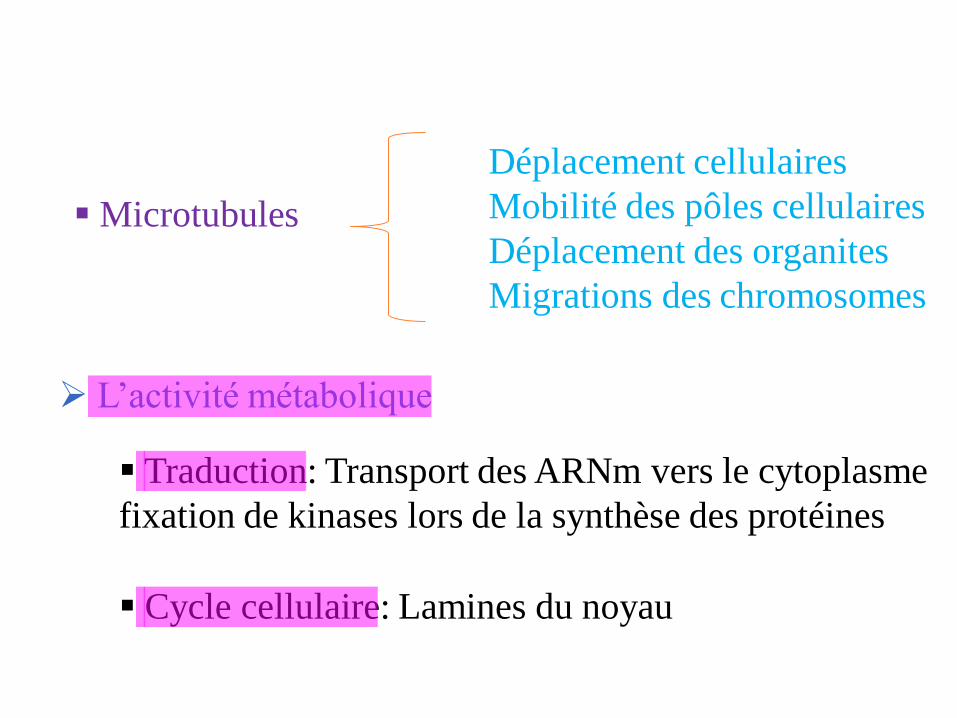
Microtubules
Déplacement cellulaires
Mobilité des pôles cellulaires
Déplacement des organites
Migrations des chromosomes
L’activité métabolique
Traduction: Transport des ARNm vers le cytoplasme
fixation de kinases lors de la synthèse des protéines
Cycle cellulaire: Lamines du noyau
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

L’activité mitotique
Réorganisation des microtubules pour créer le fuseau
mitotique ,dépolymérisation des FI
Individualisation des cellules filles par l’actine
Activation des enzymes de la mitose
La prolifération bactérienne ou virale
tokyo
Highlight
tokyo
Highlight

Les protéines de pontage (couramment désignées par les termes
anglais crosslinkers ou crosslinking proteins).
Niveau d'organisation est rendu possible par la présence de centaines de
protéines auxiliaires :
Les protéines de branchement
Les protéines capuchon capping,
Les protéines dépolymérisantes (severing proteins)
Les protéines d'ancrage
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
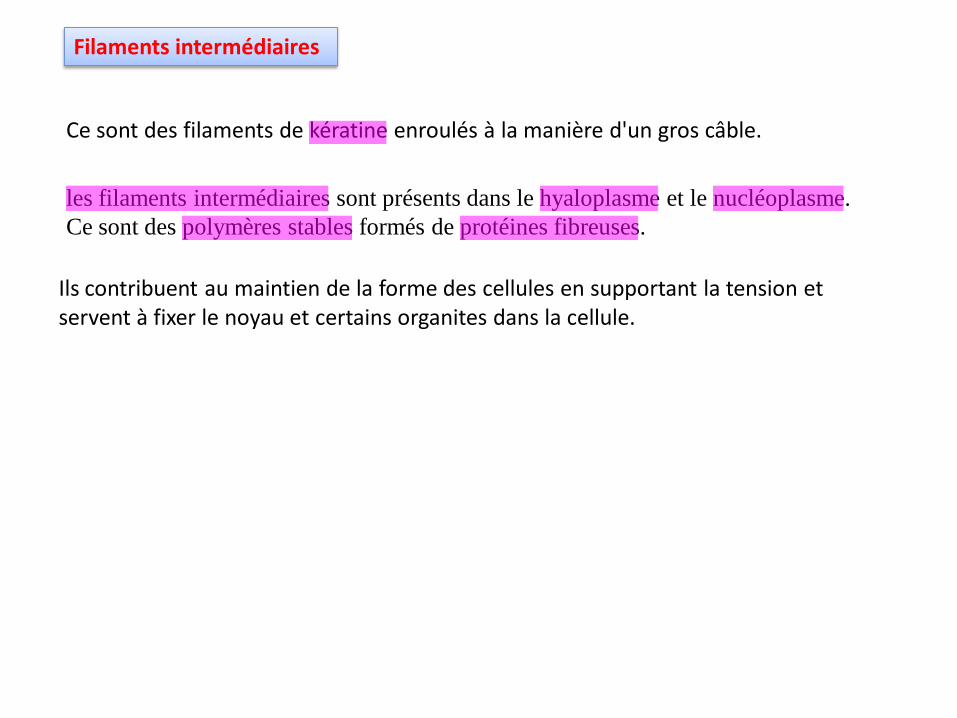
Filaments intermédiaires
Ce sont des filaments de kératine enroulés à la manière d'un gros câble.
Ils contribuent au maintien de la forme des cellules en supportant la tension et servent à fixer le noyau et certains organites dans la cellule.
les filaments intermédiaires sont présents dans le hyaloplasme et le nucléoplasme.
Ce sont des polymères stables formés de protéines fibreuses.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Structure
Ce sont des constituants du cytosquelette qui sont formés de protéines fibrillaires
assemblées de façon hélicoïdale.
Il existe 5 sous types de filaments, qui sont des polymères de différents types de
protéines fibrillaires:
kératines de type acide, (les cellules épithéliales),
kératines de type basique,
vimentine et apparentés ( les fibroblastes)
neurofilaments (neurones)
lamines (dans les noyaux, )
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
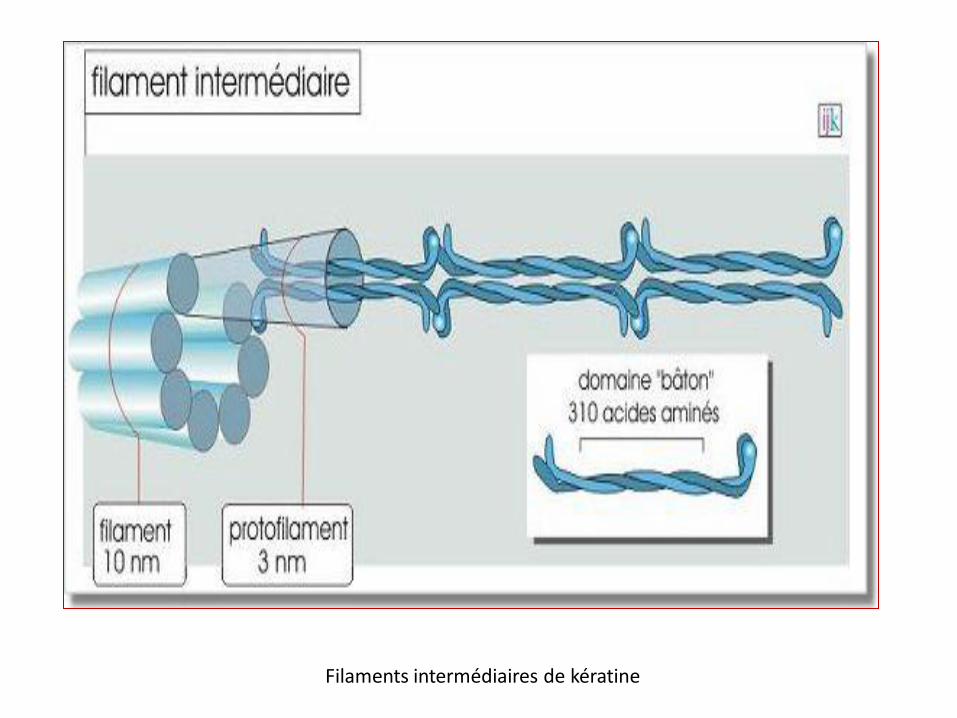
Filaments intermédiaires de kératine
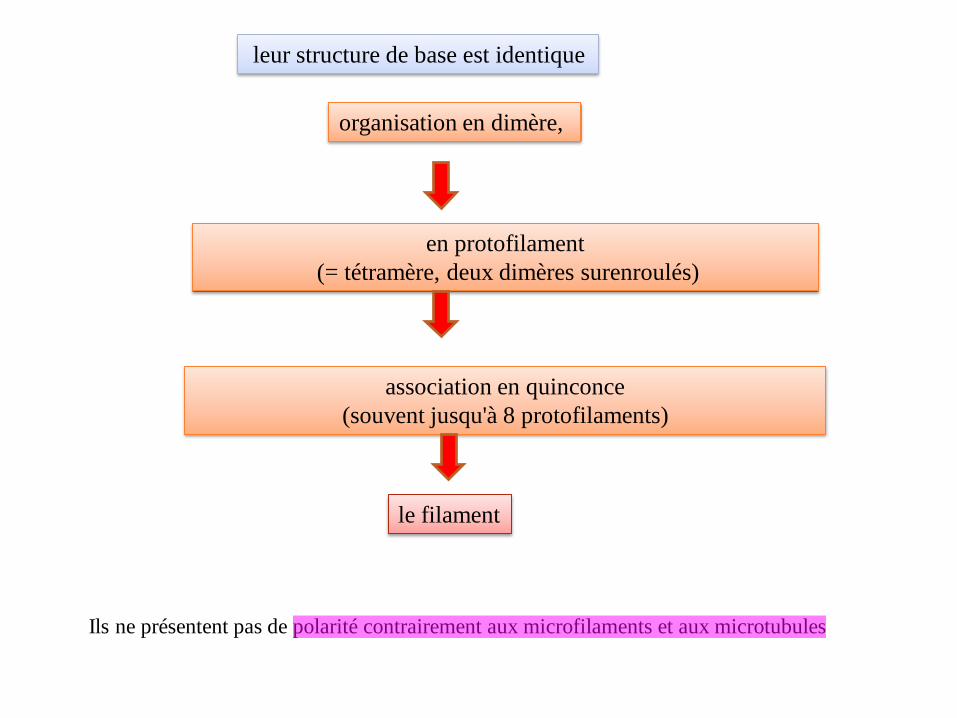
leur structure de base est identique
organisation en dimère,
en protofilament
(= tétramère, deux dimères surenroulés)
association en quinconce
(souvent jusqu'à 8 protofilaments)
le filament
Ils ne présentent pas de polarité contrairement aux microfilaments et aux microtubules
tokyo
Highlight

Fonction
au maintien de la forme cellulaire
à l'ancrage des organites cellulaires.
Ils interviennent
dans la solidité de la cellule.
dans l'adhérence et la cohésion cellulaire car ils sont en relation avec les
desmosomes et les hémidesmosomes.
Ils conférerent aux cellules de l'épiderme
une résistance aux frottements.
Ils concourent
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
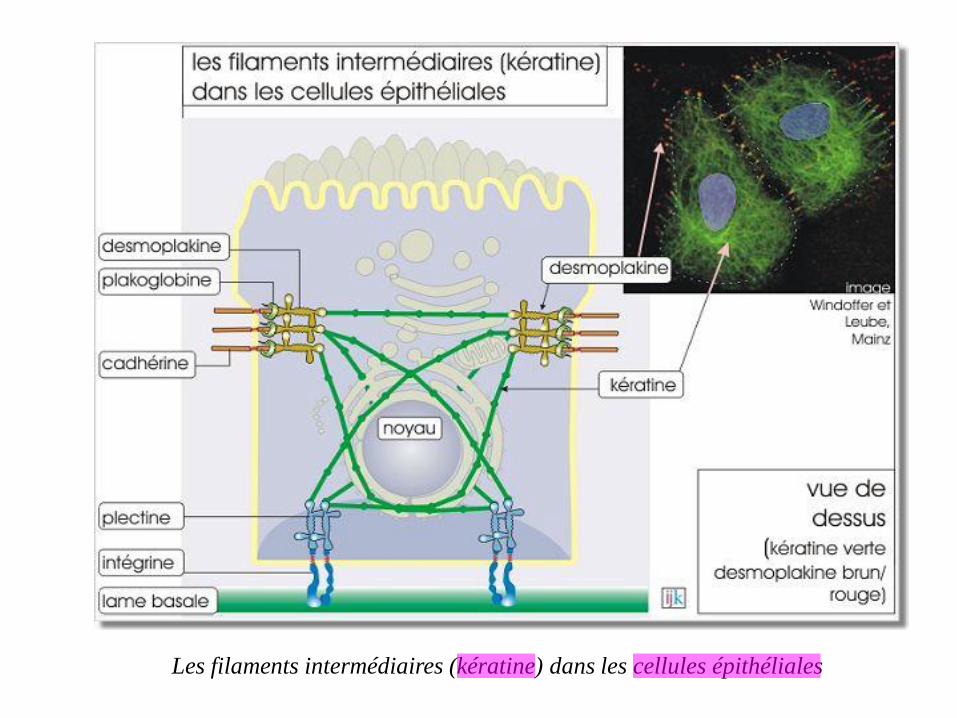
Les filaments intermédiaires (kératine) dans les cellules épithéliales
tokyo
Highlight
tokyo
Highlight

La polymérisation des filaments intermédiaires
Contrairement à l'actine et à la tubuline, qui sont des protéines globulaires, les divers types de
protéines qui constituent les filaments intermédiaires sont des molécules fibreuses très
allongées.
Leur séquence en acides aminés favorise la formation de dimères superenroulés
Au cours de l'étape d'assemblage, deux des dimères superenroulés s'associent de manière
antiparallèle pour former une sous-unité tétramérique.
C'est un protofilament (3 nm de diamètre).
Les tétramères s'ajoutent à un filament intermédiaire en cours d'élongation et
8 protofilaments forment le filament intermédiaire de 10 nm de diamètre.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Les composants des filaments intermédiaires
A l'inverse des gènes de l'actine et de la tubuline, bien conservés au cours
de l'évolution, les gènes codant les filaments intermédiaires sont diversifiés
et, à ce jour, sont regroupés en une famille d'environ 50 membres formant
6 classes différentes.

Fonctions des filaments intermédiaires
Maintien de l'intégrité cellulaire et tissulaire de l'épithélium
Dans le chapitre «les molécules d'adhérence » nous avons déjà montré l'importance
du cytosquelette dans le maintien mécanique de la cellule et du tissu.
Dans les cellules épithéliales, les filaments intermédiaires sont fortement impliqués
dans deux types de jonctions d'ancrage :
desmosome, interaction cellule-cellule, où ils sont liés avec les membres
de la famille des cadhérines
hémi-desmosome, interaction cellule-lame basale, où ils sont liés aux
intégrines
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Un treillis de filaments intermédiaires, polymères de lamine, qui double la face interne
de l'enveloppe nucléaire, forme la lamina nucléaire.
Elle soutient l'enveloppe et donne au noyau sa forme généralement globulaire.
Pendant la mitose, la lamina nucléaire se désagrège grâce à la phosphorylation des
lamines.
Sa désintégration permet l'entrée d'un autre type d'élément du cytosquelette, les
microtubules, qui, comme on le verra plus tard, participent à la séparation des
chromosomes
Soutien de l'enveloppe nucléaire
tokyo
Highlight
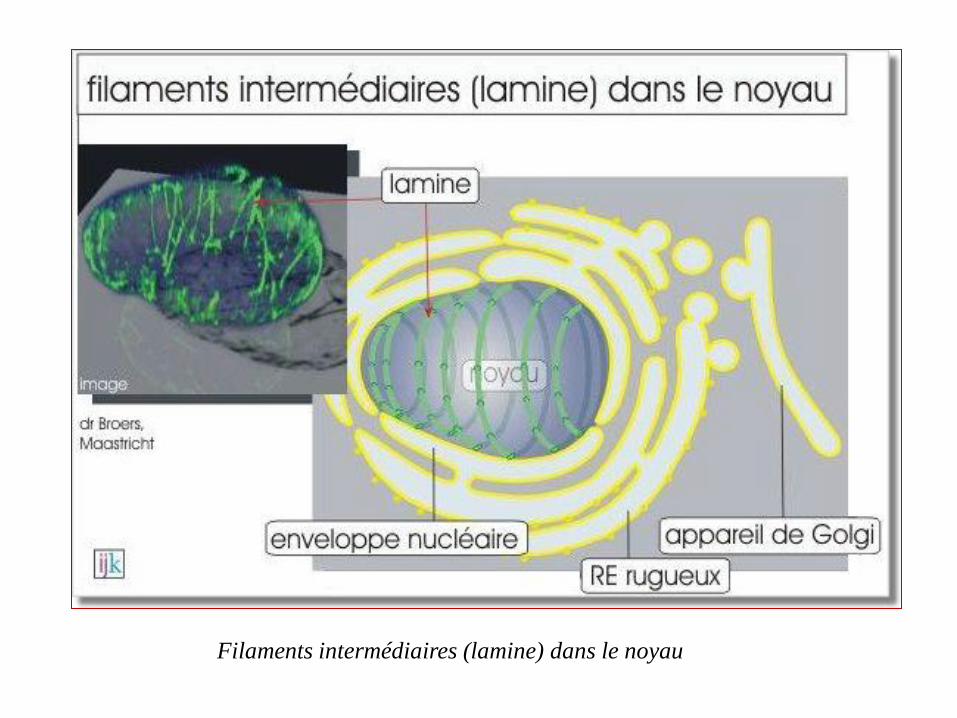
Filaments intermédiaires (lamine) dans le noyau

Les filaments de kératine sont formés en excès par les cellules épidermiques
(kératinocytes) et les cellules de l'assise génératrice dans le follicule pileux. Cette
expression excessive entraîne la mort des cellules qui restent assemblées (par des
desmosomes) et qui forment progressivement la couche cornée, un ongle ou un poil
(ou un cheveu). Les caractéristiques des filaments intermédiaires, résistance aux
tensions et aux détergents, insolubilité dans l'eau, sont donc essentielles pour une
bonne défense contre les agressions physiques et chimiques dirigées contre
l'organisme entier.
Formation des ongles, cheveux et couche cornée de la peau
tokyo
Highlight
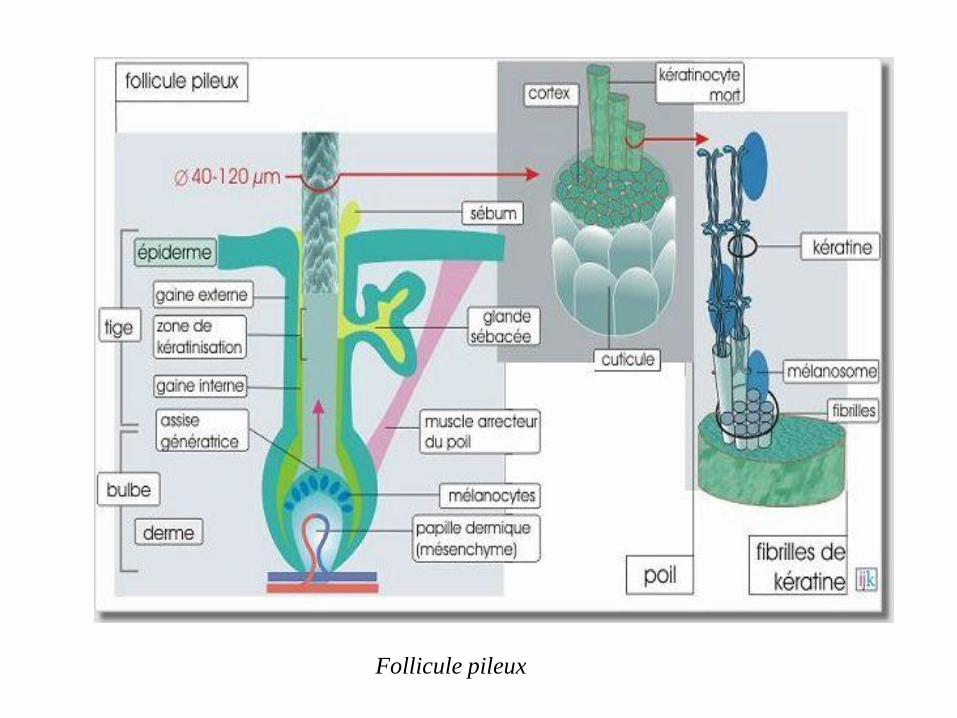
Follicule pileux
Les microtubules (MT)
Les microtubules (MT) sont des fibres constitutives du cytosquelette, au même
titre que les microfilaments d'actine et que les filaments intermédiaires.
Les microtubules sont impliqués dans la formation de la plaque équatoriale et
le déplacement pôlaire des chromosomes pendant la mitose.
tokyo
Highlight
tokyo
Highlight
Microtubules
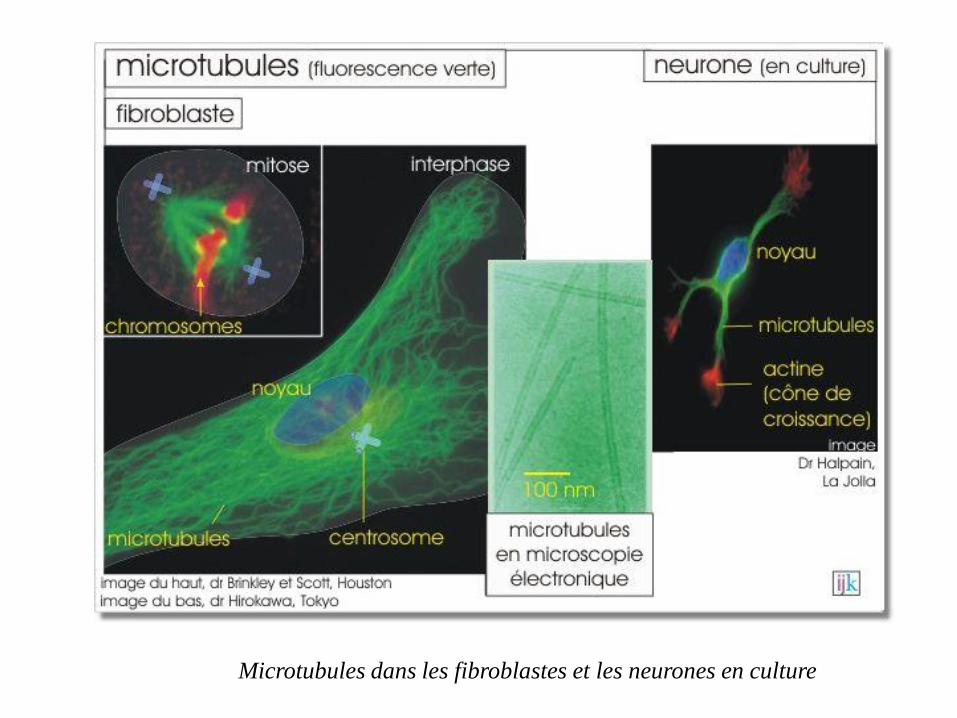
Microtubules dans les fibroblastes et les neurones en culture

Composition
Un microtubule est un tube dont la paroi est constituée de plusieurs protofilaments
de tubuline; on en compte 13 par microtubule.
Chaque protofilament est constitué d'un assemblage orienté de dimères de tubuline :
tubuline α
tubuline β
reliées par des liaisons non covalentes
Les deux extrémités des microtubules ont des propriétés différentes (polarité du
microtubule).
Cet assemblage polymérique est extrêmement dynamique : les extrémités des
microtubules polymérisent et dépolymérisent en permanence
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Phase d'équilibre : le microtubule est soumis au processus de tapis roulant décrit
plus haut.
Polymérisation
La polymérisation met en jeu l'hydrolyse de la GTP (guanosine triphosphate).
Il y a trois phases dans la mise en place du microtubule :
Phase de nucléation : plus rapide que pour l'actine, elle consiste en l'assemblage des
hétérodimères alpha et béta concomitant à l'hydrolyse de la GTP (guanosine
triphosphate) catalysée par la sous unité bêta. Ce "noyau" est la base sur laquelle le
microtubule croît.
Phase d'élongation : 13 protofilaments forment un microtubule.
In vitro, les dimères se lient aux deux extrémités libres du microtubule, bien
que la polymérisation soit plus rapide à l'extrémité (+) du microtubule.
In vivo, le pôle (-) est stabilisée pour être liée au centre de nucléation
(centrosome) du microtubule quelle que soit sa localisation dans la cellule.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
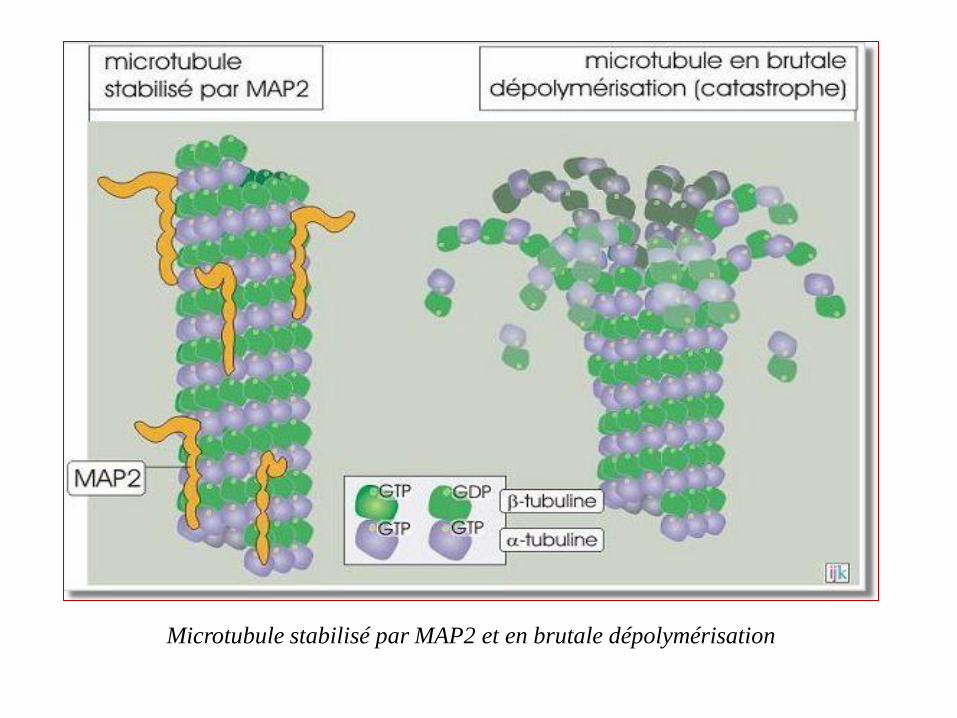
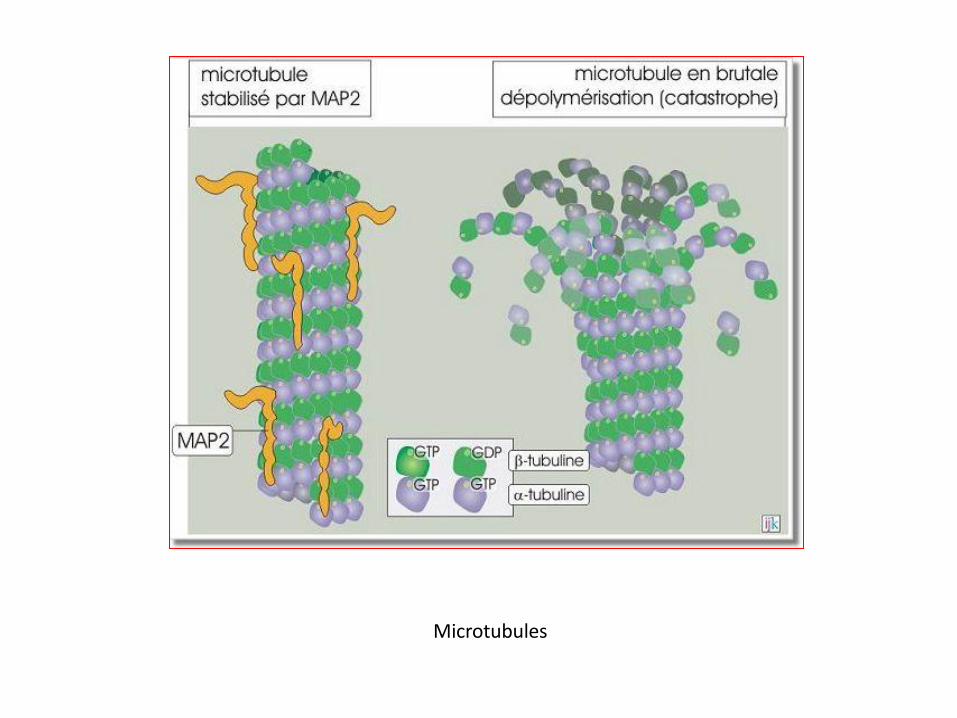
Microtubule stabilisé par MAP2 et en brutale dépolymérisation

Les microtubules sont des structures polaires : une extrémité est capable de croissance
rapide (extrémité plus), alors que l'autre extrémité se trouve le plus souvent enchâssée
dans le centrosome (extrémité moins). Le centrosome est un complexe protéique organisé
autour deux structures appelées centrioles. Les centrioles sont faits de plusieurs formes de
tubuline (α, β ,γ , δ et ε )
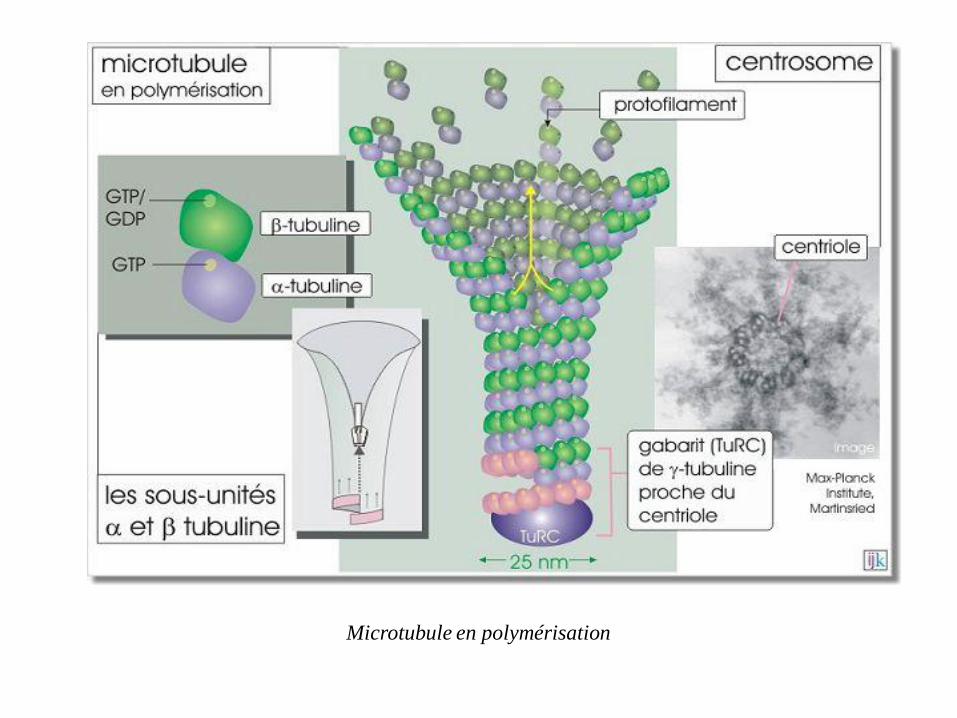
Assemblage des microtubules
A la périphérie du centrosome on trouve la tubuline- faisant partie d'un complexe
(Tubulin Ring Complex, TuRC) dont la conformation sert de gabarit au microtubule
en construction
A partir d'un centrosome, les dimères de tubuline (α et β) chargés en GTP sont ajoutés
(α du côté pôle moins et β du côté pôle plus) et élaborent des protofilaments, qui
s'assemblent latéralement entre eux, formant ainsi des feuillets.
Les feuillets se replient progressivement sur eux mêmes pour former le microtubule,
cylindre creux et rigide
Généralement le microtubule est composé de 13 protofilaments mais il existe des
microtubules qui sont le résultat de l'assemblage de 11 ou 16 protofilaments.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Les kinésines moteurs moléculaires qui se déplacent vers l'extrémité
positive (+), on parle de transport antérograde.
Les dynéines moteurs moléculaires qui se déplacent vers l'extrémité
négative (-), il s'agit d'un transport rétrograde.
Protéines associées
Les protéines associées aux microtubules ou map, les unes ont un rôle dans
la stabilisation des microtubules,
le mouvement des vésicules et des organites le long des microtubules ; ce
sont des ATPases :
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
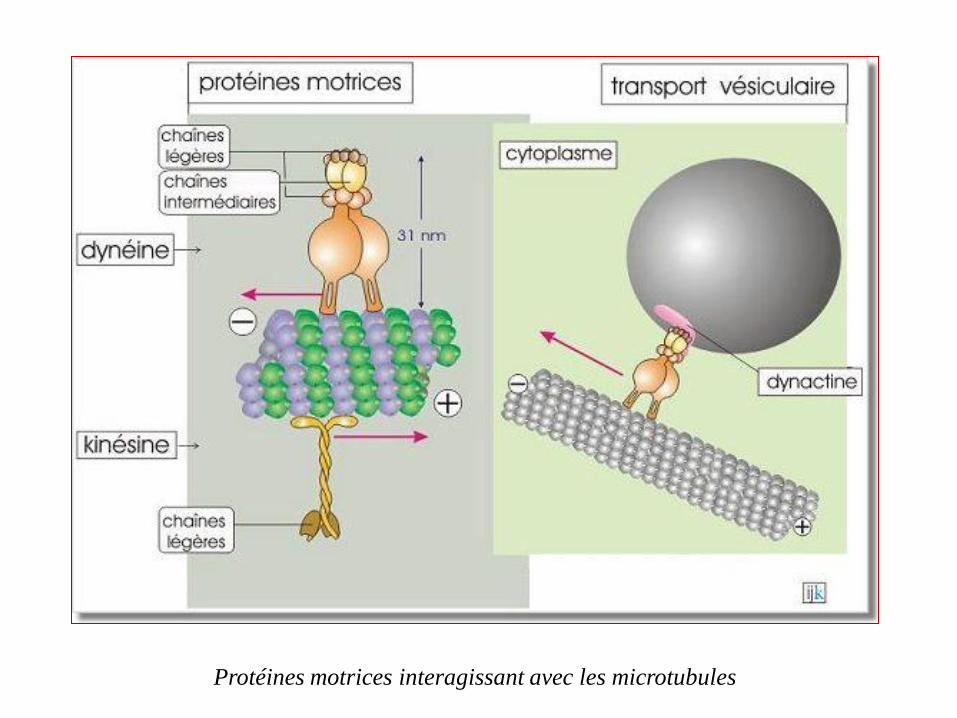
Protéines motrices interagissant avec les microtubules
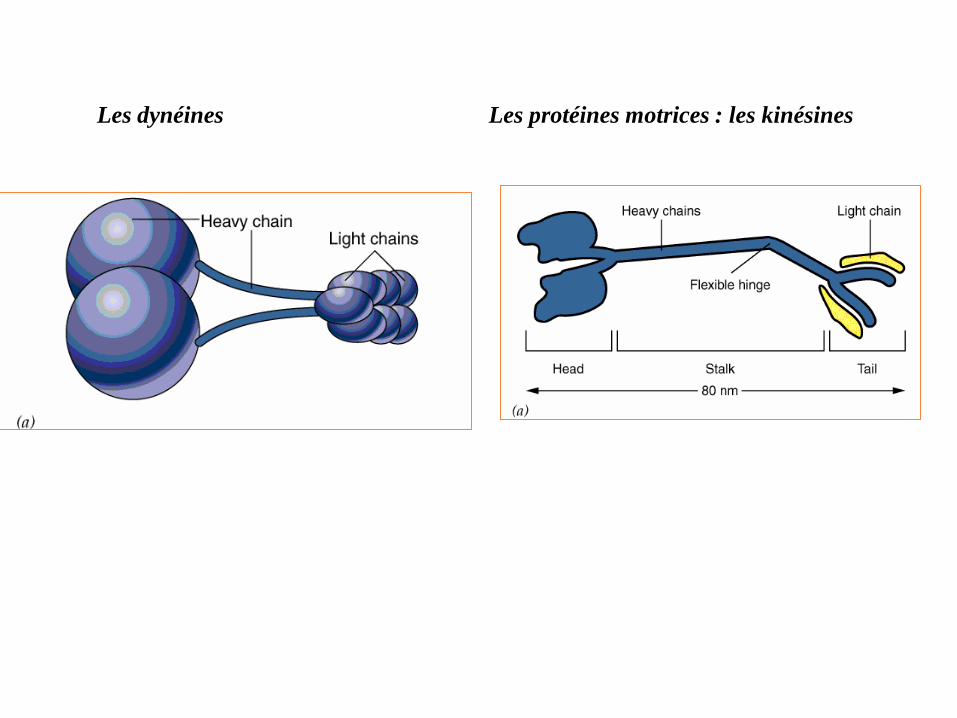
Les dynéines Les protéines motrices : les kinésines
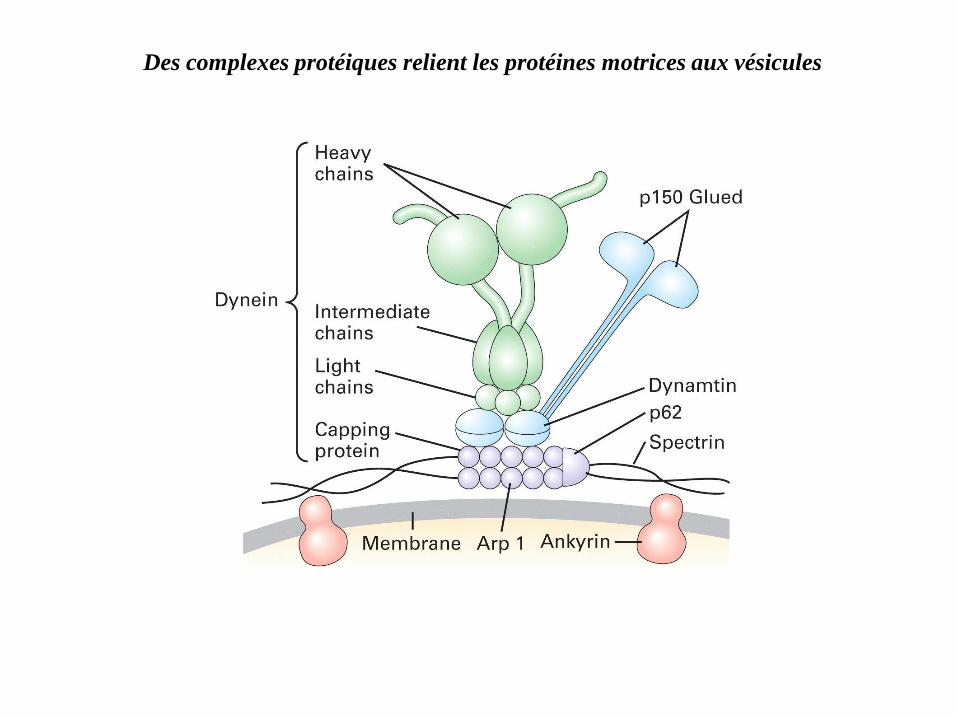
Des complexes protéiques relient les protéines motrices aux vésicules

Les microtubules kinétochoriens : relient les centrosomes aux kinétochores
(situés au niveau du centromère des chromosomes)
Rôles
Division cellulaire (la mitose)
Les microtubules jouent un rôle très important dans la division cellulaire (mitose) et
dans les courants cytoplasmiques grâce à la coopération entre les microfilaments
d'actine (le déplacement des chromosomes).
L'apparition des microtubules en phase G2 de l'interphase.
Les microtubules polaires : permettent aux 2 centrosomes de s'éloigner l'un de l'autre.
Les microtubules radiaires (également appelés microtubules astériens) : ancrent les
centrosomes aux membranes plasmiques
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Transport cytoplasmique
Les déplacements cellulaires se font avec la concurrence entre les MA
(microfilaments d'actine) et les MT
Les microtubules sont très nombreux dans les neurones, en particulier dans les
dendrites et les axones.
Ils permettent d'acheminer divers composants vers leurs extrémités, servant de
véritables rails sur lesquels des moteurs moléculaires, attachés aux composants
à transporter, se déplacent.(vésicules synaptiques ou des mitochondries).
Le transport est possible grâce à des protéines associées aux microtubules
(PAM ou MAP en anglais) :
MAP de mobilisation Entre autres la kinésine (vers l'extrémité plus),
et la dynéine (vers l'extrémité moins).
MAP d'assemblage (proteines Tau, MAP-2).
MAP de stabilisation (STOP)
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
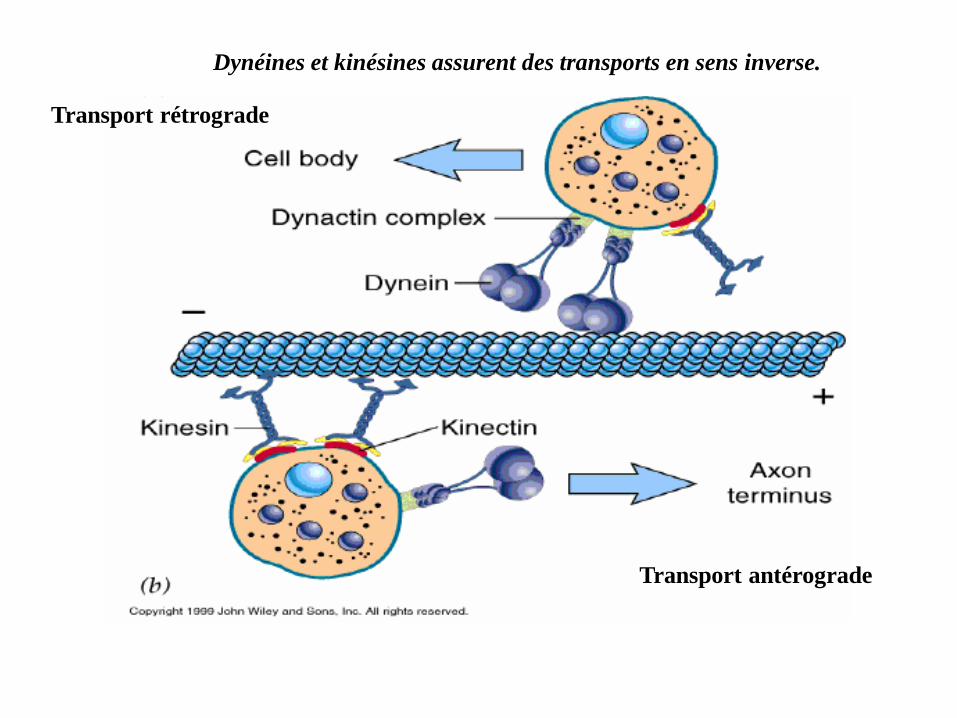
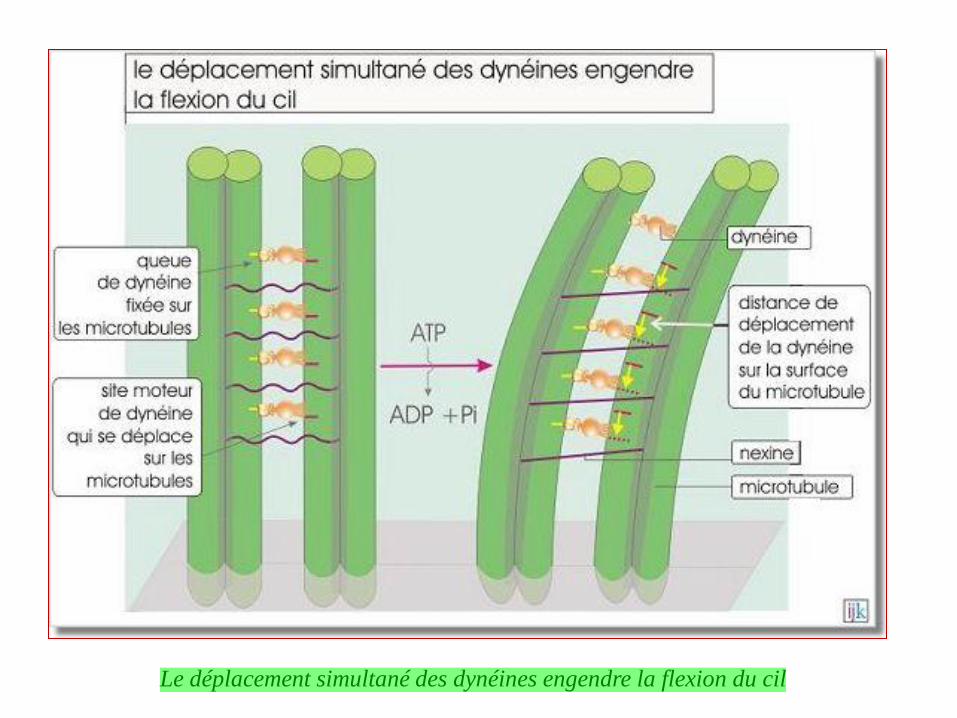
Dynéines et kinésines assurent des transports en sens inverse.
Transport rétrograde
Transport antérograde

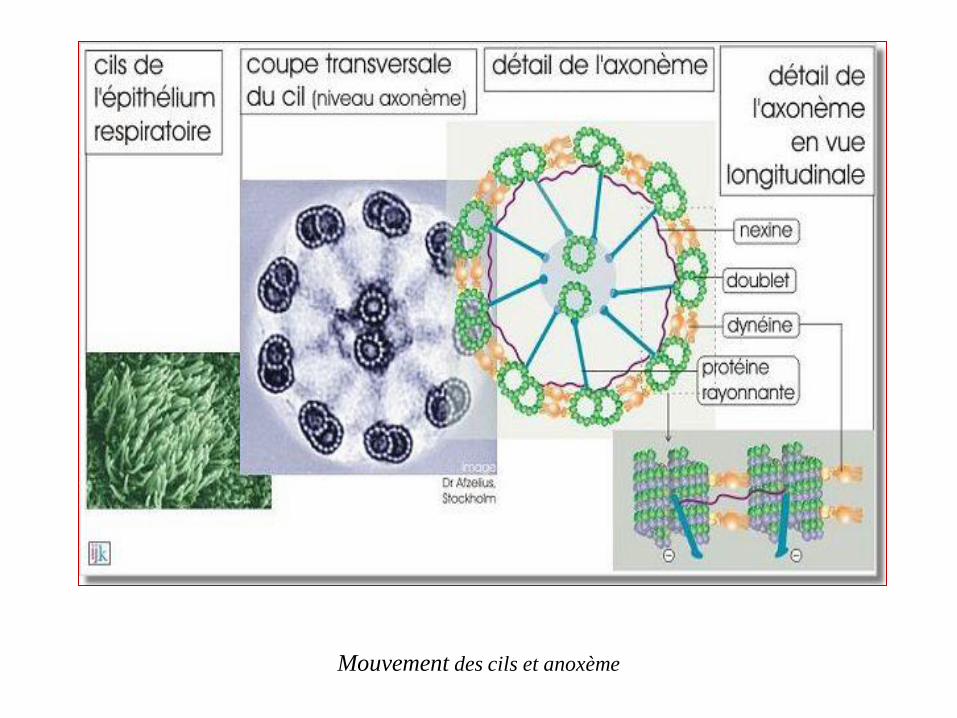
Les microtubules composent majoritairement le filament axial flagellaire des
cellules eucaryotes mobiles comme les spermatozoïdes matures et les protozoaires.
Mobilité cellulaire
Autres
Les microtubules ont d'autres rôles, parmi eux le maintien de la forme tridimensionnelle
et la diapédèse et le transport de vésicules d'endocytose et d'exocytose.
Molécules agissant sur les microtubules
Inhibiteurs de la polymérisation
la colchicine (par réaction réversible)
la vinblastine,
la vincristine Ceux-ci modifient l'équilibre de la réaction de polymérisation et
favorisent ainsi la dépolymérisation.
le nocadazole.
Activateurs de polymérisation
le Taxol.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Régulation
Les microtubules sont des structures extrêmement dynamiques. Ils sont synthétisés
et dégradés dans des temps très brefs.
Certains médicaments peuvent les déstabiliser ou, au contraire, les stabiliser.
La biogénèse des microtubules
les MT se polymérisent à partir de centres organisateurs (MTOC) :
kinétochores (division cellulaire).
corpuscule basal ou cinétosome (cils et flagelles).
la matière péricentriolaire amorphe (cellules animales) ou masse amorphe
(cellules végétales) pour la polymérisation des autres MT.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
Microtubule en polymérisation

Stabilisation des microtubules
Les cellules peuvent modifier cette instabilité dynamique par des
protéines qui s'associent aux microtubules sur toute leur longueur : les
"microtubule-associated proteins » MAP2 (200 kDa), MAP4 (135 kDa)
ou la protéine Tau (45 kDa). On estime que la présence de ces protéines
réduit 50 fois la probabilité de déclenchement d'une brutale dissociation
(catastrophe).
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
Mouvement des cils et anoxème
Le déplacement simultané des dynéines engendre la flexion du cil
tokyo
Highlight

Un filament d'actine, ou microfilament, est un homopolymère
d'actine, protéine de 42 kDa (Unité de masse atomique).
C'est un constituant essentiel du cytosquelette des cellules eucaryotes,
ainsi que des fibres musculaires.
L'actine représente ainsi environ 5% du total des protéines d'une
cellule animale typique, la moitié étant assemblée en filaments d'actine
alors que l'autre moitié est libre dans le cytosol sous forme de monomères
d'actine.
L'actine sous forme de filaments est parfois appelée actine F
(Fibrillaire), tandis que la forme monomérique est appelée actine G
(Globulaire).
Filament d'actine, ou microfilament
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

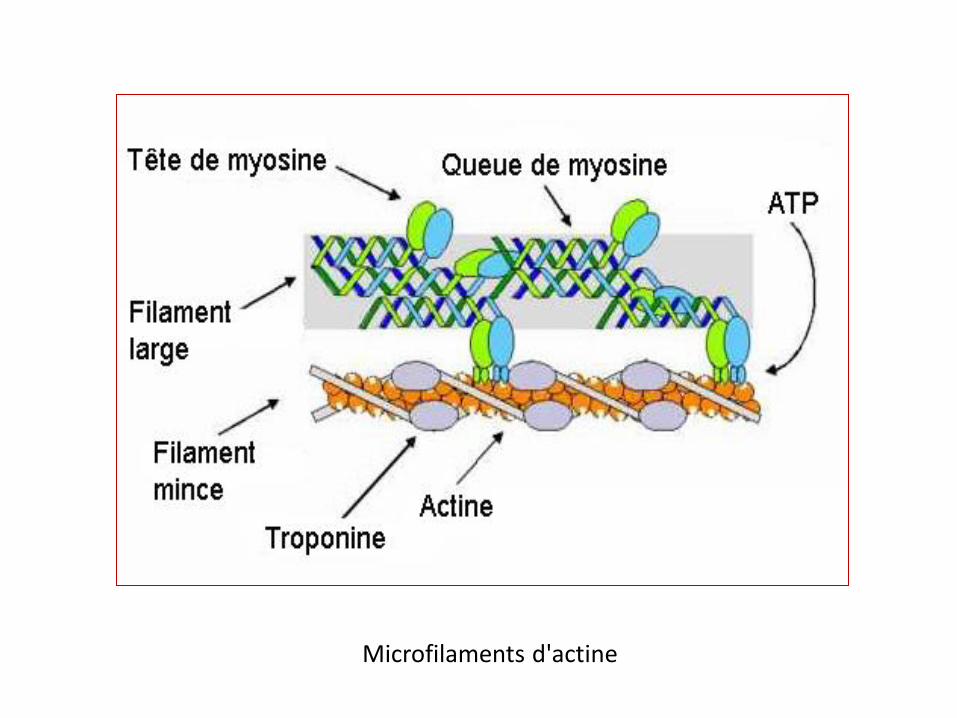
Les microfilaments d'actine sont constitués de deux chaînes d'actine entortillées.
Elles contribuent au maintien de la forme des cellules en supportant la tension.
Impliquées dans la contraction musculaire (avec la myosine).
Elles participent au transport intracellulaire
Responsables de la formation du sillon lors de la division cellulaire.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
Microfilaments d'actine

Structure d'un filament
Un filament d'actine est composé de deux brins constitués de monomères d'actine,
enroulés l'un autour de l'autre, dans une hélice double.
Chaque filament est polarisé car toutes les molécules d'actine globulaire "pointent"
dans la même direction, vers une extrémité appelée (-), l'autre extrémité étant
nommée (+).
A cette polarité structurelle s'ajoute une polarité de polymérisation.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
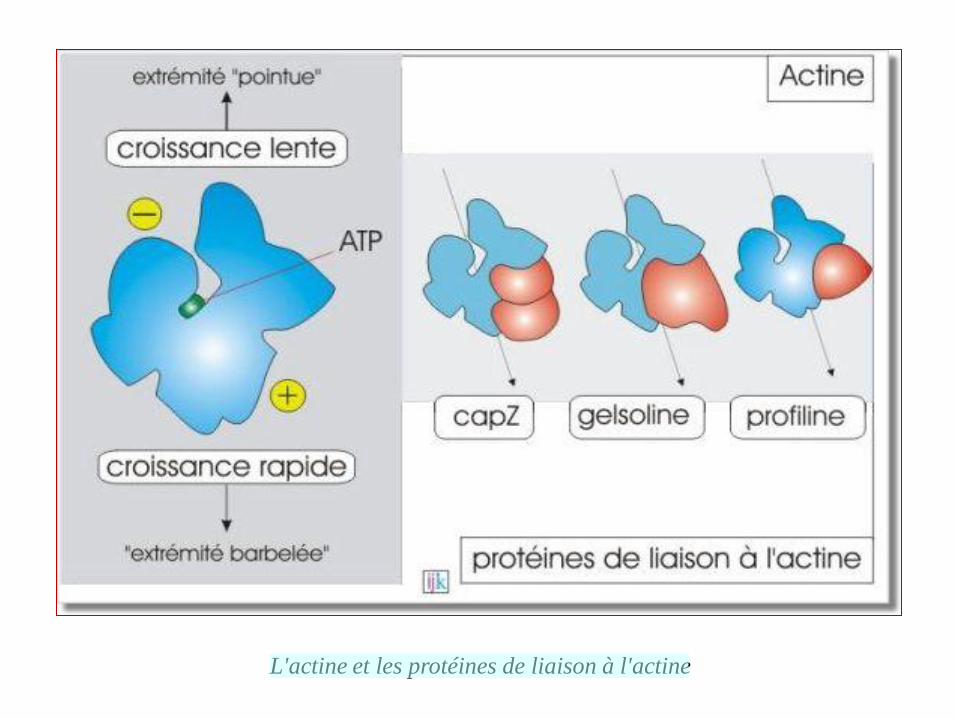
L'actine et les protéines de liaison à l'actine
tokyo
Highlight

Les monomères d'actine ainsi libérés doivent être rechargés en ATP avant
de rejoindre le filament
La polymérisation de l'actine
L'actine se polymérise (en présence d'ATP) en une hélice serrée de 5-9 nm
de diamètre formant un filament flexible et polaire
Lorsque l'on solubilise l'actine en présence de KCl, ATP, Mg2+ et d'un catalyseur tel que le complexe ARP2/3 qui permet de fixer les premiers monomères (amorce), elle forme spontanément des polymères (filament d'actine ou actine-F).
Après la polymérisation, une hydrolyse aléatoire de l'ATP a lieu, le phosphate
(Pi) est libéré et l'ADP qui en résulte reste piégé dans le polymère.
Les molécules d'actine liées à l'ADP ont tendance à se détacher du polymère
aux extrémités des filaments.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

In vivo, la polymérisation de l'actine est contrôlée par de nombreuses protéines,
comme la profiline, le complexe ARP2/3, CapZ et la gelsoline
La profiline (15 kDa) se fixe à l'actine monomérique liant alors l'ATP et
aidant à la réintégration de l'actine dans le polymère.
Le complexe protéique ARP2/3 (Actin Related Proteins 2 et 3, de 42 et
47 kDa respectivement) est impliqué dans l'initiation de la polymérisation.
Le complexe se fixe côté moins de l'actine et sa présence favorise la formation
d'une amorce constituée de trois monomères liés entre eux (site de nucléation
pour la formation de longs polymères).
L'ARP2/3 joue donc un rôle important dans la désignation des sites où l'actine
doit se polymériser.
Les pôles plus et moins des filaments peuvent être protégés par les protéines de
coiffage (capping). Ces protéines empêchent l'actine, dans son état ADP, de quitter
le polymère mais empêchent aussi sa polymérisation dans son état ATP.
CapZ, constituée d'un dimère de deux sous-unités (alpha, 34 kDa, et beta, 30 kDa),
se fixe au pôle plus, évitant ainsi la croissance rapide.
La tropomoduline (40 kDa) se fixe au pôle moins, évitant ainsi la croissance
lente.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

CapZ et tropomoduline, jouent un rôle important dans la stabilisation
des polymères d'actine- α dans les muscles striés
la gelsoline (82 kDa), en présence d'une concentration élevée de
Ca2+ cytosolique, se fixe au polymère d'actine et crée une coupure
engendrant la dislocation du filament d'actine.
La gelsoline reste fixée à l'extrémité plus, évitant ainsi la
repolymérisation rapide
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
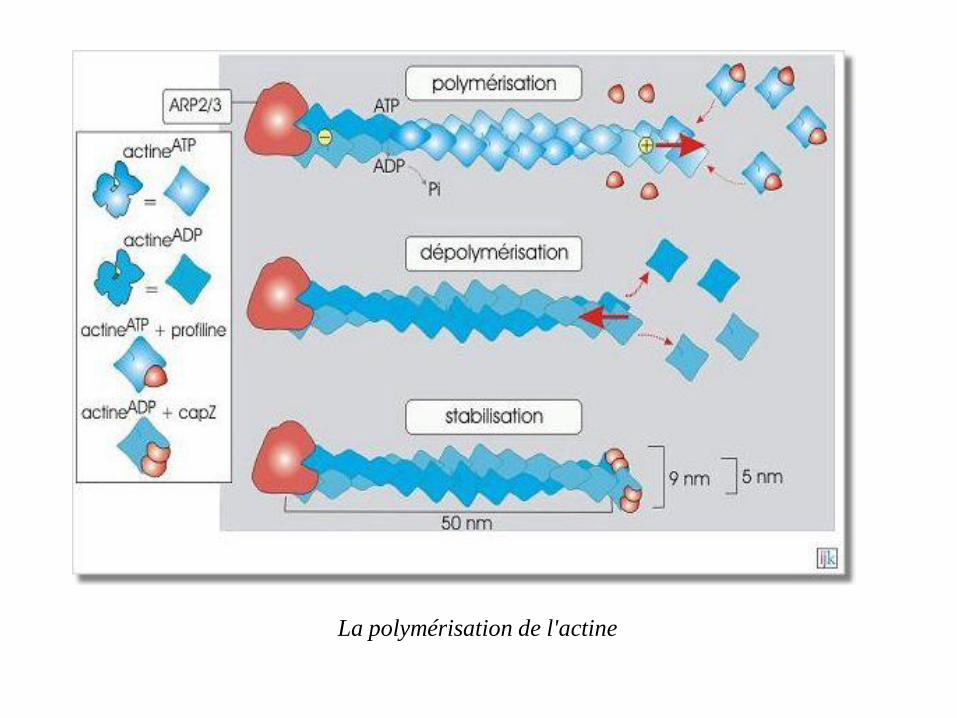
La polymérisation de l'actine
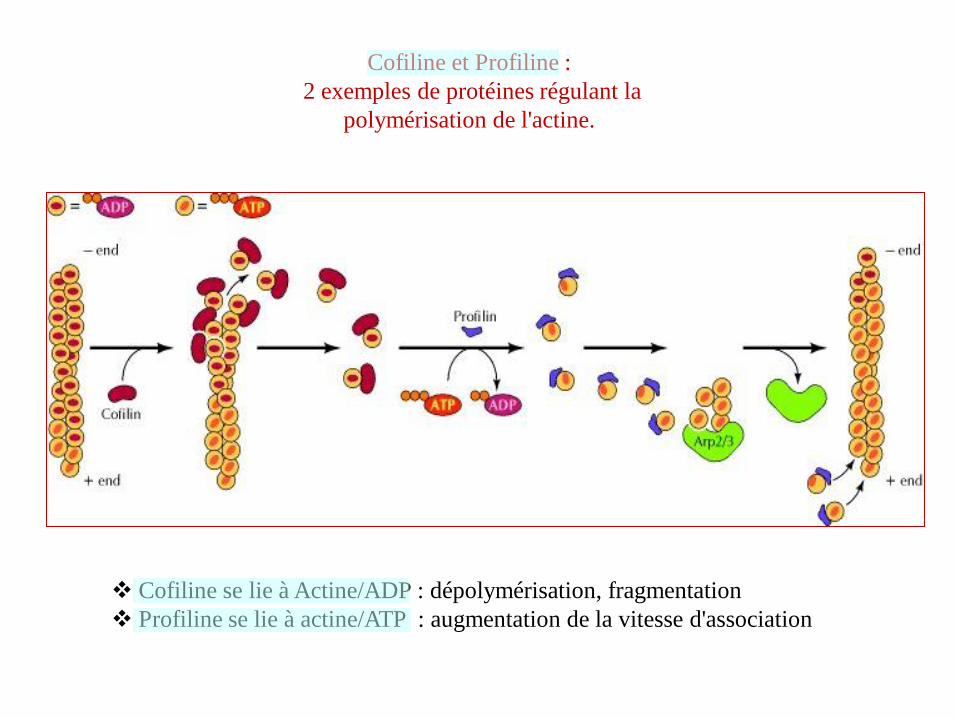
Cofiline et Profiline :
2 exemples de protéines régulant la
polymérisation de l'actine.
Cofiline se lie à Actine/ADP : dépolymérisation, fragmentation
Profiline se lie à actine/ATP : augmentation de la vitesse d'association
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
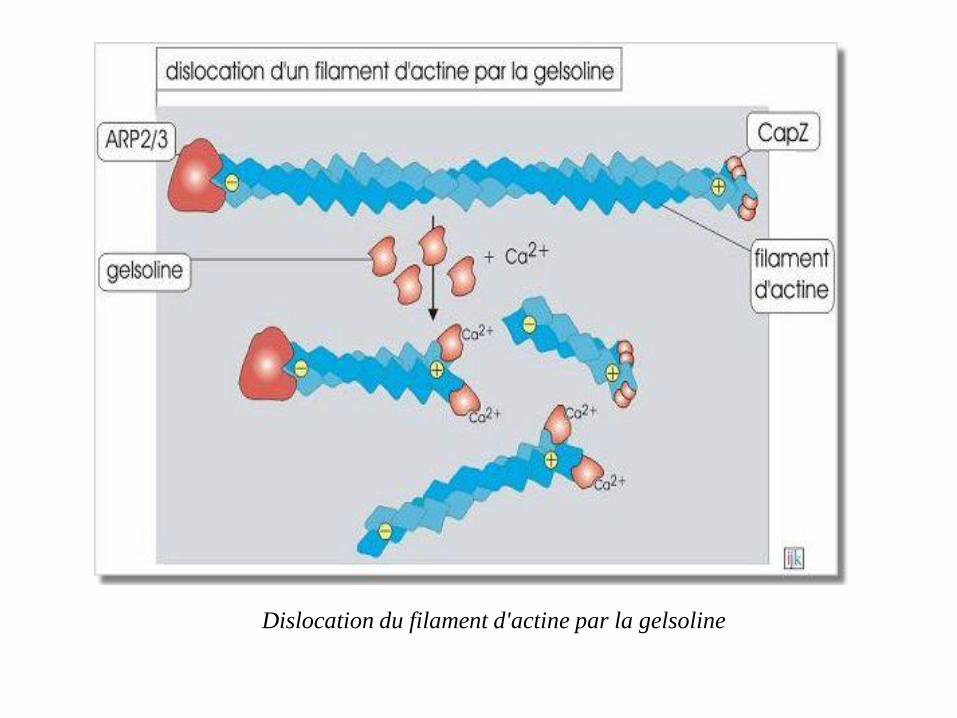
Dislocation du filament d'actine par la gelsoline

Les fonctions des filaments d'actine
Par des exemples spécifiques nous illustrons ci-dessous quelques fonctions des filaments d'actine.
Migration cellulaire
les leucocytes quittent la circulation pour s'infiltrer dans les tissus.
Les mouvements nécessaires à ce déplacement se font grâce au cytosquelette et
à l'actine en particulier.
L'actine joue un rôle dans la formation des lamellipodes résultant d'un phénomène
de protrusion membranaire
Le réseau d'actine périphérique sous-membranaire sert d'appui à la
polymérisation de nouveaux filaments qui repoussent la membrane, formant
ainsi progressivement le lamellipode.
Les sites d'initiation de la polymérisation (sites de nucléation) sont désignés
par l'activation de ARP2/3 qui pour sa part est sous l'influence des
récepteurs membranaires aux peptides N-formylés (chémokine).
Les lamellipodes sont des extensions dynamiques des leucocytes qui leur permettent de se déplacer sur une surface. Ils se forment (et disparaissent) en quelques secondes, témoignant de la dynamique rapide de la polymérisation et
dépolymérisation de l'actine.
tokyo
Highlight
tokyo
Highlight

Les faisceaux contractiles d'actine forment des fibres dites « de tension » dans les fibroblastes tissulaires (tissu conjonctif) les rendant capables de se contracter et d'exercer ainsi une traction sur la matrice extracellulaire qui les entoure. Ce processus est essentiel pour entamer la cicatrisation au cours de laquelle les deux lèvres de la blessure doivent progressivement être rapprochées. Par l'intermédiaire de complexes moléculaires d'adhérence regroupés aux sites appelés contacts focaux, les filaments d'actine sont reliés à la matrice extracellulaire (fibronectine, laminine et collagène). La molécule principalement impliquée est l'intégrine qui, grâce à un
complexe de molécules de liaison (taline, vinculine et α -actinine)
est fixée au cytosquelette d'actine
Traction sur la matrice extracellulaire
tokyo
Highlight
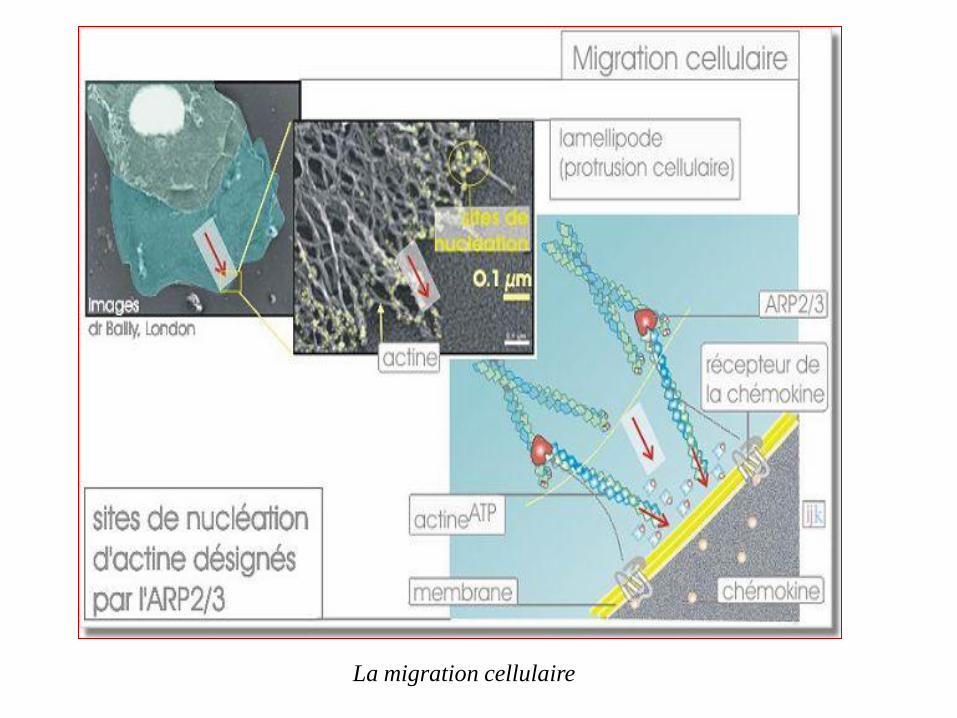
La migration cellulaire
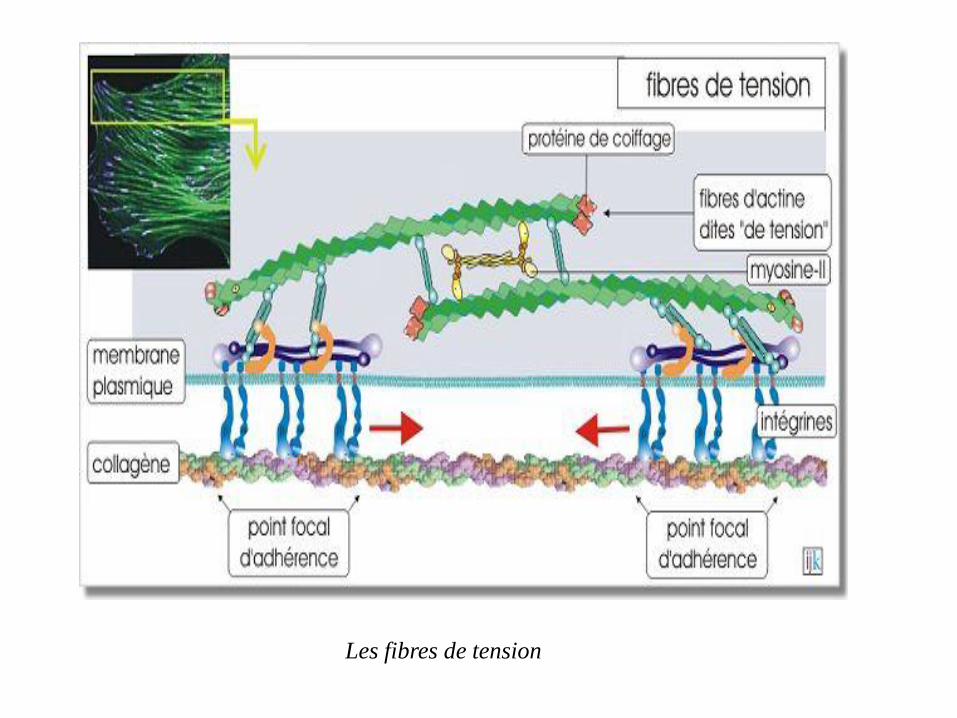
Les fibres de tension

En fin de mitose, après que les chromosomes se soient séparés grâce aux
microtubules (télophase), les filaments d'actine forment en périphérie de la
cellule et perpendiculairement à l'axe du fuseau mitotique (microtubules),
un faisceau contractile appelé anneau contractile. Quand l'anneau se
contracte (comme le cordon d'une bourse) il sépare la cellule mère en deux
cellules filles (cytodiérèse).
Cytodiérèse
Maintien de l'intégrité tissulaire et participation aux mouvements des feuillets
embryonnaires
les filaments d'actine sont un composant important de la ceinture d'adhérence.
Ces filaments sont arrangés sous forme de faisceaux contractiles. En associant
les éléments du cytosquelette d'une cellule à ceux d'une autre, cette ceinture
permet à l'épithélium de résister aux agressions mécaniques
tokyo
Highlight
tokyo
Highlight
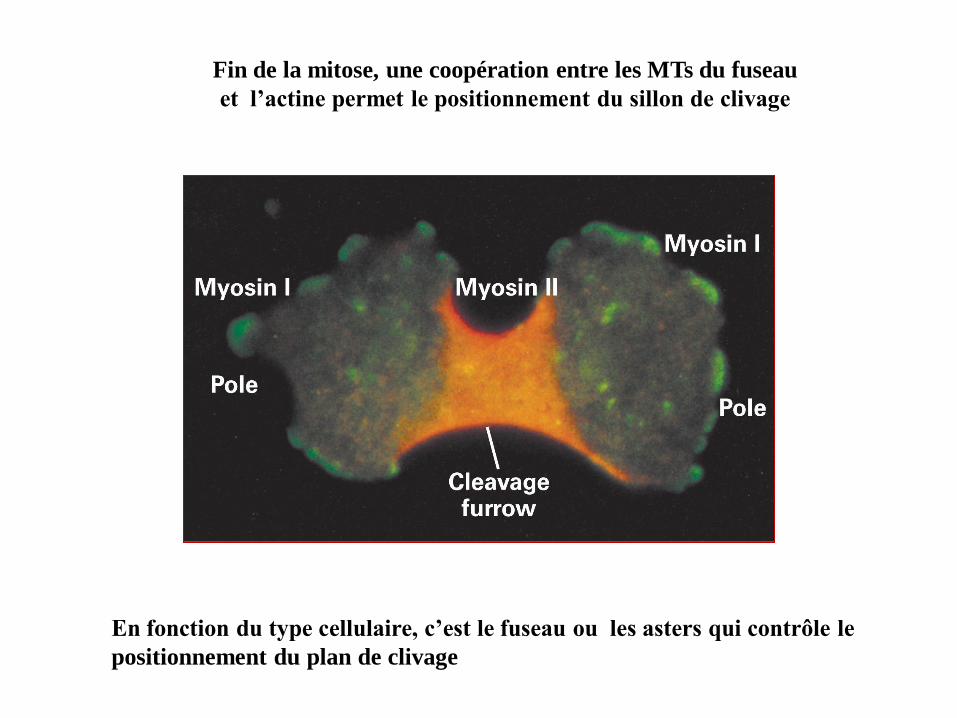
Fin de la mitose, une coopération entre les MTs du fuseau
et l’actine permet le positionnement du sillon de clivage
En fonction du type cellulaire, c’est le fuseau ou les asters qui contrôle le
positionnement du plan de clivage
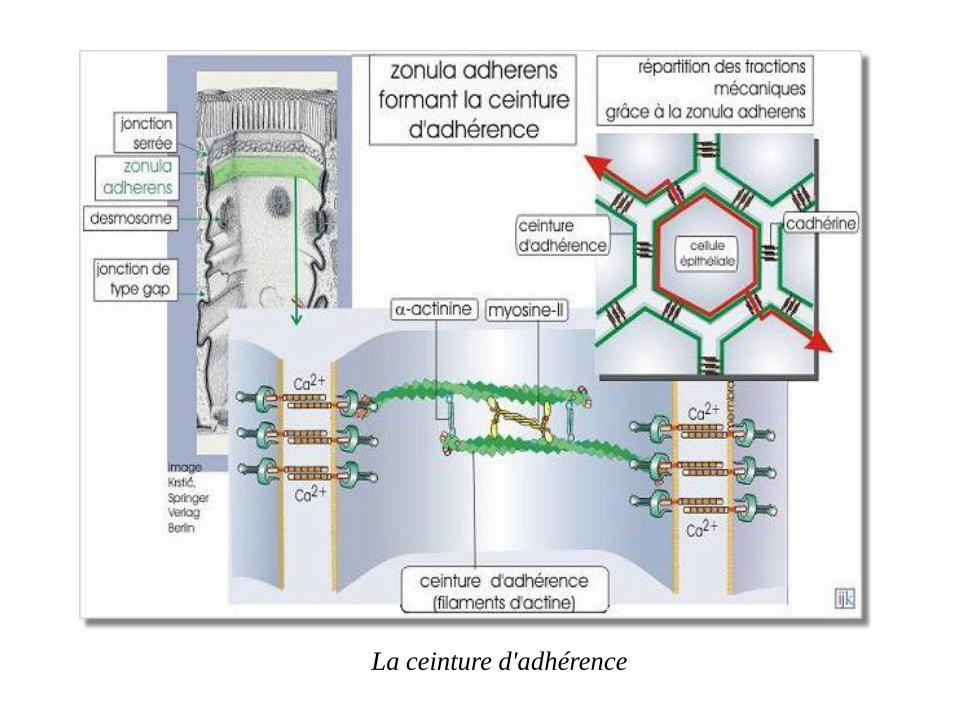
La ceinture d'adhérence

En plus de ce rôle utile de résistance tissulaire, les faisceaux
contractiles des ceintures sont à l'origine de mouvements tissulaires au
cours de l'embryogenèse. La formation du tube neural en est un
exemple représentatif. C'est en effet la contraction des ceintures qui
provoque l'affaissement du feuillet neuro-ectodermique, donnant ainsi
naissance à la gouttière neurale puis au tube neural
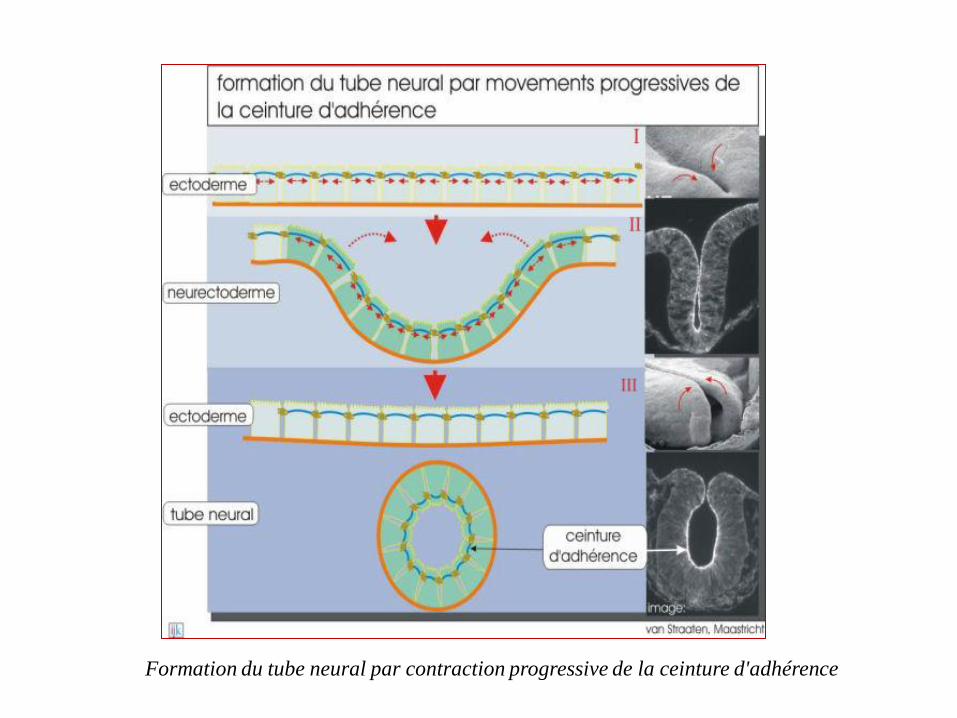
Formation du tube neural par contraction progressive de la ceinture d'adhérence

Les bordures en brosse des cellules épithéliales digestives sont formées
de microvillosités. Ces différenciations résultent en une augmentation
considérable de la surface cellulaire apicale, facilitant ainsi la capture
des nutriments dans le tube digestif (pour exemple, revoir le transport du
glucose par SGLT1
Armature des microvillosités
Ces microvillosités possèdent une armature constituée de filaments d'actine
associés en faisceaux parallèles, orientés côté + distal et liés par la fimbrine.
Ces filaments sont stabilisés par des protéines de coiffage qui se trouvent à
leurs extrémités. Les filaments sont ancrés sur un réseau de filaments
périphériques sous-membranaires (actine corticale)
tokyo
Highlight
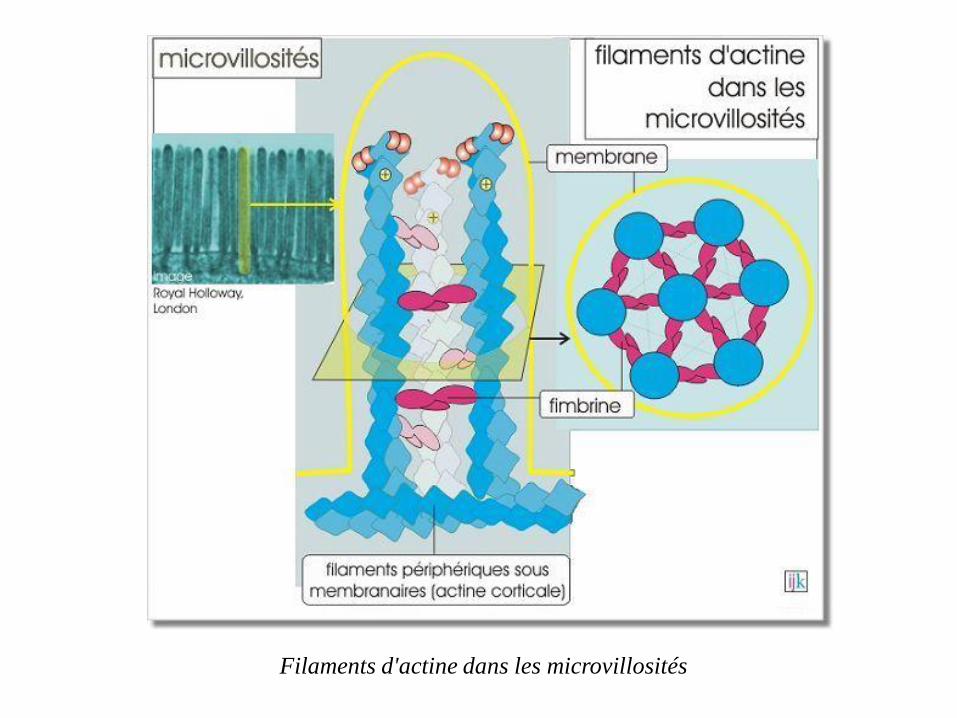
Filaments d'actine dans les microvillosités

L'actine dans les cellules musculaires
Les cellules musculaires sont des cellules où le cytosquelette
est très élaboré et dans lesquelles l'actine représente 20% de
la masse protéique totale
.
muscle squelettique est constitué de cellules géantes, les myocytes,
chaque cellule, le cytosquelette s'agence en de nombreuses
unités identiques appelées myofibrilles. Chaque myofibrille est
constituée par une juxtaposition linéaire de sarcomères,
mesurant environ 3µm, liés par leurs disques Z.
Des filaments intermédiaires, constitués de desmine (protéine de
53 kDa), entourent les myofibrilles au niveau des disques Z du
sarcomère.
Ils rendent les myofibrilles solidaires les unes des autres et de la membrane de
la cellule (géante) et réalisent l'alignement des sarcomères qui confère aux
muscles squelettique son caractéristique aspect strié en microscopie optique
tokyo
Highlight
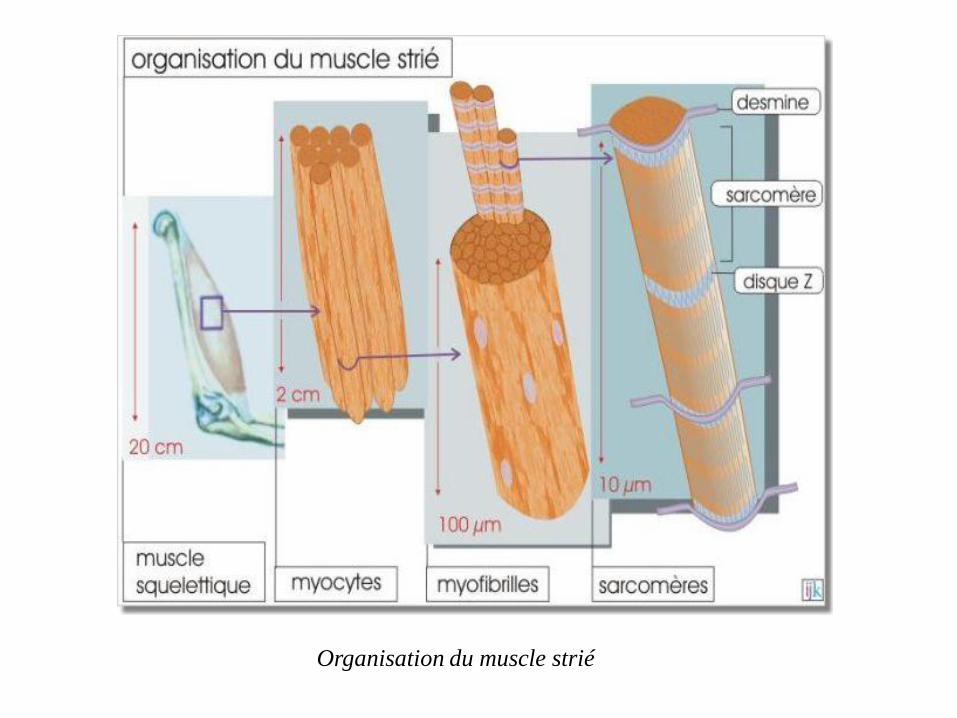
Organisation du muscle strié

Le sarcomère comme unité de contraction
L'actine et la myosine sont à la base de la contractilité des
sarcomères qui sont constitués par un assemblage de filaments
parallèles d'actine (filaments minces) et de myosine-II (filaments
épais)
Les filaments d'actine, longs d'environ 1µm sont attachés aux
disques Z par l'intermédiaire de capZ (protéine de coiffage qui se
fixe à l'extrémité plus) et de l' –α actinine.
L'extrémité moins (libre) est stabilisée par la tropomoduline. Sur sa
longueur, le filament d'actine est associé à d'autres protéines qui
interviennent dans la contraction musculaire
tokyo
Highlight
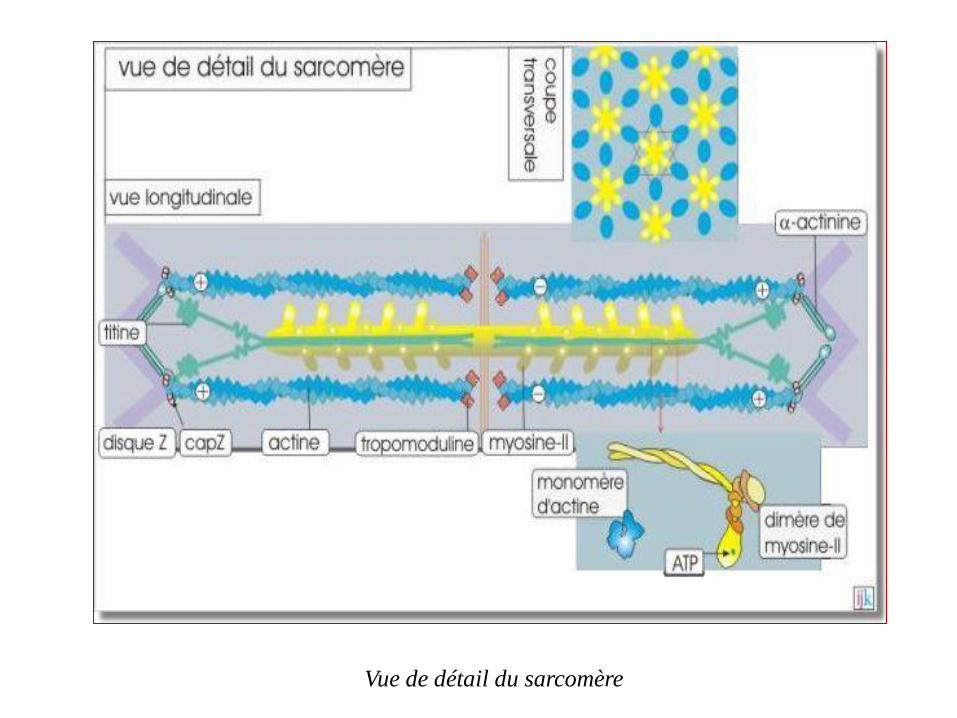
Vue de détail du sarcomère
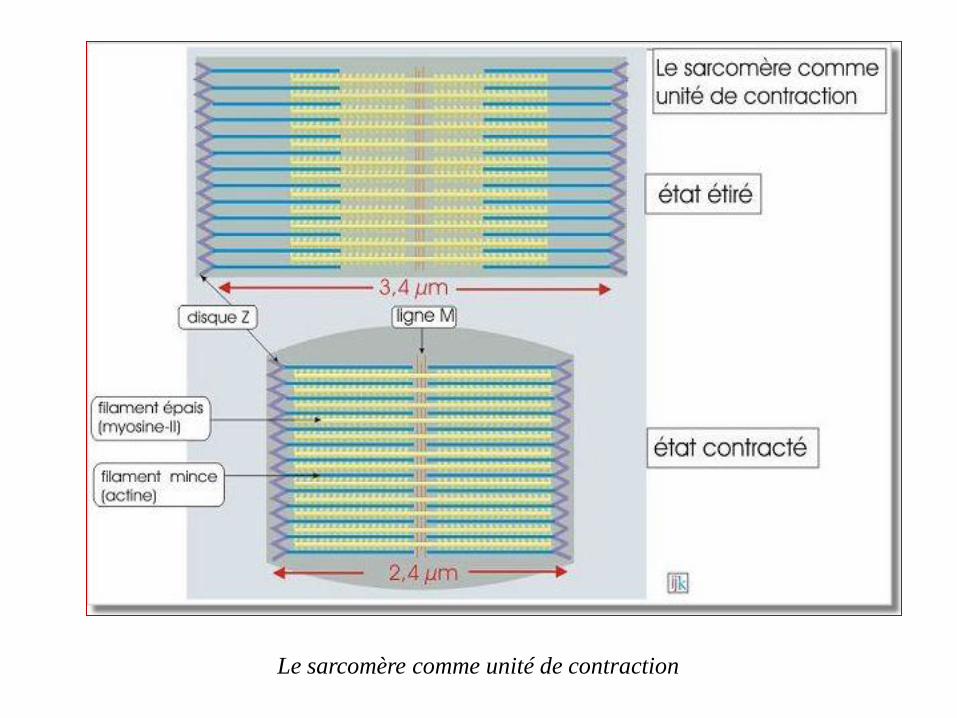
Le sarcomère comme unité de contraction
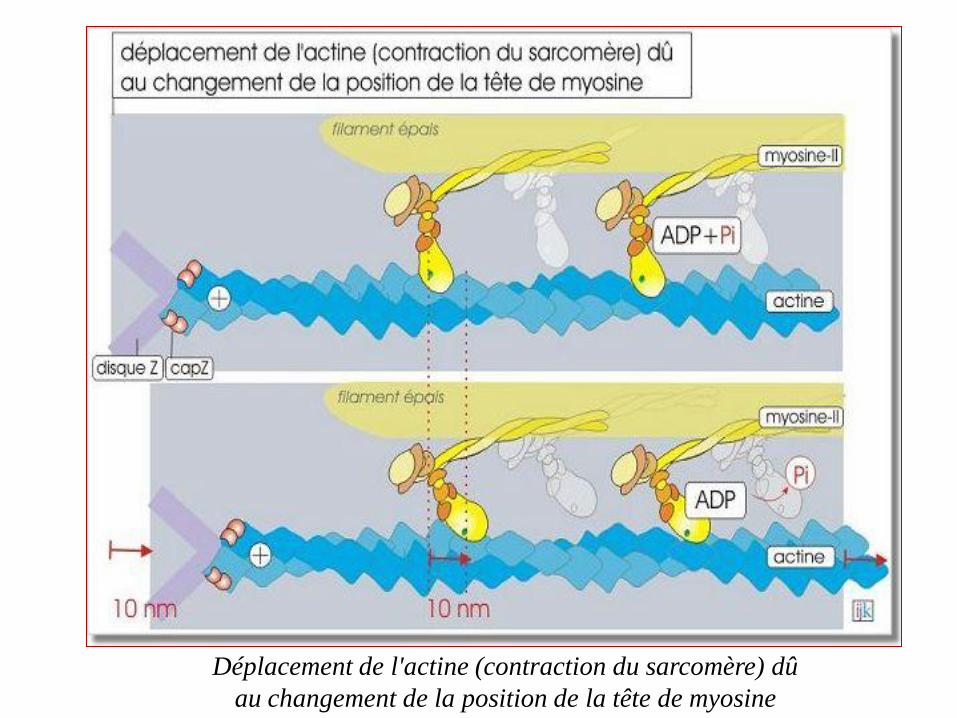
Déplacement de l'actine (contraction du sarcomère) dû
au changement de la position de la tête de myosine
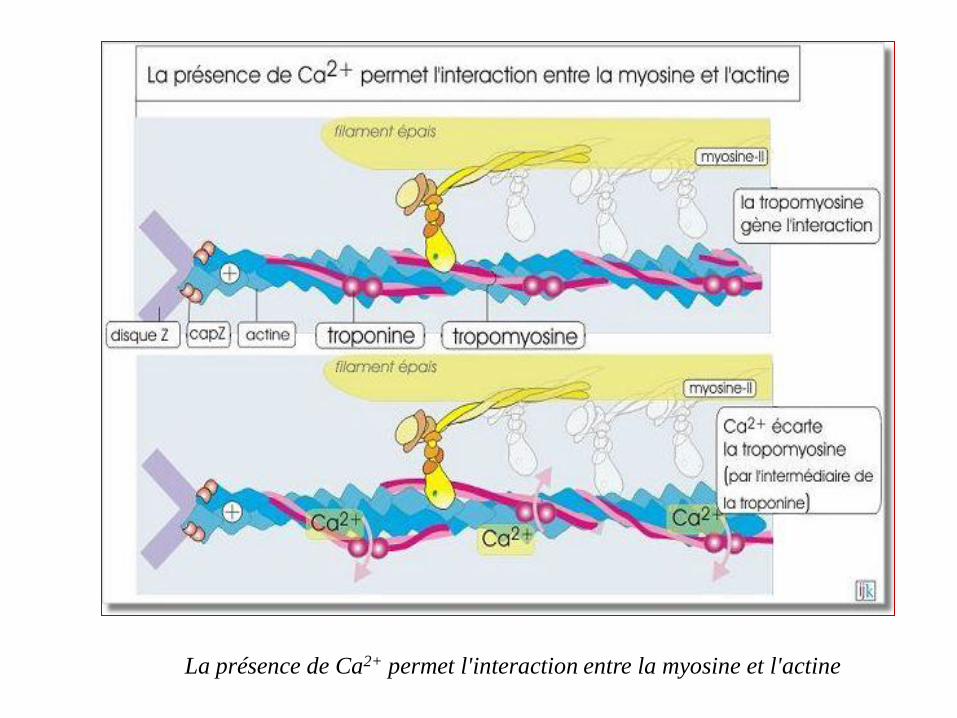
La présence de Ca2+ permet l'interaction entre la myosine et l'actine

Protéines associées
Les protéines associées à l'actine (en anglais, Actin associated proteins, ou AAP)
sont la clef du contrôle par la cellule de son stock d'actine
Elles permettent de réguler la polymérisation
d'organiser spatialement les filaments.
Elles sont à leur tour contrôlées par des protéines régulatrices qui s'insèrent dans
le réseau complexe interagissant avec toute la cellule.
Spectrine : permet l'accrochage du microfilament d'actine a la membrane plasmique
Protéines de réticulation
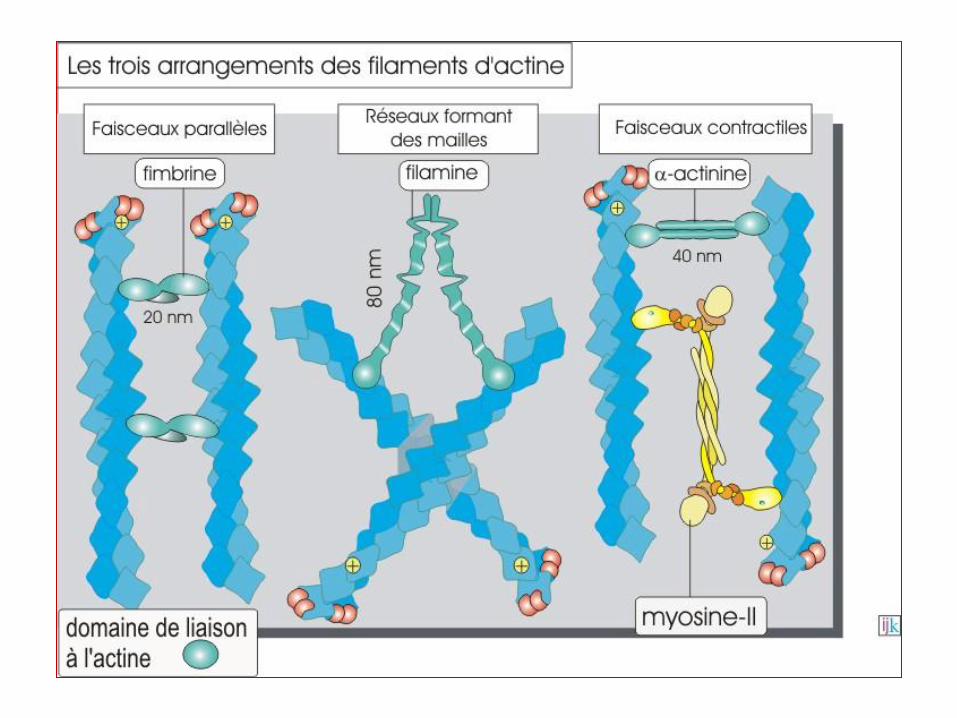
Fimbrine : maintient les filaments serrés d'actine fasciculée.
alpha actinine : protéine dimérique liant 2 microfilaments entre eux, les
maintenant parallèles entre eux. (point focal d'ancrage...).
Elle se lie au pôle + pour permettre l'intéraction actine-myosine notamment dans
le mouvement amiboïde.
Filamine : bloque l'actine réticulée pour l'empêcher de passer en actine fasciculée.
Villine : maintient les filaments serrés d'actine fasciculée, spécifique des microvillosités.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Tropomyosine
Protéines de stabilisation
Ce sont ces protéines qui permettent d'initier la polymérisation. Il en existe
plusieurs, ayant des modes d'actions différentes.
Protéines de polymérisation
Protéines de nucléation
L'ARP2/3, une protéine proche de l'actine, sert de point de départ à la
polymérisation : des monomères d'actine G viennent se lier à elle, entraînant
la formation d'un filament.
La formine possède deux domaines riches en proline. Des profilines liées
chacune à une actine G viennent se lier sur le premier ; ces actines sont alors
transférées sur l'autre domaine, où elle se lient les unes à la suite des autres
pour former le filament.
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

La profiline se fixe à l'actine G et favorise l'échange de l'ADP par de L'ATP, et
laisse le côté - de l'actine libre ; il peut ainsi s'insérer dans le polymère à
l'extrémité barbue.
Protéines de séquestration
Ces protéines servent à réguler la quantité d'actine G présente dans le cytoplasme.
La thymosine se fixe à l'extrémité pointue de l'actine G et empêche l'échange
de l'ADP liée à celle-ci par une molécule d'ATP.
Protéines de coiffe
Elles se lient à l'extrémité barbée d'un filament d'actine pour empêcher la
polymérisation (ajout d'actine G), et donc pour arrêter la croissance du filament
Elles peuvent également se lier à l'extrémité pointue pour arrêter la
dépolymérisation ou permettre la formation d'un nucléus (initiation de la
polymérisation).
CAP (capping protein).
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

La gelsoline, comme son nom l'indique, permet de passer de l'actine en gel
(dans un réseau de maille) à l'actine en solution (actine G).
La cofiline impose une torsion supplémentaire à l'actine, entraînant sa
dépolymérisation.
Protéines de fragmentation
Ces protéines permettent de cliver les filaments d'actine,
Myosine de type 1 : maintient l'actine large fasciculée avec l'aide de l'alpha-actinine.
Myosine de type 2 : contraction musculaire.
Myosine de type 5 : protéine moteur capable de se déplacer vers l'extrémité +.
Protéines motrices
Les filaments d'actine s'organisent de façons très diverses suivant leurs fonctions
biologiques. Voici quelques exemples de structures observées et leurs principales
propriétés : faisceaux, câbles de tension, réseaux, comètes.
Superstructures
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight

Le mouvement des organites : chez les végétaux supérieur le déplacement des
chloroplastes dans la cellule en fonction des conditions lumineuse fait intervenir les
filaments d'actine.
Rôles
Maintien des édifices structuraux par exemple des villosités.
Mouvements intracellulaires : contraction des cellules musculaires striées:
ces cellules musculaires sont organisées en myofibrilles (= disposition
particulière des filaments d'actines et de myosines et d'autres protéines).
L'unité fonctionnelle de la myofibrille est le sarcomère qui va se raccourcir
lors de la contraction (isotonique ou isométrique). /contraction des cellules
musculaires lisses: pas d'organisation sarcomérique, arrangement particulier
des filaments qui parcourent la cellule transversalement, longitudinalement et
en diagonale, pour s'insérer sur les corps denses.
Les mouvements cellulaires : déplacement de l'amibe, des leucocytes
(pseudopodes).
La cytodiérèse (en fin de division cellulaire).
La formation des jonctions cellulaires (y compris des neurones, via le « cône
de croissance »).
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight
tokyo
Highlight