
BANG - Paris - Inria · processus de croissance en biologie et en m edecine : division cellulaire,...
29
´ Equipe-projet INRIA BANG Biologie, Analyse Num´ erique, G´ eophysique Chef de projet : Benoˆ ıt Perthame (UPMC) Pr´ esentation : Jean Clairambault (INRIA)
Transcript of BANG - Paris - Inria · processus de croissance en biologie et en m edecine : division cellulaire,...

Equipe-projet INRIA BANG
Biologie, Analyse Numerique,Geophysique
Chef de projet : Benoıt Perthame (UPMC)
Presentation : Jean Clairambault (INRIA)

Equipe creee en 2004, alors mixte INRIA-ENS,bientot mixte INRIA-UPMC
INRIA-permanents :Marie-Odile Bristeau (DR2), Jean Clairambault (DR2), Dirk Drasdo(DR2), Americo Marrocco (responsable permanent de l’EPI, DR1)INRIA-detachee (Corps des Ponts) : Marie Doumic Jauffret (CR1)INRIA-Postdocs : Frederique Charles, Marica Pelanti, Ignacio RamisConde, Min TangINRIA-doctorants :Annabelle Ballesta, Chadha Chettaoui, Luna Dimitrio, PierreGabriel, Nick Jagiella, Thomas Lepoutre, William WeensPXIII-collaborateur : Emmanuel Audusse (MdC)UPMC-collaborateurs : Benoıt Perthame (responsable scientifique del’EPI, PR0, IUF), Nicolas Vauchelet (MdC)

Axe principal : Phenomenes de croissance en biologie
1. Modelisation par EDP/EDO et analyse mathematique de
processus de croissance en biologie et en medecine :
division cellulaire, chimiotactisme, pharmacologie moleculaire, optimisation
therapeutique, agregation et fragmentation de proteines (prion, Alzheimer)
2. Modeles individu-centres (‘IBM’) de la croissance tissulaire,
saine et tumorale : regeneration hepatique, croissance de tumeurs solides
3. Ecoulements a surface libre, fluides stratifies, couplage
hydro-biologie (EDP)
(collaboration avec J. Sainte-Marie, Lab. Saint-Venant & EPI Macs)
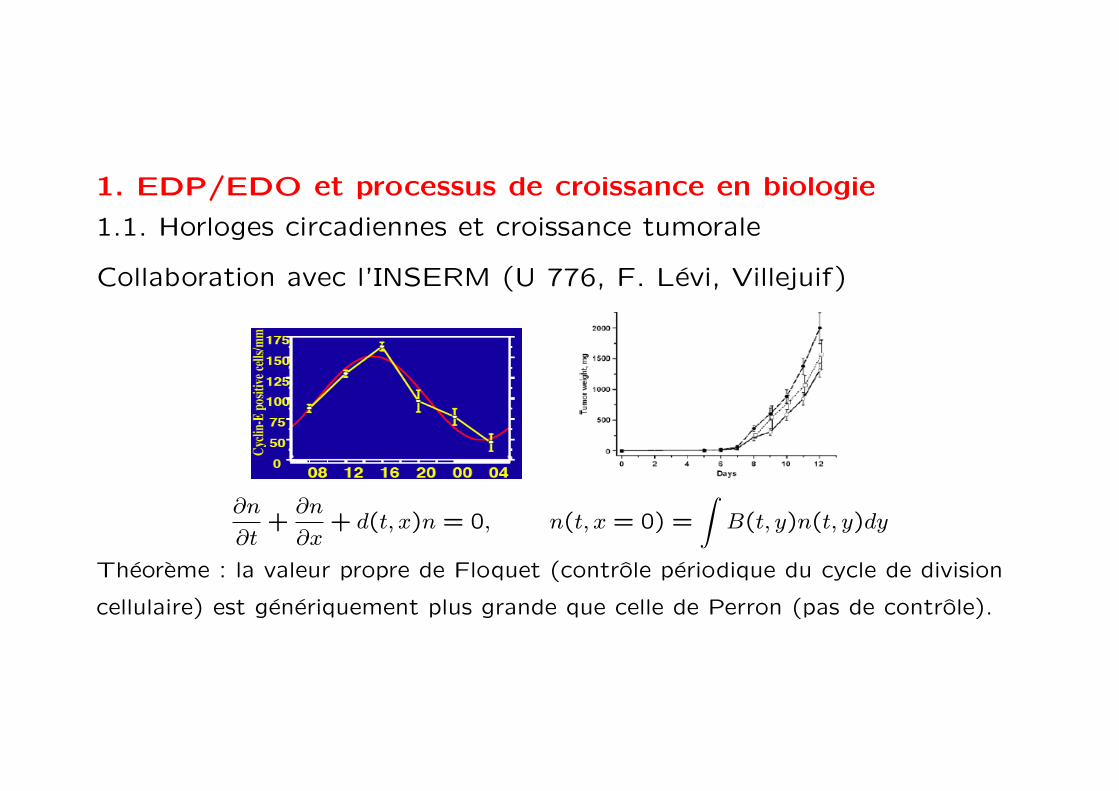
1. EDP/EDO et processus de croissance en biologie
1.1. Horloges circadiennes et croissance tumorale
Collaboration avec l’INSERM (U 776, F. Levi, Villejuif)
∂n
∂t+∂n
∂x+ d(t, x)n = 0, n(t, x = 0) =
∫B(t, y)n(t, y)dy
Theoreme : la valeur propre de Floquet (controle periodique du cycle de division
cellulaire) est generiquement plus grande que celle de Perron (pas de controle).
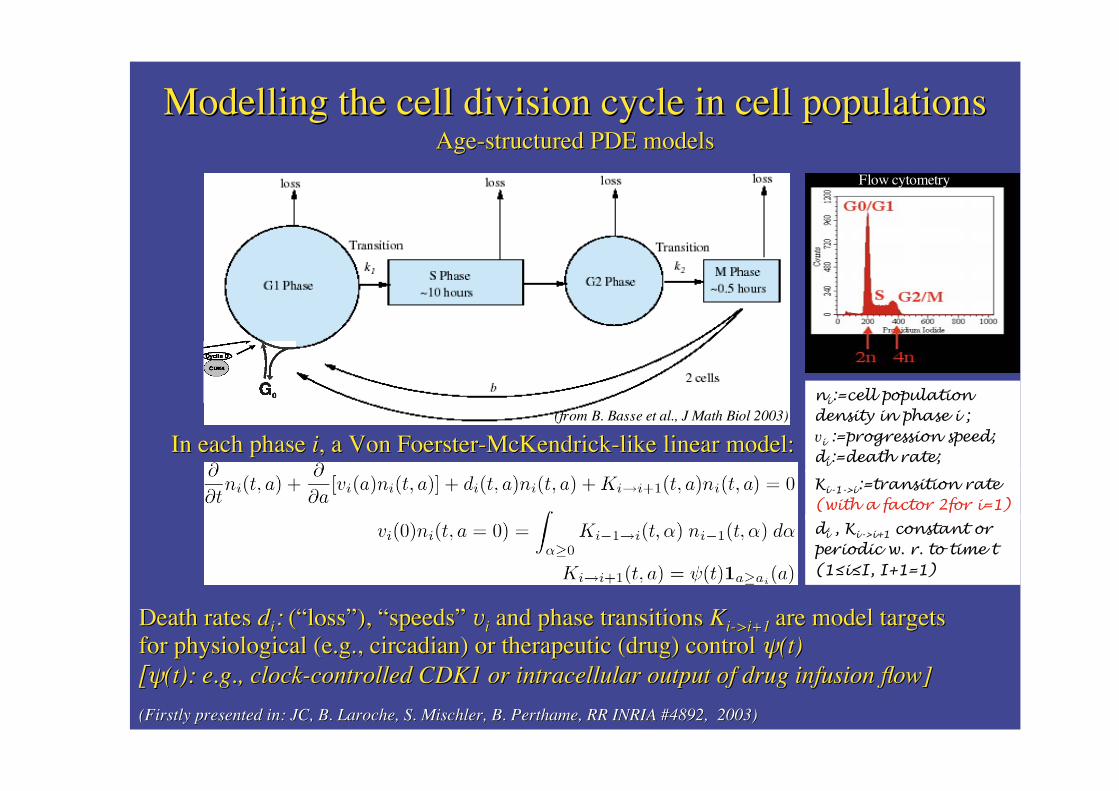
Modelling the cell division cycle in cell populationsModelling the cell division cycle in cell populationsAge-structured PDE modelsAge-structured PDE models
(from B. Basse et al., J Math Biol 2003)
In each phase In each phase ii, a Von , a Von Foerster-McKendrick-like Foerster-McKendrick-like linear model:linear model:
di , Ki->i+1 constant orperiodic w. r. to time t(1�i�I, I+1=1)
ni:=cell populationdensity in phase i ;vi :=progression speed;di:=death rate;
Ki-1->i:=transition rate(with a factor 2for i=1)
Death raDeath rattes es ddii:: ( (““lossloss””), ), ““speedsspeeds”” vvii and phase transitions and phase transitions KKi-i->i+1>i+1 are model targetsare model targetsfor physiological (e.g., circadian) or therapeutic (drug) control for physiological (e.g., circadian) or therapeutic (drug) control ��(t)(t)[�[�(t): e.g., clock-controlled CDK1 or intracellular output of drug infusion flow](t): e.g., clock-controlled CDK1 or intracellular output of drug infusion flow]
(Firstly presented in: JC, B. (Firstly presented in: JC, B. LarocheLaroche, S. , S. MischlerMischler, B. , B. PerthamePerthame, RR INRIA #4892, 2003), RR INRIA #4892, 2003)
Flow cytometry

The simplest case: 1-phase model with divisionThe simplest case: 1-phase model with division
(Here, (Here, vv(a)(a)=1=1, , a* a* is the cell cycle duration, and is the cell cycle duration, and ��(<1)(<1) is the timeis the time
during whichduring which the 1-the 1-periodic controlperiodic control � � isis actually exerted on cell division) actually exerted on cell division)
Then it can be shown that the Then it can be shown that the eigenvalue eigenvalue problem:problem:
has a unique positive 1 has a unique positive 1-periodic-periodic eigenvector eigenvector NN, with a positive , with a positive eigenvalue eigenvalue ��
andand an explicit an explicit formula can be found for formula can be found for �� when when KK00 ++ (T. (T. LepoutreLepoutre’’s s PhD work)PhD work)
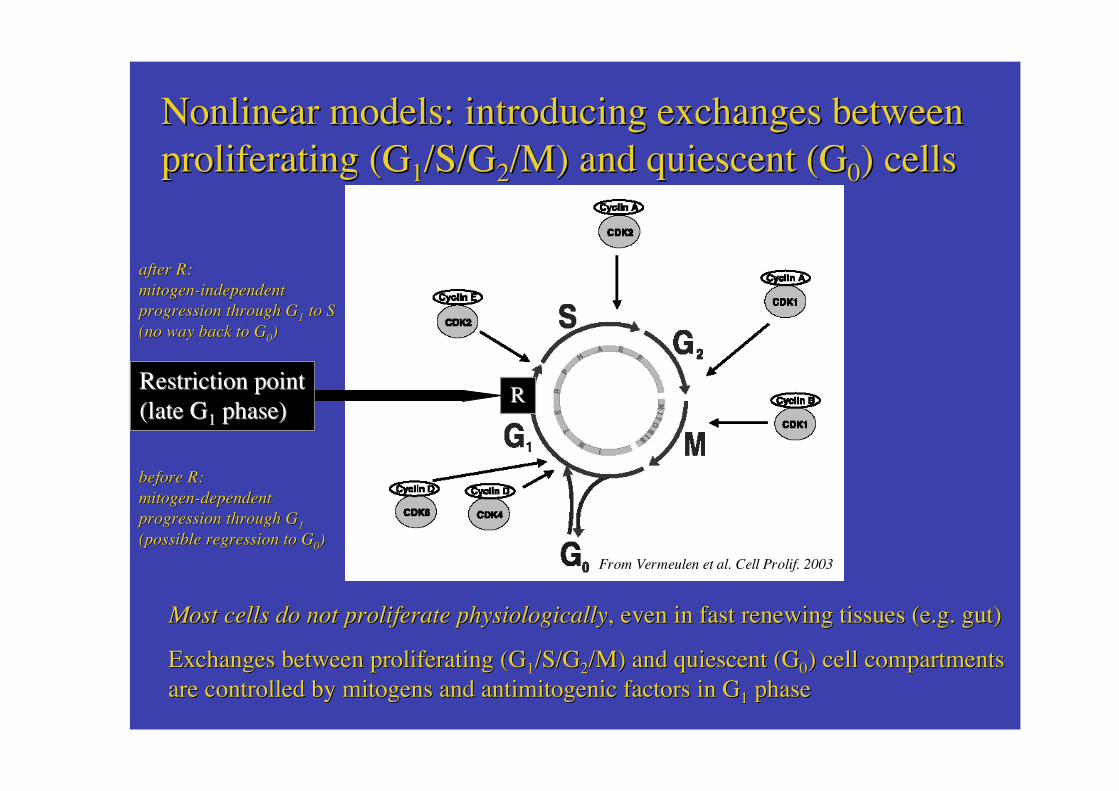
Exchanges between proliferating Exchanges between proliferating (G(G11/S/G/S/G22/M) /M) and and quiescent (Gquiescent (G00) ) cell compartmentscell compartmentsare are controlled controlled by by mitogens and antimitogenic factors mitogens and antimitogenic factors in Gin G11 phase phase
From Vermeulen From Vermeulen et al.et al. Cell Prolif Cell Prolif. 2003. 2003
Most Most cells cells do not do not proliferate physiologicallyproliferate physiologically, , even even in in fast renewing fast renewing tissues (e.g. tissues (e.g. gutgut))
Nonlinear Nonlinear models: models: introducing exchanges betweenintroducing exchanges betweenproliferating proliferating (G(G11/S/G/S/G22/M) /M) and and quiescent (Gquiescent (G00) ) cellscells
RRRestriction pointRestriction point((late late GG11 phase) phase)
before before R:R:mitogenmitogen--dependentdependentprogressionprogression through through GG11
(possible(possible regression regression to Gto G00))
after after R:R:mitogen-independentmitogen-independentprogressionprogression through through GG11 to S to S(no(no way way back to Gback to G00))

ODE models ODE models with two exchanging cell compartmentswith two exchanging cell compartments, , proliferating proliferating (P) (P) and and quiescent (Q)quiescent (Q)
((Gyllenberg Gyllenberg & Webb, & Webb, GrowthGrowth, , DevDev. & . & Aging Aging 1989; 1989; Kozusko Kozusko & & BajzerBajzer, Math , Math BioSci BioSci 2003)2003)
where, for instance:where, for instance:rr0 0 representing here the rate ofrepresenting here the rate ofinactivation of proliferating cells,inactivation of proliferating cells,and and rr
ii the rate of recruitment fromthe rate of recruitment fromquiescence to proliferationquiescence to proliferation
Initial goal: to justify Initial goal: to justify Gompertz Gompertz growthgrowth(a popular model among radiologists)(a popular model among radiologists)
Cell exchangesCell exchanges

Simple PDE models, Simple PDE models, age-structured withage-structured withexchanges between proliferation and exchanges between proliferation and quiescencequiescence
pp=density =density of proliferating cells; of proliferating cells; qq=density =density of quiescent cells; of quiescent cells; ��,,��=death terms;=death terms;KK=term describing cells leaving proliferation to quiescence, due to mitosis;=term describing cells leaving proliferation to quiescence, due to mitosis;��=term describing =term describing ““reintroductionreintroduction”” (or recruitment) from quiescence to proliferation (or recruitment) from quiescence to proliferation

DelayDelay differentialdifferential models models with two cell compartmentswith two cell compartments,,proliferatingproliferating (P)/quiescent (Q): (P)/quiescent (Q): Haematopoiesis Haematopoiesis models models
((obtained from the previous obtained from the previous model model with additional hypotheses and integration with additional hypotheses and integration in x in x along characteristicsalong characteristics))
(from Mackey, Blood 1978)(from Mackey, Blood 1978)
Properties of this model: depending on the parameters, one can have positiveProperties of this model: depending on the parameters, one can have positivestability, extinction, explosion, or sustained oscillations of both populationsstability, extinction, explosion, or sustained oscillations of both populations
(Hayes stability criteria, see Hayes, J London Math Soc 1950)(Hayes stability criteria, see Hayes, J London Math Soc 1950)
Oscillatory behaviour is observed in Oscillatory behaviour is observed in periodic periodic ChronicChronic MMyelogenous yelogenous LeukaemiaLeukaemia((CMLCML) where oscillations with limited amplitude are compatible with survival,) where oscillations with limited amplitude are compatible with survival,whereas explosion (blast crisis,whereas explosion (blast crisis, alias alias acutisationacutisation) leads to ) leads to AMLAML and death and death (Mackey and (Mackey and Bélair Bélair in Montréal; in Montréal; AdimyAdimy, Bernard, , Bernard, CrausteCrauste, , Pujo-MenjouetPujo-Menjouet, , Volpert Volpert in Lyonin Lyon))
(delay (delay �� = = cell cell division cycle time)division cycle time)
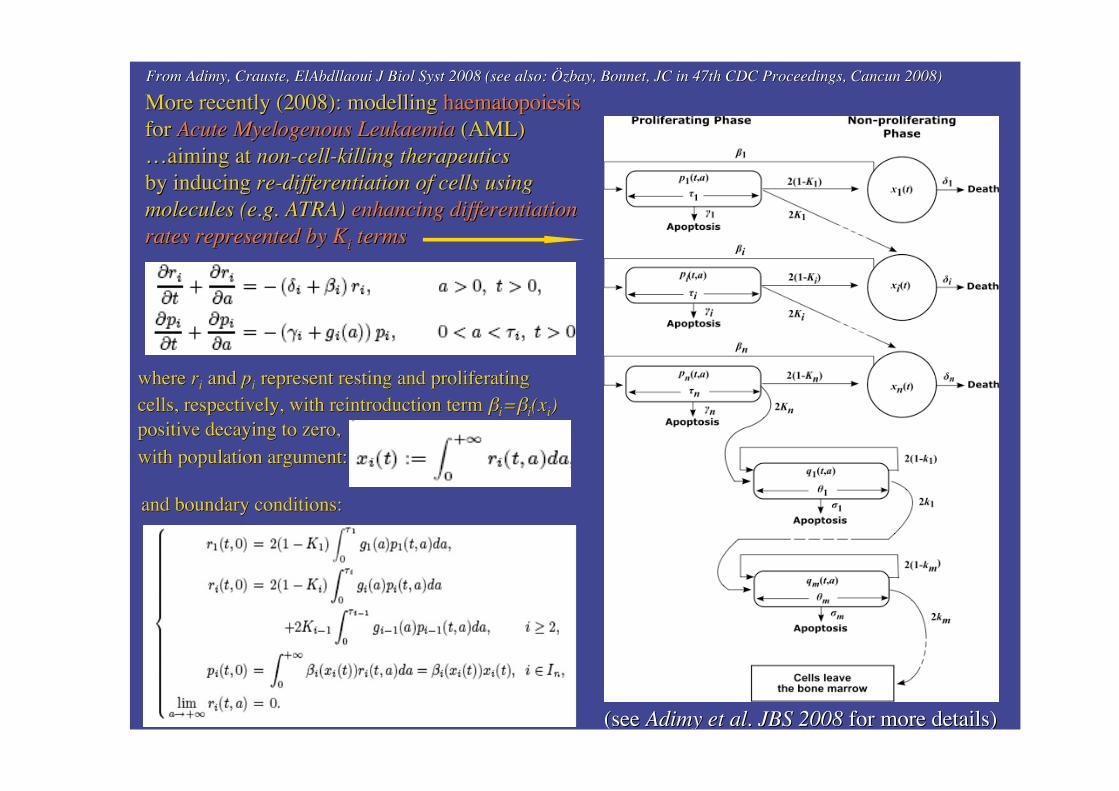
More recently (2008): modelling More recently (2008): modelling haematopoiesishaematopoiesisforfor Acute Acute Myelogenous Myelogenous LeukaemiaLeukaemia (AML) (AML)……aimingaiming at at non-cell-killing therapeuticsnon-cell-killing therapeuticsby inducingby inducing re-differentiation of cells usingre-differentiation of cells usingmolecules (e.g. ATRA)molecules (e.g. ATRA) enhancing differentiationenhancing differentiationrates represented by rates represented by KKii termsterms
(see (see Adimy Adimy et al. JBS 2008et al. JBS 2008 for more details) for more details)
where where rrii and and ppii repr represent esent resting and proliferatingresting and proliferatingcells, recells, respespectively, with reintroduction term ctively, with reintroduction term ��ii==��ii(x(xii))positive decaying to zero,positive decaying to zero,
with population argument:with population argument:
and boundary conditions:and boundary conditions:
From From AdimyAdimy, , CrausteCrauste, , ElAbdllaoui ElAbdllaoui J J Biol Syst Biol Syst 2008 (see also: 2008 (see also: ÖzbayÖzbay, Bonnet, JC in 47th CDC Proceedings, Cancun 2008), Bonnet, JC in 47th CDC Proceedings, Cancun 2008)
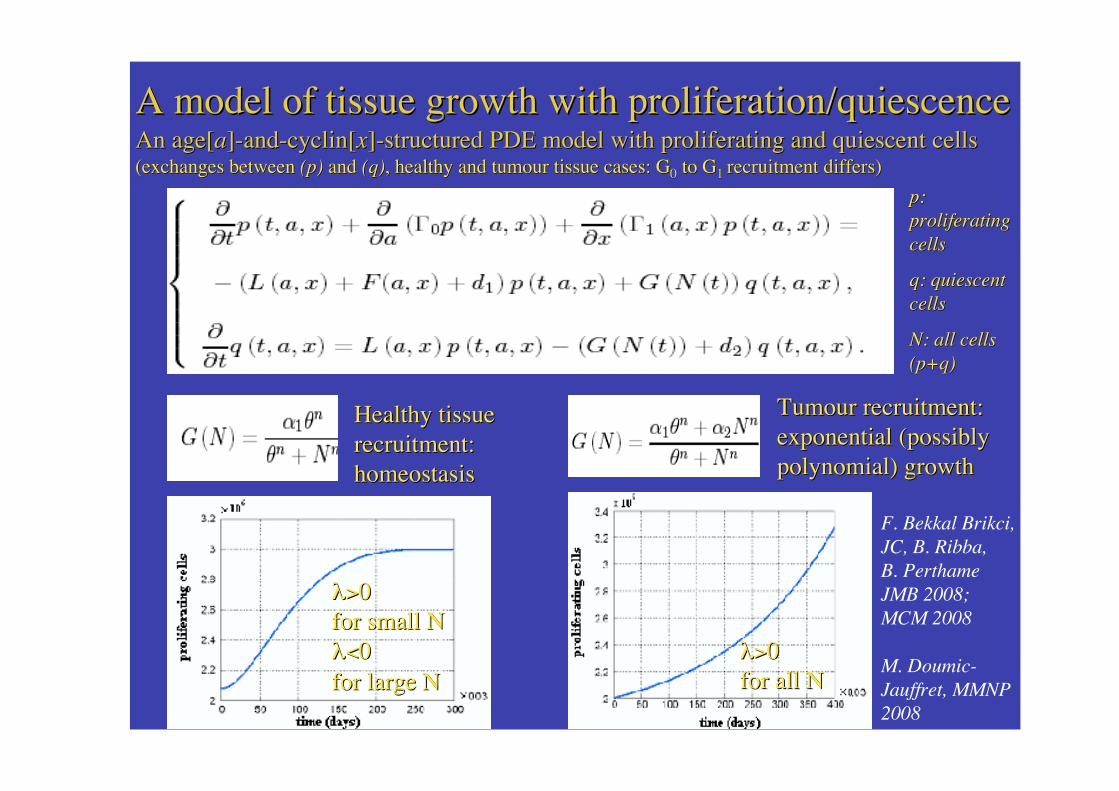
A model of tissueA model of tissue growth with proliferation/quiescencegrowth with proliferation/quiescenceAn age[An age[aa]]-and-cyclin-and-cyclin[[xx]]-structured -structured PDE model PDE model with proliferating and with proliferating and quiescent quiescent cellscells(exchanges between (exchanges between (p)(p) and and (q)(q), healthy and tumour tissue cases: G, healthy and tumour tissue cases: G00 to G to G1 1 recrurecruitmeitment differs)nt differs)
Healthy tissueHealthy tissuerecruitment:recruitment:homeostasishomeostasis
Tumour recruitment:Tumour recruitment:exponential exponential (possibly(possiblypolynomial) polynomial) growthgrowth
F. Bekkal Brikci,JC, B. Ribba,B. PerthameJMB 2008;MCM 2008
M. Doumic-Jauffret, MMNP2008
p:p:proliferatingproliferatingcellscells
q: quiescentq: quiescentcellscells
N: all cellsN: all cells(p+q)(p+q)
�>�>00for small for small ���<�<00for large for large NN
�>�>00for all for all ��

Next stepNext step:: integrating the two integrating the two models models ((Von Von Foerster-Foerster-
McKendrick-like McKendrick-like linear and nonlinear proliferation/quiescence modelslinear and nonlinear proliferation/quiescence models)) in ain acomplete cell complete cell cycle cycle model model with with phases Gphases G00-G-G11-S-G-S-G22-M-M
Keeping the same Keeping the same control control targetstargets, , adding adding control by control by growth factorsgrowth factors,,hence hence on on cyclin cyclin D, on D, on the recruitment function the recruitment function G G from from GG00 to G to G11
and and on on progression speed in progression speed in GG11
……Work Work in in progressprogress……
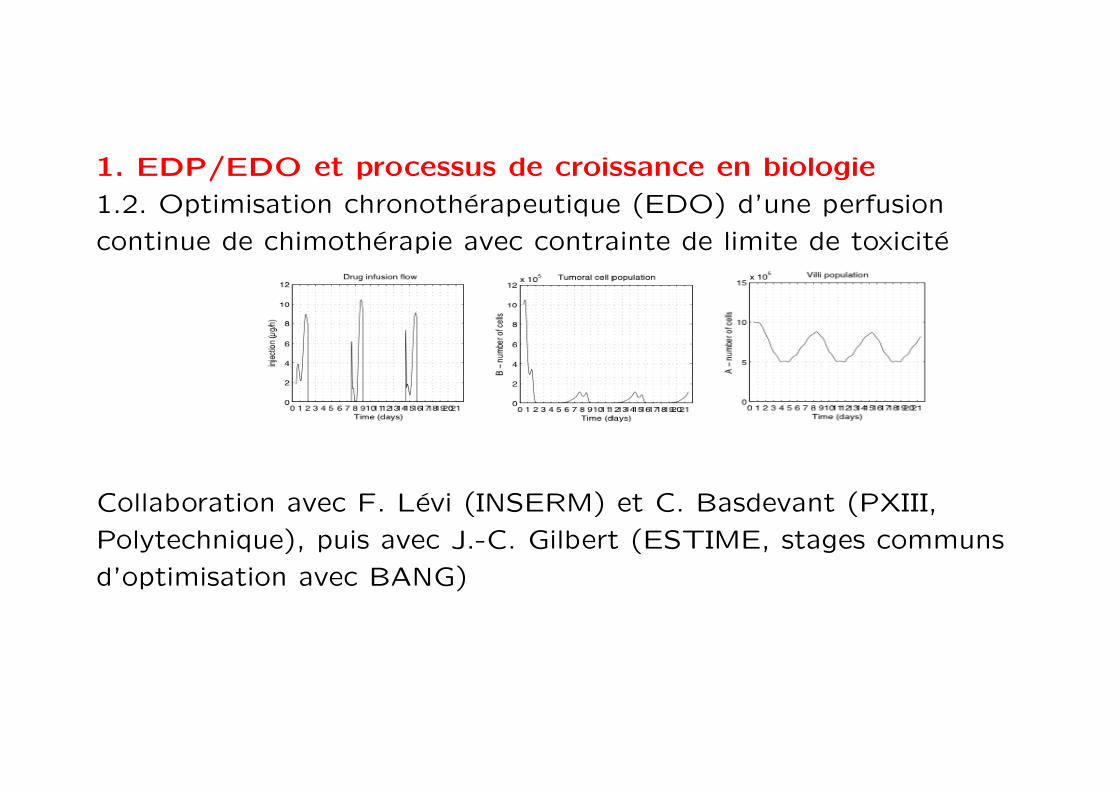
1. EDP/EDO et processus de croissance en biologie
1.2. Optimisation chronotherapeutique (EDO) d’une perfusion
continue de chimotherapie avec contrainte de limite de toxicite
Collaboration avec F. Levi (INSERM) et C. Basdevant (PXIII,
Polytechnique), puis avec J.-C. Gilbert (ESTIME, stages communs
d’optimisation avec BANG)

!"#$%&
S = Free Thymidylate Synthase (TS)
Drug output T = Blocked Thymidylate Synthase
(stable ternary FdUMP-MTHF-TS complex)!
A = ABC transporter (active drug efux)&
Input i = 5-FU infusion flow
Input j = LV infusion flow
(1)&
(2)!
(3)!
(4)!
(5)!
(6)!
(7)!
(8)!
(9)!
P&
F!
Q!
L!
N!
S!
B!
T!
A!
P = Plasma [5-FU]; F = Intracellular [FdUMP];
Q = Plasma [LV]; L =Intracellular [MTHF];
N = 5-FU-triggered Nuclear Factor; A = ABC
Transporter activity, NuclearFactor-induced;
S = Free [TS] (not FdUMP-bound);
B =[FdUMP-TS] reversible binary complex;
T =[FdUMP-TS-MTHF] stable ternary complex
i! j!
(1)! (3)!
P! Q!
(2)!
F! L!
S! B!
(7)!(9)!
(4)!
(8)!
[drug efux] N, A!
(5, 6)!
[drug inux] !
clairamb
Zone de texte
Pharmacocinétique-pharmacodynamie moléculaire

1. EDP/EDO et processus de croissance en biologie
1.3. Mouvements et auto-organisation de populations cellulaires :Qu’observe-t-on ?
En quoi est-ce utile de modeliser ces processus ?• Explication de strategies d’invasion par des organismes simples(simples par rapport aux cellules eucaryotes)• Biofilms et problemes cliniques (colonisation de catheters)• Source de defis mathematiques, numeriques et theoriques

1.3. Mouvements et auto-organisation de populations cellulaires :
Nombreux processus biologiques impliquant l’auto-oganisation de
populations de cellules : modeles EDP de Keller-Segel, Gray-Scott...
∂n
∂t−∆n+ div(n∇c) = F (n), −∆c+ αc = n.
Modeles de croissance de populations de bacteries : Mimura (g.), Gray-Scott (d.)

2. Modeles individu-centres
• Formation de structures multicellulaires (depuis l’echellemoleculaire jusqu’a l’organe).
• Analyse des donnees d’imagerie a l’interface experience / modele
Liver
regeneration
Tumor
growth

3. Ecoulements geophysiques a surface libre (EDP)
Effet du vent sur un lac stratifie par la temperature - Upwelling
• Modelisation de Saint-Venant (shallow water) a Navier-Stokesnon-hydrostatique• Vers le Navier-Stokes 3D avec maillage 2D
• Methodes numeriques (Systemes hyperboliques, schemas bases surune interpretation cinetique, schemas “well-balanced”)

Principaux resultats :
1. EDP/ EDO et processus de croissance en biologie
• Definition numerique d’un protocole optimal de perfusion chrono-therapeutique en cancerologie (JC Basdevant Levi M2AN 2005)
• EDP physiologiquement structurees : resultats d’existence etd’unicite (probleme linearise aux valeurs propres), problemes inverses
• Comparaison entre valeurs propres de Floquet et Perron (rythmecircadien et proliferation cellulaire) avec S. Gaubert, MAXPLUS
• Quelques articles : Doumic MMNP 2007, JC IEEE-EMBMag2008, Bekkal Brikci JMB 2008, Bekkal Brikci MCM 2008, JCMMNP 2009, Doumic Perthame Zubelli Inverse Prob. 2009, JCGaubert Lepoutre MMNP 2009, Doumic Kim Perthame 2009,Doumic Gabriel 2009, Doumic Goudon Lepoutre CMS 2009

Principaux resultats :
1. EDP/ EDO et processus de croissance en biologie
Mouvements cellulaires et auto-organisation spatiale :
• Approche originale utilisant des methodes de transport optimal (en
collaboration avec J. A. Carrillo - ICREA, Barcelone)
• Simulation fine de croissances dendritiques
• Developpement d’une approche multi-echelle, de la cellule a la
colonie bacterienne (N. Vauchelet)

Principaux resultats : 2. Modeles individu-centres
• Reseaux de regulation intracellulaires et expression genique (Missalet al., 2006)
• Modeles individu-centres “lattice-free” et croissancespatio-temporelle mono-couche a partir de quantites biophysiquesintracelulaires (Galle et al., 2005 ; Drasdo et al., 2007)
• Modeles multi-echelle de la croissance tumorale avec controlemoleculaire de la motilite et de l’adhesion (Ramis Conde et al. 2008)
• Modeles continus (conservation de la masse et lois de Darcy) etlien avec le ‘lattice-free agent-based model’ (Byrne et Drasdo, 2008)
• Prediction de l’organisation tissulaire pendant la regenerationhepatique apres lesions pharmaco-induites (Hoehme et al., CBI 2007(2D) et 2009 (3D))

Principaux resultats :
3. Ecoulements geophysiques a surface libre (EDP)
• Extension des equations de Saint-Venant a partir des equations de
Navier-Stokes a surface libre : multi-couches (Audusse, 2005),
non-hydrostatique (Bristeau et Sainte-Marie, 2008), Euler moyenne
verticalement, densite variable
• Resolution numerique de ces systemes, schemas cinetiques,
schemas well-balanced (Audusse et al., 2004 ; Audusse et Bristeau,
2005 ; Audusse et al., 2008)
• Application aux avalanches de debris (Mangeney et al., 2007 ;
Pirulli et al., 2007)

Objectifs : 1. EDP, EDO, processus de croissance en biologie
• Prion et maladies neurodegenatives : ANR TOPPAZ F. Charles,
M. Doumic Jauffret, P. Gabriel, T. Lepoutre, B. Perthame, avec le
CEA (N. Lenuzza, F. Mouthon, H. Rezaei) et l’IMPA (J. Zubelli)
• Pharmacocinetique-pharmacodynamie moleculaire des
anticancereux, optimisation chronotherapeutique de
polychimiotherapies. Collaboration avec F. Levi, INSERM U 776
• Cycle cellulaire et horloges circadiennes ; synchronisation des
cellules d’une population, impact sur sa proliferation : ERASysBio+
(F, NL, UK) C5Sys, F. Levi (Villejuif) et D. Rand (Warwick)
• Modelisation de l’hematopoıese (saine/leucemique). Collaboration
avec J.-P. Marie, UMRs UPMC-INSERM U 872, equipe 18

Objectifs : 2. Modeles individu-centres
• Couplage tumeur/vascularisation sur modeles individu-centres
pour la prediction par simulation de l’effet d’un medicament et son
optimisation en cancerologie (avec REO et le Deutsches
Krebsforschungszentrum)
• Modelisation multi-echelle de la structure du foie et de sa
regeneration (EU PASSPORT, BMBF HEPATOSYS) et
cancerogenese hepatique (EU CANCERSYS)

Objectifs : 3. Ecoulements geophysiques a surface libre
• Navier-Stokes 3D non-hydrostatique par approche multicouches
sur maillage 2D
• Simulation numerique d’ecoulements stratifies (densite liee a la
temperature et/ou la salinite)
• Couplage hydro-biologie, simulation des ecosystemes aquatiques

Colloques organises recemment
• Mars 2008 : Ecole CEA-EDF-INRIA “Cancer modelling”, INRIA,
Rocquencourt
• Mars 2008 : “Haematology and its disorders”, Hotel-Dieu, Paris
• Decembre 2008 : “PK-PD of anti-cancer drugs”, Cordeliers, Paris
Colloque programme
• Juin 2010 : “Role and impact of maths in medicine”, IHP, Paris

Collaborations
• INRIA : Reo, Maxplus, Macs ; Estime, Contraintes ; Simpaf
• INSERM U 776, U 872, U 972 (cancers : tumeurs solides,leucemies ; cellules souches et micro-environnement)
• IMPA (Rio de Janeiro), INRIA-MITACS (Canada), CMM(Santiago du Chili)
• Leipzig, Berlin, Dundee, Nottingham, Edimbourg, Barcelone,Turin, Chicago
• EDF-LNHE ; IPGP (ecoulements granulaires)
• Reseaux : M3CSTGT, BIOSIM et TEMPO ; PASSPORT (chir.hepatique), CANCERSYS (tumeurs hepatiques) ; ANR METHODE :Mapmo (Orleans), Cemagref, Cermics ; INRA (trophoblaste)

Positionnement international ; equipes proches
• O. Diekmann, M. Gyllenberg, M. Mackey : EDP
physiologiquement structurees (age, taille, contenu moleculaire)
• G. Webb, L. Pujo-Menjouet : prion (ANR MADCOW), F. Crauste,
M. Adimy : hematopoıese (ANR PROCELL)
• J. Glazier (Bloomington, USA), P. Hogeweg (Utrecht), A.
Deutsch (Dresde), M. Meyer-Hermann (FIAS Francfort/Main) :
modeles individu-centres
• R. Leveque (Seattle), M. Castro et C. Pares (Malaga), E.
Fernandez-Nieto (Seville), S. Noelle (Aix-la-Chapelle), S. Monismith
(Stanford), B. Hodges (Austin), J. Imberger (Australie) :
ecoulements geophysiques


















