Approche fonctionnelle diachronique et pluriscalaire des ... · précieuses relectures du rapport....
85
UNIVERSITE DES SCIENCES ET TECHNIQUES DU LANGUEDOC UNIVERSITE MONTPELLIER II - Master : Sciences et technologies – Mention : Biologie, Géoscience, Agroressources, Environnement Spécialité : Ingénierie en Ecologie et Gestion de la Biodiversité Approche fonctionnelle diachronique et pluriscalaire des queyrellins de Villar d’Arène Application à des fins de gestion intégrée des prairies de fauche subalpines Stage effectué au Laboratoire d’ECologie Alpine (LECA) / Station Alpine Joseph Fourrier (SAJF) Sous la direction de Sandra Lavorel (LECA) et Florence Baptiste (Université de Barcelone/Dpt Physiologie Végétale) CAUBET Simon Stage de Master 2 Mars 2009 – Septembre 2009
Transcript of Approche fonctionnelle diachronique et pluriscalaire des ... · précieuses relectures du rapport....

UNIVERSITE DES SCIENCES ET TECHNIQUES DU LANGUEDOC
UNIVERSITE MONTPELLIER II
- Master : Sciences et technologies – Mention : Biologie, Géoscience, Agroressources, Environnement
Spécialité : Ingénierie en Ecologie et Gestion de la Biodiversité
Approche fonctionnelle diachronique et
pluriscalaire des queyrellins de Villar d’Arène
Application à des fins de gestion intégrée des prairies de
fauche subalpines
Stage effectué au Laboratoire d’ECologie Alpine (LECA) / Station Alpine Joseph Fourrier (SAJF)
Sous la direction de Sandra Lavorel (LECA) et Florence Baptiste (Université de
Barcelone/Dpt Physiologie Végétale)
CAUBET Simon Stage de Master 2
Mars 2009 – Septembre 2009


REMERCIEMENTS
Je tiens avant tout à remercier Sandra de m’avoir fait confiance pour la réalisation de
ce stage. Mais également à Florence pour s’être investie si activement dans mon encadrement.
Merci à toutes les deux pour votre pédagogie et leur patience.
La grande charge de travail, sur le terrain et en labo, n’aurait pas pu se faire par un
seul homme (en tout cas pas moi !). Je remercie donc Marie Pascale, Karl, Manue, Sandra
pour leur aide sur le terrain. Patrick, Francesco et Cécile pour leurs coups de pouce en
statistiques. Merci à Bruno pour les analyses de sol. Un grand merci à Roland et à son
incommensurable connaissance des plantes (et cinématographique). Merci à Flore pour ces
précieuses relectures du rapport.
Les conditions de vie particulière dans ce petit chalet-labo ont fait de ce stage une
véritable aventure humaine. Tous ces repas, barbecues et chouilles partagées je les dois à
Maxime, Eric et Jonathan, Sandrine, Constance, Manue, Patrick, MP. Mais aussi à Richard,
Pascal, Rolland, Serge, Eliane, Isabelle, Harold et tous les stagiaires de passage au jardin
botaniques alpin et qui sont quelque fois venu avec moi sur le terrain. Ainsi qu’à tous ceux
qui ont été de passage dans ce chalet.
Enfin, pour ce stage de fin d’étude, une pensée envers mes parents qui m’ont permis
d’en arriver jusque là.


INTRODUCTION 1°) Contexte global...........................................................................................................1
2°) La problématique de l’adret de Vilar d’Arène .............................................................1
3°) La fauche ....................................................................................................................3
4°) Etude fonctionnelle des prairies ..................................................................................3
A) Approches d’études des prairies ..............................................................................3
B) Les traits fonctionnels .............................................................................................4
5°) Présentation de l’étude ................................................................................................5
MATERIEL ET METHODES 1°) Site d’étude.................................................................................................................6
2°) Espèce et écosystème étudiés ......................................................................................6
3°) Méthodologie..............................................................................................................7
A) Dynamique de la végétation suite à l’abandon.........................................................7
a) Détermination des parcelles d’études ...................................................................7
b) Acquisition des données floristiques ....................................................................7
B) Dynamique des communautés aux changements d’usages du sol.............................8
a) Parcelles expérimentales ......................................................................................8
b) Acquisition des données floristiques ....................................................................8
C) Mesures de croissance.............................................................................................8
D) Acquisition des variables environnementales et fonctionnelles ................................9
a) Production de biomasse épigée de F. paniculata ..................................................9
b) Acquisitions des variables fonctionnelles .............................................................9
c) Analyses des sols ...............................................................................................10
4°) La base de donnée VISTA ........................................................................................10
5°) Traitement statistique................................................................................................10
A) Approche descriptive ............................................................................................10
B) Approche comparative ..........................................................................................11
III. RESULTATS 1°) Dynamique de végétation suite à l’abandon ..............................................................12
A) Composition floristique.........................................................................................12
B) Processus écosystémiques .....................................................................................13
C) Traits fonctionnels des plantes...............................................................................14
2°) Dynamique de végétation en réponse à la fauche/abandon ........................................15
3°) Influence de la fauche sur la performance de F. paniculata .......................................16
IV. DISCUSSION 1°) Dynamiques des pelouses subalpines à F. paniculata.................................................18
2°) Réponse de la végétation aux changements d’utilisation des terres ............................20
3°) Perspectives de gestion intégrée du site .....................................................................21
V. CONCLUSION ET PERSPECTIVES ......................................................................23
BIBLIOGRAPHIE
ANNEXES
TABLE DES MATIERES

Figure 1 : Villar d’Arène : une communauté de montagne, entourée de prairies semi-naturelles dont
l’entretien repose aujourd’hui sur une agriculture économiquement marginale. Les prairies en
terrasses, témoignent d’un labour désormais arrêté.

Introduction
1°) Contexte global
Le changement d’utilisation des terres est un facteur essentiel dans la dynamique des
paysages (Vitoussek et al, 1997). Ces modifications se répercutent tant à l’échelle globale que
locale (Hooper et al, 2005), pour affecter, in fine, les populations locales qui en dépendent
(Foley et al, 2005). Ainsi furent établies les conséquences du changement que subissent les
écosystèmes pour le bien être humain et les possibilités de réagir, dans le cadre du Millenium
Ecosystem Assessment (2005). Ceci a permis une formalisation et une diffusion de la notion
de service écologique (ou écosystémique) (Daily, 1997 ; Chevassus-au-Louis et al., 2009).
Cette notion ajoute une dimension importante à la perception du fonctionnement des
écosystèmes dans un contexte anthropique (Kremen, 2005).
Bien que l’agriculture de l’arc alpin ne soit que d’une importance socio-économique
secondaire, elle exerce toujours la plus forte influence humaine directe sur l’évolution des
paysages alpins ainsi que sur la qualité et la quantité des milieux (Stöcklin & al., 2007). Elle
détermine donc une des principales ressources économiques locales au travers du tourisme et
des prestations écologiques. L’environnement tient donc une place centrale dans
l’argumentation d’une agriculture multifonctionnelle (Dobs & Pretty, 2004)
2°) La problématique de l’adret de Villar d’Arène
Villar d’Arène (Hautes Alpes) (Fig. 1), petite commune de montagne, n’échappe pas à
cette problématique actuelle. L’adret accueille une activité humaine depuis plus de 3000 ans
(Muller & al., 2000), et est essentiellement formé de prairies subalpines. Or, si ces
écosystèmes semi-naturels fournissent de nombreux services écologiques, leur entretien
repose aujourd’hui sur une agriculture économiquement marginale. Villar d’Arène constitue
donc un terrain approprié à l’étude de la vulnérabilité des écosystèmes aux changements de
l’utilisation des terres.
L’adret de Villar d’Arène est une zone aux forts enjeux économiques (tourisme et
agriculture) et écologiques (Natura 2000, Parc National des Ecrins). Les prairies subalpines de
Villar d’Arène abritent une grande diversité floristique faisant la renommée du site. Cette
flore des alpages est donc l’élément majeur de l’attrait de cette haute montagne sur les
touristes (200 000 visiteurs/an). Cet attrait, caractérisé en grande partie par une approche
hédoniste de cette zone, est donc relié à la diversité de couleurs des alpages fleuris (Quétier,
1

Figure 2 : Influence de la pratique de la
fauche sur les queyrellins (Mai 2009).
Prairie abandonnée Prairie fauchée

2006). La pérennité de cette activité socio-économique repose donc sur la diversité floristique
des prairies. Or, ce paysage s’est considérablement modifié ces 150 dernières années. Ainsi,
en 1810 les zones de labour s’étendaient jusqu’à 1900m d’altitude et les prairies de fauches
jusqu’à plus de 2100m (Mallen, 2002 ; Annexe 1 et 2). Aujourd’hui, à cette altitude, les
prairies de fauche sont délaissées (voire abandonnées) au profit de zones à vocation pastorale.
Ce changement est représentatif de la grande tendance actuelle générale à la déprise agricole
en montagne.
La communauté herbacée de ces prairies subalpines est dominée par la fétuque
paniculée (Festuca paniculata (L.) Schinz & Thell., localement dénommée « queyrel »). Sur
l’adret de Villar d’Arène, elle peut occuper jusqu’à 70% de la biomasse épigée lorsque les
prairies sont abandonnées. Cette graminée produit beaucoup de litière dont la dégradabilité
demeure limitée (Vittoz et al., 2005). De même, lorsqu’elle devient dominante, elle ralentit
les cycles des nutriments (Robson et al., 2007). Par ailleurs, elle est monopoliste et peu
appétente pour le bétail. De ce fait, aujourd’hui, ces prairies (également appelées queyrellins)
sont rarement fauchées, le plus souvent pâturées ou abandonnées. Et en l’absence
d’interventions humaines, les pelouses à fétuques paniculées évoluent vers des pelouses quasi
mono-spécifiques (Parc National des Ecrins, 2006). L’agriculture, et en particulier la fauche,
semble au contraire limiter son abondance, permettant ainsi le développement d’autres
espèces subalpines (Fig. 2).
Ainsi, l’abandon progressif de la fauche représente un risque de perte de diversité
végétale (Fischer & Wipf, 2002). Même si cette importante diversité floristique régionale est
issue d’une géologie complexe (Quétier, 2006), ce sont surtout la conjonction des facteurs
climatiques et humains qui ont conduit à l’émergence de communautés végétales
particulières. De ce fait, les agriculteurs encore présents y sont fortement subventionnés, via
des mesures agro-environnementales, afin de maintenir des pratiques traditionnelles de
gestion des écosystèmes par la fauche. Sans ces subventions, cette agriculture marginale ne
serait pas viable. Elle se justifie donc par la fourniture de services écologiques (MacDonald et
al., 2000).
Cette problématique n’est pas propre à l’adret de Villar d’Arène, et concerne
également d’autres zones de montagne de l’arc alpin. La préservation de la richesse floristique
au sein des prairies affectées par la fétuque paniculée, est un des défis des gestionnaires
locaux. Or, si l’évolution agricole de ces dernières décennies a engendré une modification des
prairies locales par le développement de cette graminée, les liens de causes à effets sont peu
connus.
2
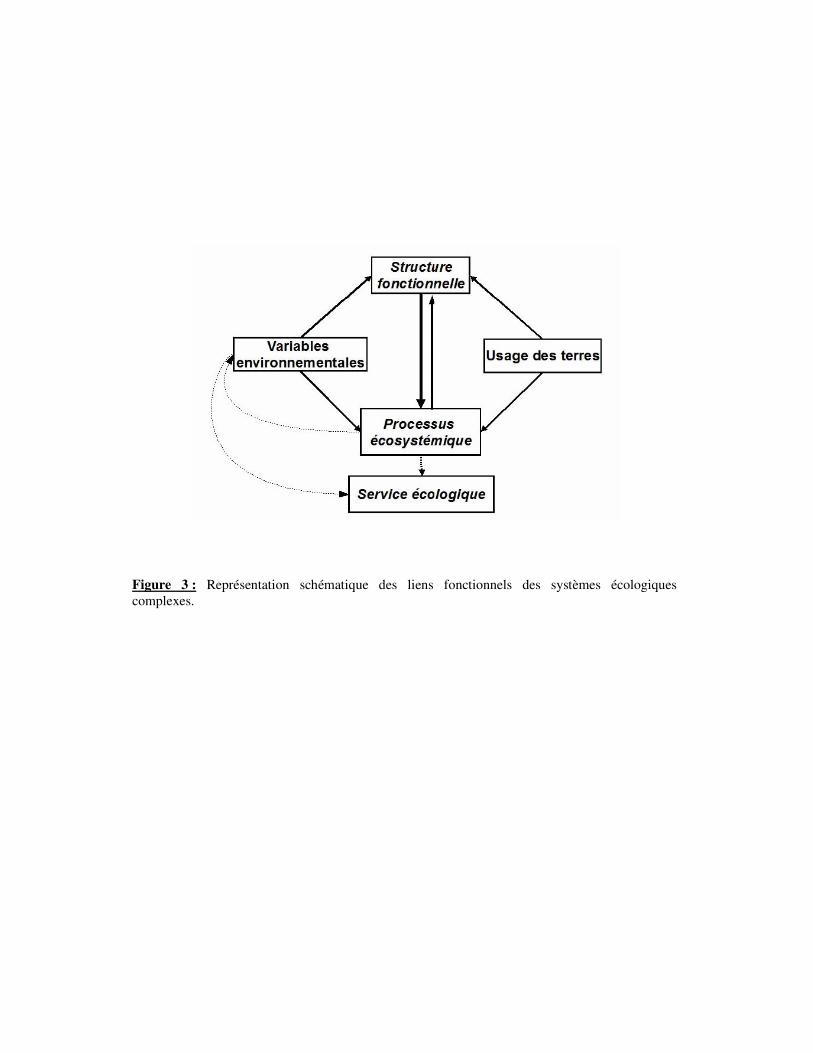
Figure 3 : Représentation schématique des liens fonctionnels des systèmes écologiques
complexes.

3°) La fauche
Cette notion de fauche, même si elle est intuitivement compréhensible, recouvre des
aspects qui peuvent paraître triviaux, mais qu’il n’est pas inutile de rappeler pour en
comprendre ses conséquences. La fauche affecte la diversité floristique, notamment en
favorisant certaines espèces au détriment d’autres. En effet, elle supprime une part importante
de la biomasse aérienne, ce qui affecte les espèces présentes et principalement celles de hautes
statures (Klimes & Klimesova, 2001). Ainsi, les espèces capables de boucler leur cycle de
reproduction en dépit de cette perturbation seront avantagées (Pausas & Lavorel, 2003).
D’autre part, cette biomasse est retirée de l’écosystème à des fins de fourrage hivernal. Elle ne
retourne pas au sol, qui s’appauvrit alors en éléments nutritifs. La fauche a donc des
conséquences importantes sur la structure et le fonctionnement de la communauté végétale et
de l’écosystème (Foster, 1999).
Néanmoins, la fauche est considérée comme une perturbation engendrant une diversité
spécifique élevée par rapport aux prairies abandonnée (Quétier et al., 2007). En effet, une
théorie classique veut que les perturbations limitent la compétition et favorisent ainsi la
diversité (Shea et al., 2004). Cette pratique est donc, de nos jours, considérée comme un outil
essentiel de conservation de la diversité végétale.
4°) Etude fonctionnelle des prairies
A) Approches d’études des prairies
Dans un contexte régional précis, la composition floristique et la performance
(production primaire, minéralisation) d’une prairie résulte, à un instant donné, d’interactions
complexes entre les facteurs du milieu (climat, sol, topographie) et les facteurs de gestion
(fauche, pâturage, fertilisation) présents ou antérieurs (Louault et al., 2002) (Fig. 3). Cette
structure dépend donc des adaptations et des réponses des organismes à l’environnement,
mais également des interactions entre organismes. Dans le cas des écosystèmes prairiaux, les
deux principaux facteurs contrôlant la dynamique de végétation (en dehors des facteurs
environnementaux) sont le mode d’exploitation (fauche et pâturage) et la fertilisation
(Ansquer et al., 2004). Ces facteurs peuvent créer une grande variabilité des caractéristiques
structurales de la prairie. Ces communautés peuvent donc être définies par un certain nombre
de descripteurs qui leurs sont propres (richesse spécifique, diversité spécifique, diversité
fonctionnelle…) (Ricklefs & Miller, 2005).
3


Les relations diversités biologiques - processus écosystémiques sont actuellement l’objet de
nombreux débats (e.g. Grime, 1997, 2002 ). Pour certains auteurs, le rôle de la diversité
fonctionnelle semble plus adapté, dès lors que l’on cherche à comprendre les processus
écosystémiques (Westoby, 1998 ; Diaz & Cabido, 2001). En effet, cette approche fait
l’hypothèse qu’une espèce peut être définie par des caractéristiques fonctionnelles, ou traits
fonctionnels, permettant ainsi de mieux appréhender la structure des communautés (et le
fonctionnement des écosystèmes) (Chapin et al., 2000 ; Diaz et al., 2006 ; Eviner & Chapin,
2003 ; Lavorel et al., 2007). De plus, cette approche fonctionnelle permettrait de réduire la
grande complexité des systèmes biologiques de façon à les rendre plus facilement
descriptibles (Smith et al., 1997).
Une approche fonctionnelle des systèmes biologiques a donc été développée pour
tenter de réduire la grande complexité des ces systèmes (Smith et al., 1997). Cette approche
fait l’hypothèse qu’une espèce peut être définie par des caractéristiques fonctionnelles, ou
traits fonctionnels. Ces traits sont ensuite utilisés pour comprendre la structure des
communautés et le fonctionnement des écosystèmes (Chapin et al., 2000 ; Diaz et al., 2006 ;
Eviner & Chapin, 2003 ; Lavorel et al., 2007).
B) Les traits fonctionnels
Les traits fonctionnels peuvent être définis comme l’ensemble des caractéristiques,
qualitatives ou quantitatives, mesurables sur un individu (de la cellule à l’organisme) (Vile,
2005). D’une manière plus formelle, les traits fonctionnels sont définis comme des caractères
« morpho-physio-phénologiques » qui ont un impact sur la valeur sélective (Violle et al.,
2007). En effet, l’hypothèse sous-jacente est que si un caractère est retrouvé chez une majorité
d’espèce, il est potentiellement utile pour ne pas être éliminé de la communauté végétale.
Ainsi, l’étude des traits fonctionnels permet de comprendre quels sont les critères de filtrage
par l’environnement et les espèces voisines. En plus de ces traits fonctionnels de réponse aux
facteurs de l'environnement, on peut distinguer les traits fonctionnels d'effet sur les processus
écosystémiques (Lavorel & Garnier 2002). En effet, le fonctionnement de l’écosystème
dépend des traits des espèces et de leur abondance relative au sein de la communauté (Grime,
1998). Ainsi, le fonctionnement de l’écosystème peut également être approché à partir des
seules caractéristiques des espèces dominantes de la communauté (Grime, 1998). Ces espèces,
parce qu’elles représentent la plus grande quantité de matière et d’énergie, auraient le plus
grand effet sur la structure et le fonctionnement de l’écosystème (Cruz & al., 2002).
4

Figure 4 : Schéma conceptuel de la démarche employée dans le cadre de cette étude

Ainsi, Les traits fonctionnels peuvent être utilisés pour décrire les communautés végétales à
différents niveaux d’agrégation : (i) à l’échelle de l’espèce et (ii) à l’échelle de la
communauté. A l’échelle de l’espèce, l’étude des traits permet de décrire la réponse des
espèces selon des gradients environnementaux. A l’échelle de la communauté, une approche
récente utilise une valeur de trait pour l’ensemble calculée comme la moyenne des valeurs de
traits des espèces, pondérée par leur abondance dans la communauté (Garnier & al., 2007 ;
Lavorel & al., 2008). C’est la valeur de trait agrégée. Elle représente la réponse de la
communauté dans son ensemble selon un gradient environnemental.
Les traits fonctionnels sont donc un excellent outil pour la compréhension de la
diversité. Ils offrent la possibilité de lier la dynamique des communautés végétales (e.g.
abondance relative des espèces et de leurs traits) au fonctionnement des écosystèmes (e.g. les
cycles biogéochimiques) (Lavorel & Garnier, 2002).
5°) Présentation de l’étude
D’une manière générale, cette étude se propose de connaître l’impact de la
pratique/abandon de la fauche sur la dynamique végétale des queyrellins. Cette problématique
est abordée au travers de deux questions sous-jacentes : (1) Comment la fauche/abandon
modifie la diversité fonctionnelle et spécifique des communautés végétales des prairies
subalpines ? (2) Comment la fauche affecte-t-elle la performance de la fétuque paniculée ?
Pour répondre à ces questions, trois volets seront utilisés (Fig. 4).
(i) Une étude de la dynamique de végétation des queyrellins sur l’ensemble de l’adret,
sera étudiée en fonction de l’arrêt de la pratique de la fauche selon un gradient temporel de
deux siècles. (ii) Une étude plus expérimentale de la pratique de la fauche, au sein de huit
communautés végétales, permettra également de quantifier la réelle influence de cette
dernière sur une dynamique de végétation à court terme. Enfin, F. paniculata étant l’espèce
dominante de ces communautés subalpines, (iii) une approche plus fine a pour objectif
d’étudier l’impact de la fauche sur sa performance.
Au travers de différentes échelles spatiales et temporelles, cette étude apportera des
éléments de compréhension sur le fonctionnement des queyrellins. Ceci afin d’utiliser au
mieux les potentialités économiques et écologiques de ces écosystèmes, au travers d’une
gestion intégrée de l’activité agro-pastorale du site.
5

Figure 5 b : Le site d’étude de l’adret de Villar d’Arène
Figure 5 a : Localisation géographique
du site d’étude

Matériel & Méthodes
1°) Site d’étude
Le site d’étude est localisé à proximité du col du Lautaret, exposé face sud (ou adret)
de la haute vallée de la Romanche, Alpes centrales françaises (Villar d’Arène, 45.04°N, 6.34°
E) (Fig. 5). Le substrat est un calcaire-schiste homogène et le climat est subalpin avec une
influence continentale. Les prairies subalpines fauchées de Villar d’Arène sont situées entre
1600m et 2100m d’altitude Elles sont soumises à des températures moyennes variant de -
1,6°C en Janvier à 14°C en Juillet (Fig. 6). L’intensité lumineuse est très intense en été et les
précipitations sont peu importantes à cette altitude. La saison de végétation ne dure que six
mois en moyenne.
2°) Espèce et écosystème étudiés
La fétuque paniculée (Festuca paniculata (L.) Schinz & Thell) est une graminée
pérenne cespiteuse géophyte à bulbe typiquement subalpine, de sol plutôt profond (Fig. 7). Sa
morphologie indique qu’elle serait une espèce compétitrice expliquée par une canopée dense
et une extension latérale importante (Gross, 2003). De plus, sa croissance printanière est
relativement rapide, ce qui peut lui conférer un avantage. Cependant, elle possède une surface
spécifique foliaire faible contrairement aux espèces compétitrices (Grime, 1977). Elle est
donc classée comme une espèce compétitrice conservatrice (Gross & al, 2007). En plus, de
ces traits caractéristiques d’espèce compétitrice, elle utilise des mécanismes allélopathiques
pour conforter sa dominance au sein de la communauté (Viard-Crétat & al, 2008). C’est donc
l’espèce dominante par excellence des prairies subalpines d’adret jamais labourées et où la
fauche a cessé (Quétier & al, 2007). Elle est présente en co-dominance dans les prés de
fauche, mais elle devient dominante lorsque cette pratique s’arrête. Elle est essentiellement
absente des anciens labours.
Les queyrellins présentent exclusivement des espèces pérennes, pour la plupart à
croissance lente et pouvant être qualifiées de conservatrices (Berendse & Elberse, 1990).
Malgré le caractère monopoliste de la fétuque paniculée, jusqu’à cinquante espèces peuvent
être trouvées par mètre carré dans les prairies fauchées (Quétier & al, 2007). Cependant, peu
de plantes poussent autour des pieds de fétuque paniculée lorsqu’ils sont très développés.
6

Figure 6 : Diagramme pluvio-thermique de Mônetier-les-Bains (1400m d’altitude, période
de référence 1926 – 1987) ; données Météo France. Pluviométrie cumulée par mois et
moyennes thermiques mensuelles
Figure 7 : Touffe de Festuca paniculata avec inflorescence.

3°) Méthodologie
A) Dynamique de la végétation suite à l’abandon
a) Détermination des parcelles d’études
Dans un premier temps, les parcelles adéquates doivent être définies selon trois
paramètres. (i) La trajectoire actuelle d’utilisation du sol (Annexe 3). Ceci a permis d’isoler
deux trajectoires en lien avec l’étude. Elles correspondent à des prairies de foin non fertilisées
et des prairies de pâtures bovines/ovines en présence de F. paniculata (trajectoires 4 et 5 de
l’annexe 1). (ii) L’aspect dynamique de végétation est abordé au travers d’un gradient
temporel de cessation de la pratique de la fauche. Des interprétations cadastrales et des photos
aériennes ont permis de dresser une cartographie de l’arrêt de la fauche par parcelle (Bautista,
2003 ; Lamarque, non soumis ; annexe 2). Ce gradient recouvre des prairies dont la fauche
s’est interrompue à partir de 1810 jusqu’à une activité encore présente de nos jours. (iii) Les
prairies doivent être caractérisées par la présence de F. paniculata.
La superposition de ces trois paramètres a permis d’isoler des parcelles propres à
l’étude. Puis, cinq classes d’âges (x) ont été établies selon le principe suivant : prairies
fauchées (x) - fauche arrêtée depuis 5ans < x < 20ans < x < 50ans < x < 100ans < x. Pour
chacune de ces classes, trois réplications (parcelles) ont été choisies à priori sur le SIG du site
d’étude, et validées en terme de conditions locales sur le terrain début Juin 2009. Soit un total
de 15 parcelles. Ces parcelles ont été choisies avec un gradient altitudinal faible et toutes en
pâturage ovins. Ceci afin de minimiser l’influence de ces deux facteurs sur leurs compositions
floristiques.
b) Acquisition des données floristiques
Les relevés floristiques sont effectués selon la méthode des points quadrats (Daget &
Poissonnet, 1969). Trois transect de 20m sont réalisés de façon aléatoire dans les parcelles de
manière à ce qu’ils ne se croisent pas. Ceci permet de considérer le groupement végétal dans
son ensemble et de traduire ainsi la variabilité due à une éventuelle hétérogénéité (Amiaud,
1998). Le long de ces transects, tous les 50cm, sont relevées toutes les espèces entrant en
contact avec une aiguille descendue verticalement dans la masse végétale. Cette méthode
permet d’évaluer les fréquences de toutes les espèces recensées. Un total de 1845 point de
relevés a été effectué selon cette approche entre le 27 Juin et le 24 Juillet. Afin de minimiser
le biais dû à la phénologie, l’échantillonnage s’est déroulé selon le gradient altitudinal et
l’exposition des parcelles.
7

Figure 8 : Schéma des traitements expérimentaux de couple de communautés (fauché / non fauché). +C : coupé ; -C : non coupé ; +L : litière laissée ou ajoutée ; -L : litière enlevée. +C-L
représente une fauche entière, tandis que –C+L correspond à un abandon total de la fauche. Les
traitements +L étaient toujours en bas des parcelles pour éviter une contamination des traitements -L,
tandis que les traitements +C/-C ont été réalisés aléatoirement à droite ou à gauche. Chaque carré (par
exemple +C+L) est une sous-parcelle (‘subplot’). Les sous parcelles –C+L et +C-L sont les témoins
respectifs en communautés non fauchées et communautés fauchées.
Figure 9 : Quadrat utilisé pour déterminer la composition floristique (50cm x 50cm, divisé en 25
carrés)

B) Dynamique des communautés aux changements d’usages du sol
a) Parcelles expérimentales
Cette démarche s’appuie sur un dispositif de terrain expérimental. Afin de tester l’effet
de la fauche sur la communauté végétale des queyrellins et sur la croissance de F. paniculata,
un dispositif de terrain a été mis en place en 2005 (Fig. 8).
Huit communautés ont été choisies au sein de prairies fauchées (4 communautés) et
non fauchées (4 communautés). Chaque communauté fauchée était adjacente à une
communauté non fauchée, permettant d’avoir des conditions édapho-climatiques similaires.
Ces communautés avaient été au préalable étudiées et correspondent à des contrastes de
gestion par la fauche (Garcia Bautista, 2004) donnant lieu à un contraste floristique important
(Clement & al, 2003). Ces parcelles, de 8m×10m, sont situées à proximité du col de Lautaret,
à une altitude comprise entre 1800m et 1900m sur le Lieu dit « la Traverse » exposé au sud.
Pour chacune de ces huit communautés, quatre traitements ont été appliqués. La moitié des
quatre sous-parcelles ont été fauchées. De façon orthogonale à cette perturbation, la litière a
été enlevée sur la moitié des sous-parcelles. Ce dispositif permet ainsi d’observer
indépendamment les effets de la fauche, de la litière et de la coupe de la biomasse.
b) Acquisition des données floristiques
La composition floristique a été déterminée selon une méthode semi-quantitative.
Dans un quadrat de 50cm×50cm, divisé en 25 carrées de 10cm de côté, un relevé d’espèces
est effectué pour chacun de ces sous-carrés (Fig. 9). Ceci permet de déterminer la proportion
des carrés dans laquelle une espèce est présente et une fréquence de présence peut ainsi être
obtenue pour l’ensemble des espèces présentes (Lavorel & al, 1999). Pour chaque sous-
parcelles, quatre quadrats ont été inventoriés, soit un total de 128 quadrats. Ces relevés ont été
effectués entre le 4 Juillet et le 23 Juillet 2009.
C) Mesures de croissance
Une fois le site d’étude déneigé, 10 individus ont été marqués dans chaque sous-
parcelles des huit communautés présentées précédemment. Les individus ont été repérés à
partir de la touffe restante de l’année précédente. Il convient de préciser qu’un individu est
représenté par une touffe qui est composée de plusieurs talles. Ces individus ont été choisis de
façon aléatoire.
Pour chacun des individus, les deux diamètres basaux ont été mesurés (longueur et
largeur) afin de calculer sa surface. Cette mesure est effectuée à une fréquence
8
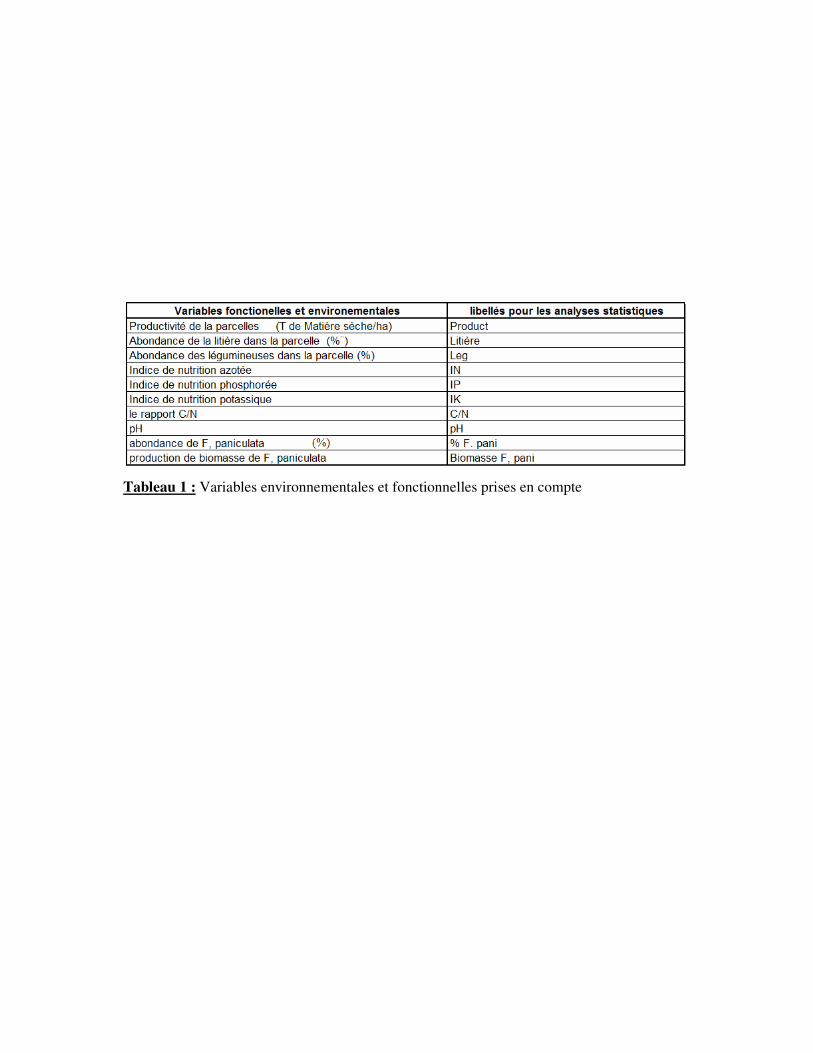
Tableau 1 : Variables environnementales et fonctionnelles prises en compte

bimensuelle à partir du mois d’Avril jusqu’à Juillet. Sur chacun des ces individus marqués, la
hauteur de trois talles choisies au hasard a été mesurée. Ces mesures sont effectuées de
manière hebdomadaire du mois d’Avril à la fin du mois de Mai, puis bimensuelle jusqu’à la
fin du mois de Juillet.
D) Acquisition des variables environnementales et fonctionnelles
Afin de mieux comprendre la composition floristique des prairies et leur
fonctionnement, plusieurs variables environnementales et fonctionnelles (production de
biomasse, indice de nutrition azotée, …) ont été mesurées en fonction de la date d’arrêt de
fauche (Tab. 1).
a) Production de biomasse épigée de F. paniculata
Afin d’éstimer sur le terrain la production de biomasse verte aérienne de F.
paniculata, une relation allométrique entre la biomasse aérienne et la surface de la touffe a
été établie (Annexe 4).
Cinq individus de F. paniculata ont été prélevés en prairies fauchées et cinq en
prairies abandonnées. Pour chacun de ces dix individus, la surface, la hauteur et le nombre de
thalle ont été mesurés. La partie aérienne des talles est séchée à l’étuve à une température de
60°C pendant 48h, puis pesée. Une régression linéaire multiple, a permis de déterminer une
relation entre la surface des individus et la production de biomasse aérienne verte. Une
dizaine d’individus a par la suite été mesurée dans chacune des parcelles.
b) Acquisition des variables fonctionnelles
Des prélèvements de biomasse aérienne ont été réalisés entre le 23 et 25 Juin 2009
pour chacune des parcelles suivies de l’adret. Ces prélèvements ont été effectués par une
tondeuse à main (WOLF SP75) dans quatre carrés de 0,25m2 (Barbaro, 1999) disposés
aléatoirement dans la communauté. Cette taille est un bon compromis entre la variabilité des
prélèvements due à la micro-hétérogénéité intra-parcelle et le temps nécessaire à la récolte
(Amiaud, 1998).
Pour chaque parcelle, les 4 prélèvements sont mélangés puis pesés. Un quart du poids
frais total a été sous-échantillonné afin d’être trié et pesé de la manière suivante :
inflorescences, partie sèche (litière), légumineuses et partie verte (Cruz & al., 2006). Ensuite
chacune de ces parties est séchée pendant 48h à l’étuve à une température de 60°C puis à
nouveau pesée. Ceci permet de caractériser la parcelle à travers le pourcentage de
9
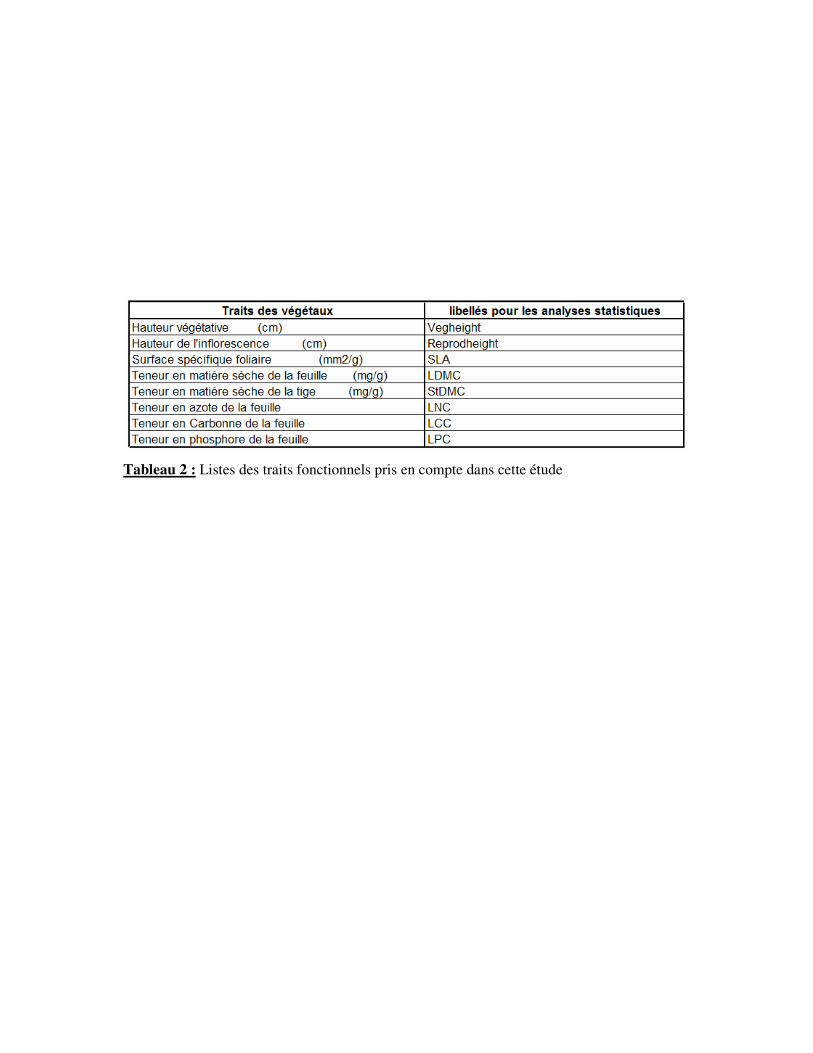
Tableau 2 : Listes des traits fonctionnels pris en compte dans cette étude

légumineuses, de litière et de matière verte aérienne. A partir de ces descripteurs, peuvent être
estimés la productivité, l’indice de nutrition azotée, potassique et phosphorée (Tab. 1). Ces
variables ont été sous-traitées par le laboratoire CESAR.
c) Analyse des sols
Sur chaque parcelle où furent effectué les relevés floristiques, un échantillon de sol a
été prélevé à l’aide d’une tarière (0-10 cm de profondeur). Cinq prélèvements sont effectués
pour chaque parcelle, et sont ensuite mélangés afin de prendre en compte l’hétérogénéité du
sol. Les prélèvements de sol ont été effectués le 27 Juin puis ont été séché à l’air libre jusqu’à
fin Juillet. Ainsi, pour chaque parcelle sont analysés le pH, le carbone (C), l’azote (N) total
(minéral et organique), le rapport C/N (Tab. 1). Ces analyses ont été réalisées au Laboratoire
d’ECologie Alpine (LECA) à Grenoble.
4°) La base de donnée VISTA
VISTA (pour Vulnerability of Ecosystem Services to land use change in Traditional
Agricultural landscape) était un projet européen cherchant à évaluer la vulnérabilité de ces
paysages traditionnels au changements d’utilisation des terres. La démarche consistait à relier
la structure et le fonctionnement des écosystèmes formant ces paysages. Dans le cadre de ce
projet, de nombreuses études sur les traits fonctionnels des espèces dominantes des prairies
subalpines de Villar d’Arène ont été effectuées et compilées dans une base de donnée sous
Access®. Ainsi pour nombre d’espèce peuvent être connus la taille, la surface foliaire et
d’autres traits (Tab. 2). Cette base de donnée a été employée pour réaliser une analyse
fonctionnelle de la composition des différentes prairies selon le gradient temporel d’arrêt de
fauche.
5°) Traitements statistiques
A) Approche descriptive
Pour les deux études de dynamique de la végétation, l’ensemble des données
floristiques relevés a été traité selon une analyse statistique mutivariée. Une Analyse
Factorielle des Correspondances (AFC) sur la composition floristique des différentes parcelles
permet de décrire les différents groupements (Barbaro, 1999). Pour l’étude de la dynamique
suite à l’abandon de la fauche, une Analyse en Composante Contrainte (ACC) (principe de
régression multiple) permet de prendre en compte simultanément l’influence floristique,
10
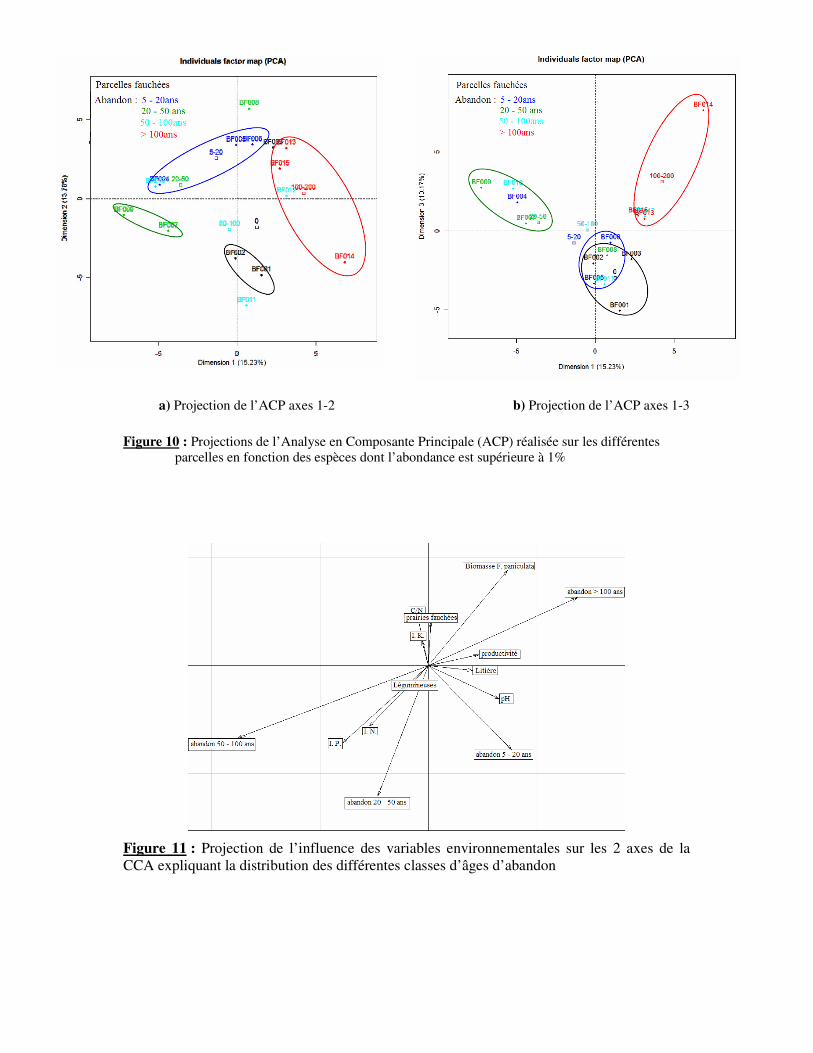
Figure 10 : Projections de l’Analyse en Composante Principale (ACP) réalisée sur les différentes
parcelles en fonction des espèces dont l’abondance est supérieure à 1%
Figure 11 : Projection de l’influence des variables environnementales sur les 2 axes de la
CCA expliquant la distribution des différentes classes d’âges d’abandon
a) Projection de l’ACP axes 1-2 b) Projection de l’ACP axes 1-3

édaphique et fonctionnelle. Ceci afin de comprendre quels sont les facteurs influant la
dynamique de végétation.
L’analyse des relations entre traits fonctionnels et variables environnementales est
réalisée au travers d’une analyse de co-inertie généralisée entre trois tableaux (Chessel & al,
1997). Cette analyse est appelée analyse RLQ (Annexe 5). Elle permet de relier,
indirectement, le milieu et son fonctionnement via les traits fonctionnels des végétaux.
L’étude de la corrélation entre les traits fonctionnels agrégés (voir Introduction) d’une
communauté végétale et les variables a été réalisée avec un test de Pearson
L’ensemble des analyses statistiques est effectué à l’aide du logiciel R (version 2.8.1).
B) Approche comparative
Des comparaisons de moyenne de l’ensemble du jeu de données récoltés (taille,
surface, biomasse, …) sont effectués par Test de Student et Wilcoxon. Ces analyses
statistiques sont effectuées à l’aide du logiciel R (version 2.8.1).
Les données récoltées dans le dispositif expérimental testant la performance de F.
paniculata ont été traitées selon une approche statistique propre au dispositif expérimental. En
effet, une analyse en split plot est la plus adaptée pour tester l’effet de plusieurs variables sur
des blocs de traitements (Dean A. & Voss D., 1999 ; Dagnelie, 2008). Ainsi, pour ce
dispositif expérimental furent testées les influences de la litière, de la coupe et de la trajectoire
d’utilisation des terres (fauchées et abandonnées) sur la hauteur et la surface de F. paniculata.
Ces analyses statistiques ont été réalisées à l’aide du logiciel JMP (version 5.0.1).
RESULTATS 1°) Dynamique de végétation suite à l’abandon
A) Composition floristique
Les différentes catégories d’âges d’abandon de la fauche sur les prairies sont
relativement bien mises en évidence par l’AFC (Fig. 10.a). Seules les parcelles de la classe
d’âge 50 – 100 ans ne sont pas structurées de manière cohérente, traduisant ainsi une forte
hétérogénéité.
La distribution observée est expliquée à hauteur de 30% par les deux premiers axes,
qui ordonnent les classes d’âge de manière relativement cohérente. L’axe 1 (15% de la
variance) différencie les parcelles de classes d’âges 5-20 ans et 20-50 ans de celle d’abandon
11
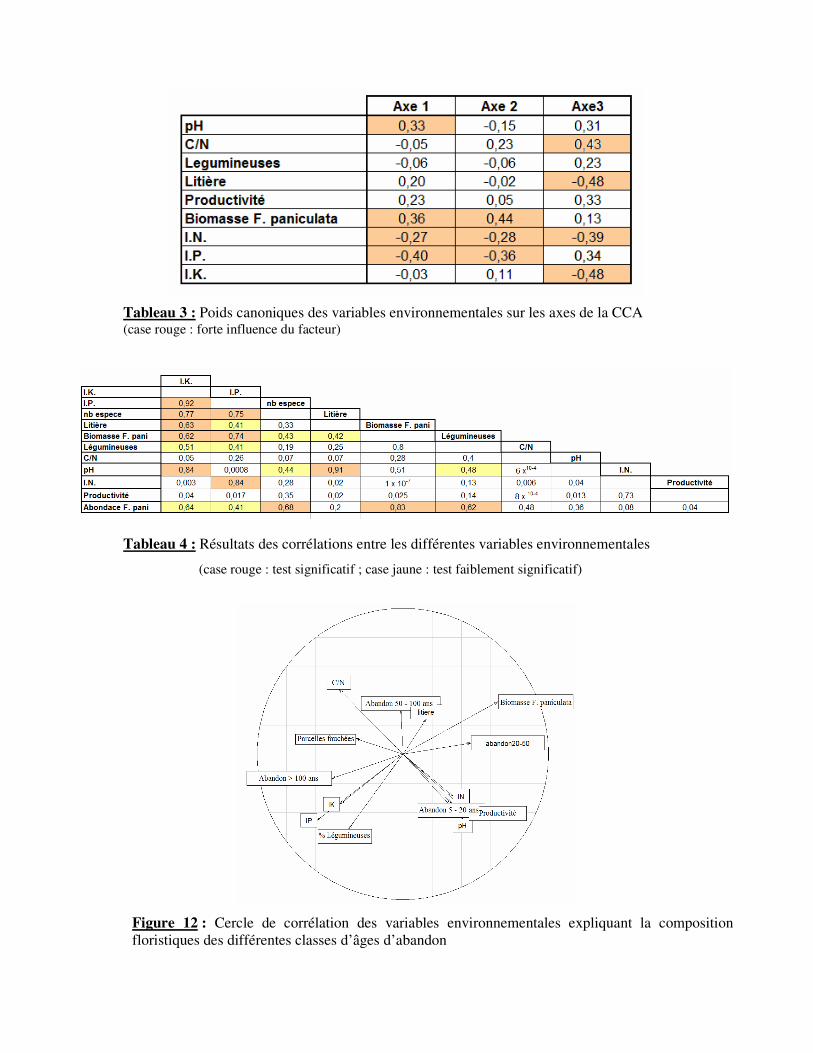
Tableau 3 : Poids canoniques des variables environnementales sur les axes de la CCA (case rouge : forte influence du facteur)
Tableau 4 : Résultats des corrélations entre les différentes variables environnementales
(case rouge : test significatif ; case jaune : test faiblement significatif)
Figure 12 : Cercle de corrélation des variables environnementales expliquant la composition
floristiques des différentes classes d’âges d’abandon

supérieur à 100 ans. Cet axe peut être expliqué par l’abondance de F. paniculata, espèce
majoritaire de cet écosystème subalpin (3% à 22% d’abondance).
L’axe 2 (14% de la variance) différencie les parcelles encore fauchées de celles dont
l’abandon est récent (5-20 ans). Cette distribution est corrélée à la hauteur de végétation. En
effet, cet axe est influencé par les parcelles BF008 et BF011. La première est caractérisée par
une végétation de couvert prairial haut (60cm – 1m) : Sanguisorba officinalis, Narcissus
poeticus, Trollius europaeus, Latyrus pratensis, Trisetum flavescens et Dactylys glomerata.
Une formation comprenant peu de graminées ou sinon de haute stature. A l’inverse, la
parcelle BF011 est majoritairement constituée par des espèces à stature plus basse : Festuca
nigrescens, Carex sempervirens, Festuca laevigata, Crocus vernus, Luzula nutans et Meum
athamanticum. Une composition majoritairement de graminées à stature plus réduite.
Enfin, une projection de l’ACP selon les axes 1-3 (25% de la variance) (Fig. 10.b) met
en avant la grande hétérogénéité des communautés végétales en terme de composition
floristique au sein des classes d’âges encore fauchées, 20-50 ans et supérieures à 100 ans. Par
exemple, la parcelle BF014 possède des espèces de prairies plus “humides” et plus grasses
telles que Gallium boreale, Gallium mollugo, Luzula nutans ou Trisetum flavescens. A
l’inverse, la parcelle BF001 est une pelouse maigre, plus sèche, avec un sol plus pauvre. Des
espèces (méso-) xérophytes comme Agrostis capillaris, Carex sempervirens, Centaurea
uniflora, Myosotis alpestre, Phyteuma michelii ou Trifolium alpinum sont caractéristiques de
ces zones.
B) Processus écosystémiques
La CCA permet de comprendre les différences floristiques inter-classes en mettant en
relation la composition floristique et les variables environnementales. Une projection de ces
dernières selon deux axes (Fig. 11) explique 35,7% de la variabilité.
L’axe 1 (20,35% de la variance) montre de fortes corrélations avec le pH (0,33), la
biomasse de F. paniculata (0.36), l’IP (-0,40) et l’IN (-0,27) (Tab. 3). Cet axe oppose donc
des prairies à production de biomasse de F. paniculata importante à des prairies dont les
ressources azotées et phosphorées sont plus importantes.
L’axe 2 (15,35% de la variance) est positivement corrélé à la biomasse de F.
paniculata (0,44), et négativement avec l’IP (-0,36) et l’IN (-0,28) (Tab. 3). Tout comme
l’axe 1, cet axe peut être lié à la fertilité du milieu en fonction de la production de biomasse
de F. paniculata. Cependant, il met en évidence la variabilité entre parcelles d’une même
classe.
12
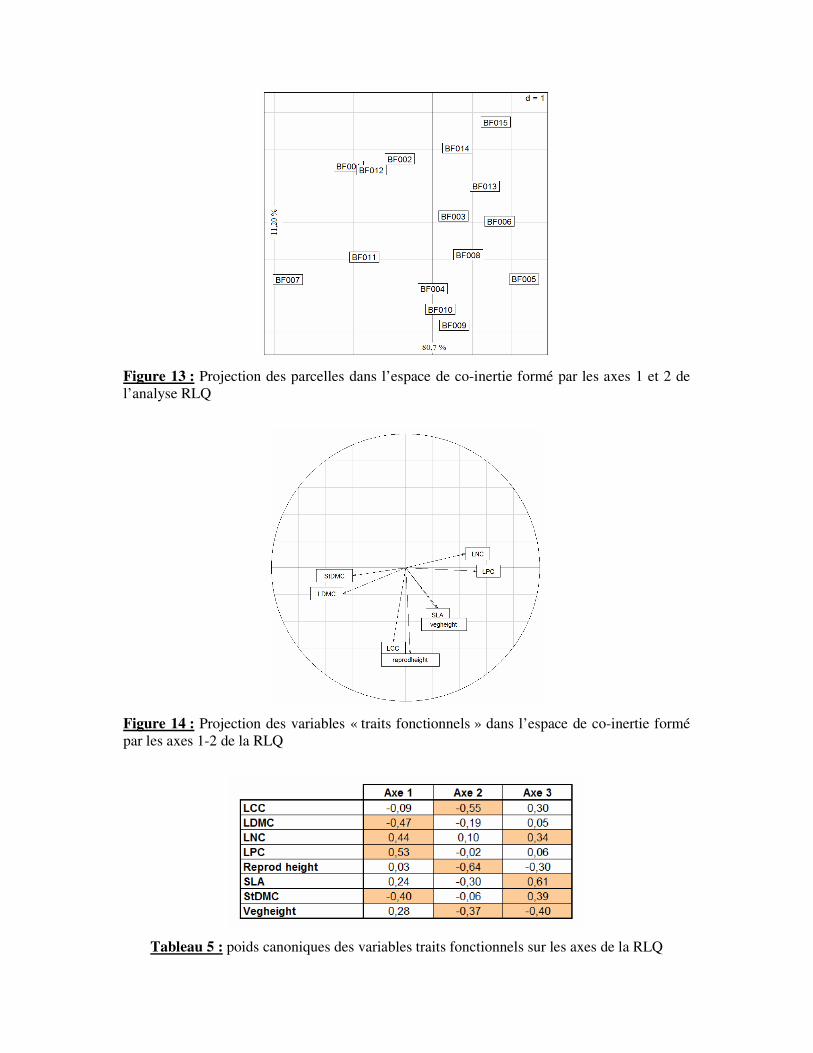
Figure 13 : Projection des parcelles dans l’espace de co-inertie formé par les axes 1 et 2 de
l’analyse RLQ
Figure 14 : Projection des variables « traits fonctionnels » dans l’espace de co-inertie formé
par les axes 1-2 de la RLQ
Tableau 5 : poids canoniques des variables traits fonctionnels sur les axes de la RLQ

Enfin, l’axe 3 (11,7% de la variance) est corrélé à l’abondance de la litière (-0,48) au sein de
la prairie, à l’IK (-0,48), l’IN (-0,39) et au rapport C/N (0,43) (Tab.3). Cet axe met en
contraste des prairies à litière importante et/ou à forte fertilité avec des prairies plus pauvres.
D’une manière générale, cette projection (Fig. 11) permet de dissocier les classes
d’âges 5–20 ans et supérieur à 100 ans, des classes 20-50 ans et 50-100 ans. Les prairies
fauchées sont toutefois proches de la classe d’âge la plus ancienne.
La significativité et l’amplitude des relations mises en évidences entre variables
environnementales sont testées au travers de régressions linéaires (Tab. 4). Le coefficient de
corrélation critique pris en compte est à R2>0,4. Tout d’abord, la productivité de la prairie est
corrélée positivement aux ressources azotées (R2=0,73). De plus, la richesse spécifique des
prairies est positivement corrélé à l’importance des ressources en IP (R2=0,77) et IK
(R2=0,75) mais diminue en fonction de l’abondance (R
2=0,68) et de la production de
biomasse (R2=0,43) de F. paniculata. Cette dernière est négativement corrélée à la proportion
de légumineuse (R2=0,80) dans la composition floristique et faiblement corrélée à
l’abondance de la litière dans l’écosystème (R2=0,42). Laquelle est négativement corrélée au
pH (R2=0,91), de même qu’avec l’IK (R
2=0,63) et l’IP (R
2=0,74). Enfin, l’IP et l’IK sont
corrélés positivement entre eux (R2=0,92) et négativement à l’abondance de F. paniculata
(respectivement R2=0,41 et R
2=0,64).
Ainsi, les caractéristiques des classes d’âges sont mises en avant par la projection des
corrélations des variables environnementales (Fig. 12). Les prairies fauchées sont
caractérisées par une richesse spécifique élevée, des ressources (phosphore et potasse) dans le
sol importantes et par conséquent une abondance et production de biomasse faibles de F.
paniculata. Les parcelles dont l’arrêt de la fauche est le plus récent (5-20 ans) sont
caractérisées par une productivité très élevée et un indice de nutrition azotée fort. Les prairies
de la classe d’âge d’abandon suivante (20-50 ans) sont caractérisées par une abondance et une
production de biomasse élevées de F. paniculata. Ces prairies présentent également la plus
faible richesse spécifique. Les prairies non fauchées depuis 50-100 ans ont une litière
importante, et par conséquent un pH acide, mais une ressource azotée importante. Enfin, les
prairies dont l’âge d’abandon est le plus ancien possèdent les mêmes caractéristiques que les
prairies fauchées. Cependant, elles possèdent une part plus importante de légumineuses.
13
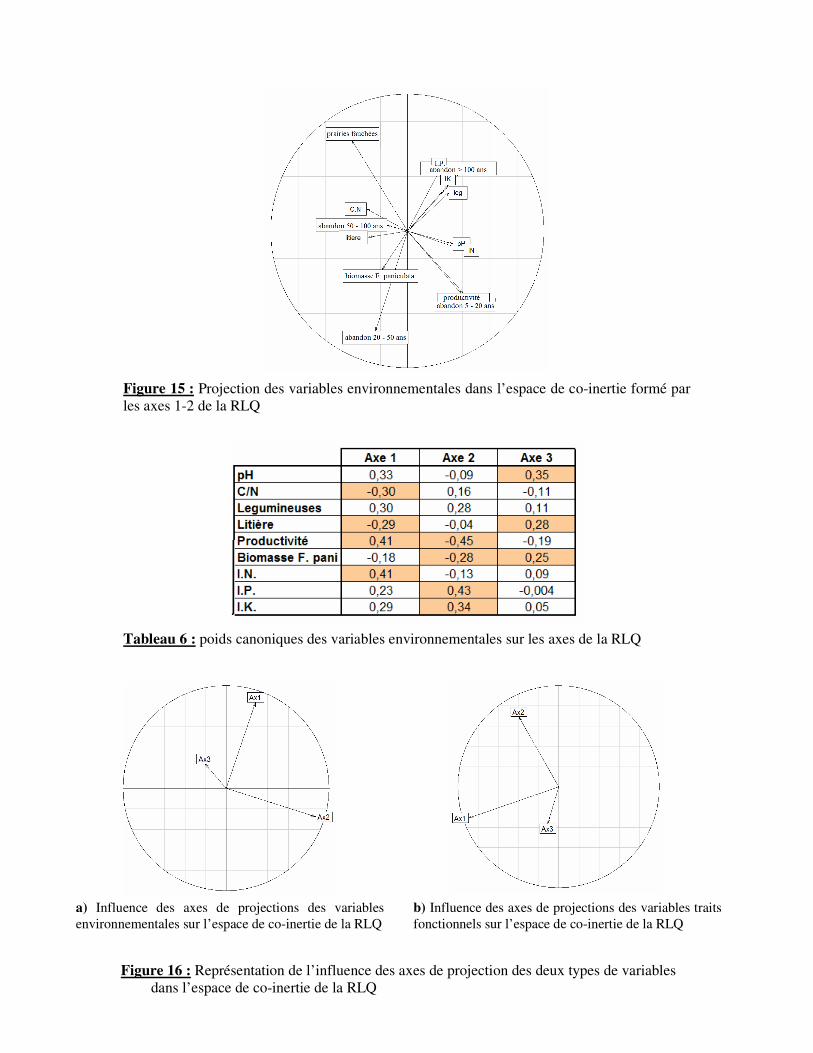
Figure 15 : Projection des variables environnementales dans l’espace de co-inertie formé par
les axes 1-2 de la RLQ
Tableau 6 : poids canoniques des variables environnementales sur les axes de la RLQ
a) Influence des axes de projections des variables
environnementales sur l’espace de co-inertie de la RLQ
b) Influence des axes de projections des variables traits
fonctionnels sur l’espace de co-inertie de la RLQ
Figure 16 : Représentation de l’influence des axes de projection des deux types de variables
dans l’espace de co-inertie de la RLQ

C) Traits fonctionnels des plantes
L’analyse RLQ a permis une projection des parcelles (Fig.13) en tenant compte des
variables environnementales et des caractéristiques fonctionnelles des communautés
végétales. Cette distribution est semblable à celle de l’AFC réalisée à partir des compositions
floristiques. L’axe 1 (80,7% de l’inertie) et l’axe 2 (11,20% de l’inertie) de la RLQ extraient
donc la quasi-totalité (92%) de la variabilité totale. Un test de Monte Carlo réalisé sur la RLQ
révèle que la relation statistique entre les variables environnementales et les variables “traits
fonctionnels” est significative (p<0,05). Il existe donc une corrélation entre les axes de projection
des variables environnementales et des variables “traits fonctionnels”. Il convient de connaître
l’influence de ces deux types de variables sur l’ordination de la RLQ.
La projection des variables traits fonctionnels dans l’espace de co-inertie de la RLQ
laisse entrevoir structuration cohérente (Fig. 14). L’axe 1 est influencé positivement par les
traits LNC (0,44) et LPC (0,53) puis négativement par StDMC (-0,40) et LDMC (-0,47) (Tab.
5). L’axe correspond donc aux stratégies de gestions des ressources par les plantes. En effet,
des valeurs élevées de LDMC et StDMC sont caractéristiques des plantes dites conservatrices
et par conséquent une croissance lente. A l’opposé, des valeurs élevés de LNC et LPC sont
caractéristiques des plantes à croissance rapide. L’axe 2 est influencé par la taille de
reproduction (-0,64), la taille végétative (-0,37), le LCC (-0,55) et la SLA (-0,30). Cet axe
peut être lié à la stature et l’architecture des plantes. Il oppose ainsi des plantes à fortes
statures et à étalement latéral important à des plantes de plus faibles statures.
Une projection des variables environnementales sur l’espace de co-inertie de la RLQ
est effectuée selon le même principe (Fig. 15). L’axe 1 est influé par la litière (-0.29), le C/N
(-0.30), la productivité (0.41) et l’IN (0.41) (Tab. 6). Cet axe est corrélé à la ressource en
azote du milieu et à l’abondance de la litière. En effet, il met en contraste entre les prairies à
forte litière et faible ressource azotée des prairies à productivité et fertilité plus élevée. L’axe
2 est influencé par la productivité (-0.45), la production de biomasse de F. paniculata (-0.28),
l’IP (0.43) et l’IK (0.34). Cet axe dissocie les prairies à forte production de biomasse de F.
paniculata des prairies à forte fertilité.
La distribution de la RLQ peut donc être expliqué par l’influence de ces deux types de
variables. L’axe 1 de la RLQ est négativement corrélée à l’axe 1 des variables traits et
positivement corrélé à l’axe 2 des variables environnementales (Fig. 16 & Tab. 7). De même,
l’axe 2 de la RLQ est positivement corrélé à l’axe 2 des variables “traits fonctionnels” et à
l’axe 1 des variables environnementales. Ainsi, les plantes dites conservatrice (StDMC &
LDMC) sont corrélées aux prairies à faible fertilité et productions de biomasse de F.
14
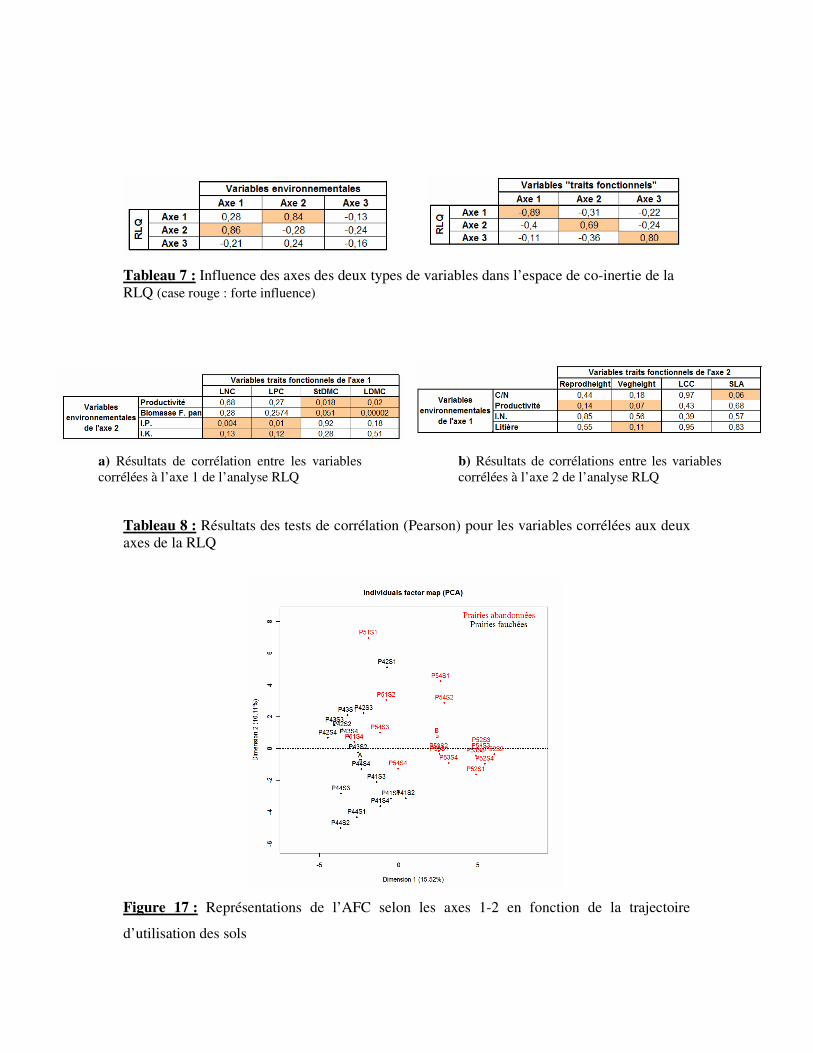
Tableau 7 : Influence des axes des deux types de variables dans l’espace de co-inertie de la
RLQ (case rouge : forte influence)
Tableau 8 : Résultats des tests de corrélation (Pearson) pour les variables corrélées aux deux
axes de la RLQ
Figure 17 : Représentations de l’AFC selon les axes 1-2 en fonction de la trajectoire
d’utilisation des sols
a) Résultats de corrélation entre les variables
corrélées à l’axe 1 de l’analyse RLQ
b) Résultats de corrélations entre les variables
corrélées à l’axe 2 de l’analyse RLQ

paniculata élevée. Les plantes à croissance rapide sont corrélées aux prairies fertiles et à
faible productivité de biomasse de F. paniculata. L’architecture et l’étalement importants de
certaines espèces sont également corrélés aux prairies aux ressources azotées importantes et à
forte productivité.
Les hypothèses de liens entre les variables de milieu et les traits fonctionnels sont
testées à l’aide de tests de corrélations (Tab. 8). Les résultats sont pris pour une valeur de
risque de p<0,15. Ainsi, les traits LDMC et StDMC sont corrélées négativement à la
productivité de la prairie (respectivement 0,02 et 0,02) et à la biomasse de F. paniculata
(respectivement 2 .10-5 et 0,05). Les traits LNC (0,004) et LPC (0,01) sont corrélés
négativement à l’IP. La productivité est corrélée positivement à la taille végétative (0,14) et
reproductive des plantes (0,07). La litière semble également corrélée positivement à la taille
végétative (0,1) des plantes.
2°) Dynamique de végétation en réponse à la fauche/abandon
L’AFC montre clairement une différence de composition floristique entre les parcelles
en prairies abandonnées et en prairies fauchées (25,63% de la variance totale ; Fig. 17). L’axe
1 de l’AFC explique (15,52%) cette distribution selon la pratique de la fauche. Les prairies
abandonnées sont caractérisées par Dactylis glomarata, Laserpitium latifolium, Trisetum
flavescens, Agrostis capillaris et Trifolium alpestre. Ces espèces caractéristiques des prairies
subalpines sur lesquelles la fauche n’est pas pratiquée. La présence de Trifolium alpestre est
également un indicateur de zone à hauteur de couvert plus élevé. Les prairies fauchées sont
caractérisées majoritairement par Carex sempervirens, Festuca laevigata, Sesleria cerulea,
Thymus serpyllum, Trifolium alpinum et Trifolium montanum. Ce sont des espèces de zones
héliophiles à faible couvert végétal et de sols secs caractéristiques des pelouses de fauches
subalpines. Deux espèces, à stature plus haute, Hieracium umbellatum et Centaurea uniflora
sont également abondantes dans ces prairies. Elles représentent des zones à faible densité et
couvert végétal. F. paniculata est présente dans les deux types de prairies, mais en abondance
différente. Ainsi, elle représente en moyenne 3,7% de la composition des parcelles en prairies
fauchées et elle est deux fois plus abondante dans les zones abandonnées.
Cependant, une analyse plus fine de cette distribution selon les différents traitements
permet d’observer un début de dynamique (Fig. 18). Pour les sous-parcelles en prairies
fauchées (Fig. 18.a), l’axe 1 (21,1%) met sensiblement en contraste les parcelles subissant
une coupe et enlèvement de litière avec les parcelles ne subissant pas de coupe et dont la
15
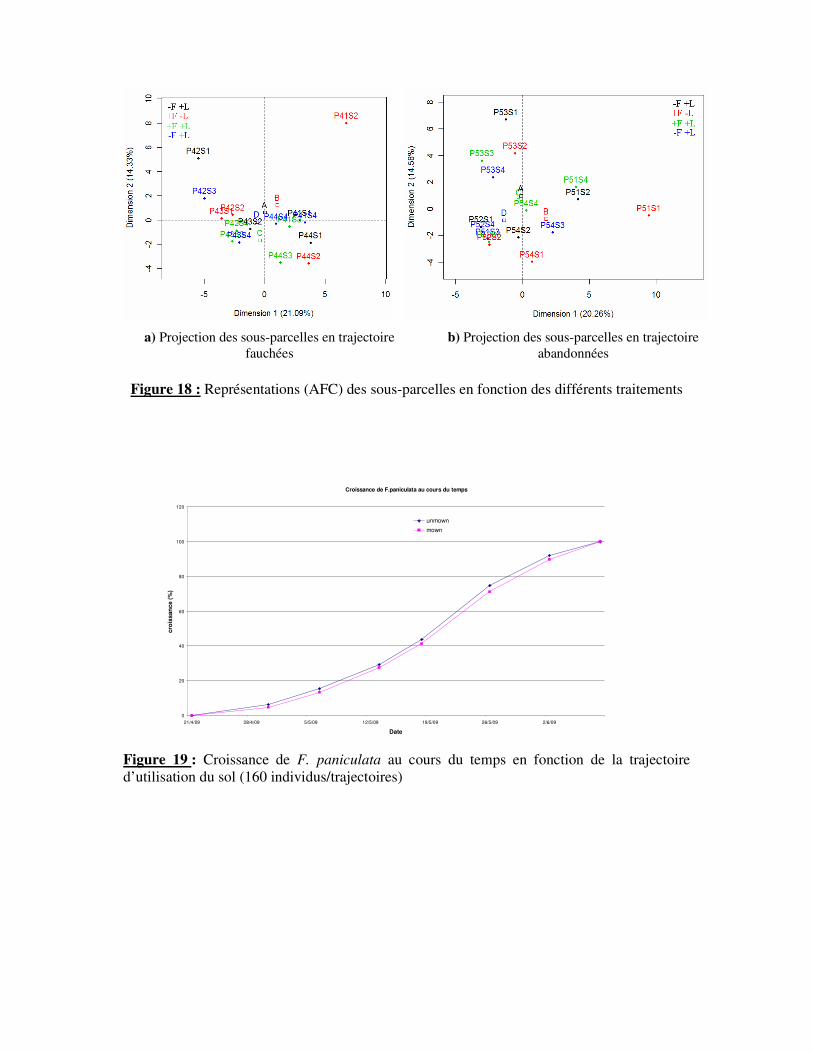
Figure 18 : Représentations (AFC) des sous-parcelles en fonction des différents traitements
Croissance de F.paniculata au cours du temps
0
20
40
60
80
100
120
21/4/09 28/4/09 5/5/09 12/5/09 19/5/09 26/5/09 2/6/09
Date
cro
issan
ce (
%)
unmown
mown
Figure 19 : Croissance de F. paniculata au cours du temps en fonction de la trajectoire
d’utilisation du sol (160 individus/trajectoires)
a) Projection des sous-parcelles en trajectoire
fauchées
b) Projection des sous-parcelles en trajectoire
abandonnées

litière est laissée. On retrouve une distribution propre à la trajectoire d’utilisation du sol.
L’axe 2 (14,33%) montre une différence entre les parcelles soumis aux traitements coupe
mais dont la litière est retirée ou laissée. En effet, des espèces à faibles statures comme
Gallium pugilum ou Hieracium umbellatum sont présents lorsque la litière est importante (à
une exception Asphodelus albus). A l’inverse, des espèces à hautes stature comme Leontodon
hispidus, Crepis pyrenaica, Crepis conyzifolia ou Hieracium villosum sont présente dans les
parcelles à peu de litière. Pour les sous-parcelles en prairies abandonnées, les deux facteurs
sont moins mis en évidence (Fig. 18.b). En effet, l’axe 1 (20,3%) est influencé par les sous-
parelles soumises à la coupe et à l’enlèvement de biomasse et par les parcelles non coupée et
où la litière n’a pas été prélevée. Il met en contraste une combinaison de facteurs,
correspondant à nouveau aux deux trajectoires d’utilisation des sols abordées dans cette étude.
L’axe 2 (14,6%) ne met pas en évidence de patron de distribution en fonction des deux
facteurs.
3°) Influence de la fauche sur la performance de F. paniculata
La représentation graphique du suivi de la croissance de F. paniculata prend la forme
d’une courbe logistique (Fig. 19). La croissance est donc caractérisée par un “plateau”
terminal marquant un ralentissement de celle-ci. Ce plateau fut atteint le 8 Juin 2009. Cette
dynamique de croissance similaire selon la trajectoire d’utilisation du sol, nous permet
d’analyser de façon conjointe l’impact de la fauche sur F. paniculata. La date choisie est au
début du « plateau » de croissance, auquel cas les individus peuvent alors être considérés
comme entièrement formés.
Une analyse en split-plot révèle une influence nettement significative (p<0,001) de la
trajectoire d’utilisation du sol (fauche/abandon) sur la hauteur des individus de F. paniculata
(Tab. 9.a). Le détail de cette analyse montre que la hauteur de F.paniculata en parcelles
fauchées est inférieure par rapport aux individus des parcelles abandonnées (Tab. 9.b). Ainsi,
en moyenne les individus en parcelles non fauchées mesurent 34,98cm de hauteur (± 0,88cm),
soit 11,6% de plus que pour les individus des parcelles fauchées. La pratique de la fauche
affecte donc la croissance de F. paniculata.
Cette démarche expérimentale permet d’étudier séparément les deux composantes de
la fauche. La coupe de biomasse est le facteur influant de façon nettement significative
(p<0,001) la stature de F. paniculata (Tab. 9.a). En effet, le facteur litière ne présente pas
d’effet significatif (p>0,05). Dans la pratique de la fauche, seule la coupe de biomasse semble
16
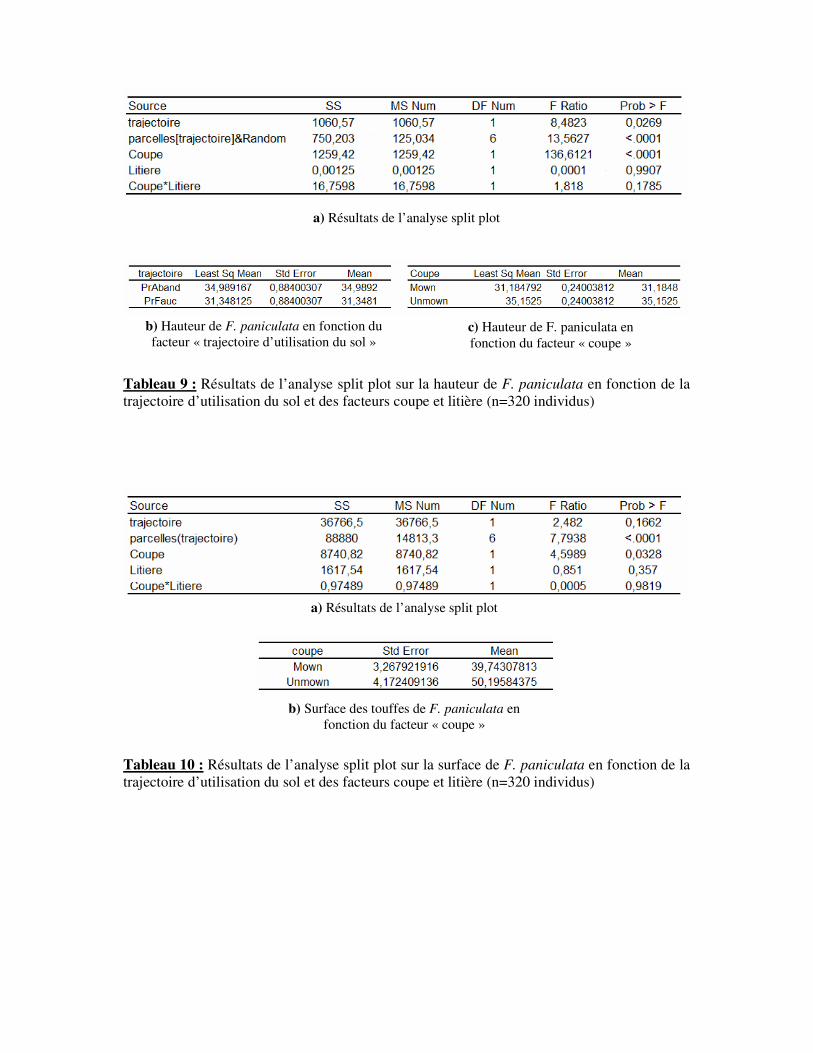
a) Résultats de l’analyse split plot
Tableau 9 : Résultats de l’analyse split plot sur la hauteur de F. paniculata en fonction de la
trajectoire d’utilisation du sol et des facteurs coupe et litière (n=320 individus)
Tableau 10 : Résultats de l’analyse split plot sur la surface de F. paniculata en fonction de la
trajectoire d’utilisation du sol et des facteurs coupe et litière (n=320 individus)
b) Hauteur de F. paniculata en fonction du
facteur « trajectoire d’utilisation du sol » c) Hauteur de F. paniculata en
fonction du facteur « coupe »
a) Résultats de l’analyse split plot
b) Surface des touffes de F. paniculata en
fonction du facteur « coupe »

affecter la croissance de F. paniculata. D’une manière générale, les individus subissant la
coupe ont une hauteur moyenne de 31,2cm (± 0,24 cm) soit 11,3% de moins que les individus
n’ayant pas leur partie aérienne coupée (hauteur moyenne = 35,15 cm ± 0,24 cm) (Tab. 9.c).
Une approche identique, réalisée sur la surface des touffes de F. paniculata, ne révèle pas une
différence significative selon la trajectoire d’utilisation (p>0,05) (Tab. 10.a). En revanche, il
existe une différence de surface des individus entre les différents traitements (p<0.0001). De
même que pour la hauteur, la surface est uniquement influencé par la coupe de biomasse
(p<0,05) (Tab. 10.a).En effet, les individus affectés par la coupe de biomasse possèdent une
superficie de 39,74 cm2 (± 3,72 cm
2), alors que les individus ne subissant pas cette
perturbation ont une superficie supérieure de 26,3% (superficie moyenne = 50,20 cm2 ± 4,17
cm2) (Tab. 10.b).
La fauche, du fait de la coupe de biomasse, affecte F. paniculata. Il convient d’évaluer la
rapidité de réponse des individus à ce stress. Une comparaison des différentes sous-parcelles
analogues permet de voir l’évolution après cinq années de pratique de ces facteurs (Tab. 11 &
Tab. 12). En raison du faible rôle de la litière démontrée précédemment, celle-ci n’est donc
pas considérée comme facteur pour cette analyse. L’étude de la surface n’ayant rien révélée
de significatif (Tab. 12), seule la hauteur est prise en compte pour cette approche (Tab. 11).
En effet, une comparaison des parcelles analogues entre prairie fauchée et abandonnée révèle
une différence de hauteur significative (p<0,0001). La comparaison au sein d’une même
trajectoire d’utilisation de sol selon la pratique ou non de la coupe révèle également une
différence de hauteur entre les individus (p < 0,0001). Comme démontré précédemment, la
pratique de la coupe affecte les individus de F. paniculata. Cependant, après cinq années de
pratique des traitements, la taille des individus pour un même traitement au sein des deux
trajectoires d’utilisation du sol n’est pas similaire. En revanche, un test de Student réalisé
entre les hauteurs d’individus des sous parcelles non coupées en prairies fauchées et les
hauteurs des individus des parcelles coupées en prairies abandonnées révèle une hauteur
moyenne identique.(p>0,05). Cette évolution de la taille peut être quantifiée. La comparaison
de hauteurs moyennes (Tab. 13) montre que les individus en parcelles fauchées ont une
hauteur moyenne supérieure de 12% suite à l’arrêt de la coupe. De même, les individus en
prairies abandonnées ont une hauteur moyenne qui a diminuée de 11,8% suite à la pratique de
la coupe. La reprise de la fauche ou l’abandon semble donc affecter la hauteur de
F.paniculata au même rythme. Après cinq ans de pratiques ou abandon de la fauche, la taille
moyenne des individus est affectée de 12%. Une comparaison des hauteurs des individus
entre parcelles témoins montre que les individus en prairies fauchées sont 20,5%
17

Tableau 11 : Résultats des tests de comparaisons de hauteur moyennes (test de Student) en
fonction des trajectoires d’utilisation du sol et de la pratique/abandon de la coupe
(n = 80 individus/modalités)
Tableau 12 : Résultats des tests de comparaisons des surfaces moyennes (test de Student) en
fonction des trajectoires d’utilisation du sol et de la pratique/abandon de la coupe
(n = 80 individus/modalités)
Tableau 13 : Hauteur moyennes de F. paniculata en fonction de la trajectoire d’utilisation du
sol et de la pratique/abandon de la coupe (n = 80 individus/modalités)

plus petits que ceux des prairies abandonnées. Ainsi, la durée pour laquelle F. paniculata
passerait de la stature élevée des prairies abandonnées à un état équivalent des prairies de
fauche peut être estimée. En supposant que ses réponses au stress soient linéaires, un peu
moins d’une dizaine d’année de fauche serait nécessaire.
Discussion
1°) Dynamique des pelouses subalpines à F. paniculata et impact sur
les propriétés écosystémiques
L’étude des traits fonctionnels des communautés révèle des stratégies d’acquisition
des ressources contrastées le long du gradient d’abandon. Toutefois ces stratégies
d’adaptation ne sont pas associées linéairement au gradient. Des espèces à fortes valeurs de
LDMC et StDMC, caractéristiques des espèces à stratégie de conservation des ressources
(Grime, 1977 ; Cunningham et al., 1999 ; Shippers & Olff, 2000 ; Cornelissen et al., 2003),
sont présentes dans les parcelles abandonnées depuis cinq à cent ans. A contrario, des espèces
déterminées par de fortes valeurs de LPC et LNC, caractéristiques des espèces à stratégie
d’exploitation des ressources, sont observées au sein des parcelles fauchées ou abandonnées
depuis plus de cent ans. Par ailleurs, s’opposent généralement des plantes à forte hauteur
végétative et reproductive dans les zones fauchée ou abandonnées depuis plus de 100 ans, à
des espèces à stature plus réduite dans les zones abandonnées (5 à 100 ans), reflètant dans
une certaine mesure l’intensité de la compétition pour la lumière (Weiher et al., 1999).
Festuca paniculata, présente également une dynamique non linéaire le long de ce gradient
temporel. Son recouvrement au sein des communautés en fonction des classes d’âges révèle
une répartition « gaussienne ». En effet, son abondance augmente jusqu’à cinquante ans
d’abandon puis semble diminuer au sein de la classe supérieure (50-100 ans), pour enfin être
très faible au sein des communautés abandonnées depuis plus de cent ans.
L’abandon semble donc être prolifique à F. paniculata sur une certaine durée.
L’analyse a permis de mettre en évidence le lien existant entre cette espèce et les stratégies
d’acquisition des nutriments des communautés végétales associées. Le rôle de la litière en lien
avec ce phénomène n’est toutefois pas démontré. En effet, la biomasse de la litière suit une
tendance « gaussienne » avec toutefois un décalage temporel par rapport à la biomasse de F.
paniculata. Au sein de la classe d’âge 50-100 an d’abandon, elle semble déjà avoir régressé
alors que la biomasse de litière est encore très importante.
18


La litière influence la régénération et la croissance des autres espèces de la
communauté végétale mais affecte aussi le recyclage des nutriments via sa quantité et sa
qualité (Güssewell & Bollens, 2003). L’analyse corrélative révèle effectivement un lien entre
les espèces de faibles statures et l’abondance de la litière. Une étude récente établit que la
litière favorise les espèces à petites graines (Viard-Cretat, 2008). Or, la masse de graine est
souvent reliée à la taille des plantes (Moles et al., 2004), bien que ce ne soit pas toujours le
cas (Westoby, 1998). Ainsi, les espèces conservatrices citées précédemment présentent de
faibles tailles végétatives, ainsi qu’une surface foliaire (SLA) relativement faible. Par ailleurs,
les analyses des stratégies fonctionnelles observées avec les variables environnementales,
établissent une corrélation négative entre la disponibilité phosphorée et potassique du sol et la
biomasse de F. Paniculata. Des résultats similaires ont été observés concernant les ressources
azotées (Robson et al., 2007). L’azote et le phosphore sont des éléments importants dans la
croissance des plantes et son exploitation peut permettre une croissance rapide des espèces
compétitrices (Wright & al., 2001).
Ainsi, les traits fonctionnels de réponse des espèces témoignent de leurs stratégies
adaptatives au milieu (Lavorel et al., 1997). Les stratégies d’acquisition des ressources sont
un indicateur des réserves nutritives du sol (Lavorel et al., 2004) et l’architecture des plantes
illustre une compétition pour la ressource lumineuse (Weiher et al., 1999). Cette étude montre
l’importance du rôle des ressources dans la dynamique de la végétation. Cependant, si un
processus est décrit, cela ne permet pas sa compréhension. En effet, un fossé d’un siècle et
demi dans le gradient temporel utilisé, ne permet pas de comprendre la très nette diminution
du recouvrement de F. paniculata au sein des prairies les plus anciennement abandonnées. Un
doute sur la véritable utilisation (labourage) de ces prairies dans le passé peut être soulevé.
Toujours est-il que les queyrellins semblent soumis à une dynamique d’embroussaillement
ralentie. Il est tout à fait probable que ces prairies puissent évoluer vers des formations
forestières, stade climacique de cet étage subalpin (Ozenda, 1985). Cependant, peu de
données expérimentales sont disponibles de nos jours pour savoir si l’installation de ligneux
est possible dans les couverts denses à Festuca paniculata (Albert, 2006).
Par ailleurs, l’interprétation de ces résultats doit être modérée. Si des caractéristiques
propres aux classes d’abandon sont mises en avant, les résultats montrent une forte
hétérogénéité intra-classe d’âge. Ainsi, des parcelles à proximité de zones humides ont une
composition floristique différente de ses consoeurs. De même, les parcelles à fortes pentes
(e.g. BF004), se différencient nettement au sein de leur classe d’age. Cette forte variabilité
19


souligne donc l’importance de prendre en compte l’état initial de la communauté végétale
pour analyser son évolution plusieurs décennies après.
2°) Réponse de la végétation aux changements d’utilisation des terres
L’arrêt de la fauche modifie donc la dynamique de végétations dans les prairies de
fauche subalpines en favorisant F. paniculata. Aussi, dans une perspective de gestion, il est
important de préciser dans quelle mesure la fauche, c'est-à-dire la coupe et/ou le retrait de la
biomasse épigée affecte la composition des communautés végétales et la croissance de F.
paniculata.
Les différents traitements expérimentaux réalisés depuis cinq ans révèlent une
discrétisation des communautés selon les deux trajectoires de changement d’utilisation des
terres (fauchée v.s. abandonnée). Les communautés subissant des traitements analogues (e.g.
traitement coupe/litière) au sein des deux trajectoires ne sont pas caractérisées par la même
composition floristique. En effet, la réponse de la végétation semble trop lente et ne permet
pas d’observer les conséquences de ces traitements sur un pas de temps court (Landsberg,
1999). En revanche, une dynamique est observée entre les différents traitements au sein d’une
même trajectoire. Ainsi, après cinq années d’abandon des communautés végétales
initialement fauchées (trajectoire fauchée) des espèces conservatrices propices aux milieux
non fauchées se sont sensiblement accrues (Gallium pugilum, Gallium verrum, Trisetum
flavescens). De plus, la litière semble avoir une faible influence sur ces communautés
indépendamment de la coupe (espèce à stature plus réduite). En effet, son retrait permet la
croissance d’espèces à hautes statures (Bakker, 1989) alors que son accumulation favorise
plutôt des espèces de plus petites tailles (Viard-Cretat, 2008). Ceci confirme les résultats déjà
observés précédemment et souligne un rôle de la litière dans la dynamique des queyrellins.
Dans les communautés situées au sein de la trajectoire abandonnée, seul le traitement
« coupe » associé au « retrait litière » affecte la structuration des communautés végétales (e.g.
Crepis conyzifolia). En effet, l’étude du fonctionnement des queyrellins a montré que
l’abandon favorisait des espèces conservatrices. Ces espèces sont généralement caractérisées
par d’importante réserves, permettant, au moins à cour terme, une meilleure tolérance en
réponse à d’éventuelles perturbations (Jouglet & Dorée, 1987). L’étude de ce protocole
confirme donc à nouveau l’importance des ressources sur la dynamique végétales des prairies
à F. paniculata.
20


Le fonctionnement des écosystèmes est déterminé, en grande partie, par les valeurs de
traits des espèces dominantes de la communauté (Garnier et al., 2004). F. paniculata est
l’espèce par excellence des prés de fauche subalpin, et sa réponse à ces deux types de
perturbations (coupe et litière) a une incidence sur les espèces compagnes. En effet, elle est
très peu tolérante à la coupe de biomasse. Après cinq années de traitement sa hauteur s’en
trouve affectée et son recouvrement a fortement diminué (entre 24% et 13%). Ainsi, d’après
les relations établies dans l’étude de la dynamique des queyrellins, la production de biomasse
de F. paniculata et l’abondance de la litière diminueront. De ce fait, ceci permet
l’augmentation de l’abondance des espèces compétitrices observée précédemment au sein des
communautés soumises à la coupe.
La coupe affecte les espèces conservatrices (comme F. paniculata) (Bakker, 1989) et
favorise des espèces à stratégie d’exploitation des ressources Par ailleurs, la litière tend à
limiter le nombre de niche de régénération et limite donc de fait la diversité spécifique de la
prairie (Jones & Hayes, 1999). F. paniculata modifie donc l’environnement physique des
écosystèmes, notemment au travers les flux de matière et d’énergie. Ceci, affecte donc la
composition des comunautés. Cette espèce apparaît donc comme une espèce ingénieur (Jones
et al., 1994) de la dynamique végétale de ces écosystèmes subalpins.
3°) Perspectives de gestion intégrée des prairies de fauche subalpines
De profonds changements amorcés depuis plus de cinquante ans menacent la pérennité
des prairies subalpines (Quétier, 2006 ; Sebastià, 2004). La restauration des prairies de fauche
est donc un des enjeux écologiques des zones de montagne (Kahmen et al., 2002). La
prolifération de F. paniculata engendre une modification des processus écosystémique et une
diminution de la diversité végétale. De plus, la production fourragère de ces prairies est de
mauvaise qualité, et les troupeaux mis en pâture dans ces milieux délaissent cette graminée
(CEMAGREF, 1981). Les difficultés d’utilisation de ces milieux ajoutés à la baisse générale
de l’agriculture alpine, les amènent donc à être abandonnés.
Le pastoralisme est considéré comme un outil important dans la gestion des habitats
(Watkinson & Ormerod, 2001). Des études sur la gestion pastorales des queyrellins ont
permis d’amorcer une réflexion sur un mode gestion approprié (Jouglet & Dorée, 1987 ;
Quiblier & Senn, 2004). Un gardiennage serré et précoce semble plus approprié pour une
diminution du recouvrement de F. paniculata et de la litière (Parc National des Ecrins, 2006).
21


Néanmoins, cette démarche engendre une charge pastorale élevée, capable de provoquer une
diminution de la richesse spécifique (Bouchard et al., 2003). Un compromis entre l’intensité
du pâturage et la préservation de la richesse floristique reste donc encore à étudier.
De nombreuses études démontrent l’importance de considérer le pâturage comme une
stratégie de restauration des prairies (WallisDeVries et al., 1998 ; Barbaro et al.,2001 ;
Pykälä, 2003). Cependant, une action uniformisée ne semble pas toujours adéquate. En effet,
une gestion appropriée de ces milieux doit considérer l’importance de l’histoire et de la
composition de la communauté (Grime et al., 2000 ; Britton et al., 2001). Ainsi, Fischer et
Wipf (2002) ont démontré qu’une gestion par la pratique de la fauche s’avère être plus
efficace pour les prairies subalpines.
Cette étude a également montré que la pratique de la fauche affecte la composition
floristique des queyrellins abandonnées et permet de réduire efficacement l’abondance de F.
paniculata. En effet, l’étude d’impact réalisée au court de cette étude révèle une réponse
rapide de F. paniculata à cette perturbation au travers d’une diminution du recouvrement et
de production de biomasse. Le surpâturage est également une perturbation adaptée pour
contenir F. paniculata. Cependant, il entraîne une nitrification du sol au travers des
excréments et par conséquent un changement marqué de la composition floristique des zones
pâturées (Leroux et al. 2003, Güssewell & Bollens 2003) et une réduction de la diversité
(Bouchard et al., 2003). La fauche apparaît donc comme un mode de gestion plus approprié,
face aux enjeux floristiques et économiques du site, pour la restauration des anciennes prairies
de fauche subalpines.
Cependant, une longue période d’abandon, des conditions édaphiques favorisant la
germination est un point essentiel du succès de la restauration de ces milieux (Pärtel et al.,
1998). Des études démontrent que le pâturage est un excellent vecteur de création de trouées
favorable à la germination (Bakker, 1989 ; Jones & Hayes, 1999). Le bétail, et
particulièrement les ovins, est également un excellent vecteur de dispersion et colonisation
(Fischer et al., 1996 ; Poschlod et al., 1998). En effet, le pâturage augmente l’hétérogénéité
des ressources (i.e., une structure hétérogène spatiale avec des zones riche ou pauvres en
nutriments) et permet une cœxistence d’espèces avec des assimilations de ressources
différentes (de Bello et al., 2006).
La prise en compte des caractéristiques de ces communautés subalpines laisse donc
entrevoir la possibilité d’une gestion au travers de la fauche et d’un pâturage peu intensif. En
effet, des études récentes sur les prairies subalpines ont démontrés qu’un pâturage de faible
intensité accompagné d’une fauche sont des alternatives possibles au surpâturage (Bakker,
22


1989 ; Rudmann-Maurer et al., 2008). Cette alternative semble donc correspondre aux enjeux
attendus de la restauration des prairies de fauches subalpines. Cependant, la courte durée de
végétation rend difficile la mise en place d’un tel mode de gestion. Car à cette altitude, ne
peut être effectuée qu’une seule fauche par an. Traditionnellement, les queyrellins sont
fauchées tardivement (mi-fin Août), dans un objectif d’une récolte fourragère importante. Or,
à cette date, les plantes commencent à rentrer en sénescence, ce qui pourrait limiter les effets
mécaniques de la fauche sur les espèces. Une fauche précoce pourrait être un compromis.
F.paniculata est une des espèces les plus précoces des prairies subalpines (Quétier, 2006). Le
suivi de croissance a révélé également une fin de croissance précoce (début Juin), alors
qu’habituellement le pic de biomasse des prairies n’est atteint que début juillet (Lavorel,
comm. pers.). Une action précoce pourrait donc affecter la performance de F. paniculata et
ainsi réduire son recouvrement. Un pâturage tardif semblerait plus approprié, et justifié, car il
permettrait alors la création de niche de régénération et induirait une dispersion des graines.
Cette stratégie de restauration soulève de nombreuses questions d’approfondissement.
Il conviendrait de quantifier l’impact d’une fauche précoce sur la performance de F.
paniculata. De même, une étude de la phénologie des espèces de ces prairies permettrait de
mieux cibler les enjeux de préservation au cours de la saison. La prise en compte de
l’importance des espèces précoces et tardives permettrait également d’orienter les stratégies
de gestion. De même, évaluer un diagnostic pastoral au pic de biomasse avec une fauche
précoce pourait fournir des informations. Enfin, il convient d’évaluer la perte du rendement
fourrager nécessaire à la viabilité de l’exploitation. Cette stratégie pourrait s’inscrire dans le
cadre des mesures agro-environnementales, justifiées par l’appartenance des prairies de
fauche du site au réseau Natura 2000.
Conclusion & perspectives
Ce travail s’inscrit dans la lignée des nombreuses études menées sur l’impact du
changement d’utilisation des terres ces dernières décennies. Cette déprise agricole affecte les
prairies de fauche subalpines par l’abondance de Festuca paniculata. L’analyse de cette
problématique à plusieurs échelles spatiales et temporelles au travers des traits fonctionnels
des plantes a permis d’apporter des éléments de réflexion sur la compréhension des règles
d’assemblages de ces communautés.
La dynamique d’abandon des prairies à F. paniculata est essentiellement engendrée
par une modification des ressources disponibles dans le sol. La production de biomasse élevée
23


de F. paniculata et la faible dégradabilité de sa litière sélectionnent les espèces végétales
entraînant une restructuration importante des communautés végétales. Ce processus est
observé sur une durée de plusieurs décennies bien que les conditions initiales (édaphique,
topographique) semblent influencer en profondeur cette dynamique. L’étude à long terme de
cette dynamique a par ailleurs permis de mettre en évidence une possible évolution de ses
communautés vers une reprise de l’enfrichement. Ces profondes mutations structurales
engendrées par F. paniculata permettent de la considérer comme une espèce ingénieur des
écosystèmes subalpins.
La diminution de la diversité végétale au sein des queyrellins est la problématique
préoccupante des gestionnaires des Alpes centrales françaises. Cette démarche fut menée en
étroite collaboration avec des gestionnaires (Centre d’Etude de Réalisation Pastorale Alpes
Méditerranée, Parc National des Ecrins et du Mercantour) au travers des ateliers de travail.
Les perspectives des gestions agro-pastorales induites par de nombreuses études vont amener
ainsi à l’élaboration d’un ouvrage technique de restauration des prairies à fétuque paniculée.
Cette démarche intégrative des problèmes issus des modifications d’utilisation des terres est
plus que jamais d’actualité, au lendemain de la parution du rapport « Chevassus » et à la veille
de la nouvelle Politique Agricole Commune (PAC) de 2013.
24


BIBLIOGRAPHIE
Albert C. (2006) Dynamique de colonisation des ligneux dans les prairies subalpines :
application à Larix decidua. Master 2, Université de Grenoble.
Amiaud B. (1998). Dynamique végétale d’un écosystème prairial soumis à différentes
modalités de pâturage. Exemple de communaux du marais poitevin. Thèse de
Doctorat. Université de Rennes 1.
Ansquer P., Theau J. P., Cruz P., Viegas J., Al Haj Khaled R. & Duru M., (2004).
Caracterisation de la diversite fonctionnelle des prairies à flore complexe : vers la
construction d’outils de gestion. Fourrages, 179: 353-368.
Bakker J.P. (1989). Nature management by grazing and cutting. Kluwer Academic
Publishers, Dordrecht, 416 p.
Barbaro L. (1999). Dynamique agro-écologique des communautés de pelouses sèches
calcicoles du Vercors méridional – Application à la gestion conservatoire de la
biodiversité par le pastoralisme. Thèse de doctorat ès biologie. Université Joseph
Fourier, Grenoble. 189p.
Barbaro L., Dutoit T. and Cozic P. 2001. A six-year experimental restoration of biodiversity
by shrub-clearing and grazing in calcareous grasslands of the French Prealps.
Biodiversity and Conservation, 10, 119–135.
Berendse F. & Elberse W. T. (1990). Competition and nutrient availability in heathland and
grassland ecosystems. In: Grace J. & Tilman D. (eds.) Perspectives on Plant
Competition, pp. 93-115. Academic Press, Florida, Orlando.
Bouchard V. Tessier M., Digaire F., Vivier J.P., Valery L., Gloaguan J.C. & Lefeuvre J.C.
(2003). Sheep grazing as management tool in Western European saltmarshes.
Biodiversity conservation and management. International Congress, Vouziers, France,
pp148-157.
Britton AJ., Pakeman RJ., Carey PD., Marss RH. (2001). Impacts of climate, management
and nitrogen deposition on the dynamics of lowland heathland. J. Veg. Sci. 12, p. 797–
806.
CEMAGREF, Centre Grenoblois, 1981. Recherches en Briançonnais: pastoralisme
montagnard. Ministère de l’agriculture.


Chapin III F.S., Zavaleta E.S., Eviner V.T., Naylor R.L., Vitousek P.M., Reynolds H.L.,
Hooper D.U., Lavorel S., Sala O.E. & Hobbie S.E. (2000). Consequences of
changing biodiversity. Nature, 405, 234.
Chessel D., Doledec S. & Chamely S. (1997). Triplet d’analyse des correspondances.
Documentation de la programmathèque ADE-4 « Analyses multivariées et expression
graphiques des variables environnementales ». Université de Lyon 1, France. 64p.
Chevassus-au-Louis B., Salles J.-M., Bielsa S., Richard D., Martin G. & Pujol J.-L (2009).
Approche économique de la biodiversité et des services liés aux écosystèmes.
Contribution à la décision publique. Rapport, Centre d’analyse stratégique, 376p.
Clément G., Quetier F., Mallen M., Choler P., Douzet R. & Lavorel S. (2003). Les
différentes trajectoires d'utilisation des terres à l'étage subalpin ont-elles laissé une
marque significative dans le paysage actuel du col du Lautaret? Laboratoire
d’Ecologie Alpine,Grenoble, France.
Cornelissen J. H. C., Lavorel S., Garnier E., Diaz S., Buchman N., Gurvich D.E., Reich P.B.,
Ter Steege H., Morgan H. D., Van Der Heidjen M. G. A., Pausas J. G. & Poorter H.
(2003). A handbook of protocols for standardized and easy measurement of plant
functional traits worldwide. Australian Journal of Botany, 51, 335-380.
Cruz P., Duru M., Therond O., Theau J. P., Ducourtieux C., Jouany C., Al Haj Khaled R. &
Ansquer P., (2002). Une nouvelle approche pour caracteriser les prairies naturelles et
leur valeur d’usage. Fourrages,172: 335-354.
Cruz P., Jouany C., Theau J. P., Petibon P., Lecloux E. & Duru M. (2006). L'utilisation de
l'indice de nutrition azotée en prairies naturelles avec présence de légumineuses.
Fourrages, 87, 369-376.
Cunningham S.A., Summerhayes B., Westoby M. (1999). Evolutionary divergence of leaf
structure and chemistry comparing rainfall and soil nutrient gradients. Ecological
Monographs, 69, 569-588.
Daget P. & Poissonnet J. (1969). Analyse phytologique des prairies. Applications
agronomiques. Montpellier, CNRS, CEPE. Doc n°48, 67p.
Dagnelie P. (2008). Le plan d’expérience évolue… Revue MODULAD, 38, 13-36.
Dean A. & Voss D. (1999). Split plot designs. In : Design and analysis of experiments (Ed.
Springer). Verlag New York, Inc., pp. 679-694.
Daily G. (1997). Nature’s Services. Societal Dependence on Natural Ecosystems, Island
Press, 412p.


de Bello F., Lepš J. & Sebastià M.-T. (2006). Variations in species and functional plant
diversity along climatic and grazing gradients. Ecography 29, 801–810.
Diaz S. and Cabido M. (2001). Vive la difference : plant functional diversity matters to
ecosystem processes. Trends in Ecology and Evolution, 16(11): 646-655.
Díaz S., Fargione J., Chapin F.S. & Tilman D. (2006). Biodiversity loss threatens human
well-being. PLoS Biology, 4, 1300.
Dobbs T. & Pretty J.N. (2004). Agri-environnemental stewardship schemes and
“multifonctionality”. Review of Agricultural Economics, 26, 220-237.
Eviner V.T. & Chapin III F.S. (2003). FUNCTIONAL MATRIX: A Conceptual Framework
for Predicting Multiple Plant Effects on Ecosystem Processes. Annual Review of
Ecology, Evolution, and Systematics, 34, 455.
Fischer SF., Poschlod P., Beinlich B. (1996). Experimental studies on the dispersal of plants
and animals on sheep in calcareous grasslands. J. Appl. Ecol. 33, p. 1206–1222.
Fischer M. & Wipf S. (2002). Effect of low-intensity grazing on the species-rich vegetation of
traditionally mown subalpine meadows. Biological Conservation, 104, 1-11.
Foley J.A., DeFries R., Asner G.P., Barford C., Bonan G., Carpenter S.R., Chapin F.S., Coe
M.T., Daily G.C., Gibbs H.K., Helkowski J.H., Holloway T., Howard E.A., Kucharik
C.J., Monfreda C., Patz J.A., Prentice I.C., Ramankutty N. & Snyder P.K., (2005).
Global Consequences of Land Use. Science, 309 : 570-574.
Foster B. L. (1999). Establishment, competition and the distribution of native grasses among
Michigan old-fields. Journal of Ecology, 87, 476-489.
Garcia Bautista A. (2004) Quels facteurs influencent la distribution spatiale et temporelle des
pratiques agropastorales au Lautaret? - Mise en place d'un SIG dans le cadre du
projet VISTA. Laboratoire d'Ecologie Alpine, Grenoble, France.
Garnier, E., Cortez, J., Billes, G., Navas, M.L., Roumet, C., Debussche, M., Laurent, G.,
Blanchard, A., Aubry, D., Bellmann, A., Neill, C. & Toussaint, J.P. (2004) Plant
functional markers capture ecosystem properties during secondary succession.
Ecology, 85, 2630-2637.
Garnier E., Lavorel S., Ansquer P., Castro H., Cruz P., Dolezal J., Eriksson O., Fortunel C.,
Freitas H., Golodets C., Grigulis K., Jouany C., Kazakou E., Kigel J., Kleyer M.,
Lehsten V., Leps J., Meier T., Pakeman R., Papadimitriou M., Papanastasis V. P.,
Quested H., Quetier F., Robson M., Roumet C., Rusch G., Skarpe C., Stenberg M.,
Theau J.P., Thebault A., Vile D., Zarovali M.P., (2007). Assessing the effects of land-
use change on plants traits, communities and ecosystem functioning in grasslands : a


standardized methodology and lessons from an application to 11 European sites.
Annals of Botany, 99 : 967-985.
Grime J. P. (1977). Evidence for the existence of three primary strategues in plants an dits
relevance to ecological and evolutionanary theory. The American Naturalist, 111,
1169-1194.
Grime J.P. (1997). ECOLOGY: Biodiversity and Ecosystem Function: The Debate Deepens.
Science, 277, 1260.
Grime J. P., (1998). Benefits of plant diversity to ecosystems: immediate, filter and founder
effects. Journal of Ecology, 86 : 902-910.
Grime JP., Brown VK., Thompson K., Masters GJ., Hillier SH., Clarke IP., Askew AP.,
Corker D., Kielty JP. (2000). The response of two contrasting limestone grasslands to
simulated climate change. Science 289, p. 762–765.
Grime J.P. (2002). Declining plant diversity: empty niches or functional shifts? Journal of
Vegetation Science, 13, 457.
Gross N. (2003). Réponse à un gradient d’utilisation des terres artificiel de six espèces de
graminées dominantes des communautés herbacées du subalpin. Rapport de DEA
Université Montpellier II, 30p.
Gross N., Suding K.N. & Lavorel S. (2007). Leaf dry matter content and lateral spread
predict response to land use change for six subalpine grassland species. Journal of
vegetation science, 18, 289-300.
Güssewell S. & Bollens U. (2003). Composition of plant species mixtures grown at various
N:P ratios and levels of nutrient supply. Basic Appl. Ecol. 4, 453–466.
Hooper D.U., Chapin F.S., Ewel J.J., Hector A., Inchausti P., Lavorel S., Lawton J.H.,
Lodge D.M., Loreau M., Naeem S., Schmid B., Setala H., Symstad A.J., Vandermeer
J., & Wardle D.A. (2005) Effects of biodiversity on ecosystem functioning: A
consensus of current knowledge. Ecological Monographs, 75 : 3-35.
Jones A.T. & Hayes M.J. (1999). Increasing floristic diversity in grassland: the effects of
management regime and provenance on species introduction. Biological Conservation,
87, 381-390.
Jouglet J.P. & Dorée A. (1987). La fauche dans le festucetum spadiceae ; son incidence sur la
dynamique et le potentiel fourrager d’une pelouse à fétuque paniculée. Rapport
Cemagref-INERM n°219, Grenoble, France. 39p.


Kahmen S., Poschlod P., Schreiber KF. (2002). Conservation management of calcareous
grasslands. Changes in plant species composition and response of functional traits
during 25 years. Biol. Conserv. 104, p. 319–328.
Klimes L. & Klimesova J. (2001). The effects of mowing and fertilization on carbohydrate
reserves and regrowth of grasses: do they promote plant coexistence in species-rich
meadows? Evolutionary Ecology, 15, 363-382.
Kremen C. (2005). Managing ecosystem services: what do we need to know about their
ecology? Ecology Letters, 8, 468.
Landsberg J., Lavorel S. & Stol J. (1999). Grazing response groups among understorey plant
in rangelands. Journal of Vegetation Science, 10, 683-696.
Lavorel S., McIntyre S., Landsberg J. & Forbes T.D.A. (1997). Plant functional
classification ; from general groups to specific groups based on response to
disturbance. Trends in Ecology and Evolution, 12, 414-424.
Lavorel S. and Garnier E. (2002). Predicting changes in community composition and
ecosystem functioning from plant traits: revisiting the Holy Grail. Functional Ecology,
16 : 545-556.
Lavorel S., Díaz S., Cornelissen J.H.C., Garnier E., Harrison S.P., McIntyre S., Pausas J.G.,
Pérez-Harguindeguy N., Roumet C. & Urcelay C. (2007). Plant functional types: are
we getting any closer to the Holy Grail. Terrestrial ecosystems in a changing world,
p.149.
Lavorel S., Grigulis K., McIntyre S., Williams N.S.G., Garden D., Dorrough J., Berman S.,
Quétier F., Thébault A. & Bonis A. (2008). Assessing functional diversity in the field
– methodology matters! Functional Ecology, 22 : 134-147.
Le Roux X., Bardy M., Loiseau P. & Louault F., 2003. Stimulation of soil nitrification and
denitrification by grazing in grasslands: do changes in plant species composition
matter ? Oecologia 137, 417-425.
Louault F., Michalet-Doreau B., Petit M. & Soussana J.F. (2002). Potentialités des prairies
permanentes de montagne pour la production fourragère et la gestion de l’espace. In
Actes du colloque INRA-ENITA, 14 et 15 Novembre 2002, Agriculture et produits
alimentaires de montagne, ENITAC, Lempdes, France. Collection Actes, 8: 33-39.
MacDonald D., Crabtree J.R., Wiesinger G., Dax T., Stamou N., Fleury P., Gutierrez Lazpita
J. & Gibon A. (2000). Agricultural abandonment in mountain areas of Europe:
Environmental consequences and policy response. Journal of Environmental
Management, 59, 47-69.


Mallen M. (2002). Fauche et pâturage sur les prés du Lautaret. 1810-2000 Villar d.Arène.
Site Pro2 – Lautaret – Combeynot - Ecrins, Parc National des Ecrins, 42 p.
Moles, A.T. & Westoby, M. (2004) Seedling survival and seed size: a synthesis of the
literature. Journal of Ecology, 92, 372-383.
Millennium Ecosystem Assessment (2005). Ecosystems and human well-being: synthesis.
Island Press. 607p.
Muller S.D., David F. & Wicha S. (2000). Impact de l’exposition des versants et de
l’anthropisation sur la dynamique forestière dans les Alpes du sud (France).
Géographie Physique et Quaternaire, 54 : 231-245.
Ozenda, P. (1985) La végétation de la chaîne alpine dans l'espace montagnard européen.
Masson, Paris, France.
Parc National des Ecrins (2006). Gestion des alpages du parc national des Ecrins. Enjeux
écologiques et pastoraux. Fiches techniques.
Pärtel M., Kalamees R., Zobel M. & Rosén E. (1998). Restoration of species-rich limestone
grassland communities from overgrown land: the importance of propagule availability.
Ecol. Eng. 10, p. 275–286.
Pausas J.G. & Lavorel S. (2003). A hierarchical deductive approach for functional types in
disturbed ecosystems. Journal of Vegetation Science, 14, 409-416.
Poschlod P., Kiefer S., Tränke U., Fischer S. & Bonn S. (1998). Plant species richness in
calcareous grasslands as affected by dispersability in space and time. Appl. Veg. Sci. 1,
p. 75–90.
Pykälä J. (2003). Effects of restoration with cattle grazing on plant species composition and
richness of semi-natural grassland. Biodiversity and Conservation, 12, 2211-2226.
Quétier F., Rivoal F., Marty P., de Chazal J. & Lavorel S. (unpublished). Descriptions of a
mountain grassland landscape by locals and visitors: an inquiry into narratives on
ecosystem services. Submitted to Landscape and Urban Planning.
Quétier F. (2006). Vulnérabilité des écosystèmes semi-naturels européens aux changements
d’utilisation des terres. Application aux prairies subalpines de Villar d’Arène, France.
Thèse Université Montpellier II, France, 269p.
Quiblier M. & Senn O. (2004). Les differents queyrellins des Alpes du sud et leur mode de
gestion. CERPAM.
Quétier F., Thebault A. & Lavorel S. (2007). Plant traits in a state and transition framework as
markers of ecosystem response to land-use change. Ecological Monographs, 77 : 33-
52.


Ricklefs R.E. & Miller G.L. (2005). Ecologie. 4eme édition, De Boeck Universite
Edition, Bruxelles, 821 p.
Robson T. M., Lavorel S., Clément J.-C. & Le Roux X. (2007). Neglect of mowing and
manuring leads to slower nitrogen cycling in subalpine grasslands. Soil Biology &
Biochemistry, 39, 930-941.
Rudmann-Maurer K.,Weyand A., Fischer M. & Stöcklin J. (2008). The role of landuse and
natural determinants for grassland vegetation composition in the Swiss Alps. Basic
Appl. Ecol,. 9, 494–503.
Schippers P. & Olff H. (2000). Biomass partitioning, architecture and turnover of six
herbaceous species from habitats with nutrient supply. Plant Ecology, 149, 219-231.
Sebastià M. T. (2004): Role of topography and soils in grassland structuring at the landscape
and community scales. Basic Applied Ecology, 5, 331–346.
Shea K., Roxburgh S.H. & Rauschert E.S.J. (2004). Moving from pattern to process:
coexistence mechanisms under intermediate disturbance regimes. Ecology Letters, 7,
491-508.
Smith T.M., Shugart H.H. and Woodward F.I. (1997). Plant functional types: their relevance
to ecosystem properties and global change. Cambridge, UK: Cambridge University
Press.
Soltner D. (1978). Les bases de la production végétale.Tome 1 – Le sol. 7ème
édition.
Collection Sciences et techniques agricoles, Angers, 311p.
Stöcklin J., Bosshard A., Klaus G., Rudmann-Maurer K. & Fischer M. (2007). Utilisation du
sol et diversité biologique dans les Alpes : faits, perspectives, recommandations. Vdf
Editions de la Haute Ecole. 191p.
Viard-Crétat F., Lefebvre M., Gallet C. & Lavorel S. (2008). A leachate a day keeps the
seedling away : mowing and the inhibitory effects of Festuca paniculata in subalpine
grasslands. Journal of Vegetation Science,
Viard-Crétat F. (2008). Mécanismes de régénération des espèces végétales dans les prairies
subalpines sur un gradient d’extensification des pratiques agricoles. Thèse Université
Montpellier II, 225p.
Vile D. (2005). Significations fonctionnelle et écologique des traits des espèces végétales :
exemple dans une succession post-culturale méditerranéenne et généralisations. These
Université Montpellier II, France, 109 p.
Violle C., Navas M-L., Vile D., Kazakou E., Fortunel C., Hummel I. & Garnier E., (2007).
Let the concept of trait be functional ! Oikos, 116 : 882-892.


Vitousek P.M., Mooney H.A., Lubchenco J., & Melillo J.M. (1997). Human domination of
Earth's ecosystems. Science, 277 : 494-499.
Vittoz P., Selldorf P., Eggenberg S. & Maire S. (2005). Festuca paniculata meadows in
Ticino (Switzerland) and their Alpine environment. Botanica Helvetica, 115, 33-48.
WallisDeVries M.F., Bakker J.P. & Van Wieren S.E. (1998). Grazing and Conservation
Management. Kluwer, Dordrecht, The Netherlands.
Watkinson A. R. & Ormerod S. J. (2001). Grasslands, grazing and biodiversity: editors’
introduction. – J. Appl.Ecol. 38: 233–237.
Weiher E., Van der Werf A., Thompson K., Roderick M., Garnier E. & Eriksson O. (1999).
Challenging Theophrastus : Acommon core list of trait for functional ecology. Journal
of Vegetation Science, 10, 609-620.
Westoby M. 1998. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and soil
199 : 213-227.
Whright I.J., Reich P.B. & Westoby M. (2001). Strategy shifts in leaf physiology, structure
and nutrient content between species of high- and low-rainfall and high- and low-
nutrient habitats. Functional Ecology, 15, 423-434.
Flore utilisées pour l’identification des espèces
Aeschimann D., Lauber K., Moser D.M. & Theurillat J-P. (2004). Flora Alpina. Editions
Belin. 2670p (3 tomes).
Aeschimann D. & Burdet H. M. (1994). Flore de la Suisse et des territories limitrophes – Le
nouveau Binz. 2ème
édition. Edition du Griffon. Neuchâtel, Suisse. 603p.
Blamey M. & Grey-Wilson C. (1991). Guide des fleurs de montagne. Delachaux et Niestlé.
Paris. 384p.
Bonnier G. & De Layens G. (1998). Flore complète portative de la France, de la Suisse et de
la Belgique. Edition Belin. 425p.
Dorée A. (1995). Flore pastorale de montagne. Tome I – Les graminées. Cemagref Editions
et Edition Boubée. Grenoble. 207p.
Dorée A. (2000). Flore pastorale de montagne. Tome II – Légumineuses et autres plantes
fourragères. Cemagref Editions. Grenoble. 227p.
Lauber L. & Wagner G. (2001). Flora Helvetica – Flore illustrée de la Suisse. 2ème
édition.
Editions Haupt. Bern, Suisse. 1615p.


Flore utilisée pour les données autoécologiques des espèces
Rameau J. C., Mansion D. & Dumé G. (1994). Flore Forestière Française. Guide écologique
illustré – Tome 2 : montagnes. Institut pour le Développement Forestier, Paris, 2421p.
Logiciels statistiques employés
R Development Core Team (2007). R: A language and environment for statistical computing.
Foundation for Statistical Computing, Vienna, Austria.
JMP (2002). SAS Institute, Cary, North Carolina.


ANNEXES
Annexe 1 : Cartographie des dates de dernier labour sur les parcelles de l’Adret de
Villar d’Arène
Annexe 2 : Cartographie des dates de dernière fauche sur les parcelles de l’Adret de
Villar d’Arène
Annexe 3 : Trajectoire d’utilisation des parcelles de l’Adret de Villar d’Arène
Annexe 4 : Résultat de la relation allométrique pour estimer la production de bimasse
de F. paniculata
Annexe 5 : Schématisation de l’analyse de co-inertie RLQ à trois tableaux
Annexe 6 : Liste des taxons identifiés sur les prairies de fauches subalpines

Annexe 1 : Cartographie des dates de dernier labour sur les parcelles de l’Adret de Villar d’Arène (P. Lamarque, 2009)

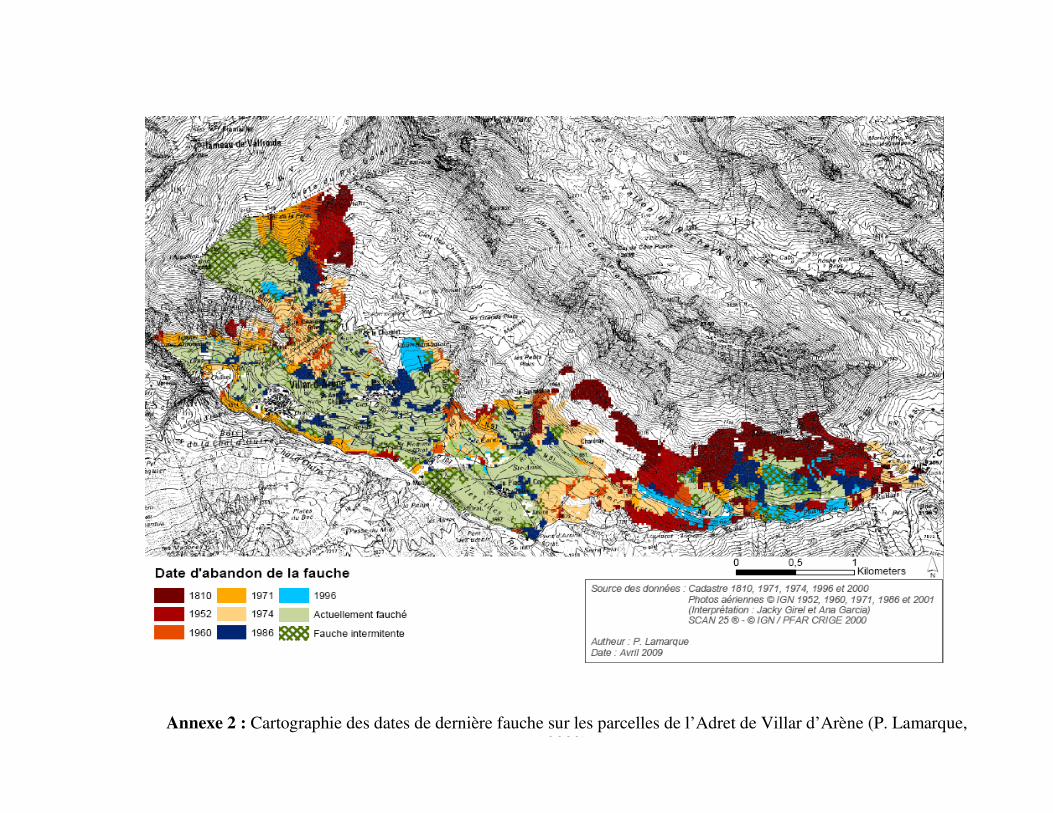
Annexe 2 : Cartographie des dates de dernière fauche sur les parcelles de l’Adret de Villar d’Arène (P. Lamarque,
2009)


Annexe 3 : Trajectoire d’utilisation des parcelles de l’Adret de Villar d’Arène (P. Lamarque, 2009)


La relation allométrique réalisée afin d’estimer la production de biomasse montre que :
- la surface des touffes est très fortement corrélée à la biomasse aérienne totale (r2=.95,
P<0.0001) (Fig. 1)
- La hauteur des touffes n’est pas corrélée significativement à la biomasse aérienne (r2
nul) (Fig. 2)
- Lorsque l’on fait une régression multiple, utiliser ces deux variables permet de passer
de r2=0.95 à r2=0.97.
- Le nombre de talles n’a pas été pris en facteur car trop difficilement utilisable.
- La production de biomasse peut donc être facilement mesurable sur le terrain rien que
par la surface. La hauteur ayant une faible corrélation.
- La quantification de la production de biomasse peut donc être calculer selon le modèle
simple suivant : Biomasse = 0.228*surface +2.0322. Si on veut y inclure la hauteur, le
modèle est : biomass= 0.246*surface -0.534*hauteur +22.574.
- Seule la surface des touffes a été mesurée pour l’estimation de la production de
biomasse.
Annexe 4 : Résultat de la relation allométrique pour estimer la production de
bimasse de F. paniculata
Fig. 1 : Corrélation entre la masse des touffes et
leur surface
Fig. 2 : Corrélation entre la masse des touffes et leur
hauteur


L’analyse des relations entre traits fonctionnels et variables environnementales peut
ëtre traiter par l’intermédiaire d’une analyse de co-inertie généralisée entre 3 tableaux,
appelée analyse RLQ. Cette méthode permet avec une ordination simultanée des 3 matrices
dans laquelle L est la matrice relevés x espèces, avec aux marges des relevés la matrice relevé
x variables (R) et la matrice espèces x traits fonctionnels Q aux marges des espèces. La
matrice centrale est analysée par une Analyse Factorielle des correspondances (AFC),
l’analyse RLQ maximise la covariance entre combinaison linéaire des colonnes de R et de Q.
Annexe 5 : Schématisation de l’analyse de co-inertie RLQ à trois tableaux.


Achillea millefolium
Aconitum anthora
Agropyron caninum
Agrostis capillaris
Alchemilla glabra
Alchemilla splendens
Alchemilla xanthochlora
Allium carinatum
Allium schoenoprasum
Anemone narcissifolia
Anthericum liliago
Arenaria ciliata subsp. multicaulis
Arnica montana
Artemisia atrata
Artemisia campestris
Asphodelus albus
Astragalus danicus
Astragalus glycyphyllos
Astragalus penduliflorus
Astrantia major
Botrychium lunaria
Briza media
Bromus erectus
Bupleurum falcatum
Bupleurum ranunculoides
Campanula barbata
Campanula rhomboidalis
Campanula rotundifolia
Campanula scheuchzeri
Campanula thyrsoides
Carduus defloratus
Carex nigra
Carex sempervirens
Carlina acaulis
Carum carvi
Centaurea montana
Centaurea scabiosa
Centaurea uniflora
Cerastium arvense subsp. strictum
Chaerophyllum villarsii
Colchicum autumnale
Cotoneaster integerrima
Crepis bocconi
Crepis conyzifolia
Crepis montana
Crepis pyrenaica
Dactylis glomerata
Deschampsia flexuosa
Dianthus pavonius
Dianthus sylvestris
Erigeron atticus
Erysimum virgatum
Euphorbia cyparissias
Euphorbia verrucosa
Euphrasia rostkoviana
Festuca laevigata
Festuca nigrescens
Festuca ovina
Festuca paniculata
Festuca rubra
Galium boreale
Galium lucidum
Galium mollugo
Galium verum
Gentiana acaulis
Gentiana campestris
Gentiana lutea
Geranium rivulare
Geranium sylvaticum
Geum montanum
Gymnadenia conopsea
Helianthemum nummularium subsp. grandiflorum
Helictotrichon pratense
Helictotrichon sedenense
Heracleum sphondylium
Hieracium amplexicaule
Hieracium corymbosum
Hieracium laggeri
Hieracium umbellatum
Hieracium villosum
Hypericum maculatum
Hypochaeris uniflora
Knautia arvensis
Knautia dipsacifolia
Laserpitium latifolium
Lathyrus occidentalis
Lathyrus pratensis
Leontodon hispidus
Leucanthemum vulgare
Lilium martagon
Lotus corniculatus
Luzula campestris
Luzula multiflora
Luzula nutans
Meum athamanticum
Molinia caerulea
Myosotis alpestris
Myosotis arvensis
Narcissus poëticus
Nigritella nigra
Onobrychis montana
Ornithogalum umbellatum
Oxytropis campestris
Oxytropis lapponica
Paradisea liliastrum
Pedicularis tuberosa
Phleum alpinum
Phleum pratense
Phyteuma betonicifolium
Phyteuma michelii
Phyteuma orbiculare
Pimpinella major
Pimpinella saxifraga
Plantago media
Plantago serpentina
Poa alpina
Poa nemoralis
Poa pratensis
Polygonum viviparum
Potentilla crantzii
Potentilla grandiflora
Potentilla recta
Primula veris
Pulmonaria angustifolia
Pulsatilla alpina
Pulsatilla vernalis
Ranunculus acris
Rhinanthus alectorolophus
Rhinanthus minor
Rumex arifolius
Salvia pratensis
Sanguisorba officinalis
Satureja alpina
Scutellaria alpina
Sedum anacampseros
Sempervivum tectorum
Senecio doronicum
Sesleria caerulea
Silene nutans
Silene vulgaris
Sisymbrium austriacum
Stachys pradica
Stachys recta
Taraxacum alpinum
Thalictrum minus
Thalictrum simplex
Thesium pyrenaicum
Thlaspi montanum
Thymus serpyllum
Tragopogon pratensis
Traunsteinera globosa
Trifolium alpestre
Trifolium alpinum
Trifolium montanum
Trifolium pratense
Trisetum flavescens
Trollius europaeus
Valeriana officinalis
Veratrum album
Veronica allionii
Vicia cracca
Viola calcarata
Annexe 6 : Liste des taxons identifiés sur les prairies de
fauches subalpines (162 taxons)