AL'UNIVERSITE F? ET M. CURIE - PARIS VI
303
THESE DE DOCTORAT D'ETAT ES SCIENCES NATURELLES présentée AL' UNIVERSITE F? ET M. CURIE - PARIS VI par Jean- Claude GAUTUN pour obtenir LE GRADE DE DOCTEUR ES SCIENCES ECOLOGIE DES RONGEURS DE SAVANE A EN MOYENNE COTE Of IVOIRE soutenue le . devant le jury composé de : MM. les Professeurs M. LAMOTTE F. BOURLIERE Dr F. PETTER M. le Professeur P. DREUX MM. R. BARBAULT A. BROSSET ORSTOM - PARIS 1981 Président, Rapporteur, Examinateur,
Transcript of AL'UNIVERSITE F? ET M. CURIE - PARIS VI
Ecologie des rongeurs de savane en moyenne Côte
d'Ivoireprésentée
par
ECOLOGIE DES RONGEURS DE SAVANE
A
soutenue le . devant le jury composé de :
MM. les Professeurs M. LAMOTTE
F. BOURLIERE
MM. R. BARBAULT
Je remercie Monsieur le Professeur G. CN~US,Directeur
Général de l'Office de la Recherche Scientifique et Technioue
Outre-Mer, de m'avoir permis d'effectuer les recherches, dont les
résultats sont présentés ici, au Laboratoire de Mammalogie du
Centre ORS TOM d'Abidjan-Adiopodoumé.
Je remercie [iIonsieur le Professeur :1. LAI·lOTTE Gui, apr è s
~'avoir conseillé au cours de l'élaboration du ~anuscrit, a accecté
d'être président du jury de cette thèse.
Je remercie le Docteur F. PETTER d'avoir bie~ voulu ~.
tous moments et,deDuis de nombreuses années, ~e f~ire profiter de
sa grande connaissance de la systématique et de la biolopie des
ron~eurs tropicaux. Ses conseils et son amitié m'ont été d'une
p:rande aide.
Je remercie r'Iessieurs les Professeurs F. BOURLIERE et
Ph. DREUX ainsi que Messieurs A. BROSSET et R. BARBAULT d'avoir
bien voulu faire partie de mon jury de thèse.
Je remercie Monsieur le Professeur Ch. TSIEAULT, mon
Directeur scientifique pendant toute la durée de cette étude.
Je remercie Monsieur le Professeur L. HIRTH, Président
du Comité Techniaue de Phytopatholosie et de Zoolorrie A9r.lir.uée
de l'ORSTQr:i, les rnembr-e s du Bureau Permanent et par-t i cu l Lè r emen t
Monsieur N. ROTH 90ur le soutien cu'ils m'ont manifesté.
i'Ionsieur L. BELLIER él. été à l'origine de ma formation
de ~arnmalogist~ qu'il soit assuré de mon amicale reconnaissance.
Je remercie I·Ionsieur i.J'iGELINI, Directeur de la station
IRCT de Bouaké (Côte d'Ivoire), d'avoir bien voulu me donner
l'autorisation de travailler au Foro.
Je remercie mes collègues A. BEAUDOU pédolo~ue à l'
ORS TOM et J. CESAR botaniste à l'IE~NT - CHEZ de Bouaké pour l'
aide qu'ils m'ont fournie dans l'êtude des sols et de la végéta
tion du Foro.
Monsieur P. CASES pour l'aide au'ils m'ont fournklors du traite
ment informaticue des données de pié~eap.e.
Je remercie ~:onsieur E. R.IO pour sa co ï.Labor-at i on
technique à cette étude.
du laboratoire de Mammalogie d'Abidjan-Adiopodoumé pour le tra
vail fourni durant l'étude, particulièrement I!;essieurs AITADOU
SIBIRI ZONE et DENIS FATTO SAGOU.
PLA N
1.1.2. Substratum.
1.1.3. Climat.
1.1.4. Végétation.
1.2.2. Manipulations.
3
3
CHAPITRE II - PEUPLE~ŒNT DES PETITS RONGEURS DE LA REGION DU FORO : IDENTIFICATION ET TRAITS GENER~UX DES ESPECES. •••• • 28
2.1. Identification des espèces . . . . 2.1.1. Tatera.
2.1.2. Taterillus.
2.1.3. Nus.
2.1.4. Arvicanthis.
2.1.5. Lemniscomys.
2.1.6. Mylomys.
2.1.7. Uranomys.
2.2.1. Peuplement dans la savane.
2.2.2. Peuplement dans les forêts-galeries.
2.2.3. Peuplement dans les cultures.
2.3. Comparaison du peuplement de la région du Foro avec ceux d'autres régions de Côte d'Ivoire. • • • • • 59
2.3.1. Comparaison qualitative.
2.3.2. Comparaison quantitative.
CHAPITRE III - REPRODUCTION ET CROISSANCE DA.NS LE lULIEU NATUREL ET EN CAPTIVITE. • • • 62
3.1. Etude de la reproduction. . . 62
3.1.1. Modalités de la reproduction dans le milieu naturel.
3.1.2. Observations sur la reproduction en captivité.
3.2. Etude de la croissance. . . . . . . . . . . . . . . . 78
3.2.1. Lemniscomys bellieri.
3.2.2. Myomys daltoni.
3.2.3. Tatera hopkinsoni.
3.2.4. Uranomys ruddi.
3.2.5. Influence des saisons sur la croissance corporelle de Lemniscomys bellieri.
CHAPITRE IV - ANALYSE DES RESULTATS DE PIEGEAGE SUR LE QUADRAT DE MARQUAGE. . . 90
4.1. Résidence. . . . . . . . 91
4.1.1. Taux de résidents dans la population de Lemniscomys bellieri.
4.1.2. Taux de résidents dans la population de Myomys daltoni.
4.1.3. Taux de résidents dans la p09ulation de Tatera h09kinsoni.
4.1.4. Taux de résidents dans la population de ['·1us spp ,
4.1.5. Origine des résidents et des passagers.
4.1.6. Conclusions.
4.2.1. Durée de persistance maximale de chaque espèce.
4.2.2. Durée de persistance moyenne de 50, 25 et 5 % de chaque population.
4.2.3. Durée de persistance moyenne de 50, 25 et 5 % de chaque p09ulation résidente.
4.2.4. Conclusions.
94
97
4.4.2. Spectres de déplacements.
4.4.4. Modifications de la mobilité des espèces selon les saisons.
4.4.5. Evolution mensuelle de la mobilité moyenne de Lemniscomys et Myomys.
4.4.6. Influence des feux de savane sur la mobilité des espèces.
4.4.7. Conclusions.
4.5. Occupation spatiale du quadrat de marquage parles principales espèces de rongeurs. ••••••••••• 103
4.5.1. Localisation des zones préférentielles des principales espèces.
4.5.2. Analyse de la répartition des captures de chaque espèce.
4.5.3. Correspondances entre la répartition numérique des principales espèces et certains éléments du milieu.
4.5.4. Conclusions.
CHAPITRE v - DENSITES ET BIQ!'.1ASSES DES POPULATIOi'!S DE RONGEURS. •••• • ••• 111
5.1. Densités et biomasses estimées sur le cuadrat de marquage du Foro. •••••• • ••• 112
5.1.1.
5.1.2.
5.1.3.
5.1.5.
5.1.6.
Méthodes d'estimation des densités.
Choix de méthodes utilisables dans le cas des ron~eurs du Foro.
Estimations des densités brutes.
Estimations de la biomasse pour 10 ha.
Discussion et conclusions.
5.2. Densités et biomasses de Lemniscomys bellieri estimées sur les lignes exhaustives de piégeaf,e.
5.2.1. Correspondance entre les effectifs pié~és
sur les li~nes et sur le quadrat.
5.2.2. Méthode d'estimation des densités sur lignes.
5.2.3. Estimation des densités mensuelles de Lemniscomys au Foro.
5.2.4. Com~araison des savanes du Foro et de Lar;;to.
5.2.5. Estimation de la biomasse mensuelle.
5.2.6. Conclusions.
CHAPITRE VI - ELEi/IENTS DI ANALYSE DEr.10GRAPHIQUE CHEZ LES ESPECES LES PLUS ABONDAi\fTES.
129
138
6.2.1. Composition par âges de l'ensemble des populations.
6.2.2. Influence des saisons sur la composition par âges de la population de Lemniscomvs.
6.3. Analyse démographique longitudinale.
6.3.1. Evolution des cohortes.
143
Jusque dans les années 60, par comparaison aux études
réalisées en milieu tempéré, un retard très important pouvait
être constaté dans l'acquisition des informations se rapportant
à la biologie et à l'écologie des rongeurs tropicaux africains.
Quelques études très ponctuelles avaient cependant été menées en
Afrique centrale (Ouganda, ZaIre) et en Afrique de l'ouest (Nigé
ria, Guinée-Mont Nimba). Pour le rest~ l'essentiel des études en
trepriseétaient des missions de courte durée réalisées dans le
but de collecter des spécimens à destination des taxinomistes et
des zoogéographes.
triel dans la plupart des pays d'Afrique tropicale, et corréla
tivement celui des recherches d'accompagnement au développement,
ont permis d'implanter différents groupes de recherches au Séné
gal, au Nigéria, en CSte d'Ivoire.
Le programme de recherches sur les rongeurs que nous
avons entrepris en moyenne cSte d'Ivoire à partir de 1971 est
complémentaire de celui réalisé précédemment par Bellier (1963
1972) à Lamto dans les savanes du sud du V baoulé. Il vise essen·
tiellement à fournir des informations sur les modalités de la re
production, la croissance, les densités, la biomasse et la démo
graphie des populations de rongeurs de savane dans la zone tropi
cale humide. Ce programme s'est déroulé en deux phases (1971-1974
et 1976-1979) séparées par un arrêt de deux ans (1974-1976) pen
dant lequel nous avons dO nous intéresser plus particulièrement
aux rongeurs nuisibles aux cultures industrielles (palmier à huile) dans la zone forestière de basse CSte d'Ivoire.
- 2 -
Le site du Foro a été choisi comme représentatif d'une
zone encore non étudiée. Il a été choisi également par suite de sa
proximité de la zone dans laquelle les cultures industrielles (can
ne à sucre, soja) ont été (ou doivent être) implantées, afin que
nos résultats puissent être utilisables s'il s'avère nécessaire
de lutter contre des rongeurs nuisibles. Enfin, il a été choisi
à la suite de l'implantation d'une équipe d'Entomologistes de
l'ORSTOM au même endroit, ce qui nous a permis de mieux renta
biliser l'infrastructure, le matériel et le personnel commun aux
deux laboratoires.
1.1. Le milieu.=========
La "Îorêt classées" de Foro-Foro - ou du Foro - (qui
est en réalité une ~ortion de savane mise en réserve avant 1947~
est située dans le centre de la C6te d'Ivoire (7° 55' N et 5° W),
à mi-chemin de Katiola au nord et de Bouaké au sud. A la limite
des secteurs des savanes guinéennes et sub-soudanaises, "elle est
comprise dans l'interfluve du Bandarna (élargi par suite de la mise
en place de la retenue d'eau du barrage de Kossou) et de son af
fluent le N'zi, dans le haut du bassin du N'zi.
La zone classée, de forme grossièrement trapézoïdale,
est limitée à·l'ouest par la route nationale Bouaké-Katiola, au
nord et au sud par deux marigots, affluents du N'zi, le Foro-Foro
et la [~'bé, oui coulent vers l'est et dont la confluence maraue
l'extrémité est de la réserve. Le domaine du Foro se présente com
me un plateau s'inclinant et se rétrécissant d'ouest en est, en
taillé par un réseau de marizots généralement orientés nord-sud
qui n'ont pas tous atteint leur profil d'équilibre (Bru~ière, 1948).
L'altitude est essentiellement de l'ordre de 200 m, avec àes col
lines et àes plateaux culminant vers 300 m.
2t "Forêt classée" est un terme normalisé de forestier qui ne préjuge pas de la nature de la vépétation du site auouel il s'adresse.
- 4 -
La zone des sav~~es auquelles appartient le Foro a
fait l'objet de trois missions photogra?hiaues aériennes réalisées
à l'intervalle de 10 ans soit
- mission 1952-53 au 1/50.000 (IGN-AOF-N032, 1952-53)
- mission 1961-62 au 1/50.000 (IGN-AOF-1961-62)
- mission 1973 au 1/20.000 (IGN-73 CI-V-072/200).
1.1.2. Le substratum.
1.1.2.1. La géologie.----------
La forêt classée du Foro est située dans la zone des
granitordes homogènes à biotite appartenant iu complexe Baoulé du
Protérozorque inférieur (Ta;;nini, 1972). Il.s'a.~it de granites
calco-alcalins faiblement orientés, à larges filons de quartz
blanc. Les cr t s t aux sont de taille variable et la s t ruc-cur-e 'la du
micro-grenu à la pegmatite 8:vec cristaux àe muscovite de 1 à 2 cm
de diamètre. Les éléments noirs y so~t bien représentés (Brugière,
1948) •
tzeuses, micacées. Parfois, certaines variétés de pegmatites don
nent localement des sols très argileux. La roche en place, lorsqu'
elle affleure, porte très souvent des l'polissoirs" restes d'une
occupation humaine ancienne (Avenard, 1975).
1.1.2.2. Les sols.
la suivante
- versants: sols ferrugineux r,ravillonnaires,
temporaire.
- 5 -
Pour Perraud (1971) l'ensemble de la séquence compre
nant plateaux et pentes est située dans la classe des sols ferra
litiques moyennement désaturés issus du ~ranite. La classification
des sols du Foro devient pour cet auteur
sur plateaux à couvert forestier: sol brun rouge, argileux,
très gravillonnaire : "remanié modal",
graveleux, à carapace à faible profondeur: "remanié induré",
- en haut de pentes, sol gris sableux à ocre rouge, argilo-sableux,
à carapace à moyenne profondeur: "remanié, recouvrerrent modal,
faciès induré",
- sur pentes, sol gris sableux à ocre jaune, sablo-argileux, gra
villonnaire, tacheté en profondeur : "remanié aopauvri Il,
- en bas de pente, sol gris sableux à ocre sableux, légèrement
tacheté en profondeur: "appauvri hydromorDhe",
- dans les bas-fonds, sol gris blanc, sablo-argileux à pseudo gley,
reposant sur des sables grossiers: "hydromporphe minéral".
1.1.3. Le climat.
Le Foro fait partie de la région appartenant au type
climatique C 1 défini par Eldin (1971) : "deux ou quatre saisons
dont une grande saison sèche d'environ cinq mois allant de novem
bre à mars, avec un à trois mois d'Harmattan (vent sec du nord),
1200 à 1600 mm de pluie, 1800 à 2300 heures d'insolation annuelle,
des températures moyennes mensuelles comprises entre 19 et 34° C,
la température moyenne annuelle se situant entre 25 et 28° C".
1.1.3.1. ~~!~~!E~~~_~!~~~~~~_~~_S!!~~~_~~_~~~~
Les informations que nous rapportons ici nous ont été
fournies par le laboratoire d'Entomologie de l'ORSTO~r qui a géré
le parc météorologique du Foro pendant la période 1971-1973, et
par le Ser~ice Agronomique de l'IRCT.
- 6 -
Les températures observées au cours de la période 1971
1973 (tab. I) sont voisines de celles rapportées ~ar Eldin (op. cit):
la température maximale moyenne ,mensuelle (Ti) a une valeur de
31,9 ~ 1,3 0 C, la température minimale moyem1e mensuelle (fn) a
une valeur de 17,6 ~ 1,1 0 C et la moyenne des températures ci-dessus Tx+Tn +( 2 ) a une valeur de 24,7 - 0,8 0 C.
Les moyennes mensuelles les plus faibles sont observées
en août pour Tx, en décembre pour Tn par suite de l'Harmattan et à
cause des minimums précédents en août et décembre pour leur ~oyenne.
Les moyennes mensuelles les plus fortes sont observées
en février pour Tx,en avril pour Tn et en février pour leur
moye~~e.
males (Tx) et des températures minimales (Tn) observées en 1971-73
et au cours de la période 1977-78 sont très voisines, ce qui nous
a permis après regroupement de l'ensemble des observations d'esti
mer des températures moyennes (tab. I).
1.1.3.1.2. Insolation (tao. II).
Durant la période 1971-1973 l'insolation totale annu
elle moyenne a été de 2390,4 heures, valeur comparable à celle
fournie ?ar Eldin. La moyenne mensuelle d'insolation passe par
des valeurs minimales entre juillet et septembre (115-117 heures)
et oar des valeurs ~aximales entre novembre et mars (231 à 252
heures).
1.1.3.1.3. Photopériode (tab. II).
Les jours les plus longs se situent normalement en juin avec 12,57 heures, les plus courts en décembre avec 11,67 heures.
L'écart entre ces deux valeurs, très faible, est de 0,9 heures soit 54 ~n.
- 7 -
La pluviosité annuelle totale a été en moyenne de 1110,0
mm pendant la période 1971-73 avec 74,3 jours de pluie en moyenne,
et de 1020,0 ± 111,6 mm pendant celle allant de 1970 à 1978. Ces
valeurs sont sensiblement plus faibles que les moyennes rapportées
par E1din pour les décennies précédentes. La répartition mensuelle
de la pluviosité est très hétéropène (tab. II) : au cours des six
mois les plus pluvieux il tombe 78,4 % de la hauteur d'eau annuelle
et au cours des quatre mois les moins pluvieux il n'en tombe que
6,3 %. Les pluies apparaissent avec une grande variabilité, géné- •
ra1ement en mars. Elles sont bien installées en avril, mai, juin.
Elles réapparaissent, toujours avec une grande variabilité, en aoOt,
sont fortes en septembre et également très variables en octobre.
Le mois de juillet, en moyenne bien arrosé, présente également une
grande variabilité dans sa pluviosité. Septembre puis avril, mai,
juin sont les mois les plus pluvieux. Novembre, puis janvier, fé
vrier et décembre sont les mois les plus secs.
1.1.3.1.5. Saisons.
faire.par l'emploi de différents indices.
- L'évapotranspiration potentielle (E.T.P.) est un indice complexe
dont le calcul nécessite la connaissance de paramètres qui carac
térisent au mieux le pouvoir évaporant de l'air qui s'exerce dans
le milieu donné. L'évapotranspiration potentielle totalise l'éva
poration physique et l'évaporation "physiologique" (Lecordier, 1974)
Les paramètres nécesaires à l'estimation de l'E.T.P.
par la formule de Turc (1961), citée par E1din (1971), sont la
température moyenne, la durée du jour, la durée quotidienne moyenne
d'insolation, l'énergie du rayonnement solaire. La valeur mensuelle
de l'E.T.P. rapportée à la pluviosité moyenne du mois (fig. 2)
permet de conclure à la présence d~ si~ mois pluvieux en moyenne
dont cinq le sont nettement (avril à juin, aoOt et septembre) et
- 8 -
un peut être pluvieux mais aussi souvent sec,juillet. La seconde
période également de six mois, se compose de deux mois humides qui
peuvent être parfois pluvieux (mars et octobre) et quatre mois net
tement secs (novembre à février).
- Un système d'indices permet de dresser le diagramme ombrothermique
de Moral (1964). Il ne prend en compte que la température mensuelle
moyenne qui, multipliée par des facteurs constants, permet de défi
nir des limites allant de l'aridité à la pluviosité et ainsi de
classer les mois selon leur pluviométrie en arides, secs, humides
ou pluvieux. Ces limites appliquées à l'année pluviométrique moyenne
(fig. 2) permettent de conclure, à la présence de cinq mois plu
vieux (avril à juin, août et septembre), de trois mois humides
(mars, juillet et octobre), de deux mois secs (décembre, janvier)
et de deux mois arides (novembre et février). On note également que
la limite des mois pluvieux déterminée par la méthode de Moral est
assez proche de celle calculée dans le cas de l'E.T.P. par la for
mule de Turc •
. - Il est enfin possible, grâce ~ un nouveau systè~e d'indices, de
dresser le diagramme ombrothermique de Lecordier (1974). Pour son
auteur ce système permet de mieux rattacher les variations biolo
giques des espèces animales aux variations du climat local. Le
classement des mois selon leur pluviométrie est enèore affiné par
la création de deux nouvelles classes (sub-humide et sub-sec).
L'application de ce procédé à la pluviométrie moyenne
mensuelle au Foro confirme le classement que nous avons obtenu
précédemment : quatre mois sont en moyenne nettement pluvieux (avril
mai, juin et septembre), août est en moyenne humide, mars et octobre
sont en moyenne sub-humides, janvier, février et novembre sont en
moyenne arides et décembre est en moyenne sec (fi~. 3).
1.1.3.1.6. Variations climatiques interannuelles.
Les variations climatiques qui se sont produites au cours de la période 1970-1978 ont trait à la oluviométrie annuelle
- 9 -
ainsi qu'à la durée et a l'intensité des différentes saisons.
- Variation de la pluviométrie annuelle.
Pendant la durée de l'étude, nous avons enregistré
(fir;. 4) deux années moyennes (1971 et 1976), une année oluvieuse. . (1974), une année très pluvieuse (1977) et cinq années déficitaires
(1970, 72, 73, 75, 78). Toutes les années sauf 1977 se situent en
deçà de l'intervalle moyen calculé par Eldin.
- Variation de la durée et de l'intensité àes saisons.
La durée et l'intensité des saisons sont très varia
bles d'une année à l'autre (fip.. 4). La grande saison des pluies
présente une très grande variabilité mais elle est toujours présentE
Normalement elle débute dans le courant de mars pour s'achever en
juin. Elle peut être précoce (1971, 1975) ou tardive (1977) ; les
pluies peuvent y @tre abondantes (1971, 1972, 1975) ou faibles
(1970, 1974). La petite -saison des pluies est plus régulière que
la grande. Dans presque tous les cas le mois de septembre est le
plus arrosé (7 cas sur 9). Dans les "conditions normales" elle se
déroule de courant août à courant octobre. Il existe toutefois des
années où la petite saison des pluies est très arrosée (1973 et
surtout 1977) et d'autres où elle est presque inexistante (1978).
L'intersaison centrée sur juillet est elle'aussi très variable;
lorsqu'elle présente une pluviosité importante, l'année se réduit
à deux saisons, l'une sèche et l'autre humide (1970, 1975) ; pré-
sentant une sécheresse marquée, elle permet le découpar.e
théorique de la saison humide en deux saisons des pluies.
La saison sèche est assez peu variable. Elle s'étend
normalement de courant octobre à courant mars, mais peut être limi
tée à la période oui va de novembre à février (1972-73, 1974-75). Le mois de décembre (exceptionnellement janvier) a une pluviosité
qui correspond, certaines années, à la classe sub-humide (1975).
- 10 -
milieux.
dans lesquelles nous avons analysé le peuplement de rongeurs. Les
savanes de Dabou, de Lamto et de Ferkéssédougou (pour le site de
Ouango-Fitini). Au niveau régional, nous avons comparé le climat
du Foro avec ceux de Bouaké et de Katio1a, où la pluviosité moyen
ne mensuelle observée est très voisine de celle du Foro (fig. 5).
Elle diffère nettement de celle constatée à Lamto, où la pluviosité
en novembre, jênvier, février et mars, très abondante, rappelle le
climat de basse Cate d'Ivoire rencontré- à Dabou.
Enfin la pluviosité du Foro se différencie nettement
de celle de Ferkéssédougou où n'existe plus qu'une seule saison des
pluies dont le maximum d'abondance se situe en août-septembre et où
la saison sèche est de plus longue durée.
La pluviométrie annuelle est voisine pour le Foro,
Katio1a et Boualcé , Celle de Larrto est un peu supérieure à celle du
Foro (+ 15 %). Pour des périodes différentes, celle de Ferkéssédou
~ou est éga1e~ent plus élevée (+ 35 %) et celle de Dabou presque
~ga1e au double (+ 94 %). Les autres variables c1imatioues caracté-- .
risant les stations retenues sont rapportées dans le tab1eauIII.
1.1.4. Végétation.
La lIforêt c1assée ll du ::'oro appartient encore aux savanes
guinéennes préforestières à Brachy~ria brachy10pha et Panicum phra~
mitoïdes. L'observation olus att~ntive de la carte de la vé~étation. - . el' Ad j ànohoùn de Cate d'Ivoire de Gui11aumet/(1971) montre qu'en fait le domaine
du Foro se situe à-la limite de- la zone de savanes sub-soudaniennes
dont la limite s'en situe à moins de 50 km au nord et à l'est.
La densité de la strate 1i~neuse décroît des plateaux
aux bas de pentes. Trente cinq espèces d'arbres et arbustes sont
- 11 -
rencont~ées couramment au Foro, en dehors des forêts de sommet et
des forêts-qaleries. Les plus abonda~tes sont Loohira lanceolata,
Afrorrnosia laxiflora, Butyroscerrnum paradoxum, ?a~inari curatel
lifolia, Pterocarpus erinaceus, Trichilia roka, Piliosti~ma thon
nins=:ii.
Si l'on compare les strates ligneuses de Lanto, du Foro
et de Ouango-Fit1ni, on constate que 18 seulement des 35 espèces
d'arbustes et arbres du Foro se retrouvent à Lamto, alors que 32
(plus trois incertaines) sont présentes à Ouango-Fitini (tab. IV).
(Menaut et Dev1neau comm. perso.).
1.1.4.2. Strate herbeuse.
Les plantes herbacées les plus courantes de la savane
du Foro appar-tLennent à six fa..,7iilles vé zé t a l.e s :
cées, ?apilionacées, Z~~!.iberacées, 2ubiacées et
Graminées, Cypera- 1"": • ~ ..Jloscoreacees. La
~lus représentée et la ~lus abonèante est celle èes Gra0inées, qui
t ~ ,.J Lnodi \00' , •• .,. ,corner-en Ancr-ooo ccn ascJ.no a s , ....... scru r-ens i s , hypar~,.en~é'. Si!1~-
thiana., n. subolumosa, HY'Perthelia dissoluta, ?s.nicum ph!"at:::~itorè.es,
Schizachyriurn san~uineum, EIJ~andra anèroor.ila et ~ott~oelia e7.al
tata. (César, comm. per-so s ) ,
(César, 1971) Comparée à celle de Lamto~ la strate herbeuse du Foro
possède un nombre important d'espèces qui lui sont propres. Parmi
celles-ci, notons cino des neuf Graminées, les deux Cypéracées,
deux des sept Papilionacées et l'espèce de Dioscoreacée (tab. V).
L'étude de la croissance globale en taille et poids de
la strate herbeuse au cours de l'année 1974 a été réalisée à partir ?
de trois prélèvements mensuels de 4 rn- de vésétation faitsdans une
parcelle de savane située sur le plateau. En plus de la croissance,
une analyse fourra~ère type a été réalisée, qlobalement, sur chaque
échantillon.
La croissance en taille de la strate herbeuse au~rnente
- 12 -
à peu près régulièrement, dans la mesure où les pluies se manifes
tent, après le passage des feux jusqu'en juillet où les Graminées
atteignent leur taille vé~étative maximale (environ 80 cm).
A partir de septembre, on observe une brusque aug~enta
tion de la taille consécutive à l'apparition des flèches portant
les épis. En octobre cette brusque croissance est achevée et les
épis murissent (fig. 6). La char~e en poids de matière sèche de la
parcelle augmente elle aussi régulièrement de janvier à novembre.
La croissance en poids au cours de la période janvier-avril est
très irrégulière par sùite de sa très forte corrélation avec les
pluies. La charge totale est passée de 150 kg/ha en janvier a
5.400 kg/ha en décembre. La teneur en eau (différence entre le
poids frais et le poids sec de l'échantillon d'herbes) est varia
ble selon les saisons. Elle est normalement très élevée au cours
des ~ériodes pluvieuses (65 à 70 %) et réduite au cours des pério
des sèches (50 à 55 ~). Elle est ~inimale (35%) juste avant les
feux (fig. 6).
Les principaux éléments minéraux ont des teneurs varia
bles selon les saisons (fig. 6). A l'exception du Sodium dont la
teneur est constante toute l'année (0,01 %), l'Azote (maximum 1 %),
le Potassium (maximum 1,3 %) et le Phosphore (maximum 0,14 %) pré
sentent les teneurs les plus élevées dans les jeunes pousses du
début de la phase végétative, en janvier, puis elles décroissent
plus ou moins régulièrement pour @tre minimales dans les chaumes
sèches de décembre. Les teneurs en Calcium (maximum 0,55 %) et en
Magnésium (maximum 0,30 %) augmentent jusqu'en mai puis diminuent
lentement et régulièrement jusqu'en décembre.
1.1.5. Les feux de brousse.
En savane, les feux de brousse sont un facteur du mi
lieu très important : en quelques heures ils peuvent éliminer inté
gralement la strate herbacée qui était à son maximum de croissance,
le feuillage des arbres et des arbustes et une vartie des ~raines
tombées sur le sol.
Des feux précoces (novembre-décembre) mannuerrt ,&,énéra
1ement d'intensité et la savane ne se trouve donc qu'imparfaitement
bru1ée. Il subsiste alors de nombreuses poches de végétation non
bru1ées dont la 'surface unitaire peut atteindre plusieurs hect~res.
Les feux tardifs, ou feux ba1ayants, se pratiouent en janvier et
surtout en février. Ils brûlent la vég,étation herbacée dans son in
tégralité par suite de son très srand état de sécheresse; ce sont
surtout les feux de mise en culture.
Dans la forêt classée du Foro, les ~ardes forestiers et
le responsable du village ont pour consi~ne de mettre en place des
feux précoces, mais à notre demande, en 1978-79 les feux ont été
tardifs (début février) dans le secteur du quadrat de marquage.
Pendant la durée de l'étude du peuplement de ronpeurs.
les feux ont eu lieu en : décembre 1971, décembre 1972, décembre
1973, janvier 1976, décembre 1977, février 1978.
1.1.6. Un milieu type, le cuadrat de marouape de 10 ha.
L'étude en continu du peuplement de ronpeurs s'est
faite dans une parcelle carrée de 10 ha, le quadrat de marGuage,
dont l'emplacement a été jugé représentatif de l'ensemble de la
forêt classée du Foro - à l'e;~ception des forêts de sommet et des
forêts-~a1eries çui ne font pas partie intégrante de la zone de
piégeage mais qui la jouxtent au nord-ouest et au nord-est.
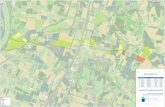
Le quadrat de marquage (QH) est situé à peu près au
centre du domaine protégé. Il est distant de 11 km du laboratoire
(fig. 7). Le dixième de sa surface correspond à l'ouest à un pla-"
teau dont l'altitude est de 280 m. La pente est douce (2 %) vers
l'est et vers le sud-est; elle est forte en suivant la dingonale
- 14 -
est et la bordure sud-est du O~ (4 ~) (fig. 8).
L'étude des sols a été faite à partir de la description
de seize fosses de 140 cm de profondeur réparties rér,ulièrement
sur le QM et d'une fosse de même dimension située dans le bas fond
à proximité de la forêt-galerie. Sur une surface aussi réduite (10
ha), les sols varient peu d'une fosse àl'autre. On peut toutefois
observer une certaine modification le long de la toposéquence qui
va du plateau situé au sud-ouest du QM au bas fond proche de la. . galerie située au nord-est. La caractérisation des sols le long de
cette toposéquence ~eut être faite à partir de.l'analyse de quatre
fosses disposées régulièrement du plateau au bas-fond dont les
caractéristiques physiochimiques sont rapportées dans le tableau VI.
Sur le plateau, où l'horizon meuble atteint 85 cm, le
profil (M) se découpe comme suit :
- de 0 à 14 cm, une couche humifère brun-noir, argilo-sableuse de
venant sablo-argileuse, s'enrichissant pro~ressivement en petits
nodules ferrusineux et en éléments quartzeux, riche en racines, de
structure continue, de cOhésion faible à moyenne, où la porosité
tubulaire fine et moyenne est très inportante,
- de 14 à 85 cm, une couche de sol brun-roupe à rou?-e sablo-ar~i
leux à arf,ilo-sableux s'enrichissent fortement en petits nodules
ferrugineux jusqu'à 53 cm, profondeur à partir de laquelle aopa
raissent des matériaux friables provenant de l'altération des ro
ches. On note la présence de quelques racines dans toute l'épRis
seur de la couche. La structure est arpileuse fine, la cohésion
moyenne, la porosité tubulaire moyenne est bien développée,
- de 85 à 140 cm, une couche d'altération des roches, friable et
riche en éléments de la couche précédente devenant pro~ressivement
dure. L'altérite y est riche en éléments de quartz de 0,2 Q 0,5 mm
de diamètre.
A mi-pente, la séparation entre l'horizon des sols meu
bles et celui des sols durs est très irré~ulière. Elle se situe
entre 59 et 103 cm. Le profil (J) se découpe co~me suit:
- de 0 à 35 cm, la couche humique appauvrie en matière or~anique de
couleur gris-brun, sableuse faiblement ar~ileuse, avec de nombreu
ses racines, s'enrichit progressivement en petits nodules ferrugni
neux et en quartz. La structure, d'abord particulaire, devient
argileuse, mal individualisée. La cohésion est très faible à faible.
La porosité importante est d'abord interstitielle puis devient tu
bulaire fine
- de 35 à 81 cm, la couche des sols brun-rouge à rouge sablo-argi
leux très riches en nodules ferrugineux de 0,5 à 2 à 3 cm, où l'on
rencontre également des éléments quartzeux de même dimension. La
structure y est anguleuse fine, mal exprimée. On y trouve un peu
d'humus et quel~ues racines. La cohésion est faible ~ moyenr.e et
la porosité interstitielle et tubuleire fine très i~~ortante ;
- de 81 à 140 cm, la couche des matériaux d'altération des roches
est dure, riche en quartz (0,2 à 0,4 ~m) et en sabl$~rossiers. 0n
y rencontre S ~ lS % de sols brun-rouge de la couche ~r~cédente.
En bas de pente, la séparation entre l'horizon des sols
meubles et la zone des sols d'altération des roches est également
irrégulière et ondulée. Le profil(D) se découpe comme suit:
- de 0 à 33 cm, la couche humique gris-brun foncé à brun foncé,
sableuse comprend une très forte proportion de racines. Elle s'en
richit progressivement en nodules ferru~ineux et quartzeux de ~etite
à moyer~e dimension (0,5 à 2 à 3 cm). La structure est particulaire
à faiblement anguleuse. La cohésion est très faible à nulle. La po
rosité interstitielle fine est très élevée ;
- de 33 à 52 cm, la couche des sols brun-rou~e sableux fai=lernent
argileux est riche en humus et en racines et com~orte une forte
;roportion de nodules ferrurrineu;: et auelques éléments cuartzeux.
La structure est particulaire et partielle~ent an~ulaire r.al indi
vidualisée. La cohésion est tr8s faible et la porosité intersti
tielle, fine ~ ~oyenne, tr~~ forte ;
- 16 -
- de 62 à 140 cm, la zone d'altération des roches se compose de
blocs de carapace et de cuirasse, de fra~~ents quartzeux et de ma
tériel de remplissage de la couche précédente tacheté et compact.
On y note la présence de placages noirs d'oxydes (fer et rnanganè se ) ,
Dans le bas fond à p~oxirnité de la r:ale~ie forestière
du marigot, le profil pédologique (0) se découpe co~rne suit:
- de 0 à 9 cm, une couche humifère grise très riche en sables et
en racines, à structure particulairs parfois en a~régats arrondis.
La cohésion est presque nulle, la porosité interstitielle et tubu
haire fine est très élevée ;
- de 9 à 111 cm, une couche sableuse Sris-clair à beise-clair conte
nant d~ns sa portion supérieure cuelques nodules ferru~ineux de
petite taille (0,2 à 0,6 cm) des racines et de l'humus ~uis, au
delà de 30 cm des blocs de cuirasse friable (juscu'à 15 cm de èia
~ètre) des éléments quartzaux açFlo~érés ~ar des oxy~es de fer e~
de ranGanèse, ,enfin au-delà de 64 cm des blocs de c,uartz de toutes
tailles (quelques centimètres à 30 à ~o cm) mélangés aux blocs èe
cuirasse friable. La structure du matériau sableux est particulaire,
sa cohésion est très faible à nulle, sa porosité interstitielle et
tubulaire fine est très importante.
- de 111 à 140 cm, la zone d'altération des sols jaune-brun à brun
très pâle est très friable. Le sol est compact et argileux. On note
la présence de fines paillettes de muscovite.
1.1.6.3. Vé~étation.--.....-------
1.1.ô.3.1. La strate li;.neuse (fip. 9).
Sur la surface de 10 ha, on a dénombré 9287 pieds, touf
fes et fourrés de li~neux soit une densité de 929 à l'ha. Les treize
espèces les plus abondantes sont Lophira lanceolata (2~,4 %), Afror mo s i a 'axi-"10ra (, .../ ~ C~) ~l'+-V1"'os-e1"'mu'" oar-adoxum (le:; .., ,":) T;)~rin""1"'i•• 1 .c. _ • ~ _ _,"'" i"J , _'.J.." .. _ ,'..,;' _ .J l'I v _ J-..ul:: _ ,~ 1'" , ;. _ ... c:.._ ...
cu .... a ....e".;-"oli'" (12 3 ~') '"I"iS ï''''n';0ll~'''' oli're1"'; (7 ,4 ~') m_ i C " i 1 '; "... 1"" __ ...L.":" ..r...c;::. ,;J ;:",\",oi"'" :.Je-.... ...L.__ ~.:....' ..... _ _ ,~ ,) , J..:., ... ... .:.__ ..... c-..;.
k ( ' 8 ") p.... . (~ - "1) ':l • , . " ....'" • "ro a L~, ~J,. l.eroc~.r!Jus er~né1.ceus ... ,0 /J , ~ ~_~osï;~~mê. "'1,onn~n1~~
( - 2 "/) .,) , '.'.', Cochlos-:;e!'r::u:7i pl3.nchoni (3,2 ;j), Annon9. sene';<:alensis (1,4;:~) •
- 17 -
Ue.naca tocroensis (1,4 %) à la li s i ère des "forêts", Terrninalia SOD
(1,1 %) et Hvrnenocardia acida (1 %). Les 17 autres espèces repré
sentent à elles toutes 3,7 %.
L'essentiel des arbrisseaux (520/ha) est constitué par
les mêmes essences que celles listées ci-dessus, excepté Uaoaca
toxoensis. A l'exception de Cochlosoermum planchoni mais avsc en
plus Bridelia ferrurr.inea, les mêmes espèces forment la ~uasi tota
lité de la strate arbustive (204 pieds/ha). Enfin, exce9té C. Dlan
choni, P. thonnin~ii, T. roka, E. ferru~inea, A. sene~alensis et
H. acida, les mêmes espèces auquelles il convient d'ajouter les
ficus représentent 97,4 % de la strate arborée (47 pieès à l'ha).
Les touffes (151 à l'ha) sont constitué$surtout par A. laxiflora
(35,8 %), L. l~"1ceolata (30,8 %) et C. plancr.oni (14,7 ;~) puis par
P. thonni:1;ü (6,7 %), i3. '::ar2.ào~('Jrr. (5,7;)), P. curatelli folia ( , 7<:'~' -, :.~ l ,
T. roka (1,4 %) et D. oliveri (1 %). Les fourrés (S,3 i l'~a) sont r 1'" '"c '" 0 1 ..,t- '" (, a ."-' =<) ,..; e~ • _ .. ;. _ _ c . .... ;;,.... __" ,,,,J ". _
1 . • • (- ., .') L -', . (O'CSC:1Y~ 0,..,;:., a zone ce p i a t aau est
la plus fournie en li;neux (1215 à l'ha) où A. laxiflora est ~ette-
t- 1 • • ( ? - .., '-') ., d -t:':en w c.Or7i.lnan'C _0 , ~ ;.: suaV:l. eL. 1 anc eo ï a t a (·7 ,.., -')_c:;;o,.l.4 _ "- ...~'- J.. ,0 ;'..; , 3. :-.:arado:-:um
(16,1 %) et P. curatellifolia (15,4 %). Les pentes bien drainées
(800 pieds à l'ha) sont surtout peuplées de L. lanceolata_,(45,4 %)
et de A. laxiflora (12,5 %). Le bas-fond (665 pieds à l'ha) est
surtout peuplé de P. curatellifolia (42,3 %) et de L. lanceolata
(14, 5 ~;).
minées (fi~. 10). Globalement, les espèces les olus aconè2.ntes sont
Androco~on ascinodis (17,6 %) et Schizachyriu~ san~uineurn (16,9 ~~),
puis Elyme.ndra anèron!"'ilê, (14,1 ~0), H:l~arrhenic?. sr.:ithi.:'tna (12,1 ;~),
". h ..... -ct ('1 C -') . d . . . ( 1) 7 c : _ ...: an.lCUI7l n ra.,<r::1. "'o~ es Jo , Q :.~ , r.n rODor:on scnl!'ensJ.s :, ,.' J '= '"
'-'v'"'ar-l-,e,..li~ subc Lurno s a (f"' 7 C')"'., '.., .1. "' ...._~. ~_ ::: ç-. =' , /0). Enfin on trouve Hyperthelia ~isso-
Lu t a (~ 1 cr ,.....eru.urn -- (? ,'1 "') et- ":lo......... o e La a :"'~1"'''''''- (0 ~ ,")~ .,- ',)1, """l",,\;;;t ...1 • ..:::1 .... _,"'" /"; •.,. ... I..tv'V _<;.0. _ ..... c::._;..,c.;.""'.,;i.. ,':J ,'J,.
Sur le plateau les principales espèces sont S. san~uineu~ (18,2 ;j),
2. androDhila -') et" ~,-,~",.,.-; i-oIr'os (, ... , C~)1...' ... ~ ..·",1"'C;:.I'.11;.l,J ....... _ . _, ,_ ; ....
- 18 -
nodis (25,4 %), H. subplumosa (18,7 %) et s. san~uineurn (16,2 %).
Le peuplement herbacé des pentes bien drainées est intermédiaire
entre ce~x des plateaux et des bas-fonds ; les princi~ales esp8ces
y sont A. ascinodis (18,4 %), H. subolumos2 (16)1 X), s. san~uineum
(15,7 %) et E. androohila (14,3 %).
1.2. Matériel et techniques. ======================J
nous ne rapportons ici que le matériel et les techniques
servant à recueillir les informations de base relatives aux Ronge~rs,
c'est à dire le 9ié~ear.e, l'éleva~e, les prises de mesures sur les
spécimens, etc •••
èe hase (méthodes de calcul des densités, analyse dérr:oSraphic:ue,
etc ••• ) seront traitées dans leurs chapitres respectifs.
1.2.1. Les oièqes et les oiépeages.
Trois modèles de pières ont été utilisés au cours de
notre étude. Deux modèles permettent la capt~re des rongeurs vi
vants (Chauvancy et piège-couloir), le troisième les tue (tapette).
1.2.1.1.1. Pièf.e Chauvancy (licence CNRS).
C'est le modèle de p1ege le plus utilis8 puisqu'il est
seul sur le quadrat de marquage (1024 pièges) et la majeure partie
des lignes exhaustives (1050 piè~es mensuellement de 1971 à 1974
et environ 400 pièges entre 1976 et 1979). Le pièpe Chauvancy se
compose d'un "couvercle" comprenant le mécanisme de fermeture 8,utO-
matique et de verrouillase de la porte lorsc:ue l'ani~al touche
l'appât ~lacé au bout d'une ti~e métallique servant de fléau et
ct' une "boîte à nid" r'e pr-é s errt é e à l' or-t r-t ne par une boite de conserve
- 19 -
d'un kilogra~me sur laquelle s'emboîte le couvercle. Pour réduire
le volume considérable représenté par plus de 1000 boites de conser
ve nécessaires au piégeage mensuel (volume très supérieur à la
contenance d'une camionnette bâchée) nous avons fait réaliser des
boîtes tronconiques en aluminium. Elles sont plus longues que les
précédentes (donc permettant la capture d'animaux de plus [rande
taille et réduisant la mortalité des captures par suite du confine
ment), mais en s'emboîtant les unes dans les autres, elles occupent
un volume extrê~ement réduit ~Rr rapport au précédent modèle (de
l'ordre de 1 m3).
Une amélioration dans le rendement des piè~es Chauv~~cy
a été obtenu par l'utilisation non plus de boîtes tronconiques en
aluminium, mais de boîtes tronconiques en grillage à petite maille,
permettant une reilleure diffusion de l'odeur de l'appât, la cap
ture plus aisée des i~dividus et des espèces présentant un co~~or
tement claustrophobe, et enfin une diminution de la nortalité dûe
au coup de chaleur dans les boîtes fermées.
1.2.1.1.2. Piège-couloir (ratière Manufrance).
C'est un parallèlépipède rectangle de 42 cm de long, de
14 cm de section carrée. Avec ses parois en fil d'acier galvanisé,
ce piè~e présente les avantages énumérés dans le cas du Chauvancy
à boîte tronconique en ~rilla~e. Afin de permettre la capture des
~us et des jeunes individus des espèces de taille moyenne (Lemnis
comys, Myomys, etc ••. ), les parois sont doublées, avec du grillaŒe
métallique du type moustiquaire. Le mécanisme de fer~eture-verrouil
la~e est constitué de deux portes placées aux deux extrémités du
parallèlépipède, bloquées en position ouverte par une tise porte
appât placée au centre du couloir et par de puissants ressorts as
surant le verrouillage en position fermée. Ce modèle de piè~e par
ses dimensions permet la capture d'individus et d'espèces de srande
taille (Tatera, Dasymys, petits carnivores, hérissons etc ••• ). De
plus, la présence des àeux ouvertures semble accroître le rendement
du pièf:e par rapport au Chauvancy et ~err.et la capture d'espèces
r-éou t é e s Lncao t ur-ab l e s avec le Chauvancy (Arvicanthi s, ~I.vloï.'Ys).
- 20 -
tellement courante qu'il n'est pas nécessaire de les décrire ici.
Notons toutefois que nous avons utilisés des tapettes à socle en
bois et à mécanisme galvanisé qui leur assure une bonne tenue en
climat tropical humide.
L'inconvénient principal de ce type de pièp.e est de
fournir des animaux morts et presque toujours décom90sés, inter
disant les prélèvements d'or~anes (yeux, organes génitaux ••• ) et
rendant aléatoires les mesures prises sur le CorDS. Le crâne étant
souvent fracassé, il ne peut plus être utilisé au cours de l'étude
taxinomicue.
Les avanta~es èe ce modèle de ~ièrre sont doubles. ~n-" n 1 - "-
tuant l'animal avant qu'il -ait consommé l'appât, on peut à partir
des captures, réaliser des analys~s de contenus stomacaux (à con
dition d'inspecter les piègeages à de très courts intervalles de
temps). De plus, on constate, à l'usage, que ce modèle de piège est
le moins sélectif, le plus commode d'emploi et celui avec lequel
les rongeurs présentent le temps de latence à la capture le plus
court.
Selon le but recherché nous avons utilisé différentes
méthodes de piégeage. Au cours de la phase d'inventaire des espè
ces nous avons utilisé le piéseage en tache, oG une quinzaine de
pièges (nombre très excédentaire par rapport au nombre de rongeurs
potentiellement espéré) sont disposés sur une surface modeste (en
viron 100 m2 ). Dans ce type de piégeage, tous les modèles de piè
~es sont utilisés afin d'augmenter les chances de collecte du plus
frand nombre d'espèces présentes dans le milieu retenu. C'est cette
méthode de piégeage aui a été employ~dans les cultures, les forêts
zaleries et de plateau, dans les villages, etc ••.
- 21 -
En savane, les piépeares exhaustifs systématiques sont
effectués sur des li~nes unitaires de 50 piè~es Chnuvancy. Entre
1971 et 1974, deu~= longueurs de lignes furent utilisées : d'une part
des 1i~nes de 245 m de long (50 p~er.es espacés de 5 m), d'autre part
des lignes de 98 m de long (50 piêges espacés de 2 m). Les pi~~es
restent tendus pendant quatre jours consécutifs, fourniss8.nt auatre
résultats quotidiens, soit un effort de piéQeage de 200 journées
pièJe par ligne unitaire conduisant à un rendement théoriouemaxi
mu~ de 200 captures. En plus du piégeage exhaustif standard, nous
avons disposé durant cette même oériode trois liBnes unitaires de
98 ~ composées de tapettes •. Au cours de la période 1976-1979, les li~nes de 245 m
de long ont seules été utilisées. Le temps de pié~ea~e a été ramené
à trois jours, soit un effort de piégeage de 150 journées-piè~e par
ligne.
Le quadrat de marquage a été mis en place dans le but
d'obtenir àes informations sur l'étho-écologie des !on~eurs et la
dynamique de leurs populations dans le même milieu~ Le site a
été choisi de façon à inté~rer sur la surface du piéfeape l'ensem
ble des grands faciès botaniques et topo-pédo1o~iaues de la station.
Nous nous sommes fixé une surfaceœenviron 10 ha (exactement 9,61
ha) qui soit compatible avec nos possibilités de piéç,ea~e (en hom
mes et en ~ntériel) Ovi puisse répondre aux conditions de milieu
i~~osêes et oui fournisse des échantillons auotidiens suffiscmment
import~nts. L'espacement entre deux pièges est de 10 m ; cette va
leur a été déterminée au cours d'un ~iépeage avec maraua~e réalisée
en 1974 sur une surface de 5 ha comptenant des piê2es disposés se
lon un espacement de 7 m. L'ensemblè du quadrat comprend 32 1i~nes
de 32 piè~es, soit un total de 1024 pièses. Les pièges restent ten
dus pendant ouatre jours consécutifs ; ils sont visités chaque ma
tin. L'effort total mensuel de piégeage a donc été constamment de
4096journées-piè~e.
1.2.1.3. L'anoât utilisé.--------------- Pendant toute l'étude et quel que soit le type de piè
Seage, l'appât utilisé a été la noix de palmier à huile. C'est un
fruit de petite taille parfaitement adapté au piégeage. Il dure
quatre, voire cinq jours dans les pié~es sans se détériorer. Il est
bien consommé par toutes les espèces de rongeurs et disponible du
rant tous les mois de l'année~
1.2.1.4. Effort de vié~ea~e.------------------ Par suite de la granàe diversité des techniques de pié
geage utilisées par chaque auteur, l'effort de pié~eage déployé au
cours d'une étude ne peut pas être caractérisé par le nombre de
pièges ou par le nombre de piégeaf:es unitaires déposés. Suiv~~t
l'école ~7.éricaine, l'intensité de ~iégeege que nous avons dé~loyée
au Foro est esti~ée par le nombre d~ pièses présents sur le terrai~
par fraction de 24 neures . L' uni té ct' intensi té de pié~ea~e est donc
la journée-piège ("night-trapll des Américains).
Dans le cas des lifnes exhaustives utilisées pour rea
liser le piégeage systématique mensuel, l'effort total a été de
162.400 journées-pièg~(812 lignes de 50 pièges posées penda~t qua
tre jours consécutifs), dur~nt la période 1971-74 et de 44.400 jour
nées-pièges (296 lignes de 50 pièges 90SéS pend~~t trois jours) au
cours de la ~ériode 1976-1979.
Le premier auadrat de marqua~e expérimental mis en place
en 1974 avait nécessité 24.000 journées-pièges et le second quadrat
de marquage, qui a fonctionné de 1976 à 1979, a nécessité 147.456
journées-pièges. L'effort total déployé au cours de cette étude a
donc été de 378.756 journées-piè~es.
Le rendement des piégeages est estimé ~ar le nombre de
captures pour 100 journées-9iè~es.Le rendement $710b2ol mo ye n des
piégeages exhaustifs a été De 2,16 captures pour 100 journées-piè~es.
- 23 -
Dans le cas des lirnes de 98 m composées de tapettes
espacées de 2 m, le rendement moyen a été de 2,8 captures/100 jour
nées-piè~es. Dans celui des li~nes de 98 m composées de pièges Chau
vancy espacés de 2 m, le rendement moyen a été de 1,88 captures pour
100 journées-pièges. Dans celui des lignes de 245 m composées de
pièses Chauvancy espacés de 5 m, le renèement a été de 2,58 captures
pour 100 journées-pièges, soit un rendement moyen par fractions de
98 m de 1,03 captures pour 100 Journées-pièges. Enfin dans le cas
des lif-nes de 245 m composées de pièges Chauvency espacés de 5 m
posés pendant trois jours, le rendement moyen a été de 2,20 captures
pour 100 journées-pièges soit 0,88 captures pour 100 journées-pièges
par fractions de 98 m•.
1.2.1.6. ~~!~~~!~!~~_~~~_~!f!~~~~~~~_~~~~~!g~~~_~~_E!~g~~E~
~!~_~_~!~_~~~_~~E~~~~_~~_~~~g~~~~·
ments, on peut définir l'action sélective des différentes techniques
de piégea~e utilisées sur les espèces piér-ées. De fait les résultats
de piéReage montrent bien que l'importance relative de chaque es
pèce dans le peuplement peut subir des variations plus ou moins im
portantes selo~ la lip,:ne de piér:.eage considérée (fiF. 11). Des dif
férences, peu importantes sont notées entre les snectres se rappor
tant aux deux modèles èe lignes ChauvancY. Elles ont trait essen
tiellement aux Benres Lemniscomys et f1yomys. Des différences bien
plus marquées s'observent au cours de l'utilisation des lignes de
t.ape t tie s , Dans ce cas, on constate une réduction irnportante du pour
centap,e de Lemniscomys leui passe de 54,5 % dans les lignes de pièges
Chauvancj' à 43~1 % dans les li~nes de .tapettes, conco~mittante cvec
un accroissement importent de celui des i~astomys qui passe de ~oins
de 1 % dans les pièges Chauvancy à 8 % dans les tapettes. Uranomys
et Granhiurus présentent une aupmentation moins marquée èans les
li~nes composées de tapettes avec respectivement 2,4 ~~ contre 1,6 % et 3,8 % contre 1,2 %.
- 24 -
Les individus capturés dans le quadrat de marcuaie sont
anesthésiés dans un bocal d'un volume de deux litres contenant un
tampon de coton imbibé de quelques gouttes d'éther éthylique. Dans
ces conditions, l'anesthésie est très rapide et dans la mesure où
l'on suit très attentivement les réactions de l'animal pendant la
phase d'aspiration de l'éther, le taux de mortalité est très infé
rieur à 1 %. Les genres ,les plus sensibles aux vapeurs d'éther et
donc chez lesouelles il y a risque de décès par surdose, 'sont Uyomys
et ~us, puis Lemniscomys, Granhiurus, Uranomys et enfin Tatera.
1.2.2.2. Observations sur les animaux caoturés dans le ouadrat -------------------------------~--------------~------
Après avoir été enesthésiés,tous les individus capturés,
marqués ou non, sont pesés et mesurés(lon~ueur tête et cor~s du
nez à l'anus, longueur de la queue et du pied avec griffes). L'état
mor-pho Log â que apparent de la sphère p:éni tale exter-ne est noté (déve
loppement et pbs1tion des testicules, coloration des bourses chez
le mâle, développement des tétines et lactation, festation visible,
ouverture vag.inale chez la femelle). Puis les individus nouveaux
sont marqués par amputation de doigts. Le code de marcua~e est un
code à base 5 pour les pieds et un code à base 4 pour les mains.
Cet artifice permet de ne Das couper plus d'un doist par membre
tout en lai~sant la ~ossibilité de numéroter un nombre suffisant
d'individus, compte-tenu des densités rencontrées. A~rRs le réveil
les individus sont relechés au point de capture. Ils sont suivis à
l'oeil afin de contrôler Qu'ils n'entrent pas à nouveau immédiate
ment dans un piè~e èu voisinase. A cette occasion, leurs terriers
ou leurs nids sont éventuellement repérés.
- 25 -
Après avoir été tués par l'inalhation d'un excès de
vapeurs d'éther éthylique ou d'éther de pétrole, les individus sont
placés en décubitus dorsal comme lors du marquarre. La longueur tête
etcorps et celle de la queue sont notées, ainsi que la lon~ueur de
la patte postérieure. Les observations morpholo~iques concernant la
sphère génitale sont notées puis l'animal est autopsié. Chez le
mâle, les testicules, les vésicules séminales, les épididymes sont
prélevés et fixés dans le liquide de Bouin. Chez la femelle oh re
cherche les traces de ~estation, on note alors le nombre d'embryon~
dans chaque corne utérine. Les cicatrices placentaires, témoIns de
gestations précédentes, sont également recherchées et dénombrées.
Puis les ovaires et les cornes utérines sont prélevées et ég,alement
placéS dans du liquide de Bouin. Pour les deux sexes les yeux sont
prélevés et fixés da~s du formol à 10 %. La ?eau est prélevée puis
traitée en vue de sa conservation en collection. La tête est é~ale
ment prélevée, débara~sée du cerveau, séchée àU. soleil ou à l'étuve
à 60 0 C puis placée dans un élevage de Dermestes qui consomment la
chair désséchée sans abimer les os du crâne. Ce dernier est blanchi
dans une solution à 30 % ct' eau oxyrréné e , séché puis mesuré et placé
en collection.
Les yeux sont fi;::és pendant 30 jours dans une solution
de formol à 10 %. Les cristallins sont alors extraits et clacés
pendant cuanze jours dans une nouvelle solution de formol 2' 10 ~:'.
Ils sont ensuite· désséchés dans une étuve ventilée à 105 0 C pendant
des durées variables selon leur taille. Pour Le~niscomys et Uranomys
la durée de dessication est de trois heures, quatre heures pour
I.1yomys et seize heures pour Tatere.• Après s'être . refroidis dans
un dessicateur contenant du silicapel, les cristallins secs sont
pesés sur une balance à nacelle précise au 5/1000 de m~.
- 26 -
L'éleva~e des ron[eurs du Foro a été réalisé dans la
salle d'élevare du laboratoire situé sur le Centre ORSTOE d'Adiopo
doumé, à proximité d'Abidj~n. Cette salle est une pièce de 50 ~2
dans l~quelle sont disposés des portoirs à cages. La température
observée est en moyenne celle de l'extérieur, mais tamponnée.
Les ca[es sont des bacs en plastique recouverts de cou
vercles en fil d'acier zinfué. Les cages individuelles sont des
ca~es à souris, qui mesurent 30 X 20 X 18 cm. Les capes pour couples
sont des cafes à rats, qui mesurent 50 X 25 X 18 cm.
Dans chaque ca~e on place de la litière composée de
sci~re de bois. Une botte de conserve d'un kilo, à laouelle est
adjointe un piè~e Chauvancy à la place du couvercle sert de botte
à nid. Le fond de cette botte est percé d'une fenêtre triançulaire
permettant aux animaux d'entrer et sortir à volonté de la boîte à
nid. Dans la botte ~ nid on dépose une p~7née de frison. La ferme
·ture de la fenêtre trianpulaire lorsque les individus sont dans le
nid permet leur manipulation ou bien le nettoyaSe de la cage olus
aisément.
granulés pour souris et de ~ranulés pour lapins en reproduction ;
chaque jour on ajoute des morceaux de papaye mûre (apport d'eau),
deux fois par semaine des noi;( de palmier à huile et une fois ~ar
se~aine une dose de mïl sermé.
Chaque jour, les naissances sont recherchées et notées.
Les jeunes sont pesés à la naissance, au sevrape (15 jours), ~uis
mensuellement à ~artir de l'â~e d'un mois.
70us les ~enres, sauf nus, ont pû être maintenus en
éleva~e sans problèmes particuliers. Nous n'avons pas obtenu de
reproduction chez Arvicanthis, Steatomys et Mylomys.
- 27 -
Une part importante de la taxinomie des ron~eurs est
fondée sur des mesures du crâne et des dents. Nous avons donc choi
si vingt mesures qui décrivent le crâne et les dents (tac.VII)
(fig. 12). Toutes les mesures sont prises à l'aide d'un ~ied à
coulisse à cadran;précis au 5/100 de millimètre.
Afin d'avoir une idée plus précise de la variation des
mesures en fonction de l'âge des individus, nous avons estimé, sur
les crânes des Lemniscomys et des Ï'Tyomys ayant servi à l'étude de
la croissance des cristallins (voir chaplII), la valeur moyenne des
mesures retenues pour des âges allant de 1 à 8 mois (excepté 7
mois). Comme Verheyen et Br-actee (1966), nous avons constaté (lue la
croissance du crâne des rongeurs est très corrélée avec l'âpe des
individus (r généralement supérieur à O,SO). Seules font excention
les r&~~ées molaires dont la taille est fixée dès l'éru0tion de la
troisième molaire. La lar2eur de la première ffiolaire supérieure
(lM1) est corrélée avec l'âse par suite de sa forme tronconique
ainsi lors de l'usure des lames d'émail la largeur de la dent au~
mente au fur et à mesure que l'on va vers la base du tronc de cône.
Enfin la largeur de l'intermaxi11aire prise derrière [:[3 - 1,j3, con
trairement à toutes les autres mesures diminue avec l'âge, le coef
ficient de corrélation est alors négatif (fig.13a).
Le taux de variation entre un et huit mois (fi~.13b),
est très différent selon la nature des mesures. Celles 'se rappor
tant aux molaires sont très stables (Rmx, Rmd, IM1). Les mesures
présentant les plus forts taux de variation se rapportent à la par
tie antérieure du crâne et particulièrement au rostre (D, l?m, P,
In, hU, LN, Fi et partiellement Cb). On note eue 10 varie ~eu et
oue Ez varie nlus que 1Bz qui lui est ~ostérieur. Enfin l'étude du
taux de variation des mesures chez l'individu p1einesent adulte
entre 4 et 8 mois (fiŒ.13c) confirme les résultats précédents. Dans
l'ensemble les mesures sont stabilisées et leur variation n'excède
pas 5 %, exceptés certaines mesures crises dans la partie antéri
eure du crâne (D, P, Fi, 12 et Ur).
- 28 -
IDENTIFICATION ET TP~ITS GENEPAUX DES ESPECES
Au cours de la période 1971-1979, on a capturé 5057
spécimens de Rongeurs de la région du Foro. Ces individus provien
nent essentiellement du milieu naturel (savane, forê~8aleries,
for@ts sèches) et pour une plus faible part des milieux cultivés.
L'ensemble des captures se répartit en quinze genres. Ce sont~pour
les Gerbillinés : Tatera. et Taterillus ; pour les ~;lurinés : I:Ius,
Arvicanthis, Lemniscomys, Mylomys, Uranornys, Thamnomys (Gremmornvs),
Praomys, nyomys, Mastomys, DasymYs et Hylomyscus ; pour les Dendro
murinés : Steatomys ; pour les Graphiurinés Graphiurus.
2.1.1. Tatera.
groupes panafricains de Gerbillidae en Afrique de l'ouest; des
individus du groupe des T. robusta représenté uniquement ~ar
T. guineae et àes individus du groupe des T. valida représenté par
trois espèces T. weï.mannt , T. hO:Jkinsoni et T. kernpL,
En 1959 ?etter a montré Gue Tetera valida n'était pas
dans l'ouest de l'Afriaue. La plus ancienne forme nommée dans cette
réfion étant T. kempi, on èoit donc parler du Sroupe kemni.
- 29 -
des deux ~roupes reposent sur des caractères de ~orpholopie cor
porelle et crânienne et sur les formules chromosomiaues. En ce
qui concerne les caractères de morphologie corporelle , on cons
tate que T. puineae est de taillelépèrement plus faible, la colo
ration dorsale est plus roussâtre devenant franchement fauve sur
les flancs. Sa queue est proportionnellement nettement plus lon
gue que le corps et elle présente un manchon de poils 'bruns sur
la moitié terminale nettement plus touffue dans sa partie terminale.
Les pieds paraissent plus allongés pa~ce que plus étroits que chez
les Tatera de l'autre groupe. Au niveau du crâne, T. F-uineae pré
sente un rostre plus étroit et plus fin par suite du développement
moins important des prémaxillaires et de la plus faible larrreur
des incisives qui les font ressembler à celles de Taterillus. En
résumé, T. çuineae est morphologiquement plus proche de Taterillus
r-racilis alors que les Tatera du ~roupe kemoi sont plus trapus et
d'aspect plus murin.
Enfin, les formules chromosomiques sont nettement dif
férentes, 2atthey (1954) et Matthey et Petter (1970) ayant dé nom-
bré :
pour Tatera hookinsoni 2N = 48 ",
pour Tatera p:uineae 2N = 50 " pour Tatera ;'!a,mbiana 2H "- 56 "
Dans une étude cytopénétioue et taxinomioue plus ré
cente, Matthey et Petter (1970), ont confirmé pour le sud de la
Haute-Volta (Bobo-Dioulasso) la présence des deux groupes rappor
tés ci-dessus, celui des T. kempi étant représenté par T. honkin
~. T. hookinsoni semble plus abondant (12 spécimens) que T. ~ui
~ (5 spécimens) parmi l'ensemble èes spécimens collectés. Pré
cédemment, Heim de Balsac et Lamotte (1958)1 pour la répion quinéen
ne du ;'1ont Ni.mba , puis Heim de Balsac (1967) pour Lamto, avaient
- 30 ~
rapporté l'ensemble àes individus de ces deux localités à Tatera
kernDi.,
Pour la C6te d'Ivoire (fig.14) nous n'avons pas eu la
confirmation cytosénétique de la présence de T. kempi. En revanche,
avec Tranier (1980) nous avons noté la présence, aussi bien au Foro
qu'à Lamto, de T. hopkinsoni (du groupe T. kempi) dont la formule
chromosomique a ét~ déterminée comme étant égale à 2N = 48 chromo
so~es. A Bouna, dans le nord de la C6te d'Ivoire; avec Bellier
(1967), nous avions obtenu, pour la majorité, des individus réfé
rables à T. hopkinsoni'et, en plus petit nombre, des individus ré
férables à T. ~uineae, ce qui est en accord avec les observations
faites par Matthey et Petter (op. cit) et ce qui tend à confirmer
les affinités nettement soudanaises de cette dernière espèce.
2.1.1.1. Descriotion.-----------
d'un Rattus~ Sa coloration àorsale brun-fauve dans la partie mé
diane s'éclaircit sur les flancs. Le ventre est blanc àe la bou
che à l'anus. Sa queue, d'une longueur au moins égale à celle du
co~ps, est brun-marron dessus et blanc-grisâtre dessous. Les oreil
les sont relativement 8randes et les yeux globuleux. Les pieds et
les mains sont blanch§tres à la partie supérieure. La sole des
pieds est brun fonc6, l'intérieur des mains est clair.
Les mensur~tions corporelles prises sur l'échê-ntillon
d'adultes sont rap~ortées d~nG le tableau VIII. La comparaison des
moyennes entre sexes fait apparaître des différences significa
tives pour le poids corporel et la taill~ tête et corps, les mâles
étant plus rrands et plus gros que les fe~elles.
Le crâne de T. honkinsoni est caractérisé par sa for~e
allonrée surtout dans sa partie antérieure, et par sa boite crani
enne large. Les bulles tympaniques sont très développées (30,4 ~~
de Cb).
Le foramen incisif est IonS ct atteint le bord anté
rieur de Ml. Le foramen oalatin s'étend de la moitié de Ml à la
moitié de M2• Les incisives supérieures relativement étroites sont
nettement opisthodontes et gravées dans le quart externe d'un pro
fond sillon longitudinal. Les rangées molaires sont composées de
dents trapues construites sur un plan de base àe Cricétidé. Les
principales mesures prises sur le crâne et les dents sont rappor
tées dans le tahleauVDI.La cornr-araison des moyennes entre sexes,
comme pour les mensurations corporelles, fait acparaître des diffé
rences sir,nificatives pour l'ensemble des mesures se rapportant à
des Lonzueur-s , Celles qui concernent des lar.rreurs ne sont pas diffé
rentes except~Io et lPm. Celles se rapportant à des hauteurs sont
significativement différentes excepté ~1. Enfin les longueurs des
rangées molaires (Rmx et Rmd) ne sont pas différentes entre sexes.
Au Foro, Tatera hookinsoni se rencontre dans tous les
types de savane, que ce soit sur les plateaux, les pentes pas trop
fortement drainées, les bas de pentes. Il n'a été piép.é eue très
occasionnellement au bord des forêts ~aleries et un peu plus fré
que~ent dans les forêts sèches de plateau. On le retrouve dans les
cultures vivrières (riz, maIs). Il se capture facilement avec les
piè~es, particulièrement avec les tapettes. Il est exclusivement
nocturne et c'est l'une des proies principales de la chouette ef
fraie. Comme tous les serbillidés, Tatera hopkinsoni est un !ouis
seur puissant ; il est ~apable de creuser ses galeries aussi bien
dans les sols latéritiques à fort pourcentape de ~raviers (concré
tions latéritiques et fragments de quartz plus ou moins ~ros), que
dans des sols plus argilo-sableux des pentes et des bas de pentes.
Son terrier se compose de plusieurs ~aleries communiquant avec la
surface par une ou plusieurs ouvertures dont l'une au moins est
particulièrement bien repérable par suite de l'accumulation des
rejets de terrassement. Les ~aleries peuvent atteindre un mètre de
profondeur. Son régime alimentaire semble se composer de rhizomes,
de bulbes et en saison de praines de graminées. On a é~alement
- 32 -
2.1.1.3. Observations biologioues. ------------------~-----
L'ensemble èe la population pié~ée se compose àe 236
mâles et de '274 femelles, soit un taux de masculinité de 0,86 non
significativement différent de l'unité. Dans le milieu naturel
les plus jeunes femelles sont actives lorsque leur poids atteint
75 g. En captivité, la seule femelle née en élevage qui ait été
.gestante l'a été à l'âge d'un an, après une durée de vie com~une
èe onze mois avec le mâle. La durée de gestation estimée par l'in
tervalle minimum entre deu;c mises-bas consécutives semble êt~e
voisine de 31 jours, ce qui est sensiblement plus 10nS que l'ob
servation de 25 jours faite par Hubert (1973) chez Tatera ~~~biana.
2.1.2. ~aterillus.
Dans les savanes guf.né ennes et soudanat ses, Ros evear'
(op. cit) ne r econna.î t cu t une vseul e e s pè c e de Taterillus! .'r. qraci
lis. Avec Tranier (1930) nous avons montré que la formule.chromo
somique des individus capturés au Foro est identi~ue à l'espèce
de référence et comporte 2N = 36 chromosomes. Ces résultats sont
en accord avec ceux rapportés précédemment par Matthey et Petter
(1970) pour des individus collectés dans le sud de la Haute-Volta
qui présentaient le même nombre de chromosomes. En Côte d'Ivoire,
le Foro est le point le plus méridional où l'on ait capturé Tate
rillus ~racilis. Il a également été rencontré à Sananférédou~ou
(fig.15). En revanche il est certainement absent de Larnto et des
savanes du Sud du V baoulé où l'on capture cependant Tatera hOj
kinsoni.
C'est t:n ron,:eur de taille ~oyenne, élancé, bon :ouis
seur et nocturne. Sa coloration générale est fauve slus so~bre sur
le èos et plus clair sur les flancs. Le ventre est blanc.
- 33 -
Les oreilles sont relativement lon~ues et les yeux
gros. La queue, nettement plus lon~ue nue le corps, présente à sa
partie distale un pinceau de poils foncés bien développé. Les
pattes postérieures sont 10nSues et étroites.
Le crâne de T. ~racilis est bombé, étroit et effilé
dans sa partie nasale et plutôt lar~e dans sa partie cervicale.
Les bulles sont bien développées (30 % de Cb). Le foramen palatin
est très développé et atteint presque la longueur de la rangée des
molaires. Les fentes incisives sont également lon?,ues et atteignent
la première molaire. Les incisives sont fines, opisthodontes et
creusées d'un profond sillon lon~itudinal. Les molaires sont
comparables à celles de T. hopkinsoni, en plus petit.
Les différentes mesures prises sur le corps, le crâne
et les dents de l'échantillon d'adultessont rapportées dans le
t abLeau IX •
2.1.2.2. Observations éthoécolo~iaues. ----------------------~-~---
T. rracilis est une espèce rare au Foro. Elle n'a été
capturée ni en for@t ni dans les cam~ements. Elle semble plus
abondante dans les cultures vi'œières telles le maïs ou le riz
pluvial. C'est l'une des proies de la chouette effraie. D'après
Hubert et Poulet (1980) T. gracilis est surtout ç,ranivore. Elle
consomme é~alement des bulbes, des rhizomes et des insectes.
2.1.2.3. Observations biolo~iaues.
et de 10 mâles. Les premières femelles ~estantes sont rencontrées
lorsoue le poids corporel atteint 25 à 30 ~. Des individus actifs
ont été capturés en avril. Deu:~ femelles restantes portaient trois
e~ auatre embryons. En élevare, nous n'avons pas obtenu de repro
duction·chez T. ~racilis. Hubert (1973) indique une durée de ~es
tation se situant entre 30 et 34 jours. Selon cet aut~ur, le nom-
- 34 -
bre moyen de jeunes est àe trois à cinq par portée et les indi
vidus sont adultes à douze semaines environ.
2.1.3. r·:rus.
Petter (1969) et Rosevear (1969) indiquent que auatre
espèces de I:Tus sont présentes en Afrique de 11 Ouest. Pus haussa 1
qui a des affinités nettement soudaniennes et saheliennes est la
seule qui n'ait pas été capturée en Côte d'Ivoire, alors que r~us
setulosus, Mus minutoïdes musculoïdes et Mus mattheyi l'ont été.
Mus setulosus se distingue facilement des deux dernières espèces
par une taille plus importante, une couleur grise, un pied plus
long et par la forme et la taille de ses molaires ; elle n'a pas
été capturée au Foro alors au'elle est présente et abondante à
Lamto et dans les friches forestières de basse Cate d'Ivoire. Nus
minutoïdes musculoïdes est l'espèce la plus abondante à Lamto et
au Foro. Elle se rencontre également plus au nord à Ouango-Fitini
(fig.16), ainsi que dans les zones défrichées et cultivées de
basse Cate d'Ivoire en particulier dans les pâturages artificiels
à Panicum maximum. Mus mattheyi n'a été captur~qulau Foro oG elle
représente moins de 10 % des captures du genre ; elle semble
absente de Larnto et de la région d'Abidjan. I:Iatthey (non publié)
a dénombré 2N = 36 chromosomes pour Mus mattheyi de la région du
Foro et 2N = 34 chromosomes pour Mus minutoïdes musculoIctes de
Bouaké.
de l'Ouest. Hus minutoïdes ~usculoïdes présente une coloration
dorsale à dominante brun-gris tiqueté de jaunâtre ; le ventre est
blanc. La séparation entre ces deux colorations est matérialisée
par une fine bordure jaunâtre. ~~s mattheyi présente une colora
tion nettement Rris-roussâtre devenant nettement roux-jaunâtre
sur les flancs. Le ventre est blanc. On ne constate pas la Drésen
ce de ligne jaunâtre séparant les deux parties du corps.
- 35 -
Chez les deux espèces, la queue, nettement plus courte
que le corps, est fine et grisâtre plus foncœdessus. Les oreilles,
brunes~sont bien développées. Pieds et mains sont blanchâtres dessus
.et dessous. Les mensurations prises sur le cor~s de l'échantillon
d' adul tes sont rapportées dans le tableau X. Le crâne des I·Ius est
caractéris&
par
ECOLOGIE DES RONGEURS DE SAVANE
A
soutenue le . devant le jury composé de :
MM. les Professeurs M. LAMOTTE
F. BOURLIERE
MM. R. BARBAULT
Je remercie Monsieur le Professeur G. CN~US,Directeur
Général de l'Office de la Recherche Scientifique et Technioue
Outre-Mer, de m'avoir permis d'effectuer les recherches, dont les
résultats sont présentés ici, au Laboratoire de Mammalogie du
Centre ORS TOM d'Abidjan-Adiopodoumé.
Je remercie [iIonsieur le Professeur :1. LAI·lOTTE Gui, apr è s
~'avoir conseillé au cours de l'élaboration du ~anuscrit, a accecté
d'être président du jury de cette thèse.
Je remercie le Docteur F. PETTER d'avoir bie~ voulu ~.
tous moments et,deDuis de nombreuses années, ~e f~ire profiter de
sa grande connaissance de la systématique et de la biolopie des
ron~eurs tropicaux. Ses conseils et son amitié m'ont été d'une
p:rande aide.
Je remercie r'Iessieurs les Professeurs F. BOURLIERE et
Ph. DREUX ainsi que Messieurs A. BROSSET et R. BARBAULT d'avoir
bien voulu faire partie de mon jury de thèse.
Je remercie Monsieur le Professeur Ch. TSIEAULT, mon
Directeur scientifique pendant toute la durée de cette étude.
Je remercie Monsieur le Professeur L. HIRTH, Président
du Comité Techniaue de Phytopatholosie et de Zoolorrie A9r.lir.uée
de l'ORSTQr:i, les rnembr-e s du Bureau Permanent et par-t i cu l Lè r emen t
Monsieur N. ROTH 90ur le soutien cu'ils m'ont manifesté.
i'Ionsieur L. BELLIER él. été à l'origine de ma formation
de ~arnmalogist~ qu'il soit assuré de mon amicale reconnaissance.
Je remercie I·Ionsieur i.J'iGELINI, Directeur de la station
IRCT de Bouaké (Côte d'Ivoire), d'avoir bien voulu me donner
l'autorisation de travailler au Foro.
Je remercie mes collègues A. BEAUDOU pédolo~ue à l'
ORS TOM et J. CESAR botaniste à l'IE~NT - CHEZ de Bouaké pour l'
aide qu'ils m'ont fournie dans l'êtude des sols et de la végéta
tion du Foro.
Monsieur P. CASES pour l'aide au'ils m'ont fournklors du traite
ment informaticue des données de pié~eap.e.
Je remercie ~:onsieur E. R.IO pour sa co ï.Labor-at i on
technique à cette étude.
du laboratoire de Mammalogie d'Abidjan-Adiopodoumé pour le tra
vail fourni durant l'étude, particulièrement I!;essieurs AITADOU
SIBIRI ZONE et DENIS FATTO SAGOU.
PLA N
1.1.2. Substratum.
1.1.3. Climat.
1.1.4. Végétation.
1.2.2. Manipulations.
3
3
CHAPITRE II - PEUPLE~ŒNT DES PETITS RONGEURS DE LA REGION DU FORO : IDENTIFICATION ET TRAITS GENER~UX DES ESPECES. •••• • 28
2.1. Identification des espèces . . . . 2.1.1. Tatera.
2.1.2. Taterillus.
2.1.3. Nus.
2.1.4. Arvicanthis.
2.1.5. Lemniscomys.
2.1.6. Mylomys.
2.1.7. Uranomys.
2.2.1. Peuplement dans la savane.
2.2.2. Peuplement dans les forêts-galeries.
2.2.3. Peuplement dans les cultures.
2.3. Comparaison du peuplement de la région du Foro avec ceux d'autres régions de Côte d'Ivoire. • • • • • 59
2.3.1. Comparaison qualitative.
2.3.2. Comparaison quantitative.
CHAPITRE III - REPRODUCTION ET CROISSANCE DA.NS LE lULIEU NATUREL ET EN CAPTIVITE. • • • 62
3.1. Etude de la reproduction. . . 62
3.1.1. Modalités de la reproduction dans le milieu naturel.
3.1.2. Observations sur la reproduction en captivité.
3.2. Etude de la croissance. . . . . . . . . . . . . . . . 78
3.2.1. Lemniscomys bellieri.
3.2.2. Myomys daltoni.
3.2.3. Tatera hopkinsoni.
3.2.4. Uranomys ruddi.
3.2.5. Influence des saisons sur la croissance corporelle de Lemniscomys bellieri.
CHAPITRE IV - ANALYSE DES RESULTATS DE PIEGEAGE SUR LE QUADRAT DE MARQUAGE. . . 90
4.1. Résidence. . . . . . . . 91
4.1.1. Taux de résidents dans la population de Lemniscomys bellieri.
4.1.2. Taux de résidents dans la population de Myomys daltoni.
4.1.3. Taux de résidents dans la p09ulation de Tatera h09kinsoni.
4.1.4. Taux de résidents dans la population de ['·1us spp ,
4.1.5. Origine des résidents et des passagers.
4.1.6. Conclusions.
4.2.1. Durée de persistance maximale de chaque espèce.
4.2.2. Durée de persistance moyenne de 50, 25 et 5 % de chaque population.
4.2.3. Durée de persistance moyenne de 50, 25 et 5 % de chaque p09ulation résidente.
4.2.4. Conclusions.
94
97
4.4.2. Spectres de déplacements.
4.4.4. Modifications de la mobilité des espèces selon les saisons.
4.4.5. Evolution mensuelle de la mobilité moyenne de Lemniscomys et Myomys.
4.4.6. Influence des feux de savane sur la mobilité des espèces.
4.4.7. Conclusions.
4.5. Occupation spatiale du quadrat de marquage parles principales espèces de rongeurs. ••••••••••• 103
4.5.1. Localisation des zones préférentielles des principales espèces.
4.5.2. Analyse de la répartition des captures de chaque espèce.
4.5.3. Correspondances entre la répartition numérique des principales espèces et certains éléments du milieu.
4.5.4. Conclusions.
CHAPITRE v - DENSITES ET BIQ!'.1ASSES DES POPULATIOi'!S DE RONGEURS. •••• • ••• 111
5.1. Densités et biomasses estimées sur le cuadrat de marquage du Foro. •••••• • ••• 112
5.1.1.
5.1.2.
5.1.3.
5.1.5.
5.1.6.
Méthodes d'estimation des densités.
Choix de méthodes utilisables dans le cas des ron~eurs du Foro.
Estimations des densités brutes.
Estimations de la biomasse pour 10 ha.
Discussion et conclusions.
5.2. Densités et biomasses de Lemniscomys bellieri estimées sur les lignes exhaustives de piégeaf,e.
5.2.1. Correspondance entre les effectifs pié~és
sur les li~nes et sur le quadrat.
5.2.2. Méthode d'estimation des densités sur lignes.
5.2.3. Estimation des densités mensuelles de Lemniscomys au Foro.
5.2.4. Com~araison des savanes du Foro et de Lar;;to.
5.2.5. Estimation de la biomasse mensuelle.
5.2.6. Conclusions.
CHAPITRE VI - ELEi/IENTS DI ANALYSE DEr.10GRAPHIQUE CHEZ LES ESPECES LES PLUS ABONDAi\fTES.
129
138
6.2.1. Composition par âges de l'ensemble des populations.
6.2.2. Influence des saisons sur la composition par âges de la population de Lemniscomvs.
6.3. Analyse démographique longitudinale.
6.3.1. Evolution des cohortes.
143
Jusque dans les années 60, par comparaison aux études
réalisées en milieu tempéré, un retard très important pouvait
être constaté dans l'acquisition des informations se rapportant
à la biologie et à l'écologie des rongeurs tropicaux africains.
Quelques études très ponctuelles avaient cependant été menées en
Afrique centrale (Ouganda, ZaIre) et en Afrique de l'ouest (Nigé
ria, Guinée-Mont Nimba). Pour le rest~ l'essentiel des études en
trepriseétaient des missions de courte durée réalisées dans le
but de collecter des spécimens à destination des taxinomistes et
des zoogéographes.
triel dans la plupart des pays d'Afrique tropicale, et corréla
tivement celui des recherches d'accompagnement au développement,
ont permis d'implanter différents groupes de recherches au Séné
gal, au Nigéria, en CSte d'Ivoire.
Le programme de recherches sur les rongeurs que nous
avons entrepris en moyenne cSte d'Ivoire à partir de 1971 est
complémentaire de celui réalisé précédemment par Bellier (1963
1972) à Lamto dans les savanes du sud du V baoulé. Il vise essen·
tiellement à fournir des informations sur les modalités de la re
production, la croissance, les densités, la biomasse et la démo
graphie des populations de rongeurs de savane dans la zone tropi
cale humide. Ce programme s'est déroulé en deux phases (1971-1974
et 1976-1979) séparées par un arrêt de deux ans (1974-1976) pen
dant lequel nous avons dO nous intéresser plus particulièrement
aux rongeurs nuisibles aux cultures industrielles (palmier à huile) dans la zone forestière de basse CSte d'Ivoire.
- 2 -
Le site du Foro a été choisi comme représentatif d'une
zone encore non étudiée. Il a été choisi également par suite de sa
proximité de la zone dans laquelle les cultures industrielles (can
ne à sucre, soja) ont été (ou doivent être) implantées, afin que
nos résultats puissent être utilisables s'il s'avère nécessaire
de lutter contre des rongeurs nuisibles. Enfin, il a été choisi
à la suite de l'implantation d'une équipe d'Entomologistes de
l'ORSTOM au même endroit, ce qui nous a permis de mieux renta
biliser l'infrastructure, le matériel et le personnel commun aux
deux laboratoires.
1.1. Le milieu.=========
La "Îorêt classées" de Foro-Foro - ou du Foro - (qui
est en réalité une ~ortion de savane mise en réserve avant 1947~
est située dans le centre de la C6te d'Ivoire (7° 55' N et 5° W),
à mi-chemin de Katiola au nord et de Bouaké au sud. A la limite
des secteurs des savanes guinéennes et sub-soudanaises, "elle est
comprise dans l'interfluve du Bandarna (élargi par suite de la mise
en place de la retenue d'eau du barrage de Kossou) et de son af
fluent le N'zi, dans le haut du bassin du N'zi.
La zone classée, de forme grossièrement trapézoïdale,
est limitée à·l'ouest par la route nationale Bouaké-Katiola, au
nord et au sud par deux marigots, affluents du N'zi, le Foro-Foro
et la [~'bé, oui coulent vers l'est et dont la confluence maraue
l'extrémité est de la réserve. Le domaine du Foro se présente com
me un plateau s'inclinant et se rétrécissant d'ouest en est, en
taillé par un réseau de marizots généralement orientés nord-sud
qui n'ont pas tous atteint leur profil d'équilibre (Bru~ière, 1948).
L'altitude est essentiellement de l'ordre de 200 m, avec àes col
lines et àes plateaux culminant vers 300 m.
2t "Forêt classée" est un terme normalisé de forestier qui ne préjuge pas de la nature de la vépétation du site auouel il s'adresse.
- 4 -
La zone des sav~~es auquelles appartient le Foro a
fait l'objet de trois missions photogra?hiaues aériennes réalisées
à l'intervalle de 10 ans soit
- mission 1952-53 au 1/50.000 (IGN-AOF-N032, 1952-53)
- mission 1961-62 au 1/50.000 (IGN-AOF-1961-62)
- mission 1973 au 1/20.000 (IGN-73 CI-V-072/200).
1.1.2. Le substratum.
1.1.2.1. La géologie.----------
La forêt classée du Foro est située dans la zone des
granitordes homogènes à biotite appartenant iu complexe Baoulé du
Protérozorque inférieur (Ta;;nini, 1972). Il.s'a.~it de granites
calco-alcalins faiblement orientés, à larges filons de quartz
blanc. Les cr t s t aux sont de taille variable et la s t ruc-cur-e 'la du
micro-grenu à la pegmatite 8:vec cristaux àe muscovite de 1 à 2 cm
de diamètre. Les éléments noirs y so~t bien représentés (Brugière,
1948) •
tzeuses, micacées. Parfois, certaines variétés de pegmatites don
nent localement des sols très argileux. La roche en place, lorsqu'
elle affleure, porte très souvent des l'polissoirs" restes d'une
occupation humaine ancienne (Avenard, 1975).
1.1.2.2. Les sols.
la suivante
- versants: sols ferrugineux r,ravillonnaires,
temporaire.
- 5 -
Pour Perraud (1971) l'ensemble de la séquence compre
nant plateaux et pentes est située dans la classe des sols ferra
litiques moyennement désaturés issus du ~ranite. La classification
des sols du Foro devient pour cet auteur
sur plateaux à couvert forestier: sol brun rouge, argileux,
très gravillonnaire : "remanié modal",
graveleux, à carapace à faible profondeur: "remanié induré",
- en haut de pentes, sol gris sableux à ocre rouge, argilo-sableux,
à carapace à moyenne profondeur: "remanié, recouvrerrent modal,
faciès induré",
- sur pentes, sol gris sableux à ocre jaune, sablo-argileux, gra
villonnaire, tacheté en profondeur : "remanié aopauvri Il,
- en bas de pente, sol gris sableux à ocre sableux, légèrement
tacheté en profondeur: "appauvri hydromorDhe",
- dans les bas-fonds, sol gris blanc, sablo-argileux à pseudo gley,
reposant sur des sables grossiers: "hydromporphe minéral".
1.1.3. Le climat.
Le Foro fait partie de la région appartenant au type
climatique C 1 défini par Eldin (1971) : "deux ou quatre saisons
dont une grande saison sèche d'environ cinq mois allant de novem
bre à mars, avec un à trois mois d'Harmattan (vent sec du nord),
1200 à 1600 mm de pluie, 1800 à 2300 heures d'insolation annuelle,
des températures moyennes mensuelles comprises entre 19 et 34° C,
la température moyenne annuelle se situant entre 25 et 28° C".
1.1.3.1. ~~!~~!E~~~_~!~~~~~~_~~_S!!~~~_~~_~~~~
Les informations que nous rapportons ici nous ont été
fournies par le laboratoire d'Entomologie de l'ORSTO~r qui a géré
le parc météorologique du Foro pendant la période 1971-1973, et
par le Ser~ice Agronomique de l'IRCT.
- 6 -
Les températures observées au cours de la période 1971
1973 (tab. I) sont voisines de celles rapportées ~ar Eldin (op. cit):
la température maximale moyenne ,mensuelle (Ti) a une valeur de
31,9 ~ 1,3 0 C, la température minimale moyem1e mensuelle (fn) a
une valeur de 17,6 ~ 1,1 0 C et la moyenne des températures ci-dessus Tx+Tn +( 2 ) a une valeur de 24,7 - 0,8 0 C.
Les moyennes mensuelles les plus faibles sont observées
en août pour Tx, en décembre pour Tn par suite de l'Harmattan et à
cause des minimums précédents en août et décembre pour leur ~oyenne.
Les moyennes mensuelles les plus fortes sont observées
en février pour Tx,en avril pour Tn et en février pour leur
moye~~e.
males (Tx) et des températures minimales (Tn) observées en 1971-73
et au cours de la période 1977-78 sont très voisines, ce qui nous
a permis après regroupement de l'ensemble des observations d'esti
mer des températures moyennes (tab. I).
1.1.3.1.2. Insolation (tao. II).
Durant la période 1971-1973 l'insolation totale annu
elle moyenne a été de 2390,4 heures, valeur comparable à celle
fournie ?ar Eldin. La moyenne mensuelle d'insolation passe par
des valeurs minimales entre juillet et septembre (115-117 heures)
et oar des valeurs ~aximales entre novembre et mars (231 à 252
heures).
1.1.3.1.3. Photopériode (tab. II).
Les jours les plus longs se situent normalement en juin avec 12,57 heures, les plus courts en décembre avec 11,67 heures.
L'écart entre ces deux valeurs, très faible, est de 0,9 heures soit 54 ~n.
- 7 -
La pluviosité annuelle totale a été en moyenne de 1110,0
mm pendant la période 1971-73 avec 74,3 jours de pluie en moyenne,
et de 1020,0 ± 111,6 mm pendant celle allant de 1970 à 1978. Ces
valeurs sont sensiblement plus faibles que les moyennes rapportées
par E1din pour les décennies précédentes. La répartition mensuelle
de la pluviosité est très hétéropène (tab. II) : au cours des six
mois les plus pluvieux il tombe 78,4 % de la hauteur d'eau annuelle
et au cours des quatre mois les moins pluvieux il n'en tombe que
6,3 %. Les pluies apparaissent avec une grande variabilité, géné- •
ra1ement en mars. Elles sont bien installées en avril, mai, juin.
Elles réapparaissent, toujours avec une grande variabilité, en aoOt,
sont fortes en septembre et également très variables en octobre.
Le mois de juillet, en moyenne bien arrosé, présente également une
grande variabilité dans sa pluviosité. Septembre puis avril, mai,
juin sont les mois les plus pluvieux. Novembre, puis janvier, fé
vrier et décembre sont les mois les plus secs.
1.1.3.1.5. Saisons.
faire.par l'emploi de différents indices.
- L'évapotranspiration potentielle (E.T.P.) est un indice complexe
dont le calcul nécessite la connaissance de paramètres qui carac
térisent au mieux le pouvoir évaporant de l'air qui s'exerce dans
le milieu donné. L'évapotranspiration potentielle totalise l'éva
poration physique et l'évaporation "physiologique" (Lecordier, 1974)
Les paramètres nécesaires à l'estimation de l'E.T.P.
par la formule de Turc (1961), citée par E1din (1971), sont la
température moyenne, la durée du jour, la durée quotidienne moyenne
d'insolation, l'énergie du rayonnement solaire. La valeur mensuelle
de l'E.T.P. rapportée à la pluviosité moyenne du mois (fig. 2)
permet de conclure à la présence d~ si~ mois pluvieux en moyenne
dont cinq le sont nettement (avril à juin, aoOt et septembre) et
- 8 -
un peut être pluvieux mais aussi souvent sec,juillet. La seconde
période également de six mois, se compose de deux mois humides qui
peuvent être parfois pluvieux (mars et octobre) et quatre mois net
tement secs (novembre à février).
- Un système d'indices permet de dresser le diagramme ombrothermique
de Moral (1964). Il ne prend en compte que la température mensuelle
moyenne qui, multipliée par des facteurs constants, permet de défi
nir des limites allant de l'aridité à la pluviosité et ainsi de
classer les mois selon leur pluviométrie en arides, secs, humides
ou pluvieux. Ces limites appliquées à l'année pluviométrique moyenne
(fig. 2) permettent de conclure, à la présence de cinq mois plu
vieux (avril à juin, août et septembre), de trois mois humides
(mars, juillet et octobre), de deux mois secs (décembre, janvier)
et de deux mois arides (novembre et février). On note également que
la limite des mois pluvieux déterminée par la méthode de Moral est
assez proche de celle calculée dans le cas de l'E.T.P. par la for
mule de Turc •
. - Il est enfin possible, grâce ~ un nouveau systè~e d'indices, de
dresser le diagramme ombrothermique de Lecordier (1974). Pour son
auteur ce système permet de mieux rattacher les variations biolo
giques des espèces animales aux variations du climat local. Le
classement des mois selon leur pluviométrie est enèore affiné par
la création de deux nouvelles classes (sub-humide et sub-sec).
L'application de ce procédé à la pluviométrie moyenne
mensuelle au Foro confirme le classement que nous avons obtenu
précédemment : quatre mois sont en moyenne nettement pluvieux (avril
mai, juin et septembre), août est en moyenne humide, mars et octobre
sont en moyenne sub-humides, janvier, février et novembre sont en
moyenne arides et décembre est en moyenne sec (fi~. 3).
1.1.3.1.6. Variations climatiques interannuelles.
Les variations climatiques qui se sont produites au cours de la période 1970-1978 ont trait à la oluviométrie annuelle
- 9 -
ainsi qu'à la durée et a l'intensité des différentes saisons.
- Variation de la pluviométrie annuelle.
Pendant la durée de l'étude, nous avons enregistré
(fir;. 4) deux années moyennes (1971 et 1976), une année oluvieuse. . (1974), une année très pluvieuse (1977) et cinq années déficitaires
(1970, 72, 73, 75, 78). Toutes les années sauf 1977 se situent en
deçà de l'intervalle moyen calculé par Eldin.
- Variation de la durée et de l'intensité àes saisons.
La durée et l'intensité des saisons sont très varia
bles d'une année à l'autre (fip.. 4). La grande saison des pluies
présente une très grande variabilité mais elle est toujours présentE
Normalement elle débute dans le courant de mars pour s'achever en
juin. Elle peut être précoce (1971, 1975) ou tardive (1977) ; les
pluies peuvent y @tre abondantes (1971, 1972, 1975) ou faibles
(1970, 1974). La petite -saison des pluies est plus régulière que
la grande. Dans presque tous les cas le mois de septembre est le
plus arrosé (7 cas sur 9). Dans les "conditions normales" elle se
déroule de courant août à courant octobre. Il existe toutefois des
années où la petite saison des pluies est très arrosée (1973 et
surtout 1977) et d'autres où elle est presque inexistante (1978).
L'intersaison centrée sur juillet est elle'aussi très variable;
lorsqu'elle présente une pluviosité importante, l'année se réduit
à deux saisons, l'une sèche et l'autre humide (1970, 1975) ; pré-
sentant une sécheresse marquée, elle permet le découpar.e
théorique de la saison humide en deux saisons des pluies.
La saison sèche est assez peu variable. Elle s'étend
normalement de courant octobre à courant mars, mais peut être limi
tée à la période oui va de novembre à février (1972-73, 1974-75). Le mois de décembre (exceptionnellement janvier) a une pluviosité
qui correspond, certaines années, à la classe sub-humide (1975).
- 10 -
milieux.
dans lesquelles nous avons analysé le peuplement de rongeurs. Les
savanes de Dabou, de Lamto et de Ferkéssédougou (pour le site de
Ouango-Fitini). Au niveau régional, nous avons comparé le climat
du Foro avec ceux de Bouaké et de Katio1a, où la pluviosité moyen
ne mensuelle observée est très voisine de celle du Foro (fig. 5).
Elle diffère nettement de celle constatée à Lamto, où la pluviosité
en novembre, jênvier, février et mars, très abondante, rappelle le
climat de basse Cate d'Ivoire rencontré- à Dabou.
Enfin la pluviosité du Foro se différencie nettement
de celle de Ferkéssédougou où n'existe plus qu'une seule saison des
pluies dont le maximum d'abondance se situe en août-septembre et où
la saison sèche est de plus longue durée.
La pluviométrie annuelle est voisine pour le Foro,
Katio1a et Boualcé , Celle de Larrto est un peu supérieure à celle du
Foro (+ 15 %). Pour des périodes différentes, celle de Ferkéssédou
~ou est éga1e~ent plus élevée (+ 35 %) et celle de Dabou presque
~ga1e au double (+ 94 %). Les autres variables c1imatioues caracté-- .
risant les stations retenues sont rapportées dans le tab1eauIII.
1.1.4. Végétation.
La lIforêt c1assée ll du ::'oro appartient encore aux savanes
guinéennes préforestières à Brachy~ria brachy10pha et Panicum phra~
mitoïdes. L'observation olus att~ntive de la carte de la vé~étation. - . el' Ad j ànohoùn de Cate d'Ivoire de Gui11aumet/(1971) montre qu'en fait le domaine
du Foro se situe à-la limite de- la zone de savanes sub-soudaniennes
dont la limite s'en situe à moins de 50 km au nord et à l'est.
La densité de la strate 1i~neuse décroît des plateaux
aux bas de pentes. Trente cinq espèces d'arbres et arbustes sont
- 11 -
rencont~ées couramment au Foro, en dehors des forêts de sommet et
des forêts-qaleries. Les plus abonda~tes sont Loohira lanceolata,
Afrorrnosia laxiflora, Butyroscerrnum paradoxum, ?a~inari curatel
lifolia, Pterocarpus erinaceus, Trichilia roka, Piliosti~ma thon
nins=:ii.
Si l'on compare les strates ligneuses de Lanto, du Foro
et de Ouango-Fit1ni, on constate que 18 seulement des 35 espèces
d'arbustes et arbres du Foro se retrouvent à Lamto, alors que 32
(plus trois incertaines) sont présentes à Ouango-Fitini (tab. IV).
(Menaut et Dev1neau comm. perso.).
1.1.4.2. Strate herbeuse.
Les plantes herbacées les plus courantes de la savane
du Foro appar-tLennent à six fa..,7iilles vé zé t a l.e s :
cées, ?apilionacées, Z~~!.iberacées, 2ubiacées et
Graminées, Cypera- 1"": • ~ ..Jloscoreacees. La
~lus représentée et la ~lus abonèante est celle èes Gra0inées, qui
t ~ ,.J Lnodi \00' , •• .,. ,corner-en Ancr-ooo ccn ascJ.no a s , ....... scru r-ens i s , hypar~,.en~é'. Si!1~-
thiana., n. subolumosa, HY'Perthelia dissoluta, ?s.nicum ph!"at:::~itorè.es,
Schizachyriurn san~uineum, EIJ~andra anèroor.ila et ~ott~oelia e7.al
tata. (César, comm. per-so s ) ,
(César, 1971) Comparée à celle de Lamto~ la strate herbeuse du Foro
possède un nombre important d'espèces qui lui sont propres. Parmi
celles-ci, notons cino des neuf Graminées, les deux Cypéracées,
deux des sept Papilionacées et l'espèce de Dioscoreacée (tab. V).
L'étude de la croissance globale en taille et poids de
la strate herbeuse au cours de l'année 1974 a été réalisée à partir ?
de trois prélèvements mensuels de 4 rn- de vésétation faitsdans une
parcelle de savane située sur le plateau. En plus de la croissance,
une analyse fourra~ère type a été réalisée, qlobalement, sur chaque
échantillon.
La croissance en taille de la strate herbeuse au~rnente
- 12 -
à peu près régulièrement, dans la mesure où les pluies se manifes
tent, après le passage des feux jusqu'en juillet où les Graminées
atteignent leur taille vé~étative maximale (environ 80 cm).
A partir de septembre, on observe une brusque aug~enta
tion de la taille consécutive à l'apparition des flèches portant
les épis. En octobre cette brusque croissance est achevée et les
épis murissent (fig. 6). La char~e en poids de matière sèche de la
parcelle augmente elle aussi régulièrement de janvier à novembre.
La croissance en poids au cours de la période janvier-avril est
très irrégulière par sùite de sa très forte corrélation avec les
pluies. La charge totale est passée de 150 kg/ha en janvier a
5.400 kg/ha en décembre. La teneur en eau (différence entre le
poids frais et le poids sec de l'échantillon d'herbes) est varia
ble selon les saisons. Elle est normalement très élevée au cours
des ~ériodes pluvieuses (65 à 70 %) et réduite au cours des pério
des sèches (50 à 55 ~). Elle est ~inimale (35%) juste avant les
feux (fig. 6).
Les principaux éléments minéraux ont des teneurs varia
bles selon les saisons (fig. 6). A l'exception du Sodium dont la
teneur est constante toute l'année (0,01 %), l'Azote (maximum 1 %),
le Potassium (maximum 1,3 %) et le Phosphore (maximum 0,14 %) pré
sentent les teneurs les plus élevées dans les jeunes pousses du
début de la phase végétative, en janvier, puis elles décroissent
plus ou moins régulièrement pour @tre minimales dans les chaumes
sèches de décembre. Les teneurs en Calcium (maximum 0,55 %) et en
Magnésium (maximum 0,30 %) augmentent jusqu'en mai puis diminuent
lentement et régulièrement jusqu'en décembre.
1.1.5. Les feux de brousse.
En savane, les feux de brousse sont un facteur du mi
lieu très important : en quelques heures ils peuvent éliminer inté
gralement la strate herbacée qui était à son maximum de croissance,
le feuillage des arbres et des arbustes et une vartie des ~raines
tombées sur le sol.
Des feux précoces (novembre-décembre) mannuerrt ,&,énéra
1ement d'intensité et la savane ne se trouve donc qu'imparfaitement
bru1ée. Il subsiste alors de nombreuses poches de végétation non
bru1ées dont la 'surface unitaire peut atteindre plusieurs hect~res.
Les feux tardifs, ou feux ba1ayants, se pratiouent en janvier et
surtout en février. Ils brûlent la vég,étation herbacée dans son in
tégralité par suite de son très srand état de sécheresse; ce sont
surtout les feux de mise en culture.
Dans la forêt classée du Foro, les ~ardes forestiers et
le responsable du village ont pour consi~ne de mettre en place des
feux précoces, mais à notre demande, en 1978-79 les feux ont été
tardifs (début février) dans le secteur du quadrat de marquage.
Pendant la durée de l'étude du peuplement de ronpeurs.
les feux ont eu lieu en : décembre 1971, décembre 1972, décembre
1973, janvier 1976, décembre 1977, février 1978.
1.1.6. Un milieu type, le cuadrat de marouape de 10 ha.
L'étude en continu du peuplement de ronpeurs s'est
faite dans une parcelle carrée de 10 ha, le quadrat de marGuage,
dont l'emplacement a été jugé représentatif de l'ensemble de la
forêt classée du Foro - à l'e;~ception des forêts de sommet et des
forêts-~a1eries çui ne font pas partie intégrante de la zone de
piégeage mais qui la jouxtent au nord-ouest et au nord-est.
Le quadrat de marquage (QH) est situé à peu près au
centre du domaine protégé. Il est distant de 11 km du laboratoire
(fig. 7). Le dixième de sa surface correspond à l'ouest à un pla-"
teau dont l'altitude est de 280 m. La pente est douce (2 %) vers
l'est et vers le sud-est; elle est forte en suivant la dingonale
- 14 -
est et la bordure sud-est du O~ (4 ~) (fig. 8).
L'étude des sols a été faite à partir de la description
de seize fosses de 140 cm de profondeur réparties rér,ulièrement
sur le QM et d'une fosse de même dimension située dans le bas fond
à proximité de la forêt-galerie. Sur une surface aussi réduite (10
ha), les sols varient peu d'une fosse àl'autre. On peut toutefois
observer une certaine modification le long de la toposéquence qui
va du plateau situé au sud-ouest du QM au bas fond proche de la. . galerie située au nord-est. La caractérisation des sols le long de
cette toposéquence ~eut être faite à partir de.l'analyse de quatre
fosses disposées régulièrement du plateau au bas-fond dont les
caractéristiques physiochimiques sont rapportées dans le tableau VI.
Sur le plateau, où l'horizon meuble atteint 85 cm, le
profil (M) se découpe comme suit :
- de 0 à 14 cm, une couche humifère brun-noir, argilo-sableuse de
venant sablo-argileuse, s'enrichissant pro~ressivement en petits
nodules ferrusineux et en éléments quartzeux, riche en racines, de
structure continue, de cOhésion faible à moyenne, où la porosité
tubulaire fine et moyenne est très inportante,
- de 14 à 85 cm, une couche de sol brun-roupe à rou?-e sablo-ar~i
leux à arf,ilo-sableux s'enrichissent fortement en petits nodules
ferrugineux jusqu'à 53 cm, profondeur à partir de laquelle aopa
raissent des matériaux friables provenant de l'altération des ro
ches. On note la présence de quelques racines dans toute l'épRis
seur de la couche. La structure est arpileuse fine, la cohésion
moyenne, la porosité tubulaire moyenne est bien développée,
- de 85 à 140 cm, une couche d'altération des roches, friable et
riche en éléments de la couche précédente devenant pro~ressivement
dure. L'altérite y est riche en éléments de quartz de 0,2 Q 0,5 mm
de diamètre.
A mi-pente, la séparation entre l'horizon des sols meu
bles et celui des sols durs est très irré~ulière. Elle se situe
entre 59 et 103 cm. Le profil (J) se découpe co~me suit:
- de 0 à 35 cm, la couche humique appauvrie en matière or~anique de
couleur gris-brun, sableuse faiblement ar~ileuse, avec de nombreu
ses racines, s'enrichit progressivement en petits nodules ferrugni
neux et en quartz. La structure, d'abord particulaire, devient
argileuse, mal individualisée. La cohésion est très faible à faible.
La porosité importante est d'abord interstitielle puis devient tu
bulaire fine
- de 35 à 81 cm, la couche des sols brun-rouge à rouge sablo-argi
leux très riches en nodules ferrugineux de 0,5 à 2 à 3 cm, où l'on
rencontre également des éléments quartzeux de même dimension. La
structure y est anguleuse fine, mal exprimée. On y trouve un peu
d'humus et quel~ues racines. La cohésion est faible ~ moyenr.e et
la porosité interstitielle et tubuleire fine très i~~ortante ;
- de 81 à 140 cm, la couche des matériaux d'altération des roches
est dure, riche en quartz (0,2 à 0,4 ~m) et en sabl$~rossiers. 0n
y rencontre S ~ lS % de sols brun-rouge de la couche ~r~cédente.
En bas de pente, la séparation entre l'horizon des sols
meubles et la zone des sols d'altération des roches est également
irrégulière et ondulée. Le profil(D) se découpe comme suit:
- de 0 à 33 cm, la couche humique gris-brun foncé à brun foncé,
sableuse comprend une très forte proportion de racines. Elle s'en
richit progressivement en nodules ferru~ineux et quartzeux de ~etite
à moyer~e dimension (0,5 à 2 à 3 cm). La structure est particulaire
à faiblement anguleuse. La cohésion est très faible à nulle. La po
rosité interstitielle fine est très élevée ;
- de 33 à 52 cm, la couche des sols brun-rou~e sableux fai=lernent
argileux est riche en humus et en racines et com~orte une forte
;roportion de nodules ferrurrineu;: et auelques éléments cuartzeux.
La structure est particulaire et partielle~ent an~ulaire r.al indi
vidualisée. La cohésion est tr8s faible et la porosité intersti
tielle, fine ~ ~oyenne, tr~~ forte ;
- 16 -
- de 62 à 140 cm, la zone d'altération des roches se compose de
blocs de carapace et de cuirasse, de fra~~ents quartzeux et de ma
tériel de remplissage de la couche précédente tacheté et compact.
On y note la présence de placages noirs d'oxydes (fer et rnanganè se ) ,
Dans le bas fond à p~oxirnité de la r:ale~ie forestière
du marigot, le profil pédologique (0) se découpe co~rne suit:
- de 0 à 9 cm, une couche humifère grise très riche en sables et
en racines, à structure particulairs parfois en a~régats arrondis.
La cohésion est presque nulle, la porosité interstitielle et tubu
haire fine est très élevée ;
- de 9 à 111 cm, une couche sableuse Sris-clair à beise-clair conte
nant d~ns sa portion supérieure cuelques nodules ferru~ineux de
petite taille (0,2 à 0,6 cm) des racines et de l'humus ~uis, au
delà de 30 cm des blocs de cuirasse friable (juscu'à 15 cm de èia
~ètre) des éléments quartzaux açFlo~érés ~ar des oxy~es de fer e~
de ranGanèse, ,enfin au-delà de 64 cm des blocs de c,uartz de toutes
tailles (quelques centimètres à 30 à ~o cm) mélangés aux blocs èe
cuirasse friable. La structure du matériau sableux est particulaire,
sa cohésion est très faible à nulle, sa porosité interstitielle et
tubulaire fine est très importante.
- de 111 à 140 cm, la zone d'altération des sols jaune-brun à brun
très pâle est très friable. Le sol est compact et argileux. On note
la présence de fines paillettes de muscovite.
1.1.6.3. Vé~étation.--.....-------
1.1.ô.3.1. La strate li;.neuse (fip. 9).
Sur la surface de 10 ha, on a dénombré 9287 pieds, touf
fes et fourrés de li~neux soit une densité de 929 à l'ha. Les treize
espèces les plus abondantes sont Lophira lanceolata (2~,4 %), Afror mo s i a 'axi-"10ra (, .../ ~ C~) ~l'+-V1"'os-e1"'mu'" oar-adoxum (le:; .., ,":) T;)~rin""1"'i•• 1 .c. _ • ~ _ _,"'" i"J , _'.J.." .. _ ,'..,;' _ .J l'I v _ J-..ul:: _ ,~ 1'" , ;. _ ... c:.._ ...
cu .... a ....e".;-"oli'" (12 3 ~') '"I"iS ï''''n';0ll~'''' oli're1"'; (7 ,4 ~') m_ i C " i 1 '; "... 1"" __ ...L.":" ..r...c;::. ,;J ;:",\",oi"'" :.Je-.... ...L.__ ~.:....' ..... _ _ ,~ ,) , J..:., ... ... .:.__ ..... c-..;.
k ( ' 8 ") p.... . (~ - "1) ':l • , . " ....'" • "ro a L~, ~J,. l.eroc~.r!Jus er~né1.ceus ... ,0 /J , ~ ~_~osï;~~mê. "'1,onn~n1~~
( - 2 "/) .,) , '.'.', Cochlos-:;e!'r::u:7i pl3.nchoni (3,2 ;j), Annon9. sene';<:alensis (1,4;:~) •
- 17 -
Ue.naca tocroensis (1,4 %) à la li s i ère des "forêts", Terrninalia SOD
(1,1 %) et Hvrnenocardia acida (1 %). Les 17 autres espèces repré
sentent à elles toutes 3,7 %.
L'essentiel des arbrisseaux (520/ha) est constitué par
les mêmes essences que celles listées ci-dessus, excepté Uaoaca
toxoensis. A l'exception de Cochlosoermum planchoni mais avsc en
plus Bridelia ferrurr.inea, les mêmes espèces forment la ~uasi tota
lité de la strate arbustive (204 pieds/ha). Enfin, exce9té C. Dlan
choni, P. thonnin~ii, T. roka, E. ferru~inea, A. sene~alensis et
H. acida, les mêmes espèces auquelles il convient d'ajouter les
ficus représentent 97,4 % de la strate arborée (47 pieès à l'ha).
Les touffes (151 à l'ha) sont constitué$surtout par A. laxiflora
(35,8 %), L. l~"1ceolata (30,8 %) et C. plancr.oni (14,7 ;~) puis par
P. thonni:1;ü (6,7 %), i3. '::ar2.ào~('Jrr. (5,7;)), P. curatelli folia ( , 7<:'~' -, :.~ l ,
T. roka (1,4 %) et D. oliveri (1 %). Les fourrés (S,3 i l'~a) sont r 1'" '"c '" 0 1 ..,t- '" (, a ."-' =<) ,..; e~ • _ .. ;. _ _ c . .... ;;,.... __" ,,,,J ". _
1 . • • (- ., .') L -', . (O'CSC:1Y~ 0,..,;:., a zone ce p i a t aau est
la plus fournie en li;neux (1215 à l'ha) où A. laxiflora est ~ette-
t- 1 • • ( ? - .., '-') ., d -t:':en w c.Or7i.lnan'C _0 , ~ ;.: suaV:l. eL. 1 anc eo ï a t a (·7 ,.., -')_c:;;o,.l.4 _ "- ...~'- J.. ,0 ;'..; , 3. :-.:arado:-:um
(16,1 %) et P. curatellifolia (15,4 %). Les pentes bien drainées
(800 pieds à l'ha) sont surtout peuplées de L. lanceolata_,(45,4 %)
et de A. laxiflora (12,5 %). Le bas-fond (665 pieds à l'ha) est
surtout peuplé de P. curatellifolia (42,3 %) et de L. lanceolata
(14, 5 ~;).
minées (fi~. 10). Globalement, les espèces les olus aconè2.ntes sont
Androco~on ascinodis (17,6 %) et Schizachyriu~ san~uineurn (16,9 ~~),
puis Elyme.ndra anèron!"'ilê, (14,1 ~0), H:l~arrhenic?. sr.:ithi.:'tna (12,1 ;~),
". h ..... -ct ('1 C -') . d . . . ( 1) 7 c : _ ...: an.lCUI7l n ra.,<r::1. "'o~ es Jo , Q :.~ , r.n rODor:on scnl!'ensJ.s :, ,.' J '= '"
'-'v'"'ar-l-,e,..li~ subc Lurno s a (f"' 7 C')"'., '.., .1. "' ...._~. ~_ ::: ç-. =' , /0). Enfin on trouve Hyperthelia ~isso-
Lu t a (~ 1 cr ,.....eru.urn -- (? ,'1 "') et- ":lo......... o e La a :"'~1"'''''''- (0 ~ ,")~ .,- ',)1, """l",,\;;;t ...1 • ..:::1 .... _,"'" /"; •.,. ... I..tv'V _<;.0. _ ..... c::._;..,c.;.""'.,;i.. ,':J ,'J,.
Sur le plateau les principales espèces sont S. san~uineu~ (18,2 ;j),
2. androDhila -') et" ~,-,~",.,.-; i-oIr'os (, ... , C~)1...' ... ~ ..·",1"'C;:.I'.11;.l,J ....... _ . _, ,_ ; ....
- 18 -
nodis (25,4 %), H. subplumosa (18,7 %) et s. san~uineurn (16,2 %).
Le peuplement herbacé des pentes bien drainées est intermédiaire
entre ce~x des plateaux et des bas-fonds ; les princi~ales esp8ces
y sont A. ascinodis (18,4 %), H. subolumos2 (16)1 X), s. san~uineum
(15,7 %) et E. androohila (14,3 %).
1.2. Matériel et techniques. ======================J
nous ne rapportons ici que le matériel et les techniques
servant à recueillir les informations de base relatives aux Ronge~rs,
c'est à dire le 9ié~ear.e, l'éleva~e, les prises de mesures sur les
spécimens, etc •••
èe hase (méthodes de calcul des densités, analyse dérr:oSraphic:ue,
etc ••• ) seront traitées dans leurs chapitres respectifs.
1.2.1. Les oièqes et les oiépeages.
Trois modèles de pières ont été utilisés au cours de
notre étude. Deux modèles permettent la capt~re des rongeurs vi
vants (Chauvancy et piège-couloir), le troisième les tue (tapette).
1.2.1.1.1. Pièf.e Chauvancy (licence CNRS).
C'est le modèle de p1ege le plus utilis8 puisqu'il est
seul sur le quadrat de marquage (1024 pièges) et la majeure partie
des lignes exhaustives (1050 piè~es mensuellement de 1971 à 1974
et environ 400 pièges entre 1976 et 1979). Le pièpe Chauvancy se
compose d'un "couvercle" comprenant le mécanisme de fermeture 8,utO-
matique et de verrouillase de la porte lorsc:ue l'ani~al touche
l'appât ~lacé au bout d'une ti~e métallique servant de fléau et
ct' une "boîte à nid" r'e pr-é s errt é e à l' or-t r-t ne par une boite de conserve
- 19 -
d'un kilogra~me sur laquelle s'emboîte le couvercle. Pour réduire
le volume considérable représenté par plus de 1000 boites de conser
ve nécessaires au piégeage mensuel (volume très supérieur à la
contenance d'une camionnette bâchée) nous avons fait réaliser des
boîtes tronconiques en aluminium. Elles sont plus longues que les
précédentes (donc permettant la capture d'animaux de plus [rande
taille et réduisant la mortalité des captures par suite du confine
ment), mais en s'emboîtant les unes dans les autres, elles occupent
un volume extrê~ement réduit ~Rr rapport au précédent modèle (de
l'ordre de 1 m3).
Une amélioration dans le rendement des piè~es Chauv~~cy
a été obtenu par l'utilisation non plus de boîtes tronconiques en
aluminium, mais de boîtes tronconiques en grillage à petite maille,
permettant une reilleure diffusion de l'odeur de l'appât, la cap
ture plus aisée des i~dividus et des espèces présentant un co~~or
tement claustrophobe, et enfin une diminution de la nortalité dûe
au coup de chaleur dans les boîtes fermées.
1.2.1.1.2. Piège-couloir (ratière Manufrance).
C'est un parallèlépipède rectangle de 42 cm de long, de
14 cm de section carrée. Avec ses parois en fil d'acier galvanisé,
ce piè~e présente les avantages énumérés dans le cas du Chauvancy
à boîte tronconique en ~rilla~e. Afin de permettre la capture des
~us et des jeunes individus des espèces de taille moyenne (Lemnis
comys, Myomys, etc ••. ), les parois sont doublées, avec du grillaŒe
métallique du type moustiquaire. Le mécanisme de fer~eture-verrouil
la~e est constitué de deux portes placées aux deux extrémités du
parallèlépipède, bloquées en position ouverte par une tise porte
appât placée au centre du couloir et par de puissants ressorts as
surant le verrouillage en position fermée. Ce modèle de piè~e par
ses dimensions permet la capture d'individus et d'espèces de srande
taille (Tatera, Dasymys, petits carnivores, hérissons etc ••• ). De
plus, la présence des àeux ouvertures semble accroître le rendement
du pièf:e par rapport au Chauvancy et ~err.et la capture d'espèces
r-éou t é e s Lncao t ur-ab l e s avec le Chauvancy (Arvicanthi s, ~I.vloï.'Ys).
- 20 -
tellement courante qu'il n'est pas nécessaire de les décrire ici.
Notons toutefois que nous avons utilisés des tapettes à socle en
bois et à mécanisme galvanisé qui leur assure une bonne tenue en
climat tropical humide.
L'inconvénient principal de ce type de pièp.e est de
fournir des animaux morts et presque toujours décom90sés, inter
disant les prélèvements d'or~anes (yeux, organes génitaux ••• ) et
rendant aléatoires les mesures prises sur le CorDS. Le crâne étant
souvent fracassé, il ne peut plus être utilisé au cours de l'étude
taxinomicue.
Les avanta~es èe ce modèle de ~ièrre sont doubles. ~n-" n 1 - "-
tuant l'animal avant qu'il -ait consommé l'appât, on peut à partir
des captures, réaliser des analys~s de contenus stomacaux (à con
dition d'inspecter les piègeages à de très courts intervalles de
temps). De plus, on constate, à l'usage, que ce modèle de piège est
le moins sélectif, le plus commode d'emploi et celui avec lequel
les rongeurs présentent le temps de latence à la capture le plus
court.
Selon le but recherché nous avons utilisé différentes
méthodes de piégeage. Au cours de la phase d'inventaire des espè
ces nous avons utilisé le piéseage en tache, oG une quinzaine de
pièges (nombre très excédentaire par rapport au nombre de rongeurs
potentiellement espéré) sont disposés sur une surface modeste (en
viron 100 m2 ). Dans ce type de piégeage, tous les modèles de piè
~es sont utilisés afin d'augmenter les chances de collecte du plus
frand nombre d'espèces présentes dans le milieu retenu. C'est cette
méthode de piégeage aui a été employ~dans les cultures, les forêts
zaleries et de plateau, dans les villages, etc ••.
- 21 -
En savane, les piépeares exhaustifs systématiques sont
effectués sur des li~nes unitaires de 50 piè~es Chnuvancy. Entre
1971 et 1974, deu~= longueurs de lignes furent utilisées : d'une part
des 1i~nes de 245 m de long (50 p~er.es espacés de 5 m), d'autre part
des lignes de 98 m de long (50 piêges espacés de 2 m). Les pi~~es
restent tendus pendant quatre jours consécutifs, fourniss8.nt auatre
résultats quotidiens, soit un effort de piéQeage de 200 journées
pièJe par ligne unitaire conduisant à un rendement théoriouemaxi
mu~ de 200 captures. En plus du piégeage exhaustif standard, nous
avons disposé durant cette même oériode trois liBnes unitaires de
98 ~ composées de tapettes •. Au cours de la période 1976-1979, les li~nes de 245 m
de long ont seules été utilisées. Le temps de pié~ea~e a été ramené
à trois jours, soit un effort de piégeage de 150 journées-piè~e par
ligne.
Le quadrat de marquage a été mis en place dans le but
d'obtenir àes informations sur l'étho-écologie des !on~eurs et la
dynamique de leurs populations dans le même milieu~ Le site a
été choisi de façon à inté~rer sur la surface du piéfeape l'ensem
ble des grands faciès botaniques et topo-pédo1o~iaues de la station.
Nous nous sommes fixé une surfaceœenviron 10 ha (exactement 9,61
ha) qui soit compatible avec nos possibilités de piéç,ea~e (en hom
mes et en ~ntériel) Ovi puisse répondre aux conditions de milieu
i~~osêes et oui fournisse des échantillons auotidiens suffiscmment
import~nts. L'espacement entre deux pièges est de 10 m ; cette va
leur a été déterminée au cours d'un ~iépeage avec maraua~e réalisée
en 1974 sur une surface de 5 ha comptenant des piê2es disposés se
lon un espacement de 7 m. L'ensemblè du quadrat comprend 32 1i~nes
de 32 piè~es, soit un total de 1024 pièses. Les pièges restent ten
dus pendant ouatre jours consécutifs ; ils sont visités chaque ma
tin. L'effort total mensuel de piégeage a donc été constamment de
4096journées-piè~e.
1.2.1.3. L'anoât utilisé.--------------- Pendant toute l'étude et quel que soit le type de piè
Seage, l'appât utilisé a été la noix de palmier à huile. C'est un
fruit de petite taille parfaitement adapté au piégeage. Il dure
quatre, voire cinq jours dans les pié~es sans se détériorer. Il est
bien consommé par toutes les espèces de rongeurs et disponible du
rant tous les mois de l'année~
1.2.1.4. Effort de vié~ea~e.------------------ Par suite de la granàe diversité des techniques de pié
geage utilisées par chaque auteur, l'effort de pié~eage déployé au
cours d'une étude ne peut pas être caractérisé par le nombre de
pièges ou par le nombre de piégeaf:es unitaires déposés. Suiv~~t
l'école ~7.éricaine, l'intensité de ~iégeege que nous avons dé~loyée
au Foro est esti~ée par le nombre d~ pièses présents sur le terrai~
par fraction de 24 neures . L' uni té ct' intensi té de pié~ea~e est donc
la journée-piège ("night-trapll des Américains).
Dans le cas des lifnes exhaustives utilisées pour rea
liser le piégeage systématique mensuel, l'effort total a été de
162.400 journées-pièg~(812 lignes de 50 pièges posées penda~t qua
tre jours consécutifs), dur~nt la période 1971-74 et de 44.400 jour
nées-pièges (296 lignes de 50 pièges 90SéS pend~~t trois jours) au
cours de la ~ériode 1976-1979.
Le premier auadrat de marqua~e expérimental mis en place
en 1974 avait nécessité 24.000 journées-pièges et le second quadrat
de marquage, qui a fonctionné de 1976 à 1979, a nécessité 147.456
journées-pièges. L'effort total déployé au cours de cette étude a
donc été de 378.756 journées-piè~es.
Le rendement des piégeages est estimé ~ar le nombre de
captures pour 100 journées-9iè~es.Le rendement $710b2ol mo ye n des
piégeages exhaustifs a été De 2,16 captures pour 100 journées-piè~es.
- 23 -
Dans le cas des lirnes de 98 m composées de tapettes
espacées de 2 m, le rendement moyen a été de 2,8 captures/100 jour
nées-piè~es. Dans celui des li~nes de 98 m composées de pièges Chau
vancy espacés de 2 m, le rendement moyen a été de 1,88 captures pour
100 journées-pièges. Dans celui des lignes de 245 m composées de
pièses Chauvancy espacés de 5 m, le renèement a été de 2,58 captures
pour 100 journées-pièges, soit un rendement moyen par fractions de
98 m de 1,03 captures pour 100 Journées-pièges. Enfin dans le cas
des lif-nes de 245 m composées de pièges Chauvency espacés de 5 m
posés pendant trois jours, le rendement moyen a été de 2,20 captures
pour 100 journées-pièges soit 0,88 captures pour 100 journées-pièges
par fractions de 98 m•.
1.2.1.6. ~~!~~~!~!~~_~~~_~!f!~~~~~~~_~~~~~!g~~~_~~_E!~g~~E~
~!~_~_~!~_~~~_~~E~~~~_~~_~~~g~~~~·
ments, on peut définir l'action sélective des différentes techniques
de piégea~e utilisées sur les espèces piér-ées. De fait les résultats
de piéReage montrent bien que l'importance relative de chaque es
pèce dans le peuplement peut subir des variations plus ou moins im
portantes selo~ la lip,:ne de piér:.eage considérée (fiF. 11). Des dif
férences, peu importantes sont notées entre les snectres se rappor
tant aux deux modèles èe lignes ChauvancY. Elles ont trait essen
tiellement aux Benres Lemniscomys et f1yomys. Des différences bien
plus marquées s'observent au cours de l'utilisation des lignes de
t.ape t tie s , Dans ce cas, on constate une réduction irnportante du pour
centap,e de Lemniscomys leui passe de 54,5 % dans les lignes de pièges
Chauvancj' à 43~1 % dans les li~nes de .tapettes, conco~mittante cvec
un accroissement importent de celui des i~astomys qui passe de ~oins
de 1 % dans les pièges Chauvancy à 8 % dans les tapettes. Uranomys
et Granhiurus présentent une aupmentation moins marquée èans les
li~nes composées de tapettes avec respectivement 2,4 ~~ contre 1,6 % et 3,8 % contre 1,2 %.
- 24 -
Les individus capturés dans le quadrat de marcuaie sont
anesthésiés dans un bocal d'un volume de deux litres contenant un
tampon de coton imbibé de quelques gouttes d'éther éthylique. Dans
ces conditions, l'anesthésie est très rapide et dans la mesure où
l'on suit très attentivement les réactions de l'animal pendant la
phase d'aspiration de l'éther, le taux de mortalité est très infé
rieur à 1 %. Les genres ,les plus sensibles aux vapeurs d'éther et
donc chez lesouelles il y a risque de décès par surdose, 'sont Uyomys
et ~us, puis Lemniscomys, Granhiurus, Uranomys et enfin Tatera.
1.2.2.2. Observations sur les animaux caoturés dans le ouadrat -------------------------------~--------------~------
Après avoir été enesthésiés,tous les individus capturés,
marqués ou non, sont pesés et mesurés(lon~ueur tête et cor~s du
nez à l'anus, longueur de la queue et du pied avec griffes). L'état
mor-pho Log â que apparent de la sphère p:éni tale exter-ne est noté (déve
loppement et pbs1tion des testicules, coloration des bourses chez
le mâle, développement des tétines et lactation, festation visible,
ouverture vag.inale chez la femelle). Puis les individus nouveaux
sont marqués par amputation de doigts. Le code de marcua~e est un
code à base 5 pour les pieds et un code à base 4 pour les mains.
Cet artifice permet de ne Das couper plus d'un doist par membre
tout en lai~sant la ~ossibilité de numéroter un nombre suffisant
d'individus, compte-tenu des densités rencontrées. A~rRs le réveil
les individus sont relechés au point de capture. Ils sont suivis à
l'oeil afin de contrôler Qu'ils n'entrent pas à nouveau immédiate
ment dans un piè~e èu voisinase. A cette occasion, leurs terriers
ou leurs nids sont éventuellement repérés.
- 25 -
Après avoir été tués par l'inalhation d'un excès de
vapeurs d'éther éthylique ou d'éther de pétrole, les individus sont
placés en décubitus dorsal comme lors du marquarre. La longueur tête
etcorps et celle de la queue sont notées, ainsi que la lon~ueur de
la patte postérieure. Les observations morpholo~iques concernant la
sphère génitale sont notées puis l'animal est autopsié. Chez le
mâle, les testicules, les vésicules séminales, les épididymes sont
prélevés et fixés dans le liquide de Bouin. Chez la femelle oh re
cherche les traces de ~estation, on note alors le nombre d'embryon~
dans chaque corne utérine. Les cicatrices placentaires, témoIns de
gestations précédentes, sont également recherchées et dénombrées.
Puis les ovaires et les cornes utérines sont prélevées et ég,alement
placéS dans du liquide de Bouin. Pour les deux sexes les yeux sont
prélevés et fixés da~s du formol à 10 %. La ?eau est prélevée puis
traitée en vue de sa conservation en collection. La tête est é~ale
ment prélevée, débara~sée du cerveau, séchée àU. soleil ou à l'étuve
à 60 0 C puis placée dans un élevage de Dermestes qui consomment la
chair désséchée sans abimer les os du crâne. Ce dernier est blanchi
dans une solution à 30 % ct' eau oxyrréné e , séché puis mesuré et placé
en collection.
Les yeux sont fi;::és pendant 30 jours dans une solution
de formol à 10 %. Les cristallins sont alors extraits et clacés
pendant cuanze jours dans une nouvelle solution de formol 2' 10 ~:'.
Ils sont ensuite· désséchés dans une étuve ventilée à 105 0 C pendant
des durées variables selon leur taille. Pour Le~niscomys et Uranomys
la durée de dessication est de trois heures, quatre heures pour
I.1yomys et seize heures pour Tatere.• Après s'être . refroidis dans
un dessicateur contenant du silicapel, les cristallins secs sont
pesés sur une balance à nacelle précise au 5/1000 de m~.
- 26 -
L'éleva~e des ron[eurs du Foro a été réalisé dans la
salle d'élevare du laboratoire situé sur le Centre ORSTOE d'Adiopo
doumé, à proximité d'Abidj~n. Cette salle est une pièce de 50 ~2
dans l~quelle sont disposés des portoirs à cages. La température
observée est en moyenne celle de l'extérieur, mais tamponnée.
Les ca[es sont des bacs en plastique recouverts de cou
vercles en fil d'acier zinfué. Les cages individuelles sont des
ca~es à souris, qui mesurent 30 X 20 X 18 cm. Les capes pour couples
sont des cafes à rats, qui mesurent 50 X 25 X 18 cm.
Dans chaque ca~e on place de la litière composée de
sci~re de bois. Une botte de conserve d'un kilo, à laouelle est
adjointe un piè~e Chauvancy à la place du couvercle sert de botte
à nid. Le fond de cette botte est percé d'une fenêtre triançulaire
permettant aux animaux d'entrer et sortir à volonté de la boîte à
nid. Dans la botte ~ nid on dépose une p~7née de frison. La ferme
·ture de la fenêtre trianpulaire lorsque les individus sont dans le
nid permet leur manipulation ou bien le nettoyaSe de la cage olus
aisément.
granulés pour souris et de ~ranulés pour lapins en reproduction ;
chaque jour on ajoute des morceaux de papaye mûre (apport d'eau),
deux fois par semaine des noi;( de palmier à huile et une fois ~ar
se~aine une dose de mïl sermé.
Chaque jour, les naissances sont recherchées et notées.
Les jeunes sont pesés à la naissance, au sevrape (15 jours), ~uis
mensuellement à ~artir de l'â~e d'un mois.
70us les ~enres, sauf nus, ont pû être maintenus en
éleva~e sans problèmes particuliers. Nous n'avons pas obtenu de
reproduction chez Arvicanthis, Steatomys et Mylomys.
- 27 -
Une part importante de la taxinomie des ron~eurs est
fondée sur des mesures du crâne et des dents. Nous avons donc choi
si vingt mesures qui décrivent le crâne et les dents (tac.VII)
(fig. 12). Toutes les mesures sont prises à l'aide d'un ~ied à
coulisse à cadran;précis au 5/100 de millimètre.
Afin d'avoir une idée plus précise de la variation des
mesures en fonction de l'âge des individus, nous avons estimé, sur
les crânes des Lemniscomys et des Ï'Tyomys ayant servi à l'étude de
la croissance des cristallins (voir chaplII), la valeur moyenne des
mesures retenues pour des âges allant de 1 à 8 mois (excepté 7
mois). Comme Verheyen et Br-actee (1966), nous avons constaté (lue la
croissance du crâne des rongeurs est très corrélée avec l'âpe des
individus (r généralement supérieur à O,SO). Seules font excention
les r&~~ées molaires dont la taille est fixée dès l'éru0tion de la
troisième molaire. La lar2eur de la première ffiolaire supérieure
(lM1) est corrélée avec l'âse par suite de sa forme tronconique
ainsi lors de l'usure des lames d'émail la largeur de la dent au~
mente au fur et à mesure que l'on va vers la base du tronc de cône.
Enfin la largeur de l'intermaxi11aire prise derrière [:[3 - 1,j3, con
trairement à toutes les autres mesures diminue avec l'âge, le coef
ficient de corrélation est alors négatif (fig.13a).
Le taux de variation entre un et huit mois (fi~.13b),
est très différent selon la nature des mesures. Celles 'se rappor
tant aux molaires sont très stables (Rmx, Rmd, IM1). Les mesures
présentant les plus forts taux de variation se rapportent à la par
tie antérieure du crâne et particulièrement au rostre (D, l?m, P,
In, hU, LN, Fi et partiellement Cb). On note eue 10 varie ~eu et
oue Ez varie nlus que 1Bz qui lui est ~ostérieur. Enfin l'étude du
taux de variation des mesures chez l'individu p1einesent adulte
entre 4 et 8 mois (fiŒ.13c) confirme les résultats précédents. Dans
l'ensemble les mesures sont stabilisées et leur variation n'excède
pas 5 %, exceptés certaines mesures crises dans la partie antéri
eure du crâne (D, P, Fi, 12 et Ur).
- 28 -
IDENTIFICATION ET TP~ITS GENEPAUX DES ESPECES
Au cours de la période 1971-1979, on a capturé 5057
spécimens de Rongeurs de la région du Foro. Ces individus provien
nent essentiellement du milieu naturel (savane, forê~8aleries,
for@ts sèches) et pour une plus faible part des milieux cultivés.
L'ensemble des captures se répartit en quinze genres. Ce sont~pour
les Gerbillinés : Tatera. et Taterillus ; pour les ~;lurinés : I:Ius,
Arvicanthis, Lemniscomys, Mylomys, Uranornys, Thamnomys (Gremmornvs),
Praomys, nyomys, Mastomys, DasymYs et Hylomyscus ; pour les Dendro
murinés : Steatomys ; pour les Graphiurinés Graphiurus.
2.1.1. Tatera.
groupes panafricains de Gerbillidae en Afrique de l'ouest; des
individus du groupe des T. robusta représenté uniquement ~ar
T. guineae et àes individus du groupe des T. valida représenté par
trois espèces T. weï.mannt , T. hO:Jkinsoni et T. kernpL,
En 1959 ?etter a montré Gue Tetera valida n'était pas
dans l'ouest de l'Afriaue. La plus ancienne forme nommée dans cette
réfion étant T. kempi, on èoit donc parler du Sroupe kemni.
- 29 -
des deux ~roupes reposent sur des caractères de ~orpholopie cor
porelle et crânienne et sur les formules chromosomiaues. En ce
qui concerne les caractères de morphologie corporelle , on cons
tate que T. puineae est de taillelépèrement plus faible, la colo
ration dorsale est plus roussâtre devenant franchement fauve sur
les flancs. Sa queue est proportionnellement nettement plus lon
gue que le corps et elle présente un manchon de poils 'bruns sur
la moitié terminale nettement plus touffue dans sa partie terminale.
Les pieds paraissent plus allongés pa~ce que plus étroits que chez
les Tatera de l'autre groupe. Au niveau du crâne, T. F-uineae pré
sente un rostre plus étroit et plus fin par suite du développement
moins important des prémaxillaires et de la plus faible larrreur
des incisives qui les font ressembler à celles de Taterillus. En
résumé, T. çuineae est morphologiquement plus proche de Taterillus
r-racilis alors que les Tatera du ~roupe kemoi sont plus trapus et
d'aspect plus murin.
Enfin, les formules chromosomiques sont nettement dif
férentes, 2atthey (1954) et Matthey et Petter (1970) ayant dé nom-
bré :
pour Tatera hookinsoni 2N = 48 ",
pour Tatera p:uineae 2N = 50 " pour Tatera ;'!a,mbiana 2H "- 56 "
Dans une étude cytopénétioue et taxinomioue plus ré
cente, Matthey et Petter (1970), ont confirmé pour le sud de la
Haute-Volta (Bobo-Dioulasso) la présence des deux groupes rappor
tés ci-dessus, celui des T. kempi étant représenté par T. honkin
~. T. hookinsoni semble plus abondant (12 spécimens) que T. ~ui
~ (5 spécimens) parmi l'ensemble èes spécimens collectés. Pré
cédemment, Heim de Balsac et Lamotte (1958)1 pour la répion quinéen
ne du ;'1ont Ni.mba , puis Heim de Balsac (1967) pour Lamto, avaient
- 30 ~
rapporté l'ensemble àes individus de ces deux localités à Tatera
kernDi.,
Pour la C6te d'Ivoire (fig.14) nous n'avons pas eu la
confirmation cytosénétique de la présence de T. kempi. En revanche,
avec Tranier (1980) nous avons noté la présence, aussi bien au Foro
qu'à Lamto, de T. hopkinsoni (du groupe T. kempi) dont la formule
chromosomique a ét~ déterminée comme étant égale à 2N = 48 chromo
so~es. A Bouna, dans le nord de la C6te d'Ivoire; avec Bellier
(1967), nous avions obtenu, pour la majorité, des individus réfé
rables à T. hopkinsoni'et, en plus petit nombre, des individus ré
férables à T. ~uineae, ce qui est en accord avec les observations
faites par Matthey et Petter (op. cit) et ce qui tend à confirmer
les affinités nettement soudanaises de cette dernière espèce.
2.1.1.1. Descriotion.-----------
d'un Rattus~ Sa coloration àorsale brun-fauve dans la partie mé
diane s'éclaircit sur les flancs. Le ventre est blanc àe la bou
che à l'anus. Sa queue, d'une longueur au moins égale à celle du
co~ps, est brun-marron dessus et blanc-grisâtre dessous. Les oreil
les sont relativement 8randes et les yeux globuleux. Les pieds et
les mains sont blanch§tres à la partie supérieure. La sole des
pieds est brun fonc6, l'intérieur des mains est clair.
Les mensur~tions corporelles prises sur l'échê-ntillon
d'adultes sont rap~ortées d~nG le tableau VIII. La comparaison des
moyennes entre sexes fait apparaître des différences significa
tives pour le poids corporel et la taill~ tête et corps, les mâles
étant plus rrands et plus gros que les fe~elles.
Le crâne de T. honkinsoni est caractérisé par sa for~e
allonrée surtout dans sa partie antérieure, et par sa boite crani
enne large. Les bulles tympaniques sont très développées (30,4 ~~
de Cb).
Le foramen incisif est IonS ct atteint le bord anté
rieur de Ml. Le foramen oalatin s'étend de la moitié de Ml à la
moitié de M2• Les incisives supérieures relativement étroites sont
nettement opisthodontes et gravées dans le quart externe d'un pro
fond sillon longitudinal. Les rangées molaires sont composées de
dents trapues construites sur un plan de base àe Cricétidé. Les
principales mesures prises sur le crâne et les dents sont rappor
tées dans le tahleauVDI.La cornr-araison des moyennes entre sexes,
comme pour les mensurations corporelles, fait acparaître des diffé
rences sir,nificatives pour l'ensemble des mesures se rapportant à
des Lonzueur-s , Celles qui concernent des lar.rreurs ne sont pas diffé
rentes except~Io et lPm. Celles se rapportant à des hauteurs sont
significativement différentes excepté ~1. Enfin les longueurs des
rangées molaires (Rmx et Rmd) ne sont pas différentes entre sexes.
Au Foro, Tatera hookinsoni se rencontre dans tous les
types de savane, que ce soit sur les plateaux, les pentes pas trop
fortement drainées, les bas de pentes. Il n'a été piép.é eue très
occasionnellement au bord des forêts ~aleries et un peu plus fré
que~ent dans les forêts sèches de plateau. On le retrouve dans les
cultures vivrières (riz, maIs). Il se capture facilement avec les
piè~es, particulièrement avec les tapettes. Il est exclusivement
nocturne et c'est l'une des proies principales de la chouette ef
fraie. Comme tous les serbillidés, Tatera hopkinsoni est un !ouis
seur puissant ; il est ~apable de creuser ses galeries aussi bien
dans les sols latéritiques à fort pourcentape de ~raviers (concré
tions latéritiques et fragments de quartz plus ou moins ~ros), que
dans des sols plus argilo-sableux des pentes et des bas de pentes.
Son terrier se compose de plusieurs ~aleries communiquant avec la
surface par une ou plusieurs ouvertures dont l'une au moins est
particulièrement bien repérable par suite de l'accumulation des
rejets de terrassement. Les ~aleries peuvent atteindre un mètre de
profondeur. Son régime alimentaire semble se composer de rhizomes,
de bulbes et en saison de praines de graminées. On a é~alement
- 32 -
2.1.1.3. Observations biologioues. ------------------~-----
L'ensemble èe la population pié~ée se compose àe 236
mâles et de '274 femelles, soit un taux de masculinité de 0,86 non
significativement différent de l'unité. Dans le milieu naturel
les plus jeunes femelles sont actives lorsque leur poids atteint
75 g. En captivité, la seule femelle née en élevage qui ait été
.gestante l'a été à l'âge d'un an, après une durée de vie com~une
èe onze mois avec le mâle. La durée de gestation estimée par l'in
tervalle minimum entre deu;c mises-bas consécutives semble êt~e
voisine de 31 jours, ce qui est sensiblement plus 10nS que l'ob
servation de 25 jours faite par Hubert (1973) chez Tatera ~~~biana.
2.1.2. ~aterillus.
Dans les savanes guf.né ennes et soudanat ses, Ros evear'
(op. cit) ne r econna.î t cu t une vseul e e s pè c e de Taterillus! .'r. qraci
lis. Avec Tranier (1930) nous avons montré que la formule.chromo
somique des individus capturés au Foro est identi~ue à l'espèce
de référence et comporte 2N = 36 chromosomes. Ces résultats sont
en accord avec ceux rapportés précédemment par Matthey et Petter
(1970) pour des individus collectés dans le sud de la Haute-Volta
qui présentaient le même nombre de chromosomes. En Côte d'Ivoire,
le Foro est le point le plus méridional où l'on ait capturé Tate
rillus ~racilis. Il a également été rencontré à Sananférédou~ou
(fig.15). En revanche il est certainement absent de Larnto et des
savanes du Sud du V baoulé où l'on capture cependant Tatera hOj
kinsoni.
C'est t:n ron,:eur de taille ~oyenne, élancé, bon :ouis
seur et nocturne. Sa coloration générale est fauve slus so~bre sur
le èos et plus clair sur les flancs. Le ventre est blanc.
- 33 -
Les oreilles sont relativement lon~ues et les yeux
gros. La queue, nettement plus lon~ue nue le corps, présente à sa
partie distale un pinceau de poils foncés bien développé. Les
pattes postérieures sont 10nSues et étroites.
Le crâne de T. ~racilis est bombé, étroit et effilé
dans sa partie nasale et plutôt lar~e dans sa partie cervicale.
Les bulles sont bien développées (30 % de Cb). Le foramen palatin
est très développé et atteint presque la longueur de la rangée des
molaires. Les fentes incisives sont également lon?,ues et atteignent
la première molaire. Les incisives sont fines, opisthodontes et
creusées d'un profond sillon lon~itudinal. Les molaires sont
comparables à celles de T. hopkinsoni, en plus petit.
Les différentes mesures prises sur le corps, le crâne
et les dents de l'échantillon d'adultessont rapportées dans le
t abLeau IX •
2.1.2.2. Observations éthoécolo~iaues. ----------------------~-~---
T. rracilis est une espèce rare au Foro. Elle n'a été
capturée ni en for@t ni dans les cam~ements. Elle semble plus
abondante dans les cultures vi'œières telles le maïs ou le riz
pluvial. C'est l'une des proies de la chouette effraie. D'après
Hubert et Poulet (1980) T. gracilis est surtout ç,ranivore. Elle
consomme é~alement des bulbes, des rhizomes et des insectes.
2.1.2.3. Observations biolo~iaues.
et de 10 mâles. Les premières femelles ~estantes sont rencontrées
lorsoue le poids corporel atteint 25 à 30 ~. Des individus actifs
ont été capturés en avril. Deu:~ femelles restantes portaient trois
e~ auatre embryons. En élevare, nous n'avons pas obtenu de repro
duction·chez T. ~racilis. Hubert (1973) indique une durée de ~es
tation se situant entre 30 et 34 jours. Selon cet aut~ur, le nom-
- 34 -
bre moyen de jeunes est àe trois à cinq par portée et les indi
vidus sont adultes à douze semaines environ.
2.1.3. r·:rus.
Petter (1969) et Rosevear (1969) indiquent que auatre
espèces de I:Tus sont présentes en Afrique de 11 Ouest. Pus haussa 1
qui a des affinités nettement soudaniennes et saheliennes est la
seule qui n'ait pas été capturée en Côte d'Ivoire, alors que r~us
setulosus, Mus minutoïdes musculoïdes et Mus mattheyi l'ont été.
Mus setulosus se distingue facilement des deux dernières espèces
par une taille plus importante, une couleur grise, un pied plus
long et par la forme et la taille de ses molaires ; elle n'a pas
été capturée au Foro alors au'elle est présente et abondante à
Lamto et dans les friches forestières de basse Cate d'Ivoire. Nus
minutoïdes musculoïdes est l'espèce la plus abondante à Lamto et
au Foro. Elle se rencontre également plus au nord à Ouango-Fitini
(fig.16), ainsi que dans les zones défrichées et cultivées de
basse Cate d'Ivoire en particulier dans les pâturages artificiels
à Panicum maximum. Mus mattheyi n'a été captur~qulau Foro oG elle
représente moins de 10 % des captures du genre ; elle semble
absente de Larnto et de la région d'Abidjan. I:Iatthey (non publié)
a dénombré 2N = 36 chromosomes pour Mus mattheyi de la région du
Foro et 2N = 34 chromosomes pour Mus minutoïdes musculoIctes de
Bouaké.
de l'Ouest. Hus minutoïdes ~usculoïdes présente une coloration
dorsale à dominante brun-gris tiqueté de jaunâtre ; le ventre est
blanc. La séparation entre ces deux colorations est matérialisée
par une fine bordure jaunâtre. ~~s mattheyi présente une colora
tion nettement Rris-roussâtre devenant nettement roux-jaunâtre
sur les flancs. Le ventre est blanc. On ne constate pas la Drésen
ce de ligne jaunâtre séparant les deux parties du corps.
- 35 -
Chez les deux espèces, la queue, nettement plus courte
que le corps, est fine et grisâtre plus foncœdessus. Les oreilles,
brunes~sont bien développées. Pieds et mains sont blanchâtres dessus
.et dessous. Les mensurations prises sur le cor~s de l'échantillon
d' adul tes sont rapportées dans le tableau X. Le crâne des I·Ius est
caractéris&