
A Le cacaoyer : stratégies de sélection -...
14
5 Novembre - Décembre 1995 Plantations, recherche, développement L e cacaoyer, Theobroma cacao L., est un arbre originaire des forêts humides d’Amérique tropicale. Sa culture se développe à partir du XIX e siècle, après l’invention de la presse à extraire le beurre des fèves, qui permet la fabrication du chocolat. Après l’introduction de la plante en Afrique, au début du XX e siècle, la production mondiale de cacao connaît un essor spectaculaire : de 115 000 t elle atteint aujourd’hui 2,5 millions de t de cacao marchand, soit une valeur d’environ 3 milliards de dollars. L’Afrique (Côte d’Ivoire, Ghana) en fournit 60 %, l’Asie (Malaisie, Indonésie) 20 % et l’Amérique (Brésil) 20 %. Certains pays comme l’Equa- teur, Trinidad, la Colombie, le Venezuela, le Mexique, l’Indonésie et la Papouasie met- tent sur le marché un cacao plus aroma- tique appelé cacao fin (environ 5 % de la production mondiale) (carte). La culture La plupart des zones cacaoyères se sont développées sur défriche de forêt. Une par- tie des grands arbres de forêt est tradition- nellement conservée pour assurer un ombrage. Cependant, un ombrage contrôlé peut être installé en plantant des légumi- neuses ligneuses après éradication totale de la forêt. La culture en plein soleil, plus récente, permet d’obtenir de meilleurs ren- dements mais impose des soins phytosani- taires plus réguliers. Durant les premières années, le cacaoyer est souvent associé à des cultures vivrières. Le cacaoyer est particulièrement sen- sible aux modifications de son environne- ment écologique. Le renouvellement in situ d’une plantation est souvent délicat, le cacao apparaissant généralement comme une culture de front pionnier (Jarrige et Ruf, 1990). Les structures de production sont variables : • en Afrique, ce sont des exploitations familiales de moins de 5 ha ; • en Amérique, les exploitations familiales atteignent environ 20 ha, mais il existe aussi de grandes plantations en Equateur, au Brésil, au Costa Rica ou à Trinidad ; • en Asie (Malaisie, Indonésie, Papouasie- Nouvelle-Guinée), où la cacaoculture est assez récente, des plantations privées ou publiques et de petites exploitations coexistent. Les problèmes de la cacaoculture Le cacaoyer est un arbre particulièrement sensible aux maladies et aux ravageurs, qui sont souvent les facteurs limitants de la production. Les principales maladies sont d’origine cryptogamique : • la pourriture brune des cabosses (Phy- tophthora spp.), est répandue dans le monde. L’espèce la plus nuisible, P. megakarya, provoque plus de 50 % de pertes en Afrique centrale ; A ugmenter la résistance aux maladies et aux insectes : pourriture brune, balai de sorcière, pod borer,... et améliorer la qualité sont les enjeux actuels pour cette importante culture traditionnelle. Le cacaoyer : stratégies de sélection Paulin D., Eskes A.B. CIRAD-CP, BP 5035, Montpellier Cedex 1, France Retour au menu
Transcript of A Le cacaoyer : stratégies de sélection -...

5
Novembre - Décembre 1995 Plantations, recherche, développement
L e cacaoyer, Theobroma cacao L., estun arbre originaire des forêtshumides d’Amérique tropicale. Sa
culture se développe à partir du XIXe siècle,après l’invention de la presse à extraire lebeurre des fèves, qui permet la fabricationdu chocolat. Après l’introduction de laplante en Afrique, au début du XXe siècle,la production mondiale de cacao connaît unessor spectaculaire : de 115 000 t elleatteint aujourd’hui 2,5 millions de t decacao marchand, soit une valeur d’environ3 milliards de dollars. L’Afrique (Côted’Ivoire, Ghana) en fournit 60 %, l’Asie(Malaisie, Indonésie) 20 % et l’Amérique(Brésil) 20 %. Certains pays comme l’Equa-teur, Trinidad, la Colombie, le Venezuela, leMexique, l’Indonésie et la Papouasie met-tent sur le marché un cacao plus aroma-tique appelé cacao fin (environ 5 % de laproduction mondiale) (carte).
La cultureLa plupart des zones cacaoyères se sontdéveloppées sur défriche de forêt. Une par-tie des grands arbres de forêt est tradition-nellement conservée pour assurer unombrage. Cependant, un ombrage contrôlépeut être installé en plantant des légumi-neuses ligneuses après éradication totalede la forêt. La culture en plein soleil, plusrécente, permet d’obtenir de meilleurs ren-dements mais impose des soins phytosani-taires plus réguliers. Durant les premières
années, le cacaoyer est souvent associé àdes cultures vivrières.
Le cacaoyer est particulièrement sen-sible aux modifications de son environne-ment écologique. Le renouvellement in situd’une plantation est souvent délicat, lecacao apparaissant généralement commeune culture de front pionnier (Jarrige etRuf, 1990). Les structures de productionsont variables : • en Afrique, ce sont des exploitations
familiales de moins de 5 ha ;• en Amérique, les exploitations familiales
atteignent environ 20 ha, mais il existeaussi de grandes plantations en Equateur,au Brésil, au Costa Rica ou à Trinidad ;
• en Asie (Malaisie, Indonésie, Papouasie-Nouvelle-Guinée), où la cacaoculture estassez récente, des plantations privées oupubliques et de petites exploitationscoexistent.
Les problèmes de la cacaocultureLe cacaoyer est un arbre particulièrementsensible aux maladies et aux ravageurs, quisont souvent les facteurs limitants de laproduction. Les principales maladies sontd’origine cryptogamique :• la pourriture brune des cabosses (Phy-
tophthora spp.), est répandue dans lemonde. L’espèce la plus nuisible,P. megakarya, provoque plus de 50 % depertes en Afrique centrale ;
Augmenter larésistance aux
maladies et auxinsectes : pourriturebrune, balai desorcière, pod borer,...et améliorer la qualitésont les enjeux actuelspour cette importanteculture traditionnelle.
Le cacaoyer :stratégies de
sélection
Paulin D., Eskes A.B.
CIRAD-CP, BP 5035, Montpellier Cedex 1, France
Retour au menu

Plantations, recherche, développement Novembre - Décembre 1995
6 CA C A O Y E R
• la maladie du balai de sorcière (Crini-pellis perniciosa), répandue dans lesCaraïbes et en Amérique du Sud, pro-voque la prolifération anarchique desrameaux et des coussinets floraux etdétruit les graines dans les jeunesfruits ;
• le «mal de machette» (Ceratocystis fim-briata), se rencontre également danscertains pays du continent américain.C’est une trachéomycose qui provoquela mort de l’arbre ;
• le vascular streak dieback, ou VSD,(Oncobasidium theobromae), répanduen Asie du Sud-Est, provoque la chutedes feuilles et entraîne la mort desrameaux.Le swollen-shoot, seule maladie à virus
répertoriée, se caractérise par un gonfle-ment des tiges et peut entraîner la mort del’arbre. Elle est présente au Nigeria, auGhana et au Togo.
Les insectes parasites les plus courantssont des piqueurs, en particulier lesmirides (Sahlbergella sp., Distantiella sp.et Helopeltis sp.) qui attaquent les jeunes
Nigeria 135
Côte d'Ivoire 830
Ghana 255
CamerounCameroon98
BrésilBrazil272
ColombieColombia51
EquateurEcuador78
MexiqueMexico31
PérouPeru11
VenezuelaVenezuela15
IndonésieIndonesia240
MalaisieMalaysia203
RépubliquedominicaineDominicanRepublic60
PapouasieNouvelle- GuinéePapua New Guinea37
19951995
Répartion de la production mondiale de cacao (t/an). / Distribution of world cocoa production (t/year).
Le cacaoyer est une espèce diploïde appartenant à la famille des Sterculia-cées. L’arbre, issu de semis, est formé d’un axe orthotrope qui donne nais-sance, à partir de 18 mois, à plusieurs couronnes successives, composées decinq branches plagiotropes. La multiplication végétative peut être réaliséepar bouturage ou par greffage de bois orthotrope et plagiotrope. La multi-plication in vitro du cacaoyer n’est pas encore bien maîtrisée, bien que desprogrès aient été obtenus avec l’embryogenèse somatique induite à partirde pièces florales (Lopez Baez, 1993). Les fleurs, de petite taille, très abon-dantes sur le tronc et les branches, sont hermaphrodites. La durée de vie dupollen est limitée à 48 heures. Cinq à 10 % des fleurs sont pollinisées par pol-linisation entomophile. Il existe un système d’incompatibilité de typegaméto-sporophytique. L’autocompatibilité est variable selon les popula-tions. Le taux d’allogamie n’est pas stable, il dépend de l’environnement,mais on l’estime généralement à 75 %. L’intensité de la pollinisation et lanutrition ont une influence sur la tenue et le remplissage en graines desfruits (Falque,1994 ; Lachenaud,1991). La production de fruits débute deux àtrois ans après la plantation. Les périodes de récoltes, généralement étaléesdans l’année, dépendent des conditions climatiques. La taille des fèves, lepourcentage de coque, leur teneur en beurre, et la dureté du beurre font lavaleur technologique du cacao marchand. La qualité organoleptique ducacao dépend de l’origine génétique et des traitements post-récolte (fer-mentation, séchage et torréfaction).
■ La plante
Retour au menu

Novembre - Décembre 1995 Plantations, recherche, développement
7CA C A O Y E R
pousses et les cabosses. En Afrique et enAsie, ils provoquent des dégâts considé-rables. En Asie du Sud-Est, un insecteforeur des cabosses (Conopomorpha cra-merella) entraîne des pertes de fruits trèsimportantes (Wood et Lass,1989).
Les rendements moyens enregistrésdans la plupart des pays (500 kg /ha) sonttrès inférieurs à ceux obtenus en stationexpérimentale, surtout à cause de la fortehétérogénéité des plantations et de l’utili-sation de matériel végétal d’origine incon-trôlée. La durée de vie économique d’uneplantation varie de 20 à 40 ans. Les vieillesplantations ne sont plus entretenues et sontprogressivement abandonnées au profit deplantations nouvelles. La recherche demodalités de rajeunissement ou de rempla-cement des anciens vergers par de nou-velles variétés est donc un enjeu important.
Les ressourcesgénétiques et la structuration despopulationsUne vingtaine d’espèces du genre Theo-broma existent à l’état naturel sur le conti-nent américain, mais seule T. cacao est cul-tivée pour l’industrie chocolatière. Lebassin de Haute-Amazonie est considérécomme le centre d’origine et de diversitéprimaire de l’espèce T. cacao, et l’Amériquecentrale, comme un centre de domestica-tion. Il existe trois grands groupes géné-tiques : les Forastero, les Criollo et les Tri-nitario. La distinction entre ces troisgroupes est basée sur leur origine géogra-phique et sur les caractéristiques morpho-logiques des cabosses et des fèves.
Les Forastero, produisant aujourd’huiplus de 80 % du cacao mondial, proviennentdu bassin amazonien. On en distingue deuxorigines, Haute-Amazonie et Basse-Amazo-nie, avec un continuum de variabilité entreles deux. Les Bas-Amazoniens (BA) sonttraditionnellement cultivés au Brésil et enAfrique de l ’Ouest. En raison de leurvigueur, de leur précocité et de leur relativerésistance à certains agents pathogènes, lesHauts-Amazoniens (HA), collectés plusrécemment et très diversifiés, sont souventutilisés dans les programmes de sélection.
Les Criollo, constitués de variétés primi-tives domestiquées par les Indiens d’Amé-
rique, donnent un cacao de bonne qualitéorganoleptique (cacao fin). Ils sont peu cul-tivés actuellement car, en général, peuvigoureux et sensibles aux insectes et auxmaladies fongiques.
Les Trinitario proviennent de l’hybrida-tion entre les deux groupes précédents. Laplupart des génotypes répertoriés sontd’origine cultivée. Ils présentent des carac-téristiques agronomiques et technologiquestrès variables. Du fait de leur vigueur élevéeet de la bonne qualité des fèves, ils ont étédistribués dans de nombreux pays.
Une grande diversitégénétique Plus de 7 300 génotypes originaux, dontplus de 2 800 d’origine sauvage, sont réper-toriés dans le monde. Les collections lesplus importantes se trouvent à Trinidad, auBrésil, au Costa Rica, en Equateur, auGhana, en Côte d’Ivoire et en Malaisie. Plu-sieurs prospections réalisées depuis 1940ont permis de collecter des populations ori-ginales de cacaoyers Forastero : au Pérou,en Equateur, au Brésil, en Colombie et plusrécemment en Guyane Française entre1987 et 1995. La caractérisation et l’évalua-tion de ce matériel sont encore très incom-
plètes. Peu de clones sauvages sont actuel-lement utilisés dans les programmes d’amé-lioration génétique.
Des études sur la diversité génétique ontété réalisées en utilisant les marqueursmoléculaires (RFLP(1) et RAPD(2)). Lastructuration génétique, mise en évidencepar ces marqueurs, se superpose globale-ment à la structuration géographique del’espèce (Laurent et al., 1994). La grandediversité des HA est confirmée (figure 1)(N’goran et al.,1994 ; Lecerteau et al.,1994). La diversité des populations localesest moins bien connue, mais chacun desgroupes présente une certaine variabilité,avec parfois des populations locales origi-nales comme les Porcelana du Venezuela oules cacaoyers spontanés prospectés enGuyane.
Les outils de la biologie moléculaire ontégalement permis de cartographier legénome du cacaoyer avec 193 locus identi-fiés (5 marqueurs isoenzymes, 160 mar-queurs RFLP et 28 marqueurs RAPD). Cettecarte constitue un outil pour la localisationde gènes d’intérêt agronomique. Des tra-vaux sont en cours pour rechercher desQTL(3) de la résistance à la pourriturebrune et de la taille des fèves (Lanaud etal.,1995).
(1) RFLP : DNA (Desoxyribonucleic acid) RestrictionFragment Length Polymorphism.(2) RAPD : Random Amplified Polymorphic DNADetection.(3) QTL : Quantitative trait loci.
Axe 2(10,6%) / Axis 2 (10.6%)
Axe 1(13,7%) / Axis 1 (13.7%)
Forastero Hauts-AmazoniensUpper Amazon Forastero
Forastero Bas-AmazoniensLower Amazon Forastero
Criollo
Trinitario
Figure 1. Diversité génétique du cacaoyer. Analyse factorielle des correspondances (AFC) réaliséeavec des marqueurs RAPD. / Genetic diversity of cocoa. Factorial correspondence analysis (FCA) usingRAPD markers.
Retour au menu

Les critères de sélectionAujourd’hui, la résistance aux fléaux parasi-taires est un objectif majeur de la sélectiondu cacaoyer. En effet, la lutte chimiquen’est pas toujours efficace et représente uncoût élevé pour le planteur. Des sources derésistance ont été identifiées pour la pour-riture brune, le balai de sorcière et le VSD.Un progrès significatif du niveau de résis-tance des variétés est attendu, grâce à l’uti-lisation de nouvelles méthodes de sélectionplus efficaces.
La résistance aux maladiesLa variabilité de la résistance intrinsèque àla pourriture brune a été mise en évidencepar des inoculations sur cabosses, feuillesou jeunes tiges (Blaha et Lotodé,1976 ;Lawrence,1978). Cependant, l’observationdes taux de pourriture en champ donne par-fois des classements différents de ceuxobtenus par ces tests d’inoculation artifi-cielle. Ces écarts pourraient être liés auxdifférences de durée de maturation descabosses et de période de fructification. Untest d’inoculation artificielle sur feuilles estaujourd’hui à l’étude pour évaluer la résis-tance intrinsèque (Nyasse et al.,1994).
Des observations au champ ont permisd’estimer le niveau de sensibilité au balaide sorcière de certaines populations decacaoyers. Des tests d’inoculation artifi-
cielle ont été utilisés en pépinière (Bartley,1981). Un test d’inoculation sur feuilles estactuellement expérimenté à Trinidad, quisemble donner des résultats positifs(Ducamp, communication personnelle).
Pour le VSD, des sources de résistance ontété identifiées par des observations au champaprès inoculation naturelle, actuellementaucun test d’inoculation artificielle n’est aupoint (Keane et Prior,1991). Mais il existedéjà des variétés commerciales résistantes.
La résistance aux insectes En Côte d’Ivoire, la sensibilité aux miridesest prise en compte dans le programme desélection de l’Idefor(4). Une note synthé-tique est attribuée, tenant compte del’intensité et de l’étendue des dégâts ainsique de l’aptitude à la régénération deszones attaquées. Cependant, ce type denotation ne permet pas de différencierattractivité, résistance et tolérance. Destests d’attractivité ont été réalisés au labo-ratoire. Ces travaux ont mis en évidence desdifférences de sensibilité et d’attractivitéentre HA, Amelonado et Trinitario.
En Asie, aucune bonne source de résis-tance au borer des cabosses n’a pu êtremise en évidence.
Le rendementL’entrée en production varie entre deux etcinq ans. Le rendement en cacao marchand(poids de fèves fermentées et séchées parhectare) se calcule en station par le cumulde cinq à six années de récolte en phase depleine production. En général, les cabossesmûres, saines, pourries et rongées sontdénombrées et les cabosses saines sontpesées. On définit également un «indexcabosse», nombre de cabosses nécessairespour obtenir un kilo de cacao marchand.
La vigueurLa vigueur se mesure au jeune âge, parl’accroissement du diamètre au collet, et àl’âge adulte par la circonférence du tronc.Pour tenir compte de l’influence du déve-loppement végétatif de l’arbre sur sa pro-duction, on calcule parfois le rapport pro-duction vigueur. Cette variable estimportante pour la sélection d’individus,dans les dispositifs expérimentaux en ran-domisation arbre par arbre.
La qualitéLes programmes de sélection d’hybridesprennent en compte la taille des fèves et letaux de beurre. L’évaluation de la qualitéaromatique est plus difficile car elle dépendbeaucoup des traitements post-récolte. Des
dégustations de liqueurs évaluent lessaveurs fondamentales (arôme cacao, amer-tume, astringence, acidité) et les goûts par-ticuliers. Une étude récente montre que legénotype du cacaoyer a une influence mar-quée sur les caractéristiques organolep-tiques et que le potentiel aromatique esthéritable. Des différences dans les arômescacao et le goût fruité ont été montréesentre des Forastero HA et entre certainsTrinitario (Clapperton et al.,1994). Contrai-rement aux caractères physiques des fèves,il n’existe pas de données fiables permet-tant d’intégrer ces caractères organolep-tiques dans un index de sélection, car lespréférences des consommateurs sontvariables selon les pays (Lockwood etEskes,1996).
Les paramètresgénétiquesL’estimation des différents paramètres géné-tiques, relatifs aux caractères à améliorer, aété possible, grâce à la réalisation de plans decroisements factoriels et diallèles. Pour laplupart des caractères étudiés, les effetsgénétiques additifs prédominent. Des effetsde dominance, généralement faibles, existentpour la vigueur, la taille des fèves et la pro-duction. La production peut être corréléepositivement avec la vigueur au jeune âge(Paulin et al., 1994). Les fortes variancesenvironnementales (20 à 78 %) sont observéespour les principaux critères de sélection dansles dispositifs expérimentaux en parcellesmono-arbre (Eskes et al., 1995).
L’héritabilité au sens strict, estimée àpartir des données individuelles des arbres,est moyenne pour la résistance à la pourri-ture brune (0,35), la production (0,30),faible pour la vigueur des arbres adultes(0,15) et très faible pour la résistance auxinsectes (Cilas et Paulin, 1994). L’héritabi-lité de la taille des fèves est forte (0,65),celles du taux de beurre et de la saveur frui-tée, moyennes (Lockwood et Eskes,1996).
Il existe un certain effet de l’interactiongénotype-milieu pour la production, lavigueur et le taux de pourriture, mais lesclassements entre hybrides dans différenteslocalités de Côte d’Ivoire sont généralementstables. A un autre niveau, les meilleursgéniteurs HA, sélectionnés à Trinidad, ontaussi présenté généralement un bon com-portement en croisement dans de nom-breux pays, ce qui suggère une certaine sta-bilité.
Des études menées à Trinidad dans lesannées 40 suggèraient une absence decorrélation entre valeurs propres et
8 CA C A O Y E R
Plantations, recherche, développement Novembre - Décembre 1995
Semis en pépinière. / Seedings in nursery.
(4) Idefor : Institut des forêts (Côte d’Ivoire).
D. P
aulin
Retour au menu

9CA C A O Y E R
Novembre - Décembre 1995 Plantations, recherche, développement
valeurs en combinaison. Cependant, deseffets additifs des caractères ayant étédémontrés, on pourrait s’attendre à trou-ver des corrélations significatives entre lavaleur des parents et les descendants. EnCôte d’Ivoire, ceci semble le cas, pour larésistance au champ à la pourriturebrune des cabosses, mais il n’y a pas decorrélation significative pour la produc-tion, la vigueur ou la tolérance auxmirides. La vigueur végétative desparents semble être corrélée négative-ment à l’efficacité de production, définiepar le rapport production/vigueur, auniveau des descendances (Sounigo etal.,1994a). Des résultats récents, obtenusen Malaisie, montrent une bonne relationentre valeur propre et valeur en croise-ment pour plusieurs caractères, dont laproduction (Lockwood et Pang, 1994).Ces résultats contrastants pourraient êtreliés aux constitutions génétiques diffé-rentes des populations étudiées (Eskes etal., 1995).
Les méthodes de sélectionLes programmes de sélection du cacaoyeront débuté en 1930 à Trinidad et au Nigeria,suivis dans les années 40 par la plupart desautres pays producteurs. La sélection mas-sale de clones et la sélection d’hybrides ontété les méthodes les plus utilisées. Les sor-
ties variétales sont principalement deshybrides de clones. Certains pays, commeTrinidad, l’Equateur, le Venezuela, l’Indo-nésie et le Came-roun (tableau) ontdiffusé des clones.
La sélection declonesLe premier pro-gramme de sélec-tion clonale fut réa-lisé à Trinidad àpartir de popula-tions de Trinitarioet de Criollo. Lesmeilleurs de cesclones ICS(5) furentdistribués aux plan-teurs, dans lesannées 1940, sousforme de boutures.Plus tard, des indi-vidus sélectionnéspour la résistanceau balai de sor-cière, obtenus àpartir de croise-ments entre HA etICS, ont été clonéset vulgarisés (clonesTSH)(6) (Freeman,1968).
La première étapedes programmes desélection de Côted’Ivoire et du Came-
roun fut une sélection clonale dans des popu-lations locales d’Amelonado (BA) et de Trini-tario.
En Côte d’Ivoire, des essais comparatifsclonaux montrent que les clones HA se clas-sent généralement en tête, suivis des clonesAmelonado locaux et enfin des clones Trini-tario, plus sensibles aux mirides. Lesmeilleurs clones produisent 1,8 à 2 t àl’hectare (Sounigo et al.,1994a).
Au Cameroun, l’objectif était de diffuserrapidement des clones sélectionnés locale-ment avec comme critères une productionde plus de 2 kg de cacao marchand/an, unemoindre sensibilité à la pourriture brune etaux mirides et l’aptitude au bouturage.Trente-cinq clones furent ensuite multipliésindustriellement par bouturage plagiotropeet vulgarisés entre 1957 et 1968. Les résul-tats au champ furent décevants en raisond’une forte mortalité des arbres au jeuneâge.
A partir des années 80, un regain d’inté-rêt pour la sélection clonale est observé,principalement en Malaisie, où des plantsgreffés sont distribués commercialement.
(5) ICS : Imperial College Selection.(6) TSH : Trinidad Selection Hybrid.
Variétés / Varieties Amérique / America Afrique / Africa Asie / Asia
Non selectionnées 55 % 80 % 65 %Non-selectedHybrides / Hybrids 40 % 20 % 30 %Clones 5 % - 5 %
TypesNon sélectionnés BA, C, T BA, HA, T, C BA, HA, TNon-selected LA, C, T LA, UA, T, C LA, UA, THybrides HA x T HA x HA HA x HAHybrids UA x T UA x UA UA x UA(ordre d’importance) HA x BA HA x BA HA x BA(order of importance) UA x LA UA x LA UA x LA
HA x C HA x T HA x TUA x C UA x T UA x TBA x T T x TLA x T T x TT x TT x T
Clones T, C T, HAT, C T, UA
C = Criollo, T = Trinitario, HA = Haut-Amazonien, BA = Bas-AmazonienC = Criollo, T = Trinitario, UA = Upper Amazon, LA = Lower Amazon
Estimation des variétés de cacaoyers cultivées dans le monde. / Estimation of thecocoa varieties grown worldwide.
Cabosses sur tronc. / Pods on trunk.
D. P
aulin
Retour au menu
La sélection d’hybridesA Trinidad, les premiers hybrides réalisésentre les clones ICS sélectionnés ne donnè-rent pas de bons résultats agronomiques.Ces clones furent croisés avec des clonesForastero HA collectés en Amazonie en1930-1940, qui possédaient un meilleurniveau de résistance à la maladie du balaide sorcière. Les descendances obtenuesmontraient une bonne valeur agronomique.Ces résultats positifs, obtenus avec deshybrides entre clones de groupes géné-tiques différents, ont été à la base de la plu-part des programmes de sélection dans lemonde.
Au Brésil, après une sélection massaledu matériel local Forastero BA, des hybri-dations furent réalisées entre ces clones etdes Forastero HA ou des Trinitario à partirde 1963 (Alvim,1975). Ces hybrides furentlargement répandus. Les nombreux géno-types amazoniens prospectés dans ce paysdans les années 1970-1980, sont encore peuutilisés dans les programmes d’hybridation.
Au Ghana et au Nigeria, les sélectionslocales de clones Forastero BA (Amelo-nado) et Trinitario se montraient très sen-sibles au swollen-shoot. Des populations deHA (G2 et G3), plus résistantes, ont été vul-garisées dans les années 50 et 60. Les
hybrides entre HA et Amelonado furentsélectionnés pour leurs meilleures homogé-néité et tolérance au swollen-shoot et à lapourriture brune. Depuis une vingtained’années, ce sont des hybrides entre HA quiont été sélectionnés et qui sont diffusés, enraison de leur meilleur niveau de résistanceau swollen-shoot (Toxopeus, 1970 ; Threshet al., 1988).
En Malaisie, du matériel Amelonado,puis Trinitario et HA, fut introduit duGhana. Des hybrides issus de croisementsentre HA et Trinitario et de croisementsentre HA furent vulgarisés, pour leursmeilleures productivité et résistance auVSD.
En Côte d’Ivoire, les premiers hybridesfurent créés en 1957, en croisant des clonesHA sélectionnés localement (UPA), avecdes clones Trinitario ou Amelonado locaux,choisis sur leur valeur propre. Les essaiscomparatifs d’hybrides confirmèrent lasupériorité des croisements intergroupesdéjà observée dans d’autres pays. A partirde 434 croisements, 12 hybrides furentsélectionnés et distribués dès 1975 auxplanteurs, sous forme de semences pro-duites dans des champs semenciers biclo-naux. En général, les hybrides issus d’uncroisement avec un parent Amelonado
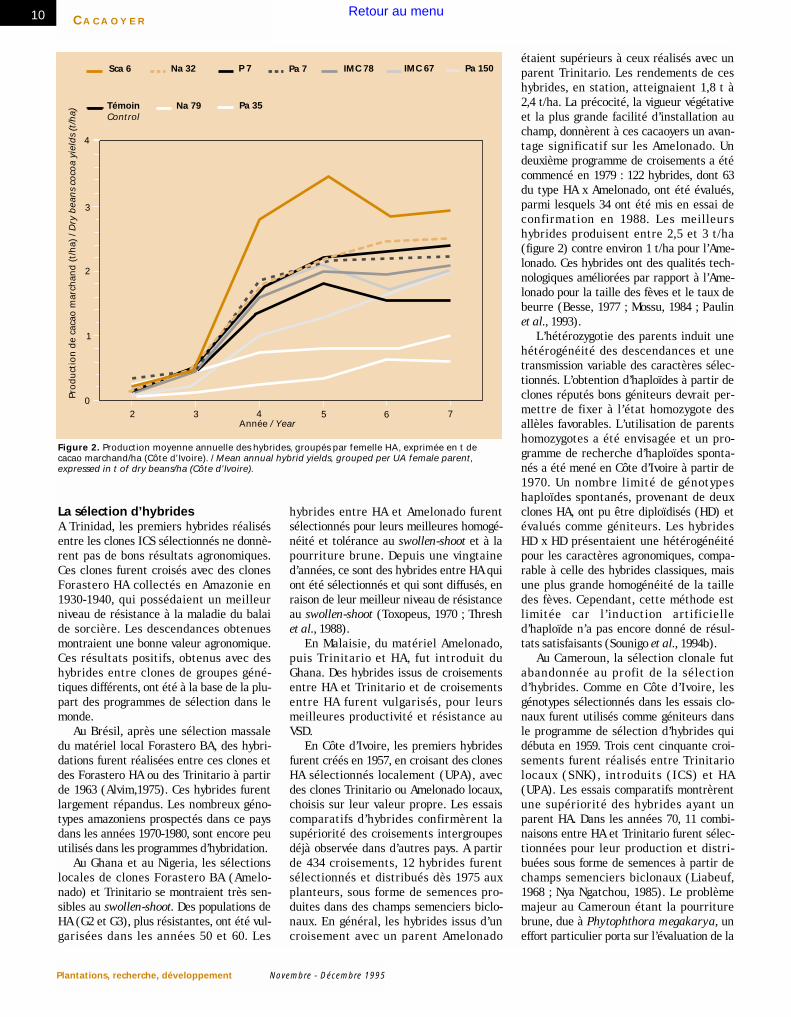
étaient supérieurs à ceux réalisés avec unparent Trinitario. Les rendements de ceshybrides, en station, atteignaient 1,8 t à2,4 t/ha. La précocité, la vigueur végétativeet la plus grande facilité d’installation auchamp, donnèrent à ces cacaoyers un avan-tage significatif sur les Amelonado. Undeuxième programme de croisements a étécommencé en 1979 : 122 hybrides, dont 63du type HA x Amelonado, ont été évalués,parmi lesquels 34 ont été mis en essai deconfirmation en 1988. Les meilleurshybrides produisent entre 2,5 et 3 t/ha(figure 2) contre environ 1 t/ha pour l’Ame-lonado. Ces hybrides ont des qualités tech-nologiques améliorées par rapport à l’Ame-lonado pour la taille des fèves et le taux debeurre (Besse, 1977 ; Mossu, 1984 ; Paulinet al., 1993).
L’hétérozygotie des parents induit unehétérogénéité des descendances et unetransmission variable des caractères sélec-tionnés. L’obtention d’haploïdes à partir declones réputés bons géniteurs devrait per-mettre de fixer à l’état homozygote desallèles favorables. L’utilisation de parentshomozygotes a été envisagée et un pro-gramme de recherche d’haploïdes sponta-nés a été mené en Côte d’Ivoire à partir de1970. Un nombre limité de génotypeshaploïdes spontanés, provenant de deuxclones HA, ont pu être diploïdisés (HD) etévalués comme géniteurs. Les hybridesHD x HD présentaient une hétérogénéitépour les caractères agronomiques, compa-rable à celle des hybrides classiques, maisune plus grande homogénéité de la tailledes fèves. Cependant, cette méthode estlimitée car l ’induction artificielled’haploïde n’a pas encore donné de résul-tats satisfaisants (Sounigo et al., 1994b).
Au Cameroun, la sélection clonale futabandonnée au profit de la sélectiond’hybrides. Comme en Côte d’Ivoire, lesgénotypes sélectionnés dans les essais clo-naux furent utilisés comme géniteurs dansle programme de sélection d’hybrides quidébuta en 1959. Trois cent cinquante croi-sements furent réalisés entre Trinitariolocaux (SNK), introduits (ICS) et HA(UPA). Les essais comparatifs montrèrentune supériorité des hybrides ayant unparent HA. Dans les années 70, 11 combi-naisons entre HA et Trinitario furent sélec-tionnées pour leur production et distri-buées sous forme de semences à partir dechamps semenciers biclonaux (Liabeuf,1968 ; Nya Ngatchou, 1985). Le problèmemajeur au Cameroun étant la pourriturebrune, due à Phytophthora megakarya, uneffort particulier porta sur l’évaluation de la
10 CA C A O Y E R
Plantations, recherche, développement Novembre - Décembre 1995
0
1
2
2
3
3
4
4 5 6 7
Pro
du
ctio
n d
e ca
cao
mar
chan
d (
t/h
a) /
Dry
bea
ns
coco
a yi
eld
s (t
/ha)
Année / Year
Sca 6 Na 32 P 7 Pa 7 IMC 78 IMC 67 Pa 150
TémoinControl
Na 79 Pa 35
Figure 2. Production moyenne annuelle des hybrides, groupés par femelle HA, exprimée en t decacao marchand/ha (Côte d’Ivoire). / Mean annual hybrid yields, grouped per UA female parent,expressed in t of dry beans/ha (Côte d’Ivoire).
Retour au menu

résistance à cette maladie. Une classifica-tion des clones fut réalisée par infectionartificielle sur cabosse. Une centaine declones furent ainsi classés (Blaha etLotodé, 1976). L’héritabilité de la résis-tance au champ fut étudiée dans un plan decroisements diallèles, qui permit aussi desupposer que la durée de maturation descabosses était une composante de la résis-tance au champ (Despréaux et al., 1989 ;Berry et Cilas, 1994).
La variabilité importante, observée ausein des familles de pleins-frères, peut êtreexploitée pour optimiser les gains géné-tiques. La sélection d’individus dans lesdescendances prend en compte la partienon-additive de la composante de lavariance génétique. Tant au Camerounqu’en Côte d’Ivoire, de très nombreuxarbres hybrides sont observés. Une sélec-tion combinée (individu-famille) sur index,basée sur l’estimation de la valeur géné-tique pour la production, la vigueur,l’encombrement et la sensibilité à la pourri-ture brune est réalisée actuellement auCameroun et le sera prochainement enCôte d’Ivoire. Pour certains caractèrescomme la résistance à la pourriture brune,on en attend un progrès important (Cilas etPaulin, 1994). Les individus sélectionnéspeuvent être multipliés végétativement ouutilisés comme nouveaux géniteurs.
La sélection récurrenteLa sélection traditionnelle d’hybrides declones présente des limites. En effet, ellen’assure pas de progrès génétique continu etl’amélioration des caractères quantitatifs, telsque la résistance aux maladies, est peu effi-cace. Pour augmenter la fréquence des géno-types cumulant des gènes favorables, les géné-ticiens proposent une amélioration despopulations de base par une sélection récur-rente. Un schéma de ce type a été lancé par leCirad(7) et l’Idefor en Côte d’Ivoire en 1990.BA et Trinitario (BA-0) et, par ailleurs, HA(HA 0) constituent les deux populations dedépart. La valeur propre, l’aptitude à la com-binaison et la représentativité de la diversitégénétique des groupes déterminent le choixdes parents. Les Trinitario sont utilisés pourintroduire des gènes de qualité dans la popu-lation Forastero BA. Dans une premièrephase, la sélection récurrente aboutira, aprèsdeux cycles, à la création de deux populationsaméliorées. Une sélection du type combinéindividu-famille sera appliquée. Ce schémapermettra de produire, dès le premier cycle,
des sorties variétales hybrides ou clones obte-nus au sein de chaque population. Une sélec-tion récurrente réciproque sera alors abordéeentre ces deux populations. Au cours de cettephase, des sorties variétales d’hybrides inter -population seront également envisagées, per-mettant d’exploiter un éventuel effet d’hété-rosis exprimé par ce type de croisement(figure 3). Les plans de croisements appliquésau premier cycle sont des plans diallèles etfactoriels ; pour les cycles suivants, des plansde croisements partiels seront utilisés afind’augmenter les chances de recombinaison etd’éviter l’érosion génétique (Clément et al.,1994 ; Eskes et al., 1995).
Aujourd’hui en Malaisie, Bal Plantationsà Sabah développe aussi un programmed’amélioration génétique par sélectionrécurrente. Il consiste à alterner essais clo-naux et essais de descendances à partir deplans de croisements factoriels entre HA.Les sorties variétales prévues sont desclones (greffes) mais aussi des hybrides(semences). Les meilleurs hybrides du pre-
mier cycle se montrent de 20 à 50 % plusproductifs que les témoins. Des informa-tions intéressantes sur les paramètres géné-tiques concernant la population des HAapparaissent déjà : corrélations significa-tives entre valeur propre et valeur en com-binaison et prédominance de l’aptitudegénérale à la combinaison (Lockwood etPang, 1993 ; Lockwood et Pang, 1994).
Conclusion etperspectives La méthode d’amélioration du cacaoyer laplus utilisée est encore la sélectiond’hybrides de clones. Les variétés hybridesproduisant du cacao ordinaire, sélection-nées depuis une trentaine d’années, semontrent bien adaptées à la plupart desrégions et en particulier aux régions de pro-duction pionnières. Pour la capacité d’adap-tation au champ, la précocité et la produc-tion, cette sélection a généralementapporté une amélioration significative par
11CA C A O Y E R
Novembre - Décembre 1995 Plantations, recherche, développement
Collection de travailBreeding population
BA0LAO
HA0UAO
sélectionselection
sélectionselection
sélectionselection
sélectionselection
sélectionselection
sortie variétalevarietytesting
sortie variétalevarietytesting
sortie variétalevarietytesting
sortie variétalevarietytesting
sortie variétalevarietaloutput
BA1 x BA1LA1 x LA1
BA2 x BA2LA2 x LA2
BA2 x HA2LA2 x UA2
HA x HAUA x UA
HA1 x HA1UA1 x UA1
HA2 x HA2UA2 x UA2
BA1LA1
HA1UA1
BA2LA2
HA2UA2
BA3LA3
HA3UA3
BA x BABA x TLA x LALA x T
Figure 3. Schéma de sélection récurrente en Côte d’Ivoire. / Recurrent selection scheme in Côted’Ivoire.
(7) Cirad : Centre de coopération internationale enrecherche agronomique pour le développement.
Retour au menu

rapport aux populations locales. Cependant,le niveau de résistance aux maladies etl’homogénéité doivent être améliorés.
Grâce à la caractérisation et à l’évalua-tion accrue des collections, l’exploitation dela diversité génétique devrait s’intensifier.Avec l’utilisation des marqueurs molécu-laires, une meilleure connaissance des dis-tances génétiques précisera les liaisonsentre distances, entre géniteurs et valeurdes hybrides, tant en intergroupe qu’enintragroupe. Le niveau d’hétérozygotie desgéniteurs pourra être également pris encompte pour améliorer l’homogénéité deshybrides. La prédominance de l’aptitudegénérale à la combinaison pour de nom-breux caractères, sera mise à profit pourcumuler des gènes impliqués dans lescaractères quantitatifs par des recombinai-sons. La mise en évidence de marqueursmoléculaires liés aux caractères d’intérêtagronomique (QTL) pourrait permettredans un avenir relativement proche d’amé-liorer l’efficacité de la sélection (sélectionassistée par marqueurs).
Certains centres de recherche utilisentactuellement de nouvelles stratégies d’amé-lioration. La sélection de clones au sein desfamilles hybrides devrait apporter des gainsgénétiques rapides, en particulier pour larésistance aux maladies et l’homogénéité.La technique de multiplication in vitro, sil’embryogenèse somatique se confirmecomme fiable, permettra de diffuser rapide-
ment des clones performants sous forme deplants orthotropes. L’amélioration despopulations par sélection récurrente assu-rera des gains génétiques significatifs àlong terme. Pour répondre à la demandespécifique en cacao fin, la revalorisation etl’amélioration génétique des populationstraditionnelles Criollo, Nacional et Trinita-rio sont actuellement à l’ordre du jour.L’objectif est précisément d’améliorer lesaptitudes agronomiques de ces cacaoyers,tout en conservant les caractères de qualitéaromatique. Les caractères recherchéspourraient être introduits par rétrocroise-ments avec des génotypes d’origine HA.
Peu de centres de recherche dans lemonde sont capables, actuellement, demaintenir un programme de sélection delongue durée. La conjoncture économiquedu cacao, plutôt défavorable durant la der-nière décennie, en est certainement lacause. Aujourd’hui cependant, la situationse montrant plus favorable, on assiste à unregain d’intérêt pour l’amélioration géné-tique du cacaoyer. Les relations entre lessélectionneurs du cacaoyer se sont accruesgrâce à la création, en 1994, d’INGENIC(International group for the GeneticImprovement of Cocoa) , une associationinternationale de généticiens du cacaoyer.Elle devrait stimuler la coopération interna-tionale dans le domaine de l’améliorationgénétique du cacaoyer. ■
12 CA C A O Y E R
Plantations, recherche, développement Novembre - Décembre 1995
Récolte à Divo (Côte d’Ivoire). / Harvesting at Divo (Côte d’Ivoire).
Le cacaoyer Criollo a été cultivé,dès la préhistoire, au Mexique eten Amérique centrale. Les Mayasle domestiquèrent pour consom-mer ses fèves et s’en servircomme monnaie d’échange. Dèsle début du XVIIe siècle, laculture s’étendit aux Caraïbes(Jamaïque, Haïti, Trinidad) et àl’Amérique du Sud (Venezuela).Les colons espagnols exportèrentcette culture en Indonésie en1560 et aux Philippines en 1670.Au XVIIIe siècle, au Brésil, laculture des Forastero, établiedans l’Etat de Para, s’étendit àBahia, où elle se développaconsidérablement. En Equateur,des cacaoyers d’origine locale, detype Nacional, furent exploitésdès la fin du XVIIe siècle, et cepays devint alors le plus grandpays producteur durant tout leXIXe siècle. Entre 1822 et 1878,des Forastero, puis des Trinitario,furent introduits en Afrique del’Ouest et en Afrique centralepar les Portugais depuis l’Amé-rique du Sud. La culture prit unessor spectaculaire au Camerounet au Nigeria (Trinitario), auGhana et surtout en Côte d’Ivoire(Forastero Amelonado) qui resteaujourd’hui le premier pays pro-ducteur au monde. La cacaocul-ture en Asie s’est développée tar-divement, mais de façon trèsrapide, à partir de la fin du XIXe
siècle, en Indonésie (Java etSumatra) avec des cacaoyersCriollo, puis Forastero, et enMalaisie (Sabah) depuis lesannées 50, avec des cacaoyersForastero Amelonado et Hauts-Amazoniens. L’extension de lacacaoculture dans le monde s’estfaite d’Ouest en Est avec descycles productifs successifs tou-chant tour à tour l’Amérique,l’Amérique du Sud, l’Afrique etaujourd’hui l’Asie du Sud-Est.Cette évolution s’est accompa-gnée du déclin de la principalezone de production que futl’Amérique centrale jusqu’auXVIIIe siècle (Jarrige et Ruf,1990).
■ Histoire de la cacaocultureD
. Pau
lin
Retour au menu

13CA C A O Y E R
Novembre - Décembre 1995 Plantations, recherche, développement
Bibliographie / References
ALVIM P. de T., 1975. Cocoa research in Brazil. Cocoa
Grower’s Bull. 23 : 18-24.
BARTLEY B.G., 1981. The status of genetic resistance in
cacao to Crinipellis perniciosa (Stahel) Singer. In :
VIe conférence internationale sur la recherche
cacaoyère, Caracas, Venezuela, nov. 1977. Lagos,
Nigeria, Cocoa Producers’ Alliance, p. 56-59.
BESSE J., 1977. La sélection générative du cacaoyer en
Côte d’Ivoire : bilan et orientation des recherches
en 1975. Ve conférence internationale sur la
recherche cacaoyère, Ibadan, Nigeria, 1-9 sept.
1977. Idaban, Nigeria, Cocoa Research Institute, p.
95-103.
BERRY D., CILAS C., 1994. Etude génétique de la réaction
à la pourriture brune des cabosses de cacaoyers
(Theobroma cacao L.) issu d’un plan de croise-
ments diallèles. Agronomie 14 (9) : 599-609.
BLAHA G., LOTODÉ R., 1976. Un critère primordial de
sélection du cacaoyer au Cameroun : la résistance à
la pourriture brune des cabosses (Phytophthora
palmivora). Variations des réactions à la maladie
en liaison avec les données écologiques et l’état
physiologique des fruits. Café Cacao Thé 20 (2) :
97-116.
CILAS C., PAULIN D., 1994. Estimation de paramètres
génétiques pour quelques caractères chez le
cacaoyer (Theobroma cacao L.). In : XIe confé-
rence internationale sur la recherche cacaoyère,
Yamoussoukro, Côte d’Ivoire, 18-24 juillet 1993.
Lagos, Nigeria, Cocoa Producers’ Alliance, p. 443-
447.
CLAPPERTON J., YOW S., CHAN J., LIM D., LOCKWOOD R.,
ROMANCZYK L., HAMMERSTONE J., 1994. The contribu-
tion of genotype to cocoa (Theobroma cacao L.)
flavour. Trop. Agr. (71) : 303-308.
CLÉMENT D., SOUNIGO O., N’GORAN J.A.K., ESKES A.B.,
1994. Amélioration génétique du cacaoyer en Côte-
d’Ivoire : présentation d’un nouveau schéma de
sélection. In : XIe conférence internationale sur la
recherche cacaoyère, Yamoussoukro, Côte d’Ivoire,
18-24 juillet 1993. Lagos, Nigeria, Cocoa Producers’
Alliance, p. 451-456.
DESPRÉAUX D., CLÉMENT D., PARTIOT M., 1989. La pourri-
ture brune des cabosses du cacaoyer au Cameroun :
mise en évidence d’un caractère de résistance au
champ. Agronomie 9 : 683-691.
ESKES A.B., PAULIN D., CLÉMENT D., N’GORAN J.A.K., SOU-
NIGO O., LACHENAUD P., CILAS C., BERRY D., YAGMPAM
A., 1995. Selection methods applied and genetic
knowledge generated in cocoa breeding in Côte
d’Ivoire and Cameroun. In : International workshop
on cocoa breeding strategies, Kuala Lumpur, Malai-
sie, 20-21 oct. 1994. Reading, Grande-Bretagne,
INGENIC, p. 41-56.
FALQUE M., 1994. Fécondation et développement des
fruits et des graines chez le cacaoyer Theobroma
cacao L. Thèse, Institut national polytechnique,
Toulouse, France, 177 p.
FREEMAN W.E., 1968. Some aspects of the cocoa bree-
ding programme. Proc. Agric. Soc. Trin. Tobago 69 :
1-15.
JARRIGE F., RUF F., 1990. Comprendre la crise du cacao.
Café Cacao Thé 34 (3) : 213-229.
KEANE P.J., PRIOR C., 1991. Vascular Streak Dieback of
Cocoa. Oxon, Grande-Bretagne, International Myco-
logical Institute, Phytopathological papers (33),
39 p.
LACHENAUD P., 1991. Facteurs de la fructification chez le
cacaoyer (Theobroma cacao L.). Thèse, Institut
national agronomique, Paris-Grignon, France,
188 p.
LANAUD C., RISTERUCCI A.M., N’GORAN J.A.K., CLÉMENT D.,
FLAMENT M.H., LAURENT V., 1995. A genetic linkage
map of Theobroma cacao L. Theor. Appl. Genet.
(sous presse)
LAURENT V., RISTERUCCI A.M., LANAUD C., 1994. Genetic
diversity in cocoa revealed by cDNA probes. Theor.
Appl. Genet. 88 : 193-198.
LAWRENCE J.S, 1978. Evaluation of methods for assessing
resistance of Theobroma cacao cultivars and
hybrids to Phytophthora palmivora Bulter. Techni-
cal Bulletin CEPEC/CEPLAC (62) : 47.
LECERTEAU E., CROUZILLAT D., PÉTIARD V., 1994. Evalua-
tion de la diversité génétique de Theobroma cacao
par les techniques de détection d’ADN polymorphe
amplifié au hasard (RAPD) et de polymorphisme de
longueur des fragments de restriction de l’ADN
(RFLP). In : XIe conférence internationale sur la
recherche cacaoyère, Yamoussoukro, Côte d’Ivoire,
18-24 juillet 1993. Lagos, Nigeria, Cocoa Producers’
Alliance, p. 337-344.
LIABEUF J., 1968. Etat des travaux effectués pour la
sélection du cacaoyer au Cameroun et perspectives
d’avenir. IIe conférence internationale sur la
recherche cacaoyère, Bahia, Brésil, 19-26 nov. 1967.
Lagos, Nigeria, Cocoa Producers’ Alliance, p. 86-92.
LOCKWOOD G., PANG J.T.Y., 1993. Utilisation of cocoa
germplasm in breeding for yield. In : International
workshop on conservation, characterisation and
utilisation of cocoa genetic resources in the 21st
century, Port of Spain, Trinidad, 13-17 sept. 1992.
Saint-Augustine, Trinidad, Cocoa Research Unit, p.
198-215.
LOCKWOOD G., PANG J.T.Y., 1994. Additive inheritance of
yield in cocoa. XIe conférence internationale sur la
recherche cacaoyère, Yamoussoukro, Côte d’Ivoire,
18-24 juillet 1993. Lagos, Nigeria, Cocoa Producers’
Alliance, p. 415-423.
LOCKWOOD G., ESKES A.B., 1996. Relationship between
cocoa variety and quality. Rencontres cacao, Mont-
pellier, 30 juin 1995. Montpellier, France, CIRAD-
CP (à paraître).
LOPEZ BAEZ O., 1993. Embryogenèse somatique de
plantes de cacaoyer Theobroma cacao L. à partir de
pièces florales. Thèse, école nationale supérieure
agronomique, Rennes, France, 137 p.
MOSSU G., 1984. Les hybrides de cacaoyers en Côte
d’Ivoire : origine et comportement. Actions priori-
taires et perspectives d’avenir en sélection. IXe
conférence internationale sur la recherche
cacaoyère, Lomé, Togo, 12-18 fév. 1984. Lagos, Nige-
ria, Cocoa Producers’ Alliance, p. 252-262.
N’GORAN J.A.K., LAURENT V., RISTERUCCI A.M., LANAUD C.,
1994. Comparative genetic diversity studies of Theo-
broma cacao L. using RFLP and RAPD markers.
Heredity 73 : 589-597.
NYA NGATCHOU J., 1985. Etat d’avancement des travaux
de génétique et d’amélioration du cacaoyer au
Cameroun. In : VIIe conférence internationale de la
recherche cacaoyère, Lomé, Togo, 12-18 fév. 1984.
Lagos, Nigeria, Cocoa Producers’ Alliance, p. 507-
511.
NYASSE S., BLAHA G., CILAS C., 1994. Pathogénie de Phy-
tophthora megakarya et son implication pour la
mise au point d’un test précoce de sensibilité. In :
XIe conférence internationale sur la recherche
cacaoyère, Yamoussoukro, Côte d’Ivoire, 18-24
juillet 1994. Lagos, Nigeria, Cocoa Producers’
Alliance, p. 83-90.
PAULIN D., MOSSU G., LACHENAUD P., CILAS C., 1993. La
sélection du cacaoyer en Côte d’Ivoire. Analyse du
comportement de soixante-deux hybrides dans
quatre localités. Café Cacao Thé 37 (1) : 3-20.
PAULIN D., MOSSU G., LACHENAUD P., ESKES A.B., 1994.
Genetic analysis of a factorial crossing scheme with
cacao hybrids tested in four locations in Ivory
Coast. In : International cocoa conference: chal-
lenge in the 90’s, Kuala Lumpur, Malaisie, 25-28
sept. 1991. Kuala Lumpur, Malaisie, Malaysian
Cocoa Board, p. 73-83.
SOUNIGO O., N’GORAN J.A.K., COULIBALY N., CLÉMENT D.,
LACHENAUD P., 1994a. Evaluation de clones de
cacaoyers pour la productivité, la résistance aux
mirides et la résistance à la pourriture des
cabosses. In : XIe conférence internationale sur la
recherche cacaoyère, Yamoussoukro, Côte d’Ivoire,
18-24 juillet 1993. Lagos, Nigeria, Cocoa Producers’
Alliance, p. 375-382.
SOUNIGO O., LANAUD C., LACHENAUD P., BASTIDE P., 1994b.
Etude du comportement en croisement de quatorze
haploïdes doublés de cacaoyer (Theobroma cacao
L.) en Côte d’Ivoire. In : XIe conférence internatio-
nale sur la recherche cacaoyère, Yamoussoukro,
Côte d’Ivoire, 18-24 juillet 1993. Lagos, Nigeria,
Cocoa Producers’ Alliance, p. 437-442.
THRESH J.M., OWUSU G.K., BOAMAH A., LOCKWOOD G., 1988.
Ghanian cocoa varieties and swollen shoot virus.
Crop Prot. 7 : 219-231.
TOXOPEUS H., 1970. Establishment ability of cacao pro-
genies in Nigeria. Euphytica 19 : 327-333.
WOOD G.A.R., LASS R.A., 1989. Cocoa. 4e édition.
Londres, Grande Bretagne, Longman, Tropical Agri-
culture Series, 620 p.
Retour au menu

Cocoa, Theobroma cacao L., originatedfrom the rainforests of TropicalAmerica. Cultivation expanded from
the 19th century onwards, following theinvention of a press to extract cocoa butterfrom beans, enabling chocolate production.When the plant was introduced in Africa, atthe start of the 20th century, world cocoaproduction really took off: from 115,000 t, itnow totals 2.5 million t of merchantablecocoa, i.e. a value of some 3 thousand milliondollars. Africa (Côte d’Ivoire, Ghana)produces 60%, Asia (Malaysia, Indonesia) 20%and America (Brazil) 20%. Some countriessuch as Ecuador, Trinidad, Colombia,Venezuela, Mexico, Indonesia and Papua NewGuinea also sell a more aromatic cocoa knownas fine cocoa (around 5% of world production)(map).
CultivationMost cocoa growing zones have developed oncleared forest land, with some forest treeskept to provide shade. However, controlledshade can be installed by planting leguminousshade trees after felling the whole forest.Growing in full sunlight, a more recentdevelopment, ensures higher yield but callsfor more regular phytosanitary treatments. Inthe first few years, cocoa is often intercroppedwith food crops. Cocoa is particularly sensitiveto changes in its ecological environment.Replanting is often difficult, and cocoagenerally has been a pioneer front crop(Jarrige and Ruf, 1990). The productionstructures vary:
• in Africa, they are family-owned farmscovering less than 5 ha;
• in America, family-owned smallholdings cancover around 20 ha, but there are also largeestates in Ecuador, Brazil, Costa Rica andTrinidad;
• in Asia (Malaysia, Indonesia, Papua NewGuinea), where cocoa growing has onlydeveloped recently, private or state-ownedestates and smallholdings exist side by side.
The hazards of cocoa growingCocoa is particularly sensitive to pests anddiseases, which often limit yield. The maindiseases are fungal:• black pod (Phytophthora spp.) is spread
worldwide. The most harmful species,P. megakarya, can cause yield losses ofover 50% in Central Africa;
• witches’ broom (Crinipellis perniciosa),which is found throughout the Caribbeanand South America, causes theuncontrolled proliferation of branches andflower cushions and destroys the seeds inthe young fruits;
• Ceratocystis wilt (Ceratocystis fimbriata)is also found in some American countries. Itis a wilting disease that eventually kills thetree;
• vascular streak dieback, or VSD, (Onco-basidium theobromae) is widespread inSoutheast Asia. It causes leaf fall and killsthe branches;
• swollen shoot, the only known virus disease,is characterized by the swelling of thestems, and can kill the tree. It is found inNigeria, Ghana and Togo.The most common insect pests are sucking
insects, particularly mirids (Sahlbergella sp.,Distantiella sp. and Helopeltis sp.), whichattack young shoots and pods. They causeconsiderable damage in Africa and Asia. InSoutheast Asia, a pod borer (Conopomorphacramerella) causes substantial fruit losses(Wood and Lass,1989).
The mean yield recorded in most countries(500 kg/ha) are well below those obtained atexperimental stations, partly due to thesubstantial heterogeneity of plantations andthe use of planting material of uncontrolledorigin. The economic life span of a plantationvaries from 20 to 40 years. Upkeep is halted inold plantings, which are gradually abandonedin favour of new plantings. The search forways of rejuvenating or replacing oldplantings with new varieties is therefore amajor challenge.
Genetic resources and population structuresThere are some twenty wild Theobromaspecies on the American continent, but only
T. cacao is grown for the chocolate industry.The Upper Amazon Basin is assumed to be thecentre of origin and of primary diversity of theT. cacao species and Central America itscentre of domestication. There are three maingenetic groups: Forastero, Criollo andTrinitario. The distinction between thesegroups is based on their geographical originand on the morpho-logical characteristics oftheir pods and beans.
The Forastero group, which currentlyaccounts for over 80% of world cocoaproduction, comes from the Am00azon Basin.There are two origins, Upper Amazon andLower Amazon, with graduated variabilitybetween the two. The Lower Amazon (LA)varieties are traditionally grown in Brazil andWest Africa. Due to their vigour, precocity andrelative resistance to certain pathogens, theUpper Amazon (UA) varieties, which werecollected more recently and varysubstantially, are often used in breedingprogrammes.
The Criollo group, which includes primitivevarieties domesticated by the AmericanIndians, produces cocoa with goodorganoleptic qualities (fine cocoa). Thesevarieties are not very widely grown nowadays,as they are generally not very vigorous and aresusceptible to insects and fungus diseases.
The Trinitario group comprises hybrids ofthe above two groups. Most of the recordedgenotypes are of cultivated origin. They havewidely varying agronomic and technologicalcharacteristics. Due to their high vigour andgood bean quality, they have been distributedin numerous countries.
Wide genetic diversity Over 7,300 original genotypes, including morethan 2,800 originally wild species, arerecorded worldwide. The largest collectionsare in Trinidad, Brazil, Costa Rica, Ecuador,Ghana, Côte d’Ivoire and Malaysia. Severalsurveys since 1940 have collected originalForastero populations: in Peru, Ecuador,Brazil and Colombia, and more recently inFrench Guiana, between 1987 and 1995. Thecharacterization and evaluation of thismaterial are still largely incomplete, and few
14 CO C O A
Plantations, recherche, développement Novembre - Décembre 1995
(1) RFLP: DNA (Desoxyribonucleic acid) RestrictionFragment Length Polymorphism.(2) RAPD: Random Amplified Polymorphic DNA Detection.
Cocoa: breeding strategies
Paulin D., Eskes A.B.CIRAD-CP, BP 5035, Montpellier Cedex 1, France
Increasing resistance to diseases and insects: black pod, witch’s broom, pod borer, etc. and improving qualityare the current challenges for this important traditional crop.
Retour au menu

15CO C O A
Novembre - Décembre 1995 Plantations, recherche, développement
wild clones are currently used in breedingprogrammes.
Genetic diversity has been studied usingmolecular markers (RFLP(1) and RAPD(2)).The genetic structure detected by thesemarkers generally corresponds to thegeographical structure of the species(Laurent et al. , 1994). The substantialdiversity amongst UA is confirmed (figure 1)(N’goran et al., 1994; Lecerteau et al., 1994).The diversity of local populations is less wellknown, but each group has a degree ofvariability, with some original localpopulations such as Porcelana fromVenezuela or the wild cocoa trees collected inFrench Guiana.
Molecular biology tools have also enabledmapping of the cocoa genome, identifying193 loci (5 isoenzyme markers, 160 RFLPmarkers and 28 RAPD markers). The map cannow be used to locate genes of agronomicvalue. Work is under way to identify QTL(3)
for black pod resistance and bean size(Lanaud et al.,1995).
Breeding criteriaOne of the main cocoa breeding objectivesnowadays is improved resistance to parasites.Chemical control is not always effective andentails high costs for growers. Sources ofresistance have been identified for black pod,witches’ broom and VSD. Significant progressis expected in terms of varietal resistance,using new and more efficient selectionmethods.
Disease resistanceThe variability of true resistance to black podwas detected by inoculating pods, leaves oryoung stems (Blaha and Lotodé, 1976;Lawrence, 1978). However, observations ofblack pod rates in the field can give differentclassifications from those obtained byartificial inoculation. These differences maybe linked to varying pod ripening times andfruiting periods. An artificial inoculation teston leaves is currently being studied, toevaluate true resistance (Nyasse et al., 1994).
The level of susceptibility to witches’ broomof certain cocoa populations has beenestimated by field observations, and artificialinoculation tests have been used in thenursery (Bartley, 1981). An inoculation teston leaves is currently being tried out inTrinidad, and has already given promisingresults (Ducamp, personal communication).
For VSD, resistance sources were identifiedby field observations after natural inoculation,
but no artificial inoculation tests have beendeveloped yet (Keane and Prior, 1991).However, resistant commercial varieties arealready available.
Resistance to insectsSusceptibility to mirids is taken into accountin the Idefor(4) breeding programme in Côted’Ivoire. A note is attributed, estimating theintensity and extent of damage and the abilityof affected zones to regenerate. However, thistype of scoring system cannot differentiatebetween preference, resistance and tolerance.Preference tests have been carried out in thelaboratory, and revealed differences insusceptibility and preference between UA,Amelonado and Trinitario types.
No effective source of resistance to podborer has yet been detected in Asia.
YieldCocoa trees begin to bear at between two andfive years of age. Dry cocoa yield (weight offermented, dried beans per hectare) perplantation are calculated by totalling five tosix years of peak production. In general, ripe,healthy, rotten and pest-damaged pods arecounted and the healthy pods weighed. A «podindex» is also determined: the number of podsneeded to obtain one kilo of dry cocoa beans.
VigourVigour is measured by the increase in stemdiameter on young trees and by stem girth onadult trees. The yield vigour ratio can be usedto express the relationship between vege-tative development and yield (yield effi-ciency). This variable is important whenselecting individuals plants in heterogeneousstands.
QualityHybrid breeding programmes take account ofbean size and butter content. Flavour qualityis more difficult to determine, as it dependsheavily on post-harvest processing. The basicflavours (cocoa flavour, bitterness, astrin-gency, acidity) and specific tastes areevaluated by liquor tastings. A recent studyshowed that genotype has a marked influenceon the organoleptic characteristics and thataroma potential is heritable. Differences incocoa and fruity flavours have been demons-trated between UA Forastero and certainTrinitario varieties (Clapperton et al., 1994).Unlike for the physical characters of thebeans, there is no reliable information thatwould enable the integration of theseorganoleptic characters into a selection index,as consumer preferences varie from one
country to another (Lockwood and Eskes,1996).
Genetic parametersThe different genetic parameters relating to thecharacters to be improved have been estimatedby carrying out factorial and diallel crossingplans. For most of the characters studied, themain genetic effects were additive. Dominanceeffects, which were generally less significant,existed for vigour, bean size and yield. Yield canbe positively correlated to early vigour (Paulin etal., 1994). There was marked environmentalvariance for the main selection criteria (20 to78%) in the single-tree plot designs (Eskes etal., 1995).
Narrow sense heritability, estimated fromindividual tree data, was moderate for black podresistance (0.35) and yield (0.30), low for adulttree vigour (0.15) and very low for insectresistance (Cilas et Paulin, 1994). Theheritability of bean size was high (0.65), and
Cocoa is a diploid species belonging to theSterculiaceae family. The tree is grownfrom seed, and comprises an orthotropicaxis that produces several successivecanopies from about eighteen monthsonwards, each with five plagiotropicbranches. Vegetative propagation ispossible by cuttings or by graftingorthotropic and plagiotropic wood. In vitromultiplication of cocoa has not yet beenmastered, although progress has beenmade in somatic embryogenesis usingfloral tissues (Lopez Baez, 1993). Theflowers are small, very numerous on thetrunk and branches, and hermaphrodite.The pollen is only viable for 48 hours. Only5% to 10% of the flowers are insect-pollinated. There is gameto-sphorophyticincompatibility, and self-compatibility variesdepending on the population. Theallogamy rate varies, depending on theenvironment, but can generally beestimated at 75%. Pollination intensity andnutrition affect pod setting and filling(Falque, 1994; Lachenaud, 1991). Fruitproduction begins two to three years afterplanting, and the harvesting period,generally spread over the year, depends onthe climatic conditions. Bean size, shellpercentage, butter content and butterhardness govern the technological value ofmerchantable cocoa, whereas itsorganoleptic quality depends on its geneticorigin and on post-harvest processing(fermentation, drying and roasting).
■ The plant
(3) QTL: Quantitative trait loci.(4) Idefor: Institut des Forêts (Côte d’Ivoire).
Retour au menu

16 CO C O A
Plantations, recherche, développement Novembre - Décembre 1995
that of butter content and fruity flavour wasmoderate (Lockwood and Eskes, 1996).
The genotype-environment interactionaffected yield, vigour and black pod incidence,but the hybrid classifications for the differentsites in Côte d’Ivoire were generally stable. Onanother level, the best UA parents, selected inTrinidad, also performed well in crosses inmany other countries, indicating somestability as well.
Early results from Trinidad indicated lackof correlation between clonal behaviour andoffspring. However, the additive effects ofmost characters suggest that significantcorrelations between genotypic andphenotypic values may be found. In Côted’Ivoire, this seemed to be the case for blackpod in the field, but there was no significantcorrelation for yield, vigour or miridtolerance. Vegetative vigour of the parentsseemed to be negatively correlated to yieldefficiency in the progenies (Sounigo et al.,1994a). Recent results from Malaysia revealeda strong correlation between clonal value andcombining ability for several characters,including yield (Lockwood and Pang, 1994).These contrasting results might be explainedby the different genetic make up of thepopulations under study (Eskes et al., 1995).
Breeding methodsCocoa breeding programmes began in 1930 inTrinidad and Nigeria, followed by most otherproducing countries in the 1940s. Massselection of clones and hybrid selection werethe most widely used methods. Most selectedvarieties were mixtures of bi-parental crosses.Certain countries, such as Trinidad, Ecuador,Venezuela, Indonesia and Cameroon (table),have distri-buted clones.
Clone selectionThe first clonal selection programme wasimplemented in Trinidad using Trinitario andCriollo populations. The best of the ICS(5)
clones were distributed to growers in the1940s, as rooted cuttings. Individuals bred forresistance to witches’ broom, obtained bycrossing UA and ICS varieties, weresubsequently cloned and distributed (TSH(6)
clones) (Freeman, 1968).The first step in the breeding programmes
in Côte d’Ivoire and Cameroon was clonalselection in local Amelonado (LA) andTrinitario populations.
In Côte d’Ivoire, clone trials have shownthat UA clones generally perform best,followed by local Amelonado clones and then
Trinitario clones, which are more susceptibleto mirids. The best clones produce 1.8 to 2 tper hectare (Sounigo et al., 1994a).
In Cameroon, the aim was to distributerapidly local clones, selected for the followingcriteria: yield of over 2 kg of dry cocoa/year,low susceptibility to black pod and mirids andsuitability for multiplication by cuttings.Thirty-five clones were distributed asplagiotropic cuttings between 1957 and 1968.The field results were disappointing, due tohigh mortality rates amongst young trees.
Since the 1980s, there has been renewedinterest in clonal selection, mainly inMalaysia, where budded plants are usedcommercially.
Hybrid breedingIn Trinidad, the first hybrids bred fromselected ICS clones failed to give goodagronomic results. These clones were crossedwith UA Forastero gentoypes collected in1930-1940, which had better witches’ broomresistance. The progenies obtained performedbetter, and the positive results obtained withhybrids between clones from different geneticgroups formed the basis for most breedingprogrammes worldwide.
In Brazil, following mass selection of localLA Forastero material, these clones werecrossed with UA Forastero and Trinitariomaterial from 1963 onwards (Alvim, 1975).These hybrids were widely distributed. Littleuse has yet been made in hybridizationprogrammes of the large numbers of Amazongenotypes collected in this country in the1970s and 80s.
In Ghana and Nigeria, the local selectionsof LA Forastero (Amelonado) and Trinitarioclones were highly susceptible to swollenshoot. Populations of more resistant UAgenotypes (F2 and F3) were distributed in the1950s and 60s. The hybrids between UA andAmelonado varieties were selected for theirgreater uniformity and tolerance to swollenshoot and black pod. For some twenty yearsnow, UA x UA hybrids have been selected anddistributed, due to their increased resistanceto swollen shoot (Toxopeus, 1970; Thresh etal., 1988).
In Malaysia, Amelonado and then Trinitarioand UA material was introduced from Ghana.Hybrids obtained by crossing UA andTrinitario and crosses between UA weredistributed for their better yield andresistance to VSD.
In Côte d’Ivoire, the first hybrids werecreated in 1957 by crossing UA clones selectedlocally (UPA) with local Trinitario orAmelonado clones, chosen for theirphenotypic value. Hybrid trials confirmed the
superiority of the crosses between clones fromdifferent groups already observed in othercountries. From 434 crosses carried out, 12hybrids were selected and distributed togrowers from 1975 onwards, in the form ofseeds produced in bi-clonal seed gardens.Generally speaking, the hybrids obtained bycrossing with an Amelonado parent weresuperior to those with a Trinitario parent. Inexperimental plots, these hybrids yielded 1.8to 2.4 t/ha. Their precocity, vegetative vigourand greater establishment ability in the field,give these trees a significant advantage overAmelonado materials. A second crossingprogramme was launched in 1979: 122hybrids, including 63 UA x Amelonado type,were evaluated and 34 were planted inconfirmation trials in 1988. The best hybridsproduce between 2.5 and 3 t/ha (figure 2),compared to around 1 t for Amelonado. Thesehybrids have better technologicalcharacteristics compared to the Amelonadomaterial as far as bean size and buttercontent are concerned (Besse, 1977; Mossu,1984; Paulin et al., 1993).
The fact that the parents are heterozygousleads to heterogeneous progenies and variableinheritance of the selected characters.Obtaining haploids from clones reputed to begood parents should make it possible to fixfavourable alleles in a homozygous state. Theuse of homozygous parents has been envi-saged, and a programme to find spontaneoushaploids was implemented in Côte d’Ivoirefrom 1970 onwards. A limited number of suchhaploid genotypes from two UA clones werediploidized (DH) and evaluated as parents.The variation for agronomic characters of theDH x DH hybrids was similar to that ofconventional hybrids, but bean size was morehomogeneous. However, this method islimited, as the artificial induction of haploidshas not yet given satisfactory results (Sounigoet al., 1994b).
In Cameroon, clone selection has beenabandoned in favour of hybrid selection. As inCôte d’Ivoire, the genotypes selected in clonaltrials were used as parents in the hybridselection programme, which began in 1959.Three hundred and fifty crosses were carriedout between local Trinitario (SNK),introduced Trinitario (ICS) and UA (UPA)material. Varietal trials revealed that hybridswith a UA parent were superior. In the 1970s,11 cross-combinations between UA andTrinitario varieties were selected for yield anddistributed in the form of seeds produced inbi-clonal seed gardens (Liabeuf, 1968; NyaNgatchou, 1985). As the main problem inCameroon is black pod, caused byPhytophthora megakarya, particular efforts
(5) ICS: Imperial College Selection.(6) TSH: Trinidad Selection Hybrid.
Retour au menu

17CO C O A
Novembre - Décembre 1995 Plantations, recherche, développement
were made to evaluate resistance to thisdisease. Around a hundred clones wereclassified by artificial pod inoculation (Blahaand Lotodé, 1976). The heritability of fieldresistance was studied in a diallel crossingscheme, which showed that pod ripening timemay also be a component of field resistance(Despréaux et al., 1989; Berry and Cilas,1994).
The substantial variability observed in full-sib families can be used to optimize geneticgains. Selecting individuals amongstprogenies takes account of the non-additivepart of the genetic variance component. Inboth Cameroon and Côte d’Ivoire, largenumbers of hybrid trees are being observed.Combined individual-family selectionaccording to indices based on estimates of thegenetic value for yield, vigour, canopy size andsusceptibility to black pod is currently beingcarried out in Cameroon and is due to belaunched shortly in Côte d’Ivoire. Substantialprogress is expected for certain characterssuch as black pod resistance (Cilas andPaulin, 1994). The individuals selected caneither be vegetatively propagated or used asnew parents.
Recurrent selectionConventional clone hybrid selection has itslimitations, since it does not ensureconsistent genetic progress, and theimprovement of quantitative characters suchas disease resistance is not very effective. Toincrease the frequency of genotypes thatcombine several favourable genes, it has beensuggested to improve base populations byrecurrent selection. A scheme of this type waslaunched by Cirad(7) and Idefor in Côted’Ivoire in 1990, using LA and Trinitario (UA-0) and UA (UA-0) as base populations. Theparents were chosen for theirphenotypic/genotypic value and representati-veness of the genetic diversity within thegroups. The Trinitario materials are beingused to introduce quality genes into the LAForastero population. In the first phase, tworecurrent selection cycles should lead to thecreation of improved populations. Combinedindividual-family selection will be applied.Right from the first cycle, this scheme willpermit variety selection (hybrids or clones)obtained within each population. Reciprocalrecurrent selection between the twopopulations will then be started. During thisphase, the possibility of producing between-population hybrids will also be looked at,since this would make use of any heterosis
effect expressed by this type of crosses (figure3). The crossing schemes applied in the firstcycle are diallel and factorial; in subsequentcycles, partial crossing schemes will beapplied to increase the chances ofrecombination and prevent genetic erosion(Clément et al., 1994; Eskes et al., 1995).
Bal Plantations in Sabah, Malaysia, iscurrently also developing a geneticimprovement programme based on recurrentselection. The programme consists inalternating clone trials and progeny trials,using factorial crossing schemes between UAgenotypes. The planned varietal outputs areclones (grafted plants), and also hybrids(seeds). The best first-cycle hybrids haveproved to be 20 to 50% more productive thanthe controls. Interesting information hasalready been obtained about the geneticparameters of the UA population: significantcorrelations between phenotypic andgenotypic values and predominance of generalcombining ability (Lockwood and Pang, 1993;Lockwood and Pang, 1994).
Conclusion and prospects The most widely used cocoa breeding methodis still selection of bi-parental crosses(hybrids). The hybrid varieties producingbulk cocoa that have been bred over the pastthirty years or so have proved suitable formost regions, particu-larly pioneer productionregions. For field establishment, precocity andyield, this breeding method has generallyresulted in a substantial improvementcompared to local populations. However,disease resistance and homogeneity still needto be improved.
Characterizing and evaluating collectionsin greater detail should enable more effectiveuse of genetic diversity. With the use ofmolecular markers, it may be possible toidentify links between genetic distancesbetween parents and the value of the hybrids,be they «between-group» or «within-group».The level of hetero-zygosity of the parentscould also be taken into account to improvehybrid homogeneity. The predominance ofgeneral combining ability for many characterscan be used to accumulate the genes involvedin quantitative characters, throughrecombinations. The detection of molecularmarkers linked to agronomically valuablecharacters (QTL) could make breeding moreeffective in the near future (marker-assistedselection).
Certain research centres are currentlyusing new breeding strategies. Clone selectionwithin hybrid families should ensure rapidgenetic progress, particularly for diseaseresistance and homogeneity. If somaticembryogenesis were to prove reliable, in vitromultiplication will permit that superior clonescan be distributed rapidly as orthotropicplants. Improving populations by recurrentselection will ensure substantial geneticprogress in the long term. In response to thespecific demand for fine cocoas, therevalorization and genetic improvement oftraditional Criollo, Nacional and Trinitariopopulations are also on the agenda. The aim isto improve the agronomic properties of thesevarieties, whilst maintaining their flavourquality characters. Desired characters couldbe introduced by backcrossing with genotypesof UA origin.
(7) CIRAD: Centre de coopération internationale enrecherche agronomique pour le développement.
The Criollo group was grown in prehistoric times in Mexico and Central America. The Mayasdomesticated it to use its beans both for consumption and as currency. At the start of the17th century, its cultivation spread to the Caribbean (Jamaica, Haiti, Trinidad) and SouthAmerica (Venezuela). Spanish settlers exported the crop to Indonesia in 1560 and thePhilippines in 1670. In the 18th century, in Brazil, Forastero cultivation, which had initiallybeen concentrated in Para State, spread to Bahia and developed rapidly. In Ecuador, a localcocoa type, «Nacional», was grown as early as the end of the 17th century, and the countrywas the world’s largest producer throughout the 19th century. Between 1822 and 1878,Forastero and then Trinitario trees were introduced in West and Central Africa from SouthAmerica by the Portuguese. The crop expanded spectacularly in Cameroon and Nigeria(Trinitario), in Ghana and above all in Côte d’Ivoire (Amelonado Forastero), which is now theworld’s largest producer. Cocoa cultivation in Asia developed late, but very rapidly, from theturn of the century in Indonesia (Java and Sumatra), with Criollo and then Forasterovarieties, and then in Malaysia (Sabah) from the 1950s onwards, with Amelonado andUpper Amazon Forastero varieties. World cocoa growing thus spread from West to East,with boom periods in America, South America, Africa and now Southeast Asia. At the sametime, production in Central America, which had been the main zone until the 18th century,declined steadily (Jarrige and Ruf, 1990).
■ History of growing cocoa
Retour au menu

Few research centres worldwide arecurrently capable of maintaining a long-termbreeding programme. The current economicsituation in the cocoa sector, which has beenunfavourable for the past decade or so, is
largely responsible for it. However, thissituation may improve and there is nowrenewed interest in cocoa geneticimprovement. The links between cocoabreeders were strengthened by the creation in
1994 of INGENIC, the International Group for
the Genetic Improvement of Cocoa, which
should stimulate international cooperation on
cocoa breeding. ■
18 CO C O A
Plantations, recherche, développement Novembre - Décembre 1995
RésuméL’amélioration génétique du cacaoyerdébute par la domestication des Criollo enAmérique centrale. Ils sont progressive-ment remplacés par les sélections de Trini-tario, puis par les Forastero, plus résistantsaux maladies et aux insectes. En 1940, descroisements entre Forastero, prospectés enHaute-Amazonie, et d’autres groupes géné-tiques, ont mis en évidence des gains signi-ficatifs de précocité et de productivité. Deshybrides entre groupes génétiques diffé-rents sont alors largement vulgarisés.Depuis les années 80, l’hybridation entrehaploïdes doublés, le choix des géniteursd’après les paramètres génétiques, la sélec-tion récurrente et la sélection sur indexd’individus dans les meilleurs descen-dances, sont mises en oeuvre. Les mar-queurs moléculaires (RFLP et RAPD) ontmieux éclairé la diversité génétique et per-mis de cartographier le génome. La résis-tance aux maladies et aux insectes, ainsique la qualité sont actuellement les objec-tifs majeurs. Une évaluation efficace dumatériel génétique, la sélection récur-rente, la sélection assistée par marqueurset la micropropagation pourraient contri-buer à les atteindre.
AbstractGenetic improvement of cocoa began withthe domestication of Criollo varieties inCentral America. They were graduallysuperseded by Trinitario selections, then byForastero trees, which have better diseaseand insect resistance. In 1940, crossingForasteros from a survey in Upper Amazo-nia with other genetic groups resulted insubstantial increases in precocity and pro-ductivity. Hybrids between different geneticgroups were distributed widely. Since the1980s, there has been a move towardshybridizing double haploids, choosingparents based on genetic parameters,recurrent selection and index selection ofindividuals amongst the best progenies.Molecular markers (RFLP and RAPD) haveprovided a clearer picture of genetic diver-sity and enabled genome mapping. Resis-tance to diseases and insects and improvedquality remain major breeding objectives.More efficient evaluation of germplasm,recurrent selection, marker-assisted selec-tion and micropropagation may contributeto solving these problems.
ResumenEl mejoramiento genético del cacaoempieza por la domesticación de los Criolloen América central. Se sustituyen progresi-vamente por las selecciones locales de Tri-nitario, y luego por los Forastero, más resis-tentes a las enfermedades y a los insectos.En 1940, cruzamientos entre Forastero,prospectados en Alta Amazonia, y otros gru-pos genéticos, evidenciaron gananciassignificativas de precocidad y de productivi-dad. Entonces, se divulgaron ampliamentehíbridos entre grupos genéticos diferentes.Desde los años 80, se pusieron en ejecuciónla hibridación entre haploides doblados, laselección de los genitores según los pará-metros genéticos, aplicándose la selecciónrecurrente e índices de selección de indivi-duos en las mejores descendencias. Losmarcadores moleculares (RFLP y RAPD)han aclarado mejor la diversidad genética ypermitido realizar mapas del genoma. Laresistencia a las enfermedades y a los insec-tos y la qualidad son actualmente objectivosmayores. Una evaluación eficaz de losrecursos genéticos, la selección recurrente,la selección asistida por marcadores y lamicropropagación podrían contribuir aresolverlos.
Retour au menu


















